
New brachyuran crabs from the Aptian–Albian Romualdo Formation, Santana Group of Brazil: Evidence for a Tethyan connection to the Araripe Basin
LUDMILA A.C. PRADO, JAVIER LUQUE, ALCINA M.F. BARRETO, and A. RICHARD PALMER
Prado, L.A.C., Luque, J., Barreto, A.M.F., and Palmer, A.R. 2018. New brachyuran crabs from the Aptian–Albian Romualdo Formation, Santana Group of Brazil: Evidence for a Tethyan connection to the Araripe Basin. Acta Palaeontologica Polonica 63 (4): 737–750.
Despite the exceptional palaeontological richness of the renowned Santana Group from the Early Cretaceous Araripe Basin of Brazil, only one specimen of true crabs (Brachyura) has been discovered to date. Here we present two new genera and species of brachyuran crabs from the upper Aptian–lower Albian Romualdo Formation of the Santana Group, represented by 30 specimens from several localities across the Araripe Basin in the state of Pernambuco, Brazil. One taxon, Exucarcinus gonzagai Prado and Luque gen. et sp. nov., is similar to several endemic stem raninoidans (Orithopsidae) from the late Aptian and early Albian of Colombia and the USA, respectively, while Romualdocarcinus salesi Prado and Luque gen. et sp. nov., seems to be closer to “higher” true crabs (Eubrachyura) from the Cenomanian of Spain. The presence of taxa similar to those from the mid-Cretaceous of northern South America, southern North America, and western Europe, reinforces the view of a Tethyan marine influence in Araripe during the breakup of South America and Africa, and adds to the growing data that suggest a generic and specific degree of endemism of true crabs in tropical South America during the mid-Cretaceous.
Key words: Decapoda, Eubrachyura, Raninoida, Exucarcinus, Romualdocarcinus, Cretaceous, South America.
Ludmila A.C. Prado [prado.lac@gmail.com; ORCID ID: http://orcid.org/0000-0002-9842-7074] and Alcina Barreto [alcinabarreto@gmail.com], Departamento de Geologia, Centro de Tecnologia e Geociências, Universidade Federal de Pernambuco, 1235, 50740-533, Recife, Pernambuco, Brazil.
Javier Luque [luque@ualberta.ca; ORCID ID: http://orcid.org/0000-0002-4391-5951], Department of Biological Sciences, University of Alberta, Edmonton, Alberta T6G 2E9, Canada and Smithsonian Tropical Research Institute, Balboa-Ancón 0843-03092, Panamá, Panamá.
A. Richard Palmer [rich.palmer@ualberta.ca], Department of Biological Sciences, University of Alberta, Edmonton, Alberta T6G 2E9, Canada.
Received 23 March 2018, accepted 4 October 2018, available online 19 November 2018.
Copyright © 2018 L.A.C. Prado et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Our understanding of the diversity and geographic distributions of fossil crabs in the tropical Americas has grown considerably in the last decades, especially regarding Late Cretaceous and Cenozoic occurrences (Luque et al. 2017). Yet, little is known about the fossil record of true crabs (Brachyura) from the Early Cretaceous in South America, with only a handful of taxa reported so far. In Brazil, several fossil decapod crustaceans have been discovered in Aptian–Albian rocks of the Santana Group, Araripe Basin (mostly shrimps, see Table 1), but so far only one crab specimen has yet been reported and described as Araripecarcinus ferreirai Martins-Neto, 1987. Although only ventral and appendicular features are preserved in the holotype of A. ferreirai, its phylogenetic position and overall anatomical characteristics suggest affinities to stem raninoidans of the superfamily Necrocarcinoidea, yet with uncertain familial affinities (Luque 2015a; Schweitzer et al. 2016b; Luque et al. 2017).
Table 1. List of decapods described from the Santana Group in the Araripe Basin.
|
Taxon |
Stratigraphy |
Locality |
References |
|
Araripecarcinus
ferreirai |
Romualdo
Formation |
near the municipality of Porteiras, state of Ceará |
|
|
Beurlenia araripensis Martins-Neto and Mezzarila, 1991 (palaemonid shrimp) |
Crato Formation (upper Aptian): laminated limestone |
uncertain |
|
|
Paleomattea deliciosa Maisey and Carvalho, 1995 (sergestid shrimp) |
Romualdo Formation (upper Aptian–lower Albian): shales and calcareous concretions with fish |
municipalities of Santana do Cariri and Missão Velha, state of Ceará, and Trindade, state of Pernambuco |
Viana
and Agostinho 1995; Saraiva et al. 2009;
|
|
Brachyuran crab zoeae indet. (brachyuran crab) |
Romualdo Formation |
uncertain |
|
|
Kellnerius jamacaruensis Santana, Pinheiro, Silva, and Saraiva, 2013 (palaemonid shrimp) |
Romualdo Formation |
municipalities of Missão Velha
|
|
|
Araripenaeus timidus Pinheiro, Saraiva, and Santana, 2014 (penaeid shrimp) |
Romualdo
Formation |
municipality of Jardim, |
|
|
Sume marcosi Saraiva, Pinheiro, and Santana, 2018 (luciferid shrimp) |
Romualdo Formation (upper |
municipality of Trindade, |
|
|
Exucarcinus gonzagai gen. et sp. nov. Prado and Luque, 2018 (orithpsid crab) |
Romualdo
Formation |
municipality of Exu, |
this paper |
|
Romualdocarcinus
salesi gen. et sp. nov. Prado and Luque, 2018 |
Romualdo Formation |
municipality of Exu, |
this paper |
Several other Early Cretaceous necrocarcinoids are known from South America, such as the orithopsid crabs Bellcarcinus Luque, 2014; Colombicarcinus Karasawa, Schweitzer, Feldmann, and Luque, 2014; and Planocarcinus Luque, Feldmann, Schweitzer, Jaramillo, and Cameron, 2012; all of them from the upper Aptian and lower Albian of Colombia (Luque et al. 2012, 2017; Bermúdez et al. 2013; Karasawa et al. 2014; Schweitzer et al. 2016b; Fig. 1). Similarly, only a handful of Early Cretaceous eubrachyurans, or “higher” true crabs, have been discovered worldwide (Klompmaker 2013; Luque 2015b), with Telamonocarcinus antiquus Luque, 2015b, from the late Aptian–early Albian of Colombia representing the oldest crown eubrachyuran (Luque et al. 2017). Here we describe two new genera and species of brachyuran crabs, Exucarcinus gonzagai gen. et sp. nov. (Raninoida: Necrocarcinoidea: Orithopsidae) and Romualdocarcinus salesi gen. et sp. nov. (Eubrachyura: ?Portunoidea: ?Eogeryonidae), from upper Aptian–lower Albian rocks of the fossiliferous Romualdo Formation, Santana Group, near the municipality of Exu in the state of Pernambuco, and discuss aspects of their systematic affinities, taphonomy, palaeoenvironment, and their palaeogeographical implications.
Institutional abbreviations.—DGEO-CTG-UFPE, Palaeontology Collection of the Geology Department, Centre of Technology and Geosciences, Federal University of Pernambuco, Brazil.
Nomenclatural acts.—This published work and the nomenclatural acts it contains have been registered in ZooBank, the online registration system for the ICZN. The electronic edition of this work was published in a journal with an ISSN 0567-7920, eISSN 1732-2421, and has been archived and is available from the following digital repository: http://www.app.pan.pl/article/item/app004802018.html.
Geographic and geological setting
During the late Early Cretaceous, rapid changes in plate tectonic reconfigurations occurred leading to the separation of South America and Africa. As a result, several small basins were formed in the interior of northeastern Brazil due to the opening of the central Atlantic Ocean (Matos 1992; Assine 1994). Among them, the Araripe Basin stands out, known internationally for the fossiliferous Lagerstätten of the Crato and Romualdo formations (Maisey 1991). The Araripe Basin, located between the states of Pernambuco, Piauí, and Ceará, is formed by five sequences: Paleozoic, Cariri Formation (upper Ordovician–lower Devonian), pre-rift Brejo Santo and Missão Velha formations (Upper Jurassic), rift Abaiara Formation (Neocomian), post-rift I Barbalha, Crato, Ipubi, and Romualdo formations (upper Aptian–lower Albian) and post-rift II Araripina and Exu formations (Albian–Cenomanian) (Assine 1992, 2007; Assine et al. 2014; Fig. 1). The Romualdo Formation is constituted by a sequence of shale intercalated with sandstone at the base, followed by black shale with usually fossiliferous calcareous nodules, and layers of calcareous sandstone with macroinvertebrates, respectively (Assine et al. 2014). At the top, fine sandstone can be found with intercalations of siltstone and shale bearing freshwater fossils such as conchostracans and molluscs (Beurlen 1971; Assine 2007).
The Romualdo Formation contains bony and cartilaginous fish, dinosaurs, pterosaurs, molluscs, echinoids, crustaceans, foraminifera, dinoflagellates, gymnosperms, and angiosperms, that suggest sedimentation in a shallow marine to transitional palaeoenvironment (Silva-Santos and Valença 1968; Mabessone and Tinoco 1973; Arai and Coimbra 1990; Berthou et al. 1990; Kellner 2002; Bruno and Hessel 2006; Lima et al. 2012; Pinheiro et al. 2014; Pereira et al. 2017). However, the palaeogeographic reconstruction of the marine transgression in the Araripe Basin is still controversial, and three directions of ingression are suggested: (i) from the Parnaíba Basin (W-NW) (Beurlen 1963, 1966, 1971; Arai et al. 1994; Arai 2014; Prado et al. 2015; Pereira et al. 2017); (ii) from the Potiguar Basin (N-NE) (Lima 1978; Petri 1987); or (iii) from de Recôncavo–Tucano–Jatobá basins (S-SE), crossing the limits of Sergipe-Alagoas Basin (Assine et al. 2014; Custódio et al. 2017; see Fig. 1 and Table 2).
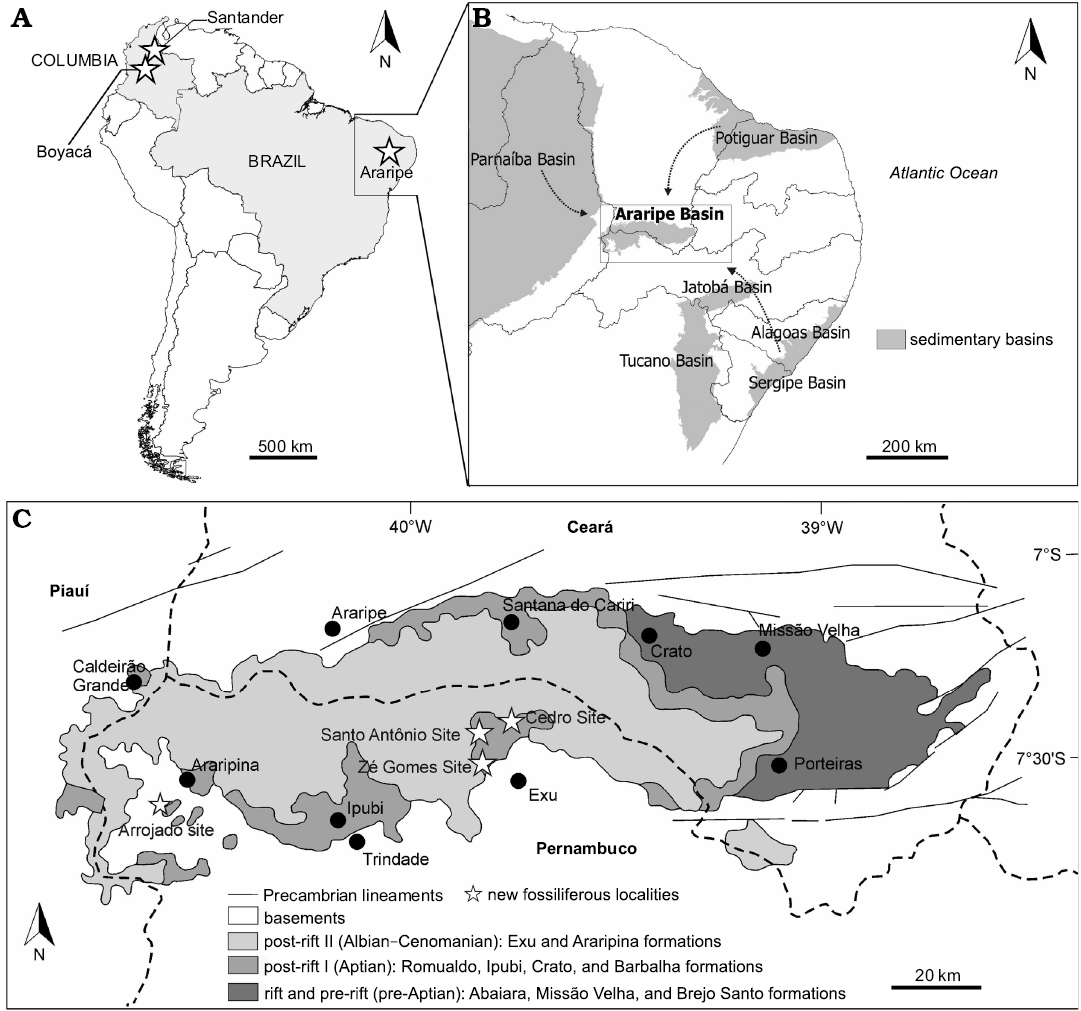
Fig. 1. A. Map showing the known occurrences (stars) of orithopsid and eubrachyuran genera in the Early Cretaceous of South America: upper Aptian–lower Albian San Gil Inferior Formation, Boyacá and upper Aptian Paja Formation, Santander, Colombia and upper Aptian–lower Albian Romualdo Formation, Araripe Basin, Brazil. B. Sedimentary basins of northeast Brazil; arrows show the three possible routes of Cretaceous marine ingression into the Araripe Basin. C. New fossiliferous localities (stars) with brachyuran crabs from upper Aptian–lower Albian Romualdo Formation, Araripe Basin, Pernambuco, Brazil (base map modified from Assine 2007).
Table 2. Key evidence advanced for three proposed marine ingression into the Araripe Basin.
|
Proposed
directions of |
Evidence |
|
Parnaíba Basin |
The
presence of autochthonous echinoids restricted to the
western portion of the Araripe Basin (Beurlen
1963, 1966, 1971;
Prado et al. 2015). |
|
Recôncavo–Tucano–Jatobá basins |
Fluvial palaeocurrents measured
in the Barbalha (Aptian, Santana Group) and Marizal
formations (Aptian, Tucano Basin), indicating south and
southeast paleoflows. As the marine transgressions normally
occur in the opposite direction to the continental
palaeodrainage, the transgression during the Aptian–Albian
in Araripe would have occurred from south-southeast to
north-northwest (Assine 1994; Assine
et al. 2014, 2016; Custódio
et al. 2017). |
|
Potiguar Basin |
Faults with SW directions that
could connect the Araripe and Potiguar basins (Lima
1978). |
Two new brachyuran crabs, Exucarcinus gonzagai Prado and Luque gen. et sp. nov. and Romualdocarcinus salesi Prado and Luque gen. et sp. nov., were collected from marine rocks of the Romualdo Formation cropping out at Cedro (39°35’32”N 7°29’35”E), Santo Antônio (39°32’51”N 7°30’31”E), and Zé Gomes (7°31’16”S 39°34’18”W) in the municipality of Exu in the State of Pernambuco (Fig. 1). All of these outcrops include calcareous sandstone layers between shale layers with calcareous nodules of the Romualdo Formation, differing from each other by the thickness of the layers and their faunistic composition (Fig. 2). From the calcareous sandstone in the municipality of Exu were collected bivalves (e.g., Corbula sp., Brachidontes araripensis Pereira, Cassab, Barreto, and Almeida, 2016b, and Aguileria dissita White, 1887), gastropods (e.g., undetermined cassiopids, Tylostoma ranchariensis Pereira, Cassab, Barreto, and Almeida, 2016b, and Cerithium sergipensis Maury, 1936), echinoids (e.g., Bothryopneustes araripensis Manso and Hessel, 2007) (Pereira et al. 2017), and the two new genera and species of true crabs described herein. In the calcareous sandstone at the Arrojado site (7°41’12”S 40°33’48”W) in the municipality of Araripina, also in the state of Pernambuco, no molluscs or echinoids have been found, but fish predominate. In one of these layers we found a crab claw associated with fossil fish (Fig. 2).
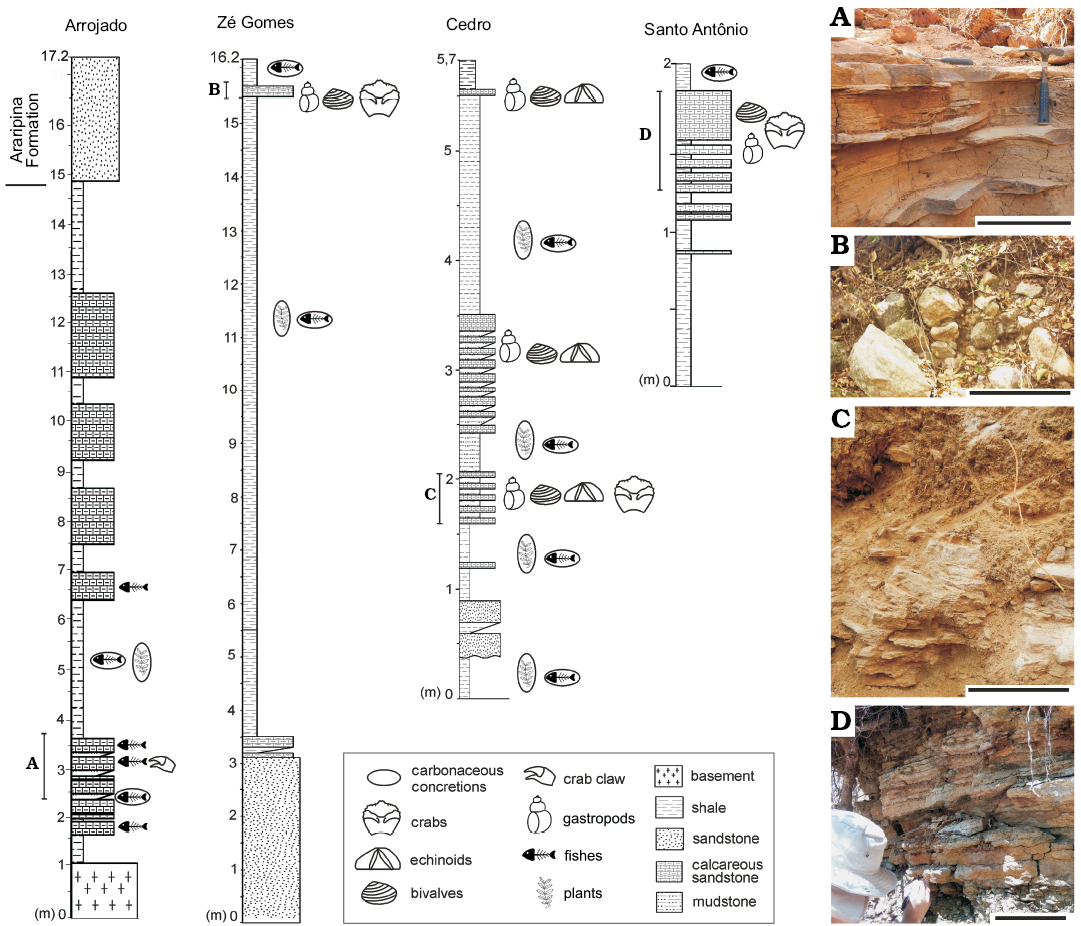
Fig. 2. Stratigraphic sections (A1–D1) and field photographs (A2–D2) of the upper Lower Cretaceous Romualdo Formation, Pernambuco, Brazil, where the new brachyuran crabs were discovered: Arrojado site (A), municipality of Araripina, Zé Gomes (B), Cedro (C), and Santo Antônio (D) sites, municipality of Exu. Scale bars 25 cm.
Material and methods
A total of 30 crab specimens, represented only by dorsal carapaces plus a single isolated cheliped, were collected in situ and ex situ from calcareous sandstone of the upper Aptian–lower Albian Romualdo Formation. The specimens were prepared mechanically and examined under a stereoscopic microscope. For a complete analysis of the material, more complete specimens (DGEO-CTG-UFPE-7747, 8109, 8116, 8122, 8127, 8137, and 8144) were imaged with a Zeiss Sigma 300 VP-FESEM scanning electron microscope at the University of Alberta, Edmonton, Canada. The type series of Exucarcinus gonzagai gen. et sp. nov. and Romualdocarcinus salesi gen. et sp. nov. are deposited in DGEO-CTG-UFPE.
Systematic palaeontology
Order Decapoda Latreille, 1802
Infraorder Brachyura Latreille, 1802
Section Raninoida Ahyong, Lai, Sharkey, Colgan, and Ng, 2007
Family Orithopsidae Schweitzer, Feldmann, Fam, Hessin, Hetrick, Nyborg, and Ross, 2003
Included genera: Aetocarcinus Schweitzer, Feldmann, Rader, and Frantescu, 2016; Bellcarcinus Luque, 2014; Cherpiocarcinus Marangon and De Angeli, 1997; Colombicarcinus Karasawa, Schweitzer, Feldmann, and Luque, 2014; Cristella Collins and Rasmussen, 1992; Exucarcinus Prado and Luque, gen. nov.; Marycarcinus Schweitzer, Feldmann, Fam, Hessin, Hetrick, Nyborg, and Ross, 2003; Orithopsis Carter, 1872; Paradoxilissopsa Schweitzer, Dworschak, and Martin, 2011 (= Lissopsis Frič and Kafka, 1887); Paradoxicarcinus Schweitzer, Feldmann, Fam, Hessin, Hetrick, Nyborg, and Ross, 2003; Planocarcinus Luque, Feldmann, Schweitzer, Jaramillo, and Cameron, 2012; Shazella Collins and Williams, 2004; Silvacarcinus Collins and Smith, 1993; Texicancer Franţescu, Feldmann, and Schweitzer, 2016.
Genus Exucarcinus Prado and Luque nov.
ZooBank LSID: urn:lsid:zoobank.org:act:1E8FAB5B-5466-4D4C-B382-99EDFE2B4E3D
Type species: Exucarcinus gonzagai Prado and Luque sp. nov.; by monotypy, see below.
Etymology: From the municipality of Exu where the material was collected, and the Greek carcinus (= karkinos), crab; gender masculine.
Diagnosis.—As for the type species by monotypy.
Exucarcinus gonzagai Prado and Luque sp. nov.
Fig. 3.
ZooBank LSID: urn:lsid:zoobank.org:act:B206279B-DB4B-4221-AB34-E8AE0D90E1E1
Etymology: In honour of Luiz Gonzaga do Nascimento (1912–1989) “the king of baião”, a singer, composer, and one of the most influential musicians of Brazilian popular music of the twentieth century, born in the Exu municipality, Pernambuco, Brazil.
Type material: Holotype (carapace): DGEO-CTG-UFPE-8114. Paratypes (carapaces): DGEO-CTG-UFPE-7745, 7748, 7749, 7751, 8103, 8105, 8107, 8109, 8115, 8116, 8133, 8106; all from the type locality.
Type locality: Zé Gomes, Santo Antônio, and Cedro sites, Araripe Basin, municipality of Exu, Pernambuco, Brazil.
Type horizon: Romualdo Formation (upper Aptian–lower Albian).
Diagnosis.—Carapace sub-hexagonal, nearly equidimensional, weakly vaulted to flattened, widest at carapace mid-length at level of epibranchial spine, finely granulated, lacking tubercles, bosses, ridges, or reniform swellings. Fronto-orbital margin slightly wider than half the carapace maximum width; rostrum long, extending beyond outer-orbital spine, sulcate axially, bifid distally, with a short lateral spine on each side of the rostrum; post-rostral slits absent. Orbits wide, horizontal, upturned, with two relatively narrow orbital fissures; innermost fissure the shortest; intra-orbital lobe short, truncated, squarish; outer orbital spine well-developed, simple, triangular, shorter than rostrum, directed anteriorly. Anterolateral margin nearly as long as posterolateral margin, with 5–7 spines excluding outer orbital spine, acute, unequal, directed anterolaterally; epibranchial spine short. Posterolateral margin slightly convex to straight, lacking spines, denticles, or tubercles. Posterior margin concave, slightly narrower than fronto-orbital margin. Cervical and branchiocardiac grooves distinct, shallow, sub-parallel to each other; cervical groove reaching anterolateral margin; secondary groove not defined; post-cervical groove absent. Dorsal regions without tubercles, bosses, reniform swellings, or ridges; intestinal region wide, depressed laterally, delimited by swollen metabranchial regions; epibranchial region swollen proximally near contact with branchial groove; meso- and metabranchial regions not differentiated, lacking conspicuous spines or tubercles, but bearing an incipient oblique ridge.
Description.—Carapace sub-hexagonal, about as long as wide in smaller specimens and slightly wider in larger specimens; dorsal carapace weakly vaulted to flattened, widest at carapace mid-length at the level of epibranchial spine. Dorsal carapace smooth, lacking conspicuous tubercles, bosses, ridges, or reniform swellings, covered with fine granulations. Fronto-orbital margin broad, slightly wider than half the carapace maximum width, and wider than posterior margin. Rostrum well-developed, long, extending well beyond outer-orbital spine, nearly one-third of fronto-orbital margin width, wider at base, with straight lateral margins converging antero-medially, bearing a pair of short but well-developed lateral spines positioned at mid-length, and a pair of distal spines forming a weakly downturned bifid rostrum; rostrum sulcate axially, with posterolateral margins at higher level than orbits; post-rostral slits absent. Orbits wide, upturned, directed forward, each orbit about one-third of fronto-orbital margin; supra-orbital margin straight, horizontal, with two relatively narrow orbital fissures parallel to each other, innermost orbital fissure shorter than outermost fissure; inner orbital lobe short, moderately developed, grading from lower margin of rostrum; intra-orbital lobe short, truncated, squarish in outline, separated from inner and outer lobes by the orbital fissures; outer orbital spine well-developed, simple, triangular, shorter than rostrum, with outer margin slightly converging anteriorly; sub-orbital margin visible in dorsal view, with one short, narrow suborbital fissure. Anterolateral margin slightly convex in smaller specimens, less so in larger specimens, nearly as long as posterolateral margin, bearing five to seven acute spines excluding outer orbital spine, of different sizes, and directed anterolaterally; epibranchial spine short, weakly to moderately developed, directed laterally. Posterolateral margin slightly convex in smaller specimens, nearly straight in larger specimen, without spines, denticles, or tubercles. Posterior margin wide, concave, slightly rimmed, much shorter than half of carapace maximum width, and slightly shorter than fronto-orbital margin. Cervical groove distinct, shallow, sinuous, arcuate, with smooth deflections, axially interrupted, distally reaching anterolateral margin; secondary groove not defined; post-cervical groove absent; branchial grooves distinct, well-developed, crescent-shaped; branchiocardic groove present, well-developed, sub-parallel to cervical groove. Epigastric region narrow, barely defined laterally by faint sub-parallel grooves; protogastric and mesogastric regions faintly tumid; hepatic region wide, depressed, without transverse ridge; epigastric, protogastric, mesogastric, and hepatic regions poorly delimited, without tubercles or bosses; metagastric and urogastric lacking tubercles or longitudinal ridge, faintly separated by a shallow depression, both regions defined laterally by crescent-shaped branchial grooves; urogastric region narrow; cardiac region well-developed, tumid, broader than meta- and urogastric regions, wider anteriorly lacking tubercles, without transverse or longitudinal ridges; intestinal region wide, depressed anteriorly by contact with posterior cardiac region, depressed laterally and delimited by swollen metabranchial regions; epibranchial region moderately defined, limited by cervical and branchiocardic grooves, swollen proximally near contact with branchial groove; meso- and metabranchial regions not differentiated, lacking conspicuous spines or tubercles, but bearing an incipient oblique ridge extending towards the posterolateral corner.
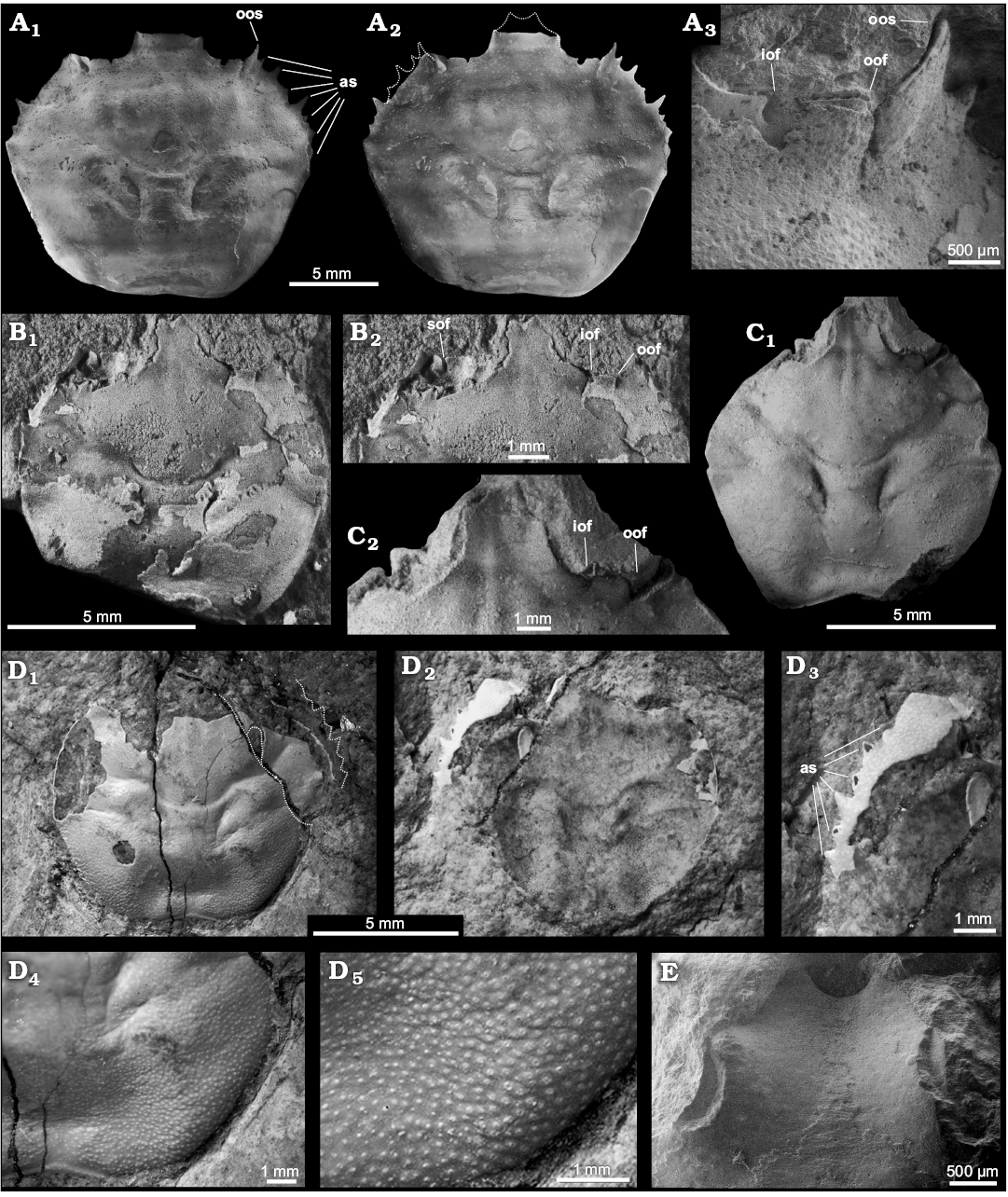
Fig. 3. Orithopsid brachyuran Exucarcinus gonzagai Prado and Luque gen. et sp. nov. from the upper Aptian–lower Albian Romualdo Formation of the Santana Group, Araripe Basin, Pernambuco, Brazil. A. Holotype DGEO-CTG-UFPE-8114; A1, SEM image of dorsal carapace; A2, negative colour image of A1; A3, close-up of right orbit, orbital fissures, and outer orbital spine. B. Paratype DGEO-CTG-UFPE-8133; B1, dorsal carapace; B2, sub-orbital margin visible in dorsal view with one short and narrow suborbital fissure. C. Paratype DGEO-CTG-UFPE-8109; C1, dorsal carapace; C2, orbit and bifid rostrum extending well beyond the outer-orbital spine. D. Paratype DGEO-CTG-UFPE-8115; D1, dorsal carapace; D2, counterpart; D3, close-up of ventral view of right anterolateral margin bearing multiple anterolateral spines; D4, close-up of posterolateral and posterior margins lacking spines; D5, close-up of dorsal carapace cuticle with fine granulations. E. Paratype DGEO-CTG-UFPE-8116, SEM image of bifid rostrum. Abbreviations: as, anterolateral spines; ios, inner orbital spine; iof, innermost orbital fissure; its, intra-orbital spine; oof, outermost orbital fissure; oos, outer orbital spine; sof, sub-orbital fissure. All specimens photographed dry and uncoated, except B, C, which are coated with ammonium chloride.
Dimensions.—For a list of measurements see Table 3.
Table 3. Measurements (in mm) of the fossil crabs Exucarcinus gonzagai gen. et sp. nov. and Romualdocarcinus salesi gen. et sp. nov. Abbreviations: AL, anterolateral length; CL, carapace length; CW, carapace width; FW, fronto-orbital width; PL, posterolateral length; PW, posterior width; RW, rostrum width.
|
Species |
DGEO-CTG-UFPE- |
Occurrence |
CL |
CW |
RW |
FW |
AL |
PL |
PW |
|
Exucarcinus gonzagai |
7745 |
Zé Gomes |
7 |
8 |
– |
5 |
3 |
4 |
4 |
|
7749 |
7.5 |
9 |
– |
– |
3.5 |
4 |
3 |
||
|
8114 |
13 |
15 |
3 |
9 |
6 |
7 |
8 |
||
|
8116 |
10 |
11 |
2.5 |
7.5 |
4 |
5 |
6 |
||
|
7748 |
Santo
|
6 |
7 |
1.5 |
4.5 |
2.5 |
3.5 |
4 |
|
|
7751 |
8.5 |
10 |
– |
6 |
4 |
4.5 |
5 |
||
|
8103 |
Cedro |
6 |
6 |
– |
4.5 |
2.5 |
3.5 |
4 |
|
|
8104 |
6.5 |
8 |
– |
6 |
3 |
3.5 |
4 |
||
|
8105 |
6.5 |
7 |
2 |
6 |
3 |
3.5 |
5 |
||
|
8106 |
6 |
7 |
– |
5 |
2.5 |
3.5 |
4 |
||
|
8107 |
4.5 |
5.5 |
– |
3.5 |
2.5 |
3 |
2.5 |
||
|
8109 |
9 |
8 |
2 |
6 |
3.5 |
5 |
5 |
||
|
8115 |
9 |
10 |
– |
6 |
4 |
5 |
5 |
||
|
8133 |
7 |
8 |
2 |
6 |
3 |
4 |
5 |
||
|
Romualdocarcinus salesi |
8108 |
Cedro |
4.5 |
6.5 |
2 |
5 |
2 |
2.5 |
4 |
|
8110 |
4.5 |
6 |
1 |
3 |
2 |
2.5 |
2 |
||
|
8111 |
– |
– |
2 |
– |
– |
– |
– |
||
|
8119 |
3.5 |
4 |
1 |
3 |
2 |
1.5 |
2 |
||
|
8122 |
5.5 |
6.5 |
2 |
6 |
1.5 |
2.5 |
3 |
||
|
8123 |
4.5 |
6 |
1.5 |
5 |
2 |
2.5 |
3.5 |
||
|
8124 |
6.5 |
7 |
2 |
6 |
3 |
3.5 |
4 |
||
|
8125 |
– |
8 |
2 |
6 |
4 |
5 |
– |
||
|
8126 |
5 |
7 |
6 |
2 |
2 |
3 |
3 |
||
|
8127 |
10 |
9 |
3 |
9 |
4 |
3 |
4.5 |
||
|
8128 |
5 |
7 |
6 |
2 |
2 |
3 |
– |
||
|
8129 |
– |
– |
2 |
– |
– |
– |
– |
||
|
8131 |
5 |
6 |
1.5 |
4.5 |
2 |
3 |
3 |
||
|
8132 |
6 |
7 |
2 |
6 |
2.5 |
3.5 |
4 |
||
|
8135 |
– |
9 |
2.5 |
7.5 |
– |
– |
– |
||
|
8137 |
7 |
8 |
2 |
6 |
4 |
3 |
4 |
Remarks.—Exucarcinus Prado and Luque gen. nov. strongly differs from Camarocarcinidae Feldmann, Li, and Schweitzer, 2007, Necrocarcinidae Förster, 1968, and several Cenomanocarcinidae Guinot, Vega, and Van Bakel, 2008, in its slightly vaulted to flattened dorsal carapace, the lack of posterolateral spines (Fig. 3A1, A2, B1, C1, D1, D4), and the absence of transverse or branchial ridges, contrasting with the strongly vaulted and ornamented carapaces seen in these families. Also, it lacks the deep cervical and branchiocardic grooves and short rostrum seen in Necrocarcinidae (Fig. 3A1, A2, B, C, E). Among stem raninoidans, the presence of a well-defined longitudinal mesial ridge, absent in Exucarcinus, is mostly seen among Palaeocorystidae Lőrenthey in Lőrenthey and Beurlen, 1929, Cenomanocarcinidae, Necrocarcinidae in part (i.e., Elektrocarcinus Schweitzer, Karasawa, Luque, and Feldmann, 2016, and Hadrocarcinus Schweitzer, Feldmann, and Lamanna, 2012), and most of the non-American Orithopsidae genera (see below). Exucarcinus Prado and Luque gen. nov. differs from Paranecrocarcinidae Fraaije, Van Bakel, Jagt, and Artal, 2008, in its flattened mesogastric region, the spinose anterolateral margin (5–7 spines) (Fig. 3A1, A2, D1–D3), and the lack of post-rostral slits, so distinctive of paranecrocarcinids. The new genus also differs from Paranecrocarcinidae and several Orithopsidae in the lack of protogastric swellings, but shares with them the wide, somewhat spatulate bifid rostrum extending well beyond the outer-orbital spine, the overall sub-hexagonal carapace with wide fronto-orbital margins, and the lack of posterolateral spines.
Exucarcinus Prado and Luque gen. nov. differs from other orithopsids (e.g., Bellcarcinus, Cherpiocarcinus, Marycarcinus, Orithopsis, Paradoxilissopsa, Silvacarcinus, Texicancer) in the lack of dorsal tubercles, reniform swellings, keels, or ridges (Fig. 3A1, A2, B1, C1, D1, D2). Other genera like Cristella and Shazella have pronounced epibranchial spines or pustulose dorsal carapaces with conspicuous dorsal grooves, none of which are apparent in the new genus. In fact, among orithopsids, Exucarcinus Prado and Luque gen. nov. seems to be more similar to some genera endemic to the Americas, like Aetocarcinus, Colombicarcinus, and Planocarcinus based on the lack of posterolateral spines, the smooth dorsal carapace lacking conspicuous dorsal ridges, keels, or tubercles, and the broad spatulate and bifid rostrum extending beyond the outer-orbital spine. However, these genera lack the numerous well-developed and acute anterolateral spines diagnostic of Exucarcinus Prado and Luque gen. nov. (Fig. 3A1, A2, D1–D3).
Stratigraphic and geographic range.—Type locality and horizon only.
Section Eubrachyura Saint Laurent, 1980
Clade Heterotremata Guinot, 1977
Family ?Eogeryonidae Ossó, 2016
Type genus: Eogeryon Ossó, 2016.
Included genera: Type genus and tentatively Romualdocarcinus Prado and Luque gen. nov.
Genus Romualdocarcinus Prado and Luque nov.
ZooBank LSID: urn:lsid:zoobank.org:act:DA947443-E1A9-485B-B1C0-F1C058DC1A6D
Type species: Romualdocarcinus salesi Prado and Luque sp. nov.; by monotypy, see below.
Etymology: From the Romualdo Formation, Santana Group; a Konservat-Lagerstätte in northeast Brazil where the material was collected, and the Greek carcinus (= karkinos), crab; gender masculine.
Diagnosis.—As for the type species by monotypy.
Romualdocarcinus salesi Prado and Luque sp. nov.
Fig. 4.
ZooBank LSID: urn:lsid:zoobank.org:act:86A8591D-A6D8-436B-AD02-8A29B36E3CF6
Etymology: In honour of the late Professor Alexandre Magno Feitosa Sales (1969–2016), an important figure in the paleontological scenario of Brazilian Northeast, mainly the Araripe Basin.
Type material: Holotype (carapace): DGEO-CTG-UFPE-8122. Paratypes (carapaces): DGEO-CTG-UFPE-8110, 8111, 8119, 8123, 8124, 8125, 8126, 8127, 8128, 8129, 8131; all from the type locality.
Type locality: Cedro site, Araripe Basin, municipality of Exu, Pernambuco, Brazil.
Type horizon: Romualdo Formation (upper Aptian–lower Albian).
Diagnosis.—Carapace sub-hexagonal to sub-square, nearly equidimensional, weakly vaulted transversely and longitudinally, widest anterior to carapace mid-length and to epibranchial spine, finely granulated, lacking ridges, bosses, tubercles, or reniform swellings. Fronto-orbital margin nearly as wide as carapace maximum width; rostrum well-developed, elongated, bifid, somewhat spatulate, axially sulcate, extending well beyond outer orbital spine, bearing a pair of short lateral spines at anterior third; post-rostral slits absent. Orbits wide, directed forward, faintly upturned; supra-orbital margin straight, sub-horizontal to slightly diverging postero-laterally, with two very short but wide orbital fissures somewhat oblique to each other; inner orbital lobe short; intra-orbital lobe short, roundish in outline; outer orbital spine well-developed, relatively long, simple, triangular, shorter than rostrum, with outer margin directed anteriorly or slightly diverging antero-laterally. Anterolateral margin shorter than posterolateral margin, with three short, broad, well-developed triangular spines excluding the outer orbital spine, first two spines the largest; epibranchial spine weakly developed, directed laterally. Posterolateral margin entire, slightly convex to nearly straight. Posterior margin slightly concave to straight, rimmed, narrower than front-orbital margin. Cervical groove moderately defined, shallow, sinuous, distally reaching anterolateral margins; secondary groove faint; postcervical groove absent. Branchiocardic groove shallower than cervical groove. Epigastric region narrow, protogastric and mesogastric regions slightly tumid; hepatic region wide, slightly tumid; intestinal region wide; epibranchial region with two swellings near contact with branchial groove; meso- and metabranchial regions not differentiated.
Description.—Carapace sub-hexagonal to sub-square, as long as wide; dorsal carapace weakly vaulted to flattened, widest anterior to carapace mid-length at level of second anterolateral spine; dorsal carapace smooth, finely granulated, with well-defined regions lacking ridges, tubercles, or reniform swellings. Fronto-orbital margin broad, nearly as wide as carapace maximum width and wider than posterior margin. Rostrum well-developed, broad, long, somewhat spatulate, extending well beyond outer orbital spine, nearly one-third of fronto-orbital margin width, wider at the base, with straight lateral margins converging antero-medially, bearing a pair of short lateral spines at anterior third, and a pair of distal spines forming a weakly downturned bifid rostrum; rostrum sulcate axially, with posterolateral margins at level of supraorbital margin; post-rostral slits absent. Orbits wide, faintly upturned, directed forward, each orbit slightly over one-third of fronto-orbital margin; supra-orbital margin straight, sub-horizontal to slightly diverging postero-laterally, with two very short but wide orbital fissures somewhat oblique to each other; innermost orbital fissure slightly wider than outermost fissure; inner orbital lobe short, moderately developed, grading from lower margin of rostrum; intra-orbital lobe short, roundish in outline, separated from inner and outer lobes by orbital fissures; outer orbital spine well-developed, simple, triangular, shorter than rostrum, with outer margin directed anteriorly or slightly diverging antero-laterally. Anterolateral margin slightly convex to straight, slightly shorter than posterolateral margin, bearing three triangular, short, well-developed spines excluding outer orbital spine; first two spines the largest, directed antero-laterally, second antero-lateral spine wider than adjacent spines; third antero-lateral spine representing the epibranchial spine, weakly developed, directed laterally. Posterolateral margin slightly convex to nearly straight, without spines, denticles, or tubercles. Posterior margin wide, slightly concave, rimmed, shorter than fronto-orbital margin and slightly wider than half of carapace maximum width. Cervical groove distinct but moderately defined, shallow, sinuous, arcuate, with smooth deflections, axially interrupted, distally reaching anterolateral margins; secondary groove present but faint; post-cervical groove absent; branchial grooves distinct, moderately developed, crescent-shaped; branchiocardic groove present, weakly developed, sub-parallel to cervical groove. Epigastric region narrow, well-defined laterally by sub-parallel grooves; protogastric and mesogastric regions slightly inflated; hepatic region wide, without transverse ridge but slightly tumid; epigastric, protogastric, mesogastric, and hepatic regions without tubercles or bosses; metagastric and urogastric regions lacking tubercles or longitudinal ridges, faintly separated by a shallow depression, both regions defined laterally by crescent-shaped branchial grooves; urogastric region narrow; cardiac region well-developed, inflated, broader than meta- and urogastric regions, wider anteriorly, lacking tubercles, without transverse or longitudinal ridges; intestinal region wide, depressed anteriorly by contact with posterior cardiac region; epibranchial region well-defined, limited by cervical and branchiocardic grooves, with two swellings near contact with branchial groove; meso- and metabranchial regions not differentiated, lacking spines, tubercles, or conspicuous ridges, but with posterior metagastric region inflated.
Dimensions.—For a list of measurements see Table 3.
Remarks.—The systematic affinities of Romualdocarcinus Prado and Luque gen. nov. are puzzling, in part due to its incompleteness (only known from dorsal carapaces), and its superficial resemblance to brachyuran crabs from different groups. For instance, Romualdocarcinus shares a sub-hexagonal carapace outline with Paranecrocarcinidae, Cenomanocarcinidae, and Orithopsidae, although Romualdocarcinus is more sub-squarish than most orithopsids, and lacks the posterolateral spines and weakly projected rostrum of Cenomanocarcinidae, or the postrostral slits seen in most Paranecrocarcinidae. Romualdocarcinus also shares with Orithopsidae, Necrocarcinidae, and Paranecrocarcinidae a fronto-orbital margin that is wider than half the carapace maximum width, but differs from Necrocarcinidae, Paranecrocarcinidae, or any other necrocarcinoid-like raninoidan, in its fronto-orbital margin being almost as wide as the carapace (Fig. 4A1, B, C, D). The subspatulate bifid rostrum that extends beyond the outer-orbital spine seen in several Paranecrocarcinidae and Orithopsidae, and the smooth dorsal carapace lacking ridges or rows of tubercles distinctive of orithopsids like Planocarcinus, Colombicarcinus, and Aetocarcinus, would advocate for affinities with the group.
Romualdocarcinus Prado and Luque gen. nov. and the orithopsid Exucarcinus Prado and Luque gen. nov., are known from the same strata and localities in the Araripe basin, yet both taxa differ in several features. In Romualdocarcinus the carapace outline is squarish, its dorsal regions are more tumid, and the posterior margin nearly straight, while in Exucarcinus the carapace is more hexagonal, has less marked dorsal regions, and the posterior margin is concave. Also, Romualdocarcinus has a shorter anterolateral margin, only three anterolateral spines (Fig. 4A1, B2, C2, D2), and a maximum width anterior to its mid-length, whereas Exucarcinus has anterolateral and posterolateral margins about the same length, has six to seven anterolateral spines, and is wider at its mid-length. Furthermore, in Romualdocarcinus the supra-orbital margins are weakly upturned and slightly divergent postero-laterally, and bear two short and wide orbital fissures faintly oblique to each other (Fig. 4A2, A3, C2, D2), which differ from the sub-horizontal margins with two narrower and sub-parallel orbital fissures seen in Exucarcinus.
Despite the superficial similarities between Romualdocarcinus and several necrocarcinoid-like raninoidans, the new genus seems to fit better within Eubrachyura, in particular Eogeryonidae, known from the Cenomanian of Spain (Ossó 2016). Both taxa share a sub-hexagonal to sub-square carapace outline, they have anterolateral margins with three spines (excluding the outer orbital spine), wide orbits with two open fissures, and a broad bifid rostrum with secondary lateral spines (Ossó 2016). However, in Eogeryonidae the rostrum is short and four tipped, while in Romualdocarcinus it is elongated and bifid (Fig. 4A, C). Other early eubrachyurans like Marocarcinidae Guinot, De Angeli, and Garassino, 2008, from the Cenomanian of Morocco, also share with Romualdocarcinus the bifid rostrum with secondary lateral spines, and the presence of three spines in the anterolateral margin, but it has smaller orbits than those of Romualdocarcinus, it lacks orbital fissures, and has cervical and branchiocardial grooves marked only towards the lateral margin. Another family from the Upper Cretaceous of the Americas, Longusorbiidae Karasawa, Schweitzer, and Feldmann, 2008, shares with Romualdocarcinus a fronto-orbital margin nearly as wide as the carapace maximum width, the broad orbits, and a rostrum elongated and axially sulcate. However, Longusorbiidae has a considerably narrower downturned rostrum and well-defined dorsal carapace regions, both absent in Romualdocarcinus.
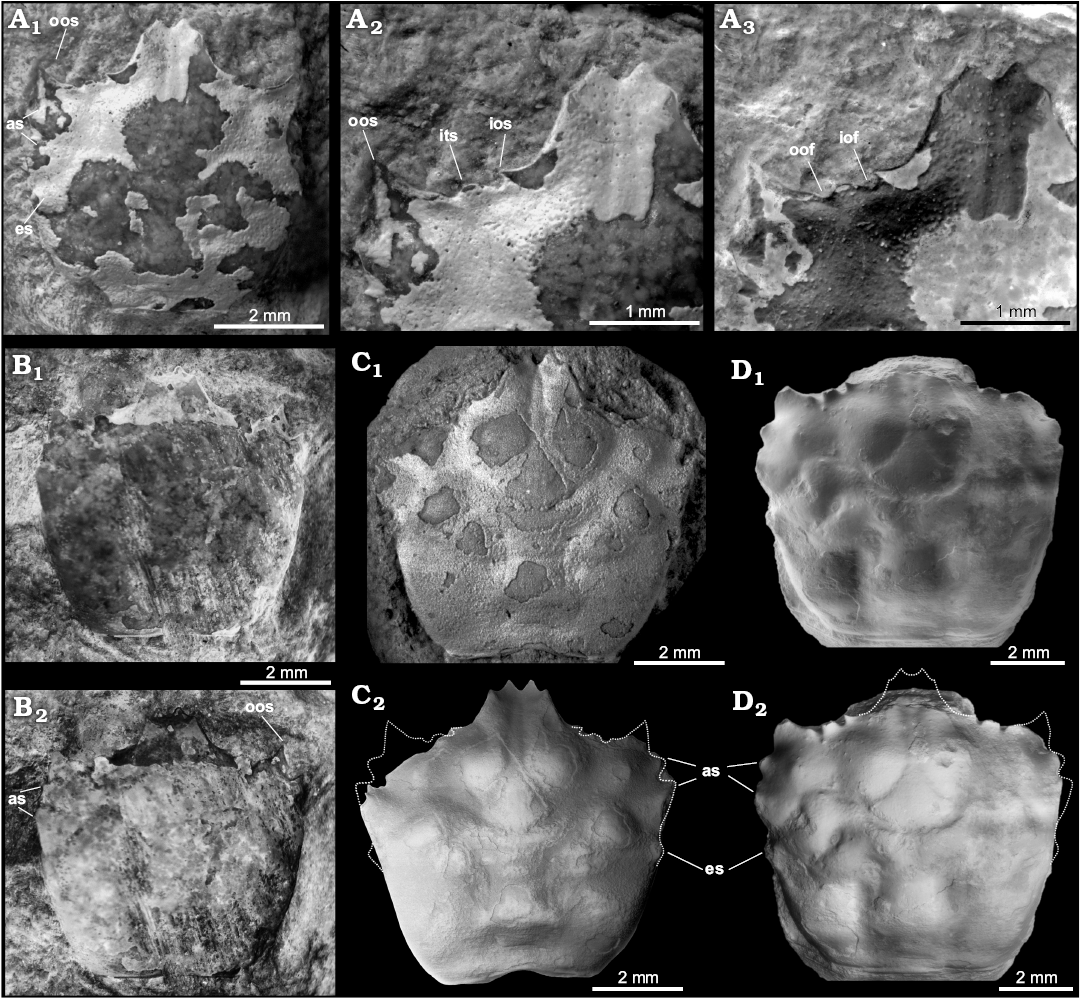
Fig. 4. ?Eogeryonid brachyuran Romualdocarcinus salesi Prado and Luque gen. et sp. nov. from the upper Aptian–lower Albian Romualdo Formation of the Santana Group, Araripe Basin, Pernambuco, Brazil. A. Holotype DGEO-CTG-UFPE-8122; A1, dorsal view showing the two anterolateral spines, the epibranchial spine, and the outer orbital spine; A2, close-up of rostrum and left orbit showing the inner-, intra-, and outer- orbital spines; A3, inverted colour image of A2, showing the short and wide inner and outer orbital fissures. B. Paratype DGEO-CTG-UFPE-8119; B1, dorsal carapace; B2, inverted colour image of B1 showing the outer orbital spine and the two anterolateral spines. C. Paratype DGEO-CTG-UFPE-8137; C1, dorsal carapace; C2, inverted colour image of C1, showing the anterolateral spines and the short epibranchial spine. D. Paratype DGEO-CTG-UFPE- 8127; D1, SEM image of dorsal carapace; D2, inverted colour image of D1, showing the anterolateral spines and the short epibranchial spine. Abbreviations: as, anterolateral spines; ios, inner orbital spine; iof, innermost orbital fissure; its, intra-orbital spine; oof, outermost orbital fissure; oos, outer orbital spine. All specimens photographed dry and uncoated, except for C1, which was coated with ammonium chloride.
Although most eubrachyuran families have their earliest fossil representatives in Upper Cretaceous or Cenozoic rocks, only a handful of eubrachyuran-like families are currently known from Lower Cretaceous units, i.e., Telamonocarcinidae Larghi, 2014, from the lower Albian of Colombia; Tepexicarcinidae Luque, 2015b, from the lower Albian of Mexico; and Componocancridae Feldmann, Schweitzer, and Green, 2008, from the lower Albian of USA (see summary in Luque 2015b). A fourth family of eubrachyurans, Priscinachidae Breton, 2009, is known from the upper Albian and Cenomanian of Spain and France (Breton 2009; Klompmaker 2013). Although only two of these families, i.e., Telamonocarcinidae and Priscinachidae, can be placed within crown superfamilies such as Dorippoidea and Majoidea, respectively, all four taxa belong to basal eubrachyuran branches, whereas Romualdocarcinus is more reminiscent of modern-looking eubrachyurans like Eogeryonidae. If Romualdocarcinus is indeed an eogeryonid, it will extend the temporal distribution of the family into the Early Cretaceous, and represents one of the oldest “higher” true crab yet known.
Stratigraphic and geographic range.—Type locality and horizon only.
Concluding remarks
Early Cretaceous crabs of the family Orithopsidae are known from the Aptian–Albian of North America, South America, and Europe (Van Bakel et al. 2012; Schweitzer et al. 2016b; Luque et al. 2017, and references therein). Exucarcinus Prado and Luque gen. nov., from the Aptian–Albian of Brazil, is very similar to other endemic South American taxa of nearly the same age such as Planocarcinus, Bellcarcinus, and Colombicarcinus.
The only brachyuran crab previously known from the Romualdo Formation and the Santana Group as a whole is Araripecarcinus ferreirai, represented by a single ventral carapace with appendages associated (Maisey 1991; Luque 2015a). In contrast, Exucarcinus gonzagai Prado and Luque gen. et sp. nov. and Romualdocarcinus salesi Prado and Luque gen. et sp. nov. are so far known exclusively from dozens of dorsal carapaces. It is possible that E. gonzagai Prado and Luque gen. et sp. nov. might correspond to the dorsal carapace of A. ferrerai, but without conclusive evidence and dorso-ventral material associated, this is just a possibility and both taxa so far cannot be compared directly or assumed as conspecific. In addition, A. ferreirai comes from a different stratigraphic interval, facies, and taphocoenosis than E. gonzagai, further suggesting a possible independent systematic placement. An isolated right brachyuran claw found in the site Arrojado (Fig. 5) was collected from the concretionary fish level from which Araripecarcinus comes (Fig. 2), and is similar to the chelipeds of A. ferreirai, suggesting that it could belong to this species. To date, all fossil and extant raninoidans, i.e., Necrocarcinoidea, Palaeocorystoidea, and Raninoidea, are exclusively known from marine subtidal settings, and there are no fossil or extant raninoidans known from brackish or freshwater habitats (Luque 2015a). Therefore, the occurrences of Exucarcinus Prado and Luque gen. nov. associated with echinoids in the municipality of Exu, Pernambuco, suggest shallow marine sedimentation in the western portion of the Araripe Basin rather than brackish depositional settings.
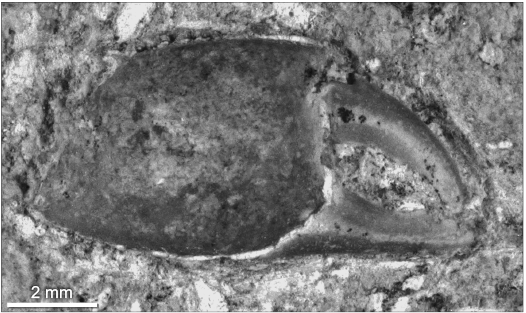
Fig. 5. Brachyura indet. (right cheliped) from the upper Aptian–lower Albian Romualdo Formation of the Santana Group, Araripe Basin, Pernambuco. DGEO-CTG-UFPE-7743, possibly akin to Araripecarcinus ferreirai Martins-Neto, 1987.
As previously indicated, the eubrachyuran-like Romualdocarcinus salesi Prado and Luque gen. et sp. nov. might be an eubrachyuran related to Eogeryon, a genus whose sternal anatomy clearly indicates an eubrachyuran affinity (Ossó 2016: fig. 5). As such, Romualdocarcinus would be expected to have a sternal construction closer to eubrachyurans and unlike that of Araripecarcinus (Luque 2015a).
Molluscs and echinoids from the same stratigraphic levels of the Romualdo Formation where the new crab faunule occurs have been interpreted as transported clasts in deposits generated by storms at different water depths (Sales 2005; Prado et al. 2015, 2016). The occurrence of mostly dorsal carapaces of crabs lacking any preferential position within the matrix of coquinas, limited at the base by abrupt contacts which exhibit fragmented bioclasts without signs of abrasion or rounding, suggest deposition under storm events where the energy was not sufficient to transport the remains over long distances. Some specimens show local fragmentation due to compaction, as the carapaces are complete but broken, and some of the fragments are only slightly displaced from the rest of the carapace. The crab carapaces here reported most likely would have been transported over short distances or remobilized within their own environment (parautochthonus sensu Kidwell et al. 1986).
To date, the genera Planocarcinus, Bellcarcinus, Colombicarcinus, Exucarcinus Prado and Luque gen. nov., and Romualdocarcinus Prado and Luque gen. nov. are restricted to northern South America, and are quite different from closer relatives in North America and Europe (Van Bakel et al. 2012; Schweitzer et al. 2016a, b; Luque et al. 2017). Our new findings indicate that the tropical Americas might have been inhabited by several early brachyuran lineages, with several endemic genera and species during the Cretaceous.
The Lower Cretaceous rocks of northeastern Brazil, including those in the Araripe Basin, record the transition from continental to marine facies (rift and post-rift sequences) linked to the separation of South America and Africa, and the birth of the Central Atlantic that connected the water masses of both hemispheres. According to Arai (2014), the Tethyan influence on these basins during the upper Aptian–lower Albian is suggested by the presence of several Tethyan faunas, such as the isopod Codoisopus brejensis in the Parnaíba Basin (Codó Formation) (Lindoso et al. 2013), the pithonellid calcispheres (calcareous dinoflagellates) in the Potiguar and Sergipe-Alagoas basins (Dias-Brito 1985a, b, 2002), bivalve Neithea (Neithea) coquandi (Péron, 1877) (Monteiro et al. 2010), planktonic foraminifera (e.g., Globigerinelloides aptiensis, G. barri, G. blowi, G. ferreolensis, G. maridalensis, Hedbergella labocaensis, H. maslakovae, H.semielongata, H. similis), and ammonoids (Epicheloniceras cf. martinioides, Vectisites simplex, and V. cf. cruzi) (Bengtson and Koutsoukos 1992; Bengtson et al. 2007) in the Sergipe-Alagoas Basin (Riachuelo Formation), and the molluscs and fishes in the Araripe Basin (see Table 2). Therefore, the marine ingression would have reached the Araripe either from the north through the Parnaíba Basin (Arai 2014) or from the northeast through the Potiguar Basin (Lima 1978) (Table 2), but not from the south (Recôncavo–Tucano–Jatobá basins).
Although the fragmentary and sparse record of fossil raninoidans and eubrachyurans from the Early Cretaceous does not help resolve this issue, the presence of taxa similar to those from the Aptian–Albian of Colombia, and the Albian–Cenomanian of USA, England, and Spain, confirm the broad distribution of these groups in the Tethys Ocean during the Early Cretaceous, and reinforces the view of a Tethyan influence in Araripe at the time.
Acknowledgements
We are grateful to the Exu city council, the friends Genilma Cordeiro, Cícero Marcelino, Juaran Rodrigues (all Pernambuco, Brazil) and the members of the palaeontology laboratory of the Universidade Federal de Pernambuco (UFPE) for field assistance. We also thank Ismar Carvalho (Universidade Federal do Rio de Janeiro, Brazil) for his suggestions in textual organization, Artur Andrade (head of DNPM’s regional office, Crato, Brazil) for his assistance in transporting fossil material abroad for study, and Jefferson Lima (Ph.D. student, UFPE, Brazil) for improvement on Figs. 1 and 2. Special thanks to Barry van Bakel (Oertijdmuseum De Groene Poort, Boxtel, Netherlands) and Àlex Ossó (independent researcher, Tarragona, Spain) for discussions on the systematic affinities of the material studied, and to the reviewers Rodney Feldmann and Carrie Schweitzer (both Kent State University, Ohio, USA) for their thoughtful and stimulating comments. This work was possible thanks to the Brazilian Federal Agency for Support and Evaluation of Graduate Education-CAPES (Programa de Doutorado Sanduíche no Exterior PDSE: 88881.134830/2016-01) to LACP, the Izaak Walton Killam Memorial Scholarship (Canada) to JL, and the Natural Science and Engineering Research Council of Canada (NSERC) Discovery Grants RGPIN 04863 to ARP.
References
Ahyong, S.T., Lai, J.C.Y., Sharkey, D., Colgan, D.J., and Ng, P.K.L. 2007. Phylogenetics of the brachyuran crabs (Crustacea: Decapoda): The status of Podotremata based on small subunit nuclear ribosomal RNA. Molecular Phylogenetics and Evolution 45, 576–586. Crossref
Alvarado-Ortega, J. and Brito, P.M. 2011. A new species of Araripichthys (Teleostei, Elopocephala) from the Tlayúa Formation (Cretaceous, Albian), México. Journal of Vertebrate Paleontology 31: 1376–1381. Crossref
Arai, M. 2014. Aptian/Albian (Early Cretaceous) paleogeography of the South Atlantic: a paleontological perspective. Brazilian Journal of Geology 44: 339–350. Crossref
Arai, M. and Coimbra, J.C. 1990. Análise paleoecológica do registro das primeiras ingressões marinhas na Formação Santana (Cretáceo Inferior da Chapada do Araripe). Atas do 1° Simpósio sobre a Bacia do Araripe e Bacias Interiores do Nordeste, 225–233. SBP, Crato-CE.
Arai, M., Lana, C.C., and Pedrão, E. 1994. Ecozona Subtilisphaera spp.: Registro eocretáceo de um importante episódio ecológico do Oceano Atlântico primitivo. Acta Geologica Leopoldensia 39: 521–538.
Assine, L.M. 1992. Análise estratigráfica da Bacia do Araripe, Nordeste do Brasil. Revista Brasileira de Geociências 22: 289–300. Crossref
Assine, L.M. 1994. Paleocorrentes e Paleogeografia na Bacia do Araripe, Nordeste do Brasil. Revista Brasileira de Geociências 24: 223–232. Crossref
Assine, L.M. 2007. Bacia do Araripe. Boletim de Geociências da Petrobrás 15: 371–389.
Assine, L.M, Perinotto, J.A.J., Custódio, M.A., Neumann, V.H., Varejão, F.G., and Mescolotti, P.C. 2014. Sequências deposicionais do Andar Alagoas da Bacia do Araripe, Nordeste do Brasil. Boletim de Geociências da Petrobrás 22: 3–28.
Assine, L.M., Quaglio, F., Warren, L.V., and Simões, M.G. 2016. Comments on paper by M. Arai “Aptian/Albian (Early Cretaceous) paleogeography of the South Atlantic: a paleontological perspective”. Brazilian Journal of Geology 46: 3–7. Crossref
Bengtson, P. and Koutsoukos, E.A.M. 1992. Ammonite and foraminiferal dating of the first marine connection between the Central and South Atlantic. In: R. Curnelle (ed.), Géologie Africaine. Elf Aquitaine Mémoire 13: 403.
Bengtson, P., Koutsoukos, E.A.M., Kakabadze, M.V., and Zucon, M.H. 2007. Ammonite and foraminiferal biogeography and the opening of the Equatorial Atlantic gateway. In: 1st International Palaeobiogeography Symposium. Paris, France, Abstracts, 12. Universite Pierre et Marie Curie and Museum national d’Historire naturelle, Paris.
Bermúdez, H.D., Gómez-Cruz, A.D.J., Hyžný, M., Moreno-Bedmar, J.A., Barragán, R., Sánchez, M.M., and Veja, F.J. 2013. Decapod crustaceans from the Cretaceous (Aptian–Albian) San Gil Group in the Villa de Leyva section, central Colombia. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 267: 255–272.
Berthou, P.Y., Viana, M.V., and Campos, D.A. 1990. Coupe da La Formation Santana dans Le secteur de “Pedra Branca” (Santana do Cariri) (Bassin D’ Araripe, NE du Brésil). Contribuition à l’ étude de La sedimentologie et des paleoenvironnements. In: L.M. Barros, P.C. Nuvens, and J.B.M Filgueira (eds.), Atas do 1° Simpósio sobre a Bacia do Araripe e Bacias Interiores do Nordeste, 173–191. SBP, Crato-CE.
Beurlen, K. 1963. Geologia e estratigrafia da Chapada do Araripe. Anais do 17° Congresso Nacional de Geologia 1: 1– 47.
Beurlen, K. 1966. Novos equinóides no Cretáceo do Nordeste do Brasil. Anais Academia Brasileira de Ciências 38: 455–464.
Beurlen, K. 1971. As condições ecológicas e faciológicas da Formação Santana na Chapada do Araripe (Nordeste do Brasil). Anais da Academia Brasileira de Ciências 43: 411–415.
Breton, G. 2009. Description of Priscinachus elongatus n. gen., n. sp., and Priscinachidae n. fam. for the earliest spider crab (Crustacea, Decapoda, Majoidea), from the French Cretaceous (Cenomanian). Geodiversitas 31: 509–523. Crossref
Bruno, A.P. and Hessel, M.P. 2006. Registros paleontológicos do Cretáceo marinho na Bacia do Araripe. Estudos Geológicos 16: 30–49.
Carter, J. 1872. On Orithopsis bonneyi, a new fossil crustacean. Geological Magazine 9: 529–532. Crossref
Collins, J.S.H. and Smith, R. 1993. Ypresian (Lower Eocene) crabs (Decapoda, Crustacea) from Belgium. Bulletin de l’Institut Royal des Sciences Naturelles de Belgique (Sciences de la Terre) 63: 261–270.
Collins, J.S.H. and Williams, R.J. 2004. A new genus and species of necrocarcinid crab (Crustacea, Brachyura) from the Upper Cretaceous of England. Bulletin of the Mizunami Fossil Museum 31: 33–35.
Collins, J.S.H. and Rasmussen, H.W. 1992. Upper Cretaceous–Lower Tertiary decapod crustaceans from west Greenland. Bulletin Grønlands Geologiske Undersøgelse 162: 1–46.
Custódio, M.A., Quaglio, F., Warren, L.V., Simões, M.G., Fürsich, F.T.J., Perinotto, J.A., and Assine, M.A. 2017. The transgressive-regressive cycle of the Romualdo Formation (Araripe Basin): Sedimentary archive of the Early Cretaceous marine ingression in the interior of Northeast Brazil. Sedimentary Geology 35: 1–15. Crossref
Dias-Brito, D. 2002. Os fósseis da bacia de Sergipe-Alagoas: calcisferas pelágicas. Phoenix 4 (39): 1–2. Available from http://www.phoenix.org.br/Phoenix39_Mar02.htm
Dias-Brito, D. 1985a. Calcisphaerulidae do Albiano da Bacia de Campos, Rio de Janeiro, Brasil. Investigações taxonômicas: biocronoestratigráficas e paleoambientais. In: D.A. Campos, C.S. Ferreira, I.M. Brito, and C.F. Viana (eds.), Coletânea de Trabalhos Paleontológicos, Geologia 27, Paleontologia e Estratigrafia 2, 295–306. Ministério das Minas e Energia, Departamento Nacional da Produção Mineral, Brasilia.
Dias-Brito, D. 1985b. Calcisphaerulidae e microfósseis associados da Formação Ponta do Mel-Bacia Potiguar, Brasil: considerações paeloecológicas e biocronoestratigráficas. In: D.A. Campos, C.S. Ferreira, I.M. Brito, and C.F. Viana (eds.), Coleção de trabalhos Paleontológicos, Geologia 27, Paleontologia e Estratigrafia 2, 307–314. Ministério das Minas e Energia, Departamento Nacional da Produção Mineral, Brasilia.
Fraaije, R.H.B., Van Bakel, B.W.M., Jagt, J.W.M., and Artal, P. 2008. New decapod crustaceans (Anomura, Brachyura) from mid-Cretaceous reefal deposits at Monte Orobe (Navarra, northern Spain), and comments on related type-Maastrichtian material. Bulletin de l’Institut Royal des Sciences Naturelles de Belgique (Sciences de la Terre) 78: 193–208.
Guinot, D., De Angeli, A., and Garassino, A. 2008. Marocarcinidae, a new eubrachyuran family, and Marocarcinus pasinii n. gen., n. sp. from the Upper Cretaceous (Cenomanian–Turonian) of Gara Sbaa, southeastern Morocco (Crustacea, Decapoda, Brachyura). Atti della Società Italiana di Scienze Naturali e del Museo Civico di Storia Naturale in Milano 149: 25–36.
Guinot, D., Vega, F.J., and Van Bakel, B. 2008. Cenomanocarcinidae n. fam., a new Cretaceous podotreme family (Crustacea, Decapoda, Brachyura, Raninoidia), with comments on related families. Geodiversitas 30: 681–719.
Karasawa, H., Schweitzer, C.E., Feldmann, R.M. 2008. Revision of the Portunoidea Rafinesque, 1815 (Decapoda: Brachyura) with emphasis on the fossil genera and families. Journal of Crustacean Biology 28: 82–127. Crossref
Karasawa, H., Schweitzer, C.E., Feldmann, R.M., and Luque, J. 2014. Systematics and phylogeny of the Raninoida (Crustacea: Brachyura). Journal of Crustacean Biology 34: 216–272. Crossref
Kellner, A.W.A. 2002. Membro Romualdo da Formação Santana, Chapada do Araripe, CE – Um dos mais importantes depósitos fossilíferos do Cretáceo brasileiro. In: C. Schobbenhaus, D.A. Campos, E.T. Queiroz, M. Winge, and M.L.C. Berbetborn (eds.), Sítios Geológicos e Paleontológicos do Brasil, 4th edition, 121–130. DNPM/CPRM – Comissão Brasileira de Sítios Geológicos e Paleontológicos (SIGEP), Brasília.
Kidwell, S.M., Fürsich, F.T., and Aigner, T. 1986. Conceptual framework for the analysis and classification of fossil concentrations. Palaios 1: 228–238. Crossref
Klompmaker, A.A. 2013. Extreme diversity of decapod crustaceans from the mid-Cretaceous (late Albian) of Spain: Implications for Cretaceous decapod paleoecology. Cretaceous Research 41: 150–185. Crossref
Larghi, C. 2004. Brachyuran decapod Crustacea from the Upper Cretaceous of Lebanon. Journal of Paleontology 78: 528–541. Crossref
Latreille, P.A. 1802. Histoire naturelle, générale et particulière des crustacés et des insectes. Ouvrage faisant suite aux œuvres de Leclerc de Buffon et partie du cours complet d’histoire naturelle rédigé par C. S. Sonnini, membre de plusieurs sociétés savantes. Vol. 1. 394 pp. F. Dufart, Paris.
Lima, F.J., Saraiva, A.A.F., and Sayão, J.M. 2012. Revisão da paleoflora das Formações Missão Velha, Crato e Romualdo, Bacia do Araripe, Nordeste do Brasil. Estudos Geológicos 22: 99–115. Crossref
Lima, M.R. 1978. Palinologia da Formação Santana (Cretáceo do Nordeste do Brasil). 337 pp. Unpublised Ph.D. Thesis, Universidade de São Paulo, São Paulo.
Lindoso, R.M., Carvalho, I.S., and Mendes, I.D. 2013. An isopod from the Codó Formation (Aptian of the Parnaíba Basin), Northeastern Brazil. Brazilian Journal of Geology 43: 16–21. Crossref
Lőrenthey, E. and Beurlen, K. 1929. Die fossilen Decapoden der Länder der Ungarischen Krone. Geologica Hungarica (Palaeontologica) 3: 1–421.
Luque, J. 2014. A new genus and species of raninoidan crab (Decapoda, Brachyura) from the Lower Cretaceous of Colombia, South America. Scripta Geologica 147: 27–34.
Luque, J. 2015a. A puzzling frog crab (Crustacea: Decapoda: Brachyura) from the Early Cretaceous Santana Group of Brazil. Frog first or crab first? Journal of Systematic Palaeontology 13: 153–166. Crossref
Luque, J. 2015b. The oldest higher true crabs (Crustacea: Decapoda: Eubrachyura): insights from the Early Cretaceous of the Americas. Palaeontology 58: 251–263. Crossref
Luque, J., Feldmann, R.M., Schweitzer, C.E., Jaramillo, C., and Cameron, C.B. 2012. The oldest frog crabs (Decapoda: Brachyura: Raninoida) from the Aptian of northern South America. Journal of Crustacean Biology 32: 405–420. Crossref
Luque, J., Schweitzer, C.E., Santana, W., Portell, R.W., Vega, F.J., and Klompmaker, A.A. 2017. Checklist of fossil decapod crustaceans from tropical America. Part I: Anomura and Brachyura. Nauplius 25: e2017025. Crossref
Mabesoone, J.M. and Tinoco, I.M. 1973. Paleoecology of the Aptian Santana Formation (Northeastern Brazil). Palaeogeography, Palaeclimatology, Palaeoecology 14: 97–118. Crossref
Maisey, J.G. 1991. Santana Fossil an Illustrated Atlas. 459 pp. Tropical Fish Hobbyist Publications, New Jersey.
Maisey, J.G. 2000. Continental break up and the distribution of fishes of Western Gondwana during the Early Cretaceous. Cretaceous Research 21: 281–314. Crossref
Maisey, J.G. and Carvalho, G.P. 1995. First records of fossil sergestid decapods and fossil brachyuran crab larvae (Arthropoda, Crustacea), with remarks on some supposed palaemonid fossils, from the Santana Formation (Aptian–Albian, NE Brazil). American Museum Novitates 3132: 1–17.
Maisey, J.G. and Moody, J.M. 2001. A review of the problematic extinct teleost fish Araripichthys, with a description of a new species from the Lower Cretaceous of Venezuela. American Museum Novitates 3324: 1–27. Crossref
Manso, C.L.M. and Hessel, M.H. 2007. Revisão sistemática Pygidiolampas araripensis (Beurlen, 1966), (Echinodermata: Cassiduloida) da Bacia do Araripe, Nordeste do Brasil. Geociências 26: 271–277.
Marangon, S. and De Angeli, A. 1997. Cherpiocarcinus, nuovo genere di brachiuro (Decapoda) dell’Oligocene del Bacino Ligure-Piemontese (Italia settentrionale). Lavori – Società Veneziana di Scienze Naturali 22: 97–106.
Martins-Neto, R.G. 1987. Primeiro registro de decápode na Formação Santana, Bacia do Araripe (Cretáceo Inferior), Brasil. Ciência e Cultura 39: 406–410.
Martins-Neto, R.G. and Mezzalira, S. 1991. Descrição de novos crustáceos (Caridea) da Formaçao Santana Cretáceo Inferior do Nordeste do Brasil. Anais da Academia Brasileira de Ciências 63: 155–160.
Matos, R.M.D. 1992. The northeast Brazilian rift system. Tectonics 11: 766–791.Crossref
Maury, C.J. 1936. O Cretáceo de Sergipe, Brasil. Monographias do Serviço Geológico e Mineralógico 11: 1–283.
Monteiro, F.A.C., Meirelles, C.A.O., and Soares, M.O. 2010. Bivalves fósseis do gênero Neithea (Bivalvia: Pectinidae) e seu significado paleogeográfico para o domínio tetiano na Bacia Potiguar. Geociências 29: 335–341.
Ossó, À. 2016. Eogeryon elegius n. gen. and n. sp. (Decapoda: Eubrachyura: Portunoidea), one of the oldest modern crabs from late Cenomanian of the Iberian Peninsula. Boletín de la Sociedad Geológica Mexicana 68: 231–246. Crossref
Pereira, P.A., Cassab, R.C.T., and Barreto, A.M.F. 2016a. Cassiopidae gastropods, influence of Tethys Sea of the Romualdo Formation (Aptiane–Albian), Araripe Basin, Brazil, Journal of South American Earth Sciences 70: 211–223. Crossref
Pereira, P.A., Cassab, R.C.T., and Barreto, A.M.F. 2017. Paleoecologia e paleogeografia dos moluscos e equinoides da Formação Romualdo, Aptiano–Albiano da Bacia do Araripe, Brasil. Anuário do Instituto de Geociências-UFRJ 40: 180–198. Crossref
Pereira, P.A., Cassab, R.C.T., Barreto, A.M.F., and Almeida, J.A.C. 2016b. Moluscos da Formação Romualdo, Aptiano–Albiano, Bacia do Araripe, Nordeste do Brasil. Boletim do Museu Paraense Emílio Goeldi, Ciências Naturais 10: 231–246.
Péron, A. 1877. Observations sur la faune des Calcaires à Echinides de Rennes-les-Bains et sur quelques fossiles du terrain crétacé supérieur. Bulletin de la Société Géologique de France 3: 499–535.
Petri, S. 1987. Cretaceous paleogeographic maps of Brazil. Palaeogeography, Palaeoclimatology, Palaeoecology 59: 117–168. Crossref
Pinheiro, A.P., Alencar, D.R., Sayão, J.M., and Santiago, A.C. 2016. Primeiro registro fossilífero de Solenoceridae Wood-Mason, 1981 (Decapoda: Penaeioidea) para bacias sedimentares do Brasil. In: D. Lima and J. Colavite (eds.), 14º Congresso Brasileiro sobre Crustáceos, Crato, Resumos, 293. Sociedade Brasileira de Carcinologia, Crato-CE.
Pinheiro, A.P., Saraiva, A.Á.F., and Santana, W. 2014. Shrimps from the Santana Group (Cretaceous: Albian): new species (Crustacea: Decapoda: Dendrobranchiata) and new record (Crustacea: Decapoda: Caridea). Anais da Academia Brasileira de Ciências 86: 663–670. Crossref
Prado, L.A.C., Pereira, P.A., Sales, A.M.F., and Barreto, A.M.F. 2016. Tafonomia dos invertebrados do Sítio Canastra, Formação Romualdo, Cretáceo Inferior, Bacia do Araripe, Araripina, Pernambuco, Brasil. Anuário do Instituto de Geociências 39: 77–87. Crossref
Prado, L.A.C., Pereira, P.A., Sales, A.M.F., and Barreto, A.M.F. 2015. Taphonomic and paleoenvironmental considerations for the concentrations of macroinvertebrate fossils in the Romualdo Member, Santana Formation, Late Aptian–Early Albian, Araripe Basin, Araripina, NE, Brazil. Journal of South American Earth Sciences 62: 218–228. Crossref
Sales, A.M.F. 2005. Análise tafonômica das ocorrências fossilíferas de macroinvertebrados do Membro Romualdo (Albiano) da Formação Santana, Bacia do Araripe, NE do Brasil: significado estratigráfico e paleoambiental. 160 pp. Unpublished Ph.D. Thesis, Universidade de São Paulo, São Paulo.
Santana, W., Pinheiro, A.P., Silva, C.M.R., and Saraiva, A.Á.F. 2013. A new fossil caridean shrimp (Crustacea: Decapoda) from the Cretaceous (Albian) of the Romualdo Formation, Araripe Basin, northeastern Brazil. Zootaxa 3620: 293–300. Crossref
Saraiva, A.Á.F., Pinheiro, A.P., and Santana, W. 2018. A remarkable new genus and species of the planktonic shrimp family Luciferidae (Crustacea, Decapoda) from the Cretaceous (Aptian/Albian) of the Araripe Sedimentary Basin, Brazil. Journal of Paleontology 92: 459–465. Crossref
Saraiva, A.Á.F., Pralon, B.G.N., and Gregati, R.A. 2009. Taxonomic remarks on Brazilian Cretaceous Decapoda from Araripe Basin, Brazil, and ecological inferences. Gaea 5: 70–74. Crossref
Schweitzer, C.E., Dworschak, P.C., and Martin, J.W. 2011. Replacement names for several fossil Decapoda. Journal of Crustacean Biology 31: 361–363. Crossref
Schweitzer, C.E., Feldmann, R.M., and Frantescu, O.D. 2016a. Early Cretaceous (Albian) decapods from the Glen Rose and Walnut formations of Texas, USA. Bulletin of the Mizunami Fossil Museum 42: 1–22.
Schweitzer, C.E., Feldmann, R.M., and Lamanna, M.C. 2012. New genus of crab (Brachyura: Raninoida: Necrocarcinidae) from the Upper Cretaceous of West Antarctica, with description of a new species. Annals of Carnegie Museum 80: 147–158. Crossref
Schweitzer, C.E., Feldmann, R.M., Fam, J., Hessin, W.A., Hetrick, S.W., Nyborg, T.G., and Ross, R.L.M. 2003. Cretaceous and Eocene Decapod Crustaceans from Southern Vancouver Island, British Columbia, Canada. 66 pp. NRC Research Press, Ontario.
Schweitzer, C.E., Karasawa, H., Luque, J., and Feldmann, R.M. 2016b. Phylogeny and classification of Necrocarcinoidea Föster, 1968 (Brachyura:Raninoida) with the description of two new genera. Journal of Crustacean Biology 36: 338–372. Crossref
Silva-Santos, R. 1985. Paleoictiofauna da Formação Codó, Bacia do Parnaíba, NE do Brasil. In: 9° Congresso Brasileiro de Paleontologia, Resumos, 11. SBP, Fortaleza.
Silva-Santos, R. and Valença, J.G. 1968. A Formação Santana e sua paleoictiofauna. Anais da Academia Brasileira de Ciências 40: 339–360.
Van Bakel, B.W.M., Guinot, D., Artal, P., Fraaije, R.H.B., and Jagt, J.W.M. 2012. A revision of the Palaeocorystoidea and the phylogeny of raninoidian crabs (Crustacea, Decapoda, Brachyura, Podotremata). Zootaxa 3215: 1–216.
Varejão, F.G., Warren, L.V., Perinotto, J.A. J., Neumann, V.H., Freitas, B.T., Almeida, R.P., and Assine, M.L. 2016. Upper Aptian mixed carbonate-siliciclastic sequences from Tucano Basin, Northeastern Brazil: implications for paleogeographic reconstructions following Gondwana break-up. Cretaceous Research 67: 44–58. Crossref
Viana, M.S.S. and Agostinho, S.M. 1995. Camarões do Membro Romualdo da Formação Santana (Cretáceo Inferior da Bacia do Araripe). Boletim do 16º Simpósio de Geologia do Nordeste Sociedade Brasileira Geologia 1: 239–243.
White, C.A. 1887. On new generic forms of Cretaceous Mollusca and their relation to other forms. Proceedings of the Academy of Natural Sciences of Philadelphia 39: 32–37.
Acta Palaeontol. Pol. 63 (4): 737–750, 2018
https://doi.org/10.4202/app.00480.2018