
A new occurrence of the Late Triassic archosaur Smok in southern Poland
GRZEGORZ NIEDŹWIEDZKI and EWA BUDZISZEWSKA-KARWOWSKA
Niedźwiedzki, G. and Budziszewska-Karwowska, E. 2018. A new occurrence of the Late Triassic archosaur Smok in southern Poland. Acta Palaeontologica Polonica 68 (4): 703–712.
Two isolated teeth, a dorsal vertebra, fragments of a humerus and femur, a fragmentary pubic “boot” and part of an ischium shaft, identified here as belonging to a large predatory archosaur were discovered in the Upper Triassic site at Marciszów near Zawiercie (southern Poland). Comparisons of the new fossils from Marciszów with the dorsal vertebrae, pubic “boot”, ischium and femur of the theropod-like Smok wawelski from Lisowice (Silesia) reveal that the two taxa are very similar. Nevertheless, due to the lack of more diagnostic elements (e.g., braincase or cranial elements), we prefer to consider all described specimens from Marciszów as Smok sp. Smok sp. shares a low mound-like, anterior trochanter with trochanteric shelf on the femur, a massive pubic “boot” with a distinct depression (= bevelled area), and a transversely lenticular ischium shaft in cross-section with S. wawelski. Some observed characters of the dorsal vertebra (e.g., lack of some lamina, shape and position of zygapophyses), however, are different from S. wawelski and may also suggest that the new findings represent a second species of the genus in the Upper Triassic of Poland. The discovery of Smok sp. at Marciszów is significant because it is the second example of the co-occurrence of this genus with: (i) bones of a large dicynodont; and (ii) record of gnawed tetrapod bones. The discovery of Smok sp. and the lack of significant morphologic divergence from S. wawelski suggest that this taxon is the only large-bodied predator currently known from the Upper Triassic of Poland. This new evidence expands the record of the genus and contributes, in some measure, to our knowledge of the stratigraphical distribution of large predatory archosaurs from the Polish Upper Triassic bone-bearing levels.
Key words: Diapsida, Archosauromorpha, Archosauria, Late Triassic, Poland, Silesia.
Grzegorz Niedźwiedzki [grzegorz.niedzwiedzki@ebc.uu.se], Subdepartment of Evolution and Development, Department of Organismal Biology, Uppsala University, Norbyvägen 18A, SE-752-36, Uppsala, Sweden.
Ewa Budziszewska-Karwowska [ebudzisz@wnoz.us.edu.pl], Museum of Earth Sciences, Faculty of Earth Sciences, University of Silesia, Będzińska 60 Street, PL-41-200 Sosnowiec, Poland.
Received 18 May 2018, accepted 31 October 2018, available online 29 November 2018.
Copyright © 2018 G. Niedźwiedzki nad E. Budziszewska-Karwowska. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
During the Late Triassic two groups of archosaurs, crocodile-line representatives called “rauisuchians” and theropod dinosaurs, filled the large-bodied predator niche in terrestrial ecosystems of Pangea (e.g., Gower 2000; Brusatte et al. 2010; Nesbitt et al. 2013). Two well-known Late Triassic tetrapod faunas from Poland demonstrate that these animals were relatively rare in their ecosystems of the region (e.g., Dzik and Sulej 2007; Dzik et al. 2008). Recent findings have revealed a range of size, dietary, and body plan diversity in the Middle–Late Triassic predatory archosaurs (e.g., Nesbitt 2003, 2011; Nesbitt et al. 2013; Zanno et al. 2015), including several forms strikingly convergent with dinosaurs (Nesbitt and Norell 2006; Nesbitt 2007). Therefore, such discoveries of dinosaur-like predatory archosaurs still arouse interest. Especially, findings of large isolated teeth of predators are not so rare as cranial or postcranial bones. Many discoveries of isolated large serrated teeth are also known from the Middle–Upper Triassic deposits of southern Poland (Kuhn 1965; Surmik and Brachaniec 2013; Skawiński et al. 2017) and among them is one of the first published remains of an archosaur from Triassic of Poland (“Megalosaurus” cloacinus Quenstedt, 1858) which represents a well-serrated and mediolaterally compressed large predator tooth (Roemer 1870: pl. 15: 5; see also Skawiński et al. 2017).
Vertebrate remains from the Upper Triassic of southern Poland have been known since the 19th century. During the last 20 years, this area has yielded new palaeontological sites with mass accumulation of fossil plants and animals (e.g., Dzik 2001; Dzik and Sulej 2007; Dzik et al. 2008; Sulej et al. 2011a, b, 2012; Niedźwiedzki et al. 2014). Most of the vertebrate fossils are attributed to typical Triassic tetrapods and are represented mainly by isolated bones and fragmentary preserved skeletons of temnospondyls, archosaurs and dicynodonts. The largest known predator from the Late Triassic faunas of Poland is the theropod-like archosaur Smok wawelski Niedźwiedzki, Sulej, and Dzik, 2012. Numerous bones (of at least three individuals) of this taxon are known from the Upper Triassic strata of the Lipie Śląskie clay-pit at Lisowice (Dzik et al. 2008; Niedźwiedzki et al. 2012; Niedźwiedzki 2013).
Between 2009 and 2012, the Museum of Earth Sciences, Faculty of Earth Sciences, University of Silesia (Sosnowiec), collected fossil vertebrates (Budziszewska-Karwowska et al. 2010) from the Marciszów near Zawiercie, south-western Poland (Fig. 1). Here we describe an isolated teeth and fragments of postcranial bones of a large predator archosaur collected at this site, which adds new facts to the discussion about the composition of the Keuper tetrapod faunas of Europe and Poland.
Two teeth, a dorsal vertebra, a fragment of a humerus, a fragment of a femur, part of an ischium shaft, and a fragmentary pubic “boot” were identified from the collected bone material as belonging to Smok sp. (Figs. 2–4). Preliminary comparisons of the femur and dorsal vertebrae of S. wawelski with the new fossil remains from Marciszów reveal that the two taxa are very similar and perhaps even represent the same species (Niedźwiedzki and Budziszewska-Karwowska 2015). It is the second record of this genus in Poland and according to first author (GN) of this report (see Niedźwiedzki et al. 2014), the Smok bone-bearing level (Lisowice and Marciszów sites) postdates all other vertebrate discoveries from the Upper Triassic of Poland (e.g., Poręba, Krasiejów, and Woźniki vertebrate faunas). The second author (EBK) follows the stratigraphical concept of Szulc et al. (2015; compare Szulc et al. 2006; Budziszewska-Karwowska et al. 2010), i.e., that the Marciszów locality correlates with Lipie Śląskie, Poręba, and Woźniki sites, as an element of mid-Norian Lisowice bone-bearing level.
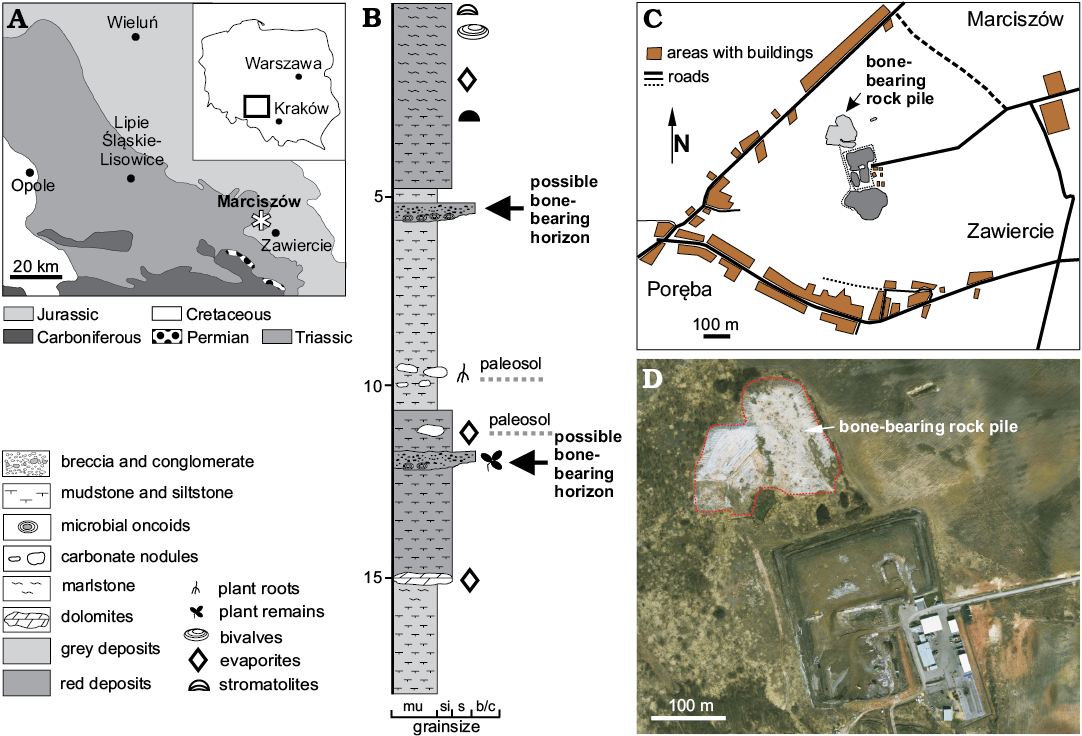
Fig. 1. A. Geological map of the Silesia showing location of the Marciszów site (asterisk), where the Smok sp. fossil remains were discovered (modified from Niedźwiedzki et al. 2014). B. The schematic section of the Marciszów site; arrows indicate two horizons suspected as being provenance of the bones; (modified from Budziszewska-Karwowska et al. 2010). C, D. Maps (C, simplified plan of Marciszów; D, satellite map of the area from Google Earth®) showing position of the bone-bearing rock pile. Abbreviations: b/c, breccia and conglomerate; mu, mudstone; s, sandstone; si, siltstone.
A recently published results (Sulej et al. 2018, in press) suggest strong similarity of the Woźniki site fauna to that from the Krasiejów site (see Dzik and Sulej 2007, 2016) assigned to the early Adamanian (late Carnian in age) land-vertebrate faunachron (Lucas 2010, 2015). The faunal assemblage from Woźniki is similar to Krasiejów in the presence of a spinicaudatan (Laxitextella), a capitosaur (Cyclotosaurus sp.), a diapsids Ozimek sp., a silesaurid dinosauriform, single-rooted postcanine teeth of a non-mammaliaform eucynodont, rich phytosaur remains (Phytosauria indet.), and the lack of more advanced dinosaurian taxa, which are present in the middle–late Norian/earliest Rhaetian of Poland (see Dzik et al. 2008; Sulej et al. 2012; Niedźwiedzki et al. 2012, 2014; Skawiński et al. 2017).
The discovery of Smok remains at Marciszów is also significant because it is the second example of the co-occurrence of this genus with a large and tooth-less dicynodont and a unique record of gnawed bones (Budziszewska-Karwowska et al. 2010; Niedźwiedzki et al. 2011, 2012).
The Marciszów site is relatively rich in fossils, with variegated terrestrial strata that preserve microbial structures (Szulc et al. 2015), plant remains (palynomorphs, charcoal and wood remains; Fijałkowska-Mader et al. 2015; Kubik et al. 2015; Philippe et al. 2015), invertebrates (freshwater bivalves; Skawina and Dzik 2011; ostracods and conchostracans; GN personal observations), vertebrate remains including sharks, actinopterygians, dipnoan fish, large dicynodonts (Budziszewska-Karwowska et al. 2010; Szulc et al. 2015; GN personal observations) and tetrapod trace fossils (Sadlok and Wawrzyniak 2013). This site is especially interesting because it yields a biota which is very similar to that described from the Lipie Śląskie clay-pit at Lisowice (Dzik et al. 2008; Niedźwiedzki et al. 2011, 2012; Niedźwiedzki 2013; Pieńkowski et al. 2014; Tałanda et al. 2017) and shows a mixture of two different environments (terrestrial and aquatic). Together with the large collection from an exceptionally fossil-rich locality at Lisowice, the Marciszów fossil collection stands out as one of the most important from the Polish Upper Triassic because it contains a unique record of the tetrapod megafauna which is still unknown in other parts of the Keuper strata of the Germanic Basin (Dzik et al. 2008).
The present paper is the first documentation of the small collection of the large archosaur fossils collected from the site and supplements the data on the Upper Triassic tetrapod faunas from southern Poland. The purpose of this paper is to describe the new Smok material, which is clearly distinct from all known Triassic archosaurs. Due to the fragmentary condition of some bones, we focus the description mainly on the dorsal vertebra and the partially preserved femur. As this is a fragmentary and isolated material, we do not extensively discuss the phylogenetic position of the genus Smok here. This task will be the subject of the senior author’s monographic paper.
Geological background
The studied material comes from a construction site of a city dump at Marciszów in the western district of the Zawiercie town (Fig. 1A, C, D) and was collected on fieldworks carried out during 2009–2012. This locality was originally identified as a fossiliferous site with palustrine, fluvial and pedogenic facies of the so-called Woźniki Limestone in earlier geological investigations of the area (see Szulc et al. 2006; Szulc and Becker 2007). A recently redefined as the Woźniki (Limestone) Member and Patoka (Marly Mudstone-Sandstone) Member of the Grabowa Variegated Mudstone-Carbonate Formation (see Szulc et al. 2015). Additional information on the geology and fossil content of this locality has been published in several previous papers (Budziszewska-Karwowska et al. 2010; Skawina and Dzik 2011; Sadlok and Wawrzyniak 2013).
The Marciszów site is located in the easternmost part of the Upper Silesian basin, close to the margin of the Kraków-Częstochowa Upland with extensive outcrops of Middle and Upper Jurassic rocks (Fig. 1A). The subsurface Upper Triassic deposits in the Upper Silesia constitute a belt that spreads out between the cities of Olkusz to the east and Opole to the west (Szulc and Becker 2007). Numerous Upper Triassic sites with well-preserved vertebrate fossils were discovered in this part of the country (for review see Sulej et al. 2011a, b, 2012) and a few new occurrences are currently being investigated. These deposits are relatively rich in fossils and the first discoveries from the Upper Silesia dates from the nineteenth century (see Roemer 1870). According to Szulc et al. (2006, 2015), the Upper Triassic clastic and carbonate deposits exposed in the Zawiercie area represent the middle Norian as indicated by palynologic Corollina meyeriana IVb Subzone (Fijałkowska-Mader et al. 2015) and some basic lithostratigraphical correlation. These deposits are succeeding in some areas, in most cases discordantly, by the uppermost Triassic or Lower Jurassic continental clastic deposits.
The section that is temporarily exposed in Marciszów shows different facies types associated with terrestrial, and in some cases, typical freshwater environments. Three basic facies types have been recognized in the section (Szulc et al. 2006). The best exposed in this profile are fluvial facies represented by fine-grained clastics, mainly grey or greenish and red mudstone along with siltstone (Fig. 1B). In some horizons they show a record of postdepositional pedogenic processes. Some of the beds have an erosional lower bounding surface and contain poorly sorted conglomeratic material with oncoids, plant fragments (wood), charcoal accumulations, bivalves, and isolated vertebrate bones and teeth. These erosional bounded layers are crucial for bone burial and preservation at Marciszów (Budziszewska-Karwowska et al. 2010; Szulc et al. 2015). However, the major part of the bone fossils collected at Lisowice come from fine-grained mudstone and siltstone (see Dzik et al. 2008).
Some carbonate concretions from Marciszów often contain rich plant remains, vertebrate fossils, as well as miospores (e.g., Corollina meyeriana, Ovalipolis ovalis, Brachysaccus neomundanus, Enzonalasporites sp.; see Sadlok and Wawrzyniak 2013), which are well-known palynomorphotypes of the Polish Upper Triassic (see Orłowska-Zwolińska 1985; Fijałkowska-Mader et al. 2015). Unfortunately, the palynomorph assemblage recognized at this locality has not yet been studied in detail and palynologic analysis was based only on a single sample (Szulc et al. 2006). This leads to difficulties in understanding the rallies between sites and correlation between bone-bearing intervals.
The section in Marciszów, also comprises playa deposits with evaporites in the lowermost part and mudstone, siltstone with rhizoliths and layers with stromatolites, conglomerate in the middle and upper parts (Szulc et al. 2006: fig. 5, 2015: fig. 14). The middle and upper parts of the section are probably the equivalents age to the fossil-bearing levels recognized in the carbonaceous grey mudstone and siltstone exposed in the upper part of the section of Lipie Śląskie clay-pit at Lisowice (Dzik et al. 2008; Niedźwiedzki et al. 2012). A recent dating of a single zircon grain, recovered from the sandstone bed positioned below the upper fossil-bearing interval in Lisowice, yielded an absolute age of 211±3 Ma (Kowal-Linka et al. 2018). This means that the upper bone-bearing interval with a large dicynodont and S. wawelski bones must be younger than the zircon grain. This provides a maximum deposition age of the layers in which it was found. The boundary between the Norian and Rhaetian stages is currently defined at the age of ~208.5 Ma (Cohen et al. 2013, updated online version 2018-08); the zircon date thus indicates no older than late Norian age for the Lisowice type of fauna from Silesia.
The section at Marciszów site comprises two discrete conglomerate horizons (Fig. 1B) that could have yielded the described material (Szulc et al. 2006, 2015; Budziszewska-Karwowska et al. 2010; Sadlok and Wawrzyniak 2013). The conglomerate fragments from the rock pile contain the remains of microbial structures (oncoids), charcoal, plants (wood and leaves), freshwater bivalves (Szulc et al. 2006; Skawina and Dzik 2011), vertebrate bones (Budziszewska-Karwowska et al. 2010; Szulc et al. 2015) and preserved tetrapod traces (Sadlok and Wawrzyniak 2013). The condition (e.g., colour, pyrite mineralization, and character of infillings) of all studied bones indicates that these come from a very similar taphonomical environment and probably represent material from the same bone accumulation or layer. Bones show some sings of abrasion and they contain also characteristic fractures (mineralized cracks or deformations). This indicates that they were transported, abraded, and probably exposed above sediment before the final burial and fossilization. Associated with isolated teeth (WNoZ/S/7/192 and 198) are fragments of host rocks that suggest poorly sorted conglomeratic layers as the source of the bones and suggest a rather high energy depositional environment for the bone-bearing deposits.
Material and methods
All specimens described here were found isolated and have been located ex situ on an old pile of post-exploitation mixed sediment, which had been accumulated during 2007–2009. Unfortunately, at present there is not a single easily accessible outcrop at Marciszów, where these bone-bearing strata are exposed and there is no suitable exposure for additional stratigraphical or taphonomical studies (Sadlok and Wawrzyniak 2013). The first bone assigned to Smok sp., a fragment of a femur (WNoZ/S/7/160), was discovered in 2009 and the other elements were collected during fieldworks in 2011 (a mid-shaft of the humerus, WNoZ/S/7/163; a middle part of the left ischium shaft, WNoZ/S/7/168; a distal part of the right pubis, a fragmentary pubic “boot”, WNoZ/S/7/170) and 2012 (isolated teeth, WNoZ/S/7/192 and WNoZ/S/7/198; a mid-dorsal vertebra, WNoZ/S/7/199).
Institutional abbreviations.—WNoZ, Museum of Earth Sciences, Faculty of Earth Sciences, University of Silesia, Sosnowiec, Poland; ZPAL, Institute of Paleobiology, Polish Academy of Sciences, Warsaw, Poland.
Systematic palaeontology
Diapsida Osborn, 1903
Archosauromorpha Huene, 1946
Archosauria Cope, 1869
Genus Smok Niedźwiedzki, Sulej, and Dzik, 2012
Type species: Smok wawelski Niedźwiedzki, Sulej, and Dzik, 2012, Lipie Śląskie clay-pit at Lisowice, Silesia, Poland, Upper Triassic.
Smok sp.
Figs. 2, 3A, 4A, B, 5A.
Material.—Two isolated teeth (WNoZ/S/7/192 and 198); dorsal vertebra (WNoZ/S/7/199); fragment of a humerus (WNoZ/S/7/163); fragment of the femur (WNoZ/S/7/160); fragment of a pubic “boot”(WNoZ/S/7/170); fragment of an ischium shaft (WNoZ/S/7/163).
Description.—Teeth: A nearly complete tooth (45 mm in length, WNoZ/S/7/198; Fig. 2A) and a broken tooth (34 mm in length, WNoZ/S/7/192; Fig. 2B) were collected in two different locations on the Marciszów rock pile but are preserved in very similar conglomeratic and calcite-pyrite mineralized mudstone. Both tooth crowns are distally curved and have some transversal enamel wrinkles (see Brusatte et al. 2007; Niedźwiedzki 2013), which sweep dorsoventrally from the serrations on the distal margins of the labial and lingual surfaces. They are robust and similar in form to the teeth of other Triassic archosaurs, such as large pseudosuchians (e.g., Batrachotomus kupferzellensis, Postosuchus kirkpatricki), as well as theropod dinosaurs (e.g., Megalosaurus bucklandi, Allosaurus fragilis, Carcharodontosaurus saharicus) from the Jurassic and Cretaceous (Brusatte et al. 2007; Niedźwiedzki 2013). The tooth WNoZ/S/7/192 is round or oval-shaped in cross-section at its base but the apical portion is clearly flattened. The broken tooth WNoZ/S/7/198 is elongated, slightly posteriorly recurved, similarly serrated on both sides, and slightly laterally compressed in the upper part of the crown. The pattern and size of the serrations in WNoZ/S/7/192 and 198 are usually very similar to the premaxillary, maxillary, and dentary teeth of Smok wawelski. Both teeth from Marciszów show serrations on the anterior and posterior carinae. The denticle density of these teeth averages 12 (WNoZ/S/7/198) to 13 (WNoZ/S/7/192) denticles per 5 mm and is comparable to S. wawelski (14–20 per 5 mm), but is clearly somewhat lower (Niedźwiedzki 2013).
Dorsal vertebrae: The nearly complete vertebra (WNoZ/ S/7/199; Fig. 2D) shows a amphicoelous centrum with well-fused neural arch and well-defined vertebral laminae. The WNoZ/S/7/199 exhibits the typical morphology of an archosaur dorsal vertebra (= size of zygapophyses, presence of the vertebral laminae and hyposphene-hypantrum intervertebral articulations). There are shallow fossae on the centrum on both the left and right lateral sides. The neural arch is extensively penetrated by deep cavities. The neural spine is broken off near its base. The centrum is as wide as tall, with oval-shaped articular surfaces. There is no ridge on the midline on the ventral surface of the trunk vertebra. Both of the diapophyses are broken off at their bases. The neural arch is fused to the centrum but there is evidence of where a neurocentral suture was, which is slightly raised in some areas. WNoZ/S/7/199 has distinct laminae between the prezygapophyses, postzygapophyses and diapophyses. On the lateral sides, the specimen has: (i) anterior centrodiapophyseal lamina, which connect the diapophyses with the the neurocentral junction; (ii) posterior centrodiapophyseal laminae, which connect the diapophyses with the posterior portion of the neurocentral junction.
The zygadiapophyseal laminae are laterally thick and pronounced and connect the diapophyses and lateral edge. The specimen WNoZ/S/7/199 is interpreted as a mid-dorsal vertebra and the following characteristics indicate this: (i) size and position of diapophyses; (ii) positions of pre- and postzygapophyses. In addition, in Smok wawelski from Lisowice, cervical and anterior dorsal vertebrae were identified by the presence of the parapophyses on the anterior rim of the centrum, whereas the parapophyses of trunk vertebrae are only present on the neural arch (see Niedźwiedzki 2013).
Shaft of the humerus: One fragment of a humerus, specimen WNoZ/S/7/163, was collected (Fig. 2C). The preserved part of the shaft is relatively long and has an approximately oval transverse section with distal ends. The shaft is longitudinally straight and has thin-wall, about 2–3 mm thick. The described shaft is similar in size and shape to that observed from humerus of Smok wawelski (see Niedźwiedzki 2013).
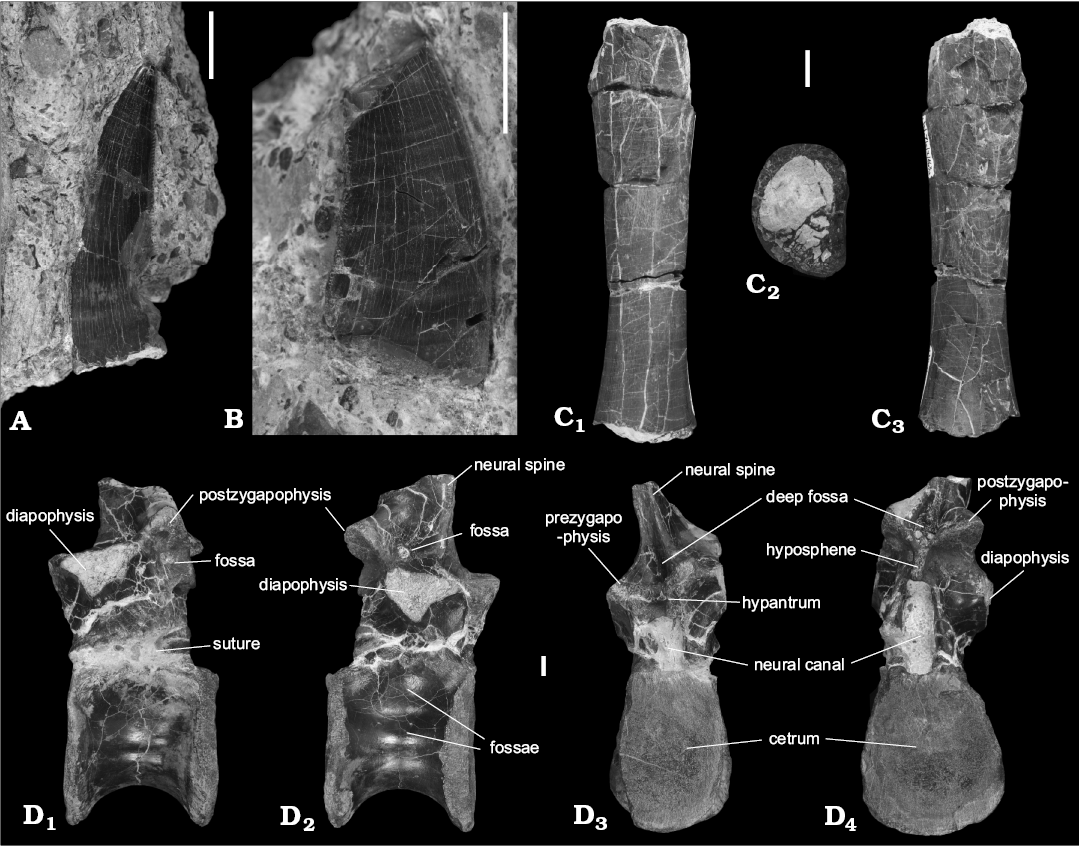
Fig. 2. Predatory archosaur Smok sp., Marciszów near Zawiercie, Poland, Late Triassic. A, B. Isolated teeth in lateral view (A, WNoZ/S/7/192; B, WNoZ/S/7/198). C. Mid-shaft of the humerus (WNoZ/S/7/163), in posterior (C1) and anterior (C2) views, distal cross section (C3). D. Mid-dorsal vertebra (WNoZ/S/7/199), in left (D1) and right (D2) lateral, anterior (D3), and posterior (D4) views. Scale bars 10 mm.
Fragment of femur: The preserved proximal fragment of the right femur, specimen WNoZ/S/7/160 (Fig. 3A), shows clear similarities to the S. wawelski femoral bones collected from Lisowice (Niedźwiedzki et al. 2012; Niedźwiedzki 2013). The proximal part of S. wawelski femur is massive, but with a slender shaft. Similarly, to S. wawelski femur, WNoZ/S/7/160 shows rather an indistinct neck separating the shaft and head. The cranial surface of the preserved part of bone, bears a mound-like muscle attachment, which is in the position of the anterior (= lesser) trochanter of many archosaurs (Ornithosuchidae and Dinosauriformes) (see Hutchinson 2001; Nesbitt 2011). There is also a discrete trochanteric shelf (= anterior trochanteric shelf) and scar linking originally the anterior trochanter with a ridge-like fourth trochanter, both structures are probably homologous with that of dinosauromorphs (see Novas 1992). The trochanteric shelf and scar extends caudally from the anterior trochanter, through the lateral surface of the femur. The same condition was observed on femur of S. wawelski (see Fig. 3B; Niedźwiedzki 2013).
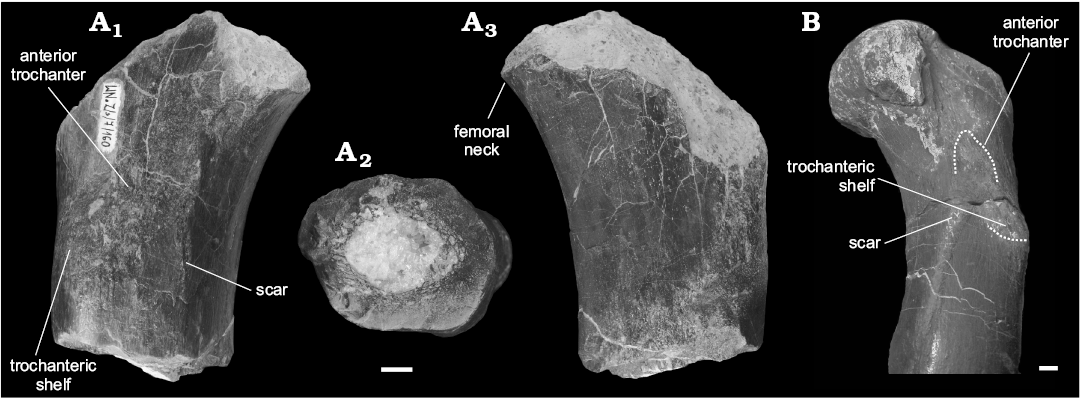
Fig. 3. Predatory archosaurs from Late Triassic of Poland, Smok sp., Marciszów near Zawiercie (A) and Smok wawelski Niedźwiedzki, Sulej, and Dzik, 2012, Lipie Śląskie clay-pit at Lisowice (B). A. Fragment of proximal region of the right femur (WNoZ/S/7/160), in anterior (A1) and posterior (A2) views, distal cross section (A3). B. Left femur (ZPAL V.33/45) in anteromedial view. Scale bars 10 mm.
Fragmentary pubic “boot”: Although not complete, the preserved fragment of a pubic “boot”, specimen WNoZ/S/ 7/170 (Fig. 4A) suggest that the distal part of pubis was relatively robust, but narrow, and laterally terminated by a large pubic “boot”. The shape and medial surface of the specimen WNoZ/S/7/170 suggests that conjoined pubes formed a long median symphysis. The ventral surface of the pubic “boot” is rugose and elongated in distal view. In the craniomedial region of the “boot” there is a distinct depression (= bevelled area) which is limited on the lateral surface of the bone by low ridge. This beveled area can be clearly observed in lateral and medial views (Fig. 4A1, A2). On the medial surface of the “boot” there is a shallow basin. This specimen (WNoZ/S/7/170) shows similarities to S. wawelski pubic “boot” (Fig. 4D, E) collected from the Smok level at Lisowice (Niedźwiedzki 2013).
Part of the ischium shaft: The collected fragment of an ischium shaft (Fig. 4B), specimen WNoZ/S/7/163, is lenticular in cross section and the preserved fragment is clearly straight in lateral view. In this respect, it resembles that of S. wawelski (Fig. 4C; see Niedźwiedzki 2013). The shaft of the ischium of S. wawelski being essentially rod-like in its mid and distal part, lenticular in cross section, clearly differs from the more plate-like ischia of “rauisuchians” (e.g., Batrachotomus kupferzellensis, Postosuchus kirkpatricki) and resembles the condition seen in basal dinosaurs (e.g., Herrerasaurus ischigualastensis).
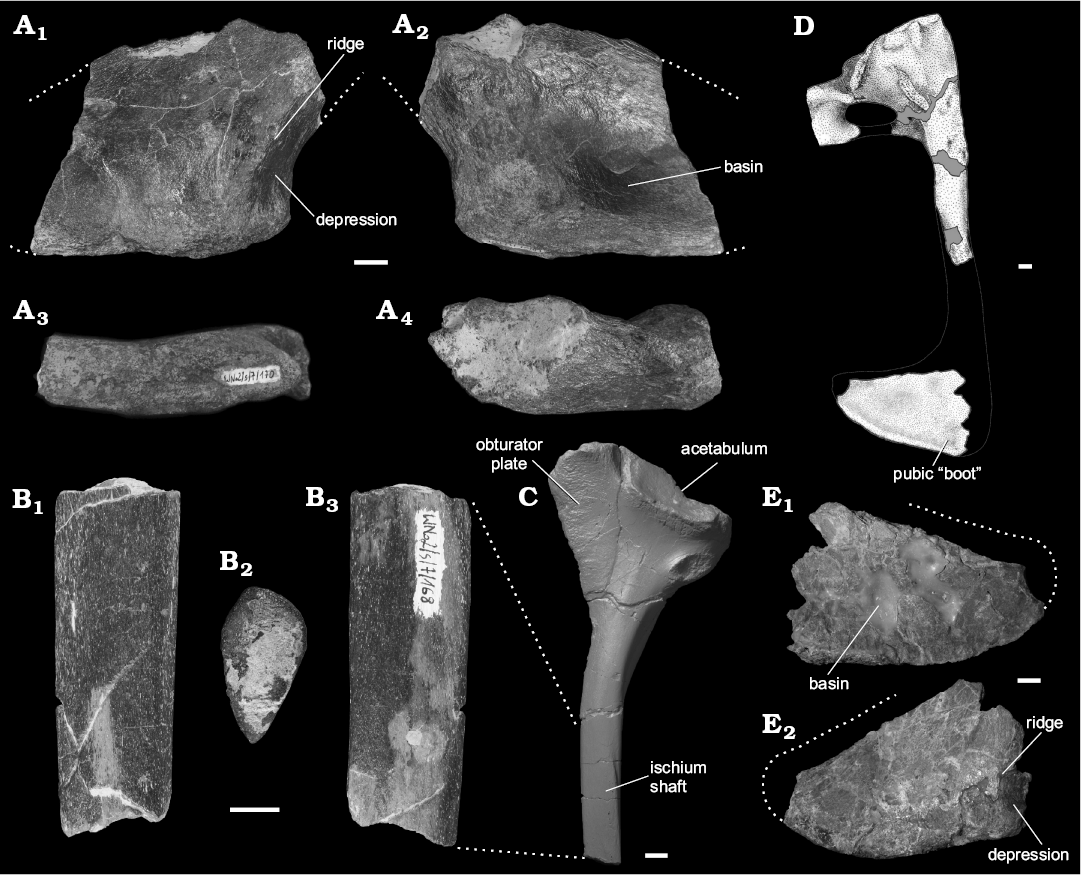
Fig. 4. Predatory archosaurs from Late Triassic of Poland, Smok sp., Marciszów near Zawiercie (A, B) and Smok wawelski Niedźwiedzki, Sulej, and Dzik, 2012, Lipie Śląskie clay-pit at Lisowice (C–E). A. Distal part of the right pubis, pubic “boot” (WNoZ/S/7/170), in lateral (A1), medial (A2), ventral (A3), and dorsal (A4) views. B. Middle part of the left ischium shaft (WNoZ/S/7/168), in lateral (B1), dorsal (B2), and medial (B3) views. C. Left ischium (ZPAL V.33/302), in lateral view. D. Reconstruction of right pubis in lateral view; based on specimens ZPAL V.33/311A, B and ZPAL V.33/298 (from Niedźwiedzki 2013). E. Distal part of the right pubis, pubic “boot” (ZPAL V.33/298), in medial (E1) and lateral (E2) views. Scale bars 10 mm.
Concluding remarks
The described isolated teeth and bone fragments were found at the same accumulation of rock material, where large bone (e.g., dicynodont bones) or bone fragments are generally very rare. Their sizes indicate that they belonged to individuals of similar size or even belong to the same individual. The collected skeletal elements include two teeth, a fragment of a humerus shaft, a fragment of the proximal part of femur, a part of an ischium shaft, a fragmentary pubic “boot”, and a nearly complete mid-dorsal vertebra. Overall, the preserved part of femur, the ischium shaft, and the mid-dorsal vertebra are from anatomical point of view very similar to that of Smok wawelski from Lisowice (Niedźwiedzki et al. 2012; Niedźwiedzki 2013). The most characteristic of these features are: (i) an indistinct neck separating the shaft and head of the femur, (ii) a mound-like anterior (= lesser) trochanter with trochanteric shelf, (iii) a rod-like ischium shaft (lenticular in cross section), (iv) a pubic “boot” with a distinct depression (= bevelled area), (v) a mid-dorsal vertebra with the shallow depressions (fossae) on centrum, (vi) a neural arch of the mid-dorsal with deep cavities and well developed laminae.
Some more or less important differences are apparent in the anatomy of the mid-dorsal vertebra (Fig. 5). The mid-dorsal vertebrae of S. wawelski (e.g., ZPAL V.33/42) show an additional lamina, which is located just below postzygapophyses. There are also differences in shape and position and of zygapophyses in dorsal vertebrae of S. wawelski and Smok sp. (Fig. 5).
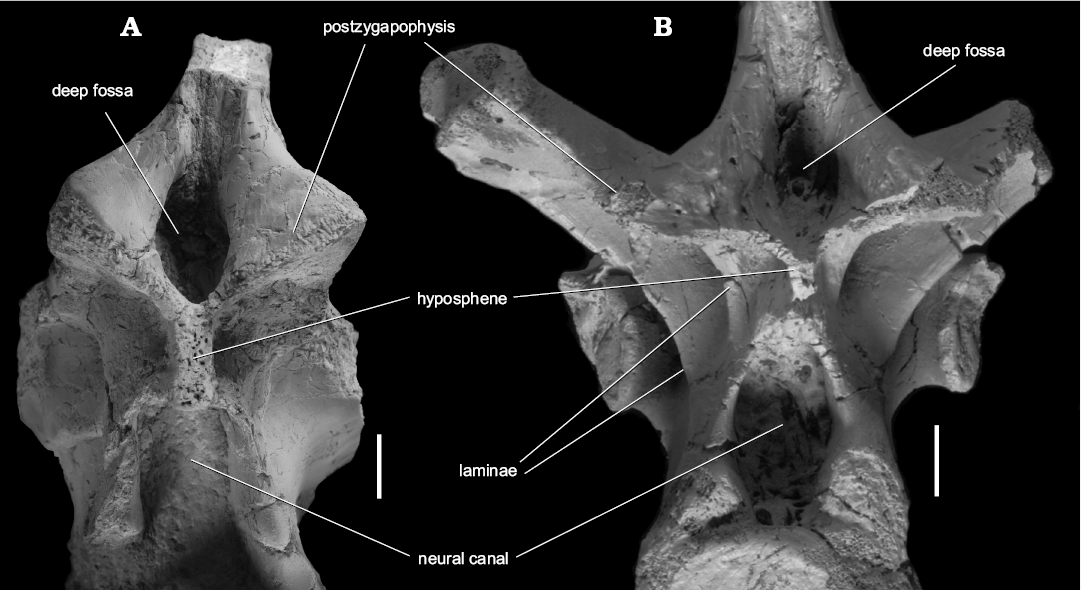
Fig. 5. Comparison and details of articular surface of the mid-dorsal vertebrae (in posterior view) of the predatory archosaurs from Late Triassic of Poland. A. Smok sp. (WNoZ/S/7/199), Marciszów near Zawiercie. B. Smok wawelski Niedźwiedzki, Sulej, and Dzik, 2012 (ZPAL V.33/42), Lipie Śląskie clay-pit at Lisowice. Scale bars 10 mm.
Although at this stage a new occurrence of the genus Smok from Marciszów has been separated from S. wawelski on a species level, it is possible that further finds may reveal more informative and diagnostic characters shared by the two taxa. The incomplete and fragmentary condition of bones from Marciszów prevents a reliable description and comparison. Without additional findings (e.g., braincase or cranial elements) and more completely preserved bones, it is impossible to state if they represent the same species as S. wawelski. Some aspects of anatomy recognized in Smok sp. from Marciszów are clearly different to that observed in postcranial material from the paratype of S. wawelski (Niedźwiedzki et al. 2012; Niedźwiedzki 2013). They may also represent anatomical features related to intraspecific variation. However, we cannot disregard that the new material of Smok sp. from the Marciszów might represent a new species which is morphologically very close to S. wawelski. This suggests a lineage persisting through the Late Triassic, with changes occurring in the organization of dorsal vertebrae.
Given our current knowledge, Smok is the only large-bodied predatory archosaur known that roamed the alluvial plains of southwestern Poland during the Late Triassic. The new finds of Smok specimens in the Upper Triassic adds to the growing record of large predatory archosaur fossils recovered in the Keuper strata of Europe. Previously described remains of large predators from this region include neotheropods such as Liliensternus liliensterni and “rauisuchians” like Batrachotomus kupferzellensis, Polonosuchus silesiacus, and Teratosaurus suevicus, but all these archosaurs were much smaller than Smok from Lisowice and Marciszów (Niedźwiedzki et al. 2012). On the basis of these discoveries, the genus Smok would have a wider palaeogeographic distribution across the Late Triassic biota of southern Poland.
New material of Smok sp. is consistent with placement of the Smok genus within the Archosauria incertae sedis. Additional material from Marciszów and a detailed description of the type material of S. wawelski from Lisowice (Niedźwiedzki 2013), as well as a more comprehensive analysis of its anatomical characters (on-going study), will help with diagnosis of the possible new species of Smok sp. discovered at Marciszów. So, the need for extensive sampling is required in order to recover more complete remains, which would provide a better understanding of the Marciszów occurrence.
Acknowledgements
Fieldwork in the Marciszów site (2010–2012) was organized by the Museum of Earth Sciences, Faculty of Earth Sciences, University of Silesia, Sosnowiec, Poland. We are grateful to the officials of Zawiercie for their logistic and financial support during the fieldworks. The fieldworks in Marciszów have been partially financed from Research Grant N N307 117037 to Grzegorz Racki. GN is currently funded by grant from the Swedish Research Council (2017-05248). We want to thank Maria Belén von Baczko and Marcel Lacerda whose comments significantly improved the final version of the paper.
References
Brusatte, S.L., Benson, R.B.J., Carr, T.D., Williamson, T.E., and Sereno, P.C. 2007. The systematic utility of theropod enamel wrinkles. Journal of Vertebrate Paleontology 27: 1052–1056. Crossref
Brusatte, S.L., Nesbitt, S.J., Irmis, R.B., Butler, R.J., Benton, M.J., and Norell, M.A. 2010. The origin and early radiation of dinosaurs. Earth-Science Reviews 101: 68–100. Crossref
Budziszewska-Karwowska, E., Bujok, A., and Sadlok, G., 2010. Bite marks on an Upper Triassic dicynodontid tibia from Zawiercie, Kraków-Częstochowa Upland, southern Poland. Palaios 25: 415–421. Crossref
Cohen, K.M., Finney, S.C., Gibbard, P.L., and Fan, J.-X. 2013. The ICS International Chronostratigraphic Chart. Episodes 36: 199–204. (updated version at http://www.stratigraphy.org/ICSchart/ChronostratChart2018-08.pdf)
Cope, E.D. 1869. Synopsis of the Extinct Batrachia, Reptilia and Aves of North America. Part I. Transactions of the American Philosophical Society, New Series 14: 1–104.
Dzik, J. 2001. A new Paleorhinus fauna in the early Late Triassic of Poland. Journal of Vertebrate Paleontology 21: 625–627. Crossref
Dzik, J. and Sulej, T. 2007. A review of the early Late Triassic Krasiejów biota from Silesia, Poland. Palaeontologia Polonica 64: 3–27.
Dzik, J. and Sulej, T. 2016. An early Late Triassic long-necked reptile with a bony pectoral shield and gracile appendages. Acta Palaeontologica Polonica 61: 805–823.
Dzik, J., Sulej, T., and Niedźwiedzki, G. 2008. A dicynodont-theropod association in the latest Triassic of Poland. Acta Palaeontologica Polonica 53: 733–738. Crossref
Fijałkowska-Mader, A., Heunisch, C., and Szulc, J., 2015. Keuper palynostratigraphy and palynofacies of the Upper Silesia (Southern Poland). Annales Societatis Geologorum Poloniae 84: 637–661.
Gower, D.J. 2000. Rauisuchian archosaurs (Reptilia, Diapsida): an overview. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 218: 447–488. Crossref
Huene, F. von 1946. Die grossen Stamme der Tetrapoden in den geologischen Zeiten. Biologische Zentralblat 65: 268–275.
Hutchinson, J.R. 2001. The evolution of femoral osteology and soft tissue on the line to extant birds (Neornithes). Zoological Journal of the Linnean Society 131: 169–197. Crossref
Kowal-Linka, M., Krzemińska, E., and Czupyt, Z. 2018. Najmłodsze detrytyczne cyrkony z utworów triasu z Lipia Śląskiego (Lisowic) koło Lublińca a wiek lisowickiego poziomu kościonośnego. In: M. Kędzierski and M. Gradziński (eds.), Polska Konferencja Sedymentologiczna POKOS 7, Góra Św. Anny, 4–7 czerwca 2018 r., Materiały konferencyjne, 77. Polskie Towarzystwo Geologiczne, Krakow.
Kubik, R., Uhl, D., and Marynowski, L. 2015. Evidence of wildfires during deposition of the upper Silesian Keuper succession, southern Poland. Annales Societatis Geologorum Poloniae 84: 685–696. Crossref
Kuhn, O. 1965. Saurischia. Fossilium Catalogus 109: 1–94.
Lucas, S.G. 2010. The Triassic timescale based on nonmarine tetrapod biostratigraphy and biochronology. In: S.G. Lucas (ed.), The Triassic Timescale. Special Publication of the Geological Society, London 334: 447–500. Crossref
Lucas, S.G. 2015. Age and correlation of Late Triassic tetrapods from southern Poland. Annales Societatis Geologorum Poloniae 85: 627–635. Crossref
Nesbitt, S.J. 2003. Arizonasaurus and its implications for archosaur divergences. Proceedings of the Royal Society of London, Series B 270 (Supplement 2): S234–S237. Crossref
Nesbitt, S.J. 2007. The anatomy of Effigia okeeffeae (Archosauria, Suchia), theropod convergence, and the distribution of related taxa. Bulletin of the American Museum of Natural History 302: 1–84. Crossref
Nesbitt, S.J. 2011. The early evolution of archosaurs: relationships and the origin of major clades. Bulletin of American Museum of Natural History 352: 1–292.Nesbitt, S.T. and Norell, M. 2006. CrossrefExtreme convergence in the body plans of an early suchian (Archosauria) and ornithomimid dinosaurs (Theropoda). Proceedings of the Royal Society of London , Series B 273: 1045–1048.
Nesbitt, S.J. and Norell, M.A. 2006. Extreme convergence in the body plans of an early suchian (Archosauria) and ornithomimid dinosaurs (Theropoda). Proceedings of the Royal Society Series B 273: 1045–1048. Crossref
Nesbitt, S.T., Brusatte, S.L., Desojo, J.B., Liparini, A. De Franca, M.A.G., Weinbaum, J.C., and Gower, D. 2013. Rauisuchia. In: S.J. Nesbitt, J.B. Desojo, and R.B. Irmis (eds.), Anatomy, Phylogeny and Palaeobiology of Early Archosaurs and their Kin. The Geological Society, London, Special Publications 379: 241–274.
Niedźwiedzki, G. 2013. The large predatory archosaur Smok wawelski from the latest Triassic of Poland. PhD Thesis, Faculty of Biology, University of Warsaw.
Niedźwiedzki, G. and Budziszewska-Karwowska, E. 2015. Smok z Zawiercia-Marciszowa – wstępne dane. In: Program Konferencji – Śląskie Dinozaury (nie tylko) z Krasiejowa. V Sesja Popularnonaukowa. Instytut Nauk Geologicznych PAN, Kraków.
Niedźwiedzki, G., Brusatte, S.L., Sulej, T., and Butler, R.J. 2014. Basal dinosauriform and theropod dinosaurs from the middle–late Norian (Late Triassic) of Poland: implications for Triassic dinosaur evolution and distribution. Palaeontology 57: 1121–1142. Crossref
Niedźwiedzki, G., Gorzelak, P., and Sulej T. 2011. Bite traces on dicynodont bones and the early evolution of large terrestrial predators. Lethaia 44: 87–92. Crossref
Niedźwiedzki, G., Sulej, T., and Dzik, J. 2012. A large predatory archosaur from the Late Triassic of Poland. Acta Palaeontologica Polonica 57: 267–276. Crossref
Novas, F.E. 1992. Phylogenetic relationships of the basal dinosaurs, the Herrerasauridae. Palaeontology 35: 51–62.
Orłowska-Zwolińska, T. 1985. Palynological zones of the Polish epicontinental Triassic. Bulletin de l’Academie Polonaise des Sciences, Serie des Sciences de la Terre 33: 107–119.
Osborn, H.F. 1903. On the primary division of the Reptilia into two sub-classes, Synapsida and Diapsida. Science 17: 275–276. Crossref
Pieńkowski, G., Niedźwiedzki, G., and Brański, P. 2014. Climatic reversals related to the Central Atlantic magmatic province caused the end-Triassic biotic crisis—evidence from continental strata in Poland. In: G. Keller and A. Kerr (eds.), Volcanism, Impacts, and Mass Extinctions: Causes and Effects. Geological Society of America Special Paper 505: 263–286. Crossref
Philippe, M., Pacyna, G., Wawrzyniak, Z., Barbacka, M., Boka, K., Filipiak, P., Marynowski, L., Thévenard, F., and Uhl, D. 2015. News from an old wood—Agathoxylon keuperianum (Unger) nov. comb. in the Keuper of Poland and France. Review of Palaeobotany and Palynology 221: 83–91. Crossref
Quenstedt, F.A. 1858. Der Jura. 842 pp. H. Laupp’schen, Tübingen.
Roemer, F. 1870. Geologie von Oberschlesien. 587 pp. Druck von Robert Nischkowsky, Breslau.
Sadlok, G. and Wawrzyniak, Z. 2013. Upper Triassic Vertebrate tracks from Kraków-Częstochowa Upland, Southern Poland. Annales Societatis Geologorum Poloniae 83: 105–111.
Skawina, A. and Dzik, J. 2011. Umbonal musculature and relationships of the Late Triassic filibranch unionoid bivalves. Zoological Journal of the Linnean Society 163: 863–883. Crossref
Skawiński, T., Ziegler, M., Czepiński, Ł., Szermański, M., Surmik, D., Tałanda, M., and Niedźwiedzki, G. 2017. A re-evaluation of the historical “dinosaur” remains from the Middle–Upper Triassic of Poland. Historical Biology 29: 442–472. Crossref
Sulej, T., Bronowicz, R., Tałanda, M., and Niedźwiedzki, G. 2011a. A new dicynodont-archosaur assemblage from the Late Triassic (Carnian) of Poland. Proceedings of the Royal Society of Edinburg 101: 261–269.
Sulej, T., Niedźwiedzki, G., and Bronowicz, R. 2012. A new Late Triassic vertebrate fauna with turtles, aetosaurs and coelophysoid dinosaurs from Poland. Journal of Vertebrate Paleontology 32: 1033–1041. Crossref
Sulej, T., Niedźwiedzki, G., Niedźwiedzki, R., Surmik, D., and Stachacz, M. 2011b. Nowy zespół kręgowców z marginalno-morskich i lądowych osadów dolnego kajpru (ladyn, środkowy trias) z Miedar na Śląsku. Przegląd Geologiczny 59: 426–430.
Sulej, T., Niedźwiedzki, G., Tałanda, M., Dróżdż, D., and Hara, E. (in press). A new early Late Triassic non-mammaliaform eucynodont from Poland. Historical Biology. Crossref
Surmik, D. and Brachaniec, T. 2013. The large superpredators’ teeth from Middle Triassic of Poland. Contemporary Trends in Geoscience 2: 91–94. Crossref
Szulc, J. and Becker, A. (eds.) 2007. Pan-European Correlation of the Epicontinental Triassic 4th Meeting. International Workshop on the Triassic of Southern Poland, September 3–8, 2007. 88 pp. Polish Geological Society, Polish Geological Institute, Institute of Geological Sciences, Jagiellonian University, Cracow.
Szulc, J., Gradziński, M., Lewandowska, A., and Heunisch, C. 2006. The Upper Triassic crenogenic lime stones in Upper Silesia (southern Poland) and their paleoenvironmental context. In: A.M. Alonso-Zarza and L.H. Tanner (ed.), Paleoenvironmental record and applications of calcretes and palustrine carbonates. Geological Society of America Special Papers 416: 133–151. Crossref
Szulc, J., Racki, G., Jewuła, K., and Środoń, J. 2015. How many Upper Triassic bone-bearing levels are there in Upper Silesia (southern Poland)? A critical overview of stratigraphy and facies. Annales Societatis Geologorum Poloniae 85: 587–626. Crossref
Tałanda, M., Bajdek, P., Niedźwiedzki, G., and Sulej, T. 2017. Upper Triassic freshwater oncoids from Silesia (southern Poland) and their microfossil biota. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 284: 43–56. Crossref
Zanno, L.E., Drymala, S., Nesbitt, S.J., and Scheider, V.P. 2015. Early crocodylomorph increases top tier predator diversity during rise of dinosaurs. Scientific Reports 5: 9276. Crossref
Acta Palaeontol. Pol. 68 (4): 703–712, 2018
https://doi.org/10.4202/app.00505.2018