
Oligocene–Miocene freshwater gastropods from the Oltu-Narman Basin in eastern Turkey
MATHIAS HARZHAUSER, THOMAS A. NEUBAUER, and İZZET HOŞGÖR
Harzhauser, M., Neubauer, T.A., and Hoşgör, İ. 2018. Oligocene–Miocene freshwater gastropods from the Oltu-Narman Basin in eastern Turkey. Acta Palaeontologica Polonica 63 (2): 357–369.
We describe the first record of a late Oligocene or early Miocene freshwater gastropod fauna from the Susuz Formation of the Oltu-Narman Basin in northeastern Anatolia. The assemblage consists of only six species, indicating the presence of a well-oxygenated lake, pond or anabranch rich in vegetation. Only two of the taxa are related to European Oligo-Miocene species. The fauna might be the first indication of a yet undocumented Eastern Anatolian freshwater biogeographic region, characterized by high endemism and minor influence from the coeval Peri-Paratethyan and Aegean–Anatolian regions in the west. The biostratigraphic value of the assemblage is low but might rather point to a Miocene age of the upper part of the Susuz Formation. Theodoxus susuzianus, Bithynia erzurumensis, and Valvata koehleri are described as new species.
Key words: Mollusca, Gastropoda, Oligocene, Miocene, Turkey, Anatolia.
Mathias Harzhauser [mathias.harzhauser@nhm-wien.ac.at], Geological-Palaeontological Department, Natural History Museum Vienna, Burgring 7, 1010 Vienna, Austria.
Thomas A. Neubauer [thomas.a.neubauer@allzool.bio.uni-giessen.de], Department of Animal Ecology & Systematics, Justus Liebig University, Heinrich-Buff-Ring 26-32 IFZ, 35392 Giessen, Germany, and Naturalis Biodiversity Center, PO Box 9517, 2300 RA Leiden, The Netherlands.
İzzet Hoşgör [izzet.hosgor@calikenerji.com], Çalık Enerji, Oil and Gas Directorate, 06520, Söğütözü-Ankara, Turkey.
Received 10 February 2018, accepted 22 April 2018, available online 29 May 2018.
Copyright © 2018 M. Harzhauser et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Central and eastern European lake mollusc faunas are well known for their high degree of endemism during the Neogene (Harzhauser and Mandic 2008; Neubauer et al. 2015 and references therein). Comparatively little is known about coeval fluvial and lacustrine faunas from the adjacent Anatolian mainland. Recently, the knowledge on marine Oligocene–Miocene mollusc faunas from Turkey was significantly enlarged by the works of İslamoğlu (2008), Landau et al. (2013), and Harzhauser et al. (2016) among others. Similarly, the Oligocene and early Miocene mollusc faunas described from eastern and northeastern Anatolia derive from marine or brackish depositional environments (Stchépinsky 1939, 1941, 1946; Hüsing et al. 2009). Contributions describing pre-Pleistocene freshwater faunas, however, are rare and focus on late Miocene and Pliocene occurrences (e.g., Taner 1974a, b; Şen 1977; Schütt and Kavuşan 1984; Wesselingh et al. 2008; Wesselingh and Alçiçek 2010). Especially Oligocene and early Miocene faunas are virtually unknown, although Anatolia harbours numerous Miocene lignite deposits (Kara-Gülbay 2015), which can be expected to yield rich mollusc faunas and vertebrates (e.g., Murray and Hoşgör 2012). Most freshwater-mollusc bearing localities, such as Harmancık, Sandıklı, and Denizli, are situated in western Anatolia (see Neubauer et al. 2015) whereas eastern Anatolia is largely unexplored. Therefore, the herein described assemblage from the upper Oligocene or lower Miocene Susuz Formation from the Oltu-Narman Basin in northeastern Turkey is a first step to decipher the biogeographic relations of eastern Anatolian fossil freshwater faunas.
Institutional abbreviations.—NHMW, Natural History Museum Vienna, Austria.
Nomenclatural acts.—The electronic edition of this article conforms to the requirements of the amended International Code of Zoological Nomenclature, and hence the new names contained herein are available under that Code from the electronic edition of this article. This published work and the nomenclatural acts it contains have been registered in ZooBank, the online registration system for the ICZN. The LSID for this publication is: http://zoobank.org/references/153CB896-6EE8-48F1-A330-80FCD3B1ED94. The electronic edition of this work was published in a journal with an ISSN 0567-7920, eISSN 1732-2421, and has been archived and is available from the following digital repository: APP archive: http://www.app.pan.pl/article/item/app004692018.html
Geological setting
The herein described material derives from the Oltu-Narman Basin in the Erzurum province in eastern Turkey (Fig. 1A, B). The Oltu-Narman Basin is a roughly SW-NE striking basin of c. 80 km length and c. 10–20 km width, which formed along the Northeast Anatolian Fault Zone close to the northeast Anatolian−Caucasus frontier. Its southwestern boundary lies about 18 km southwest of Narman, and its northeastern boundary is about 40 km northeast of Oltu (Fig. 1A, B). Palaeozoic and Upper Cretaceous rocks form the basement. Marine and volcanoclastic Eocene units represent the oldest part of the basin fill. These are unconformably overlain by Oligocene–Miocene coal-bearing fluvio-lacustrine units (Bozkuş 1990; Yilmaz and Yilmaz 2016). The surface distribution of these Paleogene and Neogene deposits is partly masked by post-Miocene volcanics (Bozkuş 1990; Yilmaz and Yilmaz 2016).
A part of the Oligocene–Miocene basin fill is represented by the about 600-m-thick Susuz Formation, which comprises conglomerate, sandstone, silt- and claystone beds, and coal seams (Bozkuş 1990). The depositional environment of the Susuz Formation was interpreted by Karayigit et al. (2002) as floodplain of meandering rivers and rarely of braided rivers with numerous channels and overbank deposits alternating with small lakes and swamps. Despite the economic importance of the coal seams, the stratigraphic correlation of the Susuz Formation is rather tentative and partly deduced from its lithostratigraphic position overlying marine Eocene units. Based on pollen and spores assemblages, Bilgiç et al. (1995) suggested a late Oligocene to early Miocene age of the formation, which is in line with the traditional concepts of Bozkuş (1990), Karayigit et al. (2002), and Kara-Gülbay (2015). For a detailed description of the tectonic and geological setting of the Oltu-Narman Basin, see Bozkuş (1990) and Yilmaz and Yilmaz (2016).
The upper part of the Susuz Formation is exposed in small outcrops near the village Kömürlü in the Balkaya district of the Erzurum province (Fig. 1A, B). One of these sections was logged and sampled by IH in 2014 (Kömürlü section; 40°46’14.74” N, 42°18’21.05” E, WGS84). The about 9-m-thick section is mostly composed of grey, thin-bedded sandstone-siltstone intercalations, at the base overlain by yellow to light green, thick-bedded sandstone and claystone on top (Fig. 1C). The described molluscs were collected at 5.2 m from thin-bedded siltstone.
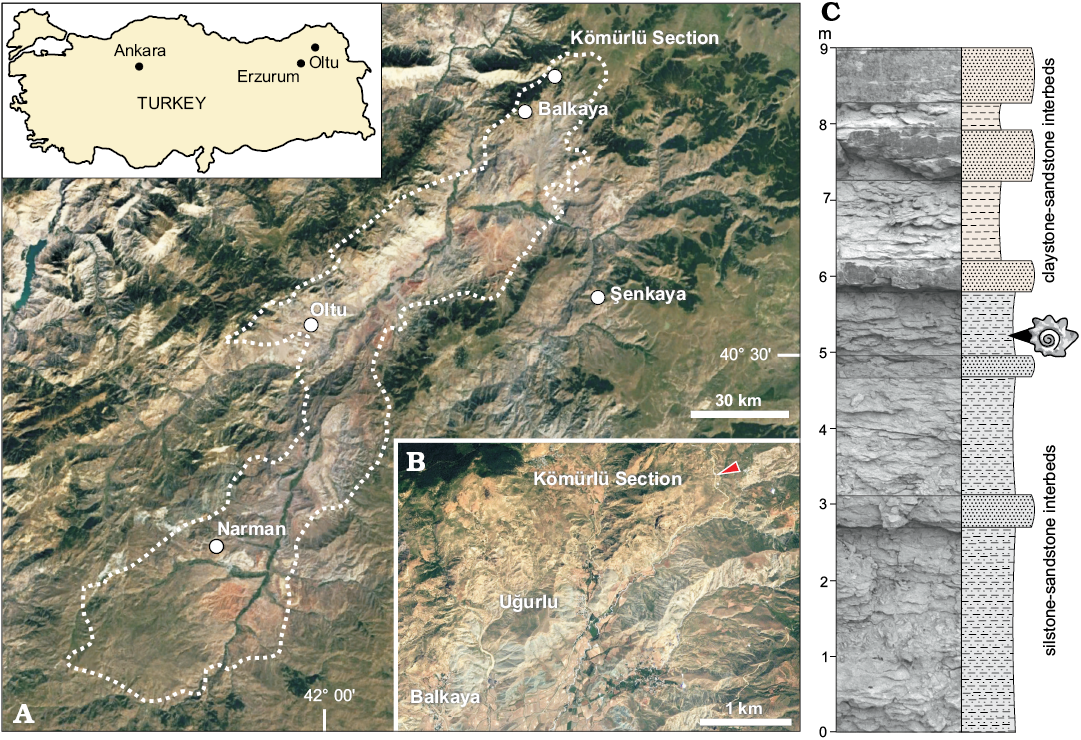
Fig. 1. A, B. Geographic position of the investigated section at the northeastern margin of the Oltu-Narman Basin in Eastern Anatolia (A), and detailed map of the position of the Kömürlü section indicated by an arrowhead (B). Outline of the Oltu-Narman Basin after Bozkuş (1990); maps generated with Google Earth Version 7.3.1, Image © 2018 Digital Globe; image taken in 7/5/2010. C. Picture and sedimentological log of the Kömürlü section (40°46’14.74” N, 42°18’21.05” E, WGS84) with sample position.
Material and methods
Most gastropod specimens are fragmented and deformed by sediment compaction. Nevertheless, the original shell is preserved in all specimens (no calcite-pseudomorphosis). The specimens were treated with diluted H2O2 to remove the consolidated sediment. All specimens are stored in the collection of the Geological-Paleontological Department of the Natural History Museum Vienna (NHMW 2018/0019/0001–0029).
Systematic palaeontology
Class Gastropoda Cuvier, 1795
Subclass Neritimorpha Golikov and Starobogatov, 1975
Superfamily Neritoidea Rafinesque, 1815
Family Neritidae Rafinesque, 1815
Genus Theodoxus Montfort, 1810
Type species: Theodoxus lutetianus Montfort, 1810, currently considered a junior synonym of Theodoxus fluviatilis (Linnaeus, 1758); by original designation. Recent, Europe.
Theodoxus susuzianus sp. nov.
Fig. 2.
ZooBank LSID: urn:lsid:zoobank.org:pub:153CB896-6EE8-48F1-A330-80FCD3B1ED94
Etymology: Referring to the upper Oligocene–lower Miocene Susuz Formation.
Type material: Holotype: NHMW 2018/0019/0001, moderately preserved, adult shell (Fig. 2A). Paratypes: NHMW 2018/0019/0002, moderately preserved, adult shell (Fig. 2B); NHMW 2018/0019/0003, moderately preserved, adult shell (Fig. 2C); NHMW 2018/0019/0004, moderately preserved, adult shell (Fig. 2D). All from type locality and horizon.
Type locality: Kömürlü (40°46’14.74” N, 42°18’21.05” E, WGS84), Oltu-Narman Basin, northeastern Turkey.
Type horizon: Marly silt and sand of the upper Susuz Formation; upper Oligocene or lower Miocene.
Material.—Type material and additional 21 specimens (NHMW 2018/0019/0005). All from type locality and horizon.

Fig. 2. Neritid gastropod Theodoxus susuzianus sp. nov. from the upper Oligocene–lower Miocene Susuz Formation, Kömürlü, Turkey. A. NHMW 2018/0019/0001, holotype in apertural (A1), oblique apical (A2), and dorsal (A3) views. B. NHMW 2018/0019/0002, paratype in apertural (B1) and oblique apical (B2) views; detail of protoconch (B3), arrowhead indicates transition into teleoconch. C. NHMW 2018/0019/0003, paratype in apertural (C1) and oblique apical (C2) views. D. NHMW 2018/0019/0004, paratype in apertural (D1) and oblique apical (D2) views; details of sculpture (D3). Scale bars 2 mm, if not stated otherwise.
Diagnosis.—Depressed drop-shaped neritid with colour pattern of regularly spaced, continuous brown and white spiral stripes. Aperture and columellar shelf simple with smooth columellar edge. Sculpture of last whorl consisting of prominent (for genus), densely spaced, prosocline axial riblets, being most prominent in adapical half of whorl.
Description.—Medium-sized shell (Table 1) with elevated spire; protoconch consisting of less than one smooth, low, moderately convex whorl of 500 μm diameter. Transition into teleoconch marked by faint rim and onset of delicate growth lines. Teleoconch comprising 2.5 whorls; first teleoconch whorl moderately high, weakly convex. Last whorl rapidly expanding; maximum diameter in lower third; whorl weakly convex to nearly flat above periphery, regularly convex below, resulting in depressed drop-shaped outline. Sculpture of last whorl consisting of densely spaced, prosocline axial riblets resulting from prominent growth lines, separated by narrower interspaces. Wide semi-circular aperture with thin outer lip. Columellar shelf broad, strongly expanding, well demarcated from base, forming thickened and convex callus in adults. Columellar edge nearly straight to faintly concave, slightly recessed behind basal lip; no denticles aside from very weak anterior swelling. Colour pattern consisting of 12–20 regularly spaced, continuous dark-brown spiral stripes separated by white stripes of about equal width. Operculum unknown.
Table 1. Measurements (in mm) of Theodoxus susuzianus sp. nov.
|
Specimen number |
Height |
Diameter |
|
NHMW 2018/0019/0001 |
6.0 |
6.1 |
|
NHMW 2018/0019/0002 |
6.0 |
6.0 |
|
NHMW 2018/0019/0003 |
4.9 |
5.2 |
|
NHMW 2018/0019/0004 |
5.8 |
5.2 |
Remarks.—The large protoconch, which consists of less than one whorl, places this species in Theodoxus and excludes a relation with other Neritininae genera, such as Clithon Montfort, 1810, Neritina Lamarck, 1816 and Pictoneritina Iredale, 1936 (see Bandel 2001). This classification is also supported by the simple columellar edge that lacks denticles, as well as the fluvial-lacustrine depositional environment, which was completely isolated from the sea. Theodoxus susuzianus displays little variation in colour pattern but is somewhat variable concerning the convexity of the adapical half of the last whorl.
Only a small number of Theodoxus species have been described so far from the Oligocene and early Miocene of central and eastern Europe and Turkey. Theodoxus supraoligocaenicus Báldi, 1973, from the Egerian (= Chattian/Aquitanian) of Hungary, differs in the presence of axial stripes on the spire and the smooth shell surface (Báldi 1973). Moreover, Báldi (1973) discusses similarities of the aperture of T. supraoligocaenicus with that of Vitta picta (Férussac, 1823), which might cast doubts about the generic placement. Theodoxus hassiacus (Wenz, 1919) sensu Esu and Girotti (2010), from the Chattian of southern Italy, is readily distinguished by its broader last whorl, slightly gradate spire and the speckled colour pattern. Theodoxus crenulatus (Klein, 1853), frequently reported from Oligocene and Miocene deposits of Europe and Turkey (e.g., Klein 1853; Báldi 1973; Kojumdgieva and Sapundgieva 1981; Harzhauser et al. 2012), is distinguished by its denticles on the columellar edge and the blunt speckles. The colour pattern of Theodoxus susuzianus is reminiscent of the late Miocene Neritina vetranici (Brusina, 1902) sensu Harzhauser et al. (2002), which differs from Theodoxus susuzianus in the presence of two protoconch whorls. Similarly, Theodoxus eugenii Jekelius, 1944 sensu Bandel (2001), from the late Miocene of Hungary, is highly reminiscent of the Turkish species concerning pattern and outline, but differs in the presence of columellar denticles. Another quite similar species in terms of colour pattern is Theodoxus bukowskii (Oppenheim, 1919) from the late Miocene of Turkey as described by Wesselingh et al. (2008). This species can be distinguished from T. susuzianus in its lower spire. None of the discussed species develops a marked axial sculpture on the last whorl.
Stratigraphic and geographic range.—Known so far only from the late Oligocene–early Miocene of the Oltu-Balkaya Basin in eastern Turkey.
Subclass Caenogastropoda Cox, 1960
Order Littorinimorpha Golikov and Starobogatov, 1975
Superfamily Truncatelloidea Gray, 1840
Family Bithyniidae Gray, 1857
Genus Bithynia Leach in Abel, 1818
Type species: Helix tentaculata Linnaeus, 1758; by subsequent designation by Herrmannsen (1846). Recent, Europe.
Bithynia erzurumensis sp. nov.
Fig. 3.
ZooBank LSID: urn:lsid:zoobank.org:act:18AD32B4-3585-47C5-8FF8-673F905AC72F
Etymology: Referring to the Erzurum province in eastern Turkey.
Type material: Holotype: NHMW 2018/0019/0006, moderately preserved, adult shell (Fig. 3A). Paratypes: NHMW 2018/0019/0007, moderately preserved, adult shell (Fig. 3B); NHMW 2018/0019/0008, moderately preserved, adult shell (Fig. 3C); NHMW 2018/0019/0009, moderately preserved, adult shell, with operculum in-situ (Fig. 3D); NHMW 2018/0019/0010, moderately preserved, adult shell (Fig. 3E). All from type locality and horizon.
Type locality: Kömürlü (40°46’14.74” N, 42°18’21.05” E, WGS84), Oltu-Narman Basin, northeastern Turkey.
Type horizon: Marly silt and sand of the upper Susuz Formation; upper Oligocene or lower Miocene.
Material.—Type material and additional 13 specimens (NHMW 2018/0019/0011) and 37 opercula (NHMW 2018/ 0019/0012–0014) (Fig. 3F, G). All from type locality and horizon.

Fig. 3. Bithyniid gastropod Bithynia erzurumensis sp. nov. from the upper Oligocene–lower Miocene Susuz Formation, Kömürlü, Turkey. A. NHMW 2018/0019/0006, holotype in apertural view. B. NHMW 2018/0019/0007, paratype in apertural view. C. NHMW 2018/0019/0008, paratype in dorsal view. D. NHMW 2018/0019/0008, paratype in apertural view showing in situ operculum. E. NHMW 2018/0019/0010, paratype in apical view. F. NHMW 2018/0019/0012, outer surface of operculum. G. NHMW 2018/0019/0013, outer surface of operculum. H. NHMW 2018/0019/0014, inner surface of operculum. Scale bars 0.5 mm, if not stated otherwise.
Diagnosis. —Small, stout shell with low conical spire, large globular last whorl, low, rapidly contracting base and deeply incised suture. Maximum diameter of last whorl in mid-height; aperture markedly angulated with posterior tip below periphery of last whorl.
Description.—Protoconch comprises very low initial part and about one strongly convex whorl of c. 950 µm diameter; transition into teleoconch unclear due to poor preservation. Low spire (Table 2) consisting of two convex teleoconch whorls, rapidly increasing in width with maximum diameter in lower third; suture deeply incised. Last whorl globular, regularly convex with maximum diameter in mid-height; base moderately convex, rapidly contracting. Aperture distinctly angulated at posterior tip, contact with last whorl below its periphery. Umbilicus narrow, partly covered by thin and narrow inner lip. Operculum solid, posteriorly angulated. Outer surface with central nucleus, prominent concentric growth lines, distinctly concave. Inner surface convex with shallow, irregular dents; height/width ratio c. 1.4.
Table 2. Measurements (in mm) of Bithynia erzurumensis sp. nov.
|
Specimen number |
Height |
Diameter |
|
NHMW 2018/0019/0006 |
4.4 |
3.5 |
|
NHMW 2018/0019/0007 |
3.1 |
1.7 |
|
NHMW 2018/0019/0008 |
3.8 |
3.2 |
|
NHMW 2018/0019/0009 |
4.1 |
3.2 |
|
NHMW 2018/0019/0010 |
3.9 |
2.5 |
Remarks.—The available material is only moderately preserved, but the characteristic outline of this stout globular species allows a clear separation from other Bithynia species and justifies its description as new species. A specimen with the operculum preserved in situ (Fig. 3D) proves that shells and opercula belong to the same species.
A globular shell shape and a low spire are also found in some species of the bithyniid genus Pseudobithynia Glöer and Pešić, 2006, e.g., in its type species Pseudobithynia irana Glöer and Pešić, 2006. As only genital features allow a separation of both genera (Glöer and Pešić 2006), we place the fossil species tentatively in Bithynia.
Although the genus was diverse during the Miocene and Pliocene in Europe (Bouchet et al. 2018 list 34 fossil species), only few have been documented from Oligocene and lower Miocene deposits (see Wenz 1928). Of these, only Bithynia globuloides (Sandberger, 1873), from the Oligocene of England, is similar to the Turkish species in outline. It differs in its ovoid aperture, which lacks a marked angulation. The middle Miocene Bithynia jurinaci Brusina, 1884, from the Dinaride Lake System, as well as B. glabra (Zieten, 1832) and B. dunkeri Gude, 1913, both from southern Germany, differ in their higher shells and less incised suture (see Schlickum 1966; Neubauer et al. 2016). The fossil species is reminiscent of the extant Turkish Pseudobithynia pentheri (Sturany, 1904) concerning the low spire and large last whorl, but differs from this species in its even more globular outline and the narrower umbilicus (see Glöer and Yildirim 2006). The extant Turkish Pseudobithynia guldeni Gürlek, 2018 lacks an umbilicus and has a low spire but differs in its higher last whorl and the smaller size (see Gürlek 2018). Similarly, the extant Bithynia mostarensis von Möllendorff, 1873, from Skadar Lake, has a rather globular outline but develops a higher last whorl, and its base is only slowly contracting compared to the fossil species (Glöer and Pešić 2007). The extant Bithynia radomani Glöer and Pešić, 2006, also from Skadar Lake, has a globular last whorl with low base but differs in its higher spire (see Glöer and Pešić 2007).
Stratigraphic and geographic range.—Known so far only from the late Oligocene–early Miocene of the Oltu-Balkaya Basin in eastern Turkey.
Subclass Heterobranchia Burmeister, 1837
Superfamily Valvatoidea Gray, 1840
Family Valvatidae Gray, 1840
Genus Valvata Müller, 1773
Type species: Valvata cristata Müller, 1774; by subsequent monotypy. Recent, Europe.
Valvata koehleri sp. nov.
Fig. 4.
ZooBank LSID: urn:lsid:zoobank.org:act:A0E6BDE6-C921-46D7-B7AA-3CA72DD43C88
Etymology: Referring to Frank Köhler (Australian Museum Sydney), in honour of his contributions to malacology.
Type material: Holotype: NHMW 2018/0019/0015, moderately preserved, fully grown specimen (Fig. 4D). Paratypes: NHMW 2018/ 0019/0016, compressed, adult specimen (Fig. 4B); NHMW 2018/ 0019/0017, slightly compressed, adult specimen (Fig. 4C); NHMW 2018/0019/0018, compressed, adult specimen (Fig. 4A); NHMW 2018/0019/0019, laterally compressed specimen. All from type locality and horizon.
Type locality: Kömürlü (40°46’14.74” N, 42°18’21.05” E, WGS84), Oltu-Narman Basin, northeastern Turkey.
Type horizon: Marly silt and sand of the upper Susuz Formation; upper Oligocene or lower Miocene.
Material.—Type material only.
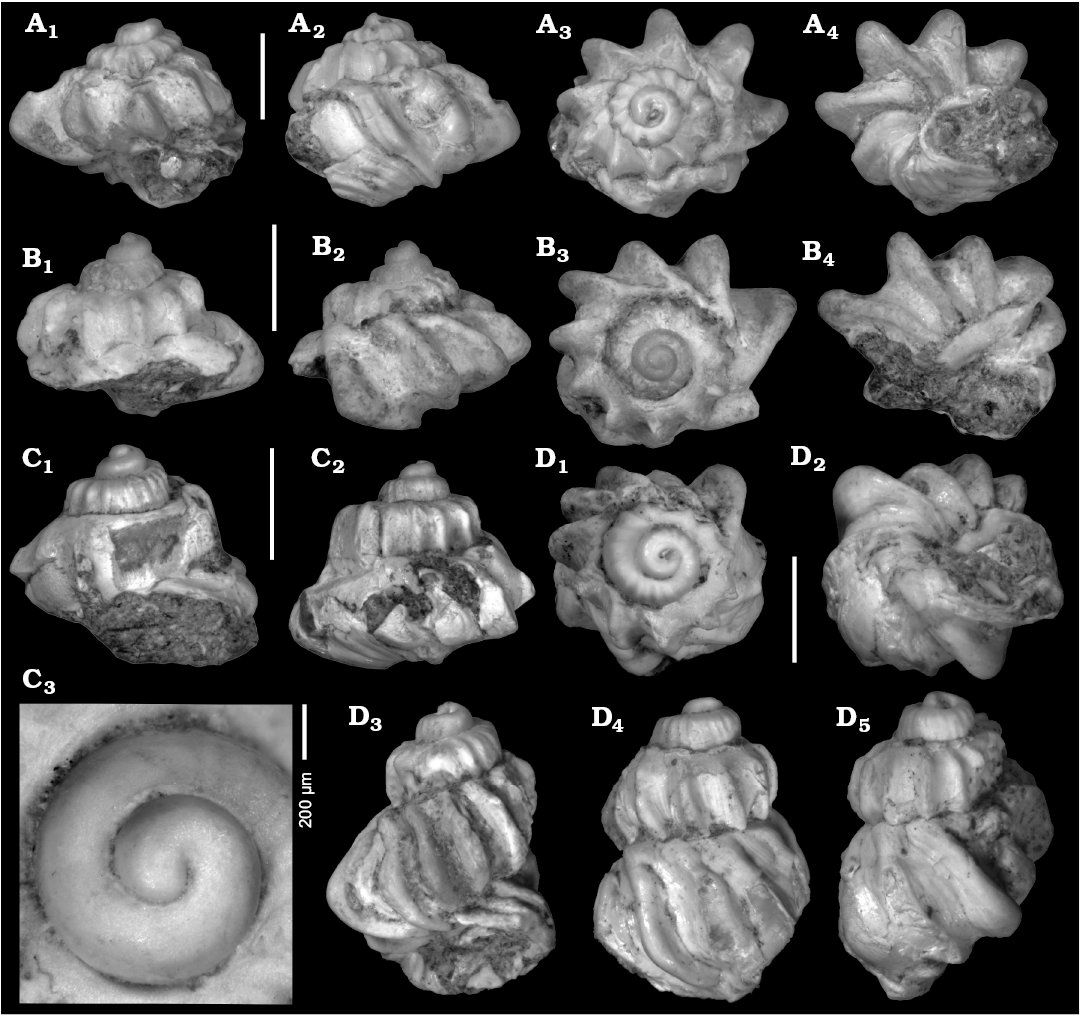
Fig. 4. Valvatid gastropod Valvata koehleri sp. nov. from the upper Oligocene–lower Miocene Susuz Formation, Kömürlü, Turkey. A. NHMW 2018/0019/0018, paratype in apertural (A1), lateral (A2), apical (A3), and umbilical (A4) views. B. NHMW 2018/0019/0016, paratype in lateral (B1, B2), apical (B3), and umbilical (B4) views. C. NHMW 2018/0019/0017, paratype in lateral views (C1, C2); protoconch (C3). D. NHMW 2018/0019/0015, holotype in apical (D1), umbilical (D2), apertural (D3), lateral (D4), and dorsal (D5) views. Scale bars 2 mm, if not stated otherwise.
Diagnosis.—Small robust shell characterized by extraordinarily blunt and protruding axial ribs and strongly changing outline of teleoconch whorls during ontogeny from gradate conical to moderately high conical. Protoconch conical, comprising 1.3 strongly convex whorls with incised suture. Early teleoconch whorls with distinct angulation and orthocline axial ribs. Second and third teleoconch whorls rapidly widening with angulation at lower suture; fourth teleoconch whorl comparatively narrow but distinctly increasing in height. Suture deeply incised. Blunt axial ribs continue on base up to strongly reduced umbilicus. Inner lip distinct, reflected; outer lip coinciding with axial rib.
Description.—Small, very robust shells with strongly allometric growth (Table 3). Conical protoconch of 950 μm diameter and 550 μm height, comprising 1.3 strongly convex whorls with incised suture. Transition into teleoconch indistinct, indicated by onset of weak axial ribs. First teleoconch whorl strongly angulated with nearly flat sutural ramp and straight flank. Weak axial riblets, bearing tiny nodes at angulation, on early whorls pass into successively more prominent orthocline axial ribs increasing in width. Cross-section of ribs changing successively from convex to trigonal. Axial ribs become blunt and wide-spaced within second teleoconch whorl; angulation shifts slightly downwards resulting in steeper sutural ramp; flank of whorl becomes more conical, with maximum diameter at lower suture. Axial ribs now distinctly recessed close to lower suture, coinciding with slight angulation. Third teleoconch whorl strongly widening, broad conical; axial ribs becoming strongly prosocline and extremely blunt, being reminiscent of spines in apical view (axial ribs account for about 1/3 of diameter at that stage); ribs partly laterally dented, forming shallow, irregular concavities. Transition into weakly convex base of third teleoconch whorl strongly accentuated by lower angulation. Fourth teleoconch whorl less expanding but increasing in height; upper and lower angulation weakening towards aperture; blunt, prosocline to slightly sigmoidal axial ribs becoming less protruding on last whorl. Aperture only partly preserved; distinct inner lip well demarcated from base; axial ribs continue on weakly convex base; umbilicus reduced to narrow chink, largely covered by reflected inner lip. Outer lip coincides with varix-like axial rib terminating in thin peristome (destroyed during preparation).
Table 3. Measurements (in mm) of Valvata koehleri sp. nov.
|
Specimen number |
Height |
Diameter |
|
NHMW 2018/0019/0015 |
5.7 |
4.5 |
|
NHMW 2018/0019/0016 |
4.5 |
5.4 |
|
NHMW 2018/0019/0017 |
3.5 |
4.8 |
|
NHMW 2018/0019/0018 |
3.9 |
4.7 |
|
NHMW 2018/0019/0019 |
5.3 |
5.8 |
Remarks.—This species is characterized by a striking allometry in whorl shape and sculpture. Especially the height of the third teleoconch whorl is variable, ranging from depressed (Fig. 4B) to moderately high (Fig. 4D). Only few valvatids with axial sculpture have been described from the Miocene and Pliocene of the eastern Mediterranean region. Of these, Valvata cincta Taner, 1974a, as described by Wesselingh et al. (2008) from the late Miocene of Turkey, has prominent axial ribs too but differs in the higher spire, more numerous and less blunt axial lips and the presence of an umbilicus. Similarly, the axial ribs of Valvata pagana Bulić and Jurišić-Polšak, 2009, from the middle Miocene of Pag Island (Croatia), are less prominent and more numerous, and the species develops a moderately wide and deep umbilicus. Among Recent valvatids, the Greek Valvata klemmi Schütt, 1962 is highly reminiscent of the Turkish fossil in respect to the strongly angulated whorls and the persisting axial ribs on the base. It differs in the much weaker axial ribs and the broader umbilical chink. This extant species was placed by Schütt (1962) in Costovalvata Poliński, 1929, with the type species Valvata (Costovalvata) hirsutecoststa Poliński, 1929. Similarly, Bulić and Jurišić-Polšak (2009) placed their Miocene V. pagana with question mark in the subgenus Costovalvata. Valvata (Costovalvata) hirsutecoststa is endemic to Lake Ohrid, has a typical valvatid shell and is characterized by a cancellate sculpture in which spiral cords are nearly as prominent as the axial ribs. The morphology and type of sculpture have little in common with V. pagana and V. klemmi. Therefore, we doubt a closer relationship. A potential genus name for these species would be Aegaea Oppenheim, 1891, which was introduced by Oppenheim (1981) for Valvata vivipariformis Oppenheim, 1891 from the Pleistocene of Kumari near Aegion (Greece). Oppenheim (1891) based the genus on the presence of prominent axial ribs and the narrow to nearly closed umbilicus, which is untypical for Valvata. As pointed out by Haszprunar (2014), however, the angulated aperture of Valvata vivipariformis rather suggests a relation with viviparids, rendering Aegaea a problematic taxon.
The excessive sculpture of V. koehleri is reminiscent of some Melanopsidae as described by Neubauer et al. (2013) from the middle Miocene of the Dinaride Lake System. Protoconch morphology and shape of the aperture exclude a relation with that family. Cretaceous to Paleocene Stomatopsidae Stache, 1889 are superficially similar concerning the prominent axial ribs but differ clearly by their elongate shells, their much larger size and the strongly callous inner lip (see Stache 1889).
Stratigraphic and geographic range.—Known so far only from the late Oligocene–early Miocene of the Oltu-Balkaya Basin in eastern Turkey.
Order Hygrophila Férussac, 1822
Suborder Branchiopulmonata Morton, 1955
Superfamily Lymnaeoidea Rafinesque, 1815
Family Physidae Fitzinger, 1833
Genus Aplexa Fleming, 1820
Type species: Bulla hypnorum Linnaeus, 1758; by subsequent designation by Herrmannsen (1846). Recent, Europe.
Aplexa cf. subhypnorum Gottschick, 1920
Fig. 5A, B.
cf. 1920 Aplexa subhypnorum n. sp.; Gottschick 1920: 116, pl. 2: 9.
Material.—Five fragmentary specimens: NHMW 2018/ 0019/0020, largest specimen (Fig. 5A); NHMW 2018/0019/ 0021 (Fig. 5B); NHMW 2018/0019/0021 (3 specimens) from the upper Susuz Formation (upper Oligocene or lower Miocene) at Kömürlü, Oltu-Narman Basin, northeastern Turkey.

Fig. 5. Physid (A, B) and planorbid (C–I) gastropods from upper Oligocene–lower Miocene Susuz Formation, Kömürlü, Turkey. A, B. Aplexa cf. subhypnorum Gottschick, 1920. A. NHMW 2018/0019/0020, in dorsal (A1) and apertural (A2) views; detail of sculpture (A3). B. NHMW 2018/0019/0021, in apertural view. C–E. Planorbarius cf. cornu (Brongniart, 1810). C. NHMW 2018/0019/0022, in apical view, specimen with characteristic protoconch microsculpture; C1 detail of C2. D. NHMW 2018/0019/0023, in apical view, specimen with prominent spiral cords. E. NHMW 2018/0019/0024, in lateral view. F–I. Gyraulus sp. F. NHMW 2018/0019/0025, in umbilical (F1), apical (F2), and lateral (F3) views. G. NHMW 2018/0019/0026, in umbilical (G1), apical (G2), and lateral (G3) views. H. NHMW 2018/0019/0027, in umbilical (H1) and apical (F2) views. I. NHMW 2018/0019/0028, in lateral view. Scale bars A1, A2, B, 5 mm; C1, 200 µm ; A3, C2, D, E, 1 mm, F–I, 500 µm.
Description.—Sinistrally coiled shell with high conical spire (Table 4) of at least three high, moderately convex teleoconch whorls (protoconch and early teleoconch not preserved). Last whorl high, moderately convex; suture deeply incised. Shell surface of spire whorls consisting of prominent, broad, densely spaced axial riblets becoming much weaker on last whorl. Axial sculpture continues on narrow subsutural cord. Aperture attaining about 50% of total height, angulated in posterior tip. Outer lip thin; inner lip delicate, narrow; columella weakly concave.
Table 4. Measurements (in mm) of Aplexa cf. subhypnorum Gottschick, 1920.
|
Specimen number |
Height |
Diameter |
|
NHMW 2018/0019/0020 |
18 |
9 |
|
NHMW 2018/0019/0021 |
12.6 |
7.5 |
Remarks.—All specimens are deformed due to sediment compaction and only fragmentarily preserved. The high and slender spire suggests a placement in Aplexa Fleming, 1820 rather than in Physa Draparnaud, 1801. Aplexa is poorly known from the Oligocene and Miocene of Europe and Turkey. So far, only Aplexa subhypnorum Gottschick, 1920 was described from the middle Miocene of Steinheim in southern Germany (Gottschick 1920). Juvenile specimens of Aplexa were also documented from the middle Miocene of Opole in Poland (Harzhauser and Neubauer 2018) and the late Miocene of the Vienna Basin (Harzhauser and Binder 2004). The Turkish shells largely match the description of Aplexa subhypnorum by Gottschick (1920) and agree in size and sculpture. The illustrations by Gottschick (1920), however, are too sketchy to support the identification of the poorly preserved Turkish material. Moreover, the large biogeographic and stratigraphic distance between the German and Turkish occurrences may give rise to doubts whether both are conspecific. Unfortunately, the syntypes of Aplexa subhypnorum are lost (Salvador et al. 2016), and a direct comparison is impossible.
Aplexa subhypnorum is known from the middle Miocene of southern Germany. It might have also occurred at that time in Poland and during the late Miocene in the Vienna Basin (Harzhauser and Neubauer 2018).
Superfamily Planorboidea Rafinesque, 1815
Family Planorbidae Rafinesque, 1815
Genus Planorbarius Duméril, 1805
Type species: Helix cornea Linnaeus, 1758; by subsequent monotypy by Froriep (1806). Recent, Europe.
Planorbarius cf. cornu (Brongniart, 1810)
Fig. 5C–E.
cf. 1810 Planorbis Cornu; Brongniart 1810: 371, pl. 22: 6.
Material.—Seven specimens: NHMW 2018/0019/0022 (Fig. 5C); NHMW 2018/0019/0023 (Fig. 5D); NHMW 2018/ 0019/0024 (Fig. 5E); NHMW 2018/0019/0024a (4 specimens) from the upper Susuz Formation (upper Oligocene or lower Miocene) at Kömürlü, Oltu-Narman Basin, northeastern Turkey.
Remarks.—Only fragmentary specimens consisting of few whorls are available showing a shell with strongly convex whorls and deep suture (Table 5). The protoconch displays traces of spiral rows of pits, which are replaced by delicate spiral cords on the first teleoconch whorl whereas the later whorls are smooth. These features correspond fully to specimens of Planorbarius cornu as described by Harzhauser et al. (2014b) from the early Miocene of the Czech Republic. Although the very high whorls support the identification, a separation from the early to late Miocene Planorbarius mantelli (Dunker, 1848) is difficult based on fragments only (see Harzhauser et al. 2014a, b). Moreover, the status of Planorbarius cornu is problematic and would need revision based on topotypic specimens from the Meulières de Montmorency in France (see Kadolsky 2015 for discussion). Therefore, we discuss the Turkish species in open nomenclature.
Planorbarius cornu was originally described from the Rupelian of the Paris Basin and is frequently recorded in Europe from Oligocene and lower Miocene deposits (Wenz 1923).
Table 5. Measurements (in mm) of Planorbarius cf. cornu (Brongniart, 1810).
|
Specimen number |
Height |
Diameter |
|
NHMW 2018/0019/0022 |
2.5 |
4.5 |
|
NHMW 2018/0019/0023 |
2.4 |
6.3 |
|
NHMW 2018/0019/0024 |
2.4 |
4.3 |
Genus Gyraulus Charpentier, 1837
Type species: Planorbis albus Müller, 1774; by subsequent designation by Dall (1870). Recent, Germany.
Gyraulus sp.
Fig. 5F–I.
Material.—Ten specimens: NHMW 2018/0019/0025 (Fig. 5F); NHMW 2018/0019/0026 (Fig. 5G); NHMW 2018/ 0019/0027 (Fig. 5H); NHMW 2018/0019/0028 (Fig. 5I); NHMW 2018/0019/0028a (6 specimens) from the upper Susuz Formation (upper Oligocene or lower Miocene) at Kömürlü, Oltu-Narman Basin, northeastern Turkey.
Description.—Small, glossy, planispiral, rather evolute shell (Table 6) comprising three whorls with moderately incised suture. Protoconch poorly preserved in all specimens. Lower and upper sides slightly concave; umbilicus and spire only weakly sunken; teleoconch whorls on lower side weakly convex in middle part of whorls, passing into nearly straight part towards periphery. Whorls flattened on upper side resulting in rather flat profile. Periphery coinciding with marked central angulation without forming keel. Distinct, prosocyrt growth lines on lower side but nearly smooth on upper side. Last whorl slightly bent towards upper side in some specimens. Aperture heart-shaped, wider than high, weakly inclined.
Table 6. Measurements (in mm) of Gyraulus sp.
|
Specimen number |
Height |
Diameter |
|
NHMW 2018/0019/0025 |
0.55 |
3.0 |
|
NHMW 2018/0019/0026 |
0.7 |
2.8 |
|
NHMW 2018/0019/0027 |
0.55 |
2.4 |
|
NHMW 2018/0019/0028 |
0.5 |
2.0 |
Remarks.—The specimens resemble several species of the European Oligocene to early Miocene. Planorbis? amblytropis Sandberger, 1871, from the Chattian of Vicques (Recolaine), Switzerland, and Gyraulus goussardianus (Noulet, 1854) (sensu Fischer 2000), from the late early to early middle Miocene of Sansan, France, both share with the present specimens the marked central angulation, the whorl expansion rate, the flat profile and the strong growth lines; only the aperture seems to be wider in both species. Similarly, Gyraulus douvillei (Saporta, 1889), from the Chattian of southern France, resembles the present specimens in terms of whorl expansion rate, growth line strength and the presence of a keel. Unfortunately, Saporta (1889) did not specify the exact location of the keel and did not provide an illustration of the shell profile. The studied specimens might be conspecific with one of these species, but the rather poor preservation of the present material, as well as the insufficient knowledge of the tentative European relatives, complicates reliable determination.
Other angulated or keeled forms comprise Gyraulus ludovici (Noulet, 1854) from Sansan, which differs in the apical angulation and the low whorl expansion rate, and Gyraulus applanatus (Thomä, 1845), from the early Miocene of the Mainz Basin (Germany), having its keel near the base. Early Miocene Gyraulus dealbatus (Braun in Walchner, 1851) differs in the only weak, subcentral angulation and the lower whorl expansion rate (see also Harzhauser et al. 2014b).
A relation with genus Hippeutis Charpentier, 1837 is unlikely. Species attributed to that genus are characterized by flatter shells with marked keel and often concave margins (e.g., Welter-Schultes 2012; Harzhauser et al. 2014b).
Concluding remarks
Biostratigraphy.—The stratigraphy of the Susuz Formation is very poorly constrained and based only on pollen data. It is only vaguely classified as Oligocene–Miocene in all stratigraphic tables (e.g., Bilgiç et al. 1995; Bozkuş 1990; Karayigit et al. 2002; Kara-Gülbay 2015; Yilmaz and Yilmaz 2016). Unfortunately, the gastropods from the Oltu-Narman Basin do not allow specifying the age of the Susuz Formation. Four of the six species are new and so far endemic to the basin. The overall character of the gastropod assemblage might rather suggest a Miocene age. Aplexa subhypnorum is a clearly Miocene species, and species presumptively related to Valvata koehleri all derive from Neogene deposits. Another, albeit much weaker argument is the presence of Theodoxus, which is rare in the Oligocene (and the generic identification of most Oligocene taxa will need confirmation), but becomes frequent during the early Miocene (e.g., Aquitanian of Greece; Harzhauser and Kowalke 2001). Finally, the sampling position in the upper part of the Susuz Formation might support a Miocene age.
Paleoenvironment and paleoecology.—All described genera are widespread in modern freshwater environments (e.g., Turner et al. 1998; Kerney 1999; Glöer 2002; Spyra 2012; Welter-Schultes 2012). According to these authors, extant Valvata species, such as V. piscinalis (Müller, 1774) and V. cristata Müller, 1774 prefer clear, well oxygenated, stagnant or slowly moving freshwater in lakes, ponds and creeks. Extant Bithynia species, such as B. leachii (Sheppard, 1823) and B. tentaculata (Linnaeus, 1758), occur in stagnant to slowly moving waters of lakes, ponds and rivers with rich vegetation. Similarly, the extant Planorbarius corneus (Linnaeus, 1758) live in standing to slowly running waters of small, densely vegetated lakes, ponds, marshes and bogs. Aplexa hypnorum (Linnaeus, 1758) prefers small water bodies rich in vegetation and tolerates even temporary droughts. Only Theodoxus has a broader habitat range and may also tolerate brackish water. Many species, such as Theodoxus fluviatilis (Linnaeus, 1758) and Theodoxus anatolicus (Récluz, 1841), prefer riverine settings, but others, such as Theodoxus heldreichi (Martens, 1879) and Theodoxus danubialis (Pfeiffer, 1828), are also adapted to lake environments.
Thus, the majority of the genera represented in the sample from the Susuz Formation indicates well-oxygenated silent or slowly moving water of a lake, pond or anabranch with rich vegetation. Theodoxus susuzianus was most probably also a lake species, as no sedimentological indication for fluvial transport is documented.
Biogeography.—The available assemblage comprises only six gastropod species. This low diversity is likely a result of undersampling, and further sampling campaigns might detect additional species. Even this small assemblage is an important contribution to our understanding of the Oligo-Miocene freshwater faunas of Eastern Anatolia. Coeval assemblages were described by Stchépinsky (1946) and İslamoğlu (2008) from western Anatolia, and they contain only one or two species each. The only possible match with those records regard P. cornu mentioned by Stchépinsky (1946) from Bucuk near Ankara. No other fauna of Oligocene or early Miocene age is known from Anatolia, the Caucasus region or the Near East. A vague similarity to European faunas is indicated by the presence of Aplexa cf. subhypnorum and Planorbarius cf. cornu. These species have been reported from early to middle Miocene localities of the Peri-Paratethyan and Aegean–Anatolian biogeographic regions (sensu Neubauer et al. 2015). The poor preservation, however, does not allow confirming that these species are indeed conspecific with their western counterparts. Bithynia erzurumensis and Valvata koehleri, in contrast, lack any related species in the early Miocene Peri-Paratethyan Region. The conspicuous morphology of Valvata koehleri is reminiscent of some Miocene and extant species from Turkey and Greece, suggesting a vague relation to the Aegean–Anatolian Region. Theodoxus susuzianus has no counterparts in the Turkish Miocene and Pliocene (although the colour pattern is found in Turkish Neogene species such as T. bukowskii). Thus, even this small sample suggests the presence of a peculiar faunal type with some relations to the Peri-Paratethyan and Aegean–Anatolian regions. The fauna from the Oltu-Narman Basin might be a first indication of the presence of a yet undefined Eastern Anatolian freshwater biogeographic region. Further data are needed to test this hypothesis.
Acknowledegments
We thank Frank Köhler (Australian Museum, Sydney, Australia) for discussions on systematics, as well as M. Cihat Alçiçek (Pamukkale University, Denizli, Turkey) and Hasan Çelik (Fırat University, Elazığ, Turkey) for sharing ideas on stratigraphy. We are grateful to Vitaliy V. Anistratenko (Schmalhausen Institute of Zoology, National Academy of Sciences of Ukraine, Kiev, Ukraine) and Michael W. Rasser (State Museum of Natural History Stuttgart, Germany) for their constructive reviews. The study contributes to the FreshGEN Project (“Freshwater systems in the Neogene and Quaternary of Europe: Gastropod biodiversity, provinciality, and faunal gradients”) funded by the Austrian Science Fund FWF (project no. P25365-B25). TAN was supported by an Alexander-von-Humboldt Scholarship.
References
Abel, C. 1818. Narrative of a Journey in the Interior of China, And of a Voyage To And From That Country, In the Years 1816 and 1817; Containing an Account of the Most Interesting Transactions of Lord Amherst’s Embassy To the Court of Pekin, and Observations on the Countries Which It Visited. Illustrated by Maps and Other Engravings. xvi + 420 pp. Longman, Hurst, Rees, Orme & Brown, London.
Báldi, T. 1973. Mollusc Fauna of the Hungarian Upper Oligocene (Egerian). 511 pp. Akadémiai Kiadó, Budapest.
Bandel, K. 2001. The history of Theodoxus and Neritina connected with description and systematic evaluation of related Neritimorpha (Gastropoda). Mitteilungen des Geologisch-Paläontologischen Institutes der Universität Hamburg 85: 65–164.
Bilgiç, T., Alişan, C., and Tulu, N. 1995. The age and the microflora of the coal beds in the Balkaya region (Erzurum-Oltu). In: Second International Turkish Geology Workshop, “Work in progress on the geology of Turkey”, 6–8 September, 1995, 76. Cumhuriyet University, Sivas.
Bouchet, P., Bank, R., and Gofas, S. 2018. Bithynia Leach, 1818. In: MolluscaBase (2017). World Register of Marine Species at http://www.marinespecies.org/aphia.php?p=taxdetails&id=182698 [accessed on 2018-01-14].
Bozkuş, C. 1990. Stratigraphy of northeast (Kömürlü) Oltu-Narman Tertiary basin. Geological Bulletin of Turkey 33: 47–56.
Brongniart, M. 1810. Sur des terrains qui paraissent avoir été formés sous l’eau douce. Annales du Muséum d’histoire naturelle 15: 357–405.
Brusina, S. 1902. Iconographia molluscorum fossilium in tellure tertiaria Hungariae, Croatiae, Slavoniae, Dalmatiae, Bosniae, Herzegovinae, Serbiae et Bulgariae inventorum. 30 pp. Officina Soc. Typographicae, Agram.
Bulić, J. and Jurišić-Polšak, Z. 2009. Macropalaeontology and stratigraphy of lacustrine Miocene deposits at Crnika beach on the Island of Pag (Croatia). Geologica Croatica 62: 135–155. Crossref
Burmeister, H. 1837. Handbuch der Naturgeschichte. Zum Gebrauch bei Vorlesungen. 2. Abt. Zoologie. 858 pp. T.C.F. Enslin, Berlin.
Charpentier, J. de 1837. Catalogue des mollusques terrestres et fluviatiles de la Suisse. Formant la seconde partie de la faune Helvétique. Nouveaux Mémoires de la Société Helvétique des Sciences Naturelles 1: 1–28.
Cox, L.R. 1960. Thoughts on the classification of the Gastropoda. Proceedings of the Zoological Society of London 33: 239–261.
Cuvier, G. 1795. Second Mémoire sur l’organisation et les rapports des animaux à sang blanc, dans lequel on traite de la structure des Mollusques et de leur division en ordre, lu à la société d’Histoire Naturelle de Paris, le 11 prairial an troisième. Magazin Encyclopédique, ou Journal des Sciences, des Lettres et des Arts 2: 433–449.
Dall, W.H. 1870. On the genus Pompholyx and its allies, with a revision of the Limnaeidae of authors. Annals of the Lyceum of Natural History of New York 9: 333–361. Crossref
Draparnaud, J.P.R. 1801. Tableau des mollusques terrestres et fluviatiles de la France. 116 pp. Renaud, Bossange, Masson & Besson, Montpellier, Paris.
Duméril, A.M.C. 1805. Zoologie analytique, ou méthode naturelle de classification des animaux, rendue plus facile à l’aide de tableaux synoptiques. 344 pp. Allais, Paris.
Dunker, W. 1848. Ueber die in der Molasse bei Günzburg unfern Ulm vorkommenden Conchylien und Pflanzenreste. Palaeontographica 1: 155–168.
Esu, D. and Girotti, O. 2010. The late Oligocene molluscan fauna from Otranto (Apulia, southern Italy): an example of alternating freshwater, lagoonal and emerged environments. Palaeontology 53: 137–174. Crossref
Férussac, A.E.J.P.J.F. d’A. de 1821–1822. Tableaux systématiques des animaux mollusques classés en familles naturelles, dans lesquels on a établi la concordance de tous les systèmes; suivis d’un prodrome général pour tous les mollusques terrestres ou fluviatiles, vivants ou fossiles. 110 pp. Bertrand, Paris.
Férussac, A.E.J.P.J.F. d’A. de 1823. Histoire naturelle générale et particulière des mollusques terrestres et fluviatiles. Livraison 20, Nérites fossiles. pl. 2. J.-B. Baillière, Paris.
Fischer, J.-C. 2000. La malacofauna de Sansan. In: L. Ginsburg (ed.), La faune miocéne de Sansan et son environment. Mémoires du Muséum national d‘Histoire naturelle 183: 129–154.
Fitzinger, L.I. 1833. Systematisches Verzeichniß der im Erzherzogthume Oesterreich vorkommenden Weichthiere, als Prodrom einer Fauna derselben. Beiträge zur Landeskunde Oesterreich’s unter der Enns 3: 88–122.
Fleming, J. 1820. Mollusca. In: D. Brewster (ed.), The Edinburgh Encyclopædia. In eighteen volumes, 14, 598–635. Blackwood, Edinburgh.
Froriep, L.F. 1806. C. Dumeril‘s Analytische Zoologie. Aus dem Französischen mit Zusätzen. 344 pp. Landes-Industrie-Comptoir, Weimar.
Glöer, P. 2002. Mollusca I. Süßwassergastropoden Nord- und Mittelweuropas. Bestimmungsschlüssel, Lebensweise, Verbreitung. 2., neubearbeitete Auflage. 327 pp. ConchBooks, Hackenheim.
Glöer, P. and Pešić, V. 2006. On the identity of Bithynia graeca Westerlund, 1879 with the description of three new Pseudobithynia n. gen. species from Iran and Greece (Gastropoda: Bithyniidae). Malakologische Abhandlungen 24: 29–36.
Glöer, P. and Pešić, V. 2007. The Bithynia species from Skadar lake (Montenegro) (Gastropoda: Bithyniidae). Mollusca 25: 7–12.
Glöer, P. and Yildirim, M.Z. 2006. Some records of Bithyniidae from Turkey with the description of Bithynia pesicii n. sp. (Gastropoda: Bithyniidae). Malakologische Abhandlungen 24: 37–42.
Golikov, A.N. and Starobogatov, Y.I. 1975. Systematics of prosobranch gastropods. Malacologia 15: 185–232.
Gottschick, F. 1920. Die Land- und Süßwassermollusken des Tertiärbeckens von Steinheim am Aalbuch. Archiv für Molluskenkunde 52: 33–66, 108–117, 163–177.
Gray, J.E. 1840. Shells of molluscous animals. Synopsis of the Contents of the British Museum 42: 105–152.
Gray, J.E. 1857. A Manual of the Land And Fresh-water Shells of the British Islands. xvi + 335 pp. Longman, Brown, Green, Longmans, and Robert, London.
Gude, G.K. 1913. The helicoid land shells of the Fiji Islands, with definitions of three new genera and descriptions of four new species. Proceedings of the Malacological Society of London 10: 325–330.
Gürlek, M.E. 2018. Pseudobithynia guldeni sp. n., a new gastropod species from the Mediterranean region of Turkey (Gastropoda: Truncatelloidea). Zoology in the Middle East 64: 64–67. Crossref
Harzhauser, M. and Binder, H. 2004. Pannonian Molluscs from the sections Richardhof and Eichkogel in the Vienna Basin (Austria, Late Miocene). Archiv für Molluskenkunde 133: 109–165. Crossref
Harzhauser, M. and Kowalke, T. 2001. Early Miocene brackish water Mollusca from the Eastern Mediterranean and from the Central Paratethys—a faunistic and ecological comparison by selected faunas. Journal of the Czech Geological Survey 46: 267–287.
Harzhauser, M. and Mandic, O. 2008. Neogene lake
systems of Central and South-Eastern Europe: Faunal diversity, gradients
and interrelations. Palaeogeography,
Palaeoclimatology, Palaeoecology 260: 417–434. Crossref
Harzhauser, M. and Neubauer, T.A. 2018. Opole (Poland)—a key locality for middle Miocene terrestrial mollusc faunas. Bulletin of Geoscience 93: 71–146. Crossref
Harzhauser, M., Kowalke, T., and Mandic, O. 2002. Late Miocene (Pannonian) gastropods of Lake Pannon with special emphasis on early ontogenetic development. Annalen des Naturhistorischen Museums Wien 103A: 75–141.
Harzhauser, M., Mandic, O., Büyükmeriç, Y., Neubauer, T.A., Kadolsky, D., and Landau, B.M. 2016. A Rupelian mangrove swamp mollusc fauna from the Thrace Basin in Turkey. Archiv für Molluskenkunde 145: 23–58. Crossref
Harzhauser, M., Neubauer, T.A., Georgopoulou, E., and Harl, J. 2014b. The Early Miocene (Burdigalian) mollusc fauna of the North Bohemian Lake (Most Basin). Bulletin of Geosciences 89: 819–908. Crossref
Harzhauser, M., Neubauer, T.A., Gross, M., and Binder, H. 2014a. The early Middle Miocene mollusc fauna of Lake Rein (Eastern Alps, Austria). Palaeontographica A 302: 1–71.
Harzhauser, M., Neubauer, T.A., Mandic, O., Zuschin, M., and Ćorić, S. 2012. A Middle Miocene freshwater mollusc assemblage from an intramontane Alpine lake (Aflenz Basin, Eastern Alps, Austria). Paläontologische Zeitschrift 86: 23–41. Crossref
Haszprunar, G. 2014. A nomenclator of extant and fossil taxa of the Valvatidae (Gastropoda, Ectobranchia). ZooKeys 377: 1–172.Crossref
Herrmannsen, A.N. 1846–1847. Indicis generum malacozoorum primordia. Nomina subgenerum, generum, familiarum, tribuum, ordinum, classium; adjectis auctoribus, temporibus, locis systematicis atque literariis, etymis, synonymis. Praetermittuntur Cirripedia, Tunicata et Rhizopoda. Vol. 1, xxvii + 637 pp. Fischer, Cassellis.
Hüsing, S.K., Zachariasse, W.-J., van Hinsbergen, D.J.J., Krijgsman, W., Inceöz, M., Harzhauser, M., Mandic, O., and Kroh, A. 2009. Oligocene Miocene basin evolution in SE Anatolia, Turkey: constraints on the closure of the eastern Tethys gateway. Geological Society, London, Special Publications 311: 107–132. Crossref
Iredale, T. 1936. Australian Molluscan Notes No. 2. Records of the Australian Museum 19: 267–340. Crossref
İslamoğlu, Y. 2008. Molluscan biostratigraphy and paleo-environmental reconstruction of Oligocene deposits in the Denizli and Kale-Tavas subbasins (SW Turkey). Geodiversitas 30: 261–285.
Jekelius, E. 1944. Sarmat und Pont von Soceni (Banat). Memoriile Institutului Geologic al Romaniei 5: 1–167.
Kadolsky, D. 2015. Eine Molluskenfauna aus der Unteren Bunten Molasse des Alpenvorlandes (Bayern, Oligozän): Die Fundstelle Habach 5. Geologica Saxonica 61: 53–72.
Kara-Gülbay, R. 2015. Organic geochemical and petrographical characteristics of coal bearing Oligo-Miocene sequence in the Oltu-Narman Basin (Erzurum), NE Turkey. International Journal of Coal Geology 149: 93–107. Crossref
Karayigit, A.İ., Kerey, İ.E., and Bozkuş, C. 2002. Depositional environments of Oligo/Miocene coal-bearing strata and coal quality from the Oltu-Balkaya Basin, northeastern Turkey. Energy Sources 24: 653–665. Crossref
Kerney, M. 1999. Atlas of the Land And Freshwater Molluscs of Britain And Ireland. 264 pp. Harley, Colchester.
Klein, A. von 1853. Conchylien der Süsswasserkalkformation Württembergs. Jahreshefte des Vereins für vaterländische Naturkunde in Württemberg 9: 203–223.
Kojumdgieva, E. and Sapundgieva, V. 1981. Biostratigraphie de l’Oligocène du bassin de la Haute Thrace d’après les Mollusques. Geologica Balcanica 11: 93–114.
Lamarck, J.B.P.A. de M. de 1816. Tableau encyclopédique et méthodique des trois règnes de la nature. Vingt-troisième partie. Mollusques et polypes divers. pp. 1–16, 391–488. Agasse, Paris.
Landau, B.M., Harzhauser, M., İslamoğlu, Y., and Da Silva, C.M. 2013. Systematics and palaeobiogeography of the gastropods of the middle Miocene (Serravallian) Karaman Basin of Turkey. Cainozoic Research 11–13: 3–576.
Linnaeus, C. 1758. Systema naturae per regna tria naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis locis. T. 1. 10th edition, 824 pp. Laurentius Salvius, Holmiae.
Martens, E. von 1879. Die Gattung Neritina. Systematisches Conchylien-Cabinet von Martini und Chemnitz 2 (10): 1–303. Crossref
Möllendorff, O. von 1873. Beiträge zur Fauna Bosniens. 73 pp. Inauguraldissertation, Görlitz.
Montfort, D. de 1810. Conchyliologie systématique, et classification méthodique des coquilles; offrant leurs figures, leur arrangement générique, leurs descriptions caractéristiques, leurs noms; ainsi que leur synonymie en plusieurs langues. Ouvrage destiné à faciliter l’étude des coquilles, ainsi que leur disposition dans les cabinets d’histoire naturelle. Coquilles univalves, non cloisonnées, 2. 676 pp. Schoell, Paris.
Morton, J.E. 1955. The evolution of the Ellobiidae with a discussion on the origin of the Pulmonata. Proceedings of the Zoological Society of London 125: 127–168. Crossref
Müller, O.F. 1773–1774. Vermivm terrestrium et fluviatilium, seu animalium infusoriorum, helminthicorum et testaceorum, non marinorum, succincta historia. xxxiii + 135, xxxvi + 214 pp. Heineck & Faber, Copenhagen & Leipzig.
Murray, A.M. and Hoşgör, I., 2012. An early Oligocene elopiform fish from a new locality in Eastern Anatolia, Turkey. Journal of Vertebrate Paleontology 32: 296–303. Crossref
Neubauer, T.A., Harzhauser, M., Kroh, A., Georgopoulou, E., and Mandic, O. 2015. A gastropod-based biogeographic scheme for the European Neogene freshwater systems. Earth-Science Reviews 143: 98–116. Crossref
Neubauer, T.A., Mandic, O., and Harzhauser, M. 2016. The freshwater mollusk fauna of the Middle Miocene Lake Drniš (Dinaride Lake System, Croatia): a taxonomic and systematic revision. Austrian Journal of Earth Sciences 108: 15–67.
Neubauer, T.A., Mandic, O., Harzhauser, M., and Hrvatović, H. 2013. A new Miocene lacustrine mollusc fauna of the Dinaride Lake System and its palaeobiogeographic, palaeoecologic, and taxonomic implications. Palaeontology 56: 129–156. Crossref
Noulet, J.-B. 1854. Mémoires sur les coquilles fossiles des terrains d’eau douce du Sud-Ouest de la France. 127 pp. Victor Masson, Paris.
Oppenheim, P. 1891. Beiträge zur Kenntniss des Neogen in Griechenland. Zeitschrift der Deutschen Geologischen Gesellschaft 43: 421–487.
Oppenheim, P. 1919. Das Neogen in Kleinasien. Zeitschrift der Deutschen Geologischen Gesellschaft 70: 1–210.
Pfeiffer, C. 1828. Naturgeschichte deutscher Land- und Süsswasser-Mollusken. Dritte Abtheilung. vi + 84 pp. Landes-Industrie-Comptoir, Weimar.
Poliński, W. 1929. Limnoloshka ispitivanja Balkanskog Poluostrva. I. Reliktna fauna gasteropoda Ochridskog Jezera. Glas Srpske Kraljevske Akademije 137: 129–182.
Rafinesque, C.S. 1815. Analyse de la nature ou tableau de l’univers et des corps organisés. 223 pp. Privately published, Palermo.
Récluz, C.A. 1841. Description de quelques espèces de nérites vivantes (2e partie, suite et fin.). Revue Zoologique par la Société Cuviérienne 1841: 337–343.
Salvador, R.B., Höltke, O., Rasser, M.W., and Kadolsky, D. 2016. Annotated type catalogue of the continental fossil gastropods in the Staatliches Museum für Naturkunde Stuttgart, Germany. Palaeodiversity 9: 15–70. Crossref
Sandberger, F. 1871–1875. Die Land-und Süßwasser-Conchylien der Vorwelt. Heft 1–3: pp. 1–96, pls. 1–12 (1870); Heft 4–5: pp. 97–160, pls. 13–20 (1871); Heft 6–8: pp. 161–256, pls. 21–32 (1872); Heft 9–10: pp. 257–352, pls. 33–36 (1873); Heft 11: pp. i–viii + 353–1000 (1875). C.W. Kreidel, Wiesbaden.
Saporta, G. de 1889. Notions stratigraphiques et paléontologiques appliquées à l’étude du gisement des plantes fossiles d’Aix-en-Provence. Annales des sciences géologiques 20: 1–60.
Schlickum, W.R. 1966. Die Molluskenfauna der Kirchberger Schichten des Jungholzes bei Leipheim/Donau. Archiv für Molluskenkunde 95: 321–335.
Schütt, H. 1962. Neue Süßwasser-Prosobranchier Griechenlands. Archiv für Molluskenkunde 91: 157–166.
Schütt, H. and Kavuşan, G. 1984. Mollusken der miozänen Süsswasserablagerungen in der Umgebung von Harmancik bei Kütahya-Bursa in Nordwestanatolien. Archiv für Molluskenkunde 114: 217–229.
Şen, Ş. 1977. La faune de rongeurs pliocenes de Çalta (Ankara, Turquie). Bulletin du Museum national d’histoire naturelle, Sciences de la Terre 61: 89–171.
Sheppard, R. 1823. Descriptions of seven new Bristish land and fresh-water shells, with observations upon many other species, including a list of such as have been found in the county of Suffolk. Transactions of the Linnean Society of London 14: 148–170. Crossref
Spyra, A. 2012. Contribution to the biology of Hippeutis complanatus (Linnaeus, 1758) (Gastropoda: Planorbidae): life cycle in Silesian woodland ponds (Southern Poland). Folia Malacologica 20: 279–287. Crossref
Stache, G. 1889. Die Liburnische Stufe und deren Grenz-Horizonte II. Nichtmarine Faunen-und Floren-Reste der Protocän-Sichten des nördlichen Verbreitungs-Gebietes. Abhandlungen der k.k. Geologischen Reichsanstalt 13: 85–170.
Stchépinsky, V. 1939. Faune miocène du vilayet de Sivas (Turquie). Maden tetkik ve arama enstitüsü yayinlarindan, Série C: Monogaphies 1: 1–63.
Stchépinsky, V. 1941. Géologie et richesses minerals de la region d’Erzincan (Turquie). Maden tetkik ve arama enstitüsü yayinlarindan, Série C: Monogaphies 2: 1–65.
Stchépinsky, V. 1946. Fossiles Caractéristiques de Turquie. Maden tetkik ve arama enstitüsä yayinlarindan, jeolojik harta materyelleri 1: 1–151.
Sturany, R. 1904. Kurze Diagnosen neuer Gastropoden. Anzeiger der kaiserlichen Akademie der Wissenschaften, mathematisch-naturwissenschaftliche Klasse 41: 115–119.
Taner, G. 1974a. Palaeontological and stratigraphical investigation of the Neogene Denizli area. Bulletin of Mineral Research and Exploration Institute of Turkey (MTA) 82: 89–126.
Taner, G. 1974b. Palaeontological and stratigraphical investigation of the Neogene Denizli area. Bulletin of Mineral Research and Exploration Institute of Turkey (MTA) 83: 145–177.
Thomä, C. 1845. Fossile Conchylien aus den Tertiärschichten bei Hochheim und Wiesbaden gesammelt und im naturhistorischen Museum zu Wiesbaden ausgestellt. Jahrbücher des Vereins für Naturkunde in Nassau 2: 125–162.
Turner, H., Kuiper, J.G.J., Thew, N., Bernasconi, R., Rüetschi, J., Wüthrich, M., and Gosteli, M. 1998. Fauna Helvetica 2. Atlas der Mollusken der Schweiz und Liechtensteins. 527 pp. Schweizerische Entomologische Gesellschaft, Neuchâtel.
Walchner, F.A. 1851. Handbuch der Geognosie zum Gebrauche bei seinen Vorlesungen und zum Selbststudium, mit besonderer Berücksichtigung der geognostischen Verhältnisse des Grossherzogthums Baden. Second Edition. 1120 pp. Christian Theodor Gross, Karlsruhe.
Welter-Schultes, F. 2012. European Non-marine Molluscs, a Guide For Species Identification. 674 pp. Planet Poster Editions, Göttingen.
Wenz, W. 1919. Zur Nomenklatur tertiärer Land- und Süßwassergastropoden. I. Nachrichtsblatt der Deutschen Malakozoologischen Gesellschaft 51: 68–76.
Wenz, W. 1923. Fossilium Catalogus I: Animalia. Gastropoda extramarina tertiaria 22: 1421–1734. W. Junk, Berlin.
Wenz, W. 1928. Fossilium Catalogus I: Animalia. Gastropoda extramarina tertiaria 38: 2231–2502. W. Junk, Berlin.
Wesselingh, F.P. and Alçiçek, H. 2010. A new cardiid bivalve from the Pliocene Baklan Basin (Turkey) and the origin of modern Ponto-Caspian taxa. Palaeontology 53: 711–719. Crossref
Wesselingh, F.P., Alçiçek, H., and Magyar, I. 2008. A Late Miocene Paratethyan type mollusc fauna from the Denizli Basin (southwestern Anatolia, Turkey) and its regional palaeobiogeographic implications. Geobios 41: 861–879. Crossref
Yilmaz, H. and Yilmaz, A. 2016. Tectonic setting and structural evolution of post-Late Cretaceous the Oltu-Balkaya basin (NE Turkey). Geological Bulletin of Turkey 59: 1–25.
Zieten, C.-H. von 1832. Die Versteinerungen Württembergs, oder naturgetreue Abbildungen der in den vollständigsten Sammlungen, namentlich der in dem Kabinet des Oberamts-Arzt Dr. Hartmann befindlichen Petrefacten, mit Angabe der Gebirgs-Formationen, in welchen dieselben vorkommen und der Fundorte, 33–64. Verlag and Lithographie der Expedition des Werkes unserer Zeit, Stuttgart.
Acta Palaeontol. Pol. 63 (2): 357–369, 2018
https://doi.org/10.4202/app.00469.2018