
A re-evaluation of the enigmatic dinosauriform Caseosaurus crosbyensis from the Late Triassic of Texas, USA and its implications for early dinosaur evolution
MATTHEW G. BARON and MEGAN E. WILLIAMS
Baron, M.G. and Williams, M.E. 2018. A re-evaluation of the enigmatic dinosauriform Caseosaurus crosbyensis from the Late Triassic of Texas, USA and its implications for early dinosaur evolution. Acta Palaeontologica Polonica 63 (1): 129–145.
The holotype specimen of the Late Triassic dinosauriform Caseosaurus crosbyensis is redescribed and evaluated phylogenetically for the first time, providing new anatomical information and data on the earliest dinosaurs and their evolution within the dinosauromorph lineage. Historically, Caseosaurus crosbyensis has been considered to represent an early saurischian dinosaur, and often a herrerasaur. More recent work on Triassic dinosaurs has cast doubt over its supposed dinosaurian affinities and uncertainty about particular features in the holotype and only known specimen has led to the species being regarded as a dinosauriform of indeterminate position. Here, we present a new diagnosis for Caseosaurus crosbyensis and refer additional material to the taxon—a partial right ilium from Snyder Quarry. Our comparisons and phylogenetic analyses suggest that Caseosaurus crosbyensis belongs in a clade with herrerasaurs and that this clade is the sister taxon of Dinosauria, rather than positioned within it. This result, along with other recent analyses of early dinosaurs, pulls apart what remains of the “traditional” group of dinosaurs collectively termed saurischians into a polyphyletic assemblage and implies that Dinosauria should be regarded as composed exclusively of Ornithoscelida (Ornithischia + Theropoda) and Sauropodomorpha. In addition, our analysis recovers the enigmatic European taxon Saltopus elginensis among herrerasaurs for the first time. This result suggests a greater body-size range for herrerasaurs than previously thought and provides further evidence for their presence in Europe during the Late Triassic. If this hypothesis is correct then this clade of herrerasaurs also represents the first clade of non-dinosaurian dinosauromorphs known to contain large-bodied carnivorous species. The results of our analyses also highlight the distinction between the clades Herrerasauridae and Herrerasauria, as they are currently defined, and necessitate a provisional revival of the latter until future works can better resolve the relationships among these important early taxa.
Key words: Archosauria, Dinosauromorpha, Herrerasauria, phylogeny, Dockum Group, Triassic, Texas, USA.
Matthew G. Baron [mgb46@cam.ac.uk], Department of Earth Science, University of Cambridge, Downing Street, Cambridge, CB2 3EQ, UK; Department of Earth Sciences, Natural History Museum, London, Cromwell Road, London SW7 5BD, UK.
Megan E. Williams [mew41@cam.ac.uk], Department of Earth Science, University of Cambridge, Downing Street, Cambridge, CB2 3EQ, UK.
Received 8 April 2017, accepted 3 October 2017, available online 10 January 2018.
Copyright © 2018 M.G. Baron and M.E. Williams. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
The Late Triassic formations of North America have yielded numerous dinosauromorph specimens, including various dinosaurs (Chatterjee 1984; Sullivan and Lucas 1999; Irmis et al. 2007a, b; Nesbitt et al. 2009a; Sarıgül 2016). Understanding the faunal composition of this spatiotemporal setting, the taxonomic diversity and disparity that is represented in this part of the fossil record, and what this can tell us about the Late Triassic world, are all important factors for gaining a better understanding of the early evolution of dinosaurs and dinosauromorphs, particularly with regard to the eventual rise in prominence of Dinosauria following this transitional period.
Both the Dockum Group and the Chinle Formation have yielded silesaurids, as well as other non-dinosaurian dinosauromorphs (Irmis et al. 2007a, b; Nesbitt et al. 2007; Sarıgül 2016); the Petrified Forest Member of the Chinle Formation of New Mexico and the Colorado City Formation of the Dockum Group of Texas have both produced specimens of the non-dinosaurian dinosauromorph Dromomeron (Irmis et al. 2007a; Nesbitt et al. 2009a). Additionally, the Petrified Forest Member of the Chinle Formation has produced specimens of the theropod dinosaur Coelophysis and the saurischian Chindesaurus bryansmalli that, along with other taxa such as Eucoelophysis baldwini, Technosaurus smalli and the controversial taxon Protoavis texensis, forms part of the rich dinosauromorph fauna of Late Triassic North America (Cope 1889; Chatterjee 1984, 1991; Long and Murry 1995; Sullivan and Lucas 1999; Nesbitt et al. 2007; Sarıgül 2016). Recently Sarıgül (2016) gave a detailed account of the fossil localities and general geological settings of the Dockum Group and other areas of palaeontological interest in the southwestern portion of North America (e.g., the Bull Canyon Formation, “siltstone member”, etc.), so we do not repeat that information here. However, we do note that a variety of dinosauromorph forms inhabited this expansive series of ancient river systems during this important stage in dinosaur and dinosauromorph evolution, with dinosaurs and non-dinosaurian dinosauromorphs clearly co-existing (Nesbitt et al. 2007; Nesbitt 2011; Sarıgül 2016).
Caseosaurus crosbyensis Hunt, Lucas, Heckert, Sullivan, and Lockley, 1998 is known from a single, almost complete right ilium (UMMP 8870; Fig. 1), which was recovered from the Tecovas Member (?upper Carnian) of the Dockum Group (Hunt et al. 1998; Nesbitt et al. 2007). The exact affinities of Caseosaurus crosbyensis have been the source of much debate, with numerous studies proposing different positions in the dinosauromorph tree for these partial remains (Case 1927; Long and Murry 1995; Hunt et al. 1998; Nesbitt et al. 2007; Ezcurra 2010). Initially, Case (1927) regarded the holotype material as referable to Coelophysis sp.; this initial assignment to Theropoda started a trend that continued to classify Caseosaurus crosbyensis as a saurischian dinosaur. This trend was followed by many subsequent studies (Long and Murry 1995; Hunt et al. 1998; Nesbitt et al. 2007; Ezcurra 2010), but has never been tested using phylogenetic methods. After Case (1927), other studies initially removed UMMP 8870 from Coelophysis or placed it within the hypodigm of Chindesaurus bryansmalli (Long and Murry 1995) but, eventually, it was referred to a new genus and species, Caseosaurus crosbyensis (Hunt et al. 1998). Long and Murry (1995) were the first to suggest a close relationship between this specimen and the South American saurischian dinosaur Herrerasaurus ischigualastensis and this idea was restated by Hunt et al. (1998). However, despite the conclusions of Hunt et al. (1998), Langer (2004) listed UMMP 8870 as a specimen of Chindesaurus bryansmalli in his discussion of basal saurischians, following Long and Murry (1995). Nesbitt et al. (2007) reassessed the material as part of a broader study of Late Triassic dinosaurs from North America and concluded that Caseosaurus crosbyensis should provisionally be considered as a valid taxon, distinct from Chindesaurus, based upon the fact that the specimens of each taxon are too incomplete to allow formal synonymisation. Nesbitt et al. (2007) also stated that Caseosaurus could not be assigned unambiguously to either Herrerasauridae or Dinosauria, due to the incomplete nature of the holotype specimen. Nesbitt et al. (2007) cited the partially open acetabulum as evidence that Caseosaurus belongs to Dinosauriformes but provided no further systematic discrimination. As a result, Caseosaurus crosbyensis is currently regarded as an indeterminate dinosauriform (Nesbitt et al. 2007). In the same study, the overall similarity between the holotype of Caseosaurus crosbyensis (UMMP 8870) and a partial ilium from the Snyder Quarry in New Mexico (NMMNH P-35995) was also highlighted (Nesbitt et al. 2007). NMMNH P-35995 was originally assigned to Eucoelophysis sp. by Heckert et al. (2000, 2003), prior to the referral of Eucoelophysis to Silesauridae (Ezcurra 2006; Nesbitt et al. 2007; Nesbitt 2011), but has received little attention in the literature since. Due to the lack of evidence for a fully perforate acetabulum in this specimen, Nesbitt et al. (2007) concluded that NMMNH P-35995 could not be referred to Dinosauria, but only to Dinosauriformes. Despite the remarkable similarity between these two specimens, Nesbitt et al. (2007) did not refer the ilium from Snyder Quarry to Caseosaurus crosbyensis due to a lack of synapomorphies between it and the holotype of Caseosaurus crosbyensis. Neither specimen was assessed phylogenetically in the study of Nesbitt et al. (2007), nor were they included in the analyses of Ezcurra (2010), Nesbitt et al. (2010), Nesbitt (2011), Novas et al. (2011) or Martinez et al. (2013). Hence, the phylogenetic affinities of neither UMMP 8870 nor NMMNH P-35995 have ever been assessed using a numerical phylogenetic analysis and both specimens currently require further evaluation if their positions within Dinosauriformes are to be better constrained. Given the high level of disagreement surrounding Caseosaurus crosbyensis and its position within Dinosauriformes, as well as the position of the important dinosauriform clade Herrerasauridae (Novas et al. 2011; Sues et al. 2011; Bittencourt et al. 2015; Baron et al. 2017a), with which Caseosaurus crosbyensis has previously been linked (Long and Murry 1995; Hunt et al. 1998), a thorough re-evaluation of this material seems necessary to help clarify some of the uncertainty around the base of the dinosaurian tree and to increase our understanding of the Late Triassic dinosauromorph fauna of western North America (Nesbitt et al. 2007; Sarıgül 2016).
The recent phylogenetic study of early dinosaurs and other dinosauromorphs by Baron et al. (2017a) presented the largest and most comprehensive early dinosaur dataset that has been assembled to date. This dataset offers a new way to assess the phylogenetic affinities of enigmatic dinosauriform taxa such as Caseosaurus crosbyensis, and what it can tell us about the base of the dinosaurian lineage, and is used for this purpose in our study.
Institutional abbreviations.—AMNH, American Museum of Natural History, New York, USA; MACN, Museo Argentino de Ciencias Naturales “Bernardino Rivadavia”, Paleontologia de Vertebrados, Buenos Aires, Argentina; MCP, Museu de Ciências e Tecnologia PUCRS, Porto Alegre, Brazil; MCN, Museu de Ciencias Naturais, Rio Grande do Sul, Brazil; MCZ, Museum of Comparative Zoology, Harvard University, Cambridge, USA; NHCC, National Heritage Conservation Commission of Zambia, Lusaka, Zambia; NHMUK, Natural History Museum, London, UK; NMMNH, New Mexico Museum of Natural History and Science, Albuquerque, USA; PULR, Universidad Nacional de La Rioja, La Rioja, Argentina; PVL, Fundación Miguel Lillo, San Miguel de Tucumán, Argentina; PVSJ, Museo de Ciencias Naturales, Universidad Nacional de San Juan, San Juan, Argentina; UMMP, University of Michigan Museum of Paleontology, Ann Arbor, USA; USNM, United States National Museum (now National Museum of Natural History), Washington D.C., USA; ZPAL, Institute of Paleobiology, Polish Academy of Sciences, Warsaw, Poland.
Other abbreviations.—MPTs, most parsimonious trees; OTUs, operational taxonomic units.

Fig. 1. The holotype of the herrerasaurian dinosauriform Caseosaurus crosbyensis Hunt, Lucas, Heckert, Sullivan, and Lockley, 1998 (UMMP 8870) from Tecovas Formation, Crosby County, Texas, USA; Carnian, Late Triassic; right ilium in lateral (A) and medial (B) views.
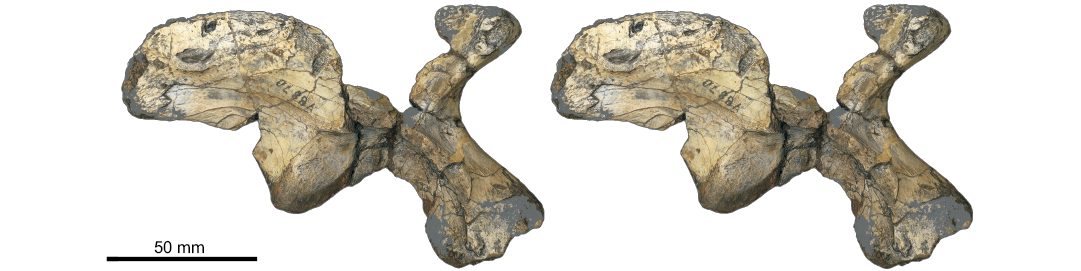
Fig. 2. The holotype of the herrerasaurian dinosauriform Caseosaurus crosbyensis Hunt, Lucas, Heckert, Sullivan, and Lockley, 1998 (UMMP 8870) from Tecovas Formation, Crosby County, Texas, USA; Carnian, Late Triassic; right ilium in lateral view; stereopair. Image credit: Adam Rountrey, Museum of Paleontology, University of Michigan, Ann Arbor, USA.
Material and methods
UMMP 8870 (Figs. 1, 2) is a single, fairly complete right ilium, which is only missing a small portion of the dorsal blade and part of the medioventral wall of the acetabulum. NMMNH P-35995 (Fig. 3) is a partial right ilium, which lacks the postacetabular process, part of the medioventral acetabular wall (although it preserves more of its medioventral wall than in UMMP 8870) and a large portion of the dorsal blade (Heckert et al. 2000, 2003; Nesbitt et al. 2007).
For the phylogenetic analyses, the holotype specimen of Caseosaurus crosbyensis and NMMNH P-35995 were added to the dataset of Baron et al. (2017a), along with several additional saurischian and theropod taxa, the Triassic sauropodomorphs Chromogisaurus novasi and Buriolestes shultzi, the Triassic theropods Daemonosaurus chauliodus and Lepidus praecisio and the Early Jurassic theropod Segisaurus halli were added in order to broaden the range of sampled taxa that would traditionally be regarded as “early saurischians”. Dimorphodon macronyx was included as an additional outgroup taxon, following its use in the study by Baron et al. (2017a: extended data fig. 2). The other difference between the treatment of operational taxonomic units (OTUs) in this study and Baron et al. (2017a) is the scoring of Agnosphitys cromhallensis. In the original study, the whole of the hypodigm of Agnosphitys cromhallensis was considered when scoring this taxon (Baron et al. 2017a) and it was recovered as a member of Silesauridae. However, the combination of anatomical characters presented by the various elements of the hypodigm of A. cromhallensis is unusual and a revision of this material is needed. For now, this study considers only the holotype specimen of A. cromhallensis, VMNH 1745—a left ilium (Fraser et al. 2002); A. cromhallensis was treated in the same way by Ezcurra (2010). Additional information on the anatomy of Guaibasaurus candelariensis was also added, based upon the observations made by Langer et al. (2011).

Fig. 3. The herrerasaurian dinosauriform Caseosaurus crosbyensis Hunt, Lucas, Heckert, Sullivan, and Lockley, 1998 (NMMNH P-35995) from Synder Quarry, Rio Arriba County, New Mexico, USA; Carnian, Late Triassic; right ilium in lateral (A) and medial (B) views.
A total of 82 taxa were scored for 457 characters. Following Baron et al. (2017a) characters 24, 35, 39, 60, 68, 71, 117, 145, 167, 169, 174, 180, 197, 199, 206, 214, 215, 222, 251, 269, 272, 286, 289, 303, 305, 307, 313, 322, 333, 334, 338, 353, 360, 376, 378, 387, 393, 442, 446 were treated as ordered.
Both Caseosaurus crosbyensis (UMMP 8870 and NMMNH P-35995) were scored as separate OTUs. Neither author has seen the specimen first hand; our description, comparisons and analyses are based upon information gathered from photographs of the material, personal communication from a number of people based at the collection in which the material is housed, as well as others who have examined the specimen first hand, from information in the literature (Hunt et al. 1998; Nesbitt et al. 2007; Ezcurra 2010), and from a high quality, manipulatable 3D digital model, which is available online (https://umorf.ummp.lsa.umich.edu/wp/specimen-data/?Model_ID=1241). In spite of this, a great deal of anatomical information could be obtained for the specimens, highlighting the utility of good descriptive work and online access to digitised representations of material in collections. Lewisuchus admixtus and Pseudolagosuchus major were scored together as a single OTU, following Nesbitt et al. (2010), Nesbitt (2011), Kammerer et al. (2012), Martinez et al. (2013), Peecook et al. (2013), and Baron et al. (2017a).
Additional phylogenetic analyses were carried out using other datasets of early dinosaurs and other archosaurs to further test the possible phylogenetic position of this taxon; Caseosaurus crosbyensis was added to the dataset of Cabreira et al. (2016), as well as to those of Ezcurra (2010), Nesbitt (2011), and Pol et al. (2011)
When using the dataset of Cabreira et al. (2016), characters 3, 4, 6, 11, 36, 60, 62, 64, 83, 115, 123, 139, 147, 148, 157, 160, 171, 173, 175, 178, 179, 182, 195, 200, 201, 202, 205, 216, 222, 240, 248 were treated as ordered. Unlike in the analyses that used the dataset of Baron et al. (2017a), the dataset of Cabreira et al. (2016) contains both Lewisuchus admixtus and Pseudolagosuchus major as separate OTUs.
When using the dataset of Ezcurra (2010), which is part based on the earlier dataset of Yates (2007), with the modifications of Smith and Pol (2007) and of Martinez et al. (2013), all multistate character were treated as unordered.
When using the dataset of Pol et al. (2011), characters 16, 18, 46, 56, 65, 97, 113, 117, 118, 125, 127, 132, 133, 135, 136, 138, 143, 147, 148, 165, 170, 174, 187, 196, 198, 210, 242, 253, and 257 were treated as ordered. All phylogenetic analyses were carried out using TNT 1.5-beta (Goloboff et al. 2008) using the New Technology Search Function, with ratchet and drift set to their default values (10 iterations and 10 cycles respectively) and with 100 random additional sequences. Character scores are presented in the SOM (Supplementary Online Material available at http://app.pan.pl/app63-Baron_etal_SOM.pdf) Bremer Support values (decay indices) were also calculated using TNT 1.5-beta (Goloboff et al. 2008).
Results
The results of the phylogenetic analyses that used the new early dinosaur dataset presented by Baron et al. (2017a) place both the holotype of Caseosaurus crosbyensis (UMMP 8870) and the Snyder Quarry specimen (NMMNH P-35995) within a clade of herrerasaurs. With the holotype of Caseosaurus crosbyensis and NMMNH P-35995 included in the dataset of Baron et al. (2017a), 85 most parsimonious trees (MPTs) were produced, each of length 1764 steps. A strict consensus of these MPTs recovers the holotype of Caseosaurus crosbyensis and NMMNH P-35995 in a polytomy with all currently recognised members of the clade Herrerasauridae (i.e., Herrerasaurus ischigualastensis, Chindesaurus bryansmalli, Sanjuansaurus gordilloi, and Staurikosaurus pricei) as well as the enigmatic taxon Saltopus elginensis. As the interrelationships within this clade cannot be yet be resolved, we must refer to this group of animals as Herrerasauria, as defined by Langer (2004) and Langer et al. (2010), i.e., all dinosaurs that share a more recent common ancestor with Herrerasaurus than with Liliensternus and Plateosaurus (see Discussion). This clade is then recovered as the sister-taxon to Dinosauria, as redefined by Baron et al. (2017a; Fig. 4). This would make Caseosaurus crosbyensis and NMMNH P-35995, as well as all other herrerasaurs, non-dinosaurian dinosauriforms. Silesauridae is recovered as the sister-taxon to the clade containing dinosaurs and herrerasaurs.
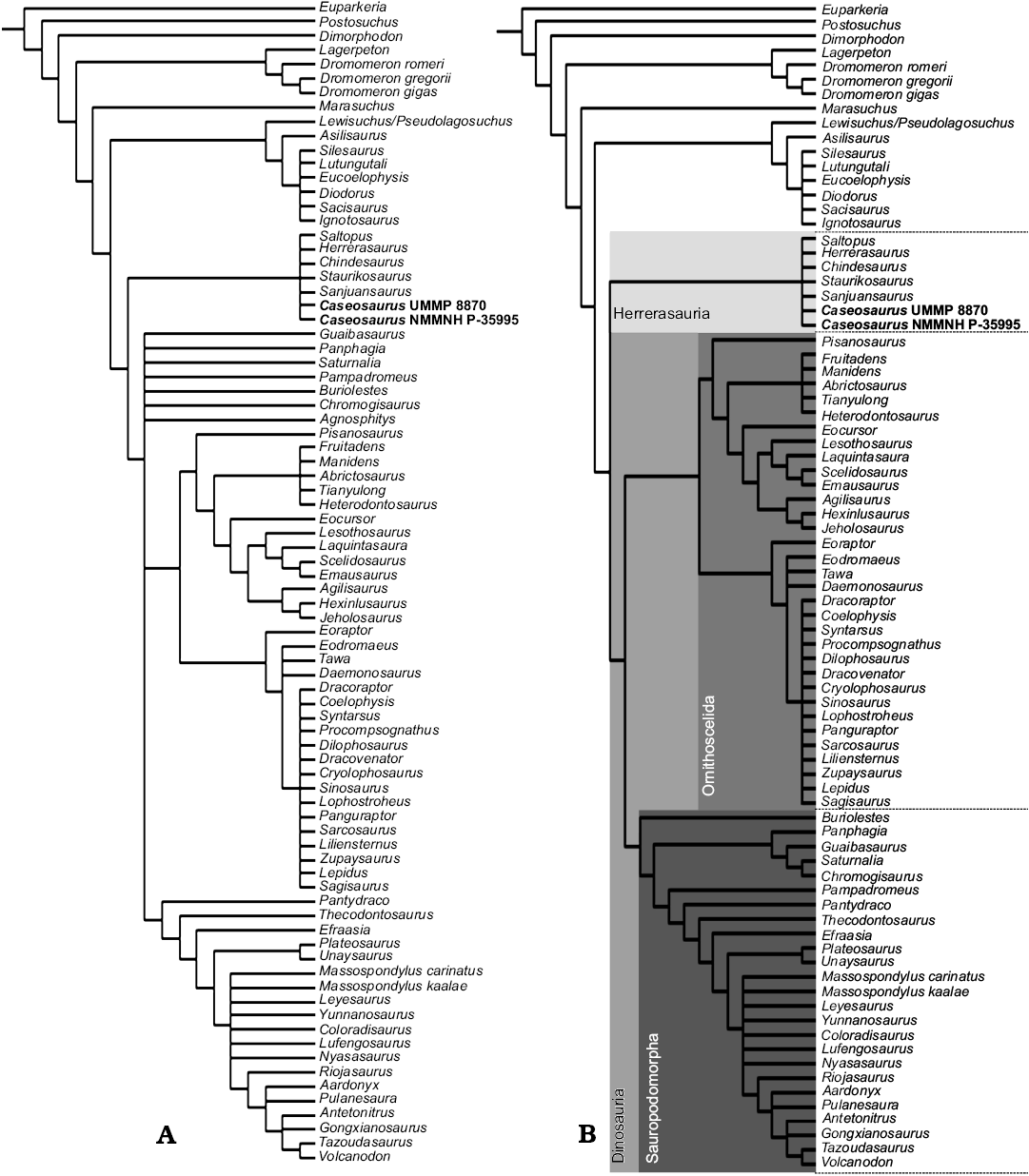
Fig. 4. A. Strict consensus tree produced from 85 MPTs, each of length 1764 steps. B. Reduced strict consensus tree produced from 91 MPTs, each of length 1764, following the removal of the possible chimera Agnosphitys cromhallenesis. For Bremer support values calculated in this analysis for each of the major nodes, see Table 1. Studied specimens in bold.
Within Dinosauria, the clades Ornithoscelida and Sauropodomorpha are recovered in a polytomy with Agnosphitys cromhallensis, Buriolestes schultzi, Chromogisaurus novasi, Guaibasaurus candelariensis, Pampadromaeus barberenai, Panphagia protos, and Saturnalia tupiniquim. Following the removal of Agnosphitys cromhallensis as an unstable or “wild-card” taxon, all of the aforementioned taxa are recovered within Sauropodomorpha in the reduced strict consensus; Sauropodomorpha and Ornithoscelida become the only constituent taxa of Dinosauria. As was also recovered in the analysis of Cabreira et al. (2016), Buriolestes schultzi is found to be the earliest diverging member of Sauropodomorpha in the reduced strict consensus tree. Sister to Buriolestes schultzi is a clade containing all other sampled sauropodomorphs and this clade is sub-divided into Guaibasauridae and a clade containing Plateosauria, Pampadromaeus barberenai, Pantydraco caducus, Thecodontosaurus antiques, and Efraasia minor (Fig. 4).
In both the strict consensus tree (Fig. 4A) and the reduced strict consensus tree (Fig. 4B), Theropoda forms the sister-taxon to Ornithischia, as part of the monophyletic Ornithoscelida, as was first recovered by Baron et al. (2017a). Theropoda contains a clade of neotheropods in a large polytomy, which includes the newly added taxa Lepidus praecisio and Segisaurus halli. This clade (Neotheropoda) is in a polytomy with Daemonosaurus chauliodus, Eodromaeus murphi, and Tawa hallae. Immediately outside of this polytomy is Eoraptor lunensis, which is recovered as the earliest diverging member of Theropoda, as in Baron et al. (2017a). In both consensus trees, Ornithischia is composed of Heterodontosauridae, Genasauria, Eocursor parvus, and Pisanosaurus mertii, the latter being recovered as the basal-most member of the clade. The reduced strict consensus tree was produced from 91 MPTs, each of length 1764 steps.
With Caseosaurus crosbyensis and NMMNH P-35995 added to the dataset of Cabreira et al. (2016), 88 MPTs each of length 848 steps were produced. In the strict consensus tree, both Caseosaurus crosbyensis and NMMNH P-35995 were recovered in a large polytomy with Herrerasaurus ischigualastensis, Staurikosaurus pricei, Sanjuansaurus gordilloi, Daemonosaurus chauliodus, Tawa hallae, Eodromaeus murphi, Lewisuchus admixtus, Pseudolagosuchus major, and Saltopus elginensis, a clade containing both Ornithischia and Silesauridae, and a clade containing all other dinosaurs (see SOM). By combining the holotype of Caseosaurus crosbyensis with NMMNH P-35995 and rerunning the analysis, 71 MPTs each of length 848 were produced, with marginally better resolution; the instability of NMMNH P-35995 being attributable to the low number of characters that it could scored for in the Cabreira et al. (2016) dataset (9/257). In the reduced strict consensus tree, Lewisuchus admixtus, Pseudolagosuchus major, or Saltopus elginensis form a polytomy with the clade containing both Silesauridae and Dinosauria, and Caseosaurus crosbyensis is recovered in a polytomy with Herrerasaurus ischigualastensis, Staurikosaurus pricei, Sanjuansaurus gordilloi, Daemonosaurus chauliodus, and Tawa hallae at the base of the dinosaurian lineage. In this analysis, Eodromaeus murphi forms the sister-taxon to the clade containing Guaibasaurus candelariensis and all more derived dinosaurs (Fig. 5). By looking at the various MPTs that were generated in this analysis, it can be observed that, based upon the character data utilised in the Cabreira et al. (2016) dataset, Caseosaurus crosbyensis can be recovered as either a herrerasaur; as a taxon outside of Eusaurischia, close to Tawa and Daemonosaurus; or as sister-taxon to all other non-eusaurischian saurischians, inducing the herrerasaur clade.

Fig. 5. Reduced strict consensus tree produced when Caseosaurus crosbyensis (in bold) was added to the dataset of Cabreira et al. (2016).
With Caseosaurus crosbyensis and NMMNH P-35995 included in the latest version of the dataset of Ezcurra (2010), 78 MPTs each of length 1215 steps were produced. A strict consensus tree produced from these MPTs recovers Caseosaurus crosbyensis and NMMNH P-35995 in a group with Herrerasaurus ischigualastensis, Staurikosaurus pricei, and an unnamed herrerasaurid specimen, MACN-Pv 18849a (see SOM: fig. S2). This provides more evidence for a close relationship between Caseosaurus crosbyensis (and NMMNH P-35995) and herrerasaurid dinosaurs such as Herrerasaurus ischigualastensis. Additional information on this result is presented in the SOM.
With Caseosaurus crosbyensis and NMMNH P-35995 added to the dataset of Pol et al. (2011), 17 MPTs were produced, each of length 617 steps. In the strict consensus tree generated from these MPTs, Caseosaurus crosbyensis and NMMNH P-35995 were recovered as sister-taxa. This pair in turn forms the sister group to Herrerasaurus ischigualastensis. This group of Herrerasaurus ischigualastensis, Caseosaurus crosbyensis, and NMMNH P-35995 then forms polytomy with Eoraptor lunensis and a group containing numerous theropod taxa. Additional information on this result is also presented in the SOM. This result supports that obtained by the analyses that used the dataset of Baron et al. (2017a), and previous assertion that Caseosaurus crosbyensis represents a herrerasaurid, and is closely related to the theropod dinosaurs. Bremer supports values were also calculated for each of the major clades in the analyses that used the datasets of Baron et al. (2017a) and Cabreira et al. (2016), and are summarised in Table 1. Bremer supports were calculated with and without the inclusion of the fragmentary taxa Saltopus elginensis and Agnosphitys cromhallensis, following Baron et al. (2017a).
Table 1. Bremmer support values calculated in the analyses that used the dataset of Baron et al. (2017a): 1, all taxa included; 2, reduced analysis, Agnosphitys cromhallensis removed as a “wild-card” taxon; 3, reduced analysis, Agnosphitys cromhallensis and Saltopus elginensis removed, following Baron et al. (2017a).
|
Clade |
Analysis |
||
|
1 |
2 |
3 |
|
|
Silesauridae+Dinosauria+Herrerasauria |
>5 |
>5 |
2 |
|
Silesauridae |
>5 |
>5 |
>5 |
|
Dinosaura+Herrerasauria |
>5 |
>5 |
>5 |
|
Dinosauria |
2 |
2 |
1 |
|
Herrerasauria |
2 |
2 |
2 |
|
Sauropodomorpha |
– |
>5 |
>5 |
|
Guaibasauridae |
– |
2 |
1 |
|
Ornithoscelida |
>5 |
>5 |
>5 |
|
Ornithischia |
1 |
1 |
>5 |
|
Theropoda |
1 |
1 |
2 |
Systematic palaeontology
Archosauria Cope, 1869
Dinosauromorpha Benton, 1984
Dinosauriformes Novas, 1992
Herrerasauria Galton, 1985 (sensu Langer et al. 2010)
Genus Caseosaurus Hunt, Lucas, Heckert, Sullivan, and Lockley, 1998
Type species: Caseosaurus crosbyensis Hunt, Lucas, Heckert, Sullivan, and Lockley, 1998, Carnian, Late Triassic, Tecovas Formation, Crosby County, Texas, USA.
Caseosaurus crosbyensis Hunt, Lucas, Heckert, Sullivan, and Lockley, 1998
Figs. 1, 2.
Holotype: UMMP 8870, a partial right ilium.
Type horizon: ?late Carnian, Late Triassic (Nesbitt et al. 2007).
Type locality: Tecovas Formation, Dockum Group, Crosby County, Texas, USA; Snyder Quarry, Petrified Forest Member, Chinle Formation, Rio Arriba County, New Mexico, USA.
Material.—NMMNH P-35995, a partial right ilium.
Emended diagnosis.—Caseosaurus crosbyensis represents a valid taxon and a most likely represents a member of the clade Herrerasauria. Caseosaurus crosbyensis is unique among herrerasaurids and dinosaurs in possessing a sharp, well developed ridge that connects the midpoint of the supraacetabular crest to the preacetabular process. Furthermore, Caseosaurus crosbyensis can be referred to Herrerasauria based upon the possession of three of the clade’s synapomorphies: a postacetabular process that is 25–35% of the total length of the length of the iliac blade and a supraacetabular crest that extends down part of pubic peduncle as a ridge without reaching the distal end*; possession of extensive rugosities on the pre- and postacetabular processes* (also present in a similar form in silesaurids, and in a slightly different form in Saturnalia tupiniquim and Coelophysis bauri). In addition to these features, Caseosaurus crosbyensis possesses a number of other features that can be observed in all other herrerasaurs,including: a preacetabular process of the ilium that expands mediolaterally towards its distal end* (also in silesaurids); absence of a brevis fossa. Furthermore, Caseosaurus crosbyensis and NMMNH P-35995 possess an ischiadic peduncle that is less ventrally extensive than the pubic peduncle in medial and lateral aspect, a condition that is also present in Herrerasaurus ischigualastensis and Staurikosaurus pricei. Caseosaurus crosbyensis can be distinguished from Herrerasaurus ischigualastensis and Staurikosaurus pricei by possessing a preacetabular process that is more than twice as long as it is deep*; having a sharp, distinct anterodorsal ridge that runs from the middle of the supraacetabular crest to the preacetabular process*; and the absence of an acetabular antitrochanter* (which also distinguishes Caseosaurus crosbyensis from Chindesaurus bryansmalli). Caseosaurus crosbyensis can be further distinguished from Staurikosaurus pricei as it possesses a preacetabular iliac process that is much shorter than the postacetabular process of the ilium (also present in Herrerasaurus ischigualastensis, Eoraptor lunensis, and Eodromaeus murphi). Further to these distinguishing features, Caseosaurus crosbyensis appears to differ from other herrerasaurs in a several other respects. For example, while Caseosaurus crosbyensis possesses rugose areas on the dorsal and lateral portions of the pre- and postacetabular processes, these areas are less extensive than they are in Herrerasaurus ischigualastensis and Staurikosaurus pricei (Fig. 6). Furthermore, the supracetabular crest of Caseosaurus crosbyensis describes a semicircle in lateral view*, differing from the condition in Herrerasaurus ischigualastensis and Staurikosaurus pricei, in which the supraacetabular crest is not semicircular but instead forms a straighter, anteroventrally oriented lip over the main body of the acetabulum (Fig. 6). For the anatomical features listed above, (*) represents those that are present in both the holotype and the referred specimen of Caseosaurus crosbyensis; the single autapomorphy of the taxon is also observed in both specimens

Fig. 6. The herrerasaurian dinosauriform Herrerasaurus ischigualastensis Reig, 1963 (PVL 2566) from Carnian, Late Triassic, Ischigualasto Formation, Hoyada de Ischigualasto, San Juan Province, Argentina; ilium in lateral view.
Description.—The holotype specimen is missing only the middle portions of the dorsal margin, as well as a small portion of the middle section of the iliac body that lies immediately above the supraacetabular rim. Further to this, judging by the shape of the preserved ventral margin of the ilium, around the acetabulum, a portion of a medioventral acetabular wall that would normally partially close the acetabulum is also missing. Towards its dorsal margin, UMMP 8870 bears rugose areas on the lateral and dorsal surfaces of the pre- and postacetabular processes. This is similar to the condition seen in Herrerasaurus ischigualastensis (PVL 2566) and Staurikosaurus pricei (Galton 1977; Bittencourt and Kellner 2009; Delcourt et al. 2012). However, these rugosities, while still quite extensive in Caseosaurus crosbyensis, appear to cover relatively smaller portions of the lateral and dorsal faces of the pre- and postacetabular processes than they do in Herrerasaurus ischigulastensis (PVL 2566), a specimen in which the rugosities form large, bulbous areas on the majority of both the lateral and dorsal surfaces of the pre- and postacetabular processes. In H. ischigualastensis, these rugosities are so pronounced that they form the dominant anatomical feature of the dorsal part of the ilium, partly obscuring other features, such as the brevis shelf (Fig. 6). Some early sauropodomorph taxa such as Saturnalia tupiniquim and Chromogisaurus novasi also possess a type of rugose area on the postacetabular process of the ilium, but in these taxa this rugose area is restricted to a small trapezoidal area near the posterior margin of the postacetabular process (Ezcurra 2010: fig. 17), rather than being developed across a broad area upon the postacetabular process, with no clear and geometric boundary. The same reduced and limited rugositites can also be observed in the ilium of Coelophysis bauri (e.g., USNM 529376). Given the differences in form and extent of the rugosities among these taxa, the potential homology of these features is difficult to determine. However, the differences in the form of this feature among these taxa is worth noting.
The preacetabular process of UMMP 8870 extends from the main body of the ilium as an anterodorsally oriented projection. The preacetabular process is mediolaterally and dorsoventrally expanded at its dorsal end, similar to the condition seen in Herrerasaurus ischigualastensis (PVL 2566) and Staurikosaurus pricei (Galton 1977; Delcourt et al. 2012). This condition is also similar to that in several silesaurid taxa, such as Silesaurus opolensis (ZPAL Ab III/361; Dzik 2003; Peecook et al. 2013) and Ignotosaurus fragilis (Martinez et al. 2013). This differs from the condition in other dinosaurian taxa. For example, in more derived sauropodomorphs, such as Efraasia minor, the preacetabular process is not expanded mediolaterally or dorsoventrally, but instead takes the form of a short, relatively flat and unexpanded triangular projection (Galton 1973; Yates 2003b; Delcourt et al. 2012); in Saturnalia tupiniquim the preacetabular process is slightly expanded mediolaterally and dorsoventrally at its dorsal point, but to a lesser degree than in herrerasaurs and silesaurids. In early ornithischians the preacetabular process is elongate and strap-like and lacks the expansion at its distal end, for example the preacetabular process of Lesothosasrus diagnosticus (NHMUK PV RU B17; Baron et al. 2017b). In theropod taxa, such as Coelophysis bauri, the preacetabular process is anteriorly oriented, dorsoventrally deep and square in lateral view, but relatively unexpanded mediolaterally at its dorsal end (AMNH 7223, AMNH 7224, USNM 529376). The lateral face of the preacetabular process in Caseosaurus crosbyensis also bears an oval-shaped rugose area, similar to that seen in some silesaurids, such as Silesaurus opolensis (ZPAL Ab III/361; Dzik 2003) and Lutungutali sitwensis (Peecook et al. 2013); this rugose area does not extend far onto the dorsal surface in Caseosaurus crosbyensis and silesaurids, contrasting with the condition in Herrerasaurus ischigualastensis (PVL 2566), in which the rugose area on the preacetabular process is more extensive and covers a larger portion of the dorsal surface. The condition in Staurikosaurus pricei appears to be somewhere between the conditions seen in Caseosaurus crosbyensis and Herrerasaurus ischigualastensis, in that in Staurikosaurus pricei the rugose area appears to extend some of the way on to the dorsal portion of the preacetabular process, but not as far as the same rugose area does in Herrerasaurus ischigualastensis (Galton 1977; Delcourt et al. 2012). In Coelophysis bauri a small rugose area is present but is restricted to the dorsal surface only, and the surface of this area is far smoother than the areas in silesaurids, herrerasaurids and Caseosaurus crosbyensis.
The postacetabular process of the holotype of Caseosaurus crosbyensis also bears rugosities on its lateral and dorsal surfaces (Figs. 1, 2, 6). These rugose areas in Caseosaurus crosbyensis are similar to those that are seen in Herrerasaurus ischigualastensis (PVL 2566) but are less extensive in the former (Fig. 6). Similar rugose areas also appear in some silesaurids (Fig. 7); Silesaurus opolensis (ZPAL Ab III/361; Dzik 2003), Lutungutali sitwensis (Peecook et al. 2013) and Ignotosaurus fragilis (Martinez et al. 2013) all have rugose areas on the dorsal and lateral surfaces of postacetabular process. Again, Staurikosaurus pricei has a condition that appears to be somewhere between Caseosaurus crosbyensis and Herrerasaurus ischigualastensis in terms of overall extent (Delcourt et al. 2012). Such laterally located rugose areas are not seen in early sauropodomorphs (e.g., Panphagia; PVSJ 874), theropods (e.g., Coelophysis bauri, AMNH 7223, AMNH 7224; Cope 1887; Colbert 1989) or ornithischians (e.g., Lesothosaurus diagnosticus, NHMUK PV RU B17; Barrett et al. 2014); some dorsally extensive rugosities can be observed in Saturnalia tupiniquim (e.g., MCP 3844-PV) and Coelophysis bauri (USNM 529376). Ventrally, the postacetabular process of the holotype of Caseosaurus crosbyensis bears a small excavation or fossa, and this feature was considered to be the brevis fossa by Hunt et al. (1998). However, Nesbitt et al. (2007) did not consider this fossa to be homologous to the brevis fossa present in a number of other dinosaurian and silesaurid taxa; these authors argued that because the fossa present in the holotype of Caseosaurus crosbyensis is not associated with a distinct ridge it does not meet the criteria given in their definition of a true brevis fossa, nor the definition given by Novas (1992). This study agrees with the observations of Nesbitt et al. (2007) and does not consider the fossa in Caseosaurus crosbyensis to represent a brevis fossa. In our phylogenetic analyses, we do not score Caseosaurus crosbyensis as possessing a brevis fossa that is homologous to those seen in a range of other dinosauriform taxa. The fossa in question is shallow and can be seen in lateral view and lies directly ventral to a slight, rounded, anteroposteriorly extending swelling on the lateral face of the postacetabular process (Fig. 1). The rugose areas are located dorsal to this swelling in Caseosaurus crosbyensis (Fig. 1), which distinguishes them from those present on the postacetabular processes of Saturnalia tupiniquim and Chromogisaurus novasi, which are ventrally located scars associated with muscle origins (Langer 2003; Ezcurra 2010). The shape of the distal end of the postacetabular process of Caseosaurus crosbyensis is also different from that in sauropodomorph taxa like Saturnalia tupiniquim and Chromogisaurus novasi. In Caseosaurus crosbyensis the postacetabular process is rounded to square in lateral and medial views, with slightly greater posterior extension occurring ventrally than dorsally, whereas in Saturnalia tupiniquim, Chromogisaurus novasi, and Panphagia protos (PVSJ 874), the postacetabular process is more trapezoidal (Langer 2003; Ezcurra 2010). In herrerasaurids, such as Herrerasaurus ischigualastensis (PVL 2566) and Staurikosaurus pricei (Galton 1977; Bittencourt and Kellner 2009; Delcourt et al. 2012), the postacetabular process is also rounded to square in lateral and medial views, which is more similar to the condition in Caseosaurus crosbyensis. In Tawa hallae (GR 155; Nesbitt et al. 2009b) the postacetabular process is also rounded to square, but with even greater posterior extension occurring ventrally; in Coelophysis bauri the postacetabular is squared posteriorly. Also in Tawa hallae (GR 155; Nesbitt et al. 2009b), the lateral face of the postacetabular process is dominated by a distinct anteroposteriorly oriented ridge. In other theropods, such as Coelophyis bauri (AMNH 7223, AMNH 7224; USNM 529376; Cope 1887; Colbert 1989; Fig. 8), the greater posterior expansion occurs dorsally rather than ventrally, a reverse of the condition in seen in Caseosaurus crosbyensis, herrerasaurids and Tawa hallae; in all other respects the morphology of the postacetabular process in early theropods is similarly round to square-ended. On its medial side, the postacetabular process bears a large longitudinal ridge in Caseosaurus crosbyensis, as in most archosaurs (Nesbitt 2011; Fig. 1).
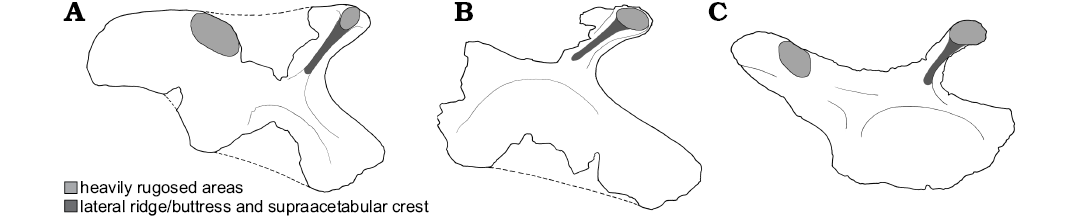
Fig. 7. Comparison of the ilia of Caseosaurus crosbyensis Hunt, Lucas, Heckert, Sullivan, and Lockley, 1998 (A, B) and Silesaurus opolensis Dzik, 2003 (C) showing locations of rugosities and lateral ridge/buttress. A. UMMP 8870, right ilium in lateral view. B. NMMNH P-35995, right ilium in lateral view. C. ZPAL AbIII404/1, left ilium in lateral view (reversed).
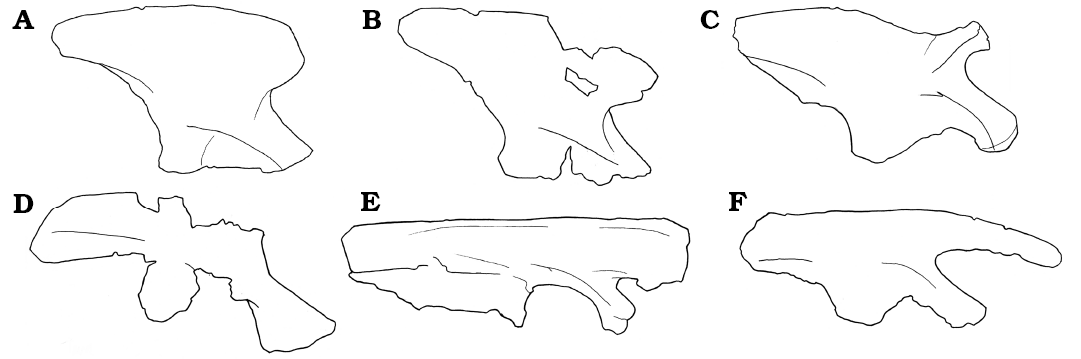
Fig. 8. Comparison of the ilia of early dinosaurs. A. Buriolestes schultzi. B. Saturnalia tupiniquim. C. Agnosphitys cromhallensis. D. Tawa hallae. E. Coelophysis bauri. F. Lesothosaurus diagnosticus.
The supraacetabular crest appears to projects laterally from the iliac body in Caseosaurus crosbyensis, although it is broken laterally. In lateral aspect the crest forms a smooth semicircle before continuing as a faint ridge along the ventral portion of the pubic peduncle, terminating dorsal to the ventral most point of the peduncle; in most theropod dinosaurs, such as Tawa hallae and Coelophysis bauri, this crest reaches the ventral most point of the peduncle. The semicircular shape of the supraacetabular crest contrasts with the condition seen in Herrerasaurus ischigualastensis and early sauropodomorphs, such as Saturnalia tupiniquim and Chromogisaurus novasi, in which the supraacetabular crest follows a less curved line along the lateral surface of the ilium and slopes more anteroventrally towards the pubic peduncle (Langer 2003; Ezcurra 2010). The condition in another herrarasaurid taxon, Staurikosaurus pricei, appears more similar to that of Caseosaurus crosbyesnsis, i.e., a smoother semicircular crest (see Delcourt et al. 2012). Outside of Herrerasauria the shape of the supraacetabular crest has a complex distribution. While some silesaurids possess a smooth semicircular crest, such as Silesaurus opolensis (ZPAL AbIII404/1; Peecook et al. 2013) and Asilisaurus kongwe (Peecook et al. 2013), others do not, as for example Lutungutali sitwensis has a straighter, sloped supraacetabular crest (Peecook et al. 2013). Extending anterodorsally from the midpoint of the supraacetabular crest in Caseosaurus crosbyensis is a distinct, sharp crest, which extends up to the anterodorsal, bulb-like tip of the preacetabular process (Figs. 1, 2). This sharp crest, or ridge, is delineated on both sides by concave surfaces and distally it merges into the oval-shaped rugose area of the preacetabular process. This is like the condition seen in many silesaurids, such as Silesaurus opolensis, Asilisaurus kongwe, Sacisaurus agudoensis, Lutungutali sitwensis, and Ignotosaurus fragilis (Dzik 2003; Ferigolo and Langer 2006; Nesbitt et al. 2010; Martinez et al. 2013; Peecook et al. 2013; Fig. 7). In Herrerasaurus ischigualastensis and Staurikosaurus pricei there is a lower, more rounded and less distinct anterodorsally oriented swelling that extends from the same point on the supraacetabular crest as the distinct, sharp ridge in Caseosaurus crosbyensis does. In these herrerasaurid taxa, there is less of a distinction between this low crest and the rest of the lateral surface of the ilium. Similar, low crests also appear in some early sauropodomorphs, such as Saturnalia tupiniquim (Langer 2003), in ornithischians, such as Lesothosaurus diagnosticus (NHMUK PV RU B17), and in theropods, such as Coelophysis bauri (AMNH 7223, AMNH7224, USNM 529376; Cope 1887; Colbert 1989). Nesbitt (2011) reported the presence of a sharp and narrow ridge in the Triassic sauropodomorph Saturnalia tupiniquim and scored this taxon for having a distinct ridge that could be considered as homologous to the ridges that are see in silesaurids. However, while a small ridge is certainly present between the supraacetabular crest and the preacetabular process in Saturnalia, this ridge does not extend from the dorsal margin of the supraacetabular crest, does not extend fully onto the lateral surface of the preacetabular process and is not bordered posterodorsally and anteroventrally by smooth concave surfaces (Langer 2003), but instead takes the form of a small, isolated, almost vertically oriented tuberosity in Saturnalia (Langer 2003). In this regard, the ridge in Saturnalia differs considerably from the ridges seen in the silesaurid taxa). We therefore do not consider this ridge in Saturnalia to be homologous to those seen in Caseosaurus crosbyensis, NMMNH P-35995, and silesaurids. As discussed above, Saturnalia appears to possess a low and indistinct anterodorsally oriented crest that does connect the supraacetabular crest to the preacetabular process, in addition to the small tuberosity described as a ridge by Nesbitt (2011). We consider the former to be homologous to the crests seen in other dinosaurs and silesaurids, and our scoring for this character (character 305; Baron et al. 2017a) reflects this.
Ventral to the supraacetabular crest, a medioventral acetabular wall extends ventrally and, while incomplete, this feature appears to be well developed and ventrally extensive in Caseosaurus crosbyensis, judging from the extent of the fragmentary remains that appear on the posterior margin of the pubic peduncle (Fig. 1). All silesaurids have extensive acetabular walls that fully close the acetabulum. Similarly, extensive acetabular walls exist in a number of early sauropodomorphs, such as Saturnalia tupiniquim and Chromogisaurus novasi (Langer 2003; Ezcurra 2010). However, in these early sauropodomorph taxa, the wall is extensive but does not fully close the acetabulum, as it does in silesaurids. The herrerasaurid Herrerasaurus ischigualastensis also has a medioventral acetabular wall (PVL 2566), but this feature is less ventrally extensive than those in silesaurids and early sauropodomorphs. In Herrerasaurus ischigualastensis the acetabulum also appears to be perforate. The exact condition in Staurikosaurus pricei is hard to assess, as the acetabular wall is broken. However, enough remains of the acetabular wall on the posterior margin of the pubic peduncle in Staurikosaurus to see that the wall is ventrally extensive. Based on what remains of the wall in Caseosaurus crosbyensis it is possible to say that a wall is present and probably quite ventrally extensive as well, but whether or not the acetabulum is fully open, partially open or imperforate is difficult to say, as in Staurikosaurus. It seems likely that the two taxa shared morphology similar to that of Herrerasaurus ischigualastensis. Nesbitt et al. (2007) regarded the acetabulum of Caseosaurus crosbyensis as being at least partially perforate, which they used as evidence when arguing that the taxon could be assigned to Dinosauriformes. We agree that, based upon the preserved material of Caseosaurus crosbyensis, a perforate acetabulum seems to be the likely morphology present in this taxon; we score this taxon for the presence of a perforate acetabulum in each of the phylogenetic analyses that we carried out.
The pubic peduncle in Caseosaurus crosbyensis is a long, anteroventrally oriented projection from the main body of the ilium. For its entire length, the posterior margin of the peduncle includes the remnants of the medioventral acetabular wall. Anteroventrally the pubic peduncle flares out anteroposteriorly, as is also the condition in some dinosaurs, including Tawa hallae, and in herrerasaurs, but not in theropods such as Coelophysis bauri (USNM 529376). In Caseosaurus crosbyensis the level of anteroposterior expansion of the anteroventral end is proportionally greater than it is in Herrerasaurus ischigualastensis (PVL 2566) and Staurikosaurus pricei (Galton 1977; Delcourt et al. 2012), as well as in silesaurids (Dzik 2003; Peecook et al. 2013), but less than it is in the early theropod Tawa hallae (GR 155; Nesbitt et al. 2009b). The ischiadic peduncle in Caseosaurus crosbyensis also includes part of the acetabular wall. The ischadic peduncle is smaller than the pubic peduncle and is almost indistinct from the acetabular wall that lies anterior to it. This is very similar to the condition in herrerasaurids, such as Herrerasaurus ischigualastensis (PVL 2566) and Staurikosaurus pricei (Galton 1977; Delcourt et al. 2012), and in silesaurids such as Ignotosaurus fragilis (Martinez et al. 2013) and Silesaurus opolensis (Dzik, 2003) (Fig. 7). In other dinosaurs, such as the theropods Coelophysis bauri (AMNH 7223, AMNH 7224; Cope 1887; Colbert 1989) and Tawa hallae (GR 155; Nesbitt et al. 2009b), and some sauropodomorphs, such as Efraasia minor (Galton 1973; Yates 2003b), the ischiadic peduncle is quite distinct from the medioventral acetabular wall. In other dinosaurian taxa, such as the early ornithischian Lesothosaurus diagnosticus (NHMUK PV RU B17; Baron et al. 2017b) and other sauropodomorphs like Saturnalia tupiniquim (Langer 2003), the ischiadic peduncle and medioventral acetabular wall do form a continuous surface but the peduncle appears a little more distinct than it does in Herrerasaurus ischigualastensis and Caseosaurus crosbyensis.
NMMNH P-35995 from the Snyder Quarry is less complete than the holotype of Caseosaurus crosbyensis (UMMP 8870); it is missing a greater proportion of its dorsal margin and almost all of its posterior section, including all of the postacetabular process (Fig. 3). However, in all of the areas that are preserved, NMMNH P-35995 bears a striking resemblance to UMMP 8870. Nesbitt et al. (2007) discussed the similarities between NMMNH P-35995 and the holotype of Caseosaurus crosbyensis in some detail, although they did not refer NMMNH P-35995 to Caseosaurus crosbyensis. The similarities that they noted were “a short, pointed anterior process of the ilium, a strong ridge trending anterodorsally from the acetabular rim to the anterior preacetabular process, a wide, open angle between the anterior process and the pubic process, a moderately developed supraacetabular rim and an ischiadic process with a rounded distal end that is dorsal to the distal end of the pubic process” (Nesbitt et al. 2007: 217). Further to this Nesbitt et al. (2007) stated that none of these features are present in the theropod taxa Coelophysis bauri (AMNH 7223), Coelophysis rhodesiensis (Raath, 1969), “Syntarsus” kayentakatae (Rowe, 1989), and Dilophosaurus wetherilli (Welles, 1954). Overall, we agree with this anatomical assessment by Nesbitt et al. (2007). We would also add that, of these distinguishing characters, a short, pointed anterior process (preacetabular process), a strong ridge trending anterodorsally from the acetabular rim to the anterior preacetabular process and a wide, open angle between the anterior process and the pubic process are also absent in Herrerasaurus ischigualastensis (PVL 2566). However, an ischiadic process with a rounded distal end that is dorsal to the ventral extent of the pubic process is present in Herrerasaurus ischigualastensis (PVL 2566) and Staurikosaurus pricei (Colbert 1970; Galton 1977; Delcourt et al. 2012) and that a wide, open angle between the anterior process and the pubic process, as described in UMMP 8870 and NMMNH P-35995, is also present in Staurikosaurus pricei (Colbert 1970; Galton 1977; Delcourt et al. 2012). Generally, we also note that the shared features of UMMP 8870 and NMMNH P-35995, as described by Nesbitt et al. (2007), are not present in ornithischian dinosaurs, such as Lesothosaurus diagnosticus (NHMUK RU B17) and Heterodontosaurus tucki (SAM-PK-K1332), in which the preacetabular process is long and “strap-like” and the ischiadic process (= peduncle) is not as reduced. Furthermore, Lesothosaurus diagnosticus and Heterodontosaurus tucki do not possess the wide, open angle between the anterior process and the pubic process that is seen in UMMP 8870, NMMNH P-35995, and Staurikosaurus pricei (Galton 1977; Delcourt et al. 2012), nor do theropods such as Coelophysis bauri (e.g., USNM 529376). Adding further comparisons to those provided by Nesbitt et al. (2007), we note that there is no strong, sharp ridge trending anterodorsally from the acetabular rim to the anterior preacetabular process or a reduction on the ischiadic process in some basal sauropodomorphs, for example Pantydrao caducus (Yates 2003a) and Efraasia minor (Galton 1973; Yates 2003b; Delcourt et al. 2012). In other sauropodomorph taxa, such as Saturnalia tupiniquim and Chromogisaurus novasi, there is a slight reduction in the ischiadic peduncle (Langer 2003; Ezcurra 2010), but the difference in the ventral extensions of the pubic and ischiadic peduncles is less than it is in UMMP 8870 and NMMNH P-35995, as well as in the herrerasaurids Herrerasaurus ischigualastensis (PVL 2566) and Staurikosaurus pricei (Galton 1977; Delcourt et al. 2012). However, we note that in Saturnalia tupiniquim (Langer 2003) and Chromogisaurus novasi (Ezcurra 2010) the pubic peduncle is not as long as it is in UMMP 8870, NMMNH P-35995, Herrerasaurus ischigualastensis (PVL 2566) and Staurikosaurus pricei (Galton 1977; Delcourt et al. 2012), which may explain why the reduction of the ischiadic peduncle in Saturnalia tupiniquim and Chromogisaurus novasi does not appear to be as great as it is in UMMP 8870, NMMNH P-35995, and herrerasaurids. We also note that, for the feature described by Nesbitt et al. (2007) as a strong ridge trending anterodorsally from the acetabular rim to the anterior preacetabular process, it is the form of this feature, rather than its absence or presence, that is the main discriminating factor between UMMP 8870/NMMNH P-35995 and other dinosaurs; there is a crest trending anterodorsally from the acetabular rim to the preacetabular process in many dinosaurian taxa, but it is only in UMMP 8870 and NMMNH P-35995 (and some silesaurids: see above) that this crest appears as a sharp, distinct ridge. In addition to the features that were discussed by Nesbitt et al. (2007), we also make the following observations about the anatomy of the specimen NMMNH P-35995.
NMMNH P-35995, much like UMMP 8870, preserves a complete preacetabular process that takes the form of a thin, anterodorsally extending process that swells dorsally, both mediolaterally and dorsoventrally, into a rounded end. The lateral surface of this process bears a distinct rugose area that also extends partly on to the dorsal surface. The rugose area on the lateral surface of the preacetabular process is connected to the middle of supraacetabular crest by a sharp, distinct ridge. The supraacetabular crest is partly broken in NMMNH P-35995 but is more complete than in UMMP 8870. In lateral view the supraacetabular crest in NMMNH P-35995 takes the form of a smooth semicircle above the acetabulum, like in Tawa hallae and Coelophysis bauri (USNM 529376), but only extends some of the way along the length of the pubic peduncle, as in UMMP 8870, but unlike in Coelophysis bauri. Also like UMMP 8870, NMMNH P-35995 shows evidence of a medioventral acetabular wall that would at least partially close the acetabulum. This wall appears to be extensive in NMMNH P-35995, but it is hard to state with confidence whether or not the acetabulum was completely imperforate. The pubic peduncle in NMMNH P-35995 is longer than the ischiadic peduncle, which is reduced and rounded distally. Unfortunately, the postacetabular process (and much of the dorsal iliac blade) is missing in NMMNH P-35995, so the presence, and possible extent, of rugose areas on the postacetabular process cannot be assessed in this specimen.
Despite the many similarities between the specimens, UMMP 8870 and NMMNH P-35995 can also be distinguished in several minor features. Firstly, the acetabulum of NMMNH P-35995 is deeper relative to the height of the iliac blade above the acetabulum than it is in the holotype of Caseosaurus crosbyensis (based upon the dorsal extent of the preacetabular process). This difference in relative dorsoventral depth of the ilia above and below the acetabulum could be related to size. NMMNH P-35995 is smaller than UMMP 8870. UMMP 8870 has a maximum dorsoventral depth of 86.9 mm whereas NMMNH P-35995 has a maximum depth of 57.8 mm. A second difference between UMMP 8870 and NMMNH P-35995 is in the angle between the preacetabular process and the pubic peduncle. In UMMP 8870 this area takes the form of a rounded, wide gap in which the preacetabular process and the pubic peduncle diverge at an obtuse angle. In NMMNH P-35995 this gap is smaller and the angle between the preacetabular process and the pubic peduncle is acute. Despite this subtle difference, Nesbitt et al. (2007) still considered a wide, open angle between the anterior process and the pubic process to be a feature uniting NMMNH P-35995 and UMMP 8870. An acute angle between the preacetabular process and the pubic peduncle is observed in basal sauropodomorphs, such as Saturnalia tupiniquim and Chromogisaurus novasi (Langer 2003; Ezcurra 2010; Delcourt et al. 2012), in ornithischians, such as Heterodontosaurus tucki (SAM-PK-K1332) and larger specimens of Lesothosaurus diagnosticus (e.g., NHMUK PV R11000; Baron et al. 2017b), in theropods, such as Coelophysis bauri (AMNH 7223, AMNH 7224; Cope 1887; Colbert 1989) and in Herrerasaurus ischigualastensis (PVL 2566). Conversely, Staurikosaurus pricei has an obtuse angle between the preacetabular process and the pubic peduncle (Galton 1977; Delcourt et al. 2012), as do the smaller specimens of the ornithischian Lesothosaurus diagnosticus (NHMUK PV RU B17; BP/1/4731; Baron et al. 2017b). Silesaurids also exhibit something of an open angle between the preacetabular process and the pubic peduncle, with most taxa having an angle close to 90° or slightly acute (e.g., Silesaurus opolensis; Dzik 2003). Currently, whether or not this feature has any taxonomic significance remains to be seen, and none of the datasets used in this study account for this feature through the inclusion of an anatomical character. The intraspecific variation that is seen in Lesothosaurus diagnosticus might suggest that this feature is size related (see Baron et al. 2017b), and this may explain the observed difference between NMMNH P-35995 and the holotype of Caseosaurus crosbyensis.
Remarks.—NMMNH P-35995 is referable to Caseosaurus crosbyensis on the basis of a shared autapomorphy, the possession of a sharp, well developed ridge connecting the midpoint of the supraacetabular crest to the preacetabular process, as well as a number of features that are unique to UMMP 8870 and NMMNH P-35995 among members of Herrerasauria: the possession of a preacetabular process that is more than twice as long as it is deep (for information on how this measurement was taken, see SOM); possession of a sharp and distinct anterodorsal ridge that runs from the middle of the supraacetabular crest to the preacetabular process of the ilium (also present in members of Silesauridae, but absent in herrerasaurs, and dinosaurs like Tawa hallae and Eodromaeus murphi); a supraacetabular crest that forms a semicircle in lateral view, rather than a straighter, anteroventrally oriented lip.
Stratigraphic and geographic range.—Tecovas Formation, Dockum Group, Crosby County, Texas, USA; Snyder Quarry, Petrified Forest Member, Chinle Formation, Rio Arriba County, New Mexico, USA.
Discussion
The results of the phylogenetic analyses carried out in this study recover Caseosaurus crosbyensis (both UMMP 8870 and NMMNH P-35995) in a close relationship with all currently recognised herrerasaurid taxa (Herrerasaurus ischigualastensis, Chindesaurus bryansmalli, Sanjuansaurus gordilloi, and Saturikosaurus pricei). These results provide good evidence in support of the hypothesis that Caseosaurus crosbyensis represents a herrerasaur, as proposed by Long and Murry (1995) and supported by Hunt et al. (1998). In addition to Caseosaurus crosbyensis, the enigmatic taxon Saltopus elginensis is also recovered as a herrerasaur. While this taxon has often been considered to fall close to the origins of dinosaurs (e.g., Benton and Walker 2011), it has, as far as we are aware, never been recovered in such a position by any previous large-scale phylogenetic analysis of early dinosaurs (or dinosauromorphs). The position of Saltopus elginensis is supported by the possession of a scapula blade that is only weakly expanded at its distal end (also present in Herrerasaurus ischigualastensis, Sanjuansaurus gordilloi, and Saturikosaurus pricei), as well as an ischial shaft that is anteriorly curved along its length (also present in Saturikosaurus pricei). If this position for Saltopus elginensis is correct then it would provide more evidence for the presence of herrerasaurs in Europe during the Late Triassic (see Niedźwiedzki et al. 2014) and would make Saltopus elginensis the first named herrerasaurian species from outside of the Americas. In addition, this result shows that the herrerasaurian clade may have also contained species that were much smaller than the large South American predators Herrerasaurus ischigualastensis, Sanjuansaurus gordilloi, and Staurikosaurus pricei, if the only known specimen of Saltopus elginensis is an adult or near adult; femoral length in Saltopus elginensis (NHMUK PV R3915, left femur) = 68.9 mm; femoral length in Herrerasaurus ischigualastensis (PVL 2566, right femur) = 473 mm (Novas 1993).
In Baron et al. (2017a), the clade Herrersauridae is the sister group to Sauropodomorpha and these two clades formed a monophyletic Saurischia, which in turn formed the sister group to Ornithoscelida. In this model, Ornithoscelida and Saurischia then formed a monophyletic Dinosauria, which was recovered as the sister group to Saltopus elginensis. As our revised analysis has recovered herrerasaurs outside of Dinosauria, essentially in the same position that was suggested for Saltopus elginensis in Baron et al. (2017a), Saurischia, in its more traditional sense, becomes a paraphyletic group. Hence, we propose that Saurischia should now be abandoned as a clade name and that Theropoda, Ornithischia and Sauropodomorpha be retained as the three subsidiary clades within Dinosauria, with Ornithischia and Theropoda forming Ornithoscelida, using the revised definitions presented by Baron et al. (2017a).
For the clade containing herrerasaurs, we suggest (provisionally, at least) reviving the name Herrerasauria. The node-based clade Herrerasauridae, which is defined as “Herrerasaurus, Staurikosaurus, their most recent common ancestor, plus all its descendants” (Novas 1992; Langer et al. 2010), while being appearing more commonly in modern studies (e.g., Ezcurra 2010; Nesbitt 2011; Niedźwiedzki et al. 2014), is limited in its scope by the fact that the taxa it encompasses is strictly dependent on the interrelationships among herrerasaurian taxa. Herrerasauridae, by its current definition, is limited to only contain taxa that are descended from the most recent common ancestor of Herrerasaurus ischigualastensis and Staurikosaurus pricei; Herrerasauridae cannot include the taxa in the herrerasaurian lineage that fall outside of the bracket defined by Herrerasaurus ischigualastensis and Staurikosaurus pricei. Herrerasauria, on the other hand, encompasses any and all taxa that are recovered in the herrerasaur lineage. Given that our main analysis recovers all herrerasaurs in a polytomy, with the relative positions of Herrerasaurus ischigualastensis and Staurikosaurus pricei within the clade unresolved, we feel that Herrerasauria serves as a more appropriate clade name than Herrerasauridae, under current definitions, when discussing the lineage containing Herrerasaurus ischigualastensis and the other taxa found within this clade in this study.
Herrerasauria was defined by Langer (2004) and Langer et al. (2010) as all dinosaurs that share a more recent common ancestor with Herrerasaurus than with Liliensternus and Plateosaurus. However, in our phylogenetic analysis, Herrerasauria is positioned just outside of Dinosauria, necessitating a slight rewording of the definition given by Langer et al. (2010) to “all taxa that share”, or, to put it into a style more consistent with other recent phylogenetic revisions (e.g., Nesbitt 2011; Baron et al. 2017a)—the most inclusive clade that includes Herrerasaurus ischigualastensis but not Liliensternus liliensterni and Plateosaurus engelhardti.
Examination of the various MPTs produced by these analyses revealed that some herrerasaurian taxa could potentially fall stem-ward of Herrerasauridae and therefore represent non-herrerasaurid herrerasaurs. For example, the holotype of Caseosaurus crosbyensis and NMMNH P-35995 were recovered outside of Herrerasauridae in 7 of the 91 MPTs in the reduced analysis (Fig. 9). Future works may better resolve the interrelationships within the herrerasaurian lineage and both Herrerasauria and Herrerasauridae may be found to be valid and necessary. Alternatively, future works may also recover a result in which Herrerasauridae does still contain all currently known members of the herrerasaur lineage and, as a result, Herrerasauridae then would, once again, take priority over Herrerasauria, as it has done in recent times (Benedetto 1973; Galton 1985; Langer 2004; Langer et al. 2010).
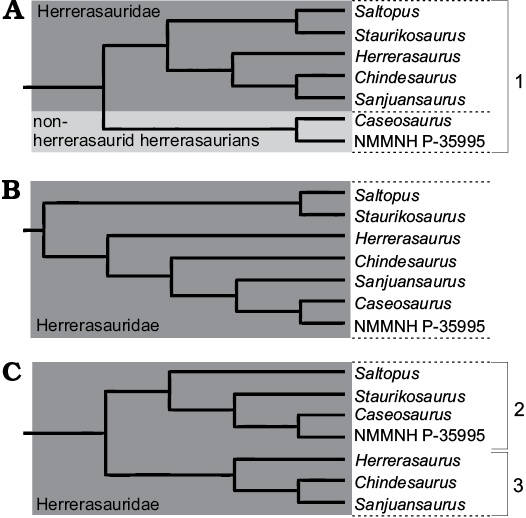
Fig. 9. Possible tree topologies recovered within the herrerasaur lineage from the MPTs produced in the analyses that used the dataset of Baron et al. (2017a). A. Tree with Caseosaurus crosbyensis inside Herrerasauria forming the sister taxon to Herrerasauridae. B. Tree with all herrerasaurian taxa included within Herrerasauridae, with Staurikosaurus pricei and Saltopus elginensis forming a clade. C. Tree with all herrerasaurian taxa included within Herrerasauridae, with two distinct clades: 1, Herrerasauria defined as the most inclusive clade that includes Herrerasaurus ischigualastensis but not Liliensternus liliensterni and Plateosaurus engelhardti; 2, the most inclusive clade that includes Staurikosaurus pricei but not Herrerasaurus ischigualastensis; 3, the most inclusive clade that includes Herrerasaurus ischigualastensis but not Staurikosaurus pricei.
In addition to the revision of relationships near the base of the dinosaur tree, this study has also noted similarities between the ilial anatomy of herrerasaurid dinosaurs, sauropodomorph dinosaurs and certain silesaurids. The rugose areas on the dorsal and lateral surfaces of the pre- and postacetabular processes of Caseosaurus crosbyensis are optimised as a synapomorphy of Herrerasauria in our analysis but also appear in several silesaurids, such as Silesaurus opolensis and Ignotosaurus fragilis (Dzik 2003; Martinez et al. 2013; Fig. 7). In fact, this character (character 318) is also optimised as synapomorphy of Silesauridae. The appearance of this character state in both clades could represent a convergence, or, given the new position suggested for Herrerasauria, a symplesiomorphy retained by herrerasaurians but lost in Dinosauria. In addition to the rugose areas on the pre- and postacetabular processes, some herrerasaurs and some silesaurids were also found to share a sharp ridge that extends anterodorsally from the middle of the supra-acetbaular crest to the preacetabular process. Almost all dinosaurs, as well as some herrerasaurs, have a low, rounded buttress in this area (see Description above), but in Caseosaurus crosbyensis and silesaurids (e.g., Silesaurus opolensis; Dzik 2003), this ridge is more distinct and sharp. This character state is currently optimised as a convergence between the silesaurids and the herrerasaurids that possess it. It may be possible that the condition in the herrerasaurian Caseosaurus crosbyensis is homologous to the condition in silesaurids and represents a plesiomorphic state lost by other herrerasaurs such as Herrerasaurus ischigualastensis. However, as the position of Caseosaurus crosbyensis within Herrerasauria is currently ambiguous, the distribution of this character state also remains ambiguous. Caseosaurus crosbyensis is currently the only known dinosauriform outside of Silesauridae that possesses such a sharp ridge in this location; the condition is not assessable in Saltopus elginensis due to the poor quality of preservation, and no such ridge is present in Marasuchus lilloensis (PVL 3871; Sereno and Arcucci 1994). Whatever the case, the possession of a ridge or crest of some kind (either low and rounded or sharp and distinct) appears to be a synapomorphy of the clade containing Silesauridae, Dinosauria and their most recent common ancestor. A straight to slightly convex ventral margin of the acetabulum is present in all silesaurid taxa in this study, as well as in the basal sauropodomorphs Buriolestes schultzi, Chromogisaurus novasi, Guaibasaurus candelariensis, Pampadromaeus barbarenai, Panphagia protos, and Saturnalia tupiniquim (Langer 2003; Martinez and Alcober 2009; Ezcurra 2010; Cabreira et al. 2011, 2016; Langer et al. 2010) but is not the condition seen in herrerasaurs, all of which appear to have concave ventral margins of the acetabulum. These character states are not currently optimised within the tree; further work (and a possible rephrasing and reordering of the character states) may be needed to help better clarify their distribution within Dinosauriformes.
Conclusions
Our phylogenetic analyses have consistently recovered the holotype of Caseosaurus crosbyensis (UMMP 8870) in a position close to Herrerasaurus ischigualastensis and other herrerasaurids. On the basis of our results and a number of anatomical observations, we have also been able to refer a right ilium from Snyder Quarry (NMMNH P-35995) to Caseosaurus crosbyensis and give an emended diagnosis for this taxon. Our analyses have, with this new anatomical information included, recovered the clade Herrerasauria outside of Dinosauria and placed Saltopus elginensis within Herrerasauria for the first time. Following the work by Baron et al. (2017a), which removed Theropoda from Saurischia, this result now breaks apart what remains of the traditional ‘saurischian’ group. We therefore propose “retirement” of the term “saurischian” in discussions of dinosaur systematics and taxonomy and suggest that Dinosauria be considered as comprising Ornithoscelida (Ornithischia+Theropoda) and Sauropodomorpha only, if our hypothesis is correct. Our result also hints further at the potentially important role of the Laurasian landmass during the early stages of dinosaur and dinosauromorph evolution and provides additional evidence that clades once thought to be of Gondwanan origin may yet prove to have undiscovered diversity within the Late Triassic faunas of the Northern Hemisphere (see Dzik 2003; Niedźwiedzki et al. 2014; Baron et al. 2017a). This work has also highlighted a number of anatomical similarities between the ilia of herrerasaurs, basal sauropodomorphs and silesaurids. It is clear that further work is needed to help clarify the distribution of a number of anatomical features within Dinosauriformes and what such distributions may mean in terms of early dinosaur evolution and dinosauromorph interrelationships.
Acknowledgements
We thank Adam Rountrey, David Vander Weele, and Jeffrey Wilson (all UMMP), for photographs of UMMP 8870 and Spencer G. Lucas (NMMNH) for photographs of NMMNH P-35995. We thank Sterling Nesbitt (Virginia Tech, Blacksburg, Virginia, USA), Christian Kammerer (Museum für Naturkunde, Berlin, Germany), Brandon Peecook (Field Museum of Natural History, Chicago, USA), and Hans-Dieter Sues (Smithsonian Institution, National Museum of Natural History, Washington D.C., USA) for their helpful comments on this manuscript. We also thank Paul M. Barrett (NHMUK) for his helpful comments on this version of the manuscript. We thank Sterling J. Nesbitt and Collin S. Van Buren (Cambridge, UK) for helpful discussions in relation to this project. We thank the Willi Hennig Society for making TNT 1.5-beta software freely available. Funding for MGB was provided by a NERC/CASE Doctoral Studentship (NE/L501578/1).
References
Baron, M.G., Norman, D.B., and Barrett, P.M. 2017a. A novel hypothesis of dinosaur relationships and early dinosaur evolution. Nature 543: 501–506. Crossref
Baron, M.G., Norman, D.B., and Barrett, P.M. 2017b. Postcranial anatomy of Lesothosaurus diagnosticus (Dinosauria: Ornithischia) from the Lower Jurassic of southern Africa: implications for basal ornithischian taxonomy and systematics. Zoological Journal of the Linnean Society. 179: 125–168.
Barrett, P.M., Butler, R.J., Mundil, R., Scheyer, T.M., Irmis, R.B., and Sanchez-Villagra, M.R. 2014. A palaeoequatiorial ornithischian and new constraints on early dinosaur diversification. Proceedings of the Royal Society B: Biological Sciences 281: 20141147. Crossref
Benedetto, J.L. 1973. Herrerasauridae, nueva familia de Saurísquios Triásicos. Ameghiniana 10: 89–102.
Benton, M.J. 1984. Classification and phylogeny of the diapsid reptiles. Philosophical Transactions of the Royal Society of London, Series B 302: 605–720. Crossref
Benton, M.J. and Walker, A.D. 2011. Saltopus, a dinosauriform from the Upper Triassic of Scotland. Earth and Environmental Science Transactions of the Royal Society of Edinburgh 101 (Special Issue 3–4): 285–299.
Bittencourt, J.S. and Kellner, A.W.A. 2009. The anatomy and phylogenetic position of the Triassic dinosaur Staurikosaurus pricei Colbert, 1970. Zootaxa 2079: 1–56.
Bittencourt, J.S., Arcucci, A.B., Marsicano, C.A., and Langer, M.C. 2015. Osteology of the Middle Triassic archosaur Lewisuchus admixtus Romer (Chanares Formation, Argentina), its inclusivity, and relationships amongst early dinosauromorphs. Journal of Systematic Palaeontology 13 (3): 189–219. Crossref
Cabreira, S.F., Kellner, A.W.A., Dias-da-Silva, S., da Silva, L.R., Bronzati, M., de Almeida Marsola, J.C., Müller, R.T., de Souza Bittencourt, J.S., Batista, B.J., Raugust, T., Carrilho, R., Brodt, A., and Langer, M.C. 2016. A Unique Late Triassic Dinosauromorph Assemblage Reveals Dinosaur Ancestral Anatomy and Diet. Current Biology 26: 309–395. Crossref
Cabreira, S.F., Schultz, C.L., Bittencourt, J.S., Soares, M.B., Fortier, D.C., da Silva L.R., and Langer, M.C. 2011. New stem-sauropodomorph (Dinosauria, Saurischia) from the Triassic of Brazil. Naturwissenschaften 98: 1035–1040. Crossref
Case, E.C. 1927. The vertebral column of Coelophysis Cope. Contributions from the Museum of Geology, University of Michigan 10: 209–222.
Chatterjee, S. 1984. A new ornithischian dinosaur from the Triassic of North America. Naturwissenschaften 71: 630–631. Crossref
Chartterjee, S. 1991. Cranial anatomy and relationships of a new Triassic bird from Texas. Philosophical Transactions of the Royal Society B: Biological Sciences 332: 277–342. Crossref
Colbert, E.H. 1970. A Saurischian dinosaur from the Triassic of Brazil. American Museum Novitates 2405: 1–39
Colbert, E.H. 1989. The Triassic dinosaur Coelophysis. Museum of Northern Arizona, Bulletin 57: 160.
Cope, E.D. 1869. Synopsis of the extinct Batrachia, Reptilia and Aves of North America. Transactions of the America Philosophical Society 14: 1–252. Crossref
Cope, E.D. 1887. On a new genus of Triassic Dinosauria. The American Naturalist 23: 626.
Delcourt, R., Azevedo, S.A.K., Grillo, O.N., and Deantoni, F.O. 2012. Biomechanical comments about Triassic dinosaurs from Brazil. Papéis Avulsos de Zoologia (São Paulo) 52: 341–347.
Dzik, J. 2003. A beaked herbivorous archosaur with dinosaur affinities from the early Late Triassic of Poland. Journal of Vertebrate Paleontology 23: 556–574. Crossref
Ezcurra, M.D. 2006. A review of the systematic position of the dinosauriform archosaur Eucoelophysis baldwini Sullivan and Lucas, 1999 from the Upper Triassic of New Mexico, USA. Geodiversitas 28: 649–684.
Ezcurra, M.D. 2010. A new early dinosaur (Saurischia: Sauropodomorpha) from the Late Triassic of Argentina: a reassessment of dinosaur origin and phylogeny. Journal of Systematic Palaeontology 8: 371–425. Crossref
Ferigolo, J. and Langer, M.C. 2006. A Late Triassic dinosauriform from south Brazil and the origin of the ornithischian predentary bone. Historical Biology 19: 1–11.
Fraser, N.C., Padian, K., Walkden, G.M., and Davis, A.L.M. 2002. Basal dinosauriform remains from Britain and the diagnosis of the Dinosauria. Palaeontology 45: 79–95. Crossref
Galton, P.M. 1973. On the anatomy and relationships of Efraasia diagnostica (Huene) n. gen., a prosauropod dinosaur (Reptilia: Saurischia) from the Upper Triassic of Germany. Paläontologische Zeitschrift 47: 229–255. Crossref
Galton, P.M. 1977. Staurikosaurus pricei, an early saurischian dinosaur from the Triassic of Brazil, with notes on the Herrerasauridae and Poposauridae. Paleontologische Zeitschrift 51: 234–245. Crossref
Galton, PM. 1985. The poposaurid thecodontian Teratosaurus suevicus v. Meyer, plus referred specimens mostly based on prosauropod dinosaurs, from the Middle Stubensandstein (Upper Triassic) of Nordwürttemberg. Stuttgarter Beiträge zur Naturkunde B 116: 139–171.
Goloboff, P., Farris, F.C., and Nixon, K.C. 2008. TNT, a free program for phylogenetic analysis. Cladistics 24: 774–786. Crossref
Heckert, A.B., Zeigler, K.E., Lucas, S.G., and Rinehart, L.F. 2003. Coelophysids (Dinosauria: Theropoda) from the Upper Triassic (Revueltian) Snyder quarry. New Mexico Museum of Natural History and Science Bulletin 24: 127–132.
Heckert, A.B., Zeigler, K.E., Lucas, S.G., Rinehart, L.F., and Harris, J.D. 2000. Preliminary description of coelophysoids (Dinosauria: Theropoda) from the Upper Triassic (Revueltian: early–mid Norian) Snyder quarry, north-central New Mexico. New Mexico Museum of Natural History and Science Bulletin 17: 27–32.
Hunt, A.P., Lucas, S.G., Heckert, A.B., Sullivan, R.M., and Lockley, M.G. 1998. Late Triassic dinosaurs from the western United States. Geobios 31: 511–531. Crossref
Irmis, R.B., Nesbitt, S.J., Padian, K., Smith, N.D., Turner, A.H., Woody, D., and Downs, A. 2007a. A Late Triassic dinosauromorph assemblage from New Mexico and the rise of dinosaurs. Science 317: 358–361. Crossref
Irmis, R.B., Parker, W.G., Nesbitt, S.J., and Liu, J. 2007b. Early ornithischian dinosaurs: the Triassic record. Historical Biology 19: 3–22. Crossref
Kammerer, C.F., Nesbitt, S.J., and Shubin, N.H. 2012. The first basal dinosauriform (Silesauridae) from the Late Triassic of Morocco. Acta Palaeontologica Polonica 57: 277–284. Crossref
Langer, M.C. 2003. The pelvic and hind limb anatomy of the stem-sauropodomorph Saturnalia tupiniquim (Late Triassic, Brazil). PaleoBios 23 (2): 1–30.
Langer, M.C. 2004. Basal Saurischia. In: D.B. Weishampel, P. Dodson, and H. Osmólska (eds.), The Dinosauria, 2nd Edition, 25–46. University of California Press, Berkley. Crossref
Langer, M.C., Bittencourt, J.S., and Schultz, C.L. 2011. A reassessment of the basal dinosaur Guaibasaurus candelariensis, from the Late Triassic Caturrita Formation of south Brazil. Earth and Environmental Science Transactions of the Royal Society of Edinburgh 101: 301–332. Crossref
Langer, M.C., Ezcurra, M.D., Bittencourt, J.S., and Novas, F.E. 2010. The origin and early evolution of dinosaurs. Biological Reviews 85: 55–110. Crossref
Long, R.A. and Murry, P.A. 1995. Late Triassic (Carnian and Norian) tetrapods from the Southwestern United States. New Mexico Museum Natural History Science Bulletin 4: 1–254.
Martinez, R.N. and Alcober, O.A. 2009. A basal sauropodomorph (Dinosauria: Saurischia) from the Ischigualasto Formation (Triassic, Carnian) and the early evolution of Sauropodomorpha. PLoS ONE 4 (2): 1–12. Crossref
Martinez, R.N., Apaldetti, C., Alcober, O.A., Colombi, C.E., Sereno, P.C., Fernandez, E., Malnis, P.S., Correa, G.A., and Abelin, D. 2013. Vertebrate succession in the Ischigualasto Formation. Journal of Vertebrate Paleontology 32: 10–30. Crossref
Nesbitt, S.J. 2011. The early evolution of archosaurs: relationships and the origin of major clades. Bulletin of the American Museum of Natural History 352: 1–292. Crossref
Nesbitt, S.J., Irmis, R.B., and Parker, W.G. 2007. A critical re-evaluation of the Late Triassic dinosaur taxa of North America. Journal of Systematic Palaeontology 5: 209–243. Crossref
Nesbitt, S.J., Irmis, R.B., Parker, W.G., Smith, N.D., Turner, A.H., and Rowe, T. 2009a. Hindlimb osteology and distribution of basal dinosauromorphs from the Late Triassic of North America. Journal of Vertebrate Paleontology 29: 498–516. Crossref
Nesbitt, S.J., Sidor, C.A., Irmis, R.B., Angielczyk, K.D., Smith, R.M.H., and Tsuji, L.A. 2010. Ecologically distinct dinosaurian sister group shows early diversification of Ornithodira. Nature 464: 95–98. Crossref
Nesbitt, S.J., Smith, N.D., Irmis, R.B., Turner, A.H., Downs, A., and Norell, M.A. 2009b. A complete skeleton of a Late Triassic saurischian and the early evolution of dinosaurs. Science 326: 1530–1533. Crossref
Niedźwiedzki, G., Brusatte, S.L., Sulej, T., and Butler, R.J. 2014. Basal dinosauriform and theropod dinosaurs from the mid–late Norian (Late Triassic) of Poland: implications for Triassic dinosaur evolution and distribution. Palaeontology 57: 1121–1142. Crossref
Novas, F.E. 1992. Phylogenetic relationships of the basal dinosaurs, the Herrerasauridae. Palaeontology 35: 51–62.
Novas, F.E. 1993. New information on the systematics and postcranial skeleton of Herrerasaurus ischigualastensis (Theropoda: Herrerasauridae) from the Ischigualasto Formation (Upper Triassic) of Argentina. Journal of Vertebrate Paleontology 13: 400–423. Crossref
Novas, F.E., Ezcurra, M.D., Chatterjee, S., and Kutty, T.S. 2011. New dinosaur species from the Upper Triassic Upper Maleri and Lower Dharmaram formations of central India. Earth and Environmental Science Transactions of the Royal Society of Edinburgh 101: 333–349. Crossref
Peecook, B.R., Sidor, C.A., Nesbitt, S.J., Smith, R.M.H., Steyer, J.S., and Angielczyk, K.D. 2013. A new silesaurid from the upper Ntawere Formation of Zambia (Middle Triassic) demonstrates the rapid diversification of Silesauridae (Avemetatarsalia, Dinosauriformes). Journal of Vertebrate Paleontology 33: 1127. Crossref
Pol, D., Garrido, A., and Cerda, I.A. 2011. A new sauropodomorph dinosaur from the Early Jurassic of Patagonia and the origin and evolution of the sauropod-type sacrum. PLoS ONE 6: e14572. Crossref
Raath, M.A. 1969. A new Coelurosaurian dinosaur from the Forest Sandstone of Rhodesia. Arnoldia Rhodesia 4: 1–25.
Reig, O.A. 1963. La presencia de dinosaurios saurisquios en los “Estratos de Ischigualasto” (Mesotriásico superior) de las provincias de San Juan y La Rioja (República Argentina)”. Ameghiniana 3: 3–20.
Rowe, T. 1989. A new species of the theropod dinosaur Syntarsus from the Early Jurassic Kayenta Formation of Arizona. Journal of Vertebrate Paleontology 9: 125–136. Crossref
Sarıgül, V. 2016. New basal dinosauromorph records from the Dockum Group of Texas, USA. Palaeontologia Electronica 19.2.21A: 1–16.
Sereno, P.C. and Arcucci, A.B. 1994. Dinosaurian precursors from the Middle Triassic of Argentina: Marasuchus lilloensis gen. nov. Journal of Vertebrate Paleontology 14: 53–73. Crossref
Smith, N.D. and Pol, D. 2007. Anatomy of a basal sauropodomorph dinosaur from the Early Jurassic Hanson Formation of Antarctica. Acta Palaeontologica Polonica 52: 657–674.
Sues, H.D., Nesbitt, S.J., Berman, D.S., and Henrici, A.C. 2011. A late-surviving basal theropod dinosaur from the latest Triassic of North America. Proceedings of the Royal Society B 278: 3459–3464. Crossref
Sullivan, R.M. and Lucas, S.G. 1999. Eucoelophysis baldwini, a new theropod dinosaur from the Upper Triassic of New Mexico, and the status of the original types of Coelophysis. Journal of Vertebrate Paleontology 19: 81–90. Crossref
Welles, S.P. 1954. New Jurassic dinosaur from the Kayenta formation of Arizona. Bulletin of the Geological Society of America 65: 591–598. Crossref
Yates, A.M. 2003a. A new species of the primitive dinosaur Thecodontosaurus (Saurischia: Sauropodomorpha) and its implications for the systematics of early dinosaurs. Journal of Systematic Palaeontology 1: 1–42. Crossref
Yates, A.M. 2003b. The species taxonomy of the sauropodomorph dinosaurs from the Löwenstein Formation (Norian, Late Triassic) of Germany. Palaeontology 46: 317–337. Crossref
Yates, A.M. 2007. Solving a dinosaurian puzzle: the identity of Aliwalia rex Galton. Historical Biology 19 (1): 93–123. Crossref
Acta Palaeontol. Pol. 63 (1): 129–145, 2018
https://doi.org/10.4202/app.00372.2017