
Anatomy and relationships of the fuxianhuiid euarthropod Guangweicaris from the early Cambrian Guanshan Biota in Kunming, Yunnan, Southwest China revisited
YICHEN WU and JIANNI LIU
The Guanshan Biota (Cambrian Series 2, Stage 4) is a typical Burgess Shale-type biota, and is one of the most significant Cambrian Konservat-Lagerstätten from China, in addition to the well-known Chengjiang and Kaili biotas. Guangweicaris is a relatively common arthropod from the Guanshan Biota. Based on new specimens from the Longbaoshan and Xinglongcun sections of the Wulongqing Formation in Kunming, we herein report novel features of Guangweicaris spinatus, such as the eye-bearing anterior sclerite that is associated with the head shield, as well as revised details of the trunk, including the ventral appendages. A detailed comparison of Guangweicaris, Fuxianhuia and Shankouia suggests a close affinity between Guangweicaris and Fuxianhuia, which improves our understanding of the evolution of early Cambrian euarthropods.
Introduction
The Guanshan Biota is a representative Burgess Shale-type biota in southwest China yielding a considerable diversity of biomineralized and soft-bodied organisms, and shares similarities with elder Chengjiang and Xiaoshiba Biotas (Hu et al. 2006, 2007, 2010a, b, 2013; Li et al. 2006, 2017; Liu et al. 2012, 2015, 2016; Luo et al. 2005, 2006, 2007; Yang et al. 2008, 2018). The more than 60 taxa described from this biota belong to over 10 groups of algae and metazoans, including sponges, chancelloriids, eldonoids, cnidarians, palaeoscolecids, hyoliths, lobopodian and sclerotized arthropods, brachiopods, eocrinoids, and vetulicolians (Hu et al. 2006, 2007, 2010a, b, 2013; Li et al. 2006, 2017; Liu et al. 2012, 2015, 2016; Luo et al. 2005, 2006, 2007; Yang et al. 2008, 2018). The Guanshan Biota is essential to elucidate the “Cambrian Explosion” of early metazoans for its exquisite soft-body preservation and its timing between the Chengjiang Fauna and the Kaili Fauna (and also the coeval Burgess Shale community) (Liu et al. 2012, 2016; Li et al. 2017).
During several excavation campaigns from 2012 to 2018, many specimens assigned to over 40 genera were collected from the pale yellowish-green, thin-bedded mudstone of the lower member of the Wulongqing Formation, with arthropods being particularly abundant. Among them, more than 400 new specimens of the fuxianhuiid euarthropod Guangweicaris spinatus Luo, Fu, and Hu, 2007 were found, of which 12 are particularly well preserved.
Since the first description of G. spinatus in 2007 (Luo et al. 2007), only one other publication (Yang et al. 2008) has provided additional morphological information on this taxon. Luo et al. (2007) described the trunk morphology and determined the stratigraphic distribution of this species, and proposed the monotypic family Guangweicarididae. Later, Yang et al. (2008) revised the morphological features of G. spinatus, including details of the head shield and tail flukes, and considered it to be the most similar to Fuxianhuia Hou, 1987. The new materials described herein not only provide new morphological information about G. spinatus, but also indicate that Guangweicaris and Fuxianhuia are indeed closely related.
Institutional abbreviations.—NWU, Northwest University, Xi’an, China
Other abbreviations.—exsag, exsagittal; sag, sagittal; tgn, the n-th trunk tergite.
Material and methods
All new specimens discussed here were observed and prepared with fine needles under high magnification using microscopes. The fossils were photographed with a Canon EOS 5D Mark II digital camera, and then the figures were prepared using graphic software. Specimens ELI-LBSG-0006B, ELI-LBSG-0046A, and ELI-LBSG-0047 were excavated from the Longbaoshan section (Cambrian Series 2, Stage 3; see Luo et al. 2008), and specimen ELI-XLCG-0022B was excavated from the Xinglongcun section (Cambrian Series 2, Stage 3; see Liu et al. 2016).
All specimens are housed at the Shaanxi Key Laboratory of Early Life and Environments, Northwest University (NWU), Xi’an, China.
Systematic palaeontology
Upper stem-group Euarthropoda Lankester, 1904
Order Fuxianhuiida Bousfield, 1995
Family Fuxianhuiidae Hou and Bergström, 1997
Genus Guangweicaris Luo, Fu, and Hu, 2007
Type species: Guangweicaris spinatus Luo, Fu, and Hu, 2007; lower Cambrian; Southwest China; monotypic.
Guangweicaris spinatus Luo, Fu, and Hu, 2007
Figs. 1, 2.
Material.—More than 400 specimens in total, 12 of them are particularly well preserved. Three specimens (ELI-LBSG-0046A, B; ELI-LBSG0006B; ELI-LBSG0357A) preserved the antennules, and only one specimen (ELI-LBSG-0046A, B) preserved the stalked-eyes.
Emended diagnosis.—Fuxianhuiid with an elongate body composed of a sub-trapezoidal head shield with eye-bearing anterior sclerite, 15 trunk tergites, and a pair of tail flukes. First three anteriormost trunk tergites relatively reduced compared to the rest of the trunk and covered by the head shield as the dorsal tergite of the second segment, with a sub-elliptical outline, extending posteriorly to cover the anterior reduced tergites. Remainder of trunk tergites subdivided into two distinct regions: tg4–tg8 being much wider (tr) than tg9–tg15; median keel developed along most of trunk, with prominent posteriorly-directed spines situated on the posteromedial margins of each tergite.
Description.—Body composed of head shield with eye-bearing anterior segment, and a trunk of 15 segments, plus a pair of tail flukes. Body dimensions typically 60–95 mm in total length and up to 30 mm in maximum width (Fig. 1A–C).
The cephalic region comprises two parts: a sub-elliptical anterior segment situated on the anteromedial margin of a subtrapezoidal head shield. A pair of lateral stalked eyes is connected to the anterior segment (Fig. 2A, B). In life position, the head shield would cover the reduced trunk segment tg1–tg3 (Figs. 1A–C, 2A, B, 4). Paired antennulae, each composed of at least 12 articles, attach immediately posterior to the anterior segment (Figs. 1B, 2A, C, D).
Trunk comprises 15 segments subdivided into a thorax (tg1–tg8) and abdomen (tg9–tg15), plus a pair of tail flukes. The anteriormost segments (tg1–tg3) are greatly reduced relative to the remainder of the trunk and preserved edge-to-edge. tg1 is the shortest (sag, exsag) and narrowest (tr), with tg2 and tg3 becoming progressively longer (sag, exsag) and wider (tr) posteriorly (Fig. 1C). tg4–tg8 are the largest trunk segments that are partially overlapping, each having spatulate tergopleurae, a median keel, and a prominent posteriorly-directed spine situated on the posteromedial margin (Fig. 1A–C). The abdomen is column-like in outline, with tg9–tg15 being much narrower (tr) than tg4–tg8, gradually decreasing in width (tr) and increasing in length (sag) posteriorly (with tg15 being one-third to one-half longer than tg14); each abdominal segment also has a median keel and a spine on the posteromedial margin. Pair of subtriangular tail flukes articulate with tg15 (Fig. 1C, D).
Thorax appears to have several sets of biramous appendages on the ventral side of the segments. Each appendage consists of a jointed endopod of at least 13 podomeres and a flap-like oval exopod (only visible in one specimen; Figs. 1A, 2A, E). Endopod podomeres sub-rectangular in outline, tapering gradually towards the distal end with no terminal spine present (Fig. 2A, E). Each large trunk segment, particularly tg4 and tg5 (Fig. 1A), appears to be associated with at least two pairs of biramous appendages.
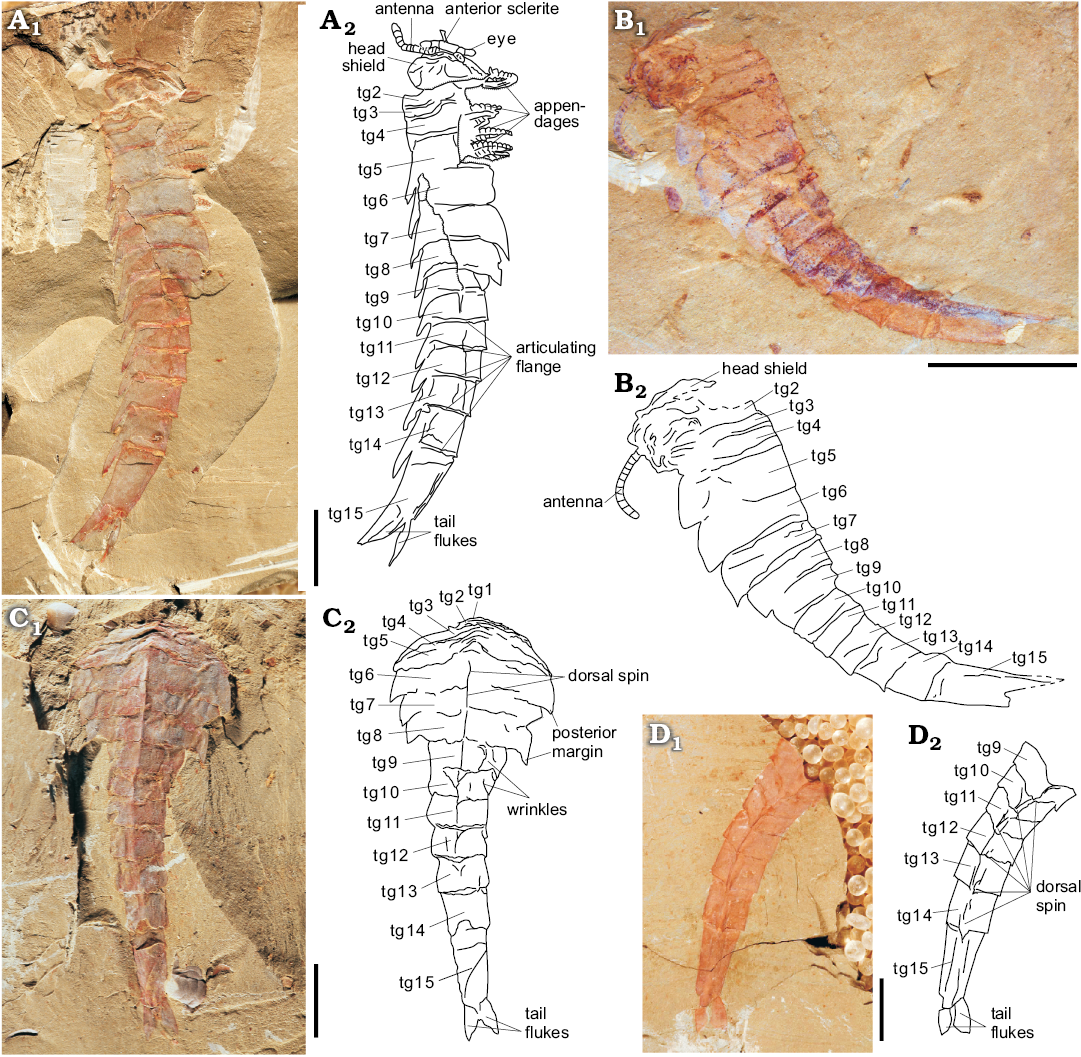
Fig. 1. Fuxianhuiid euarthropod Guangweicaris spinatus Luo, Fu, and Hu, 2007 from Guanshan Biota, Cambrian Series 2, Stage 4, Yunnan, China. A. ELI-LBSG-0046A in lateral view, showing the whole animal, including several important morphological characters of Guangweicaris: the rotated ocular segment, the pair of clavate antennules (although the right one in poorly preserved), the cephalic shield, the cephalic-appendages, the thorax, the abdomen, and the tail flukes. B. ELI-LBSG0006B in lateral view, showing the whole animal. C. ELI-LBSG-0047 in dorsal-ventral view, showing the typical Guangweicaris thorax, abdomen, and a pair of tail flukes. D. ELI-XLCG-022B in dorsal-ventral view, showing the typical Guangweicaris abdomen and the two tail flukes. Abbreviations: tg, tergite (tg1–tg3, anterior reduced segments; tg4–tg8, thoracic tergites; tg9–tg15, abdominal tergites). A1–D1, photographs; A2–D2, explanatory drawings. Scale bars 10 mm.
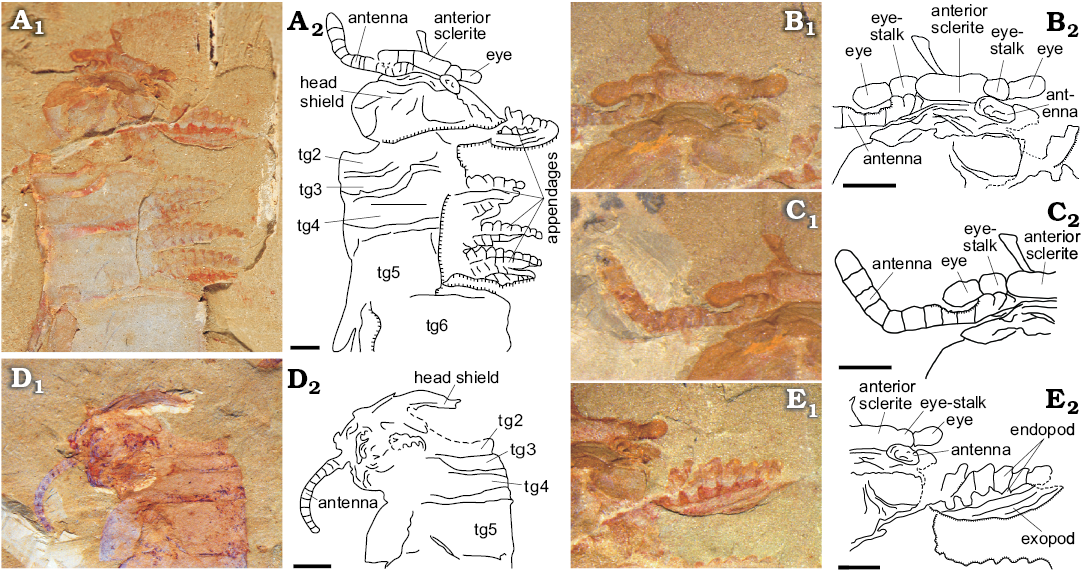
Fig. 2. Fuxianhuiid euarthropod Guangweicaris from Guanshan Biota, Cambrian Series 2, Stage 4, Yunnan, China. A. ELI-LBSG-0046A, showing the whole head part and the pairs of thoracic appendages. B. ELI-LBSG-0046A, showing the ocular segment and the stalked eyes. C. ELI-LBSG-0046A, showing the stout antennulae. D. ELI-LBSG0006B, cephalon, showing the cephalic shield and the stout antennulae. E. ELI-LBSG-0046A, showing the two pairs of cephalic appendages. All in lateral view. Abbreviations: tg, tergite (tg1–tg3, anterior reduced segments; tg4–tg6, thoracic tergites). A1–E1, photographs; A2–D2, explanatory drawings. Scale bars 2 mm.
Remarks.—Fuxianhuia protensa in the most advanced developmental phase consists of 30 segments (18 in the thorax, 12 in the abdomen) (Fu et al. 2018), which is 15 more than in Guangweicaris spinatus (10 in the thorax, five in the abdomen). Fuxianhuia xiaoshibaensis consists of 36 segments (20 in the thorax, 16 in the abdomen) (Yang et al. 2013), which is 21 more than in Guangweicaris spinatus (12 in the thorax, nine in the abdomen). Shankouia zhenghei consists of at least 41 segments (six in the thorax, 35 in the abdomen), which is 26 more than in G. spinatus (Waloszek et al. 2005).
As in Fuxianhuia protensa, F. xiaoshibaensis, and S. zhenghei, the cephalic region of G. spinatus consists of two parts: (i) the anterior segment, which is a small rectangular segment situated at the front of the body, bearing a pair of stalked eyes that insert at the lateral margins; and (ii), the head shield that covers the anteriormost (reduced) segments and their associated biramous appendages. In the ventral part of the head in all species, the proximal parts of the paired antennulae are situated close to the posterolateral corner of the anterior segment. The antennulae of F. protensa and S. zhenghei are composed of about 15 similar pivot-jointed segments slightly decreasing in size towards the distal end, while each antenna of F. xiaoshibaensis consists of more than 20 podomeres and taper distally, and the antenna of G. spinatus is composed of at least 12 clavate articles. The reduced anterior segments (of body segments tg1–tg5) beneath the head shield in F. protensa are much narrower and shorter (tr, sag) than the first uncovered tergite (tg6). These tergites do not extend into tergopleurae, as is characteristic of the remaining uncovered tergites. The anterior reduced segments and their tergites widen (tr) gradually but the tergites remain narrower (sag) than the first uncovered tergite (tg6), and the situation about the anterior reduced segments of F. protensa and F. xiaoshibaensis are the same as for G. spinatus and S. zhenghei, but G. spinatus has only three such segments, and the S. zhenghei has six. In F. protensa/F. xiaoshibaensis, from the body segment tg6/tg4 backwards, around tg12/tg17 tergites, pairs of knobs and tergopleurae are present. Tergopleurae have a convex anterior margin, curved lateral margins, and a concave posterior margin. Five thoracic tergites of G. spinatus are decorated with two symmetrical pleural spines and one triangular strong spine at the midpoint of the posterior margin. The posterior trunk of F. protensa is made of 13 similar annular segments, meanwhile in F. xiaoshibaensis it consists of 16 segments, the length (sag) of which increases posteriorly, and the width (tr) decreases gradually backward. The abdomen of G. spinatus is composed of six segments in the same shape. In S. zhenghei, however, the first uncovered tergite is as wide as the enlarged head sheild, and the subsequent trunk tergites are progressively longer and wider backwards. After approximately one half of the body, they become progressively narrower and slightly shorter. The trunk of S. zhenghei ends in a conical portion that is as long as the six preceding tergite-bearing segments, and a pair of large triangular flaps appears to insert along the whole length of this portion. Ventrally, the trunk limbs of F. protensa, F. xiaoshibaensis, S. zhenghei, and G. spinatus are biramous appendages. Each biramous appendage consists of a tubular, slowly-tapering, multi-annulated main rod and a flap-like exopod (Fig. 1C). In S. zhenghei every trunk segment bears one pair of limbs (Waloszek et al. 2005), while F. protensa, F. xiaoshibaensis and G. spinatus were described as having more than one pair of limbs per segment (Hou and Bergström 1997; Yang et al. 2013; Fu et al. 2018).
Geographic and stratigraphic range.—Longbaoshan section in Kunming, Southwest China (Cambrian Series 2, Stage 3; local Wulongqing Formation); Xinglongcun section in Wuding, Southwest China (Cambrian Series 2, Stage 3; local Wulongqing Formation).
Discussion
Most of the similarities between Fuxianhuia protensa, F. xiaoshibaensis, and Guangweicaris spinatus discussed herein are ancestral characters of the fuxianhuiids as a whole (Ortega-Hernández 2016; Yang et al. 2018), while the differences that distinguish G. spinatus from Fuxianhuia are those that specifically describe G. spinatus. According to these differences, the phylogeny of Fuxianhuiidae suggests that both Fuxianhuia and Guangweicaris have a derived organization based on the differentiation between the thorax and abdominal regions (Yang et al. 2018).
No available evidence suggests that the so-called specialized post-antennulae appendages and hypostome have been preserved in our material, which are distinctive morphological characters of the fuxianhuiids (Chen et al. 1995; Budd 2002; Waloszek et al. 2007; Yang et al. 2013, 2018). However, these structures should have existed, otherwise G. spinatus could not have taken food effectively due to the lack of mouth. More materials are needed to clarify this.
Conclusions
We added new information about the morphology of Guangweicaris spinatus—in particular on the eye, antennulae, appendages, abdomen, and tail flukes. We also revised the previous species description, and highlighted that the most important feature that connects Guangweicaris and Fuxianhuia is the trunk tagmata (thorax + abdomen) (Fig. 3). The new material also shows that Guangweicaris has an anterior sclerite with stalked eyes, and at least two biramous appendages for each thoracic tergite (like in other fuxianhuiids) (Fig. 3), what not only provide the direct evidence to support the classification of Guangweicaris and Fuxianhuia within Fuxianhuiidae Hou and Bergström 1997 (Yang et al. 2018), but also may affect the character coding and the phylogenetic position of the Guangweicaris. At last, the stalked-eyes we described here shed new light on the soft-body preservation of the fuxianhuiid in the Guanshan Biota.

Fig. 3. The comparison of fuxianhuiid euarthropods Fuxianhuia protensa Hou, 1987 from Chengjiang Biota, Cambrian Series 2, Stage 3, Yunnan, China (A) and Guangweicaris spinatus Luo, Fu, and Hu, 2007 from Guanshan Biota, Cambrian Series 2, Stage 4, Yunnan, China (B, C). A. EJ0072A in dorsal-ventral view, showing the typical F. protensa body. B. ELI-LBSG-0046A in lateral view, showing the whole head part and the pairs of thoracic appendages of Guangweicaris. C. ELI-LBSG-0003 in dorsal-ventral view, showing the typical Guangweicaris thorax and abdomena parts. Scale bars 5 mm.
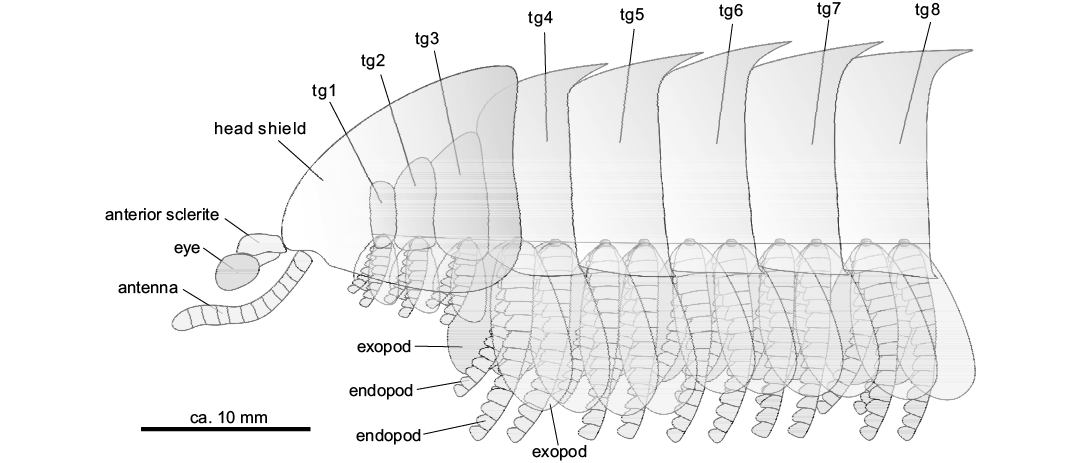
Fig. 4. Schematic reconstruction of a lateral view of the anterior body region (tg1–tg3, anterior reduced segments) of a generalized Guangweicaris. Abbreviations: tg, tergite.
Acknowledgments.—We thank referees Dieter Waloszek (Lund University, Sweden) and Javier Ortega-Hernández (University of Cambridge, UK) for their constructive comments. Xing-Liang Zhang, Zhi-Fei Zhang, Jian Han, Dong-Jing Fu, De-Gan Shu (all Early Life Institute, State Key Laboratory of Continental Dynamics, Department of Geology, Northwest University, Xi’an, China) and Richard Paterson John (University of New England, Armidale, Australia) are thanked for their kind help and valuable comments.
References
Bousfield, E.L. 1995. A Contribution to the natural classification of Lower and Middle Cambrian arthropods: food gathering and feeding mechanism. Amphipacifica 2 (1): 3–34.
Budd, G.E. 2002. A palaeontological solution to the arthropod head problem. Nature 417: 271–275. Crossref
Chen, J.Y. 2004. The Dawn of Animal World [in Chinese]. 366 pp. Jiangsu Publishing House of Science and Technology, Nanjing.
Chen, J.Y., Edgecomb, G.D, Ramsköld, L., and Zhou, G.Q. 1995. Head segmentation in Early Cambrian Fuxianhuia: Implications for arthropod evolution. Science 268: 1339–1343. Crossref
Fu, D.J., Ortega-Hernández, J., Daley, A.C., Zhang, X.L., and Shu, D.G.. 2018. Anamorphic development and extended parental care in a 520 million-year-old stem-group euarthropod from China. BMC Evolutionary Biology 18: 147. Crossref
Hou, X.G. 1987. Three new large arthropods from Lower Cambrian, Chengjiang, eastern Yunnan [in Chinese with English abstract]. Acta Palaeontologica Sinica 26: 272–285.
Hou, X.G., and Bergström, J. 1997. Arthropods from the Lower Cambrian Chengjiang Fauna, southwest China. Fossils and Strata 45: 1–116.
Hou, X.G., Bergström, J., Wang, H.F., Feng, X.H., and Chen, A.L. 1999. The Chengjiang Fauna—Exceptionally Well-Preserved Animals From 530 Million Years ago [in Chinese with English abstract]. 170 pp. Yunnan Science Technology Press, Kunming.
Hu, S.X., Li, X.K., Tan, X.H., Zhan, D.Q., and Luo, H.L. 2010a. Phlogites from the Early Cambrian Guanshan Fauna. Acta Palaeontologica Sinica 49: 360–364.
Hu, S.X., Luo, H.L., Hou, S.G., and Erdtmann, B.D. 2006. Eocrinoid echinoderms from the Lower Cambrian Guanshan Fauna in Wuding, Yunnan, China. Chinese Science Bulletin 51: 1366–1368.
Hu, S.X., Luo, H.L., Hou, S.G., and Erdtmann, B.D. 2007. Eocrinoid echinoderms from the Lower Cambrian Guanshan fauna in Wuding, Yunnan, China. Chinese Science Bulletin 52: 717–719. Crossref
Hu, S.X., Zhu, M.Y., Luo, H.L., Steiner, M., Zhao F.C., Li, G.X., Liu, Q., and Zhang, Z.F. 2013. The Guanshan Biota. 204 pp. Yunnan Science and Technology Press, Kunming.
Hu, S.X., Zhu, M.Y., Steiner, M., Luo, H.L., Zhao, F.C., and Liu, Q. 2010b. Biodiversity and taphonomy of the early Cambrian Guanshan Biota, eastern Yunnan. Science China (Earth Sciences) 53: 1765–1773. Crossref
Lankester, E.R. 1904. The structure and classification of the Arthropoda. The Quarterly Journal of Microscopical Science 47: 523–582.
Li, J.S., Liu, J.N., and Ou, Q. 2017. New observations on Vetulicola longbaoshanensis from the Lower Cambrian Guanshan Biota (Series 2, Stage 4), South China. Science China Earth Sciences 10: 1795–1804. Crossref
Li, Y., Chen, L.Z., Luo, H.L., Fu, X.P., Hu, S.X., You, T., and Liu, Q. 2006. New advances in the study of the Early Cambrian Guanshan fauna in the Kunming area, Yunnan, China. Geological Bulletin of China 25: 415–418.
Liu, J.N., Han, J., Li, J.S., Wu, Y.C., Peng, J., Qi, N., Yang, Y.Y., and Li, J. 2016. New localities and palaeoscolecid worms from the Cambrian (Stage 4, Series 2) Guanshan Biota in Kunming, Yunnan, South China. Acta Geologica Sinica (English Edition) 90: 1939–1945. Crossref
Liu, J.N., Ou, Q., Han, J., Li, J.S., Wu, Y.C., Jiao, G.X., and He, T.J. 2015. Lower Cambrian polychaete from China sheds light on early annelid evolution. The Science of Nature 102: 34. Crossref
Liu, J.N., Ou, Q., Han, J., Zhang, Z.F., He, T.J., Yao, X.Y., Fu, D.J., and Shu, D.G. 2012. New occurrence of the Cambrian (Stage 4, Series 2) Guanshan Biota in Huize, Yunnan, South China. Bulletin of Geosciences 87: 125–132. Crossref
Luo, H.L., Fu, X.P., Hu, S.X., Li, Y., Chen L.Z., You, T., and Liu Q. 2005. New vetulicoliids from the Lower Cambrian Guanshan fauna, Kunming. Acta Geologica Sinica 79: 1–6. Crossref
Luo, H.L., Fu, X.P., Hu, S.X., Li, Y., Chen, L.Z., You, T., and Liu, Q. 2006. New bivalved arthropods from the Early Cambrian Guanshan fauna in the Kunming and Wuding area. Acta Palaeontologica Sinica 45: 460–472.
Luo, H.L., Fu, X.P., Hu, S.X., Li, Y., Hou, S.G., You, T., and Liu, Q. 2007. A new Arthropod, Guangweicaris Luo, Fu et Hu gen. nov. from the Early Cambrian Guanshan Fauna, Kunming, China. Acta Geologica Sinica (English Edition) 8l: 1–7.
Luo, H.L., Li, Y., Hu, S.X., Fu, X.P., Hou, S.G., Liu, X.Y., Chen, L.Z., Li, F.J., Pang, J.Y., and Liu, Q. 2008. Early Cambrian Malong Fauna and Guanshan Fauna from Eastern Yunnan, China [in Chinese with English abstract]. 134 pp. Yunnan Science and Technology Press, Kunming.
Ortega-Hernández, J. 2016. Making sense of “lower” and “upper” stem-group Euarthropoda, with comments on the strict use of the name Arthropoda von Siebold, 1848. Biological Reviews 91: 255–273. Crossref
Waloszek, D., Chen, J.Y., Maas, A., and Wang, X.Q. 2005. Early Cambrian arthropods—new insights in to arthropod head and structural evolution. Arthropod Structure and Development 34: 189–205. Crossref
Waloszek, D., Maas, A., Chen, J.Y., and Stein, M. 2007. Evolution of cephalic feeding structures and the phylogeny of Arthopoda. Palaeogeography, Palaeoclimatology, Palaeoecology 254: 273–287. Crossref
Yang, J., Hou, X.G., and Dong, W. 2008. Restudy of Guangweicaris Luo, Fu et Hu, 2007 from the lower Cambrian Canglangpu Formation in Kunming area [in Chinese with English abstract]. Acta Palaeontogica Sinica 47: 115–122.
Yang, J., Ortega-Hernández, J., Butterfield, N.J., and Zhang, X.G. 2013. Specialized appendages in fuxianhuiids and the head organization of early euarthropods. Nature 494: 468–471. Crossref
Yang, J., Ortega-Hernández, J., Legg, D.A., Lan, T., Hou, J.B., and Zhang, X.G. 2018. Early Cambrian fuxianhuiids from China reveal origin of the gnathobasic protopodite in euarthropods. Nature Communications 9: 470. Crossref
Yichen Wu [wuyicheneli@126.com] and Jianni Liu [liujianni@126.com], State Key Laboratory of Continental Dynamics, Shaanxi Key Laboratory of Early Life and Environment, Department of Geology, Northwest University, Xi’an, China.
Received 1 September 2018, accepted 25 March 2019, available online 30 July 2019.
Copyright © 2019 Y. Wu and J. Liu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Acta Palaeontol. Pol. 64 (3): 543–548, 2019
https://doi.org/10.4202/app.00542.2018