
A new relict stem salamander from the Early Cretaceous of Yakutia, Siberian Russia
PAVEL P. SKUTSCHAS, VENIAMIN V. KOLCHANOV, ALEXANDER O. AVERIANOV, THOMAS MARTIN, RICO SCHELLHORN, PETR N. KOLOSOV, and DMITRY D. VITENKO
Skutschas, P.P. Kolchanov, V.V., Averianov, A.O., Martin, T., Schellhorn, R., Kolosov, P.N., and Vitenko, D.D. 2018. A new relict stem salamander from the Early Cretaceous of Yakutia, Siberian Russia. Acta Palaeontologica Polonica 63 (3): 519–525.
A new stem salamander, Kulgeriherpeton ultimum gen. et sp. nov., is described based on a nearly complete atlas from the Lower Cretaceous (Berriasian–Barremian) Teete vertebrate locality in southwestern Yakutia (Eastern Siberia, Russia). The new taxon is diagnosed by the following unique combination of atlantal characters: the presence of a transversal ridge and a depression on the ventral surface of the posterior portion of the centrum; ossified portions of the intercotylar tubercle represented by dorsal and ventral lips; the absence of a deep depression on the ventral surface of the anterior portion of the centrum; the absence of pronounced ventrolateral ridges; the absence of spinal nerve foramina; the presence of a pitted texture on the ventral and lateral surfaces of the centrum and lateral surfaces neural arch pedicels; the presence of a short neural arch with its anterior border situated far behind the level of the anterior cotyles; moderately dorsoventrally compressed anterior cotyles; and the absence of a deep incisure on the distal-most end of the neural spine. The internal microanatomical organization of the atlas is characterized by the presence of a thick, moderately vascularized cortex and inner cancellous endochondral bone. The recognition of stem salamanders and other vertebrates with Jurassic affinities in the Early Cretaceous high-latitude (paleolatitude estimate N 63–70°) vertebrate assemblage of Teete suggests that: (i) the large territory of present day Siberia was a refugium for Jurassic relicts; (ii) there were no striking differences in the composition of high-latitude Yakutian and mid-latitude Western Siberian Early Cretaceous vertebrate assemblages; and (iii) there was a smooth transition from the Jurassic to Cretaceous biotas in North Asia.
Key words: Amphibia, Caudata, stem salamanders, Cretaceous, Russia, Siberia.
Pavel P. Skutschas [skutchas@mail.ru, p.skutschas@spbu.ru], Veniamin V. Kolchanov [veniamin.kolchanov@mail.ru], and Dmitry D. Vitenko [mvitenko98@gmail.com], Vertebrate Zoology Department, Saint Petersburg State University, Universitetskaya nab. 7/9, 199034 Saint Petersburg, Russia.
Alexander O. Averianov [dzharakuduk@mail.ru], Zoological Institute of the Russian Academy of Sciences, Universitetskaya nab. 1, 199034 Saint Petersburg, Russia; Institute of Earth Sciences, Saint Petersburg State University, 16 Liniya VO 29, 199178 Saint Petersburg, Russia; Institute of Geology and Petroleum Technology, Kazan Federal University, Kremlevskaya str. 4/5, 420008 Kazan, Russia.
Thomas Martin [tmartin@uni-bonn.de] and Rico Schellhorn [rico.schellhorn@uni-bonn.de], Steinmann-Institut für Geologie, Mineralogie und Paläontologie, Universität Bonn, Nussallee 8, 53115 Bonn, Germany.
Petr N. Kolosov [p_kolosov@diamond.ysn.ru], Institute of Diamond and Precious Metals Geology, Siberian Branch of the Russian Academy of Sciences, pr. Lenina 39, 677980 Yakutsk, Russia.
Received 24 April 2018, accepted 1 June 2018, available online 26 July 2018.
Copyright © 2018 P.P. Skutschas et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Stem salamanders are critical for documenting primitive character states and patterns of character state transformations within Caudata. The geologically oldest stem salamanders include the following taxa: Kokartus Nesov, 1988 from the Middle Jurassic (Bathonian) of Kyrgyzstan; Marmorerpeton Evans, Milner, and Mussett, 1988 (two species M. kermacki and M. freemani) and “Kirtlington salamander A” (sensu Evans and Milner 1994), all from the Middle Jurassic (Bathonian) of Great Britain; and Urupia Skutschas and Krasnolutskii, 2011 and “Berezovsk salamander A” (sensu Skutschas 2013), both from the Middle Jurassic (Bathonian) of Russia (Evans et al. 1988; Nesov 1988; Evans and Milner 1994; Evans and Waldman 1996; Milner 2000; Skutschas and Krasnolutskii 2011; Skutschas and Martin 2011; Skutschas 2013, 2016a). The next younger stem salamanders date from the Late Jurassic. These include two Kimmeridgian occurrences, namely Karaurus Ivachnenko, 1978 from the Karabastau Formation in Kazakhstan and “cf. Marmorerpeton” (sensu Wiechmann 2000) from Portugal (Ivachnenko 1978; Milner 2000; Wiechmann 2000), plus a possible stem salamander from the Kimmeridgian–early Tithonian (Morrison Formation) of the USA (Nesov 1992; Evans and Milner 1993; Gardner and DeMar 2013). Outside China, all Bathonian vertebrate assemblages containing salamanders are dominated by stem salamanders but later, at the end of the Middle Jurassic and into the Late Jurassic, crown group salamanders became the dominant salamander component in vertebrate assemblages everywhere. Except for a possible stem salamander in the Early Cretaceous of North America (Gardner and DeMar 2013), there had been no unequivocal evidence of stem salamanders in post Jurassic deposits, and it was suggested that they became extinct before the Jurassic–Cretaceous boundary (Skutschas 2013). In 2016, a fragmentary trunk vertebra of a stem salamander from Lower Cretaceous (Aptian–Albian) deposits at Shestakovo in Western Siberia was described (Skutschas 2016b). That discovery was the first conclusive evidence that the stem salamanders survived beyond the Jurassic–Cretaceous boundary and co-existed with crown group salamanders during approximately the first 40 million years of the known history of salamanders.
In 2017 PS, RS, and DV conducted field work at the Lower Cretaceous Teete locality in southwestern Yakutia (Eastern Siberia, Russia; Fig. 1). That locality was formed close to Mesozoic polar latitudes (paleolatitude estimate 63–70° N) and has become known for its polar dinosaurs (Rich et al. 1997, 2002). On-site excavation and screen-washing of 500 kg of fossiliferous matrix produced numerous vertebrate microfossils, including remains of fishes, salamanders, turtles, choristoderes, lizards, dinosaurs, tritylodontids, and mammals. Among the vertebrate remains collected in 2017 at Teete, one atlas (ZIN PH 3/246) erected as the holotype of the new stem salamander taxon described herein. This is additional evidence for a survival of stem salamanders beyond the Jurassic–Cretaceous boundary and the presence of a vast Early Cretaceous Siberian refugium for Jurassic relicts among vertebrates.
Institutional abbreviations.—ZIN PH, Paleoherpetological collection, Zoological Institute, Russian Academy of Sciences, Saint Petersburg, Russia.
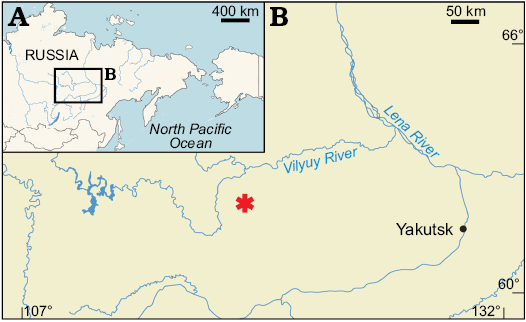
Fig. 1. A, B. Maps showing the geographic location of the Lower Cretaceous Teete locality (asterisk) in Eastern Siberia, Russia.
Material and methods
The atlas ZIN PH 3/246 (holotype of Kulgeriherpeton ultimum gen. et sp. nov.) is fragile and is partly embedded in matrix (Fig. 2), therefore some details of its structure are not visible. To overcome this problem and to gain additional morphological information, ZIN PH 3/246 was CT scanned (at 100kV and 0.1 mA, generating a resolution of 4.4 μm of pixel size and an output of 4000×4000 pixels per slice) at the Saint Petersburg State University Research Centre for X-ray Diffraction Studies (Saint Petersburg, Russia) using a Skyscan 1172. Segmentation of the CT scan data and 3D model reconstructions were made with Amira 6.3.0 (FEI-VSG Company).
ZIN PH 3/246 is stored at the Paleoherpetological collection (ZIN PH) of the Zoological Institute of the Russian Academy of Sciences, Saint Petersburg, Russia.
The CT data are deposited in the Department of Vertebrate Zoology of the Saint Petersburg State University, Saint Petersburg, Russia and can be made available by the corresponding author for the purpose of scientific study.
Systematic palaeontology
Amphibia Linnaeus, 1758
Caudata Scopoli, 1777
Genus Kulgeriherpeton nov.
Type species: Kulgeriherpeton ultimum sp. nov., monotypic; see below.
Etymology: From Yakutian kulgeri, lizard, salamander; and Greek ἑρπετόν [herpeton], a creeping animal.
Kulgeriherpeton ultimum sp. nov.
Figs. 2–4, 5A.
Etymology: From Latin ultimus, last; in reference to the relict nature of this stem salamander.
Holotype: ZIN PH 3/246, nearly complete atlas partially embedded in matrix.
Type locality: Teete, Suntar Ulus, Yakutia, Eastern Siberia, Russia.
Type horizon: Batylykh Formation, Sangar Series, Berriasian–Barremian, Lower Cretaceous, for geological settings see Kolosov et al. (2009); Averianov et al. (2018).
Diagnosis.—Placed outside the crown group (Urodela) and referred to stem group salamanders based on the absence of spinal nerve foramina in the atlas, the presence of a pitted texture on the ventral and lateral surfaces of the atlas, the presence of an anteroposteriorly short neural arch with an anterior border that is situated far from level of anterior cotyles, and its relatively large size (the maximum anterior width is about 8.8 mm and the ventral midline length, excluding the intercotylar tubercle is about 6.4 mm). Differs from all other stem salamanders for which the morphology of the atlantal centrum is known (namely Kokartus from the Middle Jurassic (Bathonian) of Kyrgyzstan, Marmorerpeton from the Middle Jurassic (Bathonian) of Great Britain, and Urupia and “Berezovsk salamander A” from the Middle Jurassic (Bathonian) of Russia) in the presence of a transversal ridge and a depression on the ventral surface of the posterior portion of the centrum. Differs further from Marmorerpeton, Urupia, and “Berezovsk salamander A” by presence of an intercotylar tubercle on the atlas with ossified dorsal and ventral lips (vs. absence of the intercotylar tubercle and presence of only the notochondral central pit in Urupia, “Berezovsk salamander A” and M. freemani; and no division of the intercotylar tubercle into dorsal and ventral lips in M. kermacki). Additionally, Kulgeriherpeton differs from Urupia by the lack of a deep depression on the ventral surface of the anterior portion of the atlantal centrum, by the lack of pronounced ventro-lateral ridges on the atlas, and by less dorso-ventrally compressed atlantal anterior cotyles (ratio of maximum height/width about 0.79 vs. about 0.5 in Urupia); additionally differs from “Berezovsk salamander A” in the atlantal centrum being relatively longer (ratio of maximum anterior width, i.e., between lateral rims of the anterior cotyles), midline length (excluding intercotylar tubercle) is about 1.37 vs. about 1.75 in “Berezovsk salamander A”; and additionally differs from Karaurus in the absence of a deep incisure on the distal-most end of the atlantal neural spine.
Description.—The atlantal centrum (Figs. 2A, 3) is slightly wider than long: the maximum anterior width (i.e., between the lateral rims of the anterior cotyles) is about 8.8 mm and the ventral midline length, excluding the intercotylar tubercle (= odontoid process), is about 6.4 mm (ratio of maximum anterior width/ventral midline length without intercotylar tubercle about 1.38). The anterior cotyles (Fig. 3K) are large and slightly dorso-ventrally compressed (ratio of maximum height/width about 0.8). The articular surfaces of the anterior cotyles are moderately concave. The intercotylar tubercle is not fully ossified and is represented by dorsal and ventral lips that are separated by a narrow strip of the articular surface extending between the anterior cotyles. The posterior cotyle is nearly circular in posterior outline (Fig. 3G). The inner surface of the posterior cotyle is deeply concave (Fig. 3G, J, L).

Fig. 2. Stem salamander Kulgeriherpeton ultimum gen. et sp. nov. from Teete locality, Yakutia, Eastern Siberia, Russia; Sangar Series, Batylykh Formation, Berriasian–Barremian, Lower Cretaceous. Atlas ZIN PH 3/246 (holotype) in dorsal (A) and ventral (B) views, with anterior end to top of figure.
The ventral surface of the centrum has a shallow median depression (Fig. 3H). The surface of the median depression is perforated by three relatively large subcentral foramina. Antero-lateral to the medial depression, there is a pair of narrow oblique grooves. Ventro-lateral ridges are absent. A distinct transverse ridge and a relatively deep transverse depression, the latter being situated just behind the transverse ridge, extend across the ventral surface of the posterior portion of the centrum.
The lateral surfaces of the centrum (Fig. 3A–D, F, I, J, L) have distinct unipartite transverse processes (the left transverse process is more prominent). The transverse processes are short and do not project far from the lateral wall of the centrum. There are relatively deep lateral depressions on the lateral surfaces of the centrum. There are three ridges on the lateral surface of the centrum; two are short (anterior and posterior alar processes) and are associated with the transverse process and one longitudinal lateral ridge flanks the dorsal border of the lateral depression. Like the transverse processes, the three ridges are more distinct and prominent on the left side. Hypapophyses and basapophyses are absent.
The neural arch is high and antero-posteriorly short, with its anterior border situated far (about one third of the length of the centrum) behind the level of the anterior cotyles (Fig. 3E, F, L). The pedicels of the neural arch are massive. The spinal nerve foramen is absent, but there is a groove for passage of the first spinal nerve on the anterior edge and the antero-lateral part of the pedicel of the neural arch (Fig. 3A, K). The neural canal is broad and low in anterior and posterior view (Fig. 3G, K). The neural arch roof is posteriorly short and extends back only to about the level of the rim of the posterior cotyle (Fig. 3F, L).
The dorso-median surface of the neural arch roof bears a high, posteriorly massive neural spine. The postero-dorsal end of the neural spine terminates in a deeply concave scar indicating that in life the distal-most end of the spine was cartilaginous. The posterior surface of the neural spine has a pair of dorso-ventrally elongated lateral depressions. The postzygapophyseal proceses project postero-laterally (approximately 45° from the midline). The postzygapophyseal facets are relatively wide, oval in outline and face ventrally and slightly laterally (Fig. 3G, L).
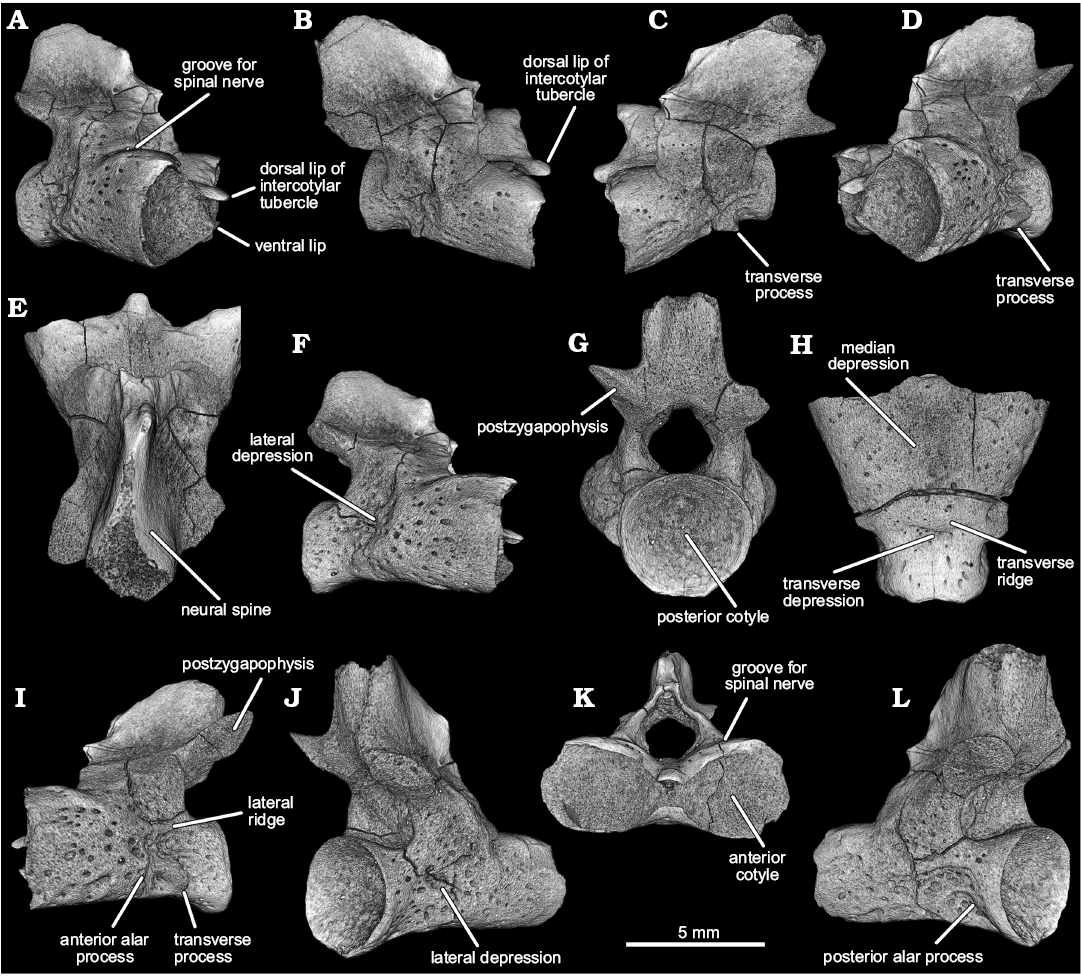
Fig. 3. Stem salamander Kulgeriherpeton ultimum gen. et sp. nov. from Teete locality, Yakutia, Eastern Siberia, Russia; Sangar Series, Batylykh Formation, Berriasian–Barremian, Lower Cretaceous. Digital restoration of atlas ZIN PH 3/246 (holotype) in right antero-lateral (A), right dorso-lateral (B), left dorso-lateral (C), left antero-lateral (D), dorsal (E), right lateral (F) posterior (G), ventral (H), left lateral (I), right postero-lateral (J), anterior (K), and left postero-lateral (L) views.
The ventral and lateral surfaces of the centrum and the lateral surfaces of the neural arch pedicels are rugouse and indented by scattered, small, rounded and oval pits.
The internal microanatomical organization of the centrum is characterized by the presence of a thick compact periosteal cortex and an inner cancellous endochondral bone with numerous erosion bays that are separated by irregularly arranged trabeculae of varying thickness (Fig. 4). The periosteal cortex is moderately vascularized by a network of short vertical and subvertical vascular canals. These vascular canals access the external surface of the bone where they are visible as rounded and oval pits forming a characteristic pitted texture. Internally, the vascular canals of the cortex are connected with erosion bays of the inner cancellous endochondral bone. A narrow notochordal canal is present and centrally located. The notochordal canal is partly infilled by a bone. The neural arch has a microanatomical organization similar to that of the centrum, but differs in a lower degree of vascularization of the cortex.
Stratigraphic and geographic range.—Type locality and horizon only.
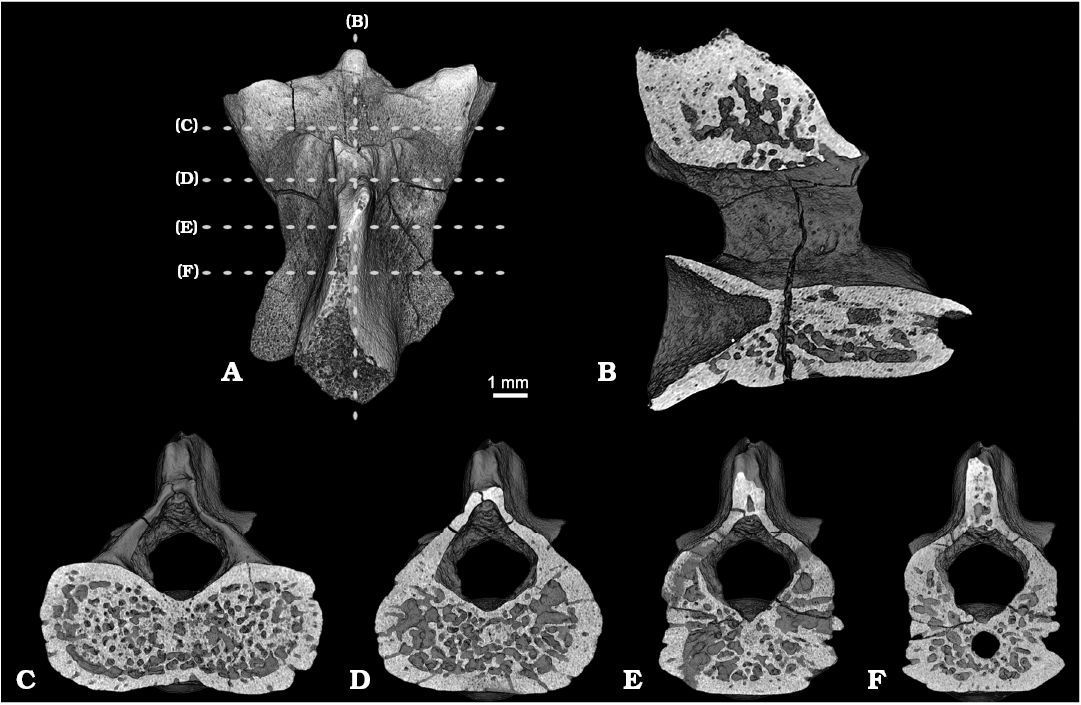
Fig. 4. Stem salamander Kulgeriherpeton ultimum gen. et sp. nov. from Teete locality, Yakutia, Eastern Siberia, Russia; Sangar Series, Batylykh Formation, Berriasian–Barremian, Lower Cretaceous. Digital restoration of atlas ZIN PH 3/246 (holotype), detailed anatomy with the locations of the microCT digital sections, dorsal view (A); longitudinal section, vertical plane (B); transverse sections (C–F).
Discussion and conclusions
Kulgeriherpeton ultimum gen. et sp. nov. is placed outside the crown group (Urodela) based on a combination of atlantal features that is primitive for crown salamanders and characteristic for stem group salamanders: the absence of spinal nerve foramina in the atlas (vs. spinal nerve foramen fully enclosed by bone in crown salamanders); the presence of pitted ventral and lateral surfaces of the atlas (vs. smooth external vertebral surface in most crown salamanders); and its relatively large size (vs. relatively smaller size in the most of early diverging crown group members (see e.g., Skutschas 2016b). Additionally, Kulgeriherpeton ultimum gen. et sp. nov., like all other stem salamanders, is characterized by an antero-posteriorly short neural arch with its anterior border situated far behind the level of the anterior cotyles (vs. long neural arch with its anterior border situated at the level of the anterior cotyles in crown salamanders) (Jim Gardner, personal communication 2016; Fig. 5). Kulgeriherpeton ultimum gen. et sp. nov. shares several atlantal features with the karaurid stem salamander Kokartus from the Middle Jurassic (Bathonian) of Kyrgyzstan (the atlas is partly visible in the articulated holotype skeleton of the karaurid Karaurus) namely: similar proportions of the anterior cotyles; an intercotylar tubercle that is represented by dorsal and ventral lips; the absence of distinct ventro-lateral ridges; and the presence of transverse processes). Those similarities indicate a possible karaurid affinity for the new stem salamander from Teete.
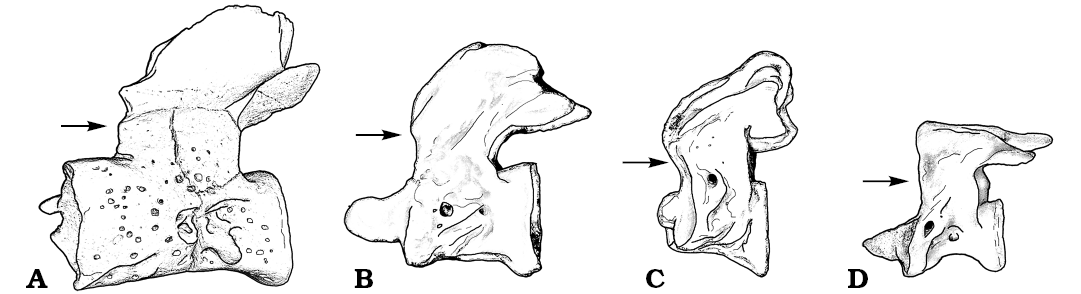
Fig. 5. Drawings of atlases of stem (A) and extant crown (B–D) salamanders in lateral views. A. Kulgeriherpeton ultimum gen. et sp. nov. B. Cryptobranchus. C. Amphiuma. D. Hynobius. Note the antero-posteriorly short neural arch with its anterior border situated far behind the level of the anterior cotyles in stem salamanders (A) and long neural arch with its anterior border is situated at the level of the anterior cotyles in crown salamanders (B–D). Arrows show the anterior border of the neural arch. Not to scale.
According to previously known and recently collected material, the high latitude vertebrate assemblage of Teete includes paleonisciform and amiid fishes, two salamander taxa (Kulgeriherpeton ultimum gen. et sp. nov. and a small undescribed crown salamander), paramacellodid lizards, primitive (= non-neochoristoderan) choristoderes, turtles, various dinosaurs (stegosaurs, ornithopods, theropods, and sauropods), the tritylodontid synapsid Stereognathus kolossovi and three mammaliaform taxa (haramiyid cf. Sineleutherus sp., a new genus of tegotheriid docodont and a new genus of eutriconodont) (Kurzanov et al. 2000, 2003; Lopatin and Agadjanian 2008; Kolosov et al. 2009; Averianov et al. 2018). In spite of the Early Cretaceous age of the fossiliferous sediments at Teete (Kolosov et al. 2009), the mammaliaform, tritylodontid, salamander and dinosaur components of the Teete vertebrate assemblage have a distinctive Jurassic appearance, being similar to Middle–Late Jurassic vertebrate assemblages known from Western Siberia (Russia), Xinjiang (China), and Great Britain (Evans and Milner 1994; Evans and Waldman 1996; Maisch et al. 2001, 2003; Martin et al. 2010; Skutschas 2013; Averianov et al. 2016, 2018; Skutschas et al. 2016a).
On the other hand, the Teete vertebrate assemblage shares many components (namely stem salamanders, primitive choristoderes, paramacellodid lizards, stegosaurs, tritylodonts and eutriconodont and docodont mammaliaforms) with the Early Cretaceous vertebrate assemblage of the Shestakovo locality (Ilek Formation) in Western Siberia (Russia) (Averianov et al. 2006; Skutschas 2016b). As suggested earlier, Western Siberia possibly was a refugium where vertebrates with Jurassic affinities (including relict salamanders) survived (Leshchinskiy et al. 2001; Skutschas 2016b). The similarity of the high-latitude vertebrate assemblage of Teete and the mid-latitude vertebrate assemblage of Shestakovo suggests that the territory of a refugium for Jurassic vertebrate relicts was not restricted to modern day Western Siberia in the Early Cretaceous, but spread farther into the northeastern part of the Asiatic continent. It also indicates the lack of striking faunistic differences between high-latitude (Teete) and some mid-latitude (Shestakovo) Early Cretaceous vertebrate assemblages of Siberia. Finally, the presence of vertebrates with Jurassic affinities (including stem salamanders) in the Early Cretaceous of Siberia supports the hypothesis (Averianov et al. 2018) that the transition from the Jurassic to Cretaceous biotas in North Asia, in contrast to the environmental perturbations that occurred in other regions at the Jurassic/Cretaceous boundary (Tennant et al. 2017), was smooth.
Acknowledgements
We thank Sergey M. Nikolaev (Suntar Inspection of Nature Protection, Suntar, Russia) for logistic support of the expedition in 2017. We are grateful to Andrey N. Nikolaev and other teachers and workers of the Khoro school (Athanasiy V. Ivanov, Sayan S. Ivanov, Elmira N. Grigorieva) as well as pupils of this school (Petr Grigoriev, Evgeniy Grigoriev, Vasiliy Samsonov, Timophey Nikolaev), and Alexey M. Ignatiev and Albert V. Stepanov (Khoro village, Russia) for their hospitality and help during fieldwork. The authors thank Jim Gardner (Royal Tyrrell Museum of Palaeontology, Drumheller, Canada) and Jianye Chen (American Museum of Natural History, New York, USA) for providing helpful comments that improved the quality of the manuscript. We thank the staff of the Saint Petersburg State University Research Centre for X-ray Diffraction Studies (Saint Petersburg, Russia) for CT scanning of the specimen. Financial support was provided by the Russian Foundation for Basic Research (project 17-54-12041-NNIO) and the Deutsche Forschungsgemeinschaft (DFG grant MA 1643/22-1). AA was also supported by Zoological Institute, Russian Academy of Sciences (project АААА-А17-117022810195-3) and by the Program of the Russian Academy of Sciences Presidium “Evolution of the organic world. The role and significance of planetary processes.” The work was performed according to the Russian Government Program of Competitive Growth of Kazan Federal University.
References
Averianov, A.O., Martin, T., Lopatin, A.V., Skutschas, P.P., Schellhorn, R., Kolosov, P.N., and Vitenko, D.D. 2018. A high-latitude fauna of mid-Mesozoic mammals from Yakutia, Russia. PLOS ONE [published online https://doi.org/10.1371/journal.pone.0199983].
Averianov, A.O., Martin, T., Skutschas, P.P., Danilov, I.G., Schultz, J., Schellhorn, R., Obraztsova, E., Lopatin, A., Sytchevskaya, E., Kuzmin, I., Krasnolutskii, S., and Ivantsov, S. 2016. Middle Jurassic vertebrate assemblage of Berezovsk coal mine in western Siberia (Russia). Global Geology 19: 187–204.
Averianov, A.O., Voronkevich, A.V., Leshchinskiy, S.V., and Fayngertz, A.V. 2006. A ceratopsian dinosaur Psittacosaurus sibiricus from the Early Cretaceous of West Siberia, Russia and its phylogenetic relationships. Journal of Systematic Paleontology 4: 359–395. Crossref
Evans, S.E. and Milner, A.R. 1993. Frogs and salamanders from the Upper Jurassic Morrison Formation (Quarry Nine, Como Bluff) of North America. Journal of Vertebrate Paleontology 13: 24–30. Crossref
Evans, S.E. and Milner, A.R. 1994. Microvertebrate faunas from the Middle Jurassic of Britain. In: N. Fraser and H.D. Sues (eds.), In the Shadow of the Dinosaurs: Early Mesozoic Tetrapods, 303–321. Cambridge University Press, Cambridge.
Evans, S.E. and Waldman, M. 1996. Small reptiles and amphibians from the Middle Jurassic of Skye, Scotland. In: M. Morales (ed.), The Continental Jurassic. Museum of Northern Arizona, Bulletin 60: 219–226.
Evans, S.E., Milner, A.R., and Mussett, F. 1988. The earliest known salamanders (Amphibia, Caudata): a record from the Middle Jurassic of England. Geobios 21: 539–552. Crossref
Gardner, J.D. and DeMar, D.G. Jr. 2013. Mesozoic and Paleocene lissamphibian assemblages of North America: a comprehensive review. In: J.D. Gardner and R.L. Nydam (eds.), Mesozoic and Cenozoic Lissamphibian and Squamate Assemblages of Laurasia. Palaeobiodiversity and Palaeoenvironments 93: 459–515.
Ivachnenko, M. 1978. Urodeles from the Triassic and Jurassic of Soviet Central Asia. [in Russian]. Paleontologičeskij žurnal 1978 (3): 84–89.
Kolosov, P.N., Ivensen, G.V., Mikhailova, T.E., Kurzanov, S.M., Efimov, M.B., and Gubin, Y.M. 2009. Taphonomy of the Upper Mesozoic tetrapod Teete locality (Yakutia). Paleontological Journal 43: 201–207. Crossref
Kurzanov, S.M., Efimov, M.B., and Gubin, Yu.M. 2000. Dinosaurs from Yakutia [in Russian]. In: A.V. Komarov (ed.), Materialy regional’noi konferentsii geologov Sibiri, Dal’nego Vostoka i Severo-Vostoka Rossii, Vol. 2, 356–357. GalaPress, Tomsk.
Kurzanov, S.M., Efimov, M.B., and Gubin, Y.M. 2003. New archosaurs from the Jurassic of Siberia and Mongolia. Paleontological Journal 37: 53–57.
Leshchinskiy, S.V., Voronkevich, A.V., Fayngertz, A.V., Maschenko, E.N., Lopatin, A.V., and Averianov, A.O. 2001. Early Cretaceous vertebrate locality Shestakovo, Western Siberia, Russia: a refugium for Jurassic relicts? Journal of Vertebrate Paleontology 21: 73A.
Linnaeus, C. 1758. Systema naturae per regna tria naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Vol. 1: Regnum animale. Editio decima, reformata. 824 pp. Laurentius Salvius, Stockholm.
Lopatin, A.V. and Agadjanian, A.K. 2008. A tritylodont (Tritylodontidae, Synapsida) from the Mesozoic of Yakutia. Doklady Biological Sciences 419: 107–110. Crossref
Maisch, M.W., Matzke, A. T., Ye, J., Pfretzschner, H.-U., and Sun, G. 2001. The fossil vertebrate faunas of the Toutunhe and Qigu Formations of the Southern Junggar Basin and their biostratigraphical and palecological implications. In: G. Sun, V. Mosbrugger, A.R. Ashraf, and Y.D. Wang (eds.), The Advanced Study of Prehistory Life and Geology of Junggar Basin, 83–94. Xinjiang, China.
Maisch, M.W., Matzke, A.T., Pfretzschner, H.-U., Sun, G., Sthr, H., and Grossmann, F. 2003. Fossil vertebrates from the Middle and Upper Jurassic of the Southern Junggar Basin (NW-China)—results of the Sino-German expeditions 1999–2000. Neues Jahrbuch für Geologie und Paläontologie, Monatshefte 2003 (5): 297–313.
Martin, T., Averianov, A.O., and Pfretzschner, H.-U. 2010. Mammals from the Late Jurassic Qigu Formation in the Southern Junggar Basin, Xinjiang, Northwest China. Palaeobiodiversity and Palaeoenvironments 90: 295–319. Crossref
Milner, A.R. 2000. Mesozoic and Tertiary Caudata and Albanerpetontidae. In: H. Heatwole and R.L. Carroll (eds.), Amphibian Biology, Vol. 4, Palaeontology, The Evolutionary History of Amphibians, 1412–1444. Surrey Beatty and Sons, Chipping Norton.
Nesov, L.A. 1988. Late Mesozoic amphibians and lizards of Soviet Middle Asia. Acta Zoologica Cracoviensia 31: 475–486.
Nesov, L.A. 1992. New salamanders from the Upper Jurassic Morrison Fm. of Dinosaur National Monument. In: V.L. Santucci (ed.), National Park Service Paleontological Research Abstract Volume, 3rd Fossil Resources Conference, 30. Fossil Butte National Monument, Wyoming.
Rich, T.H.V., Gangloff, R.A., and Hammer, W.R. 1997. Polar dinosaurs. In: P.J. Currie and K. Padian (eds.), Encyclopedia of Dinosaurs, 562–573. Academic Press, San Diego.
Rich, T.H.V., Vickers-Rich, P., and Gangloff, R.A. 2002. Polar dinosaurs. Science 295: 979–980. Crossref
Scopoli, G.A. 1777. Introductio ad historiam naturalem, sistens genera lapidum, plantarum et animalium hactenus detecta, caracteribus essentialibus donata, in tribus divisa, subinde ad leges naturae. 506 pp. Wolfgang Gerle, Prague. Crossref
Skutschas, P.P. 2013. Mesozoic salamanders and albanerpetontids of Middle Asia, Kazakhstan, and Siberia. In: J.D. Gardner and R.L. Nydam (eds.), Mesozoic and Cenozoic Lissamphibian and Squamate Assemblages of Laurasia. Palaeobiodiversity and Palaeoenvironments 93: 441–457.
Skutschas, P.P. 2016a. A new crown-group salamander from the Middle Jurassic of Western Siberia, Russia. Palaeobiodiversity and Palaeoenvironments 96: 41–48. Crossref
Skutschas, P.P. 2016b. A relict stem salamander: Evidence from the Early Cretaceous of Siberia. Acta Palaeontologica Polonica 61: 119–123.
Skutschas, P.P. and Krasnolutskii, S.A. 2011. A new genus and species of basal salamanders from the Middle Jurassic of Western Siberia, Russia. Proceedings of the Zoological Institute RAS 315: 167–175.
Skutschas, P.P. and Martin, T. 2011. Cranial anatomy of the stem salamander Kokartus honorarius (Amphibia: Caudata) from the Middle Jurassic of Kyrgyzstan. Zoological Journal of the Linnean Society 161: 816–838. Crossref
Tennant, J.P., Mannion, P.D., Upchurch, P., Sutton, M.D., and Price, G.D. 2017. Biotic and environmental dynamics through the Late Jurassic–Early Cretaceous transition: evidence for protracted faunal and ecological turnover. Biological Reviews 92: 776–814. Crossref
Wiechmann, M.F. 2000. The albanerpetontids from the Guimarota mine. In: T. Martin and B. Krebs (eds.), Guimarota: a Jurassic Ecosystem, 51–54. Verlag Dr. Friedrich, Pfeil.
Acta Palaeontol. Pol. 63 (3): 519–525, 2018
https://doi.org/10.4202/app.00498.2018