
The skull anatomy and cranial endocast of the pseudosuchid archosaur Prestosuchus chiniquensis from the Triassic of Brazil
BIANCA MARTINS MASTRANTONIO, MARÍA BELÉN VON BACZKO, JULIA BRENDA DESOJO, and CESAR L. SCHULTZ
Mastrantonio, B.M., Baczko, M.B. von, Desojo, J.B., and Schultz, C.L. 2019. The skull anatomy and cranial endocast of the pseudosuchid archosaur Prestosuchus chiniquensis from the Triassic of Brazil. Acta Palaeontologica Polonica 64 (1): 171–198.
Prestosuchus chiniquensis is the most famous “rauisuchian” described by Friedrich von Huene, eight decades ago, and several specimens have been assigned to this taxon since then. In the present contribution, we provide the first detailed description of a complete and very well preserved skull (including the braincase) assigned to Prestosuchus chiniquensis from the Dinodontosaurus Assemblage Zone of the Santa Maria Supersequence of southern Brazil. The detailed description of the skull of Prestosuchus chiniquensis, besides increasing the knowledge about this taxon, may help elucidate the taxonomic relationships of pseudosuchians even further, since most of the characters used in phylogenetic analyzes are cranial. The presence of the subnarial fenestra, a controvertial extra opening on the skull of “rauisuchians”, is thoroughly discussed considering the evidence provided by this new specimen. We consider that the small slit-opening between the premaxilla and the maxilla in Prestosuchus chiniquensis, can not safely be considered a true fenestra, but indicates more likely the existence of some degree of cranial kinesis between these elements which can result in different relative positions of the bones after definitive burial and fossilization, so that the size and shape of this opening is taphonomically controlled. Complementarily, the digital reconstruction of its cranial endocast was developed both from the observation of the preserved braincase and from CT scan images, which resulted in obtaining the first endocast known for a “rauisuchian”. The endocast of Prestosuchus chiniquensis exhibited some remarkable convergences with that of theropod dinosaurs, which could be a reflection of the similar niches they occupied, since “rauisuchians” were the top predators at the end of the Late Triassic, before the extinction of all non-crocodylomorph pseudosuchians.
Key words: Archosauria, Pseudosuchia, Rauisuchia, Loricata, neuroanatomy, Triassic, Ladinian, Carnian, Brazil.
Bianca M. Mastrantonio [bmmastrantonio@gmail.com] and Cesar L. Schultz [cesar.schultz@ufrgs.br], Laboratório de Paleovertebrados, Departamento de Paleontologia e Estratigrafia, Instituto de Geociências, Universidade Federal do Rio Grande do Sul (UFRGS), Cx. P. 15001, 91501-970, Porto Alegre, RS, Brazil.
María B. von Baczko [belenvonbaczko@gmail.com] and Julia B. Desojo [julideso@fcnym.unlp.edu.ar], Division Paleontología de Vertebrados, Museo de La Plata, Paseo del Bosque s/n, B1900FWA, La Plata, Argentina; Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET), Avenida Rivadavia 1917, C1033AAJ, Ciudad Autónoma de Buenos Aires, Argentina.
Received 17 July 2018, accepted 31 October 2018, available online 24 January 2019.
Copyright © 2019 B.M. Mastrantonio et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
The Triassic period was a crucial time in the evolution and diversification of the major archosaur lineages. Among the crocodilian-line archosaurs (Pseudosuchia), the ones traditionally considered as “rauisuchians”, (sensu Nesbitt and Desojo 2017) represents one of the most intriguing and yet enigmatic groups (e.g., Gower 2000; Lautenschlager and Rauhut 2015; Nesbitt et al. 2013). Among these, the subgroup that includes Prestosuchus chiniquensis is the so-called basal Loricata (sensu Nesbitt and Desojo 2017; i.e., excluding crocodylomorpha), so that only the taxa belonging to this group will be used here for morphological comparisons. They were the dominant quadrupedal predators and had a near cosmopolitan distribution during the Early to Late Triassic Period, prior to the dinosaur radiation (e.g., Gower 2000).
The South American records of basal Loricata include several species from Brazil and Argentina. In the latter, the following species have been described: Luperosuchus fractus Romer, 1971 from the Chañares Formation (late Middle–early Late Triassic) (Romer 1971; Desojo and Arcucci 2009; Nesbitt and Desojo 2017); Saurosuchus galilei Reig, 1959 (Sill 1974; Alcober 2000; Trotteyn et al. 2011) and Sillosuchus longicervix Alcober and Parrish, 1997 from the Ischigualasto Formation (Late Triassic); and Fasolasuchus tenax Bonaparte, 1981 from the Los Colorados Formation (Late Triassic) (Desojo and Ezcurra 2016).
In Brazil, four taxa are found, all of them in the Santa Maria Supersequence from Rio Grande do Sul State. The valid species found are: Prestosuchus chiniquensis Huene, 1938 and Decuriasuchus quartacolonia França, Langer and Ferigolo, 2011 from the Pinheiros-Chiniquá Sequence, Dinodontosaurus Assemblage Zone, Middle Triassic; Dagasuchus santacruzensis Lacerda, Schultz, and Bertoni-Machado, 2015 from the Santa Cruz Sequence, Santacruzodon Assemblage Zone, Middle Triassic; and Rauisuchus tiradentes Huene, 1938 from the Candelária Sequence, Hyperodapedon Assemblage Zone, Late Triassic. In the last decade, most of these taxa and new specimens (e.g., CPEZ-239b by Lacerda et al. 2016) were described and restudied, as well as included in updated phylogenetic contexts (França et al. 2011, 2013; Mastrantonio et al. 2013; Lacerda et al. 2015, 2016; Lautenschlager and Rauhut 2015).
Among basal Loricata (sensu Nesbitt and Desojo 2017), detailed skull descriptions have been made for Batrachotomus kupferzellensis (Gower 1999), Postosuchus kirkpatricki (Long and Murry 1995; Weinbaum 2011), Polonosuchus silesiacus (Sulej 2005; Brusatte et al. 2009), Luperosuchus fractus (Nesbitt and Desojo 2017), Saurosuchus galilei (Alcober 2000), Prestosuchus chiniquensis (Barberena 1978; Lacerda et al. 2016), Decuriasuchus quartacolonia (França et al. 2011, 2013), and Rauisuchus tiradentes (Lautenschlager and Rauhut 2015). However, only the braincases of two of these taxa were analyzed in detail, Batrachotomus kupferzellensis (Gower 2002) and Prestosuchus chiniquensis (Mastrantonio et al. 2013), but the cranial endocast anatomy is still unknown for many basal loricata.
Prestosuchus chiniquensis is represented by 10 specimens, which consist of skulls and postcranial material assigned by different authors to this species (Huene 1938; Barberena 1978; Bonaparte 1984; Parrish 1993; Nesbitt 2011; Mastrantonio et al. 2013; Lacerda et al. 2016; Roberto-da-Silva et al. 2016). The knowledge about the paleoneurology of pseudosuchians is scarce, only known from phytosaurs (Lautenschlager and Butler 2016), ornithosuchids (Baczko and Desojo 2016), erpetosuchids (Nesbitt et al. 2018), aetosaurs (Baczko et al. 2018), and crocodylomorphs (Kley et al. 2010; Bona et al. 2017). When comparing this to the information available about a wide variety of avemetatarsalians, such as pterosaurs (Witmer et al. 2003; Codorniú et al. 2016), theropods (Franzosa and Rowe 2005; Witmer and Ridgely 2009; Paulina-Carabajal and Canale 2010; Paulina-Carabajal and Currie 2012), sauropodomorphs (Paulina-Carabajal 2015, Bronzati et al. 2017), and ornithischians (Hopson 1979; Giffin 1989; Witmer and Ridgely 2008; Evans et al. 2009; Miyashita et al. 2011; Cruzado-Caballero et al. 2015), the shortage of data on pseudosuchian paleoneuroanatomy is quite evident.
The main objective of this contribution is to provide a detailed description of the skull of a new specimen of Prestosuchus chiniquensis Huene, 1938 and to present the first description of a rauisuchian cranial endocast. This study was based on a well preserved skull (UFRGS-PV-0629-T) and complemented with first hand observations of other reference specimens: UFRGS-PV-0156-T, previously described by Barberena (1978) and CPEZ-239b, recently described by Lacerda et al. (2016). Besides providing data on the early evolution of the pseudosuchian skull, this comparative description also aims to improve the knowledge of the osteology of rauisuchians and to increase the data available for future phylogenetic analyses of the crown group Archosauria.
Institutional abbreviations.—BSPG AS, Bayerische Staatssammlung für Paläontologie und Geologie, Munich, Germany; CPEZ, Paleontology Collection of the Museu Paleontológico e Arqueológico Walter Ilha, São Pedro do Sul, Brazil; MCP, Paleontology Collection of the Museu de Ciências e Tecnologia of the Pontifícia Universidade Católica do Rio Grande do Sul, Porto Alegre, Brazil; MCZ, Museum of Comparative Zoology, Harvard University, Cambridge, USA; PULR, Paleontología, Universidad Nacional de La Rioja, Argentina; PVL, Paleontología de Vertebrados, Instituto Miguel Lillo, Tucumán, Argentina; PVSJ, División de Paleontología de Vertebrados del Museo de Ciencias Naturales y Universidad Nacional de San Juan, Argentina; SMNS, Staatliches Museum für Naturkunde, Stuttgart, Germany; UFRGS-PV, Paleovertebrate Collection of the Laboratório de Paleovertebrados of the Universidade Federal do Rio Grande do Sul, Porto Alegre, Brazil; ULBRA-PVT, Paleovertebrate Collection of the Universidade Luterana do Brasil, Canoas, Rio Grande do Sul, Brazil.
Geological setting
All the material analyzed belongs to the Dinodontosaurus Assemblage Zone of the Pinheiros-Chiniquá Sequence, Santa Maria Supersequence (Horn et al. 2018) (Fig. 1).
The UFRGS-PV-0629-T (Fig. 2) was found in an outcrop located next to the highway BR 287, near the entrance of the city of Dona Francisca, about 275 km from the state capital Porto Alegre, in the central region of Rio Grande do Sul state, Brazil (coordinates UTM (22) 0270854/6720302). The outcrop is known as “Posto de Gasolina” because it is located behind a gas station.

Fig. 1. Stratigraphic framework of the Triassic package from southern Brazil. Modified from Horn et al. (2018).
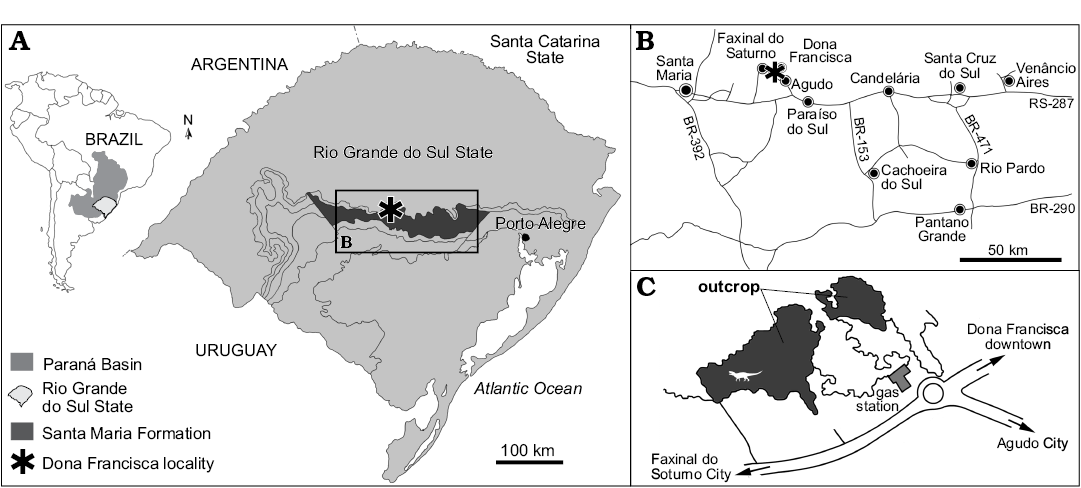
Fig. 2. Map showing location of the studied area, where the Prestosuchus chiniquensis Huene, 1938 (UFRGS-PV-0629-T) was collected, within Rio Grande do Sul State, Brazil (A) and municipality of Dona Francisca (B, C). Coordinates of the outcrop “Posto de Gasolina” UTM (22) 0270854/6720302.
Material and methods
We describe herein the skull of Prestosuchus chiniquensis based on the UFRGS-PV-0629-T (Fig. 3), which was referred to Prestosuchus chiniquensis by Mastrantonio (2010) and Mastrantonio et al. (2013). Additionally, we use data from the huge UFRGS-PV-0156-T, attributed to Prestosuchus chiniquensis by Barberena (1978) and the poorly preserved CPEZ-239b, described by Lacerda et al. (2016). All specimens are housed at the paleovertebrates collection of the Departamento de Paleontologia e Estratigrafia, Instituto de Geociências, Universidade Federal do Rio Grande do Sul, Brazil. The lectotype of Prestosuchus chiniquensis (Huene, 1938) consists of a skeleton with few skull bones: part of the lower jaw, right premaxilla, and palatal process of the maxilla, so it is only included in the comparison section for those few bones.
The UFRGS-PV-0629-T was scanned at the Hospital São José, Porto Alegre, using a medical CT scanner. Because the endocranial cavity of UFRGS-PV-0629-T was filled with ferruginous sediment, which is highly reflecting when analysing with CT scans, the images obtained were very difficult to study. The dataset consists of 168 slices with the following settings: field of view 321 mm, penetration power of 140.0 Kv and 108 mA, slice thickness of 0.8 mm and 0.4 mm of overlap. Analysis of the images and 3D reconstructions were developed with the open source software 3D Slicer v4.1.1. A complete reconstruction of the skull, in lateral view, based on the scanned bones, is provided.
Systematic palaeontology
Archosauria Cope, 1869 (sensu Gauthier and Padian 1985)
Pseudosuchia Zittel, 1887–1890 (sensu Gauthier and Padian 1985)
Loricata Merrem, 1820 (sensu Nesbitt 2011)
Genus Prestosuchus Huene, 1938
Type species: Prestosuchus chiniquensis Huene, 1938; see below.
Prestosuchus chiniquensis Huene, 1938
Figs. 3–20.
Type material: Lectotype: BSPG AS XXV 1-3/5-11/28-41/49 (Excavation 34; Huene 1942; Krebs 1976): splenial, anterior portion of the surangular, anterior portion of the angular, prearticular, right partial premaxilla, fragmentary dentary, three incomplete cervical vertebrae, fragmentary ribs, one sacral vertebrae, two sacral ribs, five anterior caudal vertebrae with chevron bones, 14 middle and posterior caudal vertebrae, right and left scapulacoracoid, interclavicle and clavicle, distal left humerus, right proximal and distal humerus, distal radius, fragmentary ulna, one manual phalanx, incomplete ilium, fragmentary ischia, pubes, and complete left hind limb. Paralectotype: BSPG AS XXV 7 (Excavation 41; Huene 1942): an articulated vertebral sequence, composed of two sacral vertebrae with sacral ribs, incomplete last dorsal and first caudal vertebrae, dorsal portion of the right ilium, a series of osteoderms articulated with the neural spines.
Type locality: “Weg Sanga”, Chiniquá District, São Pedro do Sul municipality, Rio Grande do Sul State, Brazil.
Type horizon: Pinheiros-Chiniquá Sequence of the Santa Maria Supersequence, Dinodontosaurus Assemblage Zone, Ladinian, Middle Triassic.
Material.—UFRGS-PV-0629-T: an almost complete skeleton, composed of a complete disarticulated skull, complete presacral vertebrae sequence (8 cervical, 13 dorsal), two sacral and three caudal vertebrae; complete scapular and pelvic girdle, mostly complete appendicular elements, composed of both humeri; proximal portions of the left ulna and radius; one left metacarpal; both femora, a right tibia and fibula, three isolated phalanges of a pes (Fig. 3). The description of the postcranial material and phylogenetic position is out of the scope of the present contribution, but it is part of a manuscript in process, as well as the histological information from long bones and ribs, complementarily to the microanatomy of the osteoderms previosly published (see Cerda et al. 2013). From the Dinodontosaurus Assemblage Zone, Ladinian, Middle Triassic, Dona Francisca municipality, Rio Grande do Sul State, Brazil.
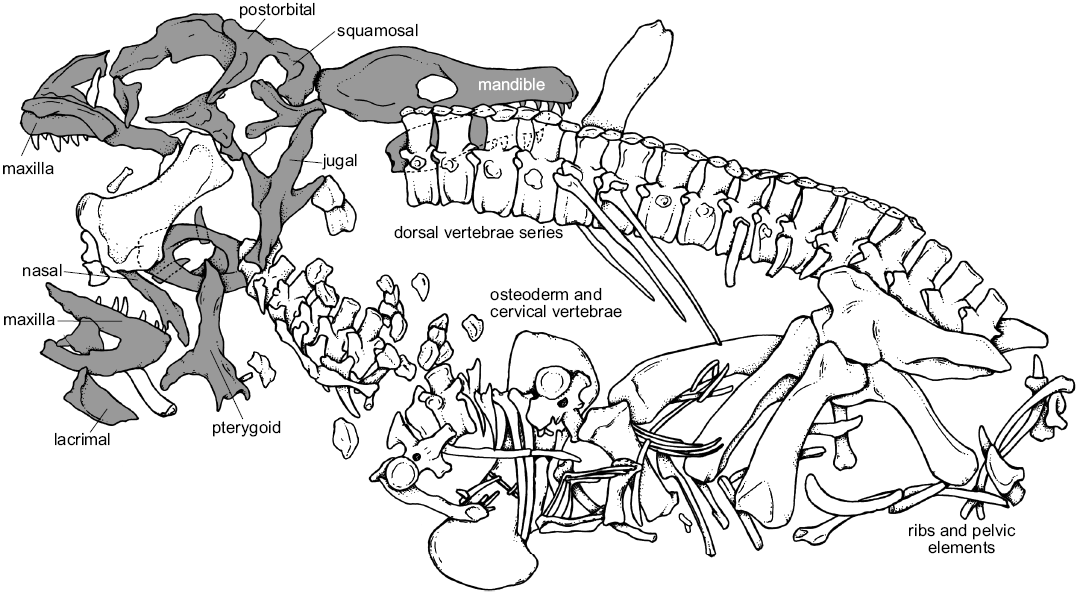
Fig. 3. Schematic drawing of the pseudosuchian archosaur Prestosuchus chiniquensis Huene, 1938 (UFRGS-PV-0629-T) from the Dinodontosaurus Assemblage Zone, Ladinian, Middle Triassic, Dona Francisca municipality, Rio Grande do Sul State, Brazil (drawing by Téo Veiga de Oliveira). Cranial elements in gray.
UFRGS-PV-0156-T: a large and complete articulated skull and associated partial axial skeleton consisting of 31 vertebrae (cervical, dorsal, sacral, and caudals, many with articulated dorsal osteoderms) along with unidentified fragmentary material. The skull of this specimen was described by Barberena (1978) and assigned to Prestosuchus chiniquensis. The complete specimen was later described by Azevedo in his unpublished Master’s thesis in 1991. From the Dinodontosaurus Assemblage Zone, Ladinian, Middle Triassic; Sanga Pascual outcrop, in the locality Pinheiros, Candelária municipality, Rio Grande do Sul State, Brazil.
UFRGS-PV-0473-T: an isolated braincase that was attributed to Prestosuchus chinquensis by Mastrantonio et al. 2013. Probably from the Dinodontosaurus Assemblage Zone, Ladinian, Middle Triassic, “Posto de Gasolina” outcrop, Dona Francisca municipality, Rio Grande do Sul State, Brazil (data regarding the collection are not clear).
UFRGS-PV-0152-T: an incomplete skull, vertebral sequence (cervical, dorsal, sacral, and caudal elements), complete scapular and pelvic girdles, humerus, left ulna, femora, left tibia and fibula, left calcaneus, fifth metatarsal and some phalanges, chevrons, and osteoderms. Nesbitt (2011: 33) considered UFRGS-PV-0152-T to be indistinguishable from UFRGS-PV-0156-T and BSPG AS XXV 1-3/5-11/28-41/49, due to overlapping features, but this specimen is not included in the present analysis because it is currently under study by Raugust (2014). From the Dinodontosaurus Assemblage Zone, Ladinian, Middle Triassic, Vale Verde municipality, Rio Grande do Sul State, Brazil (data on the exact location of the collection are not clear).
CPEZ-239b: cranial and post-cranial elements of at least two individuals of roughly the same size and very poorly preserved. There is an almost complete skull broken in two parts: the articulated rostrum and the posterior portion of the skull with associated mandibular elements, as well as two separate jaws of a second skull. This material was referred by Lacerda et al. (2016) to Prestosuchus chiniquensis. From the Dinodontosaurus Assemblage Zone, Ladinian, Middle Triassic, “Tree Sanga” outcrop, located between the cities of São Pedro do Sul, Mata and São Vicente do Sul (approximate UTM: 21J 749033 6716582), Rio Grande do Sul State, Brazil.
ULBRA-PVT-281: a large complete skull and a partial postcranial skeleton was refered to Prestosuchus chiniquensis by Roberto-Da-Silva et al. (2016). From the Dinodontosaurus Assemblage Zone, Ladinian, Middle Triassic, “Posto de Gasolina” outcrop, Dona Francisca municipality, Rio Grande doSul State, Brazil.
MCP-146: a complete pelvic girdle with the last dorsal, two sacral and three caudal vertebrae preserved in articulation that was briefly described and referred by Bonaparte (1984). Probably from the Dinodontosaurus Assemblage Zone, Ladinian, Middle Triassic, Rio Grande do Sul State, Brazil (data regarding the collection are not clear).
MCZ 4167: a poorly preserved but articulated specimen from the “Santa Maria Formation” (Santa Maria Supersequence sensu Zerfass et al. 2004), referable to Prestosuchus (see Lacerda et al. 2016 and Parrish 1993: 297). Probably from the Dinodontosaurus Assemblage Zone, Ladinian, Middle Triassic, “Posto de Gasolina” outcrop, Dona Francisca municipality, Rio Grande do Sul State, Brazil (data regarding the collection are not clear).
Diagnosis.—Two autapomorphies are described for this monospecific genus (sensu Desojo and Rauhut 2008): an anterior notch between the scapula and the coracoid; a longitudinal ridge on the dorsal surface of the ilium.
Description and comparisons.—Skull: Premaxilla (Fig. 4): Both premaxillae in the UFRGS-PV-0629-T are preserved disarticulated. In lateral view, the body of the premaxilla has a subrectangular shape, being 1.2 times longer anteroposteriorly than dorsoventrally deep. The anterior region of the main body of the premaxilla contacts its counterpart through a flat area 30 mm in length, and the posterior margin of the main body contacts the maxilla. The articulation between the premaxilla and the maxilla will be discussed later due to its implication in the diagnostic and phylogenetic context. On the lateral surface of the premaxilla, two nutrient foramina can be recognized. The ventral margin of the premaxilla has four alveoli that are oval-shaped anteriorly to round-shaped posteriorly. On the left premaxilla all teeth are well preserved, while the right premaxilla has only a broken remnant of the third tooth preserved. These teeth are labiolingually compressed, posteriorly curved, and serrated (four denticles per millimeter) on both mesial and distal surfaces.
The presence of four premaxillary teeth was reported for Saurosuchus galilei (Reig 1959; Sill 1974; Alcober 2000), Fasolasuchus tenax (Bonaparte 1981), Batrachotomus kupferzellensis (Gower 1999), Postosuchus kirkpatricki (Long and Murry 1995; Weinbaum 2011), Polonosuchus silesiacus (Sulej 2005), Rauisuchus tiradentes (Huene 1942; Lautenschlager and Rauhut 2015), Decuriasuchus quartacolonia (França et al. 2013) and all the other specimens of P. chiniquensis (UFRGS-PV-0156-T, Barberena 1978; CPEZ-239b, Lacerda et al. 2016; and ULBRA-PVT-281, Roberto-Da-Silva et al. 2016). Huene (1938) described Prestosuchus chiniquensis as having only three premaxillary, while here we identified four in the lectotype.
Posterodorsal to the main body of the premaxilla there are two processes, the nasal (= anterodorsal) process and the maxillary (= posterodorsal) process. These processes border the external naris almost completely, except by a small posterodorsal region of that aperture, which is bordered by the nasal. Consequently, the maxilla does not contribute to the margin of the external naris, as in most of the basal Loricata (e.g., Luperosuchus fractus, Saurosuchus galilei, Prestosuchus chiniquensis [UFRGS-PV-0156-T and CPEZ-239b], Fasolasuchus tenax), excepting Batrachotomus kupferzellensis in which the maxilla borders the external naris posteroventrally.
The nasal process is complete in the left premaxilla and conctacts the nasal posteriorly, but it is apparently broken in the right one (Fig. 4). On the left premaxilla, the nasal process forms a marked angle between its anterior and posterior portion resembling the condition seen in Fasolasuchus tenax (Bonaparte 1981), Decuriasuchus quartacolonia (França et al. 2013), and Saurosuchus galilei (Alcober 2000). However, the nasal process of the right premaxilla is more rounded as seen in other specimens of Prestosuchus chiniquesis (UFRGS-PV-0156-T), Rauisuchus tiradentes (Lautenschlager and Rauhut 2015), and Postosuchus kirkpatricki (Weinbaum 2011). This difference on the left and right side of the skull of the same specimen is here interpreted as a taphonomical deformation because the nasal process is thin and elongated and therefore susceptible to deformation, as seen in other specimens of Prestosuchus (UFRGS-PV-0156-T). On the other hand, a different condition is present in Batrachotomus kupferzellensis, in which there is no marked angle between the nasal process and the body of the premaxilla and this process is gently curved. The maxillary processes of UFRGS-PV-0629-T are incomplete in both premaxillae, but the one from the right premaxilla is more complete than the left one. This process contacts the maxilla posteroventrally and the nasal posterodorsally.
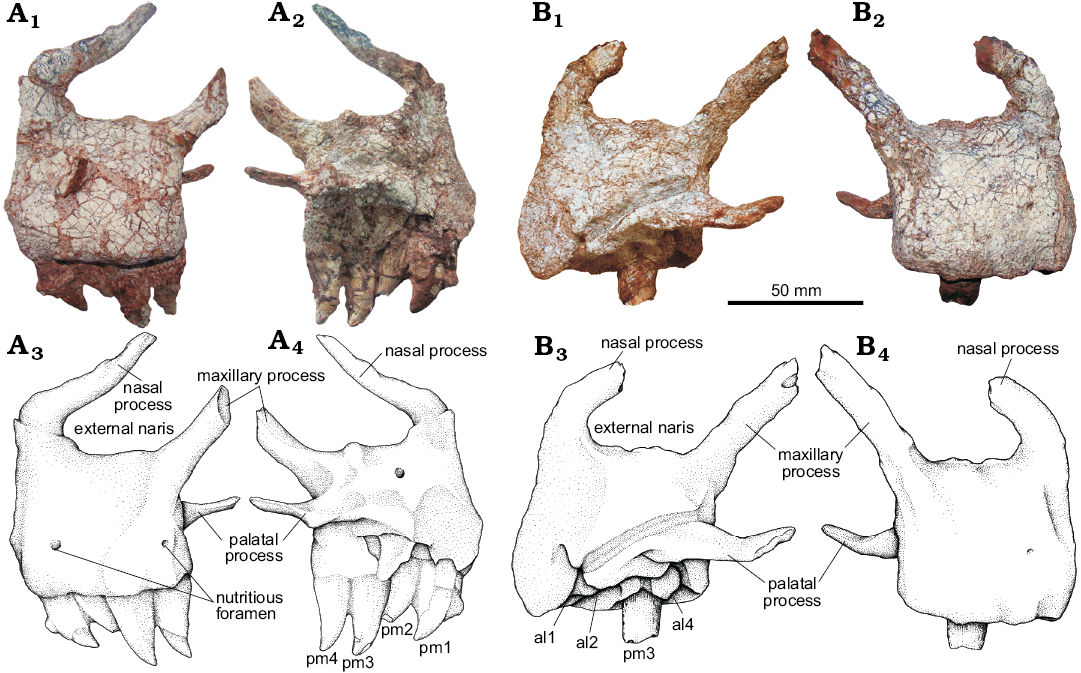
Fig. 4. The pseudosuchian archosaur Prestosuchus chiniquensis Huene, 1938 (UFRGS-PV-0629-T) from the Dinodontosaurus Assemblage Zone, Ladinian, Middle Triassic, Dona Francisca municipality, Rio Grande do Sul State, Brazil. A. Left premaxilla in lateral (A1, A3) and medial (A2, A4) views. B. Right premaxilla in medial (B1, B3) and lateral (B2, B4) views. pm3, pm4, refer to premaxillary tooth position; al1, al2, al4, refer to premaxillary empty alveoli position. Photographs (A1, A2, B1, B2) and explanatory drawings (A3, A4, B3, B4).
On the medial side of the premaxilla there is an unusually large palatal process that originates posterior to the contact area between the premaxillae and extends posteriorly beyond the main body of the premaxilla. The palatal process is dorsoventrally compressed and is covered by the palatal process of the maxilla (anteromedial process of Galton 1985) overlapping it ventrally. In Batrachotomus kupferzellensis (Gower 1999) and Postosuchus kirkpatricki (Weinbaum 2011) this process does not extend beyond the posterior margin of the premaxilla. Comparisons of the morphology of this process with other basal loricata were impaired because, in some of them, this process is broken (i.e., Rauisuchus tiradentes, Teratosaurus silesiacus) or was fossilized adpressed to the maxilla (i.e., Saurosuchus galilei, Decuriasuchus quartacolonia).
Maxilla (Fig. 5): A laterally compressed bone exposed on the lateral side of the skull. Both maxillae were preserved in the UFRGS-PV-0629-T. The lateral surface of the maxilla is rugose and has several nutritient foramina near its alveolar border. The maxilla can be divided into three portions, a main body, an ascending process, and a palatal process. The maxilla delimits the anterior and ventral margins of a large antorbital fenestra. This fenestra is surrounded by an antorbital fossa; this fossa is very shallow along the main body of the maxilla and extends ventrally, forming a platform of variable width between 15 and 20 mm from the edge of the fenestra. The antorbital fossa is delimited anteriorly by an anteroposterior ridge, which curves posterdorsally up the ascending process of the maxilla.
The main body of the maxilla of UFRGS-PV-0629-T is subrectangular in shape and contacts the premaxilla anteriorly at a concave articular surface. The main body is high on its anterior region and tapers posteriorly where it contacts the anteroventral border of the anterior process of the jugal (Fig. 5A1, A2). This contact area is flat and shows no sign of a suture. The jugal contacts the dorsolateral face of this process and some sliding in anterodorsal-posteroventral direction seems to be possible between both bones, which may suggest the possibility of cranial kinesis between these elements. On the posterior end of the maxilla, the dorsal margin is dorsally expanded into a laminar bilobated process whose posterior lobe is partially covered by the jugal through a smooth contact, which also suggests some sliding among these bones. This same structure could also contatct the lacrimal, but the descendent process of this bone is broken, and therefore it cannot be confirmed. This dorsal expansion of the maxilla is very fragile and is apparently broken in the right maxilla of UFRGS-PV-0629-T, as well as in several specimens used for comparison. The maxilla of the UFRGS-PV-0629-T and UFRGS-PV-0156-T are more triangular in shape than that of the CPEZ-239b, which is more rectangular, resembling the condition of Decuriasuchus quartacolonia (MCN-PV10.004, MCN-PV10.105a, c, d) in which the height of the main body of the maxilla is the same along its entire length. On the medial surface of the maxilla, a groove can be identified extending from the level of the sixth tooth up to the tenth tooth as the articular facet for the palatine (Fig. 5A3, A4).
The ventral margin of the maxilla is slightly concave and has 13 alveoli, with nine teeth preserved in position in the right maxilla and six in the left maxilla. These teeth are also labiolingually compressed, posteriorly curved, and serrated on both mesial and distal edges, with three to four denticles per millimeter. This same number of alveoli can also be seen in the CPEZ-239b (Prestosuchus chiniquensis), Postosuchus kirkpatricki (Long and Murry 1995; Weinbaum 2011), and Teratosaurus suevicus (Galton 1985; Benton 1986; Brusatte et al. 2009). The UFRGS-PV-0156-T (Prestosuchus chiniquensis) would have 11 or 12 teeth in each jaw according to Barberena (1978); however, the current analysis suggests that the specimen would have at least 12 teeth, or even 13. The maxillary tooth count is apparently variable amongst basal loricatans. For instance, Batrachotomus kupferzellensis (Gower 1999), Fasolasuchus tenax (Bonaparte 1981), and Polonosuchus silesiacus (Sulej 2005) were reported to have 11 maxillary teeth, although the latter two might also have more teeth according to the authors that described them. Saurosuchus galilei (Alcober 2000) has 12 maxillary teeth whereas Tikisuchus romeri (Chatterjee and Majumdar 1987) only has nine teeth. On the other hand, the tooth count of Decuriasuchus quartacolonia (MCN-PV10.004; França et al. 2013) is much higher, reporting 17 preserved alveoli. These taxa are represented by individuals of different size and probably different ages. None of them have a large enough sample size to determine if the variability in tooth count is the result of either intraspecific or ontogenetic variabilities or even sexual dimorphism.
The interdental plates are large on the anterior region of the maxilla and become smaller towards the posterior end. The contact margins between the plates are posteroventrally sloped following the orientation of the teeth and some are slightly overlapped. In UFRGS-PV-0629-T the interdental plates are not fused resembling the condition on the CPEZ-239b (Prestosuchus chiniquensis), Polonosuchus silesiacus (Sulej 2005), Saurosuchus galilei (PVL 2062; Alcober 2000), and Decuriasuchus quartacolonia (MCN-PV10.004; França et al. 2013). This condition cannot be clearly determined in Batrachotomus kupferzellensis (Gower 1999) and Luperosuchus fractus (Nesbitt and Desojo 2017), because their interdental plates are poorly preserved. The fusion of interdental plates is reported in few basal loricatans, such as Fasolasuchus tenax (PVL 3851), Postosuchus kirkpatricki, Teratosaurus suevicus, and Vivaron haydeni, a taxon based on isolated and fragmentary materials from New Mexico (Lessner et al. 2016). However, according to Weinbaum (2011: 24), the interdental plates are not fused in Postosuchus and can be distinguished from one another.
The ascending process of the maxilla is a long posterodorsally directed projection that expands distally. The process originates anterior to the antorbital fenestra and extends posterodorsally at a roughly 45° angle to the body of the maxilla and terminates in a convex articular surface for the lacrimal. The anterior border of the ascending process is overlapped by the maxillary process of the premaxilla and its dorsal border is marked by a groove where the maxillary process of the nasal articulates. The posterior end of the ascending process of the maxilla overlaps the anterior ramus of the lacrimal delimiting together the dorsal and posterior margins of the antorbital fenestra and shaping the antorbital fossa (Fig. 5A1, A2).
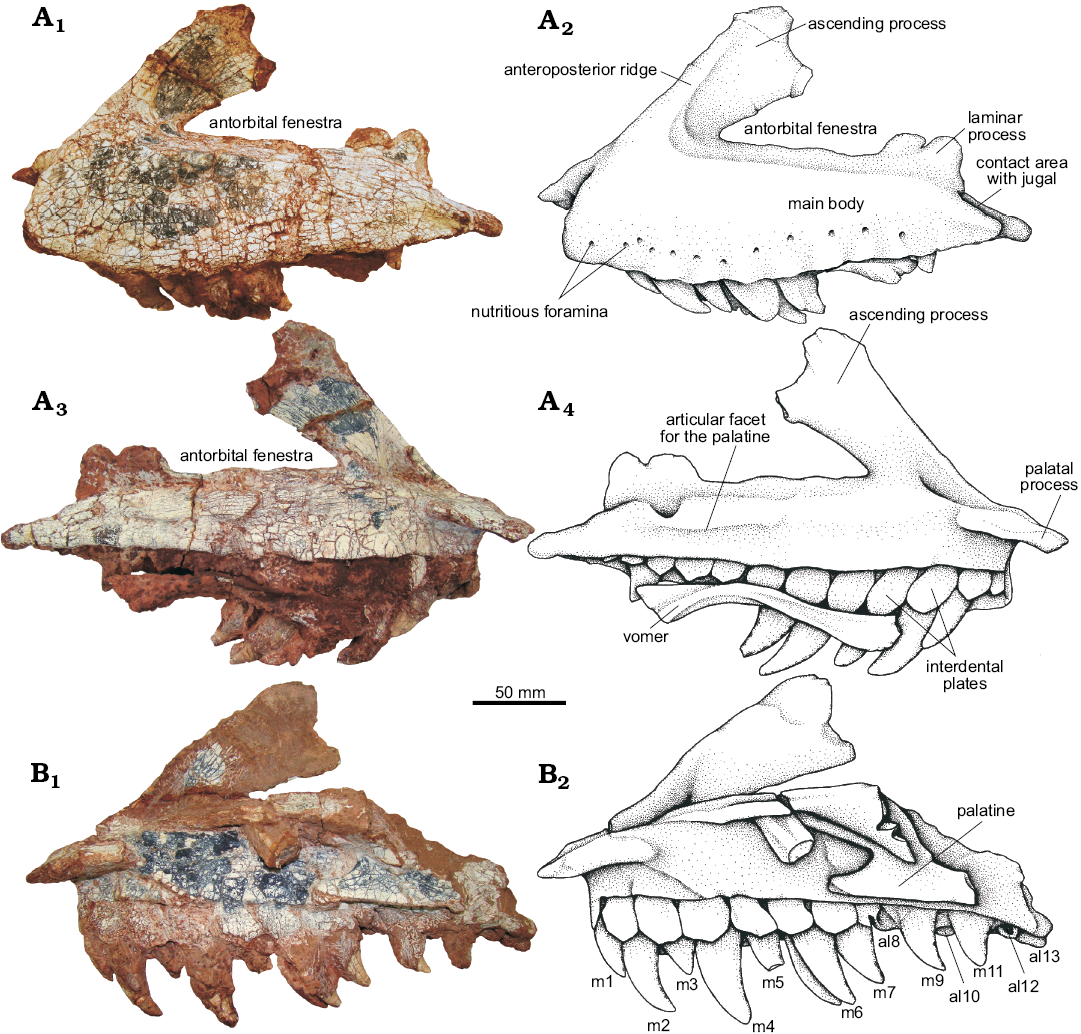
Fig. 5. The pseudosuchian archosaur Prestosuchus chiniquensis Huene, 1938 (UFRGS-PV-0629-T) from the Dinodontosaurus Assemblage Zone, Ladinian, Middle Triassic, Dona Francisca municipality, Rio Grande do Sul State, Brazil. A. Left maxilla in lateral (A1, A2) and medial (A3, A4) views. B. Right maxilla in medial view. m1–7, m9, m11, refer to maxillary tooth position; al10, al12, al13, refer to maxillary alveolus position. Photographs (A1, A3, B1) and explanatory drawings (A2, A4, B2).
The palatal process of the maxilla can be seen in medial view and projects anteromedially from the level of the second maxillary tooth and extending anteriorly to the anterior margin of the maxilla itself. The palatal process is dorsoventrally compressed, short, tapers anteriorly and overlaps dorsally the palatal process of the premaxilla, as in the CPEZ-239b (Prestosuchus chiniquensis). At the level of the palatal process of the maxilla, a foramen can be recognized on the anterodorsal surface of this bone (Fig. 6A). This foramen opens medially to the slit between premaxilla and the maxilla. A similar condition was identified in Decuriasuchus quartacolonia (França et al. 2013) and Postosuchus kirkpatricki (Weinbaum 2011). On the other hand, two foramina (probably for the same artery and nerve) were described at that region for Teratosaurus suevicus (Galton 1985), and Polonosuchus (= Teratosaurus) silesiacus (Sulej 2005; Brusatte et al. 2009), named rostromedial and rostrolateral by Brusatte et al. 2009 (respectively, medial anterior and lateral anterior sensu Galton, 1985). According to Galton (1985), both foramina were probably the route for branches of the maxillary artery and of the inferior orbital nerve (a branch of trigeminal nerve V). França et al. (2013) identified a single foramen in Decuriasuchus quartacolonia, which would be the rostrolateral (being the rostromedial absent) and the same is here stated to Prestosuchus chiniquensis.
In UFRGS-PV-0629-T (Prestosuchus chiniquensis), the premaxillae are disarticulated from the maxillae, but when they are brought into contact with one another, a small slit-opening is present in lateral view between the anterior margin of the maxilla and posterior margin of the premaxilla (Fig. 6B). This opening is also described in Postosuchus kirkpatricki (Chatterjee 1985), Saurosuchus galilei (Alcober 2000), Decuriasuchus quartacolonia (França et al. 2013), and Luperosuchus fractus (Romer 1971; Nesbitt and Desojo 2017), while its presence is controversial for other rauisuchians (see discussion). However, according to Weinbaum (2011: 22), the subnarial fenestra of Postosuchus kirkpatricki created by the articulation of the premaxilla and maxilla is much smaller than originally figured by Chatterjee (1985: fig. 2). In the reconstruction provided by Chatterjee (1985), which is based on the holotype, TTUP 9000, the palatal processes on both the left and right maxillae are broken and missing their anterior ends. Therefore, the process does not articulate with the maxillary process of the premaxilla and as a result the fenestra appears to be approximately twice its natural size. When the premaxilla and maxilla are properly articulated, the fenestra is a small and ovate opening (Weinbaum 2011).

Fig. 6. The pseudosuchian archosaur Prestosuchus chiniquensis Huene, 1938 (UFRGS-PV-0629-T) from the Dinodontosaurus Assemblage Zone, Ladinian, Middle Triassic, Dona Francisca municipality, Rio Grande do Sul State, Brazil. A. Explanatory drawing of the left maxilla in anterior view. B. Photograph of the articulated left premaxilla and maxilla in lateral view exhibiting a slit-opening between both elements.
Nasal (Fig. 7): An anteroposteriorly elongated and laterally compressed element that resembles the shape seen in Saurosuchus galilei (PVL 2062, PVSJ 32; Alcober 2000) and of Prestosuchus chiniquensis (UFRGS-PV-0156-T and CPEZ-239b). In the UFRGS-PV-0629-T both nasals are disarticulated, the right one is complete but the left one has only preserved its anterior third. The nasal can be differentiated into a main elongated body and two processes that project anteriorly from its anterior end.
The main body of the nasal articulates anterolaterally with the ascending process of the maxilla and posterolaterally with the lacrimal. The medial corner of the posterior margin of the nasal forms a small wedge that fits between the prefrontal and the frontal. In lateral view, the convex external surface of the nasals appears to form a “roman nose aspect” as that described in some taxa as Batrachotomus kupferzellensis (Gower 1999), Luperosuchus (PULR 04; Nesbitt and Desojo 2017), however, this is not the case in Prestosuchus chiniquensis (UFRGS-PV-0629-T, UFRGS-PV-0156-T, and CPEZ-239b).
Along the dorsolateral margin of the nasal, a thick rugose ridge can be recognized resembling that of Rauisuchus tiradentes (Lautenschlager and Rauhut 2015) but not reaching the development seen in Batrachotomus kupferzellensis (Gower 1999) and Postosuchus kirkpatricki (Long and Murry 1995; Weinbaum 2011). UFRGS-PV-0629-T also differs from Decuriasuchus quartacolonia (França et al. 2013) in which this ridge is completely absent. The ridge present in UFRGS-PV-0629-T is gently curved posterolaterally and delimits the lateral margin of a deep fossa on the posterior half of the nasal that extends up to the frontal (Fig. 7A).
The descending process of the nasal contacts the ascending process of the maxilla and together with the maxillary process of the premaxilla, forms the posteroventral margin of the external naris. This descending process is elongated and thin resembling that of Polonosuchus silesiacus (Sulej 2005; Brusatte et al. 2009) but differing from that of Batrachotomus kupferzellensis (Gower 1999) and Fasolasuchus tenax (Bonaparte 1981) in which it is short and wide. This process is incomplete in Saurosuchus galilei (Alcober 2000) and Rauisuchus tiradentes (Lautenschlager and Rauhut 2015).
The anterior process of the nasal of UFRGS-PV-0629-T is longer than the descending process and it is laterally compressed. It articulates with the nasal process of the premaxilla and forms the anterodorsal margin of the external naris as in all basal loricatans previously compared.
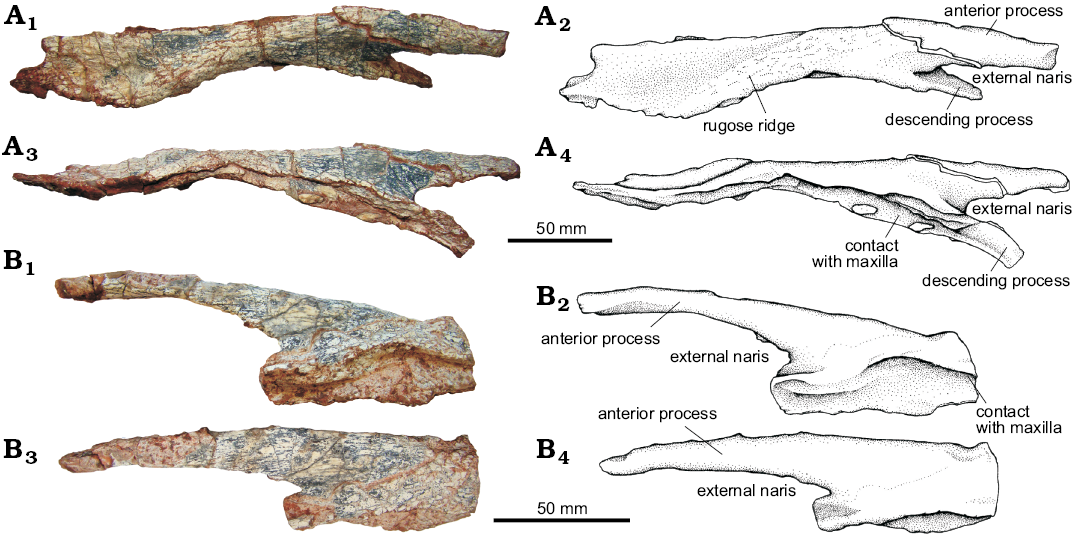
Fig. 7. The pseudosuchian archosaur Prestosuchus chiniquensis Huene, 1938 (UFRGS-PV-0629-T) from the Dinodontosaurus Assemblage Zone, Ladinian, Middle Triassic, Dona Francisca municipality, Rio Grande do Sul State, Brazil. A. Right nasal in dorsal (A1, A2) and lateral (A3, A4) views. B. Left nasal in lateral (B1, B2) and dorsal (B3, B4) views. Photographs (A1, A3, B1, B3) and explanatory drawings (A2, A4, B2, B4).
Lacrimal (Fig. 8): Only the disarticulated, but complete left lacrimal of UFRGS-PV-0629-T is preserved. This bone is anteroposteriorly elongated and laterally compressed. It is L-shaped in lateral view, with its anterior ramus approximately three times longer than the descending ramus, resembling most basal loricatans but differing from Postosuchus kirkpatricki (Weinbaum 2011) and Polonosuchus silesiacus (Brusatte et al. 2009) that have a short anterior ramus. The anterior ramus of the lacrimal of UFRGS-PV-0629-T is laminar; it overlaps the maxilla laterally and contacts the nasal dorsomedially. On the lateral surface of the posterior half of the lacrimal there is a well-developed crest that extends dorsoventrally from the dorsal margin of this element and runs along the descending ramus delimiting the posterodorsal border of the antorbital fossa. This crest does not reach the lateral extent seen in Decuriasuchus quartacolonia (MCN-PV10.105a; França et al. 2013), in which it forms a lateral expansion that covers the antorbital fenestra dorsally, an autapomorphic condition for the latter.
The posterior portion of the lacrimal is thicker than the anterior ramus and has a large lacrimal foramen. The posterior margin of the lacrimal is irregular and has a small posterior projection at the same level of the lacrimal foramen, ventral to the posterodorsal projection of the lacrimal. The descending process is located ventral to the lacrimal foramen and is dorsoventrally oriented. On its medial surface, the articular surface for the anterodorsal process of the jugal can be clearly recognized. There is no evidence of a tight suture between the lacrimal and the jugal in this region, which suggests the possibility of some degree of kinesis (Liparini 2008) as was also described for Saurosuchus galilei (Alcober 2000). On the other hand, in the specimen UFRGS-PV-0156-T (Azevedo 1991), the lacrimal and the jugal seem to be tightly linked. It could suggest that the differences among the specimens of Prestosuchus could be interpreted as result of ontogenetic variation but considering that the holotype of Saurosuchus and UFRGS-PV-0156-T have approximately the same size, appears that the “tight link” observed in the last one is more likely caused by differences in preservation.

Fig. 8. The pseudosuchian archosaur Prestosuchus chiniquensis Huene, 1938 (UFRGS-PV-0629-T) from the Dinodontosaurus Assemblage Zone, Ladinian, Middle Triassic, Dona Francisca municipality, Rio Grande do Sul State, Brazil; left lacrimal in lateral view. Photograph (A1) and explanatory drawing (A2).
Prefrontal (Figs. 9–11): Both prefrontals in UFRGS-PV-0629-T are preserved but the left one is disarticulated (Figs. 9, 10A3, A4). The prefrontal is a small element; it is subtriangular in lateral view and subrectangular in dorsal view. It is sutured anterolaterally to the lacrimal, anteromedially to the nasal, and posteromedially to the frontal, and forms the anterodorsal margin of the orbit. The morphology of the prefrontal of UFRGS-PV-0629-T is compatible with that of the UFRGS-PV-0156-T and CPEZ-239b of Prestosuchus chiniquensis. In the first (UFRGS-PV-0156-T), the prefrontals are preserved in articulation, whereas in the second (CPEZ-239b), only the left prefrontal is present but in a poor state of preservation.
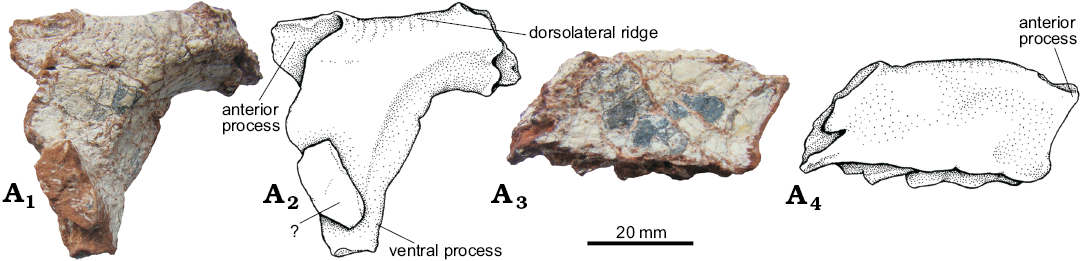
Fig. 9. The pseudosuchian archosaur Prestosuchus chiniquensis Huene, 1938 (UFRGS-PV-0629-T) from the Dinodontosaurus Assemblage Zone, Ladinian, Middle Triassic, Dona Francisca municipality, Rio Grande do Sul State, Brazil; left prefrontal in lateral (A1, A2) and dorsal (A3, A4) views. Photographs (A1, A3) and explanatory drawings (A2, A4).
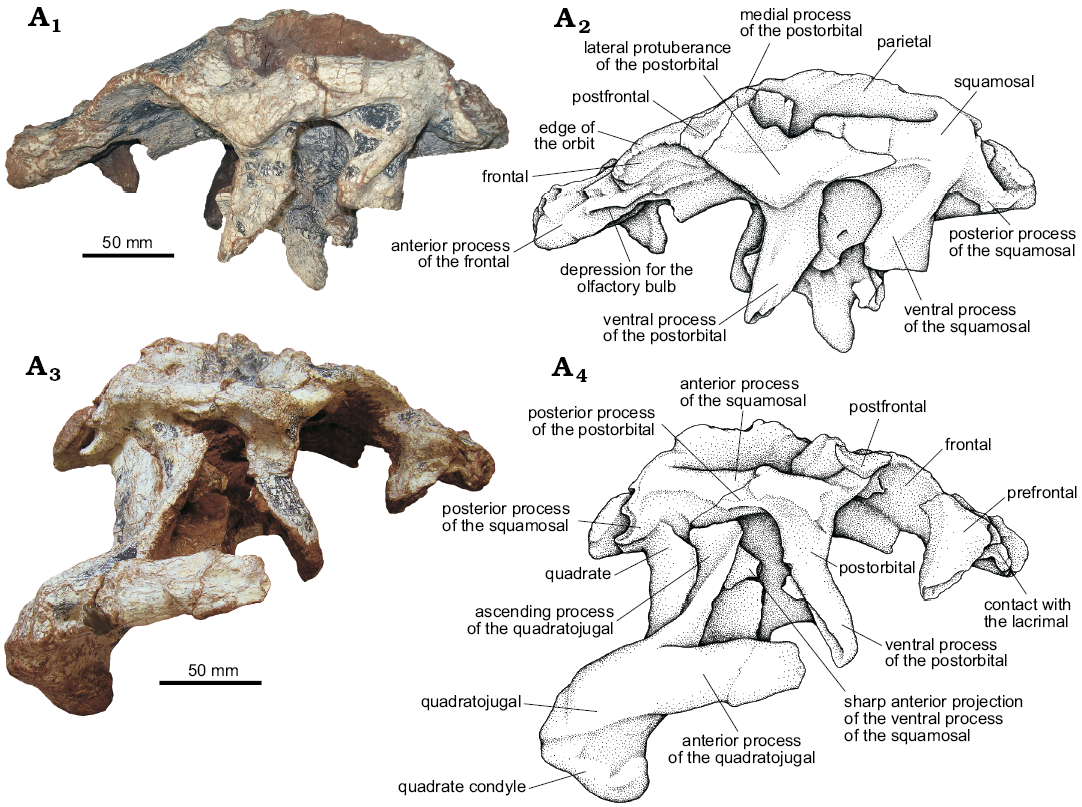
Fig. 10. The pseudosuchian archosaur Prestosuchus chiniquensis Huene, 1938 (UFRGS-PV-0629-T) from the Dinodontosaurus Assemblage Zone, Ladinian, Middle Triassic, Dona Francisca municipality, Rio Grande do Sul State, Brazil; posterior portion of the skull in left (A1, A2) and right (A3, A4) side views. Photographs (A1, A3) and explanatory drawings (A2, A4).
In the specimen UFRGS-PV-0629-T the anterior process of the prefrontal reaches the anterior extent of the frontal (Fig. 11), as in Decuriasuchus quartacolonia (MCN-PV 10105a; França et al. 2013), Batrachotomus kupferzellensis (Gower 1999; SMNS 80260), and Saurosuchus galilei (PVSJ 32 based on JBD personal observations). This condition differs from that seen in Luperosuchus fractus in which the anterior process is finger like in dorsal view fitting between the lacrimal and the frontal and does not reach the level of the anterior extent of the frontal (PULR 04; Nesbitt and Desojo 2017). The prefrontal of Prestosuchus chiniquensis also differs from that of Postosuchus kirkpatricki (Weinbaum 2011) because in the latter the prefrontal is anteroposteriorly short and does not reach the anterior extent of the frontal.
The ventral process of the prefrontals of UFRGS-PV-0629-T (Figs. 9A1, A2, 10A3, A4) is not preserved in articulation with the lacrimal, but its morphology suggests that it overlapped the posterior border of the ventral process of the latter, delimiting the anterodorsal margin of the orbit. Gower (1999) suggests that in Batrachotomus kupferzellensis the ventral process of the prefrontal would delimit the whole anterior margin of the orbit, excluding the lacrimal from this opening. Even considering that the ventral process of the prefrontal is incomplete in UFRGS-PV-0629-T, it would not reach the jugal, since there is no sign of a suture on the posterior margin of the descending process of the lacrimal, as seen in Batrachotomus.
The prefrontal of UFRGS-PV-0629-T has a low dorsolateral ridge that is a continuation of that described for the nasal and lacrimal. On the dorsal surface of the prefrontal, the posterior end of this ridge curves medially forming less than half of the dorsal margin of the orbit and reaching the frontal. This curvature is similar to that seen in the prefrontal of UFRGS-PV-0156-T (Azevedo 1991), Saurosuchus galilei (PVL 2062, PVSJ 32; Alcober 2000), and Batrachotomus kupferzellensis (Gower 1999). On the other hand, the prefrontal, the postorbital and the frontal are partially overlapped by the palpebral in Postosuchus kirkpatricki (Long and Murry 1995; Weinbaum 2011).
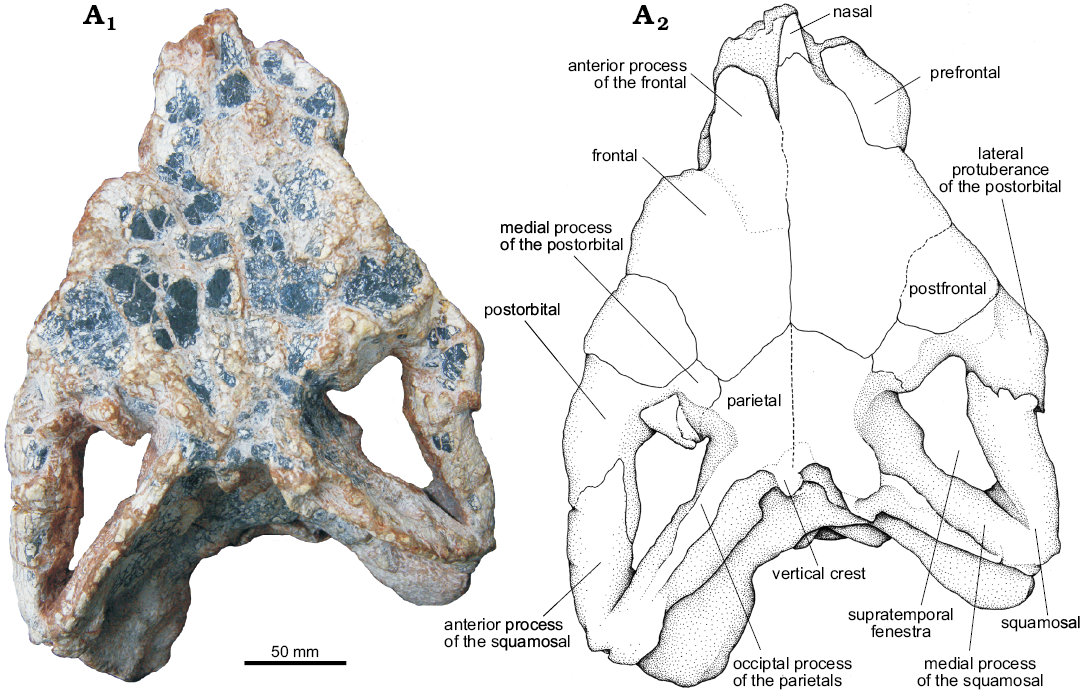
Fig. 11. The pseudosuchian archosaur Prestosuchus chiniquensis Huene, 1938 (UFRGS-PV-0629-T) from the Dinodontosaurus Assemblage Zone, Ladinian, Middle Triassic, Dona Francisca municipality, Rio Grande do Sul State, Brazil; posterior portion of the skull in dorsal view. Photograph (A1) and explanatory drawing (A2).
Frontal (Figs. 10, 11): both frontals of the UFRGS-PV-0629-T are preserved in articulation with several elements of the braincase. The frontal is a large bone that forms the largest area of the skull roof (approximately two-thirds). The frontal has an anterior process with a rectangular shape that contacts the nasal anteriorly. It also contacts the prefrontal laterally through an inverted L-shaped suture, with the major portion anteroposteriorly oriented and the smaller one lateromedially, reaching the edge of the orbit (Figs. 10, 11). Posterior to its contact with the prefrontal, the frontal extends to form part of the dorsal margin of the orbit as can also be seen in Batrachotomus kupferzellensis (Gower 1999), Saurosuchus galilei (PVL 2062, PVSJ 32; Alcober 2000), Decuriasuchus quartacolonia (França et al. 2013), and the UFRGS-PV-0156-T. On the contrary, the frontal does not participate on the margin of the orbit in Postosuchus kirkpatricki (Long and Murry 1995; Weinbaum 2011) as there is a palpebral bone present between the frontal and orbital margin. The frontal contacts the postfrontal posterolaterally, at a small elevation that occurs on the dorsal surface of the skull roof, and the parietal posteriorly, at a straight contact posterolaterally directed at 45° from the midline. The frontal also contacts a small portion of the postorbital, where the latter fits between the parietal and the postfrontal.
The dorsal surface of the frontal forms part of the dorsal depression located on the anterior region of the skull roof. In UFRGS-PV-0629-T, the frontals are slightly shifted, with the left one being more dorsally located than the right one. This dislocation makes it difficult to clearly identify the presence of a longitudinal elevation located at the midline as that recognized in the UFRGS-PV-0156-T. In Saurosuchus galilei (PVSJ 32) and Decuriasuchus quartacolonia (MCN-PV10.105a, d, MCN-PV10.004) the dorsal surface of the frontal has a small sagittal crest on its posterior third (França et al. 2013), similar to the “slightly rugose ridge” described in Luperosuchus fractus (PVLR 04; Nesbitt and Desojo 2017), but not as pronounced as the condition seen in Batrachotomus kupferzellensis (SMNS 52970, SMNS 80260; Gower 1999). The rest of the dorsal surface of the frontal of UFRGS-PV-0629-T is smooth and has no ornamentation as it also happens in Decuriasuchus quartacolonia (UFRGS-PV-0156-T; França et al. 2013) and Postosuchus kirkpatricki (Weinbaum 2011). By contrast, the dorsal surface of the frontal is rugose in Luperosuchus fractus (PULR 04; Nesbitt and Desojo 2017), Saurosuchus galilei (Alcober 2000) and Batrachotomus kupferzellensis (SMNS 52970, SMNS 80260; Gower 1999).
On the ventral surface of the anterior half of the frontals there is a well-developed depression at the midline, framed by anteroposteriorly oriented ridges (Fig. 10A1, A2). This depression records the dorsal margin of the olfactory bulbs as suggested for Batrachotomus kupferzellensis (Gower 1999), Postosuchus kirkpatricki (Weinbaum 2011), and Luperosuchus fractus (Nesbitt and Desojo 2017).
Postfrontal (Figs. 10, 11): Both postfrontals of UFRGS-PV-0629-T are preserved articulated in their natural position. They have a rectangular shape and participate on the dorsal margin of the orbit (Figs. 10, 11), similar to the condition of Saurosuchus galilei (PVL 2062, PVSJ 32; Alcober 2000), Decuriasuchus quartacolonia (França et al. 2013), and the UFRGS-PV-0156-T. It differs from Postosuchus kirkpatricki (Long and Murry 1995; Weinbaum 2011) in which the postfrontal is smaller and only visible in dorsal view, located between the parietal and the postorbital, and not reaching the dorsal margin of the orbit. Prestosuchus also differs from Luperosuchus fractus (PULR 04; Nesbitt and Desojo 2017) in which the postfrontal is completely excluded from the orbital margin by the palpebral.
The postfrontal articulates with the frontal anteromedially, and the postorbital posteromedially (Fig. 11). On its ventral surface, a semicircular depression for the articulation of the dorsolateral process of the laterosphenoid can be recognized, without evidence of mobility. By contrast, a potentially kinetic joint was suggested for Saurosuchus galilei by Alcober (2000).
Parietal (Figs. 10–12): Both parietals in UFRGS-PV-0629-T are preserved in their original position and articulated with other elements of the skull roof. These constitute approximately the posterior third of the skull roof and most of the posteror margin of the supratemporal fenestra. Each parietal contacts the frontal anteriorly, the postorbital anterolaterally, the supraoccipital posteromedially, and the laterosphenoid ventrolaterally.
The anterior portion of the parietals forms the medial plate between supratemporal fenestrae where it contacts its counterpart (Fig. 11). The sagittal crest described for Postosuchus kirkpatricki (Long and Murry 1995; Weinbaum 2011) is not present in any specimen of Prestosuchus chiniquensis (UFRGS-PV-0629-T, UFRGS-PV-0156-T, CPEZ-239b), Saurosuchus galilei (PVL 2062, PVSJ 32; Alcober 2000), Decuriasuchus quartacolonia (MCN-PV 10004, 10105a; França et al. 2013), and Luperosuchus fractus (PULR 04; Nesbitt and Desojo 2017). Moreover, the medial plate of Prestosuchus chiniquensis (UFRGS-PV-0629-T, UFRGS-PV-0156-T) does not extend up to the frontals also differing from Batrachotomus kupferzellensis (SMNS 52970, SMNS 80260; Gower 1999) in which the sagittal crest in continued as a medial elevation along the dorsal surface of the frontals.
In dorsal view, the occipital process of the parietals of UFRGS-PV-0629-T is elongated, thin, and anterolaterally to posteromedially compressed, delimiting the posteromedial margin of the supratemporal fenestrae (Fig. 11). The anterolateral surface of the occipital processes of the parietals articulates with the posterodorsal surface of the occipital processes of the squamosals through a wide contact in which both elements together form the posterior margin of the supratemporal fenestrae. The ventral borders of the occipital processes of the parietals delimit the dorsal margin of the posttemporal fenestrae (Fig. 12). These fenestrae are tight slits that are ventrally delimited by the paraoccipital processes of the opisthotics.
In occipital view, the posterior surface of the parietals of UFRGS-PV-0629-T form a well delimited vertical crest at the midline that runs from the dorsal margin of this element up to the supraoccipital (Fig. 12). This same crest can be seen in the UFRGS-PV-0156-T, but its ventral extent seems lower and shorter despite the poor preservation of that region, which in turn makes it difficult to identify in that specimen. This vertical crest cannot be recognized in Batrachotomus kupferzellensis (SMNS 52970), but two small tubercles were identified in the region where both parietals meet. Nevertheless, the vertical crest is present along its supraoccipital. The posterior region of the parietals is damaged in Saurosuchus galilei (PVSJ 32) and therefore, the presence of the vertical crest cannot be clarified within this region, although it is present in the supraoccipital.
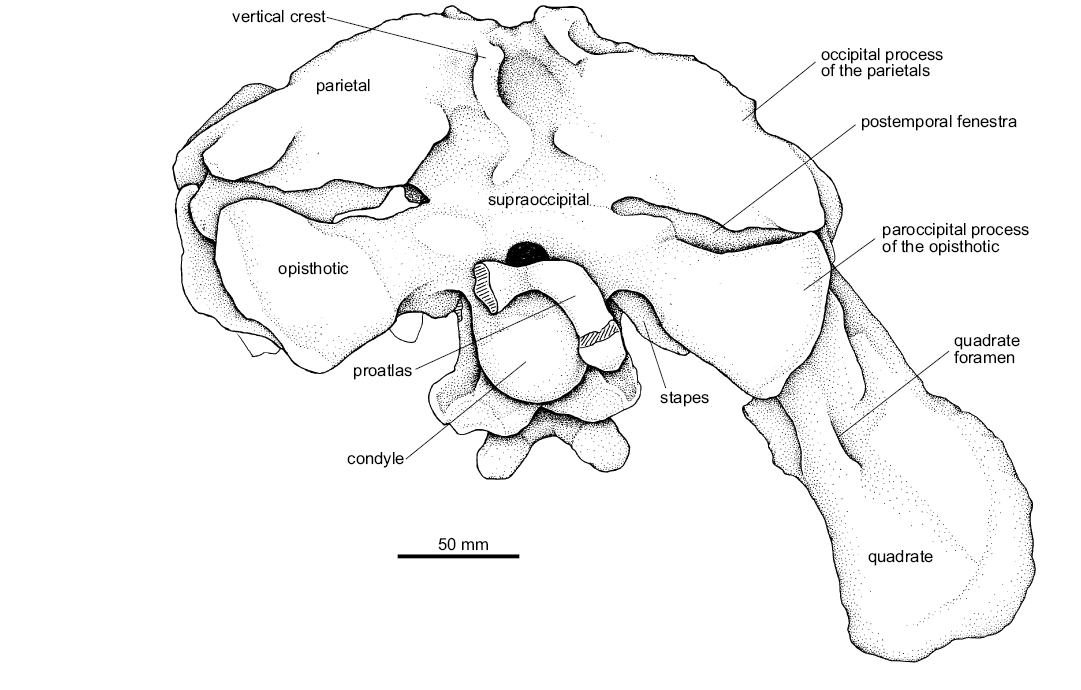
Fig. 12. The pseudosuchian archosaur Prestosuchus chiniquensis Huene, 1938 (UFRGS-PV-0629-T) from the Dinodontosaurus Assemblage Zone, Ladinian, Middle Triassic, Dona Francisca municipality, Rio Grande do Sul State, Brazil; explanatory drawing of the skull in occipital view.
Postorbital (Figs. 10, 11): Both postorbitals are preserved in their natural position and in articulation in the UFRGS-PV-0629-T. The postorbital is T-shaped in lateral view, with medial, posterior, and ventral (or descending) processes.
The short, cylindrical medial process of the postorbital protrudes anteromedially, contacts the posterior border of the frontal and postfrontal and the anterolateral margin of the parietal. This process forms the anterolateral border of the supratemporal fenestra (Fig. 11).
The posterior process of the postorbital is almost cylindrical and has a clear articular surface for anterior process of the squamosal on its dorsoposterior edge (Figs. 10, 11). The lateral surface of the postorbital and its posterior process are thick and rugose, resembling the texture seen in the dorsolateral surface of the preorbital region of the skull.
The ventral process of the postorbital of UFRGS-PV-0629-T is long, thin, and almost vertically oriented, projecting slightly anteroventrally to contact the ascending process of the jugal and therefore forming part of the postorbital bar that separates the orbit from the infratemporal fenestra (Fig. 10). The articular surface for the jugal occupies most of the posteromedial margin of the ventral process of the postorbital. In Batrachotomus kupferzellensis (SMNS 80260; Gower 1999) and Postosuchus kirkpatricki (Long and Murry 1995; Weinbaum 2011) this process was described as sinuous, granting the orbit a key-hole shape, a condition described as a diagnostic character for Rauisuchidae by some authors (e.g., Benton and Clark 1988; Alcober 2000), whereas in Saurosuchus galilei (PVL 2062, PVSJ 32; Alcober 2000), Decuriasuchus quartacolonia (França et al. 2013), and Luperosuchus fractus (PULR 04; Nesbitt and Desojo 2017) it is almost straight with a very slight concavity on its dorsal half. In the UFRGS-PV-0629-T, the ventral process of the left postorbital is straight but the right one is slightly sinuous. On the contrary, in the UFRGS-PV-0156-T this condition is reversed (with the right ventral process being straight and the left one sinuous). The presence of a slope in the postorbital bar has been scored in many morphological matrices used for phylogenetic analyses (e.g., Benton and Clark 1988; Juul 1994; Brusatte et al. 2010) but we are not confident about their natural condition in Prestosuchus.
Squamosal (Figs. 10, 11): The squamosals of UFRGS-PV-0629-T are preserved in their original position and articulated to the skull roof. They are composed of four processes that project in anterior, medial, posterior and ventral directions.
The anterior process of the squamosal is short, approximately circular in cross-section as seen in the UFRGS-PV-0156-T (Prestosuchus chiniquensis), Saurosuchus galilei (PVSJ 32), Decuriasuchus quartacolonia (França et al. 2013), but unlike the suboval shape seen in Batrachotomus kupferzellensis (SMNS 80260) and Polonosuchus silesiacus (ZPAL AbIII-563). The ventral surface of the anterior process of the squamosal of UFRGS-PV-0629-T articulates with the posterior process of the postorbital separating the supratemporal fenestra from the infratemporal fenestra (Fig. 10).
The medial process of the squamosal is laminar and anteromedially projected, it overlaps the lateral side of the occipital process of the parietal forming the posterolateral margin of the supratemporal fenestra (Fig. 11).
The ventral process of the squamosal is laminar; it laterally overlaps the quadrate on its proximal region and contacts the ascending process of the quadratojugal on its dorsal end delimiting the posterodorsal border of the infratemporal fenestra (Fig. 10). The anteroventral end of this process forms a sharp anterior projection that can also be recognized in UFRGS-PV-0156-T, Saurosuchus galilei (PVSJ 32), and Decuriasuchus quartacolonia (MCN-PV 10105 a, c, d; França et al. 2013). This condition was described as a diagnostic character for Rauisuchidae by Alcober (2000), but the ventral process was not preserved in the squamosals of Batrachotomus kupferzellensis (SMNS 52970, SMNS 80260; Gower 1999). In Polonosuchus silesiacus (Sulej 2005) and Postosuchus kirkpatricki (Weinbaum 2011), this anterior projection of the ventral process of the squamosal reaches the postorbital, dividing the infratemporal fenestra into two and forming an incipient accessory lateral temporal fenestra.
The posterior process of the squamosal of UFRGS-PV-0629-T forms two posterodorsal cavities divided by a shallow crest (Fig. 10). The dorsalmost cavity is smaller than the other one and accommodates the posterior end of the occipital process of the parietal, while the ventralmost cavity is larger and hosts the posterior process of the paroccipital process of the opisthotic. The posterior end of the posterior process of the squamosal also has a ventral concavity on its anteroventral surface in which the dorsal articular surface of the quadrate fits (“dorsal head” sensu Gower 1999).
Jugal (Fig. 13): Both jugals of UFRGS-PV-0629-T are preserved disarticulated from the skull, although the left one is preserved in articulation with the quadratojugal and ectopterygoid. The jugal is a thin, mediolaterally compressed element with three processes: the anterior, ascending, and posterior processes.
The lateral surface of the jugal body is convex and has a low anteroposteriorly directed crest, resembling that of Decuriasuchus quartacolonia (MCN-PV10.105a), Saurosuchus galilei (PVL 2062, PVSJ 32), and the UFRGS-PV-0156-T (Prestosuchus chiniquensis). The ventral margin is slightly concave and articulates with the ectopterygoid medially, at the base of the ascending and anterior processes.
The anterior process of the jugal of UFRGS-PV-0629-T articulates anteromedially with the posterodorsal margin of the maxilla and dorsally with the ventral process of the lacrimal through a dorsal laminar projection. As in Saurosuchus galilei (PVL-2062, PVSJ 32), Postosuchus kirkpatricki (Chatterjee 1985; Weinbaum 2011), and Decuriasuchus quartacolonia (França et al. 2013), this dorsal blade in UFRGS-PV-0629-T forms part of the anteroventral margin of the orbit.
The ascending process of the jugal is long, mediolaterally compressed, and tapers towards its dorsal end. All the anterior surface of this process forms the articular facet for the ventral process of the postorbital constituting the postorbital bar. It is directed posterodorsally, forming an angle of approximately 45° with the posterior process, as seen in Postosuchus kirkpatricki (Chatterjee 1985; Weinbaum 2011) and Decuriasuchus quartacolonia (França et al. 2013), contrasting with an approximately 80° angle seen in Batrachotomus kupferzellensis (SMNS 52970), Saurosuchus galilei (PVL-2062, PVSJ 32) and UFRGS-PV-0156-T (Prestosuchus chiniquensis).
The posterior process is the longest process of the jugal, it is laminar and extends laterally overlapping almost the entire extension of the anterior process of the quadratojugal. Thus, two processes form the ventral margin of the infratemporal fenestra: the posterior process of the jugal laterally and the anterior process of the quadratojugal medially. The ventral edge of the posterior process of the jugal projects posterodorsally, granting a wedge shape to its posterior end. This condition can also be seen in UFRGS-PV-0156-T (Prestosuchus chiniquensis).
Quadratojugal (Figs. 10A3, A4, 13, 14): The quadratojugals of UFRGS-PV-0629-T are V-shaped elements, with a main body that articulates posteromedially with the quadrate and bifurcates anteriorly into an anterior and an anterodorsal ascending process.
The anterior process articulates with the posterior process of the jugal dorsolaterally (Fig. 13), delimiting together the ventral margin of the infratemporal fenestra, resembling the condition of Batrachotomus kupferzellensis (Gower 1999), Decuriasuchus quartacolonia (MCN-PV10.105a), and UFRGS-PV-0156-T.
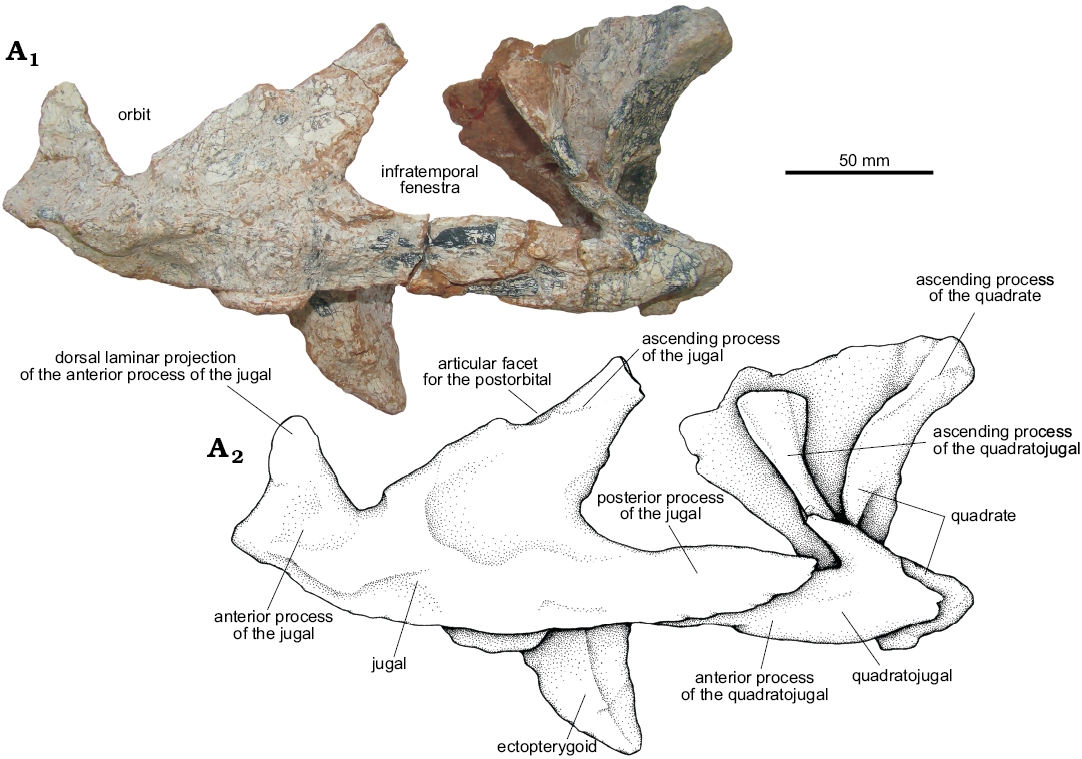
Fig. 13. The pseudosuchian archosaur Prestosuchus chiniquensis Huene, 1938 (UFRGS-PV-0629-T) from the Dinodontosaurus Assemblage Zone, Ladinian, Middle Triassic, Dona Francisca municipality, Rio Grande do Sul State, Brazil; left jugal, quadratojugal, and quadrate in lateral view. The articular condyle of the quadrate is broken. Photograph (A1) and explanatory drawing (A2).
The ascending process of the quadratojugal of UFRGS-PV-0629-T is long and laminar. Its posteromedial surface articulates with the anterolateral surface of the ascending process of the quadrate. A large quadrate foramen pierces through the contact area of these last two elements (Figs. 12, 14A1, A2). The medial surface of the dorsal end of the ascending process of the quadratojugal contacts the ventral process of the squamosal, delimiting the greater part (approximately three-quarters) of the posterior margin of the infratemporal fenestra (Fig. 10A3, A4).
Quadrate (Figs. 12–14): Both quadrates of UFRGS-PV-0629-T are preserved and only the distal condyles of the right one are broken. The quadrate is anterodorsally to posteroventrally oriented with its dorsal head articulating with the squamosal, its lateral surface with the quadratojugal, and its ventral condyles with the mandible.
The lateral and medial condyles are separated by a well-defined groove; the shape and size of the condyles have slight differences. The medial condyle is slightly larger and more oval in shape; its medial end has a more anterior position whereas the lateral end is more posterior. The lateral condyle is more subrectangular in shape and its position is more transverse. The medial condyle projects slightly more ventrally than the lateral condyle, and both condyles of the quadrate extend ventrally below the level of the quadratojugal, as also seen in Batrachotomus kupferzellensis (SMNS 80260), Saurosuchus galilei (PVSJ 32), and Decuriasuchus quartacolonia (MCN-PV10.004, MCN-PV10.105a).
From these condyles, the columnar body of the quadrate extends dorsally and expands on its proximal end forming a convex articular surface that fits into a ventral concavity of the squamosal (the “dorsal head” sensu Gower 1999) (Fig. 14). From this column, the quadrate has two distinctive anterior projections. The anteromedial projection corresponds to the pterygoid process that articulates with the quadrate process of the pterygoid. It is the largest of these projections; it is thin and triangular in medial view. The anterolateral projection corresponds to the articulation with the ascending process of the quadratojugal. The lateral margin is straight, but is interrupted on its ventral half by the quadrate foramen, which can also be recognized in the specimen UFRGS-PV-0156-T, Batrachotomus kupferzellensis (SMNS 52970, Gower 1999), Saurosuchus galilei (PVSJ 32, Alcober 2000), Polonosuchus silesiacus (Sulej 2005), Postosuchus kirkpatricki (Long and Murry 1995; Weinbaum 2011), and Decuriasuchus quartacolonia (MCN-PV 10004, França et al. 2013).
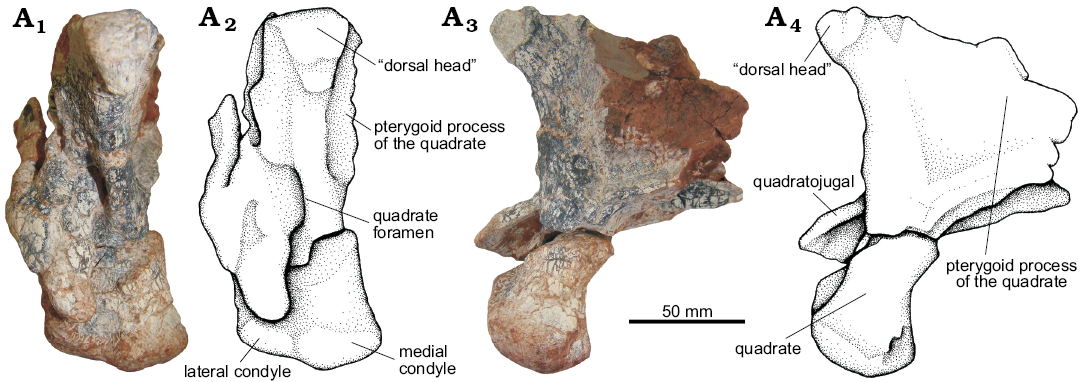
Fig. 14. The pseudosuchian archosaur Prestosuchus chiniquensis Huene, 1938 (UFRGS-PV-0629-T) from the Dinodontosaurus Assemblage Zone, Ladinian, Middle Triassic, Dona Francisca municipality, Rio Grande do Sul State, Brazil; left quadratojugal and quadrate in posterior (A1, A2) and medial (A3, A4) views. Photographs (A1, A3) and explanatory drawings (A2, A4).
Palatal complex: Vomer (Fig. 5A3, A4): The vomers of UFRGS-PV-0629-T are not preserved in their natural position, but are attached to different elements of the skull. The left one is preserved attached to the medial surface of the left maxilla (Fig. 5A3, A4), while the right one was originally preserved next to the left pterygoid, but was later removed during preparation. The vomer of UFRGS-PV-0629-T has no teeth, it is a laminar and anteroposteriorly elongated bone that would contact the anterior process of the pterygoid posteromedially and the palatine posterolaterally. The anterior region of the vomer contacts its counterpart medially and apparently both the maxilla and the premaxilla laterally, differing from the condition seen in Saurosuchus galilei (PVSJ 32; Alcober 2000) and Postosuchus kirkpatricki (Weinbaum 2011: 34) in which the vomer reaches the maxilla laterally, but not the premaxilla.
Palatine (Figs. 5B, 15A): The palatines of UFRGS-PV-0629-T are laminar, subrectangular, and edentulous elements that were not preserved in their original position. The left palatine was found isolated, while the right one was attached flat to the medial surface of the right maxilla, displaced from its natural position. The palatine would articulate with the vomer anterolaterally, the pterygoid medially, and the maxilla laterally. The anterior margin of the main body of the palatine has two anterior projections, a medial one and a lateral one, that delimit the posterior half of the choana, while the posterior margin of the palatine delimits the anterior margin of the suborbital fenestra. The anterior portion is thicker than the posterior one, as was also described for Saurosuchus galilei (PVSJ 32; Alcober 2000).
In general terms, the palatine of UFRGS-PV-0629-T is very similar to that of other loricatans such as Saurosuchus galilei (PVSJ 32; Alcober 2000), Batrachotomus kupferzellensis (Gower 1999), Polonosuchus silesiacus (Sulej 2005), and Decuriasuchus quartacolonia (França et al. 2013).
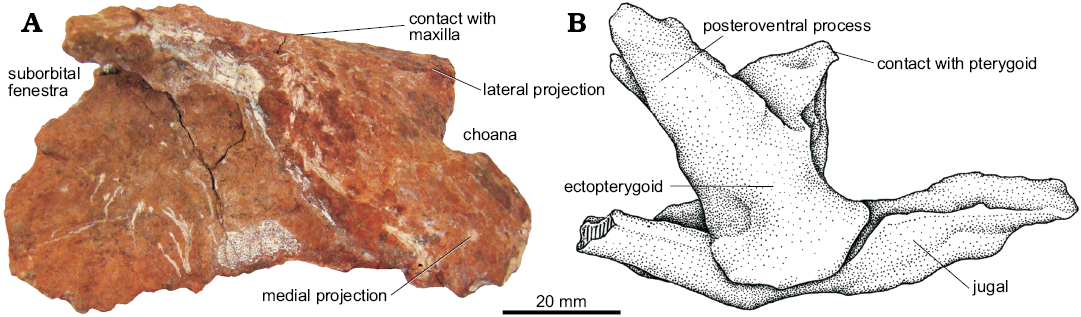
Fig. 15. The pseudosuchian archosaur Prestosuchus chiniquensis Huene, 1938 (UFRGS-PV-0629-T) from the Dinodontosaurus Assemblage Zone, Ladinian, Middle Triassic, Dona Francisca municipality, Rio Grande do Sul State, Brazil. A. Photograph of the left palatine in dorsal view. B. Explanatory drawing of the left ectopterygoid articulated with the jugal in ventral view.
Pterygoid (Fig. 16): The left pterygoid of UFRGS-PV-0629-T is preserved complete and isolated, while the right one is still articulated to the skull on its natural position, but its anterior process is incomplete. This is the first specimen that allows the detailed study of the pterygoid because in other known specimens of Prestosuchus chiniquensis since the palate is either covered by the occluded mandibles (UFRGS-PV-0156-T), or not preserved (CPEZ-239b, lectotype and paralectotype).
The pterygoid forms the largest part of the palatal complex and contacts the vomer anteriorly, the palatine laterally, the ectopterygoid posterolaterally, and the quadrate posteromedially. The pterygoid is an edentulous, elongated, and triradiate bone with an anterior palatal process, a posterodorsal quadrate process, and a lateroventral process (=ectopterygoid wing) that articulates with the ectopterygoid. It also contacts the basipterygoid through a concave articular surface that seems to be akinetic.
The palatal process of UFRGS-PV-0629-T is a laminar, concave structure that slightly curves dorsomedially reaching an almost vertical orientation at the midline. Its anterior end tapers towards its contact with the vomer (Fig. 16A1, A2).
The lateroventral process is more robust than the others and articulates with the ectopterygoid through a long suture that extend up to the suborbital fenestra. This process forms a 90° angle with the palatal process resembling the condition of Batrachotomus kupferzellensis (SMNS 80260), Saurosuchus galilei (PVSJ 32), and Decuriasuchus quartacolonia (França et al. 2013). This differs from Postosuchus kirkpatricki (Chatterjee 1985; Weinbaum 2011) and Polonosuchus silesiacus (ZPAL AbIII-563), in which this angle is wider, and probably in Rauisuchus tiradentes (Lautenschlager and Rauhut 2015) whose lateroventral process is not completely preserved but appears to form a wider angle like the latter taxa.
The quadrate process of the pterygoid of UFRGS-PV-0629-T is laminar and has two distinct projections that grant it a Y-shape form. This process is elongate and overlaps the quadrate laterally. On the anteriormost part of the quadrate process, the articular facet for the basipterygoid process of the parabasisphenoid can be identified on its anteromedial surface (Fig. 16).
The pterygoids of UFRGS-PV-0629-T do not exhibit any diagnostic features, and their morphology is very similar to that of other loricatans with known preserved pterygoids (Rauisuchus tiradentes, Postosuchus kirkpatricki, Batrachotomus kupferzellensis, Saurosuchus galilei, Polonosuchus silesiacus, and Decuriasuchus quartacolonia).
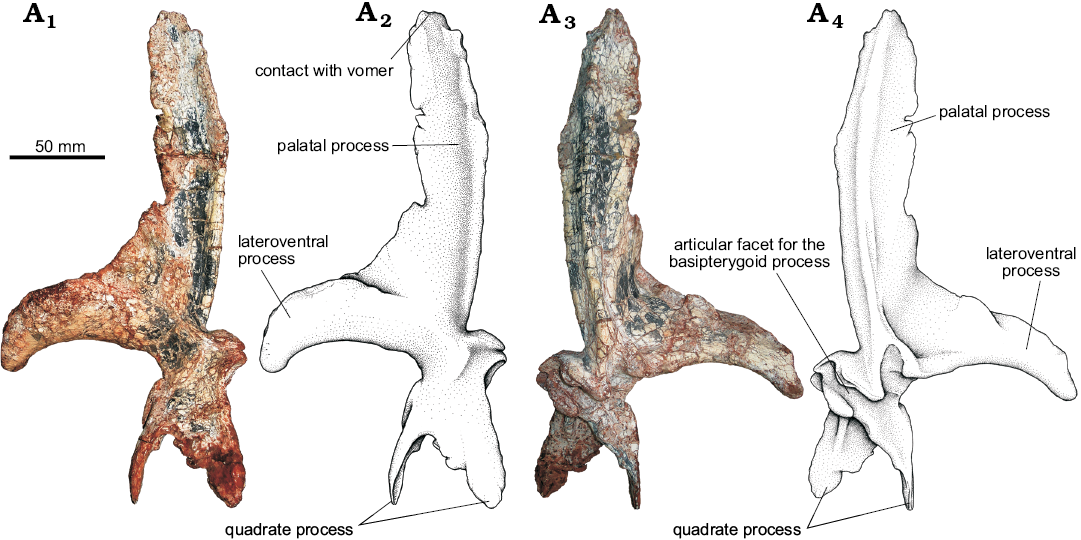
Fig. 16. The pseudosuchian archosaur Prestosuchus chiniquensis Huene, 1938 (UFRGS-PV-0629-T) from the Dinodontosaurus Assemblage Zone, Ladinian, Middle Triassic, Dona Francisca municipality, Rio Grande do Sul State, Brazil; left pterygoid in dorsal (A1, A2) and ventral (A3, A4) views. Photographs (A1, A3) and explanatory drawings (A2, A4).
Ectopterygoid (Fig. 15B): Only the left ectopterygoid of UFRGS-PV-0629-T is preserved and it is articulated in its natural position with the corresponding jugal. This is the only ectopterygoid known for Prestosuchus chiniquensis, since in other specimens it cannot be observed or it was not preserved at all (UFRGS-PV-0156-T, CPEZ-239b).
The ectopterygoid is a robust, columnar element that articulates with the jugal laterally and projects medially to contact the pterygoid. It has a long posteroventral process that articulates with the ventrolateral margin of the pterygoid as can also be seen in Postosuchus kirkpatricki (Long and Murry 1995; Weinbaum 2011), Batrachotomus kupferzellensis (SMNS 80260; Gower 1999), Saurosuchus galilei (PVSJ 32; Alcober 2000), and Decuriasuchus quartacolonia (MCN-PV 10105a; França et al. 2013). In lateral view, this process has a triangular shape with its most acute vertex at the posteroventral corner.
The ectopterygoid of UFRGS-PV-0629-T is more robust, differing from Postosuchus kirkpatricki (TTU-P 9000, TTU-P 9002; Nesbitt 2011) and Batrachotomus kupferzellensis (Gower 1999) in which this bone is more elongated.
On the UFRGS-PV-0629-T, it is not possible to determine whether the ectopterygoid has one articular facet for the jugal as in Saurosuchus galilei (Alcober 2000) and Luperosuchus fractus (Nesbitt and Desojo 2017), or two articular facets as Postosuchus kirkpatricki (TTU-P 9000, TTU-P 9002; Nesbitt 2011), Batrachotomus kupferzellensis (Gower 1999), and allegedly Rauisuchus tiradentes (Lautenschlager and Rauhut 2015), because this region is disarticulated and covered by carbonatic concretion.
Mandible (Figs. 17, 18): The UFRGS-PV-0629-T has both hemimandibles preserved; the left one has all the elements that comprise it completely articulated, while the right one preserves the dentary, splenial, and articular disarticulated from the rest of the mandible. The hemimandibles of UFRGS-PV-0629-T are elongated, being 4.66 times longer than high.
In lateral view, the anterior half of the mandible is represented by the dentary and the posterior half is occupied by the angular, surangular, and articular (Fig. 18A1, A2). In medial view, the dentary, splenial, coronoid, angular, prearticular, and articular can be identified. On the medial surface, the dentary is only partially visible at its anteriormost end because the splenial overlaps it, extending from the level of the second dentary tooth to the posterior end of the dentary (Fig. 17). Moreover, the symphyseal region is poorly defined on the medial surface of the dentary, with only a small region at the anterior end of the dentary that appears to indicate the attachment area of two ligaments that join the hemimandibles together. This feature can also be seen in other basal loricatans such as Batrachotomus kupferzellensis (SMNS 80260, Gower 1999), Saurosuchus galilei (PVSJ 32, Alcober 2000), and Postosuchus kirkpatricki (Weinbaum 2011).
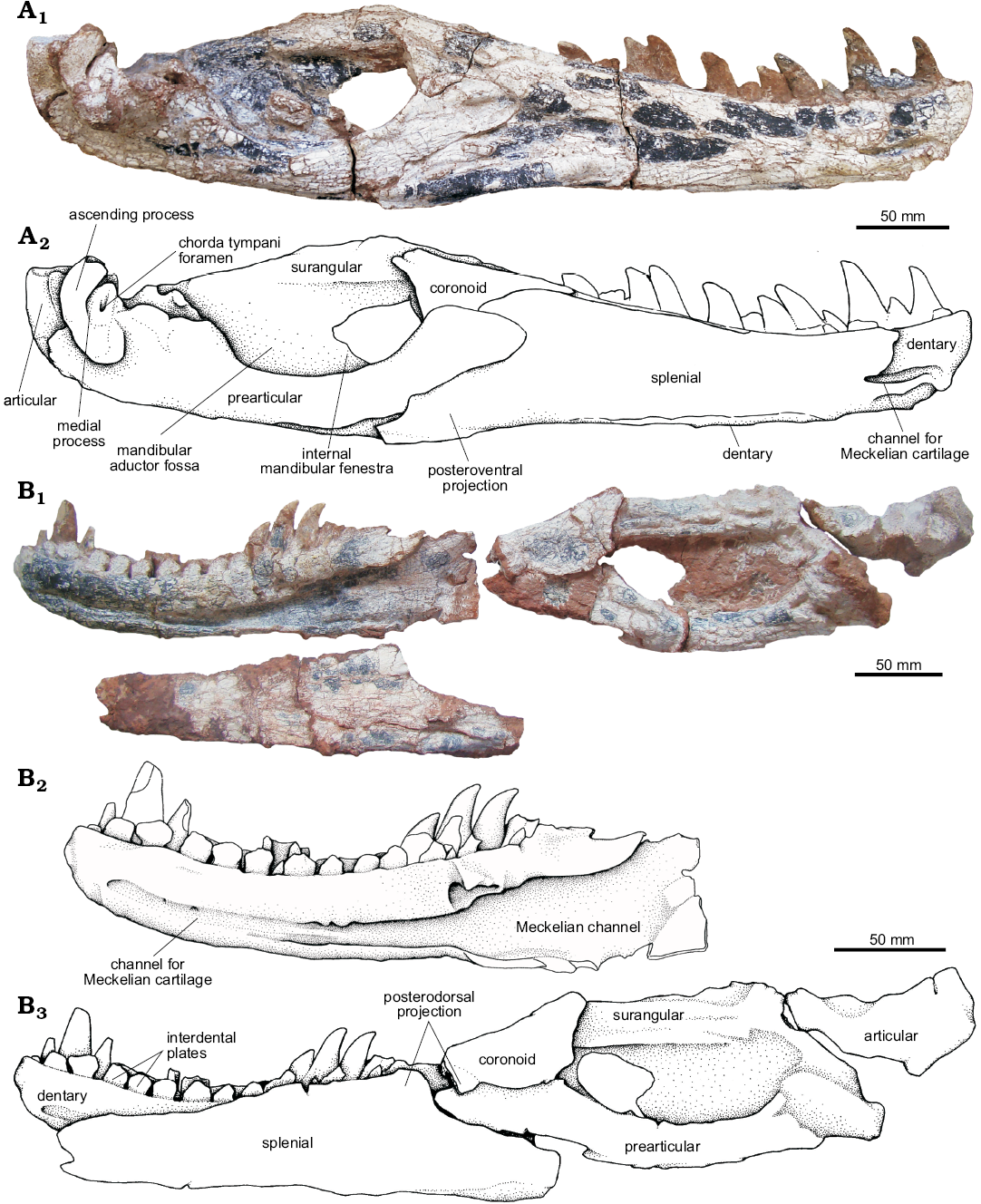
Fig. 17. The pseudosuchian archosaur Prestosuchus chiniquensis Huene, 1938 (UFRGS-PV-0629-T) from the Dinodontosaurus Assemblage Zone, Ladinian, Middle Triassic, Dona Francisca municipality, Rio Grande do Sul State, Brazil; medial view of the left (A) and right (B) mandibles. Photographs (A1, B1) and explanatory drawings (A2, B2, B3).
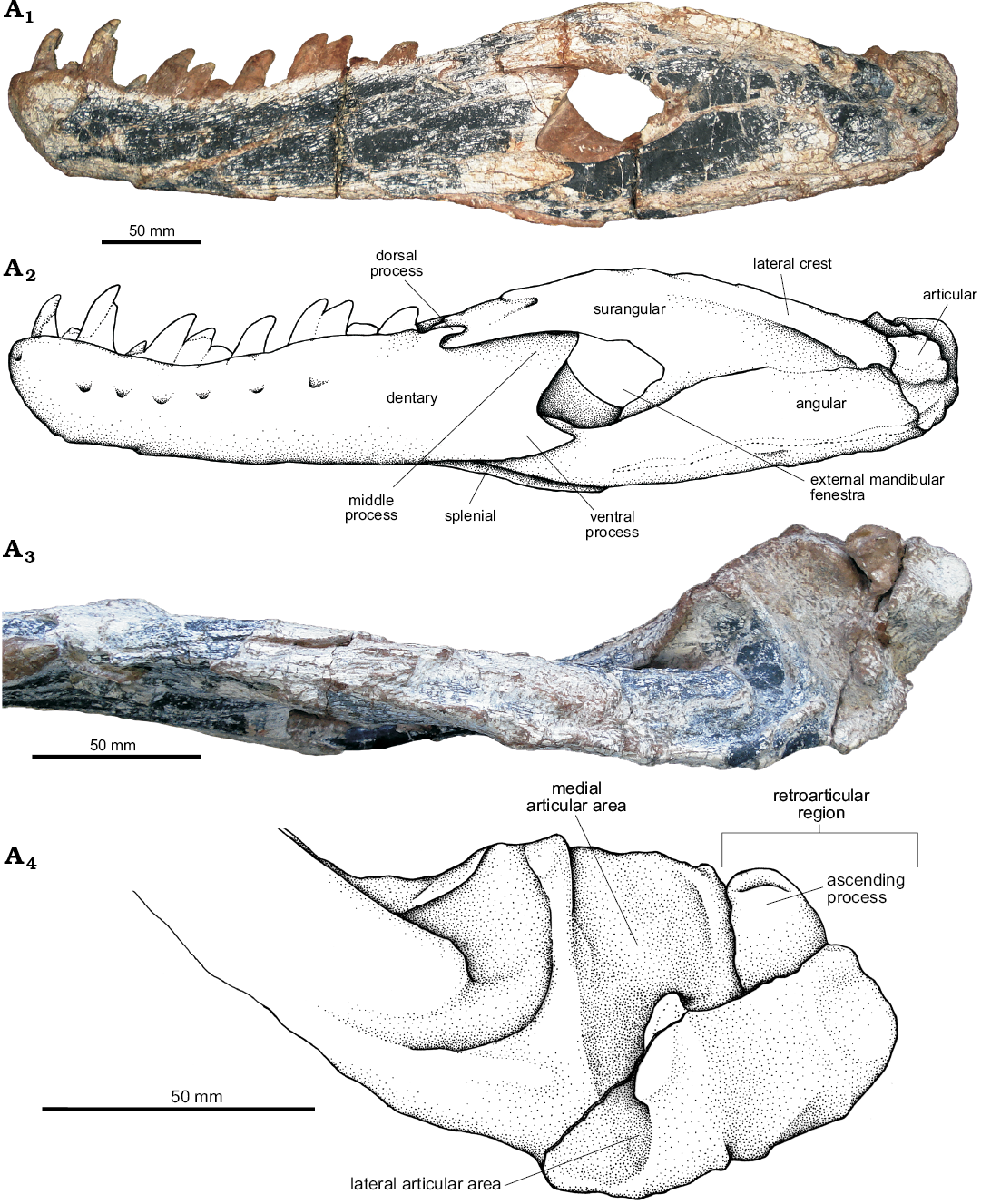
Fig. 18. The pseudosuchian archosaur Prestosuchus chiniquensis Huene, 1938 (UFRGS-PV-0629-T) from the Dinodontosaurus Assemblage Zone, Ladinian, Middle Triassic, Dona Francisca municipality, Rio Grande do Sul State, Brazil. Left mandible of in lateral view (A1, A2). Detail of the articular and retroarticular areas in dorsal view (A3, A4). Photographs (A1, A3) and explanatory drawings (A2, A4).
Dentary (Figs. 17, 18): Both dentaries of UFRGS-PV-0629-T are well preserved; they are elongated and have a slight dorsoventral expansion at the anterior end. On each dentary, 14 alveoli can be clearly identified, with 12 teeth in different eruption stages occupying the alveoli of the left dentary and only eight present in the alveoli of the right one. The dentary teeth are posteriorly curved, labiolingually compressed and serrated with three denticles per millimeter. The different eruption stages of the dentary teeth appear to reflect the wavy eruption pattern typical of reptiles (Edmund 1962, 1969), but unfortunately this pattern cannot be recognized in the maxillae to confirm this because the preserved teeth are separated by several missing ones. The lectotype of Prestosuchus chiniquensis also has 14 alveoli with 11 teeth preserved, while the alveoli and dentary teeth of the specimen UFRGS-PV-0156-T cannot be determined because the mandibles are preserved occluded. The tooth count is barely higher than on other loricatans since Fasolasuchus tenax (PVL 3851; Bonaparte 1981) has 13 alveoli on its dentary, and Batrachotomus kupferzellensis (Gower 1999) was described to have 11 or 12 alveoli. The dentary tooth series was not preserved complete in Polonosuchus silesiacus (Sulej 2005), Saurosuchus galilei (PVSJ 32; Alcober 2000) nor in Decuriasuchus quartacolonia (França et al. 2013). On the other hand, Postosuchus kirkpatricki probably has 15–16 teeth in the dentary of the holotype TTUP 9000 (Jonathan Weinbaum, personal communication 2018).
The lateral surface of the dentary of UFRGS-PV-0629-T is slightly convex, and this convexity is more pronounced on the anterior portion. This lateral surface has nine nutrient foramina, distributed in an almost straight line from the anterior end of the dentary up to the level of the eleventh mandibular teeth (Fig. 18A1, A2).
On the medial surface of the dentary, the canal for the Meckelian cartilage can be identified extending up to the level of the third dentary tooth (Fig. 17). Most of the medial surface of the dentary is covered by the splenial that overlaps it.
The ventral margin of the dentary is straight as can also be seen in the lectotype BSPG AS XXV 1 and UFRGS-PV-0156-T of Prestosuchus chiniquensis and in Batrachotomus kupferzellensis (SMNS 80260), contrasting with Saurosuchus galilei (PVSJ 32) and Decuriasuchus quartacolonia (França et al. 2013), whose dentaries have a slightly convex ventral margin or Postosuchus kirkpatricki (Weinbaum 2011) that has a sinuous ventral margin.
In lateral view, the dorsal margin of the dentary is slightly converging anteriorly with its ventral margin up to the level of the third dentary tooth, where it diverges forming the subtle dorsoventral expansion of the anterior end of the dentary. This dorsoventral expansion is much more pronounced on the alveolar margin than on the ventral margin of the dentary. The dorsoventral expansion of the dentary is also registered in Postosuchus kirkpatricki (Long and Murry 1995; Weinbaum 2011) and Batrachotomus kupferzellensis (SMNS 80260; Gower 1999), contrasting with Saurosuchus galilei (PVSJ 32; Alcober 2000) whose dentary does not expand anteriorly at all.
The posterior end of the dentary has three projections that taper posteriorly. The dorsal process is a small projection that fits into a slot of the surangular. The middle process is dorsally overlapped by the surangular in a straight and horizontal contact. The rounded ventral portion of this process delimits the anterior margin of the external mandibular fenestra. The ventral process is arrow-shaped and overlaps the angular, delimiting the anteroventral margin of the external mandibular fenestra (Fig. 18A3, A4).
Splenial (Fig. 17): The splenial of UFRGS-PV-0629-T is a laminar, elongated element that constitutes most of the medial surface of the anterior half of the hemimandible. It medially overlaps the dentary forming the medial wall of the Meckelian canal. The medial surface of the main body of the splenial contacts the dentary anteriorly, ventrally, and dorsally and extends anteriorly up to the level of the fourth dentary tooth. From this, a slender anterior projection of the splenial extends up to the level of the second dentary tooth. However, it is not possible to determine whether there is participation of the splenial in the mandibular symphysis or not (Fig. 17). The participation of the splenial in the mandibular symphisis in not very clear in basal loricatans because in Saurosuchus galilei (PVSJ 32; Alcober 2000) the anterior end of the splenial is broken beyond the level of the third dentary tooth, and in Postosuchus kirkpatricki (Weinbaum 2011) and Batrachotomus kupferzellensis (Gower 1999) the splenials preserved are fragmentary. Ventrally, the dentary and the splenial are separated by a deep anteroposteriorly elongated groove as seen in the lectotype of Prestosuchus chiniquensis (BSPG AS XXV 1) and Decuriasuchus quartacolonia (MCN-PV10.105a).
The posterior end of the splenial has two projections, a posterodorsal one and a posteroventral one. A notch is formed between these projections where the prearticular fits, as also seen in Decuriasuchus quartacolonia (MCN-PV10.105a). The posterodorsal projection of the splenial (=coronoid process) is short and tapers posteriorly, with its posterodorsal margin suturing to the coronoid and its ventral margin contacting the anterodorsal end of the angular. The posteroventral process (=angular process) is long and extends posteriorly, reaching the angular at the level of the first quarter of the internal mandibular fenestra (Fig. 17A). The splenial is sutured to the angular along its entire extent and is excluded by it from the mandibular fenestra.
Coronoid (Fig. 17): Both coronoids of UFRGS-PV-0629-T are preserved. They are laminar, subrectangular bones, located at the posterior margin of the last alveoli and the internal mandibular fenestra.
The anterior margin of the coronoid overlaps the dentary at a small area of approximately 10 mm long. The anterior half of the ventral margin of the coronoid contacts the splenial at a suture of 40 mm long and the posterior half contacts the prearticular at a suture of 32 mm long. The posterior border of the coronoid is about 30 mm high and forms the dorsal half of the anterior margin of the aductor fossa.
According to Gower (1999), some archosauriform taxa (e.g., erythrosuchids, Sphenosuchus acutus, some mesoeucrocodylian crocodylomorphs, and probably the proterochampsid Chanaresuchus bonapartei) have long and thin coronoids, which could suggest the presence of the same element in Batrachotomus kupferzellensis, although its actual shape is not clear. The presence of a coronoid was described for Saurosuchus galilei (PVSJ 32; Alcober 2000), indicated by a laminar bone located parallel and dorsal to the splenial, and extending anteriorly up to the third mandibular tooth (Alcober 2000: fig 10). This condition differs quite from that of UFRGS-PV-0629-T because in the latter the coronoid is more posteriorly located and therefore does not reach the anterior alveoli as in Saurosuchus galilei (PVSJ 32; Alcober 2000). On the contrary, the coronoid described by Chatterjee (1985) for Postosuchus kirkpatricki was identified as a small bone restricted to a small area between the prearticular, surangular, and the internal mandibular fenestra, which resembles the condition of UFRGS-PV-0629-T. However, this bone was not identified in the redescription of Postosuchus kirkpatricki carried out by Weinbaum (2011). In Decuriasuchus quartacolonia (MCN-PV10.105a) the coronoid is only partially visible because it is mostly covered by incrustation, but resembles the position seen in UFRGS-PV-0629-T, although its anterior extension cannot be fully determined.
Angular (Fig. 18A1, A2): The angular of UFRGS-PV-0629-T is a slender, elongated bone that delimits the ventral margin of the external mandibular fenestra. Medially, it forms the floor of the mandibular adductor fossa and the posterior part of the Meckelian canal. It sutures dorsally to the surangular, posteriorly and ventromedially to the articular, and anteriorly to the dentary with the splenial fitting between the two of them.
The angular of the specimen UFRGS-PV-0629-T has a subtly convex external surface as in specimen UFRGS-PV-0156-T (Prestosuchus chiniquensis) and Decuriasuchus quartacolonia (França et al. 2013), differing from Postosuchus kirkpatricki (Chatterjee 1985; Weinbaum 2011) in which the angular has a thickened and rugose ventral surface that is shorter than in Prestosuchus chiniquensis. According to Gower (1999), the angular of Batrachotomus kupferzellensis is known from only a single, incomplete, and uncertainly identified fragment from the left side of SMNS 80260.
Surangular (Figs. 17, 18): The surangular is preserved on both hemimandibles. It is an elongated bone constituted by a main body and an anterior process. It forms the dorsal and posterior margins of the external and internal mandibular fenestra, and together with the articular, prearticular and angular it constitutes the posterior half of the hemimandible. The surangular of UFRGS-PV-0629-T is complete, unlike that of the specimens BSPG AS XXV 1, which is fragmentary, and UFRGS-PV-0156-T , which cannot be observed in detail because of its poor preservation.
The main body of the surangular of UFRGS-PV-0629-T is laminar and forms the lateral wall of the mandibular adductor fossa (Figs. 17, 18). The ventral half of the lateral side is overlapped by the posteromedial surface of the angular. The surangular contacts the articular posteriorly, contributing a small portion to the articular margin of the glenoid fossa and posteriorly to the retroarticular region of the mandible, as also seen in Polonosuchus silesiacus (Sulej 2005).
The dorsal margin of the surangular of UFRGS-PV-0629-T has a marked crest (= surangular shelf) that projects laterally and extends from the anterior process of this bone up to its contact with the articular, a condition that can also be seen in Decuriasuchus quartacolonia (França et al. 2013). Ventral to the surangular shelf, there is a slightly excavated area on the lateral surface of the surangular, probably associated with the insertion of the M. adductor mandibulae externus (Sampson and Witmer 2007). The dorsomedial margin of the surangular of UFRGS-PV-0629-T has a smaller crest that runs along the anteromedial surface of the anterior process of the surangular (Fig. 18A1, A2).
The anterior process of the surangular projects anteriorly from the main body and overlaps the lateral surface of the coronoid, forming a notch for the articulation of the dorsal process of the dentary. The medial surface of the anterior process of the surangular has a depression that resembles that seen in Batrachotomus kupferzellensis (SMNS 52970; Gower 1999), which hosts the tapering end of the medial process of the dentary.
Prearticular (Fig. 17): The prearticular of UFRGS-PV-0629-T is anteroposteriorly elongated and its anterior region is curved projecting anterodorsally to fit in a notch on the splenial, delimiting the ventromedial margin of the internal mandibular fenestra (Fig. 17). It also articulates with the articular posteromedialy, although this suture is not clearly defined. An articular facet for the surangular can be recognized on its posterolateral surface and another facet for the angular on its ventral surface.
The dorsal surface of the prearticular is strongly concave and constitutes the ventromedial surface of the mandibular adductor fossa, as can be seen in Batrachotomus kupferzellensis (SMNS 80260; Gower 1999) and Postosuchus kirkpatricki (Long and Murry 1995; Weinbaum 2011). In Postosuchus alisonae (Peyer et al. 2008) the specimen is broken at that point (Jonathan Weinbaum personal communication 2018). The prearticular of UFRGS-PV-0629-T has a well defined lateral furrow for the Meckelian cartilage.
Articular (Figs. 17A, 18): The articular of UFRGS-PV-0629-T is a robust element that forms the posterior end of the hemimandibles. It sutures with the prearticular anteromedially, and the angular and surangular anterolaterally. The articular of both hemimandibles of UFRGS-PV-0629-T is preserved, although the right one is disarticulated.
The glenoid fossa of the articular is mediolaterally elongated and has two concave areas separated by a thin elevated surface. This disposition mirrors the articular surface of the quadrate, favoring an efficient articulation between the skull and the mandibles. The posterior end of the articular, posterior to the glenoid fossa, is represented by the retroarticular region (Fig. 18A3, A4).
The retroarticular portion of the articular has an ascending and a medial process (Figs. 17A, 18A3, A4). The ascending process is an elongated, almost dorsoventrally oriented, and laterally compressed structure that projects slightly anterodorsally. The medial process is ventromedially projected and is pierced by a foramen (“chorda tympani foramen”) as that seen in Batrachotomus kupferzellensis (SMNS 80260; Gower 1999), Rauisuchus tiradentes (Lautenschlager and Rauhut 2015), Postosuchus kirkpatricki (Chatterjee 1985; Weinbaum 2011), and Decuriasuchus quartacolonia (França et al. 2013). It differs from that of Fasolasuchus tenax (PVL 3850), in which the medial process is apparently not pierced by a foramen.
Cranial endocast (Fig. 19): The endocranial cavity of UFRGS-PV-0629-T was studied based on a digital reconstruction developed from CT scan images.
The digital endocast obtained is high, elongated and sinuous in lateral view, with a flexure between the forebrain and the midbrain of approximately 115° (Fig. 19). The olfactory bulbs can be clearly identified; they are elongate, separated by a deep furrow and expanded laterally, being three times wider than the olfactory tract. The latter is almost as long as the rest of the encephalon. The cerebral hemispheres are 1.2 times longer than wide and slightly expanded posteriorly, doubling their anterior width. On the posterodorsal region of the encephalon, the endocast has a dorsal projection similar to that described in some phytosaurs (e.g., Machaeroprosopus mccauleyi, Machaeroprosopus pristinus, Smilosuchus gregori; Holloway et al. 2013; Case 1928). This dorsal expansion has been interpreted either as a pineal expansion corresponding to the epiphysis, as a dural venous sinus or even as a paratympanic sinus (Lautenschlager and Butler 2016). Here, it is interpreted as the dural venous sinus because of it anteroposterior expansion, in agreement with the discussion of Lautenschlager and Butler (2016). On the ventral surface of the endocast, the hypophysis can be recognized as a dorsoventrally long and anteroposteriorly short ventral projection.
A few cranial nerves could be identified on the endocast, namely the cranial nerve II (olfactory tract) and the cranial nerves V and XII. The trigeminal nerve (cranial nerve V) appears as a single anterolateral projection where the midbrain and the hindbrain meet. It exits through a single foramen that can be seen on the lateral surface of the prootic (see Mastrantonio et al. 2013: fig. 8). The hypoglossal nerve (cranial nerve XII) can be identified on the posterolateral end of the posterior brain, but only the left exit through the exoccipital could be reconstructed (see Mastrantonio et al. 2013: fig. 7).
Stratigraphic and geographic range.—Ladinian, Middle Triassic; Dinodontosaurus Assemblage Zone; Pinheiros-Chiniquá Sequence, Santa Maria Supersequence; Rio Grande do Sul State, Brazil.
Fig. 19. The pseudosuchian archosaur Prestosuchus chiniquensis Huene, 1938 (UFRGS-PV-0629-T) from the Dinodontosaurus Assemblage Zone, Ladinian, Middle Triassic, Dona Francisca municipality, Rio Grande do Sul State, Brazil. A. Digital model of the braincase in right lateral (A1) and dorsal (A2) views, with endocast (blue) placed in its natural position. B. Digital endocast in right (B1) and left (B2) lateral and dorsal (B3) views. Not to scale.
Discussion
Paleoneuroanatomy of Prestosuchus and its implications.—The general morphology of the endocast of Prestosuchus chiniquensis (UFRGS-PV-0629-T) resembles that of theropod dinosaurs, especially due to the marked flexure between the forebrain and midbrain (approximately 115°) which differs from the wider angle (130–150°) seen in the rest of the pseudosuchians (e.g., phytosaurs, ornithosuchids, aetosaurs, and crocodylomorphs) (Baczko et al. 2018), and also a single exit for the cranial nerve XII (see below). However, the olfactory region is very similar to that of other carnivorous pseudosuchians such as phytosaurs (e.g., Parasuchus angustifrons, Ebrachosuchus neukami, Machaeroprosopus mccauleyi), the erpetosuchid Parringtonia gracilis, and most crocodylomorphs (e.g., Sebecus icaeorhinus, Caiman yacare, Alligator mississippiensis). On the other hand, this condition contrasts with that of the aetosaur Desmatosuchus spurensis (UMMP 7476), the crocodylomorph Simosuchus clarki (Kley et al. 2010), and sauropod and ornithischian dinosaurs (e.g., Diplodocus longus, Hypacrosaurus altispinus, Stegoceras validum, Euoplocephalus tutus) (Witmer et al. 2008; Evans et al. 2009; Miyashita et al. 2011; Stocker et al. 2016) in which the olfactory tracts are short and the olfactory bulbs are approximately as long as wide.
The condition of a single passage for the trigeminal nerve of UFRGS-PV-0629-T is shared with most non-archosaurian archosauriforms, pseudosuchians (aetosaurs, ornithosuchids, the loricatan Postosuchus kirkpatricki, and crocodylomorphs) excepting phytosaurs, and with sauropod and ornithischian dinosaurs (Hopson 1979; Weinbaum 2011; Baczko and Desojo 2016; Lautenschlager and Butler 2016; Baczko et al. 2018). It differs from the condition of some pterosaurs and theropod dinosaurs in which the ophthalmic branch (cranial nerve V1) and the combined canal for the maxillary (cranial nerve V2) and mandibular (cranial nerve V3) branches split inside the endocranial cavity and exit through separate passages (Witmer et al. 2003, 2008). The single passage for the cranial nerve XII seen in UFRGS-PV-0629-T is also shared with some archosauriforms, phytosaurs, pterosaurs, and theropod dinosaurs (e.g, Euparkeria capensis, Erythrosuchus africanus, Ebrachosuchus neukami, Allkaruen koi, Tyranosaurus rex, Giganotosaurus carolini) (Gower and Sennikov 1996; Witmer et al. 2008; Paulina Carabajal and Canale 2010; Codorniú et al. 2016; Lautenschlager and Butler 2016; Sobral et al. 2016). This condition differs from the multiple passages for the cranial nerve XII seen in crocodylomorphs (e.g., Sebecus icaeorhinus, Simosuchus clarki, Caiman yacare, Alligator mississippiensis, Gavialis gangeticus) and sauropodomorphs (e.g., Diplodocus longus, Saturnalia tupiniquim) (Hopson 1979; Kley et al. 2010; Bona et al. 2017; Witmer et al. 2008; Bronzati et al. 2017).
The comparison of the cranial endocast of Prestosuchus chiniquensis (UFRGS-PV-0629-T) with that of other archosaur reconstructions previously published demonstrates a susbtantial morphological variability among Archosauria. The paleoneuroanatomy of UFRGS-PV-0629-T differs with most of the pseudosuchian taxa in the overall curvature of its encephalon, but resembles them in the shape of their olfactory region (long tracts and elliptic bulbs). The endocranial reconstruction studied here supplements the osteological description of the braincase of UFRGS-PV-0629-T carried out by Mastrantonio et al. 2013.
About the presence of an opening between the premaxilla and the maxilla.—The presence of an opening and/or articulation between the premaxilla and maxilla in basal Loricata was mentioned and described as a characteristic of “rauisuchians” (e.g., Krebs 1976; Alcober 2000), and even used as a synapomorphy of the group in some phylogenetic analyses (e.g., Benton 1984; Benton and Clark 1988; Parrish 1993; Brusatte et al. 2010; Nesbitt 2011; Butler et al. 2014; Nesbitt and Desojo 2017). However, this gap between those bones varies in its relative position and size among different taxa and also can change between one side and the other of the same skull (Nesbitt and Desojo 2017).
Another problem regarding this structure is that this opening has been named differently across the literature. It has been referred to as: “slit-like opening” (Romer 1971), “accessory antorbital fenestra” (Sill 1974, Dawley et al. 1979, Long and Murry 1995), “supplementary fenestra” (Benton 1984, 1986), and “subnarial fenestra” (Chatterjee 1985; Galton 1985; Parrish 1993; Alcober 2000; França et al. 2013). Gower (1999) described this opening in Batrachotomus kupferzellensis as “a notch in the anterior margin of the maxilla that represents the anterior terminus of a short groove that has a posterolaterally-anteriomedially aligned long axis, with two small foramina lying in this groove just posterior to the notch”. According to Brusatte et al. (2009), this notch in Batrachotomus kupferzellensis would be a different, but homologous, condition regarding that of the other rauisuchians, while the presence and morphology of an opening could not be reliably assessed in Teratosaurus suevicus (Galton 1985) and Rauisuchus tiradentes (Lautenschlager and Rauhut 2015), since the premaxilla was not preserved in these taxa. In Polonosuchus (= Teratosaurus) silesiacus (Sulej 2005), the area of the articulation is smooth and rounded and the anterior border of the maxilla reaches over the premaxilla in dorsal view and forms a loose, potentially movable joint, but this connection would be less movable in T. suevicus than in Polonosuchus silesiacus (Brusatte et al. 2009).
Among basal Loricata, only Fasolasuchus tenax (Bonaparte 1981) was described as having no fenestra. More recently, Roberto-Da-Silva et al. (2016) mentioned the presence of an opening, which was interpreted as a “subnarial foramen”, in another specimen of Prestosuchus chiniquensis (ULBRA-PVT-281). Two hypotheses are in dispute about the function of this opening, according to Gower (2000). The first would be related to the air sinus system and the second to either blood vessels or nerve transmission. However, no evidence has been provided to corroborate either yet. The homology of this opening was discussed by several authors in the last twenty-five years (see Nesbitt 2011; Nesbitt and Desojo 2017). For example, Sereno and Novas (1993) have described both a “subnarial foramen” and a “premaxilla-maxilla fenestra” in Herrerasaurus ischigualastensis (Sereno and Novas 1993: fig. 2), “with the former opening below the naris and, internally, below the premaxillary palate, and the latter opening behind the naris and, internally, above the premaxillary palate” (Gower 2000: 458). The same authors interpreted the premaxilla-maxilla fenestra as an autapomorphy of H. ischigualastensis, and the subnarial foramen as a synapomorphy of H. ischigualastensis + saurischian dinosaurs that apparently evolved convergently in Saurosuchus galilei (Gower 2000). Recently, Nesbitt and Desojo (2017) analysed the presence of a slit-like gap between the posterodorsal process of the premaxilla and the maxilla in Saurosuchus galilei, Luperosuchus fractus, Prestosuchus chiniquensis (UFRGS-PV-0156-T), and Decuriasuchus quartacolonia. They proposed that the elongated gap between the premaxilla and maxilla is probably an artifact of preservation and this prompted a critical reevaluation of the putative openings in the anterior portion of the skull of other suchians.
In the described specimens of Prestosuchus chiniquensis we can find different conditions in this region of the preserved skulls. For instance, in the specimen UFRGS-PV-0156-T (Barberena 1978), this opening was reported as absent, while it is described as present (as a fenestra) in UFRGS-PV-0629-T and CPEZ-239b (Lacerda et al. 2016). According to Nesbitt and Desojo (2017), the lack of a fenestra decribed by Barberena (1978) in the UFRGS-PV-0156-T could be the result of the lateromedial compression suffered by the skull. Lacerda et al. (2016: fig. 4) described for the specimen (CPEZ-239b) that the posteroventral border of the right premaxilla is slightly curved anteriorly, forming a low, concave area on the posterior surface, followed by a thickened area that expands posteriorly on its posteroventral margin. This concave surface, sensu Lacerda et al. (2016), would be morphologically similar to the anterior margin of the subnarial fenestra described for Saurosuchus galillei (Alcober 2000), Decuriasuchus quartacolonia (França et al. 2013) and, although smaller in size, Postosuchus kirkpatricki (Weinbaum 2011).
The ULBRA-PVT-281 (Roberto-da-Silva et al. 2016) was described as having a subnarial foramen in this region; moreover, this opening was proposed as an autapomorphy for the species. In addition, Roberto-da-Silva et al. (2016) suggested that in Prestosuchus chiniquensis and Saurosuchus galilei, the subnarial foramen presents a new condition associated to its relative position on the body of the premaxilla (“located above the middle height of the main body of the maxilla, reaching the base of the ascending process”, Roberto-da-Silva et al. 2016: 15, appendix 1) and used this condition as a new character in their phylogenetic analysis.
We do not agree with the use of the term “foramen” for this strucuture because it does not have the morphology of a typical foramen, i.e., an opening that has well defined rounded margins that pierce through a bone. Besides, the use of the term foramen for this opening would be in conflict with the presence of the two true foramina that open medially to it, the rostromedial and rostrolateral foramina. We consider that the shape and size of the gap (or even its absence) between the premaxilla and the maxilla in Prestosuchus chiniquensis (and in other basal Loricata as well) can vary greatly depending on the taphonomic mode of preservation of each skull, as Nesbitt and Desojo (2017) proposed. The contact between these elements is apparently weak (probably reflecting some degree of cranial kinesis between these elements) and could result in different relative positions during the fossilization processes. Despite this, we agree that this mobile contact between the premaxilla and the maxilla is homologous to all the basal Loricata, independently of the small morphological variations that have been described between them. The presence of a notch in that area in Batrachotomus kupferzellensis is the only condition clearly distinct from other basal Loricata and can then be considered an autapomorphy for this taxon.
Conclusions
UFRGS-PV-0629-T represents the most complete specimen described for Prestosuchus chiniquensis, the greatest carnivore of the Brazilian Middle Triassic, known to date (see reconstruction of the skull in Fig. 20). In the present contribution, we described its cranial anatomy in detail and discussed some of its structures. Some characters hitherto doubtful or never mentioned before, could be described because most of the bones of this skull are disjointed, but well preserved. Among these, we emphasize the presence of the rostrolateral foramen, at the level of the palatal process of the maxilla, and the description of some palatal bones that were not possible to observe in the previously described specimens. Regarding the presence/absence of a subnarial fenestra, in this taxon and in the other basal Loricata, we understand that it is more likely related to a taphonomic artifact than to morphological variations that may have a taxonomic meaning.
Concerning the paleoneuroanatomical analysis (the first one ever published for a basal Loricata) we found that the encephalon of Prestosuchus seems strikingly convergent to that of theropod dinosaurs, which could be a reflection of their similar habits, since they occupied equivalent niches during the Mesozoic Era.
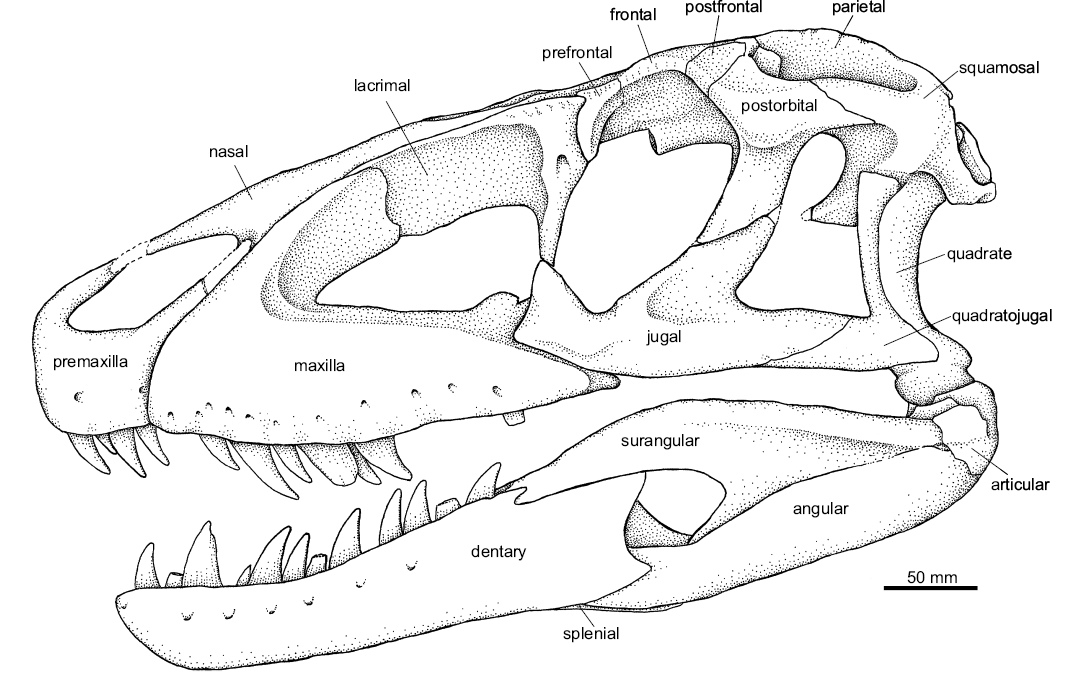
Fig. 20. Reconstruction of the pseudosuchian archosaur Prestosuchus chiniquensis Huene, 1938 (UFRGS-PV-0629-T) from the Dinodontosaurus Assemblage Zone, Ladinian, Middle Triassic, Dona Francisca municipality, Rio Grande do Sul State, Brazil; skull in lateral view.
Acknowledgements
We thank the curators Marco Brandalise de Andrade (PUCRS) and Oliver Rauhut (BSPG) for allowing us the access for studying the materials under their care. This research was partially funded by the Agencia Nacional de Promoción Científica y Técnica PICT 2014 No. 609 (to JBD). We thank the editor and two reviewers for their suggestions that improved the quality of the manuscript.
References
Alcober, O. 2000. Redescription of the skull of Saurosuchus galilei (Archosauria: Rauisuchidae). Journal of Vertebrate Paleontology 20: 302–316. Crossref
Alcober, O. and Parrish, J.M. 1997. A new poposaurid from the Upper Triassic of Argentina. Journal of Vertebrate Paleontology 17: 548–556. Crossref
Azevedo, S.A.K. 1991. Prestosuchus chinequensis Huene 1942 (Reptilia, Archosauria, Thecodontia, Proterosuchia, Rauisuchidae), da formação Santa Maria, Triássico do Estado do Rio Grande do Sul, Brasil. 157 pp. Ph.D. Thesis, Programa de Pós-Graduação em Geociências, Universidade Federal do Rio Grande do Sul, Porto Alegre.
Baczko, M.B. von and Desojo, J.B. 2016. Cranial anatomy and palaeoneurology of the archosaur Riojasuchus tenuisceps from the Los Colorados Formation, La Rioja, Argentina. PLoS ONE 11 (2): e0148575. Crossref
Baczko, M.B. von, Taborda, J.R.A., and Desojo, J.B. 2018. Paleoneuroanatomy of the aetosaur Neoaetosauroides engaeus (Archosauria: Pseudosuchia) and its paleobiological implications among archosauriforms. PeerJ 6: e5456. Crossref
Barberena, M.C. 1978. A huge Thecodont skull from Triassic of Brazil. Pesquisas 9: 62–75.
Benton, M.J. 1984. Rauisuchians and the success of dinosaurs. Nature 310: 101. Crossref
Benton, M.J. 1986. The Late Triassic reptile Teratosaurus—a rauisuchian, not a dinosaur. Palaeontology 29: 293–301.
Benton, M.J. and Clark, J.M. 1988. Archosaur phylogeny and the relationships of the Crocodilia. In: M.J. Benton (ed.), Phylogeny and Classification of the Tetrapods. Volume 1, 295–338. Clarendon Press, Oxford.
Bona, P., Paulina Carabajal, A., and Gasparini, Z. 2017. Neuroanatomy of Gryposuchus neogaeus (Crocodylia, Gavialoidea): a first integral description of the braincase and endocranial morphological variation in extinct and extant gavialoids. Earth and Environmental Science Transactions of The Royal Society of Edinburgh 106: 235–246. Crossref
Bonaparte, J.F. 1981. Descripcion de “Fasolasuchus tenax” y su significado em La sistemática y evolution de los Thecodontia. Revista Del Museo Argentino de Ciencias Naturales “Bernardino Rivadavia” 3: 55–101.
Bonaparte, J.F. 1984. Locomotion in rauisuchid thecodonts. Journal of Vertebrate Paleontology 3: 210–218. Crossref
Bronzati, M., Rauhut, O.W., Bittencourt, J.S., and Langer, M.C. 2017. Endocast of the Late Triassic (Carnian) dinosaur Saturnalia tupiniquim: implications for the evolution of brain tissue in Sauropodomorpha. Scientific Reports 7: 11931. Crossref
Brusatte, S.L., Benton, M.J., Desojo, J.B., and Langer, M.C. 2010. The higher-level phylogeny of Archosauria (Tetrapoda: Diapsida). Journal of Systematic Palaeontology 8: 3–47. Crossref
Brusatte, S.L., Butler, R.J., Sulej, T., and Niedzwiedzki, G. 2009. The taxonomy and anatomy of rauisuchian archosaurs from the Late Triassic of Germany and Poland. Acta Palaeontologica Polonica 54: 221–230. Crossref
Butler, R.J., Sullivan, C., Ezcurra, M.D., Liu, J., Lecuona, A., and Sookias, R.B. 2014. New clade of enigmatic early archosaurs yields insights into early pseudosuchian phylogeny and the biogeography of the archosaur radiation. BMC Evolutionary Biology 14: 1–16. Crossref
Case, E.C. 1928. An endocranial cast of a phytosaur from the Upper Triassic beds of western Texas. Journal of Comparative Neurology 45: 161–168. Crossref
Cerda, I., Desojo, J.B., Scheyer, T., and Schultz, C. 2013. Osteoderm microstructure of “rauisuchian” archosaurs from South America. Geobios 637: 273–283. Crossref
Chatterjee, S. 1985. Postosuchus, a new thecodontian reptile from the Triassic of Texas and the origin of tyrannosaurs. Philosophical Transaction of the Royal Society London B 309: 395–460. Crossref
Chatterjee, S. and Majumdar, P.K. 1987. Tikisuchus romeri, a new rauisuchid reptile from the Late Triassic of India. Journal of Paleontology 61: 787–793. Crossref
Codorniú, L., Paulina Carabajal, A., Pol, D., Unwin, D., and Rauhut, O.W. 2016. A Jurassic pterosaur from Patagonia and the origin of the pterodactyloid neurocranium. PeerJ 4: e2311. Crossref
Cope, E.D. 1869. Synopsis of the extinct Batrachia and Reptilia of North America. Transactions of the American Philosophical Society 14: 1–252. Crossref
Cruzado-Caballero, P., Fortuny, J., Llacer, S., and Canudo, J.I. 2015. Paleoneuroanatomy of the European lambeosaurine dinosaur Arenysaurus ardevoli. PeerJ 3: e802. Crossref
Dawley, R.M., Zawiskie, J.M., and Cosgriff, J.W. 1979. A rauisuchid thecodont from the Upper Triassic Popo Agie Formation of Wyoming. Journal of Paleonology 53: 1428–1431.
Desojo, J.B. and Arcucci, A.B. 2009. New material of Luperosuchus fractus (Archosauria: Crurotarsi) from the Middle Triassic of Argentina: the earliest known South American rauisuchian. Journal of Vertebrate Paleontology 29: 1311–1315. Crossref
Desojo, J.B. and Ezcurra, M.D. 2016. Triassic pseudosuchian archosaurs of South America. In: F.L. Agnolin, G.L. Lio, F.B. Egli, N.R. Chimento, and F.E. Novas (eds.), Historia Evolutiva y Paleobiogeografica de los vertebrados de América del Sur. Contribuciones del MACN 6: 57–66.
Desojo, J.B. and Rauhut, O. 2008. New insights on “rauisuchian” taxa (Archosauria: Crurotarsi) from Brazil. In: G. Dyke, D. Naish, and M. Parkes (eds.), SVP CA 12, Palaeontology, 31. National Museum of Ireland, Dublin.
Edmund, A.G. 1962. Sequence and rate of tooth replacement in the crocodile. Life Sciences Division of the Royal Ontario Museum of Zoology, Paleontology Contribution 56: 1–42.
Edmund, A.S. 1969. Dentition. In: C. Gans (ed.), Biology of the Reptilia. Volume 1, 117–200. Academic Press, London.
Evans, D.C., Ridgely, R., and Witmer, L.M. 2009. Endocranial anatomy of lambeosaurine hadrosaurids (Dinosauria: Ornithischia): a sensorineural perspective on cranial crest function. The Anatomical Record 292: 1315–1337. Crossref
França, M.A.G.,Langer, M.C., and Ferigolo, J. 2011. Associated skeletons of a new middle Triassic “Rauisuchia” from Brazil. Science of Nature 98: 389–395. Crossref
França, M.A.G., Langer, M.C., and Ferigolo J. 2013. The skull anatomy of Decuriasuchus quartacolonia (Pseudosuchia: Suchia: Loricata) from the middle Triassic of Brazi. In: S. Nesbitt, J.B. Desojo, and R.B. Irmis (eds.), Anatomy, Phylogeny and Palaeobiology of Early Archosaurs and Their Kin, 469–501. The Geological Society of London, London.
Franzosa, J. and Rowe, T. 2005. Cranial endocast of the Cretaceous theropod dinosaur Acrocanthosaurus atokensis. Journal of Vertebrate Paleontology 25: 859–864. Crossref
Galton, P.M. 1985. The poposaurid thecodontian Teratosaurus suevicus v. Meyer, plus referred specimens mostly based on prosauropod dinosaurs, from the Middle Stubensandstein (Upper Triassic) of Nordwürttemberg. Stuttgarter Beiträge zur Naturkunde, Serie B: Geologie und Palaontologie 116: 1–29.
Gauthier, J.A. and Padian, K. 1985. Phylogenetic, functional, and aerodynamic analyses of the origin of birds and their flight. In: M.K. Hecht, J.H. Ostrom, G. Viohl, and P. Wellnhofer (eds.), The Beginings of Birds, 185–197. Freunde des Jura-Musems, Eichstätt.
Giffin, E.B. 1989. Pachycephalosaur paleoneurology (Archosauria: Ornithischia). Journal of Vertebrate Paleontology 9: 67–77. Crossref
Gower, D.J. 1999. Cranial osteology of a new rauisuchian archosaur from the Middle Triassic of southern Germany. Stuttgarter Beiträge zur Naturkunde B 280: 1–49.
Gower, D.J. 2000. Rauisuchian archosaurs (Reptilia, Diapsida): an overview. Neues Jahrbuch für Geologie und Palaöntologie, Abhandlungen 218: 447–488. Crossref
Gower, D.J. 2002. Braincase evolution in suchian archosaurs: evidence from the rauisuchian Batrachotomus. Zoological Journal of the Linnean Society 136: 49–76. Crossref
Gower, D.J. and Sennikov, A.G. 1996. Endocranial casts of early archosaurian reptiles. Paläontologische Zeitschrift 70: 579–589. Crossref
Holloway, W.L., Claeson, K.M., and O’Keefe, F.R. 2013. A virtual phytosaur endocast and its implications for sensory system evolution in archosaurs. Journal of Vertebrate Paleontology 33: 848–857. Crossref
Hopson, J.A. 1979. Paleoneurology. Biology of the Reptilia 9: 39–146.
Horn, B.L.D., Goldberg, K., and Schultz, C.L. 2018. A loess deposit in the Late Triassic of southern Gondwana, and its significance to global paleoclimate. Journal of South American Earth Sciences 81: 189–203. Crossref
Huene, F. von 1938. Die fossilen Reptilien des südamerikanischen Gondwanalandes. Neues Jahrbuch für Mineralogie, Geologie und Paläontologie 1938: 142–151.
Huene, F. von 1942. Die fossilen Reptilien des südamerikanischen Gondwanalandes, Ergebnisse der Sauriergrabungen in Südbrasilien 1928/29. 332 pp. C.H. Becksche, München.
Juul, L. 1994. The phylogeny of basal archosaurs. Paleontologia Africana 31: 1–38.
Kley, N.J., Sertich, J.J., Turner, A.H., Krause, D.W., O’Connor, P.M., and Georgi, J.A. 2010. Craniofacial morphology of Simosuchus clarki (Crocodyliformes: Notosuchia) from the Late Cretaceous of Madagascar. Journal of Vertebrate Paleontology 30: 13–98. Crossref
Krebs, B. 1976. Pseudosuchia. In: O. Kuhn (ed.), Handbuch der Palaeoherpetologie: Thecondonti, 13: 40–98. Gustav Fischer-Verlag, Stuttgart.
Lacerda, M.B., Mastrantonio, B.M., Fortier, D.C., and Schultz C.L. 2016. New insights on Prestosuchus chiniquensis Huene, 1942 (Pseudosuchia, Loricata) based on new specimens from the “Tree Sanga” Outcrop, Chiniquá Region, Rio Grande do Sul, Brazil. PeerJ 4: e1622. Crossref
Lacerda, M.B., Schultz, C.L., and Bertoni-Machado, C. 2015. First “Rauisuchian” archosaur (Pseudosuchia, Loricata) for the Middle Triassic Santacruzodon Assemblage Zone (Santa Maria Supersequence), Rio Grande do Sul State, Brazil. PLoS ONE 10 (2): e0118563. Crossref
Lautenschlager, S. and Butler, R.J. 2016. Neural and endocranial anatomy of Triassic phytosaurian reptiles and convergence with fossil and modern crocodylians. PeerJ 4: e2251. Crossref
Lautenschlager, S. and Rauhut, O.W.M. 2015. Osteology of Rauisuchus tiradentes from the Late Triassic (Carnian) Santa Maria Formation of Brazil, and its implications for rauisuchid anatomy and phylogeny. Zoological Journal of the Linnean Society 173: 55–91. Crossref
Lessner, E.J., Stocker, M.R., Smith, N.D., Turner, A.H., Irmis, R.B., and Nesbitt, S.J. 2016. A new taxon of rauisuchid (Archosauria, Pseudosuchia) from the Upper Triassic of New Mexico increases the diversity and temporal range of the clade. PeerJ 4: e2336. Crossref
Liparini, A. 2008. Estudo da Biomecânica craniana de um rauissuquídeo a partir de tomografias computadorizadas e técnicas de imagens digitais em 3 dimensões. 122 pp. Dissertação de Mestrado, Programa de Pós-graduação em Geociências, Universidade Federal do Rio Grande do Sul, Porto Alegre.
Long, L.A. and Murry, P.A. 1995. Late Triassic (Carnian and Norian) tetrapods from the Southwestern United States. Bulletin New Mexico Museum of Natural History and Science 4: 1–254.
Mastrantonio, B.M. 2010. Descrição Osteológica de Materiais Cranianos e Pós-cranianos de Prestosuchus chiniquensis (Archosauria, Rauisuchia) do Mesotriássico do RS (Biozona de Dinodontosaurus, Formação Santa Maria) e Considerações Filogenéticas sobre Rauissúquios. Ph.D. Thesis, Programa de Pós-Graduação em Geociências, Universidade Federal do Rio Grande do Sul, Porto Alegre.
Mastrantonio, B.M., Schultz, C.L., Desojo, J.B., and Garcia, J.B. 2013. The braincase of Prestosuchus chiniquensis (Archosauria: Suchia). In: S. Nesbitt, J.B. Desojo, and R.B. Irmis (eds.), Anatomy, Phylogeny and Palaeobiology of Early Archosaurs and their Kin. Geological Society of London Special Publications 379: 425–440. Crossref
Merrem, B. 1820. Versuch eines Systems der Amphibien I (Tentamen Systematis Amphibiorum). 191 pp. J.C. Kriegeri, Marburg.
Miyashita, T., Arbour, V.M., Witmer, L.M., and Currie, P.J. 2011. The internal cranial morphology of an armoured dinosaur Euoplocephalus corroborated by X-ray computed tomographic reconstruction. Journal of Anatomy 219: 661–675. Crossref
Nesbitt, S.J. 2011. The early evolution of archosaurs: relationships and the origin of major clades. Bulletin of the American Museum of Natural History 352: 1–292. Crossref
Nesbitt, S. and Desojo, J.B. 2017. The osteology and phylogenetic position of Luperosuchus fractus (Archosauria: Loricata) from the latest Middle Triassic or earliest Late Triassic of Argentina. Ameghiniana 54: 261–282. Crossref
Nesbitt, S.J., Brusatte, S.L., Desojo, J.B., Liparini, A., De França, M.A., Weinbaum, J.C., and Gower, D.J. 2013. Rauisuchia. In: S. Nesbitt, J.B. Desojo, and R.B. Irmis (eds.), Anatomy, Phylogeny and Palaeobiology of Early Archosaurs and their Kin. Geological Society of London Special Publication? 379: 241–274. Crossref
Nesbitt, S.J., Stocker, M.R., Parker, W.G., Wood, T.A., Sidor, C.A., and Angielczyk, K.D. 2018. The braincase and endocast of Parringtonia gracilis, a Middle Triassic suchian (Archosauria: Pseudosuchia). Journal of Vertebrate Paleontology 37: 122–141. Crossref
Paulina Carabajal, A. 2015. Guía para el estudio de la neuroanatomía de dinosaurios Saurischia, con énfasis en formas sudamericanas. Publicación Electrónica de la Asociación Paleontológica Argentina 15 (1): 108–142. Crossref
Paulina Carabajal, A. and Canale, J.I. 2010. Cranial endocast of the carcharodontosaurid theropod Giganotosaurus carolinii Coria & Salgado. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 258: 249–256. Crossref
Paulina Carabajal, A. and Currie, P.J. 2012. New information on the braincase of Sinraptor dongi (Theropoda: Allosauroidea): Ethmoidal region, endocranial anatomy, and pneumaticity. Vertebrata PalAsiatica 4: 85–101.
Parrish, J.M. 1993. Phylogeny of the crocodylotarsy, with reference to archosaurian and crurotarsan monophyly. Journal of Vertebrate Paleontology 13: 287–308. Crossref
Peyer, K., Carter, J.G., Sues, H.D., Novak, S.E., and Olsen, P.E. 2008. A new suchian archosaur from the Upper Triassic of North Carolina. Journal of Vertebrate Paleontology 28: 363–381. Crossref
Raugust, T. 2014. Um novo material de rauissúquio (Archosauria, Crurotarsi) da Formação Santa Maria, Triássico Médio Sul-Rio-Grandense, Brasil. Ph.D. Thesis, Programa de Pós-Graduação em Geociências, Universidade Federal do Rio Grande do Sul, Porto Alegre.
Reig, O.A. 1959. Primeros datos descriptivos sobre nuevos reptiles arcosaurios Del Triásico de Ischigualasto. Revista de la Asociacion Geológica Argentina 13: 257–270.
Roberto-Da-Silva, L., França, M.A.G., Cabreira, S.F., Müller, R.T., and Dias-Da-Silva, S. 2016. On the presence of the subnarial foramen in Prestosuchus chiniquensis (Pseudosuchia: Loricata) with remarks on its phylogenetic distribution. Anais da Academia Brasileira de Ciências 88 (3): 1309–1323. Crossref
Romer, A.S. 1971. The Chañares (Argentina) Triassic reptile fauna. VIII. A fragmentary skull of a large thecodont, Luperosuchus fractus. Breviora 73: 1–8.
Sampson, S.D. and Witmer, L.M. 2007. Craniofacial anatomy of Majungasaurus crenatissimus (Theropoda: Abelisauridae) from the Late Cretaceous of Madagascar. In: S.D. Sampson and D.W. Krause (eds.), Majungasaurus crenatissimus (Theropoda: Abelisauridae) from the Late Cretaceous of Madagascar. Journal of Vertebrate Paleontology 27: 32–104.
Sereno, P.C. and Novas, F.E. 1993. The skull and neck of the basal theropod Herrerasaurus ischigualastensis. Journal of Vertebrate Paleontology 13: 451–476. Crossref
Sill, W.D. 1974. The anatomy of Saurosuchus galilei and the relationships of the rauisuchid thecodonts. Bulletin of the Museum of Comparative Zoology 146: 317–362.
Sobral, G., Sookias, R.B., Bhullar, B.A.S., Smith, R., Butler, R.J., and Müller, J. 2016. New information on the braincase and inner ear of Euparkeria capensis Broom: implications for diapsid and archosaur evolution. Royal Society Open Science 3 (7): 160072. Crossref
Stocker, M.R., Nesbitt, S.J., Criswell, K.E., Parker, W.G., Witmer, L.M., Rowe, T.B., Ridgely, R., and Brown, M.A. 2016. A dome-headed stem archosaur exemplifies convergence among dinosaurs and their distant relatives. Current Biology 26: 2674–2680. Crossref
Sulej, T. 2005. A new rauisuchian reptile (Diapsida: Archosauria) from the Late Triassic of Poland. Journal of Vertebrate Paleontology 25: 78–86. Crossref
Trotteyn, M.J., Desojo, J.B., and Alcober, O. 2011. Nuevo material postcraneano de Saurosuchus galilei Reig (Archosauria: Crurotarsi) del Triasico Superior del centro-oeste de Argentina. Ameghiniana 48: 605–620. Crossref
Weinbaum, J.C. 2011. The skull of Postosuchus kirkpatricki (Archosauria: Paracrocodyliformes) from the Upper Triassic of the United States. PaleoBios 30: 18–44.
Witmer, L.M. and Ridgely, R.C. 2008. The paranasal air sinuses of predatory and armored dinosaurs (Archosauria: Theropoda and Ankylosauria) and their contribution to cephalic structure. The Anatomical Record 291: 1362–1388. Crossref
Witmer, L.M. and Ridgely, R.C. 2009. New insights into the brain, braincase, and ear region of tyrannosaurs (Dinosauria, Theropoda), with implications for sensory organization and behavior. The Anatomical Record 292: 1266–1296. Crossref
Witmer, L.M., Chatterjee, S., Franzosa, J., and Rowe, T. 2003. Neuroanatomy of flying reptiles and implications for flight, posture and behaviour. Nature 425: 950. Crossref
Witmer, L.M., Ridgely, R.C., Dufeau, D.L., and Semones, M.C. 2008. Using CT to peer into the past: 3D visualization of the brain and ear regions of birds, crocodiles, and nonavian dinosaurs. In: H. Endo and R. Frey (eds.), Anatomical Imaging, 67–87. Springer, Tokyo.
Acta Palaeontol. Pol. 64 (1): 171–198, 2019
https://doi.org/10.4202/app.00527.2018