
Lopingian tetrapod footprints from the Venetian Prealps, Italy: New discoveries in a largely incomplete panorama
LORENZO MARCHETTI, MATTEO BELVEDERE, and PAOLO MIETTO
Marchetti, L., Belvedere, M., and Mietto, P. 2017. Lopingian tetrapod footprints from the Venetian Prealps, Italy: New discoveries in a largely incomplete panorama. Acta Palaeontologica Polonica 62 (4): 801–817.
After new studies were carried out in the Lopingian Val Gardena Sandstone of northern Italy, in the Recoaro area (Venetian Prealps, NE Italy), the following tetrapod ichnotaxa are identified: cf. Capitosauroides isp., cf. Merifontichnus isp., Pachypes isp., Paradoxichnium isp., and Rynchosauroides isp., probably corresponding to ?parareptile, captorhinid eureptile, pareiasaurid parareptile, archosauromorph neodiapsid, and lacertoid neodiapsid trackmakers, respectively. An undetermined track shows features consistent with possible therapsid producers. These are the first possible records of Merifontichnus and Capitosauroides in the Lopingian (late Permian) and one of the few records of Paradoxichnium worldwide. The paleoecology of the ichnoassociation highlights a relatively high diversity in the floodplain lithofacies, a monospecific association of Rhynchosauroides in distal floodplain/sabkha environments and the occurrence of Paradoxichnium isp. and cf. Capitosauroides only in the lagoon lithofacies, suggesting different habits of the trackmakers. The tetrapod ichnoassociation is characterized by eureptile and parareptile tracks, and differs from the classic Lopingian tetrapod ichnoassociation of the Dolomites mainly because of the absence of chirotheriid and small parareptile ichnotaxa. A comparison of the Italian tetrapod ichnoassociation with other Lopingian non-eolian ichnofaunas suggests a possible preference for marginal marine settings by the archosauromorph and therapsid trackmakers at low-latitudes of Pangaea.
Key words: Tetrapoda, footprints, paleoecology, Lopingian, Val Gardena Sandstone, Venetian Prealps, Italy.
Lorenzo Marchetti [lorenzo.marchetti85@gmail.com], Urweltmuseum GEOSKOP, Burg Lichtenberg (Pfalz), Burgstrasse 19, D-66871 Thallichtenberg, Germany.
Matteo Belvedere [matteo.belvedere@jura.ch], Office de la culture, Section d’archéologie et paléontologie, Paléontologie A16, Hôtel des Halles, P.O. Box 64, 2900 Porrentruy 2, Switzerland.
Paolo Mietto [paolo.mietto@unipd.it], Dipartimento di Geoscienze, Università degli Studi di Padova, Via Gradenigo 6, 35131, Padova, Italy.
Received 27 May 2017, accepted 27 July 2017, available online 7 November 2017.
Copyright © 2017 L. Marchetti et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
The Lopingian (late Permian) was a key epoch for the evolution of terrestrial vertebrates, which reached a noteworthy abundance and diversity before the Permian–Triassic mass extinction (PTME). Therapsid synapsids and parareptiles were widespread, and archosauromorph eureptiles appeared for the first time. Neodiapsid eureptiles were common, while captorhinid eureptiles and amphibians were declining.
This is mainly inferred from the skeletal record (e.g., Lucas 2006), because few Lopingian footprints are known in the World. These specifically are located in: Italy, Germany, Scotland, Russia, Morocco, Niger, Brazil, South Africa, and Australia. At present, only the ichnogenera Batrachichnus, Dicynodontipus, Dromopus, Erpetopus, Hyloidichnus, Pachypes and Paradoxichnium, and several unidentified or dubious tracks with amniote affinity are known from all of the sites except Italy (Müller 1959; Schmidt 1959; McKeever and Haubold 1996; Warren 1997; De Klerk 2002; Surkov et al. 2007; Voigt et al. 2010; Hminna et al. 2012; Costa da Silva et al. 2012; Smith et al. 2015). The Italian ichnoassociation is by far more diverse and complete (Kittl 1891; Abel 1929; Leonardi 1951, 1974; Leonardi et al. 1975; Conti et al. 1975, 1977, 1980, 2000; Ceoloni et al. 1988; Wopfner 1999; Nicosia et al. 2001; Avanzini and Tomasoni 2004; Valentini et al. 2007, 2008, 2009; Avanzini et al. 2011; Bernardi et al. 2015; Citton et al. 2016), although a comprehensive revision based on the best-preserved material is still lacking since the 1980s. Specifically, the role of footprint taphonomy and extramorphologies, a relatively recent development in tetrapod ichnology (Haubold et al. 1995; Haubold 1996) was not properly taken into account. Thus, aside from the ichnogenera Pachypes, Rhynchosauroides, Dicynodontipus, and Protochirotherium, the ichnotaxonomy of the other morphotypes is unclear.
The revision with the addition of new material from a different site in the Venetian Prealps (Recoaro area, Veneto, NE Italy), preliminarily studied by Mietto (1975, 1981), gives the opportunity to reconstruct a relatively diverse ichnofauna comparable to that of the classic localities of the Dolomites. The aim of this study is a correct ichnotaxonomy based on the most recent methods and approaches, and the reconstruction of the faunistic and paleoecological meaning of these footprints in one of very few Lopingian sites worldwide, and majority of which in a need of revision.
Institutional abbreviations.—FG, Technische Universität Bergakademie Freiberg, Germany; MCV, Museo Civico D. Dal Lago of Valdagno, Italy; MGP, Museo Geologico delle Dolomiti, Predazzo, Italy; MGP-PD, Museo di Paleontologia e Geologia of the University of Padua, Italy. Specimens with provisional number include an additional “P”.
Geological setting
The Lopingian of the Southern Alps is characterized by the deposition of alluvial red beds in an area extending from Lombardy to Slovenia, interfingering to the east into marginal marine and shallow marine lithofacies, the result of the westward-directed Paleotethys transgression (Massari and Neri 1997). Generally, the alluvial red beds are laterally continuous with an average thickness of about 200–250 m and lie unconformably on Cisuralian volcano-siliciclastic successions and on the pre-Permian metamorphic basement. Two different formations were established in the Southern Alps domain: the Verrucano Lombardo Formation from Lombardy region to the Giudicarie area in Trentino-Alto Adige region and the Val Gardena Sandstone from Trentino-Alto Adige to Friuli-Venezia Giulia regions, separated by a structural high. The latter formation interfingers with the marginal marine lithofacies of the Bellerophon Formation in an overall transgressive trend, including some significant regressive events (Massari et al. 1988, 1994). The Val Gardena Sandstone includes deposits related to alluvial fan, braided and meandering fluvial environments; no clear deltaic sequences were detected. The occurrence of calcisols and rarer vertisols points to a semi-arid environment (Massari and Neri 1997). The lithofacies of the Bellerophon Formation interfingering with the alluvial red beds are generally dolostone and evaporites, the expression of sabkha marginal marine environments, with an invertebrate fauna typical of stressed settings (Neri 2007). The Bellerophon Formation passes upwards to the shallow marine and mostly Triassic Werfen Formation, without unconformities.
The fossil content of the Val Gardena Sandstone includes abundant tetrapod footprints, plant remains and palynomorphs (e.g., Conti et al. 1977, 2000; Ceoloni et al. 1988; Pittau 2005; Avanzini et al. 2011; Kustatscher et al. 2012, 2014, 2017), whereas the Bellerophon Formation includes abundant invertebrate fossils such as nautiloids, bivalves, brachiopods, and gastropods and microfossils such as algae, foraminifera, ostracods, and palynomorphs (e.g., Accordi 1956; Broglio Loriga 1960; Broglio Loriga et al. 1988; Pasini 1985; Posenato 1988, 1998; Posenato and Prinoth 1999; Spina et al. 2015). The biostatigraphic analysis of the palynoflora and invertebrate fossils and microfossils is consistent with a Lopingian age for the Val Gardena Sandstone and Bellerophon Formation (e.g., Massari et al. 1988; Farabegoli and Perri 1998), whereas the youngest underlying volcanic unit, Ora Formation, is radiometrically dated as latest Cisuralian (274.1 ±1.6 Ma; Marocchi et al. 2008) implying a large stratigraphic gap roughly corresponding to the entire Guadalupian (Cassinis et al. 2012). The overlying shallow marine Werfen Formation includes the Permian–Triassic boundary in its basal strata (Neri 2007).
The described fossil tetrapod footprints were found in the Venetian Prealps (Italy) in the localities of Merendaore (GPS coordinates N45°43.101’ E11°11.782’ alt 824 m), Ulbe (GPS coordinates N45°42.906’ E11°12.236’ alt 771 m) and Cortiana (GPS coordinates N45°76.443’ E11°23.943’ alt 560 m), close to the village of Recoaro (Fig. 1). In this area, the Val Gardena Sandstone shows a reduced thickness, of about 20–50 m and the successions lies directly and uncomformably on the metamorphic basement (Barbieri et al. 1980). A synthetic stratigraphic column of the study area is provided in Fig. 1D. The lower half of the formation is characterized by alluvial red beds arranged in upward fining- and thinning-cycles constituted of discontinuous conglomeratic lenses with an erosive base, passing upwards to coarse-grained m-thick sandstone, red to gray in color, with cross stratification and dm-thick fine-grained sandstone and cm-thick laminated mudstone (facies association a; Fig 2A, B). The upper half of the formation is characterized by well-stratified and laterally continuous dm-thick red sandstone passing upwards to cm-thick laminated mudstone red, yellow, or gray in color. The mudstone are characterized by pedogenic horizons (facies association b; Fig. 2C, D). The mudstone are interbedded to dark or light gray dolostone in strata 10–80 cm thick with irregular bedding and arranged in upward-thickening sequence, representing the basal lithofacies of the Bellerophon Formation (facies association c; Fig. 2E, F). The upper transition of the Val Gardena Sandstone with the Bellerophon Formation is transitional; the stratigraphic limit between these two formations corresponds conventionally to the last siliciclastic layer. The tetrapod footprints occur in the mudstone of the lower part of the Val Gardena Sandstone and in the mudstone, and dolostone of the upper part of the Val Gardena Sandstone (Fig. 1).

Fig. 1. Geographic location and stratigraphy of the fossil sites (asterisked). Map showing location of the study area in North Italy (A) and Recoaro area (B). Simplified geological map (C). D. Synthetic stratigraphic log of the Permian of Venetian Prealps (a–c, facies associations). Location of Merendaore and Ulbe (E) and Cortiana (F) fossil sites. G, H. Photographs of the Ulbe outcrops, small scale transition between lithofacies (G) and large scale transition between formations (H), with indicated transition between red bed (Rb) and lagoon (Lg) lithofacies of the topmost strata of the Val Gardena Formation, immediately before the deposition of the Bellerophon Formation. BEL, Bellerophon Formation; GAR, Val Gardena Sandstone; Lg, lagoon lithofacies (dolostone); Ps, incipient paleosol with deep mudcracks; Rb, red bed lithofacies (laminated mudstone). Hammer for scale.

Fig. 2. Sedimentary structures of Val Gardena Sandstone. Facies association a, fine-grained sandstone showing cross lamination (A) and parallel ripples (B). Facies association b, reddish mudstone with paleosols (C), pedogenic veins and nodules in the mudstone (D). Note the gray dolostone strata on the top. Facies association c, gray dolostone strata interbedded in the reddish mudstone (E), invertebrate burrows in the dolostone (F). G. Bellerophon Formation, gray dolostone.
Material and methods
The studied material includes specimens with tetrapod footprints preserved in convex hyporelief or concave epirelief, coming from the localities Merendaore, Ulbe and Cortiana (Venetian Prealps, Recoaro area; Fig. 1E, F) collected in the second part of the 20th century by the amateaur collectors Ottaviano Violati Tescari, Olivo Gonzo, Antonio Rossi, Franco Slaviero, and one of the authors (PM), who published two preliminary reports (Mietto 1975, 1981). The study comprises standard techniques for tetrapod ichnology such as photos in controlled light conditions, outline drawings on transparent films, digital drawings and measurements. 3D data were acquired with NextEngine 3D Scanner HD laser scanner through the NextEngine ScanStudio HD software and then processed with CloudCompare (v.2.7.0, http://www.cloudcompare.org) to obtain the false-color depth maps. All the models used for this study can be downloaded from https://doi.org/10.6084/m9.figshare.4040205. The taphonomy of footprints was strictly considered, in order to exclude from systematic study all the morphologic features related to deformations from the optimal and anatomically-related morphology of the footprint (extramorphologies sensu Haubold et al. 1995 and Bertling et al. 2006). Measurements mainly follow the conventions of Leonardi (1987). The systematic ichnology is based on morphologic features of the best-preserved footprints. The 3D methods provided additional information about morphology, resolving the problems linked to color and lighting of the specimens and allowing a precise interpretation of the footprint depth related to a reference surface. Due to the lack of trackways and the incompleteness of some specimens, we use open nomenclature, avoiding ichnospecific assignments.
Systematic palaeoichnology
Ichnogenus Capitosauroides Haubold, 1971
Type ichnospecies: Capitosauroides bernburgensis Haubold, 1971; Bernburg (Germany), Anisian.
cf. Capitosauroides isp.
Fig. 3.
Material.—MCV 5, left manus; MCV 6, partial left manus; MCV 7, left pes; MCV 11, right manus; MCV 11/05, right manus; MCV 12, right manus and right pes, possibly of the same step cycle; MGP-PD P3; right pes; all tracks are preserved in convex hyporelief; Ulbe and Cortiana (Italy) Lopingian.
Description.—Pentadactyl footprints showing medial functionality, relatively long, thick and straight digits with rounded terminations; and short concave palm/sole impressions. Ectaxonic pes, long about 90 mm. Smaller manus, longer than wide, showing relatively thicker digits, and less marked ectaxony. All footprints show very short digit I, digit V is proximally-positioned and usually not preserved.
Remarks.—The ectaxonic pentadactyl footprints with straight thick digits ending in rounded terminations, short concave palm/sole and a very short digit I are typical of Capitosauroides Haubold, 1971. The pentadactyl manual tracks and the short palm/sole exclude a possible attribution to typical temnospondyl footprints such as Limnopus Marsh, 1894. The lack of marked impressions of digit I basal pads and the short sole exclude the possible attribution to known reptiliomorph amphibian tracks such as Amphisauropus Haubold, 1970. Digit proportions, termination and arrangement are distinct from captorhinid tracks such as Hyloidichnus Gilmore, 1927 and Merifontichnus Gand, Garric, Demathieu, and Ellenberger, 2000. However, the lack of well-preserved digit V and of trackways hampers a definitive assignment, thus we tentatively classify these tracks as cf. Capitosauroides isp.
Capitosauroides was attributed to capitosaurid amphibians (Haubold 1971) or non-archosauromorph amniotes (Klein and Lucas 2010). MCV 7 was previously attributed to procolonomorph tracks indet. (Mietto 1995). We think that these tracks are likely of non-archosauromorph amniotes, and possibly of parareptiles. Capitosauroides shows a proximally-positioned digit V not separated from the digit I–IV group, typical of other possible parareptile tracks (Erpetopus, Pachypes, Procolophonichnium). This would be consistent with Mietto (1995), but more studies are needed to confirm the attribution of Capitosauroides to parareptiles.
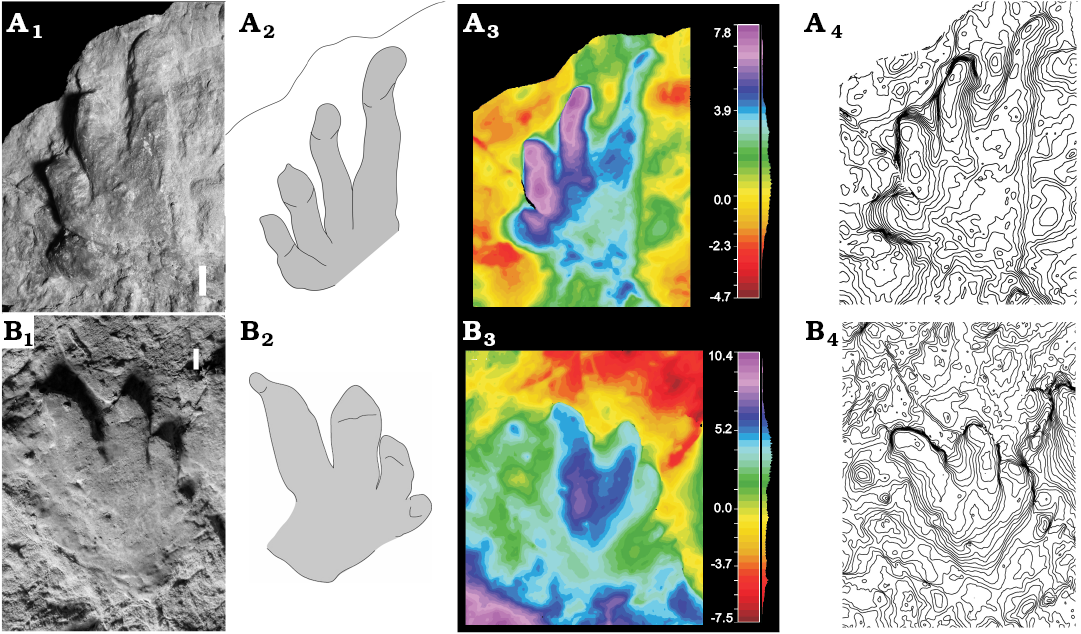
Fig. 3. Putative parareptile trace cf. Capitosauroides isp. from Ulbe (Italy), Lopingian. A. MCV 7, left pes impression showing digits I–IV. B. MCV 11/05, right manual impression showing digits I–IV. Convex hyporelief, spacing 0.5 mm. Photo (A1, B1), interpretive drawing (A2, B2), false-color depth map (A3, B3), contour lines (A4, B4). Scale bars 10 mm.
Ichnogenus Merifontichnus Gand, Garric, Demathieu, and Ellenberger, 2000
Type ichnospecies: Merifontichnus thalerius Gand, Garric, Demathieu, and Ellenberger, 2000; La Lieude (France), Guadalupian.
cf. Merifontichnus isp.
Fig. 4
Material.—MCV 2, left pes-manus couple and right pes of a different step cycle, in convex hyporelief; Ulbe (Italy), Lopingian.
Description.—Left pes-manus couple and right pedal impression preserved as convex hyporelief. The pes is long about 100 mm, pentadactyl, ectaxonic, with medial functionality, markedly wider than long. Digits are straight, relatively long, radially disposed, and show rounded terminations. Digit IV is the longest and similar in length to digit III, digit V is about as long as digit I. Digit I is parallel to the medial proximal margin of the sole. The sole is very short medially and longer in the external part; the sole proximal margin is straight to laterally convex. The manus is smaller, ectaxonic, markedly wider than long and probably pentadactyl. Digits are straight, relatively long, radially disposed, and show rounded terminations. Digit IV is the longest and similar in length to digit III, the palm is relatively short and oval in shape. The manus is inward-oriented compared to the pes.
Remarks.—The short sole/palm, the footprints wider than long, the long straight digits with enlarged terminations increasing in length from I to IV with subequal digits III–IV and relatively short pedal digit V, the disposition of pedal digit I parallel to the sole, the median lateral decrease in relief of the pes and the manus sensibly inward-oriented compared to the pes are typical of Merifontichnus Gand et al. 2000, after the preliminary revision of Marchetti (2016).
The only appreciable difference of this material with the Kungurian–Guadalupian material referable to this ichnogenus is its distinct heteropody (Newell et al. 1976; Gand et al. 2000, Marchetti et al. 2013, Marchetti 2016).
Waiting for a comprehensive ichnotaxonomic revision and because of the incomplete manual track, we tentatively classify these footprints as cf. Merifontichnus isp. A discussion of the differences with morphologically-similar ichnotaxa such as Hyloidichnus and Amphisauropus was recently exposed by Marchetti (2016). The studied material differs significantly from the ichnogenus Limnopus, too, mainly because of disposition/relative proportions of digits and sole length and shape. Moreover, a tetradactyl manus is unlikely, since the digit IV is the longest, whereas in Limnopus digit III is the longest.
Several features are consistent with derived captorhinid producers, such as the long rigid digits terminating in enlarged rounded terminations, the short palm/sole, the median-lateral decrease in relief of the pes, and the ectaxonic tracks with digits III–IV of similar length (Olson 1962; Sumida 1989; Holmes 2003). Additionally, the stout digits, the tracks wider than long and the convex proximal-lateral margin of the sole are consistent with the pedal structure of Moradisaurus. This last feature is probably due to the fusion of the calcaneum with the distal tarsal V, and the probable contact with the ground of part of the tarsus, due to a stronger vertical push on the appendicular skeleton (O’Keefe et al. 2005).
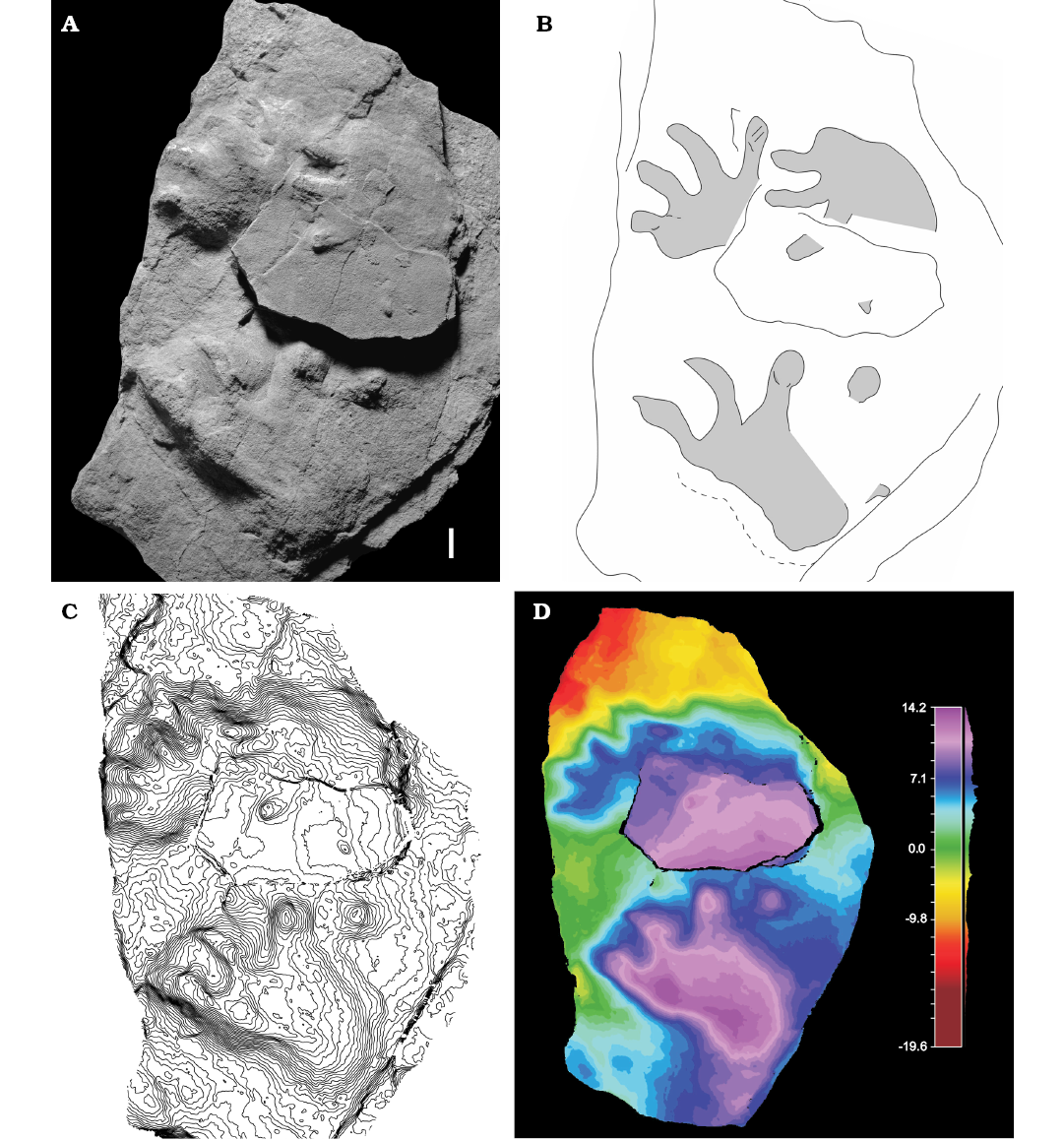
Fig. 4. Putative captorhinid trace cf. Merifontichnus isp. (MCV 2) from Ulbe (Italy), Lopingian. Left pes-manus couple and isolated right pes imprints, convex hyporelief. Photo (A), interpretive drawing (B), false-color depth map(C), contour lines (D). Scale bar 10 mm.
Ichnogenus Pachypes Leonardi, Conti, Leonardi, Mariotti, and Nicosia, 1975
Type ichnospecies: Pachypes dolomiticus Leonardi, Conti, Leonardi, Mariotti, and Nicosia, 1975; Bletterbach Gorge (Italy), Lopingian.
Pachypes isp.
Fig. 5.
Material.—MCV 3, left manual track, preserved in convex hyporelief; Ulbe (Italy), Lopingian.
Description.—Left manual imprint preserved as convex hyporelief long about 150 mm, and wide about 200 mm, with a broad palm showing a convex proximal margin, and short and sturdy digits with rounded terminations, continuously connected with the palm. The digit V is missing, digits III and IV are the longest, digits II–IV are bent inward. Footprint functionality is on digits I–III and on the medial part of the palm.
Remarks.—The broad rounded palm and the digit morphology, relative length and arrangement are characteristic of Pachypes Leonardi, Conti, Leonardi, Mariotti, and Nicosia, 1975, established from a single pedal imprint from the Val Gardena Sandstone of the Bletterbach locality of the Dolomites (Italy). Although trackways are missing, an abundant record from the type locality, including complete pes and manus, pes-manus couples and incomplete step cycles showing a complete range of preservation and size allows us to thoroughly confirm the validity of this ichnotaxon (Valentini et al. 2008, 2009). Here we confirm the determination of Mietto (1981) for the material of the Venetian Prealps. The material described from the Italian Carnian Alps (Dalla Vecchia 2013) and Russia (Surkov et al. 2007) does not show sufficient morphologic details to allow a determination, and the ichnotaxon Pachypes primus (Gubin et al. 2003) sensu Valentini et al. (2009) has to be considered a nomen dubium. Recently, Pachypes has been described from the Lopingian of Morocco (Voigt et al. 2010) and Niger (Smith et al. 2015). This type of track is confidently referred to pareiasaurian parareptile trackmakers (Leonardi et al. 1975; Valentini et al. 2008).
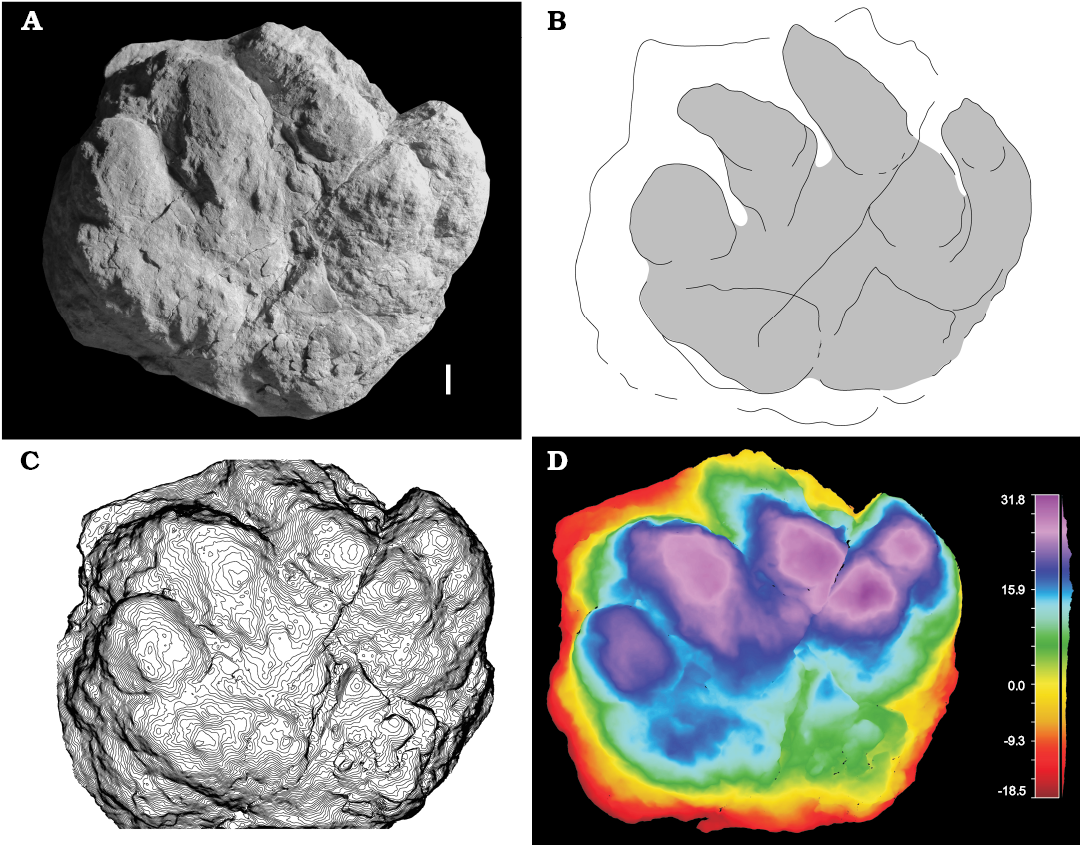
Fig. 5. Pareiasaurian parareptile trace Pachypes isp. (MCV 3) from Ulbe (Italy), Lopingian. Left manual imprint showing digits I–IV, convex hyporelief. Photo (A), interpretive drawing (B), false-color depth map (C), contour lines (D). Scale bar 10 mm.
Ichnogenus Paradoxichnium Müller, 1959
Type ichnospecies: Paradoxichnium problematicum Müller, 1959; Culmitzch (Thuringia, Germany), Lopingian.
Paradoxichnium isp.
Fig. 6.
Material.—MCV 8, possible right pes-manus couple; MCV 9, left manus; MCV 10, right manus; MCV 11/05, right manus; all tracks are preserved in convex hyporelief; Ulbe (Italy), Lopingian.
Description.—Pentadactyl manus, longer than wide and long about 70 mm, with thick and almost parallel digits increasing in length from I to IV. The digit V is relatively short, the bases of digits I and V are proximally positioned, both digits are shorter and thinner than digits II–IV. The digits II–IV terminate in large triangular claws, claws of digits I and V are smaller and acuminate. The palm is wider than long, and relatively short. The manual functionality is on digits II–IV, the palm and digits I and V can be absent. Pedal tracks larger with longer digits, lateral to the manus and usually not impressed.
Remarks.—The very characteristic morphology of the manual tracks allow a confident determination of these footprints as Paradoxichnium Müller, 1959, specifically the well-impressed and grouped digits II–IV terminating in large triangular claws and the smaller and proximally-positioned lateral digits (MCV 9, 10; Figs. 5, 6). Some specimens preserve only the digits II–IV of the manus, appearing as tridactyl impressions (MCV 8, 11, 11/05). This is clearly a taphonomic effect due to the higher functionality of digits II–IV. The rarity of pedal tracks is probably a taphonomic effect, the reason can be observed in the trackway holotype (pedal impressions are less-deeply impressed and more incomplete than manual impressions, FG 20/1; Fig. 6). The footprints classified as ?Paradoxichnium radeinensis Ceoloni, Conti, Mariotti, and Nicosia, 1988 from the same formation do not show the main features of the ichnogenus, thus they have to be assigned to a different ichnotaxon. Paradoxichnium is tentatively attributed to protorosaurid archosauromorphs, mainly because of their eureptilian affinity, the functional prevalence of digits II–IV and the trackway pattern that implies a low coupling value (Voigt 2012).
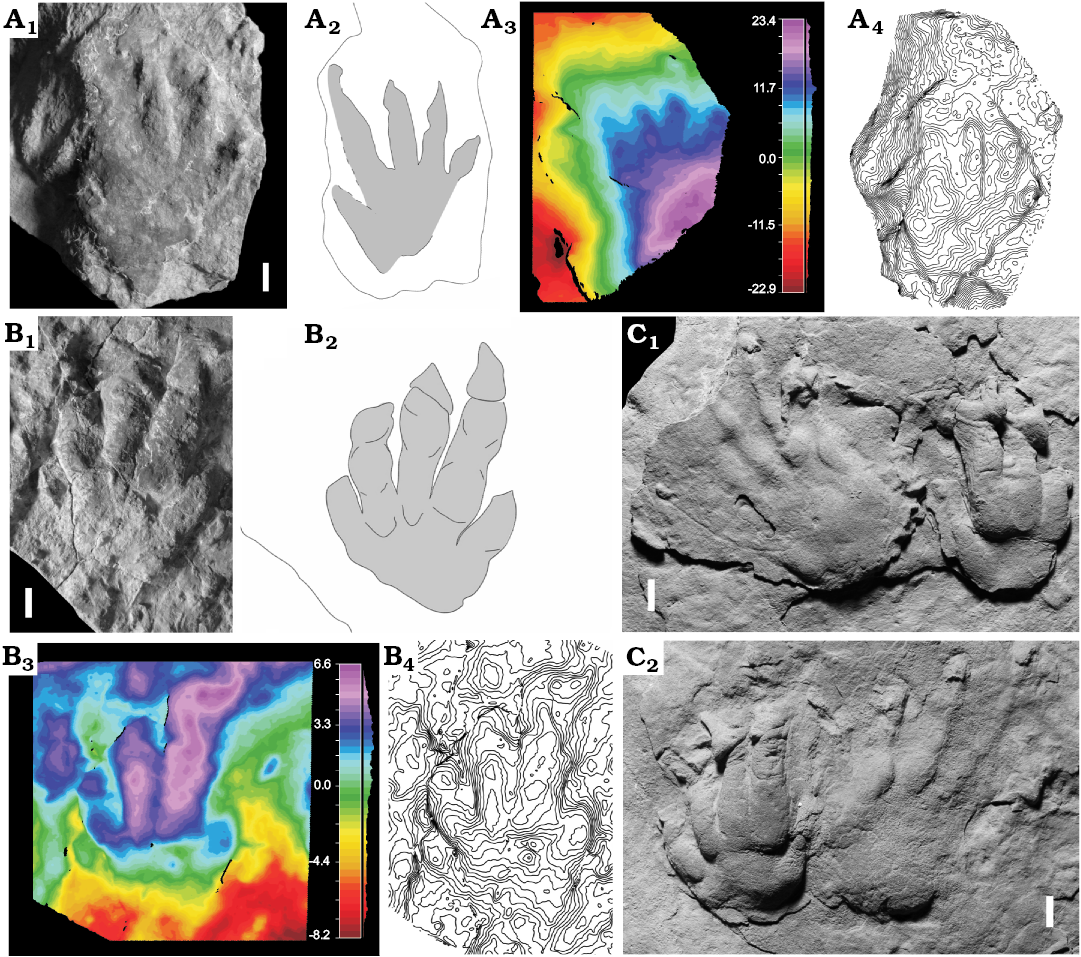
Fig. 6. Putative protorosaurid archosauromorph trace Paradoxichnium isp. A, B. Paradoxichnium isp. from Ulbe (Italy), Lopingian. A. MCV 10, right complete manus, note the proximally-positioned digits I and V and the triangular claw impressions. B. MCV 9, complete left manus impression. Note the proximally-positioned digits I and V, the parallel digits II–IV and the triangular claw impressions. C. Paradoxichnium problematicum Müller, 1959, holotype FG 20/1 from Culmitzch (Thuringia, Germany), Lopingian; right (C1) and left (C2) pes-manus couples; note the manual morphology similar to MCV 9. Convex hyporelief, spacing 0.5 mm. Photo (A1, B1), interpretive drawing (A2, B2), false-color depth map (A3, B3), contour lines (A4, B4). Scale bars 10 mm.
Ichnogenus Rhynchosauroides Maidwell, 1911
Type ichnospecies: Rhynchosauroides rectipes Maidwell, 1911; Daresbury, Cheshire (UK), Early–Middle Triassic.
Rhynchosauroides isp.
Fig. 7.
Material.—MCV 4, right track; MCV 65, right manus, right track and possible straight tail impression; MGP-PD 25510; left manus and right track of the same step cycle; left pes and other two tracks of other step cycles, possible tail/body impression; all tracks preserved in convex hyporelief; Merendaore (Italy), Lopingian.
Description.—Semiplantigrade, pentadactyl manus with long digits markedly and continuously increasing in length from I to IV, distally bent medially and terminating in sharp, thin, and triangular claws in continuity with the digits. Digit V is thin, about as long as digit II, straight, outward-oriented and its base is separated and more proximal compared to the digit IV base. The palm is short and triangular in shape. Functional prevalence is medial. Incomplete pedal tracks and possible straight tail impressions associated with footprints were observed. All the tracks are preserved in convex hyporelief.
Remarks.—The ectaxonic manus showing long digits with sharp claws, the continuous increase in length between digits I–IV, the short palm, the digit V separated from the digit IV base and outward directed are typical of Rynchosauroides Maidwell, 1911. The lack of associated pedal impressions and the unclear ichnotaxonomic status of several Rynchosauroides species is the reason why we classified these tracks as Rhynchosauroides isp. This typical Lopingian–Mesozoic ichnogenus needs a comprehensive revision, because several ichnospecies were introduced but a few might have a real ichnotaxonomic meaning. The studied material was initially classified as Eumekichnium gampsodactylum (Mietto, 1975); the genus Eumekichnium is presently considered a junior synonym of Dromopus lacertoides (Voigt, 2005). However, the manual track of Dromopus differ significantly from that of Rhynchosauroides because it shows a digit IV much longer than digit III, it is semidigitigrade and is identical in morphology with the pes. Rhynchosauroides is the most abundant tracktype of the Val Gardena Sandstone; and in the type locality is usually referred to Rhynchosauroides pallinii Conti, Leonardi, Mariotti, and Nicosia, 1977. A morphologically-similar ichnogenus is referred to Ganasauripus ladinus Valentini, Conti, and Mariotti, 2007. The trackmakers were likely lacertoid neodiapsid eureptiles (Valentini et al. 2007).
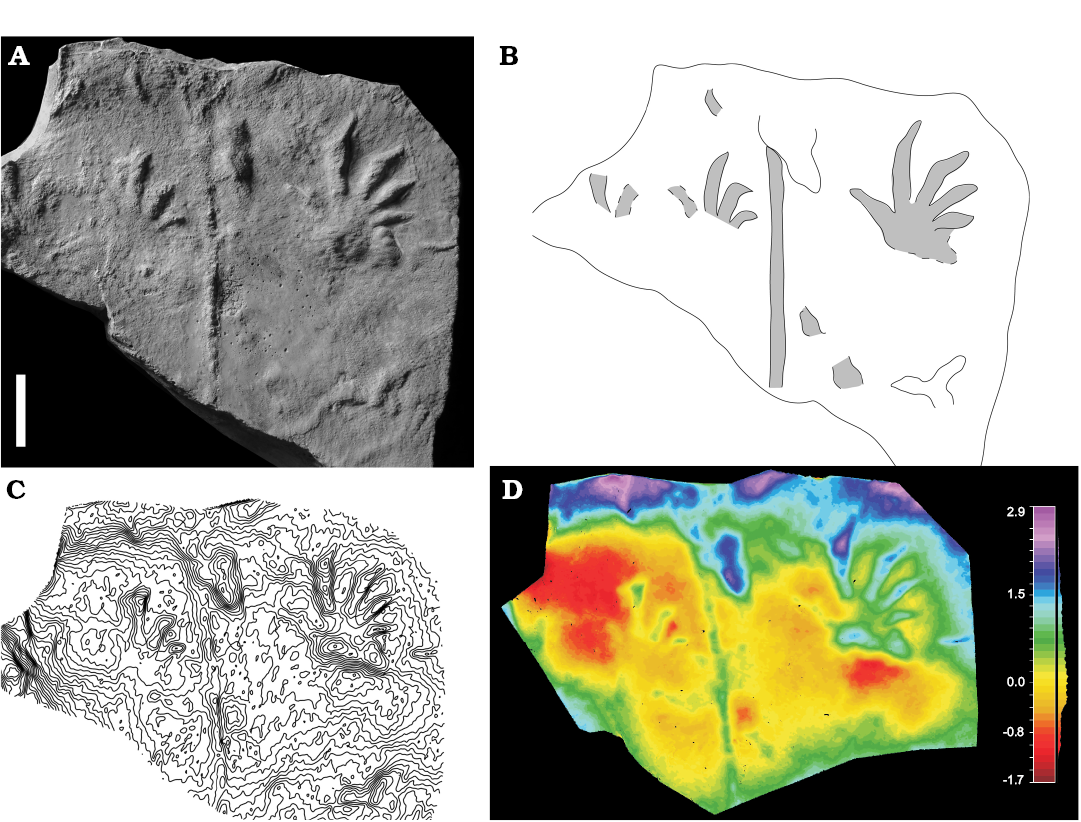
Fig. 7. Lacertoid neodiapsid eureptile trace Rhynchosauroides isp. (MCV 65) from Merendaore (Italy), Lopingian. Complete right manual impression. An incomplete track and a continuous tail are preserved on the same slab. Convex hyporelief (plaster cast), spacing 0.5 mm. Photo (A), interpretive drawing (B), contour lines (C), false-color depth map (D). Scale bar 10 mm.
Undetermined track
Fig. 8.
Material.—MCV 14/30, right track, concave epirelief; Cortiana (Italy), Lopingian.
Description.—Right footprint showing digits III–V and a partial impression of the palm/sole, with lateral functionality and expulsion rims. Foot length is about 80 mm. Digit IV is the longest, digit V is slightly shorter than digit III. Digits are relatively short and thick and terminate in rounded pads, and are more deeply impressed in their proximal and distal parts. No claws were observed. The footprint is preserved as concave epirelief.
Remarks.—This specimen, in morphology and digit arrangement, resembles the external part of a relatively large track with rounded and large digit terminations. The deeper impression of metapodial-phalangeal pads and digit tips compared to the medial part of digits is consistent with a specific skeletal feature of synapsids, the “digital arcade” sensu Kümmel and Frey (2012). Thus, these footprints are probably assignable to therapsid synapsids. In the Italian Lopingian track record, two morphotypes only show these characteristics: “Ichniotherium cottae” sensu Conti et al. (1977) and “Ichniotherium” accordi Ceoloni, Conti, Mariotti, and Nicosia, 1988. In particular, it resembles the manual track figured by Conti et al. (1977: pl. 8: 3, MGP 9/22) and attributed to “Ichniotherium cottae” (Fig. 8B). Waiting for a revision of morphologically-similar material from the Dolomites and because of the incompleteness of MCV 14/30, we do not assign it to any ichnotaxon, although it was probably produced by therapsid synapsids.
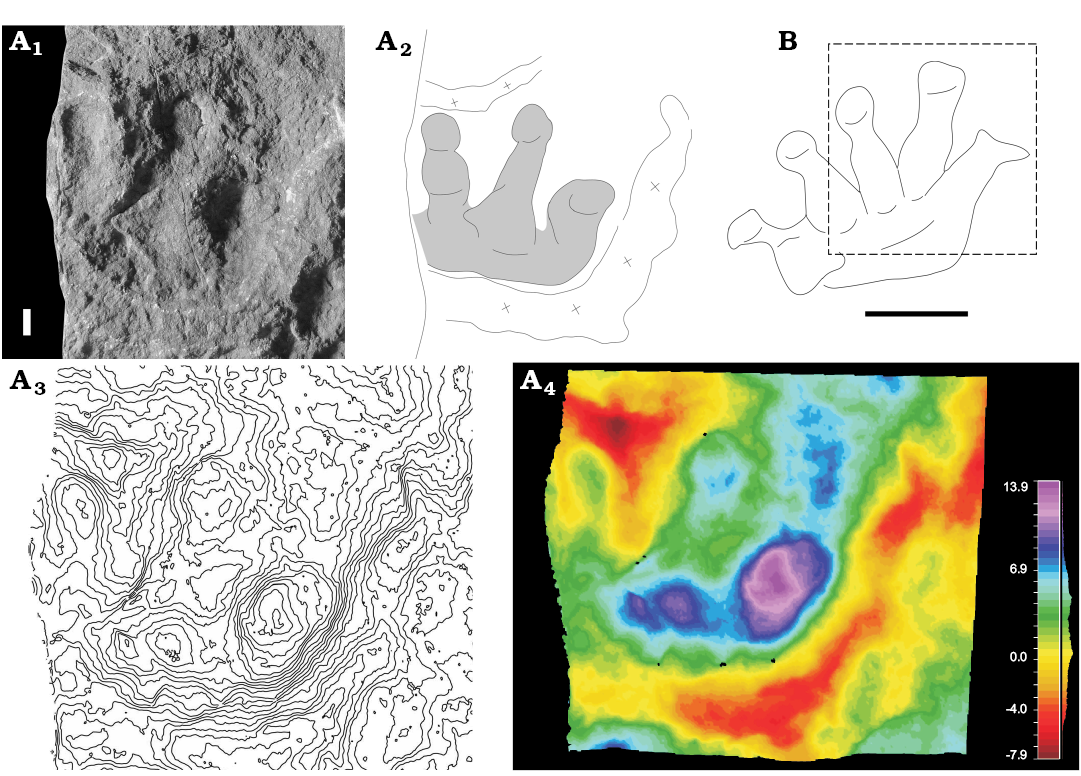
Fig. 8. A. Undetermined track (MCV 14/30) of therapsid synapsid from Cortiana (Italy), Lopingian. Incomplete right manual impression showing digits III–V and deep expulsion rims. Concave epirelief, spacing 1 mm. Photo (A1), interpretive drawing (A2), contour lines (A3), false-color depth map (A4). B. MGP 9/22, interpretive drawing of a complete left manual impression from the Bletterbach Gorge, Dolomites (Italy), Lopingian, after Conti et al. (1977). Scale bar 10 mm.
Discussion
Paleoecology.—The tracks found in the lower half of Val Gardena Sandstone in facies association a (Figs. 1, 9, Table 1) of Ulbe, Merendaore, and Cortiana localities represent the more diverse ichnoassociation recording Pachypes isp., cf. Merifontichnus isp., Rynchosauroides isp., and the large undetermined track. This interval is interpreted as a typical sequence of alluvial red beds, the occurrence of abandoned channels and lateral accretion structures suggests a meandering fluvial environment. Tetrapod footprints are preserved in concave epirelief or convex hyporelief in the laminated mudstone or in convex hyporelief in the fine-grained sandstone covering mudstone intervals. The trampling surfaces are generally interpreted as overbank or abandoned channel deposits.
The ichnoassociation found in the upper half of the Val Gardena Sandstone in facies association b of Merendaore locality (Figs. 1, 9, Table 1) includes Rynchosauroides isp. and small invertebrate burrows. These layers show varicolored and laterally-extensive laminated mudstone with pedogenic horizons and are interpreted as distal floodplain to sabkha environments.
The ichnoassociation found in the upper half of the Val Gardena Sandstone in facies association c of Ulbe and Cortiana localities (Figs. 1, 9, Table 1) includes Paradoxichnium, cf. Capitosauroides, and relatively large invertebrate burrows. The trace fossils are preserved in convex hyporelief in the dolostone strata interbedded to the mudstone and interpreted as transgressive lagoon environments passing upwards to the lagoon and marginal marine lithofacies of Bellerophon Formation.
Table 1. Specimen list from the Recoaro area, with determination, locality and facies association.
|
Specimen number |
Field number |
Locality |
Facies association |
Fossil content |
|
MCV 2 |
OG/UB 19 |
Ulbe |
a |
cf. Merifontichnus isp. |
|
MCV 3 |
AR/UB 8 |
Ulbe |
a |
Pachypes isp. |
|
MCV 4 |
OG/MR 13 |
Merendaore |
a |
Rhynchosauroides isp. |
|
MCV 66 |
MP/MR 17 |
Merendaore |
a |
coprolites |
|
MGP-PD P1 |
MP/MR 18 |
Merendaore |
a |
cf. Rhynchosauroides isp. |
|
MCV P1 |
|
Cortiana |
a |
therapsid track indet. |
|
MCV 11/02 |
OG/MR 14 |
Merendaore |
b |
lacertoid track indet. |
|
MCV 11/03 |
OG/MR 15 |
Merendaore |
b |
lacertoid track indet., burrows |
|
MCV 65 |
OV/MR 9 |
Merendaore |
b |
Rynchosauroides isp. |
|
MCV 67 |
OG/MR 20 |
Merendaore |
b |
small track indet., burrows |
|
MGP-PD 25510 |
MP/MR 10 |
Merendaore |
b |
Rynchosauroides isp. |
|
MGP-PD P2 |
MP/MR 16 |
Merendaore |
b |
tracks indet. |
|
MCV 5 |
OG/UB 3 |
Ulbe |
c |
cf. Capitosauroides isp. |
|
MCV 6 |
OG/UB 2 |
Ulbe |
c |
cf. Capitosauroides isp. |
|
MCV 7 |
OG/UB 11 |
Ulbe |
c |
cf. Capitosauroides isp. |
|
MCV 8 |
OG/UB 7 |
Ulbe |
c |
cf. Paradoxichnium isp., burrows |
|
MCV 9 |
AR/UB 12 |
Ulbe |
c |
Paradoxichnium isp. |
|
MCV 10 |
OG/UB 5 |
Ulbe |
c |
Paradoxichnium isp. |
|
MCV 11 |
OG/UB 1 |
Ulbe |
c |
cf. Capitosauroides isp., burrows |
|
MCV 11/04 |
OG/UB 21 |
Ulbe |
c |
large track indet. |
|
MCV 11/05 |
OG/UB 39 |
Ulbe |
c |
Paradoxichnium isp., cf. Capitosauroides isp. |
|
MCV 12 |
G2/CO 90 |
Cortiana |
c |
cf. Capitosauroides isp. |
|
MGP-PD P3 |
OG/UB 4 |
Ulbe |
c |
cf. Capitosauroides isp. |
|
MGP-PD P4 |
OG/UB 6 |
Ulbe |
c |
track indet. |
The overall stratigraphic sequence of the Val Gardena Sandstone in the Recoaro area is characterized by an upwards retrogradation of the alluvial environments and a contemporary marine transgression, passing from meandering fluvial to distal floodplain/sabkha to lagoon environments, prelude to the lagoon and shallow marine environments of the Bellerophon Formation. The most diverse ichnoassociation is recorded in the meandering fluvial environment (Fig. 9), including tracks of large herbivores (pareiasaurid, captorhinids, ?therapsids) and small ominivore forms (neodiapsids). This might suggest the presence of a complex trophic structure, the result of stable water source and food availability. The ichnoassociation of the terminal fan/sabkha environment (Fig. 9) includes the only Rhynchosauroides. This is consistent with a stressed environment (e.g., Neri 2007) with reduced water availability and arid conditions, more adapted to small-sized omnivore and opportunistic animals. The ichnoassociation of the lagoon environments (Fig. 9) is instead characterized by Paradoxichnum isp., probable footprints of medium-sized archosauromorph, and cf. Capitosauroides, footprint of medium-sized non-archosauromorph amniotes (?parareptiles), that are absent in the alluvial lithofacies. This might be interpreted as a paleoecological signal, because basal lagoon lithofacies at the Val Gardena Sandstone–Bellerophon Formation transition represent a brackish environment with a specific paleobiota, thus the trackmakers were probably adapted to these conditions, differently to the other tetrapod forms living in the alluvial to sabkha environments. This facies-related distribution of the ichnoassociation confirms once again the impact of the paleoenvironment on the tetrapod ichnoassociations, in terms of different occurrences and different relative abundances. More studies on Lopingian ichnoassociations are needed to eventually confirm the facies-related distribution of the tetrapod ichnotaxa found in the Recoaro area and re-define the Lopingian tetrapod ichnocoenoses (sensu Hunt and Lucas 2006, 2007) and ultimately the meaning of the tetrapod ichnofacies.
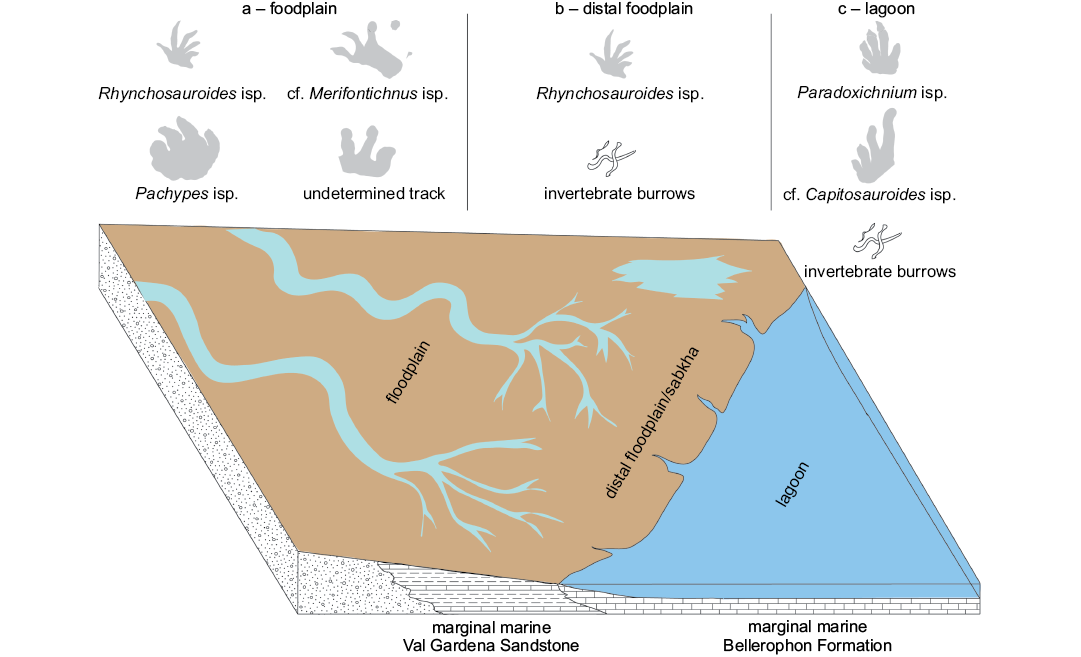
Fig. 9. Trace fossils and an interpretation of the depositional environment of the footprint localities of Recoaro (facies associations a–c).
Faunistic significance.—Through the ichnotaxonomic study of the Lopingian Val Gardena Sandstone of the Recoaro area, the following tetrapod ichnogenera are identified: cf. Capitosauroides, cf. Merifontichnus, Pachypes, Paradoxichnium, and Rynchosauroides; corresponding to ?parareptile, captorhinid eureptile, pareiasaurid parareptile, archosauromorph eureptile and lacertoid neodiapsid eureptile trackmakers, respectively. An isolated track is probably attributable to therapsid synapsids, and shares similarities with “Ichniotherium cottae” sensu Conti et al. (1977). This is the first possible record of Merifontichnus in the Lopingian, the first possible occurrence of Capitosauroides in the Paleozoic and one of the very few Lopingian localities with Paradoxichnium (Voigt and Lucas 2016).
A comparison with the ichnoassociation of the Val Gardena Sandstone of the Bletterbach site of the Dolomites is presently difficult, because of the necessity of a comprehensive taxonomic revision of these tracks (Abel 1929; Leonardi 1951; Leonardi et al. 1975; Conti et al. 1975, 1977; Ceoloni et al. 1988; Valentini et al. 2007, 2008, 2009; Bernardi et al. 2015). A list with all the ichnotaxa introduced from the Val Gardena Sandstone of Dolomites and other occurrences from the same formation of the Dolomites and Recoaro, with a provisional trackmaker attribution is provided in Table 2. At present, 5 ichnogenera and 13 ichnospecies are known, but most of them are of unclear ichnotaxonomic status, perhaps exclusive of only some intensively-studied ichnotaxa such as Pachypes dolomiticus Leonardi, Conti, Leonardi, Mariotti, and Nicosia, 1975 and Rhynchosauroides pallinii Conti, Leonardi, Mariotti, and Nicosia, 1977 (Valentini et al. 2007, 2008, 2009). The nomina dubia include: the ichnospecies Nanopus grimmi Leonardi, 1951, Ornithoidipius perwangeri Leonardi, 1951, and Phalangichnus perwangeri Conti , Leonardi, Mariotti, and Nicosia, 1977 and the ichnogenus Tridactylichnium leonardii Conti, Leonardi, Mariotti, and Nicosia, 1977 (Conti et al. 1980, 2000). We consider also the ichnogenus Prochirotherium permicum Leonardi, 1951 as nomen dubium, as it is based on a single imprint and lacks an adequate type series (Table 2). The most recent revision by Avanzini et al. (2011) highlights these ichnospecies: Chelichnus tazelwurmi, Dicynodontipus geinitzi, Ganasauripus ladinus, Hyloidichnus tirolensis, Ichniotherium accordii, Ichniotherium cottae, Janusichnus bifrons, Pachypes dolomiticus, Paradoxichnium radeinensis, Protochirotherium isp., Rhynchosauroides pallinii, Rhynchosauroides isp. 1, Rhynchosauroides isp. 2, and therapsid tracks indet. The material assigned to the Chelichnus, Hyloidichnus, Ichniotherium, and Paradoxichnium is probably not assignable to these ichnogenera, because: (i) the ichnogenus Chelichnus sensu McKeever and Haubold (1996) shows a relatively large and circular palm/sole and subequal and short digits in both the pes and the manus; this is not the case of “Chelichnus” tazelwurmi Ceoloni, Conti, Mariotti, and Nicosia, 1988, it is instead characterized by clearly ectaxonic tracks with relatively long digits and a short palm/sole; (ii) Hyloidichnus is characterized by straight digits with enlarged claws and a relatively short palm/sole and non-proximal pedal digit V, this is not the case of “Hyloidichnus” tirolensis Ceoloni, Conti, Mariotti, and Nicosia, 1988, which shows medially-bent digits terminating in acuminate claws and proximally-positioned digit V; (iii) Ichniotherium shows a marked increase of relative digit length from digit I–IV and digits more deeply impressed distally in both pes and manus; this is not the case in “Ichniotherium” accordii Ceoloni, Conti, Mariotti, and Nicosia, 1988, which is characterized by sub-equal digit length in the pes and “Ichniotherium cottae” (sensu Conti et al. 1977), characterized by a weak increase in length of digits I–IV. Moreover, both show a deeper impression of both the proximal and distal part of digits in the manual tracks; (iv) Paradoxichnium shows thick, straight and parallel manual digits II–IV with large triangular claws and proximally-positioned manual digits I and V, unlike “Paradoxichnium” radeinensis Ceoloni, Conti, Mariotti, and Nicosia, 1988. Additional occurrences from the Dolomitic area are Paradoxichnium and Hyloidichnus, supposed tracks of archosauromorph neodiapsids and captorhinid eureptiles (Voigt and Lucas 2016).
The main difference of the Val Gardena tetrapod ichnoassociation of the Dolomites with the Recoaro ichnoassociation is the occurrence of medium-sized therapsid synapsid tracks (e.g., part of the material identified as “Ichniotherium” accordii and “Ichniotherium cottae”, Dicynodontipus geinitzi), small-sized parareptile tracks (e.g., part of the material identified as “Chelichnus” tazelwurmi, Janusichnus bifrons, “Hyloidichnus” tirolensis) and chirotheriid tracks (Protochirotherium); and the non-occurrence of medium-sized ?parareptile tracks (Capitosauroides). However, a partial track from Recoaro might be tentatively attributed to therapsid producers. Derived captorhinid tracks occur in both sites, although with different genera (Hyloidichnus and Merifontichnus). The supposed archosauromorph track Paradoxichnium occurs in both sites. Rhynchosauroides and Pachypes occur in both sites and are the most abundant ichnotaxa for number of specimens. Therefore, the ichnoassociation of Val Gardena Sandstone in Dolomites and Recoaro seems rather similar, differing only in the occurrence of some ichnogenera/morphotypes such as Dicynodontipus, Protochirotherium, Hyloidichnus, and small parereptile tracks (Dolomites) or Merifontichnus and Capitosauroides (Recoaro). Both the ichnoassociations cleary belong to the Lopingian Paradoxichnium biochron sensu Voigt and Lucas (2016).
Table 2. Ichnotaxonomic status and possible attribution of the Lopingian tetrapod footprints from Italy (Val Gardena Sandstone; Recoaro and Dolomites). In bold ichnogenera and ichnospecies introduced from the Val Gardena Sandstone material. Valid ichnotaxa applied only to the ichnotaxa introduced from the Val Gardena Sandstone.
|
Locality |
Tetrapod ichnotaxa /morphotypes |
Valid |
Synapomorphy – related footprint features |
Possible trackmaker |
Reference |
|
Recoaro |
cf. Capitosauroides |
|
ectaxonic, long digits I–IV close and straight, short palm/sole, enlarged digit terminations, digit V slightly proximal |
?parareptile |
this study |
|
Recoaro |
cf. Merifontichnus isp. |
|
slightly ectaxonic, straight digits radially arranged, enlarged digit terminations, very short palm/sole |
captorhinid eureptile |
this study |
|
Recoaro |
Pachypes isp. |
|
broad, wide manus, very thick, straight, rounded digits I–IV in continuity with palm |
pareiasaurid parareptile |
this study |
|
Recoaro |
Paradoxichnium isp. |
|
ectaxonic, close goup of thick and straight digits II–IV terminating in large triangular claws, small digit V proximally-positioned (manus) |
archosauro-morph eureptile |
this study |
|
Recoaro |
Rhynchosauroides isp. |
|
long thin curved digits with sharp claws, markedly ectaxonic, digit V proximally-positioned and outwards-oriented, different morphology of pes and manus |
lacertoid neodiapsid eureptile |
this study |
|
Recoaro |
undetermined track |
|
paw-like footprints, basal digital pads deeply impressed |
therapsid synapsid |
this study |
|
Dolomites |
Prochirothrium |
nomen dubium |
|
|
|
|
Dolomites |
Nanopus grimmi |
nomen dubium |
|
|
|
|
Dolomites |
Ornithoidipus
|
nomen dubium |
|
|
|
|
Dolomites |
Pachypes dolomiticus |
valid |
ectaxonic pes and manus, very thick, straight, rounded digits I–IV in continuity with broad palm/sole, digit V small and slightly proximal |
pareiasaurid parareptile |
|
|
Dolomites |
Phalangichnus
|
nomen dubium |
|
|
|
|
Dolomites |
Rynchosauroides pallinii |
valid |
long curved digits with sharp claws, markedly ectaxonic, digit V proximally-positioned and outwards-oriented, different morphology of pes and manus |
lacertoid neodiapsid eureptile |
|
|
Dolomites |
Tridactylichnium
|
nomen dubium |
|
|
|
|
Dolomites |
“Hyloidichnus” |
valid |
ectaxonic, long digits I–IV close and straight, short palm/sole, small triangular claws, digit V slightly proximal |
small parareptile |
|
|
Dolomites |
“Ichniotherium” accordii |
valid |
Paw-like pentadactyl footprints, basal digital pads deeply impressed, mesaxonic pes and long sole |
therapsid synapsid |
|
|
Dolomites |
“Chelichnus”
|
valid |
ectaxonic, long digits I–IV close and straight, short palm/sole, small triangular claws, digit V slightly proximal |
small parareptile |
|
|
Dolomites |
“Paradoxichnium” |
valid |
ectaxonic, long digits I–IV close and straight, short palm/sole, small triangular claws, digit V slightly proximal |
parareptile |
|
|
Dolomites |
Janusichnus bifrons |
nomen dubium |
|
|
|
|
Dolomites |
Ganasauripus ladinus |
valid |
long thin curved digits with sharp claws, markedly ectaxonic, digit V proximally-positioned and outwards-oriented, different morphology of pes and manus |
lacertoid neodiapsid eureptile |
|
|
Dolomites |
Dycinodontipus geinitzi |
|
Paw-like pentadactyl footprints, similar length of digits, broad palm/sole |
therapsid synapsid |
|
|
Dolomites |
“Ichniotherium cottae” |
|
Paw-like pentadactyl footprints, basal digital pads deeply impressed, mesaxonic pes and long sole |
therapsid synapsid |
|
|
Dolomites |
cf. Protochirotherium isp. |
|
close goup of thick and straight digits I–IV, large digit V proximally-positioned and outwards-oriented, mesaxony |
archo- sauromorph eureptile |
|
|
Dolomites |
Hyloidichnus isp. |
|
slightly ectaxonic, straight digits radially arranged, enlarged digit terminations, very short palm/sole |
captorhinid eureptile |
|
|
Dolomites |
Paradoxichnium isp. |
|
ectaxonic, close goup of thick and straight digits II–IV terminating in large triangular claws, small digit V mesaxony-positioned (manus) |
archo- sauromorph eureptile |
Paleobiogeography.—The low-latitude Lopingian inland ichnoassociations of Morocco and Niger are characterized by small neodiapsid and parareptile tracks, medium-sized captorhinid tracks, large parareptile tracks, and temnospondyl tracks (Dromopus, Erpetopus, Hyloidichnus, Pachypes, Limnopus; Voigt et al. 2010; Hminna et al. 2012; Smith et al. 2015). The low-latitude Lopingian ichnoassociations of eolian environments from Germany and Scotland are dominated by supposed therapsid tracks (e.g., Chelichnus-like morphologies) and by probable small parareptile and lacertoid forms (Palmichnus and Akropus sensu Schmidt 1959) (Schmidt 1959; McKeever and Haubold 1996); whereas Lopingian tracks from low-latitude marginal marine environments of Germany only include archosauromorph tracks and small parereptile tracks to date (Paradoxichnium, Erpetopus; Müller 1959; Voigt 2012; Voigt and Lucas 2016). The mid- to high-latitude inland Lopingian ichnoassociations of South Africa and Australia are characterized by therapsid and small amniote tracks (Dicynodontipus and unidentified tracks; De Klerk 2002; Warren 1997). The preliminary study of the low-latitude marginal marine Lopingian Italian ichnofauna, especially as regards the trackmaker attribution, is provided in Table 2. Results indicate a predominance of possible parareptile (Pachypes, cf. Capitosauroides, and material attributed to “Paradoxichnium” radeinensis, “Hyloidichnus” tirolensis, “Chelichnus” tazelwurmi) and lacertoid neodiapsid tracks (Rhynchosauroides pallinii, Ganasauripus ladinus). Noteworthy the occurrence of archosauromorph tracks (cf. Protochirotherium, Paradoxichnium) and therapsid synapsid tracks (Dicynodontipus geinitzi, “Ichniotherium” accordii, “Ichniotherium cottae”, and an undetermined track). Therefore, archosauromorph tracks seem to be restricted to low-latitude marginal marine environments (Italy, Germany), and this could be linked to the trackmaker paleoecology.
Therapsid tracks seem to be reltively common in the low-latitude desert paleoenvironments of northern Pangaea. They are present in low-latitude marginal marine paleoenvironments (Italy). They are lacking in the low-latitude inland palaeoenvironments (Morocco, Niger). They were probably common in the high-latitude palaeoenvironments of southern Pangaea (South Africa). The absence of therapsid tracks in the low-latitude inland environments of Morocco and Niger is consistent with the skeletal record (Sidor et al. 2005). The hypothesis of coastal exchange of therapids between the northern and southern parts of Pangaea inferred from the skeleton record (Sidor et al. 2005), from a preliminary evaluation of the low-latitude marginal marine Val Gardena ichnotaxa/trackmakers, could be confirmed by the track record.
In order to verify and expand the discussion on these hypotheses, a comprehensive revision of the historic Bletterbach site material and a detailed study of the Lopingian tetrapod associations and ichnoassociations strictly considering paleolatitude, paleoenvironment, and paleobiogeography is the most urgent task.
Conclusions
The new study of the Lopingian tetrapod footprints of the Recoaro area of Venetian Prealps highlights a relatively diverse ichnoassociation, including the first possible occurrence of Merifontichnus and Capitosauroides in Lopingian strata and one of the few occurrences of Paradoxichnium. The comparison of the ichnoassociation in terms of different lithofacies highlights a probable paleoecologic control on the tetrapod ichnoassociation, which is relatively diverse in floodplain environments and less diverse with opportunistic/specialized forms in sabkha and lagoon environments. A comparison with other Lopingian ichnosites shows a probable preference of Permian archosauromorphs for marginal marine environments. A comparison with the ichnoassociation of the classic Lopingian site of the Dolomites (Bletterbach gorge) shows several similarities and the necessity of a comprehensive ichnotaxonomic revision of the potentially most important Lopingian site of the World either for quality of preservation, abundance of material and ichnotaxonomic diversity.
Acknowledgments
We would like to thank the late Ottaviano Violati Tescari (Vicenza, Italy), the late Olivo Gonzo (Valdagno, Italy), the late Antonio Rossi (Cornedo Vicentino, Italy), and Franco Slaviero (Valdagno, Italy) for collecting and donating the specimens; Bernardetta Pallozzi (Museo Civico D. Dal Lago, Valdagno, Italy), Mariagabriella Fornasiero (Museo di Paleontologia e Geologia of the University of Padua, Padua, Italy), Massimo Bernardi and Riccardo Tomasoni (both MUSE, Trento, Italy), Elio Dellantonio and Daniele Rossi (both Museo Geologico delle Dolomiti, Predazzo, Italy), Birgit Gaitzsch, Jörg W. Schneider, and Frank Scholze (all Technische Universität Bergakademie Freiberg, Freiberg, Germany), and Sebastian Voigt (Urweltmuseum Geoskop, Thallichtenberg, Germany) for the access to the collections and the useful discussions; the reviewers Massimo Bernardi and Grzegorz Niedźwiedzki (Uppsala University, Uppsala, Sweden). Alexander von Humboldt Foundation gave financial support to this study.
References
Abel, O. 1929. Eine Reptilien Fährte aus dem Oberen Perm (Grodener Sandstein) der Gegend von Bozen in Sudtirol. Paleobiology 2: 1–14.
Accordi, B. 1956. Calcareous algae from the Upper Permian of the Dolomites (Italy), with stratigraphy of the “Bellerophon-zone”. Journal of the Paleontological Society of India 1: 75–84.
Avanzini, M. and Tomasoni, R. 2004. Giornate di Paleontologia 2004, Bolzano 21–23 maggio 2004, Guida all’escursione: la gola del Bletterbach. Studi Trentini di Scienze naturali, Acta Geologica 79 (Supplement): 1–34.
Avanzini, M., Bernardi, M., and Nicosia, U. 2011. The Permo-Triassic tetrapod faunal diversity in the Italian Southern Alps. Earth and Environmental Sciences 25: 591–608. Crossref
Barbieri, G., De Vecchi, G., De Zanche, V., Di Lallo, E., Frizzo, P., Mietto, P., and Sedea, R. 1980. Note Illustrative della carta geologica dell’area di Recoaro alla scala 1:20000. Memorie di Scienze Geologiche 34: 23–52.
Bernardi, M., Klein, H., Petti, F.M., and Ezcurra, M.D. 2015. The origin and early radiation of archosauriforms: integrating the skeletal and footprint record. PloS One 10: e0128449. Crossref
Bertling, M., Braddy, S.J., Bromley, R.G., Demathieu, G.R., Genise, J., Mikuláš, R., Nielsen, J.K., Nielsen, K.S.S., Rindsberg, A.K., Schlirf, M., and Uchman, A. 2006. Names for trace fossils: a uniform approach. Lethaia 39: 265–286. Crossref
Broglio Loriga, C. 1960. Foraminiferi del Permiano superiore delle Dolomiti (Val Gardena, Val Badia, Val Marebbe). Bollettino della Società Paleontologica Italiana 1: 33–73.
Broglio Loriga, C., Neri, C., Pasini, M., and Posenato, R. 1988. Marine Fossil Assemblages from Upper Permian to Lowermost Triassic in the Western Dolomites (Italy). Memorie della Società Geologica Italiana 34: 5–44.
Cassinis, G., Perotti, C.R., and Ronchi, A. 2012. Permian continental basins in the Southern Alps (Italy) and peri-mediterranean correlations. International Journal of Earth Sciences 101: 129–157. Crossref
Ceoloni, P., Conti, M.A., Mariotti, N., and Nicosia, U. 1988. New Late Permian tetrapod footprints from southern Alps. Memorie della Società Geologica Italiana 34: 45–65.
Citton, P., Nicolosi, I., Carluccio, R., and Nicosia, U. 2016. Unveiling trampling history through trackway interferences and track preservational features: a case study from the Bletterbach gorge (Redagno, Western Dolomites, Italy). Palaeontologia Electronica 19: 1–20. Crossref
Conti, M.A., Leonardi, G., Mariotti, N., and Nicosia, U. 1975. Tetrapod footprints, fishes and molluscs from the Middle Permian of the Dolomites (N. Italy). Memorie Geopaleontologiche dell’Università di Ferrara 3: 139–150.
Conti, M.A., Leonardi, G., Mariotti, N., and Nicosia, U. 1977. Tetrapod footprints of the “Arenaria di Val Gardena”(North Italy). Their paleontological, stratigraphic and palaeoenvironmental meaning. Paleontographica Italica 70: 1–91.
Conti, M.A., Leonardi, G., Mariotti, N., and Nicosia, U. 1980. Tetrapodi e pesci della Val Gardena. I vertebrati fossili italiani. Catalogo della Mostra di Verona: 51–60.
Conti, M.A., Leonardi, G., Mietto, P., and Nicosia, U. 2000. Orme di tetrapodi non dinosauriani del Paleozoico e Mesozoico in Italia. In: G. Leonardi and P. Mietto (eds.), Dinosauri in Italia. Le orme giurassiche dei Lavini di Marco (Trentino) e gli altri resti fossili italiani, 297–320. Accademia Editoriale, Pisa.
Costa da Silva, R., Sedor, F.A., and Fernandes, A.C.S. 2012. Fossil footprints from the Late Permian of Brazil: an example of hidden biodiversity. Journal of South American Earth Sciences 38: 31–43. Crossref
Dalla Vecchia, F.M. 2013. Sulle tracce del passato—Pistee impronte fossili di vertebrati in Carnia e nelle areecontermini. 176 pp. Comunità Montana della Carnia, Tolmezzo.
De Klerk, W.J. 2002. A dicynodont trackway from the Cistecephalus assemblage zone in the Karoo, east of Graaff-Reinet, South Africa. Palaeontologia Africana 38: 73–91.
Farabegoli, E. and Perri, M.C. 1998. Permian/Triassic boundary and Early Triassic of the Bulla section
(Southern Alps, Italy): lithostratigraphy, facies and conodont biostratigraphy. Giornale di Geologia Ser. 3a 60: 292–311.
Gand, G., Garric, J., Demathieu, G., and Ellenberger, P. 2000. La palichnofaune de vertebres tétrapodes du Permien supérieur du bassin de Lodeve (Languedoc-France). Palaeovertebrata 29: 1–82.
Gilmore, G.W. 1927. Fossil footprints from the Grand Canyon: second contribution. Smithsonian Miscellaneous Collections 80: 1–78.
Gubin, Y.M., Golubev, V.K., Bulanov, V.V., and Petuchov, S.V. [Petuhov, S.V.] 2003. Pareiasaurian tracks from the upper Permian of Eastern Europe [in Russian with English abstract]. Palaeontological Journal 37: 514–523.
Haubold, H . 1970. Versuch der Revision der Amphibien-Fährten des Karbon und Perm. Freiberger Forschungshefte C 260: 83–117.
Haubold, H. 1971. Die Tetrapodenfährten des Buntsandsteins in der Deutschen Demokratischen Republik und in Westdeutschland und ihre Äquivalente in der gesamten Trias. Pälaontologische Abhandlungen, Abteilung A, Paläozoologie IV 3: 395–660.
Haubold, H. 1996. Ichnotaxonomie und Klassifikation von Tetrapodenfährten aus dem Perm. Hallesches Jahrbuch für Geowissenschaften B 18: 23–88.
Haubold, H., Hunt, A.P., Lucas, S.G., and Lockley, M.G. 1995. Wolfcampian (Early Permian) vertebrate tracks from Arizona and New Mexico. New Mexico Museum of Natural History and Science Bulletin 6: 135–165.
Hminna, A., Voigt, S., Saber, H., Schneider, J.W., and Hmich, D. 2012. On a moderately diverse continental ichnofauna from the Permian Ikakern Formation (Argana Basin, Western High Atlas, Morocco). Journal of African Earth Sciences 68: 15–23. Crossref
Holmes, R.B. 2003. The hind limb of Captorhinus aguti and the step cycle of basal amniotes. Canadian Journal of Earth Sciences 40: 515–526. Crossref
Hunt, A.P. and Lucas, S.G. 2006. Permian tetrapod ichnofacies. Geological Society, London, Special Publications 265: 137–156. Crossref
Hunt, A.P. and Lucas, S.G. 2007. Tetrapod ichnofacies: a new paradigm. Ichnos 14: 59–68. Crossref
Kittl, E. 1891. Saurier Fährte von Bozen. Mitteilungen Österreichischen Touristen Klub 3: 7.
Klein, H. and Lucas, S.G. 2010. Review of the tetrapod ichnofauna of the Moenkopi Formation/Group (Early–Middle Triassic) of the American Southwest. New Mexico Museum of Natural History and Science Bulletin 50: 1–67.
Kümmell, S.B. and Frey, E. 2012. Digital arcade in the autopodia of Synapsida: standard position of the digits and dorsoventral excursion angle of digital joints in the rays II–V. Palaeobiodiver Palaeoenviron 92: 171–196. Crossref
Kustatscher, E., Bauer, K., Butzmann, R., Fischer, T.C., Meller, B., van Konijnenburg-van Cittert, J.H., and Kerp, H. 2014. Sphenophytes, pteridosperms and possible cycads from the Wuchiapingian (Lopingian, Permian) of Bletterbach (Dolomites, northern Italy). Review of Palaeobotany and Palynology 208: 65–82. Crosssref
Kustatscher, E., Bernardi, M., Petti, F.M., Franz, M., van Konijnenburg-van Cittert, J. H., and Kerp, H. 2017. Sea-level changes in the Lopingian (late Permian) of the northwestern Tethys and their effects on the terrestrial palaeoenvironments, biota and fossil preservation. Global and Planetary Change 148: 166–180. Crossref
Kustatscher, E., van Konijnenburg-van Cittert, J.H., Bauer, K., Butzmann, R., Meller, B., and Fischer, T.C. 2012. A new flora from the Upper Permian of Bletterbach (Dolomites, N-Italy). Review of Palaeobotany and Palynology 182: 1–13. Crossref
Leonardi, G. 1974. Sobre una serie de rastos de tetrapodes do Permiano das Dolomitas (Italia). Anais XXVIII Congresso Sociedade Brasileira Geologia 2: 205–220.
Leonardi, G. 1987. Glossary and Manual of Tetrapod Footprint Palaeoichnology. 117 pp. Departamento Nacional de Produção Mineral, Brasilia.
Leonardi, P. 1951. Orme di tetrapodi nelle arenarie di Val Gardena (Permiano medio-inferiore) dell’Alto Adige sud-orientale. Memorie dell’Istituto di Geologia e Mineralogia dell’Università di Padova 17: 1–23.
Leonardi, P., Conti, M.A., Leonardi, G., Mariotti, N., and Nicosia, U. 1975. Pachypes dolomiticus n. gen. n. sp.; Pareiasaur footprint from the “Val Gardena Sandstone” (Middle Permian) in the Western Dolomites (N. Italy). Atti della Accademia Nazionale dei Lincei, Rendiconti—Classe di Scienze Fisiche-Matematiche e Naturali 57: 221–232.
Lucas, S.G. 2006. Global Permian tetrapod biostratigraphy and biochronology. Geological Society, London, Special Publications 265: 65–93. Crossref
Maidwell, F.T. 1911. Notes on footprints from the Keuper of Runcorn Hill. Proceedings of the Liverpool Geological Society 11: 140–152.
Marchetti, L. 2016. New occurrences of tetrapod ichnotaxa from the Permian Orobic Basin (Northern Italy) and critical discussion of the age of the ichnoassociation. Papers in Palaeontology 2: 363–386. Crossref
Marchetti, L., Bernardi, M., and Avanzini, M. 2013. Some insights on well-preserved Amphisauropus and Erpetopus trackways from the Eastern Collio basin (Trentino-Alto Adige, NE Italy). Bollettino della Società Paleontologica Italiana 52: 55–62.
Marocchi, M., Morelli, C., Mair, V., Klötzli, U., and Bargossi, G.M. 2008. Evolution of large silicic magma systems: new U-Pb zircon data on the NW Permian Athesian Volcanic Group (Southern Alps, Italy. The Journal of Geology 116: 480–498. Crossref
Marsh, O.C. 1894. I. Footprints of vertebrates in the Coal-Measures of Kansas. Geological Magazine Decade IV 1: 337–339. Crossref
Massari, F. and Neri, C. 1997. The infill of a supradetachment (?) basin: the continental to shallow-marine Upper Permian succession in the Dolomites and Carnia (Italy). Sedimentary Geology 110: 181–221. Crossref
Massari, F., Conti M.A., Fontana D., Helmod K., Mariotti N., Neri, C., Nicosia U., Ori G.G., Pasini M., and Pittau D. 1988. The val Gardena sandstone and Bellerophon Formation in the Bletterbach gorge (Alto Adige, Italy): biostratigraphy and sedimentology. Memorie di Scienze Geologiche 40: 229–273.
Massari, F., Neri, C., Pittau, P., Fontana, D., and Stefani, C. 1994. Sedimentology, palynostratigraphy and sequence stratigraphy of a continental to shallow-marine rift-related succession: Upper Permian of the eastern Southern Alps (Italy). Memorie di Scienze Geologiche 46: 119–243.
McKeever, P.J. and Haubold, H. 1996. Reclassification of vertebrate trackways from the Permian of Scotland and related forms from Arizona and Germany. Journal of Paleontology 70: 1011–1022. Crossref
Mietto, P. 1975. Orme di tetrapodi nelle arenarie permiche di Recoaro (Vicenza). Studi Trentini di Scienze Naturali 52: 57–67.
Mietto, P. 1981. Una grande impronta di pareiasauro nel Permiano di Recoaro (Vicenza). Rendiconti della Società Geologica Italiana 4: 363–364.
Mietto, P. 1995. Guida alla visita del Museo Paleontologico “Dott. Domenico Dal Lago”. 74 pp. Centro Comunale “G. Marzotto”, Valdagno.
Müller, A.H. 1959. Die erste Wirbeltierfährte (Paradoxichnium problematicum n. g. n. sp.) aus dem terrestrischen Zechstein von Thüringen. Monatsberichte der Deutschen Akademie der Wissenschaften 1: 613–623.
Neri, C. 2007. Arenaria di Val Gardena. In: M.B. Cita, E. Abbate, M. Balini, M.A. Conti, P. Falorni, D. Germani, G. Groppelli, P. Manetti, and F.M. Petti (eds.), Carta Geologica d italia 1:50000, 52–63. APAT, Quaderni del Servizio Geologico d’Italia, Rome.
Newell, N.D., Rigby, J.K., Driggs, A., Boyd, D.W., and Stehli, F.G. 1976. Permian reef complex, Tunisia. Brigham Young University Geology Studies 23: 72–112.
Nicosia, U., Sacchi, E., and Spezzamonte, M. 2001. New palaeontological data for the Val Gardena sandstone. Annali del Museo Civico di Science Naturali di Brescia 25: 83–88.
O’Keefe, F.R., Sidor, C.A., Larsson, H.C., Maga, A., and Ide, O. 2005. The vertebrate fauna of the Upper Permian of Niger—III, morphology and ontogeny of the hindlimb of Moradisaurus grandis (Reptilia, Captorhinidae). Journal of Vertebrate Paleontology 25: 309–319. Crossref
Olson, E.C. 1962. Part II. The osteology of Captorhinikos chozaensis Olson. In: E.C. Olson and H. Barghusen (eds.), Permian Vertebrates From Oklahoma and Texas (Vol. 2), 49–68. University of Oklahoma, Norman.
Pasini, M. 1985. Biostratigrafia con i foraminiferi del limite formazione a Bellerophon/Formazione di Werfen fra Recoaro e La Val Badia (Alpi Meridionali). Rivista Italiana di Paleontologia e Stratigrafia 90: 481–510.
Posenato, R. 1988. Chronological and geographic distribution of the Fam. Comelicanidae Merla, 1930 (Brachiopods). Rivista Italiana di Paleontologia e Stratigrafia 94: 383–398.
Posenato, R. 1998. The gen. Comelicania Frech, 1901 (Brachiopoda) from the Southern Alps: morphology and classification. Rivista Italiana di Paleontologia e Stratigrafia 104: 43–68.
Posenato, R. and Prinoth, H. 1999. Discovery of Paratirolites from the Bellerophon Formation (Upper Permian, Dolomites, Italy). Rivista Italiana di Paleontologia e Stratigrafia 105: 129–134.
Pittau, P. 2005. The microflora. In: P. Pittau, H. Kerp, and E. Kustatscher (eds.), The Bletterbach Canyon: “Let Us Meet Across the P/T Boundary”, Workshop on Permian and Triassic Palaeobotany and Palynology, Bozen 16–18.06.2005, Excursion Guide, 9–19. Museum of Nature South Tirol, Bozen.
Schmidt, H. 1959. Die Cornberger Fährten im Rahmen der Vierfüssler-Entwicklung. Abhandlungen hessisches Landesamt für Bodenforschung 28: 1–137.
Sidor, C.A., O’Keefe, F.R., Damiani, R., Steyer, J.S., Smith, R.M., Larsson, H.C., Sereno, P.C., Ide, O., and Maga, A. 2005. Permian tetrapods from the Sahara show climate-controlled endemism in Pangaea. Nature 434: 886–889. Crossref
Smith, R.M., Sidor, C.A., Tabor, N.J., and Steyer, J.S. 2015. Sedimentology and vertebrate taphonomy of the Moradi Formation of northern Niger: A Permian wet desert in the tropics of Pangaea. Palaeogeography, Palaeoclimatology, Palaeoecology 440: 128–141. Crossref
Spina, A., Cirilli, S., Utting, J., and Jansonius, J. 2015. Palynology of the Permian and Triassic of the Tesero and Bulla sections (Western Dolomites, Italy) and consideration about the enigmatic species Reduviasporonites chalastus. Review of Palaeobotany and Palynology 218: 3–14. Crossref
Sumida, S.S. 1989. The appendicular skeleton of the Early Permian genus Labidosaurus (Reptilia, Captorhinomorpha, Captorhinidae) and the hind limb musculature of captorhinid reptiles. Journal of Vertebrate Paleontology 9: 295–313. Crossref
Surkov, M.V., Benton, M.J., Twitchett, R.J., Tverdokhlebov, V.P., and Newell, A.J. 2007. First occurrence of footprints of large therapsids from the Upper Permian of European Russia. Palaeontology 50: 641–652. Crossref
Valentini, M., Conti, M.A., and Mariotti, N. 2007. Lacertoid footprints of the Upper Permian Arenaria di Val Gardena Formation (Northern Italy). Ichnos 14: 193–218. Crossref
Valentini, M., Conti, M.A., and Nicosia, U. 2008. Linking tetrapod tracks to the biodynamics, paleobiogeography, and paleobiology of their trackmakers: Pachypes dolomiticus Leonardi et al., 1975, a case study. Acta Geologica 83: 237–246.
Valentini, M., Nicosia, U., and Conti, M.A. 2009. A re-evaluation of Pachypes, a pareiasaurian track from the Late Permian: Neues Jahrbuch für Geologie und Paläontologie-Abhandlungen 251: 71–94. Crossref
Voigt, S. 2005. Die Tetrapodenichnofauna des kontinentalen Oberkarbon und Perm im Thüringer Wald—Ichnotaxonomie, Paläoökologie und Biostratigraphie. 179 pp. Cuvillier, Göttingen.
Voigt, S. 2012. Tetrapodenfährten im Rotliegend. Schriftenreihe der Deutschen Gesellschaft für Geowissenschaften 61: 161–175.
Voigt, S. and Lucas, S.G. 2016. Outline of a Permian tetrapod footprint ichnostratigraphy. In: S.G. Lucas and S.Z. Shen (eds.), The Permian Timescale. Geological Society, London, Special Publications 450: 1–18. Crossref
Voigt, S., Hminna, A., Saber, H., Schneider, J.W., and Klein, H. 2010. Tetrapod footprints from the uppermost level of the Permian Ikakern Formation (Argana basin, western High Atlas, Morocco). Journal of African Earth Sciences 57: 470–478. Crossref
Warren, A. 1997. A tetrapod fauna from the Permian of the Sydney Basin. Records-Australian Museum 49: 25–34. Crossref
Wopfner, H. 1999. Über Tetrapoden-Fährten, Kohlen und versteinerte Hölzer aus dem Grödner Sandstein (Perm) bei Deutschnofen. Der Schlern 73: 23–32.
Acta Palaeontol. Pol. 62 (4): 801–817, 2017
https://doi.org/10.4202/app.00392.2017