
Terreneuvian stratigraphy and faunas from the Anabar Uplift, Siberia
ARTEM KOUCHINSKY, STEFAN BENGTSON, ED LANDING, MICHAEL STEINER, MICHAEL VENDRASCO, and KAREN ZIEGLER
Kouchinsky, A., Bengtson, S., Landing, E., Steiner, M., Vendrasco, M., and Ziegler, K. 2017. Terreneuvian stratigraphy and faunas from the Anabar Uplift, Siberia. Acta Palaeontologica Polonica 62 (2): 311‒440.
Assemblages of mineralized skeletal fossils are described from limestone rocks of the lower Cambrian Nemakit-Daldyn, Medvezhya, Kugda-Yuryakh, Manykay, and lower Emyaksin formations exposed on the western and eastern flanks of the Anabar Uplift of the northern Siberian Platform. The skeletal fossil assemblages consist mainly of anabaritids, molluscs, and hyoliths, and also contain other taxa such as Blastulospongia, Chancelloria, Fomitchella, Hyolithellus, Platysolenites, Protohertzina, and Tianzhushanella. The first tianzhushanellids from Siberia, including Tianzhushanella tolli sp. nov., are described. The morphological variation of Protohertzina anabarica and Anabarites trisulcatus from their type locality is documented. Prominent longitudinal keels in the anabaritid Selindeochrea tripartita are demonstrated. Among the earliest molluscs from the Nemakit-Daldyn Formation, Purella and Yunnanopleura are interpreted as shelly parts of the same species. Fibrous microstructure of the outer layer and a wrinkled inner layer of mineralised cuticle in the organophosphatic sclerites of Fomitchella are reported. A siliceous composition of the globular fossil Blastulospongia is reported herein and a possible protistan affinity similar to Platysolenites is discussed. New carbon isotope data facilitate correlation both across the Anabar Uplift and with the Terreneuvian Series of the IUGS chronostratigraphical scheme for the Cambrian System. The base of Cambrian Stage 2 is provisionally placed herein within the Fortunian‒Cambrian Stage 2 transitional interval bracketed by the lowest appearance of Watsonella crosbyi and by a slightly higher horizon at the peak of carbon isotope excursion Iʹ from western flank of the Anabar Uplift. Correlation across the Siberian Platform of the fossiliferous Medvezhya and lower Emyaksin formations showing δ13Ccarb excursion Iʹ with the upper Sukharikha Formation containing excursion 5p and upper Ust’-Yudoma Formation containing excursion I is supported herein.
Key words: Mollusca, small shelly fossils, stratigraphy, carbon isotopes, Cambrian, Terreneuvian, Russia, Siberia, Anabar.
Artem Kouchinsky [artem.kouchinsky@nrm.se] and Stefan Bengtson [stefan.bengtson@nrm.se], Department of Palaeobiology, Swedish Museum of Natural History, Box 50007, SE-104 05 Stockholm, Sweden.
Ed Landing [ed.landing@nysed.gov], New York State Museum, 222 Madison Ave, Albany, NY 12230, USA.
Michael Steiner [michael.steiner@fu-berlin.de], Freie Universität Berlin, Malteserstrasse 74-100, Haus C Raum C.021, D-12249 Berlin, Germany.
Michael Vendrasco [mvendrasco@pasadena.edu], Department of Geology, Pasadena City College, 1570 E Colorado Blvd, Pasadena, CA 91106, USA.
Karen Ziegler [kziegler@unm.edu], Institute of Meteoritics, 1 University of New Mexico, Albuquerque, NM 87131-0001, USA.
Received 29 June 2016, accepted 28 February 2017, available online 31 May 2017.
Copyright © 2017 A. Kouchinsky et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
According to the Interdepartmental Stratigraphic Committee of Russia (Zhamoida and Petrov 2008), the base of the Cambrian System on the Siberian Platform coincides with the base of the Tommotian Stage. The underlying dolostone-dominated strata on the Siberian Platform are commonly referred to the Nemakit-Daldynian Stage of the General Stratigraphic Scale of Russia (Khomentovsky and Karlova 1993, 2005; Rozanov et al. 2008) and assigned to the Vendian System. The upper or the entire Nemakit-Daldynian is correlated with the Fortunian Stage of the Terreneuvian Series (e.g., Khomentovsky and Karlova 2005; Khomentovsky 2008; Zhamoida and Petrov 2008; Rogov et al. 2015). The latter are the lowest stage and series of the IUGS chronostratigraphical scheme for the Cambrian System, whereas informal Cambrian Stage 2 is upper Terreneuvian (e.g., Landing et al. 2007, 2013; Peng and Babcock 2011). Although the Nemakit-Daldynian Stage is traditionally and officially used in Russia, neither the Nemakit-Daldynian nor the synonymous Manykayan Stage can be formally used as regional chronostratigraphic units, as “Nemakit-Daldyn” and “Manykay,” by the international rules of stratigraphic nomenclature, are preoccupied and were earlier used as lithostratigraphic names (e.g., Salvador 1994).
Historically, “Nemakit-Daldyn” and “Manykay” have been applied to two time-equivalent lithostratigraphic units (formations) on the Anabar Uplift. The Nemakit-Daldyn Formation was defined by Savitsky (1962) at the mouth of the Nemakit-Daldyn River, a tributary of the Eriechka River, and traced in the north-western and northern flanks of the Anabar Uplift between the Kotuj and Popigaj rivers. The Manykay Formation was defined by Boris G. Lopatin and collegues in 1958–1959 (see Demokidov and Lazarenko 1964) along the middle course of the Malaya Kuonamka River (near the mouth of the Manykay Creek) on the south-eastern flank of the Anabar Uplift. The Nemakit-Daldyn and Manykay formations are respectively overlain by the Medvezhya and Emyaksin formations. The Medvezhya Formation on the western flank of the Anabar Uplift was first defined in 1959–1960 by Vladimir E. Savitsky (Egorova and Savitsky 1969) along the upper reaches of the Medvezhya River, upstream of the mouth of the Daldyn River. The Emyaksin Formation was named by Mikhail N. Zlobin (see Demokidov 1956, 1958) at the mouth of the Emyaksin-Yurege River, a tributary of the Malaya Kuonamka River, and traced along the southeastern, eastern, and northeastern flanks of the Anabar Uplift.
The name Kugda-Yuryakh Formation was coined by the geologists of the Krasnoyarsk Geological Survey (“Krasnoyarskgeologiya”) in 1979 (see Dyatlova et al. 1990). Minaeva (1985, 1991) mentioned the Kugda-Yuryakh Formation as the “Serotsvet [grey-colored] Member” or Serotsvet Formation. Missarzhevsky (1989: 66, fig. 15) placed the “Kugda beds” between the top of the Medvezhya Formation and the bottom of the overlying “Kyndyn beds”. Kaufman et al. (1996: 515) assigned the “Kugda beds” to the “Kyndyn Formation”, but noted that their lower and upper boundaries are regionally diachronous and problematic for lithostratigraphic correlation alone.
This report provides additional information on the regional and global chronostratigraphy of the lowest part of the Cambrian System, or the Terreneuvian Series. The early skeletal fossils illustrated and discussed herein along with the carbon isotope chemostratigraphy from the western and eastern flanks of the Anabar Uplift of the northern Siberian Platform facilitate comparison with Terreneuvian faunas of western Mongolia, Kazakhstan, Laurentia, Baltica, Avalonia, and Gondwanan regions as South China, India, Iran, and France. This paper continues the reports by Gubanov et al. (2004b) and Kouchinsky et al. (2011, 2015a, b; SOM 5: figs. 9–17, Suplementary Online Material available at http://app.pan.pl/SOM/app62-Kouchinsky_etal_SOM.pdf) of fossil assemblages known from younger beds on the eastern flank of the Anabar Uplift.
Institutional abbreviations.—SMNH (Mo, X), Swedish Museum of Natural History, Stockholm, Sweden (molluscs, other fossils).
Other abbreviations.—SSF, small shelly fossils.
Material and methods
The material described herein was collected during fieldworks in the Anabar Uplift of Siberia, Russia. The collections were made by AK and SB from the Manykay and Emyaksin formations along the Bol’shaya Kuonamka River in 1996 and from the Nemakit-Daldyn, Medvezhya, and Kugda-Yuryakh formations along the Kotuj River in 1992, 2007 (by AK) and 2008 (by Vladimir Pavlov) (Figs. 1–4; SOM 5).
Section 1 herein (locality M410 in Rozanov et al. 1969; locality 3 in Meshkova et al. 1976) is located at 70°08.5’ N, 114°00’ E (Figs. 1 and 4; SOM 5: figs. 1, 2; see also Landing and Kouchinsky 2016). It crops out on the left bank of the Kotuj River, upstream from the mouth of Ary-Mas-Yuryakh Creek. The lowermost sampled part of section 1 belongs to the uppermost Nemakit-Daldyn Formation (thrombolitic boundstone of the Koril Member; Kaufman et al. 1996), which is overlain above the sequence boundary by limestone beds of the Medvezhya Formation grading upwards into grey limestone with oolites attributable to the Kugda-Yuryakh Formation (e.g., Dyatlova et al. 1990; Fig. 4).
Section 2 (locality M419 in Rozanov et al. 1969; section E8 in Val’kov 1975) is situated at 70°37’ N; 103°31.5’ E, along the right bank of the Kotujkan River, ca. 2.5 km upstream its mouth (Fig. 1; SOM 5: fig. 3). The section embraces the Nemakit-Daldyn Formation (mainly limestone with minor addition of dolostone layers and interbedded siliciclastics in the lower part) separated by sequence boundaries from the underlying Staraya Rechka Formation (Ediacaran, mainly stromatolitic dolostone in the upper part) and the overlying variegated lime mudstone and wackestone of the Medvezhya Formation.
Section 3 (locality M423 in Rozanov et al. 1969; locality 9 in Meshkova et al. 1976) crops out at the mouth of the Kugda-Yuryakh Creek (70°38.6’ N, 103°25.9’ E) on the right bank of the Kotuj River, ca. 3 km downstream from the mouth of Kotujkan River (Figs. 1 and 3; SOM 5: figs. 4 and 5). The Medvezhya Formation rests on the Koril Member of the Nemakit-Daldyn Formation at section 3 and grades upward into oolitic limestone known as the Kugda-Yuryakh Formation, which is 100–130 m thick in the area and overlain by dolostone attributed to the Kyndyn Group (Minaeva 1985, 1991; Dyatlova et al. 1990; Fig. 3). Samples for this report were collected mainly from the left side of the mouth of Kugda-Yuryakh Creek canyon, from the top of the Nemakit-Daldyn Formation through the lower 75 m of the Kugda-Yuryakh Formation. A complementary set of samples from what is designated herein as sub-section 3b (70°38.5’ N; 103°25’ E) from the Medvezhya Formation was sampled from the right bank of the Kotuj River, ca. 500 m downstream from the mouth of Kugda-Yuryakh Creek. Finally, samples from sub-section 3a from the upper part of the Nemakit-Daldyn Formation (thrombolitic boundstone of the Koril Member and lime mudstone with calcisiltite of the Sub-Koril Member) were collected from a beach outcrop along the Kotuj River ca. 300 m upstream from the mouth of the Kugda-Yuryakh Creek (70°38.5’ N; 103°26.5’ E).
Section 96-4 is exposed on the left bank of the Bol’shaya Kuonamka on the eastern flank of the Anabar Uplift (section A-51 in Val’kov 1975 and 96K4 in Kouchinsky et al. 1999; 70°43.1’ N, 112°45.8’ E), 1 km upstream of the mouth of the Ulakhan-Tyulen Creek (Figs. 1 and 2; SOM 5: fig. 6). The Manykay Formation is represented here mainly by siliciclastics in its upper part, whereas a carbonate thrombolitic unit occurs in the lowermost exposed part of the section (Fig. 2). Fossils were derived from the top of the unit, which is a layer of carbonate packstone (“marker bed with angustiochreids” in Val’kov 1975: 8; see Kouchinsky et al. 1999; SOM 5: fig. 6), and from polymict gravel lenses (sections 96-4 and 96-5) of the basal Emyaksin Formation.
Adjacent sections 96-5 (70°42.4’ N, 112°47.2’ E) and 96-5a (= locality A50 in Val’kov 1975; see also Kouchinsky et al. 2001; 70°42.6’ N, 112°48’ E) are located at the Bol’shaya Kuonamka River on the eastern flank of the Anabar Uplift (Figs. 1 and 2; SOM 5: figs. 7–9). Section 96-5 is ca. 1 km upstream from the mouth and on the left bank of Ulakhan-Tyulen Creek (right tributary to Bol’shaya Kuonamka). Section 96-5a is on the right bank of Ulakhan-Tyulen Creek, at its mouth. The lower Emyaksin Formation is represented by greenish-grey and reddish-maroon lime mud- and wackestone with glauconite and polymict gravel lenses up to a few decimeters-thick at the base of formation. Section 96-6 (see Kouchinsky et al. 2001, 2015a) is the upper continuation of sections 96-5 and 96-5a (Fig. 2; SOM 5: figs. 9–11), and is located 1.0–2.5 km downstream from the mouth of Ulakhan-Tyulen, on the right bank of the Bol’shaya Kuonamka River (70°43’ N; 112°56’ E).
Included herein are also samples available from the following localities. Sample M424/10, collected by Vladimir V. Missarzhevsky from the uppermost Medvezhya or basal Kugda-Yuryakh Formation in locality M424 (Rozanov et al. 1969), which crops out on the left bank of the Kotuj River, opposite the mouth of Kugda-Yuryakh Creek, western flank of the Anabar Uplift. Samples M314/4, M314/5-10, M314/7, and M314/12, collected by Vladimir V. Missarzhevsky from the Medvezhya Formation, from locality M314 (Rozanov et al. 1969), on the right bank of the Fomich River, ca. 6 km upstream of the Afanas’ev Lakes, northwestern flank of the Anabar Uplift. Sample M321/31, collected by Vladimir V. Missarzhevsky, from the Medvezhya Formation at locality M321 (Rozanov et al. 1969), on the left bank of the Eriechka River, 1.5 km downstream of the Nemakit-Daldyn River, northwestern flank of the Anabar Uplift. Samples 769 and B-438, collected, respectively, by Anatolij K. Val’kov (Val’kov 1987: 24) and Anna R. Bokova (Bokova 1992) from the lower Emyaksin Formation, from the left bank of the Kengede River (left tributary of the Arga-Sala River), 1.8 km downstream of the Kharyjalakh Creek, southern flank of the Anabar Uplift. Samples M71-2/62 and M71-2/66, collected by Vladimir V. Missarzhevsky (Missarzhevsky 1982) from the upper Kessyusa Formation in locality M71-2, left bank of the Olenyok River, 6 km downstream of the Chuskuna Creek, Olenyok Uplift, Siberia. Sample 1571/2, derives from an unnamed formation, 2.5–4.5 m above the top of Kolosovskaya Formation (Proterozoic), at the mouth of an unnamed right tributary of the Leningradskaya River, opposite the mouth of the Stepanovo Uschel’e Creek, NE Tajmyr. Sample S1, collected by Anatolij K. Val’kov in 2002 from the uppermost Ust’-Yudoma Formation in the upper reaches of the Uchur River, Selinde River section, Siberia.
Fossils were extracted from the limestone (1–3 kg samples) with a buffered solution of 10% acetic acid. The fossils were manually sorted from the acid-resistant residues under a binocular microscope, then mounted and coated with gold for examination with a Hitachi S4300 scanning electron microscope at the Swedish Museum of Natural History. Results of carbon isotope analyses from sections 96-4, 96-5, and 96-5a were obtained at Stockholm University, Sweden, in 1999–2000 (see Kouchinsky et al. 2001 for description of methods and results). Samples from sections 1 and 3 were analysed for carbon isotopes by KZ at the University of California, Los Angeles in 2009 (see description of methods in Kouchinsky et al. 2005, 2007, 2008), and the results are available in SOM 4.

Fig. 1. A. Chemostratigraphic (δ13Ccarb) correlation between sections in the eastern (Bol’shaya Kuonamka River) and western (Kotuj River) Anabar Uplift. Nemakit-Daldyn Formation in section 2 subdivided into members 1–9 after Kaufman et al. (1996). Derivation of fossiliferous samples from section 2 shown by arrows with numbers; M419/12*, location of sampled horizons including samples M419/12, 2/4, 2/7, K1b/8, K1b/9, K1b/10, and K1b/13; 1282*, location of samples 1282, K1a/43, K1a/45, K1a/47, K1a/48, and 2/A. Correlation with the Tommotian–lower Botoman Stage carbon isotope features II–VII from stratotypes on the southeastern Siberian Platform (Brasier et al. 1994) according to Kouchinsky et al. (2001). Stratigraphic ranges and first appearances of fossils (see SOM 1): 1, first appearance of helcionellid molluscs; 2, Oelandiella korobkovi; 10, Pseudoyangtzespira selindeica; 18, Khairkhania cf. evoluta; 22, Purella antiqua; 24, Yunnanopleura accurata; 30, Protoconus? sp.; 31, first appearance of siphogonuchitids; 33, Siphogonuchites subremualis; 36, Dabashanites mirus; 37, first appearance of Halkieria sp.; 45, Paragloborilus sp.; 51, Protohertzina anabarica; 52, Fomitchella infundibuliformis; 64, Anabarites trisulcatus; 75, Cambrotubulus decurvatus; 76, Platysolenites antiquissimus. Maps of the studied areas, showing location of the sections in eastern (B, Bol’shaya Kuonamka River) and western (C, Kotuj River) Anabar Uplift. D. Map of the Siberian Platform with localities referred to in the main text indicated by circles with numbers: 1, Tajmyr, Leningradskaya River; 2, Sukharikha and Kulyumbe rivers; 3, western flanks of the Anabar Uplift, Kotuj River between the Sergej-Koril-Uoran rapids and mouth of the Kugda-Yuryakh Creek and the lowermost reaches of the Kotujkan River (see C); 4, north-western and northern flanks of the Anabar Uplift, including Medvezhya, Eriechka, Fomitch, and Rassokha rivers; 5, eastern flanks of the Anabar Uplift, including the Bol’shaya Kuonamka River lower reaches (see B), Malaya Kuonamka River middle reaches, Udzha and Emyaksin-Yurege rivers; 6, southern flanks of the Anabar Uplift, Kengede River; 7, Olenyok River middle reaches; 8, Lena River middle reaches, between sections Isit’ and Achchagyi-Kyyry-Taas; 9, Aldan River, between sections “Dvortsy” and Ulakhan-Sulugur; 10, Uchur-Maya region, sections along the Uchur River, between the Gonam and Selinde rivers, including the Mount 1291 m, Mount Konus, Nemnekey, and Selinde localities. E. Map of Eurasia showing location of the studied area. N, the strong negative basal Cambrian C-isotopic excursion (after Kaufman et al. 1996).

Fig. 2. Composite δ13Ccarb curve for the Bol’shaya Kuonamka River sections 96-4, 5, and 5a and sampled horizons with fossiliferous samples (arrows with numbers). Arrow 4/4 indicates “marker bed with angustiochreids” (Val’kov 1975; Kouchinsky et al. 1999). For section 96-6 see Kouchinsky et al. (2001, 2015a). Shift in stratigraphic column at Manykay–Emyaksin formations contact indicates position of sequence boundary. Stratigraphic ranges and first appearances of fossils (see SOM 1): 1, first appearance of helcionellid molluscs; 2, Oelandiella korobkovi; 3, Securiconus cf. simus; 6, Obtusoconus vostokovae; 7, Obtusoconus cf. mirabilis; 8, Anabarella plana; 13, Aldanella attleborensis; 14, Aldanella cf. sibirica; 22, Purella antiqua; 24, Yunnanopleura accurata; 34, Lopochites sp.; 35, Tianzhushania longa; 37, first appearance of Halkieria sp.; 38, Chancelloria symmetrica; 42, Khetatheca cotuiensis; 44, Conotheca cf. mammilata; 46, Turcutheca sp.; 47, Cupitheca sp.; 50, Orthothecid operculum type C; 51, Protohertzina anabarica; 52, Fomitchella infundibuliformis; 53, Fomitchella aff. infundibuliformis; 54, Fomitchella acinaciformis; 55, Fomitchella aff. acinaciformis; 56, Hyolithellus vladimirovae; 57, Hyolithellus sp.; 60, Selindeochrea cf. tecta; 61, Selindeochrea tripartita; 62, Selindeochrea missarzhevskyi; 63, Selindeochrea ternaria; 64, Anabarites trisulcatus; 65, Anabarites latus; 66, Anabarites natellus; 67, Anabarites kelleri; 68, Anabarites biplicatus; 69, Anabarites tristichus; 70, Anabarites compositus; 71, Anabarites hexasulcatus; 72, Anabarites hariolus; 73, Aculeochrea rugosa; 74, Tiksitheca licis; 75, Cambrotubulus decurvatus; 76, Platysolenites antiquissimus.
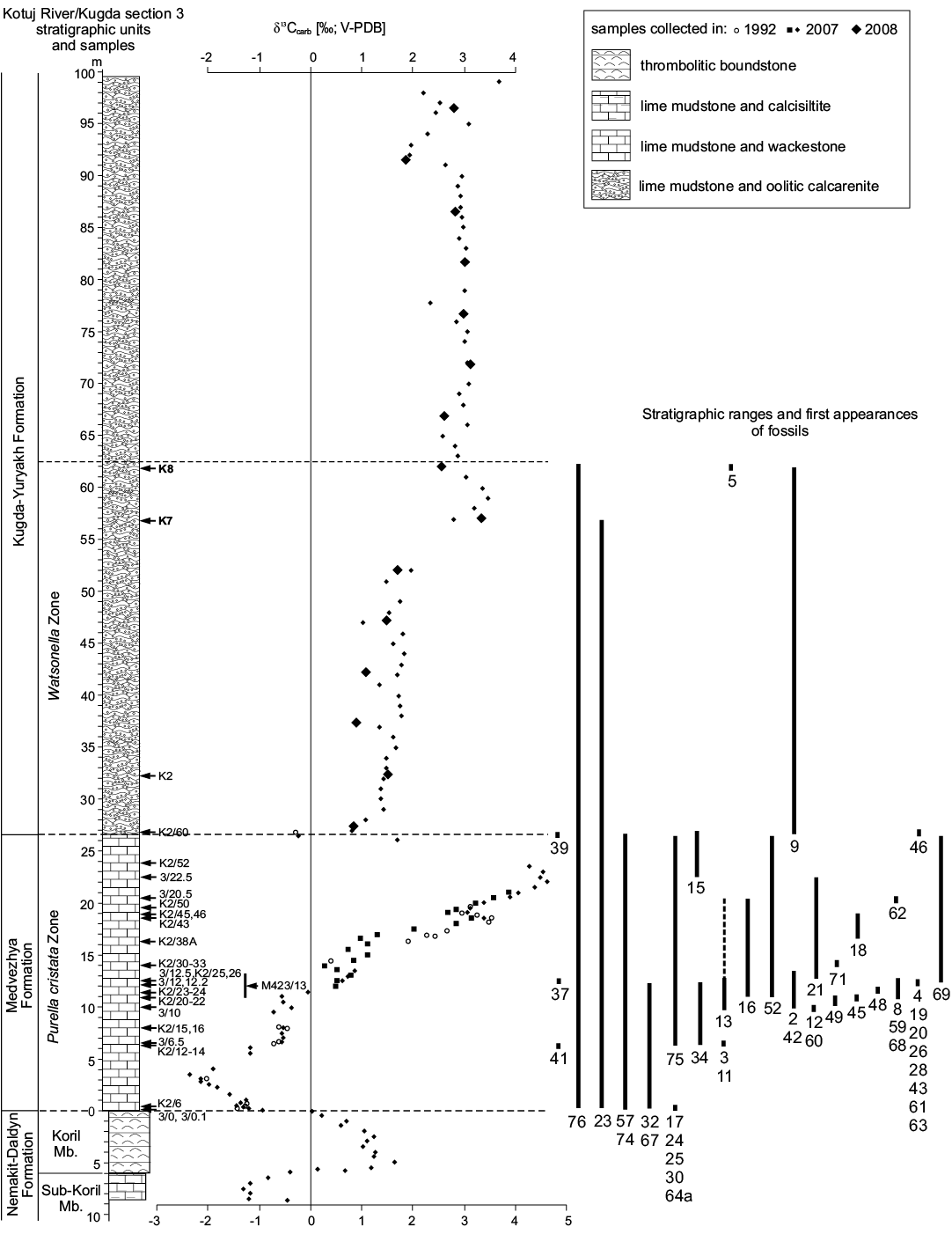
Fig. 3. The δ13Ccarb curve for Kotuj River section 3 and sampled horizons with fossiliferous samples (arrows with numbers). Shift in stratigraphic column at Nemakit-Daldyn–Medvezhya formations contact indicates position of sequence boundary. Stratigraphic ranges and first appearances of fossils (see SOM 1): 2, Oelandiella korobkovi; 3, Securiconus cf. simus; 4, Bemella jacutica?; 5, Obtusoconus cf. honorabilis; 8, Anabarella plana; 9, Watsonella crosbyi; 11, Aldanella crassa; 12, Aldanella golubevi; 13, Aldanella attleborensis; 15, Philoxenella spiralis; 16, Barskovia hemisymmetrica; 17, Khairkhania cf. rotata; 18, Khairkhania cf. evoluta; 19, Ceratoconus striatus; 20, Ceratoconus aff. striatus; 21, Salanyella cf. costulata; 23, Purella cristata; 24, Yunnanopleura accurata; 25, Yunnanopleura sp.; 26, Obscurania auriculata; 28, Algomella sp.; 30, Protoconus? sp.; 32, Siphogonuchites triangularis; 34, Lopochites sp.; 37, first appearance of Halkieria sp.; 39, first appearance of Chancelloria sp.; 41, first appearance of orthothecid hyoliths; 42, Khetatheca cotuiensis; 43, Kotujtheca curta; 45, Paragloborilus sp.; 46, Turcutheca sp.; 48, orthothecid operculum type A; 49, orthothecid operculum type B; 52, Fomitchella infundibuliformis; 57, Hyolithellus sp.; 59, Selindeochrea tricarinata; 60, Selindeochrea cf. tecta; 61, Selindeochrea tripartita; 63, Selindeochrea ternaria; 64a, Anabarites cf. trisulcatus; 67, Anabarites kelleri; 68, Anabarites biplicatus; 69, Anabarites tristichus; 71, Anabarites hexasulcatus; 74, Tiksitheca licis; 75, Cambrotubulus decurvatus; 76, Platysolenites antiquissimus.
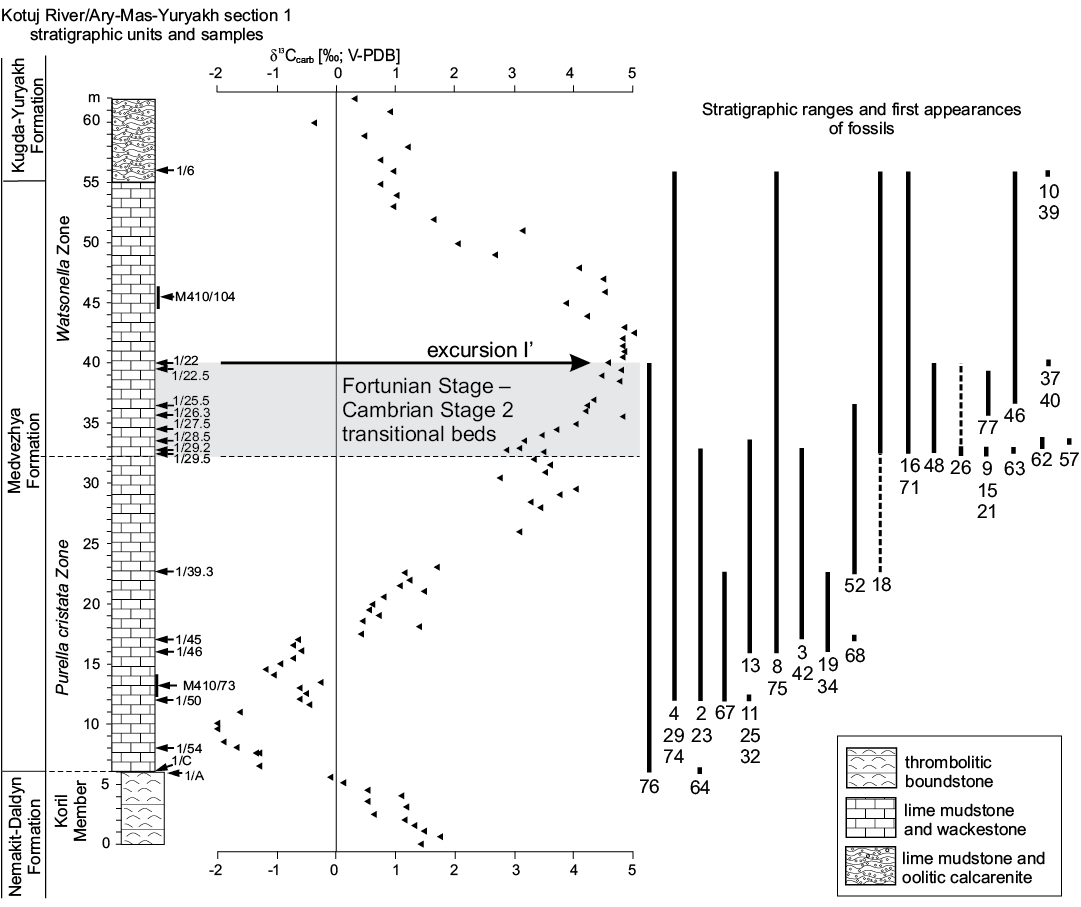
Fig. 4. The δ13Ccarb curve for Kotuj River section 1and sampled horizons with fossiliferous samples (arrows with numbers). Shift in stratigraphic column at Nemakit-Daldyn–Medvezhya formations contact indicates position of sequence boundary. Stratigraphic ranges and first appearances of fossils (see SOM 1): 2, Oelandiella korobkovi; 3, Securiconus cf. simus; 4, Bemella jacutica?; 8, Anabarella plana; 9, Watsonella crosbyi; 10, Pseudoyangtzespira selindeica; 11, Aldanella crassa; 13, Aldanella attleborensis; 15, Philoxenella spiralis; 16, Barskovia hemisymmetrica; 18, Khairkhania cf. evoluta; 19, Ceratoconus striatus; 21, Salanyella cf. costulata; 23, Purella cristata; 25, Yunnanopleura sp.; 26, Obscurania auriculata; 29, Protoconus cf. crestatus; 32, Siphogonuchites triangularis; 34, Lopochites sp.; 37, first appearance of Halkieria sp.; 39, first appearance of Chancelloria sp.; 40, Tianzhushanella sp.; 42, Khetatheca cotuiensis; 46, Turcutheca sp.; 48, Orthothecid operculum type A; 52, Fomitchella infundibuliformis; 57, Hyolithellus sp.; 62, Selindeochrea missarzhevskyi; 63, Selindeochrea ternaria; 64, Anabarites trisulcatus; 67, Anabarites kelleri; 68, Anabarites biplicatus; 71, Anabarites hexasulcatus; 74, Tiksitheca licis; 75, Cambrotubulus decurvatus; 76, Platysolenites antiquissimus; 77, Blastulospongia sp.
Fauna of the Terreneuvian Series on the flanks of the Anabar Uplift
The first descriptions of problematic skeletal fossils, particularly hyoliths, from the flanks of the Anabar Uplift appeared in the late 1950s. Hyoliths from the Kotuj River (western Anabar Uplift) were initially investigated by Sysoev (1959a, b). He described a multitude of species under Circotheca Sysoev, 1958, such as C. annae Sysoev, 1959b; C. anulata Sysoev, 1959b; C. astragaliformis Sysoev, 1959b; C. cotuiensis Sysoev, 1959b; C. limulirata Sysoev, 1959b; C. milachevae Sysoev, 1959b; C. planestria Sysoev, 1959b; C. rugata Sysoev, 1959b; and C. undata Sysoev, 1959b. Other species were assigned by Sysoev (1959a) to Orthotheca Novák, 1886, emend. Sysoev (1958). These include O. artuventrala Sysoev, 1959a; O. collatensa Sysoev, 1959a; O. dorsocava Sysoev, 1959a; O. taimyrica Sysoev, 1959a; and O. ventrocava Sysoev, 1959a. Tubular fossils described by Sysoev later included Ceratotheca cambriensis Sysoev, 1960b, and those erroneously ascribed to the originally organophosphatic Torellella Holm, 1893, such as T. duplastica Sysoev, 1963; T. rhombiformis Sysoev, 1963; T. triplastica Sysoev, 1963; and T. valguanulata Sysoev, 1963. Most of these species were never identified in collections reported by other authors. The fossils were collected in 1956 along the Kotuj River, from outcrops situated 3 km upstream to 6 km downstream from the mouth of the Kotujkan River, most likely from the Kugda-Yuryakh Formation, which is assigned herein to Cambrian Stage 2. From this stratigraphic interval, Sysoev (1965: fig. 2) also published the first figures of anabaritids known from the Siberian Platform and reported other problematic tubular fossils defined by him as hyoliths. Specimen of Hyolithellus sp. in Sysoev’s (1965: fig. 2) has been reinterpreted by Kouchinsky et al. (2009) as representing Anabarites tristichus Missarzhevsky in Rozanov et al., 1969 (see below).
Further information about the problematic skeletal fossils from the western Anabar Uplift was reported after Vladimir V. Missarzhevsky’s fieldwork in that area in 1965‒1966 (see Rozanov et al. 1969: 6). He recognized and illustrated probable orthothecids as Turcutheca annae (Sysoev, 1959b); T. cotuiensis (Sysoev, 1959b); and T. rugata (Sysoev, 1959b). Among the orthothecid hyoliths first described from the Medvezhya Formation are Allatheca concinna Missarzhevsky in Rozanov et al., 1969 (with a holotype from sample M410/104 from the Ary-Mas-Yuryakh section) and Kotuitheca curta Missarzhevsky, 1974. Korilithes bilabiatus Missarzhevsky in Rozanov et al., 1969 (with a holotype from sample M410/104) was reported and attributed by Missarzhevsky to the hyolithid hyoliths.
In 1967‒1968, Anatolij K. Val’kov obtained new material during his fieldwork on the eastern flank of the Anabar Uplift, along the Malaya Kuonamka and Bol’shaya Kuonamka rivers. He described new occurrences of orthothcid hyoliths from the Emyaksin Formation (Val’kov 1975) as Turcutheca rugata (Sysoev, 1959b) and Ladatheca annae (Sysoev, 1959b) together with new forms from higher beds of the lower Emyaksin Formation, i.e., Allatheca anabarica Val’kov, 1975 and A. cana Val’kov, 1975. A. anabarica was later considered as a probable junior synonym of A. corrugata Missarzhevsky in Rozanov et al., 1969, which is also known from that area (Anatolij K. Val’kov, personal communication 1996).
Missarzhevsky (1967) initially described anabaritids in his doctoral candidate thesis as a group of serpulid worms, and his first publications on anabaritids appeared in 1969 (Voronova and Missarzhevsky 1969; Rozanov et al. 1969). Anabarites trisulcatus Missarzhevsky in Voronova and Missarzhevsky, 1969, the first valid genus and species assigned to that problematic group was reported from the Nemakit-Daldyn Formation of the western Anabar Uplift. Anabarites ternarius, A. tricarinatus, A. tripartitus, A. tristichus, and Cambrotubulus decurvatus were among the anabaritids first described by Missarzhevsky (in Rozanov et al. 1969) from the Medvezhya Formation. The tubular fossils Kugdatheca voluta Missarzhevsky in Rozanov et al., 1969 and Tiksitheca licis Missarzhevsky in Rozanov et al., 1969 were initially regarded as orthothecid hyoliths by Missarzhevsky (in Rozanov et al. 1969), but were subsequently transferred by Missarzhevsky (1974) to the anabaritids (see also Kouchinsky et al. 2009). Missarzhevsky (1989: 191, 194) also described Anabarites kelleri Missarzhevsky, 1989 and Kotuites biplicatus Missarzhevsky, 1989 from the lower Medvezhya Formation.
Anabaritids were described from the Manykay Formation of the Malaya Kuonamka River under the new order Angustiochreida by Val’kov and Sysoev (1970), including such new forms as Angustiochrea lata Val’kov and Sysoev, 1970; Longiochrea rugosa Val’kov and Sysoev, 1970; and Aculeochrea ornata Val’kov and Sysoev, 1970 in an assemblage with Anabarites trisulcatus. The same species were also reported from the Manykay Formation at Bol’shaya Kuonamka River (Val’kov 1975: 8). Val’kov (1975: 40) reported molluscs and anabaritids such as Oelandiella korobkovi Vostokova, 1962; Anabarella cf. plana Vostokova, 1962; Cambrotubulus decurvatus Missarzhevsky in Rozanov et al., 1969; C. sibiricus (Val’kov, 1968); A. trisulcatus; “and others” from sample 1282, which he collected in 1966 from the upper Nemakit-Daldyn Formation at the mouth of the Kotujkan River, western Anabar Uplift (section 2 herein). Molluscs, anabaritids, and problematic sclerites from the same sections were collected in 1972 and confirmed but not described by Shishkin (1974). Anabarites rarus Fedorov, 1984 was additionally reported from the upper part of the Nemakit-Daldyn Formation by Fedorov (1984). From Val’kov’s sample 1282, Bokova (1985) described such new forms as Anabarites modestus Bokova, 1985; Kotyikanites sulcatus Bokova, 1985; K. vallatus Bokova, 1985 (see Kouchinsky et al. 2009), along with fossils of molluscan affinity, such as Sachites projectus Bokova, 1985; Paleosulcachites subremualis Bokova, 1985; Stenothecoides pandus Bokova, 1985; S. accuratus Bokova, 1985; and Securiconus incertus Bokova, 1985 (see below).
Molluscs from the Anabar Uplift, such as Aldanella attleborensis (Shaler and Foerste, 1888); Anabarella plana Vostokova, 1962; and Philoxenella spiralis Vostokova, 1962 were initially studied by Vostokova (1962), along with Oelandiella korobkovi Vostokova, 1962, from the Kotuj River. From the Medvezhya Formation of the western Anabar Uplift, Missarzhevsky in Rozanov et al., 1969 reported a number of molluscan fossils, such as Bemella jacutica Missarzhevsky in Rozanov and Missarzhevsky, 1966; Igorella ungulata Missarzhevsky in Rozanov et al., 1969; Latouchella korobkovi (Vostokova, 1962); L. memorabilis Missarzhevsky in Rozanov et al., 1969 (the latter species synonymized by Gubanov and Peel (1999) with Oelandiella korobkovi); Anabarella plana Vostokova, 1962; Aldanella crassa Missarzhevsky in Rozanov et al., 1969; A. utchurica Missarzhevsky in Rozanov et al., 1969; and A. attleborensis (Shaler and Foerste, 1888). Material collected by Missarzhevsky from the Medvezhya Formation of the Kotuj River yielded more molluscs. These included Securiconus costulatus Missarzhevsky, 1989 and sinistrally coiled Barskovia hemisymmetrica Golubev, 1976. Molluscan faunas from the eastern Anabar Uplift were first reported by Val’kov (1975, 1987), but no new molluscan species were published therein from the lower Emyaksin Formation of the Bol’shaya Kuonamka and Malaya Kuonamka rivers.
Tubular, originally phosphatic remains of hyolithelminths occur in the Nemakit-Daldyn, Medvezhya, and Emyaksin formations (Rozanov et al. 1969; Val’kov 1975, 1987; Meshkova et al. 1976; Khomentovsky and Karlova 1993; Vasil’eva 1998). However, they are not common and have not been previously adequately illustrated from the region.
Other phosphatic fossils, such as conoidal sclerites of the problematicum Fomitchella infundibuliformis Missarzhevsky in Rozanov et al., 1969, were initially reported from the north-western and western Anabar Uplift, including outcrops along the Kotuj River (Rozanov et al. 1969). From the lower Emyaksin Formation on the Bol’shaya Kuonamka River, Fomitchella sp. was first reported as Oneotodus? sp. (Val’kov 1975: 8). Fomitchella infundibuliformis and other species of Fomitchella reported from the Bol’shaya Kuonamka River section by Bokova (1992) included F. explicata Bokova and Val’kov in Bokova and Vasil’eva, 1990, F. acutata Bokova in Bokova and Vasil’eva, 1990, and F. oblata Bokova in Bokova and Vasil’eva, 1990.
The tommotiids Camena kozlowskii Missarzhevsky in Rozanov and Missarzhevsky, 1966 (later synonymized to Tommotia kozlowskii) and Camenella garbowskae Missarzhevsky in Rozanov and Missarzhevsky, 1966 were reported from the Medvezhya Formation of the Kotuj River from a horizon ca. 37 m above the top of the Nemakit-Daldyn Formation (sample M410/104 of Rozanov et al. 1969; Fig. 4). However, these finds have not been replicated.
Spiniform sclerites defined as “protoconodonts”, such as Protohertzina anabarica Missarzhevsky, 1973, were first discovered in the lower Nemakit-Daldyn Formation of the western Anabar Uplift by Missarzhevsky, wherefrom they are also reported herein from basal member 3 (see Kaufman et al. 1996). The sclerites have been interpreted as grasping hooks of planktonic chaetognaths (Bengtson 1983). Hertzina pensa Khomentovsky et al., 1982 from the uppermost Nemakit-Daldyn Formation (Koril Member) is probably a morphological variant of sclerites of P. anabarica and was reported by Khomentovsky et al. (1982) in association with Protohertzina sp.; Fomitchella sp.; Purella cristata Missarzhevsky, 1974; Bemella sp.; Helcionella sp.; and Torellella sp. (Khomentovsky et al. 1982).
Tubular remains generally predominate among the acid-liberated fossil assemblages from the Anabar Uplift (SOM 1). Although we were not able to identify any hyoliths from the Nemakit-Daldyn and Manykay formations, a low diversity assemblage of orthothecid hyoliths occurs in higher strata throughout the Medvezhya and lower Emyaksin formations. The upper beds of the Medvezhya Formation yielded abundant, slightly curved, and up to several cm-long Turcutheca-like tubes with oval cross-section. These tubes are not phosphatized, and their internal moulds are usually poorly preserved and cannot be chemically isolated. However, they can be isolated by mechanical preparation or are found in abundance on naturally weathered surfaces of limestone beds. The tubes are generally similar to “Ladatheca” cylindrica Grabau, 1900 from Avalonia, which was regarded as a sessile operculate “worm” by Landing (1988, 1993) and Landing et al. (1989). Abundant almost straight or irregularly curved internal moulds of problematic tubes in insoluble residues from the lower and middle Medvezhya Formation and lowermost Emyaksin Formation have rounded-triangular cross-sections and may largely be attributed to Tiksitheca licis Missarzhevsky in Rozanov et al., 1969, which is assigned herein to the anabaritids. Other forms are not that abundant and include Paragloborilus sp.; Khetatheca cotuiensis (Sysoev, 1959b); Kotujtheca curta Missarzhevsky, 1989; Turcutheca? sp.; undetermined orthothecid hyolith tubes, opercula assigned herein to types A, B, C, and D, as well as other unassigned and poorly preserved opecula. Rare Conotheca cf. mammilata Missarzhevsky in Rozanov et al., 1969 occur in the upper part of the interval studied herein in the Emyaksin Formation (see also Kouchinsky et al. 2015a).
Anabaritids, such as Anabarites trisulcatus and Cambrotubulus decurvatus are locally common in samples from the lower Nemakit-Daldyn Formation of the western Anabar Uplift. These latter species also occur in a more diverse assemblage of anabaritid tubes found in somewhat younger strata of the eastern Anabar Uplift at the top of the boundstone unit in lower section 96-4 at the Bol’shaya Kuonamka River (“marker bed with angustiochreids” in Val’kov 1975: 8; Kouchinsky et al. 1999; Fig. 2). The latter assemblage (the lower assemblage herein) includes common Anabarites latus (Val’kov and Sysoev, 1970), A. cf. trisulcatus, and A. natellus (Val’kov and Sysoev, 1970), as well as rare anabaritids Aculeochrea ornata Val’kov and Sysoev, 1970, A. rugosa (Val’kov and Sysoev, 1970) and rare disarticulated elements of the protoconodonts, such as Protohertzina. A similar association of anabaritids is reported from 30 m above the base of the Manykay Formation at the Malaya Kuonamka River (Val’kov 1970, 1975).
Younger limestone beds containing siliciclastic clay- to sand-sized material occur in the upper Nemakit-Daldyn Formation in the western Anabar Uplift (e.g., sample 1282 in Val’kov 1975; Fig. 2). These and apparently slightly younger conglomerates, that predominantly consist of siliciclastic sand- to gravel-sized clasts with carbonate cement and are attributed to the basal Emyaksin Formation on the Bol’shaya Kuonamka River, contain a more diversified shelly fossil assemblage (the upper assemblage herein). The assemblage includes shells of maikhanellids such as Purella antiqua, siphogonuchitid sclerites, and Oelandiella-like mollusc shells, along with the anabaritids cf. Anabarites trisulcatus.
The overlying Medvezhya and lower Emyaksin formations contain abundant remains attributable to Tiksitheca licis. These remains may co-occur with common Anabarites kelleri, A. biplicatus, and Cambrotubulus decurvatus, as well as relatively rare Selindeochrea tripartita and Anabarites tristichus. Originally calcareous remains also frequently include mollusc shells, notably Oelandiella, Purella, and Aldanella. Halkieriids are represented by rare phosphatized sclerites and internal moulds of Halkieria spp. The earliest halkieriid sclerites are found in the upper Nemakit-Daldyn Formation of the western Anabar Uplift. Chancelloriid sclerites represented by rare internal moulds of disarticulated and occasionally articulated sclerites with phophatized walls occur in the upper parts of the succession studied herein. They are generally referred to Chancelloria symmetrica Vasil’eva, 1985, and Chancelloria sp. (SOM 1).
Subhorizontal traces occur in the upper Manykay Formation of the Bol’shaya Kuonamka River (section 96-4; AK and SB personal observations 1996), but remain unstudied. Kouchinsky (1994) described several ichnofossils from the lower Nemakit-Daldyn Formation at the mouth of the Kotujkan River (section 2 herein). These include the horizontal traces of Olenichnus irregularis Fedonkin, 1985; Torrowangea cf. rosei Webby, 1970; Planolites cf. ballandus Webby, 1970; and Planolithes sp., as well as subvertical burrows. The latter derive from Member 3 (Kaufman et al. 1996; Fig. 1), from the Anabarites trisulcatus Zone and represent the earliest evidence of up to 20-cm deep bioturbation in the Terreneuvian Series of the Siberian Platform (SOM 2). Bioturbation is preserved as irregularly shaped, elongated concretions with a central lumen filled with crystals of dolomite (Kouchinsky 1994). These concretions were interpreted as “diffuse zones of early diagenetic cement around individual burrows” (Kaufman et al. 1996: 512). Similar bioturbation becomes widespread on the Siberian Platform in younger beds.
Faunas attributed herein to the uppermost Terreneuvian Series, upper part of Cambrian Stage 2 (i.e., regional Tommotian Stage) and Cambrian Stage 3 (~Atdabanian–lower Botoman stages) have been discussed and described by Kouchinsky et al. (2015a) from the Emyaksin Formation exposed on the eastern flank of the Anabar Uplift.
Biostratigraphic significance
A number of biozones have been introduced and used for the stratigraphic subdivision of the Terreneuvian-equivalent strata in Siberia comprising the Nemakit-Daldynian and Tommotian stages (e.g., Gubanov and Peel 1999: text-fig. 3). In contrast with three archaeocyathan zones for the Siberian Tommotian Stage, none of the “small shelly fossil” zones has formally been defined according to the concept of the International Stratigraphic Guide (Salvador 1994; Murphy and Salvador 1999) nor has a stratotype. The Interdepartmental Stratigraphic Committee of Russia (Zhamoida and Petrov 2008) accepted among them the two successive, Anabarites trisulcatus and Purella antiqua “small shelly fossil” zones for the Siberian Nemakit-Daldynian Stage. Most small shelly fossils used in regional biostratigraphy of the Terreneuvian Series in Siberia are originally calcareous, often secondarily phosphatized, such as mollusc-like shells, problematic sclerites, and tubes of hyoliths and anabaritids. A minor proportion of the assemblages consits of small skeletal fossils with primary phosphatic, organophosphatic, or entirely organic composition, such as sclerites of protoconodonts and tommotiids, shells of paterinid brachiopods and mobergellids, and tubes of hyolithelminths (e.g., Khomentovsky and Karlova 2002). Due to their original composition, phosphatic or organo-phosphatic fossils have a better biostratigraphic potential than originally calcareous “small shelly fossils” (Brasier 1986, 1989) but occur less frequently. Mollusc-like shells, hyoliths, anabaritid tubes, and problematic sclerites of originally calcareous composition are dominant components in the Fortunian and lower part of Cambrian Stage 2 assemblages discussed herein.
Anabaritids.—Anabaritids are important in the biostratigraphy of earliest Cambrian. Val’kov (1970) described an Angustiochreidae Zone (anabaritids) in the upper Manykay Formation of the eastern Anabar, later renamed by him as the Angustiochrea lata Zone (Val’kov 1975). The Anabarites trisulcatus and Cambrotubulus decurvatus zones were first proposed for the Kessyusa Formation of the Olenyok Uplift by Savitsky et al. (1972), and the C. decurvatus Zone was also recognized by Shishkin (1981) in the upper Manykay Formation on the Bol’shaya Kuonamka River. The A. trisulcatus Zone was identified by Missarzhevsky (1982) on the north-western flank of the Anabar Uplift. Within the same interval in normal marine facies on the eastern flank of the Anabar Uplift, Val’kov (1982) described the successive Anabarites trisulcatus and Angustiochrea lata zones (see also Vasil’eva 1985 and Missarzhevsky 1989).
Presently, Anabarites trisulcatus is ubiquitously used as an index fossil of the lowermost Cambrian. According to Zhamoida and Petrov (2008: 24), the Anabarites trisulcatus Zone in Siberia is characterized by A. trisulcatus, Cambrotubulus decurvatus, Protoherzinia anabarica, P. unguliformis, Chancelloria, and Protospongia. The upper and lower boundaries of the A. trisulcatus Zone are, however, uncertain (Khomentovsky et al. 1998; Khomentovsky and Karlova 2005) within the Siberian Platform. The zone has even been assigned to the upper Ediacaran (Rogov et al. 2015, see Discussion below), given the co-occurrence of anabaritids with the mostly late Ediacaran calcified tubes of Cloudina Germs, 1972 (Vidal et al. 1994; Zhuravlev et al. 2012; Wood et al. 2017; Zhu et al. 2017). Based on rare occurrences of problematic calcified tubular remains below beds assigned by them to the Anabarites trisulcatus Zone, Khomentovsky and Karlova (2005) suggested defining an additional, Cambrotubulus Zone in the lowermost part of the Siberian Nemakit-Daldynian Stage.
Above the A. trisulcatus Zone, but without definition of the stratotypes, Missarzhevsky (1982: 59) proposed the A. signatus, Tiksitheca licis, Tiksitheca korobovi–Aculeochrea composita zones and “beds with Anabaritellus isiticus” in ascending order. An alternative zonation subsequently proposed by Missarzhevsky (1989: 110), also said to be recognizable on the entire Siberian Platform, included a succession of the Anabarites latus, Selindeochrea ternaria (including the Tiksitheca licis and T. korobovi subzones), and S. isitica zones. Among these zones, the Tiksitheca licis Zone was initially equated with the Ajacycyathus sunnaginicus Zone (archaeocyaths) of the lower Tommotian Stage (Rozanov et al. 1969: 41). Assigned to this zone were the lower 30 m of the Medvezhya Formation at Ary-Mas-Yuryakh section (section 1 herein) and 25 m of the Medvezhya Formation at the Kugda (section 3 herein) and Kotujkan (section 2 herein) sections, western flank of the Anabar Uplift. Tiksitheca, then regarded as a hyolith, was subsequently transferred by Missarzhevsky (1974) to the anabaritids. Mostly represented in collections by fragmentary, smooth, and featureless internal moulds of dubious affinity, Tiksitheca licis proved, however, to be difficult to recognize and utilize as index fossil.
The majority of the anabaritid species were originally reported from the Fortunian and lower part of Cambrian Stage 2 of the Siberian Platform, where they are now also rarely known from younger beds including the lower part of the Atdabanian Stage in the eastern Anabar Uplift (Kouchinsky et al. 2015a). Most of the three-rayed, radially symmetrical anabaritid tubes from our collection can be separated into two morphogroups (genera)―those with three prominent longitudinal keels and weakly expressed transverse sculpture (Selindeochrea) and those having variously expressed transverse sculptural elements without the longitudinal keels (Anabarites). All Selindeochrea species, including S. tripartita (Anabarites tripartitus Missarzhevsky in Rozanov et al., 1969) were reported from the upper Fortunian and Cambrian Stage 2 of the Siberian Platform. Rare specimens are also present in a coeval interval in western Mongolia, South China and probably Avalonia (Kouchinsky et al. 2009).
Protoconodonts.—Primarily organo-phosphatic or entirely organic spiniform sclerites of Protohertzina were interpreted as grasping spines of chaetognaths (Szaniawski 1982, 2002; Bengtson 1983; Vannier et al. 2007) and referred to as “protoconodonts” for their microcrystalline, laminated histology (Bengtson 1976; Szaniawski and Bengtson 1993). Landing (1995) emphasized, however, the longitudinally fibrous histology of Protohertzina as “pseudoconodont” and proposed the orders Protoconodontida and Pseudoconodontida. According to Brasier (1989), Missarzhevsky (1989), and Qian and Bengtson (1989), the earliest species Protohertzina anabarica Missarzhevsky, 1973, may include P. anabarica- and P. unguliformis-type sclerites in one apparatus, although they occur in very varying propotions in the different samples and thus both forms are usually reported as separate species in the literature. They also occur in association in the topotype material of P. anabarica illustrated herein (Figs. 56, 57) and are reported from the Anabarites trisulcatus Zone of the Ust’-Yudoma and Nemakit-Daldyn formations (Missarzhevsky 1973, 1982, 1989; Khomentovsky and Karlova 1993; Brasier et al. 1993). For this interval, a Protohertzina anabarica Zone was defined by Missarzhevsky (1982, 1989) in the lower but not the lowermost Nemakit-Daldyn Formation of the Anabar Uplift.
Protohertzina anabarica sclerites may co-occur with the morphologically distinct elements of Mongolodus Missarzhevsky, 1977 (Maldeotaia Singh and Shukla, 1981), so far unknown from the Siberian Platform. Mongolodus spp. occur in the Anabarites trisulcatus–Protohertzina anabarica Assemblage Zone of South China (Qian and Bengtson 1989; Steiner et al. 2003, 2004a, 2007; Vannier et al. 2007; Yang et al. 2014), Lesser Himalayas of India (Singh and Shukla 1981; Brasier and Singh 1987; Hughes et al. 2005) and the equivalent beds of western Mongolia (Esakova and Zhegallo 1996).
Hyolithelminths.—Species-level systematics of hyolithelminths has not yet been revised properly to allow certain identification of species. The earliest forms from upper Fortunian deposits in Siberia are represented by externally smooth tubes of Hyolithellus Billings, 1872 from the upper Ust’-Yudoma (Khomentovsky et al. 1983; Varlamov et al. 2008: fig. 15) and uppermost Nemakit-Daldyn (Khomentovsky and Karlova 1993; Khomentovsky et al. 1982) formations. Rare Hyolithellus vladimirovae Missarzhevsky in Rozanov and Missarzhevsky, 1966 and Hyolithellus sp. occur in the Medvezhya and lower Emyaksin formations (herein). The oldest appearance of hyolithelminths, such as Hyolithellus, in the fossil record is not clearly defined and occurs within Fortunian Stage equivalents in western Mongolia (Voronin et al. 1982; Brasier et al. 1996; Khomentovsky and Gibsher 1996), South China (Qian and Bengtson 1989; Qian et al. 2001), India (Brasier and Singh 1987; Hughes et al. 2005), and Laurentia (Nowlan et al. 1985; Pyle et al. 2006). Torellella, such as T. laevigata (Linnarson, 1871) reliably appear in successions of the Tommotian Stage of Siberia and worldwide, in beds broadly correlated herein with the upper part of Cambrian Stage 2. The oldest undoubted Torellella in our collection is defined as T. cf. laevigata from the lower part of section 96-6 (Kouchinsky et al. 2015a), from beds correlated with the lowermost Tommotian Stage. Several species of hyolthelminths were employed in the biostratigraphy of the Siberian Platform as index fossils, although corresponding biozones lack stratotypes. Missarzhevsky (1989) specified a Torellella lentiformis to T. biconvexa zonal succession through the Tommotian Stage of the Siberian Platform. Bokova (1992) defined beds with Hyolithellus tortuosus at the base of the Tommotian Stage and correlated them with the Allatheca cana and A. anabarica zones (hyoliths; Val’kov 1987) from the Anabar Uplift.
Platysolenites.—A Platysolenites or Platysolenites antiquissimus Zone has long been recognized in Baltica, where the index species appears in Terreneuvian Series-equivalent beds and is traced up to the base of the Schmidtiellus mickwitzi Zone (trilobites; e.g., Urbanek and Rozanov 1983; Moczydlowska 1991), into the beds equivalent to the Cambrian Stage 3. In our material, Platysolenites occurs in the upper Nemakit-Daldyn through the Medvezhya formations and in the lower Emyaksin Formation. It ranges from the Anabarites trisulcatus Zone of the Fortunian Stage through the Purella Zone into beds equivalent to the lower part of Cambrian Stage 2.
Sponge spicules.—The oldest reliably reported sponge spicules have probably a hexactinellid origin and occur in the Protohertzina anabarica Zone, in the lower part of the Middle Dolomite Member, Soltanieh Formation of Iran (Antcliffe et al. 2014), which is broadly correlated with the Fortunian Stage. In South China, hexactinellid sponge spicules occur with small skeletal fossils of the Protohertzina anabarica–Kaiyangites novilis Assemblage Zone (equivalent to the Anabarites trisulcatus–Protohertzina anabarica Zone; Steiner et al. 1993, 2007) and are also reported from slightly older strata (Chang et al. 2017). Despite the evidence from the Fortunian Stage of Gondwana, undoubted originally mineralized sponge spicules are yet unknown from the equivalent deposits on the Siberian Platform (see Kouchinsky et al. 2012: online appendix 1).
Chancelloriids.—Although single sclerites attributed to problematic chancelloriids are reported from the upper Anabarites trisulcatus–lower Purella zones of Siberia (Khomentovsky et al. 1990; Khomentovsky and Karlova 2005; Fig. 5), the oldest undoubted articulated sclerites of chancelloriids are known from beds that can be correlated with the upper Fortunian Stage in Siberia (Val’kov 1983; Khomentovsky et al. 1990; this paper) and South China (Moore et al. 2014). These are reportedly preceded by such related forms as Cambrothyra ampulliformis Qian and Zhang, 1983 from the upper Anabarites trisulcatus–Protohertzina anabarica Assemblage Zone in South China (Moore et al. 2010), yet unknown in the coeval interval in Siberia.

Fig. 5. Stratigraphic ranges of selected species and groups of “small shelly fossils” (SSF) in the Terreneuvian Series of the Anabar Uplift (thick lines), combined with their ranges known elsewhere (thinner lines) and those yet uncertain (dotted lines). Grey fields represent Fortunian–Stage 2 and Stage 2–Stage 3 transitional beds, respectively. Grey line indicates position of the base of Medvezhya and Emyaksin formations. Correlation with South China adapted from Kouchinsky et al. 2012. Zones: Ns, Nochoroicyathus sunnaginicus; Dr, Dokidocyathus regularis; Dl, Dokidocyathus lenaicus–Tumuliolynthus primigenius.
Calcareous mollusc-like shells and problematic sclerites.—On the north-western flank of the Anabar Uplift, the Oelandiella korobkovi–Anabarella plana Zone (molluscs) in the Medvezhya Formation was proposed by Savitsky (1962), where Egorova and Savitsky (1969) further defined the Philoxenella spiralis–Helcionella paupera–Aldanella attleborensis Zone (molluscs). The latter was correlated by Shishkin (1981) with his Aldanella ex gr. attleborensis–Igorella ungulata Zone (molluscs), which he defined in the lower 32 m of the Emyaksin Formation along the Bol’shaya Kuonamka River.
According to Missarzhevsky (1982, 1989), the Medvezhya Formation comprises the Aldanella crassa, Anabarella plana, and Heraultipegma sibirica (now Watsonella crosbyi) zones in ascending order. The latter two zones were assigned to the Tommotian Stage by Missarzhevsky (1982), but only the highest zone was later correlated by Missarzhevsky (1989) with the lower Tommotian. In the underlying upper Nemakit-Daldyn Formation of the western Anabar Uplift, Missarzhevsky (1982, 1989) proposed the Purella cristata Zone.
The Purella antiqua Zone of the upper Nemakit-Daldynian Stage is now accepted by the Russian Stratigraphic Committee (Zhamoida and Petrov 2008). It was first proposed by Khomentovsky et al. (1983) based on the diverse fossil assemblages from ca. 25 m below the top of the Ust’-Yudoma Formation on the lower reaches of the Gonam River (Khomentovsky et al. 1983; Semikhatov and Serebryakov 1983; Val’kov and Karlova 1984) and from the upper Nemakit-Daldyn Formation of the western Anabar Uplift (Meshkova 1974; Val’kov 1975; Bokova 1985). The fossil association of the Purella antiqua Zone from the upper part of the Ust’-Yudoma Formation includes all taxa from the A. trisulcatus Zone and an additional 30 forms, notably P. antiqua, P. cristata, Latouchella korobkovi, Tiksitheca licis, T. korobovi, Archaeospira regularis, Ladatheca annae, Halkieria longa, Siphogonuchites triangularis, Hyolithellus vladimirovaе, Markuella prima, Anabarites tripartitus, and A. signatus, overlain by deposits of the first zone of the Tommotian Stage Nochorojcyathus sunnaginicus (Zhamoida and Petrov 2008: 24).
Morphological differences between the maikhanellids Purella cristata Missarzhevsky, 1974 described from the Medvezhya and upper Kessyusa formations (Missarzhevsky 1974; this paper) and P. antiqua (Abaimova, 1976) from the upper Nemakit-Daldyn Formation (Abaimova 1976; this paper) demonstrate that they represent different species. Purella antiqua occurs in the Nemakit-Daldynian Stage of the Siberian Platform and its equivalents of South China and western Mongolia. Purella cristata, which has smaller and less distinct scales, is more laterally compressed with a more extended apex than P. antiqua and first occurs in slightly younger strata than the oldest P. antiqua. Along with other species or probably synonymous forms (see Parkhaev and Demidenko 2010 and Devaere et al. 2013 for discussion of synonymies), P. cristata ranges into the Tommotian Stage of Siberia and Cambrian Stage 2 equivalents of western Mongolia, East Gondwana (South China, Iran), and West Gondwana (Armorican France). P. antiqua and Yunnanopleura accurata (Bokova, 1985), as well as P. cristata and Yunnanopleura sp. (Figs. 31–37), are tentatively considered herein as stem-group polyplacophoran molluscs, as are also the siphogonuchitids and halkieriids.
Calcareous sclerites of siphogonuchitids reported herein from the upper Nemakit-Daldyn, lower Medvezhya, and lower Emyaksin formations appeared in the Siberian fossil record along with maikhanellid shells in the basal Purella antiqua Zone (Bokova 1985; Missarzhevsky 1989; Khomentovsky et al. 1990; Brasier et al. 1993; Kaufman et al. 1996; this paper). A similar early record of siphogonuchitids and maikhanellids is also known from the lower Meishucunian Stage of South China (Qian and Bengtson 1989; Steiner et al. 2004a, 2007; Yang et al. 2014) and equivalents of western Mongolia (Voronin et al. 1982; Esakova and Zhegallo 1996) and Iran (Hamdi et al. 1989; Brasier et al. 1990) correlated with the Fortunian Stage. The first appearance of halkieriids in Siberia in the upper Nemakit-Daldynian Stage and in the upper part of the Fortunian Stage equivalent beds of western Mongolia, South China, and Avalonia is younger than that of the siphogonuchitids and maikhanellids.
Shells assigned herein to Protoconus Yu, 1979; Algomella Val’kov and Karlova, 1984; Obscurania Devaere et al., 2013; and Olenellina Vasil’eva, 1990 are probably also related to the polyplacophoran stem-group. Purella-like internal moulds, described herein as Protoconus? sp., co-occur with the earliest molluscan fauna from the upper Nemakit-Daldyn Formation on the Kotuj River belonging to the upper Nemakit-Daldynian Stage and correlated herein with the upper Fortunian Stage. Protoconus crestatus Yu, 1979, from the middle Meishucunian Stage of South China (Parkhaev and Demidenko 2010; Yang et al. 2014), Protoconus spp. from Cambrian Stage 2 equivalents of France (Devaere et al. 2013) and the internal mould defined as Purella tianzhushanensis Yu, 1979, from the Latouchella‒Obtusoconus Assemblage Zone IV of Iran (Hamdi 1995: pl. 12: 1, 2) are similar to internal moulds of Protoconus cf. crestatus Yu, 1979, as described herein from the Anabar Uplift, from beds correlated herein with the upper Fortunian‒lower part of Cambrian Stage 2. From the same beds, Obscurania auriculata (Vasil’eva, 1990) and Olenellina alutacea Vasil’eva, 1990, are also reported (see Vasil’eva 1998 and herein). Obscurania spp. are known from presumably age-equivalent beds of South China and France (Parkhaev 2006; Parkhaev and Demidenko 2010; Devaere et al. 2013).
In contrast with the shells of maikhanellids and those of Protoconus–Obscurania-type, conchs of the Ocruranus–Eohalobia group (Qian and Bengtson 1989) are not reliably reported from Siberia. They are common in the upper Fortunian–lower part of Cambrian Stage 2 equivalent strata of Gondwana, i.e., South China (Qian and Bengtson 1989) and France (Kerber 1988; Devaere et al. 2013) and western Mongolia (Esakova and Zhegallo 1996). Ocruranus finial Liu, 1979, with its close and/or synonymous forms in the Ocruranus‒Eohalobia group may also belong to the chiton stem-lineage (Vendrasco et al. 2009). Unlike maikhanellids and Protoconus‒Obscurania-type fossils, their shells consisted of crossed lamellae formed of sub-parallel acicular elements (Vendrasco et al. 2009: pl. 1: 4, 9).
Cyrtoconic shells of Oelandiella- and Bemella-type helcionellids are among the earliest known on the Siberian Platform. The oldest material is represented by internal moulds with poor preservation often referred to as Oelandiella sp. or Oelandiella cf. korobkovi. Oelandiella korobkovi Vostokova, 1962, with characteristic prominent transverse ribs are not convincingly reported from the Nemakit-Daldyn Formation and the lowest occurences of O. korobkovi thus remain questionable. They commonly occur, however, in the overlying Medvezhya and lower Emyaksin formations, in presumably upper Fortunian and lowermost part of Cambrian Stage 2.
The first assemblages of helcionellid molluscs are also reported from the age-equivalent interval in South China, which comprises the Paragloborilus subglobosus–Purella squamulosa Assemblage Zone (Zone 2 of the Meishucunian; Brasier 1989; Steiner et al. 2004a; Devaere et al. 2013). The lower part of that zone was described as the Maikhanella cambriana–Oelandiella korobkovi Subzone, where O. korobkovi and Ocruranus finial Liu, 1979 first appear (Yang et al. 2014). Oelandiella korobkovi was also identified from the Latouchella‒Obtusoconus Assemblage Zone (local Zone IV) of Iran (Hamdi et al. 1989; Hamdi 1995) and the Anabarella plana and Watsonella crosbyi zones of western Mongolia (Voronin et al. 1982; Esakova and Zhegallo 1996), which presumably represent upper Fortunian–lower part of Cambrian Stage 2 (e.g., Landing and Kouchinsky 2016), and from Cambrian Stage 2 of France (Devaere et al. 2013). In our material, the upper range of O. korobkovi extends into Cambrian Stage 2. Undoubted O. korobkovi are not, however, identified from the Atdabanian Stage on the Siberian Platform (correlated with Cambrian Stage 3) and the species disappears in the Tommotian Stage (upper part of Cambrian Stage 2 as correlated herein).
The earliest Obtusoconus honorabilis is known in the fossil record from the uppermost Fortunian Stage in South China, where it marks the Annelitellus yangtzensis–Obtusoconus honorabilis Subzone of Meishucunian Assemblage Zone 2 (Yang et al. 2014). The species reportedly ranges into Cambrian Stage 4 of North China (see Parkhaev and Demidenko 2010). It is also known from upper Fortunian–Cambrian Stage 2 of western Mongolia (Voronin et al. 1982; Esakova and Zhegallo 1996) and Iran (Hamdi 1995). Obtusoconus cf. honorabilis is herein first reported from beds correlated with lower part of Cambrian Stage 2 in Siberia.
Anabarella plana is common on the Siberian Platform, where it first occurs in beds correlated herein with the upper Fortunian Stage and co-occurs with the earliest Watsonella crosbyi in the Fortunian‒Stage 2 transitional strata of the upper Kessyusa, lower Medvezhya, and Emyaksin formations (Val’kov 1987; Bokova 1992; Kouchinsky 1999; this paper). Anabarella plana as well as morphologically similar or synonymous Anabarella spp. occur in the upper Fortunian Stage of Baltica (Mens and Isakar 1999; Isakar and Peel 2007), western Mongolia (Voronin et al. 1982; Esakova and Zhegallo 1996; Brasier et al. 1996; Gubanov 1998, 2002; Demidenko and Parkhaev 2014), South China (Luo et al. 1982; Xing et al. 1984), and Spain (Gubanov 1998, 2002; Vidal et al. 1999). In Avalonia, rare specimens identified by Landing et al. (1989) as A. plana but pending revision are reported from the uppermost Watsonella crosbyi Zone and middle Camenella baltica Zone (lower Stage 3). In Siberia, A. plana disappears from the fossil record by the end of the Tommotian Stage (upper part of Cambrian Stage 2).
Watsonella crosbyi is commonly used as an important Cambrian Stage 2 index fossil (e.g., Li et al. 2011; Peng and Babcock 2011). Its longest stratigraphic ranges are known from Avalonia (Cambrian Stage 2‒lower part of Cambrian Stage 3 according to Landing et al. 1989, 2013; but see Steiner et al. 2007 for alternative correlation). The species’ range is limited to Cambrian Stage 2 equivalents in South Austarlia (Jacquet et al. 2016; but extending into the Cambrian Stage 3 according to Gravestock et al. 2001). In South China, Watsonella occurs in the middle Meishucunian Dahai Member of Yunnan, the upper Maidiping Formation of Sichuan, and the topmost Yanjiahe Formation of Hubei, all of which are considered now as Cambrian Stage 2 (Li et al. 2011; Yang et al. 2014). In western Mongolia, Watsonella is presently known from the upper Bayan Gol Formation of the Salany Gol section (Esakova and Zhegallo 1996; Demidenko and Parkhaev 2014), where its upper range within Cambrian Stage 2 is truncated at a depositional sequence boundary. Some uncertainty existed with stratigraphic position of the tectonically truncated Heraultia Limestone in France and its associated range of W. crosbyi. The Heraultia Limestone was originally correlated with the upper Atdabanian to lower Botoman (Kerber 1988). However, Gubanov (2002: 164, fig. 2) correlated it with the lower Tommotian Stage of Siberia. A revision of the small skeletal fossil assemblage resulted in the assignment of the Heraultia Limestone to the Terreneuvian Series by Devaere et al. (2013). In the classical Aldan River sections of southern Siberia, Watsonella is known to have its FAD in the basal Tommotian (Varlamov et al. 2008) and ranges into the lower Tommotian Stage of Siberia. The oldest W. crosbyi from the western Anabar Uplift is described herein from the middle Medvezhya Formation, from beds just below δ13Ccarb excursion Iʹ (named “I prime” by Knoll et al. 1995b; Kaufman et al. 1996; Fig. 4).
Asymmetrically coiled shells of Aldanella spp. are far more common in Siberia than Watsonella crosbyi. The lower ranges of Aldanella crassa and A. attleborensis in northern Siberia are known from the beds equivalent to the upper Fortunian (Knoll et al. 1995b: fig. 2; Missarzhevsky 1989; Varlamov et al. 2008; Parkhaev and Karlova 2011; this paper) and are older than the lowest known occurrences of Watsonella crosbyi known in Siberia (this paper). Aldanella attleborensis has been suggested as an index fossil for correlation of the base of Cambrian Stage 2 (Rozanov et al. 2008; Parkhaev et al. 2011; Demidenko and Parkhaev 2014). It has been thought to be limited to the Tommotian Stage of Siberia in these reports, whereas A. crassa first appears in strata older than the basal Tommotian (Missarzhevsky 1989; Parkhaev and Karlova 2011; this paper).
Other problematic mollusc-like fossils, such as the sinistrally coiled Barskovia (morphologically close or probably synonymous to the genus Nomgoliella from western Mongolia), dextrally coiled Philoxenella and the planispiral involute and evolute forms of Khairkhania (Esakova and Zhegallo 1996) have lowest known occurences in the upper Nematit-Daldynian strata of Siberia correlated with the upper Fortunian in this report.
Hyoliths.—Since the early development of lower Cambrian biostratigraphic schemes based on hyoliths (i.e., Sysoev 1968, 1972; Meshkova 1969), this faunal group has remained a focus of research in Siberia (Val’kov 1975, 1982, 1987). In the Medvezhya and lower Emyaksin formations of the Anabar Uplift, the Spinulitheca rotunda Zone (Val’kov 1982, 1987) followed by the Allatheca cana Zone (orthothecid hyoliths; Val’kov 1975) were defined. In the western Anabar Uplift, the Ladatheca dorsocava and L. annae zones in ascending order were proposed by Missarzhevsky (1982, 1989). These biozones are, however, loosely defined and lack stratotypes.
Fossils attributable to hyoliths were not found in this study in the Nemakit-Daldyn and Manykay formations of the Anabar Uplift. Although reported in early publications from the upper Nemakit-Daldyn Formation of the western Anabar (Meshkova 1974b; Shishkin 1974; Bokova 1985; Val’kov 1987; Missarzhevsky 1989), they may represent the tubes described as anabaritids. Hyolith tubes attributed to the orthothecids become locally abundant and moderately diverse in the lower Medvezhya and Emyaksin formations, in beds correlated herein with the uppermost Fortunian Stage. They are represented therein by elongated, straight or slightly curved tubes with oval or rounded triangular cross-sections that have commonly been assigned to Turcutheca Missarzhevsky in Rozanov et al., 1969; Ladatheca Sysoev, 1968; or Khetatheca Missarzhevsky, 1989; and by shorter forms with a bluntly rounded or bulbous initial part and a gentle curvature in one plane, such as Lophotheca Qian, 1977; Paragloborilus He in Qian, 1977; and Conotheca Missarzhevsky in Rozanov et al., 1969.
Opercula of hyoliths have not yet been used in biostratigraphy. The earliest forms from the Fortunian Stage equivalents are not subdivided into dorsal and ventral lobes and thus represent orthothecid hyoliths. Opercula of hyoliths from the Fortunian Stage are internally smooth, without morphological subdivision of the inner side. These opercula are often thought to represent helcionellid-type molluscs or identified as shelly problematica (see discussions by Qian and Bengtson 1989; Parkhaev and Demidenko 2010). Their inner surface has a simpler morphology compared to that in younger claviculate forms (Kouchinsky et al. 2015a). The latter opercula appear in our material in beds attributed herein to lower part of Cambrian Stage 2 and become widespread thereafter, in the Siberian Tommotian Stage.
The association between all of the aforementioned opercula and the conchs of individual hyolith species is usually unknown or very tentative. Hyoliths belonging to the form-genus Conotheca have a circular, transverse aperture of the conchs. Their circular opercula are flat. They may have two long tooth-like cardinal processes in specimens known from the Atdabanian Stage (~Cambrian Stage 3; Kouchinsky et al. 2015a). Opercula attributed to Conotheca from the Terreneuvian Series do not, however, have the processes (e.g., Devaere et al. 2013).
The earliest appearance of hyolithid hyoliths, i.e., hyoliths with operculum subdivided into ventral and dorsal areas, or lobes, which were probably connected in such hyoliths to the retractile appendages, or helens, is in the upper part of Cambrian Stage 2 (middle‒upper Tommotian Stage of Siberia) of the southeastern Siberian Platform (e.g., Sysoev 1972). The hyolithid Burites erum Zone was named by Val’kov (1987) from the upper Tommotian Stage on the northern Siberian Platform. A problematical Korilites bilabiatus Missarzhevsky in Rozanov et al., 1969 has been regarded as an early hyolithid (Rozanov et al. 1969: 135). It was described from sample M410/104 (ca. 40 m above the base of the Medvezhya Formation; see Fig. 4 herein) in the Ary-Mas-Yuryakh section, from beds attributed herein to the Cambrian Stage 2. Meshkova et al. (1976: 9) also report undetermined poorly preserved hyolithids from the upper Medvezhya Formation in this section.
Tianzhushanellids.—Tianzhushanellids are represented by rare, disarticulated, probably originally bivalved and calcareous shells, which regarded either as problematic molluscs or stem-group brachiopods (Li and Chen 1992; Gravestock et al. 2001; Li et al. 2014). Tianzhushanella spp. or Lathamella spp. occur in the Watsonella crosbyi Assemblage Zone of middle Meishucunian Stage of South China (Li and Chen 1992) and have been recovered in this study from the coeval interval of northern Siberia attributed herein to lower part of Cambrian Stage 2.
Tommotiids and paterinids.—Phosphatic sclerites of such tommotiids as Camenella garbowskae, Lapworthella tortuosa Missarzhevsky in Rozanov and Missarzhevsky, 1966, Sunnaginia imbricata Missarzhevsky in Rozanov et al., 1969, and Tannuolina pavlovi Kouchinsky, Bengtson, and Murdock, 2010, as well as paterinid brachiopods, are known from the basal Tommotian Stage of the Siberian Platform (Sokolov and Zhuravleva 1983; Rozanov and Sokolov 1984; Rozanov et al. 1992; Khomentovsky and Karlova 1993; Rowland et al. 1998; Sipin 2001; Varlamov et al. 2008). Without description and illustration, the tommotiids Camenella spp. and Sunnaginia? sp. have been reported from outcrops of the Medvezhya Formation (Rozanov et al. 1969; Meshkova et al. 1976; Fedorov and Shishkin 1984; Khomentovsky and Karlova 1993; Knoll et al. 1995b; Luchinina et al. 1997). The same sections studied herein failed to yield tommotiids. By contrast, Camenella spp. and Lapworthella spp. as well as paterinid brachiopods commonly occur in the middle Emyaksin Formation on the eastern flank of the Anabar Uplift (Kouchinsky et al. 2015a). The tommotiids are also reported from the uppermost beds of the Ust’-Yudoma, Kessyusa, and Sukharikha formations, with ages considered to represent the Cambrian Stage 2 (see references in Kouchinsky et al. 2010, 2012).
The phosphatic remains of tommotiids have been used to define the successive Lapworthella tortuosa and L. bella subzones of the Dokidocyathus regularis Zone (archaeocyaths) of the Tommotian Stage (Rozanov et al. 1969). According to Missarzhevsky (1982, 1989), however, L. tortuosa corresponds to the Nochoricyathus sunnaginicus Zone, whereas L. bella ranges into the Atdabanian Stage. The basal Tommotian Aldanotreta sunnaginensis Zone (paterinid) is defined by Pel’man et al. (1992) within the range of the Nochoricyathus sunnaginicus Zone (archaeocyaths) and is succeeded by the Cryptotreta neguertchenensis Zone (paterinid, renamed Pelmanotreta neguertchenensis by Skovsted et al. 2015) that ranges into the Atdabanian Stage (Cambrian Stage 3).
Outside of Siberia, the first tommotiids are known from lower part of Cambrian Stage 2 of Avalonia (Landing et al. 1989), and from beds correlated herein with the upper Fortunian Stage of western Mongolia (Voronin et al. 1982; Brasier et al. 1996; Khomentovsky and Gibsher 1996; Demidenko and Parkhaev 2014; Smith et al. 2015) and South China (i.e., Porcauricula hypsilippis [Jiang, 1980], see Qian and Bengtson 1989; Li et al. 2007).
Fomitchella.—Phosphatic problematic sclerites Fomitchella infundibuliformis and F. acinaciformis are well-known from the Tommotian Stage of the Siberian Platform. The former species is recognized herein from beds correlated with the upper Fortunian–lower part of Cambrian Stage 2, whereas the latter is limited to Stage 2. A Fomitchella Zone was recognized by Missarzhevsky (1982, 1989) in the Medvezhya Formation of the Anabar Uplift, who regarded the lower part of the zone as older than the Tommotian Stage. By contrast, a Fomitchella infundibuliformis Zone was assigned to the Tommotian Stage by Khomentovsky and Karlova (1992, 2002) and Khomentovsky et al. (1998). The oldest Fomitchella sp. are reported from the upper Nemakit-Daldyn and upper Ust’-Yudoma formations, but they are poorly illustrated (Khomentovsky et al. 1982, 1983; Semikhatov and Serebryakov 1983). Fomitchella occurs in Cambrian Stage 2 of Avalonia (Landing and Brett 1982; Bengtson and Fletcher 1983; Landing 1988, 1995; Landing et al. 1989) and Kazakhstan (Missarzhevsky and Mambetov 1981; Missarzhevsky 1989). It is questionably reported from western Mongolia (Missarzhevsky 1977) and South China (Qian and Bengtson 1989; Li et al. 2007).
Mobergella.—Mobergella is a problematical, phosphatic, operculum-like fossil with muscle scars on the visceral surface (Skovsted 2003; Demidenko 2016) that occurs in the upper Tommotian of the southeastern and northeastern parts of the Siberian Platform, but is so far unknown from sections of the Anabar Uplift. The first appearance of Mobergella in the Lena-Aldan area is used to identify the base of the Dokidocyathus lenaicus–Tumuliolynthus primigenius Zone (e.g., Rozanov and Sokolov 1984: 39–40). A Mobergella or M. radiolata Zone is recognized in the upper Tommotian‒lowermost Atdabanian on the Siberian Platform (Missarzhevsky 1982; Vasil’eva 1998). Mobergella is regarded as important for definition of the base of Cambrian Series 2 and Cambrian Stage 3 (Rozanov et al. 2010). No definite record of mobergellids from the Terreneuvian Series is known outside Siberia (Qian and Bengtson 1989; Skovsted 2003; Streng and Skovsted 2006; Kouchinsky et al. 2012; Demidenko 2016).
Rhombocorniculum.—Rhombocorniculum Walliser, 1958, is known from phosphatic, horn-shaped, exteriorly sculptured sclerites with relatively thick wall composed of microtubules (e.g., Landing et al. 1980; Kouchinsky et al. 2015a). The earliest known species, R. insolutum Missarzhevsky in Missarzhevsky and Mambetov, 1981 occurs in the middle Dokidocyathus lenaicus Zone of the Tommotian Stage (Sokolov and Zhuravleva 1983; Rozanov and Zhuravlev 1992; Varlamov et al. 2008) and equivalent beds of the Emyaksin Formation of the eastern Anabar Uplift correlated with upper part of Cambrian Stage 2 (Kouchinsky et al. 2015a). A nearly equivalent first appearance of R. insolutum is also reported from the Camenella baltica Zone of Avalonia at SE Newfoundland, the lower part of which is correlated with the upper Tommotian (Hinz 1987; Landing 1988; Landing et al. 1998) or basal Atdabanian Stage (Steiner et al. 2007).
Chemostratigraphy and an integrated approach to correlation
The rationale for carbon isotope chemostratigraphy is the prediction of similar global trends in 13C/12C ratio in surface ocean waters recorded by alternating peaks and nadirs in δ13Ccarb in carbonate rock successions. This effect is explained by a relatively short ocean mixing time (ca. 1 ky) compared to the residence time of carbon in the world-ocean (ca. 100 ky). In rock successions that preserve a sufficiently long and continuous record in non-restricted marine environments, the resulting carbon isotopic signature remains interregionally recognizeable despite local sedimentary and diagenetic overprints on absolute δ13Ccarb values. This model works well in the open marine facies belt of early Cambrian age on the Siberian Platform, and has allowed high-resolution intrabasinal and global correlation (e.g., Maloof et al. 2010a, b). Carbon isotope stratigraphy and traditional biostratigraphy complement each other by providing comparative age brackets. However, caution needs to be taken with correlations based either on index taxa where the full range of fossil distribution is not discernable or on sequences of carbon isotope excursions where some peaks are omitted due to tectonic truncation or erosional unconformities.
Early Cambrian carbon isotope chemostratigraphy of the Lena-Aldan area of southeastern Siberian Platform was early elaborated by Magaritz el al. (1986) and Brasier et al. (1993, 1994) and used by Knoll et al. (1995a, b) and Kaufman et al. (1996) for chemostratigraphic correlation of the western Anabar sections with the southeastern Siberian stratotypes of the Tommotian Stage. The carbon isotope chemostratigraphy of Cambrian strata of the Siberian Platform was further refined by Kouchinsky et al. (2001, 2005, 2007, 2008, 2015a), and the detailed isotopic record through the Cambrian System is now known from Siberia (Kouchinsky et al. 2012). From depleted negative δ13Ccarb values at the Precambrian/Cambrian boundary, an overall rising trend with positive peaks reaching 5–7‰ characterises sedimentary sequences of the Terreneuvian Series in Siberia (Ripperdan 1994; Brasier et al. 1994, 1996; Kouchinsky et al. 2007), western Mongolia (Brasier et al. 1996; Smith et al. 2015; Landing and Kruse 2017), Morocco (Maloof et al. 2005, 2010a, b), and South and North China (Li et al. 2009).
Integrated correlation of the Fortunian Stage in Siberia.—The current definition of the Ediacaran‒Cambrian boundary (e.g., Landing 1994; Landing et al. 2013) means that it approximately corresponds to the lower sequence boundary of the Nemakit-Daldyn Formation, and is probably situated in the upper part of the underlying Staraya Rechka Formation of the western flanks of the Anabar Uplift, where a strong δ13Ccarb negative shift is observed (Kaufman et al. 1996; Kouchinsky et al. 2007; N in section 2 in Fig. 1). The negative excursion from the dolostone-dominated upper Staraya Rechka Formation (Anabar Uplift) was correlated by Kaufman et al. (1996) with the uppermost dolostone beds of the Turkut Formation (Olenyok Uplift), where a moderate (ca. -3‰) negative shift of δ13Ccarb values is reported by Knoll et al. (1995a). Although the globally recognized basal Cambrian deep negative trough N (after Kaufman et al. 1996; see section 2 in Fig. 1), or BACE (Basal Cambrian Carbon isotope Excursion; Zhu et al. 2006) is probably recorded in the upper Staraya Rechka and lowermost Nemakit-Daldyn formations, there is no indication of such a pronounced negative shift in the upper Turkut Formation (Nagovitsin et al. 2015; Rogov et al. 2015). Therein, a generally negative δ13Ccarb trend is variably truncated at the top of the formation by deep sub-aerial erosion that preceded deposition of the overlying Kessyusa Group (Pelechaty et al. 1996a, b). Given sporadic occurrence through the entire upper Ediacaran Turkut Formation of problematic tubes identified as Cambrotubulus sp. (Karlova and Vodanyuk 1985; Karlova 1987; Khomentovsky and Karlova 1993), the entire Anabarites trisulcatus Zone of the lower Nemakit-Daldynian Stage in Siberia was assigned to the upper Ediacaran (Rogov et al. 2015). The fossils might, however, represent other Ediacaran tubular organisms predating the Anabarites trisulcatus–Protohertzina anabarica Zone (Zhuravlev et al. 2012). Anabarites trisulcatus is unknown at present from the Turkut Formation (Nagovitsin et al. 2015), and this does not corroborate earlier reports of Anabarites sp. from the Turkut Formation by Galina A. Karlova. Anabarites trisulcatus and the second important taxon of the A. trisulcatus Zone, Protohertzina anabarica, first appear in the lower Kessyusa Group (Nagovitsin et al. 2015).
Based on unpublished material collected by Alexander B. Fedorov in 1981, Zhuravlev et al. (2012), and Zhu et al. (2017) reported the oldest co-occurrence of Cloudina and Anabarites ca. 100 m below the top of the Ust’-Yudoma Formation at the Kyyry-Ytyga (Kyra-Ytyga) section on the Yudoma River. These fossils are placed into the context of carbon isotope chemostratigraphy by Zhu et al. (2017), who show a plateau of positive values for most of the lower and middle Ust’-Yudoma Formation just above a negative excursion at the base of formation. A steady and shallow decrease of values toward the top of formation is interpreted by Zhu et al. (2017) as the onset of the BACE excursion. The Ediacaran–Cambrian boundary negative excursion can not be, however, unequivocally recognized in the Kyyry-Ytyga section. By contrast, a coarsely resolved δ13Ccarb profile from the lower–middle Ust’-Yudoma Formation at the Aldan River (Magaritz et al. 1986; Brasier et al. 1993, 1994; Varlamov et al. 2008) shows a long-lasting rising trend from negative values through the entire Ust’-Yudoma Formation reaching peak values of ca. 3.5‰ (peak of excursion I) in its upper part. Although the co-occurrence of Anabarites spp. and Cloudina reported by Zhuravlev et al. (2012) and Zhu et al. (2017) may hint at a lower stratigraphic range of anabaritids overlapping with the upper range of the typically late Ediacaran cloudinids, the latter can be found as late Ediacaran survivals into the Fortunian Age. This is supported by a report from South China and Kazakhstan (Yang et al. 2016), where cloudinids co-occur with Anabarites trisulcatus and Protohertzina anabarica in basal Cambrian strata just overlying the negative BACE carbon isotope excursion.
Rogov et al. (2015) and Nagovitsin et al. (2015) placed the upper Anabarites trisulcatus Assemblage Zone boundary in the upper Syhargalakh Formation (lower Kessyusa Group) of the Olenyok Uplift. The uppermost Syhargalakh Formation was attributed by them to the Purella antiqua Assemblage Zone. Several forms of anabaritids including annulated tubes of the Aculeochrea rugosa-type (shown but not mentioned by Rogov et al. 2015: fig. 7; cf. Fig. 78 herein) are identified from these beds just below the lowest Trichophycus pedum-type traces reported from the area (Rogov et al. 2015: fig. 2). However, such index fossils as Purella spp. or any molluscan fossils are unknown from the Syhargalakh Formation, and assignment to the Purella antiqua Zone is not substantiated biostratigraphically. Rogov et al. (2015) correlated this purported Purella antiqua Assemblage Zone with the entire Fortunian Stage and estimated the maximum age of the zonal lower boundary by a ca. 544 Ma date from the underlying tuff-breccia at the base of the Kessyusa Group (Bowring et al. 1993). Given recent advances in preparation of zircons and recalibration of standards by EARTHTIME protocols (Schmitz 2012), the latter date likely needs to be corrected and is probably a few my younger. This would then correspond to more recent estimates of the Precambrian–Cambrian boundary, as recently corrected geochronology systematically demonstrates younger ages (Compston et al. 2008; Maloof et al. 2010a; Landing et al. 2015 and references therein). Herein, we regard the A. trisulcatus Zone in Siberia to represent the lower Fortunian Stage, whereas the Purella Zone is attributed to the upper Fortunian Stage.
Transition between the Anabarites trisulcatus and Purella zones in southeastern Siberian Platform takes place in beds containing a δ13Ccarb excursion Z (Brasier et al. 1993, 1994) first described from the Ust’-Yudoma Formation at the “Dvortsy” section on the Aldan River. Fossils indicative of the Purella Zone first occur ca. 60 m below the top of the Ust’-Yudoma Formation in the nearby Uchur-Maya region of southeastern Siberian Platform (Khomentovsky et al. 1990). They are correlated by Brasier et al. (1993) with unfossiliferous beds containing excursion Z at “Dvortsy”, where the lower boundary of Purella Zone is placed in the poorly fossiliferous upper Ust’-Yudoma Formation, at the base of the succeeding excursion I (e.g., Rozanov et al. 2008). Extrapolation of peak Z from the Ust’-Yudoma Formation to a positive excursion in Member 3 of the lower Nemakit-Daldyn Formation of the western Anabar Uplift in section 2 (Fig. 1) by Kaufman et al. (1996) is, however, hampered by lack of Purella Zone fossils below Member 8 of the Nemakit-Daldyn Formation (Kaufman et al. 1996; Fig. 1). Strata between these members have not yet yielded SSF assemblages. Anabarites trisulcatus and Protohertzina anabarica first appear in lower Member 3 of the Nemakit-Daldyn Formation. Although abundant anabaritids, such as Anabarites compositus and A. ternarius typically known from the Medvezhya and Emyaksin formations, were reported by Meshkova et al. (1976: 12) from Member 2 of the lower Nemakit-Daldyn Formation, these fossil discoveries have not been replicated and need to be confirmed.
Diverse fossils of the Purella Zone from Member 8 (Kaufman et al. 1996) are located above peak values of 2.0–2.5‰ of a positive δ13Ccarb feature known from the upper Nemakit-Daldyn and Manykay formations in both areas of the western and eastern flanks of the Anabar Uplift (Fig. 1, section 2, sampled horizon 1282; Fig. 2, sections 4 and 5, samples 4/27.5 and 5/0). Correlation of this feature by Kaufman et al. (1996) with the positive δ13Ccarb excursion I from “Dvortsy” is problematical. No biomineralized fossils other than abundant anabaritids and rare Protohertzina occur in the “marker bed” (Val’kov 1970, 1982, 1987; Kouchinsky et al. 1999) at the base of that δ13Ccarb feature on the eastern flank of the Anabar Uplift (Figs. 1 and 2, section 96-4, sample 4/4). Its alternative correlation with an older excursion Z appears therefore to be more plausible (see Kaufman et al. 1996).
A boundary between the Anabarites trisulcatus and Purella antiqua zones tends to be regarded as older than the upper boundary of the Anabarites trisulcatus–Protohertzina anabarica Assemblage Zone 1 of the Meishucunian Stage. The latter boundary is placed above the δ13Ccarb excursion L2 from South China where the latter was correlated by Li et al. (2009) with excursion Z in Siberia. Compared to the A. trisulcatus Zone in Siberia, Meishucunian Assemblage Zone 1 contains a more diverse fauna, also without helcionellid molluscs but with sclerites of siphogonuchitids and maikhanellid shells. These are not present in the A. trisulcatus Zone in Siberia and appear only in the overlying Purella Zone, in assemblages with the lowest helcionellid molluscs. Thus, the Paragloborilus subglobosus–Purella squamulosa Assemblage Zone 2 in South China (Yang et al. 2014) can be correlated herein with the upper part of the Purella Zone in Siberia, where it includes the lower Medvezhya and lowermost Emyaksin formations of the Anabar Uplift (Fig. 5).
Integrated correlation of Cambrian Stage 2 in Siberia.—The upper boundary of the Purella Zone has been correlated with the Fortunian‒Cambrian Stage 2 boundary on the Siberian Platform (Zhamoida and Petrov 2008) and equated with the lower boundary of the Tommotian Stage (Rozanov et al. 2008). In addition to its biostratigraphic characteristics, the base of the Tommotian Stage is also characterised in Rozanov et al. (2008) by declining carbon isotope values through the lower Tommotian boundary in the uppermost Ust’-Yudoma Formation to a minimum in the lower Nochioriocyathus sunnaginicus Zone in the overlying basal Pestrotsvet Formation at “Dvortsy” (Magaritz el al. 1986; Brasier et al. 1993, 1994). Regionally, the diachroneity of the lower boundary of the Pestrotvet Formation (e.g., Val’kov 1987) is supported by oscillating high positive values in the lower Pestrotsvet Formation at the Selinde section in the Uchur-Maya region of the south-eastern Siberian Platform, which are unknown from “Dvortsy” (Kouchinsky et al. 2005). Thus, a relatively older age of the lower Pestrotsvet Formation at Selinde is suggested (see also Val’kov 1982, 1987).
A series of highly positive δ13Ccarb excursions, presently unknown from the Ust’-Yudoma Formation, is reported from the lithologically similar, dolostone-dominated Sukharikha Formation belonging to the same depositional sequence (Kouchinsky et al. 2007). The latter series of excursions allow global chemostratigraphic correlation with a comparable set of excursions known from Morocco (Maloof et al. 2005, 2010a, b). Among them, excursion 5p at Sukharikha was correlated by Kouchinsky et al. (2007) with excursion Iʹ, first described by Knoll et al. (1995b) and Kaufman et al. (1996) from the Medvezhya Formation of the western Anabar Uplift. Knoll et al. (1995b) and Kaufman et al. (1996) preferred correlation of their excursion Iʹ into a supposed 3‒6 Ma hiatus represented by an unconformity above a less pronounced excursion I and between the Ust’-Yudoma and Pestrotsvet formations at “Dvortsy”.
Diverse but often poorly preserved, skeletal fossils, including molluscan shells and sclerites are present in the Purella Zone (Khomentovsky and Karlova 2002), in the Ust’-Yudoma Formation of the Uchur-Maya and Yudoma-Maya regions of southeastern Siberian Platform. They were first reported by Semikhatov and Serebryakov (1983) and Khomentovsky et al. (1983) from ca. 25 m below the formation’s top at Gonam River (elevation 1291 m; see also Khomentovsky and Karlova 1989). These assemblages were later recognized from ca. 60 m below the top of the Ust’-Yudoma Formation in the Mount Konus section on the Uchur River (Khomentovsky et al. 1990) and from the Dzhanda River at a horizon ca. 40 m below the top of the Ust’-Yudoma Formation (Repina et al. 1988; Khomentovsky and Karlova 1989, 2002; Pel’man et al. 1990; Khomentovsky et al. 1990). The latter locality has such molluscs as Anabarella plana and Aldanella attleborensis (including also forms reported therein as A. rozanovi, partly synonymized by Parkhaev and Karlova 2011 with A. attleborensis). The presence of Aldanella spp. in these beds attributed by Khomentovsky and Karlova (2002) to lower Member IV of the upper Ust’-Yudoma Formation is, however, not confirmed by the latter authors. Furthermore, Parkhaev and Karlova (2011) considered the upper Purella Zone to extend into the lower beds of the Medvezhya Formation along the Kotuj River, which yield Aldanella crassa at the base of excursion Iʹ (Figs. 1, 3, 4).
Significantly, a diverse assemblage of fossils assigned herein to the upper Fortunian‒lower part of Cambrian Stage 2 interval, is also known from the uppermost 54 m of the Sukharikha Formation exposed along the Kulyumbe River (Sipin 2001). These fossils occur throughout the upper Sukharikha Formation, but are particularly abundant at the lowest fossiliferous level ca. 50 m below the Sukharikha‒Krasnoporog boundary. In contrast with the Sukharikha River section, the Sukharikha Formation at Kulyumbe River has relatively limited local exposure situated 2‒3 km downstream from Labazny Island (Datsenko et al. 1968). Its most continuous succession there (monoclinally dipping 25–35º and ca. 70 m thick) crops out on the right bank of the Kulyumbe, and passes without visible unconformities into the Krasnoporog Formation (visible thickness ca. 130 m; Sipin 2001).
Following Voronova and Rozanov (1973) and Sipin (2001: 123), the fossiliferous upper Sukharikha Formation at Kulyumbe River is regarded as younger than the equivalent unfossiliferous part of the Sukharikha Formation at Sukharikha River, with a supposed age difference explained by diachronous lithological boundaries between formations. However, given the transgressive onlap associated with a sequence boundary and the predominance of limestones (with some beds having intervening reddish colours) in the upper Sukharikha Formation at Kulyumbe, its supposedly younger age contradicts location of the Kulyumbe area in a relatively deeper normal marine facies belt as compared to the shallower-water transitional facies belt at Sukharikha.
The lowermost fauna with abundant anabaritids at Kulyumbe was assigned to the Anabarites tripartitus–Fomitchella infundibuliformis Zone and regarded as similar to Purella antiqua Zone faunas known elsewhere on the Siberian Platform (Sipin 2001). The overlying Aldanella–Fomitchella acinaciformis Zone suggested for the uppermost ca. 50 m of the Sukharikha Formation (Sipin 2001) contains the regionally lowest known hyolithelminths (Hyolithellus vladimirovae and Torellella curva) and a single reported specimen of Lapworthella tortuosa (not illustrated by Sipin 2001). These strata are overlain by the Halkieria sacciformis‒Tommotia Zone of the lower Krasnoporog Formation. If correlated lithostratigraphically with the upper Sukharikha Formation of the Sukharikha River, the corresponding beds at Kulyumbe belong to the interval with carbon isotope excursions 6p and 7p identified by Kouchinsky et al. (2007). The latter are correlated by Kouchinsky et al. (2007) with oscillating highly positive δ13Ccarb values between excursion Iʹ and lower Tommotian excursion II in the lower Emyaksin Formation (Kouchinsky et al. 2001). Chemostratigraphic data from the Sukharikha and Krasnoporog formations at Kulyumbe are, however, unavailable at present.
Excursion Iʹ of the Anabar Uplift can be correlated (Kouchinsky et al. 2007) with the lower positive δ13Ccarb excursion from the upper Mattaya Formation of the Kessyusa Group in the Olenyok Uplift (upper Kessyusa Formation in Knoll et al. 1995a). An absolute date of 529.7 ± 0.3 Ма was determined from the upper Mattaja Formation on the Khorbosuonka River near the mouth of Mattaya River, ca. 15 m below the base of the Erkeket Formation (Kaufman et al. 2012; Nagovitsin et al. 2015). The dated ash bed is situated at a position within the rising trend of carbon isotope values near the maximum of the lower prominent positive excursion in the upper Mattaja Formation (AK unpublished field and analytical data). Thus, the dated ash bed represents the maximum age of the peak and approximates the appearance of the lowest Watsonella crosbyi known from the Olenyok (Kaufman et al. 2012; Nagovitsin et al. 2015) and Anabar (herein) uplifts. The age estimate is also comparable to that on the lower Watsonella crosbyi Zone of Avalonia (Isachsen et al. 1994; Compston et al. 2008; Landing et al. 2013). Consequently, 530 Ma defines the age of the lower Watsonella crosbyi Zone boundary worldwide.
Based on the absolute age data from the Terreneuvian and its chemostratigraphic correlation, a somewhat older, ca. 531 Ma age can be estimated for the basal transgressive onlap deposits of the Medvezhya and Emyaksin formations on the western and eastern flanks of the Anabar Uplift (Fig. 5). The transgressive beds represented by the basal Pestrotsvet Formation of southeastern part of the Siberian Platform can be dated as 525‒526 Ma in accordance with the chemostratigraphy and the estimated age of the lower Tommotian boundary in Morocco (Maloof et al. 2005, 2010a, b). The basal beds of the Dokidocyathus regularis Zone in the Erkeket and middle Tyuser formations represent the same transgressive event, respectively, in the areas of the Olenyok Uplift and northeastern margin of the Siberian Platform and are ca. 523 Ma old (cf. Maloof et al. 2010b: fig. 1). Thus, the second major transgressive event in the Terreneuvian involved different parts of the Siberian Platform during ca. 8 my in the late Fortunian‒late Cambrian Age 2 interval.
The base of the undefined Cambrian Stage 2 is primarily constrained biostratigraphically by Watsonella crosbyi (Peng and Babcock 2011), which appears just below the peaks of the Siberian excursion Iʹ (Landing and Kouchinsky 2016: fig. 2; Figs. 1, 2) and South China L4/P4 (ZHUCE) carbon isotope excursions (Landing et al. 2013; Landing and Kouchinsky 2016; Fig. 1). The top of Cambrian Stage 2 is often equated with the lower occurrence of trilobites (Peng and Babcock 2011). Taking into account that these first appearances have been interpreted to be interregionally diachronous, Landing and Geyer (2012) and Landing et al. (2013) proposed that the lower and upper Cambrian Stage 2 boundaries are best defined by respectively peaks of δ13Ccarb positive excursion ZHUCE/Iʹ (South China/Siberia) and excursion IV (Siberia). This alternative integrated concept would mean that Watsonella crosbyi first appears in the uppermost Fortunian Stage and the earliest Siberian trilobites from the Profallotaspis jakutensis‒lower Repinaella zones attributed to the uppermost Terreneuvian Series and Cambrian Stage 2. However, variations in the appearance and magnitudes of the positive carbon isotope excursions require further discussion and interpretation in concert with biostratigraphy and geochronology before integrated definition of the Cambrian Stage 2 boundaries becomes acceptable.
There is still no consensus in correlation of the first prominent positive excursions known from South and North China as L4, P4, or ZHUCE (Qian et al. 2001: table 3; Zhu et al. 2001; Li et al. 2009; Maloof et al. 2010a, b; Parkhaev and Demidenko 2010: 892) as well as in interpretation of carbon isotope features D–E and F in Mongolia (Brasier et al. 1996; Smith et al. 2015; Landing and Kruse 2017). Correlation of north Siberian excursion Iʹ with south Siberian excursion I (cf. Pokrovsky and Missarzhevsky 1993; Brasier et al. 1996: 469; but Kaufman et al. 1996) needs to be also considered by taking into account that excursion I first described from “Dvortsy” occurs in dolomitic facies with relatively poorly preserved or non-extractable fossils, in contrast with normal marine facies of the Medvezhya and Emyaksin Formations on the flanks of the Anabar Uplift. The Siberian range of Watsonella crosbyi as documented in this report allows preliminary definition of the stratigraphic interval between the peak of δ13Ccarb excursion Iʹ and the lowest Watsonella crosbyi specimens below this horizon from the Anabar Uplift as the “Fortunian–Cambrian Stage 2 transitional beds”.
Chemostratigraphic correlation across the Anabar Uplift
The carbon isotope chemostratigraphy documented herein allows precise correlation of the western and eastern Anabar Uplift successions with the lower Cambrian stratotypes of the southeastern Siberian Platform and determination of an estimated age of the fossils described in this report (Fig. 1). The interval described underlies the Tommotian‒Botoman δ13Ccarb signature (peaks II‒VII) known in the middle‒upper Emyaksin Formation at Bol’shaya Kuonamka River (Kouchinsky et al. 2001, 2015a) and Pestrotsvet Formation of the Lena-Aldan area (Brasier et al. 1994). Thus, the ages of the Medvezhya Formation as well as the studied major part of the Kugda-Yuryakh and lower Emyaksin formations are older than the base of the Pestrotsvet Formation in the Tommotian Stage stratotype section at “Dvortsy” on the Aldan River (see also Knoll et al. 1995a, b; Kaufman et al. 1996; Landing and Kouchinsky 2016).
The Medvezhya Formation at section 3 is ca. 25 m thick and the lowest oolites appear at the bottom of the overlying Kugda-Yuryakh Formation (Fig. 3). In section 1, located upstream from the confluence of the Kotujkan with the Kotuj River and ca. 17 km SSW of section 3, variegated and greenish colours indicating the Medvezhya Formation persist up to ca. 40 m above the top of the Nemakit-Daldyn Formation’s Koril Member. Oolites are present further up in section 1, but the transition into these oolitic beds is not as abrupt as in section 3. The bottom of the Kugda-Yuryakh Formation in section 1 is indicated by Rozanov et al. (1969: fig. 9) and Meshkova et al. (1976: 9) at ca. 50 m above the top of the Nemakit-Daldyn Formation (Fig. 4). Consequently, the excursion Iʹ interval in section 1 is thicker than that in section 3 (Fig. 1). The stratigraphically higher δ13Ccarb oscillations Iʹa and Iʹb (Figs. 1, 2), incompletely resolved and known from localities 96-5 and 96-5a in the lower Emyaksin Formation of the Bol’shaya Kuonamka, can be correlated with the Kugda-Yuryakh Formation at locality 3 of this report, where highly positive δ13Ccarb values persist up to the top of the examined section (Fig. 3). The Kugda-Yuryakh Formation contains oolitic shoal facies, whose accumulation rate was probably much higher compared to the underlying Medvezhya (sections 1 and 3) and Emyaksin (sections 96-5, 96-5a) formations. Therefore, carbon isotope values in the Kugda-Yuryakh Formation plateau in the upper part of section 3.
Systematic palaeontology
Phylum Mollusca Cuvier, 1797
Class uncertain
Order Helcionellida Geyer, 1994
Family Helcionellidae Wenz, 1938
Genus Oelandiella Vostokova, 1962
Type species: Oelandiella korobkovi Vostokova, 1962; lower Cambrian (upper Fortunian–lower part of Cambrian Stage 2); Kotuj River, Siberia, Russia.
Oelandiella korobkovi Vostokova, 1962
Figs. 6, 7A–C, E, 8A, B.
Material.—Several hundred internal moulds, including figured SMNH Mo181931–181939, 181942–181944, from samples 1/50, 1/46, 1/45, 1/39.3, 1/29.2 (section 1, Fig. 4), K1a/27A, K1a/28A, K1a/82 (section 2, Fig. 1), K2/21, K2/22, K2/23, K2/24, K2/25, K2/26, 3/12, 3/12.2, 3/12.5 (section 3, Fig. 3), 5/10, 5/10B, 5a/1, and 5a/1.2 (sections 96-5 and 96-5a, Fig. 2), Medvezhya and Emyaksin formations (SOM 1), Anabar Uplift, Siberian Platform, Russia. Correlated with upper Fortunian and lower part of Cambrian Stage 2.
Description.—Bilaterally symmetrical, tightly coiled up to one whorl, rapidly expanding and laterally compressed shell covered with distinct transverse rugae. Aperture elongate oval in outline, with length/width ratio 1.5–2.0, gently curved in sagittal plane. Subapical margin of aperture arched. Apical part of shell curved, indistinctly separated and projects slightly beyond the apertural margin. Supra-apical side (dorsum) long and evenly convex. Inner surface of the shell (as replicated by internal moulds) with distinct straight comarginal folds that cross the dorsum.
Remarks.—A potential synonymy of Oelandiella Vostokova, 1962 to Latouchella Cobbold, 1921 was first suggested by Missarzhevsky (in Rozanov et al. 1969 and Missarzhevsky 1989) and is followed by some authors (e.g., Parkhaev and Demidenko 2010: 1054). By contrast, Devaere et al. (2013) maintained the genus Oelandiella and extended its synonymy. It is also worth mentioning that Cobbold (1935) described a new species from the Montagne Noire under the name Latouchella angusta, which Devaere et al. (2013) placed in potential synonymy with O. korobkovi. Although L. costata and O. korobkovi clearly represent two different species, the distinction between the genera Latouchella and Oelandiella is currently weakly constrained.
Latouchella costata Cobbold, 1921, was described from internal moulds from Cambrian Stage 3 of Avalonia (i.e., Shropshire, England). It differs from internal moulds of Oelandiella korobkovi Vostokova, 1962, and O. sibirica Vostokova, 1962, from Siberia mainly by the fact that the transverse ribs do not cross its dorsum (Gubanov and Peel 1998, 1999), as well as by a narrower and more extended apex (compare Gubanov and Peel 1998: fig. 1A with Figs. 6A1, B1, 7E, and 8A1 herein). The apices in L. costata from Shropshire and O. korobkovi from Siberia and China may be laterally deflected in slightly asymmetrical morphological variants (Qian and Bengtson 1989; Gubanov and Peel 1998, 1999; Parkhaev and Demidenko 2010). Gubanov and Peel (1999) also discussed variable density and regularity of the transverse ribbing on internal moulds of O. korobkovi.
Yang et al. (2014: fig. 18 C, D) and Devaere et al. (2013: fig. 4) illustrated internal moulds and conchs with coarse transverse rugae identical to those in our material. Herein, we accept that the O. korobkovi is also present in South China and France according to Devaere et al. (2013). Although Oelandiella–like forms first appear in the upper Nemakit-Daldyn Formation of the western Anabar Uplift (Fig. 9), internal moulds with coarse, high-amplitude, well-spaced corrugation (Figs. 6A–E, 8A, B) are only available herein from the upper Fortunian and Fortunian–Cambrian Stage 2 transitional beds represented by the Medvezhya and lower Emyaksin formations. These internal moulds were defined as Latouchella memorabilis (Rozanov et al. 1969; see also Parkhaev 2008: fig. 3.14 E, F), where they occur with other forms that have lower and denser transverse folds (Fig. 7A, B). By contrast, the conch of O. sibirica Vostokova, 1962, was originally characterised as having an irregular alternation of differently expressed and tightly spaced transverse rugae (see Gubanov and Peel 1999). We concur with the latter authors and find the differences insufficient to differentiate these species. Indeed, some of the moulds of O. korobkovi have irregularly spaced and differently expressed transverse rugae (Figs. 6F, 7C). The ribbing in O. korobkovi from South China and France also appears to express a high degree of variability.
The lower stratigraphic range of O. korobkovi remains questionable, because the oldest material is usually represented by internal moulds of unsatisfactory preservation often referred to as Oelandiella sp. or Bemella sp. Such forms, Oelandiella? sp. and Bemella? sp. (Fig. 9) are available in our collection from the upper Nemakit-Daldyn Formation. In our material, the upper range of O. korobkovi overlaps with Watsonella crosbyi and extends into Cambrian Stage 2 (SOM 1). Undoubted O. korobkovi are not identified from Cambrian Stage 3, or trilobite-containing beds; thus, the species probably disappears in the upper part of Cambrian Stage 2 on the Siberian Platform.
Stratigraphic and geographic range.—Upper Fortunian and Cambrian Stage 2; Siberian Platform and Altai-Sayan Folded area, western Mongolia, and Gondwana (South China, France, and Iran).

Fig. 6. Helcionellid mollusc Oelandiella korobkovi Vostokova, 1962, internal moulds, from early Cambrian Medvezhya Formation, Kotuj River, section 3, western Anabar Uplift, Siberia, Russia; samples K2/25 (A–C), 3/12.2 (D), K2/26 (E), and K2/24 (F). A–F. SMNH Mo181931–181936, respectively. A1, B1, C, D2, E2, F1, lateral views; A2, B3, D1, E3, apertural views; A3, B2, E1, subapical views; F2, upper view. Scale bar 1 mm.

Fig. 7. Helcionellid molluscs, internal moulds, from early Cambrian Medvezhya Formation, Kotuj River, section 1, western Anabar Uplift, Siberia, Russia; samples 1/50 (A, B), 1/46 (C, F), 1/45 (D), and 1/29.5 (E). A–C, E. Oelandiella korobkovi Vostokova, 1962, SMNH Mo181937–181939 (A–C, respectively), SMNH Mo181942 (E). D, F. Securiconus cf. simus Jiang, 1980, SMNH Mo181940 and 181941, respectively. A1–D1, E, F, lateral views; A2, B3–D3, apertural views; A4–C4, D2, subapical views; A3, B2, C2, upper views. Scale bar 1 mm.

Fig. 8. Helcionellid molluscs, internal moulds, from early Cambrian Emyaksin Formation, Bol’shaya Kuonamka River, section 96-5a, eastern Anabar Uplift, Siberia, Russia; sample 5a/1.2. A, B. Oelandiella korobkovi Vostokova, 1962, SMNH Mo181943 and 181944, respectively. C, E. Oelandiella korobkovi? Vostokova, 1962, SMNH Mo181945 and 181947, respectively. D. Securiconus cf. simus Jiang, 1980, SMNH Mo160411 (see also Kouchinsky 2000a: fig. 6A–F). A1, B, C2, D1, E2, lateral views; A2, E1, D2, subapical views; A3, C3, apertural views; E3, D3, upper views; C1, close-up of subapical area with polygons. Scale bar 125 µm (C1) and 1 mm (A, B, C2, C3, D, E).

Fig. 9. Helcionellid molluscs from early Cambrian Nemakit-Daldyn Formation, western Anabar Uplift (A–F) and and basal Emyaksin Formation, eastern Anabar Uplift (G), Siberia, Russia; samples 1282 (A–C) and 2/A1 (D, E), section 2; 3/0.1 (F), section 3; and 4/27.5 (G), section 96-4. A–C, F, G. Bemella? sp., phosphatized shell (A) and calcium phosphatic internal moulds (B, C, F, and G), SMNH Mo181948–181950 (A–C, respectively), SMNH Mo181953 (F), and SMNH Mo181954 (G). D, E. Oelandiella? sp., calcium phosphatic internal moulds, SMNH Mo181951 and 181952, respectively. A2, B1, C2, D2, E2, F1, G2, lateral views; A1, B3, C1, E3, F2, G1, subapical views; B2, C3, E1, F3, G3, upper views; D1, apertural view; B4, close-up of subapical area in B3. Scale bar 1 mm (D, E), 500µm (A, B1–B3, C, F, G), and 100 µm (B4).
Genus Securiconus Jiang, 1980
Type species: Securiconus simus Jiang, 1980; middle Meishucunian Stage (upper Fortunian–lower part of Cambrian Stage 2), lower Cambrian; South China.
Securiconus cf. simus Jiang, 1980
Figs. 7D, F, 8D.
Material.—About 50 internal moulds, including figured SMNH Mo160411, 181940, and 181941, from samples 1/45, 1/29.2 (section 1, Fig. 4), K2/12, 3/6.5 (section 3, Fig. 3), 5/10, 5/10B, 5a/1.2 (sections 96-5 and 96-5a, Fig. 2), Medvezhya and lower Emyaksin formations, Anabar Uplift, Siberian Platform, Russia. Correlated with the upper Fortunian and lower part of Cambrian Stage 2.
Description.—Bilaterally symmetrical, cyrtoconic (up to 1/2 revolution), rapidly expanding shells. Aperture an elongate oval, with length/width ratio ca. 2. Apical part curved with no obvious protoconch boundary and projects far beyond the aperture margin. Subapical side short. Supra-apical side (dorsum) long and evenly convex. Inner surface of the shell (as replicated by internal moulds) smooth or with irregularly spaced and low comarginal folds that cross the dorsum.
Remarks.—The internal moulds are similar in morphology to those identified by Missarzhevsky (1989: pl. 10: 5, 6) as Securiconus simus from lower Medvezhya Formation on the Kotuj River. It is not however clear if the type material of Securiconus simus Jiang, 1980, from the middle Meishucunian Stage of South China represents the same form. From probably the same locality, the holotype of Securiconus costulatus Missarzhevsky, 1989, was described (Missarzhevsky 1989: pl. 7: 5, 9, sample M421/94) and the topotype material was illustrated by Parkhaev (in Rozanov et al. 2010: pl. 33: 1–4). The latter form is wider and has more regularly and distinctly expressed transverse ribs on internal moulds. Securiconus cf. simus differs from Oelandiella korobkovi in being generally lower and cyrtoconic (up to a half revolution) rather than coiled (up to one revolution or more) and having a more tapered apical part. Bemella spp. conchs are significantly wider and have a more rounded apex. Forms from the Nemakit-Daldyn Formation described as S. incertus Bokova, 1985 (Kouchinsky 2000: fig. 6G–L), are similar to Bemella? sp. (Fig. 9A–C, F, G) or Oelandiella? sp. (Fig. 9D, E) illustrated herein, and their affinity is uncertain.
Genus Bemella Missarzhevsky in Rozanov and Missarzhevsky, 1966
Type species: Helcionella jacutica Missarzhevsky in Rozanov and Missarzhevsky, 1966; Dokidocyathus regularis Zone, Tommotian Stage (upper part of Cambrian Stage 2), lower Cambrian; Tiktirikteekh, near Churan village, middle Lena River, Siberia, Russia.
Bemella jacutica? (Missarzhevsky in Rozanov and Missarzhevsky, 1966)
Fig. 10.
Material.—Ten phosphatic internal moulds, including figured SMNH Mo181955–181958, from samples 1/50 and 1/6 (section 1, Fig. 4), 3/12, 3/12.2, and K2/25 (section 3, Fig. 3), Kugda-Yuryakh (1/6) and Medvezhya formations, western Anabar Uplift, Siberian Platform, Russia. Correlated with the upper Fortunian and lower part of Cambrian Stage 2.
Description.—Bilaterally symmetrical, low cyrtoconic (coiled less than 1/2 revolution), rapidly expanding shells. Aperture broad oval in outline, with length/width ratio ca. 1.5 and with laterally straight margins. Apical part declined and projects beyond the apertural margin, apparent protoconch rounded and blunt, hemispherical, separated by a shallow constriction on internal mould. Subapical side short and concave, dorsum long and evenly convex. The inner surface of the shell (as replicated by internal moulds) smooth and carries widely spaced irregularly distributed low comarginal folds that cross the dorsum.
Remarks.—Bemella Missarzhevsky in Rozanov et al., 1969 was described from relatively large mechanically extracted internal moulds from the Medvezhya Formation on the northwest flank of the Anabar Uplift. Bemella jacutica? described herein has diagnostic features of the genus, but has a subdued ornamentation and probably represent small specimens of the type species B. jacutica from the same area and level. Parkhaev (in Gravestock et al. 2001) and Parkhaev and Demidenko (2010) have revised Bemella. The genus is generally similar to Igorella Missarzhevsky in Rozanov et al., 1969, which was also first described from the Medvezhya Formation from mechanically prepared, mm-sized internal moulds. According to Parkhaev and Demidenko (2010) and Missarzhevsky (1989), Bemella is distinguished by having a relatively lower shell than Igorella. Bemella jacutica? is different from Bemella simplex Yu, 1979, from South China described by Parkhaev and Demidenko (2010) in having a higher shell with a more elongate, hooked apical part. Helcionellida indet. are different from Bemella jacutica? in wider shells (Fig. 11A–C) or in a more pointed hooked initial part (Fig. 11D–J).

Fig. 10. Mollusc Bemella jacutica? Missarzhevsky in Rozanov et al., 1969, internal moulds, from early Cambrian Kugda-Yuryakh (1/6) and Medvezhya formations, Kotuj River, western Anabar Uplift, Siberia, Russia; samples 3/12.2 (A), 3/12 (B), and K2/25 (D), section 3; and 1/6 (C), section 1. A–D. SMNH Mo181955–181958, respectively. A1, B2, C3, D3, subapical; A2, B1, C1, D4, lateral; A3, B3, C2, D1, upper; D2, apertural views. Scale bar 500 µm.

Fig. 11. Mollusc Helcionellida indet., internal moulds, from early Cambrian Emyaksin (A, D–J) and basal Kugda-Yuryakh (B, C) formations, Anabar Uplift, Siberia, Russia; samples 5a/34.5 (A), 5/10 (D–F, I, J), and 5/10B (G, H), sections 96-5 and 96-5a; K2/60 (B), section 3; and K1a/67 (C), section 2. A–G, I, J. SMNH Mo181959–181965 (A–G, respectively), SMNH Mo 181966 (I), and SMNH Mo 181967 (J). H. SMNH 167610. A1, B4, C2, E2, F3, upper; A2, B3, D1, E3, F2, I1, H, subapical; B1, C1, D2, E1, F1, G, I2, J3, lateral; I3, J2, apertural views; B2, apical area with polygons enlarged. Scale bar 125 µm (B2), 500 µm (A, B1, B3, B4, C, D1, I, J) and 1 mm (D2, E–G, H).
Genus Tuoraconus Missarzhevsky, 1989
Type species: Tuoraconus acutatus Missarzhevsky, 1989; Dokidocyathus regularis Zone, Tommotian Stage (upper part of Cambrian Stage 2), lower Cambrian; near Chekurovka village, lower Lena River, Siberia, Russia.
Tuoraconus sp.
Fig. 12.
Material.—Three phosphatic internal moulds, SMNH Mo181968–181970, from samples 5a/34.5 (section 96-5a, Fig. 2) and 7/31.5 (section 96-7, Kouchinsky et al. 2015a); Emyaksin Formation, eastern Anabar Uplift. Correlated with lower part of Cambrian Stage 2 (5a/34.5) and Delgadella anabara Zone, Atdabanian Stage (Cambrian Stage 3; sample 7/31.5; see Kouchinsky et al. 2015a: fig. 2).
Remarks.—Internal moulds triangular in transverse view (Fig. 12A2), narrow dorsum, laterally compressed apex projects beyond apertural margin. Imbricated micro-ornament on lateral parts of the mould near the aperture (Fig. 12A4) is similar to that in T. acutatus (Ushatinskaya and Parkhaev 2005: fig. 1) and reminiscent of foliated aragonite fabric (Vendrasco et al. 2011a, b). Specimens from samples 7/31.5 are represented by narrower internal moulds (Fig. 12B, C) than the specimen from sample 5a/34.5 and the material may therefore belong to two different species. Clear distinction is however hampered by preservation.
Genus Obtusoconus Yu, 1979
Type species: Obtusoconus paucicostatus Yu, 1979; middle Meishucunian Stage (upper Fortunian–lower part of Cambrian Stage 2), lower Cambrian; Yichang County, western Hubei Province, South China.
Remarks.—Obtusoconus spp. occur in the uppermost part of Assemblage Zone 2 in South China, where Yang et al. (2014) recognized a new Annelitellus yangtzensis‒Obtusoconus honorabilis Subzone in which the index taxa may co-occur with Obtusoconus rostriptuetus. Obtusoconus honorabilis reportedly ranges from the upper Fortunian to Cambrian Stage 4 (Anhui Province, Lengquanwang Formation; see Parkhaev and Demidenko 2010). A Latouchella‒Obtusoconus Assemblage Zone is known from Iran (upper part of Upper Shale Member, Soltanieh Formation, Alborz Mountains; Hamdi 1995) and correlated with upper Fortunian–Cambrian Stage 2. O. honorabilis also occurs in the upper Fortunian–Cambrian Stage 2 equivalents of western Mongolia (Voronin et al. 1982; Esakova and Zhegallo 1996; Parkhaev and Demidenko 2010). In our material, Obtusoconus cf .honorabilis is found above the first Watsonella crosbyi, in beds that would correspond to the Watsonella crosbyi Assemblage Zone of South China.

Fig. 12. Mollusc Tuoraconus sp., internal moulds, from early Cambrian Emyaksin Formation, Anabar Uplift, Siberia, Russia; samples 5a/34.5 (A) and 7/31.5 (B, C) from sections 96-5a and 96-7, respectively. A–C. SMNH Mo181968–181970, respectively. A1, B1, C2, lateral; A2, B3, C1, subapical; A3, B2, C3, upper views. Scale bar 125 µm (A4) and 500 µm (A2, A3, B, C).
Obtusoconus cf. honorabilis (Qian, Chen, and Chen, 1979)
Fig. 13D, E, J.
Material.—Three phosphatic internal moulds, SMNH Mo181974, 181975, and 181980, from K1a/67 and K8 (section 2, Fig. 1), Kugda-Yuryakh Formation, western Anabar Uplift. Correlated with the lower part of Cambrian Stage 2.
Description.—Bilaterally symmetrical, laterally compressed tall shell. Smaller shells nearly orthoconic (Fig. 13J1), with a slightly laterally deflected apex. Shell bent gently during later growth (Fig. 13D1, E1). Aperture straight in side view, oval in plan view, with length/width ratio of ca. 2. The inner shell surface (as replicated by the internal moulds) has broad transverse folds.
Remarks.—Obtusoconus honorabilis (Qian et al. 1979) was first described from the Meishucunian Stage, Huanshandong Member of the Dengying Formation, Yichang County of the Hubei Province, South China. The fossils also similar to those assigned to Anuliconus Parkhaev in Gravestock et al., 2001, but the uncertainty in taxonomic assignment of these specimens is due to the absence of a protoconch on the available internal moulds. In contrast with Anuliconus, the protoconch in Obtusoconus is displaced and inclined opposite to the curvature of the conch. The protoconch is evidently much smaller than the globular, large protoconch in Obtusoconus rostriptuetus (Qian, 1978). A generally similar form from Spain, O. reduncus (see Gubanov et al. 2004a) has a larger protoconch and is apparently more laterally compressed.
Obtusoconus vostokovae (Vasil’eva, 1990)
Fig. 13F.
Material.—Twelve internal moulds, including figured SMNH Mo181976, from sample 5a/34.5 (section 96-5a, Fig. 2), Emyaksin Formation (SOM 1), Anabar Uplift, Siberian Platform, Russia. Correlated with the lower part of Cambrian Stage 2.
Description.—Bilaterally symmetrical, rapidly expanding orthoconic shells with broad oval aperture (length/width ratio of ca. 1.5) and a globular slightly displaced protoconch. The inner shell surface (as replicated on the internal moulds) has wide and low concentric folds.
Remarks.—The form is originally known as Stephaconus vostokovae Vasil’eva, 1990, from the Emyaksin Formation, Udzha River, northern Siberian Platform, from the Tommotian Stage (after Vasil’eva 1990). Obtusoconus vostokovae is distinguished from other species of the genus by a relatively broader shell and larger globular protoconch.
Stratigraphic and geographic range.—Cambrian Stage 2 of the Siberian Platform.
Obtusoconus cf. mirabilis Vasil’eva, 1990
Fig. 13A, C.
Material.—Two internal moulds, SMNH Mo181971 and 181973, from sample 5a/34.75 (section 96-5a, Fig. 2), Emyaksin Formation (SOM 1), Anabar Uplift, Siberian Platform, Russia. Correlated with Cambrian Stage 2.
Description.—Orthoconic shells with laterally compressed, oval aperture (length/width ratio ca. 2) and a globular, slightly laterally displaced protoconch. The inner shell surface (as replicated on the internal moulds) is smooth.
Remarks.—Type material of Obtusoconus mirabilis Vasil’eva, 1990 derives from a drillcore from the Pestrotsvet Formation in the Lena-Aldan region, from the Tommotian Stage (Vasil’eva 1990). Specimens described herein differ from Obtusoconus mirabilis Vasil’eva, 1990 in lacking regularly spaced transverse folds, but the difference may be preservational. Obtusoconus sp. (Fig. 13B, G–I) is different from Obtusoconus cf. mirabilis in having a broader aperture, more rapidly expanding shell and a smaller, more dorso-ventrally compressed protoconch.
Genus Anabarella Vostokova, 1962
Type species: Anabarella plana Vostokova, 1962; upper Fortunian–lower part of Cambrian Stage 2; lower Cambrian; Kengede River, Siberia, Russia.
Remarks.—Although similar to stenothecids (family Stenothecidae Runnegar and Jell, 1980), Anabarella is herein assigned to the family Helcionellidae, fide Gubanov and Peel (2003) and Vendrasco et al. (2011b).

Fig. 13. Mollusc Obtusoconus Yu, 1979, internal moulds, from early Cambrian Emyaksin (A, C, F, I) and Kugda-Yuryakh (B, D, E, G, H, J) formations, Anabar Uplift, Siberian Platform, Russia; samples 5a/34.75 (A, C) and 5a/34.5 (F, I), section 96-5a, K2/60 (B, G) and K8 (D, E), section 3; K1a/67 (J), section 2; and M424/10 (H), section M424. A, C. Obtusoconus cf. mirabilis Vasil’eva, 1990, SMNH Mo181971 and 181973, respectively. B, G–I. Obtusoconus sp., SMNH Mo181972 (B), 181977–181979 (G–I, respectively). D, E, J. Obtusoconus cf. honorabilis (Qian, Chen et Chen, 1979), SMNH Mo181974 (D), SMNH Mo181975 (E), and SMNH Mo181980 (J), respectively. F. Obtusoconus vostokovae (Vasil’eva, 1990), SMNH Mo181976. A1–H1, I, J1, lateral; A2–H2, apical views; J2, view from right side as oriented in J1. Scale bar 1 mm.
Anabarella plana Vostokova, 1962
Figs. 14, 15.
Material.—About 100 phosphatic internal moulds, including figured SMNH Mo 160430, 182218–182234, from samples 5/10, 5/10B, 5a/1.2 (section 96-5 and 96-5a, Fig. 2), 1/45, 1/46, 1/29.2, 1/6 (section 1, Fig. 4), K2/21, K2/22, K2/24, K2/25, K2/26, 3/12, 3/12.2 (section 3, Fig. 3), Kugda-Yuryakh, Medvezhya, and lower Emyaksin formations, Anabar Uplift, Siberian Platform, Russia. Correlated with the upper Fortunian and lower part of Cambrian Stage 2.
Description.—Bilaterally symmetrical, coiled up to 3/4 of a whorl, rapidly expanding, laterally compressed shell, up to 5 mm in length. Aperture elongate oval with length/width ratio of ca. 3 in planar view, curved in sagittal plane. Subapical margin of aperture arched. Apex curved, no obvious protoconch boundary, projects beyond the apertural margin. Subapical side very short. Supra-apical side long and evenly convex. Inner surface of shell (replicated on internal moulds) smooth with frequent irregular, gentle comarginal folds.
Remarks.—The dorsum of the internal moulds from the Kotuj River (Fig. 14) is often longitudinally depressed (Fig. 14C4, F1) and carries a shallow polygonal microtexture (Kouchinsky 1999). Polygonal patterns were also observed on the apex, including the subapical area (Kouchinsky 1999). The shells of Anabarella plana are slightly involute and reach 10 mm in length (Gubanov and Peel 2003). The internal moulds range from an involute Oelandiella-like type (Fig. 15) to a more evolute Watsonella-like type (Fig. 14; Gubanov et al. 1999; Gubanov and Peel 2003; see the latter report for comparison between the outer shell surface and internal mould). The morphological differences between the Oelandiella- and Watsonella-like forms include a longitudinal dorsal depression in Watsonella-like internal moulds from the Medvezhya Formation compared to a smooth rounded dorsal surface in the more involute forms (Fig. 15) from the lower Emyaksin Formation. Additional material is needed, however, to determine if two different morphs or subspecies of A. plana, or even species of Anabarella are represented.
The earliest Anabarella plana is reported from the upper Nemakit-Daldyn Formation of the western Anabar Uplift (Kotuj River), from the Purella antiqua Zone by Khomentovsky and Karlova (1993), but the fossil is not illustrated. Laterally compressed internal moulds indeed occur in our material from the upper Nemakit-Daldyn Formation (see Fig. 9E), but their affinity with A. plana remains questionable. A. plana reported from the upper Ust’-Yudoma Formation (ca. 40 m below the top) from the Dzhanda River (Pel’man et al. 1990: 16: 2) can also be attributed to the Purella Zone. A. plana frequently occurs in the Medvezhya and lower Emyaksin formations and disappears from the fossil record by the upper part of Cambrian Stage 2 in Siberia. A. cf. plana also occurs in the Lontova Formation (Platysolenites antiquissimus Zone) of Baltica (Mens and Isakar 1999; Isakar and Peel 2007), which can be broadly correlated with the Terreneuvian Series (Gubanov 1998, 2002). Anabarella spp. morphologically similar to A. plana (Landing et al. 1989; Gubanov and Peel 2003) are reported from the upper Fortunian–Cambrian Stage 2 equivalent deposits of the Bayan-Gol Formation of western Mogolia (Voronin et al. 1982; Esakova and Zhegallo 1996; Brasier et al. 1996; Gubanov 1998, 2002; Demidenko and Parkhaev 2014) and from Assemblage Zones 2 and 3 of South China (Luo et al. 1982; Xing et al. 1983). In Western Gondwana (Spain), Anabarella plana Vostokova, 1962 and Oelandiella korobkovi Vostokova, 1962 are reported from the upper part of the lower Corduban Stage, probably also representing the upper Fortunian–Cambrian Stage 2 interval (Gubanov 1998, 2002; Vidal et al. 1999; Kouchinsky et al. 2012). Anabarella sp. reported from the uppermost part of the Watsonella crosbyi Zone in Avalonia ranges much higher, through the middle Camenella baltica Zone (Cambrian Stage 2–lower Cambrian Stage 3; Landing et al. 1989).
Stratigraphic and geographic range.—Upper Fortunian and lower part of Cambrian Stage 2; Siberia, western Mongolia, Baltica, Gondwana (South China, Spain), and Avalonia (eastern Newfoundland).
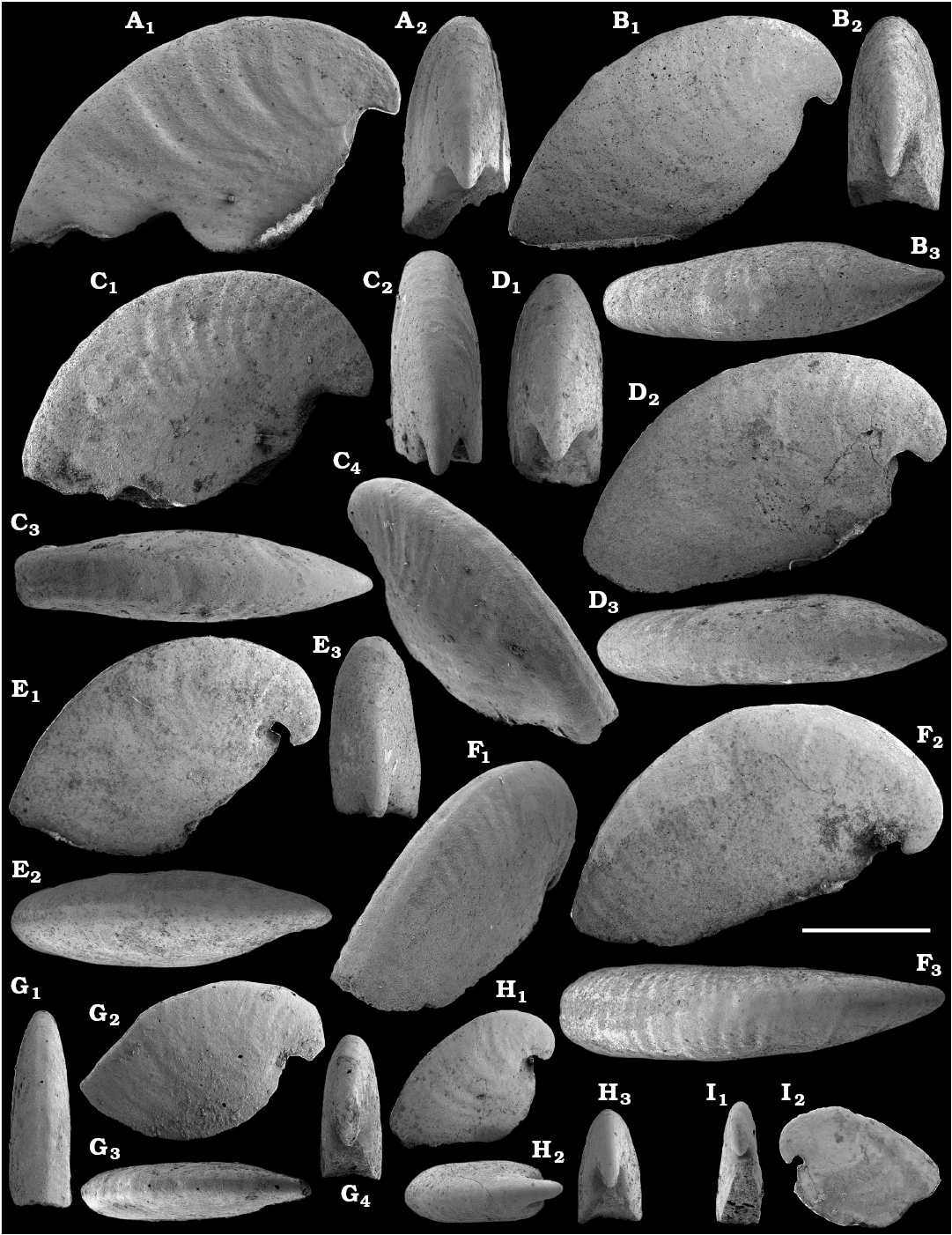
Fig. 14. Mollusc Anabarella plana Vostokova, 1962, internal moulds, from early Cambrian Medvezhya Formation, Kotuj River, western Anabar Uplift, Siberia, Russia; samples 3/12.5 (A), K2/25 (B–F, H), K2/26 (I), section 3; and 1/29.2 (G), section 1. A–I. SMNH Mo182218 (A), SMNH Mo182219 (B), SMNH Mo160430 (C), SMNH Mo182220–182225 (D–I, respectively) (compare C with Kouchinsky 1999: fig. 1A). A1–C1, D2, E1, F2, G2, H1, I2, lateral; A2–C2, D1, E3, G4, H3, I1, subapical; B3, D3, E2, F3, G3, H2, upper; G1, supra-apical view; C4, F1, oblique lateral views showing upper furrow. Scale bar 1 mm.
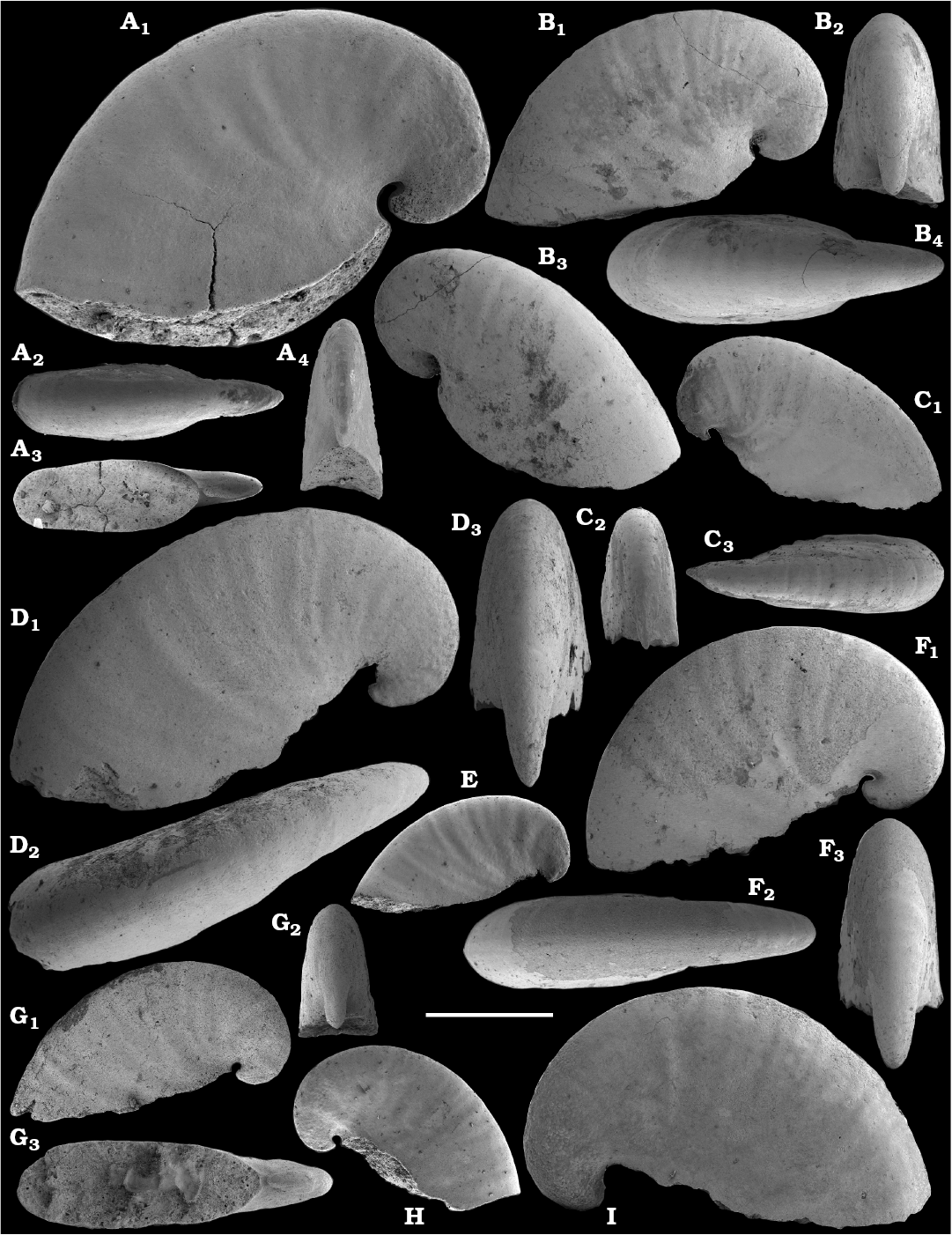
Fig. 15. Mollusc Anabarella plana Vostokova, 1962, internal moulds, from early Cambrian Emyaksin Formation, Bol’shaya Kuonamka River, eastern Anabar Uplift, Siberia, Russia; sample 5a/1.2 from section 96-5a. A–I. SMNH Mo182226–182234, respectively. A1–D1, E, F1, G1, H, I, lateral; A2, B4, C3, D2, F2, upper; A4, B2, C2, D3, F3, G2, subapical; A3, G3, apertural views. Scale bar 500 µm (A1) and 1 mm (A2–A4, B–I).
Order and family uncertain
Genus Watsonella Grabau, 1900
Type species: Watsonella crosbyi Grabau, 1900; Camenella baltica Zone?, upper part of Cambrian Stage 2–lower part of Cambrian Stage 3, lower Cambrian; boulders at Sandy Cove and Pleasant Beach, Cohasset, Massachusetts, USA.
Remarks.—Watsonella crosbyi was proposed as the oldest known rostroconch (Pojeta and Runnegar 1976; Landing 1988; Landing et al. 1989, 2013; Kouchinsky 1999). Based on morphology and shell microstructure, an alternative interpretation is that Watsonella belongs to an evolutionary lineage of helcionellid molluscs that led to the earliest bivalves (i.e., Pojetaia Jell, 1980 and Fordilla Barrande, 1881) rather than to the rostroconchs (Vendrasco et al. 2011a, b).
Watsonella crosbyi Grabau, 1900
Fig. 16.
Material.—Ten phosphatic internal moulds (and fragments), including figured SMNH Mo160433, 182235–182239, from samples 1/29.2, 1/29.5, 1/6 (section 1, Fig. 4), K8 (section 3, Fig. 3), and M424/10 (section M424); Medvezhya and Kugda-Yuryakh formations, western Anabar Uplift, Siberian Platform, Russia. Correlated with the Fortunian–Cambrian Stage 2 transitional beds and lower part of Cambrian Stage 2.
Description.—Bilaterally symmetrical, laterally compressed shell, up 4 mm long. Aperture is very elongate oval in outline, with length/width ratio of ca. 3 in planar view, curved in sagittal plane. Apex blunt evenly expands into the main part of the shell. Subapical side very short. Supra-apical side (dorsum) long, broadly convex in lateral view, flattened to gently depressed in the middle. Subapical and supra-apical margins of aperture arched towards dorsum. Inner surface of the shell (replicated on internal moulds) smooth, with irregularly spaced, gentle comarginal folds.
Remarks.—The outline and width of these internal moulds vary from broad (Fig. 16E) to relatively slim with a slightly more pronounced apex (Fig. 16A, B). The variation agrees well with that observed by Li et al. (2011) and Devaere et al. (2013) and interpreted as intraspecific, but the presence of different species or sub-species of Watsonella cannot be excluded. A medial furrow interpreted as a reflection of a hinge-like structure was observed in the apical part of a single internal mould (Dzik 1994: fig. 12G), but remains, however, questionable, because numerous other specimens from various localities display undivided juvenile shells (Vendrasco et al. 2011a, b; Li et al. 2011; Devaere et al. 2013). Its presence likely can only be confidently assessed in specimens with preserved conchs (e.g., Steiner et al. 2007: fig. 6A, D). A medial furrow along the dorsal surface of the shell was likely present (Landing et al. 1989: fig. 5.20; Steiner et al. 2007: fig. 6D; Li et al. 2011) and is reflected on the available internal moulds by a variably developed longitudinal dorsal depression (Fig. 16A3, B3). The shell microstructure of Watsonella is similar to that in Anabarella plana (see Kouchinsky 1999 and Watsonella-like forms of A. plana in Fig. 14).
Watsonella crosbyi (Grabau, 1900) occurs in the lower Emyaksin (Kengede River; Bokova 1992: 17, 46, fig. 14), Medvezhya and Kugda-Yuryakh formations of the Anabar Uplift, upper Kessyusa Group of the Olenyok Uplift, lower Pestrotsvet Formation of the Lena-Aldan and Uchur-Maya regions of the SE Siberian Platform, and lower Krasny Porog (Krasnoporog) Formation on the NW margin of the Siberian Platform (Bokova 1992; Khomentovsky and Karlova 1993, 2005; Rowland et al. 1998; Kouchinsky 1999; Sipin 2001; and herein). Isotopic data from the Kengede section are unavailable, but the lower Emyaksin Formation at Bol’shaya Kuonamka, eastern flank of the Anabar Uplift, contains prominent positive excursions (see Kouchinsky et al. 2001 and Fig. 2 herein) thought to be characteristic of Cambrian Stage 2 around the world. Herein, Watsonella crosbyi is reported from the middle Medvezhya Formation of the Ary-Mas-Yuryakh section (section 1 this report; see also Landing and Kouchinsky 2016), just below the positive peak of the excursion Iʹ (Knoll et al. 1995a, b; Kaufman et al. 1996).
As in Siberia, Watsonella crosbyi is used as an index fossil in western Mongolia. It is known to occur there in the upper Bayan-Gol Formation, near the basal part of the δ13Ccarb excursion F (Brasier et al. 1996). However, the species has a very limited stratigraphic and geographic distribution in western Mongolia (Esakova and Zhegallo 1996), which limits its use as an index fossil (Demidenko and Parkhaev 2014). A probably significantly truncated range is also known from the Heraultia Limestone, France, where a Watsonella crosbyi‒Oelandiella korobkovi Interval Zone was defined and referred to Cambrian Stage 2 (Devaere et al. 2013). Although Watsonella crosbyi first occurs in significantly younger strata than those with the earliest shelly molluscs in South China, Siberia, and western Mongolia, the Watsonella crosbyi Zone of the Chapel Island Formation on eastern Newfoundland and New Brunswick contains the earliest molluscs in Avalonia, with forms such as W. crosbyi, Aldanella attleborensis (Shaler and Foerste, 1888), and Helcionella sp. (Bengtson and Fletcher 1983; Landing et al. 1989, 1998, 2013; Kouchinsky et al. 2012).
Stratigraphic and geographic range.—Probably the uppermost Fortunian (depending on stratigraphic correlation) and Cambrian Stage 2 of Siberia, Altai-Sayan Folded area, western Mongolia, East Gondwana (South China, Australia), West Gondwana (Armorican France) and Avalonia (Massachusetts, Cape Breton Island, eastern Newfoundland). The species reportedly ranges into the lower part of Cambrian Stage 3 in Avalonia (Landing et al. 1989, 2013) and South Australia (Gravestock et al. 2001; but Jacquet et al. 2016).

Fig. 16. Mollusc Watsonella crosbyi Grabau, 1900, internal moulds, from early Cambrian Medvezhya and Kugda-Yuryakh formations, Kotuj River, western Anabar Uplift, Siberia, Russia; samples 1/29.5 (A–C) and 1/29.2 (E), section 1; K2/60 (F), section 3; and M424/10 (D) from section M424. A–F. SMNH Mo182235–182237 (A–C, respectively), SMNH Mo160433 (D), SMNH Mo182238 (E), SMNH Mo182239 (F) (compare D with Kouchinsky 1999: fig. 3A–C). A1, B2, C1, D2, E1, F, lateral; A2, B1, C2, D1, E3, subapical; A3, B3, C3, D3, E2, upper views. Scale bar 1 mm.
Pseudoyangtzespira Bokova, 1990
Type species: Pseudoyangtzespira selindeica Bokova, 1990; correlated with the uppermost Fortunian or lower part of Cambrian Stage 2, lower Cambrian; Selinde section, southeastern Siberian Platform, Russia.
Remarks.—Devaere et al. (2013) synonymized monospecific genus Pseudoyangtzespira Bokova, 1990, with Oelandiella Vostokova, 1962, whereas Parkhaev and Demidenko (2010) synonymized it with Latouchella Cobbold, 1921. Pseudoyangtzespira is, however, retained as a distinct genus by Parkhaev (2008), Rozanov et al. (2010), Parkhaev and Karlova (2011), and herein. Unlike slightly asymmetrical varieties described among Oelandiella korobkovi (Gubanov and Peel 1999; Parkhaev and Karlova 2011), dextral coiling in P. selindeica is a characteristic feature. Similarity of P. selindeica to Siberian species of Aldanella led Landing et al. (2013: 147) to regard P. selindeica as a junior synonym of A. attleborensis (Shaler and Foerste, 1888).
Pseudoyangtzespira selindeica Bokova, 1990
Fig. 17.
Material.—25 phosphatic internal moulds from sample 1/6 (section 1, Fig. 4) and a single internal mould from K1a/67 (section 2, Fig. 1), including figured SMNH Mo182240–182244, from lowermost Kugda-Yuryakh Formation (SOM 1) and P. selindeica topotype, SMNH Mo182245, from the uppermost Ust’-Yudoma Formation, Selinde section, southeastern Siberian Platform, Russia. Correlated with lower part of Cambrian Stage 2.
Description.—Slightly dextrally coiled up to one whorl, rapidly expanding shell. Spire does not project or slightly projects beyond the upper margin of the aperture (Fig. 17A2, C, D, E2, F2). Aperture extended irregularly oval or ovoid in outline, with length/width ratio ca. 3. Apical part indistinctly separated from rest of conch. Inner surface of the shell (reflected by internal moulds) smooth (Fig. 17A–E) or with low and regular comarginal folds (Fig. 17F).
Remarks.—Pseudoyangtzespira selindeica Bokova, 1990 was first described from the uppermost Ust’-Yudoma Formation of the Selinde section of the southeastern Siberian Platform (Fig. 17F). The specimens from the Anabar Uplift are represented by smooth internal moulds generally similar to A. crassa Missarzhevsky in Rozanov et al., 1969 (compare Fig. 17A–E with Fig. 18), although they derive from younger beds than those containing undoubted topotype A. crassa. P. selineica has a faster expanding, less coiled, more regularly transversely folded shell wall with a more extended apertural outline.
Stratigraphic and geographic range.—Fortunian–Cambrian Stage 2 transitional beds of Siberia.
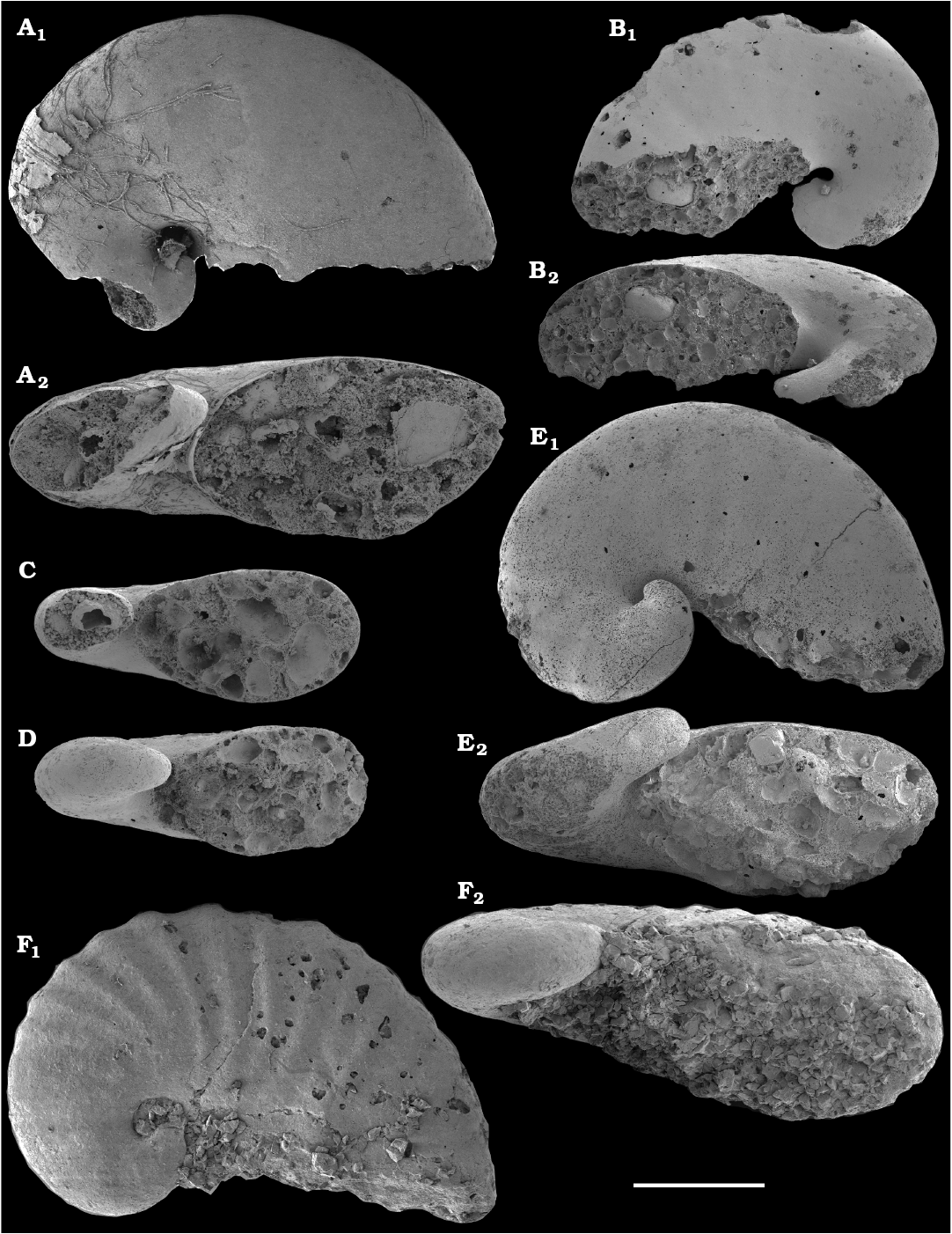
Fig. 17. Mollusc Pseudoyangtzespira selindeica Bokova, 1990 (F topotype), internal moulds, from early Cambrian Kugda-Yuryakh Formation, Kotuj River, western Anabar Uplift; samples K1a/67 (A), section 2; 1/6 (B–E), section 1; S1 (F), Selinde section, southeastern Siberian Platform, Russia. A–F. SMNH Mo182240–182245, respectively. A1, E1, F1, adapical; A2, B2, C, D, E2, F2, apertural; B1, umbilical views. Scale bar 500 µm (A, C–F) and 1 mm (B).
Order Pelagiellida Mackinnon, 1985
Remarks.—The suprageneric assignment of small gastropod-like shells such as Aldanella and Pelagiella Matthew, 1895 from the early and middle Cambrian is problematical. As summarized by Landing et al. (2002: 299), these forms, typically known from conchs with low spires and laterally to ventrolaterally elongate apertures have been variously regarded as untorted or partially torted “pelagiellid monoplacophorans” (Runnegar 1981) or partially torted “paragastropods” (Linsley and Kier 1984).
Landing et al. (1998, 2002) and Parkhaev (2002) interpreted them as archaeogastropods. Pelagiella was alternatively placed among stem-group gastropods (Thomas et al. 2010) or stem-group molluscs (Thomas and Vinther 2012, but see Dzik and Mazurek 2013) based on finds of associated paired chaetal structures.
Family Aldanellidae Linsley and Kier, 1984
Genus Aldanella Vostokova, 1962
Type species: Aldanella attleborensis (Shaler and Foerste, 1888); lower Cambrian; Hoppin Hill, Massachusetts, USA.
Remarks.—The species-level taxonomy of Aldanella is controversial. Isakar and Peel (2007) and Parkhaev and Karlova (2011) revised and compared the named species of Aldanella. The holotype of Aldanella attleborensis (Shaler and Foerste, 1888) from Avalonian eastern Massachusetts (Landing 1988: figs. 5.1, 5.5) and the available lectotype of A. kunda Öpik, 1926 from Baltica (Isakar and Peel 2007: fig. 2E, F, J) are internal moulds, and the holotype of A. attleborensis is a fragment of one. They are less well preserved than most of the material illustrated herein and by Isakar and Peel (2007: fig. 2) from Baltica, as well as by Parkhaev and Karlova (2011) from Siberia. The Avalonian material attributed to A. attleborensis from its type area in eastern Massachusetts and from eastern Newfoundland, southern New Brunswick, and Cape Breton Island ranges from the Watsonella crosbyi Zone through Camenella baltica Zone (terminal Fortunian Stage through lower part of Cambrian Series 2, Cambrian Stage 3). These specimens demonstrate variability in cross-sectional shape of the whorl, inclination and projection of the spire, and development of radial folds on the inner shell surface (Bengtson and Fletcher 1983; Landing 1988, 1991, 1996, 2004; Landing et al. 1989).
This variation has suggested extensive synonymization of Aldanella attleborensis with other named Aldanella species and with Pseudoyangtzespira selindeica in reports by EL. For example, Landing (1988; also Landing et al. 1989, 2013) regarded A. kunda as a junior synonym of A. attleborensis, which was supported by Isakar and Peel (2007) and maintained by Parkhaev and Karlova (2011). Aldanella operosa Missarzhevsky in Rozanov and Missarzhevsky, 1966 occurs through the Tommotian into the lower Atdabanian stages of the Siberian Platform. Given the better preservation of the Siberian material, recognition of such species of Aldanella from Siberia as A. attleborensis, A. crassa, A. sibirica, A. golubevi, A. utchurica, and A. operosa by Parkhaev and Karlova (2011) is maintained in this report.
Aldanella crassa is probably the oldest appearing, named species of Aldanella in Siberia (Missarzhevsky 1989; Parkhaev and Karlova 2011; Parkhaev et al. 2011). In the Anabar Uplift, A. crassa occurs at the base of the rising excursion Iʹ, whereas A. attleborensis appears a few meters above, and its lower range probably overlaps with the upper range of A. crassa (Parkhaev and Karlova 2011). The oldest Aldanella crassa and A. attleborensis are thus older than the lowest known Watsonella crosbyi from Siberia. In Avalonia, A. attleborensis co-occur with W. crosbyi from the base of the Watsonella crosbyi Zone (Landing et al. 2013). The presence of Aldanella in Mongolia is questionable (see faunal lists in Esakova and Zhegallo 1996). Reported from the Terreneuvian Stage of Baltica (Moczydłowska 1991; Gubanov 1998, 2002; Mens and Isakar 1999; Isakar and Peel 2007), A. kunda (Öpik, 1926) from Estonia was synonomised with A. attleborensis, whereas Aldanella polonica Lendzion, 1977, from Poland has been tentatively synonomized with A. crassa by Parkhaev and Karlova (2011). Aldanella attleborensis also occurs in the Dahai Member of Yunnan on the Yangtze Platform and the Yanjiahe Formation of Hubei where it has beed reported as Aldanella yanjiaheensis and co-occurs with Watsonella crosbyi (Chen 1984; Steiner et al. 2007). Pelagiella lorenzi Kobayashi, 1939 from the Upper Dolomite Member of the Soltanieh Formation of the Valiabad section of the Elburz Mountains of Iran (Hamdi 1989, 1995; Hamdi et al. 1989) was assigned by Parkhaev and Karlova (2011) to A. crassa but is somewhat less coiled and has apparently more transversely extended aperture being also similar to Pseudoyangtzespira selindeica (Fig. 17).
Aldanella crassa Missarzhevsky in Rozanov et al., 1969
Fig. 18.
Material.—About 150 phosphatic internal moulds, including figured SMNH Mo182246–182252, from samples 1/50 (section 1, Fig. 4), K2/12, K2/13, and 3/6.5 (section 3, Fig. 3), Medvezhya Formation, western Anabar Uplift, Siberian Platform, Russia. Correlated with the uppermost Fortunian Stage.
Description.—Very rapidly expanding, dextrally coiled shell, up to 3.0 mm in diameter, consists of up to 1.5 whorls. Adapical and umbilical surfaces of the whorl gently convex, spire flat or slightly protrudes above the last whorl. Lateral surface of the whorl broadly rounded. Apertural cross-section an asymmetrical broad oval (width/height ratio 1.6–1.9) that is more strongly convex on the umbilical than the adapical side. Sculpture of the shell interior (as replicated on internal moulds; Fig. 18A1, D) consists of irregular, low transverse folds. Protoconch not clearly delimited, with a diameter of 140–170 µm. Apex of internal moulds bluntly rounded and sometimes with rounded polygonal imprints 20–40 μm in diameter (Fig. 18B2).
Remarks.—Similar to Aldanella attleborensis, the species is, however, generally larger with a higher expansion rate and has a larger protoconch. Aldanella crassa in our material from sections 1 and 3 derives from older beds than A. attleborensis (see SOM 1 for species ranges). This is in agreement with Missarzhevsky (1982, 1989) and Parkhaev and Karlova (2011), who mentioned that A. crassa occurs in slightly older beds. The type material for this species was described by Missarzhevsky (in Rozanov et al. 1969) from sample M410/73 from the Ary-Mas-Yuryakh section (section 1 herein; Fig. 4).
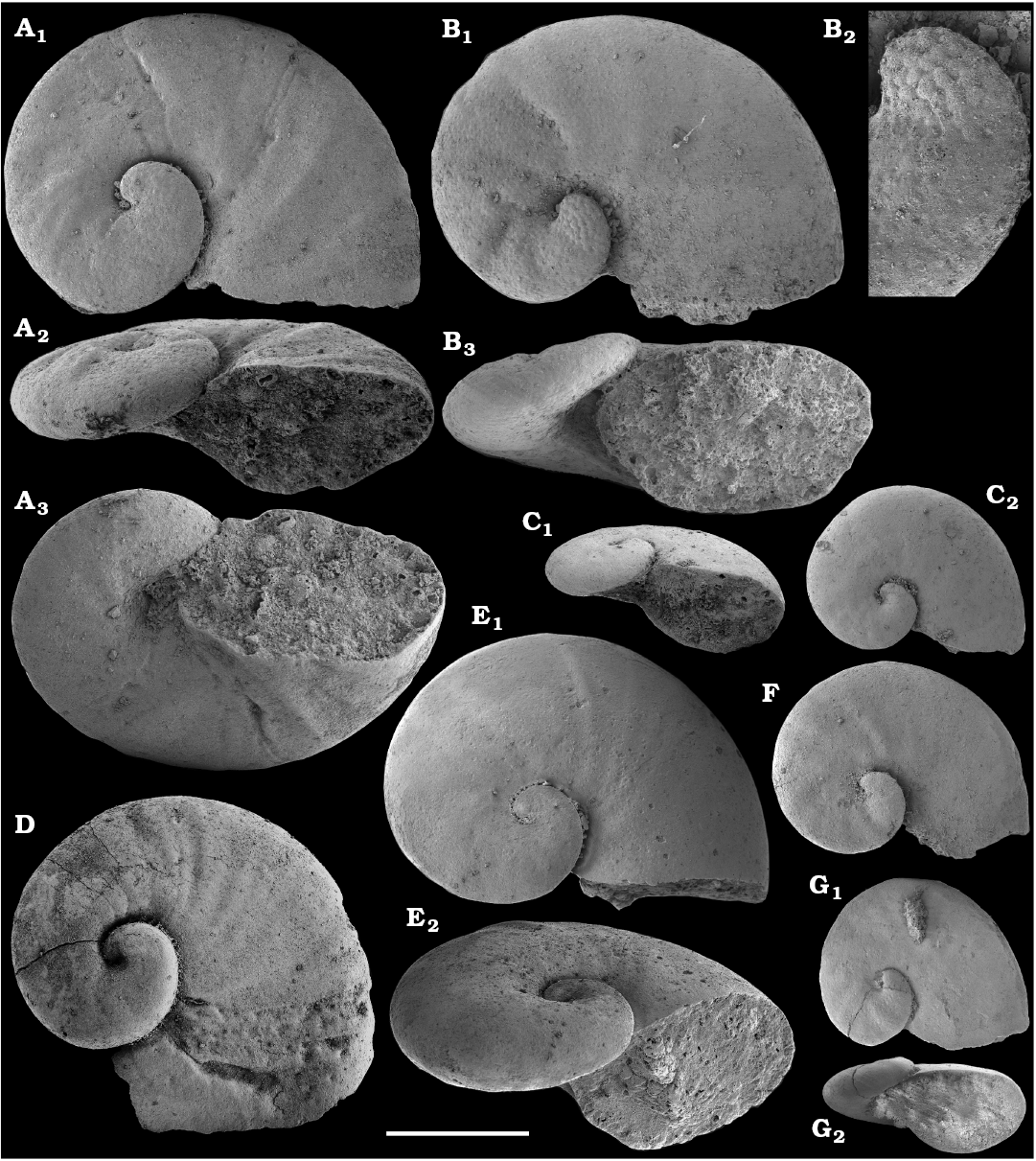
Fig. 18. Mollusc Aldanella crassa Missarzhevsky in Rozanov et al., 1969, internal moulds, from early Cambrian Medvezhya Formation, Kotuj River, western Anabar Uplift, Siberia, Russia; samples 1/50 (A, B, D–F), section 1; samples K2/13 (C) and 3/6.5 (G), section 3. A–G. SMNH Mo182246–182252, respectively. A1, B1, C2, D, E1, F, G1, adapical side; E2, oblique adapical view; A3, oblique umbilical side; A2, B3, C1, G2, apertural views; B2, close-up of apex with polygonal relief. Scale bar 200 µm (B2), 500 µm (A, B1, B3), and 1 mm (C–G).
Aldanella golubevi Parkhaev, 2007
Fig. 19.
Material.—Seven phosphatic internal moulds, including figured SMNH Mo182253–182258, from sample 3/10 (section 3, Fig. 3), Medvezhya Formation. Correlated with the uppermost Fortunian Stage.
Description.—Sinistrally coiled shell, up to 1.8 mm in diameter, consists of up to 1.5 rapidly expanding whorls. Adapical surface flattened. Apertural cross-section elongate, irregularly elliptical or ovoid, with width/height ratio ca. 2.0. Inner shell surface (as replicated on internal moulds) with smooth, broad, transverse folds.
Remarks.—Co-occurs with dextral but otherwise similar Aldanella crassa in one sample as well as at the type locality designated by Parkhaev (2007b). There are, however, several samples from the Medvezhya Formation below and above that level that are also mentioned by Parkhaev (2007b), where A. crassa occurs without A. golubevi (see SOM 1). These occurrence data suggest that A. golubevi is a separate species with a fixed sinistral coiling and doubtful affinity to dextrally coiled Aldanella, although Landing et al. (2013) regarded it as an uncommon sinistral variant of A. attleborensis. Jacquet et al. (2016) described Aldanella cf. golubevi from a horizon within the lower part of Cambrian Stage 2 of Australia.
Stratigraphic and geographic range.—Fortunian–Cambrian Stage 2 transitional beds, Siberian Platform and Australia.
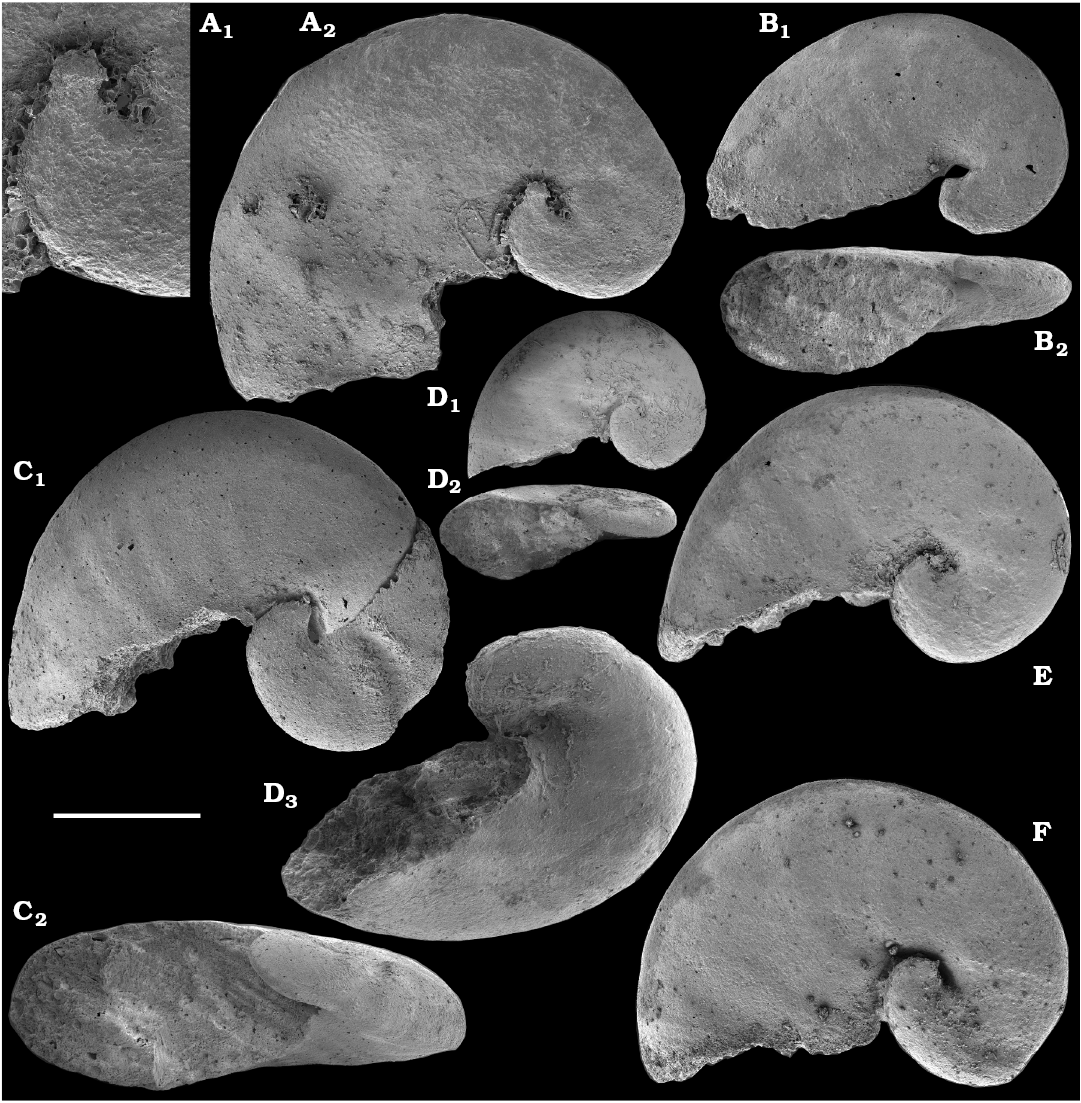
Fig. 19. Mollusc Aldanella golubevi Parkhaev, 2007, internal moulds, from early Cambrian Medvezhya Formation, Kotuj River, western Anabar Uplift, Siberia, Russia; sample 3/10, section 3. A–F. SMNH Mo182253–182258, respectively. A2, B1–D1, E, F, adapical side; D3, oblique umbilical side; B2, C2, D2, apertural views; A1, apical area enlarged. Scale bar 200 µm (A1), 500 µm (A2, B, C, D3, E, F), 1 mm (D1, D2).
Aldanella attleborensis (Shaler and Foerste, 1888)
Figs. 20–22.
Material.—Over 1000 phosphatic internal moulds, including figured SMNH Mo160428, 182260–182272, from samples 1/46, 1/45, 1/29.5 (section 1, Fig. 4), and K2/20, K2/21, K2/22, K2/23, K2/24, K2/26, 3/10, 3/12, 3/12.2, 3/12.5 (section 3, Fig. 3), and 5/10, 5/10B, 5a/1, 5a/1.2, 5a/5, 5a/6, 5a/9 (sections 96-5 and 96-5a, Fig. 2), Medvezhya and Emyaksin formations, Anabar Uplift, Siberian Platform, Russia. Correlated with uppermost Fortunian Stage and lower part of Cambrian Stage 2.
Description.—Moderately expanding shells, up to 2.0 mm in diameter, consist of up to 2.5 whorls. Spire slightly protrudes above the surface of the last whorl. Lateral surface of the whorl is broadly rounded and passes gradually into flattened or slightly convex adapical and slightly concave umbilical surfaces of the whorl. Aperture irregularly elliptical in outline, width/height ratio 1.6–1.8. Ornamentation of the shell exterior (judging from the surface of external mould; Fig. 22A) is represented by transverse growth lines. Protoconch with a diameter of 90–120 μm, not clearly separated in the available internal and external moulds (Figs. 20–22). Apex of internal moulds sometimes with polygonal to rounded impressions 20–40 μm in diameter (Figs. 20A3, F1, 21F; see also Kouchinsky 2000a; Ushatinskaya and Parkhaev 2005; Isakar and Peel 2007).
Remarks.—The specimens previously assigned to A. crassa by Kouchinsky (2000a: figs. 13A, B, L–R) are referred herein to A. attleborensis, following Parkhaev and Karlova (2011: 1187). Those authors also synonymized A. kunda (Öpik, 1926); A. rozanovi Missarzhevsky in Rozanov and Missarzhevsky, 1966 [in part]); A. yanjiahensis Chen, 1984; A. costata Missarzhevsky, 1989; and A. patelliforma Bokova, 1990, with A. attleborensis (see discussion above and more inclusive synonymy in Landing 1988; Landing et al. 1989, 2013).
Stratigraphic and geographic range.—Uppermost Fortunian and Cambrian Stage 2 of Siberia, Avalonia, North and South China, and Baltica.

Fig. 20. Mollusc Aldanella attleborensis (Shaler and Foerste, 1888), internal moulds, from early Cambrian Emyaksin Formation, Bol’shaya Kuonamka River, eastern Anabar Uplift, Siberia, Russia; sample 5a/1.2, section 96-5a. A–F. SMNH Mo160428 (A), SMNH Mo182260–182264 (B–F, respectively). A1, B, C. D, F1, adapical side; A2, F2, apertural; E, umbilical views; A3, apex enlarged. Scale bar 200 µm (A3), 500 µm (A1, A2, B, D–F), 1 mm (C).

Fig. 21. Mollusc Aldanella attleborensis (Shaler and Foerste, 1888), internal moulds, from early Cambrian Medvezhya Formation, Kotuj River, western Anabar Uplift, Siberia, Russia; samples 3/10 (A, C–F), K2/22 (B), and K2/24 (G), section 3. A–G. SMNH Mo182265–182271, respectively. A1, C, D1, E, F, G1, adapical side; A2, D2, apertural views; B, oblique adapical view; G2, apex enlarged. Scale bar 50 µm (G2), 500 µm (A–F), 1 mm (G1).

Fig. 22. Mollusc Aldanella attleborensis (Shaler and Foerste, 1888), SMNH Mo182272 (cast of the outer shell surface) from early Cambrian Emyaksin Formation, Bol’shaya Kuonamka River, eastern Anabar Uplift, Siberia, Russia; sample 5/10 from section 96-5. A. Adapical side with transverse sculpture. B. Protoconch enlarged. Scale bar 250 µm (A) and 125 µm (B).
Aldanella cf. sibirica Parkhaev and Karlova, 2011
Fig. 23.
Material.—About 80 internal moulds, including figured SMNH Mo182273–182283, from samples 5a/17.5, 5a/18.5, 5a/34.5, 5a/34.75 (section 96-5a, Fig. 2), Emyaksin Formation from the Anabar Uplift, Siberian Platform, Russia. Correlated with Cambrian Stage 2.
Description.—Dextrally coiled shell up to 1.5 mm in diameter, up to 2.5 whorls, slowly expands, with moderately projecting spire. Apertural cross-section broadly oval. Upper and lower surfaces of the last whorl convex, lateral periphery gently rounded. Protoconch not separated on the internal molds. Inner shell surface (as replicated on internal moulds) smooth, initial part with rounded polygonal impressions 20–40 μm in diameter (Fig. 23H1), apex gently pointed in some specimens.
Remarks.—Aldanella sibirica was first described by Parkhaev and Karlova (2011) from basal Erkeket Formation, section near Boroulakh on the Olenyok River, Olenyok Uplift of northern Siberian Platform, from the Dokidocyathus regularis Zone of the Tommotian Stage. The species shows variation in cross-sectional shape of the whorl, from narrower elliptical to broadly and irregularly oval. Such variation is consistent with morphological variability in internal moulds of Aldanella described by Landing (1988) from Avalonian North America. Internal moulds of A. sibirica are morphologically similar to those of A. attleborensis, as pointed out by Parkhaev and Karlova (2011: 1151), who mentioned rare “morphologically intermediate specimens”. A distinct spiral ornamentation of the shell exterior of A. sibirica (Parkhaev and Karlova 2011: pl. 11: 9) is absent in A. attleborensis (Fig. 22A herein). In the latter species, a fine spiral striation occurs on the surface of internal moulds (Fig. 20B, D). Spiral ornamentation is not developed in our material of Aldanella cf. sibirica represented by internal moulds. These are similar to specimens of A. sibirica in their broad cross-section of the last whorl and are relatively smaller than A. attleborensis.

Fig. 23. Mollusc Aldanella cf. sibirica Parkhaev and Karlova, 2011, internal moulds, from early Cambrian Emyaksin Formation, Bol’shaya Kuonamka River, eastern Anabar Uplift, Siberia, Russia; samples 5a/34.75 (A–H), 5a/17.5 (I, J), and 5a/34.5 (K), section 96-5a. A–K. SMNH Mo182273–182283, respectively. A1, D, E1–G1, H2, I, J1, K1, adapical side; A2, B2, C, E2–G2, J2, K2, apertural views; H1, apical area with polygons enlarged. Scale bar 200 µm (H1), 500 µm (A–G, H2).
Class and order uncertain
Family Khairkhaniidae Missarzhevsky, 1989
Genus Philoxenella Vostokova, 1962
Type species: Philoxenella spiralis Vostokova, 1962; upper Fortunian–lower part of Cambrian Stage 2; lower Cambrian; Rassokha River, Siberia, Russia.
Philoxenella spiralis Vostokova, 1962
Fig. 24D, E.
Material.—22 internal moulds, including figured SMNH Mo182287 and 182288, from samples 1/29.2 (section 1, Fig. 4) and 3/22.5, K2/52, and K2/60 (section 3, Fig. 3), Medvezhya and basal Kugda-Yuryakh formations, Anabar Uplift, Siberian Platform, Russia. Correlated with Cambrian Stage 2.
Description.—Shell up to 1.3 mm in diameter, dextrally coiled, up to 1.5 moderately expanding whorls with a broadly oval cross-section. The inner shell surface (as replicated by the available internal moulds) is smooth.
Remarks.—Internal moulds of Philoxenella are morphologically identical to those of Barskovia Golubev, 1976 (Fig. 24A–C, F, G), although the latter is sinistral (see below). Parkhaev (2007a, b) tentatively regarded them as dextral and sinistral variants of the same species and assigned them to the family Khairkhaniidae Missarzhevsky, 1989. This attribution is corroborated by probably similar microstructures of the conch, which is perforated by canals (Parkhaev 2006, 2007b). Philoxenella differs from Aldanella, which lacks canals in its conch walls, in being relatively loosely coiled and with lower rate of shell expansion.
Stratigraphic and geographic range.—Cambrian Stage 2 of the Siberian Platform and probably Avalonia (see Landing 1996: fig. 5b, c).
Genus Barskovia Golubev, 1976
Type species: Barskovia hemisymmetrica Golubev, 1976; upper Fortunian–lower part of Cambrian Stage 2; lower Cambrian; Plakhino Island, Enisej River, Siberia.
Barskovia hemisymmetrica Golubev, 1976
Fig. 24A–C, F, G.
Material.—17 internal moulds, including figured SMNH Mo182284–182286 and 182289, from samples 1/26.3, 1/29.2, 1/6 (section 1, Fig. 4) and K2/21, K2/50, 3/20.5 (section 3, Fig. 3), Kugda-Yuryakh (1/6) and Medvezhya formations, Anabar Uplift and SMNH Mo182290 from sample 1571/2, from Leningradskaya River, Tajmyr, Siberian Platform, Russia. Correlated with Cambrian Stage 2.
Description.—Shell up to 1.3 mm in diameter, sinistrally coiled, up to 1.5 moderately expanding whorls regularly with broadly oval, nearly circular cross-section. The inner shell surface (as replicated on the internal moulds) is smooth, conch wall transected by dense network of orthogonal canals.
Remarks.—Other such sinistral forms, as Nomgoliella Missarzhevsky, 1981, are characterized by a more tightly coiled shell and ovaliform aperture, whereas internal moulds of Nekolenia Vasil’eva, 1998, are more loosely coiled. This may be a preservational difference between shells and internal moulds. Parkhaev (2002: 34) synonymized Barskovia with Nekolenia, and both genera can probably be also synonymized with Nomgoliella. Synonymization can be justified by the facts that more and less tightly coiled forms such as Khairkhania rotata and K. evoluta appear in Khairkhania (see above) and that the shells appear to be more tightly coiled than their internal moulds. Although a circular aperture is considered diagnostic for Barskovia, broadly oval outlines are present in both B. hemisymmetrica and Philoxenella spiralis in our material (Fig. 23).
Stratigraphic and geographic range.—Upper Fortunian and Cambrian Stage 2 of the Siberian Platform and probably western Mongolia (depending on proposed synonymies).
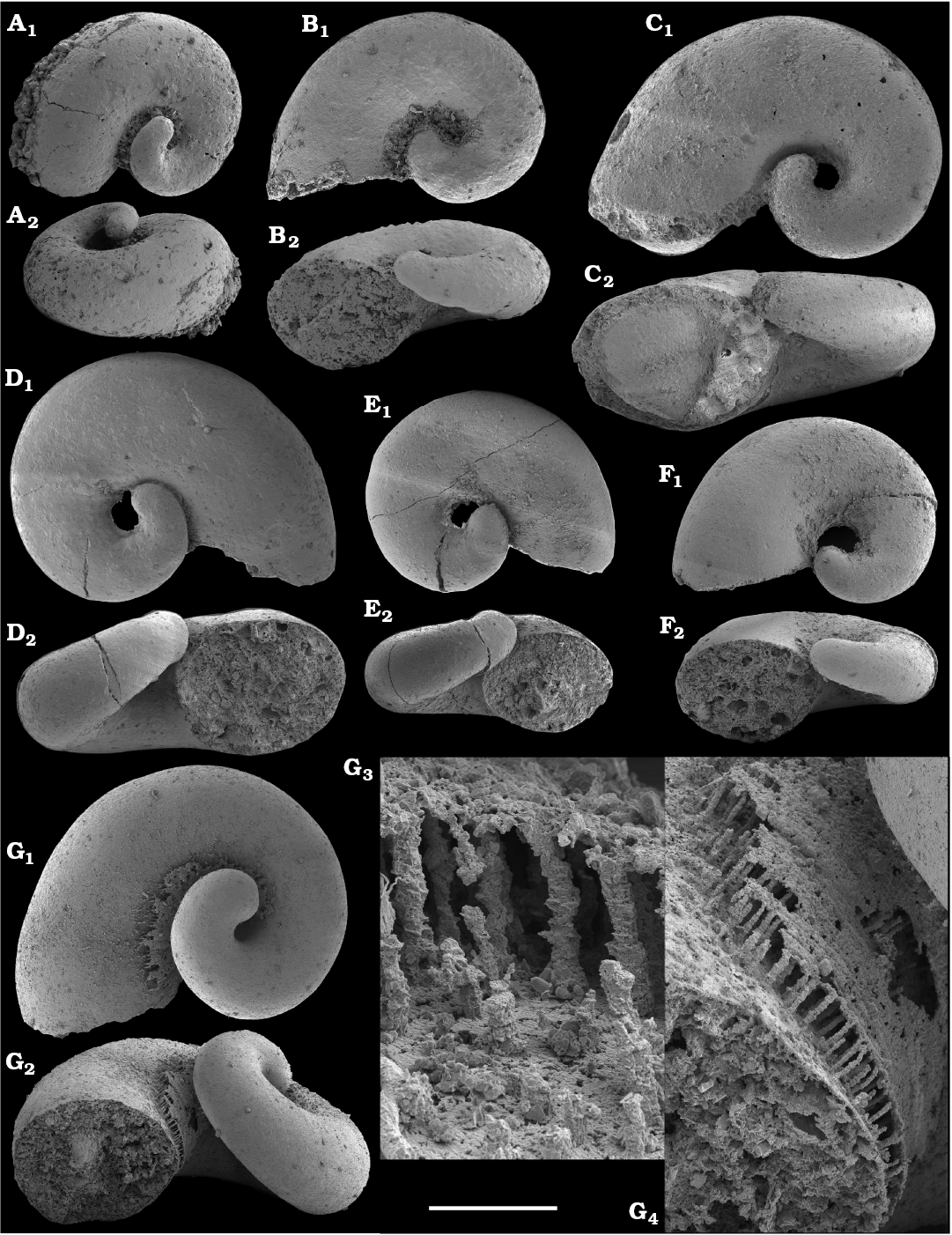
Fig. 24. Molluscs Barskovia hemisymmetrica Golubev, 1976 (A–C, F, G) and Philoxenella spiralis Vostokova, 1962 (D, E), internal moulds (G, internal mould with partly phosphatized shell), from early Cambrian Medvezhya Formation, western Anabar Uplift (A–F) and unnamed formation, NE Tajmyr (G), northern Siberia, Russia; sample K1a/76A (A), section 2; K2/21 (B) and K2/52 (D), section 3; 1/29.2 (C, E, F), section 1; 1571/2 (G), NE Tajmyr. A–G. SMNH Mo182284–182290, respectively. A1–F1, adapical side; A2, oblique adapical; G1, adapical; B2–G2, apertural views; G4, calcium phosphate cast of canals through the wall and outer shell surface; G3, cast of canals enlarged. Scale bar 25 µm (G3), 100 µm (G4), and 500 µm (A–F, G1, G2).
Genus Khairkhania Missarzhevsky, 1981
Type species: Khairkhania rotata Missarzhevsky, 1981; upper Fortunian–lower part of Cambrian Stage 2; lower Cambrian; western Mongolia.
Khairkhania cf. rotata Missarzhevsky, 1981
Fig. 25D.
Material.—Single phosphatized internal mould SMNH Mo182294 and seven fragments of internal moulds of presumably the same species from sample 3/0 (section 3, Fig. 3), basal Medvezhya Formation (see SOM 1), western Anabar Uplift, Siberian Platform, Russia. Correlated with upper Fortunian Stage.
Description.—Planispiral shell, relatively tightly coiled through ca. 1 revolution, with blunt rounded apex and indistinctly separated protoconch. Cross-section broadly oval.
Remarks.—Khairkhania rotata from western Mongolia and forms of this species described by Parkhaev (2006, 2007b, 2008) from the northwestern flanks of the Anabar Uplift are generally similar but smaller (ca. 75% in size) than the material illustrated herein as Khairkhania cf. rotata (Fig. 25D). Khairkhania sp. from the same sample are either fragmentary or open coiled with up to 1/2 revolution (Fig. 25E, F). They may represent a different species, morphological variants, or younger growth stages of the same species. Khairkhania cf. evoluta (see below) is represented by high, cyrtoconic, relatively more laterally compressed internal moulds with smaller protoconchs.
Khairkhania cf. evoluta Zhegallo in Voronin et al., 1982
Fig. 25A–C.
Material.—17 internal moulds, including figured SMNH Mo182291–182293, from samples 1/29.5 (section 1, Fig. 4), K1a/82 (section 2, Fig. 1), K2/38A, K2/43, and K2/46 (section 3, Fig. 3), from the Medvezhya Formation (SOM 1), western Anabar Uplift, Siberian Platform, Russia. Correlated with the upper Fortunian Stage.
Description.—Planispiral, slowly expanding cyrtoconic shells coiled up to 3/4 of a revolution, with rounded protoconch indistinctly separated from the rest of the shell, represented by smooth internal moulds. Cross-section broadly oval, nearly circular.
Remarks.—Khairkhania evoluta was first described from the upper Fortunian–Cambrian Stage 2 equivalent beds of western Mongolia (Voronin et al. 1982). One morphologically similar fossil, albeit with a hyolith-type microstructure of the wall, was described from the Zhongyicun Member of the Meishucunian Stage of South China (Fortunian Stage; Feng et al. 2001b). Multiple columns representing casts of canals orthogonally oriented in the shell wall were described from the probable kindred species Khairkhania rotata and from such generally similar but dextrally and sinistrally coiled forms as Philoxenella spiralis Vostokova, 1962, and Barskovia hemisymmetrica Golubev, 1976, respectively (Parkhaev 2006, 2007b; see below). The columns are also similar to those present in the tubes and opercula of hyoliths (Kouchinsky 2000b; Kouchinsky et al. 2015a). The polygonal impressions sometimes preserved on the apex of Aldanella spp. internal moulds (Figs. 18B3, 20A3, 23H1) have never been reported from Barskovia, Nomgoliella, Philoxenella, or Khairkhania. This lack of apical polygonal impressions further corroborates the taxonomic distinctiveness of the latter four genera among other coeval mollusc-like fossils.
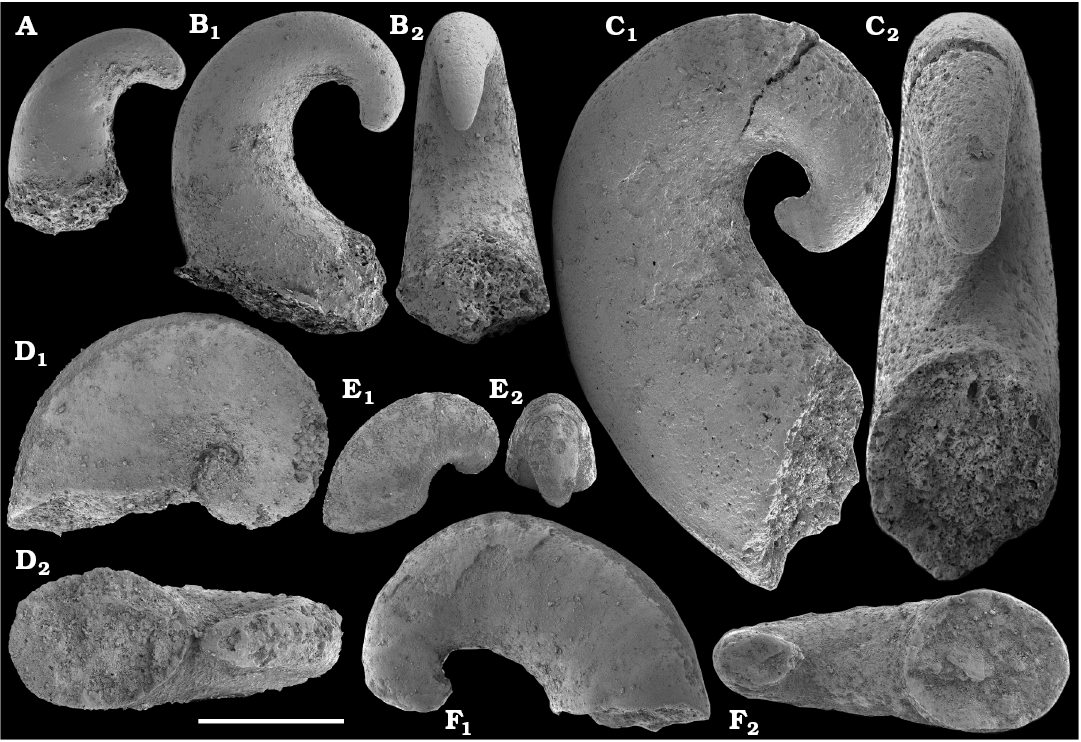
Fig. 25. Molluscs? Khairkhania Missarzhevsky, 1981, from early Cambrian Medvezhya Formation, Kotuj River, western Anabar Uplift, Siberia, Russia; samples K1a/82 (A, B), section 2; 1/29.5 (C), section 1; and 3/0 (D–F), section 3. A–C. Khairkhania cf. evoluta Zhegallo in Voronin et al., 1982, internal moulds with partly phosphatized shell, SMNH Mo182291–182293, respectively. D. Khairkhania cf. rotata Missarzhevsky, 1981, internal mould, SMNH Mo182294. E, F. Khairkhania sp., internal moulds with partly phosphatized shell, SMNH Mo182295, 182296, respectively. A, B1–F1, lateral; B2, C2, E2, subapical; D2, F2, apertural views. Scale bar 500 µm (A–C), 1 mm (D–F).
Class, order, and family uncertain
Genus Ceratoconus Chen and Zhang, 1980
Type species: Ceratoconus striatus Chen and Zhang, 1980; middle Meishucunian Stage (upper Fortunian), lower Cambrian; Yunnan Province, South China.
Remarks.—Ceratoconus is different from Salanyella Missarzhevsky, 1981 (see below), in having a more elongated shell. Yu (1987) synonymized Ceratoconus with Pollicina Holzapfel, 1895. Thus, Pollicina striata (Chen and Zhang, 1980) occurs in fossil lists from the upper Fortunian–Cambrian Stage 2 equivalents of western Mongolia (basal Unit 20 of the Bayan Gol Formation, Tiksitheca licis Zone; Voronin et al. 1982; Esakova and Zhegallo 1996; Brasier et al. 1996) and lower Medvezhya Formation of the western Anabar Uplift (Kaufman et al. 1996).
Ceratoconus striatus Chen and Zhang, 1980
Figs. 26, 27.
Material.—About 150 phosphatic internal moulds, including figured SMNH Mo182297–182302, from samples 1/39.3, 1/46 (section 1, Fig. 4), 3/12, 3/12.2, and K2/25 (section 3, Fig. 3), Medvezhya Formation, western flanks of the Anabar Uplift and 20 phosphatic internal moulds, including figured SMNH Mo182303–182306, from samples B-438 and 769, lower Emyaksin Formation, Kengede River, southern flank of the Anabar Uplift, Siberian Platform, Russia. Correlated with the upper Fortunian Stage.
Description.—Long, narrow, conical, ceratoconic shell curved up to 1/4 whorl in one plane with broad oval aperture. Apex conical with a blunt rounded tip. Protoconch not clearly separated. Inner shell surface (as replicated on internal moulds) is smooth or with low‒moderately high transverse folds. Aperture with subapical sinus (Fig. 27A2).
Remarks.—These internal moulds are identical to those of Ceratoconus rusticus (Val’kov, 1987) and C. arcuatus Missarzhevsky, 1989. Forms with larger and bulbous protoconch (see Ceratoconus aff. striatus below, Fig. 28) co-occur with those having more pointed apices (Figs. 26, 27). Both such forms were attributed to Ceratoconus striatus Chen and Zhang, 1980, by Parkhaev in Rozanov et al. (2010: pl. 29: 7, 8). As recognized herein, C. striatus has more elongated internal moulds with more subdued transverse folds than specimens of C. striatus from South China. Ceratoconus aff. striatus more likely represents a separate species and can be distinguished by a wider and more rounded initial part (protoconch?) and a greater angle of shell expansion.
Stratigraphic and geographic range.—Upper Fortunian and Cambrian Stage 2 of Siberian Platform, South China, and western Mongolia.
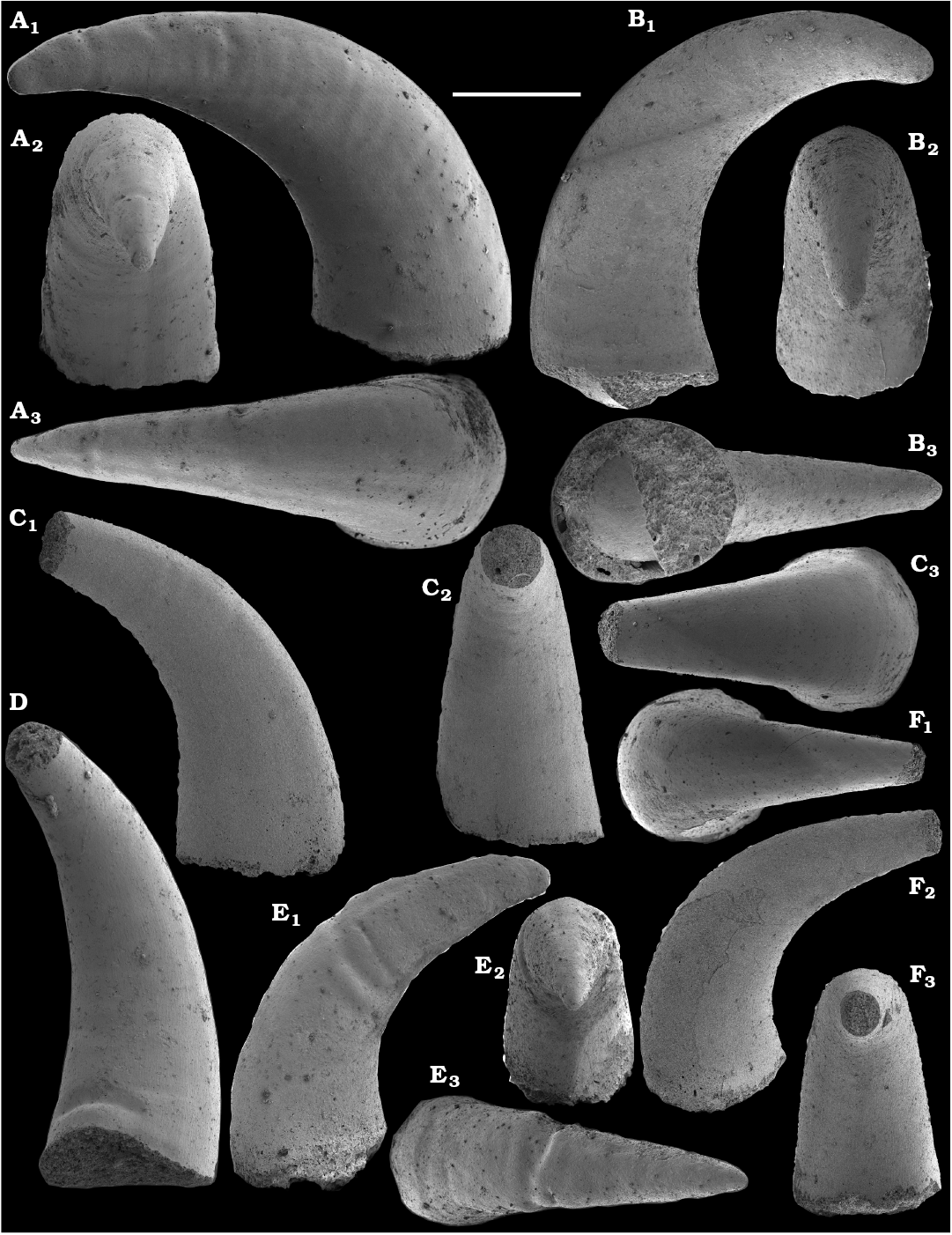
Fig. 26. Mollusc Ceratoconus striatus Chen and Zhang, 1980, internal moulds, from early Cambrian Medvezhya Formation, Kotuj River, western Anabar Uplift, Siberia, Russia; samples 1/46 (A, B, E), section 1; 3/12.2 (C, F) and K2/25 (D), section 3. A–F. SMNH Mo182297–182302, respectively. A1–C1, E1, F2, lateral; A2–C2, D, E2, F3, subapical; A3, C3, E3, F1, supra-apical; B3, apertural views. Scale bar 500 µm.

Fig. 27. Mollusc Ceratoconus striatus Chen and Zhang, 1980, internal moulds, from early Cambrian Emyaksin Formation, Kengede River section, southern Anabar Uplift, Siberia, Russia; samples B-438 (A) and 769 (B–D). A–D. SMNH Mo182303–182306, respectively. A2, subapical; A1, B, D, lateral; C, supra-apical views. Scale bar 500 µm.
Ceratoconus aff. striatus Chen and Zhang, 1980
Fig. 28.
Material.—About 50 phosphatic internal moulds, including figured SMNH Mo Mo160413, 182307–182313, from samples 3/12, 3/12.2, K2/25, and K2/26 (section 3, Fig. 3), Medvezhya Formation, western Anabar Uplift, Siberian Platform, Russia. Correlated with the uppermost Fortunian Stage.
Description.—Long narrow conical shell, curved up to 90º in one plane, with bulbous apex and rounded aperture. Protoconch not clearly separated from the rest of the shell. Inner shell surface (as replicated in internal moulds) with low transverse folds bent towards the apex on the convex margin of the conch (Fig. 28B1, C1).
Remarks.—With its more rapidly expanding shell and a larger, more bulbous apex, Ceratoconus aff. striatus has a more robust habit than C. striatus.
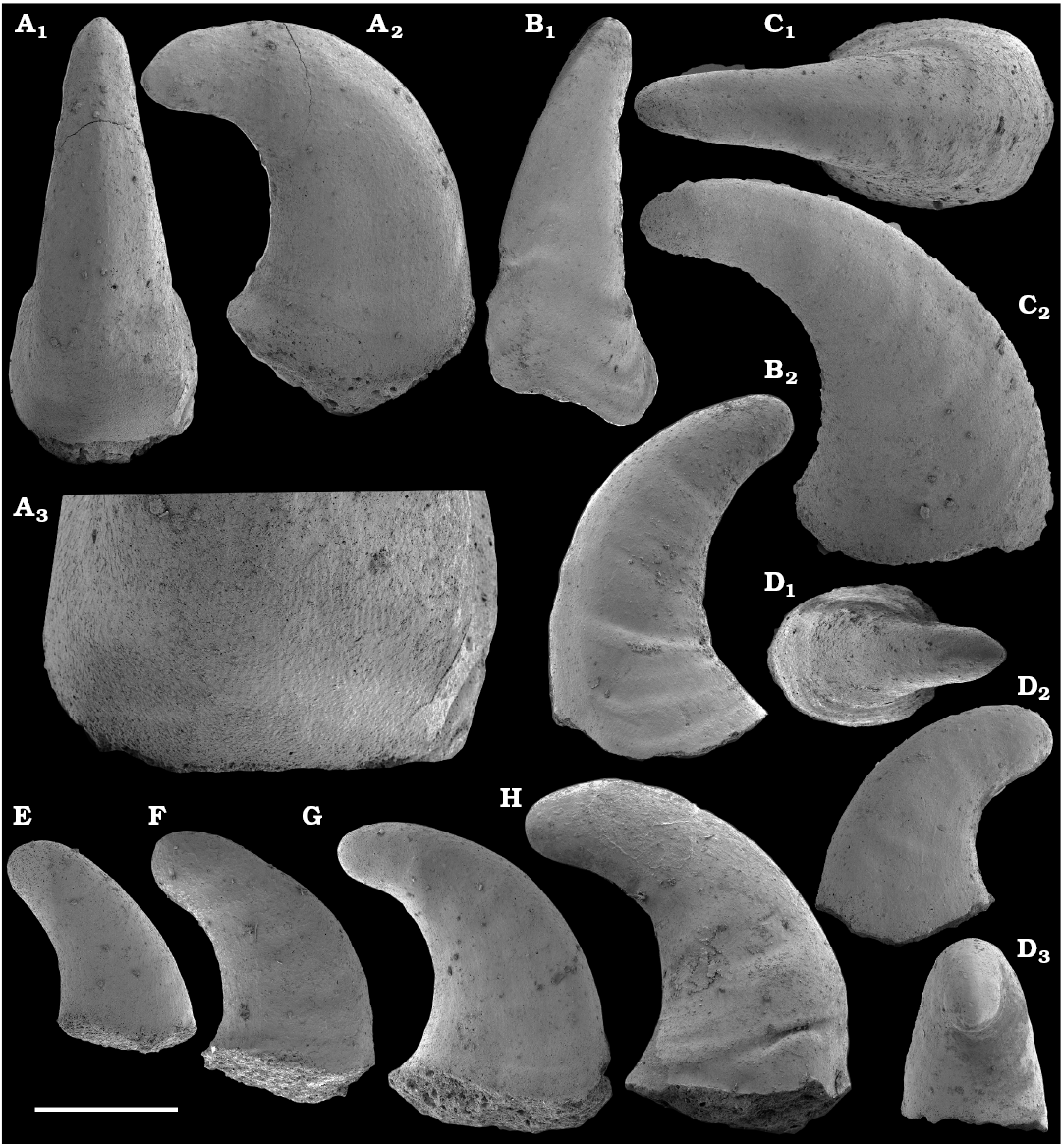
Fig. 28. Mollusc Ceratoconus aff. striatus Chen and Zhang, 1980, internal moulds, from early Cambrian Medvezhya Formation, Kotuj River, western Anabar Uplift, Siberia, Russia; samples 3/12.2 (A–C, E), 3/12 (D), and K2/25 (F–H) from section 3. A–H. SMNH Mo182307–182313 (A–G respectively), SMNH Mo160413 (H) (compare H with Kouchinsky 2000a: fig. 7A–G). A1–D1, supra-apical; A2–D2, E–H, lateral; D3, subapical views; A3, apertural area with polygons enlarged. Scale bar 200 µm (A3), 500 µm (A1, A2, B–H).
Genus Salanyella Missarzhevsky, 1981
Type species: Salanyella costulata Missarzhevsky, 1981; Watsonella crosbyi Zone, lower Cambrian (lower part of Cambrian Stage 2); Salany-Gol section, western Mongolia.
Salanyella cf. costulata Missarzhevsky, 1981
Figs. 29A–C, E–H, 30.
Material.—About 50 phosphatic internal moulds, including figured SMNH Mo160414, 182314–182316, 182318–182321, and a single partially preserved phosphatized conch with internal mould, SMNH Mo182322, from samples 1/29.2 (section 1, Fig. 4), 3/12, 3/12.2, 3/22.5, K2/25, and K2/26 (section 3, Fig. 3), from the Medvezhya Formation, western flanks of the Anabar Uplift, Siberian Platform, Russia. Correlated with the uppermost Fortunian Stage.
Description.—Moderately expanding, short conical shell curved up to 45º, with broadly oval, straight aperture. Apical region blunt, rounded, not clearly separated from the rest of the shell. Outer shell surface smooth, with thin closely spaced growth lines (Fig. 30B1, B2).
Remarks.—Moulds are covered with tiny, densely arranged, rounded bumps near the aperture and larger, round to elongate bumps on the adapical surface (Fig. 29E2). Similar forms were described under the monospecfic genus Lenoconus Vasil’eva, 1990, as Lenoconus sulcatus Vasil’eva, 1990, from the Dockidocyathus regularis Zone of the Tommotian Stage (upper part of Cambrian Stage 2) on the lower reaches of the Lena River at the Ulakhan-Ald’yarkhaj section. The type specimen of L. sulcatus designated by Vasil’eva (1990: pl. 2: 3, 4) has a hooked apical part. It is twice as large as Lenoconus sp. from the Tommotian Stage on the Aldan River (Vasil’eva 1990: pl. 2: 7), which likely represents a juvenive form of the same species. The apical part of the latter is not so curved.
Lenoconus is synonymized herein with Salanyella. The specimens described herein are generally similar in shape and treated as growth stages probably of a single species reported herein as Salanyella cf. costulata. A likely similar form is reported without illustration as S. costulata from the Anabar Uplift and elsewhere on the Siberian Platform by Khomentovsky and Karlova (2005). Phosphatized shells of S. costulata Missarzhevsky, 1981, from Cambrian Stage 2 of western Mongolia have, however, numerous thin transverse ribs not present on the internal moulds from the Anabar Uplift (Figs. 29, 30A). The outer surface of the S. cf. costulata shell is preserved as a phosphatic crust (Fig. 30B) that has regularly spaced growth lines, although they are less distinct than the ribs in the topotype material from western Mongolia. In addition, steinkerns from western Mongolia bear a regular, transverse, pustulouse ornament, which is probably a result of diagenetic phosphatisation of shell pores.
An apical tuft of acicular elements present between the phosphatized shell and internal mould (Figs. 29F1, 30B4) probably represents phosphatic casts of early diagenetic crystals of fibrous aragonite that grew in spaces between the inner surface of the shell and its sediment infill. Similar casts are observed in other parts of the internal moulds where they may have a different orientation, either parallel to or inclined to the surface of the mould. Such features are observed on internal moulds of maikhanellids (Fig. 32B1, 34A1, A4), septa of hyoliths replicated by phosphate (Fig. 53I), as well as on the internal moulds of chancelloriid sclerites from Siberia and Laurentia (Kouchinsky et al. 2015a: fig. 34A).
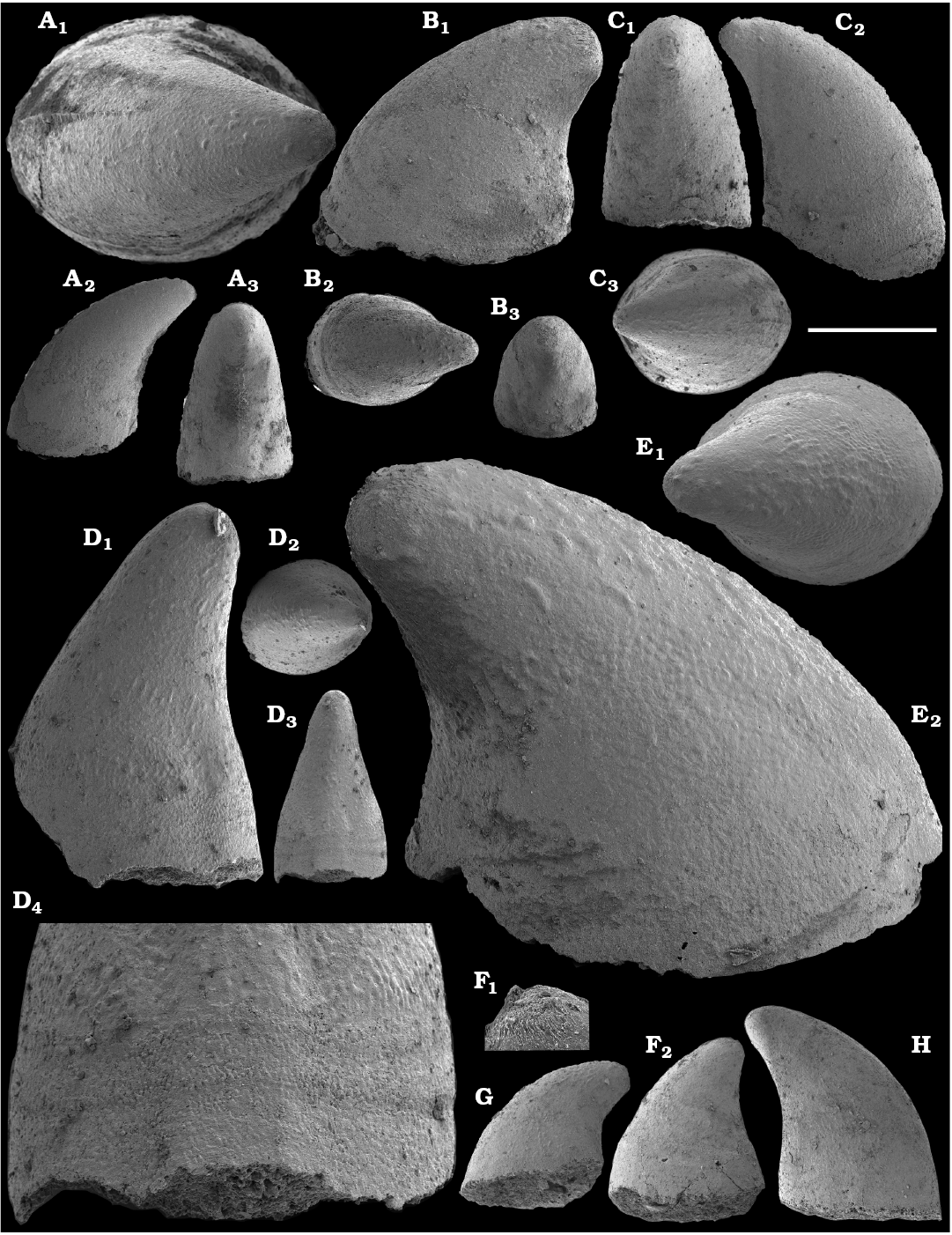
Fig. 29. Molluscs Salanyella cf. costulata Missarzhevsky, 1981 (A–C, E–H) and Salanyella sp. (D), internal moulds, from early Cambrian Medvezhya Formation, Kotuj River, western Anabar Uplift, Siberia, Russia; samples 3/12.2 (A–D), 3/12 (E), and K2/25 (F–H) from section 3. A–H. SMNH Mo182314–182318 (A–E, respectively), 160414 (F; see also Kouchinsky 2000a: fig. 7H–J), 182319 (G), 182320 (H). A1, B2, C3, D2, E1, supra-apical; A2, B1, C2, D1, E2, F2, G, H, lateral; A3, B3, C1, D3, subapical views; F1, apex with spiny structures enlarged; D4, apertural area in D3 enlarged. Scale bar 125 µm (D4, E2, F1), 250 µm (A1, B1, D1, E1), 500 µm (A2, A3, B2, B3, C, D2, D3, F2, G, H).

Fig. 30. Mollusc Salanyella cf. costulata Missarzhevsky, 1981, from early Cambrian Medvezhya Formation, Kotuj River, western Anabar Uplift, Siberia, Russia; samples 3/22.5 (A) and K2/26 (B) from section 3. A. SMNH Mo182321, internal mould. B. SMNH Mo182322, internal mould with phosphatized shell. A1, apical; A2, subapical; A3, B2, lateral; B1, oblique supra-apical; B3, oblique subapical; B5, apertural views; B4, enlargement of apex with spiny structures on the internal mould. Scale bar 100 µm (B4) and 250 µm (A, B1–B3, B5).
Class and order uncertain
Family Maikhanellidae Missarzhevsky, 1989
Remarks.—The family Maikhanellidae was initially assigned by Missarzhevsky (1989: 24, 179) to the monoplacophorans. Ponder et al. (2007) supported that assignment but also discussed the similarity of the cap-like shell and its scaly ornamentation to those of some patelliform gastropods (see also Parkhaev and Demidenko 2010). An alternative phylogenetic interpretation by Bengtson (1992) is based on evidence for maikhanellids merging siphogonuchitid sclerites into their shells. Based on younger Wiwaxia-like organisms with non-mineralized scleritomes and on complete scleritomes of halkieriids, which have two terminal shells and rows of intermediate sclerites (Conway Morris and Peel 1995), maikhanellids were assigned to a phylum-level, lophotrochozoan stem-group called Halwaxiida Conway Morris and Caron, 2007. Butterfield (2006) suggested, however, that Wiwaxia and halkieriids have markedly different sclerite construction and composition and cannot be assigned to one phyletic grouping. This conclusion was also supported by comparison of ontogenetic series of wiwaxiid scleritomes from Cambrian Stage 3 deposits of the Chengjiang Lagerstätte with the Halkieria scleritome (Zhang et al. 2015).
An alternative concept of a new molluscan class, Diplacophora Vinther and Nielsen, 2005, a sister group to the Polyplacophora De Blainville, 1816, was advanced by Vinther and Nielsen (2005). Parkhaev and Demidenko (2010: 955) further discussed the affinity of sachitids (halkieriids and siphogonuchitids) to chitons and placed them in a subclass Diplacophora of the class Polyplacophora. The presence of two shell-plates (anterior and posterior) may however not characterize all members of the halkieriid stock, since only a single shell plate is present in the probably closely related Orthrozanclus Conway Morris and Caron, 2007. In this report, we consider an assignement of the maikhanellids and sachitids to the stem-lineage of the Polyplacophora, although persuasive evidence is still lacking.
New evidence on the widespread occurrence of multi-unit, shell-bearing scleritomes in the Terreneuvian Series emerged from studies of the Ocruranus–Eohalobia group of fossils (Qian and Bengtson 1989; Siegmund 1997), which were interpreted to represent components of the same organism from the chiton stem lineage (Vendrasco et al. 2009). Similarly, halkieriids allow comparisons with the shells of Ocruranus Liu, 1979, and Eohalobia Jiang in Luo et al., 1982 (synonymized under Ocruranus by Vendrasco et al. 2009). However, there is no evidence of merged sclerites in the latter genera, and their shells have prismatic and lamello-fibrillar microstructures similar to those known from early Cambrian helcionellid molluscs (Vendrasco et al. 2009).
Genus Purella Missarzhevsky, 1974
Type species: Purella cristata Missarzhevsky, 1974; correlated with the upper Fortunian–lower part of Cambrian Stage 2; lower Cambrian; left bank on the middle reaches of the Olenyok River, 6 km downstream of the mouth of Chuskuna Creek, Siberia, Russia.
Purella antiqua (Abaimova, 1976)
Figs. 31, 32, 33A–F, H.
Material.—Over 500 phospatic internal moulds with fragments of phosphatized shells, including figured SMNH Mo160403, 160406, 182323–183337, from samples 4/27.5, 5/0, 5/0.25 (sections 96-4 and 96-5, Fig. 2), 2/A, 1282, K1a/43, and K1a/47 (section 2, Fig. 1), from the upper Manykay, Nemakit-Daldyn, and basal Emyaksin formations (SOM 1), Anabar Uplift, Siberian Platform, Russia. Correlated with the Fortunian Stage.
Description.—Shell low, conical, with blunt apex that does not project beyond the apertural margin. Aperture ovoid with length/width ratio 1.3–1.5. Dorsal side with a broad longitudinal fold (Fig. 34C2). The shell is composed of comarginally elongated scales (Fig. 31). The apical area is externally smooth, without scales, and is composed of densely juxtaposed microtubules with radial orientation (Kouchinsky 2000a; Figs. 31A, 32A).
Remarks.—Abaimova (1976) described the morphological variation of Purella antiqua conchs from a single sample collected from the same horizon and locality as samples 1282, 2/A, K1a/43, and K1a/47 (SOM 1). The assemblage of remains includes internal moulds with fragmentary phosphatized shells up to 1.5 mm long, sometimes bearing a median buttress rimmed by two shallow furrows. These molds are smooth at the apex, but concentrically sculptured distally, with a different shape of the aperture. The apices range from those with an almost central position of the apex to those with the apex overhanging the apertural margin (see variable position of the apex in Abaimova 1976: figs. 5, 8, 11, 15). Such a variation was suggested to distinguish Purella antiqua from “other helcionellids” (Abaimova 1976: 175). One of the specimens illustrated by Abaimova (1976: pl. 20: 15) is similar to Stenothecoides accuratus Bokova, 1985, described from the same locality and bed, but is identified in this report as Yunnanopleura accurata (Bokova, 1985) (discussed below).
Stratigraphic and geographic range.—Fortunian Stage of Siberian Platform, South China, and western Mongolia.

Fig. 31. Mollusc Purella antiqua (Abaimova, 1976), calcium phosphate coatings of the shells, from early Cambrian Nemakit-Daldyn (D, E) and basal Emyaksin (A–C) formations, Anabar Uplift, Siberia, Russia; samples 4/27.5 (A), 5/0 (B, C), and K1a/47 (D, E) from sections 96-4, 96-5, and section 2, respectively. A–E. SMNH Mo160403 (A; see also Kouchinsky 2000a: fig. 3), 182323 (B), SMNH Mo182324 (C), SMNH Mo160406 (D; see also Kouchinsky 2000a: fig. 4G), SMNH Mo182325 (E). A1, A4, views of inner shell surface; A2, close-up of inner shell surface in A1; A3, microtubules of the initial shell enlarged; B1, lateral; B2, apical; B3, subapical views; C, oblique subapical view showing apically smooth external shell surface and distal shell composed of circumferentially elongated scales; external (D) and inner (E) surfaces of shell fragment. Scale bar 50 µm (A3), 100 µm (A2), 250 µm (A1, A4, B–E).

Fig. 32. Mollusc Purella antiqua (Abaimova, 1976), from early Cambrian Nemakit-Daldyn Formation, western Anabar Uplift, Siberia, Russia; sample 1282, section 2. A. SMNH Mo182326; A1, partial external mould of shell; A2, enlarged apical area shows radial microtubules and circumferential scales. B. SMNH Mo182327, internal mould; B1, oblique lateral; B2, upper views. C. SMNH Mo182328, lateral view of internal mould separated by a narrow slit (arrowed) from external phosphatic coat of a dissolved shell. D. SMNH Mo182329, upper view of internal mould that underlies external phosphatic coating of a dissolved shell. E. SMNH Mo182330, internal mould; E1, subapical; E2, upper views. Scale bar 125 µm (A2), 250 µm (B–E), 500 µm (A1).

Fig. 33. Molluscs Purella antiqua (Abaimova, 1976) (A–F, H), Protoconus? sp. (G), and Yunnanopleura sp. (I), from early Cambrian Emyaksin (A–F) and Medvezhya (H–I) formations, Anabar Uplift, Siberia, Russia; samples 4/27.5 (A–D), section 96-4; 5/0 (E, F), section 96-5; 3/0 (H) and 3/0.1 (G, I), section 3. A–F, H. Internal moulds with partly phosphatized shell (B and D), SMNH Mo182331–182336 (A–F, respectively), SMNH Mo182337 (H). G, I. Internal moulds, SMNH Mo182338, 182339, respectively. A1, B, C1, E, F, G2, H3, I2, upper; A2, C2, G1, H1, I3, lateral; C3, D, G3, H2, I1, subapical views. Scale bar 250 µm (C1), 500 µm (A, B, C2, C3, D–I).

Fig. 34. Molluscs Protoconus? sp. (A, B) and Purella antiqua? (Abaimova, 1976) (C), internal moulds, from early Cambrian Nemakit-Daldyn Formation, western Anabar Uplift, Siberia, Russia; sample 2/A, section 2. A–C. SMNH Mo182340–182342, respectively. A1, B1, C2, upper; A2, C3, supra-apical; A3, B3, subapical; A4, B2, C1, lateral views. Scale bar 250 µm (A, C), 500 µm (B).
Purella cristata Missarzhevsky, 1974
Figs. 35.
Material.—Over 80 internal moulds with fragments of phosphatized shells, including figured SMNH Mo182343‒182348, from samples 1/29.2, 1/29.5, 1/50 (section 1, Fig. 4), K2/12, K2/13, K2/25, K7, 3/0, 3/0.1, 3/6.5, 3/12, 3/12.2 (section 3, Fig. 3), Medvezhya and Kugda-Yuryakh formations, western Anabar Uplift, Siberian Platform, Russia. Correlated with upper Fortunian and lower part of Cambrian Stage 2.
Description.—Shell low to high conical, with blunt apex slightly inclined but not projecting beyond the apertural margin. Aperture wide oval to broadly rounded rhomboid, with narrower subapical part, length/width ratio 1.5–1.8. Supra-apical side of the shell may bear a broad longitudinal buttress between apex and apertural margin. Shell composed of comarginally elongated scales.
Remarks.—Purella cristata Missarzhevsky, 1974, was first described from limestones (sample M71-2/66 collected by Vladimir V. Missarzhevsky) in the upper Kessyusa Formation of the Olenyok Uplift. Purella cristata is more laterally compressed than P. antiqua and has a more elongated shape (Fig. 35). Parkhaev (in Varlamov et al. 2008: pl. 12: 3a, 3b; in Rozanov et al. 2010: pl. 26: 1) described internal moulds of P. cristata with preserved scales. Parkhaev and Demidenko (2010) suggested its synonymy with P. dzhandica Yermak in Pel’man et al., 1990, and P. rudavskae Vasil’eva, 1998, from Siberia. The taxonomy of Purella was discussed by Parkhaev and Demidenko (2010) and Devaere et al. (2013). The co-occurrence of more elongate forms comparable with P. arcana (Val’kov 1987: pl. 16: 4; Fig. 35A, herein) with those identified in this report as P. cristata from sample 1/50 (Kotuj River, section 1; Figs. 4, 35) suggests that P. arcana Val’kov, 1987 is probably synonymous with P. cristata. In P. gracilis Zhegallo in Voronin et al., 1982, hollow spines are present in the subapical area (Devaere et al. 2013), as in P. squamulosa Qian and Bengtson, 1989 (i.e., Yang et al. 2014). These spines are unknown in other species and in our material. Parkhaev and Demidenko (2010) followed Missarzhevsky (1989) and synonymized Rozanoviella Missarzhevsky, 1981, from the Terreneuvian beds of western Mongolia with Purella. Parkhaev and Demidenko (2010) also concluded a probable synonymy of R. atypica Missarzhevsky, 1981 (Missarzhevsky 1981: text-fig. 2, pl. 3: 9; Esakova and Zhegallo 1996: pl. 20: 4), with Purella antiqua based on general morphology and shell structure (Abaimova 1976: pl. 20: 1, 4; Kouchinsky 2000a: fig. 3).
Stratigraphic and geographic range.—Fortunian Stage and lower part of Cambrian Stage 2; South China and Siberian Platform, probably western Mongolia.

Fig. 35. Mollusc Purella cristata Missarzhevsky, 1974, internal moulds with partly phosphatized shells (B, C), from early Cambrian Medvezhya Formation, western Anabar Uplift, Siberia, Russia; sample 1/50, section 1. A–F. SMNH Mo182343‒182348, respectively. A1, B3, C1, D2, E3, F1, upper; A2, B2, C3, D1, E2, F2, lateral; B1, C2, E1, F3, subapical views; F4, enlargement of F2 showing tuberculate texture. Scale bar 125 µm (F4), 250 µm (C3), 500 µm (A, B, C1, C2, D, E, F1‒F3).
Yunnanopleura Yu, 1987
Type species: Yunnanopleura biformis Yu, 1987; middle Meishucunian Stage, lower Cambrian (upper Fortunian); Yunnan, South China.
Yunnanopleura accurata (Bokova, 1985)
Fig. 36.
Material.—Several hundred internal moulds with fragments of phosphatized conchs, including figured SMNH Mo160405, 182349–182355, from samples 2A, 1282, K1a/43, K1a/47 (section 2, Fig. 1), 3/0 (section 3, Fig. 3), 4/27.5, 5/0, 5/0.25 (sections 96-4, 96-5, Fig. 2), Nemakit-Daldyn, Medvezhya, lower Emyaksin formations, Anabar Uplift, Siberian Platform, Russia. Correlated herein with the upper Fortunian Stage.
Description.—Low cap-shaped shells with ovoid aperture (length/width ratio ca. 1.5). Apex blunt, smooth, overhangs the apertural margin and projects slightly beyond it. Scales oriented parallel to the apertural margin appear on the shell surface distal to the apex (Fig. 36B3) and at the subapical apertural margin (Fig. 36H3).
Remarks.—These fossils were first described by Bokova (1985) as Stenothecoides accuratus Bokova, 1985, and S. pandus Bokova, 1985. The material derives from sample 1282, collected by Anatolij K. Val’kov from a section at the mouth of the Kotujkan River in the western Anabar Uplift that is equivalent to section 2 of this report. Bokova’s (1985) fossils are very similar to Yunnanopleura biformis Yu, 1987, and Y. longidens Feng, Sun, and Qian, 2001a, from South China, which similarly co-occur with scaly shells of Purella (Qian and Bengtson 1989; Feng et al. 2001a; Parkhaev and Demidenko 2010). Yunnanopleura longidens was considered synonymous with Y. biformis by Parkhaev and Demidenko (2010). Thus, Y. accurata (Bokova, 1985) can be considered as a senior synonym of Y. biformis. Other samples containing Purella antiqua and P. cristata also contain Yunnanopleura shells (SOM 1). Yunnanopleura sp. (Fig. 37) is generally similar to Y. accurata, but has a more extended apex which overhangs the apertural margin. The fossils occur as internal moulds with concentric folds covered with minute tubercules. Yunnanopleura sp. co-occurs with P. cristata (Fig. 35).
Given the widespread co-occurrence of generally higher Purella-like shells with more flattened Yunnanopleura-like forms in Siberia, western Mongolia, France, and South China, and including similarity in their preservation, shell structure, and textures on the surface of internal moulds, it seems likely that both form genera derive from the same, probably Halkieria-like scleritome with two terminal shells and, probably, siphogonuchitid-type sclerites (Bengtson 1992; Yang et al. 2014).
We hypothesize herein therefore that Purella antiqua and Yunnanopleura accurata, as well as P. cristata and Yunnanopleura sp., are components of the same scleritomes, respectively P. antiqua and P. cristata, where they represented the opposing, terminal shells/sclerites of a slug-like organism. A polyplacophoran affinity of Yunnanopleura was also discussed by Yu (2014). Without direct evidence from complete maikhanellid scleritomes, however, these forms are described in this report as form taxa under their original or revised generic names as the conchs of single-shelled organisms.
Stratigraphic and geographic range.—Fortunian Stage of Siberia and South China.

Fig. 36. Molluscs Yunnanopleura accurata (Bokova, 1985), internal moulds with partly phosphatized shells (B, H), from early Cambrian Nemakit-Daldyn (A, E, H) and Emyaksin (B–D, F, G) formations, Anabar Uplift, Siberia, Russia; samples K1a/47 (A) and 2/A (E), section 2; 3/0 (H), section 3; 4/27.5 (C, G), section 96-4, and 5/0 (B, D, F), section 96-5. A–H. SMNH Mo182349 (A), SMNH Mo160405 (B; see also Kouchinsky 2000a: fig. 4I), SMNH Mo182350–182355 (C–H, respectively). A1, B1, C2, D, E1, F1, G2, H1, upper; A2, B3, E2, F3, H3, subapical; A3, B2, C1, F2, G1, H2, lateral views. Scale bar 500 µm (A, B, C2, D–G, H3), 1 mm (C1, H1, H2).

Fig. 37. Mollusc Yunnanopleura sp., internal moulds, from early Cambrian Medvezhya Formation, western Anabar Uplift, Siberia, Russia; sample 1/50, section 1. A–C. SMNH Mo182356–182358, respectively. A1–C1, upper; A2, B3, C3, lateral; A3, B2, C2, subapical views. Scale bar 500 µm.
Family uncertain
Genus Obscurania Devaere, Clausen, and Steiner, 2013
Type species: Obscurania auriculata (Vasil’eva, 1990); lower Cambrian; Udzha River, Siberia, Russia.
Remarks.—See Devaere et al. 2013 for discussion of this new generic name that replaces the preoccupied designation Obscurella Vasil’eva, 1990, and for synonymy of the latter with Auricullina Vasil’eva, 1998. The type species was assigned to Punctella Zhong, 1977, by Kouchinsky (2000a: 138) and Feng and Sun (2003: 24).
Obscurania auriculata (Vasil’eva, 1990)
Fig. 38A–E.
Material.—13 internal moulds, including figured SMNH Mo182359–182363, from samples 1/29.5 and 1/29.2 (section 1, Fig. 4), 3/12.2, K2/25, K2/26 (section 3, Fig. 3), Medvezhya Formation, western Anabar Uplift, Siberian Platform, Russia. Correlated with the uppermost Fortunian and lower part of Cambrian Stage 2.
Description.—Low cap-shaped shell with broadly oval, slightly flared aperture. Apex rounded, bent, somewhat overhangs arched subapical margin of the aperture and projects slightly beyond it. Internal mould covered by small scattered tubercules (ca. 10 μm wide and 20 μm long) inclined towards the apex.
Remarks.—Parkhaev (2006) and Parkhaev and Demidenko (2010) noted a polygonal texture at the apex. Kouchinsky (2000a) and Parkhaev (2006) interpreted the tubercules as casts of pores in the shell. Obscurania tormoi Devaere, Clausen, and Steiner in Devaere et al., 2013, was distinguished from O. auriculata by a wider and more pronounced subapical sinus, a nearly circular aperture, a more inclined apex, and a flattened subapical field (Devaere et al. 2013). Other forms with a lower shell with tuberculate ornament on the surface of internal moulds, more elongated aperture, a beak-shaped apex that overhangs the apertural margin, and flattened subapical field were referred to Olenellina alutacea Vasil’eva, 1990 (Figs. 38H and probably Fig. 40D). The type and single species of Olenellina Vasil’eva, 1990, was described from the upper Kessyusa Formation of the Olenyok Uplift, Siberia, which is correlated herein with Cambrian Stage 2. Obscurania auriculata is reported to be different in having a higher shell, flared aperture, and tubular tubercules distributed en echelon over the surface of internal moulds. In contrast, irregular small tubercles distributed circumferentially on the apex occur in Olenellina (see Vasil’eva 1998: 78, 79). The species is likely also synonymous with material described from Cambrian Stage 2 strata of South China (Xing et al. 1984; Steiner et al. 2004a) as “Tuberoconus” paucipapillae Yue in Xing et al., 1984.
Stratigraphic and geographic range.—Uppermost Fortunian and Cambrian Stage 2 of Siberia, east Gondwana (South China), and, probably, West Gondwana (Armorican France).
Genus Algomella Val’kov and Karlova, 1984
Type species: Algomella calyptrata Val’kov and Karlova, 1984; Purella antiqua Zone, lower Cambrian (correlated with upper Fortunian); elevation 1291m, Gonam River, Siberia, Russia.
Algomella sp.
Fig. 38F, G.
Material.—Two internal moulds SMNH Mo182364 and 182365, from sample 3/12.2 (section 3, Fig. 3), Medvezhya Formation, western Anabar Uplift, Siberian Platform, Russia. Correlated with Fortunian–Cambrian Stage 2 transitional beds.
Description.—The moulds are bilaterally symmetrical, cone-shaped, with very broad, irregularly oval aperture. Apex blunt, not clearly separated from the rest of the shell, slightly displaced towards one side of the aperture. Subapical side long, with a broad, shallow median fold covered with faint polygons. The surface of the mould (Fig. 38G) is covered with scattered tubercules.
Remarks.—The fossils are generally similar to Algomella spp. (see Val’kov and Karlova 1984: 27–30, pl. 4: 1–3), which lack, however, a recognizable tuberculate ornamentation. Internal moulds of Tannuella taishirica Zhegallo in Esakova and Zhegallo, 1996 from western Mongolia are different in being smooth and lack the subapical fold.

Fig. 38. Problematic molluscs from early Cambrian Medvezhya Formation, western Anabar Uplift, Siberia, Russia; samples 1/29.5 (C, D, F), section 1; K2/25 (A), K2/26 (B), and 3/12.2 (E, G, H), section 3. A–E. Obscurania auriculata (Vasil’eva, 1990), internal moulds, SMNH Mo182359 (A), SMNH Mo182360 (B), SMNH Mo182361a (C), SMNH Mo182363 (D), SMNH Mo182362 (E). F, G. Algomella sp., internal moulds, SMNH Mo182361b (F, lateral view arrowed in C2), SMNH Mo182365 (G). H. Olenellina alutacea Vasil’eva, 1990, internal mould, SMNH Mo182366. A1, B1, C1, D2, E1, G1, H1, lateral; A2, B3, D3, E2, G2, H2, upper; A3, B2, C2, D1, E3, F, H3, subapical views. Scale bar 500 µm (A, B, E, F, G), 1 mm (C, D, H).

Fig. 39. Problematic molluscs from early Cambrian Medvezhya Formation, western Anabar Uplift, Siberia, Russia; samples 3/12.2 (A, B), K2/25 (C), 3/12 (D), and 3/10 (E) from section 3. A, B, D, E. Purella? sp., calcium phosphatic internal moulds, SMNH Mo182367 (A), SMNH Mo182368 (B), SMNH Mo182370 (D), SMNH Mo182371 (E). C. Purella cristata Missarzhevsky, 1974, calcium phosphatic internal mould, SMNH Mo182369. A1–D1, E4, lateral; A2, B2, C3, D3, E2, upper; A3, C2, D2, E3, subapical; E1, supra-apical views. Scale bar 500 µm.

Fig. 40. Problematic molluscs from early Cambrian Medvezhya (E–G) and Kugda-Yuryakh (A–D, H) formations, western Anabar Uplift, Siberia, Russia; samples K1a/67 (A–D), section 2; 1/22 (E–G), section 1; and K2/60 (H), section 3. A, B, G. Purella sp., calcium phosphatic internal moulds, SMNH Mo182372 (A), SMNH Mo182373 (B), SMNH Mo182377 (G). C, E, F, H. Obscurania? sp., calcium phosphatic internal moulds, SMNH Mo182374 (C), SMNH Mo182376 (E), SMNH Mo182364 (F), SMNH Mo182378 (H). D. Olenellina alutacea? Vasil’eva, 1990, calcium phosphatic internal mould, SMNH Mo182375. A1, B3, C1, D1, G2, H3, upper; A2, B1, C2, D3, F, G3, H2, subapical; A3, B2, C3, D2, E1–H1, lateral; E2, apical views. Scale bar 500 µm (A–C, G), 1 mm (E, F, H), 2.5 mm (D).
Genus Protoconus Yu, 1979
Type species: Protoconus crestatus Yu, 1979; Paragloborilus subglobosus‒Purella squamulosa Assemblage Zone, middle Meishucunian Stage (Zone 2), lower Cambrian (upper Fortunian Stage); Yichang County, western Hubei Province, South China.
Remarks.—See Parkhaev and Demidenko (2010), Devaere et al. (2013), and Yang et al. (2014) for discussion of the genus and species.
Protoconus cf. crestatus Yu, 1979
Fig. 41.
Material.—Three internal moulds, SMNH Mo182379–182381, from samples 1/50 and 1/6 (section 1, Fig. 4), Medvezhya and Kugda-Yuryakh formations, western Anabar Uplift, Siberian Platform, Russia. Correlated with upper Fortunian and lower part of Cambrian Stage 2.
Description.—Bilaterally symmetrical, cap-shaped shell with broad oval aperture. Apex rounded, not clearly separated from the rest of the shell. Inner surface of the shell (replicated by the internal mould) with two parallel longitudinal thickenings that flank a wide median buttress that defines the entire dorsal side of the internal mould (Fig. 41A1, B3, C1).
Remarks.—Subapical side short with the subapical shelf that is diagnostic for the genus Protoconus (Devaere et al. 2013; Fig. 41A3). The furrows and median buttress are covered with a low nodular texture with positive relief on the mould (Fig. 41C1, C4). The nodules are ca. 5 μm in diameter. Yang et al. (2014) reported moulds and partial conchs of P. crestatus from the late Fortunian (Assemblage Zone 2) of South China. The latter species is similar and probably synonymous with Purella tianzhushanensis Yu, 1979. The latter was also described by Hamdi (1995: pl. 12: 1, 2) from the Latouchella–Obtusoconus Assemblage Zone (Upper Shale Member, Soltanieh Formation, Albortz Mountains, Iran).

Fig. 41. Problematic mollusc Protoconus cf. crestatus Yu, 1979, calcium phosphatic internal moulds, from early Cambrian Medvezhya and Kugda-Yuryakh formations of western Anabar Uplift, Siberia, Russia; samples 1/50 (A) and 1/6 (B, C) from section 1. A–C. SMNH Mo182379 (A), SMNH Mo182380 (B), SMNH Mo182381 (C). A1, B3, C1, upper; A2, B1, C3, subapical; A3, B2, C2, lateral; A4, supra-apical views; C4, close-up of lateral surface in C3. Scale bar 125 µm (C4), 500 µm (A, B, C1–C3).
Protoconus? sp.
Figs. 33G, 34A, B.
Material.—Three internal moulds, SMNH Mo182338, 182340, 182341, from samples 2/A (section 2, Fig. 1) and 3/0.1 (section 3, Fig. 3), upper Nemakit-Daldyn and basal Medvezhya formations, western Anabar Uplift, Siberian Platform, Russia. Correlated with the upper Fortunian Stage.
Description.—Bilaterally symmetrical, low cap-shaped shell with irregularly ovoid, broad aperture. Apex not clearly distinct from rest of conch, situated near the middle of the shell in plan view, displaced towards the narrower side of the aperture. Inner surface of the shell (replicated by the internal mould) with two parallel longitudinal thickenings that define a wide median buttress flanked by furrows that extend from the apex to the wider side of the aperture. The subapical side is laterally slightly concave and forms a low broad, somewhat arched margin. The opposite side is gently convex.
Remarks.—The apex has a somewhat nodose texture (Fig. 34A1) similar to that in co-occurring Purella antiqua moulds. The same texture occurs laterally to the apex. The distal parts of the mould are covered with a polygonal texture. The top of the mould is smooth with rare tubercules. These types of textures are distributed on the internal mould in a bilaterally symmetrical pattern (Fig. 34A1). Protoconus cf. crestatus (see above) is different in having a more marginally located apex and broader median buttress.
Problematic shells
Figs. 42, 43.
Material.—Specimens: SMNH X5789, from sample K2/26 (section 3, Fig. 3), Medvezhya Formation, and SMNH X5790, from sample 5a/34.5 (section 96-5a, Fig. 2), Emyaksin Formation, Anabar Uplift, Siberian Platform, Russia. Correlated with the upper Fortunian and Cambrian Stage 2, respectively.
Remarks.—The specimen from the Medvezhya Formation (Fig. 42) is a flat fragment of a shell divisible into proximal and distal areas. The proximal area is slightly depressed in the middle and presumably represents the initial part of the shell. It lacks growth lines and consists of granules of probably phosphatized original shell material (Fig. 42C). This proximal part is separated along a sharp margin from the distal area of the shell. The distal shell is covered with numerous concentric growth lines that curve around the proximal part and are superimposed on a microstructure of subradially extended laths composed of parallel aciculate subunits. The latter show a differential orientation, and neighboring laths lie at oblique angles to each other. The general shape of the broken fragment and the microstructure of acicular laths are similar to those of Eohalobia diandongensis Jiang in Luo et al., 1982, which was interpreted by Vendrasco et al. (2009: pl. 1) as part of the chiton-like scleritome of Ocruranus finial (Liu, 1979). The fragment in Fig. 42 may represent the inner surface of a shell similar to that in Fig. 43. They both may belong to the same group of chiton-like molluscs with shells microstructurally different from those of maikhanellids.
The specimen from the Emyaksin Formation (Fig. 43) is a dome-shaped shell with a deep subapical reentrant (Fig. 43B, D). The apex is slightly adapically shifted and forms an elevated flattened disk with a granulated surface (Fig. 43C) separated by a circular rim and suture from the rest of the shell. The outer surface of the shell is covered with concentric growth lines. Although the shell is reminiscent in plan view to a hyolith operulum, the deep reentrant and apparent lack of clavicules make interpretation of the fossil as a hyolith operculum difficult. On the contrary, a general similarity to the shell plates of Ocruranus is more pronounced (e.g., Qian and Bengtson 1989: fig. 67).

Fig. 42. Problematic phosphatized shell fragment (SMNH X5789), from early Cambrian Medvezhya Formation, western Anabar Uplift, Siberia, Russia; sample K2/26. A. Oblique transversal view. B. Plain view on the fragment. C. Apical part enlarged. Scale bar 25 µm (C), 125 µm (A, B).

Fig. 43. Problematic phosphatized shell fragment with internal mould (SMNH X5790), from early Cambrian Emyaksin Formation, eastern Anabar Uplift, Siberia, Russia; sample 5a/34.5. A. Upper view. B. Subapical view; a deep subapical reentrant is covered with an adhered shell fragment. C. Apical part enlarged. D. Lateral view. Scale bar 50 µm (C), 250 µm (A, B, D).
Order Sachitida He in Yin et al., 1980
Family Siphogonuchitidae Qian, 1977
Genus Siphogonuchites Qian, 1977
Type species: Siphogonuchites triangularis Qian, 1977; Watsonella crosbyi Assemblage Zone, middle Meishucunian Stage, lower Cambrian (lower part of Cambrian Stage 2); Maidiping, Emei County, Sichuan Province, South China.
Siphogonuchites triangularis Qian, 1977
Figs. 44C, 45A, B.
Material.—Over 25 phosphatized fragments of sclerites and phosphatic internal moulds, including figured SMNH X5793, 5801, 5802, from samples 1/50 (section 1, Fig. 4), 3/0, 3/0.1, and 3/12.2 (section 3, Fig. 3), Medvezhya Formation and pyritised sclerites/internal moulds from samples K1a/36A and K1a/41A (section 2, Fig. 1), upper Nemakit-Daldyn Formation, western Anabar Uplift, Siberia, Russia. Correlated with the upper Fortunian Stage.
Description.—Elongated asymmetrical sclerites, curved irregularly with curvature either almost in one plane (sometimes S-shaped) but often helically twisted up to 90°. Cross-section irregularly triangular, trapezoidal, or polygonal. Upper/convex sclerite surface transversely arched, with one to three pronounced longitudinal ridges separated by shallow longitudinal furrows. Longitudinal and transverse rugae with a regular pattern of low bumps where rugae intersect. Lower/concave surface smooth and flattened transversely. Sharp lateral ridges at intersection of upper and lower surfaces.
Remarks.—Initially described from the Watsonella crosbyi Assemblage Zone (Zone 3 of the Meishucunian), the type species Siphogonuchites triangularis Qian, 1977, is known to range from Meishucunian Assemblage Zone 1 in South China, where it co-occurs with maikhanellids (e.g., Maikhanella multa) and other siphogonuchitids, such as Dabashanites mirus Chen, 1979, and Lopochites spp. (Qian and Bengtson 1989; Steiner et al. 2004a, 2007; Yang et al. 2014; see below). Qian and Bengtson (1989) discussed variability and provided a comprehensive synonymy of the species and genus (see also Devaere et al. 2013). Siphogonuchites triangularis was first reported in Siberia from the basal Pestrotsvet Formation at Selinde (Khomentovsky and Karlova 1993, not illustrated therein) and, questionably, as Siphogonuchites aff. S. triangulariformis [sic!, should be triangularis] from 60 m below the top of the Ust’-Yudoma Formation at the Mount Konus section (Khomentovsky et al. 1990: 34, pl. 8: 6) on the southeastern Siberian Platform. This occurrence was assigned to the Purella antiqua Zone (Khomentovsky and Karlova 1993).
Stratigraphic and geographic range.—Terreneuvian Series of Siberia (Khomentovsky et al. 1990), western Mongolia (Voronin et al. 1982; Esakova and Zhegallo 1996), South China (Steiner et al. 2007; Yang et al. 2014), and Cambrian Stage 2 of France (Devaere et al. 2013).
Siphogonuchites subremualis (Bokova, 1985)
Fig. 44A, D–G.
Material.—Over 100 phosphatized sclerites and phosphatic internal moulds, including figured SMNH X5791, 5794–5797, from samples 2/A, 1282, K1a/43, and K1a/47 (section 2, Fig. 1), upper Nemakit-Daldyn Formation, western Anabar Uplift, Siberia, Russia. Correlated with the upper Fortunian Stage.
Description.—Flattened sclerites curved in one plane. Cross-section low triangular to trapezoid. Sclerites taper towards the base and form a shaft with irregular cross-section (Fig. 44A, F, G). Upper/convex surface of sclerite with two longitudinal ridges separated by a shallow furrow. The ridges gradually merge into a single ridge that continues towards the acute distal end of the sclerite (Fig. 44E). The ridges are transversed by faint folds that laterally continue at an angle towards the axis of the sclerite (Fig. 44F2). The lower/concave surface of the sclerite is smooth with faint longitudinal striation and rugae. Articulated sclerites are of different thicknesses and packed next to each other along their shaft-like basal parts (Fig. 44A).
Remarks.—Articulated sclerites are also known from Siphogonuchites triangularis (Esakova and Zhegallo 1996: pl. 11). The scleritome of S. triangularis was probably composed of broader and slim sclerites that were adjacent to each other (Esakova and Zhegallo 1996). We agree with Esakova and Zhegallo (1996), who considered Siphogonuchites subremualis a distinctive species. S. subremualis sclerites have two prominent ridges along their almost bilaterally symmetrical blades that are much shorter and taper much faster proximally (to the base) and distally (to the pointed tip) than those of S. triangularis.
Stratigraphic and geographic range.—Upper Fortunian Stage of the Siberian Platform.
Genus Lopochites Qian, 1977
Type species: Lopochites latazonalis Qian, 1977; Paragloborilus subglobosus–Purella squamulosa Assemblage Zone, middle Meishucunian Stage (Zone 2), lower Cambrian (upper Fortunian Stage); Shaanxi Province, South China.
Lopochites sp.
Fig. 45C–K.
Material.—Over 30 internal moulds with fragments of phosphatized walls, including SMNH X5803–5811, from samples 1/46, 1/45, 1/39.3 (section 1, Fig. 4), K2/12, K2/20, K2/21, K2/24, 3/10, 3/12.2 (section 3, Fig. 3), 4/27.5, 5/10, 5/10B, 5a/1.2 (sections 96-4, 96-5, 96-5a), Medvezhya and lower Emyaksin formations, Anabar Uplift, Siberia, Russia; upper Fortunian Stage.
Description.—Elongate, narrow, helically twisted (up to 90º) and gently longitudinally bent sclerites. Cross-section narrow oval at the base and broader, irregular oval distally. Foramen narrow, oval, and slightly bent, separated from the rest of sclerite by a shallow circumferential depression. Surface of sclerite covered with longitudinal striae; fine transverse striae at the base. Wall composed of longitudinally oriented fibres (Fig. 45D).
Remarks.—Lopochites spp. occur in the Terreneuvian Series of South China, western Mongolia, and Iran (Esakova and Zhegallo 1996; Steiner et al. 2004a). Specimens of Lopochites spp. described from South China (Qian and Bengtson 1989) are phosphatized sclerites, in contrast with our material which is comprised of phosphatic internal moulds. Occasionally, preserved walls of the Siberian sclerites suggest that the outer surface was smooth and lacked the prominent longitudinal ridges present in species from South China and western Mongolia. The outer wall is however not completely preserved and may show a smooth inner surface just above the internal moulds (Fig. 45D). The sclerites are similar to Lopochites in being helically twisted longitudinally and in having a narrow irregularly oval foramen, a rounded cross-section of the distal part (e.g., Qian and Bengtson 1989: fig. 13E6, E7) and similar microstructure of the wall (Qian and Bengtson 1989: fig. 14). Lopochites sp. is generally more similar to L. curtus Grigorieva (in Voronin et al. 1982), whereas L. latazonalis is broader and shorter.
Genus Tianzhushania Qian, Chen, and Chen, 1979
Type species: Tianzhushania longa Qian, Chen, and Chen, 1979; Watsonella crosbyi Assemblage Zone, middle Meishucunian Stage (Zone 3), lower Cambrian (lower part of Cambrian Stage 2); Tianzhushan, Yichang, Hubei Province, South China.
Tianzhushania longa Qian, Chen, and Chen, 1979
Fig. 44H–J.
Material.—Eight fragments of internal moulds from sample K1a/47 (section 2, Fig. 1) and a single internal mould from 4/27.5 (section 96-4, Fig. 2), including figured SMNH X5798–5800, from the upper Nemakit-Daldyn and basal Emyaksin formations, respectively, Anabar Uplift, Siberian Platform, Russia. Correlated with the upper Fortunian Stage.
Description.—Horn-shaped sclerites, gently curved in one plane, with a broadly oval cross-section. Upper/convex surface of sclerite with two to four distinct longitudinal ridges, tranverse folds trend laterally at an angle to the axis of sclerite (Fig. 44J). Lower/concave surface flattened, convex in cross-section with faint longitudinal sculpture (as replicated on internal moulds). Intersection of longitudinal and transverse sculpture produces regularly distributed low bumps (Fig. 44I).
Remarks.—The material includes internal moulds similar to sclerites and internal moulds referred to Lomasulcachites macrus by Devaere et al. (2013: fig. 9). The proximal and distal portions of the sclerites are broken off, but cross-section, regular curvature, and sculpture are similar to the latter species’ sclerites. The Siberian material is also similar to phosphatized sclerites of Tianzhushania tetramera Esakova (in Esakova and Zhegallo 1996: pl. 12: 1‒3) from western Mongolia. Because of their morphological similarity, we consider herein Lomasulcachites Qian and Jiang in Luo et al., 1982 to be a junior synonym of Tianzhushania. Tianzhushania has not been previously reported from the Siberian Platform.
Stratigraphic and geographic range.—Upper Fortunian Stage of Siberian Platform, upper Fortunian (Paragloborilus subglobosus–Purella squamulosa Assemblage Zone) and Fortunian–Cambrian Stage 2 transitional interval (Watsonella crosbyi Assemblage Zone) of South China (Qian and Bengtson 1989; Parkhaev and Demidenko 2010; Devaere et al. 2013; Yang et al. 2014), upper Fortunian of Iran (upper part of Upper Shale Member, Soltanieh Formation, Elborz Mountains; Latouchella–Obtusoconus Assemblage Zone [Zone IV]; Hamdi 1995), upper Fortunian–lower part of Cambrian Stage 2 of western Mongolia (Esakova and Zhegallo 1996) and France (Kerber 1988; Devaere et al. 2013).
Genus Dabashanites Chen, 1979
Type species: Dabashanites mirus Chen, 1979; Paragloborilus subglobosus–Purella squamulosa Assemblage Zone 2, middle Meishucunian Stage (Zone 2), lower Cambrian (upper Fortunian Stage); Shaanxi Province, South China.
Dabashanites mirus Chen, 1979
Fig. 44B.
Material.—Single phosphatized sclerite SMNH X5792 from sample 1282 (section 2, Fig. 1), upper Nemakit-Daldyn Formation, western Anabar Uplift, Siberia, Russia. Correlated with the upper Fortunian Stage.
Description.—Aggregate (1 mm long and ca. 0.4 mm wide) of closely spaced, hollow, blade-like sclerites that gradually merge along their basal parts. Base of the merged group is rectangular in cross-section. Distal parts of the sclerites are gently curved. Sclerites display inner longitudinal fibrosity and transverse striation on the surface.
Remarks.—Steiner et al. (2004a) synonymized Dabashanites with Siphogonuchites. Dabashanites differs, however, from other siphogonuchitids in being composed of merged sclerites with a common base. Dabashanites may actually be a central branched sclerite of a Siphogonuchites-scleritome. We maintain Dabashanites as a separate taxon until the association of siphogonuchitid sclerites and branched Dabashanites is demonstrated. Dabashanites lemidiensis Kerber, 1988, from the upper Fortunian–lower part of Cambrian Stage 2 of France probably represents a fragment of a Maikhanella-like scleritome. A certain degree of similarity exists between fused sclerites of Dabashanites and the blind canals that surround the lumen in the sclerites of halkieriid Thambetolepis (Bengtson et al. 1990). Ontogenetically merged siphogonuchitid sclerites and cap-like shells composed of merged sclerites support the suggestion that the sclerites and shells may have belonged to the same organism (Bengtson 1992).
Stratigraphic and geographic range.—Upper Fortunian Stage of Siberia (Purella Zone), South China (Anabarites trisulcatus–Protohertzina anabarica Assemblage Zone; see Chen 1979; Steiner et al. 2004a); occurrence in the Paragloborilus subglobosus–Purella squamulosa Assemblage Zone of Meishucunian Stage of South China mentioned by Parkhaev and Demidenko (2010) but may reflect reworking. Also occurs in western Mongolia (Mongolodus rostriformis and Halkieria amorpha–Tianzhushania tetramera zones; Esakova and Zhegallo 1996), and Iran (Middle Dolomite Member, Soltanieh Formation, Elborz Mountains; Hamdi 1995).
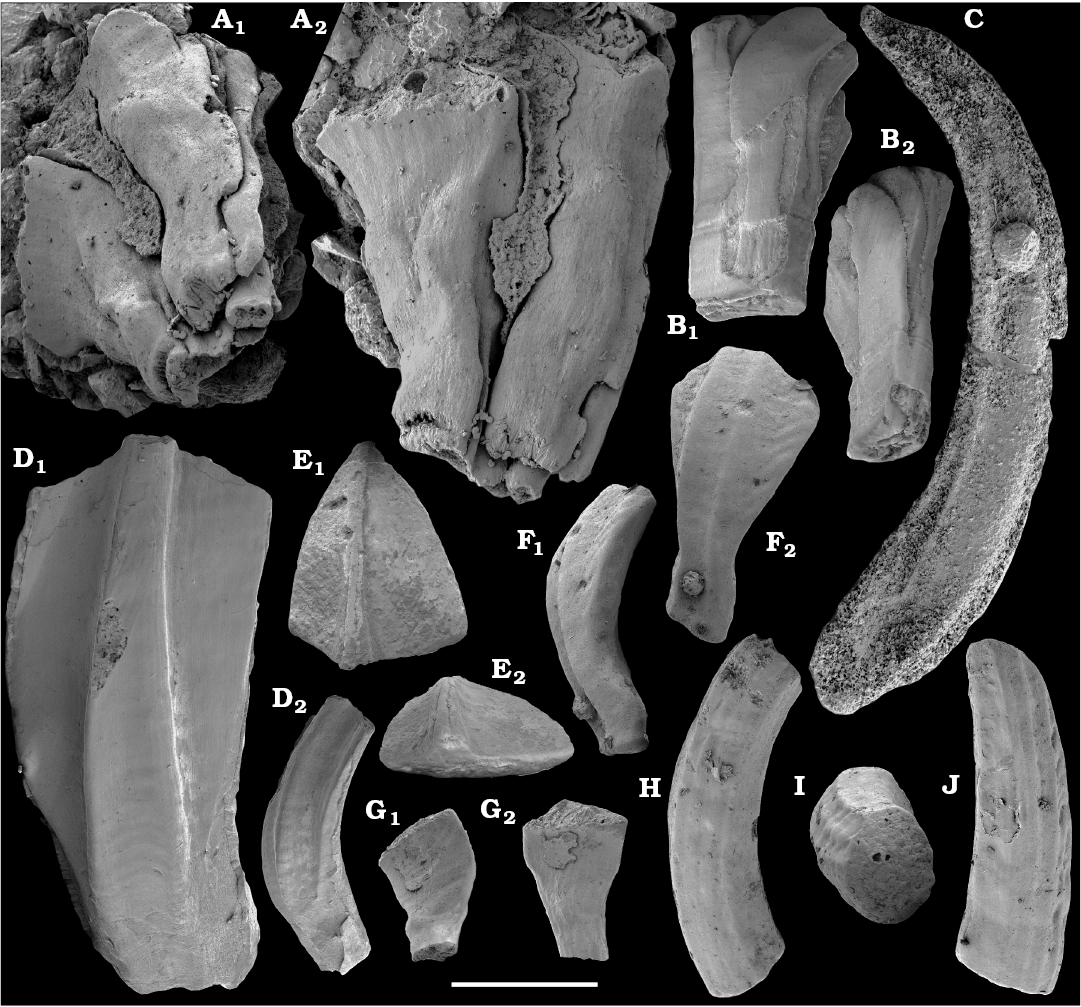
Fig. 44. Siphogonuchitid sclerites from early Cambrian Nemakit-Daldyn Formation of western Anabar Uplift, Siberia, Russia; samples K1a/43 (A), 1282 (B), K1a/36A (C), 2/A (D, E, G), and K1a/47 (F, H–J) from section 2. A, D–G. Siphogonuchites subremualis (Bokova, 1985), calcium phosphatic internal moulds, SMNH X5791 (A), SMNH X5794–5797 (D–G, respectively). A1, oblique lateral view of basal part of articulated sclerite group; A2, lateral view of sclerites; D1, view of lower surface of sclerite; D2, lateral view; E1, view of upper surface at distal end; E2, transversal view; G1, G2, views of lower side of basal part of sclerite. B. Dabashanites mirus Chen, 1979, group of articulated phosphatized sclerites, SMNH X5792; B1, B2, oblique lateral views. C. Siphogonuchites triangularis Qian, 1977; pyritised internal mould in lateral view, SMNH X5793. H–J. Tianzhushania longa Qian, Chen, and Chen, 1979; calcium phosphatic internal moulds, SMNH X5798–5800, respectively; H, J, lateral; I, transverse views. Scale bar 250 µm (A), 500 µm (B, D1, E–J), 1 mm (C, D2).
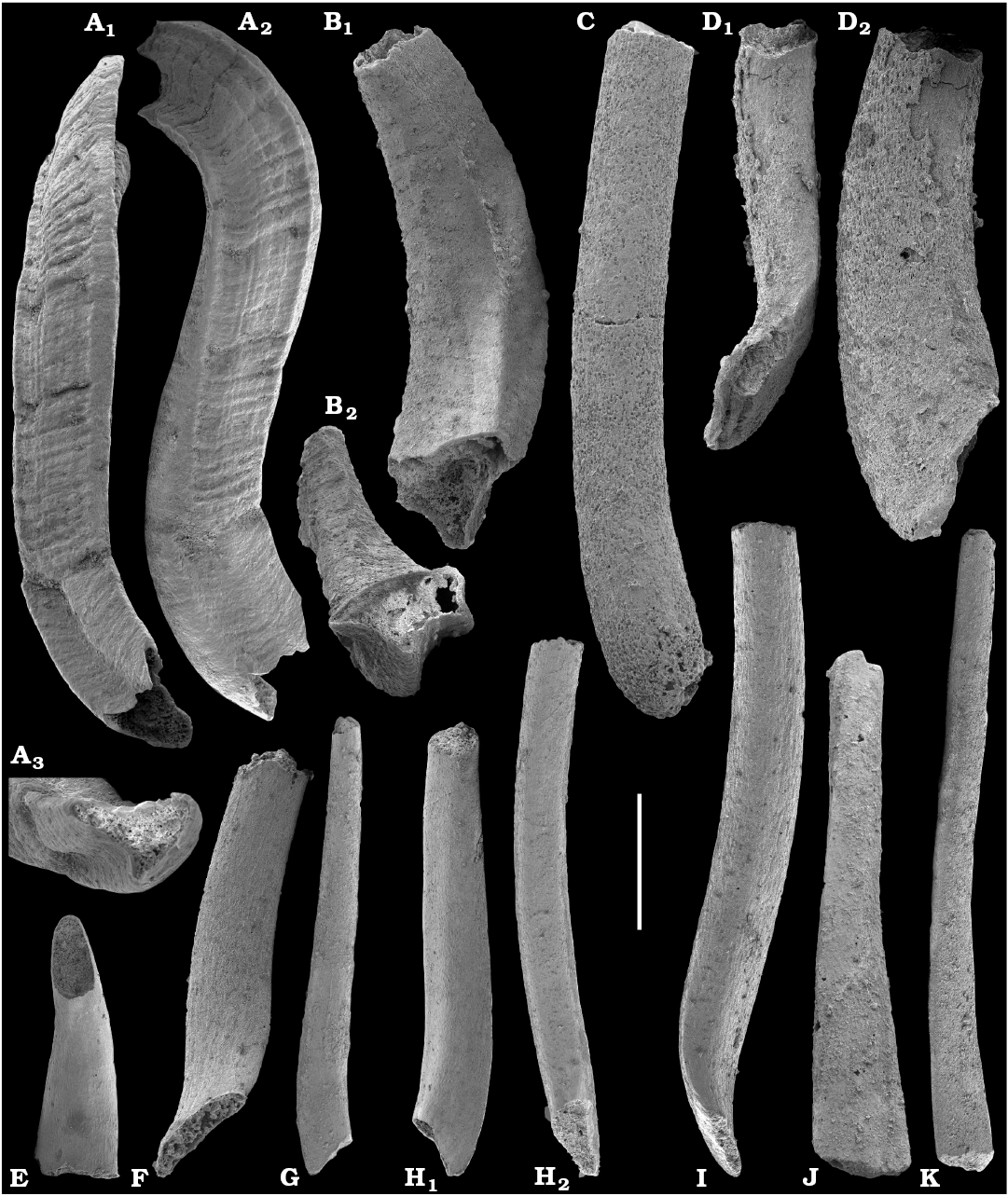
Fig. 45. Siphogonuchitid sclerites from early Cambrian Medvezhya (A, B, D, E, G, H, J, K) and Emyaksin (C, F, I) formations, Anabar Uplift, Siberia, Russia; samples 1/50 (A), 1/45 (H), 1/39.3 (E), section 1; 3/0 (B) and 3/10 (D, G, J, K), section 3; 4/27.5 (C), section 96-4; and 5/10 (F, I), section 96-5. A, B. Siphogonuchites triangularis Qian, 1977, phosphatized sclerites, SMNH X5801, 5802, respectively. A1, A2, B1, lateral views; A3, B2, transverse views of basal end. C–K. Lopochites sp., calcium phosphatic internal moulds with partly preserved phosphatized walls (D), SMNH X5803–5811, respectively. Lateral views with basal end toward bottom of figure. Scale bar 200 µm (D2), 250 µm (C, D1, F, I), 500 µm (A, B, E, G, H, J, K).
Family Halkieriidae Poulsen, 1967
Genus Halkieria Poulsen, 1967
Type species: Halkieria obliqua Poulsen, 1967; lower Cambrian; Bornholm Island, Denmark.
Halkieria spp.
Fig. 46.
Material.—Over 50 fragmentary phosphatized sclerites and internal moulds, including figured SMNH X5812–5827, from samples 1/29.2, 1/22, 1/6 (section 1, Fig. 4), K1a/35A, K1a/67 (section 2, Fig. 1), K2/26, K2/60, K2, 3/12.2, 3/22.5 (section 3, Fig. 3), 5a/6, 5a/18.5, 5a/34.5, 5a/34.75 (section 96-5a, Fig. 2), upper Nemakit-Daldyn, Medvezhya, Kugda-Yuryakh, and Emyaksin formations, Anabar Uplift, Siberian Platform, Russia; correlated with the upper Fortunian and lower part of Cambrian Stage 2.
Discussion.—The majority of sclerites, represented by 35 internal and external moulds and fragments of the wall, are identified as cultrate sclerites referred to Halkieria sp. type A (Fig. 46A, B, I, M, N). They are 2.5–3 times longer than wide, with a flattened broadly triangular cross-section through the sclerites and an irregularly narrow oval cross-section through the lumen (as replicated by internal moulds). The sclerites are gently S-shaped, with the distal end directed dorsally. A distinct median ridge extends from the basal foramen to the pointed distal tip on the upper (dorsal) side, where it is joined by longitudinal ribs and striae. Whereas the dorsal surface is convex in cross-section, the ventral surface is flattened to slightly concave with thin and frequent transverse terraces oriented subparallel to the edge of the foramen. The oval foramen is 1/4 of the maximal sclerite width. The basal part of many sclerites is deflected 60‒90° to the blade of the sclerite. The inner surface of the wall may also be covered with small pits, as replicated by tubercles on the internal moulds (Fig. 46A2). The sclerites are similar to those described as the cultrate sclerites of H. sacciformis (Meshkova, 1969) by Parkhaev and Demidenko (2010) and of H. trianguliformis Mambetov in Missarzhevsky and Mambetov, 1981 from Malyi Karatau (Kazakhstan).
The five cultrate sclerites of Halkieria sp. type B from samples 5a/34.5 and 5a/34.75 (Fig. 46E, F) are fragments of internal moulds (length/width ratio ˃ 3.0). The main part (i.e., blade) is slightly arcuate, has a broadly lenticular cross-section, and intersects the elongate basal part at a right angle. The bases are asymmetrical and rounded triangular or rhomboidal in cross-section. Their foramen is probably elongately elliptical.
Halkieria sp. type C (Fig. 46K, P) is represented by three triangular, flattened, arcuate internal moulds of presumably palmate sclerites with slightly twisted distal parts. The basal part is narrow and elongate, but the detailed structure of this part of the sclerite is unknown. The moulds are covered with fine longitudinal striae.
Several sclerites of Halkieria sp. have a convex upper surface and a flattened lower surface. They are longitudinally bent up to 90° and have a blunt distal end. An oval foramen is situated in the middle of the rhomboidal base. The sclerites are divided by a low median ridge into two almost bilaterally symmetrical sides. A single elongate specimen (Fig. 46C1) has two additional low ridges parallel to the median ridge and a wall composed of transversely elongate laths (Fig. 46C2). The oldest Halkieria sp. in the collections is from the upper Nemakit-Daldyn Formation (sample K1a/35A). These specimens are represented by two internal moulds composed of blocky dolomite (Fig. 46H, L).
Remarks.—Halkieria sp. is known from the upper Nemakit-Daldyn Formation (Fig. 46H, L) correlated with the upper Fortunian. Halkieria spp. are also reported from the upper Ust’-Yudoma Formation (30 m below its top, middle part of the Purella Zone) in the Uchur-Maya region (Khomentovsky et al. 1990: pl. 4: 8, pl. 5: 1–4, pl. 6: 1, 2; Khomentovsky and Karlova 2005). The latter forms are reminiscent of Halkieria sp. type A sclerites from the Medvezhya Formation illustrated herein (see Fig. 46A, B, I, M, N). In South China, the oldest known halkieriids are from upper Assemblage Zone 2 (Annelitellus yangtzensis–Obtusoconus honorabilis Subzone; Yang et al. 2014). In Avalonia (southeast Newfoundland), Halkieria stonei Landing in Landing et al., 1989 occurs in the regionally oldest assemblages of the lower Watsonella crosbyi Zone (Landing 1992; Landing et al. 1989, 1998).

Fig. 46. Sclerites of Halkieria spp. from early Cambrian Nemakit-Daldyn (H, L), Medvezhya (I, G, M–O), Kugda-Yuryakh (A, B, J), and Emyaksin (C–F, K, P) formations, Anabar Uplift, Siberia, Russia; samples K2/60 (A, B, J) and K2/26 (G), section 3; K1a/35A (H, L), section 2; 1/22 (I, M, O) and 1/29.2 (N), section 1; and 5a/34.5 (C–F, K, P), section 96-5a. A, B, I, M, N. Halkieria sp. type A, cultrate sclerites, SMNH X5812, 5813 (A, B, respectively), SMNH X5816 (I), SMNH X5822 (M), SMNH X5823 (N). A1, transversal view of internal mould; A2, close-up showing tuberculate ornamentation of internal mould; A3, I1, M, view of upper (dorsal) side of calcium phosphatic internal moulds; I2, lateral view; B, upper view of internal mould fragment; N1, view of lower (ventral) side of phosphatized sclerite; N2, view of base; N3, transverse view of opposite broken end. C, D, G, J, O. Halkieria sp., phosphatized sclerites, SMNH X5814, 5815 (C, D, respectively), SMNH X5819 (G), SMNH X5825 (J), SMNH X5824 (O). E, F. Halkieria sp. type B, calcium phosphatic internal moulds of cultrate sclerites, SMNH X5817, 5818, respectively. E1, lateral view; E2, F1, view of lower (ventral) side; F2, basal part enlarged. H, L. Halkieria sp., dolomitised internal moulds, SMNH X5820, 5821, respectively. H1, L1, lateral views; H2, L2, views of lower (ventral) side. K, P. Halkieria sp. type C, calcium phosphatic internal moulds of presumed palmate sclerites, SMNH X5826, 5827, respectively. K1, view of apical part; K2, view of lower (ventral) side; P1, lateral view; P2, view of upper (dorsal) side. C1, apical; C2, D1, O1, lateral; C3, D2, G, J, O2, lower (ventral) side views; . Scale bar 100 µm (A2), 250 µm (C2, D1, G), 500 µm (A1, A3, C1, C3, D2, E1, F2, J, K, P), 1 mm (H, L, M, N).
Phylum and Class uncertain
Order Chancelloriida Walcott, 1920
Family Chancelloriidae Walcott, 1920
Genus Chancelloria Walcott, 1920
Type species: Chancelloria eros Walcott, 1920; middle Cambrian; Burgess Shale, British Columbia, Canada.
Remarks.—The earliest undoubted articulated sclerites of Chancelloria sp. in Siberia are reported from the Purella Zone. These sclerites occur in the upper Ust’-Yudoma Formation (30 m below its top) in the Uchur-Maya region (Khomentovsky et al. 1990: pl. 8: 5) and 22–23 m below the top of the Ust’-Yudoma Formation at “Dvortsy” (Val’kov 1983; Khomentovsky et al. 1990: pl. 10: 4?, 5, 6; Varlamov et al. 2008: 29). These occurrences are correlated herein with the upper part of the Fortunian Stage. The locally oldest Chancelloria sp. sclerites are reported from the Ilsanella compressa–Anabarella plana Zone of western Mongolia (Voronin et al. 1982; Brasier et al. 1996) and Watsonella crosbyi Zone of Avalonia (Bengtson and Fletcher 1983; Landing 1988; Landing and Murphy 1991).
Chancelloria symmetrica, reinterpreted as Chancelloriella irregularis by Moore et al. (2014), first appears in upper Assemblage Zone 2 of the Meishucunian Stage (Annelitellus yangtzensis–Obtusoconus honorabilis Subzone, upper Fortunian Stage) in South China. Apart from the oldest chancelloriids with central rays from the upper Fortunian, a probable relative of the chancelloriids is Cambrothyra ampulliformis (= truncata) Qian and Zhang, 1983, with single-rayed sclerites, is known from the lower‒middle Fortunian Assemblage Zone 1 of South China (Steiner et al. 2004a: fig. 2; Moore et al. 2010).
Chancelloria symmetrica Vasil’eva, 1985
Fig. 47A, B, F.
Material.—Single 7+1 sclerite from sample 5a/18.5; single 8+1 and three 6–7+1 sclerites from 5a/34.5; 7+1, 8+1 and 9+1 sclerites from 5a/34.75 (section 96-5a, Fig. 2); single 7+1 sclerites from samples 1/6 (section 1, Fig. 4) and K2, as well as several disarticulated sclerites from K2/60 (section 3, Fig. 3), including figured SMNH X5828, 5829, and 5833, from the Kugda-Yuryakh and Emyaksin formations, Anabar Uplift, Siberia, Russia. Correlated with Cambrian Stage 2.
Description.—Articulated sclerites consist of 6–8 lateral rays gently curved away from the plane of the basal facet and the one central ray (Fig. 47A, F). The rays are enlarged at their bases. Base of the central ray 1.5‒2.0 times greater in diameter than the lateral ray bases. Basal facet of each ray flattened, covered with a polygonal network (Fig. 47B), and has a central foramen. Facets of the rays lie on a shared, flattened basal surface.
Remarks.—Chancelloria symmetrica was described by Vasil’eva (1985) from the Cambrian Stage 2 equivalent beds of the Siberian Platform. Conspecific forms are reported from western Mongolia, Australia, and South China (Parkhaev and Demidenko 2010: 901), although the assignment of sclerites from South China to C. symmetrica by Parkhaev and Demidenko (2010) was questioned by Moore et al. (2014). Chancelloria cf. eros Walcott, 1920, described from the Emyaksin Formation (Kouchinsky et al. 2015a), is different from C. symmetrica in having relatively longer, slim rays which are much less inflated at their bases. A polygonal microstructure on the basal facet of one articulated sclerite (Fig. 47B) is similar to the botryoidal microstructure observed in other chancelloriid sclerites (see Bengtson et al. 1990; Moore et al. 2014).
Phylum, class, order, and family uncertain
Problematic sclerite
Fig. 47C.
Material.—Single internal mould, SMNH X5831, from sample K2/21 (section 3, Fig. 3), Medvezhya Formation, western Anabar Uplift, Siberia, Russia. Correlated with the upper Fortunian Stage.
Description.—Elongate, slightly curved sclerite with nearly circular cross-section. Distal termination bluntly rounded. The sclerite becomes slightly wider towards the basal end, where it abruptly narrows into a tubular structure about half the diameter of the widest part of the sclerite. The tubular base has one prominent and several less distinct transverse, circular folds.
Remarks.—The fossil may represent an internal mould of the monorayed chancelloriid Eremactis Bengtson and Conway Morris in Bengtson et al., 1990. It is somewhat reminiscent of spine-shaped sclerites that form the apical tuft in chancelloriids (Bengtson and Collins 2015). An alternative interpretation as a siphogonuchitid or halkieriid sclerite is difficult because of the lack of dorso-ventrally defined morphology.

Fig. 47. Sclerites of Chancelloria from early Cambrian Emyaksin (A, B) and Kugda-Yuryakh (D, E, F) formations, and a problematic sclerite from Medvezhya Formation (C), Anabar Uplift, Siberia, Russia; samples 5a/34.5 (A, B), section 96-5a; K2/21 (C), K2/60 (D, E), and K2 (F), section 3. A, B, F. Chancelloria symmetrica Vasil’eva, 1985, SMNH X5828, 5829 (A, B, respectively), SMNH X5833 (F). A, F, calcium phosphatic internal moulds; B, basal surface of internal mould with outer phosphatic coating of the wall. C. Calcium phosphatic internal mould of a problematic sclerite, SMNH X5831. C1, lateral view; C2, basal part enlarged. D, E. Chancelloria sp., fragmentary internal moulds of central rays, SMNH X5830 (D), SMNH X5832 (E). Scale bar 125 µm (C2), 250 µm (A, B, C1, E), 1 mm (D, F).
Phylum, class, and order uncertain (stem-group Brachiopoda?)
Family Tianzhushanellidae Conway Morris in Bengtson et al., 1990
Remarks.—Tianzhushanellids are represented by disarticulated, bilaterally symmetrical, and probably originally bivalved (although no firm evidence exists, see Li et al. 2014) and aragonitic shells assignable to stem-group brachiopods. The type genus, Tianzhushanella Liu, 1979, was synonymized with Lathamella Liu, 1979 (Esakova and Zhegallo 1996; Parkhaev in Gravestock et al. 2001; Li et al. 2014; compare Li 2012). Other genera attributed to tianzhushanellids are the apistoconchs (Apistoconcha spp.) from Cambrian Stage 3 of Gondwana (Australia and Turkey; Bengtson et al. 1990; Gravestock et al. 2001; Sarmiento et al. 2001; Elicki and Gürsu 2009), North China (Li et al. 2014), Mongolia (Esakova and Zhegallo 1996), and Laurentia (Greenland; Skovsted 2006). Apistoconchs have been discussed as stem-group brachiopods (Balthasar 2008; Skovsted et al. 2010) or referred to the molluscs (Parkhaev 1998; Parkhaev in Gravestock et al. 2001). Although Tianzhushanella ovata is often attributed to paterinid brachiopods, the shell composition of this form was originally calcareous, probably aragonitic (Li and Chen 1992), and is thus different from the primarily phosphatic paterinids.
The first appearance of tianzhushanellids (Tianzhushanella) predates the first brachiopods (Kouchinsky et al. 2010, 2012) such as the organophosphatic paterinids (e.g., Aldanotreta sunnaginensis Pel’man, 1977, from the Nochoroicyathus sunnaginicus Zone of the lower Tommotian Stage of Siberia) and calcareous obolellids (Nochoroiella isitica Pel’man, 1983, and Obolella sp. from the Dokidocyathus lenaicus Zone of the middle Tommotian; see Bengtson et al. 1987; Ushatinskaya and Malakhovskaya 2001) from the upper part of Cambrian Stage 2. Tianzhushanellids (= lathamellids) are reported from lower part of Cambrian Stage 2 of South China, where their first undoubted representatives are known from the Watsonella crosbyi Assemblage Zone (Li and Chen 1992; Steiner et al. 2007; see Kouchinsky et al. 2012: online appendix 1). Besides the poorly illustrated Tianzhushanella ovata Liu, 1979, redescribed by Xing and Yue (in Xing et al. 1984), such forms are also known from South China as Lathamella caeca Liu, 1979, and L. symmetrica Li and Chen, 1992. Tianzhushanella sp. and T. tolli sp. nov. are herein described from the age-equivalent beds of the Siberian Platform. It appears, therefore, that tianzhushanellids are widespread in the terminal Fortunian–lower part of Cambrian Stage 2 interval.
Genus Tianzhushanella Liu, 1979
Type species: Tianzhushanella ovata Liu, 1979; Watsonella crosbyi Assemblage Zone, middle Meishucunian Stage (Zone 3), lower Cambrian (lower part of Cambrian Stage 2); Yangtze Gorges, West Hubei Province and Yuanjiaping section, Ningqiang, Shaanxi Province, South China.
Tianzhushanella sp.
Fig. 48.
Material.—Four internal moulds SMNH X5834, 5835, 5837, 5838, from samples 1/22 (section 1, Fig. 4), K1a/67 (section 2, Fig. 1), and K2/60 (section 3, Fig. 3), from the Medvezhya and Kugda-Yuryakh formations, western Anabar Uplift; and SMNH X5836, from sample M71-2/62, from the upper Kessyusa Formation, Olenyok Uplift, Siberian Platform. Correlated with lower part of Cambrian Stage 2.
Description.—Bilaterally symmetrical shell with broad oval aperture and submarginal apex. Subapical margin of aperture is broadly and strongly arched. Inner surface of apical area has a pair of radially elongated, closely spaced, bilaterally symmetrically situated cavities replicated by a pair of spine-like protrusions on the internal mould (Fig. 48C). The elevated median part of the inner surface near the apex is covered with polygonal imprints, whereas larger imprints occur distally (Fig. 48E).

Fig. 48. Stem-group brachiopod? Tianzhushanella sp., internal moulds, from early Cambrian Medvezhya (A, B) and Kugda-Yuryakh (D, E) formations, western Anabar Uplift, Siberia, Russia; samples 1/22 (A, B), section 1; K2/60 (D), section 3; and K1a/67 (E), section 2; and upper Kessyusa Formation, Olenyok Uplift, Siberia, Russia, sample M71-2/62 (C). A–E. SMNH X5834–5838, respectively. A1, B3–D3, E1, upper; A2, B2–E2, subapical; A3, B1, C1, D4, lateral views; D1, subapical area enlarged; E3, supra-apical proximal area with tubercles enlarged; E4, polygonal texture on surface of mould. Scale bar 50 µm (E3), 100 µm (D1, E4), 500 µm (A–C, D2–D4, E1, E2).
Tianzhushanella tolli sp. nov.
Fig. 49.
Etymology: Named for Siberian Arctic explorer Eduard Gustav von Toll (1858–1902).
Type material: Holotype: SMNH X5839 (Fig. 49A) and paratype: SMNH X5840 (Fig. 49B), collected by Vladimir V. Missarzhevsky.
Type locality: 70°42’34’’ N, 112°47’58’’ E, locality M314 (Rozanov et al. 1969; Fig. 1) on the right bank of the Fomich River, ca. 6 km upstream from the Afanas’ev Lakes, northwestern flank of the Anabar Uplift.
Type horizon: Medvezhya Formation, variegated limestones with glauconite, 7 m above the top of the Nemakit-Daldyn Formation, sample M314/7 (see Rozanov et al. 1969: 44, fig. 9). Correlated herein with the Fortunian‒Cambrian Stage 2 transitional beds.
Material.—The species is known at present only from the type series.
Diagnosis.—Species of Tianzhushanella characterized by shells with a median septum that subdivides the apex into two broad halves that face each other.
Description.—Bilaterally symmetrical shell with broad, oval, nearly circular aperture and with apex which overhangs subapical margin. Subapical margin of the aperture gently concave, broadly and slightly arched (as replicated on the internal moulds). Inner surface of the mould of the apical area is subdivided into two bilaterally symmetrical lobes separated by a broad, median depression that arises at the subapical side (Fig. 49A3, B2), crosses the umbo (Fig. 49A1, B1) and becomes obsolescent on the supra-apical side. Two lateral shallow linear depressions on the mould originate at the subapical margin of the aperture, diverge at ca. 30° to cross lateral parts of the mould, and become obsolescent before reaching the supra-apical margin of the aperture. The apical part and median zone of the mould are covered with an aciculate texture (Fig. 49A1).
Remarks.—The new species differs from other forms of Tianzhushanella and Lathamella in having the apex subdivided into two broad halves by a longer and narrower, median, linear septum (furrow on internal moulds). Other named species of the two genera have smaller umbonal cavities with a relatively broader median depression.
Stratigraphic and geographic range.—Type locality and horizon only.

Fig. 49. Stem-group brachiopod? Tianzhushanella tolli sp. nov., internal moulds, from early Cambrian Medvezhya Formation, northern flank of the Anabar Uplift, Siberia, Russia; sample M314/7 from the Fomitch River section. A. Holotype, SMNH X5839. A1, upper; A2, supra-apical; A3, subapical; A4, lateral views. B. Paratype, SMNH X5840. B1, oblique upper; B2, subapical; B3, lateral views. Scale bar 250 µm (A1, A4), 500 µm (A2, A3, B).
Phylum uncertain
Class Hyolitha Marek, 1963
Order Orthothecida Marek, 1966
Family uncertain
Remarks.—Probably the oldest known orthothecid hyoliths in Siberia have been recovered ca. 60 m below the top of the Ust’-Yudoma Formation from the Mount Konus section on the Uchur River (Fig. 1). These specimens were assigned to Lophotheca socialis Val’kov, 1987, and Lophotheca sp. They derive from the Purella Zone of the Nemakit-Daldynian Stage (Khomentovsky et al. 1990: pl. 3: 6–8; Brasier et al. 1993). Hyoliths have not previously been conclusively identified from the upper Nemakit-Daldyn Formation on the western flank of the Anabar Uplift, although tubular remains referred to as orthothecid hyoliths have been reported there, but not illustrated. The supposed orthothecids include Ladatheca dorsocava (Sysoev, 1959a) (i.e., Bokova 1985; Missarzhevsky 1989), Loculitheca sp. and Spinulitheca sp. (i.e., Bokova 1985), Circotheca kuteinikovi Missarzhevsky in Rozanov et al., 1969 (i.e., Meshkova 1974), and Conotheca and Circotheca (i.e., Shishkin 1974). However, abundant tubular remains with circular to rounded triangular cross-section from these beds were described as Anabarites spp. by Bokova (1985), which makes the earlier assignment of the tubes from the Nemakit-Daldyn Formation to the hyoliths questionable.
In western Mongolia, Pseudorthotheca bistriata Qian, 1978, is found in the basal Purella Zone (Khomentovsky and Gibsher 1996; Brasier et al. 1996). From South China, Conotheca subcurvata (Yu, 1974) is reported from Meishucunian Assemblage Zone 1 (Qian and Bengtson 1989; Steiner et al. 2004a). Other reports of early orthothecids in Meishucunian Assemblage Zone 1 include Kunyangotheca ostiola Qian, 1978; Spinulitheca billingsi (Sysoev, 1962) and Turcutheca crasseocochlia (Sysoev, 1962), in beds 3–4 of the Zhujiaqing Formation at the Meishucun section, eastern Yunnan (Luo et al. 1982) but are not well documented (Kouchinsky et al. 2012). Lophotheca cf. multicostata Qian, 1977 and Conotheca-like forms are also reported from the Lesser Himalaya in an interval correlated with Meishucunian Assemblage Zone 1 of China (Brasier and Singh 1987; Hughes et al. 2005). “Ladatheca” cylindrica (Grabau, 1900) from the upper Fortunian Stage (“Ladatheca” cylindrica Zone) and higher strata is the oldest presumed orthothecid hyolith from Avalonia (Landing et al. 1988, 1989; Landing 1993). However, it is alternatively regarded as operculate tubular problematicum (Landing 1988; Landing et al. 1989) or an annelid (Landing 1993).
Genus Khetatheca Missarzhevsky, 1989
Type species: Circotheca cotuiensis Sysoev, 1959b; lower Cambrian (upper Fortunian–lower part of Cambrian Stage 2); Kotuj River, Siberia, Russia.
Khetatheca cotuiensis (Sysoev, 1959a)
Figs. 50A–C, F–H, 53C.
Material.—About 100 internal and external moulds, including figured SMNH X5841–5843, 5846–5848, 5866, from samples 1/45, 1/39.3, 1/29.5, 1/29.2 (section 1, Fig. 4), K2/21, K2/22, K2/23, K2/24, K2/25, K2/26, 3/10, 3/12, 3/12.2, 3/12.5 (section 3, Fig. 3), 5/10, 5/10B, 5a/1, 5a/1.2 (sections 96-5, 96-5a), Medvezhya and Emyaksin formations, Anabar Uplift, Siberian Platform, Russia. Correlated with the upper Fortunian Stage.
Description.—Conch gently curved and slightly longitudinally helically twisted, with subcircular to broadly oval cross-section. Outer surface covered with widely spaced, weakly to strongly developed transverse folds with flattened tops and numerous superimposed frequent growth lines (Fig. 50A1, A2). Inner surface of the conch (replicated on internal mould) covered with transverse folds. Rare septa concave toward the aperture (Figs. 50F, 53C1, apical parts). Early growth stage (as replicated on internal moulds) with regularly spaced transverse folds separated by relatively broader interspaces (Fig. 50G).
Remarks.—The gently helically twisted transversely oval conchs are a diagnostic feature of the monospecific genus Khetatheca, according to Missarzhevsky (1989). The holotype came from locality M424 along the Kotuj River. Sysoev (1959b: 89) attributed these beds to the Atdabanian Stage, but Missarzhevsky (1989: 160) regarded them as sub-Tommotian or lower Tommotian. According to the chemostratigraphy of the Medvezhya and Kugda-Yuryakh formations (Fig. 1), these beds belong to lower part of Cambrian Stage 2. A single specimen of Khetatheca cotuiensis from sample M424/10 is from the type locality (Fig. 53C).
Stratigraphic and geographic range.—Upper Fortunian and lower part of Cambrian Stage 2 of the Siberian Platform and probably western Mongolia (Missarzhevsky 1989).
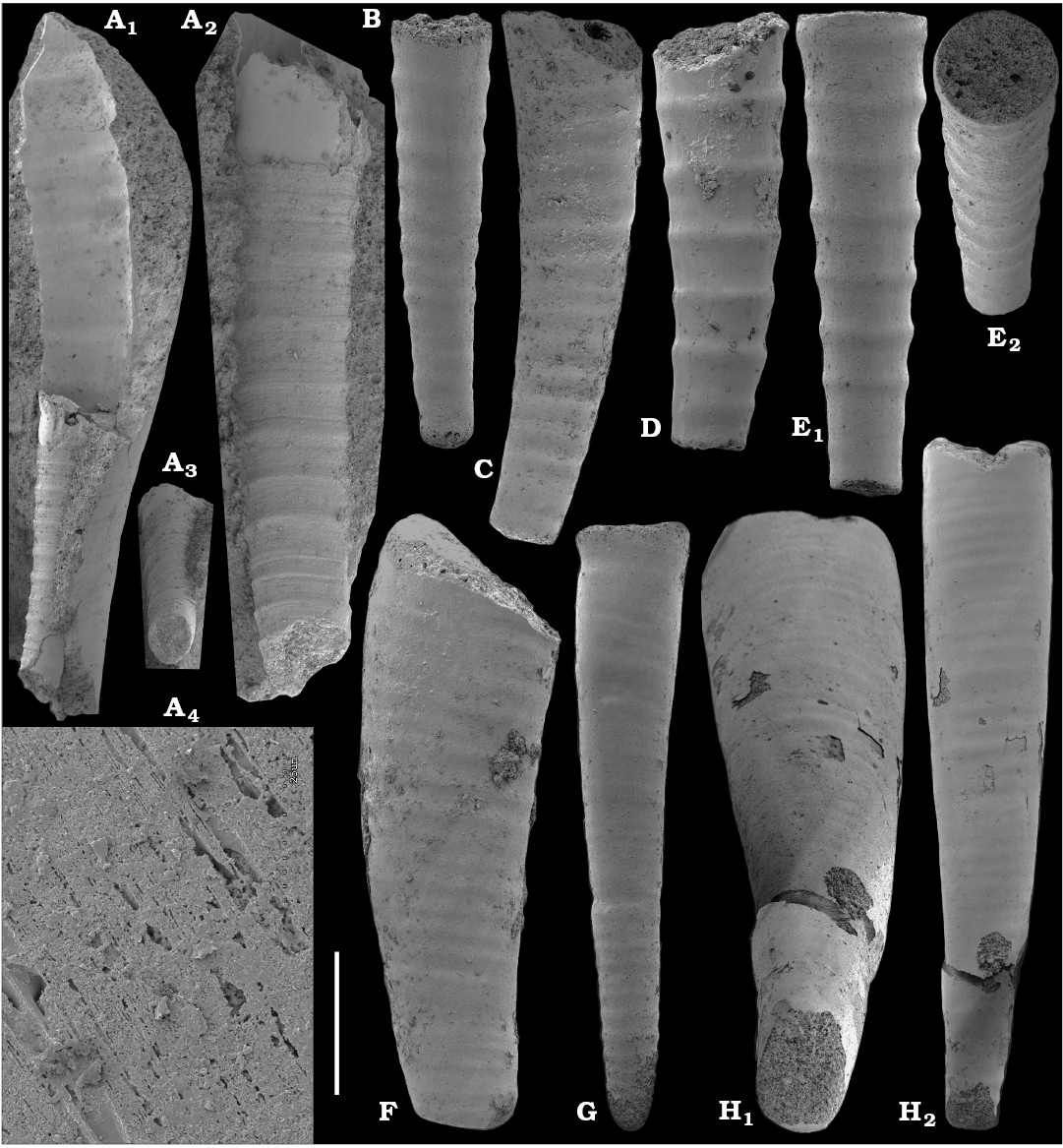
Fig. 50. Orthothecid hyoliths from early Cambrian Medvezhya Formation, western Anabar Uplift, Siberia, Russia; samples K2/21 (A), 3/12 (E), 3/12.2 (B, G, H) and K2/25 (C, D, F) from section 3. A–C, F–H. Khetatheca cotuiensis (Sysoev, 1959b), calcium phosphatic internal moulds, SMNH X5841–5843 (A–C, respectively), SMNH X5846–5848 (F–H, respectively). A1, lateral view; A2, internal mould with fragment of phosphatized wall enlarged; A3, oblique transversal view of apical end; A4, inner surface of phosphatized fibrous wall enlarged; B, C, lateral views of smaller specimens; F, G, H2, lateral; H1, oblique lateral views of larger specimens. D, E. Khetatheca cotuiensis? (Sysoev, 1959b) with wide oval cross-section and wider spaces between transverse folds than in B and C, calcium phosphatic internal moulds; SMNH X5844, 5845, respectively. D, E1, lateral views; E2, oblique transversal view of apertural end. Scale bar 25 µm (A4), 500 µm (A2, B–F), 1 mm (A1, A3, G, H1).
Genus Kotuitheca Missarzhevsky, 1974
Type species: Kotuitheca curta Missarzhevsky, 1974; lower Cambrian (upper Fortunian–lower part of Cambrian Stage 2); Kotuj River, Siberia, Russia.
Kotuitheca curta Missarzhevsky, 1974
Fig. 51A–E.
Material.—About 20 internal moulds, including SMNH X5849–5853, from samples 3/12.2 (section 3, Fig. 3) and M321/31 (see below), Medvezhya Formation, Anabar Uplift, Siberian Platform, Russia. Correlated with the upper Fortunian Stage.
Description.—Short conch with a broadly oval, nearly circular cross-section, bluntly pointed apex weakly separated from the rest of the conch. Conch generally unevenly curved in one plane. It is gently bent or almost straight up to the middle of the conch or even closer to the aperture, where the curvature becomes more pronounced.
Remarks.—Several topotypes representing internal moulds were recovered from sample M321/31 collected from the Rassokha River (section M321 in Rozanov et al. 1969; Missarzhevsky 1974: 181, pl. 23: 1, 2; Missarzhevsky 1989: 158, pl. 3: 5, pl. 4). The name spelling of the Kotuyitheca in Missarzhevsky (1974) was corrected to Kotuitheca in Missarzhevsky (1989).
Stratigraphic and geographic range.—Upper Fortunian Stage of the Siberian Platform.
Genus Paragloborilus He in Qian, 1977
Type species: Paragloborilus subglobosus He in Qian, 1977; Watsonella crosbyi Assemblage Zone, middle Meishucunian Stage, lower Cambrian (upper Fortunian–lower part of Cambrian Stage 2); Gaoqiao section, Emei County, Sichuan Province, South China.
Paragloborilus sp.
Fig. 51I, J.
Material.—Two internal moulds, SMNH X5857, 5858, from samples K2/21 (section 3, Fig. 3) and K1a/80 (section 2, Fig. 1), respectively, from the Medvezhya Formation, western Anabar Uplift, Siberian Platform, Russia. Correlated with the upper Fortunian Stage.
Description.—Conch short, curved in one plane, with circular cross-section and bulbous protoconch separated by a shallow circular constriction (as replicated on smooth internal moulds).
Remarks.—The fossil is similar in general shape to Paragloborilus subglobosus, but is represented by smooth internal moulds. By contrast, P. subglobosus typically shows faint transverse growth lines on the inner molds. However, the syntypes of this species also include relatively smooth steinkerns (see Steiner et al. 2007: fig. 6L, M). Paragloborilus subglobosus is described as having a broadly oval (e.g., Parkhaev and Demidenko 2010) or circular (Qian and Bengtson 1989) transverse cross-section. Salanytheca Missarzhevsky, 1981, was synonymized with Paragloborilus by Qian and Bengtson (1989). Except for one species, Paragloborilus papillaris (Missarzhevsky, 1981) with prominent transverse ribs from the lower Cambrian of the Siberian Platform and western Mongolia, other known species of Paragloborilus are only known from the middle Meishucunian Stage of South China (see Parkhaev and Demidenko 2010; but note statement in Yang et al. 2014 on an erroneous report of Paragloborilus sp. from the Fortunian Anabarites trisulcatus–Protohertzina anabarica Assemblage Zone).
Genus Cupitheca Duan in Xing et al., 1984
Type species: Paragloborilus mirus He in Qian, 1977; middle Meishucunian Stage, lower Cambrian (upper Fortunian–lower part of Cambrian Stage 2); South China.
Cupitheca sp.
Fig. 51K.
Material.—Two internal moulds, including figured SMNH X5859, from sample 5a/17.5 (section 96-5a, Fig. 2), Emyaksin Formation, eastern Anabar Uplift, Siberian Platform, Russia. Correlated with lower part of Cambrian Stage 2.
Remarks.—The fossils are represented by fragments of internal moulds with longitudinal striation, broadly oval to nearly circular cross-section, and hemispherical narrower end formed by a septum. They are most similar to Actinotheca mira (He in Qian, 1977) from the middle Meishucunian Stage of South China (see discussion of the genera Actinotheca Xiao and Zhou, 1984 and Cupitheca by Bengtson et al. 1990: 203; Gravestock et al. 2001: 97), but have a less prominent blunt termination. Cupitheca mira is also reported from the Tommotian and Atdabanian stages of the Siberian Platform (Parkhaev and Demidenko 2010: 951). Cupitheca sp. is herein interpreted as an orthothecid hyolith with a hemispherical, apically convex septum. A similar fragment of an internal mould, albeit with an oval and less prominent terminal septum, was illustrated by Kouchinsky et al. (2015a: fig. 25D) as Loculitheca? sp. from the Atdabanian beds of the Emyaksin Formation (section 96-7 therein).
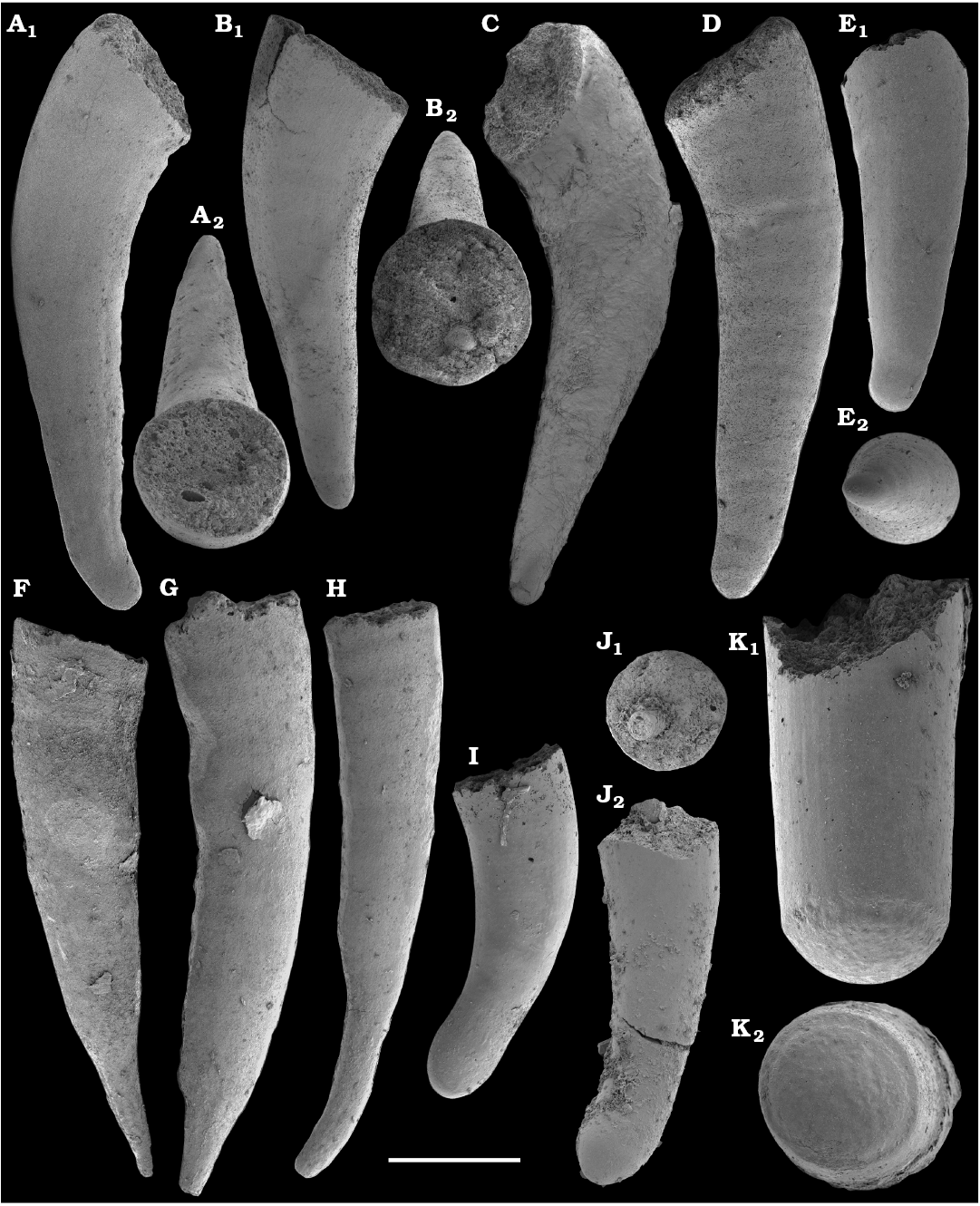
Fig. 51. Orthothecid hyoliths, internal moulds, from early Cambrian Medvezhya (A–G, I, J) and Emyaksin (H) formations, Anabar Uplift, Siberia, Russia; samples M321/31 (A–C), Rassokha River section; 3/12.2 (D, E), K2/21 (F– I), section 3; K1a/80 (J), section 2; 5a/17.5 (K), section 96-5a. A–E. Kotuitheca curta Missarzhevsky, 1974; SMNH X5849–5853, respectively. A1, B1, C, D, E1, lateral; A2, B2, apertural; E2, apical views. F–H. Orthothecida sp. indet. 1, SMNH X5854–5856, respectively. Lateral views. I, J. Paragloborilus sp., SMNH X5857, 5858, respectively. I, J2, lateral views; J1, transverse view of apertural end. K. Cupitheca sp., SMNH X5859. K1, lateral view; K2, apical view of septum. Scale bar 250 µm (K), 500 µm (A–J).
Genus Conotheca Missarzhevsky in Rozanov et al., 1969
Type species: Conotheca mammilata Missarzhevsky in Rozanov et al., 1969; upper Tommotian Stage, lower Cambrian (upper part of Cambrian Stage 2); Tiktirikteekh, near Churan village, middle Lena River, Siberian Platform, Russia.
Conotheca cf. mammilata Missarzhevsky in Rozanov et al., 1969
Fig. 52.
Material.—Four phosphatized conchs, SMNH X5860–5862 and Mo160438 (Kouchinsky 2000b: fig. 2), from samples 5a/34.5 and 5a/34.75 (section 96-5a, Fig. 2), Emyaksin Formation, eastern Anabar Uplift, Siberian Platform, Russia. Correlated with Cambrian Stage 2.
Description.—Gently dorso-ventrally curved conchs with circular sross-section, up to several mm long, with transverse growth lines. In dorsal view, apical angle is ca. 15°. Conch wall consists of fibres (microstructure is detailed by Kouchinsky 2000b). These may intensely branch and anastomose, and their typical orientation in the available broken sections is transverse (Fig. 52C).
Remarks.—Identical phosphatized conchs occur in the Atdabanian portion of the Emyaksin Formation (sections 96-3 and 96-7; Kouchinsky et al. 2015a: fig. 27A–E, H).
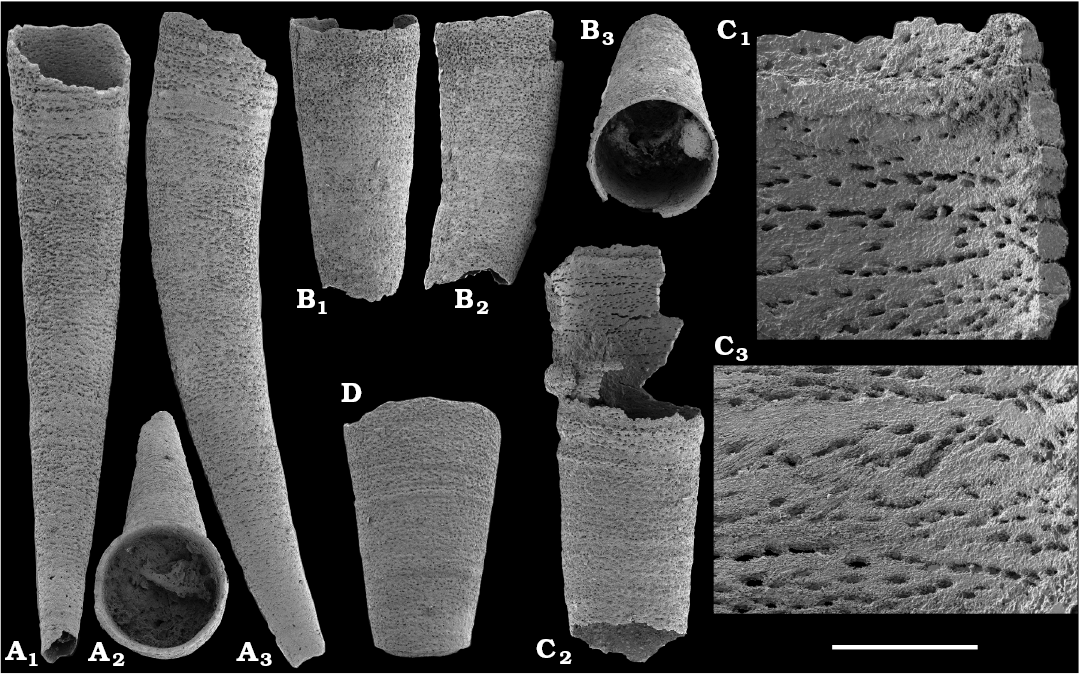
Fig. 52. Orthothecid hyoliths Conotheca cf. mammilata Missarzhevsky in Rozanov et al., 1969, phosphatized tubes, from early Cambrian Emyaksin Formation, eastern Anabar Uplift, Siberia, Russia; samples 5a/34.5 (A), 5a/34.75 (B–D), section 96-5a. A, B, D. SMNH X5860 (A), SMNH X5861 (B), SMNH X5862 (D). C. SMNH Mo160438 (see Kouchinsky 2000b: fig. 2). A1, A3, B1, B2, C2, D, lateral views; A2, B3, transversal views of apertural end; C1, C3, fibrous microstructure of tubular wall, enlarged. Scale bar 100 µm (C1, C3), 500 µm (A, B, C2, D).
Genus Turcutheca Missarzhevsky in Rozanov et al., 1969
Type species: Torellella crassecochlia Sysoev, 1962; Tommotian Stage, lower Cambrian (upper part of Cambrian Stage 2); 1 km downstream from the village of Krestyakh, middle reaches of the Lena River, Siberia, Russia.
Turcutheca sp.
Fig. 53A, B, D–G.
Material.—Over 20 internal moulds, including figured SMNH X5864, 5865, 5867–5870, from samples 1/25.5, 1/22.5, 1/22, and 1/6 (section 1, Fig. 4), K2/60 (section 3, Fig. 3), 5a/34.5 and 5a/34.75 (section 96-5a, Fig. 2), Medvezhya, Kugda-Yuryakh, and lower Emyaksin formations, Anabar Uplift, Siberian Platform, Russia. Correlated with the Cambrian Stage 2.
Remarks.—The fossils are represented by phosphatic, smooth and gently bent internal moulds (Fig. 53A, B, D, E, G) and phosphatic coatings (Fig. 53F1, F2) of septate conchs with the broadly oval cross-section. They include elongate (up to several cm), slowly expanding conchs with rare oval septa convex towards the apex. From the same horizon as sample K2/60 (Fig. 53E, G), from the left bank of the Kotuj River, 6 km downstream the mouth of the Kotujkan River, similar moulds that are larger and have transverse rugae were described by Missarzhevsky (in Rozanov et al. 1969: 108, pl. 1: 8) as Turcutheca rugata (Sysoev, 1959b).
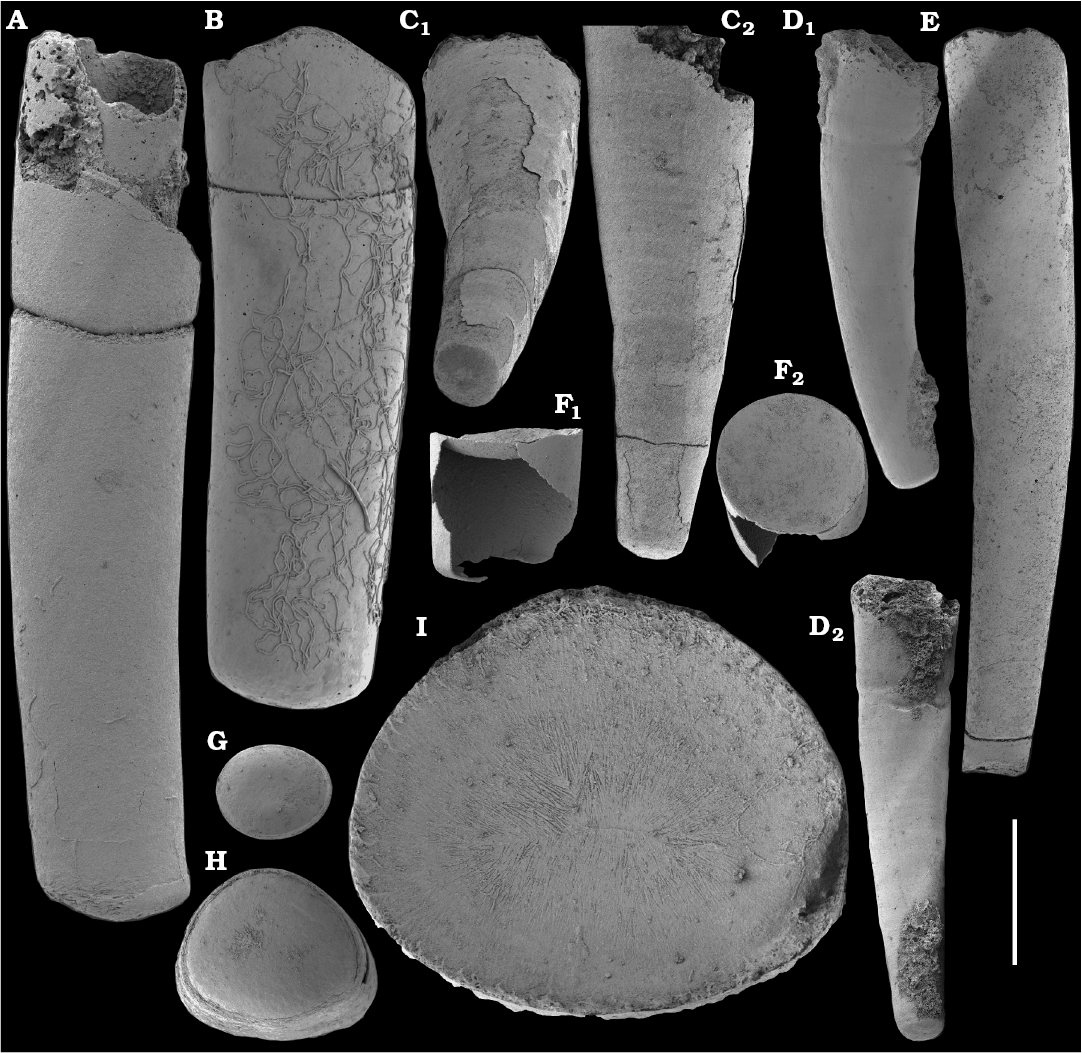
Fig. 53. Orthothecid hyoliths Turcutheca sp. (A, B, D–G) and Khetatheca cotuiensis (Sysoev, 1959b) (C), from early Cambrian Emyaksin (A, B, F, H, I), Medvezhya (C, D), and basal Kugda-Yuryakh (E, G) formations, Anabar Uplift, Siberia, Russia; samples 1/22 (D), section 1; K2/60 (E, G), section 3; 5a/34.5 (F, H, I) and 5a/34.75 (A, B), section 96-5a; M424/10 (C), section M424. A, B, D–G. SMNH X5864, 5865 (A, B, respectively), SMNH X5867–5870 (D–G, respectively). A, B, D, E, lateral views of calcium phosphatic internal moulds with transverse septae; F1, lateral view of phosphatic coating of tube fragment with two septae; F2, oblique transverse view of septum near aperture of fragment; G, transverse view of oval septum from apical side of internal mould. C. SMNH X5866, calcium phosphatic internal mould. C1, oblique lateral view of blunt apical end with septum; C2, lateral view of mould with two transverse septae at apical end. H, I. SMNH X5871, 5872, respectively, calcium phosphatic inner coatings of the hyolith conch wall and septae. Transverse views of rounded triangular septum from the apical (H) and apertural (I) sides. Scale bar 500 µm (A, B, I), 1 mm (C–H).
Orthothecida sp. indet. 1
Fig. 51F–H.
Material.—Three phosphatic internal moulds, SMNH X5854–5856, from sample K2/21 (section 3, Fig. 3), Medvezhya Formation, western Anabar Uplift, Siberian Platform, Russia. Correlated with the upper Fortunian Stage.
Remarks.—The fossils are internal moulds of thecae with low transverse rugae, a moderate apical angle and more rapidly tapered extended apex with a rounded end. The moulds are slightly bent dorso-ventrally (?) in a single plane, which passes through the narrower sides of the broadly oval cross-section.
Orthothecid operculum type A
Fig. 54A–D.
Material.—Four partly phosphatized opercula, SMNH X5873–5876, from samples 1/29.5 and 1/22 (section 1, Fig. 4), K2/23 and 3/10 (section 3, Fig. 3), Medvezhya Formation, western Anabar Uplift, Siberia, Russia. Correlated with the upper Fortunian‒lower part of the Cambrian Stage 2.
Description.—Operculum with domal shape in lateral view and broadly oval in planar view. Concave inner surface smooth and featureless. Outer surface with concentric folds and growth lines; a single, low radial buttress extends from the apex to the apertural margin along one of the shorter sides (dorsal?) of the operculum and emphasises its bilateral symmetry. Presumed dorsal margin of aperture gently protrudes.
Remarks.—The smaller opercula (Fig. 54C2, D2) are more flattened than the larger one (Fig. 54A2) from the same sample 3/10.
Orthothecid operculum type B
Fig. 54E, F.
Material.—Ten phosphatized opercula, including figured SMNH X5877, 5878, from samples 3/10 and K2/21 (section 3, Fig. 3), Medvezhya Formation, western Anabar Uplift, Siberian Platform, Russia, with a broad circular to elongate ovoid outline in planar view. Middle part of the Platform. Correlated with the upper Fortunian Stage.
Description.—Thick-walled operculum with domal lateral profile. Broad rounded apex of operculum smooth, slightly shifted towards the narrower, presumably dorsal side of the margin. Margin of the operculum slightly flared with faint growth lines. Deeply concave inner surface smooth with flattened marginal rim.
Remarks.—A presumably smaller specimen of the same species (Fig. 54E) is nearly circular in planar view.
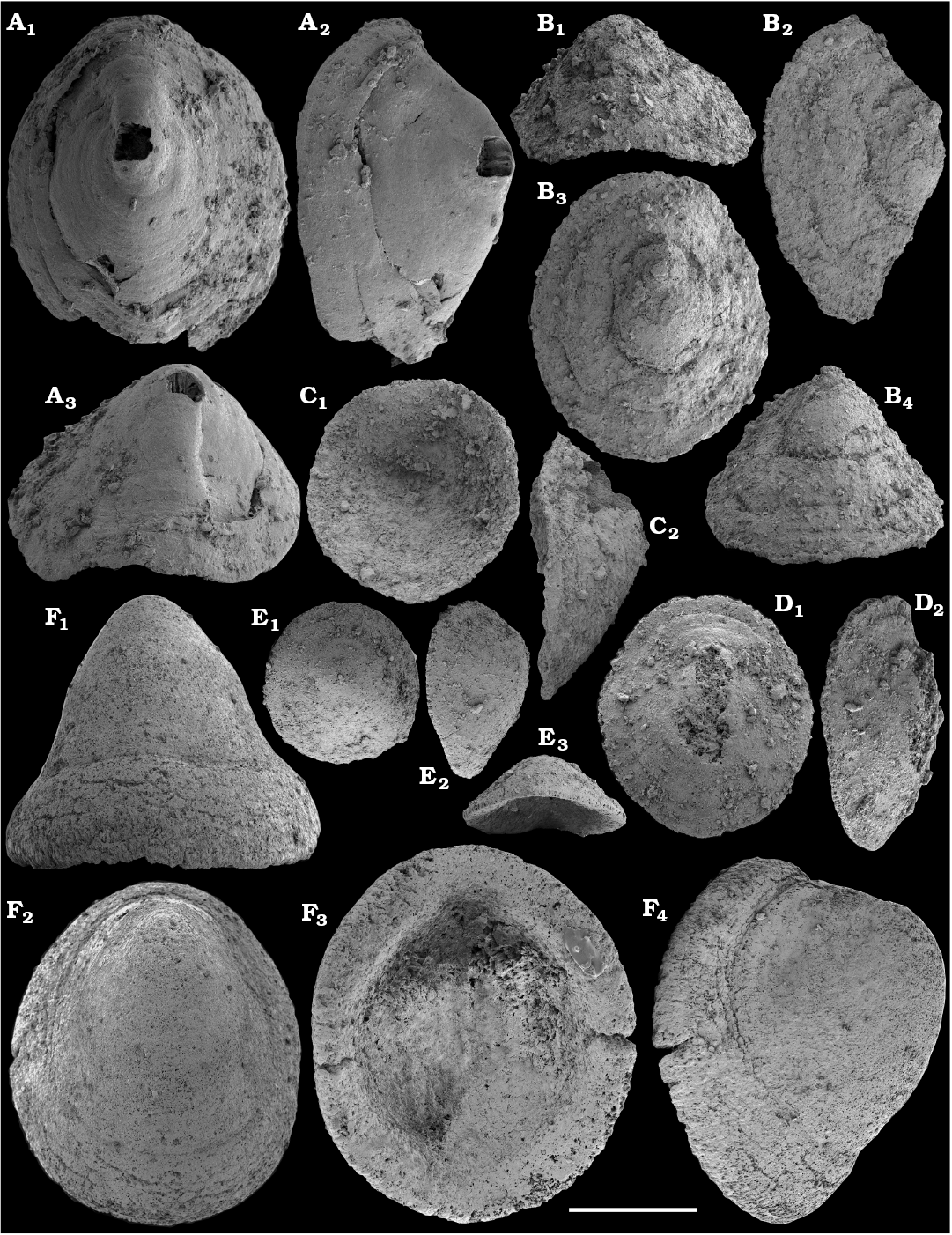
Fig. 54. Opercula of orthothecid hyoliths type A (A–D) and type B (E, F), from early Cambrian Medvezhya Formation, western Anabar Uplift, Siberia, Russia; sample 3/10 from section 3. A–D. SMNH X5873–5876, respectively. A1, B3, D1, apical; A2–D2, lateral views; A3, B1, views of upper margin of operculum (presumably dorsal); B4, view of lower margin of operculum (presumably ventral); C1, view of inner surface of operculum. E, F. SMNH X5877, 5878, respectively. E1, F2, apical; E2, F4, lateral views; E3, view of upper margin of operculum (presumably dorsal); F1, view of lower margin of operculum (presumably ventral); F3, view of inner surface of operculum. Scale bar 250 µm (A), 500 µm (B–F).
Orthothecid operculum type C
Fig. 55A, B, D, F, H.
Material.—Partly phosphatized opercula, SMNH Mo160464 and SMNH X5886, from samples 5/10B and 5a/34.75 (sections 96-5 and 96-5a, Fig. 2), from the lower Emyaksin Formation, eastern Anabar Uplift, and SMNH X5880, 5882, 5883, from sample M314/12, collected by Vladimir V. Missarzhevsky from the Medvezhya Formation, northern Anabar Uplift (see Rozanov et al. 1969). Correlated with the lower part of Cambrian Stage 2.
Description.—Operculum slightly convex, shield-like in lateral view and rounded trapezoidal in planar view, with narrower, presumed dorsal margin. Elevated apex ranges from central location (Fig. 55C2) or lies closer to dorsum. Two low blade-like clavicles on the visceral surface roughly parallel the dorsal margin and form a very wide, oblique angle (Fig. 55B2, D, F). Outer surface with concentric growth lines. Wall relatively thick, consists of radially oriented aciculate elements.
Remarks.—This type of operculum (Fig. 55A) was illustrated by Kouchinsky (2000b: fig. 7B) as an allathecid? operculum, and its microstructure was discussed. Similar microstructures are observed in the opercula from sample M314/12 (Fig. 55B–D, F, H). Among them, the largest operculum (Fig. 55B) preserves the outermost layer with concentric growth lines and underlying radially oriented fibre bundles. The specimens in Fig. 55D and F feature phosphatic coatings of a now-dissolved, strongly raised inner surface of the operculum.
Orthothecid operculum type D
Fig. 55G, H.
Material.—Over 20 dolomitized opercula, including SMNH X5885, from sample K2/22 (section 3, Fig. 3), Medvezhya Formation, western Anabar Uplift, Siberia, Russia. Correlated with the Fortunian–Cambrian Stage 2 transitional beds.
Description.—Thick walled opercula with a domal shape in lateral view and broadly oval shape in planar view. The apical part is almost central, but the more marginal surface slopes evenly towards the periphery. Approximately midway to the margin, the slope abruptly becomes steeper and produces a slightly concave marginal rim. Outer surface with concentric growth lines.
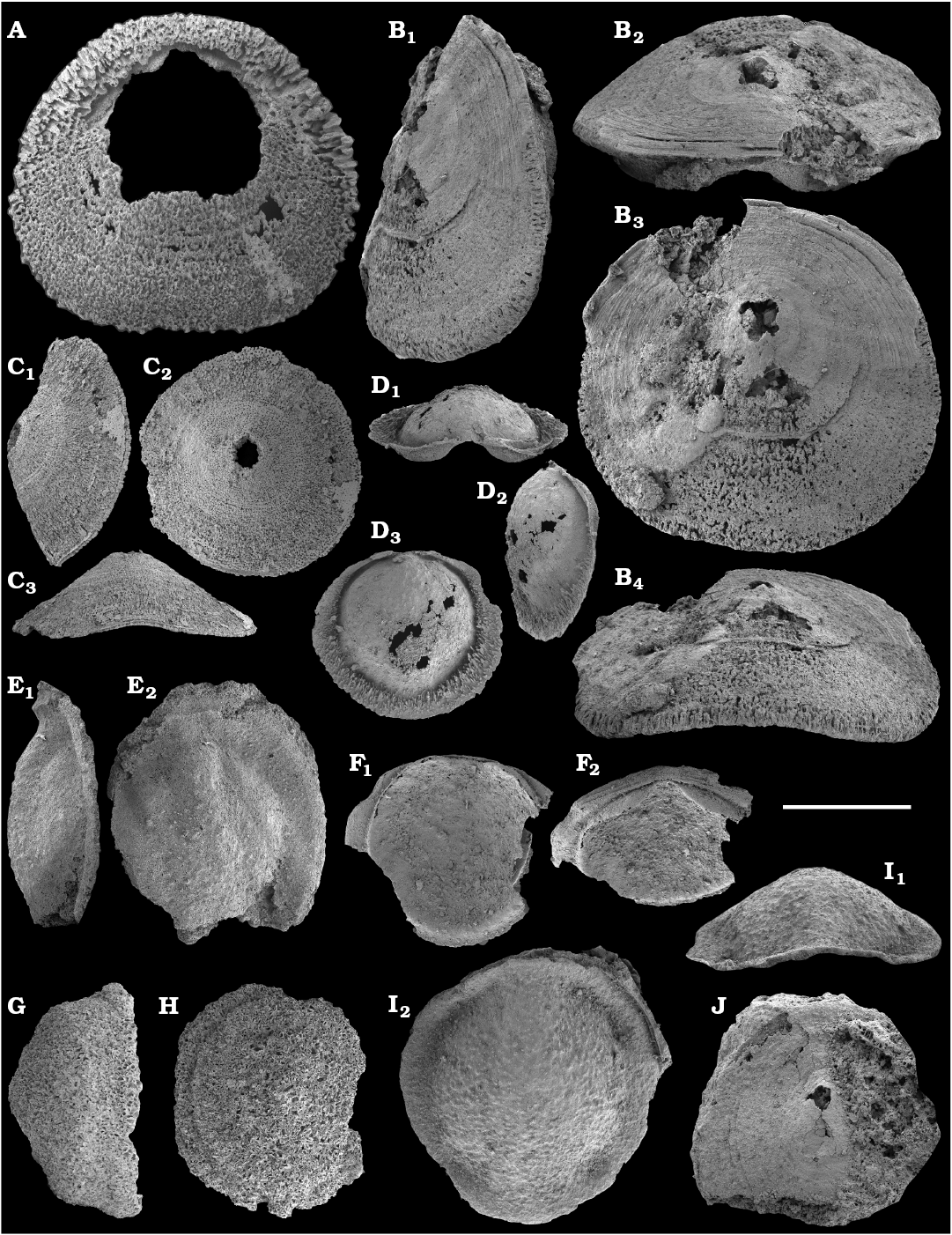
Fig. 55. Opercula of orthothecid hyoliths type C (A, B, D, F, J) and type D (G, H), and undetermined (C, E, I), from early Cambrian Medvezhya (B–D, F, G) and Emyaksin (A, E, H) formations, Anabar Uplift, Siberia, Russia; samples K2/22 (G, H), section 3; 5/10 (F), 5/10B (J), section 96-5; 5a/1.2 (I), 5a/34.75 (A), section 96-5a; M314/12 (B–D, F), Fomitch River section. A, B, D, F, J. Partly phosphatized of opercula, SMNH Mo160464 (A; see also Kouchinsky 2000b: fig. 7A), SMNH X5880 (B), SMNH X5882 (D), SMNH X5883 (F), SMNH X5886 (J). D and F represent calcium phosphatic coatings of inner surface of opercula viewed from external (D) and visceral (F) surfaces; B1, D2, lateral; A, B3, D3, J, apical views; views of dorsal (B2, D1) and ventral (B4) margins; F1, view of inner surface of operculum; F2, oblique view of dorsal margin. C. Partly phosphatized operculum, SMNH Mo160469 (see also Kouchinsky 2000b: fig. 7M). C1, lateral; C2, apical views; C3, view of presumably ventral margin. E, I. Calcium phosphatic internal moulds of opercula, SMNH X5884 (E), SMNH X5887 (I). E2, I2, apical; E1, lateral views; I1, view on the lower (presumably ventral) margin, as in I2. G, H. Dolomitized internal mould of opercula, SMNH X5885 (G), SMNH X5881 (H). G, lateral; H, apical views. Scale bar 500 µm (A–F, I, J), 1 mm (G, H).
Phylum Chaetognatha? Leuckart, 1854
Class, order, and family uncertain
Genus Protohertzina Missarzhevsky, 1973
Type species: Protohertzina anabarica Missarzhevsky, 1973; Anabarites trisulcatus Zone, lower Cambrian (lower Fortunian Stage); mouth of the Kotujkan River, Siberia, Russia.
Remarks.—A longitudinal orientation of phosphatic fibres has been used to assign Protohertzina to the order Pseudoconodontida, whereas the order Protoconodontida was suggested for sclerites composed of basally accreted layers of microcrystalline calcium phosphate (Landing 1995).
Protohertzina anabarica Missarzhevsky, 1973
Figs. 56–58.
Material.—Thirty-five phosphatic sclerites from the topotype locality M419 (section 2 herein), at the mouth of the Kotujkan River, including SMNH X5888–5906, from samples M419/12, K1b/8, K1b/13, 2/4, and 2/7 (section 2, Fig. 1), lower Nemakit-Daldyn Formation, Anabarites trisulcatus Zone; and three specimens, SMNH X5907–5909, from sample 96-4/4 (section 96-4, Fig. 2), middle Manykay Formation, Angustiochrea lata Zone (Val’kov 1975), Anabar Uplift, Siberian Platform, Russia. Correlated with the lower part of the Fortunian Stage.
Description.—Simple, bilaterally symmetrical, spine-shaped apatitic sclerites, up to 3 mm long, gently curved in the sagittal plane (fragments without basal part) but up to 90° curvature in base–tip transition (in more complete, longer specimens), cross-section continuously expands from the tip towards the flared basal part. Internal cavity extends into the tip. Convex side of sclerite is rounded, smooth, with single, shallow, basal furrow (Figs. 56C2, 57G1). Opposite, longitudinally concave side of sclerite with well-defined median ridge bordered by two usually distinct lateral ridges. Cross-section, with more or less pronounced ridges, varies from broad to narrow depending on growth stage. Ridges become more prominent gradually towards the broader basal part of sclerite, apex (tip), where the cross-section is somewhat elongated in the sagittal plane. Wall composed of multiple layers of fibres longitudinally oriented on outer surface of sclerites (Figs. 56E, 57F1).
Remarks.—According to Missarzhevsky (1973: 54), the holotype came from sample M41/12 from the Nemakit-Daldynian Stage in the western Anabar Uplift. However, this number is a typographic error, because section M41 is located on the middle Lena River (Churan) and its basal beds belong to the Dokidocyathus regularis Zone of the middle Tommotian Stage (Rozanov et al. 1969). The correct sample number is thus M419/12 (section 2 herein, see Fig. 1, SOM 1). The 1.2 mm-long holotype illustrated by Missarzhevsky (1973: pl. 9: 1, 2) is now lost and probably represented a fragment with a broken-off basal part. Missarzhevsky (1973) mentioned the variable morphology of Protohertzina anabarica in the first description of the species. The morphological variations of P. anabarica sclerites from the topotype material illustrated in this study (Figs. 56 and 57; samples M419/12, K1b/8, K1b/13, and 2/7) support these earlier observations and are assigned to that same species.
In the topotype material, more compressed forms co-occur with more rounded ones as described by Missarzhevsky (1973: fig. 3). The flared basal part is often not preserved and thus only slender spinose distal fragments remain. Those with broad basal parts are similar to P. robusta Qian, 1977, and apparently, Hertzina pensa Khomentovsky, Didenko, and Pyatiletov, 1982, which was described from the uppermost Nemakit-Daldyn Formation (Koril Member). Protohertzina anabarica internal moulds also occur in the middle Manykay Formation of the eastern Anabar Uplift (Fig. 58). The slender forms with a less robust appearance and narrower rounded basal part (Figs. 56A–E, H, 57A–G, K, 58B) are similar to P. unguliformis Missarzhevsky, 1973, first described in the same publication by Vladimir V. Missarzhevsky from the basal Pestrotsvet Formation in the Yudoma-Maya region, southeastern Siberian Platform, and subsequently reported worldwide (P. unguliformis-type sclerites) often in association with P. anabarica (or P. anabarica-type sclerites). According to the original descriptions by Missarzhevsky (1973: 55), P. anabarica differs from P. unguliformis in having a more isometrical cross-section, a less strongly defined median keel, and distinctly expressed lateral ribs. More or less slender topotype specimens are however treated herein under P. anabarica, given the morphological variability of P. anabarica sclerites noted in their original description, and a significantly younger age of the topotype material of P. unguliformis from the southeastern Siberian Platform which needs further study. The wall of P. anabarica elements is composed of very elongate, needle-like crystallites with a longitudinal orientation (e.g., Fig. 57F1, I1, I2).
Stratigraphic and geographic range.—Terreneuvian Series; Siberia, Laurentia (Conway Morris and Fritz 1980; Nowlan et al. 1985; McIlroy and Szaniawski 2000; Pyle et al. 2006), Kazakh Terrains, western Mongolia, Gondwana (South China, Iran, India; Missarzhevsky 1973; Qian and Bengtson 1989; Brasier and Singh 1987), and Avalonia (Landing et al. 1989). See also Parkhaev and Demidenko (2010) and Kouchinsky et al. (2012: online appendix 1).
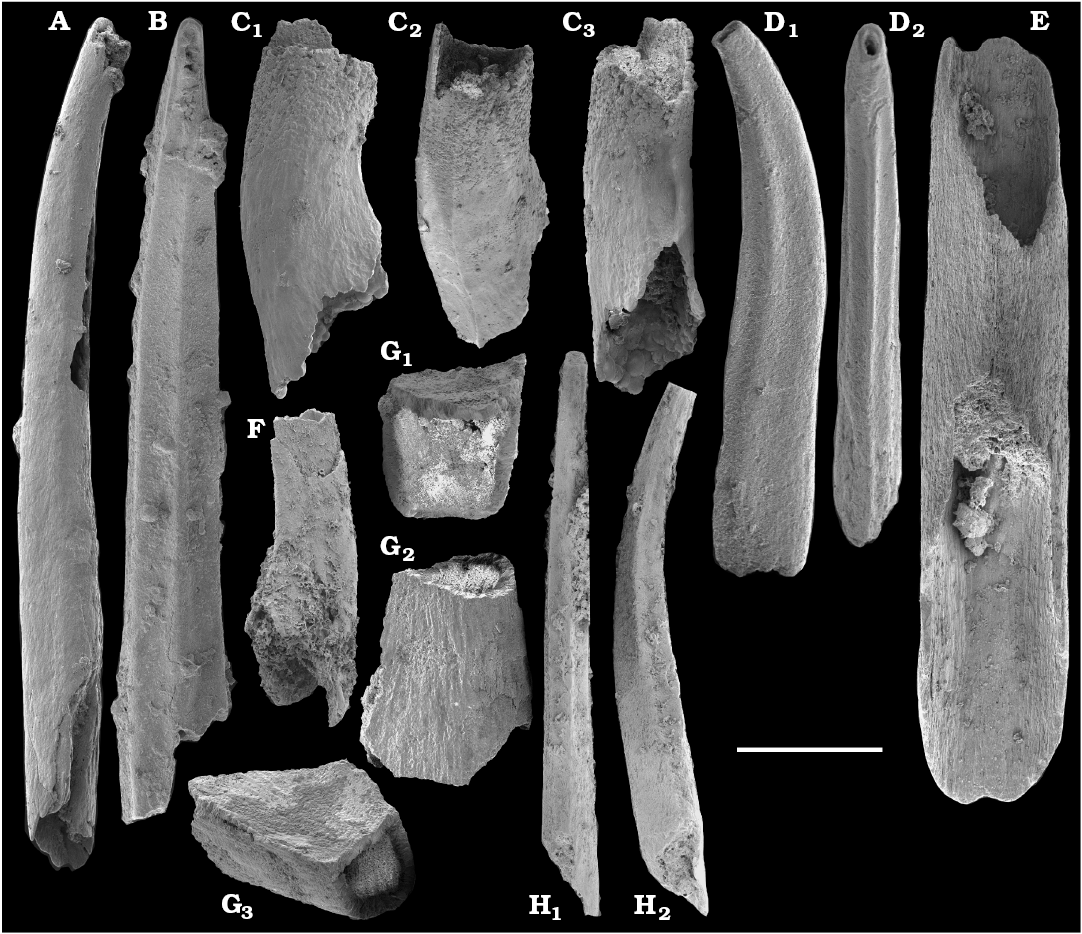
Fig. 56. Protoconodonts Protohertzina anabarica Missarzhevsky, 1973. P. anabarica-type sclerites (F, G) and P. unguliformis-type sclerites (A–E, H), from early Cambrian Nemakit-Daldyn Formation, western flank of the Anabar Uplift, Siberia, Russia; samples M419/12 (A–C, F, G) and K1b/13 (D, E, H), section 2. A–C. SMNH X5888–5890, respectively. D, E. SMNH X5894, 5895, respectively. F–H. SMNH X5891–5893, respectively. Scale bar 250 µm (A–C, E, G), 500 µm (D, F, H).
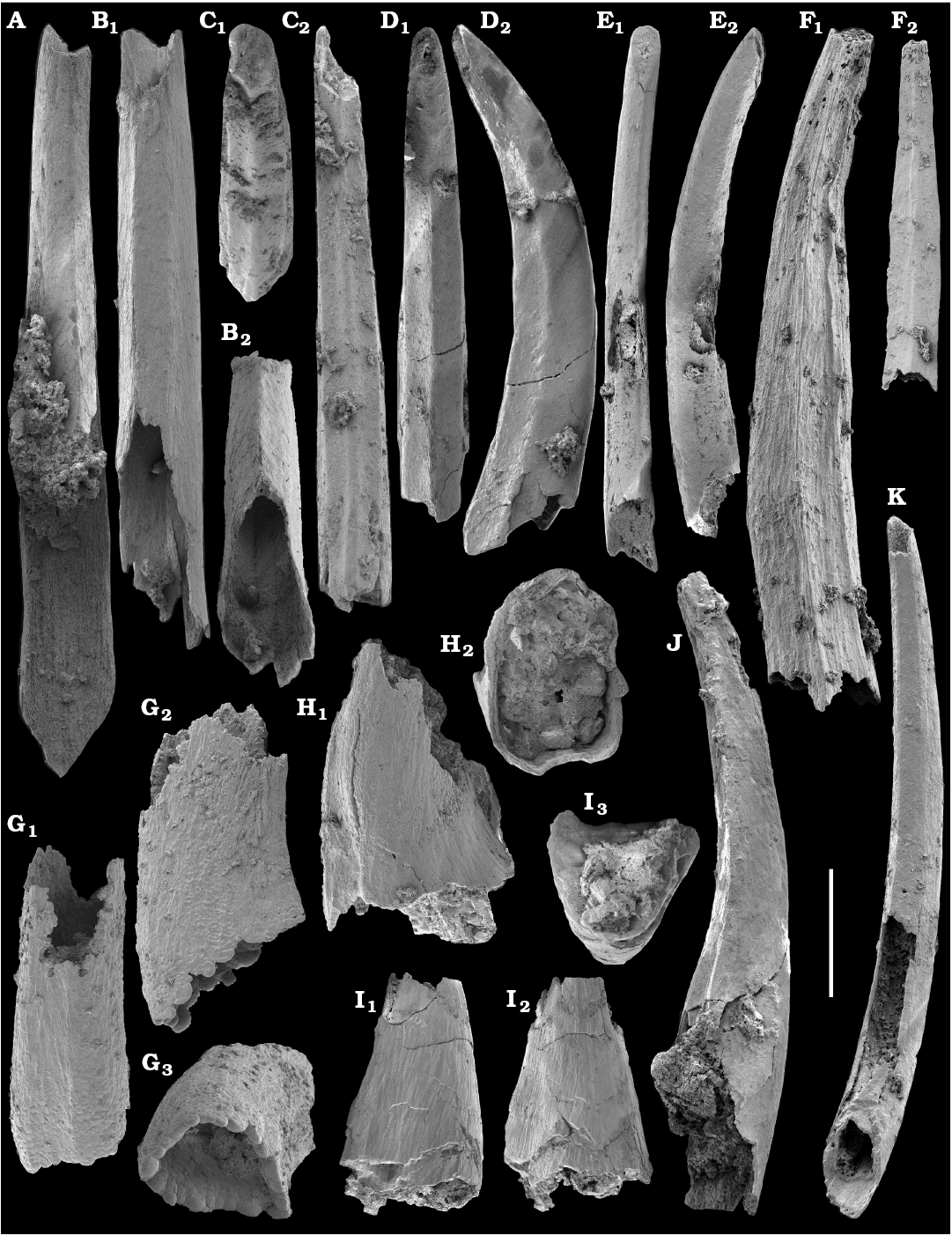
Fig. 57. Protoconodonts Protohertzina anabarica Missarzhevsky, 1973. P. anabarica-type sclerites (H–J) and P. unguliformis-type sclerites (A–G, K), from early Cambrian Nemakit-Daldyn Formation, western flank of the Anabar Uplift, Siberia, Russia; sample 2/7, section 2. A–K. SMNH X5896–5906, respectively. Note longitudinal fibres exposed on the lateral side (F1) of sclerite. Scale bar 250 µm (A–D, F1, G, H, J), 500 µm (E, F2, I, K).
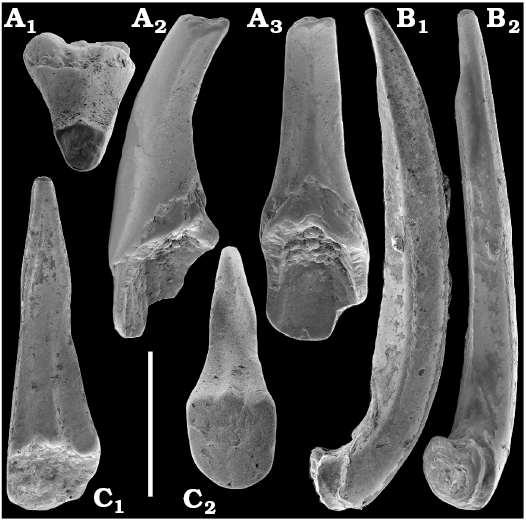
Fig. 58. Protoconodonts Protohertzina anabarica Missarzhevsky, 1973. P. anabarica-type sclerites (A, C) and P. unguliformis-type sclerites (B), from early Cambrian Manykay Formation, eastern flank of the Anabar Uplift, Siberia, Russia; sample 4/4, section 96-4. A, C. SMNH X5907, 5908, respectively. B. SMNH X5909. Scale bar 500 µm.
Phylum, class, order, and family uncertain
Genus Fomitchella Missarzhevsky in Rozanov et al., 1969
Type species: Fomitchella infundibuliformis Missarzhevsky in Rozanov et al., 1969; lower Cambrian (upper Fortunian–lower part of Cambrian Stage 2); 6 km upstream of the Afanas’ev Lakes, middle reaches of the Fomitch River, Siberia, Russia.
Included species: In addition to the type species from the northern flank of the Anabar Uplift, from sample M314/4, collected by Vladimir V. Missarzhevsky from red-orange limestone 9‒10 m above the base of the Medvezhya Formation (Rozanov et al. 1969), other species were formally described as Fomitchella acinaciformis Missarzhevsky, 1977; F. acutata Bokova in Bokova and Vasil’eva, 1990; F. explicata Bokova and Val’kov in Bokova and Vasil’eva, 1990; F. inchoata Yang and He, 1984; F. oblata Bokova in Bokova and Vasil’eva, 1990; F. rugosus Jiang in Luo et al., 1982; and F. yankonensis Yuan and Zhang, 1983. F. grandilica Bokova, 1990 and F. ordinata Bokova, 1990 are informally named by Bokova (1992).
Remarks.—The oldest appearances of Fomitchella from the Fortunian Stage of the Siberian Platform are poorly known. Those include Fomitchella cf. infundibuliformis from 25 m below the top of the Ust’-Yudoma Formation from the “Mount 1291 m” section on the Gonam River, southeastern Siberian Platform (reported without illustration in Khomentovsky et al. 1983 and Semikhatov and Serebryakov 1983, but not reported in Khomentovsky and Karlova 1993) and Fomitchella sp. (4 fragments) from the basal Koril Member of the upper Nemakit-Daldyn Formation at the Ary-Mas-Yuryakh section on the Kotuj River (see Khomentovsky et al. 1982: 18, pl. 1: 5). In our material, the oldest F. infundibuliformis occurs in the lower (but not the basal) parts of the Medvezhya and Emyaksin formations (SOM 1), which can be assigned to the upper Purella Zone and correlated with the upper Fortunian Stage.
Fomitchella aff. F. infundibuliformis was reported from the Karatau Member of the Chulaktau Formation at the Aktugaj section in the Lesser Karatau Range. This occurrence is in the Pseudorthotheca costata Zone, which has been correlated by Missarzhevsky and Mambetov (1981) and Missarzhevsky (1989) with younger strata of the Tommotian Stage.
Fomitchella is shown by Li et al. (2007) to range from the middle Meishucunian to the Qiongzhusian stages. Three species of Fomitchella, F. rugosus Jiang in Luo et al., 1982; F. yankonensis Yuan and Zhang, 1983; and F. inchoata Yang and He, 1984, were described from South China, but the validity of these species and their affinity to Fomitchella Missarzhevsky in Rozanov et al., 1969 were questioned by Bengtson (1983) and Qian and Bengtson (1989). Variable helmet- or trumpet-shaped sclerites occasionally merged at bases were synonymized under Kaiyangites novilis Qian and Yin, 1984a (Steiner et al. 2003, 2007) and compared to Fomitchella by Steiner et al. (2003). They occur in the Assemblage Zone 1 of South China. The wall of K. novilis is probably organic or calcareous, unlike Fomitchella. Alternatively, the fossil was attributed to the cambroclaves (Yao et al. 2005).
Trumpet-shaped elements are also known from the Chert-Phosphate Member, Lower Tal Formation at Madleota in the Lesser Himalayas, which is correlated with Assemblage Zone 1 of South China (Brasier and Singh 1987; Brasier 1989). These elements were compared with Fomitchella, but were found to be morphologically different (Brasier and Singh 1987). They may represent fragments of Mongolodus spp. (Maldeotaia Singh and Shukla, 1981, was synonymized by Azmi [1996: 458] and McIlroy and Szaniawski [2000] with Mongolodus Missarzhevsky, 1977).
Fomitchella indundibuliformis was reported to range from the middle Watsonella crosbyi Zone (i.e., lower part of Cambrian Stage 2) into the lower Camenella baltica Zone (Terreneuvian‒Cambrian Series 2 boundary interval) in Avalonia (Landing and Brett 1982; Bengtson and Fletcher 1983; Landing 1988, 1995; Landing et al. 1989).
Remarks.—Fomitchella elements are conical, hollow phosphatic sclerites with a multilayered wall (see Bengtson 1983). Despite their overall conodont-like shape, the affinity of Fomitchella with conodonts was questioned by Bengtson (1983), who suggested that their overall similarity to euconodonts is “convergent or fortuitous” (Bengtson 1983: 12). The wall in Fomitchella is up to 15 µm thick and consists of thin (0.5–2 µm) lamellae that are subparallel to the surface of the sclerites and are interpreted as growth increments. These growth increments are apparently expressed on the exterior of the sclerites by transverse, weakly defined terraces that extend to the tip of some of the sclerites (e.g., Figs. 59B1, 61G2). The apical part exhibits a concentric lamination and its narrow tip is filled with mineralized material (Fig. 65C4). The lamellae of the wall are finely granular. The innermost lamellae are restricted to the apical part. Hence, according to Bengtson (1983), the sclerites grew by addition of lamellae to the outside of the sclerite. The inner and outer surfaces are generally smooth and without noticeable growth lines. Bengtson (1983: 10) also mentioned a fine radial ornamentation on the outer surface of sclerites. Consequently, Fomitchella was treated as a conodont-like problematic fossil (e.g., Landing 1988).
The outermost layer is thin (several µm thick), covers the entire outer surface of the sclerite without traces of growth increments, and is built of radially oriented ca. 0.1 µm fibres (Figs. 60A5, B2, 65C3) grouped in radially extended bundles (several µm thick). The fibre bundles terminate at the external side of the apertural margin with indentations/depressions that are several µm wide and deep (Fig. 60A5). Such fibrosity is not observed on the inner surface of Fomitchella cones nor within the lamellar wall. This histology composed of radial fibres precludes assignment of Fomitchella to the order Pseudoconodontida as proposed by Landing (1995).
In some specimens (Figs. 61F3, 62E2, G2), a circumferential wrinkled zone is exposed distally to the margin of the fibrous layer. The wrinkles are 5–10 µm wide in radial direction of sclerite and extend subparallel to the margin. They are separated from each other by irregular sutures. On the external side of the sclerite margin, the proximal wrinkles tend to overlap the distal wrinkles. On the inner side of the margin, the distal wrinkles overlap the more proximal wrinkles (Fig. 61C3). Internal moulds bear casts of the sutures between wrinkles that are subparallel to the margin of sclerite, as well as casts of scattered tubercules (1–10 µm diameter; Fig. 61G3). The tubercules represent pits in the inner surface of the wall that are also observed in the margin of the wall (Fig. 59B2) and have been reported from cross-sections (Bengtson 1983). Bengtson (1983: 10) noted fine (ca. 1.0 µm diameter) canals that penetrate through the lamellae and suggested that the canals may either be original features of the wall or were produced by boring organisms. Except for the marginal zone with wrinkles, no growth increments are observed on the inner surface of the sclerite, which is generally smooth but can be irregularly folded. Growth increments of the inner layer are exposed on the outer surface of some specimens from samples 5a/1 and 5a/1.2 (Fig. 64F, G), presumably with a partially or completely exfoliated outer layer.
Thus, the inner layer (up to 10 µm thick) is probably composed of growth increments expressed as wrinkles that were added to the apertural margin circumferentially by basal marginal accretion. The increments merge with each other and produce a continuous lamination of the wall. The outer layer was probably continuously built by setoblasts at the margin of the sclerite. Both layers were readily phosphatized during diagenesis. The original mineralization of the sclerites was probably weak as evidenced by elastic deformation of some sclerites during early sediment compaction (Fig. 63F). On the other hand, brittle deformation and cracks occur (Figs. 59E, 63F) and indicate a more thorough degree of mineralization. Given the mode of secretion of the inner layer by marginal accretion, it is similar to that observed in tommotiids (Landing 1984; Kouchinsky et al. 2015a: fig. 41), although fibrosity and setae are unknown in morphologically similar mitral sclerites of Camenella-like or in Lapworthella-like tommotiids (Landing 1984; Skovsted et al. 2009; Kouchinsky et al. 2010).
Fomitchella infundibuliformis Missarzhevsky in Rozanov et al., 1969
Figs. 59–64.
Material.—Seven topotype specimens, SMNH X5910–5916, from sample M314/4 and over 200 sclerites from samples 1/39.3, 1/29.5, 1/29.2, 1/28.5, 1/27.5, 1/26.3, 1/25.5 (section 1, Fig. 4), K1a/80 and K1a/82 (section 2, Fig. 1), K2/21, K2/22, K2/23, K2/26, K2/33, K2/38A, K2/43, K2/45, K2/50, K2/52, K2/60?, 3/12.2, 3/20.5, 3/22.5 (section 3, Fig. 3), 5/10, 5/10B, 5a/1, 5a/1.2, 5a/5, 5a/6, 5a/8, 5a/9 (sections 96-5, 96-5a, Fig. 2), including figured SMNH X5917–5926, 5928–5931, 5933–5935, 5937–5941, from other samples (see SOM 1) from the Medvezhya and lower Emyaksin formations, Anabar Uplift, Siberian Platform, Russia. Correlated with the upper Fortunian and lower part of the Cambrian Stage 2.
Description.—Phosphatic, conical, hollow sclerites with elongate, usually recurved tip and flared base. Basal margin irregularly rounded in planar view and undulating, often with circumferential wrinkles. Outer surface with radial striation, smooth or occasionally bumpy. Inner surface smooth, often with scattered pits and closely spaced wrinkles at the basal margin. Wall with thin growth lamellae, of which the innermost ones are restricted to the apical part.
Remarks.—Topotype specimens of F. infundibuliformis from sample M314/4 (Figs. 59, 60) have a broad rounded aperture with the two opposing sides of the margin arched. One of the specimens is compacted perpendicular to the plane of its broken aperture (Fig. 59E). A certain similarity exists between the illustrated topotype material and the elements from southeastern Newfoundland and eastern Massachusetts (Landing 1988; Landing et al. 1989). The Avalonian material has a subcircular outline of the aperture and includes specimens ranging from gently to rapidly tapering, are more or less recurved, and have acicular longer or shorter tips (“cusps” sensu Landing 1988).
The species differs from Fomitchella acinaciformis Missarzhevsky, 1977, in having a flared base, higher rate of expansion, and relatively broad circumferential wrinkled margin. Fomitchella aff. infundibuliformis has a wider apex (Fig. 65); Fomitchella aff. acinaciformis (Fig. 66A–E) has narrower, less flared, and more elongate sclerites. Fomitchella sp. (Fig. 66H) sclerites are much smaller than the other specimens. A comparison with other, earlier described species is hampered by their inadequate illustration.
Stratigraphic and geographic range.—Upper Fortunian–Cambrian Stage 2 of the Siberian Platform and probably Kazakhstan (Missarzhevsky and Mambetov 1981), Upper Fortunian–lower part of Cambrian Stage 3 of Avalonia (Landing and Brett 1982; Bengtson and Fletcher 1983; Landing 1988, 1995; Landing et al. 1989); doubtful reports from the Terreneuvian Series of western Mongolia (Missarzhevsky 1977) and South China (Qian and Bengtson 1989).

Fig. 59. Problematic calcium phosphatic sclerites Fomitchella infundibuliformis Missarzhevsky in Rozanov et al., 1969, from early Cambrian Medvezhya Formation, northern flank of the Anabar Uplift, Siberia, Russia; topotypes from sample M314/4, Fomitch River section. A. SMNH X5910. B. SMNH X5912. C. SMNH X5911. D. SMNH X5913. E. SMNH X5914. A1, B3, C2, D1, E1, lateral; A2, D2, apertural; B1, C1, E2, apical views; B2, close-up of inner surface showing comarginal folds. Scale bar 100 µm (B2), 250 µm (A, B1, B3, C–E).

Fig. 60. Problematic calcium phosphatic sclerites Fomitchella infundibuliformis Missarzhevsky in Rozanov et al., 1969, from early Cambrian Medvezhya Formation, northern flank of the Anabar Uplift, Siberia, Russia; topotypes from sample M314/4, Fomitch River section. A, B. SMNH X5915, 5916, respectively. A2, A4, B1, lateral views; A1, B3, apertural views; A3, A5, B2, enlargements of the apertural margin showing fibrous composition of outer layer. Scale bar 12.5 µm (B2), 25 µm (A5), 100 µm (A3), 250 µm (A1, A2, A4, B1, B3).
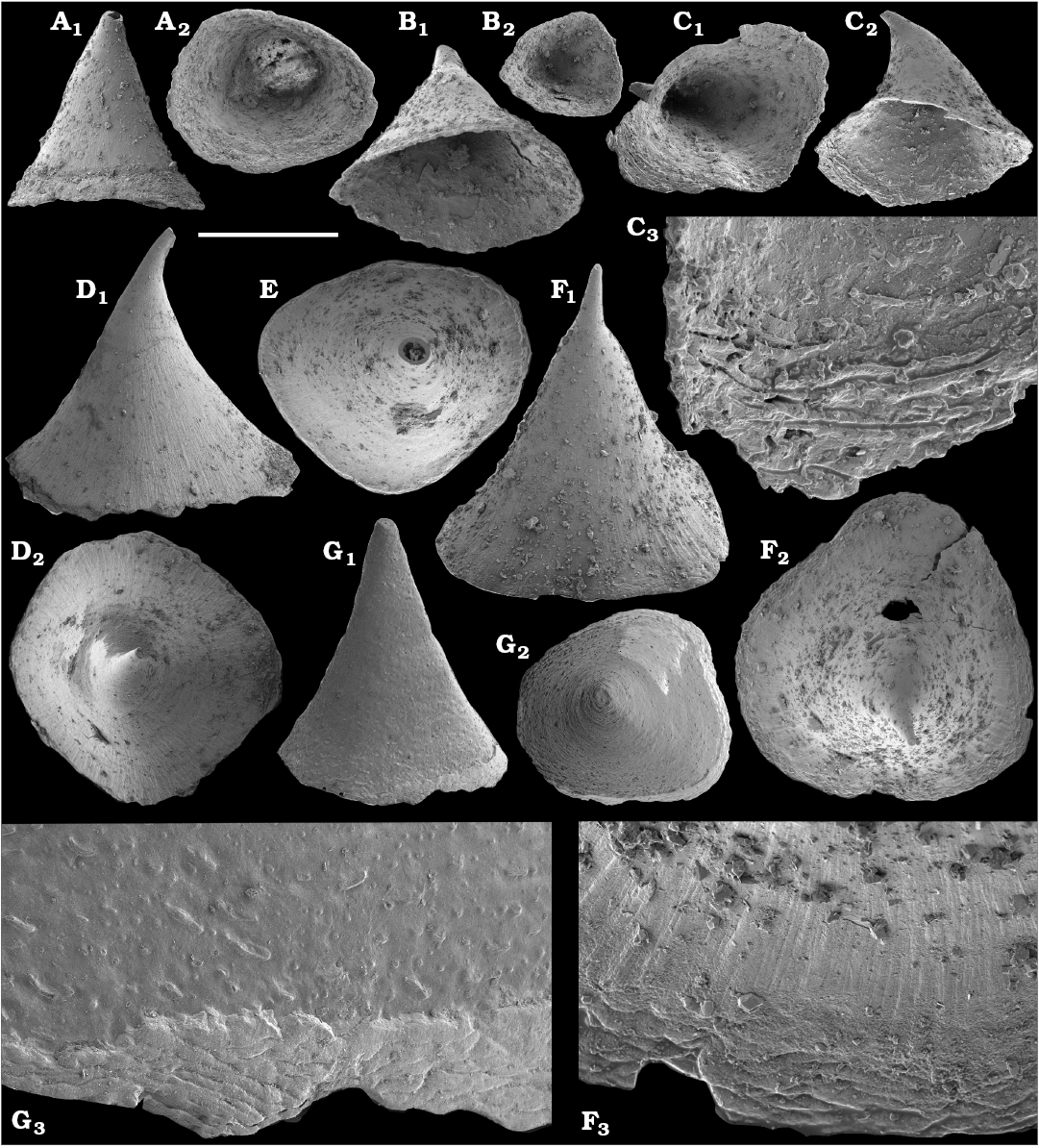
Fig. 61. Problematic calcium phosphatic sclerites Fomitchella infundibuliformis Missarzhevsky in Rozanov et al., 1969, from early Cambrian Medvezhya Formation, western flank of the Anabar Uplift, Siberia, Russia; samples K2/26 (A–C, E, F), K2/21 (D), 3/12.2 (G), section 3. A–G. SMNH X5917–5923, respectively. A1, D1, F1, G1, lateral; B1, C2, oblique lateral; A2, B2, C1, apertural; D2, E, F2, G2, apical views; C3, enlargement of C2 shows comarginally folded inner apertural margin; G3, enlargement of outer surface with comarginal folds and tuberculate outer surface of inner layer; F3, outer surface of sclerite margin with comarginal folds and fibrous outer layer. Scale bar 50 µm (C3, F3), 100 µm (G3), 250 µm (A, B1, C1, C2, D, E, F1, F2), 500 µm (B2, G1, G2).

Fig. 62. Problematic calcium phosphatic sclerites Fomitchella infundibuliformis Missarzhevsky in Rozanov et al., 1969 (A–C, E–G) and Fomitchella sp. (D), from early Cambrian Medvezhya Formation, western flank of the Anabar Uplift, Siberia, Russia; samples K2/33 (A, E–G), section 3; 1/39.3 (B), section 1; and K1a/82 (C, D), section 2. A–G. SMNH X5924–5930, respectively. A1, B1, C, D, E1, F, G1, lateral; A2, apertural; B2, apical views; E2, G2, enlargements show marginal folds in the wall and fibres of the outer layer. Scale bar 50 µm (G2), 100 µm (E2), 250 µm (A, B1, F, G1), 500 µm (B2, C, D, E1).
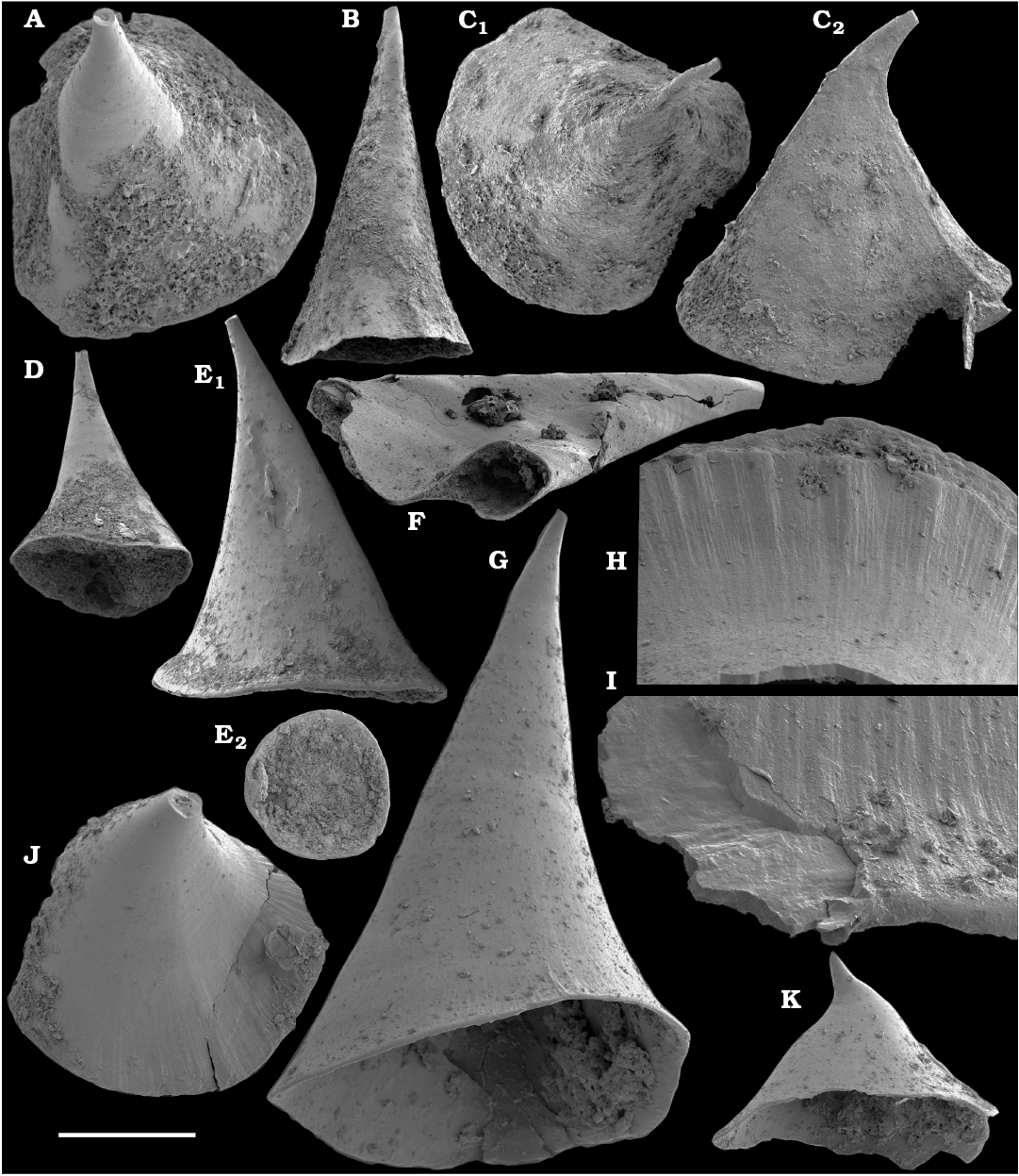
Fig. 63. Problematic calcium phosphatic sclerites Fomitchella infundibuliformis Missarzhevsky in Rozanov et al., 1969 (A, C–E, G–K) and Fomitchella sp. (B, F), from early Cambrian Medvezhya Formation, western flank of the Anabar Uplift, Siberia, Russia; samples 3/22.5 (F), K2/45 (G–K), and K2/50 (A, D), section 3; 1/29.2 (B, C), section 1; and K1a/73 (E), section 2. A–K. SMNH X5931–5941, respectively. A, C1, J, apical; B, C2, D, E1, F, G, K, lateral; E2, apertural views; H, I, close-ups of margin with fibres in outer layer. Scale bar 50 µm (I), 100 µm (H), 250 µm (A, C, E1, G, J, K), 500 µm (B, D, E2, F).
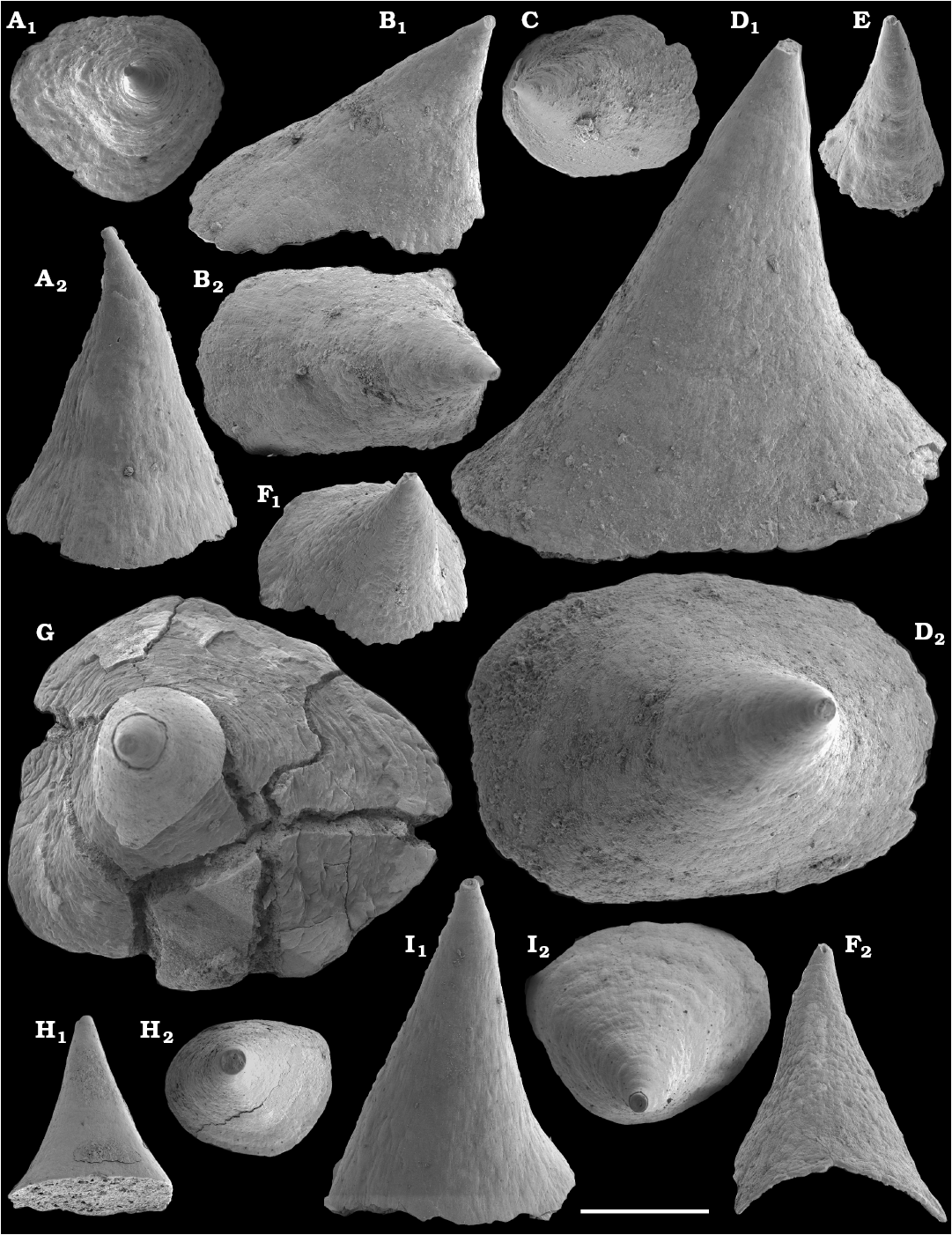
Fig. 64. Problematic calcium phosphatic sclerites (with outer fibrous layer eroded) Fomitchella infundibuliformis? Missarzhevsky in Rozanov et al., 1969, from early Cambrian Emyaksin Formation, eastern flank of the Anabar Uplift, Siberia, Russia; samples 5a/1 (A–F), 5a/1.2 (G–I), section 96-5a. A–I. SMNH X5942–5950, respectively. A1, B2, C, D2, F1, G, H2, I2, apical; A2, B1, D1, E, F2, H1, I1, lateral views. Scale bar 250 µm (A–G, I), 500 µm (H).
Fomitchella aff. infundibuliformis Missarzhevsky in Rozanov et al., 1969
Fig. 65.
Material.—Four phosphatic sclerites, including figured SMNH X5951–5953, from samples 5a/10.5 and 5a/15 (section 96-5a, Fig. 2), Emyaksin Formation, eastern Anabar Uplift, Siberian Platform, Russia. Correlated with lower part of Cambrian Stage 2.
Description.—Calcium-phosphatic, conical, hollow sclerites with flared and slightly undulating basal margin, irregularly rounded in apical view and wide, laterally compressed tip recurved in a single plane, with oval cross-section. Outer surface with radial and fine transverse, concentric striation. Wall shows thin growth lamellae.
Remarks.—See F. infundibuliformis for comparison.

Fig. 65. Problematic calcium phosphatic sclerites Fomitchella aff. infundibuliformis from early Cambrian Emyaksin Formation, eastern flank of the Anabar Uplift, Siberia, Russia; samples 5a/10.5 (A, B) and 5a/15 (C), section 96-5a. A–C. SMNH X5951–5953, respectively. A1, A3, B1, B3, C2, lateral; A2, B2, C1, apical views; C3, fibres in outer layer enlarged; C4, layered wall at the apex, enlarged. Scale bar 12.5 µm (C3), 25 µm (C4), and 250 µm (A, B, C1, C2).
Fomitchella acinaciformis Missarzhevsky, 1977
Figs. 66F, G, 67.
Material.—About 50 phosphatic sclerites, including figured SMNH X5959, 5960, 5962–5968, from samples 5a /17.5 and 5a/18.5 (section 96-5a, Fig. 2), Emyaksin Formation, eastern Anabar Uplift, Siberian Platform, Russia. Correlated with the lower part of Cambrian Stage 2.
Description.—Phosphatic, elongated, conical, hollow sclerites, gently curved and slightly compressed in a single plane. Apex somewhat more elongate, almost straight. Basal margin ovoid, lateral sides may have gentle longitudinal depression. Outer surface with radial striation or smooth. Inner surface smooth.
Remarks.—See Fomitchella infundibuliformis for comparison. The holotype of F. acinaciformis came from sample M314/5-10, a mixed sample collected by Vladimir V. Missarzhevsky from the lower Medvezhya Formation at a horizon located 5‒10 m above the base of section M314 (Rozanov et al. 1969: 44, fig. 9). The type material of F. infundibuliformis (see above) was collected ca. 4 m above the base of section M314.
Two sclerites illustrated as Maldeotaia bandalica Singh and Shukla, 1981, from the Watsonella crosbyi Zone of Avalonian southeast Newfoundland (Landing et al. 1989: fig. 7.5, 7.6) are morphologically comparable to Fomitchella acinaciformis and are brought to the latter species. This lowest occurrence in the Fortunian‒Cambrian Stage 2 transition interval is comparable to the species’ lowest known appearance in Siberia.
Stratigraphic and geographic range.—Terminal Fortunian‒Cambrian Stage 2 of the Siberian Platform and, probably, western Mongolia (see Missarzhevsky 1977) and Avalonia (Landing et al. 1989, discussed above).
Fomitchella aff. acinaciformis Missarzhevsky, 1977
Fig. 66A–E.
Material.—About 50 phosphatic sclerites, including figured SMNH X5954–5958, from samples 5a/9, 5a/10.5, and 5a/17.5 (section 96-5a, Fig. 2), from the lower Emyaksin Formation, eastern Anabar Uplift, Siberian Platform, Russia. Correlated with the lower part of Cambrian Stage 2.
Description.—Relatively tall conical phosphatic sclerites with somewhat flared apertural margin, rounded in apical view. The middle part of the sclerite is oval in cross-section and tapers into a gently recurved narrow tip with rounded cross-section. Outer surface with fine transverse concentric striation and with radial striation at the apertural margin.
Remarks.—This form differs from Fomitchella acinaciformis in having a wider, almost circular aperture and less lateral compression of the upper part of the sclerite.

Fig. 66. Problematic calcium phosphatic sclerites Fomitchella aff. acinaciformis Missarzhevsky, 1977 (A–E), Fomitchella acinaciformis Missarzhevsky, 1977 (F, G), and Fomitchella sp. (H), from early Cambrian Emyaksin Formation, eastern flank of the Anabar Uplift, Siberia, Russia; samples 5a/10.5 (A, B, E), 5a /17.5 (C, F–H), 5a/9 (D), section 96-5a. A–H. SMNH X5954–5961, respectively. A1, oblique apical; A3, C2, D2, H3, apical; A2, B, C3, D1, E1, F, G2, G3, H1, H2, lateral; E2, apertural; G1, oblique apertural views; C1, close-up of apertural margin with fibres (note two inserted sclerites). Scale bar 50 µm (C1), 250 µm (A1, B, C2, C3, E, H), 500 µm (A2, A3, D, F, G).
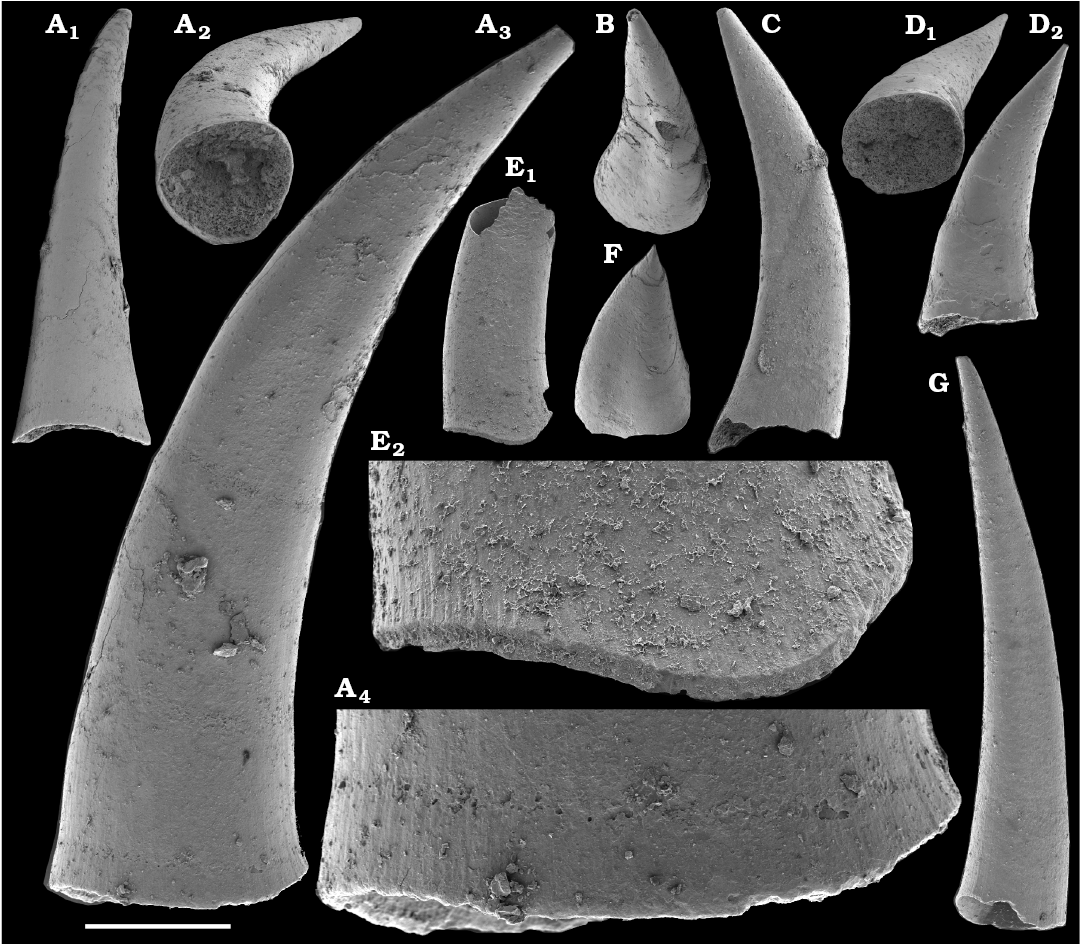
Fig. 67. Problematic calcium phosphatic sclerites Fomitchella acinaciformis Missarzhevsky, 1977, from early Cambrian Emyaksin Formation, eastern flank of the Anabar Uplift, Siberia, Russia; sample 5a/18.5, section 96-5a. A SMNH X5962. B. SMNH X5964. C. SMNH X5966. D. SMNH X5967. E. SMNH X5963. F. SMNH X5965. G. SMNH X5968. A1, A3, C, D2, G, lateral; A2, D1, oblique apertural; B, F, oblique apical views; A4, E2, outer surface with fibres at apertural margin. Scale bar 100 µm (A4, E2), 250 µm (A3), 500 µm (A1, A2, B–D, E1, F, G).
Phylum and class uncertain
Remarks.—The affinity of hyolithelminths is uncertain and usually regarded either within the cnidarians (e.g., Vinn and Kirsimäe 2015) or stem group annelids (e.g., Skovsted and Peel 2011).
Order Hyolithelminthida Fisher, 1962
Family Hyolithellidae Walcott, 1886
Genus Hyolithellus Billings, 1872
Type species: Hyolithes micans Billings, 1872; lower Elliptocephala asaphoides assemblage, Dyeran Stage, lower Cambrian; Troy, New York State, USA.
Hyolithellus vladimirovae Missarzhevsky in Rozanov and Missarzhevsky, 1966
Fig. 68A, C, G.
Material.—Ten fragmentary phosphatic tubes, including SMNH X5969, 5971, 5974, from samples 5a/10.5, 5a/15.5, 5a/17.5, and 5a/34.5 (section 96-5a, Fig. 2), Emyaksin Formation, eastern Anabar Uplift, Siberian Platform, Russia. Correlated with the lower part of Cambrian Stage 2.
Description.—Slowly expanding slightly irregularly curved, phosphatic tubes with circular cross-section, 250–750 µm in diameter. External surface covered with low but distinct straight folds, regularly spaced at 100–200 µm and transverse to the longitudinal axis of the tube.
Remarks.—The holotype derives from the middle course of the Lena River at Churan village from sample M42e of the Pestrotsvet Formation (Dokidocyathus regularis Zone, Tommotian Stage; Rozanov and Missarzhevsky 1966). The fine longitudinal or transverse striation described by Rozanov and Missarzhevsky (1966) from the type material is not observed on the tubes. Hyolithellus grandis Missarzhevsky in Rozanov et al., 1969 only differs in that is several times larger in size. However, it tends to co-occur with H. vladimirovae, which may justify synonomy of the two forms (Sipin 2001). Hyolithellus isiticus Missarzhevsky in Rozanov et al., 1969 mainly differs in having numerous, well-developed flanges in the later growth stages, compared to a smooth initial part and an intermediate part with moderately high ribs. Thus, the middle part of its conch is, according to Missarzhevsky in Rozanov et al. (1969: 150, 151), similar to H. vladimirovae. The narrower smooth tubes described below may represent ontogenetic variants of H. vladimirovae. The external ornamentation of H. insolitus Grigor’eva in Voronin et al., 1982 (see Kouchinsky et al. 2015a) features weaker concentric ribs, faint longitudinal ridges and furrows that undulate between the ribs.
Stratigraphic and geographic range.—Upper Fortunian–Cambrian Stage 2 of Siberia, Laurentia (as Hyolithellus cf. isiticus in Pyle et al. 2006), Kazakhstan (Missarzhevsky and Mambetov 1981), South China (Steiner et al. 2007), and northern India (Brasier and Singh 1987: fig. 8.3–8.5).
Hyolithellus sp.
Fig. 68B, D–F, H, I.
Material.—25 fragmentary phosphatic tubes, including SMNH X5970, 5972, 5973, 5975–5977, from samples K2/6 and K2/60 (section 2, Fig. 1), 1/28.5 (section 1, Fig. 4), 5a/6, 5a/34.5, and 5a/34.75 (section 96-5a, Fig. 2), Medvezhya and Emyaksin formations, Anabar Uplift, Siberian Platform, Russia. Correlated with the upper Fortunian and lower part of Cambrian Stage 2.
Description.—Slightly irregularly curved tubes with circular cross-section, 150–200 µm in diameter. External surface smooth, with faint transverse folds perpendicular to the longitudinal axis of the tube.
Remarks.—The form is distinguished from Hyolithellus vladimirovae by the smaller size of its tubes and subdued transverse rugae. Deformation of some tubes (Fig. 68D) indicates that the original material of the tube was relatively flexible and, thus, phosphatization likely was, in part, secondary. See Kouchinsky et al. (2012: online appendix 1) for the first occurrences of hyolithelminths worldwide.

Fig. 68. Calcium phosphatic tubes of hyolithelminths Hyolithellus vladimirovae Missarzhevsky in Rozanov and Missarzhevsky, 1966 (A, C, G) and Hyolithellus sp. (B, D, E, F, H, I), from early Cambrian Emyaksin (A–D, G, I), Medvezhya (E, F), and Kugda-Yuryakh (H) formations, Anabar Uplift, Siberia, Russia; samples 5a/34.5 (A, B), 5a/15.5 (C), 5a/34.75 (D, I), 5a/10.5 (G), section 96-5a; K2/6 (E, F), K2/60 (H), section 3. A–E. SMNH X5969–5973, respectively. G–I. SMNH X5974–5976, respectively. F. SMNH X5977. Scale bar 500 µm.
Phylum Cnidaria? Hatschek, 1888
Class and order uncertain
Family Anabaritidae Missarzhevsky, 1974
Remarks.—Anabaritids are attributed, in agreement with most other reports (see Kouchinsky et al. 2009), to diploblastic animals, and are probably cnidarians. Given the uncertainties in assignment of the Cambrian cnidariomorphs and the fact that anabaritid tubes exhibit an unusual type of tri-radial symmetry, they are placed among stem-group cnidarians in which a derived and “modern” type of radial symmetry was not yet established. It is also important to consider that an extant type of tissue differentiation present in crown-group cnidarians may not have been achieved in lower–middle Cambrian stem-group cnidarians (Park et al. 2011). The affinities, occurrences and systematics of anabaritids are reviewed by Kouchinsky et al. (2009). Kouchinsky et al. (2009: appendix 1 in the supplementary material) list 72 species (including 10 nomina nuda) and 19 genera (including 2 nomina nuda) of anabaritids that have been formally described. In addition, two formally described species can be added to the list. These include Aculeochrea mesezhnikovi Vasil’eva, 1998, and A. trilamellosa Vasil’eva, 1998 (by original designation; see remarks in the description of Anabarites compositus Missarzhevsky in Rozanov et al., 1969, below). The majority of the anabaritid species were originally reported from the Terreneuvian Series of the Siberian Platform, where they are now also known from the younger Atdabanian Stage (lower part of Cambrian Stage 3) of the eastern Anabar Uplift (see Kouchinsky et al. 2015a). Most of the tri-radial symmetrical anabaritid tubes from our collection can be separated into two groups (genera), those with three prominent longitudinal keels and weakly expressed transverse sculpture (Selindeochrea) and those with variously expressed, transverse sculptural elements (Anabarites).
Genus Selindeochrea Val’kov, 1982
Type species: Selindeochrea tecta Val’kov, 1982; lower Cambrian (lower part of Cambrian Stage 2); upper reaches of the Selinde River, Siberia, Russia.
Remarks.—Anabarites tricarinatus Missarzhevsky in Rozanov et al., 1969 was erroneously indicated as the type species of Selindeochrea by Kouchinsky et al. (2009: 286). The latter genus includes anabaritid species with tri-radial symmetrical conchs with prominent longitudinal keels and weakly expressed transverse sculpture. The genus includes S. tecta Val’kov, 1982; S. tricarinata (Missarzhevsky in Rozanov et al., 1969); S. tripartita (Missarzhevsky in Rozanov et al., 1969); S. missarzhevskyi (Vasil’eva, 1986); and probably S. ternaria (Missarzhevsky in Rozanov et al., 1969).
Selindeochrea tricarinata (Missarzhevsky in Rozanov et al., 1969)
Fig. 69.
Material.—Over 50 specimens, including figured SMNH X3412, 5978, 5980–5982, from samples K2/21, K2/25, K2/50, 3/12, and 3/12.2 (section 3, Fig. 3), Medvezhya Formation, western Anabar Uplift, Siberian Platform, Russia. Correlated with the upper Fortunian and lowermost Cambrian Stage 2.
Description.—Slowly expanding slightly curved tubes with tri-lobate cross-sections, slightly twisted clockwise (from apex to the aperture, in the direction of growth). Grooves (depressions) between extended lobes are wide and shallow with flattened middle part. There is a narrow median ridge on the flattened depressions between the lobes (as replicated by internal moulds). In the flattened depressions, the growth lines are arched and point at an acute angle towards the aperture. In preserved thecae, the lobes are extended distally by prominent keels. The keels undulate, with folds running towards the aperture at ca. 45°. The lobes of the internal moulds are rounded or somewhat swollen and almost straight.
Stratigraphic and geographic range.—Upper Fortunian–lower part of Cambrian Stage 2 of the Siberian Platform.
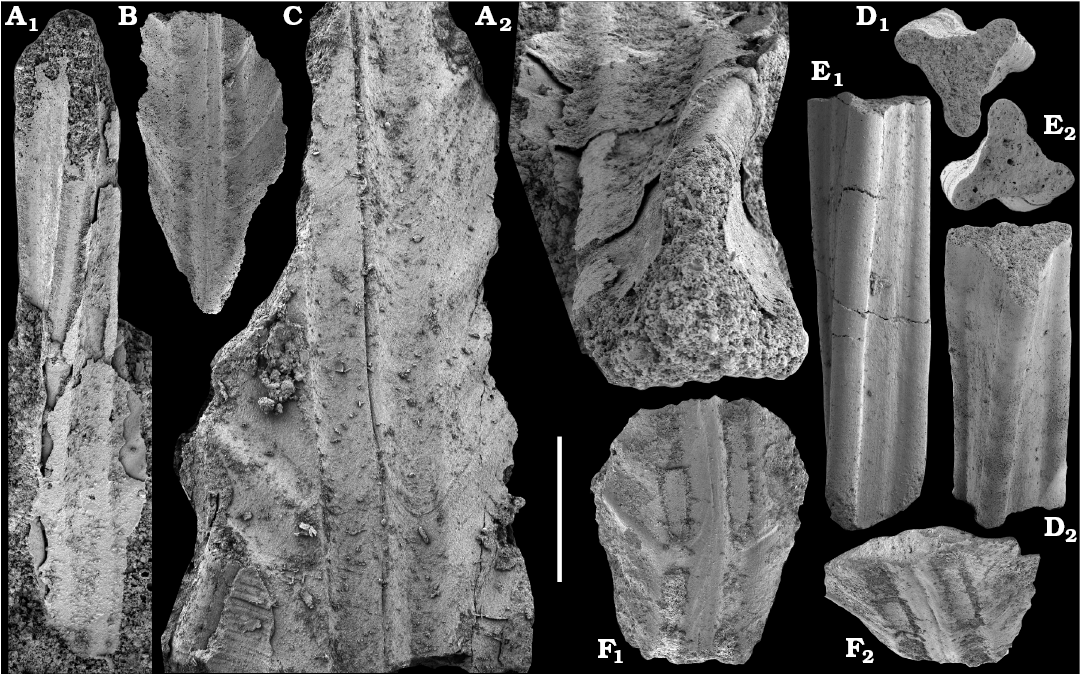
Fig. 69. Anabaritid Selindeochrea tricarinata (Missarzhevsky in Rozanov et al., 1969), from early Cambrian Medvezhya Formation, western Anabar Uplift, Siberia, Russia; samples K2/25 (A, C), 3/12.2 (B, D), and 3/12 (E, F), section 3. A. SMNH X3412-part (see also Kouchinsky et al. 2009: fig. 48C); A1, phosphatic internal mould with external phosphatic coating of longitudinal lobes and their distal extensions, keels; A2, cross-section enlarged. B, F. SMNH X5978, 5982, respectively; phosphatic external moulds with longitudinal keels; B, F1, lateral; F2, oblique views. C. SMNH X3412-counterpart (see also Kouchinsky and Bengtson 2002: fig. 9A and Kouchinsky et al. 2009: fig. 48D), cast of the outer surface. D, E. SMNH X5980 and 5981, respectively; phosphatic internal moulds; D1, E2, transverse; D2, E1, lateral views. Scale bar 250 µm (A2) and 500 µm (A1, B–F).
Selindeochrea cf. tecta Val’kov, 1982
Fig. 70.
Material.—About 20 phosphatic and glauconitised internal moulds, including figured SMNH X3778‒3780, 5986, from samples 3/10 (section 3, Fig. 3), 5a/34.5 and 5a/34.75 (section 96-5a, Fig. 2), Medvezhya and Emyaksin formations. Correlated with the upper Fortunian and lower part of the Cambrian Stage 2.
Description.—Tubes with three prominent longitudinal lobes twisted clockwise (from apex to the aperture, in the direction of growth) through several revolutions. Outer surface covered with undulating growth lines. Lumen (internal mould) with three elongated lobes with distal parts twisted up to 90°. Adapically, the lobes are lower and not twisted (Fig. 70C2). The lobes are separated by V-shaped grooves that are shallow adapically.
Remarks.—Selindeocrea tecta has straight trapezoidal lobes similar to those in Fig. 70D (see Kouchinsky et al. 2009). Unlike S. tecta, the direction of twist as described herein is clockwise in all of the available S. cf. tecta specimens, but this feature does not necessarily have to be an invariant character. Val’kov (1987: 109, pl. 13: 4‒13) described similar forms with curved distal parts of the lobes as observed in cross-sections as Anabarites cf. tricarinatus Missarzhevsky in Rozanov et al., 1969 from the lower Erkeket Formation of the Olenyok Uplift, northern Siberia. Examination of the fossils chemically extracted by Val’kov from sample 846 yielded ca. 20 poorly preserved internal moulds (AK personal observations) that are twisted clockwise in the direction of growth and similar to Selindeochrea cf. tecta.

Fig. 70. Anabaritid Selindeochrea cf. tecta Val’kov, 1982, phosphatic internal moulds, from early Cambrian Emyaksin (A–C) and Medvezhya (D) formations, Anabar Uplift, Siberia, Russia; samples 5a/34.75 (A, C), 5a/34.5 (B), section 96-5a; and 3/10 (D), section 3. A. SMNH X3779 (see also Kouchinsky et al. 2009: fig. 47G). B. SMNH X3778 (see also Kouchinsky et al. 2009: fig. 47C). C. SMNH X3780 (see also Kouchinsky et al. 2009: fig. 47H). D. SMNH X5986. A1, B2, C1, D1, D3, lateral; A2, B1, D2, transverse views; C2, transverse view of apex. Scale bar 500 µm.
Selindeochrea tripartita (Missarzhevsky in Rozanov et al., 1969)
Fig. 71.
Material.—Several hundred internal moulds, including figured SMNH X5988–5991, and one internal with external mould, SMNH X5987, from samples 3/12, 3/12.2, K2/25, K2/26 (section 3, Fig. 3), 5a/6 and 5a/9 (section 96-5a, Fig. 2), Medvezhya and Emyaksin formations. Correlated with the upper Fortunian and lowermost Cambrian Stage 2.
Description.—Slowly expanding, slightly curved tubes with three radially extended lobes separated by deep grooves. Externally, each lobe carries a prominent median longitudinal keel (Fig. 71A). Each keel may exceed the internal diameter of the tube and regularly undulates with folds oriented ca. 45° towards the aperture. The lobes become more prominent aperturally. Along the middle part of each lobe (on internal moulds) there is a narrow depression (Fig. 71B, D, white arrow). Within the grooves, the growth lines are gently curved towards the aperture.
Remarks.—The presence of prominent longitudinal keels is diagnostic for Selindeochrea and distinguishes it from Anabarites.
Stratigraphic and geographic range.—Upper Fortunian Stage and lower part of Cambrian Stage 2; Siberia (Rozanov et al. 1969: 158, pl. 8: 11, 16, 20; Matthews and Missarzhevsky 1975: pl. 2: 3; Val’kov 1982: 75, pl. 12: 1‒6; Missarzhevsky 1989: pl. 13: 8; Khomentovsky and Karlova 1989: 57, pl. 4: 6; Khomentovsky et al. 1990: pl. 11: 4; Pel’man et al. 1990: pl. 3: 8; Kouchinsky et al. 2009: fig. 20), probably western Mongolia (Esakova and Zhegallo 1996: 92, 93, pl. 3: 5‒8) and probably Avalonia (Landing 1988: 690, fig. 9, fig. 13, in part; Landing et al. 1989: 757, fig. 5, figs. 13, 14, in part).
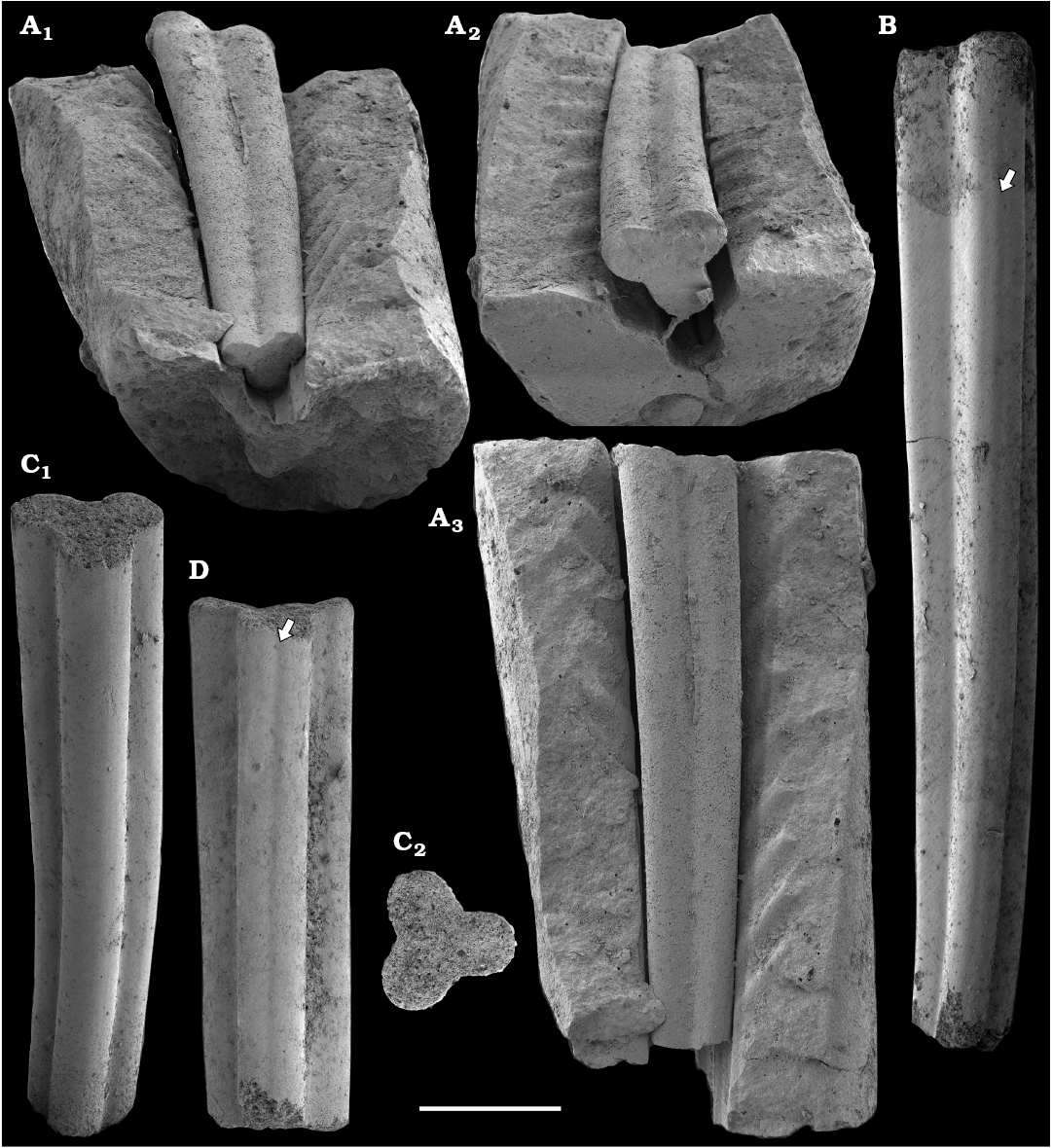
Fig. 71. Anabaritid Selindeochrea tripartita (Missarzhevsky in Rozanov et al., 1969), from early Cambrian Medvezhya Formation, western Anabar Uplift, Siberia, Russia; samples 3/12 (A), K2/26 (B), 3/12.2 (C, E), K2/25 (D), section 3. A. SMNH X5987, calcium phosphatic internal mould with external mould of longitudinal keels; A1, transverse apical view; A2, view of apertural end; A3, lateral view. B–D. SMNH X5988–5990, respectively. B, C1, D, lateral; C2, transversal views of calcium phosphatic internal moulds. Scale bar 500 µm.
Selindeochrea missarzhevskyi (Vasil’eva, 1986)
Fig. 72.
Material.—30 specimens, including figured SMNH X5993–5996, from sample 5a/34.75 (section 96-5a, Fig. 2), Emyaksin Formation, and seven internal moulds and one external mould, including figured SMNH X5992, 5997, from samples 1/28.5 and 1/29.2 (section 1, Fig. 4), from the Medvezhya Formation, Anabar Uplift, Siberian Platform, Russia. Correlated with the uppermost Fortunian and lower part of Cambrian Stage 2.
Description.—Tubes somewhat irregularly curved and longitudinally folded in three wide laterally extended lobes that have median keels. Outer surface with undulating growth lines. Lumen (internal mould) with three longitudinal lobes, each with rhomboidal or domal transverse profile. Lobes separated from each other by deep narrow grooves that shallow apically. In this direction, the lobes also acquire a rounded shape. Cross-section rounded at the apex.
Remarks.—The new material assigned to Selindocrea missarzhevskyi includes internal moulds very similar to those of other Selindeochrea fossils, i.e., with casts of prominent longitudinal keels and undulatory, transverse growth lines. The presence of longitudinal keels was also noted by Vasil’eva (1986). Selindeochrea missarzhevskyi is different from most other fossils attributed to Selindeochrea herein in having wider and straight lobes of the internal molds that are not longitudinally twisted. It most closely resembles S. tripartita, in particular apically, but S. missarzhevskyi differs in having more acute terminations of the lobes (as shown by internal moulds) and a greater apical angle of the thecae.
Stratigraphic and geographic range.—Cambrian Stage 2 (lower part) of the Siberian Platform.
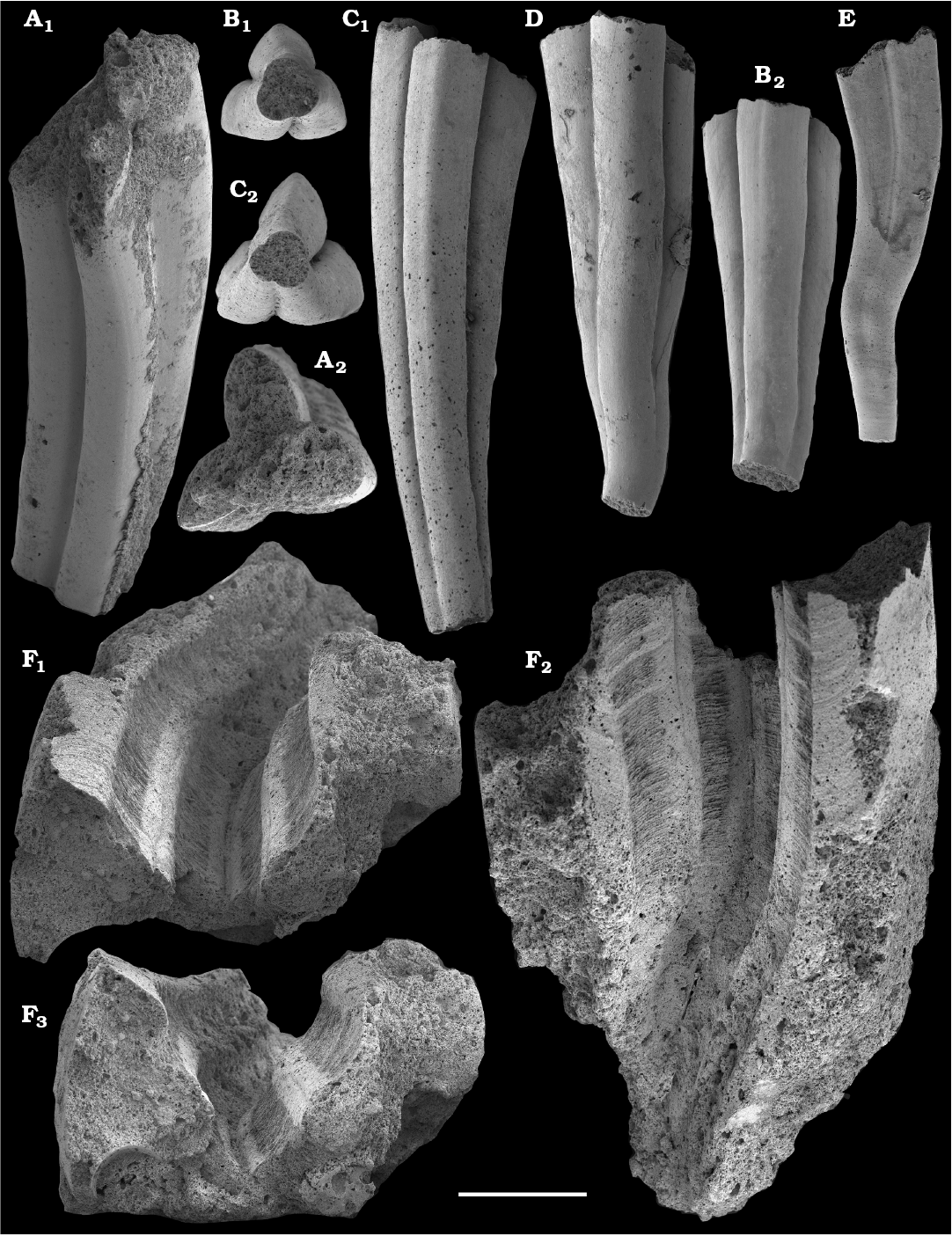
Fig. 72. Anabaritid Selindeochrea missarzhevskyi (Vasil’eva, 1986) from early Cambrian Medvezhya (A, F) and Emyaksin (B–E) formations, Anabar Uplift, Siberia, Russia; samples 1/28.5 (A, F), section 1; and 5a/34.75 (B–E), section 96-5a. A–E. SMNH X5992–5996, respectively; calcium phosphatic internal moulds. A1, B2, C1, D, E, lateral views; A2, transverse view of apertural end; B1, C2, transverse views of apical end. F. SMNH X5997, calcium phosphatic external mould; F1, oblique transverse; F2, lateral; F3, transverse views. Scale bar 500 µm.
Selindeochrea ternaria (Missarzhevsky in Rozanov et al., 1969)
Fig. 73.
Material.—Over ten internal moulds, including figured SMNH X5998–6001, from samples 1/29.2 (section 1, Fig. 4), 3/12.2, K2/25, K2/26 (section 3, Fig. 3), and 5a/18.5 (section 96-5a, Fig. 2), from the lower Emyaksin and Medvezhya formations, Anabar Uplift, Siberian Platform, Russia. Correlated with the upper Fortunian Stage and lower part of Cambrian Stage 2.
Description.—Tubes almost straight or somewhat curved, torted clockwise or counterclockwise, narrow, longitudinally folded in laterally elongate lobes separated by wide V-shaped grooves. Inner surface of the tubes (as replicated by internal moulds) with growth lines.
Remarks.—The narrow blade-like lobes are somewhat similar to the longitudinal keels described above in other species of Selindeochrea. Given that proposed similarity and lack of any prominent transverse sculptural elements, the fossil is tentatively attributed to Selindeochrea.
Stratigraphic and geographic range.—Upper Fortunian Stage and lower part of Cambrian Stage 2 of the Siberian Platform and South China.

Fig. 73. Anabaritid Selindeochrea ternaria (Missarzhevsky in Rozanov et al., 1969), calcium phosphatic internal moulds, from early Cambrian Medvezhya (A, C, D) and Emyaksin (B) formations, Anabar Uplift, Siberia, Russia; samples K2/25 (A), K2/26 (D), 3/12.2 (C), section 3; 5a/18.5 (B), section 96-5a. A. SMNH X5998. B. SMNH X6000. C. SMNH X5999. D. SMNH X6001. A1, C, D, lateral; A2, B, transverse views. Scale bar 500 µm.
Genus Anabarites Missarzhevsky in Voronova and Missarzhevsky, 1969
Type species: Anabarites trisulcatus Missarzhevsky in Voronova and Missarzhevsky, 1969; Anabarites trisulcatus Zone, lower Cambrian (lower Fortunian Stage); mouth of the Kotujkan River, Siberia, Russia.
Anabarites trisulcatus Missarzhevsky in Voronova and Missarzhevsky, 1969
Fig. 74A–F, H, I, K.
Material.—Several hundred fragmentary tubes and internal moulds, including figured SMNH X6002–6007, 6009, 6010, 6012, from samples 4/4, 1/C, 2/4, K1b/8, K1b/10, K1b/13, 2/7, and M419/12 (section 2, Fig. 1), Manykay and Nemakit-Daldyn formations, Anabar Uplift, Siberian Platform, Russia. Correlated with the Fortunian Stage.
Description.—Irregularly curved, slowly expanding tubes with triquetrous transverse profile. Three rounded lobes are separated by narrow and shallow grooves that are commonly distinctively defined on the internal and external surfaces of the tube. Thin transverse growth lines curve towards the aperture in the grooves and form wedge-like protrusions of the aperture (Fig. 74E).
Remarks.—Distinct wedge-like protrusions are observed among the type material from the Nemakit-Daldyn Formation. Specimens from the Medvezhya and Emyaksin formations have an apparently straight apertural margin and may represent, therefore, different species (see remarks on Anabarites cf. trisulcatus by Kouchinsky et al. 2009: 257). Internal moulds of Anabarites aff. trisulcatus (Fig. 74Q, R) from the Medvezhya Formation differ from the topotypes of A. trisulcatus in having broader, shallow longitudinal grooves. Growth lines are not expressed on the available internal moulds (see SOM 1 for other species associated with Anabarites aff. trisulcatus).
The limited number of distinctive features and difference in preservation of the tubes makes recognition of Anabarites trisulcatus and the distinction of potentially close species difficult. Several other forms of Anabarites co-occur in the upper Manykay Formation at the Bol’shaya Kuonamka River (sample 96-4/4; see also descriptions in Kouchinsky et al. 2009; SOM 1). These named species include rapidly expanding Anabarites latus Val’kov and Sysoev, 1970 (Fig. 74M) and Anabarites natellus Val’kov and Sysoev, 1970 (Fig. 74P) with rounded hexagonal cross-section.
Stratigraphic and geographic range.—The species is questionably reported (see discussion on carbon isotope trends above) from the uppermost Ediacaran of the Siberian Platform, occurs through the Terreneuvian Series worldwide and ranges into Cambrian Stage 3 of the Siberian Platform, South China and Tarim (see Kouchinsky et al. 2009, 2015a and references therein).
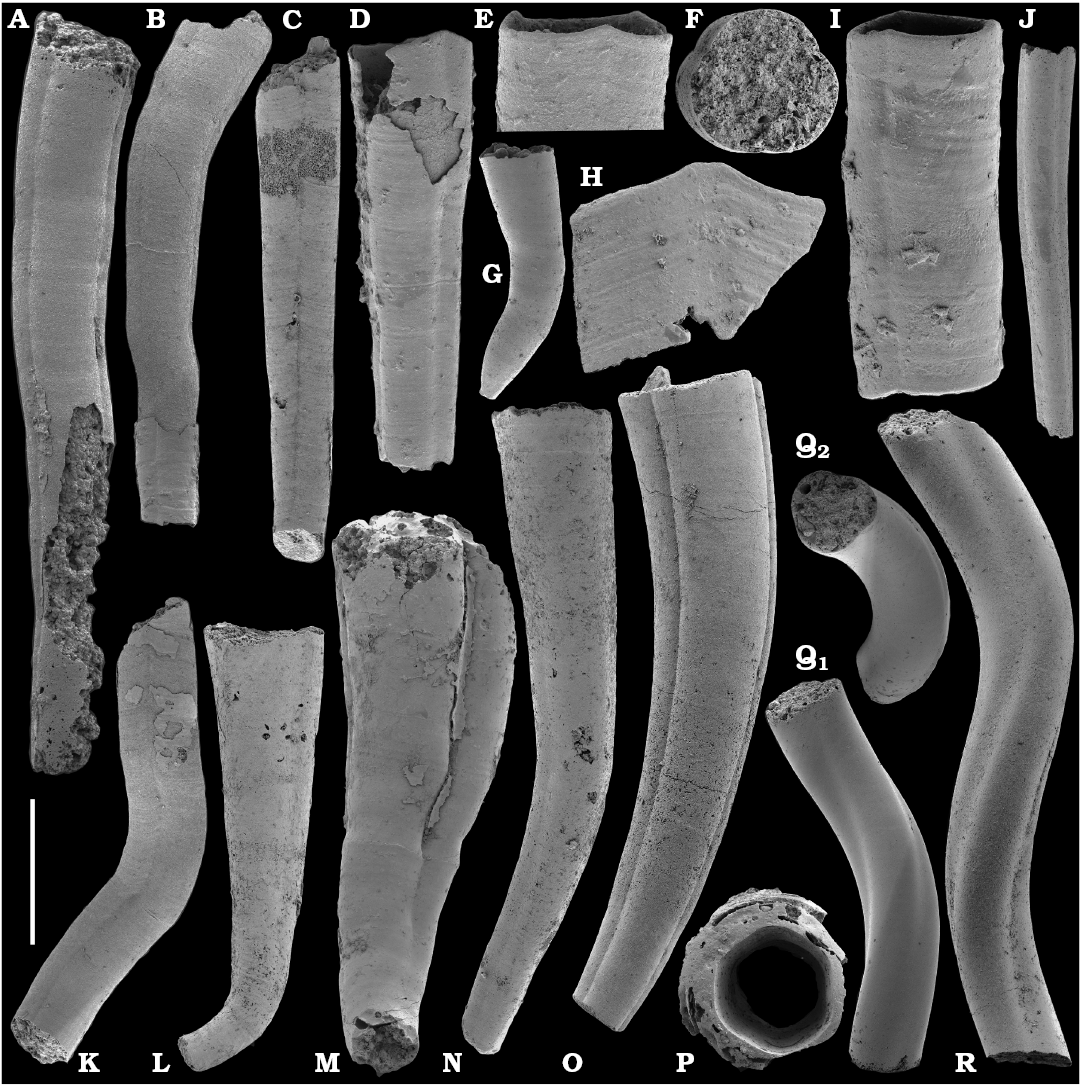
Fig. 74. Anabaritids from early Cambrian Nemakit-Daldyn (A–F, H, I, K, L, N), Manykay (G, M, P), Medvezhya (Q, R), and Emyaksin (J, O) formations, Anabar Uplift, Siberia, Russia; samples 2/7 (A, B, K), K1b/10 (C), M419/12 (D–F, H, I), 1282 (L, N), section 2; 1/29.2 (Q, R), section 1; 4/4 (G, M, P), 5/0 (O), and 5a/34.75 (J), from sections 96-4, 96-5, and 96-5a, respectively. A–F, H, I, K. Anabarites trisulcatus Missarzhevsky in Voronova and Missarzhevsky, 1969, SMNH X6002–6007 (A–F, respectively), SMNH X6009 (H), SMNH X6010 (I), SMNH X6012 (K), phosphatized tubes with internal moulds. A–D, I, K, lateral views; E, lateral view of apertural region; F, transverse view; H, apertural fragment of relatively large tube. G, J, L, N. Anabarites cf. trisulcatus, SMNH X6008 (G), SMNH X6011 (J), SMNH X6013 (L), SMNH X6015 (N), internal moulds, lateral views. M. Anabarites latus (Val’kov and Sysoev, 1970), SMNH X6014, lateral view. O. Anabarites cf. signatus Mambetov in Missarzhevsky and Mambetov, 1981, SMNH X6016, lateral view. P. Anabarites natellus (Val’kov and Sysoev, 1970), SMNH X6017, transverse view of etched external mould;. Q, R. Anabarites aff. trisulcatus Missarzhevsky in Voronova and Missarzhevsky, 1969, SMNH X6018, 6019, respectively; calcium phosphatic internal moulds, lateral views. Scale bar 250 µm (E, I), 500 µm (A–D, G, H, J, L, N–P), 1 mm (F, K, M, Q, R).
Anabarites kelleri Missarzhevsky, 1989
Fig. 75H–J.
Material.—Several thousand internal moulds, including figured SMNH X6024–6026, from samples 1/50, 1/46, 1/45, 1/39.3, 1/29.5 (section 1, Fig. 4), 3/0.1, 3/6.5, 3/12, 3/12.2, K2/12, K2/13, K2/20, K2/21, K2/22, K2/23, K2/24, K2/25, K2/26 (section 3, Fig. 3), 5/10, 5/10B, 5a/1, 5a/1.2, 5a/5, 5a/6, 5a/10.5, 5a/17.5 (section 96-5a, Fig. 2), Medvezhya Formation, western Anabar Uplift, Siberian Platform, Russia (SOM 1). Correlated with the uppermost Fortunian Stage.
Description.—Small, almost straight conchs with rounded triangular or tri-lobate cross-section; weakly developed longitudinal grooves; thin, straight growth lines present (see also Kouchinsky et al. 2009).
Remarks.—See Kouchinsky et al. (2009) for discussion of the validity of the species and Kouchinsky et al. (2015a) for probable lower part of Cambrian Stage 3 occurrence in the Emyaksin Formation.
Stratigraphic and geographic range.—Upper Fortunian Stage–Cambrian Stage 2 and probably lower part of Cambrian Stage 3, Siberian Platform (Kouchinsky et al. 2015a).
Anabarites biplicatus (Missarzhevsky, 1989)
Fig. 75A–G.
Material.—Several thousand internal moulds, including figured SMNH X3707, 3712, 3713, 6020–6023, from samples 1/45 (section 1, Fig. 4), K2/21, K2/22, K2/23, K2/24, K2/25, 3/12, 3/12.2, 3/12.5 (section 3, Fig. 3), 5/10, 5/10B, 5a/1.2, 5a/5 (sections 96-5 and 96-5a, Fig. 2), Medvezhya and Emyaksin formations, Anabar Uplift, Siberian Platform, Russia (SOM 1). Correlated with the uppermost Fortunian Stage.
Description.—Thecae up to 2 mm long, laterally flattened and curved in one plane, with two shallow, wide longitudinal depressions. Cross-section ovoid in later growth stage but rounded triangular apically.
Remarks.—See Kouchinsky et al. (2009) for discussion of the validity of the species and Kouchinsky et al. (2015a) for probable lower part of Cambrian Stage 3 occurrence in the Emyaksin Formation.
Stratigraphic and geographic range.—Upper Fortunian Stage–Cambrian Stage 2 and probably lower part of Cambrian Stage 3, Siberian Platform (Kouchinsky et al. 2015a).

Fig. 75. Anabaritids Anabarites biplicatus (Missarzhevsky, 1989) (A–G) and A. kelleri Missarzhevsky, 1989 (H–J), calcium phosphatic internal moulds, from early Cambrian Medvezhya (A–E, H–J) and Emyaksin (F, G) formations, Anabar Uplift, Siberia, Russia; samples K2/21 (A–C, E, J), K2/26 (H), K2/13 (I), 3/12 (D), section 3; K5/10 (F), 5a/1.2 (G), sections 96-5 and 96-5a, respectively. A–J. SMNH X3713 (A; see also Kouchinsky et al. 2009: fig. 38R), SMNH X3712 (B; see also Kouchinsky et al. 2009, fig. 38P), SMNH X6020 (C), SMNH X6021 (D), SMNH X3707 (E; see also Kouchinsky et al. 2009: fig. F–G), SMNH X6022 (F), SMNH X6023 (G), SMNH X6024–6026 (H–J, respectively). A–C, D2, E–G, H–J, lateral views; D1, transverse view of apertural end. Scale bar 500 µm (A–F, H–J), 1 mm (G).
Anabarites tristichus Missarzhevsky in Rozanov et al., 1969
Fig. 76A–C, F.
Material.—Several hundred internal moulds, including figured SMNH X3671, 3672, 6027, from samples 3/12, 3/12.2, K2/25 (section 3, Fig. 3), 5/0 and 5a/34.75 (sections 96-5 and 96-5a, Fig. 2), from the Medvezhya and Emyaksin formations, and two celestite-barite replaced tubes with internal moulds, including figured SMNH X3409, from sample K2/26, Medvezhya Formation, Anabar Uplift, Siberian Platform, Russia. Correlated with the upper Fortunian and lower part of Cambrian Stage 2.
Description.—Almost straight, slightly torted tubes with rounded hexagonal aperture. Walls with transverse flanges and thin, shallow growth lines. Three broad, flattened longitudinal lobes separated by shallow depressions. Internal, short protrusions of the wall are impressed into each longitudinal depression and essentially extend as a dashed line on the internal moulds from the apex to the aperture along the three longitudinal depressions (Fig. 76A, B, F).
Remarks.—Anabarites tristichus tubes preserved in celestite and barite are discussed by Kouchinsky and Bengtson (2002) and Kouchinsky et al. (2009). The first illustration of anabaritids from Siberia was published by Sysoev (1965: fig. 2) under the name Hyolithellus sp. (recognized by Kouchinsky et al. [2009] as A. tristichus Missarzhevsky in Rozanov et al., 1969). The age of A. tristichus (Hyolithellus sp. Sysoev [1965: fig. 2]) from “the Atdabanian Horizon” from a locality on the left bank of the Kotuj River, 6 km downstream from the mouth of the Kotujkan River on the western flank of the Anabar Uplift) is questionable. It is not clear from which level of the outcrop the specimen was collected. It probably came from the Kugda-Yuryakh Formation, and, thus, can be correlated with lower part of Cambrian Stage 2.
Stratigraphic and geographic range.—Terreneuvian Series and lower Stage 3; Siberia and Gondwana (South China).
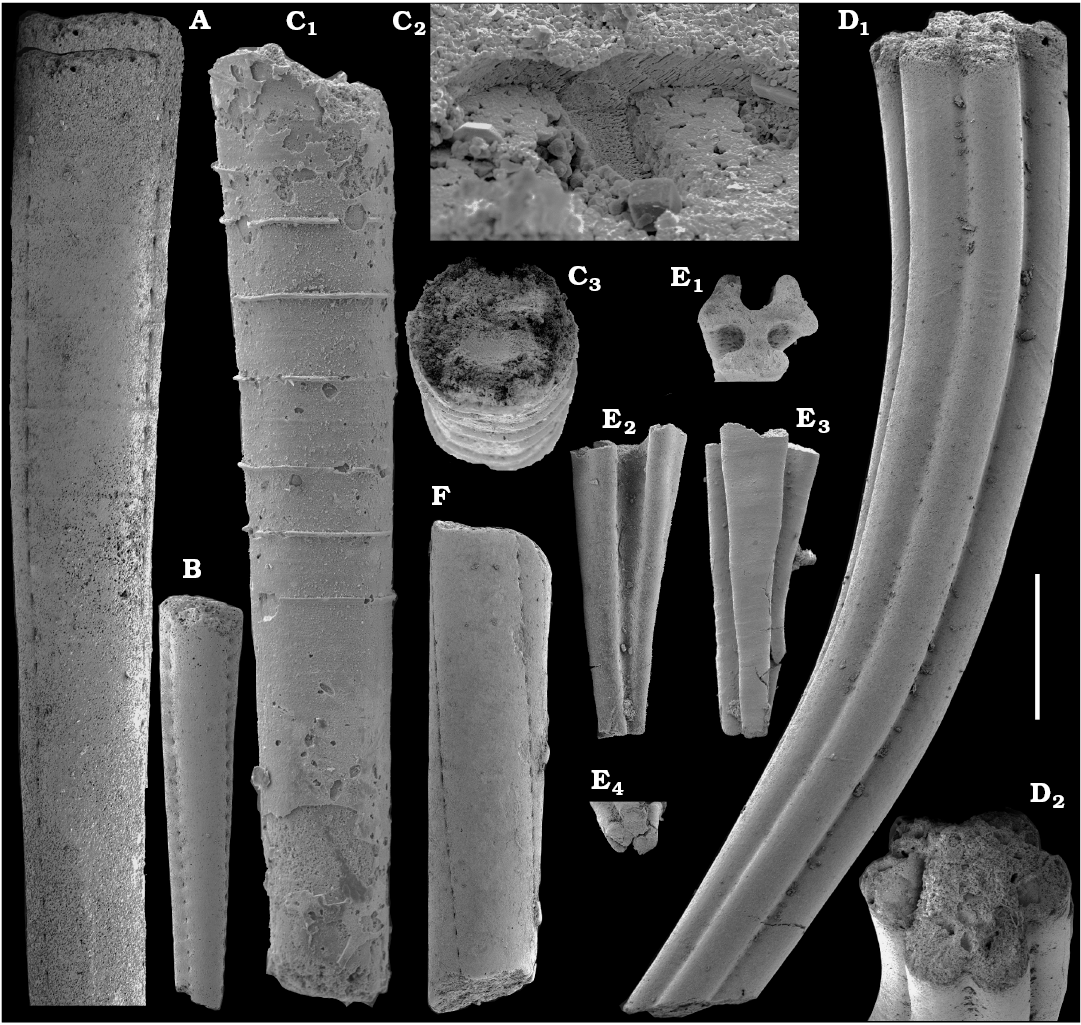
Fig. 76. Anabaritids from early Cambrian Medvezhya (A–D) and Emyaksin (F, E) formations, Anabar Uplift, Siberia, Russia; samples K2/25 (A, B), K2/26 (C), section 3; 5/0 (F), section 96-5; 5a/34.5 (E), section 96-5a; and 1/29.2 (D), section 1. A–C, F. Anabarites tristichus Missarzhevsky in Rozanov et al., 1969, SMNH X3671 (A; see also Kouchinsky et al. 2009: fig. 26C), SMNH X3672 (B; see also Kouchinsky et al. 2009: fig. 26D), SMNH X3409 (C; see also Kouchinsky and Bengtson 2002: figs. 2, 3 and Kouchinsky et al. 2009: fig. 26A, B), SMNH X6027 (F). A, B, F, lateral views of calcium phosphatic internal moulds; C1, lateral view of celestite-barite replaced tube with calcium phosphatic internal mould; C2, internal protrusion of the wall enlarged; C3, transverse view. D. Anabarites hexasulcatus (Missarzhevsky, 1974), SMNH X6028; D1, lateral; D2, transverse views. E. Anabarites hariolus (Val’kov, 1987), SMNH X3697 (see also Kouchinsky et al. 2009: fig. 36K–M). E1, transverse view of apertural end; E2, E3, lateral views; E4, transverse view of apical end. Scale bar 25 µm (C2), 500 µm (A, B, C1, C3, D–F).
Anabarites compositus Missarzhevsky in Rozanov et al., 1969
Fig. 77.
Material.—Over one hundred internal moulds with rare fragments of phosphatized walls, including figured SMNH X3677–3679, 3681–3684, 6029, 6030, from sample 5a/34.5 (see Kouchinsky et al. 2009; Fig. 77A–I), and probably also several glauconitised internal moulds, including figured SMNH X6031, 6032, from sample 5a/18.5, Emyaksin Formation, eastern Anabar Uplift, Siberian Platform, Russia. Correlated with lower part of Cambrian Stage 2.
Description.—Almost straight tubes with rounded hexagonal inner cross-section (Fig. 77G, J) and transverse, flanged growth lines. Internal protrusions of the conch wall are obliquely inclined towards the aperture (Fig. 77H1) and replicated as three rows of notches in the middle of each depression on the internal mould (Fig. 77F2, I, K). A semi-circular pocket in the inner surface of the tube wall is associated with each protrusion (Fig. 76E, H1, D2). These structures produce chevron-like elements on the internal moulds.
Remarks.—The internal indentations on Anabarites compositus tubes are spine-like and rounded in cross-section (Kouchinsky et al. 2009: fig. 30A, B). They differ from the blade-like, longitudinally elongate indentations along the axis of the tube moulds in A. tristichus (compare Fig. 77K with Fig. 76A). The pocket-like structures in the wall of A. compositus are reminiscent of the circumferential internal folds in the walls of Aculeochrea rugosa Val’kov and Sysoev, 1970 (Fig. 78) and A. ornata Val’kov and Sysoev, 1970 (see Kouchinsky et al. 2009: fig. 49). For this reason, Missarzhevsky (1989: 193) assigned A. compositus to Aculeochrea Val’kov and Sysoev, 1970.
Spine-like protrusions covered by “lamellae” are apparently present in Aculeochrea trilamellosa Vasil’eva, 1998, and Aculeochrea mesezhnikovi Vasil’eva, 1998 (Vasil’eva 1998: pl. 16: 1–5, pl. 17: 1, 2, 5). The “lamellae” presumably represent similar structures on the internal moulds as the chevrons in A. compositus. Aculeochrea mesezhnikovi is reportedly different from A. compositus in having “lamellae” situated in depressed parts of the mould (grooves), whereas the more prominent and wider lobes are devoid of such “laminae” (Vasil’eva 1998). Aculeochrea trilamellosa similarly has three longitudinal rows of “lamellae” and a circular cross-section of the internal mould. The illustrations by Vasil’eva (1998) do not, however, allow a more detailed comparison with Anabarites compositus. A fossil from the upper lower Cambrian of England misidentified (Hinz 1987: pl. 14: 37, 38) as A. compositus actually resembles Selindeochrea tricarinata (Missarzhevsky; see also Brasier 1989: 135), but most likely represents the orthothecid hyolith Gracilitheca.
Stratigraphic and geographic range.—Lower part of Cambrian Stage 2, Siberian Platform.
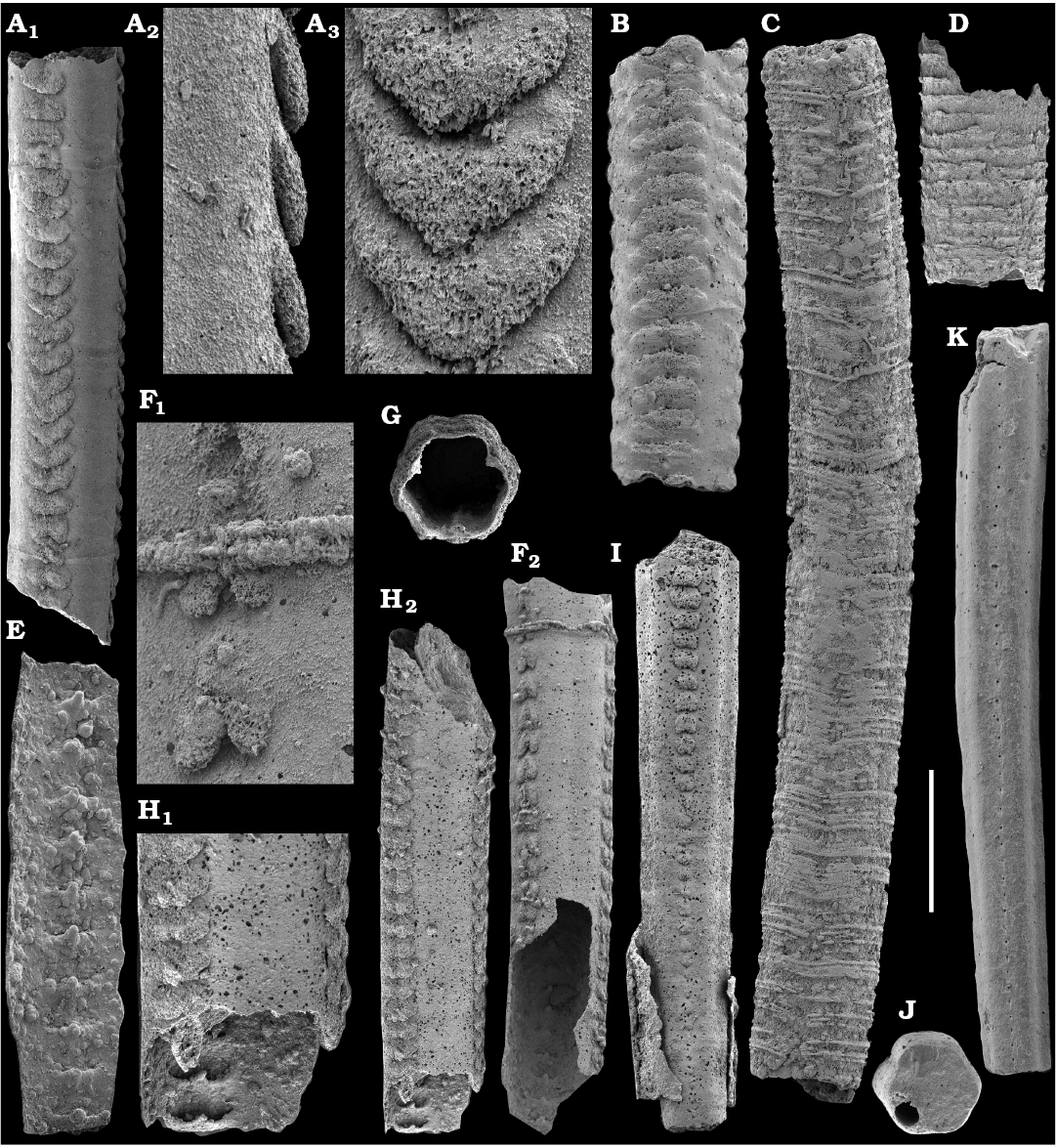
Fig. 77. Anabaritid Anabarites compositus Missarzhevsky in Rozanov et al., 1969, from early Cambrian Emyaksin Formation, Anabar Uplift, Siberia, Russia; samples 5a/34.5 (A–I) and 5a/18.5 (J, K), section 96-5a. A–K. SMNH X3677 (A; see also Kouchinsky et al. 2009: fig. 29A–C), SMNH X3679 (B; see also Kouchinsky et al. 2009: fig. 29F, G), SMNH X3678 (C; see also Kouchinsky et al. 2009: fig. 29D–E), X6029 (D), SMNH X3682 (E; see also Kouchinsky et al. 2009: fig. 30A–E), SMNH X3684 (F; see also Kouchinsky et al. 2009: fig. 31), SMNH X3683 (G; see also Kouchinsky et al. 2009: fig. 30F–H), SMNH X6030 (H), X3681 (I; see also Kouchinsky et al. 2009: fig. 29J), SMNH X6031 (J), and SMNH X6032 (K). A1, B–D, F2, H2, I, K, lateral views of internal moulds and calcium phosphatic inner and outer coats on the tube walls; A2, A3, enlargement of “chevrons” on surface of internal mould; E, fragment of inner coating with protrusions directed towards apertural end of tube; F1, enlargement of F2 showing external surface of inner coating with “chevrons” and partly preserved phosphatized apertural flanges; G, transverse view of partly phosphatized tube; H1, enlargement of H2 showing “chevrons” and internal protrusions of the wall preserved by inner phosphatic coating of conch; J, transverse view of internal mould. Scale bar 100 µm (A2, A3, F1), 250 µm (H1), and 500 µm (A1, B–D, F2, G, H2, I–K).
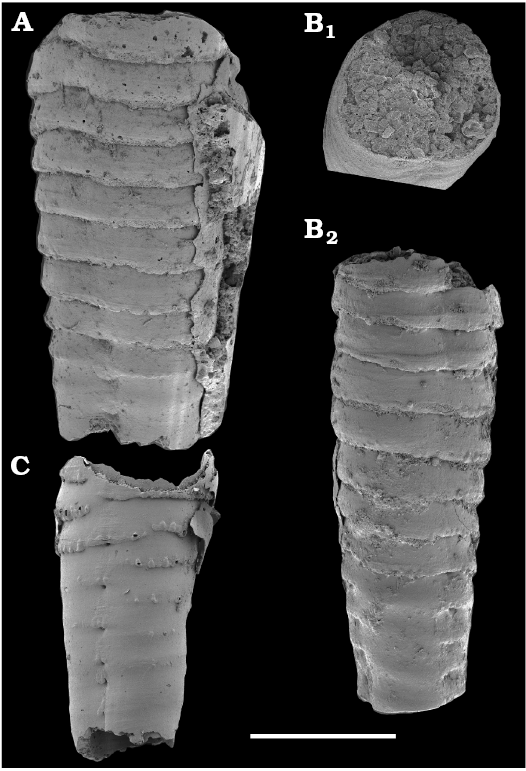
Fig. 78. Anabaritid Aculeochrea rugosa (Val’kov and Sysoev, 1970), internal moulds, from early Cambrian Manykay Formation, eastern Anabar Uplift, Siberia, Russia; sample 4/4, section 96-4. A. SMNH X6033. B. SMNH X3773 (see also Kouchinsky et al. 2009: fig. 50G). C. SMNH X3768 (see also Kouchinsky et al. 2009: fig. 50A–B). A, B2, C, lateral; B1, transverse views. Scale bar 500 µm.
Anabarites hexasulcatus (Missarzhevsky, 1974)
Fig. 76D.
Material.—About 30 internal moulds from samples K2/30 (section 3, Fig. 3), 1/29.2, 1/28.5, 1/6 (section 1, Fig. 4), 5a/6, 5a/17.5, and 5a/18.5 (section 96-5a, Fig. 2), including figured SMNH X6028, from the Medvezhya, Kugda-Yuryakh, and Emyaksin formations, Anabar Uplift, Siberian Platform, Russia. Correlated with the uppermost Fortunian and lower part of Cambrian Stage 2.
Description.—Conchs usually curved in one plane or irregularly without noticeable torsion, with rounded hexagonal aperture and thin growth lines (Fig. 76D1). Lumen (as replicated by internal moulds) subdivided into six rounded longitudinal lobes and intervening grooves. Three of the grooves are usually somewhat deeper than the other three; the shallower grooves split each of the main lobes into a pair of secondary ones. Second-order grooves appear early on the lobes which are delimited by the first-order (i.e., deeper) grooves.
Remarks.—See Kouchinsky et al. (2009) for discussion.
Stratigraphic and geographic range.—Uppermost Fortunian and lower part of Cambrian Stage 2, Siberian Platform, western Mongolia (Esakova and Zhegallo 1996: 96, pl. 3.17), and probably South China (Kouchinsky et al. 2009).
Anabarites hariolus (Val’kov, 1987)
Fig. 76E.
Material.—Single specimen, SMNH X3697, from sample 96-5a/34.5, Emyaksin Formation, eastern Anabar Uplift, Siberian Platform, Russia. Correlated with lower part of Cambrian Stage 2.
Description.—Tube almost straight, expands rapidly, with three prominent T- or Y-shaped longitudinal lobes separated by deep grooves. Each of the lobes apically has a second-order groove that shallows and disappears apically.
Remarks.—See Kouchinsky et al. (2009) for discussion.
Stratigraphic and geographic range.—Cambrian Stage 2, Siberian Platform.
Genus Tiksitheca Missarzhevsky in Rozanov et al., 1969
Type species: Tiksitheca licis Missarzhevsky in Rozanov et al., 1969; lower Cambrian (upper Fortunian–lower part of Cambrian Stage 2); mouth of the Kugda-Yuryakh Creek, Kotuj River, Siberia, Russia.
Remarks.—Landing (1988) and Landing et al. (1989) discussed the generally similar, rounded-triangular, elongate conchs of Anabarites and Tiksitheca. These reports also noted the range of variability in the depth or even presence of longitudinal grooves in large collections of Anabarites tubes and/or moulds. For this reason, the two genera of which Anabarites was actually proposed by Voronova and Missarzhevsky (1969), were considered synonymous (Landing et al. 1989). In this report, the traditional distinctions of earlier named Anabarites and Tiksitheca species are however maintained (see also Kouchinsky et al. 2009).
Tiksitheca licis Missarzhevsky in Rozanov et al., 1969
Fig. 79A–C, K–M.
Material.—Thousands of internal moulds and partially phosphatized tubes, including figured SMNH X6034–6038, from the Manykay, Nemakit-Daldyn, Medvezhya, and lower Emyaksin formations, Anabar Uplift, Siberian Platform, Russia. Correlated with the upper Fortunian and lower part of Cambrian Stage 2.
Description.—Curved to almost straight calcareous tubes with rounded triangular, radially symmetrical cross-section, slowly expanding from the apical part. The wall may include relatively prominent, transverse flanges and intervening growth lines (Kouchinsky and Bengtson 2002). Inner surface of the tube with transverse growth lines (as reflected on internal moulds).
Remarks.—The presence of flanges in a wall with anabaritid-type microstructure is also known from Anabarites tristichus (Kouchinsky and Bengtson 2002), and is compatible with the interpretation of Tiksitheca licis as an anabaritid (see remarks by Missarzhevsky in Rozanov et al. 1969: 114). The type material of T. licis from sample M423/13 (Fig. 3), which is a horizon in section 3 of this report, was collected by Vladimir V. Missarzhevsky from the Medvezhya Formation.
Partly phospatized slightly irregularly curved tubes preserved with diagenetic phosphatic inner and outer crusts are found in sample 3/10 from the Medvezhya Formation (Fig. 80A–E, G). Numerous distinct straight or slightly undulating growth increments are replicated on their outer surfaces. The tubes have rounded triangular outer cross-sections, and their internal moulds would normally be identified as Tiksitheca licis. The latter occur in the same beds of the Medvezhya Formation (SOM 1). In addition to T. licis, rare problematical, longitudinally twisted tubes with rounded triangular cross-section, known as Kugdatheca voluta Missarzhevsky in Rozanov et al., 1969, were extracted from several samples (Fig. 79N, O, SOM 1).
Stratigraphic and geographic range.—The type species Tiksitheca licis is known from the upper Fortunian Stage and commonly occurs throughout the strata correlated herein with lower part of Cambrian Stage 2 in Siberia and reportedly ranges into the upper part of Cambrian Stage 2 (Kouchinsky et al. 2009 and references therein). The species is often listed with T. korobovi and T. voluta (see Kouchinsky et al. 2009). Tiksitheca licis and Tiksitheca spp. are also reported from the upper Fortunian–lower part of Cambrian Stage 3 of South China (Li et al. 2007). Tiksitheca licis occur in the Terreneuvian strata of western Mongolia, Iran, India, and Kazakhstan (see Kouchinsky et al. 2009 and references therein).
Genus Cambrotubulus Missarzhevsky in Rozanov et al., 1969
Type species: Cambrotubulus decurvatus Missarzhevsky in Rozanov et al., 1969; lower Cambrian (upper Fortunian–lower part of Cambrian Stage 2); mouth of the Ary-Mas-Yuryakh Creek, Kotuj River, Siberia, Russia.
Cambrotubulus decurvatus Missarzhevsky in Rozanov et al., 1969
Fig. 79D–J, P.
Material.—Hundreds of internal moulds (SOM 1), including figured SMNH X6039–6045, X6048, from the Manykay, Nemakit-Daldyn, Medvezhya, and lower Emyaksin formations, Anabar Uplift, Siberian Platform, Russia. Correlated with the Fortunian and lower part of Cambrian Stage 2.
Description.—Variably curved to almost straight calcareous tubes with variable expansion rates, with circular transverse cross-section and straight thin growth lines. Aperture straight, apical part tapered open (where completely preserved).
Remarks.—The type material of Cambrotubulus decurvatus came from sample M410/73 (Fig. 4). It was collected by Vladimir V. Missarzhevsky from the Medvezhya Formation at section 1 of this report. Tubular straight to broader conical internal moulds with circular cross-section co-occur in the type material and topotype sample 1/46 (SOM 1). This variability is consistant with the original diagnosis of the genus and species. Rapidly expanding funnel shaped internal moulds were later referred to as Cambrotubulus conicus Missarzhevsky, 1989, whereas coiled tubular moulds were termed C. plicativus Val’kov, 1987. Both were first described from the upper Fortunian strata of the same locality at the Olenyok River section on the Olenyok Uplift, northern Siberian Platform (see Cambrotubulus spp. in Kouchinsky et al. 2009).
In Siberia, Cambrotubulus-like fossils are reported from the uppermost Ediacaran (Khomentovsky and Karlova 2005; Zhuravlev et al. 2012; Nagovitsin et al. 2015; Rogov et al. 2015), contrary to Zhuravlev et al. (2012: 219), who tend to identify these fossils in the late Ediacaran as Chenmengella Zhuravlev, Liñán, and Gámez Vintaned, 2012. Cambrotubulus does not have a tri-radial cross-section and does have irregularly curved tubes. It, thus, is very similar to Chenmengella. Cambrotubulus decurvatus and other Cambrotubulus species reported from Siberia commonly appear in the Terreneuvian Series, although they are reliably known only from strata correlated herein with the middle part of the Fortunian Stage though the lower Cambrian Stage 2. Cambrotubulus tubes from Siberia exhibit the same microstructure of the wall as observed in such other anabaritid species as Anabarites tristichus and Tiksitheca licis in cases of exceptional preservation (Kouchinsky and Bengtson 2002).
Cambrotubulus decurvatus also occurs in the lower and middle Bayan Gol Formation of western Mongolia (Esakova and Zhegallo 1996), which was correlated by Demidenko and Parkhaev (2014), Landing and Kouchinsky (2016), and in this report with the pre-Tommotian beds of Siberia, and is referable to the Fortunian and, pending further study, to the lower part of Cambrian Stage 2 (Brasier et al. 1996; Kouchinsky et al. 2012). It also occurs through Meishucunian Assemblage Zone 1 of South China (e.g., Yang et al. 2014). Cambrotubulus decurvatus was also reported from the Mongolian Khairkhan Formation (Esakova and Zhegallo 1996, not illustrated). C. corniformis Elicki, 1994, was first described from the Ludwigsdorf Member in the Görlitz Syncline of Germany. Additional material is, however, needed to corroborate the identification of these fossils. Thus, the upper range of Cambrotubulus tubes is uncertain and may extend into Cambrian Stage 3.
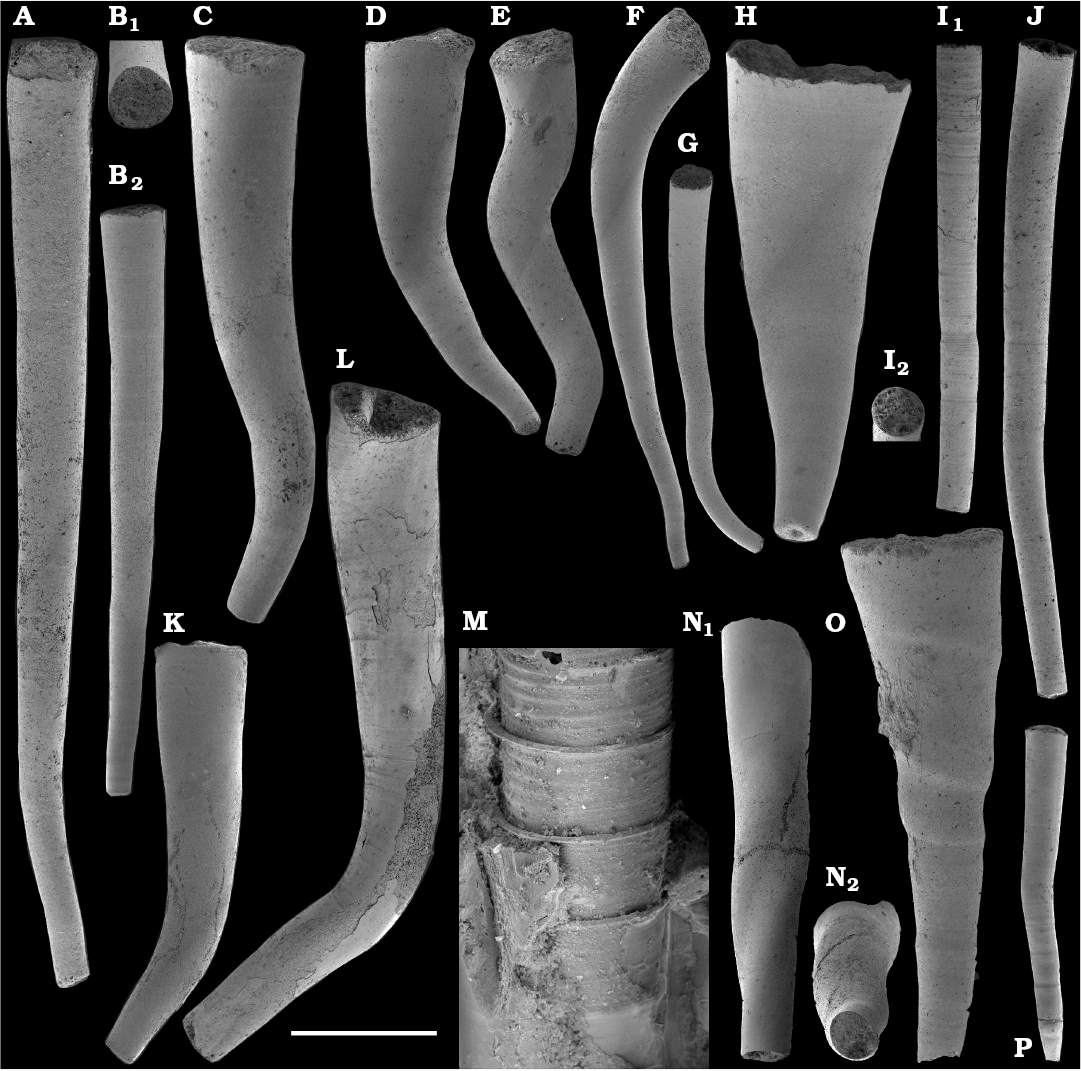
Fig. 79. Anabaritids from early Cambrian Medvezhya (A, B, D–G, I, J, L, M, P) and Emyaksin (C, H, K, N, O) formations of Anabar Uplift, Siberia, Russia; samples 3/12.2 (A, B, D, F, G), K2/21 (L), K2/26 (E, M), section 3; 5a/1.2 (C, H, K, N, O), section 96-5a; 1/22 (I, J), 1/29.5 (P), section 1. A–C, K, L. Tiksitheca licis Missarzhevsky in Rozanov et al., 1969, internal moulds, SMNH X6034, 6035 (A, B, respectively), SMNH X6037 (C), SMNH X6036 (K), SMNH X6038 (L). A, B2, C, K, L, lateral views; B1, transverse view of apertural end. M. T. licis, SMNH X3410 (see also Kouchinsky and Bengtson 2002: fig. 6 and Kouchinsky et al. 2009: fig. 39E), lateral view of tube replaced by celestite and barite. D–J, P. Cambrotubulus decurvatus Missarzhevsky in Rozanov et al., 1969, internal moulds, SMNH X6039–6045 (D–J, respectively), SMNH X6048 (P). D–H, I1, J, P, lateral views; I2, transverse view of apertural end. N. Kugdatheca voluta Missarzhevsky in Rozanov et al., 1969, SMNH X6046. N1, lateral view of internal mould; N2, oblique transverse view of apical end. O. K. voluta?, lateral view of internal mould, SMNH X6047. Scale bar 500 µm (M, O), and 1 mm (A–L, N, P).
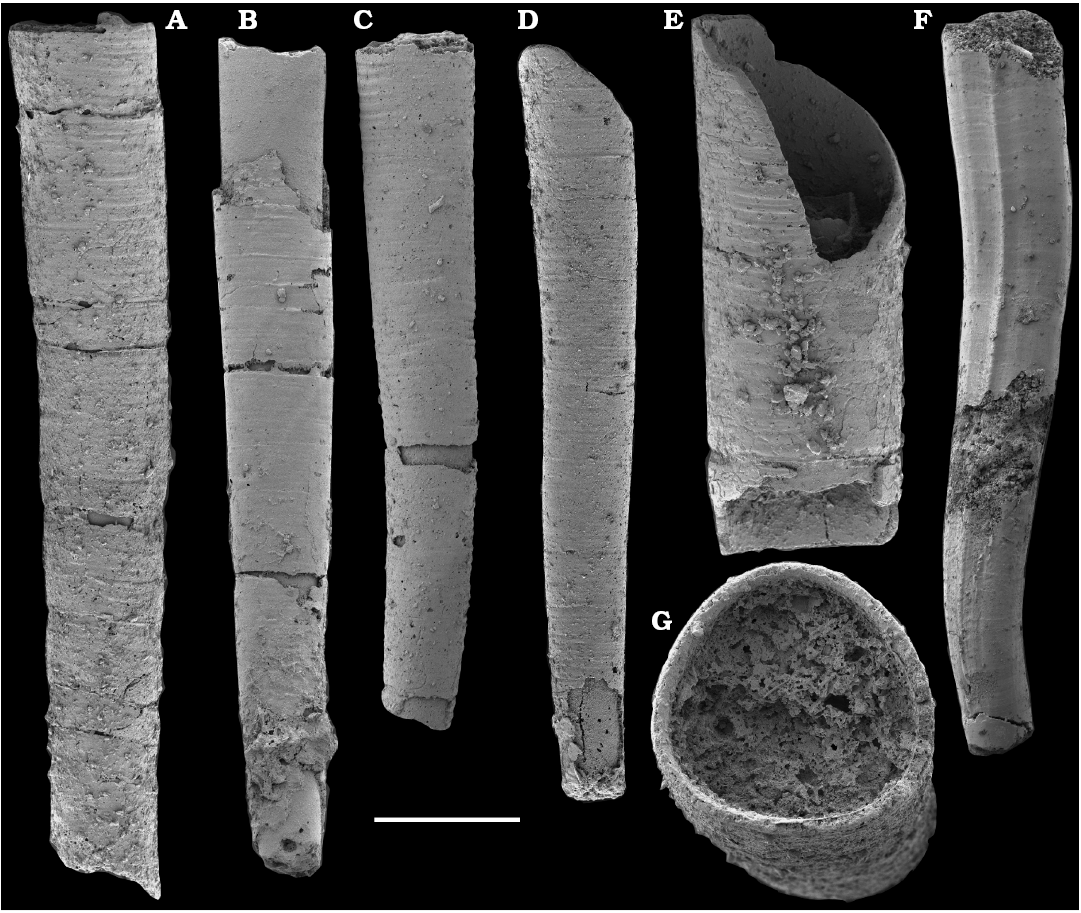
Fig. 80. Problematic tubes from early Cambrian Medvezhya Formation, western Anabar Uplift, Siberia, Russia; samples 3/10 (A–E, G) and K2/26 (F), section 3. A–E, G. Tiksitheca licis? Missarzhevsky in Rozanov et al., 1969, SMNH X6049–6053 (A–E, respectively), SMNH X6054 (G). Calcium phosphatic internal moulds and outer coatings of conchs. F. Problematic tube or sclerite, lateral view of internal mould, SMNH X6055. Scale bar 250 µm (G), 500 µm (A–E), 1 mm (F).
Phylum Foraminifera? d’Orbigny, 1826
Class, order, and family uncertain
Genus Platysolenites Pander, 1851
Type species: Platysolenites antiquissimus Eichwald, 1860; lower Cambrian (Terreneuvian Series); near Kunda, Estonia.
Platysolenites antiquissimus Eichwald, 1860
Figs. 81, 82H, SOM 3.
Material.—More than 140 specimens, including figured SMNH X6056–6064, 6072, from samples 1/A, 1/54, 1/46, 1/45, 1/29.5, 1/29.2, 1/27.5, 1/25.5, 1/22.5, 1/22 (section 1, Fig. 4), K1b/9, 2/A, K1a/24, K1a/25, K1a/26, K1a/27, K1a/28, K1a/42A, K1a/45, K1a/47, K1a/48, K1a/82 (section 2, Fig. 1), 3/0, 3/6.5, 3/10, 3/12, 3/12.5, 3/20.5, K2/12, K2/20, K2/21, K2/22, K2/24, K2/30, K2/33, K2/43, K2/50 (section 3, Fig. 3), 5/0.5, 5a/1, 5a/5, 5a/6, 5a/8, 5a/9, 5a/18.5, and 5a/34.5 (sections 96-5 and 96-5a, Fig. 2), Nemakit-Daldyn, Medvezhya, and Emayksin formations. Correlated with the Fortunian and lower part of Cambrian Stage 2.
Description.—Tubes composed of siliceous or clay grains (up to 5 mm long and up to 0.7 mm in diameter) with circular cross-section and straight aperture. The tubes have straight tranverse annulation (Fig. 81I) and granular composition of the wall, which is up to 70 µm thick.
Remarks.—Platysolenites is interpreted as a sedentary foraminiferan with agglutinated tests that commonly contain grains of quartz and K-feldspar (e.g., Glaessner 1978; Felitzyn 1992; McIlroy et al. 1994). The tests show an elastic deformation that suggests that they were not rigid in life (Fig. 81C–F). A bulbous initial part (proloculus of McIlroy et al. 2001) is invariably missing from our material. Some of the fragments presumably preserve a circular aperture (Fig. 81A, D, G). Finds of agglutinated spherical tests of Blastulospongia sp. (see below) similar in composition to tubular Platysolenites antiquissimus are consistent with a protistan affinity of both of these fossils.
Tubular fossils defined as Platysolenites sp. are illustrated from drill cores from the subsurface margins of the Siberian Platform (east Western Siberia; Kontorovich et al. 2008), where they co-occur with the upper Ediacaran Cloudina–Namacalathus assemblage (Zhuravlev et al. 2012). Platysolenites is also reported from Purella Zone equivalent beds on the northwestern and southern flanks of the Anabar Uplift by Missarzhevsky (1989: 212). The species is known from drill cores in the northern inner part of the Siberian Platform (Khomentovsky et al. 1998), although never previously illustrated or adequately described from the Cambrian deposits of the Siberian Platform. This report shows that P. antiquissimus commonly occurs in the Nemakit-Daldyn, Medvezhya and lower Emyaksin formations of the Anabar Uplift. Thus, most occurrences of the species from Siberia are from the Fortunian Stage. In contrast with terrigenous deposits of Laurentia, Baltica and Avalonia, the species in Siberia occurs mainly in carbonate deposits.
Stratigraphic and geographic range.—Uppermost Ediacaran? of Siberia (Kontorovich et al. 2008; Zhuravlev et al. 2012); Terreneuvian Series of Siberia, Baltica, Laurentia, and Avalonia; Cambrian Stage 3 of Baltica, Laurentia, Avalonia, and Armorica (McIlroy et al. 2001; Streng et al. 2005, and references therein).
Genus Blastulospongia Pickett and Jell, 1983
Type species: Blastulospongia monothalamos Pickett and Jell, 1983; middle Cambrian; Australia.
Blastulospongia sp.
Fig. 82A, B, SOM 3.
Material.—Two almost complete specimens, SMNH X6065 and 6066, and a single fragment from samples 1/22.5 and 1/26.3 (section 1, Fig. 4), Medvezhya Formation, western Anabar Uplift. Correlated with the lowermost Cambrian Stage 2.
Description.—Hollow spherical tests perforated with numerous rounded pores. The smaller specimen (Fig. 82A) is ca. 350 µm and the larger one (Fig. 82B) ca. 800 µm in diameter. The pores are 20–50 µm in diameter and randomly scattered on the surface with a density of 150 pores/mm2. Thickness of the wall is 10–20 µm. Wall of the larger specimen is deformed with isometrical folds.
Remarks.—Blastulospongia sp. is represented by two specimens that are markedly different in diameter, although having the same density and diameter of pores. They are different from all known species of Blastulospongia in their diameter, pore sizes and pore density. Blastulospongia monothalamos from the middle Cambrian of Australia are 1.0–1.9 mm in diameter and with pores ca. 50 µm in diameter. Blastulospongia mindyallica Bengtson, 1986, from the upper Cambrian of Australia is 280–520 µm in diameter, with 15–20 µm perforations and a few µm-thick wall (Bengtson 1986) and has ca. 300 pores/mm2. Blastulospongia polytreta Conway Morris and Chen, 1990, from the lower Cambrian of South China has a maximum diameter of 350–370 µm and ca. 7 µm-thick walls with ca. 1500 pores/mm2 that range in diameter from 9 to 21 µm (Conway Morris and Chen 1990). Aetholicopalla adnata from Meishucunian Assemblage Zone 2 of South China (Yang et al. 2014: fig. 13P) is probably also a Blastulospongia that is similar to the form described herein. It is ca. 700 µm in diameter with pores larger than 50 µm. From the Fortunian strata of northwestern Canada, Pyle et al. (2006) reported two “indeterminate spheroids” 425 µm in diameter with pores from 3 to 10–15 µm in diameter and compared these to B. polytreta. Blastulospongia minima (nomen nudum) referred to as planktonic Sarcodina incertae sedis represents the main component (50–90%) of presumably lower–middle Atdabanian-age, pelagic bedded cherts in olistostome-containing flysch of the Spassky terrain (Russian Far East), where they co-occur with siliceous spicules (Panasenko 1998). Blastulospongia minima spherical tests are different from other species of Blastulospongia in being much smaller. They are reportedly 15–60 µm in diameter and are perforated by pores of 1 µm (in smaller specimens) to 3 µm (in larger ones) in diameter and may represent a protistan genus different from Blastulospongia.
Several fossils attributable to Blastulospongia were previously described from Siberia. A hemispherical problematicum, ca. 0.85 mm in diameter with pores 20–40 µm in diameter, is known from the lower Pestrotsvet Formation (Spinulitheca billingsi Zone) at the Gonam section on Mount 1291 m (sample 1605 of Val’kov 1987: pl. 14: 15). Irregularly spherical, rarely elongated fossils 1–2 mm in diameter with pores up to 0.5 mm in diameter (Khomentovsky et al. 1990: 34, but up to 40 µm in diameter in the illustrations) from the upper Kessyusa Formation on the Olenyok Uplift (i.e., on the Kyursyuke River ca. 17 km upstream of the mouth) (12 specimens; Khomentovsky et al. 1990: pl. 6: 5–7, pl. 7: 1) and from the lowermost Pestrotsvet Formation at the Aldan River at Byukteleekh (single specimen; Khomentovsky et al. 1990: pl. 7: 2) were referred to as Blastulospongia monothalamos. A globula from the lower Emyaksin Formation at the Udzha River reported as Markuelia sp. by Vasil’eva (1998: pl. 44: 10) is ca. 0.55 mm in diameter with pores up to 30 µm in diameter.
Deformation of the wall in the larger specimen suggests that it was not completely mineralized in life but rather contained a significant amount of organic material. Pickett and Jell (1983) first suggested a sphinctozoan or foraminiferan affinity of Blastulospongia. A radiolarian affinity of Blastulospongia was discussed by Bengtson (1986) and White (1986). It was also regarded as a problematicum variously assigned to the sponges, radiolarians or uncertain protists (Conway Morris and Chen 1990). A protozoan affinity may be supported by a silicified and supposedly agglutinated nature of the tests studied herein, in which energy-dispersive X-ray analyses suggest the presence of detrital minerals incorporated into the wall and cemented with silica. The wall consists of 1–10 µm crystals having the following elemental composition: O (60–75 atomic %), Si (ca 10–25 atomic %), K (2–10 atomic %), Al (2–10 atomic %) with trace amounts of Fe, Mg, and Ca (SOM 3). Co-occurrence with calcitic shelly fossils (now dissolved) preserved as phosphatic internal moulds as well as with organophosphatic Fomitchella sclerites and siliceous/agglutinated Platysolenites (SOM 3b) is compatible with an originally siliceous composition of the Blastulospongia wall (SOM 3a), rather than reflecting selective replacement by silica during diagenesis. The wall of Platysolenites antiquissimus consists of up to 5 µm crystals displaying the same elemental composition (SOM 3).
Based on these results, Blastulospongia is herein interpreted as a silicified and probably agglutinating organism similar in composition to Platysolenites. They may represent unilocular tests of benthic or semi-bentic foraminiferans. Some were probably sessile because of the flattened hemispherical shape and evidence of attachment (i.e., B. polytreta from the Shuijingtuo Formation; see Conway Morris and Chen 1990: fig. 4.5, 4.6).
Stratigraphic and geographic range.—Blastulospongia is originally known from the middle and upper Cambrian of Australia (Pickett and Jell 1983; Bengtson 1986). It was subsequently reported from the Terreneuvian Series of South China (Conway Morris and Chen 1990), Siberia (Val’kov 1987; Khomentovsky et al. 1990; Vasil’eva 1998), and Laurentia (Pyle et al. 2006).
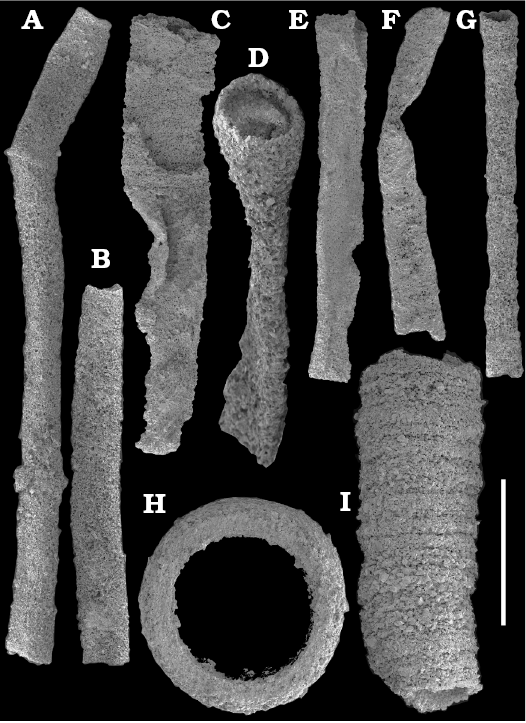
Fig. 81. Foraminifer? Platysolenites antiquissimus Eichwald, 1860, siliceous tubes from early Cambrian Nemakit-Daldyn (A, B, E), Medvezhya (D, F–I), and Emyaksin (C) formations, Anabar Uplift, Siberia, Russia; samples K1a/26A (A, B), K1a/28A (F), section 2; 5a/1 (C), section 96-5a; 1/A (E), 1/22 (H, I), section 1; K2/21 (D, G), section 3. A–I. SMNH X6056–6064, respectively. Scale bar 500 µm (D, H, I), 1 mm (A–C, E–G).

Fig. 82. Globular and other problematic fossils from early Cambrian Nemakit-Daldyn (D), Manykay (E), and Medvezhya (A–C, F–H) formations, Anabar Uplift, Siberia, Russia; samples 1/26.3 (A, F), 1/22.5 (B, H), 1/22 (C), 1/50 (G), section 1; 1282 (D), section 2; 4/4 (E), section 96-4. A, B. Blastulospongia sp., siliceous tests with terrigenous material (agglutinated?), SMNH X6065, 6066, respectively. B1, general view; B2, B3, close-ups of the outer surface. C–F. Undetermined phosphatized spheroids, SMNH X6067–6070, respectively. G. Archaeooides granulatus Qian, 1977, phosphatized, SMNH X6071. H. Platysolenites antiquissimus Eichwald, 1860, siliceous tube, SMNH X6072; H1, general view of the tube; H2, close-up of the outer surface. Scale bar 20 µm (B3, H2), 60 µm (B2), 300 µm (A, B1, C–G), 600 µm (H1).
Problematicum
Fig. 83.
Material.—SMNH X6073, from sample 3/10 (section 3, Fig. 3), Medvezhya Formation, western Anabar Uplift, Siberian Platform, Russia. Correlated with the uppermost Fortunian Stage.
Description.—The fossil is a hollow, cap-like phosphatic crust, 0.1–0.5 mm thick, with a radially folded surface that is elliptically shaped in upper view. The major and minor axes of the ellipse measure ca. 2.6 and 1.9 mm, respectively, and the structure is 1.2–1.5 mm tall. The fossil is interpreted herein as a phosphatized cuticular fragment of the external surface of an animal with uncertain affinity. Fossilization occurred probably during a post-embryonic stage, because the fragment is relatively large compared to phosphatized globular embryos known from the lower Cambrian and lacks enveloping egg membranes (e.g., Dong et al. 2013; Steiner et al. 2014).
The fossil apparently shows an original bilateral symmetry with the plane of symmetry cutting dorso-ventrally (?) along the minor axis. This symmetry plane separates eight radial lobes on each side of the fossil. The bilaterally symmetrical appearance may be due to diagenetic deformation, although it may also reflect an original biradially symmetrical oral (?) surface. The lobes feature radial and transverse striation.
Remarks.—The fossil is interpreted as a fragment of cuticularised structure similar to that known from olivooids from the Anabarites trisulcatus–Protohertzina anabarica Assemblage Zone of South China (Steiner et al. 2014). The life-cycle of such animals was initially reconstructed by Bengtson and Yue (1997) and Yue and Bengtson (1999), who suggested that it represents the larval development of directly developing cnidarians that are probably best assigned to stem group scyphozoans, an interpretation that was further supported by the discovery of associated pentaradial ephyra-like fossils (Dong et al. 2013). Fossilized embryos with tetraradial lobes at one pole were also described by Kouchinsky et al. (1999) from the lower Fortunian middle Manykay Formation at Bol’shaya Kuonamka (section 96-4) and interpreted as cnidarian embryos with eight incipient tentacles (Bengtson and Kouchinsky 2009).
Steiner et al. (2014) drew attention to the similarity of these alleged tentacles to the tetramerous structures in the embryonic stages of olivooids (i.e., Quadrapyrgites). Steiner et al. (2004a, b, 2014) reinterpreted olivooids as stem-group cycloneuralians with tetramerous or pentamerous oral structures in embryos and tubular hatchlings, similar to those in priapulids. Thus, they represent non-feeding priapulid-type larvae possibly undergoing metamorphosis through successive molt stages. A cnidarian affinity was, however, maintained by Liu et al. (2014) and Dong et al. (2016). Olivooid tubular hatchlings have a closed oral end with 12 (Quadrapyrgites), 10 (Olivooides), or 15 (a potential new species of olivooid) radial oral lobes (Steiner et al. 2014). A similar radially folded structure is also visible on Eolarva Zhang and Dong, 2015, from the same beds of South China. Eolarva was interpreted, however, as a cnidarian-grade organism that show indirect development (Zhang and Dong 2015). Although having a generally radially symmetrical arrangement of its 16 lobes, the fossil discussed herein lacks, however, an obvious pentamerous or tetramerous radial symmetry of the type known from olivooids and is larger than the known oral structures in olivooids.

Fig. 83. Problematicum, phosphatized external surface of an undetermined organism (SMNH X6073) from early Cambrian Medvezhya Formation, western Anabar Uplift, Siberia, Russia; sample 3/10, section 3. A. Upper view (on the oral? surface). B–D. Lateral views. Scale bar 500 µm (A), 1 mm (B–D).
Conclusions
The skeletal microstructural and morphological details of “small shelly fossils” are described in this report mainly from the Fortunian Stage. The variety of topotype Protohertzina anabarica sclerites and Anabarites trisulcatus tubes is documented. The recovery of Selindeochrea tripartita with longitudinal keels shows the clear distinction between Selindeochrea spp. anabaritids and Anabarites spp. Blastulospongia is preserved as siliceous spheres, which may have been agglutinated and sharing protistan affinity with Platysolenites. The latter is common in siliciclastic deposits of Baltica and is first described and shown in this report to be continuously distributed through the Fortunian carbonate sections of the Siberian Platform. The maikhanellids, Purella and Yunnanopleura are reinterpreted to be parts of the same scleritome. Fomitchella sclerites demonstrate incremental growth of the inner layer at the edge of the aperture similar to that in Lapworthella- and Camenella-like tommotiids. The outer layer with a longitudinal fibrous microstructure and minute pores in the inner layer of Fomitchella may indicate the presence of setae at the edge of the sclerite. Probable stem-group brachiopods, the tianzhushanellids (lathamellids), are now described from lower part of Cambrian Stage 2 of Siberia. Along with the preservation of fine microstructure details in a number of skeletal fossils, the phosphatisation of the cuticular surface of a problematicum (Fig. 83) and intricate details of globular fossils interpreted as cnidarian embryos (Kouchinsky et al. 1999; Bengtson and Kouchinsky 2009) demonstrate that the flanks of Anabar Uplift contain Fortunian Age localities with exceptionally preserved skeletal parts and body fossils.
The lower Cambrian fossil assemblages described in this report belong to the Fortunian and lower part of Cambrian Stage 2 of the Terreneuvian Series on the northern Siberian Platform. They record the first phase of the early Cambrian radiation of bilaterians (Kouchinsky et al. 2012) that was accompanied by biomineralization among emerging lophotrochozoan groups. The widespread biomineralization was preceded by the onset of significantly deeper and behaviorly more complex bioturbation than in the upper Ediacaran (Rogov et al. 2012; Brasier et al. 2013). This more intense bioturbation first appears on the Siberian Platform in the Anabarites trisulcatus Zone (this paper). Although common or morphologically close fossil species occur in the Terreneuvian Series of western Mongolia (Esakova and Zhegallo 1996), Avalonia (Landing et al. 2013), and such Gondwanan regions as Iran (Hamdi 1995), France (Devaere et al. 2013), and South China (Yang et al. 2014; Yu 2014), the difference in taxonomic composition between these and Siberian assemblages can be explained by their palaeogeographic separation as well as by the limited dispersal capabilities and rapid evolutionary radiation of the constituent organisms.
The lower and upper parts of the Fortunian Stage are represented on the Siberian Platform by the Anabarites trisulcatus and Purella zones, respectively. Transition between them is probably marked by δ13Ccarb excursion Z. As the horizon defining the global lower Cambrian Stage 2 boundary is not formally decided by the ICS and IUGS, we suggest that two alternative boundary horizons, that involve the range of Watsonella crosbyi, are available in Siberia as documented in this report and by Landing and Kouchinsky (2016). These can be defined either as (i) a horizon in the Ary-Mas-Yuryakh section (section 1 herein) defined by the peak of the δ13Ccarb excursion Iʹ above the lowest Watsonella crosbyi specimens (e.g., Landing et al. 2013) or at (ii) a lower level at Ary-Mas-Yuryakh defined by the horizon with the lowest W. crosbyi specimens below the peak of the excursion Iʹ. The stratigraphic interval between these two horizons is herein designated as the “Fortunian–Cambrian Stage 2 transitional beds”. Excursions Iʹ and 5p from northern Siberian Platform and excursion I known from southern Siberian Platform likely represent the same δ13Ccarb excursion expressed in different facies belts with different assemblages and preservation of fossils.
Cambrian Stage 2 is characterised in its lower part by the lowest known appearances of such fossils as orthothecid hyoliths with a relatively complex morphology of the inner surface of the opercula and tianzhushanellids (lathamellids). The absence of brachiopods (paterinids) and reliably reported bimembrate tommotiids (such as Camenella) and hyolithid hyoliths represent a noticeable difference of upper Fortunian‒lower part of Cambrian Stage 2 biotas from those of the overlying deposits of upper part of Cambrian Stage 2‒Cambrian Stage 3 in Siberia (Kouchinsky et al. 2012, 2015a) and globally in coeval rock units (Landing 1994; Landing et al. 2013; Landing and Kouchinsky 2016). By contrast, the younger deposits differ in the rarity or even complete absence of maikhanellids, siphogonuchitids and anabaritids. The lack of biomineralized ecdysozoan remains in the Terreneuvian interval studied in this report contrasts with the overlying Cambrian Stage 3 (Atdabanian–lower Botoman) strata of the Emyaksin Formation where they are common (Kouchinsky et al. 2015a). The upper part of Cambrian Stage 2‒Cambrian Stage 3 interval is correlated herein with the regional Tommotian‒lower Botoman stages as defined in their stratotypes on the Siberian Platform.
The upper boundary of Cambrian Stage 2 can conventionally be placed above the earliest Mobergella spp. and Rhombocorniculum insolutum and below the lowest known bivalved mollusc Fordilla and pelagiellid mollusc Pelagiella (Fig. 5). These Cambrian Stage 2‒Cambrian Stage 3 transitional beds are also situated between the lowest appearances of ecdysozoan mineralized remains in the fossil record (e.g., trilobites and bradoriids) and the first post-Terreneuvian maximum of the δ13Ccarb excursion above. The latter represents carbon isotope peak IV located in the Repinaella Zone of the Siberian Platform and middle part of the Emyaksin Formation (Landing et al. 2013; Kouchinsky et al. 2015a). Thus, Cambrian Stage 2 can be recognized by standard stratigraphic correlation techniques (i.e., biostratigraphy and carbon isotope chemostratigraphy) and evaluated temporally by geochronological data available from the lower Cambrian.
The Fortunian‒Cambrian Stage 2 transitional beds on the flanks of the Anabar Uplift are readily accessible in continuous sections with well-preserved and abundant fossils (SOM 5). In addition, carbon isotope chemostratigraphy can be successfully applied for precise correlation between these sections due to excellent preservation of the original δ13Ccarb record in normal marine carbonates of the Emyaksin and Medvezhya formations (Kouchinsky et al. 2001, 2015a; this paper). Carbon isotope chemostratigraphy can further be applied across the Siberian Platform with correlation by the younger Tommotian‒Botoman δ13Ccarb signature that is shared with the Lena-Aldan stratotype area in the southeastern Siberian Platform (Kouchinsky et al. 2001, 2005, 2007, 2012, 2015a). In general, the δ13Ccarb record through the entire Cambrian of Siberia can be most probably correlated globally (Maloof et al. 2005, 2010; Kouchinsky et al. 2007, 2008, 2012).
Acknowledgements
AK thanks Mikhail Drozhin (Saint Petersburg, Russia) for the company and assistance in the field trip to Kotuj in 2007. The work is dedicated to the memory of the late Vladimir V. Missarzhevsky and Anatolij K. Val’kov, who participated in the field trip to Bol’shaya Kuonamka in 1996. We are grateful to Vladimir Pavlov (Institute of Physics of the Earth, Moscow, Russia) for providing additional samples from the Kugda-Yuryakh Formation and to Bruce Runnegar and Edward Young (both Department of Earth and Space Sciences, University of California, Los Angeles, USA) for support, access to facilities and laboratory analyses of carbon isotopes. The authors thank Pavel Parkhaev (Paleontological Institute, Moscow, Russia) and Thomas Wotte (University of Cologne, Cologne, Germany) for detailed and critical reviews of the manuscript. AK is indebted to the Russian Academy of Sciences for the opportunity to join the field trip to the Kotuj River in 1991 as a student at the Moscow State University and in 1992 as a junior researcher at the Geological Institute (Moscow, Russia). The fieldwork was financially supported by grants from the Royal Swedish Academy of Sciences (KVA), the Swedish Natural Science Research Council (NFR), and the Swedish Research Council (VR). AK also acknowledges support from the Nordic Center for Earth Evolution project (grant DNRF53) and from the Swedish Research Council (grants to SB and Lars Holmer) during preparation of the material and writing of the manuscript. MS is grateful for financial support by Deutsche Forschungsgesellschaft (grant STE 814/5-1).
References
Abaimova, G.P. 1976. The oldest gastropods of Siberia [in Russian]. In: I.T. Žuravleva (ed.), Stratigrafiâ i paleontologiâ nižnego kembriâ SSSR. Trudy Instituta Geologii i Geofiziki SO AN SSSR 296: 174–175.
Antcliffe, J.B., Callow, R.H., and Brasier, M.D. 2014. Giving the early fossil record of sponges a squeeze. Biological Reviews of the Cambridge Philosophical Society 89 (4): 972–1004. Crossref
Azmi, R.J. 1996. Evidence for soft tissue basal support in earliest Cambrian protoconodonts from the Lesser Himalaya: conodont function and affinity. In: J. Pandey, R.J. Azmi, A. Bhandari, and A. Dave (eds.), Contributions to the XV Indian Colloqium on Micropaleontology and Stratigraphy, 457–461. Wadia Institute of Himalayan Geology, Dehradun.
Balthasar, U. 2008. Mummpikia gen. nov. and the origin of calcitic−shelled brachiopods. Palaeontology 51: 263–279. Crossref
Barrande, J. 1881. Systême Siluriené du centre de la Bohême, Vol. 6, Classe des Mollusques. Ordre des Acéphalés. 342 pp. Chez l’auteur et éditeur, Paris.
Bengtson, S. 1976. The structure of some Middle Cambrian conodonts, and the early evolution of conodont structure and function. Lethaia 9: 185–206. Crossref
Bengtson, S. 1983. The early history of the Conodonta. Fossils and Strata 15: 5–19.
Bengtson, S. 1986. Siliceous microfossils from the Upper Cambrian of Queensland. Alcheringa 10: 195–216. Crossref
Bengtson, S. 1992. The cap-shaped Cambrian fossil Maikhanella and the relationship between coeloscleritophorans and molluscs. Lethaia 25: 401–420. Crossref
Bengtson, S. and Collins, D. 2015. Chancelloriids of the Cambrian Burgess Shale. Palaeontologia Electronica 18.1.6A: 1–67. http://palaeo-electronica.org/content/2015/1031-chancelloriids
Bengtson, S. and Fletcher, T.P. 1983. The oldest sequence of skeletal fossils in the Lower Cambrian of southeastern Newfoundland. Canadian Journal of Earth Sciences 20: 525–536. Crossref
Bengtson, S. and Kouchinsky, A. 2009. Virtual dissection of an early cnidarian embryo. 9th North American Paleontological Convention, 21th–26th June, 123. University of Cincinnati, Cincinnati, Ohio.
Bengtson, S. and Yue, Z. 1997. Fossilized metazoan embryos from the earliest Cambrian. Science 277: 1645–1648. Crossref
Bengtson, S., Conway Morris, S., Cooper, B.J., Jell, P.A., and Runnegar, B.N. 1990. Early Cambrian fossils from South Australia. Association of Australasian Palaeontologists, Memoir 9: 1–364.
Bengtson, S., Fedorov, A.B., Missarzhevsky, V.V., Rozanov, A.Y., Zhegallo, E.A., and Zhuravlev, A.Y. 1987. Tumulduria incomperta and the case for Tommotian trilobites. Lethaia 20: 361–370. Crossref
Billings, E. 1872. On some new species of Palaeozoic fossils. The Canadian Naturalist and Quarterly Journal of Science. With Proceedings of the Natural History Society of Montreal 6: 213–222, 240.
Bokova, A.R. 1985. The oldest complex of the Cambrian organisms of western Anabar region [in Russian]. In: V.V. Homentovskij (ed.), Stratigrafiâ pozdnego dokembriâ i rannego paleozoâ Sibiri. Vend i rifej, 13–28. Institut Geologii i Geofiziki SO AN SSSR, Novosibirsk.
Bokova, A.R. 1990. New Lower Cambrian gastropods from the Siberian Platform. Paleontological Journal 24: 134–136.
Bokova, A.R. 1992. Biostratigrafiâ i skeletnye zooproblematiki nižnego kembriâ vostoka Sibirskoj platformy. 147 pp. Unpublished doctoral candidate thesis, Âkutskij institut geologičeskih nauk, SO AN SSSR.
Bokova, A.R. and Vasil’eva, N.I. 1990. Some new species of skeletal problematics from the Lower Cambrian of the Olenyok Uplift [in Russian]. Trudy Instituta Geologii i Geofiziki SO AN SSSR 783: 1–159.
Bowring, S.A., Grotzinger, J.P., Isachsen, C.E., Knoll, A.H., Pelechaty, S.M., and Kolosov, P. 1993. Calibrating rates of Early Cambrian evolution. Science 261: 1293–1298. Crossref
Brasier, M.D. 1986. The sequence of small shelly fossils (especially conoidal microfossils) from English Precambrian–Cambrian boundary beds. Geological Magazine 123: 237–256. Crossref
Brasier, M.D. 1989. Towards a biostratigraphy of the earliest skeletal biotas. In: J.W. Cowie and M.D. Brasier (eds.), The Precambrian–Cambrian Boundary, 117–165. Clarendon Press, Oxford.
Brasier, M.D. and Singh, P. 1987. Microfossils and Precambrian–Cambrian boundary stratigraphy at Maldeota, Lesser Himalaya. Geological Magazine 124: 323–345. Crossref
Brasier, M.D., Khomentovsky, V.V., and Corfield, R.M. 1993. Stable isotopic calibration of the earliest skeletal fossil assemblages in eastern Siberia (Precambrian–Cambrian boundary). Terra Nova 5: 225–232. Crossref
Brasier, M.D., Magaritz, M., Corfield, R., Luo, H., Wu, X., Ouyang, L., Jiang, Z., Hamdi, B., He, T., and Frazier, A.G. 1990. The carbon- and oxygen-isotopic record of the Precambrian–Cambrian boundary interval in China and Iran and their correlation. Geological Magazine 127: 319–332. Crossref
Brasier, M.D., McIlroy, D., Liu, A.G., Antcliffe, J.B., and Menon, L.R. 2013. The oldest evidence of bioturbation on Earth: comment. Geology 41: e289. Crossref
Brasier, M.D., Rozanov, A.Y., Zhuravlev, A.Y., Corfield, R.M., and Derry, L.A. 1994. A carbon isotope reference scale for the Lower Cambrian succession in Siberia: report of IGCP project 303. Geological Magazine 131: 767–783. Crossref
Brasier, M.D., Shields, G., Kuleshov, V.N., and Zhegallo, E.A. 1996. Integrated chemo- and biostratigraphic calibration of early animal evolution: Neoproterozoic–early Cambrian of southwest Mongolia. Geological Magazine 133: 445–485. Crossref
Butterfield, N.J. 2006. Hooking some stem-group “worms”: fossil lophotrochozoans in the Burgess Shale. BioEssays 28: 1161–1166. Crossref
Chang, S., Feng, Q., Clausen, S., and Zhang, L. 2017. Sponge spicules from the lower Cambrian in the Yanjiahe Formation, South China: The earliest biomineralizing sponge record. Palaeogeography, Palaeoclimatology, Palaeoecology 474: 36–44. Crossref
Chen, M. 1979. Some skeletal fossils from the phosphatic sequence, early Lower Cambrian, South China [in Chinese]. Scientia Geologica Sinica 2: 187–189.
Chen, P. 1984. Discovery of Lower Cambrian Small Shelly Fossils from Jijiapo, Yichang, West Hubei and its significance [in Chinese with English abstract]. Professional Papers of Stratigraphy and Palaeontology 13: 49–64.
Chen, Y. and Zhang, S. 1980. Small shelly fossils from the early Lower Cambrian, Sonlingpo, eastern Yangtze Gorges [in Chinese]. Geological Review 26: 190–197.
Cobbold, E.S. 1921. The Cambrian horizons of Comley (Shropshire) and their Brachiopoda, Pteropoda, Gasteropoda, etc. Quarterly Journal of the Geological Society 76: 325–386. Crossref
Cobbold, E.S. 1935. Lower Cambrian from Herault, France. The Annals and Magazine of Natural History 16: 25–48. Crossref
Compston, W., Zhang, Z., Cooper, J.A., Ma, G., and Jenkins, R.J.F. 2008. Further SHRIMP geochronology on the early Cambrian of South China. American Journal of Science 308: 399–420. Crossref
Conway Morris, S. and Caron, J.-B. 2007. Halwaxiids and the early evolution of lophotrochozoans. Science 315: 1255–1258. Crossref
Conway Morris, S. and Chen, M. 1990. Blastulospongia polytreta n. sp., an enigmatic organism from the Lower Cambrian of Hubei, China. Journal of Paleontology 64: 26–30. Crossref
Conway Morris, S. and Fritz, W.A. 1980. Shelly microfossils near the Precambrian–Cambrian boundary, Mackenzie Montains, northwestern Canada. Nature 286: 381–384. Crossref
Conway Morris, S. and Peel, J.S. 1995. Articulated halkieriids from the Lower Cambrian of North Greenland and their role in early protostome evolution. Philosophical Transactions of the Royal Society of London B 347: 305–358. Crossref
Cuvier, G. 1797. Tableau élementaire de l’historie naturelle des animaux. 710 pp. Baudouin, Paris.
d’Orbigny, A.D. 1826. Tableau méthodique de la classe des Céphalopodes: Annales des Sciences Naturelles, sér. 1 7: 245–314.
Datsenko, V.A. [Dacenko, V.A.], Zhuravleva, I.T. [Žuravleva, I.T.], Lazarenko, N.P., Popov, Y.N. [Popov, Û.N.], and Chernysheva, N.E. [Černyševa, N.E.] 1968. Biostratigraphy and fauna of the Cambrian deposits of north-western Siberian platform [in Russian]. Trudy NIIGA 155: 1–242.
De Blainville, H.M. 1816. Prodrome d’une nouvelle distribution systématique du règne animal. Bulletin de la Société Philomatique de Paris 8: 113–124.
Demidenko, Y.E. 2016. Morphology, taxonomic position, and stratigraphic distribution of the early Cambrian skeletal problematics Mobergella radiolata Bengtson, 1968. Paleontological Journal 50: 435–449. Crossref
Demidenko, Y.E. and Parkhaev, P.Y. 2014. On the problem of recognition of the lower Tommotian boundary using the SSF. In: R. Zhan and B. Huang (eds.), Extended Summary, IGCP Project 591 Field Workshop 2014, 26–31. Nanjing University Press, Nanjing.
Demokidov, K.K. 1956. General aspects of geological structure of the Lena-Olenyok region [in Russian]. Trudy NIIGA 89: 22–37.
Demokidov, K.K. 1958. On stratigraphic subdivision of Cambrian deposits of northern Siberian Platform [in Russian]. Trudy NIIGA 67: 3–12.
Demokidov, K.K. and Lazarenko, N.P. 1964. Stratigrafiâ verhnego dokembriâ i kembriâ i nižnekembrijskie trilobity severnoj časti Srednej Sibiri i ostrovov Sovetskoj Arktiki. 288 pp. Nedra, Moskva.
Devaere, L., Clausen, S., Steiner, M., Álvaro, J.J., and Vachard, D. 2013. Chronostratigraphic and palaeogeographic significance of an early Cambrian microfauna from the Heraultia Limestone, northern Montagne Noire, France. Palaeontologia Electronica 16 (2): 17A. http://palaeo-electronica.org/content/2013/298-french-cambrian-microfauna
Dong, X.-P., Cunningham, J.A., Bengtson, S., Thomas, C.-W., Liu, J., Stampanoni, M., and Donoghue, P.C.J. 2013. Embryos, polyps and medusae of the Early Cambrian scyphozoan Olivooides. Proceedings of the Royal Society B 280: 20130071. Crossref
Dong, X.-P., Vargas, K., Cunningham, J.A., Zhang, H., Liu, T., Chen, F., Liu, J., Bengtson, S., and Donoghue, P.C.J. 2016. Developmental biology of the early Cambrian cnidarian Olivooides. Palaeontology 59: 387–407. Crossref
Dyatlova, I.N. [Dâtlova, I.N.], Dronov, A.N., and Trofimov, V.R. 1990. New data on stratigraphy of the Lower Cambrian of western flanks of the Anabar Uplift [in Russian]. In: R.S. Matuhin and V.Y. Šenfil’ (eds.), Biostratigrafiâ i paleontologiâ kembriâ Severnoj Asii, 123–135. Nauka, Novosibirsk.
Dzik, J. 1994. Evolution of “small shelly fossils” assemblages of the Early Paleozoic. Acta Palaeontologica Polonica 39: 247–313.
Dzik, J. and Mazurek, D. 2013. Affinities of the alleged earliest Cambrian gastropod Aldanella. Canadian Journal of Zoology 91: 914–923. Crossref
Egorova, L.I. and Savitzky, V.E. [Savickij, V.E.] 1969. Stratigraphy and biofacies of the Cambrian of the Siberian Platform (western flanks of the Anabar Uplift) [in Russian]. Trudy SNIIGGIMS 43: 1–408.
Eichwald, E. 1860. Lethaea rossica ou palaeontologie de la Russie. 1657 pp. Schweizerbart, Stuttgart.
Elicki, O. and Gürsu, S. 2009. First record of Pojetaia runnegari Jell, 1980 and Fordilla Barrande, 1881 from the Middle East (Taurus Mountains, Turkey) and critical review of Cambrian bivalves. Paläontologische Zeitschrift 83: 267–291. Crossref
Esakova, N.V. and Zhegallo, E.A. [Žegallo, E.A.] 1996. Biostratigraphy and fauna of the Lower Cambrian of Mongolia [in Russian]. In: A.Û. Rozanov (ed.), Trudy Sovmestnoj Rossijsko-Mongol’skoj Paleontologičeskoj Ekspeditsii 46: 1–216.
Fedonkin, M.A. 1985. Palaeoichnology of the Vendian Metazoa [in Russian]. In: B.S. Sokolov and A.B. Ivanovskij (eds.), Vendskaâ sistema. Istoriko-geologičeskoe i paleontologičeskoe obosnovanie 1. Paleontologiâ, 112–117. Nauka, Moskva.
Fedorov, A.B. 1984. New representatives of the skeletal biota from stratotype sections of the Precambrian–Cambrian of the Siberian Platform (Aldan and Kotuj rivers) [in Russian]. In: V.S. Surkov (ed.), Novye vidy drevnih bespozvonočnyh i rastenij neftegazonosnyh provintsij Sibiri, 5–9. SNIIGGiMS, Novosibirsk.
Fedorov, A.B. and Shishkin, B.B. [Šiškin, B.B.] 1984. Lower Cambrian boundary in the northern part of the Siberian Platform [in Russian]. In: V.I. Krasnov (ed.), Problemy ârusnogo rasčleneniâ sistem fanerozoâ Sibiri, 5–14. Nauka, Novosibirsk.
Felitsyn, S. 1992. Rare element concentrations indicating the agglutinating character of Lower Cambrian Platysolenites tubes. Lethaia 25: 131–133. Crossref
Feng,W.M. and Sun,W.G. 2003. Phosphate replicated and replaced microstructure of molluscan shells from the earliest Cambrian of China. Acta Palaeontologica Polonica 48: 21–30.
Feng,W., Sun,W., and Qian, Y. 2001a. Skeletalization characters, classification and evolutionary significance of Early Cambrian monoplacophoran maikhanellids. Acta Palaeontologica Sinica 40: 195–213.
Feng, W., Xinan, M., and Kouchinsky, A. 2001b. Hyolith-type microstructure in a mollusc-like fossil from the Early Cambrian of Yunnan, China. Lethaia 34: 303–308. Crossref
Fisher, D.W. 1962. Other small conoidal shells. In: R.C. Moore (ed.), Treatise on Invertebrate Paleontology. Part W. Miscellanea, W98–W143. Geological Society of America, Boulder and University of Kansas Press, Lawrence.
Germs, G.L. 1972. New shelly fossils from the Nama Group, South West Africa. American Journal of Science 272: 752–761. Crossref
Geyer, G. 1994. Middle Cambrian mollusks from Idaho and early conchiferan evolution. In: E. Landing (ed.), Studies in Stratigraphy and Paleontology in Honor of Donald W. Fisher. Bulletin of the New York State Museum 481: 69–86.
Glaessner, M.F. 1978. The oldest Foraminifera. Australian Bureau for Mineral Research, Geology and Geophysics Bulletin 192: 61–65.
Golubev, S.N. 1976. Ontogenetic changes and evolutionary trends in Early Cambrian spiral gastropods of the superfamily Pelagiellacea. Paleontological Journal 10: 143–149.
Grabau, A.W. 1900. Pa1aeontology of the Cambrian terranes of the Boston Basin. Occasional Papers of the Boston Society of Natural History 4: 601–694.
Gravestock, D.I., Alexander, E.M., Demidenko, Y.E., Esakova, N.V., Holmer, L.E., Jago, J.B., Lin, T., Melnikova, L.M., Parkhaev, P.Y., Rozanov, A.Y., Ushatinskaya, G.T., Zang, W., Zhegallo, E.A., and Zhuravlev, A.Y. 2001. The Cambrian biostratigraphy of the Stansbury Basin, South Australia. Transactions of the Palaeontological Institute 282: 1–344.
Gubanov, A.P. 1998. The Early Cambrian molluscan evolution and its palaeogeographic implications. Acta Universitatis Carolinae, Geologica 42: 419–422.
Gubanov, A.P. 2002. Early Cambrian palaeogeography and the Iberia–Siberia connection. Tectonophysics 352: 153–168. Crossref
Gubanov, A.P. and Peel, J.S. 1998. Redescription of the type species of Latouchella Cobbold, 1921 (Mollusca) from the Lower Cambrian of Comley (England). GFF 120: 17–20. Crossref
Gubanov, A.P. and Peel, J.S. 1999. Oelandiella, the earliest Cambrian helcionelloid mollusc from Siberia. Palaeontology 42: 211–222. Crossref
Gubanov, A.P. and Peel, J.S. 2003. The Early Cambrian helcionelloid mollusk Anabarella Vostokova. Palaeontology 46: 1073–1087. Crossref
Gubanov, A.P., Fernández Remolar, D.C., and Peel, J.S. 2004a. Early Cambrian molluscs from Sierra de Córdoba (Spain). Geobios 37: 199–215. Crossref
Gubanov, A.P., Kouchinsky, A., and Peel, J.S. 1999. The first evolutionary adaptive lineage within fossil molluscs. Lethaia 32: 155–157. Crossref
Gubanov, A.P., Kouchinsky, A., Peel, J.S., and Bengtson, S. 2004b. Middle Cambrian molluscs of “Australian” aspect from northern Siberia. Alcheringa 28: 1–20. Crossref
Hamdi, B. 1989. Stratigraphy and Palaeontology of the Late Precambrian to Early Cambrian in the Alborz Mountains, Northern Iran. Geological Survey of Iran, Report 59: 1–41.
Hamdi, B. 1995. Precambriam‒Cambrian deposits in Iran [in Farsi]. In: A. Hushmandzadeh (ed.), Treatise on the Geology of Iran 20: 1–303.
Hamdi, B., Brasier, M.D., and Jiang, Z. 1989. Earliest skeletal fossils from Precambrian–Cambrian boundary strata, Elburz Mountains, Iran. Geological Magazine 126: 283–289. Crossref
Hatschek, B. 1888. Lehrbuch der Zoologie: eine morphologische Übersicht des Thierreiches zur Einführung in das Studium dieser Wissenschaft. 450 pp. G. Fischer, Jena. Crossref
Hinz, I. 1987. The Lower Cambrian microfauna of Comley and Rushton, Shropshire/England. Palaeontographica A 198: 41–100.
Holm, G. 1893. Sveriges kambrisk-siluriska Hyolithidae och Conulariidae. Sveriges Geologiska Undersökning, Afhandlingar och Uppsatser C 112: 1–172.
Holzapfel, E. 1895. Das Obere Mitteldevon (Schichten mit Stringocephalus Burtini un Maeneceras terbratum) in Rheinischen Gebirge. Abhandlungen der Königlich Preussischen Geologischen Landesanstalt, Neue Folge 16: 1–460.
Hughes, N.C., Peng, S., Bhargava, O.N., Ahluwalia, A.D., Walia, S., Myrow, P.M., and Parcha, S.K. 2005. Cambrian biostratigraphy of the Tal Group, Lesser Himalaya, India, and early Tsanglangpuan (late early Cambrian) trilobites from the Nigali Dhar syncline. Geological Magazine 142: 57–80. Crossref
Isakar, M. and Peel, J.S. 2007. Lower Cambrian helcionelloid molluscs from Estonia. GFF 129: 255–262. Crossref
Isachsen, C.E., Bowring, S.A., Landing, E., and Samson, S.D. 1994. New constraint on the division of Cambrian time. Geology 22: 496–498. Crossref
Jacquet, S.M., Brougham, T., Skovsted, C.B., Jago, J.B., Laurie, J.R., Betts, M.J., and Topper, T.P. 2016. Watsonella crosbyi from the lower Cambrian (Terreneuvian, Stage 2) Normanville Group in South Australia. Geological Magazine 131: 767–783. Crossref
Jell, P. 1980. Earliest known pelecypod on Earth—a new Cambrian genus from South Australia. Alcheringa 4: 233‒239. Crossref
Jiang, Z. 1980. The Meishucun Stage and fauna of the jinning county, Yunnan [in Chinese, with English summary]. Bulletin of the Chinese Academy of Geological Sciences, Series I 2: 1, 75‒92.
Karlova, G.A. 1987. First finds of skeletal fauna in the Turkut Formation of the Olenyok Uplift [in Russian]. Doklady Akademii Nauk SSSR 292: 204–205.
Karlova, G.A. and Vodanyuk, S.A. [Vodanûk, S.A.] 1985. New data on the deposits transitional to the Cambrian from the Khorbosuonka River basin (Olenyok Uplift) [in Russian]. In: V.V. Homentovskij (ed.), Stratigrafiâ pozdnego dokembriâ i rannego paleozoâ Sibiri. Vend i rifej, 3–13. Institut Geologii i Geofiziki SO AN SSSR, Novosibirsk.
Kaufman, A.J., Knoll, A.H., Semikhatov, M.A., Grotzinger, J.P., Jacobsen, S.B., and Adams, W. 1996. Integrated chronostratigraphy of Proterozoic–Cambrian boundary beds in the western Anabar region, northern Siberia. Geological Magazine 133: 509–533. Crossref
Kaufman, A.J., Peek, S., Martin, A.J., Cui, H., Grazhdankin, D., Rogov, V., Xiao, S., Buchwaldt, R., and Bowring, S. 2012. A shorter fuse for the Cambrian Explosion? Geological Society of America Abstracts with Programs 44 (7): 326.
Kerber, M. 1988. Mikrofossilien aus unterkambrischen Gesteinen der Montagne Noire, Frankreich. Palaeontographica Abteilung A 202: 127–203.
Khomentovsky, V.V. [Homentovskij, V.V.] 2008. The Yudomian of Siberia, Vendian and Ediacaran Systems of the International Stratigraphic Scale. Stratigraphy and Geological Correlation 16: 581–598. Crossref
Khomentovsky, V.V. [Homentovskij, V.V.] and Gibsher, A.S. [Gibšer, A.S.] 1996. The Neoproterozoic–lower Cambrian in northern Govi-Altay, western Mongolia: regional setting, lithostratigraphy and biostratigraphy. Geological Magazine 133: 371–390. Crossref
Khomentovsky, V.V. [Homentovskij, V.V.] and Karlova, G.A. 1989. Vendian–Cambrian beds of the Dzhandy River section and their analogs in the East Siberian reference sections [in Russian]. In: V.V. Homentovskij, Û.K. Sovetov, V.Û. Šenfil’, M.S. Âkšin, and V.B. Morâkin (eds.), Pozdnij dokembrij i rannij paleozoj Sibiri. Aktual’nye voprosy stratigrafii, 23–61. IGiG SO AN SSSR, Novosibirsk.
Khomentovsky, V.V. [Homentovskij, V.V.] and Karlova, G.A. 1992. The Lower Cambrian boundary and its justification in Siberia [in Russian]. Geologiâ i Geofizika 11: 3–26.
Khomentovsky, V.V. and Karlova, G.A. 1993. Biostratigraphy of the Vendian–Cambrian boundary beds and the Lower Cambrian boundary in Siberia. Geological Magazine 130 (1): 25–45. Crossref
Khomentovsky, V.V. and Karlova, G.A. 1994. Pecularities of ecology of the Vendian–Cambrian small shelly biotas of the Siberian Platform. Stratigraphy and Geological Correlation 2 (3): 8–17.
Khomentovsky, V.V. and Karlova, G.A. 2002. The boundary between the Nemakit-Daldynian and Tommotian Stages (Vendian–Cambrian Systems) of Siberia. Stratigraphy and Geological Correlation 10: 217–238.
Khomentovsky, V.V. and Karlova, G.A. 2005. The Tommotian Stage base as the Cambrian lower boundary in Siberia. Stratigraphy and Geological Correlation 13: 21–34.
Khomentovsky, V.V. [Homentovskij, V.V.], Didenko, A.N., and Pyatiletov, V.G. [Pâtiletov, V.G.] 1982. General conclusions on stratigraphy of the Vendian of the western flanks of the Anabar Uplift [in Russian]. In: V.V. Homentovskij, M.S. Âkšin, and S.S. Bragin (eds.), Novye dannye po stratigrafii pozdnego dokembriâ Sibiri, 3–20. Institut Geologii i Geofiziki SO AN SSSR, Novosibirsk.
Khomentovsky, V.V., Fedorov, A.B., and Karlova, G.A. 1998. The lower Cambrian boundary in the inner areas of the north Siberian Platform. Stratigraphy and Geological Correlation 6: 293–302.
Khomentovsky, V.V. [Homentovskij, V.V.], Val’kov, A.K., and Karlova, G.A. 1990. New data on the biostratigraphy of transitional Vendian–Cambrian strata on the middle reaches of the Aldan River [in Russian]. In: V.V. Homentovskij and A.S. Gibšer (eds.), Pozdnij dokembrij i rannij paleozoj Sibiri. Voprosy regional’noj stratigrafii, 3–57. Institut Geologii i Geofiziki SO AN SSSR, Novosibirsk.
Khomentovsky, V.V. [Homentovskij, V.V.], Val’kov, A.K., Karlova, G.A., and Nuzhnov, S.V. [Nužnov, S.V.] 1983. Reference section of the Precambrian–Cambrian deposits on the Gonam River [in Russian]. In: V.V. Homentovskij, M.S. Âkšin, and G.A. Karlova (eds.), Pozdnij dokembrij i rannij paleozoj Sibiri. Vendskie otloženiâ, 29–44. Institut Geologii i Geofiziki SO AN SSSR, Novosibirsk.
Knoll, A.H., Grotzinger, J.P., Kaufman, A.J., and Kolosov, P. 1995a. Integrated approaches to terminal Proterozoic stratigraphy: An example from the Olenek Uplift, northeastern Siberia. Precambrian Research 73: 251–270. Crossref
Knoll, A.H., Kaufman, A.J., Semikhatov, M.A., Grotzinger, J.P., and Adams, W. 1995b. Sizing up the sub-Tommotian unconformity in Siberia. Geology 23: 1139–1143. Crossref
Kobayashi, T. 1939. Restudy of Lorenz’s Raphistoma bröggeri from Shantung with a note on Pelagiella. In: Jubilee Publication in Commemoration of Professor H. Yabe’s 60th Birthday, 283–288. Yabe kyoju kanreki kinen kai, Sendai.
Kontorovich, A.E., Varlamov, A.I., Grazhdankin, D.V., Karlova, G.A., Klets, A.G., Kontorovich, V.A., Saraev, S.V., Terleev, A.A., Belyaev, S.Y., Varaksina, I.V., Efimov, A.S., Kochnev, B.B., Nagovitsin, K.E., Postnikov, A.A., and Filippov, Y.F. 2008. A section of Vendian in the east of the West Siberian Plate (based on data from the Vostok 3 Borehole). Russian Geology and Geophysics 49: 932–939. Crossref
Kouchinsky, A. [Kučinskij, A.] 1994. Ichnofossils from the Manykay Formation (Lower Cambrian) in northwestern Anabar Uplift [in Russian]. Bûlleten’ Moskovskogo Obŝestva Ispytatelej Prirody, seriâ geologičeskaâ 69 (4): 55–62.
Kouchinsky, A. 1999. Shell microstructures of the Early Cambrian Anabarella and Watsonella as new evidence on the origin of the Rostroconchia. Lethaia 32: 173–180. Crossref
Kouchinsky, A. 2000a. Shell microstructures in Early Cambrian molluscs. Acta Palaeontologica Polonica 45: 119–150.
Kouchinsky, A. 2000b. Skeletal microstructures of hyoliths from the Early Cambrian of Siberia. Alcheringa 24: 65–81. Crossref
Kouchinsky, A. and Bengtson, S. 2002. The tube wall of Cambrian anabaritids. Acta Palaeontologica Polonica 47: 431–444.
Kouchinsky, A., Bengtson, S., and Gershwin, L. 1999. Cnidarian-like embryos associated with the first shelly fossils in Siberia. Geology 27: 609–612. Crossref
Kouchinsky, A., Bengtson, S., and Murdock, D. 2010. A new tannuolinid problematic from the lower Cambrian of the Sukharikha River in northern Siberia. Acta Palaeontologica Polonica 55: 321–331. Crossref
Kouchinsky, A., Bengtson, S., Clausen, S., and Vendrasco, M.J. 2015a. An early Cambrian fauna of skeletal fossils from the Emyaksin Formation, northern Siberia. Acta Palaeontologica Polonica 60: 421–512. Crossref
Kouchinsky, A., Bengtson, S., Clausen, S., Gubanov, A. P., Malinky, J.M., and Peel, J.S. 2011. A Middle Cambrian fauna of skeletal fossils from the Kuonamka Formation, northern Siberia. Alcheringa 35: 123–189. Crossref
Kouchinsky, A., Bengtson S., Feng W., Kutygin R., and Val’kov A. 2009. The Lower Cambrian fossil anabaritids: affinities, occurrences and systematics. Journal of Systematic Palaeontology 7: 241–298. Crossref
Kouchinsky, A., Bengtson, S., Gallet, Y., Korovnikov, I., Pavlov, V., Runnegar, B., Shields, G., Veizer, J., Young, E., and Ziegler, K. 2008. The SPICE carbon isotope excursion in Siberia: a combined study of the upper Middle Cambrian–lowermost Ordovician Kulyumbe River section, northwestern Siberian Platform. Geological Magazine 145: 609–622. Crossref
Kouchinsky, A., Bengtson, S., Missarzhevsky, V., Pelechaty, S., Torssander, P., and Val’kov, A. 2001. Carbon isotope stratigraphy and the problem of a pre-Tommotian Stage in Siberia. Geological Magazine 138: 387–396. Crossref
Kouchinsky, A., Bengtson, S., Pavlov, V., Runnegar, B., Torssander, P., Young, E., and Ziegler, K. 2007. Carbon isotope stratigraphy of the Precambrian–Cambrian Sukharikha River section, northwestern Siberian platform. Geological Magazine 144: 1–10. Crossref
Kouchinsky, A., Bengtson, S., Pavlov, V., Runnegar, B., Val’kov, A., and Young, E. 2005. Pre-Tommotian age of the lower Pestrotsvet Formation in the Selinde section on the Siberian platform: Carbon isotopic evidence. Geological Magazine 142: 319–325. Crossref
Kouchinsky, A., Bengtson, S., Runnegar, B., Skovsted, C., Steiner, M., and Vendrasco, M.J. 2012. Chronology of early Cambrian biomineralisation. Geological Magazine 149: 221–251. Crossref
Kouchinsky, A., Holmer, L.E., Steiner, M., and Ushatinskaya, G.T. 2015b. The new stem-group brachiopod Oymurania from the lower Cambrian of Siberia. Acta Palaeontologica Polonica 60: 963–980. Crossref
Landing, E. 1984. Skeleton of lapworthellids and the suprageneric classification of tommotiids (Early and Middle Cambrian phosphatic problematica). Journal of Paleontology 58: 1380–1398.
Landing, E. 1988. Lower Cambrian of eastern Massachusetts: stratigraphy and small shelly fossils. Journal of Paleontology 62: 661–695. Crossref
Landing, E. 1991. Upper Precambrian through Lower Cambrian of Cape Breton Island: faunas, paleoenvironments, and stratigraphic revision. Journal of Paleontology 65: 570–595. Crossref
Landing, E. 1993. In situ earliest Cambrian tube worms and the oldest metazoan-constructed biostrome (Placentian Series, Southeastern Newfoundland). Journal of Paleontology 67: 333–342. Crossref
Landing, E. 1994. Precambrian–Cambrian global stratotype ratified and a new perspective of Cambrian time. Geology 22: 179–182. Crossref
Landing, E. 1995. Upper Placentian‒Branchian Series of mainland Nova Scotia (middle‒upper Lower Cambrian): faunas, paleoenvironments, and stratigraphic revision. Journal of Paleontology 69: 475‒495. Crossref
Landing, E. 1996. Reconstructing the Avalon continent: marginal-to-inner platform transition in the Cambrian of Avalonian New Brunswick. Canadian Journal of Earth Sciences 33: 623–632.
Landing, E. 2004. Precambrian–Cambrian boundary interval deposition and the marginal platform of the Avalon microcontinent. Journal of Geodynamics 37: 411–435. Crossref
Landing, E. and Brett, C.E. 1982. Lower Cambrian of eastern Massachusetts: microfaunal sequence and the oldest known borings. Geological Society of America Abstracts with Programs 14: 33.
Landing, E. and Geyer, G. 2012. Misplaced faith—limitations of the first appearance datum (FAD) in chronostratigraphy and proposal of more robust Lower Cambrian correlation standards. In: Y. Zhao, M.Y. Zhu, J. Peng, R.R. Gaines, and R.L. Parsley (eds.), Cryogenian–Ediacaran to Cambrian Stratigraphy and Paleontology of Guizhou, China. Journal of Guizhou University (Natural Sciences) 29 (Supplement 1): 170–171.
Landing, E. and Kouchinsky, A. 2016. Correlation of the Cambrian Evolutionary Radiation: geochronology, evolutionary stasis of earliest Cambrian (Terreneuvian) small shelly fossil (SSF) taxa, and chronostratigraphic significance. Geological Magazine 153: 750–756. Crossref
Landing, E. and Kruse, P.D. 2017. Integrated stratigraphic, geochemical, and paleontological late Ediacaran to early Cambrian records from southwestern Mongolia. Comment. Geological Society of America Bulletin 129: 1–15. Crossref
Landing, E. and Murphy, J.B. 1991. Uppermost Precambrian(?)–Lower Cambrian of mainland Nova Scotia: faunas, depositional environment, and stratigraphic revision. Journal of Paleontology 65: 382–396. Crossref
Landing, E. and Westrop, S.R. 2004. Environmental patterns in the origin and evolution and diversification loci of Early Cambrian skeletalized Metazoa: evidence from the Avalon microcontinent. In: J.H. Lipps and B. Wagoner (eds.), Neoproterozoic–Cambrian Biological Revolutions. Paleontological Society Paper 10: 93–105.
Landing, E., Bowring, S.A., Davidek, K., Westrop, S.R., Geyer, G., and Heldmaier, W. 1998. Duration of the Early Cambrian: U-Pb ages of volcanic ashes from Avalon and Gondwana. Canadian Journal of Earth Sciences 35: 329–338. Crossref
Landing, E., Geyer, G., and Bartowski, K.E. 2002. Latest Early Cambrian small shelly fossils, trilobites, and Hatch Hill dysoxic interval on the Quebec continental slope. Journal of Paleontology 76: 287‒305. Crossref
Landing, E., Geyer, G., Brasier, M.D., and Bowring, S.A. 2013. Cambrian Evolutionary Radiation: context, correlation, and chronostratigraphy–overcoming deficiencies of the first appearance datum (FAD) concept. Earth-Science Reviews 123: 133–172. Crossref
Landing, E., Myrow, P., Benus, A.P., and Narbonne, G.M. 1989. The Placentian Series: appearance of the oldest skeletalized faunas in southeastern Newfoundland. Journal of Paleontology 63: 739‒769. Crossref
Landing, E., Narbonne, G.M., Myrow, P., Benus, A.P., and Anderson, M.M. 1988. Faunas and depositional environments of the upper Precambrian through Lower Cambrian, southeastern Newfoundland. In: E. Landing, G.M. Narbonne, and P. Myrow (eds.), Trace Fossils, Small Shelly Fossils, and the Precambrian–Cambrian Boundary. New York State Museum Bulletin 463: 18–52.
Landing, E., Nowlan, G.S., and Fletcher, T.P. 1980. A microfauna associated with Early Cambrian trilobites of the Callavia Zone, norther Antigonish Highlands, Nova Scotia. Canadian Journal of Earth Sciences 17: 400–418. Crossref
Landing, E., Peng, S.C., Babcock, L.E., Geyer, G., and Moczydłowska-Vidal, M. 2007. Global standard names for the lowermost Cambrian series and stage. Episodes 30: 283–289.
Landing, E., Rushton, A.W.A., Fortey, R.A., and Bowring, S.A. 2015. Improved geochronologic accuracy and precision for the ICS chronostratigraphic charts: Examples from the late Cambrian–Early Ordovician. Episodes 38: 154–161.
Lendzion, K. 1977. First gastropod fauna from the Klimontovian Stage (Lower Cambrian) of southeastern Poland. Kwartalnik Geologiczny 21: 239–243.
Leuckart, R. 1854. Salpen und Verwandte. Zoologische Untersuchungen Giessen 2: 47–63.
Li, D., Ling, H.F., Jiang, S.Y., Pan, J.Y., Chen, Y.Q., Cai, Y.F., and Feng, H.Z. 2009. New carbon isotope stratigraphy of the Ediacaran–Cambrian boundary interval from SW China: implications for global correlation. Geological Magazine 146: 465–484. Crossref
Li, G. 2012. Diverse stem-group brachiopod fossils from the lower Cambrian of South China. 2012 Geological Society of America Annual Meeting in Charlotte. Abstracts with Programs 44: 326.
Li, G. and Chen, J. 1992. Early Cambrian cap-shaped lathamellids: their microstructures and systematic [in Chinese]. Acta Palaeontologica Sinica 31: 467–480.
Li, G., Steiner, M., Zhu, M., Zhu, X., and Erdtmann, B.-D. 2007. Early Cambrian fossil record of metazoans in South China: generic diversity and radiation patterns. Palaeogeography, Palaeoclimatology, Palaeoecology 254: 226–246. Crossref
Li, G., Zhang, Z., Hua, H., and Yang, H. 2014. Occurrence of the enigmatic bivalve fossil Apistoconcha in the lower Cambrian of southeast Shaanxi, North China Platform. Journal of Paleontology 88: 359–366. Crossref
Li, G., Zhao, X., Gubanov, A., Zhu, M., and Na, L. 2011. Early Cambrian Mollusc Watsonella crosbyi: A potential GSSP index fossil for the base of the Cambrian Stage 2. Acta Geologica Sinica 85: 309–319. Crossref
Linnarsson, J.G.O. 1871. Om några försteningar från Sveriges och Norges “Primordialzon”. Öfversikt af Kongliga Vetenskaps-Akademiens Förhandlingar 6: 789–796.
Linsley, R.M. and Kier, W.M. 1984. The Paragastropoda: A proposal for a new class of Paleozoic Mollusca. Malacologia 25: 241–254.
Liu, D. 1979. Earliest Cambrian brachiopods from southwest China [in Chinese]. Acta Palaeontologica Sinica 18: 505–511.
Liu, Y., Li, Y., Shao, T., Zhang, H., Wang, Q., and Qiao, J. 2014. Quadrapyrgites from the lower Cambrian of South China: growth pattern, post-embryonic development, and affinity. Chinese Science Bulletin 59: 4086–4095. Crossref
Luo, H., Jiang, Z., Wu, X., Song, X., Ouyang, L., Zhang, S., Luo, W., Xue, X., Li, P., Liang, Q., Xie, Y., and Li, X. 1982. The Sinian–Cambrian boundary in eastern Yunnan, China [in Chinese, with English summary]. 265 pp. People’s Publishing House, Yunnan.
Luchinina, V.A. [Lučinina, V.A.], Korovnikov, I.V., Sipin, D.P., and Fedoseev, A.V. 1997. Biostratigraphy of the Upper Vendian–Lower Cambrian Sukharikha River section (Siberian platform) [in Russian]. Geologiâ i geofizika 38: 1346–1358.
MacKinnon, D.I. 1985. New Zealand late Middle Cambrian molluscs and the origin of Rostroconchia and Bivalvia. Alcheringa 9: 65–81. Crossref
Magaritz, M. 1989. δ13C minima follow extinction events: A clue to faunal radiation. Geology 17: 337–340. Crossref
Magaritz, M., Holser, W.T., and Kirschvink, J. L. 1986. Carbon isotope events across the Precambrian/Cambrian boundary on the Siberian Platform. Nature 320: 258–259. Crossref
Magaritz, M., Latham, A.J., Kirschvink, J.L., Zhuravlev, A.Y., and Rozanov, A.Y. 1991. Precambrian–Cambrian boundary problem I: Carbon isotope correlations for Vendian and Tommotian time between Siberia and Morocco. Geology 19: 847–850. Crossref
Maloof, A.C., Porter, S.M., More, J.L., Dudás, F.Ö., Bowring, S.A., Higgins, J.A., Fike, D.A., and Eddy, M.P. 2010a. The earliest Cambrian record of animals and ocean geochemical change. Geological Society of America Bulletin 122: 1731–1774. Crossref
Maloof, A.C., Ramezani, J., Bowring, S.A., Fike, D.A., Porter, S.M., and Mazouad, M. 2010b. Constraints on early carbon cycling from the duration of the Nemakit-Daldynian–Tommotian boundary δ13C shift, Morocco. Geology 38: 623–626. Crossref
Maloof, A., Schrag, D., Crowley, J., and Bowring, S. 2005. An expanded record of early Cambrian carbon cycling from the Anti-Atlas margin, Morocco. Canadian Journal of Earth Sciences 42: 2195–2216. Crossref
Marek, L. 1963. New knowledge on the morphology of Hyolithes. Sborník geologickych ved, řada Paleontologie 1: 53–72.
Marek, L. 1966. New hyolithid genera from the Ordovician of Bohemia. Časopis národního muzea 135: 89–92.
Matthew, G.F. 1895. The Protolenus fauna. Transactions of the New York Academy of Sciences 14: 101‒153. Crossref
Matthews, S.C. and Missarzhevsky, V.V. 1975. Small shelly fossils of late Precambrian and early Cambrian age: a review of recent work. Journal of the Geological Society 131: 289–304. Crossref
McIlroy, D. and Szaniawski, H. 2000. A lower Cambrian protoconodont apparatus from the Placentian of southeastern Newfoundland. Lethaia 33: 95–102. Crossref
McIlroy, D., Green, O.R., and Brasier, M.D. 1994: The world’s oldest foraminiferans. Microscopy and Analysis 44: 13–15.
McIlroy, D., Green, O.R., and Brasier, M.D. 2001. Palaeobiology and evolution of the earliest agglutinated Foraminifera: Platysolenites, Spirosolenites and related forms. Lethaia 34: 13–29. Crossref
Mens, K. and Isakar, M. 1999. Facies distribution of early Cambrian molluscs in Estonia. Proceedings of the Estonian Academy of Sciences, Geology 48: 110–115.
Meshkova, N.P. [Meškova, N.P.] 1969. On the paleontological characteristics of the Lower Cambrian deposits of the Siberian Platform [in Russian]. In: I.T. Žuravleva (ed.), Biostratigrafiâ i paleontologiâ nižnego kembriâ Sibiri i Dal’nego Vostoka, 158–172. Nauka, Moskva.
Meshkova, N.P. [Meškova, N.P.] 1974. Colloquia on hyoliths and remains of problematic animals from the Precambrian–Cambrian transitional deposits of the Siberian Platform [in Russian]. Geologiâ i geofizika 2: 143–146.
Meshkova, N.P. [Meškova, N.P.], Nikolaeva, I.V., Kulikov, Y.P. [Kulikov, Û.P.], Zhuravleva, I.T. [Žuravleva, I.T.], Luchinina, V.A. [Lučinina, V.A.], Musatov, D.I., and Sidoras, S.D. 1976. Stratigraphy of transitional Precambrian–Cambrian deposits of northern Anabar Uplift [in Russian]. In: I.T. Žuravleva (ed.), Stratigrafiâ i paleontologiâ nižnego i srednego kembriâ SSSR, 3–22. Novosibirsk, Nauka.
Minaeva, M.A. 1985. Type section of the Kyndyn Group of the Cambrian on the flanks of the Anabar Uplift [in Russian]. Sovetskaâ geologiâ 1985 (1): 54‒64.
Minaeva, M.A. 1991. Types of sections through the Cambrian on the flanks of the Anabar Uplift and sedimentation environments [in Russian]. Sovetskaâ geologiâ 1991 (4): 36‒45.
Missarzhevsky, V.V. [Missarževskij, V.V.] 1967. Zonal’naâ stratigrafiâ drevnejših otloženij kembriâ Sibirskoj platformy. 23 pp. Unpublished Doktoral Candidate Thesis, Geological Institute AN SSSR, Moskva.
Missarzhevsky, V.V. [Missarževskij, V.V.] 1973. Conodontomorph organisms from the Precambrian‒Cambrian boundary beds of the Siberian Platform and Kazakhstan [in Russian]. In: I.T. Žuravleva (ed.), Problemy paleontologii i biostratigrafii nižnego kembriâ Sibiri i Dal’nego Vostoka, 53–58. Nauka, Novosibirsk.
Missarzhevsky, V.V. [Missarževskij, V.V.] 1974. New data on the oldest fossils of the Early Cambrian of the Siberian Platform [in Russian]. In: I.T. Žuravleva and A.Û. Rozanov (eds.), Biostratigrafiâ i paleontologiâ nižnego kembriâ Evropii i severnoi Azii, 179–189. Nauka, Novosibirsk.
Missarzhevsky, V.V. [Missarževskij, V.V.] 1977. Conodonts (?) and phosphatic problematica from the Cambrian of Mongolia and Siberia [in Russian]. In: L.P. Tatarinov (ed.), Bespozvonočnye paleozoâ Mongolii, 10–19. Nauka, Moskva.
Missarzhevsky, V.V. [Missarževskij, V.V.] 1981. Early Cambrian hyoliths and gastropods from Mongolia [in Russian] Paleontologičeskij žurna1 1981 (1): 21–28.
Missarzhevsky, V.V. [Missarževskij, V.V.] 1982. Subdivision and correlation of the Precambrian‒Cambrian boundary beds based on some ancient groups of skeletal organisms [in Russian]. Bûlleten’ Moskovskogo obŝestva ispytatelej prirody. Otdelenie geologii 57 (5): 52–67.
Missarzhevsky, V.V. [Missarževskij, V.V.] 1989. Oldest skeletal fossils and stratigraphy of Precambrian and Cambrian boundary beds [in Russian]. Trudy Geologičeskogo Instituta, Akademiâ Nauk SSSR 443: 1–237.
Missarzhevsky, V.V. [Missarževskij, V.V.] 1993. Precambrian–Cambrian boundary stratigraphy in the south-east of the Siberian Platform [in Russian]. Bûlleten’ Moskovskogo obŝestva ispytatelej prirody. Otdelenie geologii 68 (5): 60–71.
Missarzhevsky, V.V. [Missarževskij, V.V.] and Mambetov, A.M. 1981. Stratigraphy and fauna of the Cambrian and Precambrian boundary beds of the Maly Karatau Range [in Russian]. Trudy Geologičeskogo Instituta AN SSSR 326: 1–92.
Moczydłowska, M. 1991. Acritarch biostratigraphy of the Lower Cambrian and the Precambrian–Cambrian boundary in southeastern Poland. Fossils and Strata 29: 1–127.
Moore, J.L., Porter, S.M., Steiner, M., and Li, G. 2010. Cambrothyra ampulliformis, an unusual coeloscleritophoran from the Lower Cambrian of Shaanxi Province, China. Journal of Paleontology 84: 1040–1060. Crossref
Moore, J.L., Li, G., and Porter, S.M. 2014. Chancelloriid sclerites from the Lower Cambrian (Meishucunian) of eastern Yunnan, China, and the early history of the group. Palaeontology 57: 833–878. Crossref
Müller, K.J. 1959. Kambrische Conodonten. Deutsche Geologischce Gesellschaftung Zeitschrift 111: 434‒485.
Müller, K.J. 1962. Supplement to systematics of conodonts. In: R.C. Moore (ed.), Treatise on Invertebrate Paleontology, Part W, Miscellanea, W246‒W249. Geological Society of America, Boulder and University of Kansas Press, Lawrence.
Murphy, M.A. and Salvador, A. (eds.) 1999. International stratigraphic guide—an abridged edition. Episodes 22: 255–271.
Nagovitsin, K.E., Rogov, V.I., Marusin, V.V.,Karlova, G.A., Kolesnikov, A.V., Bykova, N.V., and Grazhdankin, D.V. 2015. Revised Neoproterozoic and Terreneuvian stratigraphy of the Lena-Anabar Basin and north-western slope of the Olenek Uplift, Siberian Platform. Precambrian Research 270: 226–245. Crossref
Novák, O. 1886. Zur Kenntniss der Fauna der Etage F-fl in der palaeozischen Schichtengruppe Böhmens. 27 pp. Aus den Sitzungsberichten der königlichen böhmens Gesellschaft der Wissenschaften, Böhmen.
Novák, O. 1891. Revision der palaeozoischen Hyolithiden Böhmens. Abhandlungen der mathematisch-naturwisschenschaftlichen Classe der böhmischen Gesellschaft der Wissenschaften, series 7 4: 1–48.
Nowlan, G.S., Narbonne, G.M., and Fritz, W.H. 1985. Small shelly fossils and trace fossils near the Precambrian–Cambrian boundary in the Yukon Territory, Canada. Lethaia 18: 233–256. Crossref
Öpik, A. 1926. Über den estländischen blauen Ton. Publications of the Geological Institution of the University of Tartu 6: 39–46.
Panasenko, E.S. 1998. Novye neobyčnye mikrofossilii iz pelagičeskih kremnistyh otloženij kembriâ. Dal’nevostočnyj geologičeskij institut, Rossijskaâ Akademiâ Hauk, Vladivostok. http://www.fegi.ru/primorye/geology/panas.htm
Pander, C.H. 1851. Sur une découverte de fossiles faite dans la partie inférieure du terrain silurien de Russie. Bulletin de la Société Géologique de France 8: 251–259.
Park, T., Woo, J., Lee, D.-J., Lee, D.-C., Lee, S., Han, Z., Chough, S.K., and Choi, D.K. 2011. A stem-group cnidarian described from the mid-Cambrian of China and its significance for cnidarian evolution. Nature Communications 2: 1–6. Crossref
Parkhaev, P.Y. 1998. Siphonoconcha—a new class of early Cambrian bivalved organisms. Paleontological Journal 32 (1): 1–15.
Parkhaev, P.Y. 2002. Muscle scars of Cambrian univalved mollusks and their systematic importance. Paläontologische Zeitschrift 5: 15–19.
Parkhaev, P.Y. 2006. New data on the morphology of the earliest gastropod mollusks of the genus Aldanella Vostokova, 1962 (Archaeobranchia, Pelagielliformes). Paläontologische Zeitschrift 3: 15–21.
Parkhaev, P.Y. 2007a. The Cambrian “basement” of gastropod evolution. Geological Society of London, Special Publications 286: 415–421. Crossref
Parkhaev, P.Y. 2007b. Shell chirality in Cambrian gastropods and sinistral members of the genus Aldanella Vostokova, 1962. Paleontological Journal 41: 233–240. Crossref
Parkhaev, P.Y. 2008. The Early Cambrian radiation of Mollusca. In: W.F. Ponder and D.R. Lindberg (eds.), Phylogeny and Evolution of the Mollusca, 33–69.University California Press, Berkeley. Crossref
Parkhaev, P.Y. and Demidenko, Y.E. 2010. Zooproblematica and Mollusca from the Lower Cambrian Meishucun section (Yunnan, China) and taxonomy and systematics of the Cambrian small shelly fossils of China. Paleontological Journal 44: 883–1161. Crossref
Parkhaev, P.Y. and Karlova, G.A. 2011. Taxonomic revision and evolution of Cambrian mollusks of the genus Aldanella Vostokova, 1962 (Gastropoda: Archaeobranchia). Paleontological Journal 45: 1145–1205. Crossref
Parkhaev, P.Y., Karlova, G.A., and Rozanov, A.Y. 2011. Taxonomy, stratigraphy and biogeography of Aldanella attleborensis—a possible candidate for defining the base of Cambrian Stage 2. Museum of Northern Arizona Bulletin 67: 298–300.
Pelechaty, S.M., Grotzinger, J. P., Kashirtsev, V. A., and Zhernovsky, V. P. 1996a. Chemostratigraphic and sequence stratigraphic constraints on Vendian–Cambrian basin dynamics, northeast Siberian Craton. Journal of Geology 104: 543–564. Crossref
Pelechaty, S.M., Kaufman, A.J., and Grotzinger, J.P. 1996b. Evaluation of δ13C chemostratigraphy for intrabasinal correlation: Vendian strata of northeast Siberia. GSA Bulletin 108: 992–1003. Crossref
Pel’man, Y.L. [Pel’man, Û.L.] 1977. Early and Middle Cambrian inarticulate brachiopods of the Siberian Platform [in Russian]. Trudy Instituta Geologii i Geofiziki AN SSSR, Sibirskoe Otdelnie 316: 1–168.
Pel’man Y.L. [Pel’man, Û.L.] 1983. Phylum Brachiopoda [in Russian.]. In: B.S. Sokolov and I.T. Žuravleva (eds.), Ârusnoe rasčlenenie nižnego kembriâ. Atlas okamenelostej. Trudy Instituta Geologii i Geofiziki AN SSSR, Sibirskoe Otdelenie 558: 147–153.
Pel’man, Y.L. [Pel’man, Û.L.], Ermak, V.V., Fedorov, A.B., Luchinina, V.A. [Lučinina, V.A.], Zhuravleva, I.T. [Žuravleva, I.T.], Repina, L.N., Bondarev, V.I., and Borodaevskaya, Z.V. [Borodaevskaâ, Z.V.] 1990. New data on stratigraphy and palaeontology of the upper Precambrian and lower Cambrian of the Dzhanda River (right tributary of the Aldan River) [in Russian.]. In: L.N. Repina (ed.), Biostratigrafiâ i paleontologiâ kembriâ Severnoj Azii, 3–32. Nauka, Novosibirsk.
Pel’man, Y.L. [Pel’man, Û.L.], Aksarina, N.A., Koneva, S.P. Popov, L.E., Sobolev, L.P., and Ushatinskaya, G. T. [Ušatinskaâ, G. T.]. 1992. Drevnejšie brahiopody territorii severnoj Evrazii. 145 pp. OIGGiM RAN, Novosibirsk.
Peng, S.C. and Babcock, L.E. 2011. Continuing progress on chronostratigraphic subdivision of the Cambrian System. Czech Bulletin of Geosciences 86: 391–396. Crossref
Pickett, J. and Jell, P.A. 1983. Middle Cambrian Sphinctozoa (Porifera) from New South Wales. Memoirs of the Association of Australasian Palaeontologists 1: 85–92.
Pojeta, J., Jr., and Runnegar, B. 1976. The paleontology of rostroconch molluscs and the early history of the phylum Mollusca. U.S. Geological Survey Professional Paper 968: 1–88.
Ponder, W.F., Parkhaev, P.Y., and Beechey, D.L. 2007. A remarkable similarity in scaly shell structure in early Cambrian univalved limpets (Monoplacophora; Maikhanellidae) and a recent fissurellid limpet (Gastropoda: Vetigastropoda) with a review of Maikhanellidae. Molluscan Research 27: 153–163.
Poulsen, C. 1967. Fossils from the Lower Cambrian of Bornholm. Danske Videnskabernes Selskab, Matematisk-Fysiske Meddelelser 36 (2): 1–46.
Pyle, L.J., Narbonne, G.M., Nowlan, G.S., Xiao, S., and James, N.P. 2006. Early Cambrian metazoan eggs, embryos, and phosphatic microfossils from northwestern Canada. Journal of Paleontology 80: 811–825. Crossref
Qian, Y. 1977. Hyolitha and some problematica from the Lower Cambrian Meichucun Stage in central and SW China. Acta Palaeontologica Sinica 16: 255–278.
Qian, Y. 1978. The Early Cambrian hyolithids in central and southwest China and their stratigraphical significance. Memoirs of Nanjing Institute of Geology and Palaeontology. Academia Sinica 11: 1–50.
Qian, Y. and Bengtson, S. 1989. Palaeontology and biostratigraphy of the Early Cambrian Meishucunian Stage in Yunnan Province, south China. Fossils and Strata 24: 1–156.
Qian, Y. and Yin, G. 1984. Small shelly fossils from the lowest Cambrian in Guizhou. Professional Papers of Stratigraphy and Palaeontology 13: 91–124.
Qian, Y. and Zhang, S.-B. 1983. Small shelly fossils from the Xihaoping Member of the Tongying Formation in Fangxian County of Hubei Province and their stratigraphical significance [in Chinese]. Acta Palaeontologica Sinica 22: 82–94.
Qian, Y., Chen, M., and Chen, Y. 1979. Hyolithids and other small shelly fossils from the Lower Cambrian Huangshandong Formation in the eastern part of the Yangtze Gorge [in Chinese]. Acta Palaeontologica Sinica 18: 207–232.
Qian, Y., Li, G., and Zhu, M. 2001. The Meishucunian Stage and its small shelly fossils sequence in China. Acta Paleontologica Sinica 40 (Supplement): 54–62.
Repina, L.N., Borodaevskaya, Z.V. [Borodaevskaâ, Z.V.], and Ermak, V.V. 1988. Reference section of the Selinde river (southeastern margin of the Aldan shield) [in Russian]. Trudy Instituta Geologii i Geofiziki AN SSSR, Sibirskoe Otdelnie 720: 3–31.
Ripperdan, R. 1994. Global variations in carbon isotope composition during the latest Neoproterozoic and earliest Cambrian. Annual Review of Earth and Planetary Sciences 22: 385–417. Crossref
Rogov, V.I., Karlova, G.A., Marusin, V.V., Kochnev, B.B., Nagovitsin, K.E., and Grazhdankin, D.V. 2015. Duration of the first biozone in the Siberian hypostratotype of the Vendian. Russian Geology and Geophysics 56: 501–511. Crossref
Rogov, V.I., Marusin, V., Bykova, N., Goy, Yu., Nagovitsin, K., Kochnev, B., Karlova, G., and Grazhdankin, D. 2012. The oldest evidence of bioturbation on Earth. Geology 40: 395–398. Crossref
Rowland, S.M., Luchinina, V.A., Korrovnikov, I.V., Sipin, D.P., Tarletskov, A., and Fedoseev, A.V. 1998. Biostratigraphy of the Vendian–Cambrian Sukharikha River section, northwestern Siberian Platform. Canadian Journal of Earth Sciences 35: 339–352. Crossref
Rozanov, A.Y. [Rozanov, A.Û.] and Missarzhevsky, V.V. [Missarževskij, V.V.] 1966. Biostratigraphy and fauna of the lower strata of the Cambrian [in Russian]. Trudy Geologičeskogo Instituta AN SSSR 148: 1–126.
Rozanov, A.Y. [Rozanov, A.Û.] and Sokolov, B.S. (eds.) 1984. Ârusnoe rasčlenenie nižnego kembriâ. Stratigrafiâ. 184 pp. Nauka, Moskva.
Rozanov, A.Y. and Zhuravlev, A.Y. 1992. The Lower Cambrian fossil record of the Soviet Union. In: J.H. Lipps and P.W. Signor (eds.), Origin and Early Evolution of the Metazoa, 205–82. Plenum, New York. Crossref
Rozanov, A.Y., Khomentovsky, V.V., Shabanov, Y.Y., Karlova, G.A., Varlamov, A.I., Luchinina, V.A., Pegel’, T.V., Demidenko, Y.E., Parkhaev, P.Y., Korovnikov, I.V., and Skorlotova, N.A. 2008. To the problem of stage subdivision of the Lower Cambrian. Stratigraphy and Geological Correlation 16: 1–19.
Rozanov, A.Y. [Rozanov, A.Û.], Missarzhevsky, V.V. [Missarževskij, V.V.], Volkova, N.A., Voronova, L.G., Krylov, I.N., Keller, B.M., Korolyuk, I.K. [Korolûk, I.K.], Lendzion, K., Michniak, R., Pyhova, N.G., and Sidorov, A.D. 1969. The Tommotian Stage and the Cambrian lower Boundary problem [in Russian]. Trudy Geologičeskogo Instituta, Akademiâ Nauk SSSR 206: 1–380.
Rozanov, A.Y. [Rozanov, A.Û.], Parkhaev, P.Y. [Parhaev, P.Y.], Demidenko, Y.E. [Demidenko, Û.E.], Karlova, G.A., Korovnikov, I.V., Shabanov, Y.Y. [Šabanov, Û.Û.], Ivantsov, A.Y. [Ivancov, A.Û.], Luchinina, V.A. [Lučinina, V.A.], Malakhovskaya, Y.E. [Malahovskaâ, Û.E.], Mel’nikova, L.M., Naimark, E.B., Ponomarenko, A.G., Skorlotova, N.A., Sundukov, V.M., Tokarev, D.A., Ushatinskaya, G.T. [Ušatinskaâ, G.T.], and Kipriyanova, L.D. [Kipriânova, L.D.] 2010. Iskopaemye stratotipov ârusov nižnego kembriâ. 228 pp. PIN RAN, Moskva.
Rozanov, A.Y., Parkhaev, P.Y, Demidenko, Y.E., and Skorlotova, N.A. 2011. Mobergella radiolata—a possible candidate for defining the base of Cambrian Series 2 and Stage 3. Museum of Northern Arizona Bulletin 67: 304–306.
Rozanov, A.Y. [Rozanov, A.Û.], Repina, L.N., Apollonov, M.K., Shabanov, Y.Y. [Šabanov, Û.Â], Zhuravlev, A.Y. [Žuravlev, A. Û.], Pegel’, T.V., Fedorov, A.B., Astashkin, V.A. [Astaškin, V.A.], Zhuravleva, I.T. [Žuravleva, I.T.], Egorova, L.I., Chugaeva, M.N. [Čugaeva, M.N.], Dubinina, S.V., Ermak, V.V., Esakova, N.V., Sundukov, V.V., Sukhov, S.S. [Suhov, S.S.], and Zhemchuzhnikov, V.G. [Žemchuzhnikov, V.G.] 1992. Kembrij Sibiri. 135 pp. Nauka, Novosibirsk.
Runnegar, B. 1981. Muscle scars, shell form and torsion in Cambrian and Ordovician molluscs. Lethaia 14: 311–322. Crossref
Runnegar, B. and Jell, P.A. 1980. Australian Middle Cambrian molluscs: corrections and additions. Alcheringa 4: 111–113. Crossref
Sarmiento, G.N., Fernández-Remolar, D., and Göncüoglu, M.C. 2001. Cambrian small shelly fossils from the Çal Tepe Formation, Taurus Mountains, Turkey. Coloquios de Paleontología 52: 117–134.
Savitsky, V.E. [Savickij, V.E.] 1962. New data on stratigraphy of the Aldanian Stage of the Anabar anticlyse and its boundary with the Precambrian [in Russian]. Informacionnyj sbornik Instituta Geologii Arktiki 27: 14–21.
Savitsky, V.E. [Savickij, V.E.], Shabanov, Y.Y. [Šabanov, Û.Â.], Evtushenko, V.M. [Evtušenko, V.M.], and Shishkin, B.B. [Šiškin, B.B.] 1972. Key section of the Cambrian of the Khorbosuonka facial region (north-east of the Siberian Platform). Material to the stratigraphy and palaeontology of the Siberian Platform [in Russian]. Trudy SNIIGiMS 32: 53–60.
Salvador, A. (ed.). 1994. International Stratigraphic Guide. A Guide to Stratigraphic Classification, Terminology, and Procedure. Second Edition. 214 pp. International Union of Geologists and the Geological, Society of America, Boulder, Colorado.
Schmitz, M.D. 2012, Radiometric ages used in GTS 2012. In: F.M. Gradstein, J.G. Ogg, M.D. Schmitz, and G.M. Ogg (eds.), The Geologic Time Scale. Volume 2, 1045–1082. Elsevier, Amsterdam. Crossref
Semikhatov, M.A. [Semihatov, M.A.] and Serebryakov, S.N. [Serebrâkov, S.N.] 1983. The Siberian hypostratotype of the Riphean [in Russian]. Trudy Geologisčeskogo Instituta AN SSSR 210: 1–207.
Shaler, N.S. and Foerste, A.F. 1888. Preliminary description of North Attleborough fossils. Bulletin of the Museum of Comparative Zoology 16: 27–41.
Shishkin, B.B. [Šiškin, B.B.] 1974. Shelly fauna in the Nemakit-Daldyn Formation (northwestern Anabar Uplift) [in Russian]. Geologiâ i geofizika 4: 111–114.
Shishkin, B.B. [Šiškin, B.B.] 1981. New data on stratigraphy of the early Cambrian deposits of the northeastern flank of the Anabar Uplift (Bol’shaya Kuonamka River) [in Russian]. In: N.P. Meškova and I.V. Nikolaeva (eds.), Pograničnye otloženiâ dokembriâ i kembriâ Sibirskoj platformy (biostratigrafiâ, paleontologiâ, usloviâ obrazovaniâ). Trudy Instituta Geologii i Geofiziki AN SSSR, Sibirskoe Otdelnie 475: 20–27.
Siegmund, H. 1997. The Ocruranus‒Eohalobia group of small shelly fossils from the Lower Cambrian of Yunnan. Lethaia 30: 285–291. Crossref
Singh, P. and Shukla, S.D. 1981. Fossils from the Lower Tal: their age and bearing on the stratigraphy of Lesser Himalaya. Geoscience Journal 2: 157–176.
Sipin, D.P. 2001. Hiolity, problematičnye organizmy i stratigrafiâ nižnego kembriâ severo-zapada Sibirskoj platformy. 201 pp. Unpublished Doktoral Candidate Thesis, Institut Geologii Nefti i Gaza SO AN SSSR, Novosibirsk.
Skovsted, C.B. 2003. Mobergellans (Problematica) from the Cambrian of Greenland, Siberia and Kazakhstan. Paläontologische Zeitschrift 77: 429–443. Crossref
Skovsted, C.B. 2006. Small Shelly fauna from the upper Lower Cambrian Bastion and Ella Island formations, north-east Greenland. Journal of Paleontology 80: 1087–1112. Crossref
Skovsted, C.B. and Peel, J.S. 2011. Hyolithellus in life position from the lower Cambrian of North Greenland. Journal of Paleontology 85: 37–47. Crossref
Skovsted, C.B., Balthasar, U., Brock, G.A., and Paterson, J.R. 2009. The tommotiid Camenella reticulosa from the early Cambrian of South Australia: Morphology, scleritome reconstruction, and phylogeny. Acta Palaeontologica Polonica 54: 525–540. Crossref
Skovsted, C.B., Holmer, L.E., Topper, T.P., and Brock, G.A. 2010. Brachiopod affinity of the Lower Cambrian problematic bivalve Apistoconcha. In: International Palaeontological Congress, London, June, 28–July, 3, Abstracts with Programme, 356.
Skovsted, C.B., Ushatinskaya, G., Holmer, L.E., Popov, L.E., and Kouchinsky, A. 2015. Taxonomy, morphology, shell structure and early ontogeny of Pelmanotreta nom. nov. from the lower Cambrian of Siberia. GFF 137: 1–8. Crossref
Smith, E.F., Macdonald, F.A., Petach, T.N., Bold, U., and Schrag, D.P. 2015. Integrated stratigraphic, geochemical, and paleontological late Ediacaran to early Cambrian records from southwestern Mongolia. Geological Society of America Bulletin 128: 442–468. Crossref
Sokolov, B.S. and Zhuravleva, I.T. [Žuravleva, I.T.] (eds.) 1983. Stage subdivision of the Lower Cambrian of Siberia. Atlas of fossils [in Russian]. Trudy Instituta Geologii i Geofiziki SO AN SSSR 558: 1–216.
Steiner, M., Li, G., Qian, Y., and Zhu, M. 2004a. Lower Cambrian small shelly fossils of northern Sichuan and southern Shaanxi (China), and their biostratigraphic importance. Geobios 37: 259–275. Crossref
Steiner, M., Li, G., Qian, Y., Zhu, M., and Erdtmann, B.D. 2003. Lower Cambrian small shelly faunas from Zhejiang (China) and their biostratigraphical implications. Progress in Natural Science 13: 852–860. Crossref
Steiner, M., Li, G., Qian, Y., Zhu, M., and Erdtmann, B.D. 2007. Neoproterozoic to early Cambrian small shelly fossil assemblages and a revised biostratigraphic correlation of the Yangtze Platform (China). Palaeogeography, Palaeoclimatology, Palaeoecology 254: 67–99. Crossref
Steiner, M., Qian, Y., Li, G., Hagadorn, J.W., and Zhu, M. 2014. The developmental cycles of early Cambrian Olivooidae fam. nov. (?Cycloneuralia) from the Yangtze Platform (China). Palaeogeography, Palaeoclimatology, Palaeoecology 398: 97–124. Crossref
Steiner, M., Zhu, M., Li, G., Qian, Y., and Erdtmann, B.D. 2004b. New Early Cambrian bilaterian embryos and larvae from China. Geology 32: 833–836. Crossref
Streng, M. and Skovsted, C.B. 2006. A new mobergellan (small shelly fossils) from the early Middle Cambrian of Morocco and its significance. Paläontologische Zeitschrift 80: 209–220. Crossref
Streng, M., Babcock, L.E., and Hollingsworth, J.S. 2005. Agglutinated protists from the Lower Cambrian of Nevada. Journal of Paleontology 79: 1214–1218. Crossref
Sysoev, V.A. 1958. Superorder Hyolithoidea [in Russian]. In: Û.A. Orlov, N.P. Luppov, and V.V. Druŝic (eds.), Osnovy paleontologii. Mollûski-golovonogie P. Ammonoidei (ceratity i ammonity), Vnutrirakovinnye. Priloženie: Konikonhii, 184–190. Gosgeoltehizdat, Moskva.
Sysoev, V.A. 1959a. Hyoliths of the genera Circotheca and Orthotheca from the Lower Cambrian of the Siberian Platform [in Russian]. Paleontologičeskij žurnal 1959 (1): 68–75.
Sysoev, V.A. 1959b. Hyoliths of the genus Circotheca from the Lower Cambrian of the Tajmyr National District [in Russian]. Paleontologičeskij žurnal 1959 (1): 84–94.
Sysoev, V.A. 1960a. Classification of the family Hyolithidae [in Russian]. Âkutskij Filial Naučnyj soobŝinâ, SO AN SSSR 4: 43–53.
Sysoev, V.A. 1960b. On the find of lower Cambrian hyolith Ceratotheca [in Russian]. Âkutskij Filial Naučnyj soobŝinâ, SO AN SSSR 4: 124–125.
Sysoev, V.A. 1962. Hiolity kembriâ severnogo sklona Aldanskogo ŝita. 66 pp. Âkutskij Filial SO AN SSSR, Âkutsk.
Sysoev, V.A. 1963. Hyoliths of the genus Torellella from the Lower Cambrian of the Anabar anticline [in Russian]. Paleontologičeskij žurnal 1963 (1): 49–55.
Sysoev, V.A. 1965. Main features of evolution of hyoliths [in Russian]. In: V.F. Vozin (ed.), Paleontologiâ i biostratigrafiâ paleozojskih i triasovyh otloženij Âkutii, 5–20. Nauka, Moskva.
Sysoev, V.A. 1968. Stratigrafiâ i hiolity drevnejših sloev nižnego kembriâ Sibirskoj platformy. 67 pp. Âkutskij Filial SO AN SSSR, Âkutsk.
Sysoev, V.A. 1972. Biostratigrafiâ i ortotecimorfnye hiolity nižnego kembriâ Sibiri.152 pp. Âkutskij Filial SO AN SSSR, Âkutsk.
Szaniawski, H. 1982. Chaetognath grasping spines recognized among Cambrian protoconodonts. Journal of Paleontology 56: 806–810.
Szaniawski, H. 2002. New evidence for the protoconodont origin of chaetognaths. Acta Palaeontologica Polonica 47: 405–19.
Szaniawski, H. and Bengtson, S. 1993. Origin of euconodont elements. Journal of Paleontology 67: 640‒654. Crossref
Thomas, R.D.K. and Vinther, J. 2012. Implications of the occurrence of paired anterior chaetae in the late Early Cambrian mollusc Pelagiella from the Kinzers Formation of Pennsylvania for relationships among taxa and early evolution of the Mollusca. Geological Society of America Abstracts with Programs 44 (7): 326. https://gsa.confex.com/gsa/2012AM/webprogram/Paper212374.html.
Thomas, R.D.K., Vinther, J., and Matt, K. 2010. Paired chaetae associated with spiral shells of the late Early Cambrian mollusc Pelagiella from the Kinzers Formation: taphonomy, functional morphology, and potential evolutionary relationships. Geological Society of America Abstracts and Program 42 (5): 633.
Urbanek, A. and Rozanov, A.Y. 1983. Upper Precambrian and Cambrian Palaeontology of the East-European Platform. 158 pp. Wydawnictwa Geologiczne, Warszawa
Ushatinskaya, G.T. and Malakhovskaya, Y.E. 2001. Origin and development of the Cambrian brachiopod biochores. Stratigraphy and Geological Correlation 9: 540–556.
Ushatinskaya, G.T. and Parkhaev, P.Y. 2005. Preservation of imprints and casts of cells of the outer mantle epithelium in the shells of Cambrian brachiopods, mollusks, and problematics. Paleontological Journal 39: 251–263.
Val’kov, A.K. 1968. To the fauna of the Kessyuse Formation of the Lower Cambrian of the Olenyok Uplift [in Russian]. In: K.B. Mokšancev, V.A. Sysoev, I.I. Tučkov, G.S. Fradkin, and G.I. Šteh (eds.), Tektonika, stratigrafiâ i litologiâ osadočnyh formacij Âkutii, 115–119. Âkutsk Publishing House, Âkutsk.
Val’kov, A.K. 1970. On the oldest beds of the Lower Cambrian of north-eastern Siberian Platform [in Russian]. In: A.K. Bobrov (ed.), Stratigrafiâ i paleontologiâ proterozoâ i kembriâ vostoka Sibirskoj Platformy, 91–93. Knižnoe izdatel’stvo, Âkutsk.
Val’kov, A.K. 1975. Biostratigrafiâ i hiolity kembriâ severo-vostoka Sibirskoj platformy. 140 pp. Nauka, Moskva.
Val’kov, A.K. 1982. Biostratigrafiâ nižnego kembriâ vostoka Sibirskoj platformy (Učuro-Majskij rajon. 91 pp. Nauka, Moskva.
Val’kov, A.K. 1983. Distribution of the oldest skeletal fossils and correlation of the Cambrian lower boundary in south-eastern part of the Siberian Platform [in Russian]. In: V.V. Homentovskij, M.S. Âkšin, and G.A. Karlova (eds.), Pozdnij dokembrij i rannij paleozoj Sibiri. Vendskie otloženiâ, 37–48. Institut Geologii i Geofiziki SO AN SSSR, Novosibirsk.
Val’kov, A.K. 1987. Biostratigrafiâ nižnego kembriâ vostoka Sibirskoj platformy (Ûdoma-Oleniokskij region). 136 pp. Nauka, Moskva.
Val’kov, A.K. and Karlova, G.A. 1984. Fauna from the Vendian–Cambrian transition beds in the lower reaches of the Gonam River [in Russian]. In: V.V. Homentovskij, V.Y. Šenfil’, and M.S. Âkšin (eds.), Stratigrafiâ pozdnego dokembriâ i rannego paleozoâ. Srednââ Sibir’, 12–41. Institut Geologii i Geofiziki SO AN SSSR, Novosibirsk.
Val’kov, A.K. and Sysoev, V.A. 1970. Cambrian angustiochreids of Siberia [in Russian]. In: A.K. Bobrov (ed.), Stratigrafiâ i paleontologiâ proterozoâ i kembriâ vostoka Sibirskoj platformy, 94–100. Knižnoe izdatel’stvo, Âkutsk.
Vannier, J., Steiner, M., Renvoise, E., Hu, S.-X., and Casanova, J.-P. 2007. Early Cambrian origin ofmodern food webs: evidence from predator arrow worms. Proceedings of the Royal Society B 274: 627–633. Crossref
Varlamov, A.I., Rozanov, A.Y., Khomentovsky, V.V., Shabanov, Y.Y., Abaimova, G.P., Demidenko, Y.E., Karlova, G.A., Korovnikov, I.V., Luchinina, V.A., Malakhovskaya, Y.E., Parkhaev, P.Y., Pegel, T.V., Skorlotova, N.A., Sundukov, V.M., Sukhov, S.S., Fedorov, A.B., and Kipriyanova, L.D. 2008. The Cambrian System of the Siberian Platform. Part 1: The Aldan-Lena region. 300 pp. PIN RAN, Moscow.
Vasil’eva, N.I. 1985. A contribution to the systematics of the order Chancelloriida Walcott, 1920 (incertae sedis) from Lower Cambrian deposits of the East Siberian Platform [in Russian]. In: B.S. Sokolov and I.T. Žuravleva (eds.), Problematiki pozdnego dokembriâ i paleozoâ. Trudy IGiG AN SSSR 632: 115–126.
Vasil’eva, N.I. 1986. A new genus of anabaritids from the Lower Cambrian of the Siberian Platform [in Russian]. Paleontologičeskij žurnal 1986 (2): 103–104.
Vasil’eva, N.I. 1990. New Early Cambrian gastropod mollusks from the Siberian Platform and some questions of their systematic [in Russian]. In: A.I. Nikolaev (ed.), Mikrofauna SSSR: sistematika i voprosy biostratigrafii, 4–21. VNIGRI, Leningrad.
Vasil’eva, N.I. 1998. Melkaâ rakovinnaâ fauna i biostratigrafiâ nižnego kembriâ Sibirskoj platformy. 139 pp. VNIGRI, St. Petersburg.
Vendrasco, M.J., Checa, A.G., and Kouchinsky, A.V. 2011a. Shell microstructure of the early bivalve Pojetaia and the independent origin of nacre within the Mollusca. Palaeontology 54: 825–850. Crossref
Vendrasco, M.J., Kouchinsky, A., Porter, S.M., and Fernandez, C.Z. 2011b. Phylogeny and escalation in Mellopegma and other Cambrian molluscs. Palaeontologia Electronica 14 (2): 1–44.
Vendrasco, M.J., Li, G., Porter, S.M., and Fernandez, C.Z. 2009. New data on the enigmatic Ocruranus‒Eohalobia group of early Cambrian small skeletal fossils. Palaeontology 52: 1373–1396. Crossref
Vendrasco, M.J., Porter, S.M., Kouchinsky, A., Li, G., and Fernandez, C.Z. 2010. New data on molluscs and their shell microstructures from the Middle Cambrian Gowers Formation, Australia. Palaeontology 53: 97–135. Crossref
Vidal, G., Palacios, T., Gámez-Vintaned, J.A., Díez Balda, M.A., and Grant, S.W.F. 1994. Neoproterozoic–early Cambrian geology and palaeontology of Iberia. Geological Magazine 131: 729–765. Crossref
Vidal, G., Palacios, T., Moczydłowska, M., and Gubanov, A.P. 1999. Age constraints from small shelly fossils on the early Cambrian terminal Cadomian Phase in Iberia. GFF 121: 137–143. Crossref
Vinn, O. and Kirsimäe, K. 2015. Alleged cnidarian Sphenothallus in the Late Ordovician of Baltica, its mineral composition and microstructure. Acta Palaeontologica Polonica 60: 1001–1008.
Vinther, J. and Nielsen, C. 2005. The Early Cambrian Halkieria is a mollusk. Zoologica Scripta 34: 81–89. Crossref
Voronin, Y.I. [Voronin, Û.I.], Voronova, L.G., Grigorieva, N.V., Drozdova, N.A., Zhegallo, E.A. [Žegallo, E.A.], Zhuravlev, A.Y. [Žuravlev, A.Û.], Ragozina, A.L., Rozanov, A.Y. [Rozanov, A.Û.], Sayutina, T.A. [Saûtina, T.A.], Sysoev, V.A., and Fonin, V.D. 1982. The Precambrian–Cambrian boundary in the geosynclinal regions (reference section Salany-Gol, MNR) [in Russian]. Trudy Sovmestnoj sovetsko-mongol’skoj paleontologičeskoj èkspedicii 18: 1–150.
Voronova, L.G. and Missarzhevsky, V.V. [Missarževskij, V.V.] 1969. Finds of algae and worm tubes in the Precambrian–Cambrian boundary beds of the northern part of the Siberian Platform [in Russian]. Doklady AN SSSR 184: 207–210.
Voronova, L.G. and Rozanov, A.Y. [Rozanov, A.Û.] 1973. On relationship of litho- and biostratigraphic boundaries at the example of the Cambrian of the Igarka region. Problems of the palaeontology and stratigraphy of Siberia and Far East [in Russian]. Trudy IGiG SO AN SSSR 49: 220–224.
Vostokova, V.A. 1962. The Cambrian gastropods from Siberia and Tajmyr [in Russian]. Trudy Naučno-Issledovatel’skogo Instituta Geologii Arktiki 28: 51–74.
Walcott, C.D. 1886. Second contribution to the studies on the Cambrian faunas of North America. Bulletin of the United States Geological Survey 30: 1–369.
Walcott, C.D. 1920. Middle Cambrian Spongiae. Smithsonian Miscellaneous Collections 67: 261–364.
Walliser, O.H. 1958. Rhombocorniculum comleyense n. gen. n. sp. (Incertae sedis, Unterkambrium, Shropshire). Paläontologisches Zeitschrift 32: 176–180. Crossref
Webby, B.D. 1970. Late Precambrian trace fossils from New South Wales. Lethaia 3: 79–109. Crossref
Wenz, W. 1938. Gastropoda. Allgemeiner Teil und Prosobranchia. In: O.H. Schindewolf (ed.), Handbuch der Paläozoologie. Band 6, 1–720. Bornträger, Berlin.
White, R.D. 1986. Cambrian Radiolaria from Utah. Journal of Paleontology 60: 778–780. Crossref
Wood, R.A., Zhuravlev, A.Y., Sukhov, S.S., Zhu, M., and Zhao, F. 2017. Demise of Ediacaran dolomitic seas marks widespread biomineralization on the Siberian Platform. Geology 45 (1): 27–30. Crossref
Xing, Y., Ding, Q., Luo, H., He, T., Wang, Y., Jiang, Z., Chen, Y., Zhang, S., Liu, G., Xiong, X., Chen, P., Wang, Y., Yin, G., Zheng, S., Qian, Y., Yue, Z., Wu, X., Song, X., Ouyang, L., and Ding, L. 1984. The Sinian‒Cambrian boundary of China [in Chinese, with an English summary]. Bulletin of the Institute of Geology, Chinese Academy of Geological Sciences 10: 1–262
Yang, B., Steiner, M., Li, G., and Keupp, H. 2014. Terreneuvian small shelly faunas of east Yunnan (South China) and their biostratigraphic implications. Palaeogeography, Palaeoclimatology, Palaeoecology 398: 28–58. Crossref
Yang, B., Steiner, M., Zhu, M., Li, G., Liu, J., and Liu, P. 2016. Transitional Ediacaran‒Cambrian small skeletal fossil assemblages from South China and Kazakhstan: Implications for chronostratigraphy and metazoan evolution. Precambrian Research 285: 202–215. Crossref
Yang, X. and He, T. 1984. New small shelly fossils from Lower Cambrian Meishucun stage of Nanjiang area, northern Sichuan. Professional Papers of Stratigraphy and Palaeontology 13: 35–48.
Yao, J., Xiao, S., Yin, L., Li, G., and Yuan, X. 2005. Basal Cambrian microfossils from the Yurtus and Xishanblaq formations (Tarim, north-west China): systematic revision and biostratigraphic correlation of Micrhystridium-like acritarchs. Palaeontology 48: 687–708. Crossref
Yin, J., Ding, L., He, T., Li, S., and Shen, L. 1980. The Palaeontology and Sedimentary Environment of the Sinian System in Emei-Ganluo Area, Sichuan [in Chinese, with English abstract]. 231 pp. Sichuan People’s Publishing House, Chengdu.
Yu, W. 1974. Cambrian hyolithids [in Chinese]. In: Handbook of the Stratigraphy and Paleontology of Southwest China, 111–113. Science Press, Beijing.
Yu, W. 1979. Earliest Cambrian monoplacophorans and gastropods from western Hubei with their biostratigraphical significance [in Chinese, with English abstract]. Acta Palaeontologica Sinica 18: 233–270.
Yu, W. 1987. Yangtze micromolluscan fauna in Yangtze region of China with notes on the Precambrian–Cambrian boundary [in Chinese, with English abstract]. In: Stratigraphy and Palaeontology of Systemic Boundaries in China: Precambrian–Cambrian Boundary, 19–344. Nanjing University Publishing House, Nanjing.
Yu, W. 2014. On the Yangtzeconus priscus‒Archaeospira ornata Assemblage (Mollusca) of the earliest Cambrian of China. Acta Geologica Sinica 88: 1262–1287. Crossref
Yuan, K. and Zhang, S. 1983. Discovery of the Tommotian fauna in SW China [in Chinese]. Acta Palaeontologica Sinica 22: 31–41.
Yue, Z. and Bengtson, S. 1999. Embryonic and post-embryonic development of the Early Cambrian cnidarian Olivooides. Lethaia 32: 181–195.
Zhamoida, A.I. [Žamojda, A.I.] and Petrov, O.V. (eds.) 2008. Status of knowledge of the Precambrian and Phanerozoic stratigraphy in Russia. Goals of further studies [in Russian.]. Postanovleniâ mežvedomstvennogo stratigrafičeskogo komiteta i ego postojannyh komissij 38: 1–131.
Zhang, H. and Dong, X. 2015. The oldest known larva and its implications for the plesiomorphy of metazoan development. Chinese Science Bulletin 60: 1947–1953. Crossref
Zhang, Z., Smith, M.R., and Shu, D. 2015. New reconstruction of the Wiwaxia scleritome, with data from Chengjiang juveniles. Scientific Reports 5: 14810. Crossref
Zhong, H. 1977. Preliminary study on the ancient fauna of South China and its stratigraphic significance [in Chinese]. Scientia Geologica Sinica 2: 118–128.
Zhu, M., Babcock, L.E., and Peng, S.C. 2006. Advances in Cambrian stratigraphy and paleontology: Integrating correlation techniques, palaeobiology, taphonomy and paleoenvironmental reconstruction. Palaeoworld 15: 217–222. Crossref
Zhu,M., Zhuravlev, A.Y., Wood, R.A., Zhao, F., and Sukhov, S.S. 2017. A deep root for the Cambrian explosion: Implications of new bio- and chemostratigraphy from the Siberian Platform. Geology 45: 455–458. Crossref
Zhu, R.-X., Li, X.-H., Hou, X., Pan, Y.-X., Wang, F., Deng, C.-L., and He, H.-Y. 2009. SIMS U-Pb zircon age of a tuff layer in the Meishucun section, Yunnan, southwest China: constraint on the age of the Precambrian‒Cambrian boundary. Science in China, Series D-Earth Sciences 52: 1385–1392. Crossref
Zhuravlev, A.Y., Liñán, E., Gámez Vintaned, J.A., Debrenne, F., and Fedorov, A.B. 2012. New finds of skeletal fossils in the terminal Neoproterozoic of the Siberian Platform and Spain. Acta Palaeontologica Polonica 57 (1): 205–224. Crossref
Acta Palaeontol. Pol. 62 (2):
311‒440, 2017
https://doi.org/10.4202/app.00289.2016