
An Early Triassic gladius associated with soft tissue remains from Idaho, USA—a squid-like coleoid cephalopod at the onset of Mesozoic Era
LARISA A. DOGUZHAEVA, ARNAUD BRAYARD, NICOLAS GOUDEMAND, LAUREL J. KRUMENACKER, JAMES F. JENKS, KEVIN G. BYLUND, EMMANUEL FARA, NICOLAS OLIVIER, EMMANUELLE VENNIN, and GILLES ESCARGUEL
Doguzhaeva, L.A., Brayard, A., Goudemand, N., Krumenacker, L.J., Jenks, J.F., Bylund, K.G., Fara, E., Olivier, N., Vennin, E., and Escarguel, G. 2018. An Early Triassic gladius associated with soft tissue remains from Idaho, USA— a squid-like coleoid cephalopod at the onset of Mesozoic Era. Acta Palaeontologica Polonica 63 (2): 341–355.
We describe an Olenekian (Early Triassic) “fossil squid” belonging to the oldest complex Mesozoic marine biota collected in the Lower Shale unit of the Lower Triassic Thaynes Group in Idaho, USA. The studied specimen shows a tapered structure embedded in a cylindrical soft body. Morphological, ultrastructural and geochemical features of the specimen suggest that it corresponds to an internally-shelled cephalopod exhibiting a tapered micro-laminated gladius with rachis, narrow median and lateral fields and a large conus; a pair of posterior large fin-supported cartilages and fins; ventral and dorsal mantle band-shape structures, the dorsal one being cartilaginous; mantle patches; a stomach containing undigested arm-hooks and sheet-like pieces of potential flooded ink. Coupled SEM/EDS analyses show that (i) arm-hooks and ink were pseudomorphed by nanoparticles (less than 0.6 mm in diameter) of carbon, (ii) gladius and soft tissues were substituted by granules of calcium phosphate, (iii) cartilage canalicula’s were partially filled with calcium phosphate grains and crystals of Zn- and S-containing minerals. The specimen was hence probably fossilized due to metabolism of P- and C-accumulating bacteria. Based on this specimen, Idahoteuthis parisiana Doguzhaeva and Brayard gen. et sp. nov. and Idahoteuthidae Doguzhaeva and Brayard fam. nov. are erected. This family is characterized by an elongated, cylindrical, dorsally cartilaginous muscular mantle; well-developed, about 0.2 mantle length, rounded anteriorly and acute posteriorly, fin-supported cartilages and similarly shaped two fins at conical mantle termination, and thin slender gladius with narrow median and lateral fields, rachis and breviconic conus. This family assumedly falls in Myopsida (Decabrachia). A streamlined body, large fin-supported cartilages and eroded arm-hooks in the stomach of Idahoteuthis Doguzhaeva and Brayard gen. nov. suggest that this was a maneuverable cannibal predator that dwelled in the subequatorial shallow sea of the west coast of Pangaea.
Key words: Cephalopoda, Coleoidea, phylogeny, gladius, soft tissue preservation, Triassic, USA, Idaho.
Larisa A. Doguzhaeva [larisa.doguzhaeva@gmail.com; larisa.doguzhaeva@nrm.se], Department of Palaeobiology, Swedish Museum of Natural History, P.O. Box 50007, SE-10405 Stockholm, Sweden.
Arnaud Brayard [arnaud.brayard@bourgogne.fr], Emmanuel Fara [emmanuel.fara@u-bourgogne.fr], and Emmanuelle Vennin [Emmanuelle.vennin@u-bourgogne.fr], Biogéosciences UMR6282, CNRS, Univ. Bourgogne Franche-Comté, 6 Boulevard Gabriel, 21000 Dijon, France.
Nicolas Goudemand [nicolas.goudemand@ens-lyon.fr], Université de Lyon, ENS de Lyon, CNRS, Université Claude Bernard Lyon 1, Institut de Génomique Fonctionnelle de Lyon, UMR 5242, 46 Allée d’Italie, F-69364 Lyon Cedex 07, France;
Laurel J. Krumenacker [ljkrumenacker@gmail.com], Department of Earth Sciences, Montana State University, P.O. Box 173480, Bozeman, Montana 59717-3480, USA.
James F. Jenks [jenksjimruby@gmail.com], 1134 Johnson Ridge Lane, West Jordan, Utah 84084, USA.
Kevin G. Bylund [kevin@ammonoid.com], 140 South 700 East, Spanish Fork, Utah 84660, USA.
Nicolas Olivier [nicolas. olivier@univ-lyon1.fr], Institute of Functional Genomics of Lyon, ENS Lyon-CNRS 5242-INRA USC 1370, 46 allée d’Italie, 69364 Lyon Cedex 07, France.
Gilles Escarguel [Gilles.Escarguel@univ-lyon1.fr], Laboratoire d’Ecologie des Hydrosystèmes Naturels et Anthropisés, UMR CNRS 5023, Université Claude Bernard Lyon 1, 27-43 Boulevard du 11 novembre 1918, 69622 Villeurbanne Cedex, France.
Received 29 May 2017, accepted 5 April 2018, available online 5 June 2018.
Copyright © 2018 L.A. Doguzhaeva et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
A ~250.6 Myr-old middle Olenekian (= early Spathian, Early Triassic) complex marine ecosystem named the Paris Biota, has been recently discovered in southern Idaho, USA (Brayard et al. 2017). One of the most intriguing specimens of this exceptional fossil assemblage is represented by an elongated structure interpreted as a coleoid gladius associated with soft tissues (Brayard et al. 2017: fig. 4G). This gladius is the oldest one that reveals similarities to gladii of some extant coleoid taxa and therefore it can serve as a basis to propose a new scenario on the evolutionary development of a gladius. Based on this specimen, Idahoteuthis parisiana gen. et sp. nov. and Idahoteuthidae Doguzhaeva and Brayard fam. nov. are erected and their systematic position is analyzed. The preservation of the non-biomineralized structures, a gladius, arm-hooks and sheet patches of a potential ink in the stomach, mantle and fin-supported cartilages, as well as the position of this coleoid in the Early Triassic food web are also discussed.
Institutional abbreviations.—NRM, Swedish Museum of Natural History, Stockholm, Sweden; UBGD, Université de Bourgogne, Géologie Dijon, France.
Geological setting
The gladius specimen comes from the Lower Shale unit of the Lower Triassic Thaynes Group, west of the city of Paris, Bear Lake County, southeastern Idaho, western USA (Fig. 1; Brayard et al. 2017: figs. 1–4). This unit is represented by alternating limestone and shale beds, which were deposited during the middle Olenekian in the shallow Western USA Basin. This epicontinental sea was connected to the Panthalassic Ocean to the west and located at a near equatorial position on the western margin of Pangaea (Fig. 1). Exposures containing this specimen are approximately 20 m thick and consists of mainly grey to blue, thin-bedded silty limestone, deposited in a distal upper offshore environment on a very flat platform. The early Spathian (base of the middle Olenekian) ammonoid Tirolites abundantly occurs throughout these exposures (Brayard et al. 2017). A radiometric age of about 250.6 Mya (i.e., only about 1.3 Mya after the end-Permian mass extinction) is assigned to the immediately overlying Middle Limestone unit and was obtained from the coeval Tirolites–Columbites beds in South China (Galfetti et al. 2007; Jenks et al. 2013).
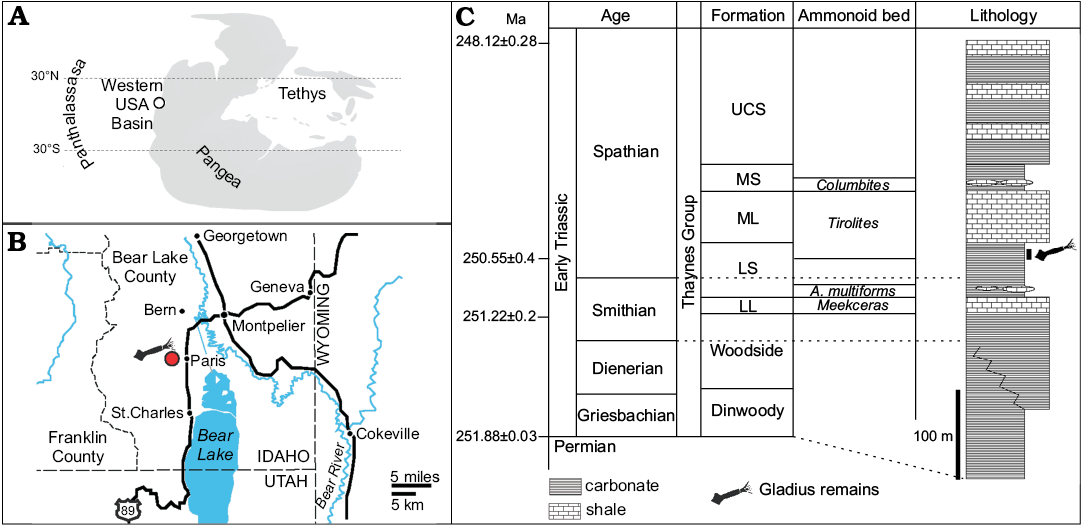
Fig. 1. A. Palaeogeographic map of the western USA basin during the Early Triassic. B. Present-day location of the studied exposure. C. Synthetic stratigraphical and lithological succession of the Bear Lake area (after Romano et al. 2013) with position of the studied specimen. Ammonoid zonation after Jenks et al. (2013) and Jattiot et al. (2016). Stratigraphy follows the main units defined by Kummel (1954). Radiometric ages after Galfetti et al. (2007) and Burgess et al. (2014). Abbreviations: LL, Lower Limestone; LS,Lower Shale; ML, Middle Limestone; MS, Middle Shale; UCS, Upper Calcareous Siltstone.
Material and methods
The studied specimen is ca. 48.7 mm long and ca. 7.4 mm large fossil embedded in a compact silty limestone split into two slabs (Figs. 2A, 3). The specimen is compressed because of compaction and deformed according to original differences in morphology and composition observed between its anterior, middle and posterior parts (Figs. 2, 3). This specimen displays non-biomineralized structures, the original dislocation and symmetry of which may be changed. Considering its tapered end (Figs. 2A, 3) as indicating the anterior part, the specimen is exposed on its left side (Fig. 2A). The tapered end is likely folded and compressed due to compaction (Figs. 2A, 3). The middle part of the left side of the gladius is removed that allows to see the material beneath (Figs. 2A, C, 4). The middle part of the specimen is cylindrical in shape (Fig. 2A); one side (dorsal; at the top on pictures) being more straight and less deformed than the opposite one (ventral; at the bottom on pictures). The latter shows a wavy outline indicative of elastic soft tissue deformations (Fig. 2A). The posterior end of the specimen shows a flattened triangular structure (Fig. 2) and a pair of pear-like structures with a metallic luster (Fig. 2A). At the anterior end of the specimen (Fig. 2A1), an uncertain organic structure distantly resembling longitudinally fractured coleoid-type jaws is present (see Doguzhaeva et al. 2007b: figs. 11.1A, B, 11.7A, 11.8A–F; Doguzhaeva and Mapes 2017: figs. 2A, B, 3A, B for comparison). It is formed by set of long curved, black interrupted lines, some of which are jointed and form a beak-like structure (Fig. 2A1).
The specimen was studied using a Nikon S522249 light microscope and a Wild M400 light photomicroscope. Ultrastructural and elemental analyses were carried out by means of a Scanning Electron Microscope Hitachi-4300 equipped with an energy dispersive spectrometer (EDS) at the Swedish Museum of Natural History in Stockholm. Ultrastructures were examined under magnifications up to 70 000, using a gold coating (Figs. 4–9). Elemental analyses (SOM: table 1, Supplementary Online Material available at http://app.pan.pl/SOM/app63-Doguzhaeva_etal_SOM.pdf) were performed at an accelerated voltage of 15 kV, and we used energy calibration as measured on standard minerals of known accuracy. EDS analyses were made on several points of each morphological structure and matrix (SOM: table 1). All elements were analyzed and no peak was omitted. The contents of the elements are given in per cent to total weight.
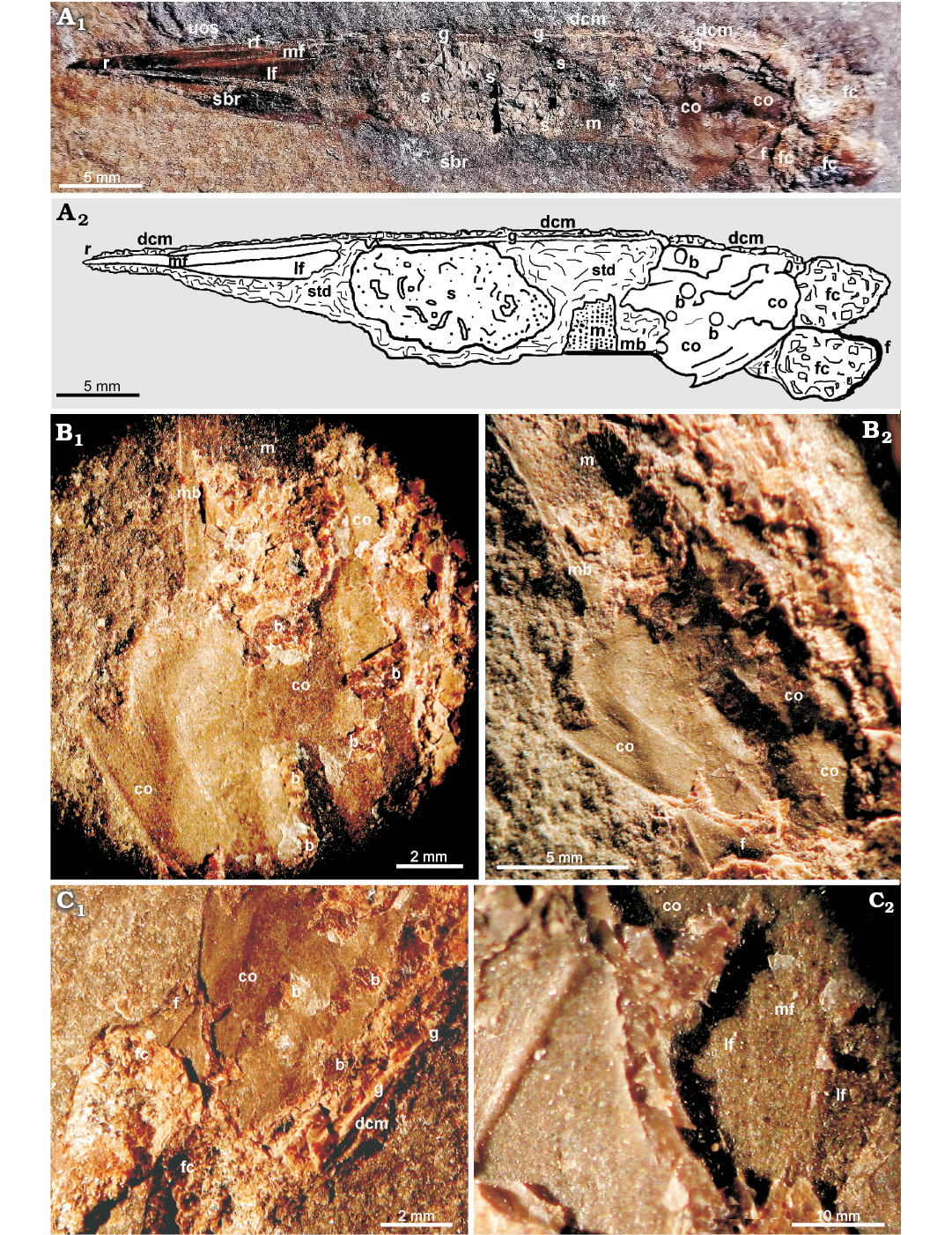
Fig. 2. Coleoid cephalopod Idahoteuthis parisiana Doguzhaeva and Brayard gen. et sp. nov. (holotype, UBGD 30545); middle Olenekian, Early Triassic; Idaho, USA. UBGD 30545. A. Overall view of the specimen showing the anteriorly tapered gladius embedded in soft tissues and uncertain organic structure positioned at its tapered end. Photograph (A1) and explanatory drawing (A2). B. Conus (B1) and its counterpart (B2) showing the outer and inner surfaces, respectively, as well as adjacent part of soft body to show a band-like mantle structure bordering a body ventrally and continuing from the conus forward and a mantle coating the ventral side of the body and showing a mantle rippled pattern (see A2). C. Posterior portion of the specimen showing position (C1) and structute (C2) of tiny fragment of the apical gladius showing three narrow slowly expended fields with rounded anterior outlines. Abbreviations: b, bite mark; co, conus; dcm, dorsal cartilaginous mantle; f, fin; fc, fin cartilage; g, gladius; lf, left lateral field; m, mantle; mb, mantle band-like structure; mf, median field; r, rachis; rf, right lateral field; s, stomach; sbr, soft body remains; std, soft tissue debris; uos, uncertain organic structure.

Fig. 3. Coleoid cephalopod Idahoteuthis parisiana Doguzhaeva and Brayard gen. et sp. nov. (holotype, UBGD 30545); middle Olenekian, Early Triassic; Idaho, USA; a folded tapered anterior part of the gladius with a rachis, and narrow median and acute left and right lateral fields. Abbreviations: lf, left lateral field; mf, median field; rf, right lateral field; sbr, soft body remains.
Results
Morphology and ultrastructure.—The studied specimen shows the following structures; their initial location and symmetry may have been modified because of various potential postmortem deformations.
Gladius: The total length of the exposed part of the gladius is 42.2 mm, and the total preserved length of the fossil is 48.7 mm. The actual length of the gladius is likely longer as the length of the rachis (a free continuation of the median field) is incomplete (Figs. 2A1, 3). The gladius is slender, tapered, with three narrow longitudinal areas (median and lateral fields), a rachis, and a relatively long conus that is ~0.15% of the gladius length (without the rachis) (Fig. 2A1). The gladius structure exhibits five preserved parts: (i) a short piece of a broken rachis, (ii) an anterior tapered part (Figs. 2A, 3), (iii) a dorsal, thin longitudinal fracture between the tapered part and the end of the conus (Fig. 2A), (iv) a conus (Fig. 2), and (v) a displaced tiny ~3 mm long fragment of an apical part (Fig. 2C). It shows three narrow fields that are broadly rounded anteriorly; the median one being slightly longer than lateral fields (Fig. 2C2). The gladius is thin-walled, micro-laminated, non-biomineralized and may have been constituted by chitin (Fig. 9C).
Fin-supported cartilages: These are large, about 0.15% of the gladius length, paired, pear-like structures adjacent to the conus (Fig. 2A). They are formed by a canalicular cartilage (Bairati et al. 1987) characterized by a well developed matrix and the canalicular network made of canals with individualized walls (Figs. 5–7, 8A).
Fins: These are ~10 mm long, partially preserved paired non-biomineralized structures located near the posterior end of the conus. They surround the pear-shaped canalicular cartilages. They are ~0.2 of the preserved length of the specimen (Fig. 2A).
Stomach: This structure corresponds to a large, oval, fractured sac located in the central part of the specimen, close to its ventral side (Fig. 2A). It exhibits a lighter color than the surrounding material and provides access to numerous inclusions. The stomach shows a broken folded edge on its ventral side, excepting on the posterior part where it adjoins a preserved fragment of the band-like mantle structure that may have protected it. The stomach is, however, not broken dorsally and laterally. It contains diverse, supposedly undigested hard remains as well as soft tissue debris; about ten arm hooks and tiny black sheet patches of ink material are identified (Figs. 4, 9A, B, D, F). For comparison, the stomach described in the Late Cretaceous “fossil squid” Dorateuthis contains undigested fish remains (Lukeneder and Harzhauzer 2004).
Arm-hooks: These are found accumulated in the stomach (Fig. 4) and are not observed around the specimen. They are ~2–2.4 mm in maximum size (distance between the distal end of the basement and the acute tip of the hook), black, compressed, fractured, eroded, and lack of typical luster. They have a relatively long and thick basement and display a nearly straight, thick shaft inclined towards a strongly curved and long hook (for comparisons see Rieber 1970; Kulicki and Szaniawski 1972; Engeser and Clarke 1988; Doguzhaeva et al. 2007a: fig. 6.6; Doguzhaeva et al. 2007c: figs. 2B, C, 5A; Doguzhaeva et al. 2010a: figs. 6, 8; Doguzhaeva et al. 2010b: figs. 6A–E, 9C, D).
Mantle: Along the ventral side, the mantle is represented by a short portion of a straight band continuing from the conus forward along the body (Fig. 2A, B) and an adjacent patch with thin wavy longitudinal ridges (Fig. 2B). On the dorsal side of the specimen, it is represented by a dark wavy cartilaginous band, consisted from a canalicular cartilaginous (Figs. 2A2, 7, 8) (for comparison see Doguzhaeva et al. 2007b: figs. 11.2C, 11.3–11.6, 11.7A–D; Doguzhaeva and Mapes 2018: figs. 2.6, 3.2, 3.3, 4.1–4.6).
Soft body remains: This category comprises non-biomineralized remains attached to the gladius anteriorly (Figs. 2A, 3).
Soft tissue debris: Sporadically-dispersed soft tissue remains are identified because of their micro-granular ultrastructure and plastic deformation typical for fossilized non-biomineralized material (Fig. 9A, B, D, E, F). They are preserved between the folded sides of the gladius (for comparison see Doguzhaeva et al. 2007c: figs 6.8–6.11; Doguzhaeva et al. 2010b: figs. 2–9).
Ink: Despite the fact that the ink sac is not preserved, potential flooded ink is irregularly disseminated in the specimen as tiny pieces of black sheets (Figs. 4A, 9A, B). Similar black sheets of flooded ink were observed in some specimens of the Late Triassic coleoid Phragmoteuthis, while some other specimens show flask-shape ink sacs (Doguzhaeva et al. 2007c: figs. 2A, 3B, C). The patches of black sheets were especially identified in the stomach. Like the arm-hooks, they consist of carbon micro-granules (Fig. 9A) (for comparison see Doguzhaeva et al. 2004: figs. 1A–C, 2A, B; Doguzhaeva et al. 2007c: figs. 3B, C, 6B, E).
Bite marks: These marks are rounded or oval, irregularly spaced pits about 1–2 mm in diameter largely observed in the conus (Fig. 2A2, B1, C1).
The uncertain organic structure: This structure consists of dark chitinous laminas (Fig. 2A1). It distantly resembles a longitudinally sectioned coleoid beak (for comparison see Doguzhaeva et al. 2006: figs. 2, 7A–C, 8; Klug et al. 2010: figs. 2A–H; Doguzhaeva and Mapes 2017: figs. 2A, B, 3).
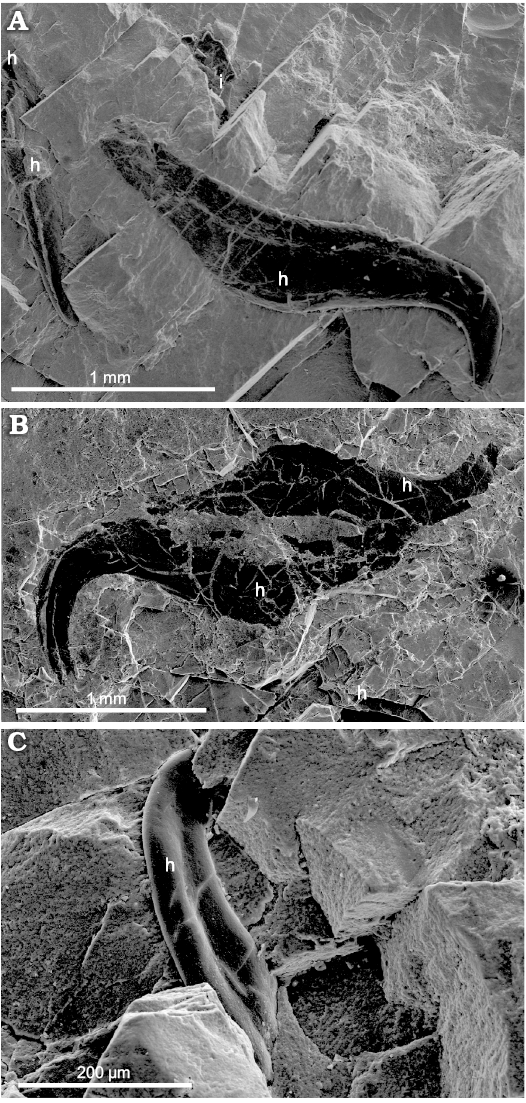
Fig. 4. Coleoid cephalopod Idahoteuthis parisiana Doguzhaeva and Brayard gen. et sp. nov. (holotype, UBGD 30545); middle Olenekian, Early Triassic; Idaho, USA; arm-hooks in the stomach. A. A well exposed arm-hook with deformed shaft and a broken distal end of the basement, tip not exposed; another arm-hook less visible and fractured. B. Three deformed arm-hooks. C. A deformed arm-hook showing a depression along the compressed shaft. Abbreviations: h, arm-hook; i, ink sheet-like patch.
Original composition and fossilization of the specimen (SEM/EDS data).—Gladius: The gladius shows a micro-laminated ultrastructure (Fig. 9C) and is uniformly dominated by calcium phosphate: calcium is about 20–28%; phosphorus is 3–13%, oxygen is 35–50%, and carbon is 10–17% (SOM: table 1). At some places, the gladius is weakly silicified (silicon is up to about 2%) and contains antimony (up to 8%), iodine (about 3%) and sodium, platinum, and magnesium (less than 1%). The gladius lacks sulfur and iron (SOM: table 1). This chemical composition points towards a diagenetic phosphatization of chitin, which is in living coleoids also micro-laminated (Doguzhaeva and Mutvei 2006: figs. 4A–G, 5B). Chitinous fossil gladii have been the place for fossilization processes mediated by phosphorus-accumulating bacteria, which enhance the preservation of gladii as phosphatized pseudomorphs (Doguzhaeva and Mutvei 2003), especially in anoxic paleoenvironments (see Weaver et al. 2011). An experimental study on the micro-laminated pen of Loligo showed that an alkaline treatment changes porosity, wettability, swelling and the crystalline packing of β-chitin by means of intercalation of water molecules between the chitin laminas (Ianiro et al. 2014). Similar ways of degradation of the studied gladius may have preceded its diagenetic phosphatization.
Fin cartilages: The external coating of fin cartilages shows highly variable values of carbon (about 13–40%), oxygen (about 8–48%), phosphorus (up to 9%), sulfur (up to 28%), calcium (from less than 1–22%), and zinc (up to about 50%). It also contains antimony (about 6%), mercury (up to about 4%), iodine (about 2%), silicon (up to 2%) and sodium, aluminum, and iron (less that 1%) (SOM: table 1). The cartilage matrix (its original elastic components were likely elastin and collagen) is uniformly dominated by calcium phosphate, like the gladius: calcium is about 20–28%, phosphorus is up to 12%, oxygen is 36–47%, antimony is 6–9%, iodine is less than 3%, sodium, magnesium, and aluminum are less than 1% (SOM: table 1). However, the cartilage matrix shows small amounts of zinc (up to 1.2%), sulfur (up to 0.4%), potassium (up to 0.6%), and iron (up to 0.9%) which are not detected in the gladius. The canal wall exhibits a composition similar to the surface coating while the canal filling shows large content of zinc (up to 63.5%) and sulfur (up to 19%); carbon is up to about 47%, oxygen is 1–48%, and calcium is 1–24%. Besides, there are mercury (up to about 7%), platinum (up to about 5%), silicon (up to about 6%), antimony (2–6%), sodium (up to about 3%), phosphorus (up to 3%), iron (up to 3%), iodine (up to about 2%), aluminum (up to about 2%), magnesium (about 1%), and potassium (less than 1%). Thus, EDS data indicate that fin cartilages, which have a more complex in vivo histological structure, show a more diversified, diagenetic chemical composition than the micro-laminated gladius, including a specific distribution of heavy metals.
Matrix adjoining the gladius: The matrix shows values of carbon similar to those of the gladius (about 17%) while the oxygen content is slightly higher (63% against 51%) and the maximum values of phosphorus are twice lower than in the gladius (about 6% against 13% in the gladius; SOM: table 1), potentially indicating differential fossilization and diagenetic pathways. Calcium is highly variable (4–23%) but its highest values are only slightly lower than in the gladius (about 28%). Silicon maximum values are five times higher than in the gladius. Potassium, sodium, and sulfur, which are absent in the gladius, are detected in the matrix but values remain anecdotal (less that 1%). Antimony and iodine are slightly lower in the contacts than in the gladius in the matrix but values remain anecdotal (7% and 2% against 8% and 3%, respectively). The composition of the matrix adjoining the gladius was thus probably in situ poorly influenced by the decay of soft tissues.
Arm-hooks: They are uniformly dominated by high carbon content (about 70%). Oxygen is about 25%, calcium 3%, and chromium 1% (SOM: table 1). This composition points towards a fossilization by means of carbonization. The carbonization of arm-hooks was previously reported in the Late Triassic Phragmoteuthis (Doguzhaeva et al. 2007c). The differing composition of the observed chitin structures, like the gladius and the arm-hooks, may result from minor variations of their original chemical composition, as well as the potential different packing of the chitin sheets and the crystalline packing of β-chitin. This observation agrees with the micro-granules of carbon found in the Late Triassic Phragmoteuthis, which are smaller in the arm-hooks than in the gladius suggesting some ultrastructural differences (Doguzhaeva et al. 2007c).
Ink: Observed potential ink tiny patches show high content of carbon (up to about 46%) and oxygen (up to about 44%), and low content of calcium (up to 6%), phosphorus (up to 3%), titanium (up to 3%), and sulfur (about 2%) (SOM: table 1). Therefore, the ink (originally melanin) was fossilized by means of carbonization, like the arm-hooks. The ink is the only substance showing a peak of titanium. This may correspond to observations in vivo on melanin from coleoid ink, which absorbs heavy metals (Chen et al. 2009).
Overall, the SEM/EDS analyses (SOM: table 1) highlight the main diagenetic processes responsible for preservation of the organic matter-rich structures; they are: (i) phosphatization of the gladius, cartilages, mantle, stomach mass and soft tissue debris, (ii) zinc-sulfur mineralization of the canalicular system in fin cartilages, and (iii) carbonization of arm hooks and ink. In addition to zinc, the absorption of heavy metals such as platinum, mercury, and titanium occurred during the fossilization of the non-biomineralized structures (SOM: table 1). The observed differential soft tissue preservation could result from the combined effects of the metabolism of anaerobic P-, C-, and S-accumulating bacteria and seawater potentially characterized by a high content of sulfur and heavy metals. These observations are in accordance with previous experimental studies showing, e.g., the capacity of the invertebrate cartilage tissues to form hydroxyapatite (Eilberg et al. 1975), and the demonstrated in vivo adsorption and absorption of heavy metals by soft tissues of modern coleoids (see Miramand and Guary 1980; Smith et al. 1984; Chen et al. 2009; Jamil et al. 2014).
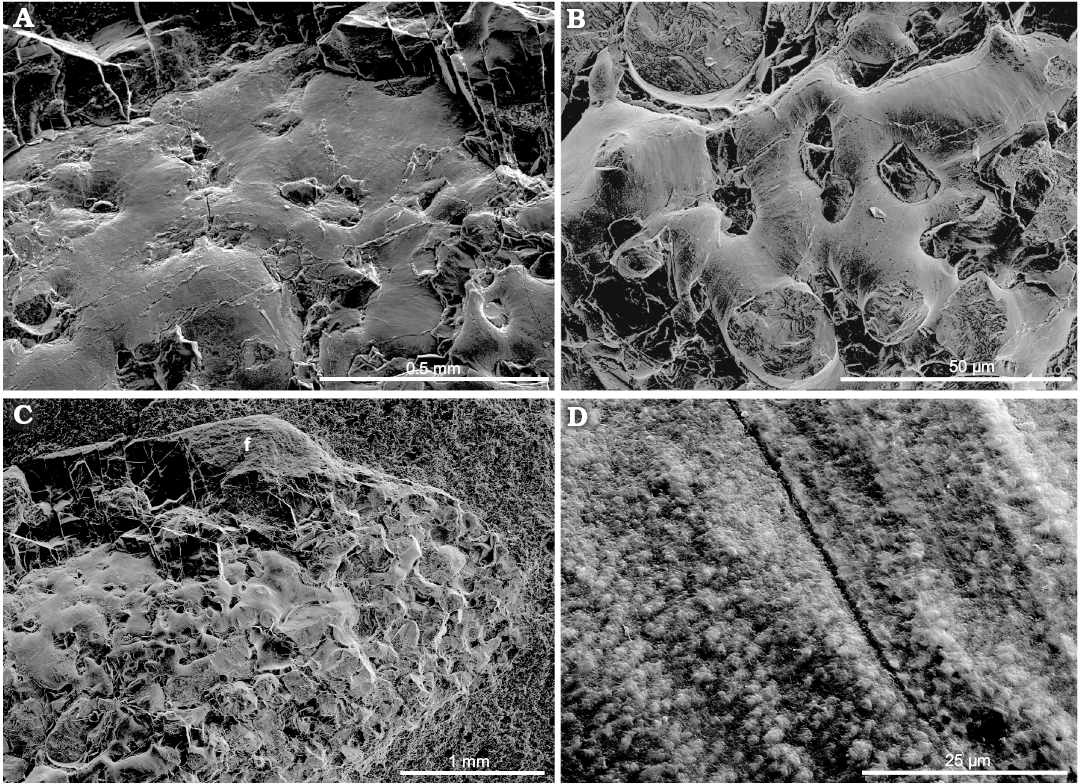
Fig. 5. Coleoid cephalopod Idahoteuthis parisiana Doguzhaeva and Brayard gen. et sp. nov. (holotype, UBGD 30545); middle Olenekian, Early Triassic; Idaho, USA; fin-supported cartilages. The surfaces of fractured cartilages (A–C), outer surface (D). Abbreviation: f, fin.
Systematic palaeontology
Class Cephalopoda Cuvier, 1797
Subclass Coleoidea Bather, 1888
Superorder Decabrachia Boettger, 1952
?Order Myopsida Orbigny, 1841
Family Idahoteuthidae Doguzhaeva and Brayard nov.
Type genus: Idahoteuthis Doguzhaeva and Brayard gen. nov.; see below, monogeneric.
Diagnosis.—Cylindrical, stream-line, dorsally cartilaginous muscular mantle; two well-developed, ca. 0.2–0.3 mantle length, pear-shape fin-supported cartilages and fins at conical mantle termination; thin slender gladius with narrow median and lateral fields, rachis; breviconic conus as long as fin-supported cartilages and fins.
Geographic and stratigraphic range.—Middle Olenekian (early Spathian) Lower Shale unit of the Lower Triassic Thaynes Group; west of the city of Paris, Bear Lake County, southeastern Idaho, western USA.
Genus Idahoteuthis Doguzhaeva and Brayard nov.
Type species: Idahoteuthis parisiana Doguzhaeva and Brayard gen. et sp. nov., see below, monotypic.
Etymology: Named after the state of Idaho, USA where the specimen was found.
Diagnosis.—Slender gladius; maximum width/length ratio, ca. 0.2, in middle part; long, narrow, anteriorly angulated lateral fields; median and lateral keels; conus large, ca. 0.25 gladius length without rachis length.
Remarks.—At the juvenile stage, the gladius is narrow, with three narrow fields each of which shows a broadly rounded anterior outline (Fig. 2C). The median field is slightly longer that lateral fields. This morphology suggests that the rachis and anteriorly tapered median and angulated lateral fields appeared at later ontogenetic stage. The new family Idahoteuthidae assumedly falls in the order Myopsida. Extant myopsids are neritic, often dwelling in very shallow waters, or are upper slope demersal species; many species are also efficient swimmers (Vecchione and Young 2008). Combined observed features in Idahoteuhis gen. nov. and comparisons with present-day coleoids therefore suggest that members of Idahoteuthidae had a streamlined body, a micro-laminated ultrastructure and a non-biomineralized, supposedly chitinous composition of gladius providing a light and flexible skeleton, as well as well-developed fin cartilages favoring effective maneuvering in swimming.
Idahoteuthis parisiana Doguzhaeva and Brayard sp. nov.
Figs. 2–9.
Etymology: Named after the city of Paris, Idaho (USA), where the specimen was found.
Holotype: UBGD 30545. The specimen is exposed on two split surfaces of the broken concretion. It is compressed because of compaction and strongly deformed according to original differences in morphology and composition observed between its anterior, middle and posterior parts. It reveals a multi-way fossilization resulted in different preservation of chitin gladius, and arm hooks and soft tissue remains..
Type horizon: Early Triassic, middle Olenekian (early Spathian).
Type locality: West of the city of Paris, southeastern Idaho, western USA.
Diagnosis.—Tapered, slender, ca. 40–50 mm long, gladius with maximum width/length ratio, ca. 0.2, in its middle part; narrow median field with rachis; narrow, anteriorly angulated long lateral fields; median and lateral keels; conus large, ca. 0.25 gladius length without rachis length.
Description.—The holotype is a 48.7 mm long specimen showing non-biomineralized structures. Its anterior, central and posterior parts are differently deformed. The middle part of the gladius is embedded in soft body that retains its cylindrical shape in the middle part of the specimen; the cephalic area is not preserved (Fig. 2A). The gladius is thin-walled, micro-laminated, flexible, diagenetically phosphatized and apparently originally chitinous (Fig. 9C; SOM: table 1). It is ca. 42.2 mm long, anteriorly tapered and exhibits a large conus at its posterior end (Figs. 2, 3). The median field is narrow and anteriorly ended by a free rachis (Figs. 2A, 3). The extremity of the rachis is broken and its actual length is unknown; the measured length is about 4 mm. The lateral fields are narrow, long, and acute anteriorly (Fig. 2A1). The conus is ca. 8.3 mm long, that is ca. 0.25 of the gladius length (Fig. 2A, C1). The posterior extremity of the conus approximately corresponds to the top of the V-shape “sinus” between fins (Fig. 2C). The conus shows a small lateral tooth-like structure (Fig. 2A2, B, C1). The smooth outer surface of the gladius is exposed in its most anterior tapered part (Fig. 2A), in the conus (Fig. 2A1, C1) and in a tiny ~3 mm apical part of the gladius that is displaced from its original position and seen in the marginal part of the conus (Fig. 2C). The apical gladius has slowly expanded narrow median and two lateral fields with broadly rounded anterior outline; the central field being slightly longer and broader that lateral ones. This highlights the presumed ontogenetic transformation of the rounded frontal outline of the juvenile gladius into the tapered frontal outline in later ontogenetic stages. It is worth noting that the ontogenetic stage that would be characterized by a Loligosepia-type gladius showing a broad triangular median field, is missing in the ontogeny of Idahoteuthis parisiana Doguzhaeva and Brayard gen. et sp. nov. In the middle part, the gladius is deformed in a way that its sides are brought together and overlap one another. Because the left half of the gladius is broken and removed, soft tissue remains are exposed there (Fig. 2A, B). This part of the specimen is occupied by a large, oval, phosphatized stomach (Fig. 2A1). Due to compaction, the latter likely looks larger, than it was originally. The stomach exhibits a lighter color than other structures in this area. It has a gently folded periphery along the ventral side but it is less deformed along the dorsal side (Fig. 2A). The stomach contains irregularly dispersed deformed arm hooks, ten of which have been found on a fracture plan (Fig. 4). It is worth to note that the stomach of the Late Cretaceous “fossil squid” Dorateuthis contained the undigested fish remains (Lukeneder and Harzhauzer 2004). In addition to the arm hooks, the exposed surface of the stomach displays scattered tiny, black sheet fragments of potential flooded ink. Like the arm hooks, ink exhibits a micro-globular ultrastructure and a carbon composition (Fig. 9A; SOM: table 1). Posteriorly, a pair of pear-like, fin-supported cartilages and fins complete the specimen (Figs. 2A, C1, 5, 6, 8A). They adjoin the conus; both being approximately equally long. The mantle is exposed like a ventral band-shape structure extending from the conus along the ventral margin of the soft body (Fig. 2A2, B). It might provide postmortem fixation of the cylindrical shape of the body on the ventral side where other supporting structures were missing. The ventral mantle is also represented by a patch preserved in front of the conus; it has thin, wavy longitudinal ridges (Fig. 2B). On the dorsal side, the gladius is coated by a band-shape structure of thin cartilaginous mantle (Figs. 2A2, C1, 7, 8). This cartilage tissue is canalicular (Figs. 7B, 8), like in fin cartilages (Figs. 5C, 6, 8A). Additionally, sporadically-dispersed mantle debris are identified by their plastic micro-deformation and micro-granular ultrastructures typical for fossilized non-biomineralized materials (Fig. 9A, E, F; for comparison see Doguzhaeva et al. 2004b: figs. 2A, B; Doguzhaeva et al. 2007a: figs. 6.7A–E; Doguzhaeva et al. 2007b: figs. 11.3–11.7; Doguzhaeva et al. 2010b: figs. 2–9).
Stratigraphic and geographic range.—Type locality and horizon only.
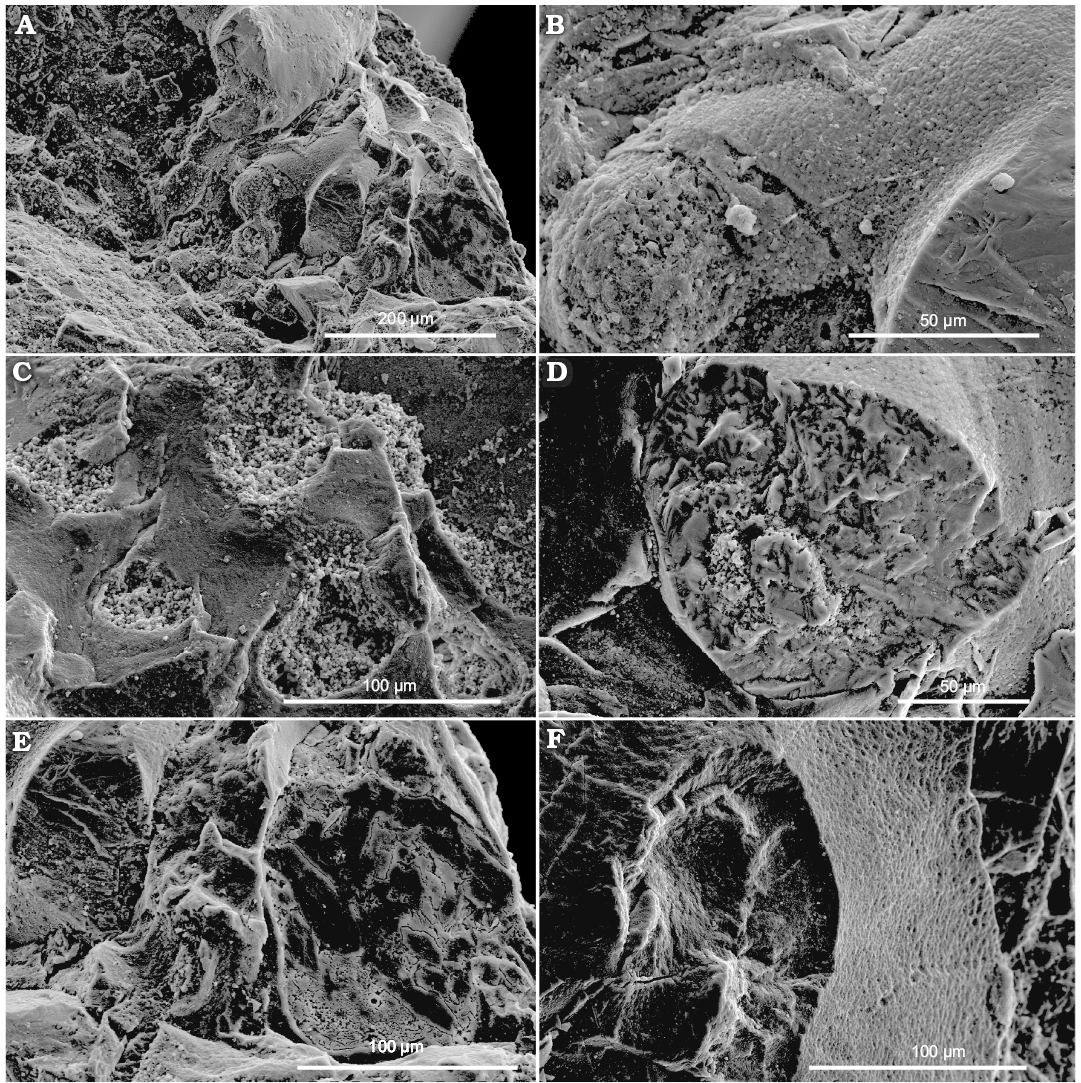
Fig. 6. Coleoid cephalopod Idahoteuthis parisiana Doguzhaeva and Brayard gen. et sp. nov. (holotype, UBGD 30545); middle Olenekian, Early Triassic; Idaho, USA; views on the canalicular structure of fin-supported cartilages (A–D).
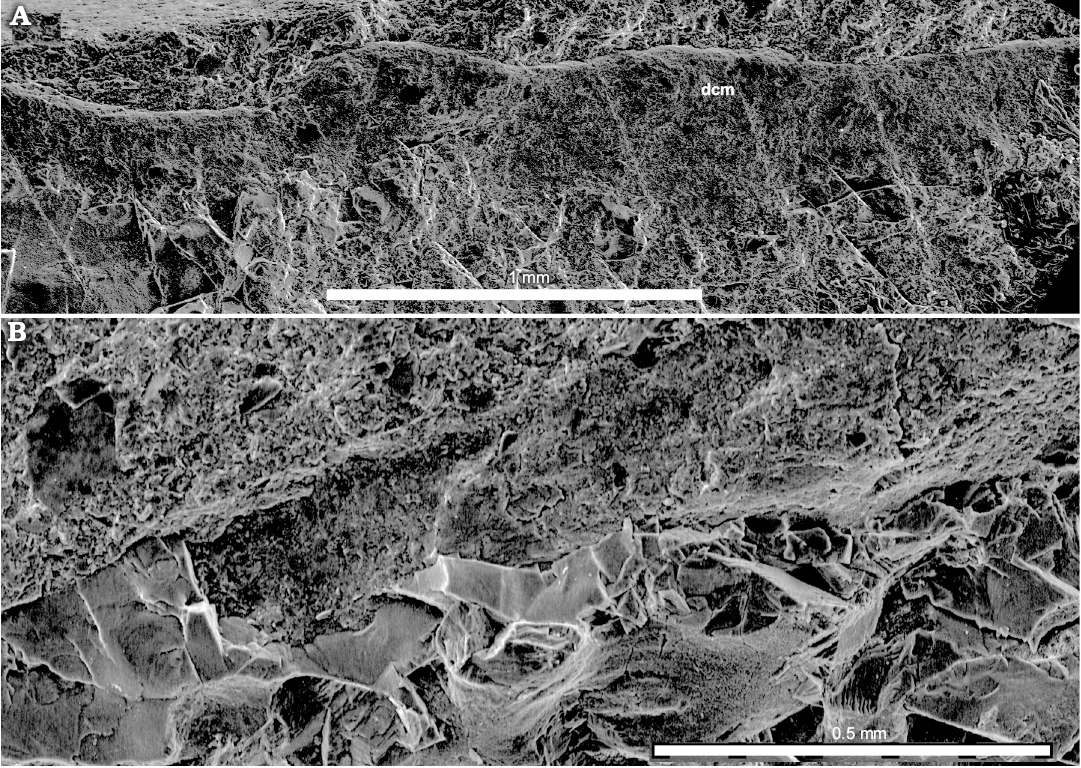
Fig. 7. Coleoid cephalopod Idahoteuthis parisiana Doguzhaeva and Brayard gen. et sp. nov. (holotype, UBGD 30545); middle Olenekian, Early Triassic; Idaho, USA; cartilaginous mantle band-shape structure on the dorsal side of soft body. Overview (A) and detail of analicular cartilage (B). Abbreviation: dcm, dorsal cartilaginous mantle.
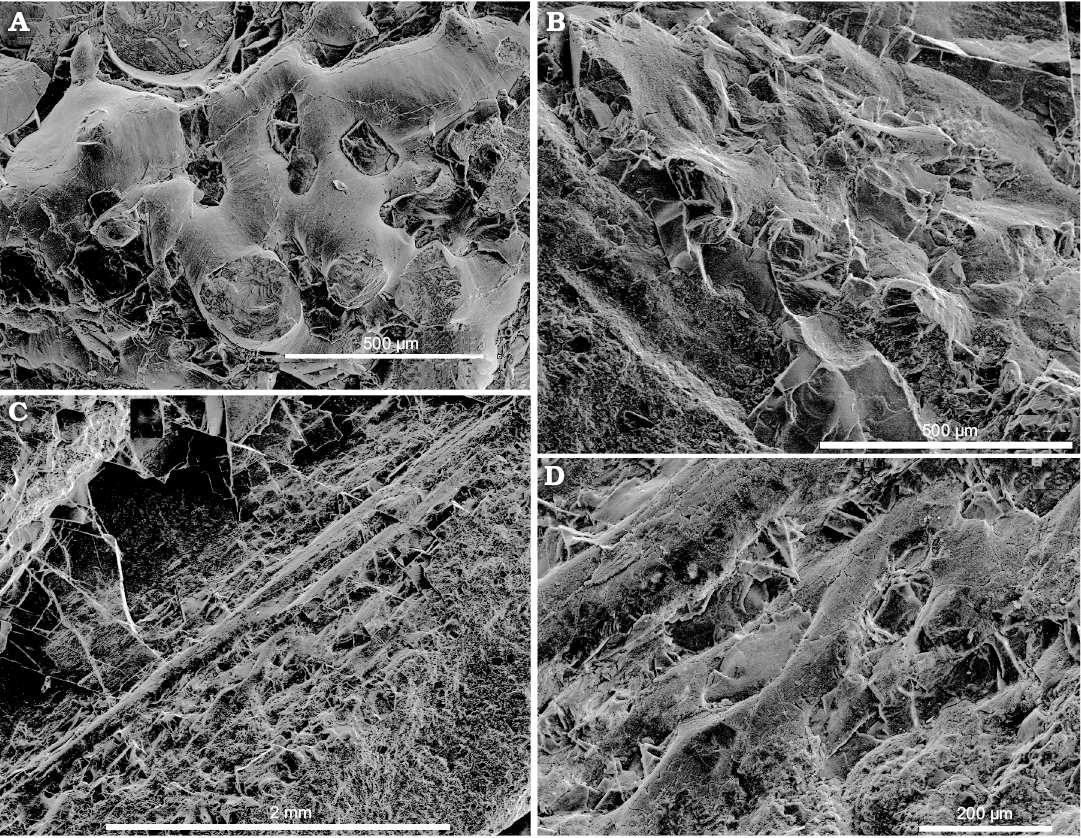
Fig. 8. Coleoid cephalopod Idahoteuthis parisiana Doguzhaeva and Brayard gen. et sp. nov. (holotype, UBGD 30545); middle Olenekian, Early Triassic; Idaho, USA. A. Canalicular cartilage of fin-supported structure. B–D. Cartilaginous mantle band-shape structure on dorsal side showing a canalicular type of cartilage.

Fig. 9. Coleoid cephalopod Idahoteuthis parisiana Doguzhaeva and Brayard gen. et sp. nov. (holotype, UBGD 30545); middle Olenekian, Early Triassic; Idaho, USA; globular ultrastructure of non-biomineralized structures. A. Sheet-like patch of flooded ink (top) and soft tissue debris (bottom); a site of the geochemical analysis is marked with a square. B. Undetermined long soft tissue piece showing a micro-granular ultrastructure on the dorsal side of the soft body. C. Micro-laminated ultrastructure of the gladius. D. Undetermined non-biomineralized micro-laminated fragment from the stomach. E. A linear fragment of soft tissue on dorsal side of the body. F. Micro-globular ultrastructure. Abbreviations: i, ink sheet-like patch; sbt, soft body tissues; std, soft tissue debris.
Discussion
Phylogenetic implications.—Naef (1922) suggested that the evolutionary development of the gladius started based on a broad Loligosepia-type gladius progressively narrowing, with a median field reducing up to a thin rib or rachis leading to the narrow gladii of extant taxa. In this way, Jeletzky (1966) considered the Suborder Loligosepiina Jeletzky, 1965 as the stem group of modern oegopsid and myopsid squids. Morphological similarities between the gladii of the Early Jurassic Loligosepia, characterized by a broad triangular median field (see Fuchs and Weis 2008; Donovan and Boletzky 2014), and extant squid Thysanoteuthis were also interpreted as evidence of an ancestor-descendant relationship between these two genera by Starobogatov (1983). These similarities were oppositely considered as having no phylogenetic value by Nesis (1992). Young et al. (1998) assumed that the gladius appeared at least four times in extant taxa Vampyromorpha, Oegopsida, Myopsida, and Sepiolidae. Vecchione et al. (1999) also suggested that a three-part gladius with a broad median field and wings has no value for determining phylogenetic relationships. Later, Kröger et al. (2011) endorsed this view. Fuchs et al. (2013) described a Loligosepia specimen showing eight, rather than ten, arms in an arm crown. Since this discovery, loligosepiids have been considered separately from modern Decabrachia (Fuchs et al. 2013) and Donovan and Boletzky (2014) suggested that Loligosepia lies near the origin of Recent Octobrachia during the Early Jurassic. Following this view, fossil gladii with a Loligosepia-type gladius fall in Octobrachia rather than Decabrachia. This might be the case for the Middle Triassic Reitnerteuthis and the Late Triassic Germanoteuthis in which the median field is breviconic and triangular (Schweigert and Fuchs 2012). Contrary to these two Middle and Late Triassic genera, the Early Triassic Idahoteuthis Doguzhaeva and Brayard gen. nov. may fall in Decabrachia because of its narrow median field. Narrow gladii are well documented in e.g., younger Plesioteuthis from the Late Jurassic (Donovan and Toll 1988; Fuchs et al. 2007, 2015; Klug et al. 2010, 2015). Based on morphological similarities with gladii of extant squids (Ommastrephidae) (i.e., a narrow and long rachis, median and lateral keels, and the conus), Plesioteuthis was considered as the rootstock of oegopsid squids (Jeletzky 1966; Donovan and Toll 1988). Based on recently recovered beaks Plesioteuthis was assigned to Decabrachia (Klug et al. 2010), however based on two pairs of fins and cirrate arms, Plesioteuthis has been assigned to octobrachians (Klug et al. 2015).
A narrow gladius is also known in e.g., the Early Cretaceous Nesisoteuthis. Judging on a shape of growth lines, it has a pointed median field and is even more similar to ommastrephids than the Jurassic Plesioteuthis. However, whether Nesisoteuthis has a conus remains unknown as a single specimen of this genus is available and does not show a preserved posterior part (Doguzhaeva 2005: figs. 1A–C).
Idahoteuthis Doguzhaeva and Brayard gen. nov. is preceded by the Early Permian gladius-bearing coleoid Glochinomorpha stifeli (Gordon 1971) that shows limited morphological similarity with the other known gladii (Doguzhaeva and Mapes 2015: figs. 1–3). Recently obtained data on Glochinomorpha via ultrastructural and chemical approaches, however, suggests that: (i) the gladius appeared in the late Paleozoic, rather than in the Middle–Late Triassic, (ii) its appearance preceded Middle–Late Triassic phragmoteuthids still having a phragmocone, (iii) the biochemical development of shell material (sensu secretion of non-biomineralized, apparently chitinous, shell material as opposed to aragonite in the phragmocone of Phragmoteuthis) preceded its morphological transformation (sensu typical gladius structure with a limited number of longitudinal sectors and the presence of a conus), and (iv) the combination of the archaic gladius morphology and advanced gladius composition results from the asynchronous appearance of biochemical and morphological innovations in early evolutionary stages of squids (Doguzhaeva and Mapes 2015). Thus, a high evolutionary rate corresponding to the assumed morphologic transformation of the Glochinomorpha-type gladius (see Doguzhaeva and Mapes 2015: figs. 1A–H, 2–4) into a tapered, narrow Idahoteuthis-type gladius (Figs. 2A) have taken place during the Early Permian–Early Triassic interval. The Early Triassic occurrence of the Idahoteuthis-type gladius supports a scenario where gladius-bearing coleoids evolved independently from the proostracum-bearing belemnoids, including phragmoteuthids (see Doguzhaeva 2012). Even if Phragmoteuthis-type proostraca are found in older intervals than the middle Olenekian (see Rosenkrantz 1946), a direct evolutionary transformation of this shell type into a gladius would request the unlikely short-time combination of the following processes: (i) the elimination of a phragmocone comprising about 20 camerae, septa, septal necks, connecting rings and conotheca; (ii) the elimination of a soft siphuncle, including blood vessels and connecting tissues, running through approximately 20 camerae of the phragmocone; (iii) the re-organization of the soft tissues and muscular system; (iv) the development of physiological processes that can have enhanced secretion of chitin and associated proteins, instead of aragonite shell material; and (v) the appearance of a new locomotion system. Keeping this in mind, a direct transformation of the Phragmoteuthis-type shell into an Idahoteuthis-type gladius is hardly conceivable in a relatively short geological time interval. For the same reasons, the Phragmoteuthis lineage can hardly be considered, in our opinion, as the rootstock of Decabrachia or Octobrachia (Fuchs and Iba 2015).
To sum up, the earliest known gladii are presently known due to the Early Permian Glochinomorpha and the Early Triassic Idahoteuthis. Contrary to Glochinomorpha-type gladius, the Idahoteuthis-type gladius exhibits advanced morphological characters: three narrow longitudinal fields and a conus that are developed in extant Myopsida and Oegopsida. Based on molecular data, the divergence time for Myopsida appears close to the Late Permian/Early Triassic interval (Strugnell et al. 2006). The divergence time for Decabrachia is older (Strugnell et al. 2006), which is in agreement with the Late Carboniferous occurrence of the spirulid Shimanskya (Doguzhaeva et al. 1999).
Implications for Early Triassic food webs.—Numerous arm hooks have been collected in coprolites from the same exposures where Idahoteuthis Doguzhaeva and Brayard gen. nov. was found. Thus, in the middle Olenekian of the western USA basin, some coleoids were a prey for indeterminate predators, likely large-sized vertebrates such as fishes or ichthyosaurs (Brayard et al. 2017). For instance, chondrichthyan remains corresponding to specimens of about 1 m in length are common in some coeval exposures from Idaho (Romano et al. 2013). Ichthyosaur remains are also known from neighboring localities, and can have also been potential hunters for coleoids (Brayard et al. 2017). A strong prey-predator trophic relationship between coleoids and vertebrates thus may have existed during the middle Olenekian, at least in the Western USA basin. Based on the chitinous, thin-walled, tapered narrow gladius indicative of a streamlined body, the well-developed fin cartilages, the posterior fins, and the arm-hooks in the stomach, I. parisiana Doguzhaeva and Brayard gen. et sp. nov. was probably well adapted for active swimming and may have also been a small size predator inhabiting the subequatorial shallow warm sea on the west coast of Pangaea. Overall, the presence of eroded and deformed arm-hooks in the stomach of Idahoteuthis (Fig. 4) suggests cannibalism among Early Triassic coleoids, as frequently encountered in extant squids (Ibánez and Keyl 2010). Such trophic links among coleoids and between coleoids and other taxa highlight the multi-layered marine trophic network in the subequatorial warm shallow water marine environment of the west coast of Pangaea soon after the end-Permian mass extinction.
Conclusions
An Olenekian (Early Triassic) small size squid Idahoteuthis parisiana Doguzhaeva and Brayard gen. et sp. nov. and Idahoteuthidae Doguzhaeva and Brayard fam. nov. are described on the basis of a narrow, tapered, thin-walled gladius embedded in cylindrical body showing a pair of fins, large fin-supported cartilages, a dorsal cartilaginous band-like mantle structure, ventral non-cartilaginous band-like mantle structure and a stomach containing undigested arm hooks and flat patches of ink. The family Idahoteuthidae assumedly belongs to order Myopsida.
Idahoteuthis parisiana evidences the fossil squids inhabited the subequatorial warm shallow sea on the west coast of Pangaea in the Early Triassic; they were mobile predators, but also a prey for larger predators, including coleoids. Cannibalism of I. parisiana is deduced from the stomach content comprising undigested arm hooks. The micro-laminated lifetime chitin material of the gladius, microenvironments linked to diverse microbial blooms, and transient anoxic/disoxic conditions are considered as the main potential factors leading to the formation of the multiple fossilization types of the gladius and soft tissue body remains.
Acknowledgements
We thank all these persons and organizations that supported us and improved the paper. Dirk Fuchs (Bayerische Staatsammlung für Paläontologie und Geologie, Berlin, Germany), Viacheslav Bizikov (Russian Institute of Fisheries and Oceanography, Moscow, Russia) and three anonymous referees reviewed the paper. Daniel A. Stephen (Department of Earth Science, Utah Valley University, Orem, USA) helped during fieldwork. The Bear Lake County Road Department allowed access to the outcrop yielding the studied gladius. This study was supported by the Arnaud Brayard projects AFTER (ANR-13-JS06-0001-01), EvoDevOdonto (ANR-14-ACHN-0010), and the Royal Swedish Academy of Sciences, personally by Vivi Vajda, a Head of the Department of Palaebiology of the Swedish Museum of Natural History.
References
Bairati, A., De Biasi, S., Cheli, F., and Oggioni, A. 1987. The head cartilage of cephalopods. I. Architecture and ultrastructure of the extracellular matrix. Tissue cell 19: 673–685. Crossref
Bather, F.A. 1888. Shell-growth in Cephalopoda (Siphonopoda). Annals and Magazine of Natural History 6: 421–427. Crossref
Boettger, C.B. 1952. Die Stämme des Tierreichs in ihrer systematischen Gliederung. Abhandlungen der Braunschweigischen Wissenschaftlichen Gesellschaft 4: 238–300.
Brayard, A., Krumenacker, L.J., Botting, J.P., Jenks, J.F., Bylund, K. G., Fara, E., Vennin, E., Olivier, N., Goudemand, N., Saucède, T., Charbonnier, S., Romano, C., Doguzhaeva, L., Thuy, B., Hautmann, M., Stephen, D.A., Thomazo, C., and Escarguel, G. 2017. Unexpected Early Triassic marine ecosystem and the rise of the Modern evolutionary fauna. Science Advances 3: e1602159. Crossref
Chen,S., Xue,C., Wang, J., Feng, H., Wang, Y., Ma, Q., and Wang, D. 2009. Adsorption of Pb (II) and Cd (II) by squid Ommastrephes bartrami melanin. Bioinorganic Chemistry and Applications 2009: 901563. Crossref
Cuvier, G. 1797. Tableau elementaire de l’histoire naturelle des animaux. 710 pp. Baudoin, Paris.
Doguzhaeva, L.A. 2005. A gladius-bearing coleoid cephalopod from the Aptian of Central Russia. Mitteilungen des Geologisch-Paläontologischen Institutes der Universität Hamburg 89: 41–48.
Doguzhaeva, L.A. 2012. The original composition of the pro-ostracum of an early Sinemurian belemnite from Belgium deduced from mode of fossilization and ultrastructure. Palaeontology 55: 249–260. Crossref
Doguzhaeva, L.A. and Mutvei, H. 2003. Gladius composition and ultrastructure in extinct squid-like coleoids: Loligosepia, Trachyteuthis and Teudopsis. Revue de Paléobiologie 22: 877–894.
Doguzhaeva, L.A. and Mapes, R.H. 2015. Arm hooks and structural features in the Early Permian Glochinomorpha Gordon 1971, indicative of its coleoid affiliation. Lethaia 48: 100–114. Crossref
Doguzhaeva, L.A. and Mapes, R.H. 2017. Beak from the body chamber of the Early Carboniferous (Visean) shelled longiconic coleoid (Cephalopoda) from Arkansas, USA. Lethaia 50 (4): 540–547. Crossref
Doguzhaeva, L.A. and Mapes, R.H. 2018. A new late Carboniferous coleoid from Oklahoma, USA: implications for the early evolutionary history of the subclass Coleoidea (Cephalopoda). Journal of Paleontology 92 (2): 157–169. Crossref
Doguzhaeva, L.A. and Mutvei, H. 2006. Ultrastructural and chemical comparison between gladii in living coleoids and Aptian coleoids from Central Russia. Acta Universitatis Carolinae—Geologica 49: 83–93.
Doguzhaeva, L.A., Mapes, R.H., and Mutvei, H. 1999. A Late Carboniferous spirulid coleoid from the southern mid-continent (USA): Shell wall ultrastructure and evolutionary implications. In: F. Oloriz and F.J. Rodrigez-Tovar (eds.), Advancing Research on Living and Fossil Cephalopods, 47–57. Kluver Academic Plenum Publishers, New York.
Doguzhaeva, L.A., Mapes, R.H., and Mutvei, H. 2004. Occurrence of ink in Paleozoic and Mesozoic coleoids (Cephalopoda). Mitteilungen des Geologisch-Paläontologischen Institutes der Universität Hamburg 88: 145–156.
Doguzhaeva, L.A., Mapes, R.H., and Mutvei, H. 2007a. A Late Carboniferous coleoid cephalopod from the Mazon Creek Lagerstätte (USA), with a radula, arm hooks, mantle tissues, and ink. In: N.H. Landman, R.A. Davis, and R.H. Mapes (eds.), Cephalopods Present and Past. New Insights And Fresh Perspectives, 121–143. Springer, The Netherlands.
Doguzhaeva, L.A., Mapes, R.H., and Mutvei, H. 2007b. The preservation of body tissues, shell, and mandibles in the ceratitid ammonoid Austrotrachyceras (Late Triassic), Austria. In: N.H. Landman, R.A. Davis, and R.H. Mapes (eds.), Cephalopods Present and Past: New Insights and Fresh Perspectives. 221–238. Springer, Dordrecht.
Doguzhaeva, L.A., Mapes, R.H., and Mutvei, H. 2010a. Evolutionary patterns of Carboniferous coleoid cephalopods based on their diversity and morphological plasticity. In: K. Tanabe, Y. Shigeta, T. Sasaki, and H. Hirano (eds.), Cephalopods Present and Past, 171–180. Tokai University Press, Tokyo.
Doguzhaeva, L.A., Mapes, R.H., Bengtson, S., and Mutvei, H. 2010b. A radula and associated cephalic remains of a Late Carboniferous coleoid from Idaho, USA. Ferrantia 59: 37–50.
Doguzhaeva, L.A., Summesberger, H., and Mutvei, H. 2006. A unique Upper Triassic coleoid from the Austrian Alps reveals proostracum and jaw ultrastructure. Acta Universitatis Carolinae—Geologica 49: 69–82.
Doguzhaeva, L.A., Summesberger, H., Mutvei, H., and Brandstaetter, F. 2007c. The mantle, ink sac, ink, arm hooks and soft body debris associated with the shells in Late Triassic coleoid cephalopod Phragmoteuthis from the Austrian Alps. Palaeoworld 16: 272–284. Crossref
Donovan, D.T. and Boletzky, S.V. 2014. Loligosepia (Cephalopoda: Coleoidea) from the Lower Jurassic of the Dorset coast, England. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 273: 45–63.
Donovan, D.T. and Toll, R.B. 1988. The gladius in coleoid (Cephalopoda) evolution. In: M.R. Clarke and E.R. Trueman (eds.), The Mollusca, Paleontology and Neontology of Cephalopods 12, 89–101. Academic Press, San Diego.
Eilberg, R.G., Zuckerberg, D.A., and Person, N.J.P. 1975. Mineralization of invertebrate cartilage. Calcified Tissue Reseach 19: 85–90. Crossref
Engeser, T. and Clarke, M.R. 1988. Cephalopod hooks, both recent and fossil. In: M.R. Clarke and E.R. Trueman (eds.), The Mollusca, Paleontology and Neontology of Cephalopods 12, 133–151. Academic Press, San Diego. Crossref
Fuchs, D. and Iba, Y. 2015. The gladiuses in coleoid cephalopods: homology, parallelism, or convergence? Swiss Journal of Palaeontology 134: 187–197. Crossref
Fuchs, D. and Weis, R. 2008. Taxonomy, morphology and phylogeny of Lower Jurassic loligosepiid coleoids (Cephalopoda). Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 249: 93–112. Crossref
Fuchs,D., Iba, Y., Tischlinger, H., and Klug, C. 2015.·The locomotion system of Mesozoic Coleoidea (Cephalopoda) and its phylogenetic significance. Lethaia 49: 433–454. Crossref
Fuchs, D., Keupp, H., and Schweigert, G. 2013. First record of a complete arm crown of the Early Jurassic coleoid Loligosepia (Cephalopoda). Paläontologische Zeitschrift 87: 431–435. Crossref
Fuchs, D., Klinghammer, A., and Keupp, H. 2007. Taxonomy, morphology and phylogeny of plesioteuthidid coleoids from the Upper Jurassic (Tithonian) Plattenkalks of Solnhofen. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 245: 239–252. Crossref
Galfetti, T., Bucher, H., Ovtcharova, M., Schaltegger, U., Brayard A., Brühwiller, T., Goudemand, N., Weissert, H., Hochuli, P.A., and Guodun, F.C.K. 2007. Timing of the Early Triassic carbon cycle perturbations inferred from new U-Pb ages and ammonoid biochronozones. Earth and Planetary Science Letters 258: 593–604. Crossref
Gordon, M. Jr. 1971. Primitive squid gladii from the Permian of Utah. US Geological Survey Professional Paper 750C: C34–C38.
Ianiro, A., Di Giosia, M., Fermani, S., Samorì, C., Barbalinardo, M., Valle, F., Pellegrini, G., Biscarini, F., Zerbetto, F., Calvaresi, M., and Falini, G. 2014. Customizing properties of β-chitin in squid pen (gladius) by chemical treatments. Marine Drugs 12: 5979–5992. Crossref
Ibánez, C. and Keyl, F. 2010.
Cannibalism in cephalopods. Reviews in Fish
Biology and Fisheries 20: 123–136. Crossref
Jamil, T., Lias, K., Norsila, D., and Syafinaz, N.S. 2014. Assessment of heavy metal contamination in squid (Loligo spp.) tissues of Kedah-Perlis waters, Malaysia. The Malaysian Journal of Analytical Sciences 18: 195–203.
Jattiot, R., Brayard, A., Fara, E., and Charbonnier, S. 2015. Gladius-bearing coleoids from the Upper Cretaceous Lebanese Lagerstätten: Diversity, morphology, and phylogenetic implications. Journal of Paleontology 89:148–167. Crossref
Jeletzky, J.A. 1965. Taxonomy and phylogeny of fossil Coleoidea (= Dibranchiata). Geological Survey of Canada Papers 65: 72–76 Crossref.
Jeletzky, J.A. 1966. Comparative morphology, phylogeny and classification of fossil Coleoidea. Paleontological Contributions, University of Kansas, Mollusca 7: 1–16.
Jenks, J., Guex, J., Hungerbühler, A., Taylor, D., and Bucher, H. 2013. Ammonoid biostratigraphy of the early Spathian Columbites parisianus zone (Early Triassic) at Bear Lake Hot Springs, Idaho, USA. New Mexico Museum of Natural History and Science 61: 268–283.
Klug, C., Fuchs, D., Schweigert, G., Röper, M., and Tischlinger, H. 2015. New anatomical information on arms and fins from exceptionally preserved Plesioteuthis (Coleoidea) from the Late Jurassic of Germany. Swiss Journal Palaeontology 134: 245–255. Crossref
Klug, C., Schweigert, G., and Dietl, G. 2010. A new Plesioteuthis with beak from the Kimmeridgian of Nusplingen (Germany). Ferrantia 59: 73–77.
Kröger, B., Vinther, J., and Fuchs, D. 2011. Cephalopod origin and evolution: a congruent picture emerging from fossils, development and molecules. BioEssays 33: 602–613. Crossref
Kulicki, C. and Szaniawski, H. 1972. Cephalopod arm hooks from the Jurassic of Poland. Acta Palaeontologica Polonica 17: 379–426.
Kummel, B. 1954. Triassic stratigraphy of southeastern Idaho and adjacent areas. U.S. Geoogical Survey Professional Paper 254: 165–194.
Lukeneder, A. and Harzhauzer, M. 2004. The Cretaceous coleoid Dorateuthis syriaca Woodward: morphology, feeding habits and phylogenetic implications. Annalen des Naturhistorischen Museums in Wien 106 A: 213–225.
Miramand, P. and Guary, J.C. 1980. High concentrations of some heavy metals in tissues of the Mediterranean octopus. Bulletin of Environmental Contamination and Toxicology 24: 783–788. Crossref
Naef, A. 1922. Die fossilen Tintenfische. Ein paläozoologische Monographie. 322 pp. Fischerverlag, Jena.
Nesis, K.N. 1992. The diamondback squid, Thysanoteuthis rhombus Troschel, 1857: a “living fossil”? Ruthenica 2: 91–103.
d’Orbigny, A. 1835–1843. Mollusques. In Voyage dans l’Amérique Méridionale 50: 1–758.
Rieber, H. 1970. Breviconoteuthis breviconus (Reis), ein Phragmoteuthide aus der Mittleren Trias des Monte San Giorgio (Kanton Tessin Schweiz). Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 7: 415–421.
Romano, C., Kogan, I., Jenks, J., Jerjen, I. and Brinkmann, W. 2013. Saurichthys and other fossil fishes from the late Smithian (Early Triassic) of Bear Late Country (Idaho, USA), with a discussion of saurichthyid palaeogeography and evolution. Bulletin of Geosciences 87: 543–570.
Rosenkrantz, A. 1946. Hook-bearing cephalopods from the East Greenland Permian. Meddelelser fra Dansk Geologisk Forening 11: 160–161.
Schweigert, G. and Fuchs, D. 2012.
First record of a true coleoid cephalopod from the Germanic Triassic
(Ladinian). Neues Jahrbuch für Geologie und
Paläontologie, Abhandlungen 266: 19–30.
Crossref
Smith, J.D., Plues, L., Heyraud, M., and Cherry, R.D. 1984. Concentrations of the elements Ag, Al, Ca, Cd, Cu, Fe, Mg, Pb and Zn, and the radionuclides 210Pb and 210Po in the digestive gland of the squid Nototodarus gouldi. Marine Environmental Research 13: 55–68. Crossref
Starobogatov, Y.I. 1983. The system of the Cephalopoda [in Russian]. In: J.I. Starobogatov and K.N. Nesis (eds.), Sistematika i ekologia golovonogih molluskov, 4–7. Zoological Institute USSR Academy of Sciences, Leningrad.
Strugnell, J., Jackson, J., Drummond, A.J., and Cooper, A. 2006. Divergence time estimates for major cephalopod groups: evidence from multiple genes. Cladistics 22: 89–96. Crossref
Vecchione, M., Young, R.E., Donovan, D.T., and Rodhouse, P. 1999. Reevaluation of coleoid cephalopod relationships based on modified arms in the Jurassic coleoid Mastigophora. Lethaia 32: 113–118. Crossref
Vecchione, M. and Young, R.E. 2008. Myopsida Naef, 1916. The Tree of Life Web Project (version 21 April 2008) http://tolweb.org/Myopsida/52670/2008.04.21
Young, R.E., Vecchione, M., and Donovan, D. 1998. The evolution of coleoid cephalopods and their present biodiversity and ecology. South African Journal of Marine Scienses 20: 393–420. Crossref
Weaver, P.G., Doguzhaeva, L.A., Lawver, D.R., Tacker, R.C., and Ciampaglio, C.N. 2011. Characterization of organics consistent with ß-chitin preserved in the Late Eocene cuttlefish Mississaepia mississippiensis. PLoS ONE 6 (11): e28195. Crossref
Acta Palaeontol. Pol. 63 (2): 341–355, 2018
https://doi.org/10.4202/app.00393.2017