
Chemosymbiotic bivalves from the late Pliocene Stirone River hydrocarbon seep complex in northern Italy
STEFFEN KIEL and MARCO TAVIANI
Kiel, S. and Taviani, M. 2018. Chemosymbiotic bivalves from the late Pliocene Stirone River hydrocarbon seep complex in northern Italy. Acta Palaeontologica Polonica 63 (3): 557–568.
Seven species of chemosymbiotic bivalves are described from the late Pliocene Stirone River hydrocarbon seep complex in northern Italy, including one new species and two in open nomenclature. The known species are the solemyid Acharax doderleini, the lucinids Lucinoma persolida and Megaxinus ellipticus, and the vesicomyid Isorropodon aff. perplexum; in open nomenclature we report two lucinids, including the largest species of Lucinoma known from the Italian Pliocene to date, and a strongly inflated, large Anodontia sp. The most abundant species at the Stirone seep complex is the lucinid Megaxinus stironensis sp. nov. This Pliocene seep fauna differs from that of the well-known Miocene “Calcari a Lucina” seep deposits by lacking large bathymodiolin mussels and vesicomyid clams; instead, the dominance of the lucinid Megaxinus stironensis gives this fauna a unique character. We speculate that at the Stirone seep complex, Megaxinus had occupied the ecological niche that Meganodontia occupied at the Miocene “Calcari a Lucina” seep sites in the Mediterranean basin, and that the dominance of Megaxinus could be a wide-spread feature of Pliocene chemosynthesis-based ecosystems in Mediterranean Pliocene.
Key words: Bivalvia, Lucinidae, Vesicomyidae, hydrocarbon seep, chemosymbiosis, Pliocene, Italy, Apennines.
Steffen Kiel [steffen.kiel@nrm.se], Swedish Museum of Natural History, Department of Palaeobiology, Box 50007, 10405 Stockholm, Sweden.
Marco Taviani [marco.taviani@bo.ismar.cnr.it], Institute of Marine Sciences, Italian National Research Council, Via Gobetti 101, 40129 Bologna, Italy; Stazione Zoologica Anton Dohrn, Villa Comunale, 80121 Napoli, Italy, and Biology Department, Woods Hole Oceanographic Institution, 266 Woods Hole Rd, Woods Hole, MA 02543, USA.
Received 1 March 2018, accepted 25 April 2018, available online 26 June 2018.
Copyright © 2018 S. Kiel and M. Taviani. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Hydrocarbon seeps are places on the seafloor where fluids rich in methane and hydrogen sulfide reach the seabed (Suess 2014). These sites are often inhabited by a highly specialized macrofauna of low diversity but high biomass, which takes advantage of nutrients provided by chemoautotrophic bacteria that use methane and sulfide to synthesize organic matter (Paull et al. 1985; Childress et al. 1986; Sibuet and Olu 1998). The fauna inhabiting hydrocarbon-seeps in the Mediterranean Sea has an impoverished character (Olu et al. 2004; Ritt et al. 2010, 2011; Taviani 2011; Taviani et al. 2013) compared to those known from the continental margins of the Atlantic Ocean proper or from the Indian and Pacific oceans (Van Dover et al. 2002). In the Mediterranean basin, seep deposits and associated faunal communities have a long geologic history, with several Mesozoic examples (Lemoine et al. 1982; Gaillard et al. 1992; Kiel and Peckmann 2008), a few Eocene examples (Venturini et al. 1998; Natalicchio et al. 2015), and the well-known “Calcari a Lucina” sites of the Italian Miocene, which host a fauna with “oceanic” character (Moroni 1966; Taviani 1994, 2011, 2014; Kiel and Taviani 2017). However, the timing of the faunal change from an “oceanic” fauna in the Miocene to the present-day impoverished fauna is poorly constrained due to the scarcity of fossil examples from the intervening time interval. A hydrocarbon seep complex of late Pliocene age has been recently described in detail from strata of the Argille Azzurre (Lugagnano Formation) along the Stirone River (Fig. 1) in the Italian Northern Apennines (Cau et al. 2015). The purpose of the present contribution is to provide a detailed report of the associated chemosymbiotic bivalve fauna.
Institutional abbreviations.—MGGC, Museo Geologico Giovanni Capellini, University of Bologna, Italy; MSF, Museo Civico di Scienze Naturali, Faenza, Italy; NRM, Swedish Museum of Natural History, Stockholm, Sweden; ZMB, Zoological Museum, University of Bologna, Italy.
Geological setting
This remarkable and to-date unique Pliocene-age situation was first reported on by Taviani et al. (1997) who also provided information on its chemosymbiotic bivalve fauna (Solemya doderleini, Megaxinus cf. incrassatuts, and ?Lucinoma sp.). Geological and stratigraphic information on the Stirone River hydrocarbon seep has been added over the years (Monegatti et al. 2001; Barbieri and Cavalazzi 2005; Taviani 2014; Capozzi et al. 2013, 2017) with also some fragmentary information and figures on its chemosymbiotic bivalve fauna (Taviani 2001, 2011; Raineri 2007; Taviani and Ceregato 2009). The detailed work by Cau et al. (2015) summarized and implemented all previous information, further providing detailed lithostratigraphic logs. Discrete hydrocarbon-imprinted carbonate bodies of different thickness and degree of lithification and host muddy sediment punctuate the Stirone section at the Zanclean/Piacenzian boundary, and the benthic foraminifera and mollusk fauna indicate a depositional depth not exceeding 500 m (Cau et al. 2015). Such horizons are the source of the macropaleontological material discussed here (Fig. 1).
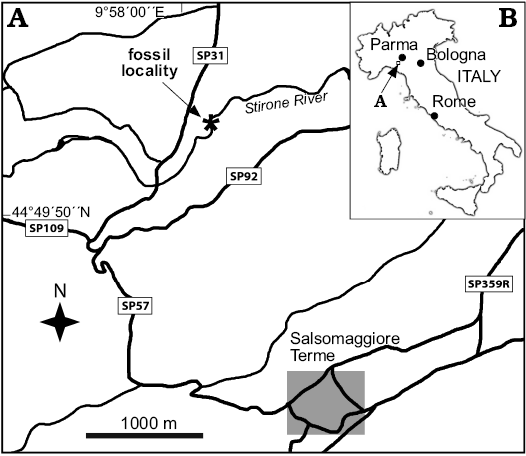
Fig. 1. Map showing location of Stirone River seep deposit on the map of Italy (B) and Salsomaggiore Terme region (A).
Material and methods
The material was collected by the authors and co-workers over many years since 1997. Specimens were coated with ammonium chloride prior to photography.
Systematic palaeontology
Class Bivalvia Linnaeus, 1758
Subclass Protobranchia Pelseneer, 1889
Family Solemyidae Gray, 1840
Genus Acharax Dall, 1908
Type species: Solemya johnsoni Dall, 1891 (by original designation); Recent, Pacific.
Acharax doderleini (Mayer, 1861)
Fig. 2.
1861 Solenomya doderleini sp. nov; Mayer 1861: 364.
1901 Solenomya doderleini Mayer; Sacco 1901: 128–129, pl. 27: 1–4.
2011 Acharax doderleini (Mayer); Taviani et al. 2011: figs. 3.3, 3.4, 3.8, 4.1, 4.2.
Material.—Three specimens: the most complete is a right valve embedded in rock matrix (MGGC.22007), 41 mm long; further complete and fragmentary specimens were seen in the field. All from the late Pliocene of Italy, Stirone seep complex.
Remarks.—The first documentation for this species at the Stirone seep complex is an articulated shell with remains of periostracum found in situ next to the main limestone block in 1997 (Taviani et al. 2011: fig. 6); other shells, highly decalcified, have been later recorded within smaller poorly-lithified lucinid mudstone a few meters above (Cau et al 2015: fig. 8). This species and other deep-water Acharax from the Italian Neogene have been discussed in detail earlier (Taviani et al. 2011).
Stratigraphic and geographic range.—Miocene to late Pliocene of northern Italy (Taviani et al. 2011).

Fig. 2. The solemyid bivalve Acharax doderleini (Mayer, 1861) from the late Pliocene of Italy, Stirone River seep deposit; MGGC.22007 view on exterior of right valve.
Subclass Heterodonta Neumayr, 1884
Family Lucinidae Fleming, 1828
Genus Megaxinus Brugnone, 1880
Type species: Lucina transversa Bronn, 1831 (subsequent designation by Pallary 1904, see Glover and Taylor 1997 for a detailed discussion); Pliocene, Italy.
Megaxinus ellipticus (Borson, 1825)
Fig. 3.
1825 Lucina elliptica sp. nov.; Borson 1825: 140, pl. 19: 5.
Material.―A single, large, articulated shell (MGGC.22008), 51 mm long and 53 mm high; from the late Pliocene of Italy, Stirone seep complex.
Remarks.—This species is common in the Mediterranean Pliocene (Chirli 2015) and our specimen agrees well with those illustrated by Sacco (1901: pl. 17). It has also been reported from the Pleistocene of Italy (Chirli 2015) but many records may prove in the future to be misidentifications since encompassing too many varieties of habitats, shallow to deep. A similar species is Megaxinus rostratus (Pecchioli, 1864), also from the Pliocene of northern Italy, which may or may not be synonym with M. ellipticus (Sacco 1901; Glover and Taylor 1997).
Stratigraphic and geographic range.—According to Sacco (1901) this species is found in late Miocene to late Pliocene sediments in northern Italy, and there are unconfirmed reports from the middle Miocene of Poland (Studencka and Studencki 1988), the late Miocene of southern Turkey (Islamoglu and Taner 2003), and the late Pliocene of Crete (Drinia et al. 2005).
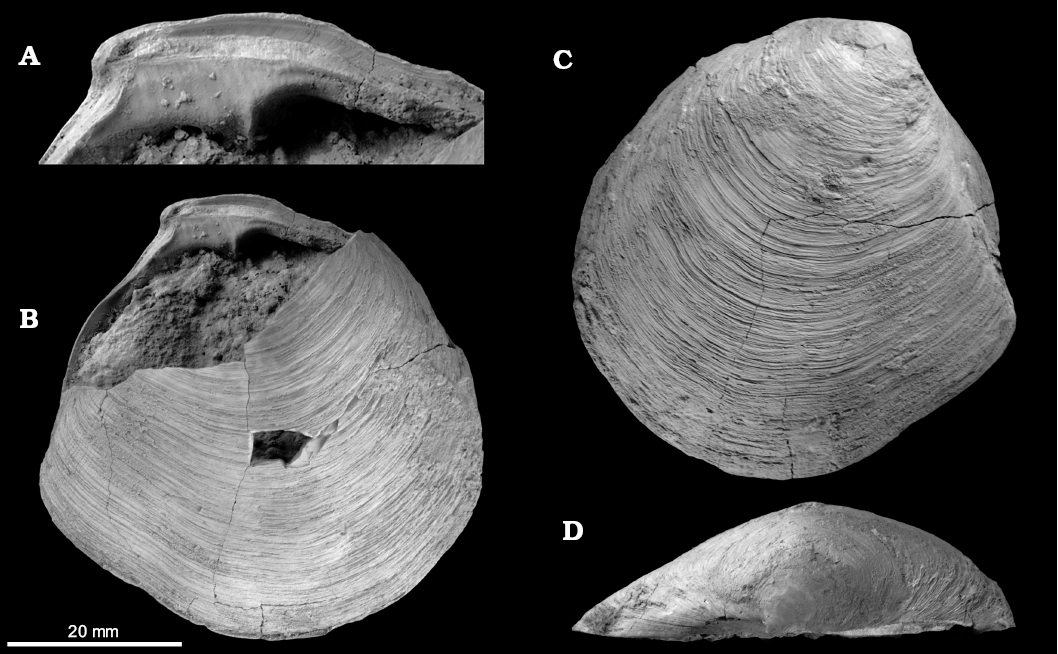
Fig. 3. The lucinid bivalve Megaxinus ellipticus (Borson, 1825) from the late Pliocene of Italy, Stirone River seep deposit. MGGC.22008, close-up of hinge region of RV (A), view of left valve exterior and right valve hinge (B), view of exterior of right valve (C); dorsal view on right valve (D).
Megaxinus stironensis sp. nov.
Figs. 4, 5.
1901 Megaxinus bellardianus (Mayer); Sacco 1901: 75–76, pl. 17: 30, 36; not 29, 32–35, 37.
Etymology: For the Stirone River.
Type material: Holotype: MGGC.22009, a shell with both valves preserved. Paratypes: articulated and single shells MGGC.22010–22013, MSF 2143, NRM Mo184001–184003; all from the type locality and horizon.
Type locality: The Stirone River seep deposit, Northern Apennines, Italy.
Type horizon: Late Pliocene part of the Lugagnano Formation.
Material.—The type material; hundreds more seen in the field.
Dimensions.—The largest specimens including the holotype reach 45 mm in length and height, individual valves are 13 mm thick.
Diagnosis.—Average-sized, moderately inflated Megaxinus, of equal height and length, with small lunule, two incisions on anterodorsal margin, strong and narrow hinge plate, and very long anterior adductor muscle scar.
Description.—Shells of average size for the genus, almost circular, of equal height and length, moderately inflated (T/L = 0.29), with blunt, elevated, prosogyrate umbo. Posterodorsal area broad, bordered by indistinct, broad, shallow groove resulting in a slight angulation on the posterior margin, another indistinct, broad and shallow groove running from the umbo to the center of the ventral margin, where it results in a shallow indentation. Lunule very small, surrounded by two incisions on the anteroventral margin; ligament external, elongate in a sunken groove. Outer shell surface with irregular commarginal growth increments. Hinge plate strong and narrow, edentulous; nymph plate long and thin. Inner shell surface with few or many pustules that are arranged radially toward the ventral margin; in some specimens the pustules are so numerous that they give the interior a granulate appearance; pallial blood vessel scar usually distinct; anterior adductor muscle scar broad and very elongate, tapering ventrally, reaching the ventral quarter of the shell, close to pallial but detached from it for ¾ of its length; posterior adductor muscle scar indistinct, irregular ovate in outline. Pallial line entire, crenulate; area beyond pallial line crenulate close to pallial line but mostly smooth.
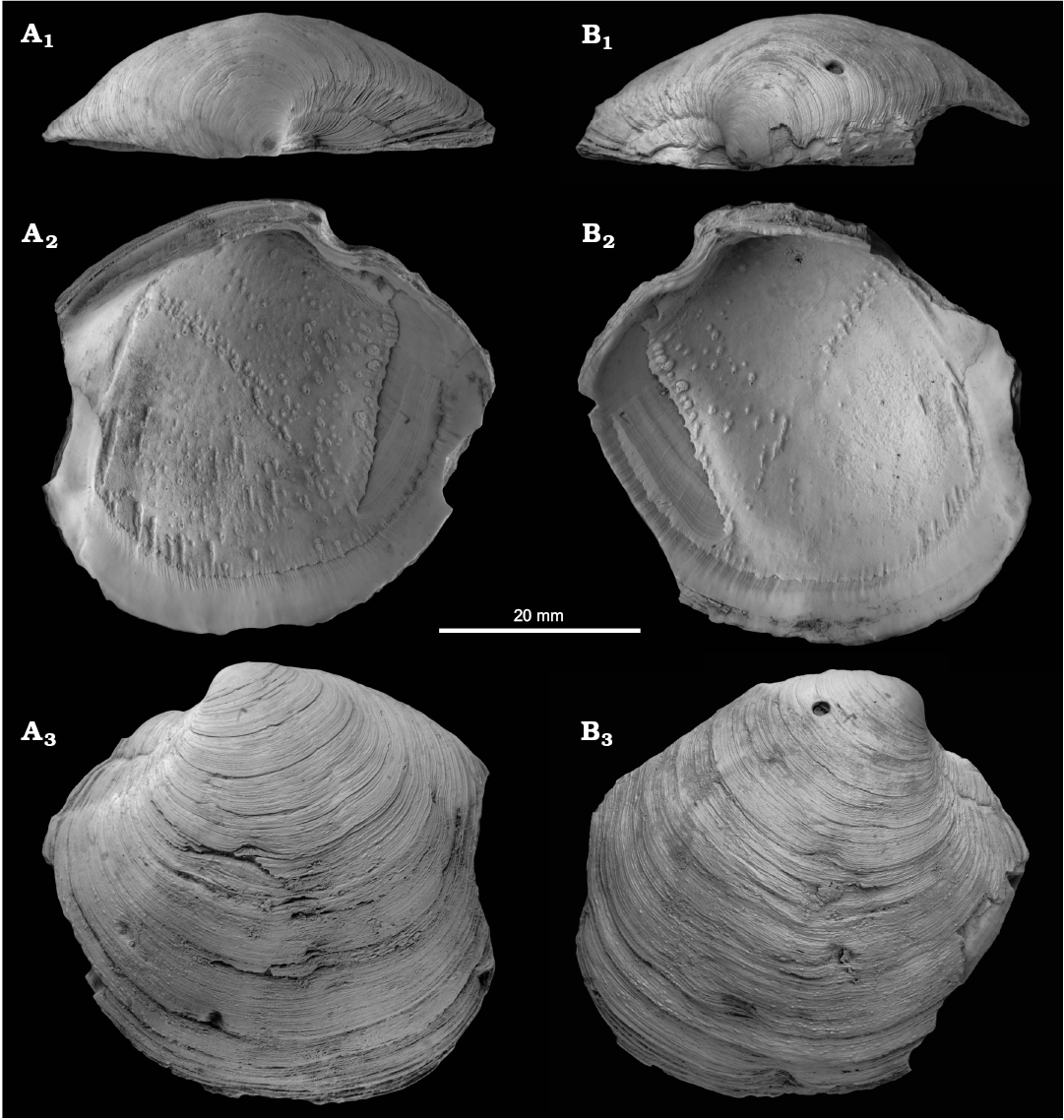
Fig. 4. The lucinid bivalve Megaxinus stironensis sp. nov. from the late Pliocene of Italy, Stirone River seep deposit. MGGC.22009, holotype, left (A) and right (B) valves in dorsal views (A1, B1), views on the interior including hinge, muscle attachment scars and blood vessel attachment scar (A2, B2), and views of exterior (A3, B3).
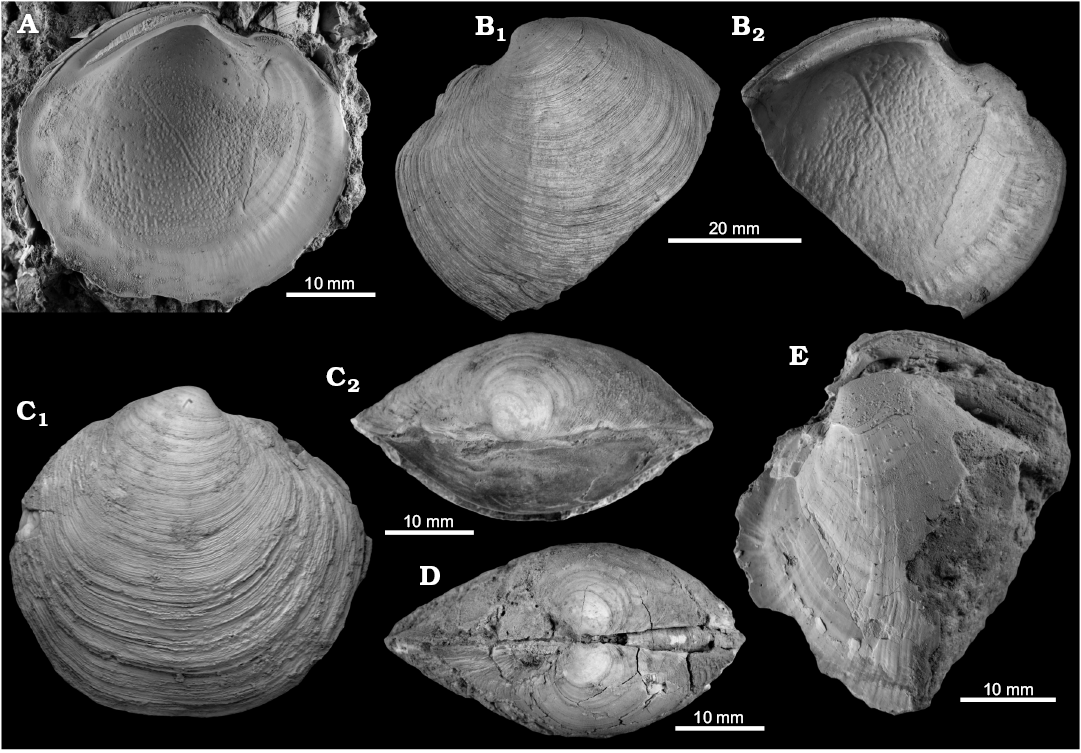
Fig. 5. The lucinid bivalve Megaxinus stironensis sp. nov. from the late Pliocene of Italy, Stirone River seep deposit. A. MGGC.22010, interior of left valve. B. MGGC.22011, exterior (B1) and interior (B2) of left valve of a large specimen. C. MGGC.22012, exterior of right valve (C1) and dorsal view (C2) of articulated specimen. D. NRM Mo184001, apical view of articulated specimen. E. MGGC.22013, internal mold showing strong radial striation on the thin veneer of remaining shell; this type of preservation is not uncommon at the Stirone River.
Remarks.—This is the most common species at the Stirone River seep deposit and it also occurs in large numbers in shell beds in the surrounding mudstone. Sacco (1901: 75–76) assigned numerous specimens of middle Miocene to late Pliocene age to Megaxinus bellardianus (Mayer, 1864). These specimens show quite a diversity of shapes, some being similar and others being dissimilar to the specimens from the Stirone seep deposit. Given the rich and cryptic present-day diversity of this genus and related lucinid genera (Glover and Taylor 1997; Taylor and Glover 2005) we prefer to apply to a more narrow species concept to the Neogene Megaxinus than Sacco’s (1901) approach of lumping most specimens into M. bellardianus, although a full revision of Neogene Megaxinus is beyond the scope of this study. Mayer (1864: 27) wrote that the type specimen of M. bellardianus is the specimen illustrated by Michelotti (1847: pl. 4: 10). That specimen is of Miocene age and the provided drawing indicates that it has a more pronounced posterodorsal ridge, and a more pointed and less prosogyrate beak than the specimens from the Stirone seep deposit concerned here. Thus due to the differences in shell shape, we consider the Stirone specimens to be distinct from M. bellardianus and introduce the new species M. stironensis for them. Among the many specimens of M. bellardianus illustrated by Sacco (1901: pl. 17: 30, 36) resemble the Stirone specimens in every aspect, and interestingly are from the late Pliocene (Piacenzian) of Vezza d’Alba, and hence of similar age as the Stirone specimen. We include those specimens from Vezza d’Alba in Megaxinus stironensis sp. nov.
The extant Megaxinus yemenensis Glover and Taylor, 1997 differs from M. stironensis by being elongate-oval rather than round, the extant M. appendiculatus, M. unguiculus, and the Neogene Italian M. rostratus (Pecchioli, 1864) and M. transversus (Bronn, 1831) differ from M. stironensis in having a truncate posterior margin and a more pointed anterior margin (i.e., Sacco 1901; Glover and Taylor 1997). Among the geographically close, extant species from the Gulf of Oman, M. omanensis (Smith, 1906) differs from M. stironensis by having a higher and more angular shell and a smaller and less prosogyrate beak, whereas M. arabicus Glover and Taylor, 1997 has a lower shell with pointed anterior margin and a narrower anterior adductor muscle scar (Glover and Taylor 1997).
Similar specimens associated with Pliocene whale-fall communities around Italy (Dominici et al. 2009; Danise et al. 2010, 2014) have been assigned to Megaxinus incrassatus du Bois de Montpéreux, 1831. However, there are three reasons why we find this questionable. First, the type specimen of M. incrassatus is from a middle Miocene locality on the Volyn’-Podollya Plateau in the western Ukraine. The specimen was described as “not well preserved” but the illustration shows a very nicely preserved shell (du Bois de Montpéreux 1831: 58, pl. 6: 1–3). The illustration is thus most likely an idealized reconstruction that should not be taken at face value. Second, specimens of Megaxinus incrassatus from the type locality or nearby illustrated by Friedberg (1934–1936: pl. 20: 1, 2), which are probably the best available approximation to the type specimens, have a shorter anterior adductor muscle scar and much smaller umbones. Third, the type locality is of Miocene rather than Pliocene age, and Sacco (1901) noted that M. incrassatus is rare in the Miocene of Italy, and was unknown to him from Pliocene deposits. The specimen illustrated from the Orciano Pisano whale fall (Danise et al. 2010: fig. 7A) has a shallower lunule and a longer anterodorsal margin than Megaxinus stironensis, and may be yet a different (or new) species.
Stratigraphic and geographic range.—Late Pliocene of northern Italy.
Genus Lucinoma Dall, 1901
Type species: Lucina filosa Stimpson, 1851 (by original designation); Recent, northwest Atlantic.
Lucinoma persolida (Sacco, 1901)
Figs. 6, 7.
1901 Dentilucina persolida sp. nov.; Sacco 1901: 82–83, pl. 19: 1, 2.
2011 Lucinoma sp.; Taviani 2011: fig. 3h.
Material.—Four specimens; the largest specimen (MGGC. 22014) consists of two disarticulated valves, 50 mm long and 46 mm high; the others (MGGC.22015, NRM Mo184004, NRM Mo184005) are articulated and single valves partly embedded in rock matrix; from the late Pliocene of Italy, Stirone seep complex.
Remarks.—Lucinoma persolida is distinct from the two extant Mediterranean Lucinoma species, L. kazani and L. borealis, by being less inflated and by having a more distinctive lunula. Among the two, L. borealis is more similar to Lucinoma persolida because of its similar outline and its straight anterior adductor muscle scar that stays close to the pallial line. The extant L. gagei Oliver and Holmes, 2006 from the Oman margin differs from L. persolida by its narrower anterior adductor muscle scar and its much more distinct posterodorsal groove.
Among fossil species, Lucinoma perusina (Sacco, 1901) from the classical Miocene “Calcare a Lucina” deposits throughout Italy (cf. Moroni 1966; Taviani 1994; Kiel and Taviani 2017) is more inflated, has less protruding umbones, and its posterodorsal area is more well-defined, by being broader and flatter, and more angular. Quite similar to L. persolida regarding shell outline is the early Miocene Japanese Lucinoma shinokii Hirayama, 1954, but it is more strongly inflated than L. persolida and its internal features are unknown (Hirayama 1954). Lucinoma acutilineata (Conrad 1849), the most common lucinid at Pliocene seep deposits in Japan (Majima et al. 2005a) differs from L. persolida by its more angular outline of the whorl. Compared to the recently described Lucinoma saetheri from Miocene seep deposits in New Zealand (Amano et al. 2018), L. persolida has sharper ribs and a lower shell with more prominent umbones.
Stratigraphic and geographic range.—Late Pliocene of northern Italy.
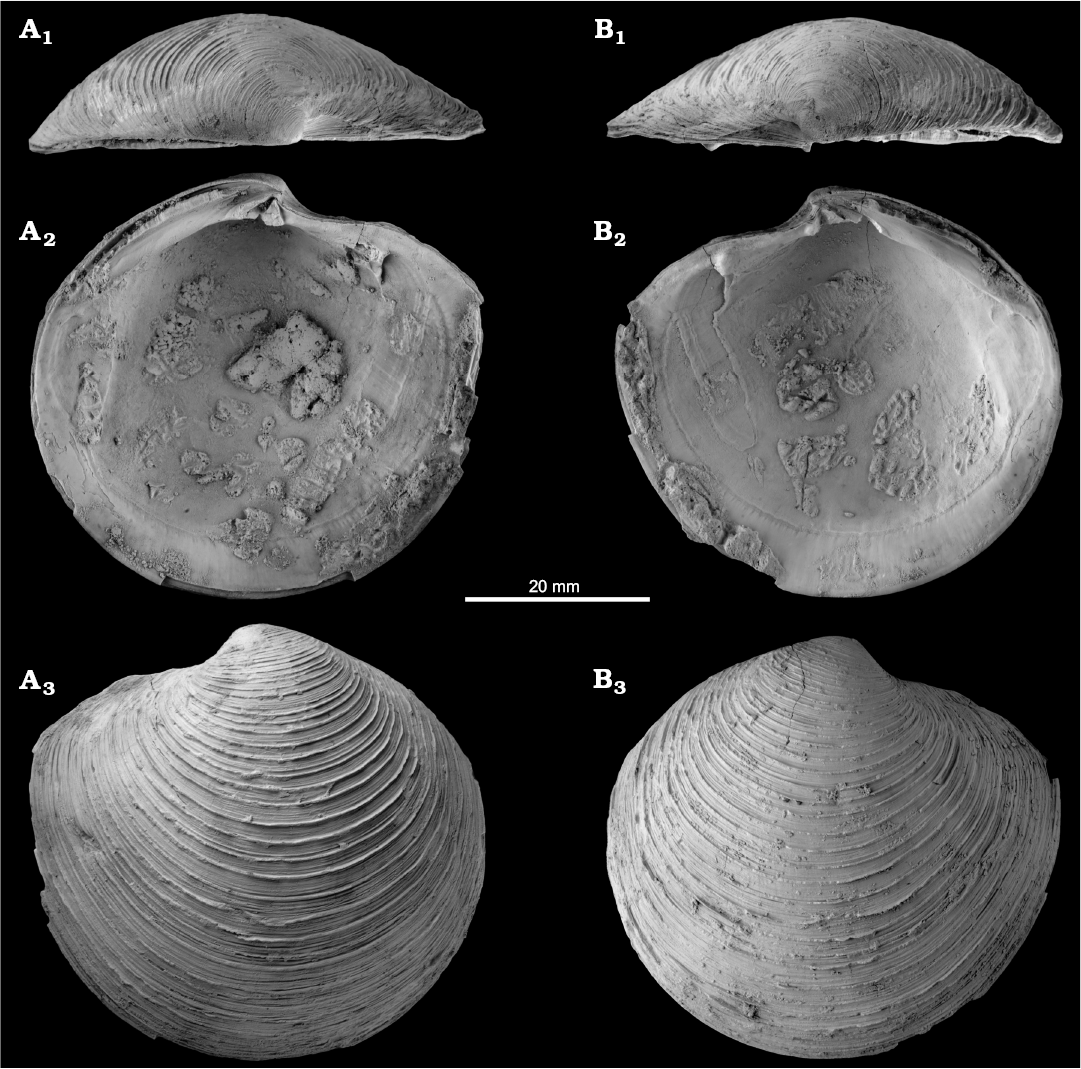
Fig. 6. The lucinid bivalve Lucinoma persolida (Sacco, 1901) from the late Pliocene of Italy, Stirone River seep deposit. MGGC.22014, left (A) and right (B) valves; in dorsal views (A1, B1), views of the interior including hinge and muscle attachment scars (A2, B2), and views of exterior (A3, B3).
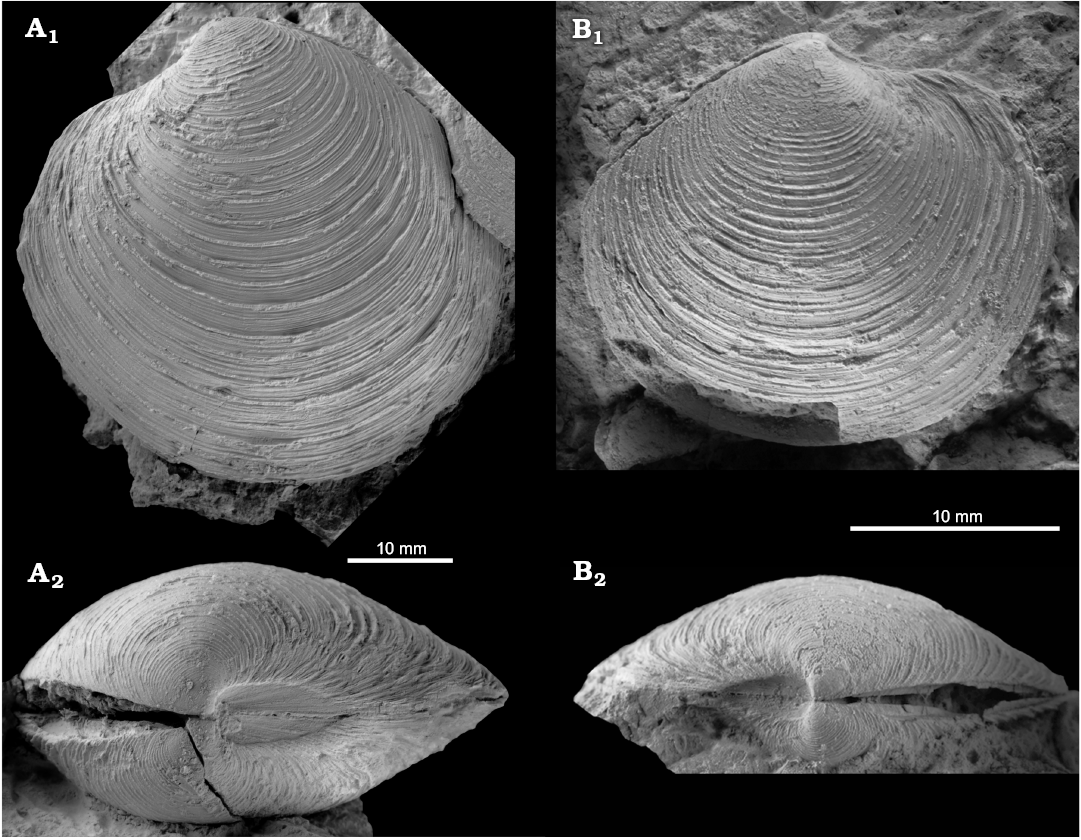
Fig. 7. The lucinid bivalve Lucinoma persolida (Sacco, 1901) from the late Pliocene of Italy, Stirone River seep deposit. A. MGGC.22015, small articulated specimen, view of right valve (A1), dorsal view (A2). B. NRM Mo184004, articulated specimen, view of left valve (B1), dorsal view (B2).
Lucinoma sp.
Fig. 8.
Material.—MGGC.22016, a single articulated, thick-shelled specimen, 70 mm long, 67 mm high, and 39 mm thick; from the late Pliocene of Italy, Stirone seep complex.
Remarks.—The only known specimen was previously figured as Lucina sp. by Taviani and Ceregato (2009). It is very similar to the middle–late Miocene Lucinoma perusina, from which it may differ by its broader and shorter lunule and less distinct posterodorsal area. To date, this is the largest deep-water chemosymbiotic lucinid known in the Mediterranean after the Miocene. It could be a new species but we prefer at present to leave in open nomenclature.
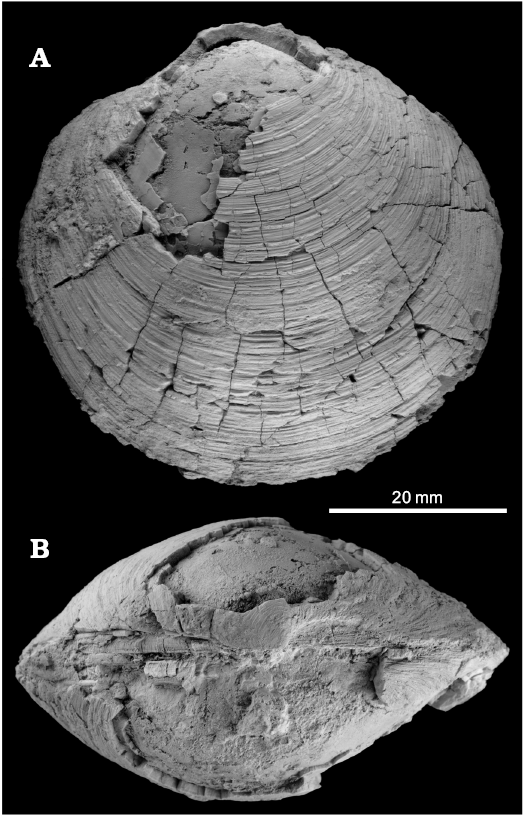
Fig. 8. The large lucinid bivalve Lucinoma sp. from the late Pliocene of Italy, Stirone River seep deposit. MGGC.22016, view of left valve (A), dorsal view (B).
Genus Anodontia Link, 1807
Type species: Anodontia alba Link, 1807 (by monotypy); Recent, Caribbean Sea.
Anodontia sp.
Fig. 9.
Material.―Two specimens: MGGC.22018 is an articulated shell reaching 25 mm in length, MGGC.22017 is a 23 mm long left valve; from the late Pliocene of Italy, Stirone seep complex.
Description.—Small to medium-sized Anodontia (sensu lato), strongly inflated (T/L ca. 0.35), umbones elevated, somewhat prosogyrate; posterodorsal area poorly defined; no lunule; external surface with irregular, commarginal growth lines. Hinge plate very narrow and edentulous, ligament long and external, sunken in narrow groove, ligament plate thin.
Remarks.—The Anodontia group is known to host a remarkable cryptic diversity (Taylor and Glover 2005). Sacco (1901) and Chirli (2015) assigned specimens of Anodontia (sensu lato) from the Pliocene of Italy to the extant shallow-water species Anodontia fragilis (Philippi, 1836). But most likely the Stirone representative is a new taxon; unfortunately, features of the pallial line and muscle scars are unknown, hence we keep these specimens in open nomenclature. Anodontia has been also reported from a Miocene seep deposit in northern Italy (Kiel et al. 2018). However, it is apparently excluded from modern deep-water cold seep situations in the eastern Atlantic Ocean and Mediterranean.

Fig. 9. The lucinid bivalve Anodontia sp. from the late Pliocene of Italy, Stirone River seep deposit. A. MGGC.22017, smaller specimen, close-up of edentulous hinge area of left valve (A1), dorsal view (A2), and view of outer side of left valve (A3). B. MGGC.22018, larger specimen, left valve, exterior view.
Family Vesicomyidae Dall and Simpson, 1901
Genus Isorropodon Sturany, 1896
Type species: Isorropodon perplexum Sturany, 1896 (by monotypy); Recent, eastern Mediterranean Sea.
Isorropodon aff. perplexum Sturany, 1896
Fig. 10.
Material.—Two fragmentary specimens, preserving the hinge area of a left and a right valve (MGGC.22019 and MGGC.22020); from the late Pliocene of Italy, Stirone seep complex.
Description.—Prodissoconch roundish, about 220 µm wide. Beak somewhat elevated, slightly prosogyrate; surface smooth; lunular incision present. Hinge plate narrow, three teeth in each valve; in right valve, cardinal 1 thin just anterior of beak, elongate, subparallel to dorsal margin, cardinal 3a commencing slightly anterior of beak, elongate and very thin, slightly thickened anteriorly, parallel to dorsal shell margin, bends slightly ventrally at its posterior end; cardinal 3b just posterior of 3a, equally thin, very slightly oblique to dorsal shell margin; in left valve, cardinal 2a short, parallel to anterior shell margin, connected through a thin ridge with the stout, triangular cardinal 2b; 4b elongate, thin, parallel to dorsal shell margin.
Remarks.—The hinge dentition of the two available fragmentary specimens agrees well with that of extant Mediterranean Isorropodon perplexum (Sturany, 1896) as figured by Cosel and Salas (2001) and also the prodissoconch shown here on Fig. 9B2 is similar in size and shape to that of Isorropodon perplexum. A single left valve identified as Isorropodon aff. perplexum has been reported from late Miocene (Messinian) sediments near Torino, northern Italy (Janssen and Krylova 2012) and seems to have a slightly shorter hinge plate than the late Pliocene specimens figured here. Two extant Isorropodon species are known from mud volcanoes in the Gulf of Cadiz in the NE Atlantic (Oliver et al. 2011). Among them, Isorropodon megadesmus Oliver, Rodrigues, and Cunha, 2011 differs from the Pliocene Isorropodon aff. perplexum by its higher hinge plate and a curved cardinal 3a; and Isorropodon sp. indet. has a hinge similar to that from the Stirone River but its cardinal 3b appears more undulating. Also the hinge dentition of the extant Isorropodon nyeggaensis from the North Atlantic off of northern Norway (Krylova et al. 2011) is similar to that of the Stirone specimens, although its nymph appears to be broader. At this Norwegian cold seep site, this vesicomyid (as Calyptogena sp.) occurs with Acharax and thyasirids in ca. 700 m water depth (Ivanov et al. 2010).
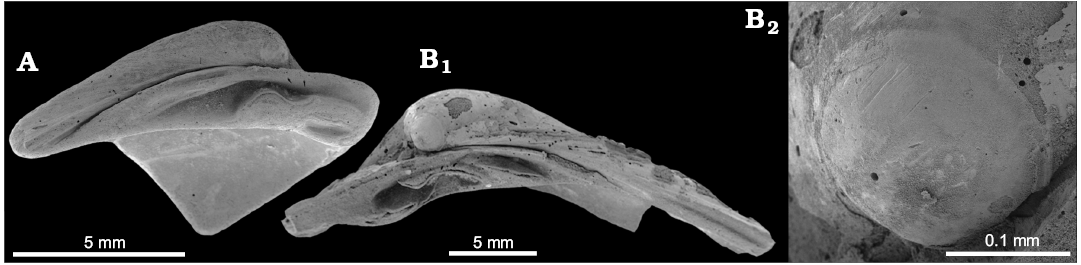
Fig. 10. The vesicomyid bivalve Isorropodon sp. from the late Pliocene of Italy, Stirone River seep deposit. A. MGGC.22019, hinge area of left valve. B. MGGC.22020, hinge area of right valve (B1), close-up of prodissoconch (B2).
Concluding remarks
The scarcity of Lucinoma at the Stirone River seep is remarkable considering that Lucinoma is very abundant at many Miocene “Calcari a Lucina” sites (Taviani 1994, 2014) as well as at many seep sites in the present-day Mediterranean Sea (Salas and Woodside 2002; Olu et al. 2004; Taviani et al. 2013). The Stirone River seep fauna is also quite distinct from the middle to late Miocene “Calcari a Lucina” fauna in northern Italy due to the lack of large vesicomyid clams and bathymodiolin mussels. These large “oceanic” species likely disappeared from the Mediterranean basin due to the Messinian Salinity crisis (cf. Taviani 2001, 2003, 2011; Olu et al. 2004; Cau et al. 2015).
The lucinid genus Megaxinus that dominates the Stirone seep deposit is still living in the Mediterranean Sea today (Glover and Taylor 1997) but has never been reported from any Mediterranean seep site (Olu et al. 2004; Ritt et al. 2010, 2011; Taviani et al. 2013; Taviani 2014). Although the precise habitat of extant Megaxinus is unknown, the lack of extant specimens with preserved soft anatomy might indicate that it is a deep burrower. It could potentially have occupied a similar ecologic niche as Meganodontia had in the Miocene “Calcari a Lucina” sites, because also Meganodontia is a deep burrower and the extant M. acetabulum lives at depths down to 472 m (Bouchet and Cosel 2004). This depth range is similar to both that of extant Megaxinus (Glover and Taylor 1997; Cosel and Bouchet 2008) and the estimated paleodepth of the Stirone seep complex (Cau et al. 2015). Whether the dominance of Megaxinus is a unique feature of this particular seep complex, or has occurred more widespread in the Mediterranean Pliocene, remains to be tested. However, remarkable in this context is the abundance of a species of Megaxinus associated with a Pliocene whale fall in northern Italy (Dominici et al. 2009; Danise et al. 2010; Danise and Dominici 2014). Lucinids are generally uncommon at whale falls, both fossil (Goedert et al. 1995; Amano and Little 2005; Kiel and Goedert 2006; Amano et al. 2007; Jenkins et al. 2017) and Recent (Smith and Baco 2003). Thus the abundance of Megaxinus at both the Stirone seep complex and the Orciano Pisano whale fall support the hypothesis that Megaxinus had a prominent role in chemosynthesis-based ecosystems in the Mediterranean Pliocene.
Pliocene seep deposits are not only rare in the Mediterranean Basin but also world-wide. Most records are from the western Pacific and are dominated by “oceanic” taxa such as large vesicomyids, bathymodiolins and lucinids, as well as the thyasirid Conchocele (Nobuhara 2003; Majima et al. 2005a, b, 2007, 2010; Wang et al. 2006; Amano and Kiel 2010, 2012). The same applies to three potential Pliocene seep deposits from the eastern Pacific (Olsson 1942; Squires 1991; Campbell 1992). Thus with Megaxinus being unknown not only from Miocene and present-day Mediterranean seeps, but also from extant and fossil seeps world-wide, the Stirone seep fauna has a unique “Mediterranean Pliocene” character.
The uniqueness of the Stirone seep complex and of its valuable accompanying fauna deserves proper measures of protection under the status of a dedicated geosite (Taviani 2014). Fortunately, the site is already located within the Stirone River Regional Park (Raineri 2007) what guarantees its safeguard for the time being.
Acknowledgements
Marco Roveri, Massimo Salmi, Simone Cau, Lorenzo Angeletti, and Alessandro Ceregato (Bologna, Italy) contributed over the years to sample the Stirone section for its macropaleontological content. We thank Kazutaka Amano (Joetsu University of Education, Japan) for insights into the Japanese seep fauna, Krzysztof Hryniewicz (Institute of Paleobiology Polish Academy of Sciences, Warsaw, Poland) and Elena Krylova (Russian Academy of Sciences, Moscow, Russia) for their helpful reviews. Financial support for field work by SK was provided by the Deutsche Forschungsgemeinschaft through grant Ki802/6-1 and the Alméns fund of the Kungliga Vetenskaps-Akademien. This is ISMAR-CNR Bologna scientific contribution n. 1956.
References
Amano, K. and Kiel, S. 2010. Taxonomy and distribution of fossil Archivesica (Vesicomyidae, Bivalvia) in Japan. The Nautilus 124: 155–165.
Amano, K. and Kiel, S. 2012. Two Neogene vesicomyid species (Bivalvia) from Japan and their biogeographic implications. The Nautilus 126: 79–85.
Amano, K. and Little, C.T.S. 2005. Miocene whale-fall community from Hokkaido, northern Japan. Palaeogeography, Palaeoclimatology, Palaeoecology 215: 345–356. Crossref
Amano, K., Little, C.T.S., and Campbell, K.A. 2018. Lucinid bivalves from Miocene hydrocarbon seep sites of eastern North Island, New Zealand, with comments on Miocene New Zealand seep faunas. Acta Palaeontologica Polonica 63: 371–382. Crossref
Amano, K., Little, C.T.S., and Inoue, K. 2007. A new Miocene whale-fall community from Japan. Palaeogeography, Palaeoclimatology, Palaeoecology 247: 236–242. Crossref
Barbieri, R. and Cavalazzi, B. 2005. Microbial fabrics from Neogene cold seep carbonates, Northern Apennine, Italy. Palaeogeography, Palaeoclimatology, Palaeoecology 227: 143–155. Crossref
Borson, S. 1820–1825. Saggio di orittografia piemontese. Memorie della Reale Accademia delle Scienze di Torino: 25, 180–229; 26, 297–364; 29, 251–318.
Bouchet, P. and Cosel, R. von 2004. The world’s largest lucinid is an undescribed species from Taiwan (Mollusca: Bivalvia). Zoological Studies 43: 704–711.
Bronn, H.G. 1831. Italiens Tertiär-Gebilde und deren organische Einschlüsse. 176 pp. Groos, Heidelberg. Crossref
Brugnone, G. 1880. Le conchiglie plioceniche selle vicinaze di Caltanisetta. Bolletino Societá Malacologia Italiana 6: 85–158.
Campbell, K.A. 1992. Recognition of a Mio-Pliocene cold seep setting from the Northeast Pacific Convergent Margin, Washington, U.S.A. Palaios 7: 422–433.
Capozzi, R., Negri, A., Reitner, J., Taviani, M., Franchi, F., and Oppo, D. (eds.) 2013. Conference Carbonate Conduits Linked to Hydrocarbon-enriched Fluid Escape, Workshop and Field Seminar, Bologna, June 28th–July 1st, 2013. 51 pp. ISMAR, Bologna.
Capozzi, R., Oppo, D., and Taviani, M. 2017. Cold seepages: An economic tool for hydrocarbon appraisal. American Association of Petroleum Geologists Bulletin 101: 617–623.
Cau, S., Franchi, F., Roveri, M., and Taviani, M. 2015. The Pliocene-age Stirone River hydrocarbon chemoherm complex (Northern Apennines, Italy). Marine and Petroleum Geology 66: 582–595. Crossref
Childress, J.J., Fisher, C.R., Brooks, J.M., Kennicutt, M.C.I., Bidigare, R.R., and Anderson, A.E. 1986. A methanotrophic marine molluscan (Bivalvia, Mytilidae) symbiosis: mussels fueled by gas. Science 233: 1306–1308. Crossref
Chirli, C. 2015. Malacofauna Pliocenica Toscana, Vol. 11. Bivalvia, Heteroconcha Hertwig, 1895. 234 pp. Published by the author, Firenze.
Conrad, R.A. 1849. Fossils from the northwestern America. In: J.D. Dana (ed.), U.S. Exploration Expedition, 1838–1842, under Charles Wilkes. Geology, Volume 10, 723–728. C. Sherman, Philadelphia.
Cosel, R. von and Bouchet, P. 2008. Tropical deep-water lucinids (Mollusca: Bivalvia) from the Indo-Pacific: essentially unknown, but diverse and occasionally gigantic. Tropical Deep-Sea Benthos, volume 25. Mémoires du Muséum National d’Histoire Naturelle 196: 115–213.
Cosel, R. von and Salas, C. 2001. Vesicomyidae (Mollusca: Bivalvia) of the genera Vesicomya, Waisiuconcha, Isorropodon and Callogonia in the eastern Atlantic and the Mediterranean. Sarsia 86: 333–366. Crossref
Dall, W.H. 1891. On some new or interesting west American shells obtained from the dredgings of the U.S. Fish Commission steamer Albatross in 1888, and from other sources. U.S. National Museum Proceedings 14: 173–191.
Dall, W.H. 1901. Synopsis of the Lucinacea and of the American species. Proceedings of the U.S. National Museum of Natural History 23: 779–833.
Dall, W.H. 1908. The Mollusca and the Brachiopoda. Bulletin of the Museum of Comparative Zoology, Harvard University 43: 205–487.
Dall, W.H. and Simpson, C.T. 1901. The Mollusca of Porto Rico. United States Fishery Commission, Bulletin 20: 351–524.
Danise, S. and Dominici, S. 2014. A record of fossil shallow-water whale falls from Italy. Lethaia 47: 229–243. Crossref
Danise, S., Dominici, S., and Betocchi, U. 2010. Mollusk species at a Pliocene shelf whale fall (Orciano Pisano, Tuscany). Palaios 25: 449–456. Crossref
Danise, S., Dominici, S., Glover, A.G., and Dahlgren, T.G. 2014. Molluscs from a shallow-water whale-fall and their affinities with adjacent benthic communities on the Swedish west coast. Marine Biology Research 10: 3–16. Crossref
Dominici, S., Cioppi, E., Danise, S., Betocchi, U., Gallai, G., Tangocci, F., Valleri, G., and Monechi, S. 2009. Mediterranean fossil whale falls and the adaptation of mollusks to extreme habitats. Geology 37: 815–818. Crossref
Drinia, H., Koskeridou, E., and Antonarakou, A. 2005. Late Pliocene benthic foraminifera and mollusks from the Atsipades Section, Central Crete; Palaeoecological distribution and use in palaeoenvironmental assessment. Geobios 38: 315–324. Crossref
du Bois de Montpéreux, F. 1831. Conchiologie fossile et aperçu géognostique des formations du plateau Wolhyni-Podolien. 76 pp. Simon Schropp & Co., Berlin.
Fleming, J. 1828. A History of British Animals, Exhibiting the Descriptive Characters and Systematical Arrangement of the Genera and Species of Quadrupeds, Birds, Reptiles, Fishes, Mollusca and Radiata of the United Kingdom; Including the Indigenous, Extirpated, and Extinct Kinds; Together with Periodical and Occasional Visitants. xxiii + 554 pp. Bell & Bradfute, Edinburgh.
Friedberg, W. 1934–1936. Mięczaki mioceńskie ziem Polskich. (Mollusca miocaenica Poloniae), Pars II. Lamellibranchiata. 283 pp. Polskie Towarzystwo Geologiczne, Kraków.
Gaillard, C., Rio, M., and Rolin, Y. 1992. Fossil chemosynthetic communities related to vents or seeps in sedimentary basins: the pseudobioherms of southeastern France compared to other world examples. Palaios 7: 451–465. Crossref
Glover, E.A. and Taylor, J.D. 1997. New species and records of Rastafaria and Megaxinus (Bivalvia: Lucinidae) from the western Indian Ocean and Red Sea, with a reappraisal of Megaxinus. Journal of Conchology 36: 1–18.
Goedert, J.L., Squires, R.L., and Barnes, L.G. 1995. Paleoecology of whale-fall habitats from deep-water Oligocene rocks, Olympic Peninsula, Washington State. Palaeogeography, Palaeoclimatology, Palaeoecology 118: 151–158. Crossref
Gray, J.E. 1840. Synopsis of the Contents of the British Museum. 370 pp. Trustees of the British Museum, London.
Hirayama, K. 1954. On some Miocene species of Lucinoma from Japan, with description of two new species. Japanese Journal of Geology and Geography 25: 101–115.
Islamoglu, Y. and Taner, G. 2003. Antalya Miyosen havzasinin Bivalvia faunasi (Bati-orta Toroslar, GB Türkiye). MTA Dergisi 127: 1–27.
Ivanov, M.V., Mazzini, A., Blinova, V., Kozlova, E., Laberg, J.-S., Matveeva, T., Taviani, M., and Kaskov, N. 2010. Seep mounds on the Southern Vøring Plateau (offshore Norway). Marine and Petroleum Geology 27: 1235–1261. Crossref
Janssen, R. and Krylova, E.M. 2012. Bivalves of the family Vesicomyidae from the Neogene Mediterranean basin (Bivalvia: Vesicomyidae). Archiv für Molluskenkunde 141: 87–113. Crossref
Jenkins, R.G., Kaim, A., Amano, K., Sakurai, K., and Matsubara, K. 2018. A new Miocene whale-fall community dominated by bathymodiolin mussel Adipicola from Hobetsu area, Hokkaido, Japan. Paleontological Research 22: 105–111. Crossref
Kiel, S. and Goedert, J.L. 2006. Deep-sea food bonanzas: Early Cenozoic whale-fall communities resemble wood-fall rather than seep communities. Proceedings of the Royal Society B 273: 2625–2631. Crossref
Kiel, S. and Peckmann, J. 2008. Paleoecology and evolutionary significance of an Early Cretaceous Peregrinella-dominated hydrocarbon-seep deposit on the Crimean Peninsula. Palaios 23: 751–759. Crossref
Kiel, S., Sami, M., and Taviani, M. 2018. A serpulid-Anodontia-dominated methane-seep deposit from the Miocene of northern Italy. Acta Palaeontologica Polonica 63: 567–575. Crossref
Kiel, S. and Taviani, M. 2017. Chemosymbiotic bivalves from Miocene methane-seep carbonates in Italy. Journal of Paleontology 91: 444–466. Crossref
Krylova, E.M., Gebruk, A.V., Portnova, D.A., Todt, C., and Haflidason, H. 2011. New species of the genus Isorropodon (Bivalvia: Vesicomyidae: Pliocardiinae) from cold methane seeps at Nyegga (Norwegian Sea, Vøring Plateau, Storrega Slide). Journal of the Marine Biological Association of the U.K. 91: 1135–1144. Crossref
Lemoine, M., Arnaud-Vanneau, A., Arnaud, H., Létolle, R., Mevel, C., and Thieuloy, J.-P. 1982. Indices possibles de paléo-hydrothermalisme marin dans le Jurassique et le Crétacé des Alpes occidentales (océan téthysien et sa marge continentale européenne): essai d’inventaire. Bulletin de la Société Géologique de France 24: 641–647. Crossref
Link, H.F. 1806–1808. Beschreibung der Naturalien. 1: 160 pp., 2: 30 pp., 3: 38 pp. Sammlung der Universität zu Rostock, Rostock.
Linnaeus, C. 1758. Systema naturæ per regna tria naturæ, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis,locis. Vol. 1. Editio decima. 824 pp. Laurentius Salvius, Holmiae.
Majima, R., Jenkins, R.G., Kase, T., Aguilar, Y.M., Nanjo, T., Wani, R., Wada, H., Fernando, A.G.S., and Hayashi, H. 2010. In situ Calyptogena colonies from Pliocene back-arc basin fills in Leyte Island, Philippines. Journal of the Geological Society of Japan 116: 15–16. Crossref
Majima, R., Kase, T., Kawagata, S., Aguilar, Y.M., Hagino, K., and Maeda, M. 2007. Fossil cold-seep assemblages from Leyte Island, Philippines. Journal of Geography 116: 643–652. Crossref
Majima, R., Nobuhara, T., and Kitazaki, T. 2005a. Review of fossil chemosynthetic assemblages in Japan. Palaeogeography, Palaeoclimatology, Palaeoecology 227: 86–123. Crossref
Majima, R., Shibata, T., Taguchi, K., Saito, H., and Wada, H. 2005b. New knowledge of a cold-seep assemblage in the Pliocene Takanabe Formation, Miyazaki Group. Fossils, The Palaeontological Society of Japan 78: 59–63.
Mayer, K. 1864. Systematisches Verzeichniss der fossilen Reste von Madeira, Porto Santo und Santo Maria. 95 pp. Published by the author, Zürich.
Mayer, M.C. 1861. Description de Coquilles fossiles des terrains tertiaires supérieurs. Journal de Conchyliologie 9: 358–373.
Michelotti, G. 1847. Description des fossiles des terrains miocènes de l’Italie septentrionale. 408 pp. Société Hollandaise des Sciences, Leiden.
Monegatti, P., Raffi, S., Roveri, M., and Taviani, M. 2001. One day trip in the outcrops of Castell’Arquato Plio-Pleistocene Basin: from the Badland of Monte Giogo to the Stirone River. In: P. Monegatti, F. Cecca, and S. Raffi (eds.), International Conference Paleobiogeography and Paleoecology (2001: Piacenza, Italy, and Castell’Arquato, Italy), May 31–June 2 2001, Excursion Guidebook, 1–22. Elsevier, Paris.
Moroni, M.A. 1966. Malacofauna del “Calcare a Lucine” di S. Sofia–Forlì. Palaeontographica Italica 60: 69–87.
Natalicchio, M., Peckmann, J., Birgel, D., and Kiel, S. 2015. Seep deposits from northern Istria, Croatia: a first glimpse into the Eocene seep fauna of the Tethys region. Geological Magazine 152: 444–459. Crossref
Neumayr, M. 1884. Zur Morphologie des Bivalvenschlosses. Sitzungsberichte der Kaiserlichen Akademie der Wissenschaften 88: 385–419.
Nobuhara, T. 2003. Cold seep carbonate mounds with Vesicomya (Calyptogena) kawamurai in slope-mud facies of the Pliocene forearc basin of the Sagara-Kakegawa area, central Japan. Paleontological Research 7: 313–328. Crossref
Oliver, P.G. and Holmes, A.M. 2006. A new species of Lucinoma (Bivalvia: Lucinoidea) from the oxygen minimum zone of the Oman Margin, Arabian Sea. Journal of Conchology 39: 63–77.
Oliver, P.G., Rodrigues, C.F., and Cunha, M.R. 2011. Chemosymbiotic bivalves from the mud volcanoes of the Gulf of Cadiz, NE Atlantic, with descriptions of new species of Solemyidae, Lucinidae and Vesicomyidae. Zookeys 113: 1–38. Crossref
Olsson, A.A. 1942. Tertiary and Quaternary fossils from the Burica Peninsula of Panama and Costa Rica. Bulletins of American Paleontology 27: 1–106.
Olu, K., Sibuet, M., Fiala-Médoni, A., Gofas, S., Salas, C., Mariotti, A., Foucher, J.-P., and Woodside, J. 2004. Cold seep communities in the deep eastern Mediterranean Sea: composition, symbiosis and spatial distribution on mud volcanoes. Deep-Sea Research I 51: 1915–1936. Crossref
Pallary, P. 1904. Addition à la faune Malacologique de la Golfe de Gabès. Journal de Conchyliologie 52: 212–248.
Paull, C.K., Jull, A.J.T., Toolin, L.J., and Linick, T. 1985. Stable isotope evidence for chemosynthesis in an abyssal seep community. Nature 317: 709–711. Crossref
Pecchioli, V. 1864. Descrizione di alcuni nuovi fossili delle argille subappennine toscane. Atti della Società Toscana di Scienze Naturali 6: 1–32.
Pelseneer, P. 1889. Sur la classification phylogenetique des pélécypodes. Bulletin scientifique de la France et de la Belgique 20: 27–52.
Philippi, R.A. 1836. Enumeratio Molluscorum Siciliae cum viventium tum in tellure Tertiaria fossilium quae in itinere suo observavit. 303 pp. Simonis Schroppii et Sociorum, Berolini.
Raineri, G. 2007. Il Parco dello Stirone un museo all’aperto. Le rocce e i fossili raccontano. 52 pp. Consorzio Parco Fluviale Regionale dello Stirone, Salsomaggiore Terme.
Ritt, B., Pierre, C., Gauthier, O., Wenzhöfer, F., Boetius, A., and Sarrazin, J. 2011. Diversity and distribution of cold-seep fauna associated with different geological and environmental settings at mud volcanoes and pockmarks of the Nile Deep-Sea Fan. Marine Biology Research 158: 1187–1210. Crossref
Ritt, B., Sarrazin, J., Caprais, J.-C., Noël, P., Gauthier, O., Pierre, C., Henry, P., and Desbruyères, D. 2010. First insights into the structure and environmental setting of cold-seep communities in the Marmara Sea. Deep-Sea Research I 57: 1120–1136. Crossref
Sacco, F. 1901. I molluschi dei terreni terziarii del Piemonte e della Liguria. Parte XXIX. 217 pp. Carlo Clausen, Torino.
Salas, C. and Woodside, J. 2002. Lucinoma kazani n. sp (Mollusca: Bivalvia): evidence of a living benthic community associated with a cold seep in the Eastern Mediterranean Sea. Deep-Sea Research I 49: 991–1005. Crossref
Sibuet, M. and Olu, K. 1998. Biogeography, biodiversity and fluid dependence of deep-sea cold-seep communities at active and passive margins. Deep-Sea Research II 45: 517–567. Crossref
Smith, C.R. and Baco, A.R. 2003. Ecology of whale falls at the deep-sea floor. Oceanography and Marine Biology: an Annual Review 41: 311–354.
Smith, E.A. 1906. Natural History Notes from R.I.M.S. “Investigator”. Series III., No.10. On Mollusca from the Bay of Bengal and the Arabian Sea. Annals and Magazine of Natural History 18: 157–175, 245–264. Crossref
Squires, R.L. 1991. New morphologic and stratigraphic information on Calyptogena (Calyptogena) gibbera Crickmay, 1929, (Bivalvia: Vesicomyidae), from the Pliocene and Pleistocene of southern California. The Veliger 34: 73–77.
Stimpson, W. 1851. A Revision of the Synonymy of the Testaceous Mollusks of New England, With Notes on Their Structure, and Their Geographical and Bathymetrical Distribution. 58 pp. Phillips, Sampson, and company, Boston.
Studencka, B. and Studencki, W. 1988. Middle Miocene (Badenian) bivalves from the carbonate deposits of the Mójcza-Pinczów Range (southern slopes of the Holy Cross Mountains. Central Poland). Acta Geologica Polonica 38: 1–44.
Sturany, R. 1896. Mollusken I (Prosobranchier und Opisthobranchier; Scaphopoden; Lamellibranchier) gesammelt von S.M. Schiff ‘Pola’ 1890–94. Denkschriften der mathematisch-naturwissenschaftlichen Klasse der Kaiserlichen Akademie der Wissenschaften 63: 1–36.
Suess, E. 2014. Marine cold seeps and their manifestations: geological control, biogeochemical criteria and environmental conditions. International Journal of Earth Sciences 103: 1889–1916. Crossref
Taviani, M. 1994. The “calcari a Lucina” macrofauna reconsidered: Deep-sea faunal oases from Miocene-age cold vents in the Romagna Apennine, Italy. Geo-Marine Letters 14: 185–191. Crossref
Taviani, M. 2001. Fluid venting and associated processes. In: G.B. Vai and I.P. Martini (eds.), Anatomy of an Orogen: the Apennines and Adjacent Mediterranean Basins, 351–366. Kluwer Academic Publishers, Dordrecht.
Taviani, M. 2003. Shaping the biogeography of the Mediterranean basin: one geologist’s perspective. Biogeographia 24: 15–22.
Taviani, M. 2011. The deep-sea chemoautotroph microbial world as experienced by the Mediterranean metazoans through time. In: J. Reitner, N.-V. Quéric, and G. Arp (eds.), Advances in Stromatolite Geobiology. Lecture Notes in Earth Sciences 131, 277–295. Springer, Berlin.
Taviani, M. 2014. Marine chemosynthesis in the Mediterranean Sea. In: S. Goffredo and Z. Dubinsky (eds.), The Mediterranean Sea: Its History and Present Challenges, 69–83. Springer, Dordrecht.
Taviani, M. and Ceregato, A. 2009. Vite primordiali negli abissi. Darwin 32: 82–87.
Taviani, M., Angeletti, L., and Ceregato, A. 2011. Chemosynthetic bivalves of the family Solemyidae (Bivalvia, Protobranchia) in the Neogene of the Mediterranean Basin. Journal of Paleontology 85: 1067–1076. Crossref
Taviani, M., Angeletti, L., Ceregato, A., Foglini, F., Froglia, C., and Trincardi, F. 2013. The Gela Basin pockmark field in the strait of Sicily (Mediterranean Sea): chemosymbiotic faunal and carbonate signatures of postglacial to modern cold seepage. Biogeosciences 10: 4653–4671. Crossref
Taviani, M., Roveri, M., Aharon, P., and Zibrowius, H. 1997. A Pliocene deepwater cold seep (Stirone River, N. Italy). In: G.B. Vai, M. Taviani, S. Conti, and P. Aharon (eds.), Cold-E-Vent. Hydrocarbon Seepage and Chemosynthesis in Tethyan Relic Basins: Products, Processes and Causes. An International Field Workshop to be held in Bologna and nearby Apennines. June 23–26/1997. Abstract with Program, 20. ISMAR, Bologna.
Taylor, J.D. and Glover, E.A. 2005. Cryptic diversity of chemosymbiotic bivalves: a systematic revision of worldwide Anodontia (Mollusca: Bivalvia: Lucinidae). Systematics and Biodiversity 3: 281–338. Crossref
Van Dover, C.L., German, C.R., Speer, K.G., Parson, L.M., and Vrijenhoek, R.C. 2002. Evolution and biogeography of deep-sea vent and seep invertebrates. Science 295: 1253–1257. Crossref
Wang, S.-W., Gong, S.-Y., Mii, H.-S., and Dai, C.-F. 2006. Cold-seep carbonate hardgrounds as the initial substrata of coral reef development in a siliciclastic paleoenvironment of southwestern Taiwan. Terrestrial, Atmospheric & Oceanic Sciences 17: 405–427. Crossref
Venturini, S., Selmo, E., Tarlao, A., and Tunis, G. 1998. Fossiliferous methanogenic limestones in the Eocene flysch of Istria (Croatia). Giornale di Geologia, serie 3a 60: 219–234.
Acta Palaeontol. Pol. 63 (3): 557–568, 2018
https://doi.org/10.4202/app.00473.2018