

New partial dentaries of amphitheriid mammal Palaeoxonodon ooliticus from Scotland, and posterior dentary morphology in early cladotherians
ELSA PANCIROLI, ROGER B.J. BENSON, and RICHARD J. BUTLER
Panciroli, E., Benson, R.B.J., and Butler, R.J. 2018. New partial dentaries of amphitheriid mammal Palaeoxonodon ooliticus from Scotland, and posterior dentary morphology in early cladotherians. Acta Palaeontologica Polonica 63 (2): 197–206.
We describe two partial dentaries of mammals from the Middle Jurassic of Scotland. They belong to the early cladotherian Palaeoxonodon ooliticus. These dentaries comprise the first specimen of P. ooliticus ever found—although its significance was initially unrecognised so it remained undescribed until now—and the most recently discovered specimen, found during fieldwork in 2017. The new specimen preserves part of the coronoid process of the dentary, previously unknown for P. ooliticus, demonstrating the presence of a deep masseteric fossa, with a prominent crest enclosing the fossa anteriorly, and a masseteric foramen, located in the masseteric fossa on the buccal surface of the dentary. On the lingual surface, the mandibular foramen is offset from the Meckel’s sulcus, and positioned below the alveolar plane. These morphologies allow an updated analysis of the phylogenetic position of P. ooliticus, confirming a sister-taxa relationship between Palaeoxonodon and Amphitherium. The position of the mandibular foramen, and the slight extension of the masseteric fossa into the body of the dentary are new autapomorphies for Palaeoxonodon.
Key words: Mammalia, Cladotheria, Palaeoxonodon ooliticus, Jurassic, Bathonian, Kilmaluag Formation, UK, Scotland.
Elsa Panciroli [elsa.panciroli@ed.ac.uk], School of Geosciences, University of Edinburgh, Grant Institute, Kings Buildings, Edinburgh, EH9 3FE, UK; National Museum of Scotland, Chambers St, Edinburgh, EH1 1JF, UK.
Roger B.J. Benson [roger.benson@earth.ox.ac.uk], Department of Earth Sciences, University of Oxford, South Parks Road, Oxford, OX1 3AN, UK.
Richard J. Butler [r.butler.1@bham.ac.uk], School of Geography, Earth and Environmental Sciences, University of Birmingham, Birmingham, B15 2TT, UK.
Received 23 October 2017, accepted 17 February 2018, available online 20 March 2018.
Copyright © 2018 E. Panciroli et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
The Middle Jurassic was a key period in the evolutionary history of mammals. Recent research suggests a global radiation of early mammals beginning in the Early Jurassic, and entailing large increases in diversity and phenotypic disparity (Luo 2007; Close et al. 2016). This gave rise to docodontans, shuotheriids, australosphenidans, multituberculates, and cladotherians in the Early to Middle Jurassic (Luo et al. 2002; Kielan-Jaworowska et al. 2004). Continued fossil discoveries are critical to understanding this radiation. Furthermore, Cladotheria includes Theria, the clade comprising living marsupials and placentals and their stem lineages. Therefore, early cladotherian fossils provide information on the ancestral morphologies of the group, from which the more derived structures of extant therians evolved.
The early cladotherian Palaeoxonodon was originally reported based on isolated teeth from the Middle Jurassic of England (Freeman 1976a, b, 1979; Sigogneau-Russell 2003), and its detailed phylogenetic affinities remained uncertain. An almost complete dentary from the Isle of Skye, Scotland (NMS G. 2015.17.1), described by Close et al. (2016), revealed that the lower tooth row of Palaeoxonodon shows a gradient of variation in several dental characters that were used to delimit species in previous taxonomy. Based on these new observations, a previously erected Palaeoxonodon species, Palaeoxonodon freemani, and the genus Kennetheridium (Sigogneau-Russell 2003), were both synonymised with the type species of Palaeoxonodon, Palaeoxonodon ooliticus. This dentary also provided additional characters for a more informative phylogenetic analysis, supporting previous work that suggested Palaeoxonodon is the sister taxon to Amphitherium (Close et al. 2016; but see also Averianov et al. 2015 for an alternative hypothesis).
Here, we report two further specimens of Palaeoxonodon from the Isle of Skye. A newly discovered specimen, NMS G.2017.37.1, was found during fieldwork in 2017. Although it is not as complete as that found by Close et al. (2016), it preserves the base of the coronoid process and adjacent regions of the dentary. These features were missing from previous specimens and allow additional characters to be scored for phylogenetic analysis. Another specimen, NMS G.1992.47.123, was recovered during fieldwork by Michael Waldman and Robert J.G. Savage in 1973. It had previously been mentioned as an undescribed pantotherian by Clemens (1986) and Evans et al. (2006). The discovery of this specimen pre-dates all other known specimens of Palaoxonodon, including the holotype of P. ooliticus (Freeman 1976a). However, for reasons unknown, it was never described. After being transferred posthumously from the late RJG Savage of the University of Bristol to the National Museum of Scotland, it was rediscovered by EP in the collections at NMS in 2016.
Institutional abbreviations.—NHMUK PV, Natural History Museum, London, UK; NMS, National Museums Scotland, Edinburgh, UK.
Other abbreviations.—MPTs, most parsimonious trees; TBR, tree bisection and re-connection.
Material and methods
The two specimens described here are both in the collections at the NMS: NMS G.1992.47.123 and NMS G.2017.37.1. Both were collected from the Kilmaluag Formation of the Great Estuarine Group (Bathonian, Middle Jurassic) of the Strathaird Peninsula, north of Elgol, Isle of Skye, Scotland. NMS G.2017.37.1 was collected at Cladach a’ Ghlinne, whereas NMS G.1992.47.123 was collected approximately 0.8 km south of Cladach a’ Ghlinne along the coastline. The Kilmaluag Formation comprises dolomitised blue-grey limestone, interbedded with calcareous siltstone and shale. The presence of freshwater gastropods and bivalves (Viviparus, Neomidon, and Unio), and abundant freshwater ostracods, such as Darwinula and Theriosynoecum, support the environmental interpretation that this was a freshwater lagoon environment (Andrews 1985; British Geological Survey 2011). The strata yielding the new specimens are frequently referred to as “the vertebrate beds”. Also known from this formation are fossil crocodylomorphs, squamates, choristoderes, salamanders, fishes, mammals, and tritylodontids, as well as rare archosaur remains (Waldman and Savage 1972; Evans et al. 2006; Close et al. 2016; Panciroli et al. 2017a, b).
Micro-CT scan data were obtained for NMS G.1992.47.123 using the PCT scanner built in-house at the University of Edinburgh, School of Geosciences Experimental Geoscience Facility. The scanner comprises a Feinfocus 10-160kV dual transmission/reflection source, Perkin Elmer XRD0822 amorphous silicon x-ray flat panel detection, and terbium doped gadolinium oxy-sulfide scintillator. Data acquisition software was written in-house, and scans were reconstructed using Octopus 8.7 software. The scan resolution for this scan is 20 µm. NMS G.2017.37.1 micro-CT scan data were obtained at Cambridge Biotomography Centre (http://www.cbc.zoo.cam.ac.uk/) using a Nikon Metrology XT H 225 ST micro-CT scanner. The scan resolution for this scan is 7.64 µm. All microCT scans were digitally reconstructed, image processed, and measured using Mimics 19.0 at the National Museum of Scotland. Digital reconstructions are available in SOM (Supplementary Online Material available at http://app.pan.pl/SOM/app63-Panciroli_etal_SOM.pdf).
Lower molar terminology follows that by Sigogneau-Russell (2003) with the following amendments: the talonid cusp is renamed the hypoconid, and the crest running from the hypoconid to the metaconid is the cristid obliqua (as in Davis 2011); the paraconal sulcus is renamed the hypoflexid (as in Close et al. 2016); cusp e is not present in Palaeoxonodon.
Phylogenetic analysis was carried out using TNT 1.5, with settings as in Close et al. (2016): all analyses were carried out by first using the new technology search with default command settings, and stipulating that the shortest tree was to be found 10 times. This was followed by TBR branch swapping on the recovered most parsimonious trees (MPTs), and then calculating a strict consensus. Jackknife resampling statistics were calculated using 1000 replicates, and values are displayed as absolute frequencies. Bremer support values were obtained by TBR branch swapping on the MPTs, incrementally increasing suboptimal trees from one to eight steps (see SOM for TNT script and amended data matrix).
Systematic palaeontology
Mammalia Linnaeus, 1758
Cladotheria McKenna, 1975
Amphitheriidae Owen, 1846
Genus Palaeoxonodon Freeman, 1976b
Type species: Palaeoxonodon ooliticus Freeman, 1976b; Forest Marble Formation, Bathonian, Kirtlington, Oxfordshire, England, UK.
Palaeoxonodon ooliticus Freeman, 1976b
Fig. 1.
Holotype: NHMUK PV M36508, right lower molar (Freeman 1976b: pl. 17: 1–4).
Type locality: Kirtlington, Oxfordshire, England, UK.
Type horizon: Forest Marble Formation, Bathonian (Mammal Bed).
Material.—NMS G. 2015.17.10, partial left dentary referred to this species by Close et al. (2016); NMS G.1992.47.123 and NMS G.2017.37.1, two partial left dentaries, described here; from the Kilmaluag Formation (Bathonian), Isle of Skye, Scotland, UK.
Specimens originally referred to Palaeoxonodon freemani, Palaeoxonodon sp., Kennetheridium leesi, and Kennetheridium sp. (Close et al. 2016). Right lower molars: NHMUK PV MJ.59, MJ.196, ?MJ.242, M36507 (Freeman 1979: pl. 17: 1–4, but now lost), MJ.197, MJ.213, MJ.388, MJ.530, MJ.569, MJ.593, MJ.715, ?M44303, MJ.702, MJ.290, MJ.117, MJ.379, ?MJ.430, MJ.514, MJ.428, MJ.515, MJ.532, MJ.801; left lower molars: NHMUK PV ?MJ.200, MJ.236, MJ.619, MJ.639, MJ.849, ?MJ.220, ?MJ.350, ?MJ.618, MJ.626, MJ.628, MJ.657, MJ.701, MJ.727, MJ.825, MJ.827, MJ.837, ?M51823, MJ.802, MJ.53, MJ.746, MJ.289, ?MJ.518, MJ.824, MJ.214, MJ.846); right upper molars: NHMUK PV MJ.146, MJ.524, MJ.749, MJ.754, MJ.792, M36512, MJ.99, MJ.238, MJ.241, MJ.458, MJ.817, M36532. More questionably, Sigogneau-Russell (2003), also referred right upper molars: NHMUK PV MJ.44, MJ.231, MJ.669, M34994, MJ.32, MJ.506, MJ.508, MJ.512bis, MJ.742, MJ.743, MJ.788; and left upper molars NHMUK PV MJ.137, MJ.392, MJ.436, MJ.636, M36504, M36530, M36526, MJ.25, MJ.294, MJ.627. All from the Forest Marble Formation (Bathonian), Kirtlington, Oxfordshire, England, UK.
Emended diagnosis (modified from Close et al. 2016).—Tribosphenic cladotherian mammal differing from dryolestoids in possessing an elongate lower molar talonid with hypoconid placed buccal to the midline of the crown, well-developed upper molar metacone, and winglike parastylar region. Differs from all other non-zatherian cladotherians in possession of a mesiolingual lower molar cingulid. Differs from Amphitherium in possession of five molars instead of six or seven, the more lingual position of the hypoconid, and presence of an entocristid partially enclosing an incipiently basined talonid (sensu Sigogneau-Russell 2003). Differs from Nanolestes in possessing less cuspidate upper molars with weaker development of cusp B, and relatively taller and less procumbent lower molar paraconid. Differs from Arguimus in the presence of a more fully triangulated trigonid of m1 with a larger paraconid, more spirelike lower molar cusps, absence of a distinct cusp e, and larger metaconid on ultimate molar. Resembles Amphitherium and Nanolestes but differs from zatherians in the absence of a distinct cusp e on lower molars, absence of hypoconulid, and buccally positioned metacone. Relatively slender dentary unlike dryolestids, with a deep masseteric fossa and coronoid scar as in other amphitheriids. The masseteric fossa intrudes slightly into the body of the dentary below the ultimate molar, unlike all other early cladotherians. The posterior opening of the mandibular canal, the mandibular foramen, is offset from the Meckel’s sulcus, unlike Amphitherium or Peramus. The posteriormost mental foramen is positioned in the canine/incisor region of the dentary.
Description.—NMS G.1992.47.123 is a fragment of left dentary containing m2, m3, and m4 in position, with roots, and the anterior part of the alveolus for m5 (Fig. 1). The molar crowns were broken and detached from their roots, and the latter remained inside the dentary fragment. The crowns have been restored to their original positions digitally to facilitate description and comparison. The dentary fragment measures 2.5 mm in anteroposterior length, 0.62 mm in width at the widest point, and the dentary dorsoventral depth below m3 is 1.3 mm (for tooth measurements see Table 1).
Table 1. Dental measurements (in mm) for Palaeoxonodon
ooliticus Freeman, 1976b, NMS G.1992.47.123 and NMS
G.2017.37.1.
Measurements follow Sigogneau-Russell (2003) and
Close et al. (2016).
|
Specimen |
Tooth |
Total length |
Total width |
Total crown height |
Trigonid |
Talonid |
Protoconid height |
||
|
length |
angle |
length |
cusp to metacristid |
||||||
|
NMS G.1992.47.123 |
m2 |
0.88 |
0.62 |
0.91 |
0.63 |
60° |
0.25 |
0.19 |
0.21 |
|
m3 |
0.88 |
0.60 |
0.98 |
0.61 |
49° |
0.27 |
0.11 |
0.31 |
|
|
m4 |
0.89 |
0.64 |
|
0.61 |
50° |
0.27 |
0.12 |
|
|
|
NMS G.2017.37.1 |
m3 |
1.00 |
0.78 |
|
0.75 |
47° |
0.25 |
0.15 |
|
|
m4 |
0.89 |
0.84 |
|
0.60 |
58° |
0.40 |
0.15 |
|
|
All three crowns of NMS G.1992.47.123 are well preserved, except for the following damage: the protoconid, metaconid, and paraconid cusp tips of m4 are missing; the hypoconid tip and tip of cusp f of m3 are missing; and the hypoconid and buccal apical portion of the protoconid are missing from m2 (Fig. 1). The surface of the paraconid of m3 is damaged, missing enamel on the lingual surface of the paraconid, metaconid, and cingulid, and some of the anterobuccal enamel surface is also missing apically on the protoconid. The cristid obliqua of m3 is well preserved, revealing a distinct incipient talonid basin in occlusal view (Fig. 1C), defined lingually by the entocristid. The incipient talonid basin of m4 is well preserved, with a clear entocristid on the lingual edge.
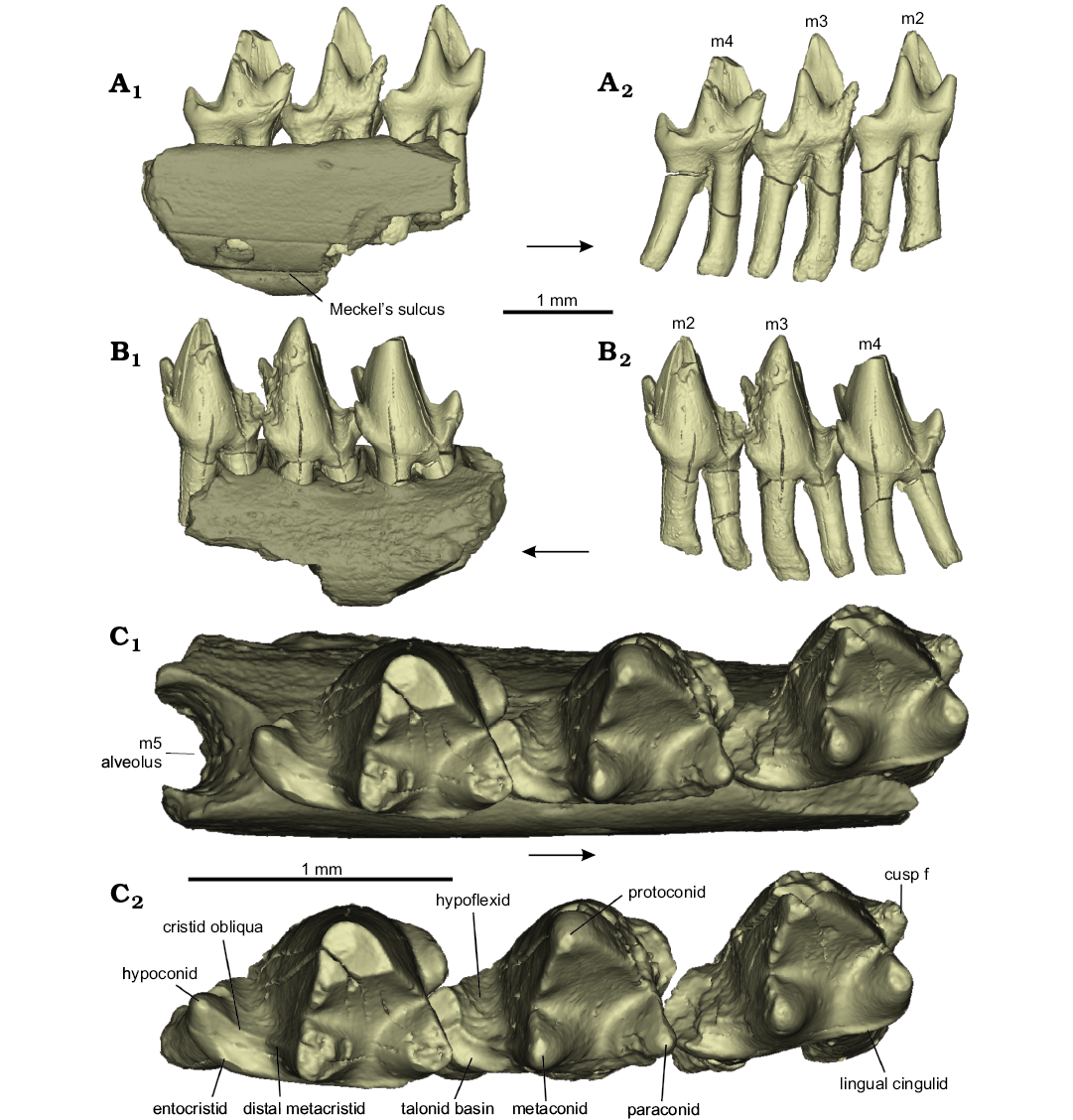
Fig. 1. Amphitheriid mammalian Palaeoxonodon ooliticus Freeman, 1976b (NMS G.1992.47.123) from the Kilmaluag Formation, Bathonian, in lingual (A), buccal (B), and occlusal (C) views; partial left dentary (A1–C1); dentition only (A2–C2). Arrows indicate anterior direction.
NMS G.2017.37.1 is a fragment of left dentary 7.2 mm long with an approximate dorsoventral depth of 1.6 mm below m3 (Fig. 2) (for tooth measurements see Table 1). The teeth m3 and m4 are preserved in position. Most of m2 is also preserved in position, but was fragmented prior to discovery. A fragment of the m1 root is also present. The alveolus for m5 is present but empty (Fig. 2B1). The cusp tips are missing from both m3 and m4, but the protoconid, paraconid, and metaconid positions are clear on both molars, as are the hypoflexid, hypoconid, and incipient talonid basin, defined lingually by the entocristid. None of these features are easily identified in m2, in which the hypoconid and hypoflexid are missing, and only the base of the anterior half of the crown is present.
Multiple fractures are present on the dentary, but the general osteology is evident. The Meckel’s sulcus is preserved on the lingual surface of the dentary, extending anteriorly from a point just ventral to the mandibular foramen, up to the fractured region ventral to m2 (Fig. 2C1). The mandibular foramen is present posteriorly on the lingual surface of the dentary, posteroventral to the empty alveolus for m5 and offset from the Meckel’s sulcus. The pterygoid shelf is unclear from the preserved morphology.
The masseteric fossa, located posteriorly on the buccal surface of the dentary, is deep and is defined by ridges anterodorsally and ventrally. A masseteric foramen is located anteroventrally within the masseteric fossa. The anterior margin of the masseteric fossa is well-developed and extends anteriorly, ventral to the alveolus for m5. It extends just slightly into the body of the mandible ventral to m5 (Fig. 2A1, B1).
Stratigraphic and geographic range.—Middle to Late Bathonian, England and Scotland.
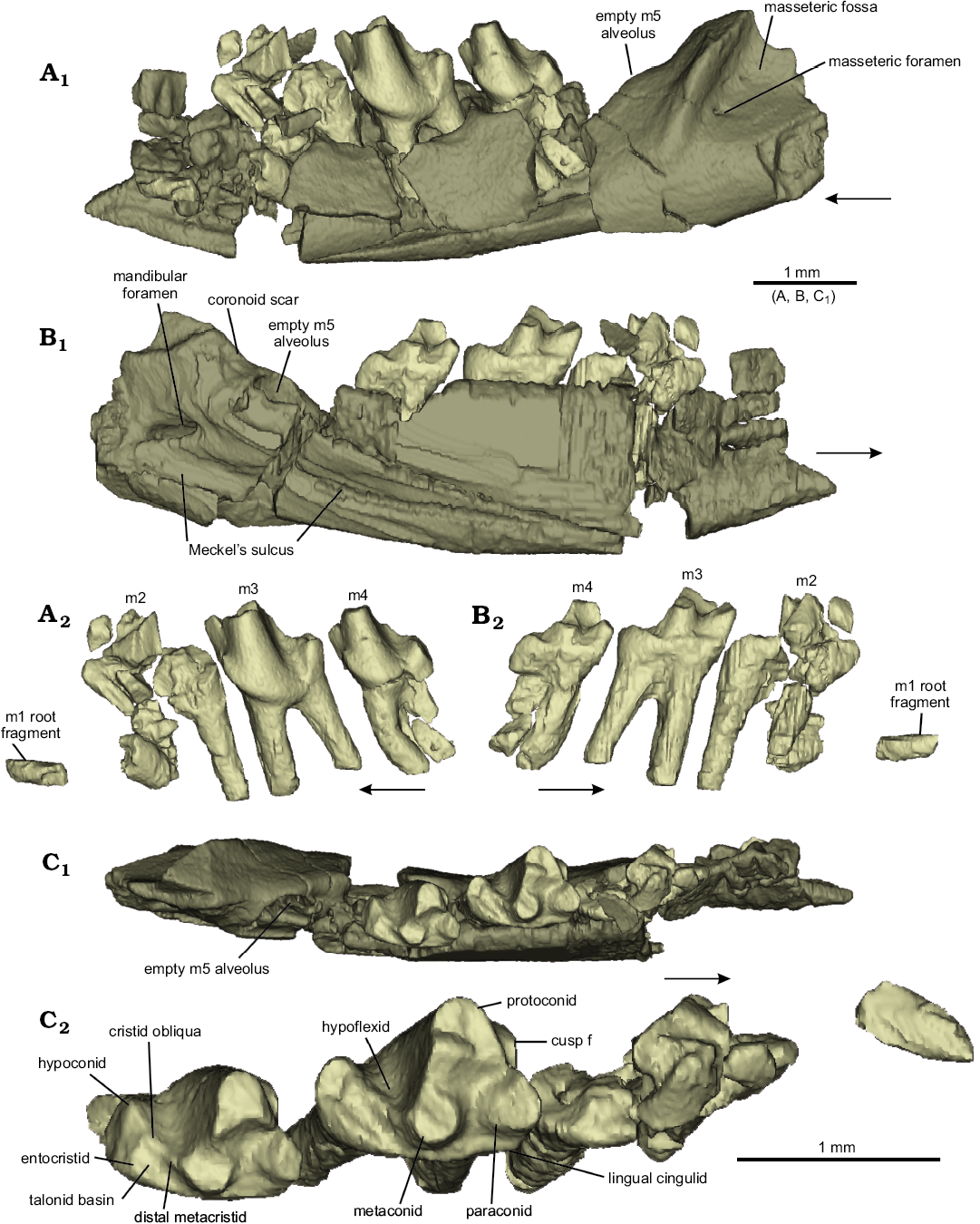
Fig. 2. Amphitheriid mammalian Palaeoxonodon ooliticus Freeman, 1976b (NMS G.2017.37.1) from the Kilmaluag Formation, Bathonian, in lingual (A), buccal (B), and occlusal (C) views; partial left dentary (A1–C1), dentition only (A2–C2). Arrows indicate anterior direction.
Phylogenetic analysis
We reran the phylogenetic analysis of Close et al. (2016), which was based on the matrix of Zhou et al. (2013), updating scores for characters of the posterior portion of the dentary present in NMS G.2017.37.1. These included the position of the mandibular foramen, the morphology of the anterior margin of the masseteric fossa, the presence of the masseteric foramen, and the presence of a coronoid scar (see Appendix 1 for newly scored characters and SOM for full matrix and TNT script). We also re-evaluated and amended six characters present in NMS G. 2015.17.1 that were either not scored by, or that we interpreted differently from, Close et al. (2016). These include confirming the absence of Crompton’s groove, the absence of a premolar or precanine diastema, confirming the presence of the hypoconid, and the morphology of the postcanine roots. Additionally, we re-evaluated and rescored six characters present in Amphitherium, Peramus, and Arguimus (see Appendix 1 for justification for rescoring, and Zhou et al. 2013 for full character list).
The phylogenetic analysis recovered 5376 most parsimonious trees. Despite the additional character scores, we found no change in the relationships between Palaeoxonodon, Amphitherium, and Peramus from previous analyses, with Palaeoxonodon and Amphitherium placed in a sister-group relationship. These relationships have weak Bremer support values of 1, and this clade (Amphitheriidae) forms an unresolved polytomy with Nanolestes, Arguimus, and the clade including Kielantherium + Aegialodon + Theria, as in Close et al. (2016).
Five autapomorphies were recovered for Palaeoxonodon by our phylogenetic analysis, the first three of which were already identified by Close et al. (2016), and the last two of which are identified for the first time here: (i) the posteriormost mental foramen being positioned in the canine/incisor region of the dentary (more anteriorly than in Amphitherium or Peramus); (ii) the absence of a mesio-lingual cingular cuspule e; and (iii) an absent/weak ectoflexus in the second upper molar; (iv) the slight extension of the masseteric fossa onto the body of the mandible; and (v) the position of the mandibular foramen offset from the Meckel’s sulcus. Palaeoxonodon and Amphitherium share synapomorphic characters in the presence of a narrow labial stylar shelf of the penultimate upper molar, and the more labial position of the metacone relative to the paracone. Palaeoxonodon, Amphitherium, and Peramus share the following synapomorphies: the convergence of the Meckel’s sulcus with the ventral border of the mandible; and possessing open rooted postcanines.
Discussion
Most previous cladistic analyses support a sister group relationship between Palaeoxondon and Amphitherium (Prothero 1981; Close et al. 2016; but see Averianov et al. 2015 for an alternative view). In these previous analyses this was based on (i) the large talonid (accounting for one-third of molar length), (ii) possessing a buccally positioned principal hypoconid, (iii) roots that are mostly equal in diameter along their length except for a slight narrowing distally, and (iv) the possession of five premolars with (v) moderately well-developed lingual cingulids.
Close et al. (2016) identified the variably present cuspule on the cristid obliqua (mesoconid) in their specimen NMS G. 2015.17.1 as a feature shared with the holotype NHMUK PV M36508 (and two other specimens according to Sigogneau-Russell 2003: NHMUK PV MJ530 and NHMUK PV MJ618) from the Forest Marble Formation. The mesoconid is not present in either of our specimens, NMS G.1992.47.123 or NMS G.2017.37.1. Therefore we support the interpretation of this character as variable among specimens of Palaeoxonodon.
Close et al. (2016) also suggested that the different dental characters used to erect the species P. freemani and Kennetheridium leesi on the basis of isolated teeth (Sigogneau-Russell 2003) are the morphological differences between tooth positions in the same tooth row of their specimen, relegating the latter two taxa to junior subjective synonyms of P. ooliticus. This character variation includes differences in talonid length, prominence of the cingulid, size of the paraconid, and height and cusp sharpness of the trigonid, among different molars in the same individual. Similarly, we see some of this variation in NMS G.1992.47.123: for example the paraconid is much smaller in m3 and m4 than in m2, and slopes at a shallower angle in m2; and the cingulid is more prominent in m2 than m3 or m4. We can add that the entocristid is more prominent in m2, and that the protoconid is broader both anteroposteriorly and buccolingually in m4. This adds to the evidence for variation along the tooth row, and emphasises the need for caution in erecting new species based on isolated teeth.
A posterior portion of the dentary including the base of the coronoid process, masseteric fossa, and mandibular foramen are preserved in NMS G.2017.37.1 and have not previously been preserved in any other specimen of Palaeoxonodon. The preserved morphology suggests a sloping coronoid process, as in other early cladotherians. Strong masseteric ridges and a deep masseteric fossa are also present, as in other early cladotherians including other amphitheriids such as Amphitherium (Butler and Clemens 2001), other peramurids, dryolestids, and paurodontids (Martin 1999; Kielan-Jaworowska et al. 2004; Davis 2012). Unlike paurodontids, the dentary is relatively slender. The preserved morphology of NMS G.2017.37.1 suggests that a pterygoid shelf may have been present on the lingual surface of the dentary; however, its extent is not clear from this specimen. The pterygoid shelf is strongly protruding in many other early cladotherians (Martin 1999; Kielan-Jaworowska et al. 2004). The anterior position of the posteriormost mental foramen, in the canine/incisor region of the dentary, is recovered here as an autapomorphy of Palaeoxonodon. While there is a possibility that a more posterior foramen may have been present in the fractured portion of the dentary below p1 in NMS G. 2015.17.1, we find no evidence for this at present, pending the recovery of new specimens.
Close et al. (2016) identified a possible coronoid scar in NMS G. 2015.17.1, which we can confirm in NMS G.2017.37.1. This is absent in Peramus according to previous analyses (Clemens and Mills 1971; Davis 2012), but was coded as present by Zhou et al. (2013). We have rescored this character as absent in Peramus in our analysis. The coronoid scar is present in Amphitherium (Butler and Clemens 2001) and many other early cladotherians, including the zatherian Nanolestes (Martin 2002). However, it is absent in another zatherian, Arguimus (Lopatin and Averianov 2006). Close at al. (2016) also inferred a lower incisor count of four based on comparison to Amphitherium. Although we cannot verify this feature using NMS G.1992.47.123 or NMS G.2017.37.1, we consider it likely to have been present, as this character state is common among early Cladotheria. As such, we have used it in our reconstruction (Fig. 3C).
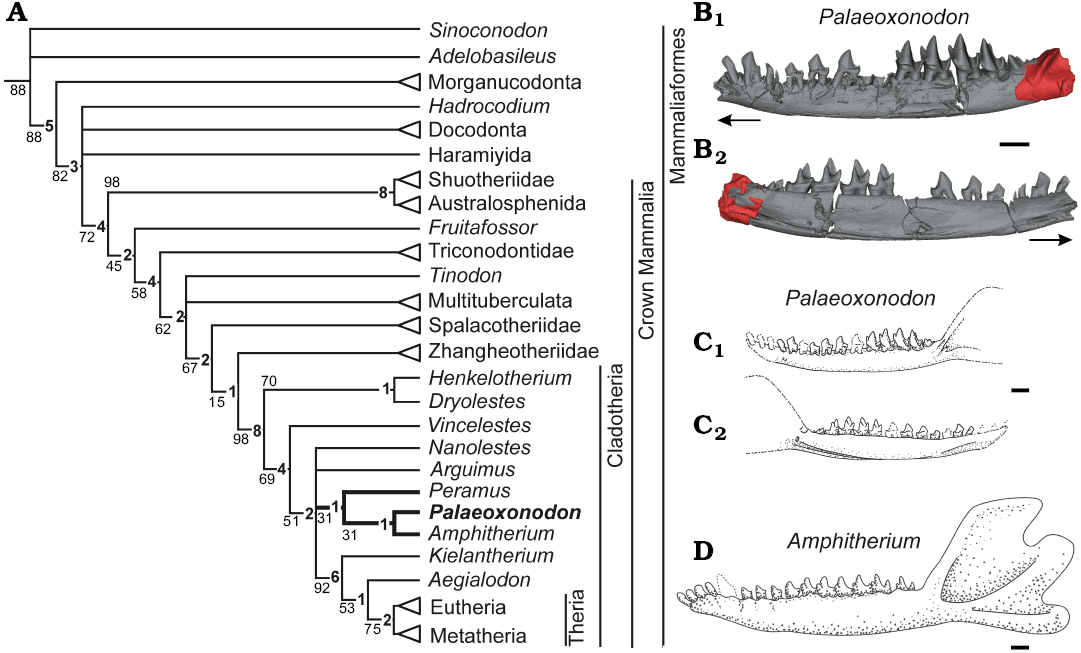
Fig. 3. Parsimony analysis and reconstructions of dentary of Palaeoxonodon ooliticus (B, C), with Amphitherium (D) for comparison. A. Equally weighted parsimony analysis with additional mandibular characters for the early cladotherian Palaeoxonodon ooliticus, and rescored and updated matrix from Close et al. 2016 (adapted from Zhou et al. 2013). Jackknife resampling statistics and Bremer values (bold) are shown adjacent to individual nodes. B. Composite of NMS G. 2015.17.1 (grey) and coronoid of NMS G.2017.37.1 (red) in buccal (B1) and lingual (B2) views (arrows indicate anterior direction). Data for NMS G. 2015.17.1 from Close et al. (2016). C. Reconstruction of jaw in buccal (C1) and lingual (C2) views. D. Reconstruction based on composite created by EP from NHMUK PV M.36822 and OUMNH J.20075 in buccal view. Scale bars 1 mm.
The presence of a Meckel’s sulcus is common among stem mammals. In NMS G.2017.37.1, the mandibular foramen is preserved, indicating that Palaeoxonodon had a Meckel’s sulcus that was offset from the foramen, a feature not shared with either Amphitherium (Butler and Clemens 2001) or Peramus (Clemens and Mills 1971; Davis 2012). The mandibular foramen in Palaeoxonodon also does not appear to be confluent with the pterygoid shelf, unlike in Amphitherium and Peramus, although damage to this part of the dentary in NMS G.2017.37.1 makes this interpretation uncertain. The position of the mandibular foramen ventral to the alveolar plane, seen in Palaeoxonodon and Amphitherium, is also shared with other early cladotherians, and the basal metatherian Sinodelphys. The clade comprising Palaeoxonodon + Amphitherium + Peramus is in a polytomy with Nanolestes, the clade comprising Kielantherium + Aegialodon + Theria, and Arguimus. Arguimus shares few dentary character states with Palaeoxonodon. Arguimus has no Meckel’s sulcus (the absence of Meckel’s sulcus is an autapomorphy of Arguimus; see Lopatin and Averianov 2006) and no coronoid scar. Vincelestes also has no Meckel’s sulcus or coronoid scar (Bonaparte and Rougier 1987). While other analyses have recovered the relationship of Vincelestes as closer to Theria than either Peramus or Amphitherium (Kielan-Jaworowska et al. 2004; Bonaparte 2008), our results, like those of Close et al. (2016) and Zhou et al. (2013) place Vincelestes as a sister group to the polytomy comprising Palaeoxonodon + Amphitherium + Peramus, Nanolestes, Arguimus, and Kielantherium + Aegialodon + Theria.
The phylogenetic affinities of Palaeoxondon have been contested due to the presence of an “incipient basin” (Sigogneau-Russell 1999, 2003) on the talonid, referred to as the incipient talonid basin herein, and the variable presence of a cuspule on the cristid obliqua. In an alternative hypothesis, Averianov et al. (2015) recovered a close relationship between Amphibetulimus and Palaeoxonodon, placing them in a clade with Nanolestes. This clade was found to be phylogenetically distinct from, and crownward to, Amphitherium (Averianov et al. 2015). Support for this relationship with Nanolestes, rather than with Amphitherium and Peramus was based on the absence of a “retromolar space”, the proportional size of the stylocone relative to the metacone in the upper molars, and absence of a coronoid scar. We now confirm the coronoid scar is in fact present in Palaeoxonodon. The analysis by Averianov et al. (2015) also included Mozomus, which was found to form a polytomy with Zatheria (including Peramus and Arguimus), and the clade comprising Palaeoxonodon + Nanolestes + Amphibetulimus. Neither Mozomus nor Amphibetuliumus were included in our analysis and so we did not test the relationships between these taxa and other early cladotherians.
Characters of the talonid basin have previously been used to argue for a close affinity between Palaeoxonodon and Peramus, to the exclusion of Amphitherium (Freeman 1976b, 1979; Kielan-Jaworowska et al. 2004). However, the presence of plesiomorphies such as the principal talonid cusp (hypoconid) with no embrasure for occlusal contact with the metacone of the upper molar may suggest Palaeoxonodon is more stemward than Peramus, and indeed several authors have suggested that Palaeoxonodon and the morphologically similar Amphitherium are sister-taxa, occuring stemward of Peramus (Sigogneau-Russell 2003; Davis 2011, 2012). Close et al. (2016) suggested that the “incipient” talonid basin of Palaeoxonondon and the talonid basin of Peramus could either be homoplastic, or a synapomorphy of the clade Palaeoxonodon + Amphitherium + Peramus, and this currently cannot be resolved. We agree with Close et al. (2016) that the phylogenetic analyses carried out in their paper and herein cannot resolve this unambiguously.
Palaeoxonodon is the only taxon in our analysis in which the mandibular foramen is located in the pterygoid fossa and offset from the Meckel’s sulcus. In Amphitherium, the foramen is aligned with the posterior end of the Meckel’s sulcus, as it is for the most closely related taxa among early Cladotheria (Butler and Clemens 2001; Kielan-Jaworowska et al. 2004). This is a condition seen in crown Marsupialia and some basal metatherians, such as Didelphodon (Fox and Naylor 1986; Wilson et al. 2016), but not in more crownward Metatheria. However, there is some damage to the posterolingual portion of the dentary in NMS G.2017.37.1. More complete, undamaged specimens may shed further light on the placement of the mandibular foramen. The very slight extension of the anterior crest of the masseteric fossa into the body of the dentary is not shared with any other taxon in this analysis, except the extant Oryctolagus (European rabbit). However the more extreme extension of the anterior crest of the masseteric fossa, where it extends below the ultimate pre-molar or first molar, is commonly seen in multituberculates (e.g., Gambaryan and Kielan-Jaworowska 1995). This may suggest Palaeoxonodon had a more extensively developed superficial masseteric muscle than other early cladotherians. Without knowing the position and morphology of the condylar process, which is crucial for determining the in-lever for the dentary (Gill et al. 2014), it is not possible to make informed inferences about the biomechanics of mastication in this genus. This would require more complete mandibles that are amenable for muscle reconstruction, as shown in other studies (Lautenschlager et al. 2017).
The presence of a masseteric foramen (also referred to as the labial mandibular foramen), located in the masseteric fossa on the buccal side of the dentary, is a newly identified feature in Palaeoxonodon. A deep masseteric fossa with a distinct masseteric foramen is a feature shared with peramurans, but is also present in numerous taxa across different lineages, including other zatherians, triconodontids, and basal metatherians and eutherians (Davis 2012). In Palaeoxonodon the foramen is not as large as in some specimens of Peramus, but it quickly merges with the mandibular canal inside the dentary, posteroventrally to the position of m5. The morphology of this foramen in Palaeoxonodon is similar to that described by Davis (2012) for the specimens of Peramus that lack an enlarged masseteric foramen: the mandibular canal is enlarged mesial to the masseteric fossa, and is separated by only a thin wall of bone (Davis 2012: 813–814). Davis suggests the variability in the size of the masseteric foramen in specimens of Peramus may be attributed to post-depositional crushing and infill. Until further specimens of Palaeoxondon are found possessing the posterior portion of the dentary, it is not possible to say if the size of this feature similarly varies in this taxon.
Conclusions
Newly described specimens of Palaeoxonodon ooliticus from the Isle of Skye add valuable new information for morphological comparison, and new data for phylogenetic analysis. The additional features of the masseteric fossa, and the position of the mandibular foramen identified in NMS G.2017.37.1, while not altering the phylogenetic relationships between Palaeoxonodon and its nearest relatives among cladotherians, are new autapomorphies for this genus. They provide additional character scores for future analysis, reducing the amount of missing information for this taxon.
The “re-discovery” of NMS G.1992.47.123 provides a cautionary tale for palaeontologists who delay in the publication of specimens. By not pursuing identification and description of this specimen, an opportunity was missed to name a second Mesozoic mammal from Scotland, an important taxon for understanding the morphology of early Cladotheria. This more complete specimen would also have provided a holotype with some variation in the tooth row, perhaps preventing the erection of taxa that are now suggested to be junior synonyms. This is an informative lesson from the palaeontological history of Mesozoic mammals. NMS G.1992.47.123 also contributes to the description of rich Scottish Mesozoic mammal specimens that have, until now, remained undescribed, and therefore unrecognised scientifically.
Although a relatively large number of upper teeth and lower dentitions from Palaeoxonodon are now known, they have not yet been found in association, and postcrania have yet to be identified. This provides a goal for further discoveries in the rich microvertebrate beds of the Isle of Skye in the coming years.
Acknowledgements
We are grateful to the John Muir Trust and Scottish Natural Heritage, who allow us access and give permission for prospecting and collection on the Isle of Skye. Thanks to Roger Close (University of Birmingham, UK) for sharing his data and digital visualisations of the previously recovered Palaeoxonodon specimen, and many helpful discussions, including with regard to TNT. We are also indebted to Stig Walsh (Natural Sciences Department at National Museums Scotland, Edinburgh, UK) for access to the specimens, fieldwork collaboration, and his advice and discussions. Our reviewers, Zhe-Xi Luo (University of Chicago, USA) and Alexander Averianov (Russian Academy of Sciences, Petersburg, Russia), provided invaluable input for the improvement of this manuscript, for which we are also grateful. EP is funded by NERC (NE/L002558/1). NMS G.2017.37.1 was scanned using funding from the European Union’s Horizon 2020 research and innovation programme 2014–2018, under grant agreement 677774 (European Research Council starting grant: TEMPO) to RBJB. Participation of RJB in fieldwork was supported by a Marie Curie Career Integration Grant (grant number 630123).
References
Andrews, J.E. 1985. The sedimentary facies of a late Bathonian regressive episode: the Kilmaluag and Skidiburgh Formations of the Great Estuarine Group, Inner Hebrides, Scotland. Journal of the Geological Society 142: 1119–1137. Crossref
Averianov, A., Martin, T., Lopatin, A., and Krasnolutskii, S. 2015. Stem therian mammal Amphibetulimus from the Middle Jurassic of Siberia. Paläontologische Zeitschrift 89: 197–206. Crossref
Bonaparte, J.F. 2008. On the phylogenetic relationships of Vincelestes neuquenianus. Historical Biology 20: 81–86. Crossref
Bonaparte, J.F. and Rougier, G.W. 1987. Mamíferos del Cretácico Inferior de Patagonia. IV Congresso Latinomericano de Paleontologia 1: 343–359.
British Geological Survey 2011. Stratigraphic Framework for the Middle Jurassic Strata of Great Britain and the Adjoining Continental Shelf: Research Report RR/11/06. 177 pp. British Geological Survey, Keyworth.
Butler, P.M. and Clemens, W.A. 2001. Dental morphology of the Jurassic holotherian mammal Amphitherium, with a discussion of the evolution of mammalian post-canine dental formulae. Palaeontology 44: 1–20. Crossref
Clemens, W.A. 1986. On Triassic and Jurassic mammals. In: K. Padian (ed.). The Beginning of the Age of Dinosaurs, 237–246. Cambridge University Press, Cambridge.
Clemens, W.A. and Mills, J.R.E. 1971. Review of Peramus tenuirostris. Bulletin of the British Museum (Natural History) Geology 20: 89–113.
Close, R.A., Davis, B.M., Walsh, S., Woloniewicz, A.S., Friedman, M., and Benson, R.B.J. 2016. A lower jaw of Palaeoxonodon from the Middle Jurassic of the Isle of Skye, Scotland, sheds new light on the diversity of British stem therians. Palaeontology 59: 155–169. Crossref
Davis, B.M. 2011. Evolution of the tribosphenic molar pattern in early mammals, with comments on the “dual-origin” hypothesis. Journal of Mammalian Evolution 18: 227–244. Crossref
Davis, B.M. 2012. Micro-computed tomography reveals a diversity of peramuran mammals from the Purbeck Group (Berriasian) of England. Palaeontology 55: 789–817. Crossref
Evans, S., Barrett, P., Hilton, J., Butler R.J., Jones, M.E.H., Liang, M.-M., Parrish, J.C., Rayfield, E.J., Sigogneau-Russell, D., and Underwood, C.J. 2006. The Middle Jurassic vertebrate assemblage of Skye, Scotland. In: P. Barrett and S. Evans (eds), Proceedings of the Ninth Symposium on Mesozoic Terrestrial Ecosystems and Biota, 36–39. Natural History Museum, London.
Freeman, E. 1976a. A mammalian fossil from the Forest Marble (Middle Jurassic) of Dorset. Proceedings of the Geologists’ Association 87: 231–235.
Freeman, E. 1976b. Mammal teeth from the Forest Marble (Middle Jurassic) of Oxfordshire, England. Science 194: 1053–1055. Crossref
Freeman, E. 1979. A Middle Jurassic mammal bed from Oxfordshire. Palaeontology 22: 135–166.
Fox, R.C. and Naylor, B.G. 1986. A new species of Didelphodon Marsh (Marsupialia) from the Upper Cretaceous of Alberta, Canada: palaeobiology and phylogeny. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 172: 357–380.
Gambaryan, P.P. and Kielan-Jaworowska, Z. 1995. Masticatory musculature of Asian taeniolabidoid multituberculate mammals. Acta Palaeontologica Polonica 40: 45–80.
Gill, P.G., Purnell, M.A., Crumpton, N., Brown, K.R., Gostling, N.J., Stampanoni, M., and Rayfield, E.J. 2014. Dietary specializations and diversity in feeding ecology of the earliest stem mammals. Nature 512: 303–305. Crossref
Kielan-Jaworowska, Z., Cifelli, R.L., and Luo, Z.-X. 2004. Mammals from the Age of Dinosaurs: Origins, Evolution, and Structure. 630 pp. Columbia University Press, New York. Crossref
Lautenschlager, S., Gill, P., Luo, Z.-X., Fagen, M.J. and Rayfield, E.J. 2017. Morphological evolution of the mammalian jaw adductor complex. Biological Reviews 92: 1910–1940. Crossref
Linnaeus, C. 1758. Systema naturae per regna tria naturae: secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis, 824 pp. Tenth edition. Laurentius Salvius, Stockholm.
Lopatin, A. and Averianov, A. 2006. Revision of a pretribosphenic mammal Arguimus from the Early Cretaceous of Mongolia. Acta Palaeontologica Polonica 51: 339–349.
Luo, Z.-X. 2007. Transformation and diversification in early mammal evolution. Nature 450: 1011–1019. Crossref
Luo, Z.-X., Kielan-Jaworowska, Z., and Cifelli, R.L. 2002. In quest for a phylogeny of Mesozoic mammals. Acta Palaeontologica Polonica 47: 1–78.
Martin, T. 1999. Dryolestidae (Dryolestoidea, Mammalia) aus dem Oberen Jura von Portugal. Abhandlungen der senckenbergischen naturforschenden Gesellschaft 550: 1–119.
Martin, T. 2002. New stem-lineage representatives of Zatheria (Mammalia) from the Late Jurassic of Portugal. Journal of Vertebrate Paleontology 22: 332–348. Crossref
McKenna, M.C. 1975. Toward a phylogenetic classification of the Mammalia. In: W.P. Luckett and F.S. Szalay (eds), Phylogeny of the primates, 21–43. Plenum Publishing Corporation, New York. Crossref
Owen, R. 1846. History of British Fossil Mammals and Birds. 560 pp. John Van Voorst, London.
Panciroli, E., Benson, R.B.J., and Walsh, S. 2017a. The dentary of Wareolestes rex (Megazostrodontidae): a new specimen from Scotland and implications for morganucodontan tooth replacement. Papers in Palaeontology 3: 373–386. Crossref
Panciroli, E., Walsh, S., Fraser, N., Brusatte, S. L., and Corfe, I. 2017b. A reassessment of the postcanine dentition and systematics of the tritylodontid Stereognathus (Cynodontia, Tritylodontidae, Mammaliamorpha), from the Middle Jurassic of the UK. Journal of Vertebrate Paleontology 37 (5): 1351448. Crossref
Prothero, D.R. 1981. New Jurassic mammals from the Como Bluff, Wyoming, and the interrelationships of non-tribosphenic theria. Bulletin of the American Museum of Natural History 176: 281–168.
Sigogneau-Russell, D. 1999. Réévaluation des Peramura (Mammalia, Cladotheria) sur la base de nouveaux spécimens du Crétacé inférieur d’Angleterre et du Maroc. Geodiversitas 21: 93–127.
Sigogneau-Russell, D. 2003. Holotherian mammals from the Forest Marble (Middle Jurassic of England). Geodiversitas 25: 501–537.
Waldman, M. and Savage, R.J.G. 1972. The first Jurassic mammal from Scotland. Journal of the Geological Society of London 128: 119–125. Crossref
Wilson, G.P., Ekdale, E.G., Hoganson, J.W., Calede, J.J., and Vander Linden, A. 2016. A large carnivorous mammal from the Late Cretaceous and the North American origin of marsupials. Nature Communications 7: 13734. Crossref
Zhou, C.F., Wu, S., Martin, T., and Luo, Z.-X. 2013. A Jurassic mammaliaform and the earliest mammalian evolutionary adaptations. Nature 500: 163–167. Crossref
Appendix 1
Newly scored characters for Palaeoxonodon—the following six characters (from Zhou et al. 2013) were newly scored based on morphology present in specimen NMS G.2017.37.1:
12. Coronoid bone (or its attachment scar): (1) absent to (0) present
13. Location of the mandibular foramen (posterior opening of the mandibular canal): ? to (1) in the pterygoid fossa and offset from Meckel’s sulcus (the intersection of Meckel’s sulcus at the pterygoid margin is ventral and posterior to the foramen).
14. Vertical position of the mandibular foramen: ? to (0) below the alveolar plane.
22. Crest of the masseteric fossa along the anterior border of the coronoid process: ? to (1) present and distinctive.
23. Anteroventral extension of the masseteric fossa: ? to (1) extending anteriorly onto the body of the mandible.
24. Labial mandibular foramen (masseteric foramen) inside the masseteric fossa: ? to (1) present.
Re-scored characters for Palaeoxonodon—we re-scored and added the following characters (from Zhou et al. 2013) for Palaeoxonodon differently from Close et al. (2016):
6. Groove for the replacement dental lamina (Crompton’s groove): (1) absent. Crompton’s groove for the replacement dental lamina, as seen in Morganucodon and some other stemward mammaliaformes, is not present on any of the specimens of Palaeoxonodon ooliticus so far recovered from England or Scotland. We therefore consider it justifiable to score this character as absent.
97. Hypoconid (we designate the distal cingulid cuspule d as the homolog to the hypoconid in the teeth with linear alignment of the main cusps; we assume the cusp to be the hypoconid if there is only a single cusp on the talonid in the teeth with reversed triangulation): (1) present (as distal cusp d, sensu Crompton 1971), elevated above the cingulid level, labially positioned (or tilted in the lingual direction). We score this based on the morphology of all known material so far.
174. Diastema separating the lower first and second premolars (defined as the first and second functioning premolar or premolariform postcanine): (0) absent (gap less than one tooth root for whichever is smaller of the adjacent teeth). No diastema separates the premolars pm1 and pm2, nor is there a diastema pre- or post-canine, in this taxon in any specimens known so far.
182. Enlarged diastema in the lower incisor-canine region (better developed in older individuals): (0) absent; as for 174.
195. Open root end of the postcanines: (1) present; based on NMS G. 2015.17.1.
196. Degrees of root division: (2) two or three complete divided roots. All specimens now recovered confirm that Palaeoxonodon ooliticus has two completely divided roots in all teeth except incisors, and that they are not connected by dentine sheets.
Re-scored characters for Arguimus—we re-scored and added the following characters for Arguimus differently from Close et al. (2016):
13. Location of the mandibular foramen (posterior opening of the mandibular canal): 4 to (?) the location of this structure cannot be confirmed in Arguimus.
14. Vertical position of the mandibular foramen: 1 to (0) below the alveolar plane.
24. Labial mandibular foramen inside the masseteric fossa: ? to (1) present.
26. Posterior-most mental foramen: ? to variable (1, 2, 3, 4).
Rescored characters for Amphitherium and Peramus—we re-scored and added the following characters for Amphitherium and Peramus from Zhou et al. (2013):
12. Coronoid bone (or its attachment scar): (1) absent in Peramus.
195. Open root end of the postcanines: (1) present (for both taxa). This character was previously scored as (2) for Amphitherium and Peramus, which is not an option for this character.
Acta Palaeontol. Pol. 63 (2): 197–206,
2018 https://doi.org/10.4202/app.00434.2017