
New ischnacanthiform jaw bones from the Lower Devonian of Podolia, Ukraine
VICTOR VOICHYSHYN and HUBERT SZANIAWSKI
Voichyshyn, V. and Szaniawski, H. 2018. New ischnacanthiform jaw bones from the Lower Devonian of Podolia, Ukraine. Acta Palaeontologica Polonica 63 (2): 327–339.
Investigation of fish fauna assemblages obtained by dissolution of calcareous rock samples from Early Devonian marine deposits of Podolia revealed new material of ischnacanthiform jaw bones. One family Podoliacanthidae fam. nov. and two new genera and species, Drygantacanthus semirotunda gen. et sp. nov. and Kasperacanthus serratus gen. et sp. nov., are established. The new family is based on one main key feature, the presence of denticle groups of Podoliacanthus type situated on the lingual tooth row. The family comprises three genera, Podoliacanthus, Drygantacanthus gen. nov., and Kasperacanthus gen. nov., as well as one new form undetermined to generic level. Another new form described in open nomenclature displays the remains of the most powerful known jaws among Podolian ischnacanthids known to now. The new forms have diverse main teeth morphology, which probably reflect differentiated hunting methods.
Key words: Acanthodii, Ischnacanthiformes, dentigerous jaw bone, Devonian, Lochkovian, Podolia.
Victor Voichyshyn [victor@smnh.org], State Museum of Natural History NASU, Teatralna Str. 18, 79008, L’viv, Ukraine.
Hubert Szaniawski [szaniaw@twarda.pan.pl], Institute of Paleobiology, Polish Academy of Sciences, ul. Twarda 51/55, 00-818 Warszawa, Poland.
Received 11 January 2018, accepted 19 March 2018, available online 17 May 2018.
Copyright © 2018 V. Voichyshyn and H. Szaniawski. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Fossil remains of Acanthodii are widely distributed within all sections of the marine Lower Devonian deposits in Podolia (Western Ukraine). However, they were usually only mentioned in publications concerning other Devonian fishes (Novitskaya and Obruchev 1964; Miles 1973; Karatajūtė-Talimaa 1978; Denison 1979; Valiukevičius 2000; Voichyshyn 2001, 2010, 2011). Few works, however, regarding the ischnacanthiforms (order Ischnacanthiformes) from the region have been undertaken to date, and referred to mainly in passing (Ørvig 1967; Valiukevičius 2000). Until now only one publication has been devoted to representatives of this group found in Podolia (Voichyshyn and Szaniawski 2012). As a result, only a few forms of ischnacanthiform dentigerous jaw bones from the Tyver Series of the Lower Devonian deposits of the region were known until now. They belong to Podoliacanthus and Ischnacanthiformes indet. The genus Podoliacanthus was described from Ivanie Zolote, Zalishchyky and Ustechko, and has also recently been discovered in Dobrivlany, Gorodok, Ugrynkivtsi (Fig. 1C), and Jagilnytsia Stara (Fig. 1B). It has also been found in the Peremyshlany-1 borehole (Fig. 1A). As a consequence, its stratigraphic distribution is expanded down to the middle part of the Chortkiv Horizon of the Tyver Series (Fig. 2: point 1). In addition, the investigations show that the Podolian marine deposits seem to have a variety of Podoliacanthus-like and related forms. The specimen which had been referred to Ischnacanthiformes fam., gen. et sp. indet. (Voichyshyn and Szaniawski 2012: fig. 8D) still remains as the oldest jaw bone fragment within the Podolian Lower Devonian section, but its stratigraphic position is presently corrected as being within the approximate middle part of the Chortkiv Horizon of the Tyver Series (Fig. 2: point 2).
Several Podoliacanthus forms were described in open nomenclature because they are known from singular specimens only (Voichyshyn and Szaniawski 2012). Owing to that, we have continued the search in different Podolian localities. Examination of the numerous chemically processed samples led to the discovery of new material of ischnacanthiform dentigerous jaw bones. It should be particularly emphasized that the search for ischnacantiform jaw bones by the dissolution of limestone samples shows considerable promise, especially for the study of small adult and juvenile ischnacanthiform specimens.
Institutional abbreviations.—SMNH, State Museum of Natural History, National Academy of Sciences of Ukraine, Lviv, Ukraine; ZPAL, Institute of Paleobiology, Polish Academy of Sciences, Warsaw, Poland.
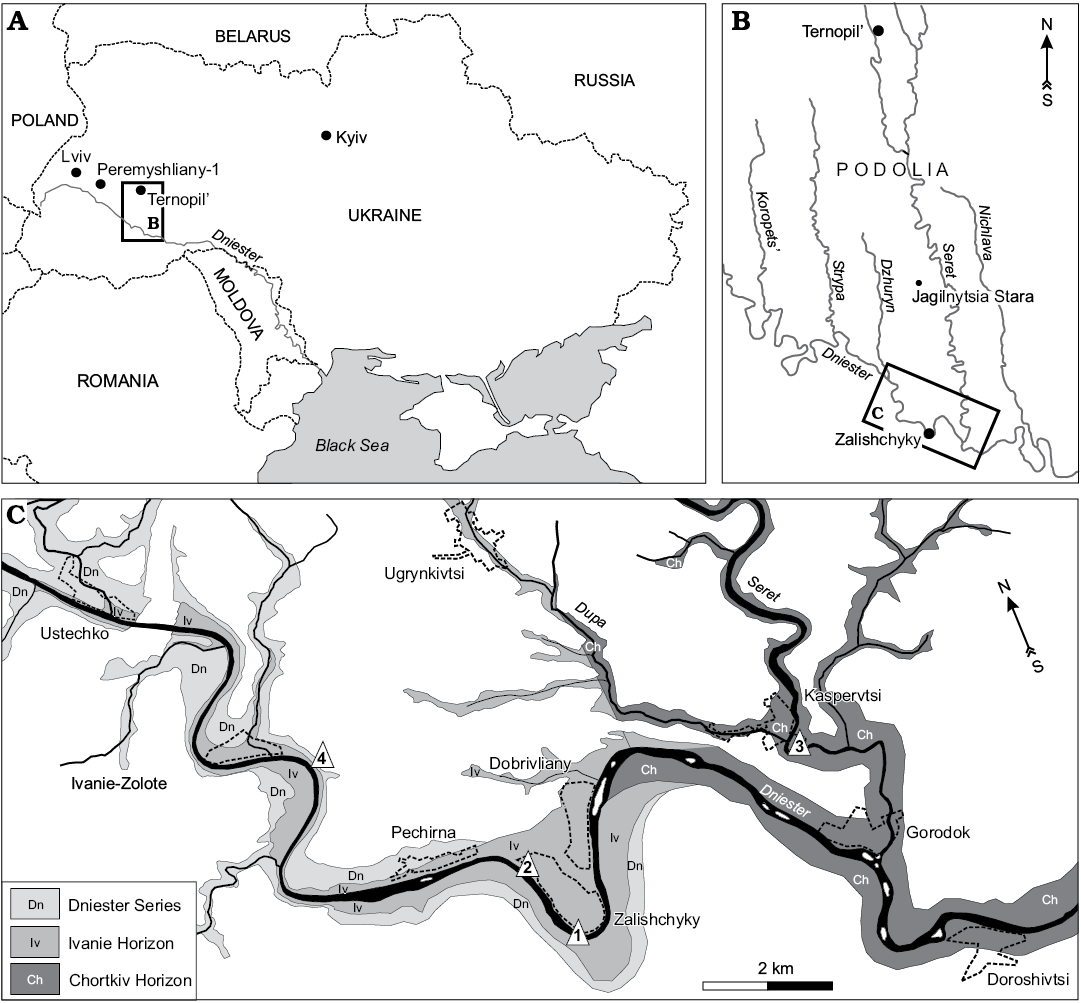
Fig. 1. A. Location of the study area in Ukraine with position of Podolia. B. Map of the studied region. C. Lower Devonian exposures (modified from Voichyshyn and Szaniawski 2012) with points of sampled sections (triangles 1–4).
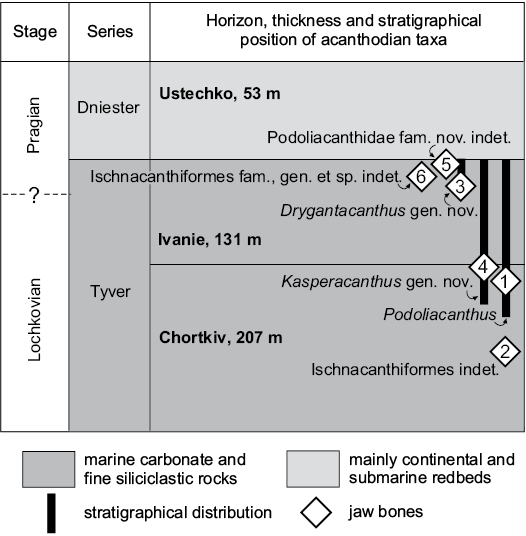
Fig. 2. Simplified stratigraphy of the marine Lower Devonian beds in Podolia showing distribution of Ischnacanthiformes jaw bones (modified from Drygant and Szaniawski 2012; Voichyshyn and Szaniawski 2012).
Material and methods
The jaw bone remains described in this paper are derived from dissolved calcareous rocks collected by Daniel Drygant (SMNH) and Andrzej Baliński (ZPAL) from the Kasperivtsi (Fig. 1C: point 3) and Zalishchyky (Fig. 1C: points 1, 2) vicinities, and by the second author of the paper from the Ivanie Zolote vicinity (Fig. 1C: point 4). The material under description was found in the samples taken from the middle part of the Chortkiv Horizon to the upper part of the Ivanie Horizon of the Tyver Series (Fig. 2).
The samples have been dissolved in 10–15% acetic acid solution. The residuum has been washed, dried, and viewed under binocular microscope. The illustrated fish fossils are glued to the SEM stubs. The SEM photographs were taken in the laboratory of ZPAL using a Philips XL-20 scanning electron microscope.
The terminology used in the paper follows the previous publication devoted to acanthodian jaw bones from Podolia (Voichyshyn and Szaniawski 2012). Nevertheless, since Kasperacanthus gen. nov. jaw bone demonstrates clear limits between “multicuspidate teeth” in the meaning of Ørvig (1967: 147) we accept some additional terms in the description of the genus. The lateral tooth row consists of several “complex-shaped teeth” which include a tooth proper and two denticulated flanges attached to it. Inter-tooth denticles are an integral part of the denticulated flange of the complex tooth. The term “tooth/teeth” remains for tooth/teeth proper. For the “flanges” of tooth proper we use term “ridges” to avoid the confusion of terms in the genus description.
Geological setting
A brief description of the geological setting of Podolian Early Devonian marine deposits has been published by Małkowski et al. (2009), Drygant and Szaniawski (2012), Filipiak et al. (2012), Racki et al. (2012), Szaniawski (2012), and Voichyshyn and Szaniawski (2012). The boundary between the Lochkovian and Pragian in the region is still uncertain (Fig. 1). The most recent review estimated it to be within the upper third of the Ivanie Horizon (Drygant and Szaniawski 2012: fig. 2). Jaw bone fragments from Kasperivtsi (Fig. 1C: point 3) have been found in residuum which contained numerous broken shells of bivalves. The fish fossil assemblage of the sample includes scales of thelodonts (Apalolepis obruchevi Karatajūtė-Talimaa, 1978, Turinia polita Karatajūtė-Talimaa, 1978, and Nikolivia elongata Karatajūtė-Talimaa, 1978), rare fragments of pteraspids and poraspid armour, as well as acanthodian scales, spines, bones, and tooth whorls (with a jaw bone fragment of Podoliacanthus-like form). Scales of the chondrichthyan Seretolepis elegans Karatajūtė-Talimaa, 1978, tesserae of the tesselated heterostracan Corvaspis kingi Woodward, 1934, and some unrecognized fish remains were also discovered.
Systematic palaeontology
Class Acanthodii Owen, 1846
Order Ischnacanthiformes Berg, 1940
Family Podoliacanthidae nov.
Type genus: Podoliacanthus Voichyshyn and Szaniawski, 2012; Early Devonian, Podolia, Ukraine.
Genera included: The type genus, Drygantacanthus gen. nov., and Kasperacanthus gen. nov.
Diagnosis.—Small ischnacanthiforms. Lingual tooth row consists of groups of denticles, with one bigger (central) denticle and usually two or more smaller ones. The denticle groups of the lingual row have a common elongate base which is surrounded by small pores of the vascular canal system. Variable additional dentiform structures frequently, but not always, occur on the medial teeth side, at the base or lower third. These structures are a small robust denticle, a set of these, or a short vertical ridge.
Remarks.—Judging by the diagnostic features of both established families of the order Ischnacanthiformes, Ischnacanthidae and Poracanthodidae, as well as Acritolepidae (family of uncertain affinity), none of them is adequate to accommodate Podoliacanthus-like taxa (see Discussion below). Considering that we are dealing with related but strongly diversified material, which evidently includes more than one genus we are of the opinion that it is necessary to establish a new family.
Stratigraphic and geographic range.—Lower Devonian of Podolia (Ukraine) and Greenland (Denmark).
Genus Drygantacanthus nov.
Type species: Drygantacanthus semirotunda sp. nov., by monotypy, see below.
Species included: The type species only.
Etymology: In honour of Daniel Drygant (SMNH) who found the material under description, and from Latin acanthus, thorn.
Diagnosis.—Medium sized podoliacanthids with dentigerous jaw bone possessing bulky dome-shaped teeth of the lateral tooth row with a set of 4–6 denticles in a semicircular arrangement medially.
Remarks.—Drygantacanthus gen. nov. differs from Podoliacanthus by the possession of more massive teeth of the lateral tooth row, with a posterior and anterior flange of equal size, as well as the different type of additional dentition on their medial side (see the genus diagnosis above). Differences between the genera also occur in terms of the shape of teeth, as well as their parabasal section and inter-tooth pits. Judging by the available material of both genera, another possible difference can be observed in general jaw bone size.
The new genus differs from Kasperacanthus gen. nov. in possessing a different teeth shape within the lateral tooth row, with additional dentition on their medial side, and a solid inter-tooth wall without gaps.
Stratigraphic and geographic range.—Lower Devonian, (Early Pragian?) of Podolia, Ukraine.
Drygantacanthus semirotunda sp. nov.
Figs. 3, 4.
Etymology: After the semicircular-like arrangement of the tooth additional dentition.
Holotype: SMNH BP.1581/5, a jaw bone fragment, 1.63 mm long, with two teeth of the lateral tooth row.
Type locality: Right bank of the Dniester River opposite Zalishchyky, Zastavna (Chernivtsi) region, Podolia, Ukraine; sample 2a/11 (after Drygant 2010); Fig. 1C: point 1.
Type horizon: Upper part of Ivanie Horizon of the Tyver Series, Lower Devonian.
Material.—Except the holotype, two jaw bone fragments with one tooth of the lateral tooth row, paratypes: SMNH BP.1587/3 (1.2 mm long) and SMNH BP.1590/11 (0.65 mm long). Both the specimens are from the same rock sample which was found on the left bank of the Dniester River, north-west Zalishchyky outskirts (Fig. 1C: point 2), Podolia, Ukraine; Upper Lochkovian, Ivanie Horizon.
Diagnosis.—Same as for the genus.
Description.—The teeth bodies are bulky, especially in their lower half, with one medial, vertically oriented, ridge, curved forward (Figs. 3A, C, 4B1, B2). The ridge almost reaches the tooth tip and divides the tooth into two parts. The posterior surface is slightly concave linguolabially, increasing the size of the adjoined inter-tooth pit. The teeth parabasal section generally looks like a circular sector with the central angle of about 140° or less. Anterior part of tooth is convex linguolabially and outlined at its base (under the ridge lower end) by a set of 4–6 denticles in a semicircular arrangement (Figs. 3A, B, 4A, B2, B3). The denticle set is arranged horizontally on the medial tooth side but can rise up on the anterior side. The denticles have a hollow interior (Fig. 3B). A border between crown and base of tooth is visible at its base, beneath the denticle set (Fig. 3E). The teeth are somewhat inclined backwards and have medium-sized anterior and posterior flanges of equal dimension (Figs. 3C, 4A2). One can see at least five broken inter-tooth denticles in the holotype but, judging by the distance between the teeth, there could be twice as many of them (Figs. 3A, B, D, 4A1, A3). The inter-tooth pit is clearly defined. Pores of the vascular canal are hardly visible within the pit area of the holotype, possibly because they are encrusted with silt. There are some sporadic pores on the lateral jaw bone side (Figs. 3D, 4A4).
The lingual ridge is hardly distinguishable along the two preserved teeth of the holotype (Fig. 3C), which has at least one remnant of the lingual denticle group (Fig. 3B, C, E). The remnant looks like a hollow base of multiple denticles, being somewhat elongated at a small angle to the longitudinal jaw bone axis. It is surrounded by pores of the vascular canal system. The appearance of the remnant is similar to the lingual denticle groups of Podoliacanthus (cf. Voichyshyn and Szaniawski 2012: figs. 5F, 8C). The lingual ridge of the specimen BP.1587/3 has no denticle group (Fig. 4A1, A3), most likely because the fragment represents the anterior part of the jaw bone where the denticle groups are usually not developed (cf. Podoliacanthus zychi jaw bone fragments, Voichyshyn and Szaniawski 2012: figs. 4B, C, E, 5A, 6B). The width/height tooth ratio of the species is estimated at 0.8 to 1.0.
The specimen BP.1587/3, in comparison with the others, has some differing features. It does not have the vertical medial ridge on the tooth. Instead, its tooth profile has a slight concavity on both the posterior and anterior sides (Fig. 4A2). The inter-tooth pit part before the tooth in BP.1587/3 specimen reveals numerous pores, up to 65 µm in diameter (Fig. 4A3).
Remarks.—The shape of the new species’ teeth, in occlusal and lingual views, is somewhat similar to those of Grenfellacanthus zerinae Long, Burrow, and Ritchie, 2004 (Long et al. 2004: fig. 1A, C) but this similarity has no systematic importance. The main difference is in presence of nine vertical striae in Grenfellacanthus teeth of the lateral tooth row and in number of denticles in lingual tooth row.
Stratigraphic and geographic range.—North-western and southern vicinities of Zalishchyky. Upper part of the Ivanie Horizon of Tyver Series (Late Lochkovian or Early Pragian).
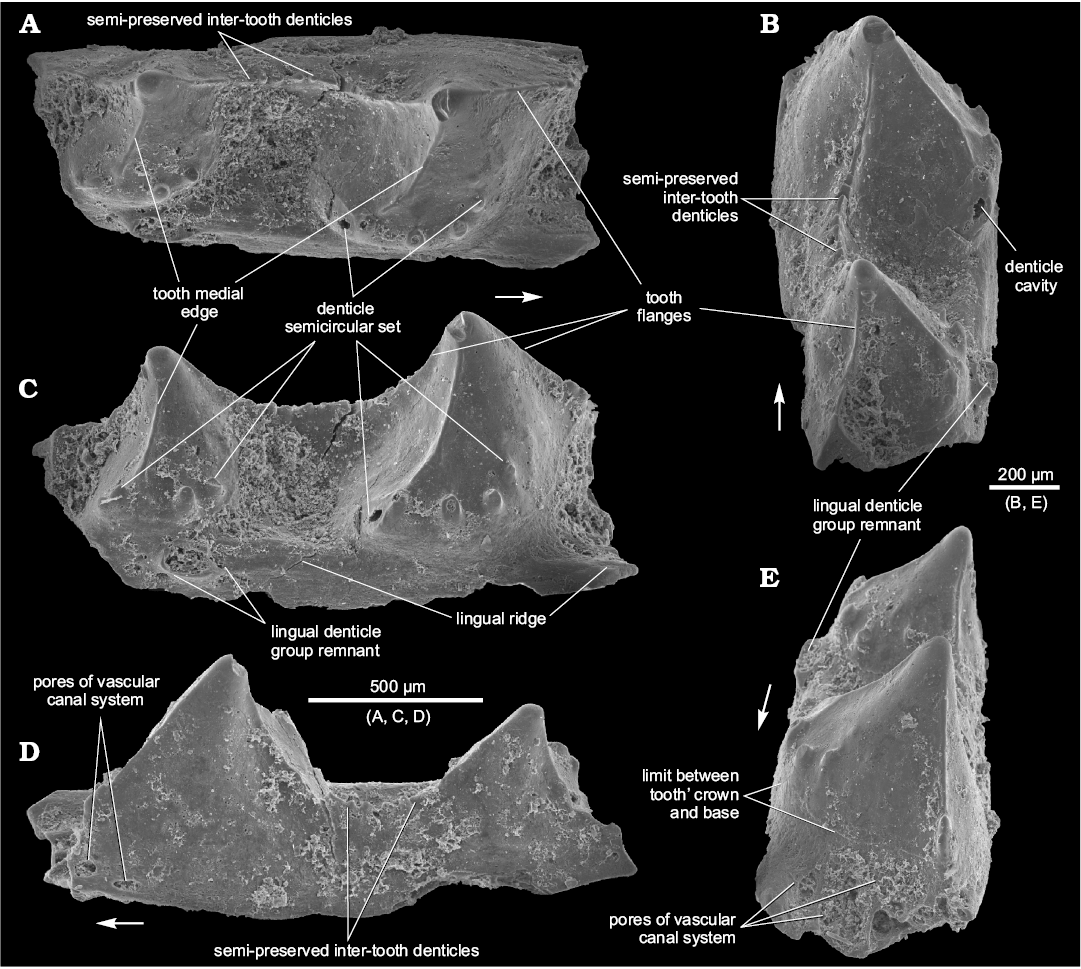
Fig. 3. Ischnacanthiform fish Drygantacanthus semirotunda gen. et sp. nov., the holotype, SMNH BP.1581/5, Dniester River opposite Zalishchyky, Zastavna (Chernivtsi) region, Podolia, Ukraine; Upper part of Ivanie Horizon of the Tyver Series, Lower Devonian. Jaw bone fragment in occlusal (A), postero-occlusal (B), lingual (C), lateral (D), and antero-occlusal (E) views. Arrows indicate rostral direction.
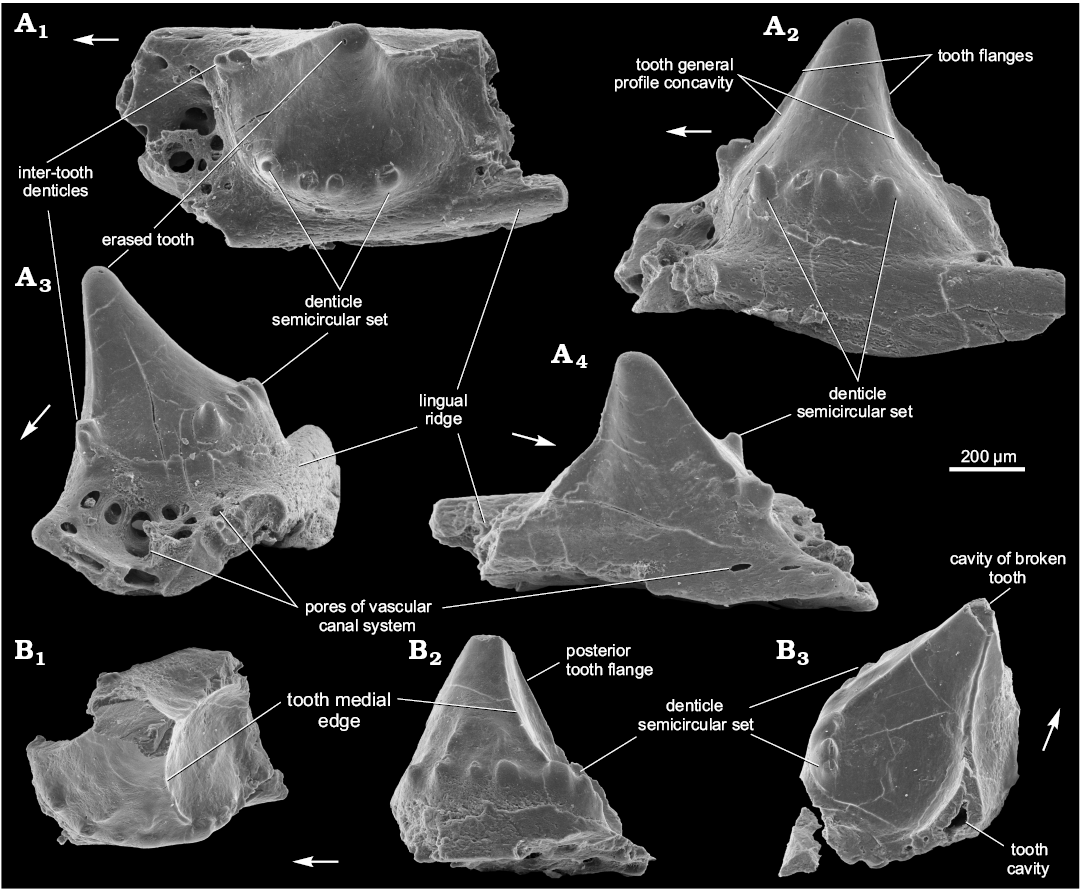
Fig. 4. Ischnacanthiform fish Drygantacanthus semirotunda gen. et sp. nov., Dniester River, north-west Zalishchyky outskirts, Podolia, Ukraine, Upper Lochkovian, Ivanie Horizon; the paratypes, SMNH BP.1587/3 (A) and SMNH BP.1590/11 (B). Jaw bone fragments in occlusal (A1, B1), lingual (A2, B2), anterior (A3), lateral (A4), and posterior (B3) views. Arrows indicate rostral direction.
Genus Kasperacanthus nov.
Type species: Kasperacanthus serratus sp. nov.; see below.
Species included: The type species only.
Etymology: After the type locality name, and from Latin acanthus, thorn.
Diagnosis.—Small- to medium-sized podoliacanthids. The dentigerous jaw bone has caniniform teeth within the halfmoon to circular parabasal section, and a totally smooth tooth surface. The lateral tooth row has gaps which separate the complex-shaped teeth. Relatively large inter-tooth denticles increase in size in the anterior direction within the posterior denticulate flange of the complex tooth.
Remarks.—Kasperacanthus gen. nov. differs from Podoliacanthus in having no dentition attached to the teeth of the lateral tooth row, relatively large inter-tooth denticles, and characteristic gaps between the complex teeth (see the type species description below).
The new genus differs from Drygantacanthus gen. nov. in having differently shaped teeth along the lateral tooth row, without any dentition on their medial side.
Stratigraphic and geographic range.—Lower Devonian (Late Lochkovian or Early Pragian) of Podolia, Ukraine.
Kasperacanthus serratus sp. nov.
Figs. 5, 6.
Etymology: After the serrated profile of lateral tooth rows and inter-teeth denticles of the jaw bone.
Holotype: SMNH BP.1595/14, a jaw bone fragment, 1.2 mm long, with three teeth of the lateral tooth row.
Type locality: Left bank of the Seret River near Kasperivtsi, downstream a dam, Zalishchyky (Ternopil) region, Podolia, Ukraine; sample 3/11г (Daniel Drygant collection); Fig. 1C: point 3.
Type horizon: Middle part of the Chortkiv Horizon, Tyver Series, Lower Devonian.
Material.—Aside from the holotype, three other jaw bone fragments: SMNH BP.1601/14, 1.07 mm long, with one preserved tooth of the lateral tooth row and three teeth remnants, ZPAL P.14/5.5, 2.42 mm long with two teeth, and ZPAL P.14/7.6, 2.81 mm long, with one tooth of the lateral tooth row. The last two specimens have been found in the same rock sample taken from the left bank of the Dniester River, on the East of Ivanie Zolote. Its stratigraphic level corresponds to upper part of the Ivanie Horizon of the Tyver Series, 34 m below the boundary with the Dniester Series, Zalishchyky (Ternopil) region, Podolia, Ukraine (Fig. 2C). The SMNH BP.1601/14 was found in the same residuum as the holotype. It is a part of another jaw bone but possibly belongs to the same fish individual as the holotype.
Diagnosis.—Same as for the genus.
Description.—Teeth of the lateral tooth row are conical in shape and smooth. Their parabasal section is from half-moon to almost circular (Figs. 5A1, B1, 6A1, B1). No additional dentition on the teeth is found. The teeth are hollow (Figs. 5B5, 6A3) and the holotype does not show any ridges on the teeth of the lateral tooth row. However, the teeth of the other specimens, including SMNH BP.1601/14 from the same rock sample as the holotype, have a narrow posterior ridge and traces of an anterior one (Figs. 5B2, B3, 6A2, B2). The teeth are slightly inclined backwards and inwardly relative to the jaw bone axis (Figs. 5A1, A2, B1, B2, 6A1, B1). They are relatively high; their width/height ratio is 0.5–0.7. In the ZPAL P.14/5.5 rare pores of vascular canals can be observed within the lower third of the medial surface (Fig. 6A4) but there are no such pores in the other specimens (Figs. 5A2, A3, B2, B3, 6B2). As in previously described species, there is a crown-base border on the medial side of the tooth (Fig. 6A3, B4).
There is no continuous inter-tooth wall between the teeth. Instead one can see the gaps which emphasize the complex tooth within the lateral tooth row (Fig. 5A1, B1, B3, B5). The complex tooth consists of the tooth proper and two denticulated flanges, long posterior and short anterior (Fig. 5A1, A2, B2, B5). There are 5–9 denticles in the posterior complex tooth flange (Figs. 5A3, B1, B5, 6A2) and 1–2 denticle(s) in the anterior flange (Figs. 5A1, A3, 6A1). The denticles are relatively large and, in the posterior flange, increase in size in anterior direction (Figs. 5A3, 6A1, B1, B2). The biggest have their own posterior and anterior ridges and an even posterior flange (Fig. 6B4). Tips of the denticles seem to be slightly inclined backwards, in the same manner as the teeth (Fig. 6B3). Inter-tooth pits are clearly outlined along the lateral tooth row only (Fig. 5A1, A2, B1, B2).
The lingual ridge is weakly expressed (Fig. 5A1, A2, B1, B2). The denticle groups can be observed on the lingual ridge of SMNH BP.1601/14 (Fig. 5B1–B4) but do not occur in the holotype (Fig. 5A1–A3). Vascular canal pores are situated mainly along the inter-tooth pit perimeter in the specimens from Kasperivtsi. The biggest of them is situated on the posterior part of the base of the teeth (Fig. 5A3, B2). Numerous, though mainly smaller vascular canal pores are scattered over the inter-tooth pit area in the other specimens (Fig. 6A4, B1). The labial side of the jaw bone also possesses vascular canal pores (Fig. 5A4).
The way of growth of the lateral tooth row has engaged our attention. One can see teeth remnants at the posterior (older) part of the SMNH BP.1601/14 (Fig. 5B1, B3, B5). The distance between them at the beginning of the jaw bone growth enlarged significantly but then the growth slowed down. Setting the distance between first two smallest tooth remnants at 1, the next tooth-to-tooth distance ratios are 2.3, 1.9, and 1.2 (the last value is taken from the holotype). The first inter-tooth interval has no denticles. In essence it might be equated to the lateral tooth row gap (Fig. 5B5). The second shows a relatively large gap and the first three inter-tooth denticles which represent the posterior flange of the first complex tooth (Fig. 5B3, B5). At the same time, ZPAL P.14/7.6, which evidently possesses the species’ largest jaw bone has no lateral tooth row gap visible (Figs. 6B2, B3). Possibly, this reflects a change of prey and/or a hunting manner associated with fish size increase.
Remarks.—The described species material resembles the jaw bone of Gomphonchus (Gross 1971: fig. 9H). Nonetheless, judging by the figure in Gross’ publication, Gomphonchus has very high teeth (the width/height tooth ratio is about 0.36) without teeth ridges.
Stratigraphic and geographic range.—Kasperivtsi and Ivanie Zolote vicinities; upper third of the Chortkiv Horizon to upper part of the Ivanie Horizon of the Tyver Series (late Lochkovian to early Pragian?).
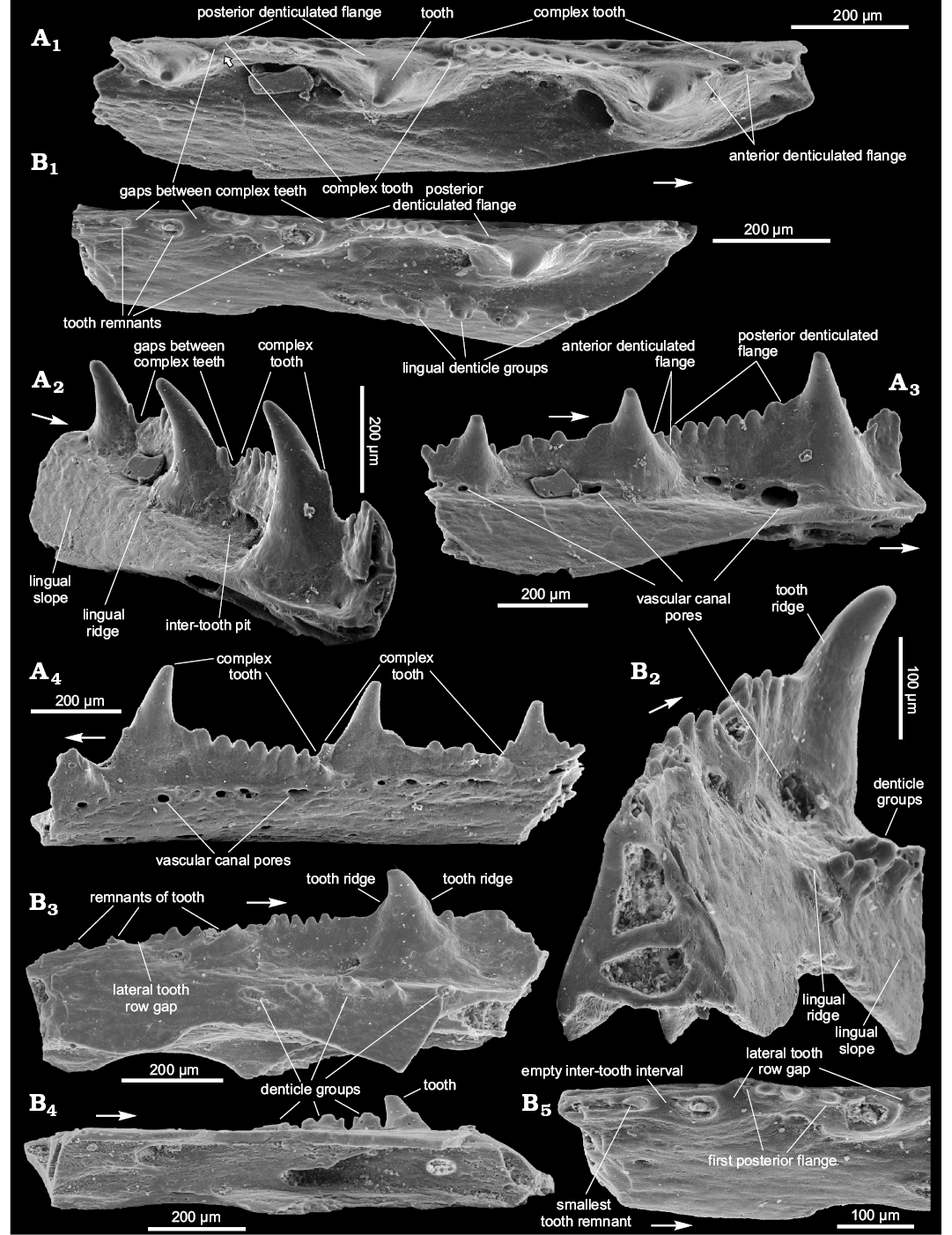
Fig. 5. Ischnacanthiform fish Kasperacanthus serratus gen. et sp. nov., Seret River near Kasperivtsi, downstream a dam, Zalishchyky (Ternopil) region, Podolia, Ukraine; Middle part of the Chortkiv Horizon, Tyver Series, Lower Devonian; the holotype, SMNH BP.1595/14 (A) and paratype, SMNH BP.1601/14 (B). Jaw bone fragments in occlusal (A1, B1, B5), anterolingual (A2), lingual (A3, B3), lateral (A4), posterolingual (B2), and bottom (B4) views. Arrows indicate rostral direction.
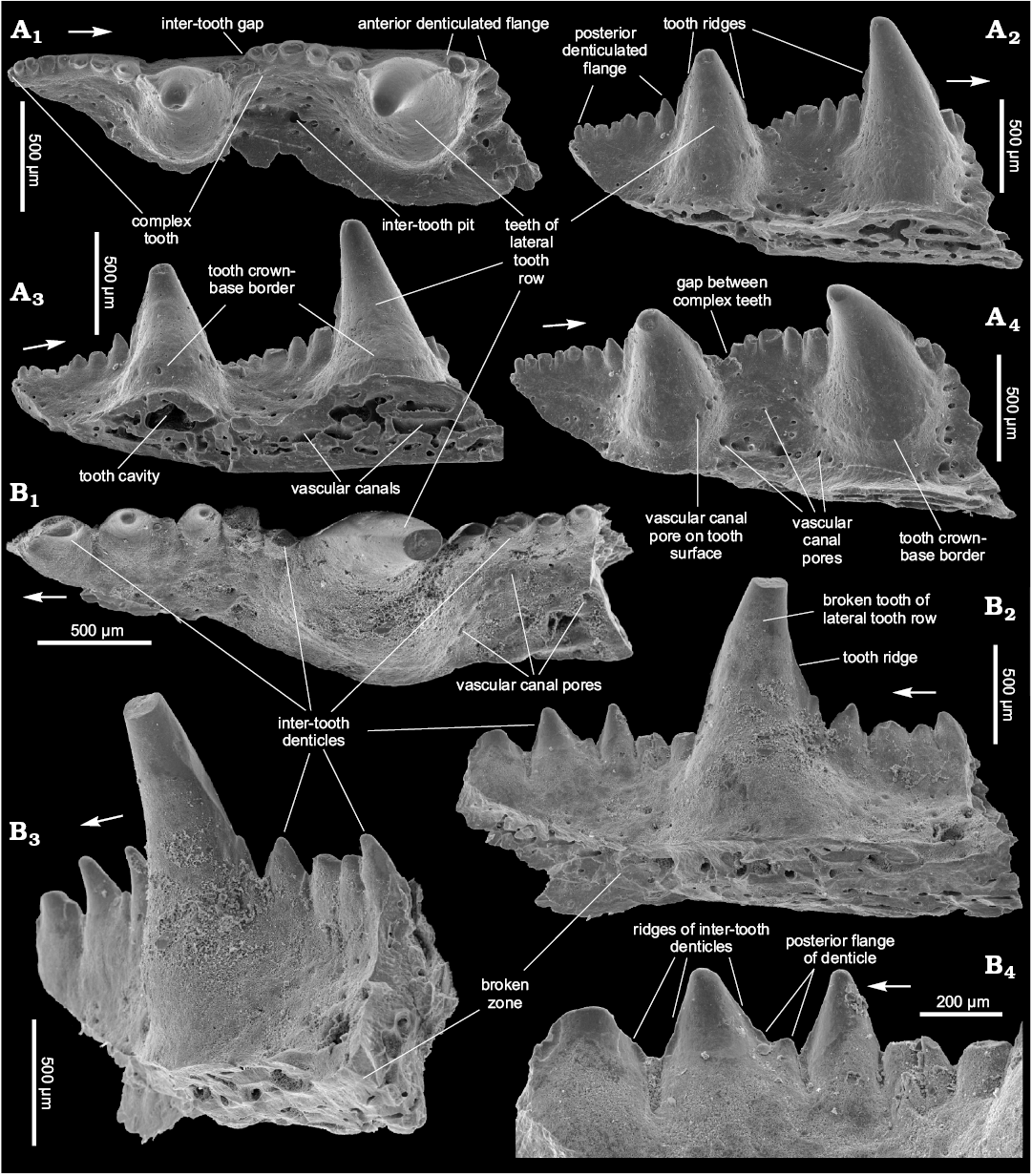
Fig. 6. Ischnacanthiform fish Kasperacanthus serratus gen. et sp. nov., Dniester River, on the East of Ivanie Zolote; upper part of the Ivanie Horizon of the Tyver Series, Lower Devonian; the paratypes ZPAL P.14/5.5 (A) and ZPAL P.14/7.6 (B). Jaw bone fragments in occlusal (A1, B1), lingual (A2, B2, B4), bottom-lingual (A3), top-lingual (A4), and posterolingual (B3) views. Arrows indicate rostral direction.
Podoliacanthidae gen. et sp. indet.
Fig. 7.
Material.—The illustrated specimen only, ZPAL P.14/12.10, a fragment of jaw bone, 3.25 mm long, with 2 broken teeth of the lateral tooth row. The specimen was found on the left bank of the Dniester River, on the east outskirts of Ivanie Zolote (Fig. 1C: point 4) in the deposits belonging to the upper part of the Ivanie Horizon of the Tyver Series, about 6 m below the boundary with the Dniester Series (Fig. 2: point 5).
Description.—The fragment, evidently from a middle part of the jaw bone, represents a relatively large and mature jaw bone. It is one of the biggest known specimens within the proposed Podoliacanthidae fam. nov. The jaw bone seems to be narrow. The width of the preserved fragment increases forward from about 0.7–1.0 mm. The height of the jaw bone, except for that of the lateral tooth row, is about 1.5 mm. Both teeth have their lower half preserved, where they have a short vertical ridge on the medial tooth side (Fig. 7A, B). The ridge width, in profile view, increases downward (Fig. 7C). The teeth are triangular in parabasal section. There is a crown-base border on the medial side, beneath the vertical ridge (Fig. 7A). The lingual tooth row resembles that of Podoliacanthus. The denticle groups are arranged diagonally in irregular transversal rows extending far onto the lingual slope of the jaw bone (Fig. 7A, B). The degree of their erasure is an evidence of the specimen maturity. The upper groups are somewhat larger than those situated on the slope. In general, it is particularly notable that the size difference between the denticle groups and the teeth of the lateral tooth row is significantly bigger than those known in other podoliacanthid forms. Specifically, along with the degree of erosion of the denticle groups, it likely indicates the maturity of the jaw bone. Nothing is known about posterior/anterior flanges of the teeth. Possibly, there are 3–5 inter-tooth denticles between the teeth of the lateral tooth row, but due to the poor state of preservation their presence can be known from indirect evidence only (Fig. 7C). The inter-tooth pits of the lateral tooth row are clearly defined. They are nearly oval in shape and rapidly increase in size forward, are rather deep and have pores of vascular canals mainly along their perimeter (Fig. 7D). There is a horizontally-arranged set of pores on the upper half of the lateral jaw bone surface (Fig. 7B).
Remarks.—In comparison with other podoliacanthid forms the specimen represents the largest jaw bone and is distinguished by the possession of lingual dentition of the teeth as a short vertical ridge. Additional data are needed for determining its place within the family.

Fig. 7. Podoliacanthidae fam. nov. gen. et sp. indet., ZPAL P.14/12.10, Dniester River, on the east outskirts of Ivanie Zolote; upper part of the Ivanie Horizon of the Tyver Series, Lower Devonian; jaw bone fragment in lingual (A), occlusal (B), and linguo-occlusal (D) views, postero-lingual view of lateral tooth row (C). Arrows indicate rostral direction.
Ischnacanthiformes fam., gen. et sp. indet.
Fig. 8.
Material.—SMNH BP.1586/4, a jaw bone fragment, 3.4 mm long, with three teeth of the lateral tooth row in different states of preservation (the smallest has a complete but erased tip, though lacks a side tooth part, the two other teeth have lost their tips); right bank of the Dniester River, opposite Zalishchyky, Zastavna (Chernivtsi) region (Podolia), Ukraine (Fig. 1C: point 1); upper part of Ivanie Horizon of the Tyver Series, Lower Devonian (Fig. 2: point 6).
Description.—The teeth are triangular in profile view with their tips slightly inclined backwards. The teeth seem to be heavy, massive and probably have no tooth cavity as, at least, the smallest of them (Fig. 8D, E). The broken tip of the largest tooth also does not reveal the tooth cavity (Fig. 8D, E). All broken zones show only vascular canals 20–30 µm in diameter (Fig. 8B, E). There are no clearly visible anterior or posterior flanges on the teeth. The largest of the preserved teeth has a crown-base border on the medial side, which comprises the lower third of the overall tooth height (Fig. 8B, E). The lingual tooth side forms a buttress that juts out in a medial-posterior direction (Fig. 8A–C). Thus, a parabasal section of the tooth looks like an “arrow” directed forward. The tooth size increases forward rapidly. The teeth have a smooth surface with no tooth side dentition visible. The width/height tooth ratio can be estimated as 1. The teeth are relatively tight. The linear size of the teeth in their base plane is much bigger than size of the inter-tooth pits on this level. Inter-tooth pits are funnel-shaped with a narrow lingual outlet (Fig. 8A, C, E). The lingual ridge is poorly preserved (Fig. 6C) and is weakly developed along the two smaller teeth of the jaw bone fragment and slightly marked out by pores of the vascular canal system near the largest (Fig. 8B). Pores of about 30 µm in diameter appear in the second lingual outlet, while those along the base of the largest tooth are 50–70 µm in diameter (Fig. 8A, B). No pores are present around the two smaller teeth and in the inter-tooth pit between them. Small pores are randomly distributed on the lateral/labial side of the jaw bone (Fig. 8D) and there are no inter-tooth denticles between the first two teeth of the jaw bone fragment. They are possibly broken off between the second and the third (the largest) tooth. One or two broken inter-tooth denticles appear immediately ahead the third tooth (Fig. 8A, C). The inter-tooth walls are slightly curved outwards and, together with the curved lateral side of the teeth, form a sinusoidal contour of the lateral tooth row (Fig. 8A).
Remarks.—The new form can be distinguished by massive solid teeth which have a smooth surface, prominent lingual buttress and arrow-like parabasal section. The specimen evidently represents a mature jaw bone. We leave this form in open nomenclature as only one jaw bone fragment is known.
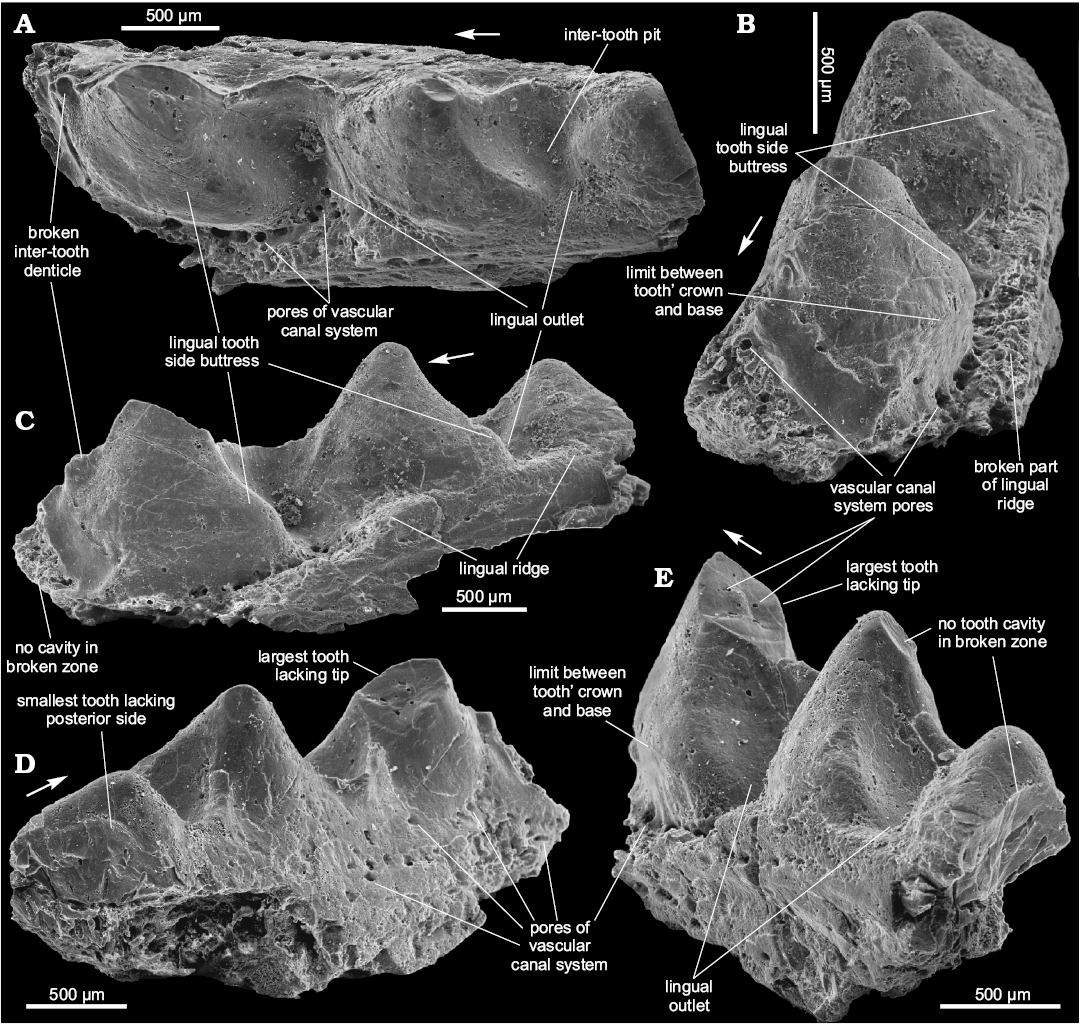
Fig. 8. Ischnacanthiformes fam. gen. et sp. indet., SMNH BP.1586/4, Dniester River, opposite Zalishchyky, Zastavna (Chernivtsi) region (Podolia), Ukraine; upper part of Ivanie Horizon of the Tyver Series, Lower Devonian; jaw bone fragment in occlusal (A), antero-linguo-occlusal (B), antero-lingual (C), postero-lateral (D), and postero-lingual (E) views. Arrows indicate rostral direction.
Discussion
The description of the new acanthodian taxa is based on a very limited number of fragmentary specimens. However, Early Devonian jaw bones of ischnacanthiform acanthodians are still poorly known, and thus worthy of publication. Their jaw bones are especially rare in comparison with other acanthodian remains, which is not surprising if one takes into account how many scales, tooth whorls and even spines are on one fish, with only four jaw bones. Additionally, many of the remains belong to acanthodians which did not possess jaw bones at all. Furthermore, the bones are very fragile due to their elongated structure (Voichyshyn and Szaniawski 2012). Nevertheless, all described taxa have clear features to distinguish them from other fossils and are characteristic enough to be recognised.
The concept of Podoliacanthidae fam. nov. is based on one main feature, the presence of characteristic denticle groups of Podoliacanthus type situated on the lingual tooth row (Figs. 3B, C, E, 4C1, C2, 5B1–B4; Voichyshyn and Szaniawski 2012: figs. 3F, 4A, 5F–H, 6C, E, H, 7C3, C4, 8A–C). Considering that there are different morphological forms within the plurality of jaw bones characterised by this feature we decided to establish a new family to keep them closely related. The majority of podoliacanthids also possess variable medial side dentition of the lateral tooth row teeth. But, as we pointed out previously (Voichyshyn and Szaniawski 2012), the morphology of lingual tooth row seems to have diagnostic importance of higher taxonomical level than the tooth medial side dentition.
The family Ischnacanthidae includes a majority of the newly discovered acanthodians with dentigerous jaw bones, regardless their particular morphology. The situation is similar to that of the genus Cephalaspis of Osteostraci. The large jaw bones of Ischnacanthus lack a lingual tooth row. Instead its representatives have well developed anterolabial and posterolabial flanges on their teeth (Ørvig 1967; Denison 1979; Long 1986; Hanke et al. 2001; Burrow 2002, 2004, 2007; Blais et al. 2015) that differentiate them from podoliacanthid morphology. The family Poracanthodidae, established on the base of scale histology, has evidently larger jaw bones with striated teeth of the lateral tooth row by wavy groovelets (Poracanthodes, Valiukevičius 2003) or sharp vertical keels/ridges (Zemlyacanthus, Valiukevičius 1992), and different morphology of the lingual tooth row surface. Other acanthodian family possessing jaw bones and being of uncertain affinity is Acritolepidae which has been erected based on the scale morphology, and has “jaw bones with large triangular tooth cusps and smaller secondary cusps along the main lateral ridge and striated denticles on the mesial ledge” (Valiukevičius and Burrow 2005). In fact, concerning the jaw bone, the family diagnosis is based on the genus Acritolepis which differs from representatives of Podoliacanthidae fam. nov. (Voichyshyn and Szaniawski 2012).
The key feature of Drygantacanthus gen. nov. is the set of denticles on the medial side of the teeth. Having larger teeth body, jaws of this genus seem to be stronger in comparison to Podoliacanthus. Although Drygantacanthus gen. nov. is represented by one- or two-teeth jaw bone fragments they have been found in different localities (Fig. 1C: points 1, 2) which supports our opinion that they belong to the distinct taxon.
Kasperacanthus gen. nov. has revealed some intriguing features. First of all, it has lingual denticle groups of Podoliacanthus type but, at the same time, it does not show any medial tooth side dentition, which distinguishes this genus from all other podoliacanthids known at present. The second remarkable feature is the presence of the regular gaps in the lateral tooth row, which mark the boundary between the complex-shaped teeth. Additionally, the taxon’s jaw bones demonstrate significant development of inter-tooth denticles. Kasperacanthus serratus sp. nov. is recorded from two widely-separated localities (Fig. 1C: points 3 and 4) with different deposit ages determining the species’ stratigraphic range. The specimens from Ivanie Zolote probably belong to mature jaw bones while the others represent possibly semi-adult specimens.
The specimen of Podoliacanthidae gen. et sp. indet. is interesting from several points of view. On one hand it evidently belonged to a taxon related to Podoliacanthus, which supports the establishment of the new family. On the other hand, it demonstrates potential jaw bone size, which can be reached by podoliacanthid representatives. Additionally, it is an example of a mature jaw bone fragment showing some peculiarities of age.
Ischnacanthiformes fam., gen. et sp. indet. differs from other Podolian ischnacanthiforms in having very solid but relatively short teeth. Their strength is supported by an expansion of the medial tooth side into a prominent buttress. It is possible that the form, being known from single jaw bone fragment, was rather scarce in the local fish fauna assemblages.
According to the recently established position of the boundary between Lochkovian and Pragian in Podolian Lower Devonian section, Drygantacanthus gen. nov. and Ischnacanthiformes fam., gen. et sp. indet. are here treated as of early Pragian age, while Kasperacanthus gen. nov., and Podoliacanthus seem to be distributed from mid-late Lochkovian to early Pragian. All of them are known from marine deposits.
Small jaw bones studied under magnification show a light structure with numerous canals and cavities causing general lightness of the bone (Figs. 4B3, 6A3; Voichyshyn and Szaniawski 2012: figs. 4F, 6G). Large jaw bones become more solid and heavy, probably independently of their systematic position. It can be seen on the largest forms described in this paper. The specimen of Podoliacanthidae fam. nov. gen. et sp. indet. does not exhibit any cavities on the both broken teeth and jaw bone section (Fig. 7A, B, D). Ischnacanthiformes fam., gen. et sp. indet. possess jaws with really heavy and massive teeth. This is evidenced by a lack of internal tooth cavity which does not occur in broken teeth (Fig. 8D, E), or broken jaw bone zones (Fig. 8C–E) and even in broken inter-tooth denticle (Fig. 8A). Additionally, the vascular canals and pores on these areas are small and relatively rare (Fig. 8B, E). The largest Kasperacanthus gen. nov. specimens likewise show almost continuous bone tissue with relatively rare and small vascular canals on broken jaw bone parts (Fig. 6B2, B3).
The similarity between morphological features of teeth of the lateral tooth row and of the denticles of their flanges in Kasperacanthus gen. nov. biggest specimen (Fig. 6B2–B4) deserves attention. The denticles look like the teeth on a reduced scale. Such similarity has been noted between teeth of the lateral tooth row and denticles of the lingual tooth row of several ischnacanthiform specimen/species (Voichyshyn and Szaniawski 2012: 895). These are evidence of some self-similar (fractal) patterns in tooth rows morphology of the ischnacanthiforms.
The morphology of the teeth of Drygantacanthus gen. nov. suggests their function was to break shells or other defensive structures of potential prey. Kasperacanthus gen. nov. has its teeth and significant denticles of the inter-tooth wall evidently for capturing and consuming its prey. The powerful jaws of Ischnacanthiformes fam., gen. et sp. indet. probably could crush more hard coverings than the Drygantacanthus gen. nov. ones. The strength of acanthodian jaw bones increased with ontogeny, according to their size, along with bone solidity and changing of diet preferences.
Conclusions
In spite of the fragility and rareness of ischnacanthiform jaw bones, it is worth considering using a method of dissolution of marine calcareous rocks to uncover them. Such investigation can help to explain the early ontogenetic stages of ischnacanthiform growth and to discover small sized ischnacanthiform species within a local fish fauna.
Despite the comparatively poor preservation of the material described in this paper it shows distinct features of the new taxa, different from other ischnacanthiforms known to date. It illustrates the potential ischnacanthiform taxonomical variety among marine fish fauna of Early Devonian age. The presence of several forms related to Podoliacanthus allows for the establishment of a new family reflecting their variability.
It is probable that growth of ischnacanthiform jaw bones coincided with increase of their solidity and degree of bone tissue continuity. Evidently, such an increase of jaw varieties must be accompanied by an enlargement of potential prey. The conjectured modes of nutrition i.e., crushing certain defensive covers or capturing and biting prey may explain the diverse tooth morphology among the described taxa.
Acknowledgements
We thank Andrzej Baliński (ZPAL) and Daniel Drygant (SMNH) for donating some of the specimens described in this paper. We thank also to both reviewers of the manuscript Carole Burrow (Queensland Museum, Brisbane, Australia) and the anonymous referee, for constructive comments, which greatly improved our paper. This research has been supported by the National Science Center of Poland, grant no 2016/21/B/ST10/02334.
References
Blais, S.A., Hermus C.R., and Wilson, M.V.H. 2015. Four new Early Devonian ischnacanthid acanthodians from the Mackenzie Mountains, Northwest Territories, Canada: an early experiment in dental diversity. Journal of Vertebrate Paleontology 35: 1189–1199. Crossref
Burrow, C.J. 2002. Lower Devonian acanthodian biostratigraphy of south-eastern Australia. Memoirs of the Association of Australasian Palaeontologists 27: 75–137.
Burrow, C.J. 2004. Acanthodian fishes with dentigerous jaw bones: the Ischnacanthiformes and Acanthodopsis. Fossils and Strata 50: 8–22.
Burrow, C.J. 2007. Early Devonian (Emsian) acanthodian faunas of the western USA. Journal of Paleontology 81: 824–840. Crossref
Denison, R. 1979. Acanthodii. In: H.-P. Schultze (ed.), Handbook of Paleoichthyology. Part 5, 1–62. Gustav Fisher Verlag, Stuttgart.
Drygant, D.M. 2010. Devonski konodonty pivdenno-zahidnoi okrainy Shidno-evropejskoî platformy (Volyno-Podillâ, Ukraîna). 191 pp. Naukova Dumka, Kyiv.
Drygant, D.M. and Szaniawski, H. 2012. Lochkovian conodonts from Podolia, Ukraine and their stratigraphic significance. Acta Palaeontologica Polonica 57: 833–861. Crossref
Filipiak, P., Zatoń, M., Szaniawski, H., Wrona, R., and Racki, G. 2012. Palynology and microfacies of Lower Devonian mixed carbonate-siliciclastic deposits in Podolia, Ukraine. Acta Palaeontologica Polonica 57: 863–877. Crossref
Gross, W. 1971. Downtonische und dittonische Acanthodier-Reste des Ostseegebietes. Palaeontographica, Abteilung A136 (1–6): 1–82.
Hanke, G.F., Davis, S.P., and Wilson, M.V H. 2001. The new species of the acanthodian genus Tetanopsyrus from northern Canada, and comments on related taxa. Journal of Vertebrate Paleontology 21: 740–753. Crossref
Karatajūtė-Talimaa, V.N. [Karataûtė-Talimaa, V.N.] 1978. Telodonty silura i devona SSSR i Špicbergena. 336 pp. Mokslas, Vilnius.
Long, J.A. 1986. New ischnacanthid acanthodians from the Early Devonian of Australia, with a discussion of acanthodian interrelationships. Journal of the Linnean Society (Zoology) 87: 321–339. Crossref
Long, J.A., Burrow, C.J., and Ritchie, A. 2004. A new Late Devonian acanthodian fish from the Hunter Formation near Grenfell, New South Wales. Alcheringa 28: 147–156. Crossref
Małkowski, K., Racki, G., Drygant, D., and Szaniawski, H. 2009. Carbon isotope stratigraphy across the Silurian–Devonian transition in Podolia, Ukraine: evidence for a global biogeochemical perturbation. Geological Magazine 146: 674–689. Crossref
Miles, R.S. 1973. Articulated acanthodian fishes from the Old Red Sandstone of England, with a review of the structure and evolution of the acanthodian shoulder-girdle. Bulletin of the British Museum (Natural History). Geology 24: 113–213.
Novitskaya, L.I. [Novitskaâ, L.I.] and Obruchev, D.V. [Obručev, D.V.] 1964. Class Acanthodei [in Russian]. In: D.V. Obručev (ed.), Osnovy paleontologii. Spravočnik dlâ paleontologov i geologov SSSR. Besčelûstnye, ryby, 175–194. Nauka, Moskva.
Ørvig, T. 1967. Some new acanthodian material from the Lower Devonian of Europe. In: C. Patterson and P.H. Greenwood (eds.), Fossil Vertebrates. Journal of the Linnean Society (Zoology) 47: 131–153. Crossref
Racki, G., Baliński, A., Wrona, R., Małkowski, K., Drygant, D., and Szaniawski, H. 2012. Faunal dynamics across the Silurian–Devonian positive isotope excursion (δ13C, δ18O) in Podolia, Ukraine: Comparative analysis of the Ireviken and Klonk events. Acta Palaeontologica Polonica 57: 795–832. Crossref
Szaniawski, H. 2012. Siluro-Devonian of Podolia, Ukraine: Paleobiological, biostratigraphic, and geochemical aspects. Acta Palaeontologica Polonica 57: 793–794. Crossref
Valiukevičius, J. 1992. First articulated Poracanthodes from the Lower Devonian of Severnya Zemlya [in Russian]. In: E. Mark-Kurik (ed.), Fossil Fishes as Living Animals, 193–213. Institute of Geology, Academy of Sciences of Estonia, Tallinn.
Valiukevičius, J. 2000. Acanthodian biostratigraphy and interregional correlations of the Devonian of the Baltic States, Belarus, Ukraine and Russia. Courier Forshungsinstitut Senckenberg 223: 271–289.
Valiukevičius, J. 2003. Devonian acanthodians from Severnaya Zemlya Archipelago (Russia). Geodiversitas 25: 131–204.
Valiukevičius, J. and Burrow, C.J. 2005. Diversity of tissues in acanthodians with Nostolepis-type histological structure. Acta Palaeontologica Polonica 50: 635–649.
Voichyshyn, V. [Vojčyšyn, V.] 2001. Distribution of fossil remains of Agnatha and accompanying vertebrate groups in deposits of Lower Devonian of Podolia [in Ukrainian]. Naukovi zapysky Deržavnogo pryrodoznavčogo muzeû 16: 47–58.
Voichyshyn, V.K. [Vojčyšyn, V.K.] 2010. Peculiarities of vertebrate fauna distribution in Early Devonian of Podolia [in Ukrainian]. Naukovi zapysky Deržavnogo pryrodoznavčogo muzeû 26: 205–218.
Voichyshyn, V. 2011. The Early Devonian armoured agnathans of Podolia, Ukraine. Palaeontologia Polonica 65: 1–211. Crossref
Voichyshyn, V. and Szaniawski, H. 2012. Acanthodian jaw bones from Lower Devonian marine deposits of Podolia, Ukraine. Acta Palaeontologica Polonica 57: 879–896. Crossref
Acta Palaeontol. Pol. 63 (2): 327–339, 2018
https://doi.org/10.4202/app.00456.2018