
Morphological and systematic re-assessment of the late Oligocene “Halitherium” bellunense reveals a new crown group genus of Sirenia
MANJA VOSS, SILVIA SORBI, and DARYL P. DOMNING
Voss, M., Sorbi, S., and Domning, D.P. 2017. Morphological and systematic re-assessment of the late Oligocene “Halitherium” bellunense reveals a new crown group genus of Sirenia. Acta Palaeontologica Polonica 62 (1): 163–172.
“Halitherium” bellunense is exclusively known from a single individual from upper Oligocene glauconitic sandstone near Belluno, northern Italy. According to a review of its morphological basis, which consists of associated cranial elements, some vertebrae and ribs, this specimen is identified as a juvenile, because the first upper incisor (I1) and supposedly second upper molar (M2) are not fully erupted. However its juvenile status allowed only cautious conclusions on its taxonomy and systematic affinity. The presence of a nasal process of the premaxilla with a broadened and bulbous posterior end, and a lens-shaped I1, corroborate an evolutionarily-derived status of this species that places it well within the sirenian crown group Dugonginae. Considering these new data and in order to avoid continued misuse of the inappropriate generic name of Halitherium, a new generic name, Italosiren gen. nov., and emended species diagnosis are supplied for this taxon.
Key words: Mammalia, Tethytheria, Sirenia, Dugonginae, evolution, Oligocene, Italy.
Manja Voss [manja.voss@mfn-berlin.de], Museum für Naturkunde, Leibniz Institute for Evolution and Biodiversity Science, Invalidenstraße 43, 10115 Berlin, Germany.
Silvia Sorbi [silviasorbi@gmail.com], Museo di Storia Naturale, Università di Pisa, Via Roma 79, 56011 Calci, Pisa, Italy.
Daryl P. Domning [ddomning@Howard.edu], Laboratory of Evolutionary Biology, Department of Anatomy, Howard University, Washington, District of Columbia 20059,USA.
Received 22 June 2016, accepted 12 December 2016, available online 6 March 2017.
Copyright © 2017 M. Voss et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
The Dugonginae are one of the most diverse groups amongst Sirenia, or sea cows. They comprise a single living species today, the dugong (Dugong dugon) from the Indo-Pacific region, but, in contrast, their fossil record suggests a high taxonomic diversity worldwide, often characterised by sympatric offshoots (Domning 2001; Vélez-Juarbe et al. 2012). Crenatosiren olseni from the Oligocene of the southeastern USA is consistently regarded as the most plesiomorphic representative of this group (e.g., Domning 1994, 1997; Bajpai and Domning 1997; Vélez-Juarbe et al. 2012; Vélez-Juarbe and Domning 2015), indicating that this subfamily apparently had its origin in the West Atlantic and Caribbean region. In this region, dugongines reached their highest diversity from the late Oligocene until the early Pliocene, documented by taxa like Dioplotherium (Domning 1978, 1989a; Toledo and Domning 1989), Xenosiren (Domning 1989b), Nanosiren (Domning and Aguilera 2008), Callistosiren (Vélez-Juarbe and Domning 2015), and Corystosiren (Domning 1990). Considering sea cows as predominantly feeding on seagrasses, most of these genera evolved large, blade-like and self-sharpening tusks, which in contrast to the smaller, sub-conical tusks of Miocene taxa like Metaxytherium, may have been used to dig up rhizomes of robust seagrasses (Domning and Beatty 2007). Fossil dugongine members outside the West Atlantic and Caribbean region are known by the diverse Oligocene–Miocene sirenian fauna from India with Bharatisiren (Bajpai and Domning 1997; Bajpai et al. 2006), Kutchisiren (Bajpai et al. 2010) and Domningia (Thewissen and Bajpai 2009), and also from the early Miocene of France and Libya by taxa of the genus Rytiodus (Lartet 1866; Delfortrie 1880; Domning and Sorbi 2011).
Although the dugongine fossil record includes a number of species established on the basis of well-preserved specimens, the phylogenetic interrelationships within this group are unresolved. This is most evident by the different topographies of the Dugonginae in each of the recent cladistic approaches, such as Domning (1994), Bajpai and Domning (1997), Sorbi (2008), Vélez-Juarbe et al. (2012), Vélez-Juarbe and Domning (2015), and Springer et al. (2015). Whereas other parts of the sirenian phylogeny are pretty stable, a reliable dugongine phylogeny is not available at present. Nevertheless, the monophyly of this group is well supported by synapomorphies referring to the morphology of the premaxillary nasal process, the supraorbital process of the frontal, and the preorbital process of the jugal, for example (e.g., Domning 1994; Domning and Aguilera 2008; Vélez-Juarbe and Domning 2015).
In this study, we conclude that a further species from Italy hitherto known as “Halitherium” bellunense De Zigno, 1875 belongs to the European dugongine assemblage. This taxon has long been regarded as being part of the “Halitherium”-species assemblage, which however, comprises taxa that are less derived than the Dugonginae (Voss 2013). Moreover, from its establishment and for much of the ensuing century, the genus Halitherium Kaup, 1838 served as a catchall for poorly-known sirenian fossils from various parts of the world (Domning et al. 2010). For the reasons mentioned above, the need for revision of this genus has become clear, nowhere more so than in the case of “H.” bellunense. Although Domning (1989a, 1996) already postulated dugongine affinities for “H.” bellunense, a hypothesis that was recently corroborated by Sorbi (2008) and Voss (2013), its nomenclatural status is unchanged until today. Therefore, the present paper aims at testing Domning’s (1989a, 1996) initial hypothesis by providing a morphological re-evaluation of the material assigned to “H.” bellunense.
Although the ideal would be to provide a phylogeny showing the systematic position of “H.” bellunense, we refrain from doing so for the above-mentioned reason of the topological instability of the dugongine clade. Since the focus of the present paper does not rest on issues such as taxon sampling and character choice for clarifying this instability, we instead point out that “H.” bellunense unambiguously belongs to the Dugonginae by discussing some possible systematic alternatives on the basis of previous cladistic approaches from Sorbi (2008) and Voss (2013). A phylogenetic analysis of Sirenia as part of a broader framework and focussing on all hitherto known valid taxa, including the new genus established here, is in progress and supposed to be published at a later stage once the genus Halitherium is revised. Hence, the present study primarily addresses the morphology and revised taxonomy of “H.” bellunense by establishing a new and clearly defined genus as well as an up-to-date synonymy.
Institutional abbreviations.—JCU, James Cook University, Townsville, Queensland, Australia; MGPD, Museo dell’Istituto di Geologia e Paleontologia, Padova, Italy.
Other abbreviations.—DP1–5, upper deciduous premolar or alveolus 1–5; I1, first upper incisor; lSYM/lPM, ratio for the value of the symphysis length divided by the total length of the premaxilla; M1–3, upper molar or alveolus 1–3.
Geological setting
According to De Zigno (1875), “Halitherium” bellunense was found in Cavarzana, near Belluno in northern Italy (Fig. 1), the spelling of which was emended to Cavarzano in Sorbi (2008). The holotype was embedded in dark green, glauconitic, and shell-rich sandstone overlain by grey sandstone layers. The glauconitic sandstone was considered to be early Miocene in age (De Zigno 1875) and this stratigraphic assignment was maintained until quite recently (e.g., Lepsius 1882; Abel 1905; Pilleri 1985; Domning 1996). On the basis of our review of the geology, the stratigraphic provenance of “H.” bellunense is now determined to be Chattian (upper Oligocene). Today, these sediments are identified as belonging to the lowermost sequence (upper Chattian–lower Aquitanian) of the Venetian Molasse Basin in the Belluno syncline, between the Trento Plateau in the northwest and the Friuli Platform in the east (Bosellini et al. 1981; Dallanave et al. 2009). The transgressive systems tract of the lower sequence comprises a basal, condensed, glauconitic and fossiliferous sand sheet, the so-called Belluno Glauconitic Sandstone, which corresponds to De Zigno’s (1875) type horizon of “H.” bellunense. The top of the transgressive event is represented by the lower part of the Bastia Siltstone, De Zigno’s (1875) overlying grey sandstone that represents a finer-grained, essentially muddy unit.
The determination of the type stratum of “H.” bellunense is uncertain, but can be limited. According to Ghibaudo et al. (1996), the Belluno Glauconitic Sandstone is 11m thick and can be subdivided, from base to top, into a shell-rich, gravelly sandstone facies followed by sandstone characterised by an alignment of bioclasts, which are topped by a bioturbated sandstone facies. Only the first and second facies of the Belluno Glauconitic Sandstone contain bioclasts, mainly pectinids, and therefore have relevance as to the type stratum of “H.” bellunense. De Zigno (1875) did not mention the shell-pavements characteristic of the 68 cm thick second facies. Based on that, it could be inferred that “H.” bellunense comes from the first facies, which ranges in thickness 15–35 cm depending on the depth of infilled incisive shallow channels in the underlying turbidite substrate. Either way, this stratigraphic distinction is only subtle and the type stratum can be limited at least to the basal 83 cm to 103 cm of the Belluno Glauconitic Sandstone.
Both units, the Belluno Glauconitic Sandstone and the lower part of the Bastia Siltstone, are shown as representing the upper section of the Chattian stage (Ghibaudo et al. 1996). This precise and clearly defined stratigraphic determination is supported by the subsidence analysis of Mellere et al. (2000), who date the molasse deposits of the Belluno Glauconitic Sandstone to 27–25 Ma and the Bastia Siltstone to 25–22.5 Ma on the basis of planktonic foraminifera. Foraminiferal assemblages and facies associations also reveal both units to represent deposits of a shallow marine environment, placing paleobathymetrically the Belluno Glauconitic Sandstone in the inner neritic and the Bastia Siltstone in the offshore inner-shelf level (Mellere et al. 2000). In conclusion, “H.” bellunense comes from the basal part of the Belluno Glauconitic Sandstone Formation and is late Chattian in age.
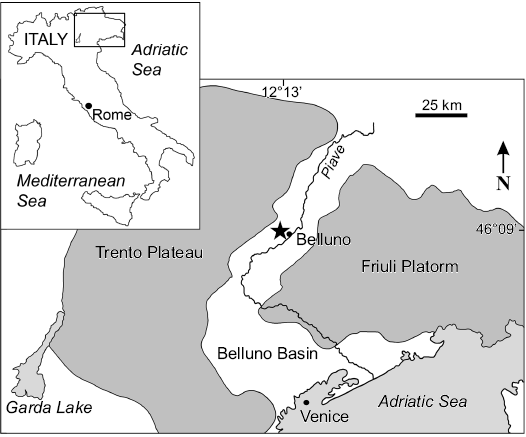
Fig. 1. Geographic location of Cavarzano, Valle delle Guglie, near Belluno, Italy (modified after Bosellini et al. 1981 and Dallanave et al. 2009). Asterisk indicates estimated type locality of Italosiren bellunensis.
Material and methods
“Halitherium” bellunense is known from a single specimen comprising the skullcap, left premaxilla, maxillary fragments including teeth, zygomatic processes of the squamosals, a jugal fragment, and remains of vertebrae and ribs representing a juvenile, which is stored in the MGPD. The premaxilla was originally embedded in sedimentary matrix medial to the rostrum and posteriorly, partly surrounding the rear end of the nasal process. Since this element provides critical characters for assigning this species to the Dugonginae, it was further prepared and large parts of the matrix were removed by Letizia Del Favero (MGPD), which greatly enhanced its morphological interpretation. The macroscopic descriptions of the personally investigated holotype consider the preserved cranial and postcranial osteology which provides the basis for the determination of the species’ systematic affinities. Measurements were carried out with either a digital caliper of standard size (max. 150 mm) or a larger caliper (up to 200 cm). All measurements are quoted in millimetres, unless otherwise stated, and listed in Tables 1, 2. We follow standard convention in abbreviating tooth families as I, C, P, and M, with upper and lower case letters referring to upper and lower teeth, respectively. Images were taken with a digital camera in all relevant views, using the macro-function under normal light. Line drawings of skeletal elements in different views complement the documentation of “H.” bellunense. We follow standard convention in abbreviating tooth families as I, C, P, and M, with upper and lower case letters referring to upper and lower teeth, respectively.
Table 1. Measurements (in mm if not otherwise stated) of the cranium of Italosiren bellunensis (MGPD-18Z–23Z, 7384Z). Letters in parentheses denote standard dimensions established by Domning (1978: fig. 7, table 2). [], preserved lengths; “e” estimated dimensions; l, left; r, right.
|
Length of premaxillary symphysis (AH) |
[94] |
|
Length of premaxilla |
[178] |
|
Height of jugal below orbit (ab) |
40 |
|
Length of mesorostral fossa (HI) |
80e |
|
Anteroposterior length of zygomatic-orbital bridge of maxilla (no) |
34 |
|
Dorsoventral thickness of zygomatic-orbital bridge of maxilla (T) |
17 |
|
Length of zygomatic process of squamosal (OP) |
101l, 99r |
|
Dorsoventral height of zygomatic process of squamosal (WX) |
43l+r |
|
Anteroposterior length of root of zygomatic process of squamosal (QR) |
[53] |
|
Length of parietals, frontoparietal suture to rear of external occipital protuberance (P) |
[77] |
|
Width of cranium at frontoparietal suture (GG’) |
[65] |
|
Minimum width of parietals |
63 |
|
Maximum width of parietals |
79 |
|
Maximum height of rostrum (KL) |
[54] |
|
Height of supraoccipital (HSo) |
[61] |
|
Width of supraoccipital (WSo) |
[83] |
|
Deflection of masticating surface of rostrum from occlusal plane (RD) |
60°e |
|
Angle between supraoccipital and parietal |
115° |
Table 2. Linear dimensions (in mm) of cheek teeth of Italosiren bellunensis (MGPD-20/21Z). [], preserved crown length; e, estimated dimensions.
| |
Length |
Anterior width |
Posterior width |
|
DP5 |
[21] |
21e |
18.5e |
|
M1 |
21.9 |
22 |
19.5 |
|
M2 |
22.5 |
22.7 |
20 |
Systematic palaeontology
Class Mammalia Linnaeus, 1758
Superorder Afrotheria Stanhope, Waddell, Madsen, de Jong, Hedges, Cleven, Kao, and Springer, 1998
Mirorder Tethytheria McKenna, 1975
Order Sirenia Illiger, 1811
Family Dugongidae Gray, 1821
Subfamily Dugonginae Gray, 1821
Genus Italosiren nov.
Type species: Halitherium bellunense (De Zigno, 1875), monotypic; see below.
Etymology: From Latin Italia, Italy; and siren, siren (f.).
Diagnosis.—As for the type species by monotypy.
Stratigraphic and geographic range.—Upper Chattian (latest late Oligocene), northern Italy.
Italosiren bellunensis (De Zigno, 1875) comb. nov.
Fig. 2.
1875 Halitherium bellunense sp. nov.; De Zigno 1875: 438, pls. 14–15.
1878 Halitherium bellunense De Zigno, 1875; De Zigno 1878: 68.
1882 Metaxytherium bellunense (De Zigno, 1875); Lepsius 1882: 180.
1887 Halitherium bellunense De Zigno, 1875; De Zigno 1887: 728.
1905 Halitherium bellunense De Zigno, 1875; Abel 1905: 393, text-fig. 1.
1987 Halitherium bellunense De Zigno, 1875; Domning and Thomas 1987: 207.
1989a Halitherium bellunense De Zigno, 1875; Domning 1989a: 425, text-fig. 6.
Holotype: MGPD-18Z skullcap, MGPD-19Z left premaxilla with tusk, MGPD-20/21Z fragment of left maxilla with DP5–M2, MGPD-22Z left and MGPD-23Z right zygomatic processes of squamosals, MGPD-7384Z fragment of left jugal, MGPD-7385/6Z fragment of right maxilla with DP5 and M1, MGPD-7387Z an isolated fragment of a molar, MGPD-7383Z a fragment of a lower molar, seven fragments of vertebrae (MGPD-7362Z, 7367Z, 7368Z, 7369Z, 7374Z, 7375Z, 7376Z), and six rib fragments (MGPD-7358/9Z, 7363Z, 7364Z, 7366Z, 7381Z) of a juvenile individual.
Type locality: Cavarzano, Valle delle Guglie, near Belluno, Italy.
Type horizon: Upper Chattian (latest late Oligocene), 27–25 Ma; basal portion of the Belluno Glauconitic Sandstone Formation.
Material.—Only the type material.
Emended diagnosis.—Dugongine sirenian having the following unique combination of synapomorphies: nasal process of premaxilla broadened and bulbous at posterior end, having more or less vertical joint surface in contact with frontal (shared with Dioplotherium, Xenosiren, and Rytiodus heali); zygomatic-orbital bridge of maxilla shortened and thickened posteriorly, but not transformed into vertical wall (shared with Dugong and Nanosiren); ventral extremity of jugal lies ventral to orbit (shared with all dugongines except for Xenosiren, Crenatosiren, and Nanosiren); processus retroversus of squamosal moderately inflected (shared with Crenatosiren, Callistosiren, Rytiodus, Kutchisiren, Dioplotherium allisoni, and Bharatisiren indica); supraoccipital wider ventrally than dorsally (shared with Nanosiren, Dugong, Kutchisiren, and Bharatisiren kachchhensis). Differs from various other Dugonginae in retaining the following unique combination of plesiomorphies: frontal roof flat without knoblike bosses medial to temporal crests (character combination shared with Bharatisiren indica); squamosals do not reach temporal crests (shared with Corystosiren and Dioplotherium cf. allisoni from Brazil); ventral rim of orbit not overhanging (shared with Corystosiren and Callistosiren); first upper incisor (I1) present with I1 crown having enamel on all sides (shared with Crenatosiren, Nanosiren sanchezi, and Dioplotherium manigaulti). Differs further from the similar genus Rytiodus in lacking a distinct intertemporal constriction of the cranial roof, an overhanging orbital rim of the jugal, large and blade-like tusks, and differs at least from R. heali in having a supraoccipital that is only slightly wider than high.
Description.—This specimen was previously described and illustrated by De Zigno (1875) and Abel (1905). Here, the holotype material is re-described on the basis of a re-investigation contributing to new and more comprehensive indications of the taxon’s skeletal morphology. The anatomical terminology mentiond here follows Domning (1978).
Premaxilla: The left premaxilla (Fig. 2A) is almost completely preserved and measures 178 mm in total length with the symphysis reaching a length of 94 mm. The value of the symphysis length divided by the total length of the premaxilla, i.e., the ratio lSYM/lPM, is 0.53, indicating that the symphysis is longer than half the total length of the premaxilla. Taking into account the conditions in other dugongine taxa with similar ratios like Rytiodus capgrandi or Crenatosiren olseni, the symphysis is considered to be enlarged relative to the cranium. In lateral view (Fig. 2A1), the premaxillary symphysis is compressed to form a low middorsal ridge with a slight boss at the summit. The first incisor tusk (I1) is preserved in the symphysis, indicating that a medial dentiform process like that in Hydrodamalis is missing. In lateroposterior view, the maxillopremaxillary suture is indicated and perpendicular to the posterior end of the symphysis. Although the angle of deflection of the rostrum cannot be unambiguously determined, it is estimated to be strong, about 60°, as in most other dugongines. Anteromedial to the nasal process, the mesorostral fossa shows no indentations and extends, retracted and enlarged, in a dorsoposterior direction. Whether the external nares surpass the level of the anterior margin of the orbit cannot be determined due to the missing facial part of the skull. The posterior end of the nasal process is broadened and bulbous, having a posterolateral oblique joint surface in contact with the frontal and/or nasal (Fig. 2A2).
Frontal: MGPD-18Z preserves the posterior part of the frontal roof, which is flat without knoblike bosses (Fig. 2B1). The dorsolateral edges that presumably bore the temporal crests are broken. The interior aspect of the preserved skullcap remains undescribed due to adhering sediment.
Parietal: The parietal roof (Fig. 2B1) is more or less flat and slightly concave centrally where the temporal crests reach their maximum height and constriction. Generally, the intertemporal constriction is weak, with the temporal plane of the parietal sloping as a flat wall laterally. The temporal crests form prominent and thickened lyriform keels that bulge medially at the centre of the parietal roof. They diverge and decrease in height anteriorly and posteriorly. The course of the temporal crests is not interrupted by the squamosals, so they reach the nuchal crest. Anteriorly, the frontal processes of the parietal are separated by an angle of about 90°, indicating the course of the frontoparietal suture, and presumably extended only a short distance on the frontal roof.
Supraoccipital: Small parts of the supraoccipital are missing on the left dorsolateral and dorsomedial sides, and its left and right thirds are separated from the mid-portion by fractures (Fig. 2B2). Nevertheless, it can be determined to form a compact wall, with a width/height ratio of about 1.36, which indicates that the supraoccipital is only slightly wider than high. It is wider ventrally than dorsally, and, in lateral view, it meets the parietal roof at an angle of about 115°. The nuchal crest is narrow and sharp-edged, and extends as a convex ridge dorsolaterad. Though partially eroded, the nuchal crest shows no notch in the median plane, but rather an external occipital protuberance that forms a prominent knob rising above the parietal roof. The muscle insertions for the attachment of the semispinalis capitis muscles are rounded. The external occipital protuberance extends ventrad as a distinct but low median ridge, somewhat shifted to the left side. It exceeds half the height of the supraoccipital and extends almost to its ventral tip. The ventral margin is pointed in the median plane, with the exoccipital sutures meeting at an angle of about 132°.
Maxilla: Two maxillary fragments associated with the cheek dentition are preserved. On the lateral side of the left fragment (MGPD-20/21Z; De Zigno 1875: pl. 15: 4), the broken base of the zygomatic-orbital bridge is observable, indicating that it lies slightly above the alveolar margin, elevated about 10 mm. Where broken, the bridge is dorsoventrally thicker posteriorly (17 mm) than anteriorly (12 mm). According to its length (about 34 mm) and height (17 mm), the zygomatic-orbital bridge is evaluated as being shortened (thickness ≥ 0.40 × length) anteroposteriorly, but not transformed into a transverse vertical wall.
Squamosal: The cranial portion of the squamosal is not preserved, but its attachment areas posterolateral to the parietal show that it did not extend to the temporal crests (Fig. 2B1). Only the left and right zygomatic processes (MGPD-22Z and 23Z; Fig. 2C, D) are present, with the left one having preserved the posterolateral part of the zygomatic root, indicating a distinct notch (Fig. 2D). The processus retroversus is moderately inflected without projecting below the jugosquamosal suture (Fig. 2C). In lateral view, the zygomatic process has a flat surface and is roughly triangular in shape, dorsoventrally high posteriorly and tapering anteriorly. Its posterodorsal end is convex in outline, the ventral margin is straight, except for a distinct convexity just in front of the processus retroversus. Medially, the zygomatic process is slightly concave with the dorsal margin inclined inward, forming a sigmoid margin.
Jugal: The middle part of the left jugal is preserved (Fig. 2C) with the preorbital and zygomatic processes present, but broken anteriorly and posteriorly, respectively. The zygomatic process may have been as long as the diameter of the orbit, judging from the corresponding attachment area on the ventral margin of the zygomatic process of the squamosal, which reaches as far posteriorly as the beginning of its ventral convexity. However, the orbital area of the skull is too incompletely preserved for an unambiguous conclusion on that character. Ventral to the orbit, the dorsal margin is thick, but not overhanging laterally. The ventralmost extremity is positioned under the orbit. Anterior to the tip of the squamosal zygomatic process, the postorbital process bulges upwards slightly, but without rising high enough to form a postorbital bar with the frontal.
Mandible: A bone fragment of about 40 mm in its maximum dimensions is referred to the lower jaw due to the presence of a two-rooted molar or molariform premolar (MGPD-7383Z). The tooth crown is only fragmentarily preserved, hampering any detailed descriptions.
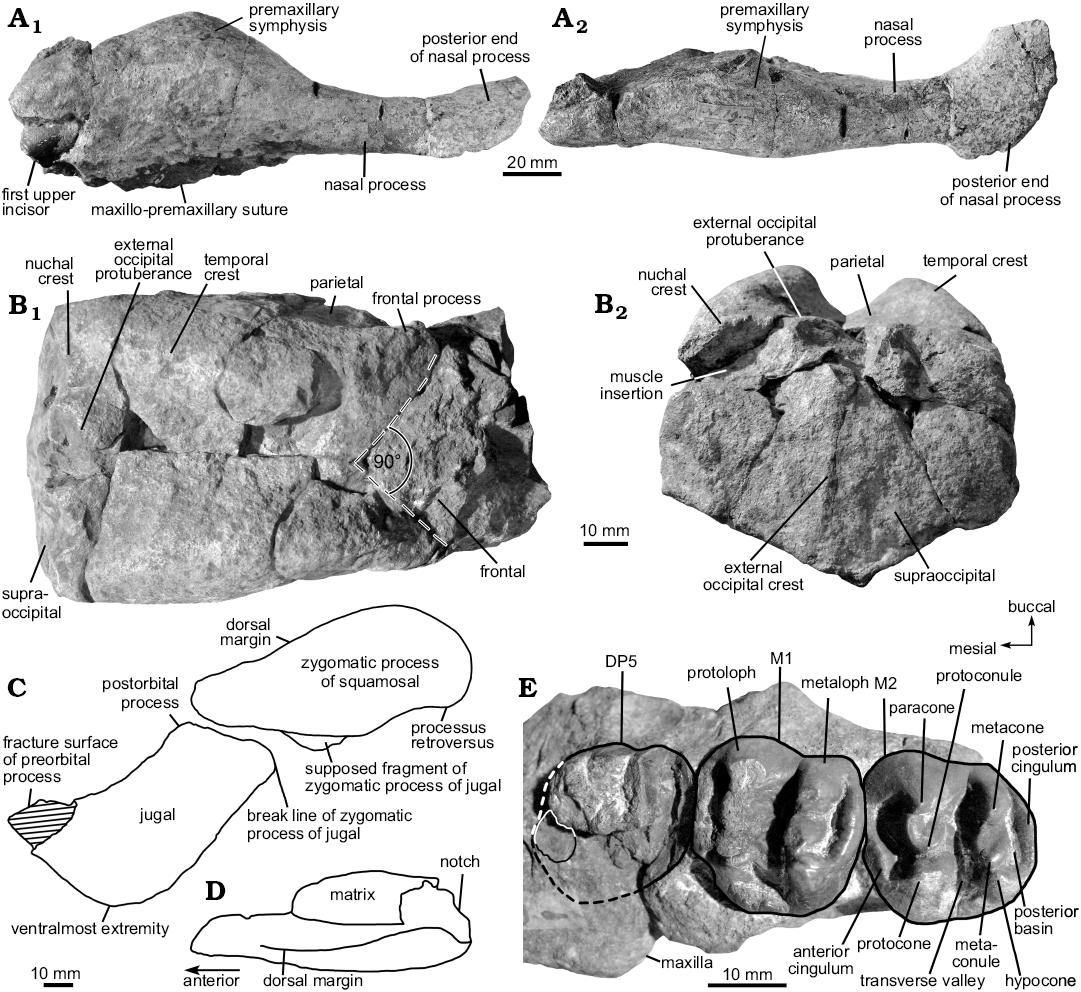
Fig. 2. Cranial and dental elements of the dugongine sirenian Italosiren bellunensis (De Zigno, 1875) comb. nov. from Cavarzano, Italy; upper Chattian, late Oligocene. A. Left premaxilla (MGPD-19Z) in lateral (A1) and dorsal (A2) views. B. Parietal-supraoccipital skullcap (MGPD-18Z) in dorsal (B1) and posterior (B2) views. C. Outline drawing of left zygomatic process of squamosal (MGPD-22Z) and jugal (MGPD-7384Z) in lateral view; shaded area indicates missing part. D. Outline drawing of left zygomatic process of squamosal (MGPD-22Z) in dorsal view. E. Left maxillary fragment (MGPD-20/21Z) in occlusal view; dashed lines indicate the estimated outline of DP5, outlined area indicates portion of controversial interpretation and is considered here as representing remnants of the DP5 crown.
Dentition: Besides two poorly preserved dental remains (MGPD-7383Z and 7387Z) that do not provide any information, morphologically valuable records are the left premaxilla including the first incisor tusk and the left and right maxillary fragments. The upper dental formula is interpreted to be I1, C0, P0, DP5, M1–3.
De Zigno (1875: pl. 15: 1) illustrated the first incisor tusk (I1) as being complete and unworn with a lateral exposure in the premaxilla of about 30 mm. Though the anteriormost tip of about 10 mm is broken and missing today (Fig. 2A1), the original state of preservation reveals this tusk to be most likely unerupted. In cross section, the crown is lens-shaped with sharp anterior and posterior edges, measuring 17 × 8 mm in diameters. The crown is covered by black enamel on all sides, but no conclusions can be drawn on the extent of the enamel, i.e., either along the entire length of the tusk or being absent from a distinct root. The slightly swollen lateral sides of the premaxillary symphysis indicate a length of the tusk alveolus that is not restricted to less than half the length of the premaxillary symphysis. However, whether the alveolus extends about half the length of the symphysis or even exceeds that level remains uncertain. The clarification of these characters must wait until more and better preserved material is known, particularly from adult specimens, because the extent of the I1 alveolus increases with the individual’s age, as can be observed in the living dugong (Dugong dugon) (Marsh 1980; personal observations by MV, SS, DD). In any case, interpretation of an expanded nasal process of the premaxilla as indicating forceful use of large tusks (Domning and Beatty 2007) argues in favour of the I1 being large in adult Italosiren.
The cheek dentition is qualitatively and quantitatively best preserved in the left maxillary fragment MGPD-20/21Z, and therefore this is the basis for the following description (Fig. 2E). Three cheek teeth are preserved, but their identification is uncertain; two possible interpretations are considered here:
(i) They may represent DP5, M1, and M2 (opinion of MV). Abel (1905) and Sorbi (2008) interpret a small rounded tooth fragment mesiolingual to the incomplete DP5 crown (Fig. 2E: outlined area) as remains of the fifth premolar or the fourth deciduous premolar, respectively. Here, this tooth fragment, which is mesially slightly elevated above the level of the bone, is considered to be part of the irregularly broken DP5 crown. This observation is additionally supported by the original extent of the DP5 alveolus still discernible mesially, lingually and distally by imprints in the maxillary bone, and by De Zigno (1875: pl. 15: 3), who illustrates the anteriormost “molar” as a single tooth. As shown in Fig. 2D, the area that might have been covered by the complete crown includes that fragment, which additionally corresponds to the tight positioning of the cheek teeth that barely would have given space for a further premolar at this place. Accordingly, DP5 is only slightly smaller than the subsequent molars (Table 2) and similar in having a slightly heart-shaped crown as also can be generally observed in related taxa. The fracture surfaces of the crown provide insights to the broken lingual and distolabial roots, identifying this tooth as a three-rooted molariform or submolariform premolar.
(ii) Alternatively, these three teeth may represent DP4, DP5, and M1 (possibility suggested by DD). If the fragment mesiolingual to the anterior tooth is part of its broken crown, then this crown was longer than either of the more posterior teeth, not smaller. However, Abel (1905) regards the fragment as part of a root; and De Zigno’s (1875: pl. 15: 4) shows an overhang of the crown at the front end, suggesting that the fragment is level with the bone and not high enough to be part of the crown. However, a molariform tooth has a single large root on the lingual side, not a small mesiolingual one (see also the cross section of this tooth in Zigno’s 1875: pl. 15: 7). Furthermore, the crown of the anterior tooth seems relatively long and narrowed toward the front, so perhaps not heart-shaped, even though it is so worn that its length is significantly diminished. It could therefore be the submolariform DP4, which should be proportionately longer than the fully molariform DP5 or M1. That would make the following two teeth DP5 and M1. Then the fragment might be a remnant (root) of DP3 (P5 of Abel 1905) that has been encroached on by mesial drift of DP4.
Along the small bony shelf anterior to the molar teeth, there are no traces of alveoli. Although this left maxillary fragment is incompletely preserved, and it is likely that additional deciduous premolars where present considering I. bellunensis is a juvenile, we therefore assume that permanent premolars were not developed in the animal’s lifetime, nor were likewise the canines or the second and third incisors. This would also be in agreement with other dugongid taxa showing similarly derived conditions such as Dioplotherium manigaulti (Domning 1989a) and Callistosiren boriquensis (Vélez-Juarbe and Domning 2015).
The left M1 (according to MV; Fig. 2E) is fully erupted and only slightly worn, with the uppermost tips of the main cusps flattened. Its cusp pattern does not differ from that of M2 and is summarised in the description below. The left M2 (according to MV; Fig. 2E) is incompletely erupted, occupying a slightly lower position relative to M1. Its crown is entirely unworn. The precingulum transversally slopes from the lingual to the labial (or buccal) side and encloses a narrow anterior cingular valley that opens anterolabially. The protoloph is composed of the paracone and protocone, both bracketing the protoconule to form a transverse ridge. While the protocone forms a prominent vertically-rising cusp, the paracone and protoconule slope in the lingual direction. The transverse valley is deep and not obstructed by the metaconule, whereas the latter is nearly transversally aligned with the hypocone and metacone to form the metaloph. As on the protoloph, the metacone and metaconule are inclined lingually, but the hypocone slopes labially. A posterior cingulum is attached to the hypocone and encloses a posterior basin of moderate size that opens labially.
Vertebral column: Besides four vertebral fragments (MDPD-7362Z, 7374Z, 7375Z, 7376Z) that are not assignable to a specific region of the vertebral column, three centra (MGPD-7367Z, 7368Z, 7369Z) are identified as thoracics and one centrum (MGPD-7383Z) is allocated to the lumbars. All centra are lacking the transverse processes and neural arches. The thoracic centra show a slightly heart-shaped outline, while the lumbar centrum is oval. The proximal and distal extremities are flat and slightly concave medially.
Ribs: Six rib fragments are present, of which MGPD-7358/9Z is the largest. It is composed of originally two fragments that are now glued together, representing the distal part of the rib shaft. This element measures about 240 mm in maximum preserved length and has a more or less constant anteroposterior width that diminishes slightly in the last 70 mm of the somewhat tapered distal extremity. Cemented gravel and shells cover the rib fragment, but the fracture surface of its shaft reveals an elliptical cross section of 50 × 30 mm. According to the curvature of the distal end, the rib is allocated to the left side of the thorax and may have occupied an anterior position. Another noteworthy element is a 128 mm long proximal fragment, most likely belonging to the left first rib (MGPD-7366Z). The capitulum and tuberculum are separated by 42 mm, indicating a long collum typical for the anteriormost ribs. Ventral to the capitulum, a protuberance is not developed.
Discussion
Based on the re-examination of “Halitherium” bellunense from Italy we confirm its stratigraphic provenance from the Chattian portion of the Belluno Glauconitic Sandstone Formation. Most importantly, we corroborate Domning’s (1989a, 1996) hypothesis on the dugongine affinities of this species. This new subfamily assignment requires that we establish a new genus, Italosiren, for “H.” bellunense.
Although the morphology of Italosiren bellunensis is inadequately known up to now due to the incomplete preservation of the holotype and only known specimen, this taxon can be clearly assigned to the crown group subfamily Dugonginae on the basis of the following derived characters: nasal process of premaxilla with broadened and bulbous posterior end having a more or less vertical joint surface with the frontal and/or nasal, and I1 mediolaterally compressed (lens-shaped with sharp anterior and posterior edges).
Referring to Domning (1994), who presented the first computer-based cladistic analysis of sirenians, the specific morphology of the posterior end of the premaxillary nasal process and an either lens-shaped, lozenge-shaped, or flattened I1 crown are synapomorphic traits that define a clade uniting the genera Rytiodus Lartet, 1866, Corystosiren Domning, 1990, Xenosiren Domning, 1989b, and Dioplotherium Cope, 1883. A close relationship of I. bellunensis to either Rytiodus or Corystosiren finds support from Sorbi (2008), who phylogenetically analysed this species for the first time by resolving it as a member of a monophyletic group including Rytiodus and Corystosiren, all supposed to uniquely share a short nasal process of the premaxilla (character 7 in Domning 1994 and later studies, e.g., Vélez-Juarbe and Domning 2014). However, the character definition for the scoring of the length of the nasal process was modified by Vélez-Juarbe et al. (2012: character 7), who evaluate this process as being “very short” when the “distance from rear of symphysis to posterior end of premaxilla < ½ length of symphysis”. Therefore, the nasal process of I. bellunensis must be considered as being long by this definition. In addition to this morphological difference of I. bellunensis compared to Rytiodus capgrandi and Corystosiren varguezi, the latter two species are no longer recovered as sister taxa in later studies (e.g., Vélez-Juarbe et al. 2012; Vélez-Juarbe and Domning 2015), but instead have a variable position within the Dugonginae or, as in Springer et al. (2015), plot within a polytomy. Hence, it is difficult to draw reliable conclusions on the phylogenetic relationships of I. bellunensis.
Domning and Sorbi (2011) discussed the potential phylogenetic position of I. bellunensis along with the introduction of the new species Rytiodus heali from the Miocene of Libya, and finally considered this taxon as most likely related to Rytiodus. Morphological features that corroborate this hypothesis in addition to the above-mentioned synapomorphies refer to the squamosal that does not reach the temporal crests and does not significantly indent the posterolateral corners of the parietal. Although these characters are likewise present in Corystosiren and Dioplotherium cf. allisoni (Toledo and Domning 1989), both known from the West Atlantic and Caribbean region, one can argue for a closer relationship of I. bellunensis with Rytiodus on the basis of their similar paleobiogeographical distribution (in Europe and North Africa, respectively).
Voss (2013) provided an alternative phylogenetic hypothesis and revealed the clade [D. manigaulti + [D. allisoni + [I. bellunensis + X. yucateca]]] by recovering I. bellunensis as the sister taxon to Xenosiren yucateca on the basis of a single homoplasy. Apart from differences on the species level, this result corroborates the hypothesis of Sorbi (2008) insofar as I. bellunensis represents a dugongine, hence providing objective support for Domning’s (1989a, 1996) previous suggestion. Interestingly, Corystosiren and Rytiodus are exactly those taxa that plot in a polytomy (Voss 2013) comparable to the results in Springer et al. (2015), indicating that further morphological investigations and cladistic studies on these two species and other dugongines may contribute to a better resolution of their interrelationships.
The cladistic approaches by Sorbi (2008) and Voss (2013) including I. bellunensis highlight the above-mentioned problem of the inconsistently resolved Dugonginae. The need for further systematic revisions was already pointed out by Domning (1994), who outlined the dugongine clade as one of the most unstable parts of his phylogeny, which is supported in all later studies (e.g., Vélez-Juarbe et al. 2012; Vélez-Juarbe and Domning 2015; Springer et al. 2015). Nevertheless, our study unambiguously recovers I. bellunensis as belonging to the Dugonginae and hence provides strong support for our initially developed hypothesis.
Although I. bellunensis displays a combination of characters that indicate an evolutionarily-derived status, further conclusions on its taxonomic and systematic position require caution for two reasons. On the one hand, the morphology of this species is not well known, due to the incomplete preservation of its holotype, lacking particularly relevant parts of the skull. On the other hand, the type of I. bellunensis is identified as representing a young animal. This is indicated by the most likely unerupted and unworn incisor tusk (Fig. 2A1), and by the early state of wear and eruption of the molars (Fig. 2D). As mentioned above and in contrast to Abel (1905), the preserved molariform teeth most likely do not represent the left DP5–M3 (MGPD-20/21Z) and right M1–2 (MGPD-7385/6Z), but the left DP5–M2 and the right DP5-M1 (opinion on tooth interpretation by MV). A tooth fragment directly in front of the remains of the left DP5 in MGPD-20/21Z is interpreted here as belonging to that same DP5 (Fig. 2E). The interpretation of the last preserved molar as not representing the left M3 is supported by the fact that the upper M3 in dugongids is usually larger than M2 and has an elongated shape with the metaloph transversally shorter than the protoloph. However, this is not the case in the present material. Here, the left M1 is nearly as large as M2, both showing a slightly heart-shaped outline with the protoloph as large as the metaloph. Moreover and considering the tooth eruption sequence, the presence of an unerupted tusk is inconsistent with a partially erupted M3. Following the interpretation of the maxillary teeth provided above, the left M2 occupies a position behind the zygomatic-orbital bridge, but would be more or less at the level of the bridge’s posterior edge in adults. Although it is difficult to judge the absolute age, I. bellunensis represents an immature specimen, considering the cheek tooth succession in young Dugong reported for example by Mitchell (1973) and Marsh (1980).
One other possibility is that this specimen has a small and incompletely-erupted tusk because it represents a female individual, by analogy with females of the modern Dugong dugon (cf. Marsh 1980). However, we consider this unlikely, because the sexual dimorphism of the tusks seen in D. dugon has not been demonstrated in any other sirenian (Sorbi et al. 2012); there is no evidence of dimorphism in Italosiren or any other fossil dugongine; and the sex of the studied specimen is unknown.
Against that background, it is possible that the specific shape of the I1 crown in I. bellunensis only reflects the morphology of the tip of an unworn tusk in a juvenile, which still may change during ontogeny and tooth wear, and should become much larger in adults and hence similar to the large blade-like tusks observable in its above-hypothesised closest relative Rytiodus. Domning and Sorbi (2011) already stated that such differences could be due to ontogeny and that I. bellunensis may even represent a young individual of Rytiodus sp. when some or all of the other morphological differences of I. bellunensis from that genus are also discounted as ontogenetic. However, since this cannot be ascertained at this time, because ontogenetic series from Rytiodus or any other closely-related taxon are not sufficiently known, we stick to the present available knowledge by treating I. bellunensis as a taxon distinct from all other known dugongines. This becomes most obvious by significant morphological differences observable between I. bellunensis and its hypothesised closest dugongine relatives. For example, I. bellunensis remarkably differs from Xenosiren in lacking a zygomatic-orbital bridge of the maxilla that is transformed into a vertical wall. In comparison with Corystosiren, Dioplotherium, and Rytiodus, I. bellunensis has not developed a deeply concave frontal roof. Moreover, I. bellunensis significantly differs from Rytiodus in lacking a distinct intertemporal constriction of the cranial roof, an overhanging orbital rim of the jugal, and large, blade-like tusks. In particular, the characters referring to the anteroposterior length of the zygomatic-orbital bridge and the overhanging orbital rim of the jugal do not display significant ontogenetic changes in the living dugong. According to personal observations by MV, the zygomatic-orbital bridge is shortened and the jugal orbital rim does distinctly overhang in juvenile (JCU MM019, 053, 181) and adult specimens (e.g., JCU MM192, 020). Considering the unique combination of morphological features in I. bellunensis, the evaluation of its systematic position is therefore postponed pending further investigations.
Conclusions
The paleogeographic and stratigraphic distribution of the sirenian specimen now assigned to Italosiren bellunensis remain restricted to the Belluno Basin and to Chattian sediments according to our present knowledge (although the species doubtless had a wider distribution in the Tethyan/Paratethyan region). The morphology of I. bellunensis confirms that the Dugonginae lived along the Euro-North African coasts during the late Oligocene through the early Miocene. The stratigraphical and taxonomical review of this species is here placed on an objective geological and morphological basis for the first time, and corroborates the hypothesis of its dugongine affinities already postulated by Domning (1989a, 1996).
Acknowledgements
We gratefully acknowledge Letizia Del Favero (MGPD) for the preparation of the left premaxilla, Stefano Castelli (MGPD) for providing us with photos of this element, and Mariagabriella Fornasiero (MGPD) for providing access to the fossil sirenian material. Additionally, we thank Stefanie Klug (The University of Manchester, UK) and Jason Dunlop (Museum für Naturkunde, Berlin, Germany) for their helpful comments on the manuscript and for checking the spelling and grammar. Finally, we are grateful for the constructive reviews by Iyad Zalmout (Saudi Geological Survey, Jeddah, Saudi Arabia) and Jorge Velez-Juarbe (Natural History Museum of Los Angeles County, USA), who helped to improve the present paper. The present study received funding from the SYNTHESYS Project http://www.synthesys.info/ which is financed by European Community Research Infrastructure Action under the FP7 “Capacities” Program (GB-TAF-5171, HU-TAF-5158, both to MV). We also thank the Deutsche Forschungsgemeinschaft (DFG, HA 1776/11-1 to O. Hampe) and Oliver Hampe (Berlin) for supporting this project.
References
Abel, O. 1905. Über Halitherium bellunense, eine Übergangsform zur Gattung Metaxytherium. Jahrbuch der Kaiserlich-Königlichen Geologischen Reichsanstalt 55: 393–398.
Bajpai, S. and Domning, D.P. 1997. A new dugongine sirenian from the early Miocene of India. Journal of Vertebrate Paleontology 17: 219–228. Crossref
Bajpai, S., Domning, D.P., Das, D.P., Vélez-Juarbe, J., and Mishra, V.P. 2010. A new fossil sirenian (Mammalia, Dugonginae) from the Miocene of India. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 258: 39–50. Crossref
Bajpai, S., Thewissen, J.G.M., Kapur, V.V., Tiwari, B.N., and Sahni, A. 2006. Eocene and Oligocene sirenians (Mammalia) from Kachchh, India. Journal of Vertebrate Paleontology 26: 400–410. Crossref
Bosellini, A., Masetti, D., and Sarti, M. 1981. A Jurassic “Tongue of the Ocean” infilled with oolitic sands: The Belluno Trough, Venetian Alps, Italy. Marine Geology 44: 59–95. Crossref
Cope, E.D. 1889. Synopsis of the families of Vertebrata. American Naturalist 23: 849–877. Crossref
Dallanave, E., Agnini, C., Muttoni, G., and Rio, D. 2009. Magneto-biostratigraphy of the Cicogna section (Italy): Implications for the late Paleocene–early Eocene time scale. Earth and Planetary Science Letters 285: 39–51. Crossref
Delfortrie, E. 1880. Découverte d’un squelette entier de Rytiodus dans le falun Aquitanien. Actes de la Société Linnéenne de Bordeaux 34:131–144.
De Zigno, A. 1875. Sirenii fossil itrovati nel Veneto. Memorie del Reale Istituto Veneto di Scienze, Lettre ed Arti 18: 427–456.
De Zigno, A. 1878. Sopra un nuovo sirenio fossile scopertonelle colline di Bra in Piemonte. Atti del Reale Accademia dei Lincei, Memorie della Classe di scienze fisiche matematiche e naturali 3 (2): 939–949.
De Zigno, A. 1887. Quelques observations sur les siréniens fossiles. Bulletin de la Société Geologique de France 3 (15): 728–732.
Domning, D.P. 1978. Sirenian evolution in the North Pacific Ocean. University of California Publications in Geological Sciences 118: 1–176.
Domning, D.P. 1989a. Fossil Sirenia of the West Atlantic and Caribbean region. II. Dioplotherium manigaulti Cope, 1883. Journal of Vertebrate Paleontology 9: 415–428. Crossref
Domning, D.P. 1989b. Fossil Sirenia of the West Atlantic and Caribbean region. III. Xenosiren yucateca, gen. et sp. nov. Journal of Vertebrate Paleontology 9: 429–437. Crossref
Domning, D.P. 1990. Fossil Sirenia of the West Atlantic and Caribbean region. IV. Corystosiren varguezi, gen. et sp. nov. Journal of Vertebrate Paleontology 10: 361–371. Crossref
Domning, D.P. 1994. A phylogenetic analysis of the Sirenia. In: A. Berta and T. Deméré (eds.), Contributions in Marine Mammal Paleontology Honoring Frank C. Whitmore, Jr. Proceedings of the San Diego Society of Natural History 29: 177–189.
Domning, D.P. 1996. Bibliography and index of the Sirenia and Desmostylia. Smithsonian Contributions to Paleobiology 80: 1–611.
Domning, D.P. 1997. Fossil Sirenia of the West Atlantic and Caribbean region. VI. Crenatosiren olseni Reinhart, 1976. Journal of Vertebrate Paleontology 17: 397–412. Crossref
Domning, D.P. 2001. Sirenians, seagrasses, and Cenozoic ecological change in the Caribbean. Palaeogeography, Palaeoclimatology, Palaeoecology 166: 27–50. Crossref
Domning, D.P. and Aguilera, O.A. 2008. Fossil Sirenia of the West Atlantic and Caribbean region. VIII. Nanosiren garciae, gen. et sp. nov. and Nanosiren sanchezi, sp. nov. Journal of Vertebrate Paleontology 28: 479–500. Crossref
Domning, D.P. and Beatty, B.L. 2007. Use of tusks in feeding by dugongid sirenians: observations and tests of hypotheses. The Anatomical Record 290: 523–538. Crossref
Domning, D.P. and Sorbi, S. 2011. Rytiodus heali, sp. nov., a new sirenian (Mammalia, Dugonginae) from the Miocene of Libya. Journal of Vertebrate Paleontology 31: 1338–1355. Crossref
Domning, D.P. and Thomas, H. 1987. Metaxytherium serresii (Mammalia: Sirenia) from the early Pliocene of Libya and France: a reevaluation of its morphology, phyletic position, and biostratigraphic and paleoecological significance. In: N. Boaz, A. ElArnauti, A.W. Gaziry, J. de Heinzelin, and D.D. Boaz (eds.), Neogene Paleontology and Geology of Sahabi, 205–232. John Wiley & Sons Inc, New York.
Domning, D.P., Zalmout, I.S., and Gingerich, P.D. 2010. Sirenia. In: L. Werdelin and W.J. Sanders (eds.), Cenozoic Mammals of Africa, 147–160. University of California Press, Berkeley.
Ghibaudo, G., Grandesso, P., Massari, F., and Uchman, A. 1996. Use of trace fossils in delineating sequence stratigraphic surfaces (Tertiary Venetian Basin, northeastern Italy). Palaeogeography, Palaeoclimatology, Palaeoecology 120: 261–279. Crossref
Gray, J.E. 1821. On the natural arrangement of vertebrose animals. London Medical Repository 15: 296–310.
Illiger, J.K.W. (ed.) 1811. Prodomus systematis mammalium et avium: additis terminis zoographicis utriusque classis eorumque versione Germanica. 301 pp. Berolini, Salfeld.
Kaup, J.J. 1838. Halytherium und Pugmeodon im Maynzer Becken. Neues Jahrbuch für Mineralogie, Geognosie, Geologie und Petrefaktenkunde 1838: 318–320.
Lartet, M. 1866. Note sur deux nouveaux siréniens fossiles des terrains tertiaires du bassin de la Garonne. Bulletin de la Société Geologique de France 23: 673–686.
Lepsius, G.R. (ed.) 1882. Halitherium Schinzi, die fossile Sirene des Mainzer Beckens. Abhandlungen des Mittelrheinischen geologischen Vereins 1: 1–200.
Linnaeus, C. 1758. Systema naturae per regna tria naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Editio 10 (reformata), 823 pp. Laurentii Salvii, Holmiae.
Marsh, H. 1980. Age determination of the dugong (Dugong dugon (Müller)) in northern Australia and its biological implications. Reports of the International Whaling Commission, Special Issue 3: 181–201.
McKenna, M.C. 1975. Toward a phylogenetic classification of the Mammalia. In: W.P. Luckett and F.S. Szalay (eds.), Phylogeny of the Primates: A Multidisciplinary Approach, 21–46. Plenum, New York. Crossref
Mellere, D., Stefani, C., and Angevine, C. 2000. Polyphase tectonics through subsidence analysis: the Oligo-Miocene Venetian and Friuli Basin, north-east Italy. Basin Research 12: 159–182. Crossref
Mitchell, J. 1973. Determination of relative age in the dugong (Dugong dugon, Müller) from a study of skulls and teeth. Zoological Journal of the Linnean Society 53: 1–23. Crossref
Pilleri, G. 1985. The Miocene Cetacea of the Belluno sandstones (eastern southern Alps). Memoire di Scienze Geologiche 36: 1–250.
Simpson, G.G. 1932. Fossil Sirenia of Florida and the evolution of the Sirenia. Bulletin of the American Museum of Natural History 59: 419–503.
Sorbi, S. 2008. The Neogene Euro-North African Sirenia: Phylogenetic and Systematic Study, Palaeoecological and Palaeobiogeographic Considerations. 396 pp. Unpublished Ph. D. dissertation, Università di Pisa, Pisa.
Sorbi, S., Domning, D.P., Vaiani, S.C., and Bianucci, G. 2012. Metaxytherium subapenninum (Bruno, 1839) (Mammalia, Dugongidae), the latest sirenian of the Mediterranean Basin. Journal of Vertebrate Paleontology 32: 686–707. Crossref
Springer, M.S., Signore, A.V., Paijmans, J.L.A., Vélez-Juarbe, J., Domning, D.P., Bauer, C.E., He, K., Crerar, L., Campos, P.F., Muphy, W.J., Meredith, R.W., Gatesy, J., Willerslev, E., MacPhee, R.D.E., Hofreiter, M., and Campbell, K.L. 2015. Interordinal gene capture, the phylogenetic position of Steller’s sea cow based on molecular and morphological data, and the macroevolutionary history of Sirenia. Molecular Phylogenetics and Evolution 91: 178–193. Crossref
Stanhope, M.J., Waddell, V.G., Madsen, O., de Jong, W.W., Hedges, S.B., Cleven, G.C., Kao, D., and Springer, M.S. 1998. Molecular evidence for multiple origins of the Insectivora and for a new order of endemic African mammals. Proceedings of the National Academy of Sciences USA 95: 9967–9972. Crossref
Thewissen, J.G.M. and Bajpai, S. 2009. A new Miocene sirenian from Kutch, India. Acta Palaeontologica Polonica 54: 7–13. Crossref
Toledo, P.M. de and Domning, D.P. 1989. Fossil Sirenia (Mammalia: Dugongidae) from the Pirabas Formation (Early Miocene), northern Brazil. Boletim do Museu Paraense Emilio Goeldi. Série Ciências da Terra 1: 119–146.
Vélez-Juarbe, J. and Domning, D.P. 2014. Fossil Sirenia of the West Atlantic and Caribbean region. IX. Metaxytherium albifontanum, sp. nov. Journal of Vertebrate Paleontology 34: 444–464. Crossref
Vélez-Juarbe, J. and Domning, D.P. 2015. Fossil Sirenia of the West Atlantic and Caribbean Region. XI. Callistosiren boriquensis, gen. et sp. nov. Journal of Vertebrate Paleontology 35: 1–16. Crossref
Vélez-Juarbe, J., Domning, D.P., and Pyenson, N.D. 2012. Iterative evolution of sympatric seacow (Dugongidae, Sirenia) assemblages during the past ~26 Million years. PLoS ONE 7: 1–8. Crossref
Voss, M. 2013. Revision of the Halitherium-species Complex (Mammalia, Sirenia) from the Late Eocene to Early Miocene of Central Europe and North America. 298 pp. Unpublished Ph.D. Dissertation, Humboldt-Universität zu Berlin, Berlin.