
A new triadotypid insect from the Late Triassic of South Africa
ISABLELLE DEREGNAUCOURT, TORSTEN WAPPLER, JOHN M. ANDERSON, and OLIVIER BÉTHOUX
Deregnaucourt, I., Wappler, T., Anderson, J.M., and Béthoux, O. 2017. A new triadotypid insect from the Late Triassic of South Africa. Acta Palaeontologica Polonica 62 (3): 613–618.
Extant odonates (damsel- and dragonflies) represent a small subset of the historical biodiversity of this group. Among their successive stem-groups, the Triadotypomorpha are poorly documented. Herein we describe a new species Reisia rieki from the Molteno Formation (South Africa, Upper Triassic) belonging to this taxon. The comparatively large sample allows a relatively complete description of the wing venation in Triadotypomorpha. We noticed the occurrence of a strongly oblique crossvein located between RA and RP1, a condition documented in some other Pandiscoidalia and which might be of phylogenetic importance. The new species probably inhabited open landscapes and foraged above large water bodies. The documentation of a Gondwanian Triadotypomorpha demonstrates that the group had a worldwide distribution by the Triassic.
Key words: Arthropoda, Odonata, Pandiscoidalia, Triassic, Carnian, Molteno Formation, South Africa.
Isabelle Deregnaucourt [isabelle.deregnaucourt@edu.mnhn.fr] and Olivier Béthoux [obethoux@mnhn.fr] (corresponding author), Sorbonne Universités, UPMC Univ Paris 06, MNHN, CNRS, Centre de recherche sur la paléontologie-Paris (CR2P), 57 rue Cuvier, CP38, F-75005 Paris, France.
Torsten Wappler [twappler@uni-bonn.de], Steinmann Institute, University of Bonn, Nussallee 8, 53115 Bonn, Germany; and [wappler@hlmd.de], Hessisches Landesmuseum, Friedensplatz 1, 64283 Darmstadt, Germany.
John M. Anderson [jmanderson.gondwana@gmail.com], Environmental Studies Institute, Witwatersrand University, 1 Jan Smuts Ave., Braamfontein, Johannesburg 2000, South Africa.
Received 28 January 2017, accepted 21 April 2017, available online 17 July 2017.
Copyright © 2017 I. Deregnaucourt et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Compared to other insect orders, the fossil record of odonates (damsel- and dragonflies) is comparatively rich. Many of the successive fossil sister-groups of crown-Odonata have been documented and classified, essentially based on wing venation, which is very elaborate and disparate in these insects (Riek and Kukalová-Peck 1984; Bechly 1996). Yet, some groups are known from few species only, and the documented Southern hemisphere fossil odonates are few. For example, only four species of Odonatoptera (understood as odonates + “protodonates”) are known from Southern Palaeozoic localities (Riek and Kukalová-Peck 1984; Martins-Neto 1996; Schlüter 2003; Nel et al. 2012), a figure in contrast with the tens of species known from contemporaneous, Northern Hemisphere localities (e.g., Beckemeyer 2000; Nel et al. 2008, 2012; Li et al. 2013; see also Carpenter 1992).
Herein we describe a new species, from the Molteno Formation (South Africa; Triassic) belonging to the taxon Triadotypomorpha. The comparatively large sample allows a relatively complete description of the wing venation of a species belonging to this taxon. Palaeoecological and palaeobiogeographical aspects are briefly discussed.
Institutional abbreviations.—PRE/F, “Pretoria/Fossil”, National Botanical Institute [now South African National Biodiversity Institute], Pretoria, South Africa, relocated to Bernard Price Institute for Palaeontology (now Evolutionary Studies Institute), University of the Witwatersrand, Johannesburg, South Africa.
Other abbreviations.—AA, anterior Analis; Cu, Cubitus; CuA, anterior Cubitus; CuP, posterior Cubitus; Irp1–rp2, Irp1+2–rp3+4 and Irp3–rp4, intercalary veins occurring between RP1 and RP2, between RP1+2 (then RP2 only) and RP3+4 (then RP3 only), and between RP3 and RP4, respectively; M, Media (red); MA, anterior Media; MP, posterior Media; R, Radius (blue in the figures); RA, anterior Radius; RP, posterior Radius; RP3+4, first posterior branch of RP; RP1, anterior branch resulting from the second fork of RP; RP2, posterior branch resulting from the second fork of RP; ScP, posterior Subcosta; Sn, subnodal crossvein.
Material and methods
The fossils described are housed at the Bernard Price Institute (School of Geosciences, University of the Witwatersrand; Johannesburg, South Africa). They bear the specimen numbers PRE/F/17569 (negative and positive imprints; Figs. 1A, 2A, 3), PRE/F/16442 (negative and positive imprints; Figs. 1B, 2B), PRE/F/10616 (negative and positive imprints; Figs. 1C, 2C) and PRE/F/17499 (negative imprint; Figs. 1D, 2D).
All specimens were collected from the Molteno Formation, which forms an outcrop area of 25 000 km² within the Karoo Basin, in South Africa. The Basin lies in North of the Cape Fold Belt and unites the deposits of the Palaeozoic up to the Mesozoic, with an accumulation reaching up to 5500 m (Lanci et al. 2013). The Karoo Basin is interpreted as a retro-arc foreland basin which subsided during a vertical motion of rigid basement blocks and intervening crustal faults (Tankard et al. 2009). It is subdivided into the Stormberg, Beaufort, Ecca, and Dwyka groups (Prevec et al. 2009). Together with the Drakensberg, Elliot, and Clarens formations, the Molteno Formation belongs to the Stormberg Group consisting of the deposits from the Upper Triassic to Jurassic. They have been formed during the final unloading phase of the Belt (Bordy et al. 2004). The Molteno Formation itself belongs to the early Carnian age (Anderson et al. 1998).
During the Upper Triassic the investigated area was a part of the supercontinent Gondwana and belonged to an extensive inland basin. It has been located between latitude 50° and 55° S and there are comparable fossil faunas in the Transantarctic Mountains and the Sydney Basin in Australia. The climate was characterized by strong seasonal differences with dry, hot summers and cold, dark winters (Cairncross et al. 1995). The layers, which have been deposited in the area, are remains of large braided river systems with their northerly prograding floodplains. They consist of conglomerate, sandstone, shale and rare coal layers which are subdivided into three primary facies: large-scale fining-up, coarse grained sequences in the river zone; upward-coarsening crevasse-splay and sheet flood sequences and at last laminated lacustrine and marsh shale deposits in the floodplain (Anderson et al. 1998). The Molteno biome is separated in 7 primary ecozones. The habitats reach from Dicroidium riparian forests over Dicroidium and Sphenobaiera woodlands to Heidiphyllum thicket, Equisetum marsh and fern/Ginkgophytopsis meadows (Anderson et al. 1998).
Draft drawings were prepared with the aid of a Zeiss SteREO Discovery V8 Stereomicroscope equipped with a pair of W-PL 10x/23 eye pieces, a Plan Apo S 1.0x FWD objective, and a drawing tube (Jena, Germany). Photographs were taken using a Canon EOS 5D Mark III equipped with Canon 50 mm or MP-E 65 mm macro lenses. The light-mirror technique has been used to provide positive views of the specimens. Photographs were optimised using Adobe Photoshop CS6. Draft drawings were inked using Adobe Illustrator CS6 using both scanned draft drawings and photographs.
We follow the so-called “serial insect wing venation groundplan” (Lameere 1922, 1923). Essentially based on it, we follow the Odonata wing venation groundplan elaborated by Riek and Kukalová-Peck (1984) and its successive implementations at various levels of inclusiveness (Nel et al. 1993; Bechly 1995; Béthoux 2015). In addition to this standard terminology, we propose to use additional terms to indicate branches of RP3+4, which branching patterns are relevant to the following discussion, as follows: RP3a and RP3b are the anterior and posterior branches of RP3, respectively; RP4a and RP4b are the anterior and posterior branches of RP4, respectively (RP4a and RP4b are further split into RP4aa, RP4ab, etc. accordingly).
Systematic palaeontology
Order Odonata Fabricius, 1793
Taxon Pandiscoidalia Nel, Béthoux, Bechly, Martínez-Delclòs, and Papier, 2001
Taxon Triadotypomorpha Bechly, 1996
Family Triadotypidae Grauvogel and Laurentiaux, 1952
Genus Reisia Handlirsch, 1912
= Triadotypus Grauvogel and Laurentiaux, 1952
= Rabru Béthoux, de la Horra, Benito, Barrenechea, Galán, and López-Gómez, 2009
Type species: Reisia gelasii (Reis, 1909); Lower “Muschelkalk”, Middle Triassic; Münnerstadt, Lower Franconia, Germany.
Included species: Type species and Reisia sogdiana (Pritykina, 1981) (combination by Bechly 1997, but misspelled “Reisia sogdianus”; same combination claimed as new by Nel et al. 2001; specific epithet “sogdianus” erroneously reported as occurring as “sodiques” in Carpenter 1992 and Bechly 1997—where it occurs as “sogdianus”—by Nel et al. 2001); Reisia nana Bechly, 1997 (initially regarded as a variety of Reisia guillaumei by Laurentiaux-Vieira et al. 1952); Reisia rubra (Béthoux, de la Horra, Benito, Barrenechea, Galán, and López-Gómez, 2009) comb. nov.; Reisia rieki sp. nov.
Emended diagnosis.—Long RP+MA stem basal to the arculus.
Remarks.—As a preliminary remark, the assignment of Iverya averyi Béthoux and Beattie, 2010 to the Triadotypidae (see original description) is to be discarded, as the species possesses the specialised “MAb” crossvein, regarded by Nel et al. (2001) as diagnostic of the clade Discoidalia (sister-group of the Triadotypomorpha, itself including the Triadotypidae; see below).
The systematics of Triadotypomorpha (related to the new material herein investigated) is confused. The last account on the topic has been given by Nel et al. (2001), who recognised two genera, Triadotypus Grauvogel and Laurentiaux, 1952 and Reisia Handlirsch, 1912, the former being considered distinct owing to a simple MP. Nel et al. (2001), who last revised the type species of Triadotypus, Triadotypus guillaumei, essentially relied on a reconstruction very similar to that provided in the original description (redrawn with minor modifications as Nel et al. 2001: fig. 1), and on a sketch of the basal area of the wing (Nel et al. 2001: fig. 2) and a photograph (Nel et al. 2001: fig. 3). However, Grauvogel and Laurentiaux (1952: 129) mentioned that the state of preservation of the holotype of Triadotypus guillaumei is insufficient to allow a positive statement on the branching pattern (or lack thereof) of MP (their “CuP”). Our examination of photographs of the holotype provided by Léa Grauvogel-Stamm let us believe that MP is actually branched in this specimen, in a very distal position, as in Reisia gelasii. The distinction between Reisia and Triadotypus therefore is not evident. We disagree with Grauvogel and Laurentiaux (1952) who considered “Reisia gelasii” as a nomen nudum, and concur with Bechly (1997) who considered Triadotypus guillaumei and Reisia gelasii as synonyms. As a consequence, both genera should be considered synonyms, and Reisia given a priority.
The species Rabru rubra Béthoux, de la Horra, Benito, Barrenechea, Galán, and López-Gómez, 2009 is the most recently described triadotypomorph. The species is very poorly known, just as many Reisia species. Indeed, we failed to identify a suitable, diagnostic character state for the corresponding genus, which we then propose to synonymise under Reisia. The validity of the species is not obvious; it might differ from R. gelasii owing to more developed branches of RP2 (diagnostic states listed by Béthoux et al. 2009 are shared with other species).
In summary, we propose that the Triadotypomorpha include a single family, the Triadotypidae, itself including a single genus, Reisia.
Reisia rieki sp. nov.
Figs. 1–3.
Etymology: Named after Edgar Frederick Riek (1920–2016), who described the earliest collections of Molteno insects in three papers in the mid-1970’s.
Type material: Holotype: PRE/F/17569 (positive and negative imprints). Paratypes: PRE/F/16442 (negative and positive imprints) and PRE/F/17499 (negative imprint); all from the type locality.
Type locality: Kapokkraal (locality code “Kap 111”; see Anderson and Anderson 1984), Karoo Basin, South Africa.
Type horizon: Molteno Formation, Carnian, Triassic.
Material.—Type material and specimen PRE/F/10616 (negative and positive imprints) from the Aasvoëlberg locality (locality code “Aas 411”; see Anderson and Anderson 1984), Carnian (Triassic), Molteno Formation, Karoo Basin, South Africa.
Diagnosis.—MP area moderately developed (as opposed to “small” in Reisia gelasii and R. nana, and “large” in R. sogdianus; unknown in R. rubra); sparse crossveins network (as opposed to dense in R. sogdianus); main portion of Irp1+2–rp3+4 and RP3/3a straight (regularly bent in all other species of Reisia, but unknown in R. nana); clear origin of RP3b (no clear differentiation of RP3b in R. sogdianus); RP2, Irp1+2–rp3+4 and RP3a with a strong bending near their end (putative apomorphy; unknown in R. nana); RP2 with a few, very distal branches (as opposed to more basal origin, as in R. rubra and, to some extent, in R. gelasii—unknown in R. nana and R. sogdianus).
Description.—General features: wing total length and maximum width unknown, about 108–118 mm, and 15–17 mm, respectively; anterior wing margin slightly thicker near the apex (i.e., in the area where a pterostigma occurs in extant relatives); area between the anterior wing margin and ScP large, with many crossveins, distally broadened in the “pterostigmal area”; in the area between the anterior wing margin and RA (distal to the nodus), crossveins stronger than in other areas; occurrence of a Sn; in the area between RA and RP1, occurrence of a distinctively oblique crossvein (known only in the holotype; see asterisk on Fig. 1A and Fig. 3); RP+MA divided into RP and MA basal to the nodus; RP divided into RP1+2 and RP3+4 opposite Sn; RP1+2 divided into RP1 and RP2 shortly before wing mid-length; Irp1–rp2 (convex) occupying a large area filled with convex branches (of Irp1–rp2) and supplementary concave intercalaries, all sub-parallel; RP3+4 divided into RP3 and RP4 basal to the fork of RP1+2; areas between RP1+2/2, Irp1+2–rp3+4 and RP3+4/3/3a each with a single row of cells; RP3 divided in RP3a and RP3b, without obvious further forks; RP2, Irp1+2–rp3+4 and RP3a strongly bent close to the posterior wing margin, which they reach at a nearly right angle; large area between RP3a and RP3b; occurrence of Irp3–rp4, simple; areas between RP3/3b, Irp3–rp4 and RP4/4a/4aa each with a single row of cells; RP4 dichotomously divided, with three orders of branching for RP4a and RP4b (RP4baa and RP4bab visible); a single row of cells occurs between RP3+4/4 and MA; MA simple; MA and MP parallel, area between these two veins with a single row of cells, longer than broad; MP with no evident fork, but delimiting a large area filled with several concave MP branches and convex intercalary veins, subparallel; CuA with numerous branches (convex) and intercalary veins (concave) between them; some of these intercalaries are forked and additional, convex intercalaries occur between them.
Specimen PRE/F/17569 (Figs. 1A, 2A, 3): right wing, median part preserved; preserved length 61 mm, maximum width 16.6 mm; basal part of the wing partly distorted, with ScP most probably concealed by RA (the termination of ScP not visible, occurrence of the nodal crossvein cannot be verified); apex and base of the wing missing.
Specimen PRE/F/16442 (Figs. 1B, 2B): fragment of a left wing; preserved length 36.2 mm, width of 16.9 mm; apex and first half of the wing missing; specimen slightly larger than others.
Specimen PRE/F/10616 (Figs. 1C, 2C): fragment of the apex of a right wing; preserved length 29.5 mm, maximum width 13 mm; distal part of RP2 preserved, with few, short branches.
Specimen PRE/F/17499 (Figs. 1D, 2D): fragment of the distal third of a right wing; preserved length 43.2 mm, maximum width 13.8 mm.
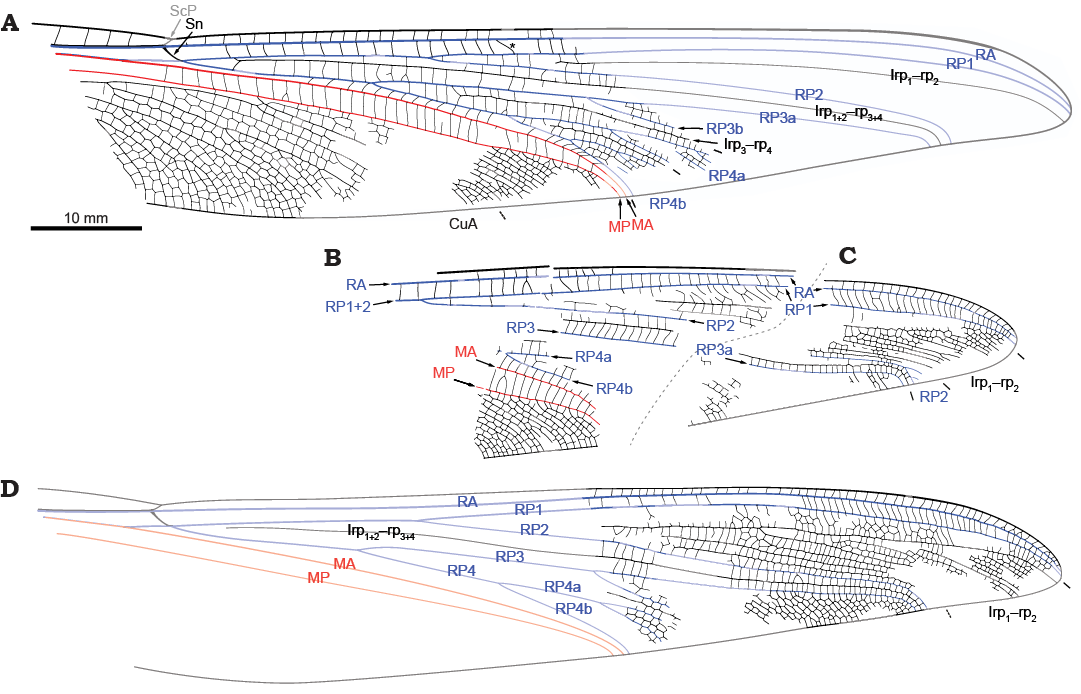
Fig. 1. Triadotypid insect Reisia rieki sp. nov. from Carnian (Triassic) of Kapokkraal (A, B, D) and Aasvoëlberg (C) localities, Molteno Formation, Karoo Basin, South Africa. A. PRE/F/17569, right wing. B. PRE/F/16442, left wing. C. PRE/F/10616, right wing. D. PRE/F/17499, right wing.

Fig. 2. Triadotypid insect Reisia rieki sp. nov. from Carnian (Triassic) of Kapokkraal (A, B, D) and Aasvoëlberg (C) localities, Molteno Formation, Karoo Basin, South Africa. A. PRE/F/17569, right wing (light-mirorred, flipped horizontally; the white frame indicates the area magnified in Fig. 3). B. PRE/F/16442, left wing (light-mirrored). C. PRE/F/ 10616, right wing (light-mirrored, flipped horizontally). D. PRE/F/17499, right wing (light-mirrored, flipped horizontally).
Remarks.—Because of the elaborate system of intercalary veins it proved impossible to identify CuP and AA branches with certainty in the only specimen displaying this area (Figs. 1A, 2A). For similar reasons, the delimitation of CuA is uncertain.
We hypothesise that all specimens herein documented belong to a single species. Although they are very incomplete, there are sufficient portions in common to allow comparison in venation and size. A good match was obtained for the specimens PRE/F/17569 (Figs. 1A, 2A) and PRE/F/16442 (Figs. 1B, 2B) based on the RP1–RP2 and RP4a–RP4b forks. Considering these specimens altogether, the specimen PRE/F/17499 (Figs. 1D, 2D) perfectly matches them (in particular the outline, and the peculiar, straight course of RP2, Irp1+2–rp3+4, and RP3/3a). Finally, the specimen PRE/F/10616 (Figs. 1C, 2C) is very similar to the specimen PRE/F/17499, but also complement the specimen PRE/F/16442 (with a minor difference in size, the latter being the largest of the set). The range of intra-specific variability is unknown in related species (all are known from singletons but one, R. guillaumei, known thanks to two incomplete specimens), but the observed differences are minimal. In conclusion there is no ground to assume that the specimens belong to different species. Until new specimens are discovered and constitute a challenge to this proposition, it should be considered valid.
The species possesses a long RP + MA stem basal to the arculus (Figs. 1A, 2A), a trait distinctive of Triadotypomorpha (see above). Our diagnosis demonstrates that this species differs from those previously known. Its size is similar to that of other Reisia species, as far as it can be inferred from the limited data on these species.
Stratigraphic and geographic range.—Putatively early Carnian, Late Triassic (Anderson and Anderson 1983); Karoo Basin, Molteno Formation, localities Kapokkraal and Aasvoëlberg (respective locality codes “Kap 111” and “Aas 411”; see Anderson and Anderson 1984), South Africa.

Fig. 3. Triadotypid insect Reisia rieki sp. nov. from Carnian (Triassic) of Kapokkraal locality, Molteno Formation, Karoo Basin, South Africa, PRE/F/17569a, detail of the oblique crossvein between RA and RP1 (opposite the first fork of RP4, indicated by an asterisk), as indicated on Fig. 2A (light-mirrored, flipped horizontally).
Discussion
Thanks to a modest, yet comparatively large, set of specimens, we could investigate the wing morphology of a new species of Triadotypomorpha with greater details than most previous authors who contributed on this group. We noticed the occurrence of a strongly oblique crossvein located between RA and RP1. The same condition has been documented in the genera Triadophlebia, Neritophlebia, Cladophlebia, and Paurophlebia (see original descriptions in Pritykina 1981), which are all Triadophlebiomorpha. This character state is therefore a putative apomorphy of the Pandiscoidalia (which includes the clade Triadotypomorpha, sister-group to Triadophlebiomorpha and Stigmoptera—the latter including crown-Odonata and their closest stem-groups), or of a subgroup within this clade. Indeed, not all Triadophlebiomorpha possess this crossvein, and it is absent in Stigmoptera (Nel et al. 2012). Therefore, its phylogenetic significance is not evident, but it is worth being carefully scrutinised.
Thanks to previous palaeoecological reconstructions (Cairncross et al. 1995; Anderson et al. 1998) and occurrence data of R. rieki sp. nov. specimens, it can be deduced that the species populated wide, open areas, including the vicinity of river channels (locality “Kap 111”) and floodplain lakes (locality “Aas 411”). This is consistent with the large size of the species, suggestive of low manoeuvrability, balanced by comparatively high flight speeds, beneficial for dispersal. Indeed, the group was previously documented in Laurasia only. Our discovery suggests that it had spread worldwide by the Triassic.
Conclusions
Prior to the current contribution, the clade Triadotypomorpha was documented thanks to five incomplete specimens only, belonging to four species. Reisia rieki sp. nov. is amongst the best documented species of the group, which is composed of remote stem-Odonata. The new species probably inhabited open landscapes and foraged above large water bodies. The documentation of a Gondwanian Triadotypomorpha demonstrates that the group had a worldwide distribution by the Triassic.
Acknowledgements
We are grateful to the Heidi M. Anderson (Honorary Research Associate, Evolutionary Studies Institute, Johannesburg, South Africa) for her dedication with collecting fossil insects from the Molteno Formation, and to Léa Grauvogel-Stamm for providing photographs of the holotype of R. guillaumei. We thank André Nel (Institut de Systématique, Evolution, Biodiversité, ISYEB, UMR 7205, Paris, France) and Alexander P. Rasnitsyn (Borissak Paleontological Institute of the Russian Academy of Sciences, Moscow, Russia) for prompt and useful comments on a previous version of this contribution. Data on the “Anderson collection” and the new fossil species in particular were collected during two visits at Evolutionary Studies Institute in 2014 and 2015. We are gratefull to Marion Bamford, Bernhard Zipfel, and Tandi Scott-Turner (all Evolutionary Studies Institute) who proved very helpful during these visits. Visits were supported by two grants to OB from the “Action Transversale Muséum Emergences” (2014, 2015) and by a grant to TW and OB from the DFG (WA 1492/12-1; 2014).
References
Anderson, J.M. and Anderson, H.M. 1983. Palaeoflora of Southern Africa. Molteno Formation (Triassic). Vol 1: Part 1, Introduction. Part 2, Dicroidium. 228 pp. Botanical Research Institute, Pretoria.
Anderson, J.M. and Anderson, H.M. 1984. The fossil content of the Upper Triassic Molteno Formation, South Africa. Palaeontologia Africana 25: 39–59.
Anderson, J.M., Anderson, H.M., and Cruickshank, A.R.I. 1998. Late Triassic Ecosystems of the Molteno/Lower Elliot Biome of Southern Africa. Palaeontology 41: 387–421.
Bechly, G. 1995. Morphologische Untersuchungen am Flügelgeäder der rezenten Libellen und deren Stammgruppenvertreter (Insecta; Pterygota; Odonata) unter besonderer Berücksichtigung der Phylogenetischen Systematik und des Grundplanes der Odonata. Petalura special volume 1: 1–341.
Bechly, G. 1996. Morphologische Untersuchungen am Flügelgeäder der rezenten Libellen und deren Stammgruppenvertreter (Insecta; Pterygota; Odonata) unter besonderer Berücksichtigung der Phylogenetischen Systematik und des Grundplanes der Odonata [revised edition including appendix in English]. Petalura Special Volume 2: 1–402.
Bechly, G. 1997. New fossil odonates from the Upper Triassic of Italy, with a redescription of Italophlebia gervasuttii, and a reclassification of Triassic dragonflies (Insecta: Odonata). Rivista del Museo civico di Scienze Naturali “Enrico Caffi” 19: 31–70.
Beckemeyer, R.J. 2000. The Permian Insect fossils of Elmo, Kansas. The Kansas School Naturalist 46: 3–16.
Béthoux, O. 2015. The Late Carboniferous Triplosoba pulchella is not a fly in the ointment but a stem-mayfly. Systematic Entomology 40: 342–356. Crossref
Béthoux, O. and Beattie, R.G. 2010. Iverya averyi gen. nov. and sp. nov., a new Triadotypomorphan species from the Middle Triassic at Picton, New South Wales, Australia. Acta Geologica Sinica 84: 688–692.Béthoux, O., de la Horra, R., Benito, I.M., Barrenechea, J.F., Galán, A.B., and López-Gómez, J. 2009. A new triadotypomorphan insect from the Anisian (Middle Triassic), Buntsandstein facies, Spain. Journal of Iberian Geology 35: 179–184.
Bordy, E.M., Hancox, P.J., and Rubidge, B.S. 2004. Fluvial style variations in the Late Triassic–Early Jurassic Elliot formation, main Karoo Basin, South Africa. Journal of African Earth Sciences 38: 383–400. Crossref
Cairncross, B., Anderson, J.M., and Anderson, H.M. 1995. Palaeoecology of the Triassic Molteno Formation, Karoo Basin, South Africa-sedimentological and palaeological evidence. South African Journal of Geology 98: 452–478.
Carpenter, F.M. 1992. Superclass Hexapoda. In: R.L. Kaesler (ed.), Treatise on Invertebrate Paleontology, xxii+655 pp. The Geological Society of America, Lawrence and the University of Kansas, Boulder.
Fabricius, J.C. 1793. Entomologia systematica emendata et aucta. Secundum classes, ordines, genera, species adjectis synonimis, locis, observationibus, descriptionibus. Tome 2. viii+519 pp. Proft, C.G., Copenhagen.
Grauvogel, L. and Laurentiaux, D. 1952. Un protodonate du trias des Vosges. Annales de Paleontologie, Invertébrés 38: 121–129.
Handlirsch, A. 1912. Über Insektenrest aus der Trias Frankens. Abhandlungen der Naturhistorischen Gesellschaft zu Nürnberg 18: 79–82.
Lameere, A. 1922. Sur la nervation alaire des insectes. Bulletin de la Classe des Sciences de l’Académie Royale de Belgique 8: 138–149.
Lameere, A. 1923. On the wing-venation of insects. Psyche 30: 123–132. Crossref
Lanci, L., Tohver, E., Wilson, A., and Flint, S. 2013. Upper Permian magnetic stratigraphy of the lower Beaufort Group, Karoo Basin. Earth and Planetary Science Letters 375: 123–134. Crossref
Laurentiaux-Vieira, F., Ricour, J., and Laurentiaux, D. 1952. Un Protodonate du Trias de la Dent de Villard. Bulletin de la Société Géologique de France (6) 1: 319–324.
Li, Y., Béthoux, O., Pang, H., and Ren, D. 2013. Early Pennsylvanian Odonatoptera from the Xiaheyan locality (Ningxia, China): new material, taxa, and perspectives. Fossil Record 16: 117–139, 244. Crossref
Martins-Neto, R.G. 1996. Reinterpretação da venação e revisão das categorias taxonômicas superiores de Gondvanoptilon brasiliensis (= G. brasiliense nom. transl.) Rösler, Rohn & Albamonte, inseto do Paleozóico da bacia do Paraná e Saucrolus silvai Santos artrópode do Cretáceo da bacia Sanfranciscana. Revista Universidade Guarulhos, Série Geociências 1: 42–45.
Nel, A., Bechly, G., Prokop, J., Béthoux, O., and Fleck, G. 2012. Systematics and evolution of Palaeozoic and Mesozoic damselfly-like Odonatoptera of the “protozygopteran” grade. Journal of Paleontology 86: 81–104. Crossref
Nel, A., Béthoux, O., Bechly, G., Martínez-Delclòs, X., and Papier, F. 2001. The Permo-Triassic Odonatoptera of the “protodonate” grade (Insecta: Odonatoptera). Annales de la Société Entomologique de France (N.S.) 37: 501–525.
Nel, A., Fleck, G., Garrouste, R., and Gand, G. 2008. The Odonatoptera of the Late Permian Lodève basin (Insecta). Journal of Iberian Geology 34: 115–122.
Nel, A., Martínez-Delclòs, X., Paicheler, J.C., and Henrotay, M. 1993. Les “Anisozygoptera” fossiles. Phylogénie et classification (Odonata). Martinia hors-série 3: 1–311.
Prevec, R., Labandeira, C.C., Neveling, J., Gastaldo, R.A., Looy, C.V., and Bamford, M. 2009. Portrait of a Gondwanan ecosystem: A new late Permian fossil locality from KwaZulu-Natal, South Africa. Review of Palaeobotany and Palynology 156: 454–493. Crossref
Pritykina, L.N. 1981. New Triassic odonates from Middle Asia [in Russian]. In: V.N. Višniakova, G.M. Dlussky, and L.N. Pritykina (eds.), Novye iskopaemye nasekomye s Territorii SSSR. Trudy Paleontologičeskogo Instituta, Akademiâ Nauk SSSR 183: 5–42.
Reis, O.M. 1909. Handlirschia gelasii nov. gen. et spec. aus dem Schaumkalk Frankens. Abhandlungen der Koeniglich Bayerischen Akademie der Wissenschaften, Mathematisch-Physikalischen Klasse 23: 659–694.
Riek, E.F. and Kukalová-Peck, J. 1984. A new interpretation of dragonfly wing venation based upon Early Upper Carboniferous fossils from Argentina (Insecta, Odonatoidea) and basic character states in pterygota wings. Canadian Journal of Zoology 62: 1150–1166. Crossref
Schlüter, T. 2003. Fossil insects in Gondwana—localities and palaeodiversity trends. Acta Zoologica Cracoviensia 46 (Supplement Fossil Insects): 345–371.
Tankard, A., Welsink, H., Aukes, P., Newton, R., and Stettler, E. 2009. Tectonic evolution of the Cape and Karoo basins of South Africa. Marine and Petroleum Geology 26: 1379–1412. Crossref
Acta Palaeontol. Pol. 62 (3): 613–618, 2017
https://doi.org/10.4202/app.00345.2017