
The Interatheriinae notoungulates from the middle Miocene Collón Curá Formation in Argentina
BÁRBARA VERA, MARCELO REGUERO, and LAUREANO GONZÁLEZ-RUIZ
Vera, B., Reguero, M., and González-Ruiz, L. 2017. The Interatheriinae notoungulates from the middle Miocene Collón Curá Formation in Argentina. Acta Palaeontologica Polonica 62 (4): 845–863.
The Interatheriinae (Notoungulata, Interatheriidae) from the Collón Curá Formation (Colloncuran South American Land Mammal Age, SALMA) are revised here, based on old and new collections from western Neuquén, Río Negro, and Chubut provinces where this geologic unit crops out. After a detailed study of the holotype of Icochilus endiadys, we conclude that its cranial and dental morphology are diagnostic of the genus Protypotherium, and as a result we include I. endiadys in this genus (P. endiadys comb. nov.). Deciduous dentition and postcranial remains are also ascribed to P. endiadys, which allows us to determine its pattern of dental eruption and describe part of its limbs, expanding its diagnosis. In addition, we describe a new species of Protypotherium, P. colloncurensis sp. nov., which differs from P. endiadys in having larger size, a more robust mandible, strongly imbricate upper molars, a well-developed parastyle on P1, a subcircular and non-overlapping p1, a much reduced p2, and a smaller talonid on p3–4. Based on the revision, we identified only one genus of Interatheriinae in the Collón Curá Formation (i.e., Protypotherium). Protypotherium endiadys extends its distribution from Neuquén to Chubut provinces, including Río Negro; the new species, in turn, was only recognized in Río Negro Province, appearing together with P. endiadys in Estancia El Criado, Comallo, and Chico River localities. Discriminant and cladistic analyses were performed including P. endiadys, the new taxon, and other middle Miocene interatheres, and yielded strong support for our systematic conclusions. A thorough phylogenetic analysis of Protypotherium is provided for the first time. Based on this analysis, Protypotherium and Miocochilius form a monophyletic group supported by four synapomorphies.
Key words: Notoungulata, Interatheriidae, Neogene, Patagonia, Argentina, South America.
Bárbara Vera [bvera@mendoza-conicet.gob.ar], Instituto Argentino de Nivología, Glaciología y Ciencias Ambientales (IANIGLA), CONICET, Avenida Ruiz Leal s/n, 5500 Mendoza, Argentina.
Marcelo Reguero [regui@fcnym.unlp.edu.ar], División Paleontología de Vertebrados, Museo de La Plata, Paseo del Bosque s/n. B1900FWA La Plata, Argentina.
Laureano González-Ruiz [gonzalezlaureano@yahoo.com.ar], Centro de Investigación Esquel de Montaña y Estepa Patagónica (CIEMEP), CONICET, UNPSJB, Roca 780, 9200 Esquel, Chubut, Argentina.
Received 17 April 2017, accepted 8 August 2017, available online 27 October 2017.
Copyright © 2017 B. Vera et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Interatheriinae Ameghino, 1897 is a group of small to middle-sized endemic ungulates from South America, which traditionally formed, together with the Notopithecinae, the family Interatheriidae (Simpson 1945; but see Vera 2016). The Interatheriinae are broadly known from South America and their stratigraphic record extends from the early Oligocene (Abanico Formation; Hitz et al. 2000; Flynn et al. 2003) to the late Miocene (Ituzaingó Formation; Cione et al. 2000). Protypotherium Ameghino, 1885 is the most diverse and widely distributed Neogene genus in the southern part of South America. Following the first phylogenetic analysis by Cifelli (1993), who considered the Interatheriinae as a clade, many other subsequent analyses have attempted to resolve interatheriid (and interatheriine) relationships (Hitz et al. 2000, 2006; Reguero et al. 2003; Reguero and Prevosti 2010; Vera 2016; Vera et al. 2017), the most widely accepted hypothesis being that put forward by Hitz et al. (2000, 2006), who proposed a phylogenetic-based node definition for the Interatheriinae, separating this group from other basal interatheriids (i.e., notopithecids such as Ignigena and Johnbell).
Several faunas formed the basis of the middle Miocene (15.97–11.83 Ma) Friasian SALMA (South American Land Mammal Age), an interval that documents the evolution of mammals between the early Miocene (Santacrucian SALMA) and the late Miocene (Chasicoan SALMA) (see Marshall et al. 1983). Use of the Friasian SALMA or Friasian sensu lato was relegated by use of the Colloncuran (including Friasian sensu stricto), Laventan, and Mayoan faunas as successive independent SALMAS (Marshall and Salinas 1990; Vucetich et al. 1993; Flynn and Swisher 1995; Madden et al. 1997; Croft et al. 2016).
Four genera of interatheriids have been mentioned for the middle Miocene faunas (Friasian sensu stricto, Colloncuran, and Mayoan) from high latitudes of South America: Protypotherium Ameghino, 1885; Icochilus Ameghino, 1889; Epipatriarchus Ameghino, 1903; Caenophilus Ameghino, 1903; and Interatherium Ameghino, 1887 (see Bondesio et al. 1980; Pascual et al. 1984). However, these taxa have not been studied at the same level of detail as coeval interatheriids from mid- and low-latitude faunas, such as La Venta in Colombia, and Quebrada Honda and Cerdas in Bolivia (Stirton 1953; Croft 2007; Croft et al. 2009).
The Interatheriinae from the Collón Curá Formation (Colloncuran SALMA) were first described by Roth (1899), who erected two species. This author described Icochilus endiadys based on a nearly complete cranium, but he also mentioned several mandibular fragments, isolated teeth, and a fragment of maxilla, indicating some differences from the holotype, such as the shape of molars which have a more open lingual sulcus and a face nearly straight and extended more anteriorly. Additionally, Roth (1899) described a second species, Nesciotherium indiculus, based on an upper molar similar to that of Interatherium, but he expressed some doubts about its inclusion in the Interatheriidae. This species was later recognized as a mesotheriid (Pascual et al. 1978; Kramarz et al. 2011).
After the initial descriptions by Roth (1899), middle Miocene interatheres from the Collón Curá Formation were only mentioned in faunal lists but not subjected to detailed study. According to the latest works, the Interatheriinae from the Collón Curá Formation include Icochilus endiadys in Collón Curá River in Neuquén Province (see Kramarz et al. 2011 and references herein); Protypotherium sp., Interatherium sp., and Epipatriarchus sp. in Pilcaniyeu Viejo; Protypotherium australe in Cantera Lif Mahuida; Protypotherium sp. at Cerro Castillo; and Protypotherium praerutilum at Cerro Mesa de Trapalcó, all localities in Río Negro Province (Pascual et al. 1984 and references herein; Rolleri et al. 1984; Pardiñas and Reguero 1987). Interatheriinae indet. and Protypotherium sp. were mentioned as occurring in Chico River in Chubut Province (González Ruiz et al. 2012; Vera et al. 2016).
In this contribution, we revise and describe the Interatheriinae from the Collón Curá Formation (Colloncuran SALMA, middle Miocene) of north-western Patagonia (Argentina), including Roth’s (1899) old collection curated at Museo de La Plata, and other old and new specimens recovered from several localities in Neuquén, Río Negro, and Chubut provinces, the details of which remain unpublished. Based on the restudy of the holotype of Icochilus endiadys, we propose taxonomic changes, expand its diagnosis and assign new specimens to this species. In addition, we describe a new species from the Collón Curá Formation in Río Negro Province. Applying multivariate statistics allowed us to assess morphological differences among Miocene species. In addition, a cladistic analysis was performed in order to study the relationships of the Colloncuran species within the Interatheriidae in a broad sense, as well as within the genus Protypotherium and with respect to other middle Miocene interatheriine taxa.
Institutional abbreviations.—LIEB-PV, Laboratorio de Investigaciones en Evolución y Biodiversidad, Paleovertebrados, Esquel, Argentina; MACN-PV, Colección Paleontología de Vertebrados del Museo Argentino de Ciencias Naturales “Bernardino Rivadavia”, Buenos Aires, Argentina; MAPBAR, Museo de la Asociación Paleontológica de Bariloche, San Carlos de Bariloche, Argentina; MLP, Museo de La Plata, Argentina; UCMP, University of California, Museum of Paleontology, Berkeley, USA; YPM-VPPU, Yale Peabody Museum, Vertebrate Paleontology Princeton University Collection, New Haven, USA.
Other abbreviations.—APD, anteroposterior diameter; C/c, upper/lower canine; DP/dp, upper/lower deciduous teeth; H, height; I/i, upper/lower incisor; L, length; M/m, upper/lower molar; P/p, upper/lower premolar; TD, transverse diameter; W, width.
Material and methods
Descriptions and comparisons are based on both specialized bibliographic and personal observations (BV). For teeth, we follow Smith and Dodson (2003). Postcranial bones are described on a single vertical axis irrespective of their anatomical position, defining proximal and distal ends with respect to the distance from the skull, and considering the sagittal plane (lateromedial axis) and transverse plane (anteroposterior axis). Anatomical terminology mainly follows the International Committee on Veterinary Gross Anatomical Nomenclature (2005).
Cranial, bone, and teeth measurements were taken with a Mitutoyo digital calliper (0.01 mm accuracy). Two variables were measured for each tooth: length, corresponds to maximum mesiodistal diameter; width, measured perpendicular to length. We plotted length vs. width of M/m1 in order to compare relative sizes between Protypotherium endiadys and the new species.
With the objective of identifying a priori morphological groups, but based on mathematical algorithms (i.e., the discriminant functions), we performed a discriminant analysis using our set of dental measurements, as well as metrics of specimens from other collections. The six variables included are related to teeth measurements: width of M2, M3, m1, and m3, and the log-transformed length of m2 and m3. Non-normally distributed variables were verified by Kolmogorov-Smirnov test and log-transformed. For each variable, we considered 13 values for Protypotherium endiadys, four values for the new species, P. australe (YPM-VPPU 015349, YPM-VPPU 015350, YPM-VPPU 015357, YPM-VPPU 015598) and Interatherium (YPM-VPPU 015036, YPM-VPPU 015074, YPM-VPPU 015100, YPM-VPPU 015201), and three values for Miocochilius anomopodus (UCMP 39244, UCMP 39231, UCMP 38409). The analysis was conducted using Infostat v. 2016 (Di Rienzo et al. 2016).
The phylogenetic relationships of the new species were assessed using a modified version of the dataset of Vera et al. (2017). This dataset (see SOM 1, Supplementary Online Material available at http://app.pan.pl/SOM/app62-Vera_etal_SOM.pdf) encompassed a total of 21 taxa scored for 51 morphological characters (see SOM 2). Four characters were modified from the original dataset and eight new characters were added (character 43, presence or absence of fossettes on upper molars; character 44, spaces among i3, c and p1; character 45, entepicondylar foramen of humerus; character 46, supratrochlear foramen of humerus; 47, maxilla forms the dorsal border of the orbit; 48, sliver of frontal bone anteriorly projected; 49, parastyle of M1–2; 50, size of M1–3). The characters were considered non-additive and scored for six species of Protypotherium, including P. endiadys and the new taxon, and other interatheriines. In addition, some representatives of Henricosborniidae (Simpsonotus praecursor Pascual, Vucetich, and Fernández, 1978), Notostylopidae (Notostylops murinus Ameghino, 1897), Notopithecidae (Notopithecus adapinus Ameghino, 1897), and Oldfieldthomasiidae (Oldfieldthomasia debilitata Ameghino, 1901) were used as outgroup taxa. An equally weighted parsimony analysis was conducted using TNT 1.1 (Goloboff et al. 2008a, b), a method that minimizes the number of postulated evolutionary transformations. We performed a branch-and-bound search algorithm (implicit enumeration in TNT), which warrants finding all of the most parsimonious trees for a given dataset. Support values were calculated using Bremer and Jackknife indices (the latter is a resampling technique that is summarized using absolute frequencies and GC frequencies; see Goloboff et al. 2003). Nodal support was estimated using the script BREMER.RUN provided by TNT, GC frequency and symmetric resampling (Goloboff et al. 2003).
Geological and geographic setting
The gray tuffaceous sandstone beds outcropping along the inferior valley of the Collón Curá River were first mentioned by Roth (1899) and formally described as the Collón Curá Formation by Yrigoyen (1969); its associated mammalian fauna was the basis for recognition of the Colloncuran SALMA (Marshall and Salinas 1990; Vucetich et al. 1993; Flynn and Swisher 1995).
Later, Rabassa (1975, 1978) and Bondesio et al. (1980) presented a geological description of the Collón Curá Formation in the Pilcaniyeu Viejo area (including Estancia El Criado, Pichileufú River, and Comallo localities) and recognized three members, Caruhué Member at the base, Pilcaniyeu Ignimbrite Member, and Las Bayas Member at the top. In the Ingeniero Jacobacci area, Coira (1979) recognized the Collón Curá Formation formed by La Pava (La Pava Formation = Caruhué Member), Las Chacras and Marileo members, while in Paso Flores and Lipetrén areas, Nullo (1978, 1979) identified the La Pava Formation (La Pava Member = Caruhué Member) and the Collón Curá Formation, which was divided into three facies: lower and upper clastic facies plus a pyroclastic and ignimbrite facies in the middle (Pilcaniyeu Ignimbrite Member = Pilcaniyeu). In turn, Vucetich et al. (1993) exhaustively described the fossiliferous outcrops from the Collón Curán Formation at Cañadón del Tordillo (Neuquén); these authors identified a lower Member which was informally divided into a lower section (including La Pava Member = La Pava Formation = part of Caruhué Member) and an upper section (part of Caruhué Member = lower clastic facies), with the Pilcaniyeu Ignimbrite Member spanning the middle and upper Member. However, other authors (Cucchi et al. 1998; González et al. 2000; Bilmes et al. 2013) deem La Pava and Collón Curá formations as independent units, although González et al. (2000) consider that there could be a gap between both formations.
In general terms, the Collón Curá Formation is characterized by tuff, sandstone, and ignimbrite, bearing fossil mammals below and above the Pilcaniyeu Ignimbrite Member (Vucetich et al. 1993; Cucchi et al. 1998; Escosteguy et al. 2013). A recent 40Ar/39Ar analysis in amphibole crystals collected from fresh pumice clasts revealed an age of 14.86 ± 0.13 Ma for the middle section of the Collón Curá Formation in the Gastre Basin (Bilmes et al. 2013), placing this unit and its fauna in the middle Miocene (Langhian).
The Interatheriinae described here were collected from several localities in Nequén, Río Negro, and Chubut provinces (Fig. 1). In the valley of the Collón Curá River, Cañadón del Tordillo, Pilcaniyeu Viejo, Comallo, and the valley of the Pilcaniyeu River, outcrops are recognized as being from the Collón Curá Formation by the presence of the Pilcaniyeu Ignimbrite Member (Rabassa 1975, 1978; Bondesio et al. 1980; Mazzoni and Stura 1990; Vucetich et al. 1993). In turn, to the south (e.g., Chico River), this marked bed (Pilcaniyeu Ignimbrite Member) is not present and the Collón Curá Formation is identified by its lithological characteristics and relationships with the underlying and overlying units (Ravazzoli and Sesana 1977; Volkheimer and Lage 1981; Mazzoni 1993; González et al. 2000; Bilmes et al. 2013).
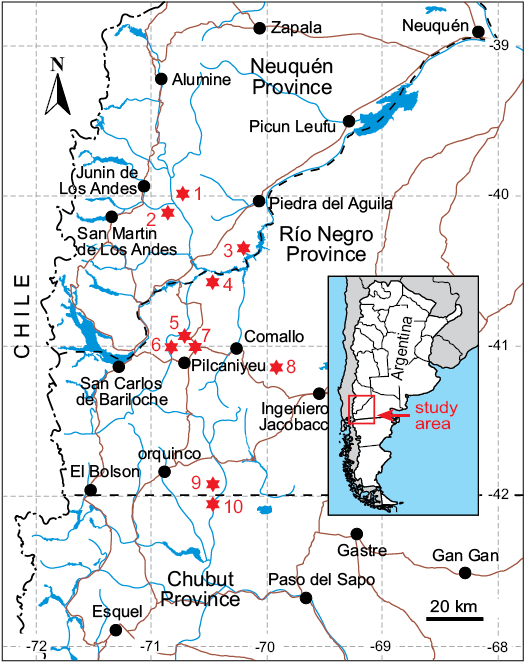
Fig. 1. Map showing localities with outcrops of the Collón Curá Formation. 1, Estancia Collón Curá; 2, Collón Curá River; 3, Cañadón del Tordillo; 4, Cerro Castillo; 5, Estancia El Criado; 6, Pichileufú River; 7, Cerro San Antonio; 8, Comallo; 9, Chico River (Río Negro Province); 10, Chico River (Chubut Province).
Systematic palaeontology
Notoungulata Roth, 1903
Interatheriidae Ameghino, 1897
Interatheriinae Ameghino, 1897
Genus Protypotherium Ameghino, 1885
Type species: Protypotherium antiquum Ameghino, 1885; Huayquerian SALMA, Entre Ríos Province.
Emended diagnosis (from Ameghino 1885 and Sinclair 1909).—Protypotherium differs from Interatherium and Cochilius in having P/p3–P/p4 completely differentiated from molars, presence of mesial sulcus on P2–4, P4 nearly square or wider than long, upper molars usually coequal in size but may slightly diminish in size posteriorly, i3 ≥ i2, lower canine similarly sized to i3, smaller talonid than trigonid on p3–4, sulcus shallow in the talonid of m3, and little or moderately developed descending process of the maxilla. Protypotherium is characterized by having P-shaped trigonid and triangular talonid on m1–2 (in contrast to Interatherium, Cochilius, and Miocochilius), and an entepicondylar foramen in the humerus (absent in Interatherium). Unlike Miocochilius, Protypotherium has a caniniform p1 and a shorter m3.
Protypotherium endiadys (Roth, 1899) comb. nov.
Figs. 2–6.
1899 Icochilus endiadys; Roth 1899: 38–41, pl. 6: 1–4.
1978 Interatherium endiadys; Pascual et al. 1978: 179.
1986 Icochilus andiadys Roth, 1899; Mones 1986: 153.
2011 Icochilus andiadys Roth, 1899; Kramarz et al. 2011: 559, fig. 2F.
Holotype: MLP 12-2886, skull with left I1–M3 and right P2–M3 (Fig. 2A).
Type locality: Collón Curá River, Neuquén Province, Argentina (Fig. 1).
Type horizon: Collón Curá Formation, middle Miocene (Langhian, ages ranging 14.0–15.5 Ma, according to Flynn and Swisher 1995).
Material.—Collón Curá River: MLP 12-2887: a, right mandible with c–p3; b, right mandible with m2–3; c, left mandible with m1–2 (a, b, same individual); MLP 12-2916: a, right mandible with m2–3; b, left mandible with m1–3; c, right i1–2 and left i2; d, right lower canine; e, right astragalus (a–e, same individual). Estancia Collón Curá: MLP 90-IX-2-170, mandible with left m3 and right i1–c and p2–3; MLP 90-XII-17-20, left maxilla with P3–M3. Sur balsa Collón Curá: MLP 90-XII-22-1, left mandible with p4–m3. Cañadón del Tordillo: MLP 91-IV-1-47, right mandible with m1–3. Pichileufú River: MLP 73-VII-7-1, right maxilla with P3–M3, mandible with right i3–p2 and left p1–2, left mandible with m1–2, and proximal of ulna; MLP 73-VII-8-4, right mandible with p3–m2. Estancia El Criado: MLP 73-V-III-63, skull with right P1–M3 and left P3–M3; MLP 73-VII-6-3, palate with right P1–M3 and left P3–M3; MLP 73-VII-6-5, right mandible with p4–m1; MLP 73-VII-6-6, right mandible with m1–2; MLP 73-VII-6-8, left maxilla with P3–M3; MLP 73-VII-6-9, mandible with left i1 and right i1–m1; MLP 73-VII-8-6, right mandible with m1–3; MLP 73-VII-9-1, skull with left I1–M1 and right I1–C, P3–M3; MLP 74-II-1-4, left mandible with p2–m3; MLP 74-II-1-16 (originally assigned to Interatherium sp.), left mandible with m2–3; MLP 92-V-10-106: a, left mandible with m1–3; b, right mandible with p4–m2; c, right i3–p2 and left p2; d, left maxilla with DP2–DP4 and M1; e, mandible with right dp4–m1 and left dp3–4–m1; MLP 92-V-10-108, left hemipalate with P1–M3; MLP 92-X-10-119: 1, right mandible with m1–3; 2, right maxilla with P3–M2; 6, left maxilla with P3–M3; 7, right maxilla with M2–3; 8, left maxilla with M2–3; 9, left maxilla with M2–3; 10, right mandible with p2–4; 12, left mandible with m3; 14, left mandible with p2–m3; 19, right mandible with m1–3; 23, right mandible with p3–4 and erupting m1; 24, mandible with left i1–c, p2–m1, and right i1–c, p2–3; 25, right maxilla with M1–3; 26, left mandible with p4–m3; 27, left mandible with p4–m3; 28, left maxilla with M1–3; 30, left mandible with m1–3; 32, left mandible with m2–3; 33, left mandible with p2–m1; 34, right mandible with p4–m2; 36, left mandible with p2–m2; 38, right maxilla with P1–DP2 and P3–M1; 43, right maxilla with P3–M2; 44, left mandible with p2–m1; 45, left mandible with m1–3; 46, left mandible with p4–m1; 47, left mandible with m2–3; 48, right mandible with p2–4; 49, left mandible with p4–m3. Cerro Castillo: MLP 90-XII-24-34 (originally assigned to Protypotherium sp.): a, left mandible with m1–2; b, left mandible with m3; c, right mandible with p3–4; d, left mandible with pd2–dp4. Comallo: MAPBAR 947, left hemipalate with P1–M3; MAPBAR 953, left mandible with m1–3; MAPBAR 955, right maxilla with P3–M3; MAPBAR 958, left mandible with m1–3; MAPBAR 959, left maxilla with P1–M3; MAPBAR 960, left mandible with m1–3; MAPBAR 965, mandible with right i1, p1–m3, and left i1–c, p2–m3; MAPBAR 1381, skull with left I3–M3 and right I1–P2, P4–M3, and distal of humerus. Chico River (Río Negro Province): MLP 92-XI-16-17, right mandible with p2–m2. Chico River (Chubut Province): LIEBPV 7002, right maxilla with P3–M3. Collón Curá Formation (Neuquén, Río Negro, and Chubut provinces, Argentina), middle Miocene. Figure 1 details the localities referenced here, except for Sur balsa Collón Curá (from Roth 1899), which was not precisely located.
Emended diagnosis.—Protypotherium endiadys is characterized by: maxilla extended dorsally over the orbit; anterior process of the frontal bone is lacking; small descending process of the maxilla; slender and relatively low mandible; reduced P/p1; nearly equally-sized and barely imbricated upper molars, which have fossettes and a deep and simple lingual sulcus; well-developed posterior lobe in M3; no diastemata between the i3 and canine, or between c and p1; i1–c are moderately procumbent; p1 caniniform; p2–3 with talonid much smaller than trigonid; and lower molars with a similarly-sized trigonid. It differs from P. australe in being smaller, having a shorter rostrum and longer basicranium in relation to skull length, less imbricated upper incisors, non-overlapping P/p1, humerus with deeper trochlea and supratrochlear foramen, ulna with larger trochlear notch/olecranon ratio, and less concave articular surface of the astragalar trochlea.
Description.—Skull and upper dentition: The MLP 12-2886 specimen is a nearly complete cranium with left I1–M3 and right P2–M3; except for the broken zygomatic arches, it is in good condition, preserving part of the palate, basicranium, nasals, frontal bones, fragmentary parietals and posterior part of skull (Fig. 2A; Table 1). It should be noted that the maxilla in MLP 12-2886 extends dorsally over the orbit (here named dorsal process of the maxilla) and no anterior process of the frontal bone is observed (Fig. 2A1, A2), such as was also noted by Roth (1899). This feature differs from other Miocene interatheres, such as Protypotherium autrale, Interatherium robustum and Miocochilius anomopodus, where the frontal bone projects anteriorly like a sliver, separating the maxilla from the orbit (Vera 2016). The rostrum of MLP 12-2886 is shorter and wider than that of Miocochilius anomopodus (UCPM 38091), which is narrow and elongated. The bullae of MLP 12-2886 are small relative to skull size (Fig. 2A2, A4); the antorbital foramen is placed at M1 level; orbits are large, having a diameter of 1/3 the length of the skull. The descending process of the maxilla is little developed (Fig. 2A2, A4), differing from the moderate process in Protypotherium australe and the much more developed process of Interatherium. In ventral view, the palate has a transverse fracture causing a separation between I2–3 (Fig. 2A4). Permanent upper and lower cheek-teeth are euhypsodont, as is typical of Miocene interatheres.
The I1 is the largest of incisors (Fig. 2B; Table 1); I1–I3 are elongated, labiolingualy flattened, and slightly narrowing posteriorly. The canine is labiolingually flattened, with a wide and labial convex anterior part, and a narrow, flattened and longer posterior part; it differs from the incisors because it is labially bilobed and lingually convex, instead of having the straighter faces of incisors (Fig. 2B).
In the holotype, an old individual, P1 is a reduced, subcircular in outline and non-overlapping tooth (Fig. 2A3, A4), differing from the canine and the other premolars (see below). In younger individuals (e.g., MLP 73-V-III-63 and MLP 92-V-10-108), however, P1 is not as reduced as in MLP 12-2886, and it has an incipient parastyle, like in P2. P2–4 (Fig. 2A3, A4, C) are premolariform, nearly square, and completely differentiable from molars; in addition, they have a strongly folded ectoloph, and a mesial sulcus that forms two small lobes in the mesial face; this sulcus is more evident in P3–4. A tiny fossette is present in P2–4 (Fig. 2A3, A4).
Molars, in turn, are rectangular and equally sized (Fig. 2A3, A4, C; Table 1); they have a smoothly undulate ectoloph and a deep lingual sulcus divides them into two lobes. In M1, the lingual sulcus is deeper than in M2–3. In M1–2, the posterior lobe is longer than the anterior one, whereas the condition is opposite in M3. The imbrication is remarkable between P4–M1 and M1–2. There are two fossettes, one in each lobe of the molars (Fig. 2A3, A4). There is a thick layer of cementum on the lingual face of P2–M3.

Fig. 2. Skull and upper dentition of interatheriine notoungulate Protypotherium endiadys (Roth, 1899) from the Collón Curá Formation, middle Miocene, Patagonia, Argentina. A. MLP 12-2886 (holotype), skull with left I1 (broken), I2–M3 and right P2–M3 in dorsal (A1), lateral (A2), and occlusal (A4) views; A3, explanatory drawing. B. MLP 73-VII-9-1, left I1–C in occlusal view (B2); B1, explanatory drawing. C. LIEBPV 7002, maxilla with right P3–M3 in occlusal view.
LIEBPV 7002 (Fig. 2C) is the only specimen recovered from exposures of the Collón Curá Formation at Chico River locality, Chubut Province (Fig. 1). It is a fragment of maxilla with P3–M3 comparable in size and morphology to MLP 12-2886 and other specimens associated with the Protypotherium endiadys-morphotype, such as MAPBAR 959 and MAPBAR 1381 (Table 1).
Protypotherium endiadys differs from the Laventan species of the genus Miocochilius; M. federicoi Croft, 2007 has a better developed P1 and a lingual sulcus internally bifurcated on M1–2; in contrast, M. anomopodus Stirton, 1953 has more elongated premolars and a shallow lingual sulcus on molars. In addition, the early/middle Miocene interathere specimen referred to as Protypotherium sp. nov. from Cerdas (Bolivia; Croft et al. 2009: fig. 8C) has longer than wide P2–3 and a tiny middle lobe in M1. Regarding younger Chasicoan interatheres, Protypotherium endiadys has a square and better developed posterior lobe in M3, whereas the MLP 12-2177 specimen (P. minutum Cabrera and Kraglievich, 1931) is smaller and its M3 has a posterior lobe triangular-shaped, which is much larger than the anterior lobe; in turn, P. distinctum Cabrera and Kraglievich, 1931 (MLP 12-2178) is characterized by its larger size (Table 1) and more imbricated upper molars, which have a peculiar bifurcated lingual sulcus.
Table 1. Upper tooth dimensions (in mm) of Protypotherium endiadys. *deciduous teeth; approximate values in parentheses; L, length; W, width.
| |
I1 |
I2 |
I3 |
C |
P1 |
P2 |
P3 |
P4 |
M1 |
M2 |
M3 |
|||||||||||
|
L |
W |
L |
W |
L |
W |
L |
W |
L |
W |
L |
W |
L |
W |
L |
W |
L |
W |
L |
W |
L |
W |
|
|
MLP 12-2886 |
6.0 |
2.0 |
4.0 |
2.0 |
3.0 |
2.0 |
5.0 |
2.0 |
3.0 |
2.0 |
3.0 |
3.0 |
4.0 |
3.0 |
4.0 |
3.0 |
6.0 |
4.0 |
– |
– |
6.0 |
3.0 |
|
MLP 73-V-III-63 |
|
|
|
|
|
|
|
|
3.0 |
2.2 |
3.3 |
2.7 |
2.9 |
3.1 |
3.4 |
3.1 |
5.3 |
3.4 |
5.0 |
3.2 |
5.0 |
2.7 |
|
MLP 73-VII-6-3 |
|
|
|
|
|
|
|
|
2.9 |
2.0 |
3.6 |
2.7 |
3.2 |
3.0 |
3.8 |
3.3 |
5.4 |
3.4 |
5.0 |
3.2 |
5.1 |
2.7 |
|
MLP 73-VII-6-8 |
|
|
|
|
|
|
|
|
|
|
|
|
(3.1) |
3.3 |
3.7 |
3.5 |
5.4 |
3.4 |
4.6 |
3.2 |
4.5 |
2.9 |
|
MLP 73-VII-7-1 |
|
|
|
|
|
|
|
|
|
|
|
|
– |
3.4 |
3.6 |
4.0 |
6.1 |
4.6 |
|
|
|
|
|
MLP 73-VII-9-1 |
5.4 |
– |
4.2 |
1.8 |
4.2 |
1.8 |
4.3 |
2.0 |
3.1 |
3.6 |
3.9 |
4.1 |
3.6 |
3.4 |
4.4 |
3.9 |
7.0 |
4.2 |
6.0 |
3.9 |
6.3 |
3.5 |
|
MLP 90-XII-17-20 |
|
|
|
|
|
|
4.0 |
(2.2) |
3.0 |
(2.2) |
– |
– |
3.2 |
4.2 |
4.4 |
– |
6.7 |
4.7 |
6.1 |
4.0 |
6.4 |
3.8 |
|
MLP 92-V-10-106d |
|
|
|
|
|
|
|
|
|
|
3.4* |
2.2* |
4.0* |
2.6* |
4.4* |
2.4* |
4.8 |
2.3 |
|
|
|
|
|
MLP 92-V-10-108 |
|
|
|
|
|
|
|
|
2.7 |
2.9 |
3.3 |
3.3 |
3.1 |
3.9 |
4.1 |
4.0 |
6.0 |
4.2 |
6.0 |
4.1 |
4.9 |
3.4 |
|
MLP 92-X-II-119.2 |
|
|
|
|
|
|
|
|
|
|
|
|
(3.9) |
4.1 |
4.9 |
4.3 |
6.8 |
4.3 |
6.3 |
4.5 |
|
|
|
MLP 92-X-II-119.6 |
|
|
|
|
|
|
|
|
|
|
|
|
3.9 |
4.4 |
6.9 |
4.6 |
6.1 |
3.9 |
6.1 |
3.4 |
|
|
|
MLP 92-X-II-119.7 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
5.0 |
3.0 |
|
MLP 92-X-II-119.8 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
6.4 |
3.8 |
|
MLP 92-X-II-119.9 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
5.1 |
3.6 |
5.3 |
3.4 |
|
MLP 92-X-II-119.25 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
6.6 |
3.4 |
6.0 |
3.8 |
5.9 |
4.5 |
|
MLP 92-X-II-119.28 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
5.4 |
3.6 |
4.4 |
3.0 |
|
|
|
MLP 92-X-II-119.38 |
|
|
|
|
|
|
|
|
3.4 |
2.2 |
3.6* |
3.1* |
– |
– |
(3.8) |
(2.5) |
(4.3) |
– |
|
|
|
|
|
MLP 92-X-II-119.43 |
|
|
|
|
|
|
|
|
|
|
|
|
3.8 |
3.5 |
4.2 |
3.6 |
6.9 |
4.4 |
|
|
|
|
|
MAPBAR-955 |
|
|
|
|
|
|
|
|
|
|
|
|
4.1 |
3.6 |
3.8 |
3.6 |
– |
– |
6.0 |
3.6 |
|
|
|
MAPBAR-959 |
|
|
|
|
|
|
|
|
2.8 |
2.5 |
3.16 |
3.4 |
3.6 |
3.9 |
3.9 |
4.3 |
5.9 |
4.3 |
5.8 |
4.5 |
5.6 |
4.0 |
|
MAPBAR-1381 |
4.8 |
1.2 |
3.6 |
1.7 |
3.5 |
1.8 |
3.6 |
1.7 |
3.6 |
1.9 |
3.2 |
2.6 |
– |
– |
3.8 |
3.6 |
– |
3.6 |
5.2 |
3.3 |
5.0 |
3.0 |
|
LIEBPV 7002 |
|
|
|
|
|
|
|
|
|
|
|
|
3.4 |
3.2 |
3.9 |
3.4 |
5.1 |
3.5 |
4.8 |
3.2 |
– |
2.8 |
|
Data sample |
3 |
2 |
3 |
3 |
3 |
3 |
4 |
4 |
9 |
9 |
7 |
7 |
12 |
13 |
15 |
14 |
16 |
16 |
14 |
14 |
13 |
13 |
|
Mean |
5.4 |
1.6 |
3.9 |
1.8 |
3.6 |
1.9 |
4.2 |
2.0 |
3.1 |
2.4 |
3.4 |
3.1 |
3.5 |
3.6 |
4.2 |
3.7 |
5.9 |
3.9 |
5.5 |
3.7 |
5.5 |
3.4 |
Mandible and lower dentition: The lots MLP 12-2887 and MLP 12-2916 were both collected by Santiago Roth in Collón Curá River, the type locality of the Collón Curá Formation (Neuquén Province). Indeed, Roth (1899: 40) mentioned in the description of “Icochilus” endiadys that he had also found several fragments of mandibles and isolated teeth; however, he did not describe these specimens. Based on that, we infer that MLP 12-2887 and MLP 12-2916 are most likely associated with the MLP 12-2886 skull (holotype of “Icochilus” endiadys, see above). In addition, the finding of associated upper and lower dentitions in the sample herein presented (e.g., MLP 73-VII-7-1) allows us to recognize and describe the lower dentition for Protypotherium endiadys.
MLP 12-2887 comprises three fragments of mandible. In MLP 12-2887a (Fig. 3A), the canine is V-shaped and labiolingually flattened; p1 is mesially broken, wider than the canine (Table 2); and p2 is similar to that of MLP 74-II-1-4 from Río Negro. MLP 12-2887b is probably associated with MLP 12-2887a and comparable to MLP 12-2887c (left m1–2; Fig. 3C). MLP 12-2916a and MLP 12-2916b (Fig. 3B) are very similar to MLP 74-II-I-4 (Fig. 3F; Table 2). In MLP 12-2916c, i1 has its tip broken and the base is circular in outline; i2 is proximally flattened and the outline of the base is circular, a shallow lingual sulcus divides the occlusal surface in a short mesial part and in a longer distal part. The p2–3 of MLP 12-2887a (Fig. 3A) have the talonid much more reduced in relation to the trigonid compared to Miocochilius anomopodus (UCPM 38409), where the difference in size is not so evident between trigonid and talonid, which are separated by a wider sulcus. Another difference is a simpler and more conical p1 in MLP 12-2887a versus a larger and better developed p1 in the Laventan specimen. The interatheres from Cerdas (Croft et al. 2009) differ from Protypotherium endiadys in having a more rudimentary p2 like in MLP 12-2887a, and m3 lacks labial sulcus on the talonid (present in MLP 12-2887b and MLP 12-2916b).
The i1–2 are circular in outline, with i1 being more flattened laterally and smaller than i2 (Fig. 3D; Table 2); i3, in turn, is longer than i2 and compressed labiolingually (Fig. 3D, E; Table 2); the c is V-shaped, labiolingualy flattened and longer than i3 (Fig. 3A, D, E; Table 2). The i1–c are moderately procumbent. The mandibular symphysis has a triangular shape, very narrow at incisor level (Fig. 3E). The p1 is similar to the canine, but labially more convex with a shallow deeper lingual sulcus (Fig. 3E); p1 is overlapped by the canine (Fig. 3D).
Table 2. Lower tooth dimensions (in mm) of Protypotherium endyadys. *deciduous teeth; approximate values in parentheses; L, length; W, width.
| |
i1 |
i2 |
i3 |
c |
p1 |
p2 |
p3 |
p4 |
m1 |
m2 |
m3 |
|||||||||||
|
L |
W |
L |
W |
L |
W |
L |
W |
L |
W |
L |
W |
L |
W |
L |
W |
L |
W |
L |
W |
L |
W |
|
|
MLP 12-2887a |
|
|
|
|
|
|
3.5 |
1.3 |
(2.7) |
1.9 |
3.8 |
2.4 |
4.2 |
2.8 |
4.4 |
– |
|
|
|
|
|
|
|
MLP 12-2887b |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
5.9 |
3.6 |
7.4 |
2.9 |
|
MLP 12-2887c |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
5.0 |
3.1 |
5.1 |
3.2 |
|
|
|
MLP 12-2916a |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
5.2 |
3.5 |
7.6 |
2.7 |
|
MLP 73-VII-6-5 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
4.3 |
3.3 |
6.1 |
3.8 |
|
|
|
|
|
MLP 73-VII-6-6 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
– |
3.6 |
5.7 |
3.5 |
|
|
|
MLP 73-VII-6-9 |
1.6 |
1.8 |
2.2 |
1.9 |
3.3 |
2.3 |
3.7 |
2.0 |
3.4 |
1.6 |
4.1 |
2.4 |
3.7 |
– |
4.3 |
3.4 |
|
|
|
|
|
|
|
MLP 73-VII-7-1 |
|
|
1.5 |
1.7 |
2.7 |
1.6 |
3.7 |
1.8 |
3.2 |
1.6 |
|
|
|
|
|
|
5.8 |
3.0 |
5.5 |
3.2 |
|
|
|
MLP 73-VII-8-6 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
6.2 |
3.5 |
6.3 |
3.3 |
8.1 |
2.8 |
|
MLP 73-VII-8-4 |
|
|
|
|
|
|
|
|
|
|
|
|
– |
2.4 |
4.1 |
2.6 |
5.4 |
3.1 |
5.4 |
2.9 |
|
|
|
MLP 74-II-1-4 |
|
|
|
|
|
|
|
|
|
|
2.9 |
2.1 |
3.8 |
2.9 |
3.8 |
3.4 |
5.4 |
3.4 |
5.6 |
3.4 |
7.6 |
3.2 |
|
MLP 74-II-1-16 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
5.3 |
3.4 |
7.0 |
3.2 |
|
MLP 90-IX-2-170 |
1.5 |
2.4 |
1.4 |
2.6 |
2.2 |
2.8 |
3.4 |
2.2 |
– |
– |
3.8 |
2.4 |
4.2 |
2.9 |
|
|
|
|
|
|
6.6 |
2.8 |
|
MLP 90-XII-22-1 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
4.1 |
2.9 |
5.7 |
3.1 |
5.3 |
3.3 |
8.2 |
2.7 |
|
MLP 90-XII-24-34a |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
5.8 |
3.4 |
5.6 |
3.2 |
|
|
|
MLP 90-XII-24-34b |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
6.7 |
2.9 |
|
MLP 90-XII-24-34c |
|
|
|
|
|
|
|
|
|
|
|
|
4.4 |
2.5 |
4.0 |
3.0 |
|
|
|
|
|
|
|
MLP 91-IV-1-47 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
6.4 |
3.9 |
5.7 |
3.7 |
7.6 |
3.4 |
|
MLP 92-V-10-106a |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
4.6 |
3.1 |
4.5 |
3.1 |
6.2 |
2.4 |
|
MLP 92-V-10-106b |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
5.7 |
3.3 |
|
|
|
|
|
MLP 92-V-10-106c |
1.2 |
1.7 |
3.2 |
1.8 |
4.6 |
1.5 |
4.7 |
1.8 |
4.1 |
1.7 |
4.5 |
2.0 |
|
|
|
|
|
|
|
|
|
|
|
MLP 92-V-10-106e |
|
|
|
|
|
|
|
|
|
|
|
|
3.9* |
2.1* |
4.0* |
2.2* |
4.0 |
1.9 |
|
|
|
|
|
MLP 92-X-10-119.1 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
5.6 |
3.9 |
– |
– |
7.9 |
3.2 |
|
MLP 92-X-10-119.10 |
|
|
|
|
|
|
|
|
|
|
|
|
– |
2.5 |
4.1 |
2.9 |
|
|
|
|
|
|
|
MLP 92-X-10-119.12 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
8.0 |
3.5 |
|
MLP 92-X-10-119.14 |
1.6 |
1.9 |
2.2 |
2.2 |
2.9 |
2.0 |
3.4 |
1.8 |
– |
– |
3.5 |
2.3 |
4.0 |
3.2 |
4.5 |
3.4 |
5.7 |
3.6 |
6.1 |
3.6 |
7.6 |
3.2 |
|
MLP 92-X-10-119.19 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
5.8 |
3.3 |
5.3 |
3.5 |
|
|
|
MLP 92-X-10-119.23 |
|
|
|
|
|
|
|
|
|
|
|
|
3.2 |
2.2 |
4.0 |
– |
|
|
|
|
|
|
|
MLP 92-X-10-119.24 |
1.5 |
2.1 |
1.8 |
2.3 |
2.4 |
2.4 |
3.4 |
2.3 |
– |
– |
3.4 |
2.3 |
4.0 |
2.7 |
4.0 |
3.2 |
5.7 |
3.4 |
|
|
|
|
|
MLP 92-X-10-119.26 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
5.8 |
3.7 |
5.4 |
3.8 |
7.3 |
3.1 |
|
MLP 92-X-10-119.27 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
5.9 |
3.5 |
5.4 |
3.4 |
|
|
|
MLP 92-X-10-119.30 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
4.5 |
2.4 |
4.3 |
2.4 |
|
|
|
MLP 92-X-10-119.32 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
5.7 |
3.3 |
6.7 |
3.1 |
|
MLP 92-X-10-119.33 |
|
|
|
|
|
|
|
|
|
|
3.7 |
2.4 |
4.0 |
2.6 |
3.4 |
3.1 |
5.5 |
3.5 |
|
|
|
|
|
MLP 92-X-10-119.34 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
3.7 |
2.6 |
4.9 |
3.3 |
5.5 |
3.3 |
|
|
|
MLP 92-X-10-119.36 |
|
|
|
|
|
|
|
|
|
|
3.6 |
2.3 |
3.6 |
2.4 |
4.6 |
2.9 |
– |
– |
5.8 |
3.2 |
|
|
|
MLP 92-X-10-119.44 |
|
|
|
|
|
|
|
|
|
|
4.1 |
1.9 |
3.9 |
2.6 |
4.2 |
3.1 |
5.7 |
3.2 |
|
|
|
|
|
MLP 92-X-10-119.45 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
5.2 |
3.3 |
5.4 |
3.4 |
– |
3.0 |
|
MLP 92-X-10-119.47 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
5.7 |
3.0 |
6.5 |
2.5 |
|
MLP 92-X-10-119.48 |
|
|
|
|
|
|
|
|
|
|
3.7 |
2.3 |
3.9 |
3.0 |
4.3 |
3.4 |
|
|
|
|
|
|
|
MLP 92-X-10-119.49 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
5.9 |
2.9 |
5.8 |
2.7 |
7.3 |
2.2 |
|
MLP 92-XI-16-17 |
|
|
|
|
|
|
|
|
|
|
4.0 |
2.5 |
4.3 |
2.9 |
|
|
|
|
|
|
|
|
|
MAPBAR 953 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
5.5 |
3.8 |
5.7 |
3.8 |
7.1 |
3.4 |
|
MAPBAR 960 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
5.7 |
3.6 |
5.4 |
3.6 |
7.0 |
3.6 |
|
MAPBAR 965 |
|
|
|
|
|
|
|
|
|
|
2.8 |
2.0 |
3.3 |
2.6 |
4.2 |
3.1 |
5.2 |
3.5 |
5.8 |
3.5 |
7.5 |
2.8 |
|
Data sample |
5 |
5 |
6 |
6 |
6 |
6 |
7 |
7 |
4 |
4 |
13 |
13 |
14 |
15 |
17 |
15 |
27 |
28 |
27 |
27 |
20 |
21 |
|
Mean |
1.5 |
2.0 |
2.1 |
2.1 |
3.0 |
2.1 |
3.7 |
1.9 |
3.4 |
1.7 |
3.7 |
2.3 |
3.9 |
2.7 |
4.1 |
3.1 |
5.5 |
3.3 |
5.5 |
3.3 |
7.3 |
3.0 |
The p2 of MLP 74-II-1-4 is a small version of p3–4 (Fig. 3F), with a narrower trigonid, reduced talonid, and shallower labial and lingual sulci. The same characteristics are present in the same tooth of MLP 90-IX-2-170. The p3–4 are larger and better developed than p2, mainly in the trigonid (Fig. 3F). The trigonid has a P-shaped outline with a very convex and inclined labial face, a concave distal face, and a smoothly undulated mesial face; the talonid, in contrast, is reduced and smaller than the trigonid. Labial and lingual sulci in p3–4 are deeper than in p2.
The m1–2 are indistinguishable from each other; they differ from p3–4 in having a larger talonid, and deeper labial and lingual sulci (Fig. 3F1, F2); like p3–4, they have a P-shaped trigonid, but shorter in comparison; the talonid is triangular in outline, with the distal face concave at the point where it contacts the following trigonid tooth; in turn, the lingual face of the talonid is slightly concave. The m3 has a trigonid similar to that of m1–2, but differs from them in having a longer talonid, which is lingually and labially trilobed (Fig. 3F1, F2). In premolars and molars, the cement mainly covers labial faces and the enamel is lacking in the mesial face of the trigonid and in the distal face of the talonid (Fig. 3F2). The height of the horizontal ramus is nearly constant anteroposteriorly (Fig. 3F3).
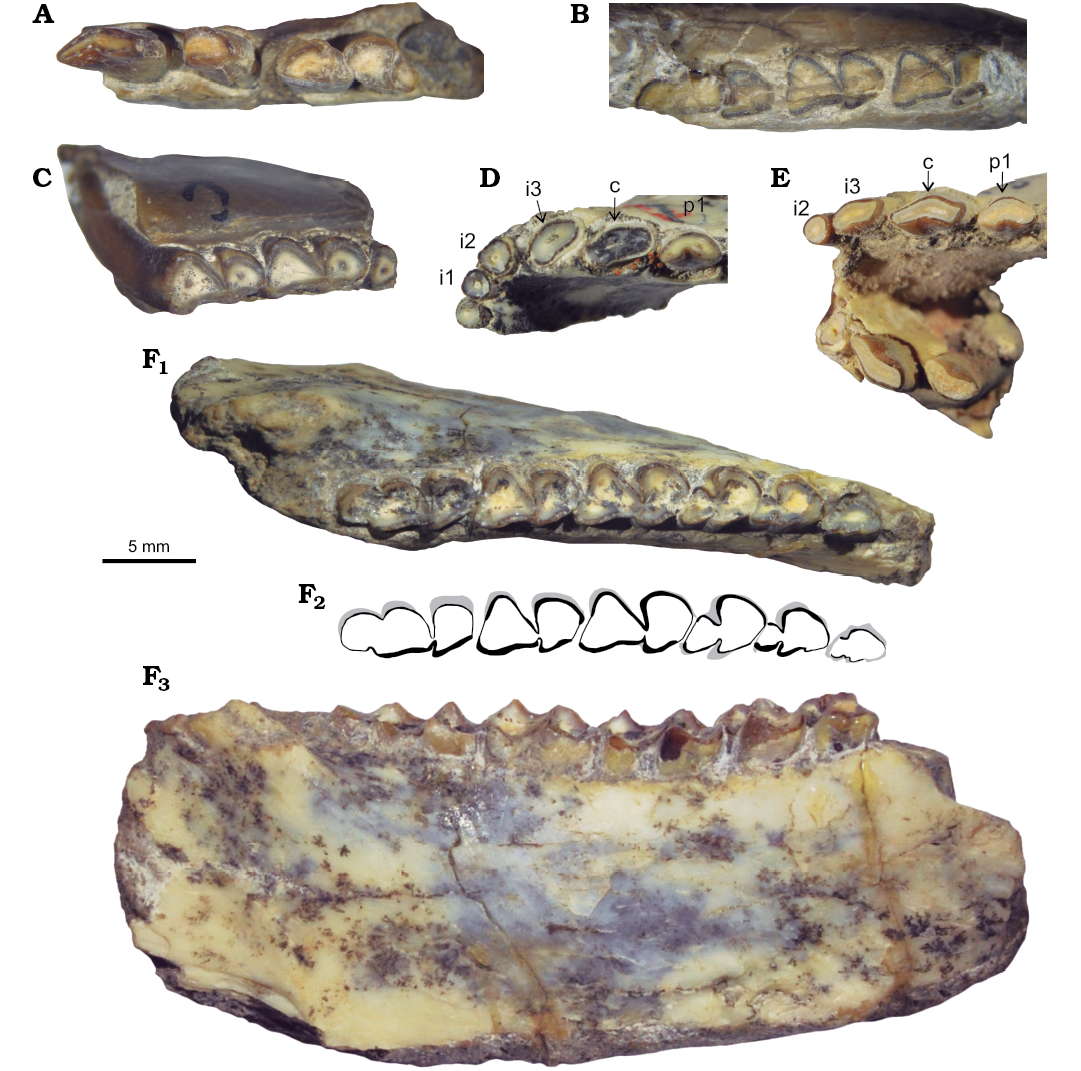
Fig. 3. Lower dentition of interatheriine notoungulate Protypotherium endiadys (Roth, 1899) from the Collón Curá Formation, middle Miocene, Patagonia, Argentina. A. MLP 12-2887a, mandible with right c–p3 in occlusal view. B. MLP 12-2916b, mandible with left m1–3 in occlusal view. C. MLP 12-2887c, mandible with left m1–2 in occlusal view. D. MLP 73-VII-6-9, mandible with left i1 and right i1–p1 in occlusal view. E. MLP 73-VII-7-1, mandible with right i3–p2 in occlusal view. F. MLP 74-II-I-4, mandible with left p2–m3 in occlusal (F1) and lateral (F3) views; F2, explanatory drawing.
Deciduous dentition and pattern of eruption: Unlike permanent dentition, deciduous teeth are low-crowned and develop roots. MLP 92-V-10-106d (DP2–4 and M1; Fig. 4A) and MLP 92-X-10-119.40 (DP2–4) are very similar to each other (Table 1). DP2–3 share with the permanent homologous teeth a deep mesial sulcus and a much folded ectoloph, but differ from them in being mesiodistally elongated and narrower (Table 1). DP4, in turn, resembles a molar, with a rectangular outline, being larger than wide (Fig. 4A1, A2; Table 1); it has a lingual sulcus (instead of the mesially placed sulcus in DP2–3) dividing the tooth into two lobes, but the anterior lobe is much more reduced than the posterior lobe, contrasting with M1. The M1 of MLP 92-V-10-106d is a not fully-erupted tooth, having one of the smallest dimension values of the sample (Fig. 5); it has two very shallow and small fossettes, one in each lobe. MLP 92-X-10-119.38 (Fig. 4C) is also a juvenile specimen, but it is in a bit more mature stage than MLP 92-V-10-106d. MLP 92-X-10-119.38 has its P1, P2, and M1 fully erupted (but broken), and P3–4 are still inside the maxilla, with P3 being lower than P4 (Fig. 4C). In lateral view, we can observe the germ of P2 below DP2. It is interesting to note that MLP 92-X-10-119.38 corroborates that the sequence of eruption for permanent premolars is in posterior-anterior direction, from P4 to P2. A similar pattern was described for some notopithecids (Vera and Cerdeño 2014), as well as for Oligocene and Miocene interatheres (Vera 2016; Vera et al. 2017).
Regarding lower dentition, MLP 92-V-10-106e comprises two fragments of mandible with right dp4–m1 (Fig. 4B1, B2, B5) and left dp3–4 (Fig. 4B3, B4). They are very probably associated with the MLP 92-V-10-106d maxillary fragment (see above). The dp3 has a larger and more developed talonid than p3; in turn, dp4 has a molarized talonid, larger and wider than the talonid of p4, and its trigonid is longer and narrower than in m1. In MLP 92-V-10-106e, m1 is little worn; it has a shallow, long and narrow sulcus placed at the distolingual corner of the talonid, which quickly disappears with wear (Fig. 4B1, B2). Both MLP 92-V-10-106e and MLP 92-X-10-119.30 (m1–3) are juvenile individuals, showing smaller measurements for m1 (Fig. 5; Table 2). The MLP 92-X-10-119.18 specimen has its p3 inside the bone and p4 nearly fully-erupted, corroborating that p4 erupts before p3, as it was described for upper teeth (see above). In addition, MLP 92-X-10-119.49 (Fig. 4D) has its p4 (partially broken) inside the bone, while m3 is erupting, indicating that dp4 is maintained throughout m3 eruption, such as happens in other interatheriines with a similar pattern of permanent premolar eruption (Vera 2016).
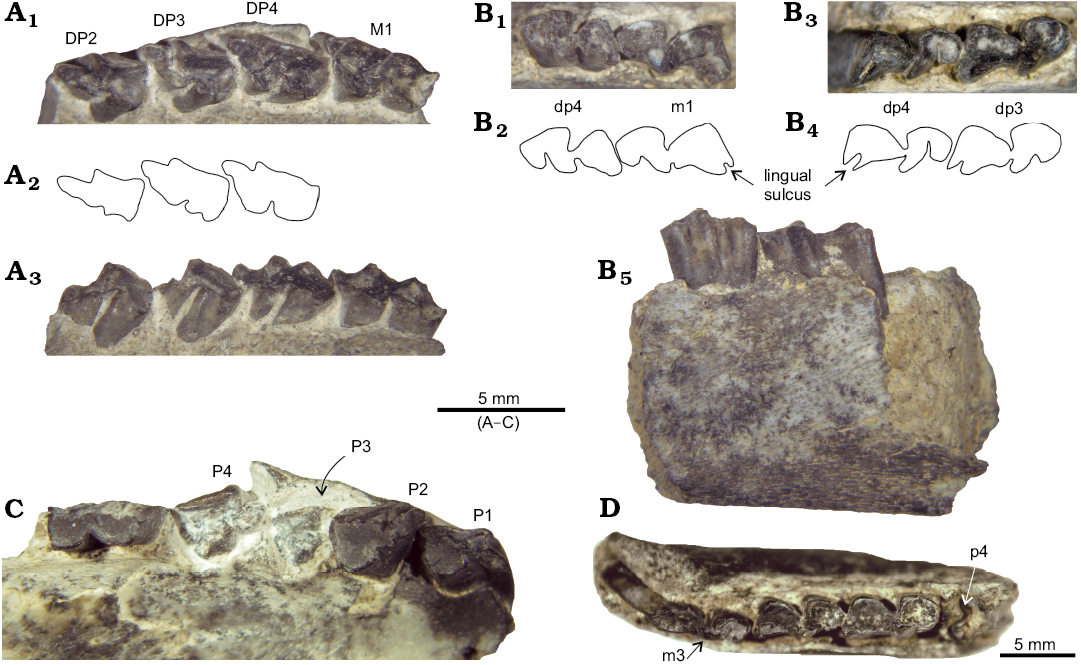
Fig. 4. Deciduous dentition of interatheriine notoungulate Protypotherium endiadys (Roth, 1899) from the Collón Curá Formation, middle Miocene, Patagonia, Argentina. A. MLP 92-V-10-106d, maxilla with left DP2–DP4 and M1 in occlusal (A1) and lingual (A3) views; A2, explanatory drawing. B. MLP 92-V-10-106e, mandible with right dp4–m1 in occlusal (B1) and lingual (B5) views and mandible with left dp3–dp4 in occlusal (B3) view; B2, B4, explanatory drawings. C. MLP 92-X-10-119.38, maxilla with right P1–DP2, P3–M1 in occlusal view. D. MLP 92-X-10-119.49, mandible with left p4 (inside bone, broken), m1–3 in occlusal view.
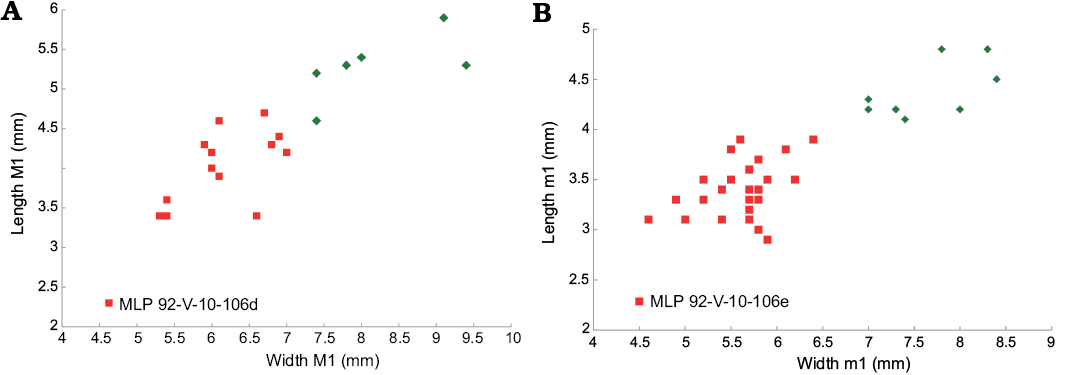
Fig. 5. Bivariate plots (length vs width) for the upper (A) and lower (B) first molar, comparing Protypotherium endiadys (square) and P. colloncurensis (diamond).
Appendicular skeleton: MAPBAR 1381 includes a distal fragment of left humerus, whose distal epiphysis (W, 19.5 mm; TD, 9.5 mm) is anteroposteriorly flattened and lateromedially enlarged (Fig. 6A). The ectepicondyle is incipient, whereas the entepicondyle is externally much expanded, having a well-developed supracondylar crest (rather broken) and a large entepicondylar foramen (Fig. 6A1). This configuration and the foramen are characteristics of Protypotherium australe (e.g., YPM-VPPU 015386, W, 18.2 mm; TD, 11.0 mm), but they are absent in Interatherium robustum (YPM-VPPU 015036). The medial crest of the trochlea is a prominent keel externally oriented and distally protruding (Fig. 6A3); in comparison, the capitulum is relatively small. The olecranon fossa is deep and there is a circular and small supratrochlear foramen, placed medially (Fig. 6A2). The coronoid fossa is not conspicuous. The deep trochlea, high medial crest, and supratrochlear foramen are features shared with Miocochilius anomopodus (UCMP 38091, W, 28.6 mm; T, 19.0 mm).
The left ulna MLP 73-VII-7-1 (Fig. 6B) is associated with upper and lower dentition. The olecranon (APD, 10.64 mm; TD, 6.13 mm; Fig. 6B2) is proximally rough, laterally flattened, and medially curved; in lateral view, it has a rectangular outline. The trochlear notch is obliquely oriented with respect to the proximodistal axis; laterally, it forms a high and continuous border. The maximum diameter of the trochlear notch is larger than the olecranon length. The anconeal process is mediolaterally wide and has a robust border anterodistally pointed. The medial coronoid process is wide, robust, and anteriorly well expanded, while the lateral coronoid process is less developed in comparison (Fig. 6B1). The radial notch is subrectangular and slightly convex. In general terms, the morphology of MLP 73-VII-7-1 agrees with the genus Protypotherium, although specific differences cannot be definitively established considering the fragmentary state of the bone; however, the ulna of P. endiadys has a less curved olecranon and a larger trochlear notch/olecranon ratio than P. australe (e.g., YPM-VPPU 015339, YPM-VPPU 015341).
The MLP 12-2916e right astragalus (L, 15.1 mm; Fig. 6C) is associated with lower dentition. As in most interatheres, the lateral and medial crests of the trochlea (H, 8.5 mm; W, 6.4 mm) are vertical and parallel to each other, aligned with the axis of neck and head (Fig. 6C1). The articular surface of the trochlea narrows posteriorly and is less concave than in Protypotherium australe (YPM-VPPU 015077) or Miochochilius anomopodus (UCMP 37913). The sustentacular facet is convex and proximolaterally-distomedially oriented, covering most of the posterior face (Fig. 6C2). The head is broken.
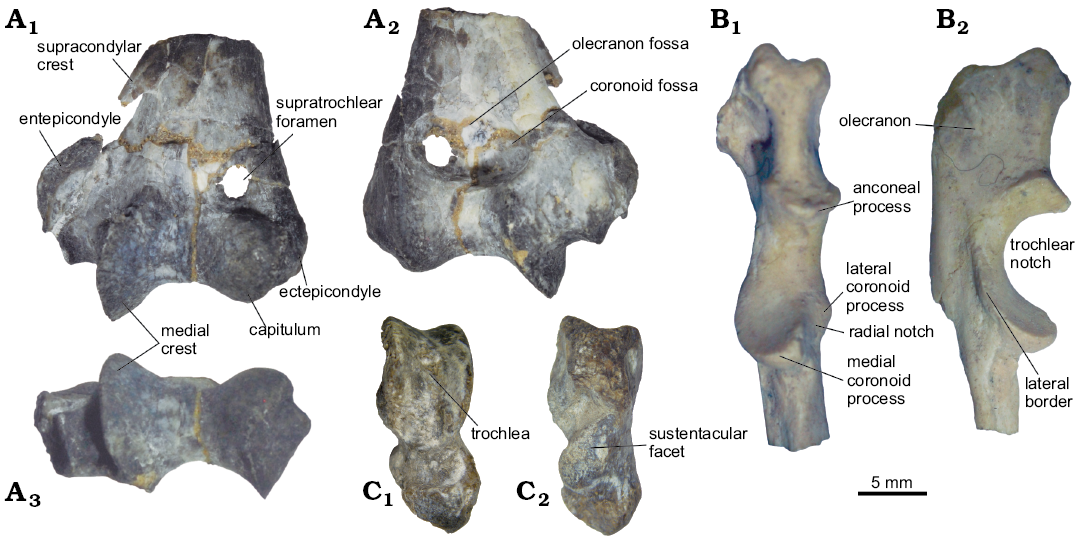
Fig. 6. Postcranial bones of interatheriine notoungulate Protypotherium endiadys (Roth, 1899) from the Collón Curá Formation, middle Miocene, Patagonia, Argentina. A. MAPBAR 1381, distal fragment of left humerus in anterior (A1), posterior (A2), and distal (A3) views. B. MLP 73-VII-7-1, proximal fragment of left ulna in anterior (B1) and lateral (B2) views. C. MLP 12-2916e, right astragalus in anterior (C1) and posterior (C2) views.
Remarks.—Icochilus endiadys is the only species of the Interatheriinae formally described by Roth (1899) from the Collón Curá Formation in Neuquén Province. According to Roth (1899), I. endiadys exhibits cranial similarities to both I. extensus Ameghino, 1889 and the genus Interatherium, although Roth (1899) recognized it differs from them in its dentition, which resembles, in turn, that of Protypotherium praerutilum Ameghino, 1887. In spite of that, Roth (1899) placed his species into the genus Icochilus Ameghino, 1889, which was later synonymized with Interatherium (Sinclair, 1909); both genera are under study at present (Fernández et al. 2015). Based on the restudy of the holotype of Icochilus endiadys (MLP 12-2886) plus the new material presented here and referred to the same taxon, we consider that the characteristics observed on these specimens are diagnostic of the genus Protypotherium instead of Interatherium (= Icochilus), agreeing with Roth’s (1899) inclination.
Stratigraphic and geographic range.—Collón Curá Formation, middle Miocene; Neuquén, Río Negro, and Chubut provinces, Argentina.
Protypotherium colloncurensis sp. nov.
Figs. 5, 7.
Etymology: In reference to the name of the geological unit, the Collón Curá Formation, where the new taxon comes from.
Holotype: MLP 91-IX-4-26, skull fragmented in several pieces: hemipalates with right and left M1–3, part of basicranium and posterodorsal fragment of vault (Fig. 7A).
Type locality: Chico River, Río Negro Province, Argentina (Fig. 1).
Type horizon: Collón Curá Formation, middle Miocene (Langhian, 14.86 ± 0.13 Ma; Bilmes et al. 2013).
Material.—Estancia El Criado: MLP 91-V-1-111, right mandible with m1–3; MLP 91-V-1-129, mandible with left and right p1–m3; MLP 92-V-10-105, right mandible with p3–m3; MLP 92-X-10-119: 3, right maxilla with P3–M2; 4, right mandible with m1–3; 5, right maxilla with M1–3; 13, right maxilla with P3–M2 and broken I3–P1; 15, right mandible with p3–4; 16, left mandible with p3–m2; 18, palate with left P1–M3 and right P2–M3, and left mandible with i1–m3; 31, right mandible with p2–m1; 35, left mandible with p4–m2; 36, left mandible with p2–m2; 46, left mandible with p4–m1; 50, right maxilla with M1–2. Cerro San Antonio: MLP 74-II-1-1 (originally assigned to Epipatriarchus sp.), left maxilla with P1–M2. Comallo: MLP 92-XI-18-31: a, right maxilla with M1–3; b, right maxilla with M3; c, right and left i1–3; d, left lower molar; e, M3 (a, c–e, probably same individual). Collón Curá Formation (Río Negro Province, Argentina), middle Miocene. Figure 1 details the localities referenced here.
Diagnosis.—Protypotherium colloncurensis is larger (38% in linear measurements) than Protypotherium endiadys and differs from it because of its higher and more robust mandible; wider upper canine; well-developed parastyle in P1; P2 overlaps P1; strongly imbricated upper molars, which have a well-developed and forwardly extended parastyle, a more strongly folded ectoloph, curved lingual sulcus, and without fossettes; molar size decreases markedly from M1 to M3; very procumbent lower incisors; p1 is subcircular and different from premolars, non-overlapping; very reduced p2; much smaller talonid in relation to trigonid in p3–4; and trigonid of molars narrows and shortens from m1 to m3.
Description.—Skull and upper dentition: The MLP 91-IX-4-26 holotype preserves the zygomatic arches, which are dorsoventrally low, and the anterior part of the zygomatic arch is laterally wide (Fig. 7A). In ventral view, the maxillary process of the zygomatic arch is a moderately concave triangular surface widening posteriorly. The descending process of the maxilla is barely developed (Fig. 7A2), as in P. endiadys.
Upper incisors are not known for P. colloncurensis. In comparison with P. endiadys, P. colloncurensis has a wider C (Table 3), and P1 has a well-developed parastyle and is overlapped by P2 (Fig. 7B). P2–4 have a strongly folded ectoloph, with a peculiar wider and labially more extended parastyle than the paracone fold (Fig. 7B).
Molars are larger than in P. endiadys, strongly imbricated and have a labially undulated ectoloph, with a well-developed and forwardly extended parastyle, and a deep labial sulcus (Figs. 5, 7A, C); no fossettes are observed. M1–2 have a posterior lobe wider than the anterior lobe which has a nearly straight distal face forming an angle of 90º with the ectoloph; in contrast, in M3 the posterior lobe is triangular-shaped with the distal face drawing an angle < 90°. M1 is the largest of the molar series, especially in the transverse diameter of the anterior lobe, which gives a peculiar trapezoid shape to the molar (Fig. 7C). Molar size decreases markedly from M1 to M3 (Table 3) and the lingual sulcus, which divides the anterior and posterior lobes, is curve-shaped and mesially pointed. These features are also observed in Protypotherium minutum (MLP 12-2177; Table 3); in turn, P. endiadys has nearly equally-sized upper molars with transversely straight lingual sulcus (Fig. 2A3, C). The most overlapped molars are present in P. distictum (MLP 12-2178; Table 3), but the latter differs from P. colloncurensis in having a bifid lingual sulcus.
Table 3. Upper tooth dimensions (in mm) of Protypotherium colloncurensis. Approximate values in parentheses; L, length; W, width.
| |
I3 |
C |
P1 |
P2 |
P3 |
P4 |
M1 |
M2 |
M3 |
|||||||||
|
L |
W |
L |
W |
L |
W |
L |
W |
L |
W |
L |
W |
L |
W |
L |
W |
L |
W |
|
|
MLP 91-IX-4-26 (holotype) |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
8.1 |
5.3 |
7.6 |
4.2 |
|
MLP 74-II-1-1 |
|
|
|
|
2.7 |
2.5 |
4.1 |
3.7 |
4.1 |
4.4 |
5.1 |
4.7 |
7.4 |
4.6 |
6.5 |
4.3 |
|
|
|
MLP 92-X-II-119.3 |
|
|
|
|
|
|
|
|
4.0 |
4.9 |
4.7 |
5.3 |
7.8 |
5.3 |
7.0 |
4.9 |
|
|
|
MLP 92-X-II-119.5 |
|
|
|
|
|
|
|
|
|
|
|
|
8.0 |
5.4 |
7.9 |
– |
|
|
|
MLP 92-X-II-119.13 |
4.7 |
2.2 |
– |
3.3 |
– |
3.9 |
4.4 |
3.8 |
4.4 |
4.6 |
4.8 |
4.7 |
7.4 |
5.2 |
6.8 |
4.5 |
6.5 |
3.8 |
|
MLP 92-X-II-119.50 |
|
|
|
|
|
|
|
|
|
|
|
|
9.4 |
5.3 |
7.3 |
4.7 |
|
|
|
MLP 92-XI-18-31a |
|
|
|
|
|
|
|
|
|
|
|
|
9.1 |
5.9 |
7.7 |
5.4 |
6.7 |
(4.0) |
|
MLP 92-XI-18-31b |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
7.6 |
4.2 |
|
Data sample |
1 |
1 |
0 |
1 |
1 |
2 |
2 |
2 |
3 |
3 |
3 |
3 |
6 |
6 |
7 |
6 |
4 |
4 |
|
Mean |
4.7 |
2.2 |
– |
3.3 |
2.7 |
3.2 |
4.3 |
3.8 |
4.2 |
4.6 |
4.9 |
4.9 |
8.2 |
5.3 |
7.3 |
4.9 |
7.1 |
4.1 |
|
Protypotherium minutum MLP 12-2177 |
|
|
|
|
3.1 |
3.2 |
3.8 |
3.6 |
5.2 |
3.8 |
5.0 |
3.3 |
5.1 |
2.8 |
|
|
|
|
|
Protypotherium distinctum MLP 12-2178 |
|
|
|
|
|
|
4.7 |
4.2 |
7.5 |
5.2 |
7.3 |
4.5 |
|
|
|
|
|
|
Mandible and lower teeth: Incisors are very procumbent. The i1–2 are sub-circular in cross-section, i1 being the smallest of incisors and laterally flattened (Fig. 7E; Table 4). The i3 and c are ellipsoidal in cross-section, labiolingually flattened (Fig. 7E); in contrast, p1 is shorter and wider than the canine, subcircular in cross-section and non-overlapping (Fig. 7D1, D2), differing from the laterally compressed p1 in P. endiadys. There are short spaces separating c from i3 and p1 (Fig. 7D4), a feature also observed in P. endiadys and Miocochilius anomopodus. Protypotherium colloncurensis has a higher and more robust horizontal ramus compared to P. endiadys (Figs. 5, 7D3).
The p2 differs from p1 in being larger and labiolingually flattened but, unlike p3–4, it does not have a differentiated talonid. In comparison, p3–4 are significantly larger than p2, both with well-defined trigonid and talonid (Fig. 7D1, D2; Table 4); p3 differs from p4 in having a longer trigonid. Compared to P. endiadys, premolars in the new species are clearly different in size and morphology from p2 to p4; in p3–4 the talonid is much smaller than the trigonid; and p2 is a very reduced tooth. In turn, in P. endiadys premolars are similar to one another, the talonid is larger (in relation to the trigonid) than in the other species, and p2 is not so reduced, being a small version of p3–4.
Table 4. Lower tooth dimensions (in mm) of Protypotherium colloncurensis. Approximate values in parentheses; L, length; W, width.
| |
i1 |
i2 |
i3 |
c |
p1 |
p2 |
p3 |
p4 |
m1 |
m2 |
m3 |
|||||||||||
|
L |
W |
L |
W |
L |
W |
L |
W |
L |
W |
L |
W |
L |
W |
L |
W |
L |
W |
L |
W |
L |
W |
|
|
MLP 91-V-1-111 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
7.8 |
4.8 |
7.8 |
4.4 |
11.1 |
4.0 |
|
MLP 91-V-1-129 |
|
|
|
|
|
|
5.1 |
3.5 |
3.6 |
3.9 |
– |
– |
6.2 |
3.7 |
6.7 |
4.2 |
8.4 |
4.5 |
8.3 |
4.3 |
11.5 |
3.6 |
|
MLP 92-V-10-105 |
|
|
|
|
|
|
|
|
|
|
|
|
4.9 |
2.9 |
– |
3.4 |
7.0 |
4.3 |
7.1 |
3.9 |
9.0 |
3.5 |
|
MLP 92-X-10-119.4 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
8.0 |
4.2 |
7.8 |
4.1 |
12.3 |
3.6 |
|
MLP 92-X-10-119.15 |
|
|
|
|
|
|
|
|
|
|
|
|
4.7 |
2.9 |
5.9 |
3.6 |
|
|
|
|
|
|
|
MLP 92-X-10-119.16 |
|
|
|
|
|
|
|
|
|
|
|
|
5.3 |
3.1 |
5.9 |
3.8 |
7.3 |
4.2 |
7.5 |
4.6 |
|
|
|
MLP 92-X-10-119.18 |
1.9 |
2.3 |
1.9 |
2.5 |
3.4 |
2.1 |
4.1 |
2.2 |
4.0 |
2.4 |
4.5 |
3.3 |
4.7 |
3.1 |
– |
4.3 |
7.4 |
4.1 |
7.2 |
4.0 |
– |
– |
|
MLP 92-X-10-119.31 |
|
|
|
|
|
|
|
|
|
|
4.0 |
2.6 |
5.8 |
3.4 |
6.6 |
4.3 |
8.3 |
4.8 |
|
|
|
|
|
MLP 92-X-10-119.35 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
– |
3.5 |
6.2 |
- |
5.6 |
– |
|
|
|
MLP 92-X-10-119.46 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
5.4 |
3.1 |
7.0 |
4.2 |
|
|
|
|
|
Data sample |
1 |
1 |
1 |
1 |
1 |
1 |
2 |
2 |
2 |
2 |
2 |
2 |
6 |
6 |
5 |
8 |
9 |
8 |
7 |
6 |
4 |
4 |
|
Mean |
1.9 |
2.3 |
1.9 |
2.5 |
3.4 |
2.1 |
4.6 |
2.9 |
3.8 |
3.2 |
4.3 |
3.0 |
5.3 |
3.2 |
6.1 |
3.8 |
7.5 |
4.4 |
7.3 |
4.2 |
11.0 |
3.7 |
The m1 and m2 are very similar to each other; they have a triangular talonid, which is slightly concave on the lingual face, becoming heart-shaped (Fig. 7D1, D2). The m1 has a longer mesial face than m2. The m3 differs from m1–2 in having a longer talonid, which is labially bilobed due to a vertical sulcus (Fig. 7D1, D2). All molars have a square trigonid, which becomes narrower and shorter from m1 to m3; this is a difference with respect to P. endiadys, in which the trigonid of successive molars remains nearly equally-sized. Premolars and molars are covered by a thick layer of cementum, mainly on labial side (Fig. 7D1, D2). No deciduous dentition was identified for P. colloncurensis.
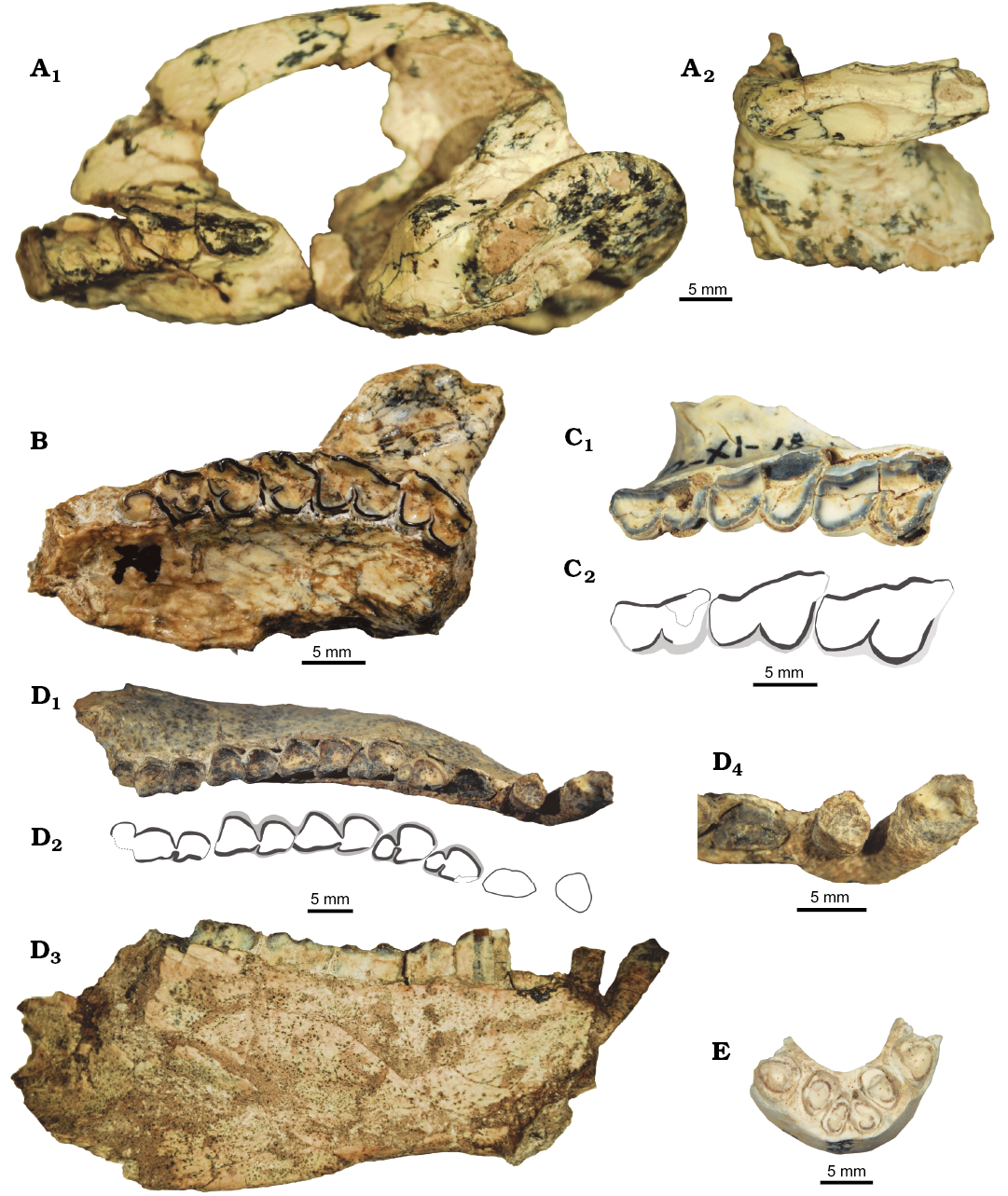
Fig. 7. Upper and lower dentition of interatheriine notoungulate Protypotherium colloncurensis, new species from the Collón Curá Formation, middle Miocene, Río Negro Province, Argentina. A. MLP 91-IX-4-26 (holotype), left hemipalates with M1–3, part of basicranium and posterodorsal fragment of vault in occlusal (A1) and lateral (A2) views. B. MLP 74-II-1-1, maxilla with left P1–M2 in occlusal view. C. MLP 92-XI-18-31a, maxilla with right M1–3 in occlusal view (C1); C2, explanatory drawing. D. MLP 91-V-1-129, mandible with left and right p1–m3 in occlusal (D1) and lingual (D3) views and detail of p1–3 (D4); D2, explanatory drawing. E. MLP 92-XI-18-31c, right and left i1–3 in occlusal view.
Multivariate statistics: The results of a discriminant analysis based on six variables of dentition showed a statistically significant separation among Miocene interatheres (Fig. 8). The analysis yielded two discriminant functions that accounted for 95.42% of variance in the data set. Excepting width of m3, which negatively influenced scores, all variables influenced scores positively. The first discriminant function (85.78%) mainly separated the morphospace of Protypotherium endiadys, which occupies intermediate values, from Protypotherium colloncurensis, which is placed to the right of the axis. Interatherium had the lowest (negatives) values for the first discriminant function, while Miocochilius and Protypotherium colloncurensis partially overlap at the right extreme (highest positive values); these latter taxa, however, had different values for the second discriminant function. Interestingly, Protypotherium australe had a wide range of values for the first discriminant function, partially overlapping P. endiadys in the negative values and P. colloncurensis in the positive values. This indicates variability between the Santacrucian specimens of Protypotherium.
Stratigraphic and geographic range.—Collón Curá Formation, middle Miocene; Río Negro Province, Argentina.
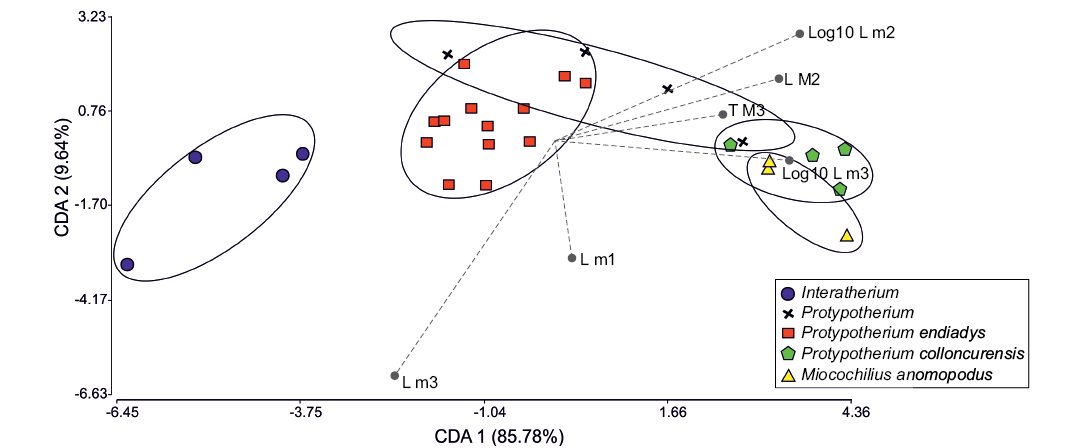
Fig. 8. Scatterplot of a discriminant analysis of Miocene interatheres based on six dental dimensions. Abbreviations: CDA, canonical discriminant analysis; L, length; M/m, upper/lower molar; T, transverse diameter.
Phylogenetic analysis
The analysis resulted in nine most parsimonious trees of 143 steps (CI = 0.47; RI = 0.65). The strict consensus tree is shown in Fig. 9. Node A is here considered as the Interatheriinae sensu stricto, a well-supported clade including the most recent common ancestor of Eopachyrucos pliciferus and Interatherium robustum, and all of its descendants, in agreement with previous works (Cifelli 1993; Hitz et al. 2000; Reguero et al. 2003; Reguero and Prevosti 2010). This monophyletic group (Fig. 9, node A) has high support values (Relative Bremer support 4; GC frequency 44; symmetric resampling 91) and five synapomorphies: high-crowned cheek teeth (character 1[1, 2]), mesial cingulum is absent in M1–3 (character 13[1]), rectangular M1–2, W < L (character 18[1]), sulcus dividing lophs on worn M1–2 (character 20[1, 2]), and the ectoflexid reaches the base of the crown of m1–2 (character 35[1]).
Within the Interatheriinae, three well-supported clades were recovered. The clade including Deseadan and post-Deseadan representatives (Fig. 9, node B), with Santiagorothia chiliensis (a pre-Deseadan species) as sister taxon, is supported by four synapomorphies: presence of cement (character 0[1]), absence of distal cingulum in P3–4 (character 14[1]), fossettes on molars disappear very early (character 43[1]), and upper molar size decreases from M1 to M3 (character 50[1], unambiguous).
The clade of Interatherium + Cochilius (Fig. 9, node C) is well supported by high support values (Relative Bremer support, 2; GC frequency, 67; symmetric resampling, 84) and five synapomorphies: P1 overlapped only by P2 (character 8[2], unambiguous), i3 < i2 (character 21[1]), canine absent or reduced with respect to incisors (character 24[0]), the mandible at m2 level is at least twice as high as at incisor level (character 39[1], unambiguous), and very prominent descending process of the maxilla (character 40[2]). This assemblage agrees with the results of Vera et al. (2017).
Finally, four synapomorphies were recovered for the group including the species of Protypotherium plus the genus Miocochilius (Fig. 9, node D): P4 W ≥ L (character 10[0]), mesial sulcus on P3–4 (character 19[1]), talonid < trigonid on p4 (character 32[0]), and nearly equally-sized upper molars (character 50[0], unambiguous); however, this node has low support values. It should be noted that Protypotherium endiadys and P. colloncurensis are included within this clade, justifying the assignment of both taxa to the same genus. Within this clade, Protypotherium australe is the sister taxon of a less inclusive group (Fig. 9, node E) supported by three unambiguous synapomorphies: P1 overlapped only by P2 (character 8[2]), i1 bifid (character 22[2]), which characterizes P. australe, and the supratrochlear foramen of humerus (character 46[0]) (lacking in Protypotherium australe and Interatherium robustum, both Santacrucian taxa).
Protypotherium endiadys is the sister group of the node including other middle Miocene taxa (Fig. 9, node F) and the early Miocene P. sinclairi. This clade (Fig. 9, node F) is a polytomy supported by one unambiguous synapomorphy, p1 not overlapped (character 26[0]), and includes the two species of the Laventan genus Miocochilius. Within this group, Protypotherium colloncurensis forms a clade with the Chasicoan Protypotherium minutum (Fig. 9, node G), which is supported by one unambiguous synapomorphy: upper molar size decreasing from M1 to M3 (character 50[1]). The unresolved relationships within node F are due to the poorly represented taxa, such as Protypotherium distinctum, P. sinclairi, and Miocochilius federicoi, for which only 15, 16, and 20 characters have been respectively scored (SOM 2).
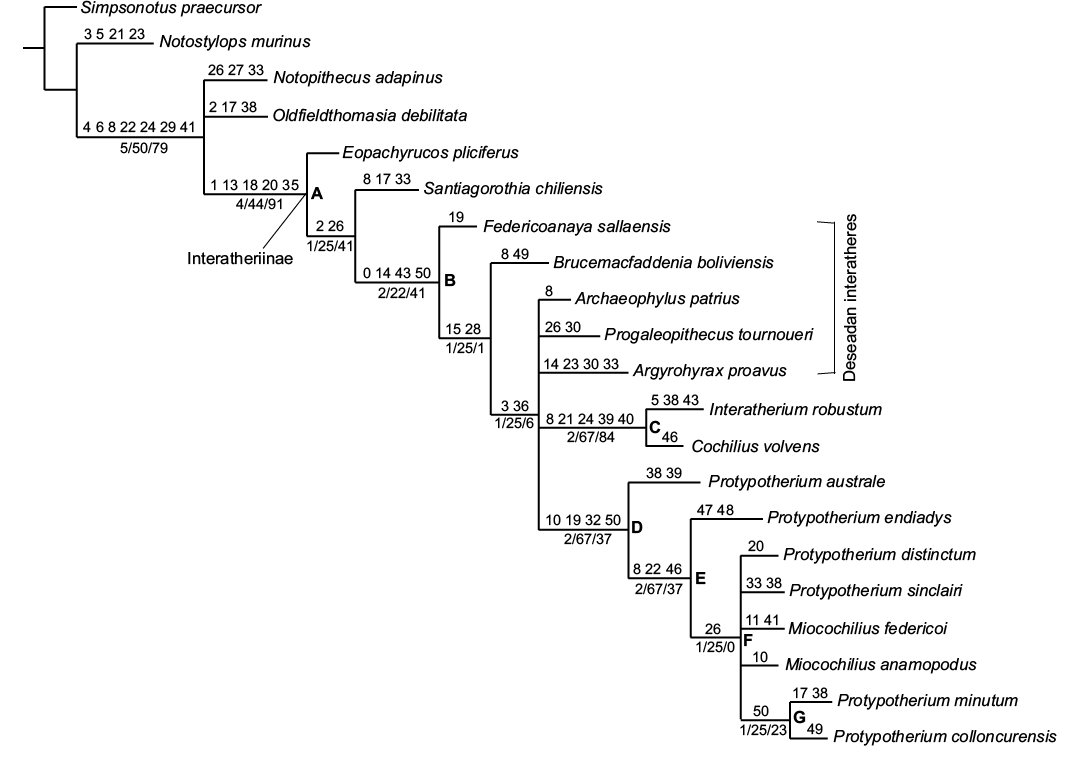
Fig. 9. Strict consensus of nine most parsimonious trees (L = 143; CI = 0.47; RI = 0.65), resulting from a cladistic analysis of a modified matrix taken from Vera et al. (2017). Letters in bold are the nodes explained in the text. Numbers above branches represent synapomorphies, while numbers below branches indicate support values: Bremer indices, GC frequency and symmetric resampling.
Discussion
Based on the revision of Roth’s (1899) collection curated at the MLP from Collón Curá River (Neuquén), besides the holotype (MLP 12-2886) of “Icochilus” endiadys, we identified six mandibular fragments, one tooth and an astragalus (MLP 12-2887 and MLP 12-2916), all of them assigned here to Protypotherium endiadys; however, we could not find the fragment of maxilla that Roth (1899: 40) mentioned for the same species. After Roth (1899), no new specimens were described for “Icochilus” endiadys from Neuquén province, but taxonomic modifications were introduced for this taxon. Following Sinclair’s (1909) synonymy between Interatherium and Icochilus, Pascual et al. (1978) changed the generic name of Icochilus endiadys to Interatherium endiadys, a proposal followed also by Bondesio et al. (1980). In turn, Kramarz et al. (2011) maintained Roth’s (1899) original generic name but alluded to a different specific epithet as Icochilus andiadys.
Concerning the Interatheriinae from Río Negro Province, Casamiquela (1963) mentioned Protypotherium australe at the Cantera Lif Mahuida (10 km west from Ingeniero Jacobacci) from the La Pava Formation or base of the Collón Curá Formation; unfortunately, we did not identify any specimen from this locality in the MLP collection. Bondesio et al. (1980) cited Protypotherium, Interatherium, and Epipatriarchus for Pilcaniyeu Viejo, references which were also listed by Pascual et al. (1984), but who erroneously cited Ganduglia (1977) and Coira (1979) instead of Bondesio et al. (1980) regarding the presence of Protypotherium from this locality. None of all these citations included descriptions or collection numbers. In the MLP collection, we identified the specimen assigned to Epipatriarchus sp. (MLP 74-II-1-1; Fig. 6B), which is reassigned here to Protypotherium colloncurensis, and the specimen assigned to Interatherium sp. (MLP 74-II-1-16), which is regarded here as P. endiadys.
Rolleri et al. (1984) mentioned Protypotherium praerutilum (MLP 79-XII-17-1) from Cerro Mesa de Trapalcó, but this specimen was not located at the MLP repository. Pardiñas and Reguero (1987) cited Protypotherium sp. for Cerro Castillo; this MLP 90-XII-24-34 specimen is here assigned to Protypotherium endiadys.
Finally, González Ruiz et al. (2012) reported an Interatheriinae indet. from the Collón Curá Formation at Chico River locality in Chubut Province; this specimen (LIEB PV 7002) was later identified as Protypotherium sp. (Vera et al. 2016) and here assigned to Protypotherium endiadys.
In sum, prior to this work and according to the latest updates, the fauna of the Interatheriinae from the Collón Curá Formation was composed of a single formally recognized species, Icochilus endiadys, plus references to Protypotherium australe, Protypotherium praerutilum, Protypotherium sp., Interatherium sp., Epipatriarchus, and Interatheriinae indet. However, after thorough revisions of new and old collections, two distinguishable morphotypes of interatheres are recognized for the Collón Curá Formation whose morphological characteristics are diagnostic for the genus Protypotherium: P/p3–P/p4 completely differentiated from molars, nearly square or wider than long P4, i3 larger than i2, lower canine similarly sized to i3, p3–4 with smaller talonid than trigonid, and little or moderately developed descending process of the maxilla.
Based on that, we reassigned “Icochilus” endiadys to Protypotherium endiadys, which was recognized in Colloncuran localities from Neuquén, Río Negro, and northern Chubut, extending its distribution to southern latitudes. The most peculiar features of Protypotherium endiadys are that the maxilla extends dorsally over the orbit and the anterior process of the frontal bone is lacking. Both conditions are present in Eocene notoungulates, such as Notopithecus and Oldfieldthomasia, but contrast with the Miocene interatheres Protypotherium australe, Interatherium, and Miocochilius, where the frontal bone projects anteriorly like a sliver, separating the maxilla from the orbit. This latter feature was considered diagnostic for the Interatheriinae (Vera 2016); however, it was not recovered as a synapomorphy in our phylogenetic analysis. Protypotherium endiadys is a smaller species than P. australe and P. distinctum; it differs from the Laventan species of the genus Miocochilius in having a reduced and subcircularly outlined P1 and deep and simple lingual sulcus on M1–2. With respect to Protypotherium australe, P. endiadys has a humerus with deeper trochlea and supratrochlear foramen.
Additionally, we recognized a new species of Protypotherium, P. colloncurensis, which was identified only from Río Negro Province, appearing together with P. endiadys at Estancia El Criado, Comallo, and Chico River localities. This species differs from P. endiadys because of its larger size; a well-developed parastyle on P1; strongly imbricated upper molars with a mesially well-extended parastyle, curved lingual sulcus, and lacking fossettes; a subcircular and non-overlapping p1; and a very reduced p2. It should be noted that some features of the dentition are shared with the younger Chasicoan interatheres, such as the strongly imbricated upper molars present in Protypotherium distictum (MLP 12-2178), while the marked decrease in the size of molars from M1 to M3 and their curved lingual sulcus are shared with P. minutum (MLP 12-2177).
The discriminant analysis confirms our differentiation between Protypotherium endiadys and P. colloncurensis based on tooth morphology, as well as the separation of other Miocene taxa considered here.
The phylogenetic analysis provides a good support for the new species Protypotherium colloncurensis and its distinction from P. endiadys. In the topology obtained, both species are included within the clade including the species of Protypotherium (Fig. 9, node D), justifying their generic assignment, which is supported by four synapomorphies. Within clade E (Fig. 9), Protypotherium endiadys branches earlier with respect to Protypotherium colloncurensis, which is grouped with the Chasicoan Protypotherium minutum (Fig. 9, node G).
It is interesting to note that node D (Fig. 9) groups both the species of Protypotherium and Miocochilius. On the one hand, this hypothesis agrees with the morphological similarities between the Laventan taxon Miocochilius, in particular the better known species M. anomopodus, and Protypotherium, but, at the same time, implies a paraphyletic condition for the genus Protypotherium. On the other hand, most of these Miocene interatheres, such as Protypotherium sinclairi, P. minutum, P. distinctum, and Miocochilius federicoi, are known only from fragmentary remains, which is predictably reflected in their unresolved relationships as a polytomy. In summary, on the basis of this comprehensive review of the middle Miocene species of Protypotherium, results suggest the need to focus on a more complete revision, including early and late Miocene species, and to carry out research addressing the validity of this taxon.
The reassignment of Icochilus endiadys to Protypotherium and the identification of a new species increase the diversity of the genus for the middle Miocene, filling the gap between the Santacrucian and Chasicoan species of Protypotherium (see Kramarz et al. 2015). Regarding other genera of the Interatheriinae in the Collón Curá Formation, as was previously mentioned, we reject the presence of Interatherium, which seems to be restricted to the early Miocene, while in the case of Epipatriarchus we provisionally do not recognize this taxon in the formation, pending a thorough revision of the material.
Concerning faunal affinities, the presence of Protypotherium in the Collón Curá Formation is confirmed, although expected, since it is a widely distributed genus throughout the Miocene in Argentina, Chile, and Bolivia (Ameghino 1885, 1887; Cione et al. 2000; Flynn et al. 2002; Croft et al. 2009). At species level, however, both taxa here described are exclusive to the Collón Curá Formation (middle Miocene); Protypotherium endiadys displays peculiar features that differ markedly from older (Santacrucian) and younger (Laventan) taxa, whereas P. colloncurensis shows some similarities with the Chasicoan species. Nevertheless, these affinities should be considered provisional because most of the middle Miocene interatheres, such as those from Frías, Huemules, Senguer, Guenguel, and Mayo rivers, remain poorly known.
Conclusions
After an exhaustive revision of the Interatherinae specimens from several collections, we identified only one genus, Protypotherium, in the Collón Curá Formation, ruling out previous references to Interatherium and Epipatriarchus in this unit.
The features of “Icochilus” endiadys are diagnostic of the genus Protypotherium rather than Icochilus or Interatherium. P/p3–P/p4 are fundamentally different from molars, the ectoloph undulates moderately on upper molars, p3–4 have talonid smaller than trigonid, and the descending maxillary process is little to moderately developed. Consequently, we propose regarding “Icochilus” endiadys as P. endiadys, a distinct species in the genus Protypotherium. Additionally, the diagnosis of P. endiadys is expanded and now includes characters from both deciduous dentition and postcranial remains (astragalus and forelimb bones), which are described for the first time for this species.
We identified a new species from the Collón Curá Formation, Protypotherium colloncurensis, which differs from P. endiadys mainly by its larger size, well-developed parastyle in P1, strongly imbricated upper molars, which have a well-developed and forwardly extended parastyle, very procumbent lower incisors, very reduced p2, and a much smaller talonid than trigonid in p3–4; in turn, P. colloncurensis shares with the Chasicoan Protypotherium species the more overlapped upper molars, which markedly decrease in size from M1 to M3.
The geographic distribution of P. endiadys extends from Neuquén to Chubut provinces, including Río Negro; P. colloncurensis, in turn, is only recognized in Río Negro Province, appearing together with P. endiadys in Estancia El Criado, Comallo, and Chico River localities.
Both the multivariate and phylogenetic analyses provided strong support for our systematic conclusions. The multivariate approach proved to be appropriate for extinct species with well-preserved dental series.
A thorough phylogenetic analysis of Protypotherium is provided for the first time. Based on our analysis, Protypotherium is a paraphyletic group including the species of Miocochilius. This agrees with morphological similarities between both taxa, although most of these Miocene interatheres are barely known.
This contribution expands on the understanding of the diversity of the Interatheriinae from the Collón Curá Formation and increases our knowledge of the record of Protypotherium for the Colloncuran age and the middle Miocene of high latitudes of South America.
Acknowledgements
We gratefully thank Ralph Hitz (Tacoma Community College, Tacoma, Washington, USA) and an anonymous reviewer for their constructive suggestions that improved the manuscript. Thanks are due to Ari Iglesias, Nicolás Ferreiro, and the staff of the MPB; Patricia Holroyd (UCMP), Alejandro Kramarz (MACN), and Daniel Brinkman (YPM-VPPU), who facilitated the access to collections under their care. Oscar Martínez (Universidad Nacional de la Patagonia San Juan Bosco, Sede Esquel, Chubut, Argentina) and Gabriel Martin (Centro de Investigación Esquel de Montaña y Estepa Patagónica CIEMEP, Esquel, Chubut, Argentina), and Teresa Dozo and Nelson Novo (both Centro Científico Tecnológico CONICET, Puerto Madryn, Chubut, Argentina) kindly lent the LIEBPV 7002 specimen. We specially thank Dirección Patrimonio y Museos de la Secretaría de Cultura (Ministerio de Turismo, Cultura y Deporte, Viedma, Río Negro, Argentina), which granted the loan of fossil specimens from MPB. Rafael Bottero (Instituto Argentino de Nivología, Glaciología y Ciencias Ambientales IANIGLA, Mendoza, Argentina) created the map in Fig. 1, Remedios Marín (IANIGLA) helped with the edition of the photographs and Nélida Horak (Mendoza, Argentina) revised the English grammar. Most of the specimens from MLP were collected in Río Negro and Neuquén provinces during the fieldtrips (1990–1993) organized between Museo de La Plata and the Duke University. This research was financially supported by CONICET and the project ANPCyT-PICT 2013-2270 (to LGR).
References
Ameghino, F. 1885. Nuevos restos de mamíferos fósiles oligocenos recogidos por el Profesor Pedro Scalabrini y pertenecientes al Museo Provincial de la ciudad de Paraná. Boletin Academia Nacional de Ciencias de Córdoba 8: 5–207.
Ameghino, F. 1887. Enumeración sistemática de las especies de mamíferos fósiles coleccionados por Carlos Ameghino en los terrenos eocenos de la Patagonia austral y depositados en el Museo La Plata. Boletín del Museo La Plata 1: 1–26.
Ameghino, F. 1889. Contribución al conocimiento de los mamíferos fósiles de la República Argentina. Actas de la Academia Nacional de Ciencias de Córdoba 6: 1–1027.
Ameghino, F. 1897. Les mammifères crétacés de l’Argentine. Deuxième contribution à la connaissance de la faune mammalogique des couches à Pyrotherium. Boletín del Instituto Geográfico Argentino 18: 1–117.
Ameghino, F. 1901. Notices préliminaires sur les ongulés nouveaux des terrains crétacés de Patagonie. Boletín de la Academia Nacional de Ciencias de Córdoba 16: 348–426.
Ameghino, F. 1903. Nuevas especies de mamíferos cretáceos y terciarios de la República Argentina. Anales de la Sociedad Científica Argentina 56: 193–208.
Bilmes, A., D’Elia, L., Franzese, J.R., Veiga, G.D., and Hernández, M. 2013. Miocene block uplift and basin formation in the Patagonian foreland: the Gastre Basin, Argentina. Tectonophysics 601: 98–111. Crossref
Bondesio, P., Rabassa, J., Pascual, R., Vucetich, M.G., and Scillato Yané, G.J. 1980. La Formación Collón-Curá de Pilcaniyeu Viejo y sus alrededores (Río Negro, Argentina). Su antigüedad y las condiciones ambientales según su distribución de litogénesis y sus vertebrados. II Congreso Argentino de Paleontología y Bioestratigrafía y I Congreso Latinoamericano de Paleontología, Actas 3: 85–99.
Cabrera, A. and Kraglievich, L. 1931. Diagnosis previas de los ungulados fósiles del Arroyo Chasicó. Notas Preliminares Museo de La Plata 1: 107–113.
Casamiquela, R.M. 1963. Sobre un par de anuros del Mioceno de Río Negro (Patagonia): Wawelia gerholdi n. gen. et sp. (Ceratophrydidae) y Gigantobatrachus parodii (Leptodactylidae). Ameghiniana 3(5): 141–160.
Cifelli, R.L. 1993. The phylogeny of the native South American ungulates. In: F.S. Szalay, M.J. Novacek, and M.C. McKenna (eds.), Mammal Phylogeny: Placentals, 195–216. Springer-Verlag, New York. Crossref
Cione, A.L., Azpelicueta, M.M., Bond, M., Carlini, A.A., Casciotta, J.R., Cozzuol, M.A., de la Fuente, M., Gasparini, Z., Goin, F.J., and Noriega, J. 2000. In: F.G. Aceñolaza and R. Herbst (eds.), Miocene Vertebrates from Entre Ríos Province, Eastern Argentina, El Neógeno de Argentina. Serie Correlación Geológica 14: 191–237.
Coira, B.L. 1979. Descripción Geológica de la Hoja 40d, Ingeniero Jacobacci, Provincia de Río Negro. SEGEMAR, Boletín 168: 1–94.
Croft, D.A. 2007. The Middle Miocene (Laventan) Quebrada Honda fauna, Southern Bolivia and a descripción of its notoungulates. Palaeontology 50: 277–303. Crossref
Croft, D.A., Anaya, F., Auerbach, D., Garzione, C., and MacFadden, B.J. 2009. New Data on Miocene Neotropical Provinciality from Cerdas, Bolivia. Journal of Mammalian Evolution 16: 175–198. Crossref
Croft, D.A., Carlini, A.A., Ciancio, M.R., Brandoni, D., Drew, N.E., Engelman, R.K., and Anaya, F. 2016. New mammal faunal data from Cerdas, Bolivia, a middle-latitude neotropical site that chronicles the end of the middle Miocene climatic optimum in South America. Journal of Vertebrate Paleontology: e1163574. Crossref
Cucchi, R., Espejo, P., and González, R. 1998. Hoja Geológica 4169-I, Piedra del Águila. Provincias del Neuquén y Río Negro. SEGEMAR, Boletín 424: 1–74.
Di Rienzo, J.A., Casanoves, F., Balzarini, M.G., Gonzalez, L., Tablada, M., and Robledo, C.W. 2016. InfoStat versión 2016. Grupo InfoStat, FCA, Universidad Nacional de Córdoba, Cordoba. http://www.infostat.com.ar
Escosteguy, L., Geuna, S., Franchi, M., González Díaz, E., and Dal Molín, C.N. 2013. Hoja Geológica 4172-II, San Martín de los Andes. Provincias de Río Negro y Neuquén. SEGEMAR, Boletín 409: 1–92.
Fernández, M., Fernicola, J.C., and Cerdeño, E. 2015. Revisión taxonómica de Interatherium Ameghino 1887 e Icochilus Ameghino 1889 (Interatheriidae, Typotheria, Notoungulata) del Santacrucense (Mioceno Temprano tardío) de la provincia de Santa Cruz, Argentina. Ameghiniana 53: 19R.
Flynn, J.J. and Swisher III, C.C. 1995. Cenozoic South American land mammal ages: correlation to global geochronologies. In: W.A. Berggren, D.V. Kent, M.P. Aubry, and J. Hardenbol (eds.), Geochronology, Time Scales, and Global Stratigraphic Correlation. Society for Sedimentary Geology Special Publication 54: 317–333. Crossref
Flynn, J.J., Novacek, M.J., Dodson, H.E., Franssinetti, D., McKenna, M.C., Norell, M.A., Sears, K.E., Swisher III, C.C., and Wyss, A.R. 2002. A new fossil mammal assemblage from the southern Chilean Andes: implications for geology, geochronology, and tectonics. Journal of South American Earth Sciences 15: 285–302. Crossref
Flynn, J.J., Wyss, A.R., Croft, D.A., and Charrier, R. 2003. The Tinguiririca Fauna, Chile: biochronology, paleoecology, biogeography, and a new earliest Oligocene South American Land Mammal “Age”. Palaeogeography, Palaeoclimatology, Palaeoecology 195: 229–259. Crossref
Ganduglia, P.R. 1977. Observaciones geológicas en la región de Ingeniero Jacobacci provincia de Río Negro (con énfasis en el Cretácico y Terciario). 86 pp. Unpublished B.Sc. Dissertation, Facultad de Ciencias Naturales, Universidad Nacional de Buenos Aires.
Goloboff, P.A., Farris, J.S., Källersjö, M., Oxelman, B., Ramírez, M.J., and Szumik, C.A. 2003. Improvements to resampling measures of group support. Cladistics 19: 324–332. Crossref
Goloboff, P.A., Farris, J., and Nixon, K. 2008a. TNT: Tree Analysis Using New Technology, version 1.1 (Willi Hennig Society Edition). Program and documentation. http://www.zmuc.dk/public/phylogeny/tnt
Goloboff, P.A., Farris, J., and Nixon, K. 2008b. A free program for phylogenetic analysis. Cladistics 24: 774–786. Crossref
González, P., Coluccia, A., Franchi, M., Caba, R., and Dalponte, M. 2000. Hoja Geológica 4169-III, Ingeniero Jacobacci, Provincia de Río Negro. Mapa y texto preliminar. SEGEMAR, Boletín 311: 1–97.
González Ruiz, L.R., Novo, N., Martínez, O., Tejedor, M.F., Dozo, M.T., Pérez, M.E., Sterli, J., and Bogan, S. 2012. Nuevas localidades de vertebrados fósiles del Mioceno medio del noroeste del Chubut, Argentina. Ameghiniana 49 (4): R29.
Hitz, R.B., Flynn. J.J., and Wyss, A.R. 2006. New Basal Interatheriidae (Typotheria, Notoungulata, Mammalia) from the Paleogene of central Chile. American Museum Novitates 3520: 1–32. Crossref
Hitz, R.B., Reguero, M.A., Wyss, A.R., and Flynn, J.J. 2000. New interatheriines (Interatheriidae, Notoungulata) from the Paleogene of central Chile and southern Argentina. Fieldiana, Geology 42: 1–26.
International Committe on Veterinary Gross Anatomical Nomenclature. 2005. Nomina Anatomica Veterinaria, 5th Edition. 166 pp. World Association of Veterinary Anatomists, New York.
Kramarz, A.G., Forasiepi, A.M., and Bond, M. 2011. Vertebrados Cenozoicos. In: H.A. Leanza, C. Arregui, O. Carbone, J.C. Danieli, and J.M. Vallés (eds.), Relatorio del XVIII Congreso Geológico Argentino, Abstracts, 557–572. Asociación Geológica Argentina, Buenos Aires.
Kramarz, A.G., Bond, M., and Arnal, M. 2015. Systematic description of three new mammals (Notoungulata and Rodentia) from the early Miocene Cerro Bandera Formation, northern Patagonia, Argentina. Ameghiniana 52: 585–597. Crossref
Madden, R.H., Guerrero, J., Kay, R.F., Flynn, J.J., Swisher, C.C. III, and Walton, A.H. 1997. The Laventan stage and age. In: R.F. Kay, R.H. Madden, R.L. Cifelli, and J.J. Flynn (eds.), Vertebrate Paleontology in the Neotropics. The Miocene Fauna of La Venta, 499–519. Smithsonian Institution Press, Washington.
Marshall, L.G. and Salinas, P. 1990. Stratigraphy of the Río Frías Formation (Miocene), along the alto Río Cisnes, Aisen, Chile. Revista Geológica de Chile 17: 57–87.
Marshall, L.G., Hoffstetter, R., and Pascual, R. 1983. Mammals and stratigraphy: geochronology of the continental mammal-bearing Tertiary of South America. Paleovertebrata, Mémoire Extraordinaire: 1–93.
Mazzoni, M.M. 1993. Sedimentación Colloncurense en las provincias de Río Negro y Neuquén. Ameghiniana 30: 351.
Mazzoni, M.M. and Stura, S. 1990. El Miembro Ignimbrítico Pilcaniyeu, Formación Collón-Curá (Mioceno), provincias de Río Negro y Neuquén. Reunión Argentina de Sedimentología (San Juan), Actas 3: 187–192.
Mones, A. 1986. Paleovertebrata Sudamericana. Catálogo sistemático de los vertebrados fósiles de América del Sur. Parte I: Lista preliminar y bibliografía. Courier Forschungsinstitut Senckemberg 82: 1–625.
Nullo, F.E. 1978. Descripción Geológica de la Hoja 41d, Lipetrén. Provincia de Río Negro. SEGEMAR, Boletín 158: 1–88.
Nullo, F.E. 1979. Descripción Geológica de la Hoja 39c, Paso Flores. Provincia de Río Negro. SEGEMAR, Boletín 167: 1–70.
Pardiñas, U.F.J. and Reguero, M. 1987. Vertebrados fósiles de Cerro Castillo (Departamento Pilcaniyeu). Nueva localidad para el Neógeno de Río Negro. In: M. Abril, G. Bochatey, V. Escribano, O. Giménez, R. Martínez, and J. Rodríguez (eds.), IV Jornadas Argentinas de Paleontología de Vertebrados, Abstracts, 28. Asociación Paleontológica Argentina, Buenos Aires.
Pascual, R., Vucetich, M.G., and Fernández, J. 1978. Los primeros mamíferos (Notoungulata, Henricosborniidae) de la Formación Mealla (Grupo Salta, Subgrupo Santa Bárbara): sus implicancias filogenéticas, taxonómicas y cronológicas. Ameghiniana 15: 366–390.
Pascual, R., Bondesio, P., Scillato-Yané, G.J., Vucetich, M.G., and Gasparini Z.B. 1978. Vertebrados. In: Geología y Recursos Naturales del Neuquén, VII Congreso Geológico Argentino, Relatorio, 177–185. Buenos Aires.
Pascual, R., Bondesio, P., Vucetich, M.G., Scillato-Yané, G.J., Bond, M., and Tonni, E.P. 1984. Vertebrados fósiles Cenozoicos. In: V. Ramos (ed.), IX Congreso Geológico Argentino (San Carlos de Bariloche), Relatorio II (9), 439–461. Asociación Geológica Argentina, Buenos Aires.
Rabassa, J. 1975. Geología de la region de Pilcaniyeu-Comallo, provincial de Río Negro, Argentina. Fundación Bariloche. Publicaciones del Departamento de Recursos Naturales y Energía 17: 1–128.
Rabassa, J. 1978. Estratigrafía de la región Pilcaniyeu-Comallo, provincia de Río Negro. VII Congreso Geológico Argentino (Neuquén), Actas 1: 731–746.
Ravazzoli, I.A. and Sesana, F.L. 1977. Descripción Geológica de la Hoja 41c, Río Chico. Provincia de Río Negro. SEGEMAR, Boletín 148: 1–77.
Rolleri, E.O, Dellapé, D.A., and Manceñido, M.O. 1984. Relaciones estratigráficas y correlación de las formaciones Pichi Picún Leufú y Collón Curá (Miembro Naupa Huen), aflorantes en el curso inferior y medio del cañadón Michihuau, provincia de Río Negro. IX Congreso Geológico Argentino (San Carlos de Bariloche), Actas 1: 458–477.
Reguero, M.A. and Prevosti, F.J. 2010. Rodent-like notoungulates (Typotheria) from Gran Barranca, Chubut Province, Argentina: phylogeny and systematics. In: R.H. Madden, A.A. Carlini, M.G. Vucetich, and R.F. Kay (eds.), The Paleontology of Gran Barranca, 148–165. Cambridge University Press, New York.
Reguero, M.A., Ubilla, M., and Perea, D. 2003. A new species of Eopachyrucos (Mammalia, Notoungulata, Interatheriidae) from the Late Oligocene of Uruguay. Journal of Vertebrate Paleontology 23: 445–457. Crossref
Roth, S. 1899. Apuntes sobre la geología y la paleontología de los territorios del Río Negro y Neuquén. Revista del Museo de La Plata 9: 1–56.
Roth, S. 1903. Los ungulados sudamericanos. Anales del Museo La Plata, Paleontología Argentina 5: 1–36.
Simpson, G.G. 1945. The principles of classification and a classification of mammals. Bulletin of the American Museum of Natural History 85: 1–350.
Sinclair, W.J. 1909. Typotheria of the Santa Cruz beds. In: W.B. Scott (ed.), Reports of the Princeton University Expedition to Patagonia 1896–1899, 6, 1–110. Schweizerbart’sche Verlagshandlung, Stuttgart.
Smith, J.B. and Dodson, P. 2003. A proposal for a standard terminology of anatomical notation and orientation in fossil vertebrate dentitions. Journal of Vertebrate Paleontology 23: 1–12. Crossref
Stirton, R.A. 1953. A new genus of Interatheres from the Miocene of Colombia. University of California Publications in Geological Sciences 29: 265–348.
Vera, B. 2016. Phylogenetic revision of the South American notopithecines (Mammalia, Notoungulata). Journal of Systematic Palaeontology 14: 461–480. Crossref
Vera, B. and Cerdeño, E. 2014. New insights on Antepithecus brachystephanus Ameghino, 1901 and dental eruption sequence in “notopithecines” (Mammalia, Notoungulata) from the Eocene of Patagonia, Argentina. Geobios 47: 165–181. Crossref
Vera, B., Cerdeño, E., and Reguero, M.A. 2017. The Interatheriinae from the Late Oligocene of Mendoza (Argentina), with comments on some Deseadan Interatheriidae. Historical Biology 29: 607–626. Crossref
Vera, B., González Ruiz, L., Martin, G., and Tejedor, M. 2016. Interatheriinae (Notoungulata) del Mioceno medio de la provincia del Chubut (Argentina). XI Congreso de la Asociación Paleontológica Argentina. Ameghiniana 54 (4) (Abstracts Supplement): 54.
Volkheimer, W. and Lage, J. 1981. Descripción geológica de la Hoja 42c, Cerro Mirador, provincia del Chubut. SEGEMAR, Boletín 181: 1–71.
Vucetich, M.G., Mazzoni, M.M., and Pardiñas, U.F.J. 1993. Los roedores de la Formación Collón Curá (Mioceno Medio), y la ignimbrita Pilcaniyeu, Cañadón Tordillo, Neuquén. Ameghiniana 30: 361–381.
Yrigoyen, M. 1969. Problemas estratigráficos del Terciario de Argentina. Ameghiniana 6: 349–356.
Acta Palaeontol. Pol. 62 (4): 845–863, 2017
https://doi.org/10.4202/app.00373.2017