

Diversity in rhynchocephalian Clevosaurus skulls based on CT reconstruction of two Late Triassic species from Great Britain
SOFIA A.V. CHAMBI-TROWELL, DAVID I. WHITESIDE, and MICHAEL J. BENTON
Chambi-Trowell, S.A.V., Whiteside, D.I., and Benton M.J. 2019. Diversity in rhynchocephalian Clevosaurus skulls based on CT reconstruction of two Late Triassic species from Great Britain. Acta Palaeontologica Polonica 64 (1): 41–64.
Clevosaurus was an early sphenodontian, known from many specimens and species from the Late Triassic and Early Jurassic. Here we describe and compare the cranial morphology of Clevosaurus hudsoni, the type species, and Clevosaurus cambrica, both from the UK, based on digital reconstructions from segmented CT scan data of two skulls. C. hudsoni has been described in some detail, but the CT data reveal new information on cranial anatomy, and C. cambrica is a newly described species for which the CT scans enable us to identify many previously undescribed bones, re-identify bones whose identity had previously been uncertain, and refine certain cranial elements. The near complete preservation of the left side of the skull of C. cambrica, and the preservation of much of the left side of the skull of C. hudsoni, provided us with the opportunity to fully reconstruct both species’ skulls in three dimensions. For the first time, coronoid bones are identified in Clevosaurus species from the British Isles. These two species show the diversity of morphology in the genus Clevosaurus. We provide evidence for two morphotypes of C. hudsoni based on study of the dentary dentition of the syntypes and other attributed specimens.
Key words: Rhynchocephalia, Lepidosauria, Clevosaurus, anatomical description, 3D skull reconstruction, CT scanning, Triassic, UK.
Sofia Chambi-Trowell [sc14927@my.bristol.ac.uk], David I. Whiteside [David.Whiteside@bristol.ac.uk], and Michael J. Benton [mike.benton@bristol.ac.uk], School of Earth Sciences, University of Bristol, Bristol, BS8 1TF, UK.
Received 13 November 2018, accepted 6 January 2019, available online 20 February 2019.
Copyright © 2019 S. Chambi-Trowell et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Sphenodontia are rhynchocephalian lepidosaurs, which is sister group to Squamata (lizards, snakes and relatives), that originated in the early Mesozoic (Jones et al. 2013). Today, sphenodontians are represented solely by the tuatara (Sphenodon punctatus) of New Zealand, but in the Triassic and Jurassic, sphenodontians were a morphologically disparate and diverse group (Jones 2008; Herrera-Flores et al. 2017). Mesozoic sphenodontians show particularly high diversity in their dental morphology (Jones 2009). The genus Clevosaurus, which first appeared in the Late Triassic and was globally distributed by the Early Jurassic (Hsiou et al. 2015), is notable for its mesiodistally elongated teeth, which occluded precisely, resulting in a self-sharpening cutting surface (Whiteside and Duffin 2017) and leaving conspicuous wear facets on the opposing surfaces of the dentary and maxilla (Fraser 1988). The genus was named for the type species Clevosaurus hudsoni (Swinton 1939) from the Late Triassic fissure infills of Cromhall Quarry, England. Since 1939, nine species of Clevosaurus have been identified (Fraser 1988; Sues et al. 1994; Wu 1994; Säilä 2005; Bonaparte and Sues 2006; Klein et al. 2015; Keeble et al. 2018), five of them from the British Isles. A tenth species, Clevosaurus latidens (Fraser 1993), considered as closer to opisthodontians than to other species of Clevosaurus by Martinez et al. (2013) and Hsiou et al. (2015), was recently reassigned to the distantly related genus Fraserosphenodon (Herrera-Flores et al. 2018).
Recently, the anatomy of a Clevosaurus hudsoni specimen was described from CT scan data by O’Brien et al. (2018), and likewise for the new species Clevosaurus cambrica from Pant-y-ffynnon quarry, Wales, by Keeble et al. (2018). Both species are from fissure localities dated as Rhaetian (Whiteside et al. 2016) but on separate palaeo-islands. The CT scans reveal detailed cranial anatomy that had previously been poorly known for Clevosaurus. Both species differ in dentition, both in terms of tooth number and, as revealed here, tooth shape. Because of the prevalence of Clevosaurus in their faunas, comprising sphenodontians and in some deposits mammals (e.g., in Pant quarry where Clevosaurus convallis co-occurs; Säilä 2005), as well as early crocodylomorphs and dinosaurs, and the suggestion of diverse diets and possible dietary partitioning in these assemblages (Jones 2008; Gill et al. 2014), it is possible that these differing dentitions indicate differing dietary ecologies.
Here, we reconstruct the cranial morphology of these two species in three dimensions using the CT scan data sets, and where appropriate amending previous work. From additional work on the CT scans, we reveal new details of the braincase of C. hudsoni, and we show that NHMUK PV R37014 (C. cambrica), previously thought to comprise fragmentary remains, in fact comprises a near complete skull. The reconstructed skulls of these two specimens are compared with each other and with modern Sphenodon. In addition, we re-examine syntype dentary material of C. hudsoni described by W.E. Swinton and specimens from the collection of P.L. Robinson (both in the NHMUK) and compare them to NHMUK PV R36832.
Institutional abbreviations.—NHMUK, Natural History Museum, London, UK.
Material and methods
Data sets.—We examined two specimens of Clevosaurus, a partial disarticulated skull, cervical vertebrae and forelimb of C. hudsoni (NHMUK PV R36832) and material from the type series of C. cambrica (NHMUK PV R37013, R37014). These specimens had previously been CT-scanned and segmented, and then described by O’Brien et al. (2018) and Keeble et al. (2018), respectively (Fig. 1). Additionally, a CT scan surface data .stl file for the skull of an adult Sphenodon (YPM 9194) was taken from the Digital Morphology database (Maisano 2001; http://www.digimorph.org) for comparison.
Using Avizo 8.0 (Visualization Sciences Group) for the duration of the project, we completely re-segmented the cranial remains of C. cambrica. The right palatine was revealed to be complete, with the maxillary process and facet intact. Many undocumented bones were also found in the CT scan dataset and were segmented, revealing that rather than consisting merely of some partial mandibles, maxillae and palatal bones, as shown in Keeble et al. (2018), NHMUK PV R37014 consists of a near complete skull. The dermal roofing bones and palate were found to be complete for both sides, and almost the entire left side of the skull was found to be present, including a quadrate, squamosal, postorbital, postfrontal, prefrontal, supratemporal, both premaxillae, as well as much of the braincase. The only bones absent, or unidentifiable in the scans, were the jugals and stapes. Also unrecorded, coronoid bones were identified in both specimens. The data for C. hudsoni was also partially re-segmented where bones were found to be incompletely segmented, especially around the braincase and quadrate.
Surfaces were then generated for both updated datasets and reduced to 500 000 faces in Avizo (Visualization Sciences Group), and then exported as .stl files, following the protocol of Lautenschlager (2016) for use in Blender (Blender foundation, https://www.blender.org/).
Details of the skull morphology revealed by CT scans of NHMUK PV R36832 were confirmed by examination of the type specimen of C. hudsoni to verify the specimen’s identity. We examined the dentaries belonging to ten specimens of C. hudsoni in the collections of Pamela L. Robinson and William E. Swinton (eight and two specimens, respectively) under a light microscope, in addition to a maxilla and palatine from W.E. Swinton’s collection (all in NHMUK).
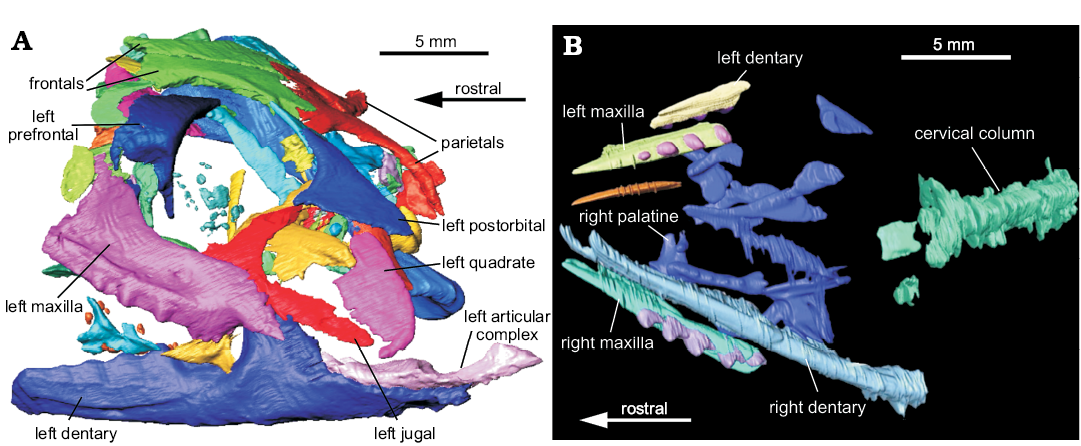
Fig. 1. The two surface data-sets of lepidosaur rynchocephalians. A. Clevosaurus hudsoni Swinton, 1939 (NHMUK PV R36832) from Rhaetian, Cromhall Quarry, England, UK, the left side of the digitally segmented skull in lateral view, adapted from O’Brien et al. (2018). B. Clevosaurus cambrica Keeble, Whiteside, and Benton, 2018 (NHMUK PV R37014) from Late Triassic, Pant-y-ffynnon, Wales, UK, available prior to this paper, the digitally segmented skull in ventral view, from Keeble et al. (2018).
Digital restoration of the skulls of Clevosaurus hudsoni and C. cambrica.—The surface files generated for the data set of Clevosaurus hudsoni were opened in Blender. Individual cranial bones were isolated and labelled, and all postcranial bones were removed from the dataset, as were the sclerotic ossicles, as they were too fragmentary to form part of the reconstruction. Repairs were made to minor holes and breaks in bones, as well as to remove segmentation artefacts where possible. The bones of the skull were reassembled in life position. Following the digital restoration methods of Lautenschlager (2016), bones that were either missing or incomplete on one side of the skull, but present on the other, were duplicated and mirrored. Because much of the right side of the skull is missing, many of the bones were duplicated from the left. Due to noticeable asymmetry in the dermal roofing bones (parietals and frontals), which were still articulated and complete on both left and right sides of the skull, these bones were divided down the middle, with the right parietal and frontal being replaced with mirrored duplicates from the left side. This was necessary, as their previous asymmetry had posed difficulties in spatial positioning and articulation on the right side of the skull.
Some flattening of skull bones was inferred, although this mostly had to be ignored because of the difficulty of confirming the extent of the deformation (see Lautenschlager 2016), but some corrections were made to the frontals and parietals. The rostral-most portions of the frontals were selected and bent ventrally to meet the maxillae as they would have in life, and the parietal process was shifted back to meet the posterior process of the squamosal.
Some bones were fragmentary in the original dataset, such as the left maxilla and dentary, both of whose anterior tips were broken and missing in NHMUK PV R36832. These were repaired with scaled-up portions of the more complete bones of C. cambrica. Other bones were so poorly preserved, damaged or missing in their entirety that they could not be repaired in this way, and these include the premaxillae, nasals, supratemporals, palatines and vomers. These bones were reconstructed by taking the same bones in C. cambrica and manually adjusting them with reference to the illustrations of Fraser (1988) and Robinson (1973), using the modelling tools in Blender. For example, the complete palatine of C. cambrica was scaled up and imported into the reconstruction of the C. hudsoni skull. The completed skull of C. hudsoni was then exported from Blender as a surface .stl file.
Following the same approach, we also reconstructed the skull of C. cambrica. Because it was much more complete, the only bones to be replaced with those of C. hudsoni were the jugal and stapes. The postorbital and prefrontal of C. cambrica had both become fragmented into smaller pieces, but these fragments were large enough to be restored manually, with reference to C. hudsoni.
Images of both reassembled skulls (see SOM 1–3, Supplementary Online Material available at http://app.pan.pl/SOM/app64-Chambi_Trowell_etal_SOM.pdf) were then imported into Artweaver (Boris Eyrich Software), where they were drawn over to form fully reconstructed skull. Measurements were taken from the reconstructed skulls of C. hudsoni and C. cambrica, as well as Sphenodon punctatus (from Maisano 2001), and these were converted to three ratios for comparison: maximum cranial height to cranial length, maximum cranial height to maximum cranial width, and maximum cranial width to cranial length.
Measurements of second moment of area of the ultimate dentary tooth for Clevosaurus hudsoni, C. cambrica and Sphenodon.—The ultimate (= most posterior) mandibular tooth of Clevosaurus cambrica, C. hudsoni and Sphenodon was isolated for biomechanical study. Cross sections were taken down the height of each of these teeth, with the base defined by the lowest points on either side of the tooth where it meets the dentary bone. Cross sections were separated by 10% of total tooth height, and starting 10% down from the tooth apex, resulting in a total of ten measurements for each tooth, following the methodology of Gignac and Erickson (2015). These measurements are independent of tooth height and provide a direct comparison between changing tooth shape and morphology dorso-ventrally. The section measurements were taken in Blender, using the ruler tool and we generated planar surfaces to indicate where each of these measurements would be taken. Dorsal screen shots were then taken at the level of each of these measurements, each featuring the ruler tool recalibrated for each shot to account for changes in the distance of the viewer. These images were then opened in Artweaver, and the cross-sectional area of each of the teeth was converted into a single solid black surface. Images were then converted into 8-bit TIFF files in ImageJ. The area of each was calculated using the measurement tool in ImageJ, each time calibrating for the differences in the distance for the teeth using the values obtained earlier from the ruler tool. Here we treat the teeth as having homogeneous material properties and calculated for each of the teeth the second moment of area (Ix, Iy). To calculate the change in the distribution of a tooth around the central point of mass down the tooth height, Ix was divided by Iy.
Forelimb bone measurements for Clevosaurus hudsoni and C. cambrica.—The forelimb of Clevosaurus cambrica was resegmented so that the individual bones could be separated and measured in Blender, while measurements for C. hudsoni were taken directly from the graphs provided in O’Brien et al. (2018) using the ruler tool in ImageJ. Data from both species were then compared with data on forelimb bone ratios (between the length of the ulna, radius and humerus, and with total forelimb length and skull length) for C. hudsoni provided by Fraser (1988: 157, tables 2, 3). The measurements were converted into ratios to allow for size-independent comparison. The data set was too small for statistical comparison, so direct comparisons were instead made.
Systematic palaeontology
Class Reptilia Laurenti, 1768
Order Rhynchocephalia, Günther, 1867
Family Sphenodontidae Cope, 1870
Subfamily Clevosaurinae Bonaparte and Sues, 2006
Genus Clevosaurus Swinton, 1939
Type species: Clevosaurus hudsoni Swinton, 1939; see below.
Clevosaurus hudsoni Swinton, 1939
Material.—Referred specimen, NHMUK PV R36832, is a near complete skull, with much of the left side of the skull still articulated.
Description.—The scans of Clevosaurus hudsoni (NHMUK PV R36832) showed a few bones that had been incompletely segmented or overlooked, including the right postorbital, both quadrates, the right pterygoid, the braincase, and the coronoid process.
Postorbital: The right postorbital (Fig. 2A) had been segmented but not identified. Its morphology was a near perfect mirror of the left postorbital (which is described in O’Brien et al. 2018), but it has a damaged postero-ventral edge and a better-preserved ventral process. The ventral process bends anteriorly at a roughly 90° angle and extends medially to form a shelf that would have partially wrapped around the jugal when articulated, and would have formed the posteroventral rim of the orbit.
Squamosal: Both squamosals have been identified (Fig. 2B, C), with the left squamosal being better preserved than the right. The left squamosal had been correctly identified by O’Brien et al. (2018), though it was later labelled as a left pterygoid flange; notably, the left squamosal had undergone more rotation than any of the other bones in the skull, nearly ~180°, which probably led to this misidentification. Its identity as a squamosal was confirmed based on its differences from the complete right pterygoid, including being much thicker than the delicate pterygoid flange, its matching mirrored morphology with a similar bone that we identify as the right squamosal, and its defined postorbital facet, quadrate process and medially recurved posterior process. The anterior process is damaged for both sides, but part of the postorbital facet on the left squamosal is well defined, the ventral process is long and would have extended three-quarters of the height of the quadrate to which it articulated. The posterior process curves posteromedially towards the posterior process of the pterygoid and bears a rough uneven surface that would have likely formed an articulation site for the supratemporal.
Quadrate: The left quadrate of NHMUK PV R36832 (Fig. 2D) was found to be more complete than previously described, and its morphology appears to be typical of C. hudsoni. The right quadrate (Fig. 2E) was also located out of place, thrust upwards and out of position by the forelimb bones that were found within the skull (O’Brien et al. 2018); the posterior region of the “lateral plate” appears to have been damaged. The condyles for both quadrates are preserved and they are separated by a deeply concave cotyle. Dorsally, the quadrate extends into two connected structures either side of the condyles, the “lateral plate” which anteriorly bears the quadrato-jugal foramen, and the medially placed “vertical strut”, which is a tall columnar structure with expanded ends. Both converge anteriorly, and extend to form the quadrato-pterygoid process, which is a thin flange, reinforced by a robust ventral ridge.
Palatine: On re-examination of NHMUK PV R36832, we find the left palatine possesses nine teeth (Fig. 2F), with a thin surface of bone running beneath them all, possibly representing the raised lateral ridge typically found on the palatine of C. hudsoni, but at the resolution in this part of the CT scans, it is hard to discern bone from matrix. In O’Brien et al. (2018), these teeth are segmented but some were suggested to be pterygoidal teeth, though the left pterygoid is absent. We confirm that the teeth are from the palatine. Nine palatine teeth is more than the five to six teeth noted for C. hudsoni by Fraser (1988: 137), and suggests there may be a greater range in morphology for this species than previously described.
Pterygoid: The right pterygoid, already partially segmented, was re-examined, and we found the thin quadrato-pterygoidal flange (Fig. 2G) and lateral ectopterygoidal-pterygoidal process still articulated with the right ectopterygoid. The complete pterygoid matches the description given for this species by Fraser (1988: 138, fig. 16). The anterior region of the pterygoid bearing two rows of teeth is described in O’Brien et al. 2018. The two tooth rows of the left pterygoid, and its lateral process were also located, still articulated with the left ectopterygoid, but the resolution in this region of the scans was too low to discern anything more.
Parabasisphenoid: The basipterygoid processes in Clevosaurus hudsoni (NHMUK PV R36832) had previously been segmented but had been left unlabelled, whereas the parasphenoid, basisphenoid and left pila antotica process had not been segmented. The basisphenoid and parasphenoid are fused with no sutures discerned between them (Fig. 2H). Anteriorly, the basisphenoid forms the paired basipterygoid processes, both of which were preserved intact, having been previously segmented but left unidentified by O’Brien et al. (2018). Located between the two basipterygoid processes, the cultriform process is preserved (Fig. 2H) and is elongate compared to that of previous reconstructions of C. hudsoni (Fraser 1988: fig. 17B). Also, its base is nearly level with the basipterygoid processes rather than positioned anteriorly, as in C. brasiliensis (Bonaparte and Sues 2006).
Prootic: The left pila antotica process was identified from amongst previously partially segmented bone. It is a tall, flattened slender process that has an antero-posteriorly expanded dorsal head (Fig. 2H).
Coronoid: The left coronoid bone is preserved (Fig. 2I) and is reported here for the first time in Clevosaurus hudsoni. The coronoid bone had become vertically and medially displaced from the coronoid process and was segmented as part of the dentary in O’Brien et al. 2018, artificially raising its height in the original model. It is a small latero-medially compressed bone that forms the medial portion of the coronoid eminence.
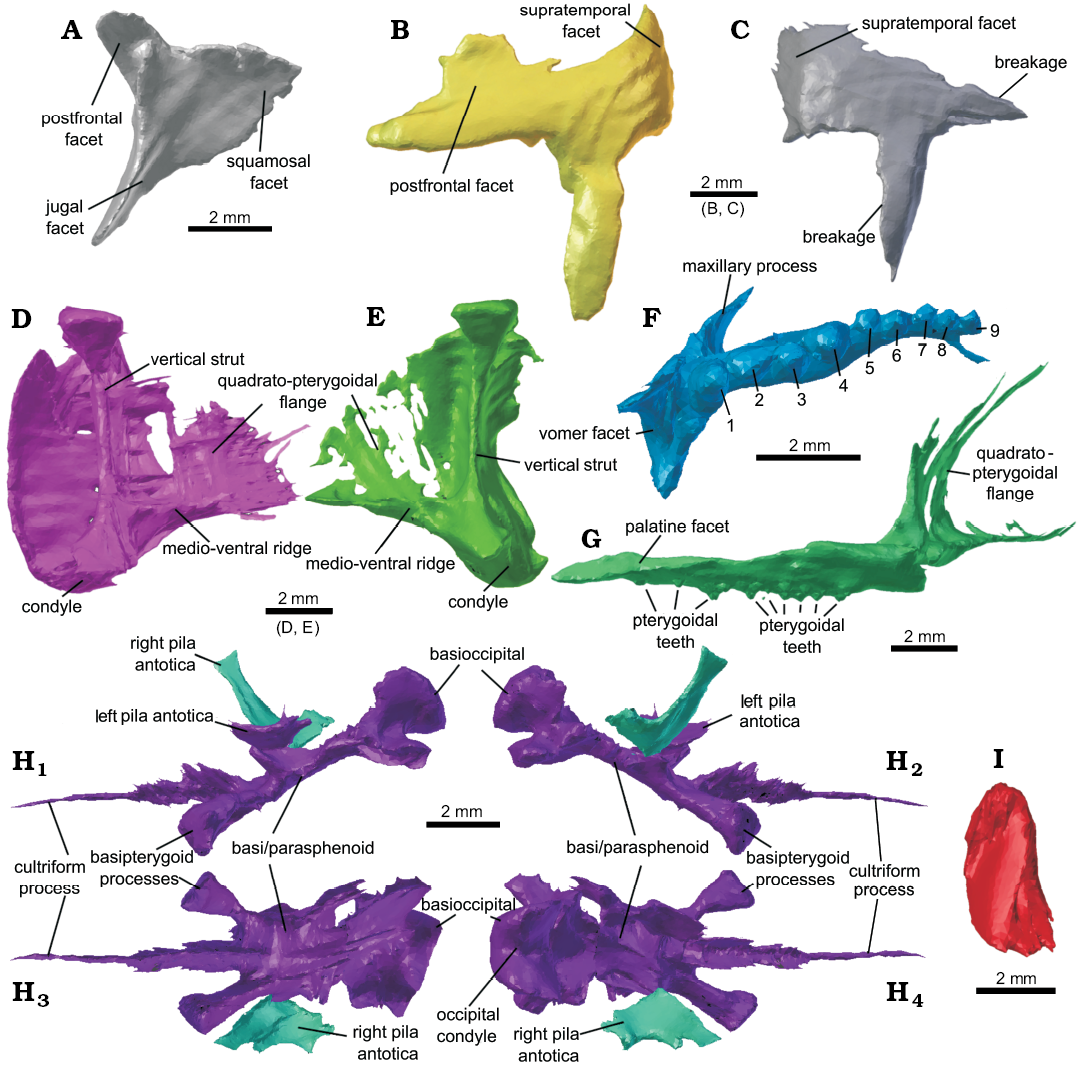
Fig. 2. Newly identified cranial bones present in lepidosaur rhynchocephalian Clevosaurus hudsoni Swinton, 1939 (NHMUK PV R36832) from Rhaetian, Cromhall Quarry, England, UK. A. The right postorbital in medial view, identified from amongst unidentified bones from the original surface dataset created by O’Brien et al. (2018). B. The left squamosal in lateral view, re-identified here from its previous suggested identity of a pterygoid flange. C. The right squamosal in lateral view, like A identified here from amongst previously unidentified bones from the original dataset. D. The left quadrate in medial view, partially resegmented from the original dataset to be more complete. E. The right quadrate in medial view, newly segmented and identified here. F. Left palatine in ventral view, partially resegmented from the original data set, note the number of palatine teeth (nine). G. The right pterygoid in medial view, partially resegmented from the original dataset, seen here with a newly segmented pterygoid flange. H. The newly identified pila antotica, and newly segmented para- and basisphenoids in left (H1) and right (H2) lateral; ventral (H3), and dorsal (H4) views. I. The left coronoid in medial view.
Remarks.—Examination of the Clevosaurus hudsoni dentaries at the NHMUK revealed two possible morphotypes. The additional dentary teeth of NHMUK PV R36832 and the syntypes from W.E. Swinton’s original description (NHMUK PV R5939a and R5939b) have a simple structure, where the tooth shape is triangular in labial profile with the “extensive antero-lateral flange” described by Fraser (1988: 144) reduced or absent in the syntypes. There is also little or no overlap between teeth and with the reduced antero-lingual escape structures, together forming a straighter cutting edge (Fig. 3A–C), which could be related to ontogenetic change that is beyond the scope of this study. However, we note that the C. hudsoni specimen NHMUK PV R36832, which, although smaller, shares a similar tooth morphology to the syntypes and is reported to be an adult specimen (O’Brien et al. 2018: 191; it also lacks any remnant hatchling dentition). Furthermore, although a similar change occurs in some of the most heavily worn of the dentaries of C. convallis, with a reduction in anterolateral flange length and tooth overlap, it only affects the anteriormost of the teeth, and not the entire row (Säilä 2005). Several of the dentaries in the P.L. Robinson collection (NHMUK PV R37270, R37271, and R37272) show a greater resemblance to the reconstructions made by Fraser (1988: 142), with extensive antero-lateral flanges, noticeable overlap between teeth and large antero-lingual escape structures, so that the cutting edge of the teeth was en echelon (Figs. 3D–F). The syntype palatine in W.E. Swinton’s collection (NHMUK PV R5939c) bears four (possibly a fifth broken) lateral teeth that scale down in size posteriorly and bear posterolateral wear facets, and the maxilla (NHMUK PV R9249) bearing four additional teeth and a morphology closely resembling that of NHMUK PV R36832 (see SOM 4 and 5). Together, this information confirms that our specimen is conspecific with the syntype specimens of C. hudsoni.
Stratigraphic and geographic range.—Upper Triassic (Rhaetian) fissure infill within Lower Carboniferous Limestone, Cromhall Quarry, near Cromhall Village, Gloucestershire, South-West England, UK (NGR ST 704916).
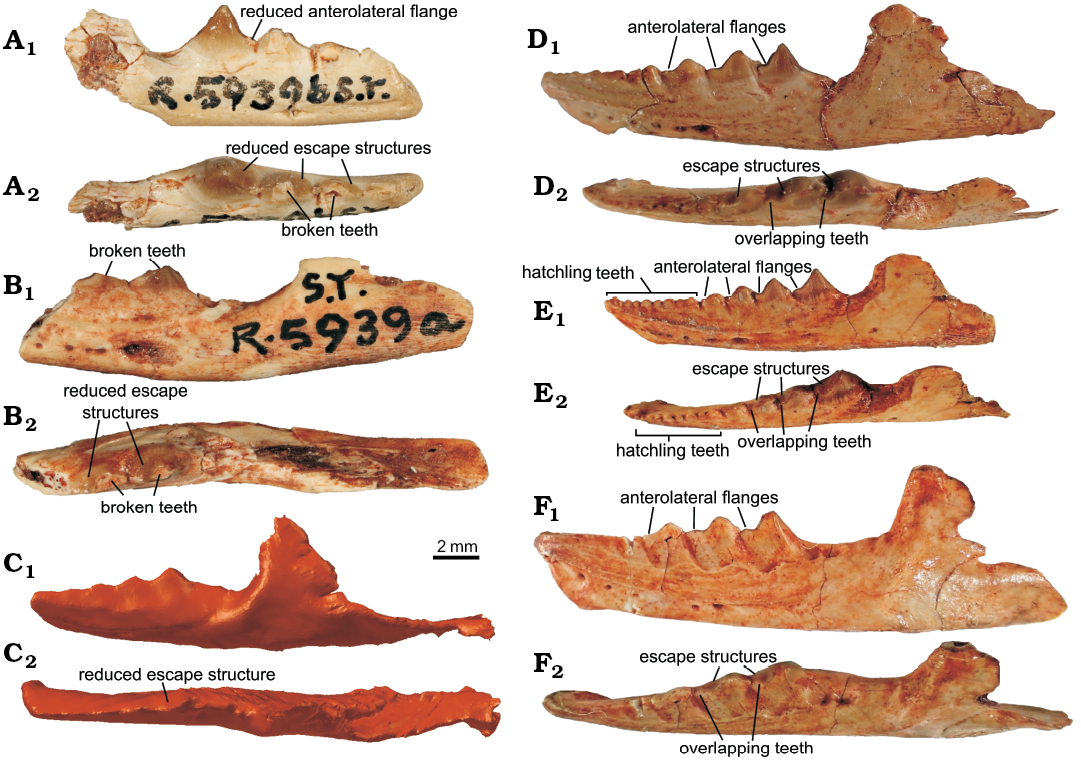
Fig. 3. Dentaries of lepidosaur rynchocephalian Clevosaurus hudsoni Swinton, 1939 from Rhaetian, Cromhall Quarry, England, UK, including the two syntypes (A, B) belonging to W.E. Swinton’s collection. A. NHMUK ST R5939b. B. NHMUK ST R5939a. C. NHMUK PV R36832. D. R 37270, note the highly worn hatchling teeth. E. NHMUK PV R 37271, note the presence of hatchling teeth. F. NHMUK PV R 37272. In lateral (A1–F1) and dorsal (A2–F2) views.
Clevosaurus cambrica Keeble, Whiteside and Benton, 2018
Material.—Referred specimens include a partially disarticulated near complete skull and at least five cervical vertebra, NHMUK PV R37014, and a left forelimb associated with shoulder elements and dorsal vertebra, NHMUK PV R37013.
Description.—The original report of Clevosaurus cambrica (NHMUK PV R37014; Keeble et al. 2018) identified the scanned materials as a few cervical vertebrae and the skull below the palate level, comprising portions of both maxillae, mandibles, palatines and pterygoids. A close inspection of these scans showed that the left side of the partial skull of C. cambrica is near complete (Fig. 4), though missing several of the postero-lateral bones, including the jugal, postfrontal, postorbital, quadrate, squamosal and prefrontal. The right side of the skull has sustained more physical damage, with several bones (e.g., left maxilla, dentary, prefrontal, and postorbital) fragmented. The bones are all closely associated, and in a few cases show some articulation. The whole skull was ventro-dorsally compressed post-mortem, but was only minimally deformed, and the bones appear to have fallen between and on either side of the mandibles before fossilisation took place. There are also five articulated elements originally identified as cervical vertebrae by Keeble et al. (2018), though they considered that the most anterior element might be a basioccipital, which we confirm here. Much of the braincase of C. cambrica is also present. The only missing bones, or at least not visible at the resolution of the scans, are the jugals and stapes.
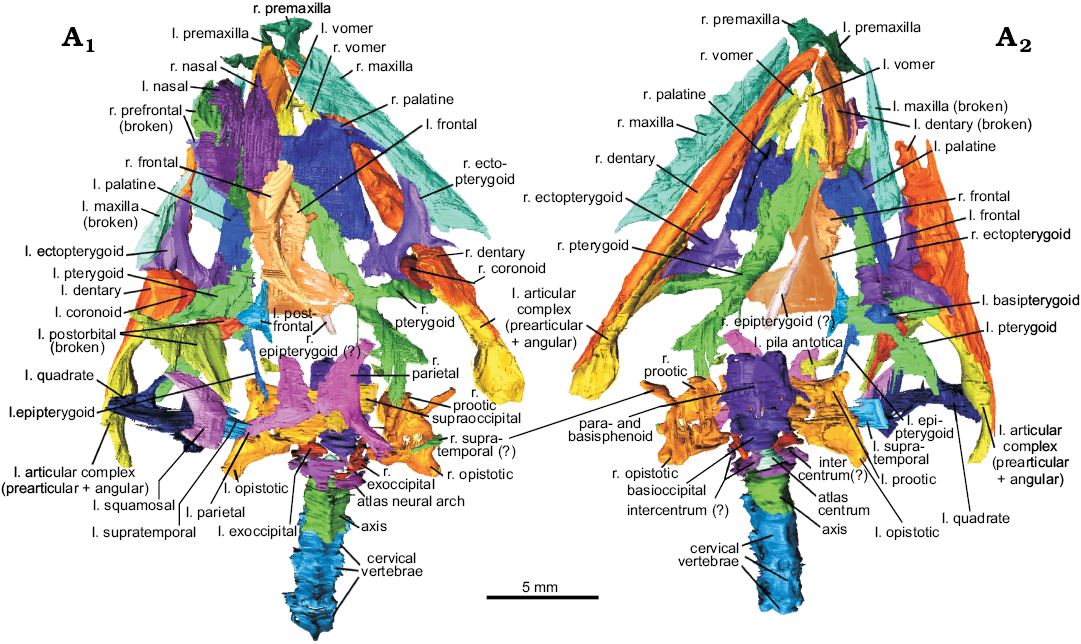
Fig. 4. The resegmented bones of lepidosaur rynchocephalian Clevosaurus cambrica Keeble, Whiteside, and Benton, 2018 (NHMUK PV R37014) from Late Triassic, Pant-y-ffynnon, Wales, UK; in dorsal (A1) and ventral (A2) views. Abbreviations: l., left; r., right.
The left forelimb (humerus, ulna, and radius) and partial pectoral girdle (scapulae and coracoids) of this same specimen were also resegmented to refine the resolution (Fig. 5). In addition to the scapula already identified, two previously uncertain elements were identified as a second scapula and a coracoid, and a fourth previously unsegmented bone was identified as a second coracoid. Further dorsal vertebrae were identified running in a column beneath the forelimb, forming a total of six articulated vertebrae. The carpal mentioned by Keeble et al. (2018) could not be located in this reassessment of the CT scans.
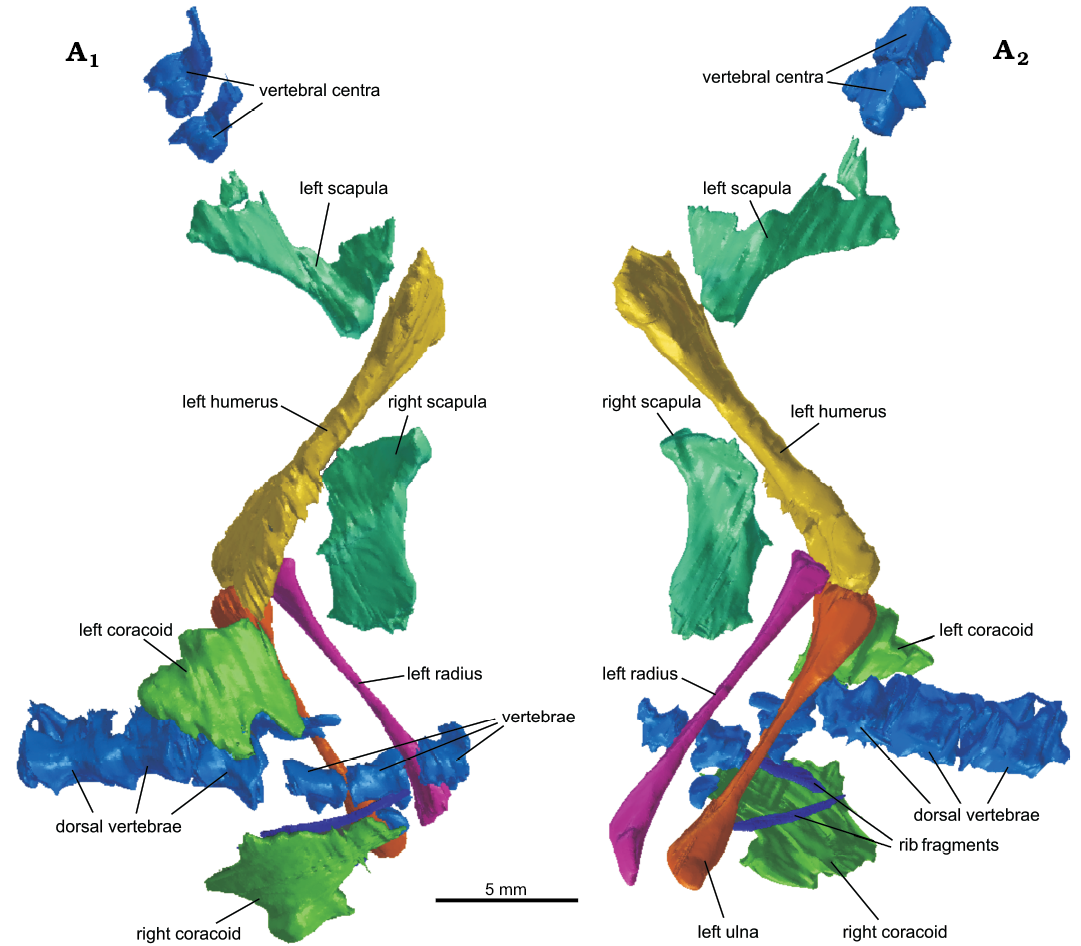
Fig. 5. The newly re-segmented surface of the shoulder elements, left forelimb and partial vertebral column of lepidosaur rynchocephalian Clevosaurus cambrica Keeble, Whiteside, and Benton, 2018 (NHMUK PV R37013) from Late Triassic, Pant-y-ffynnon, Wales, UK; in medial (A1) and lateral (A2) views.
Premaxilla: The left and right premaxillae of Clevosaurus cambrica (NHMUK PV R37014) are preserved, with the right showing a more complete dorsal process (Fig. 6A). The two premaxillae are found in close association with the anterior edge of the maxilla. The internarial process is relatively short but robust and tapers to a point. The dorsal process makes the posterior edge of the nasal openings, and there the maxillary facet is a posterior concavity that bifurcates to form two flattened processes which extend beneath the medial surface of the maxilla, as in most Clevosaurus. The exception is C. convallis which lacks the bifurcating dorsal maxillary process and is more robust (Säilä 2005: 820). The unexpectedly short length of both the internarial and dorsal maxillary processes preserved here suggests they might have been broken but the internarial process of C. convallis is similarly short (Säilä 2005: 820). The portion of the dorsal maxillary process ventral to the start of the bifurcation is relatively longer than in C. hudsoni (Fraser 1988: fig. 5) and C. sectumsemper (Klein et al. 2015: fig. 3A, B). The number of teeth present on the premaxilla is not clear, either due to the resolution of the scans or perhaps indicating that the worn premaxilla teeth are already encased in secondary bone and dentine so that they form a single conical structure, much like that observed in other species of Clevosaurus (Fraser 1988; Sues et al. 1994; Säilä 2005; Bonaparte and Sues 2006). However, in C. sectumsemper this overgrowth is not observed and this is a diagnostic feature of the species (Klein et al. 2015: 409).
Maxilla: While the holotype maxilla (NHMUK PV R37016) has been described (Keeble et al. 2018), the maxillae in NHMUK PV R37014 were incompletely segmented (Fig. 6B). The right maxilla is complete and bears six acrodont teeth. The left maxilla has broken in two, at approximately two-thirds along its length, and its anterior portion bears four acrodont teeth. The posterior portion remains articulated with the left ectopterygoid, and it appears to be damaged and teeth cannot be clearly discerned. The maxilla is as described by Keeble et al. (2018), but bears a small sixth additional tooth, adding to the ‘five prominently flanged teeth’ of Keeble et al. (2018), and is approximately 12 mm in length. The marginal tooth row is offset posteromedially so that it is inset from the lateral margin of the maxilla posteriorly, as in C. bairdii (Sues et al. 1994: 329). However, unlike C. bairdii, the premaxillary process is pronounced in C. cambrica. A lip of secondary bone appears above the 2nd to 5th maxillary tooth (Fig. 6B2) and no hatchling teeth appear to be present. The maxilla of C. cambrica is similar in shape to that of C. hudsoni (Fraser 1988: fig. 6; O’Brien et al. 2018: fig. 5A), although superficially the suborbital margin formed by the dorsal process of C. cambrica is more recurved, resembling that of C. bairdii (Sues et al. 1994: fig. 1B). It is also posteriorly longer, with the facial process of the maxilla being less than half the length of the suborbital length, suggesting larger orbits, and this morphology is similar to C. brasiliensis albeit with a shallower dorsal process (Hsiou et al. 2015: fig. 2C).
Nasal: Nasals are rarely preserved in clevosaurs as they are fragile (Fraser 1988: 131–132), but here both nasals are present as large, thin, concave and simple structures (Fig. 6C). The left nasal has some damage to its lateral edge where the left prefrontal has been thrust upwards through it. It is likely both nasals were damaged, as the lateral and anterior edges of both appear to be incomplete. The nasals in C. brasiliensis are steep and fused to the frontals (Bonaparte and Sues 2006: fig. 1A, B; Hsiou et al. 2015: 7), but here we find both nasals isolated from the frontals and their length suggests that they could not have been angled as steeply or fused to the frontals.
Frontal: Both frontals are present (Fig. 6D) and they are broken apart at an asymmetrical angle so that the left frontal posteriorly is still sutured with a portion of the right frontal, while a larger portion of the right frontal has become displaced and lies beneath the left frontal. While some detail like the prefrontal facets is preserved, finer detail such as the sutures between the two frontals does not appear in these scans. When their positions are corrected, the two frontals together form an hour-glass shape in dorsal view.
Parietal: Both parietals (Fig. 6E) are preserved in close association, but there appears to be some asymmetry in the parietal processes, and it is possible that the left parietal sustained damage that is not clear on the scans. The parietals have disassociated from the frontals, which appear to have been moved anteriorly with the rest of the skull. The left parietal process is positioned at a ~90° angle to the left parietal table horizontally, and is vertical and flat, and this is likely the result of a break. The right parietal process is angled further backward at about 120° horizontally and slopes ventrally at a shallow angle vertically; the right parietal process also bears a bump on its ventral surface where it joins the right parietal. There are distinct slot-like facets for the supratemporals on the posterior-most ends of both parietal processes. The pineal foramen is not visible at current resolution, but there may have been damage in this region. As with the frontals, the lateral parietal margins are much more robust than the medial ones. As in Clevosaurus hudsoni and C. brasiliensis (Fraser 1988: 174; Hsiou et al. 2015), the parietal table of C. cambrica lacks a true parietal crest, unlike Sphenodon.
Prefrontal: The left prefrontal (Fig. 7A) has broken into at least three pieces, perhaps when the prefrontal was forced upwards through the left nasal. The largest fragment is the keeled ventral portion of the prefrontal, which would have formed the anterior margin of the orbital rim, a large flat bone that is angled at roughly 90° at its keel. The second fragment is slightly smaller and comprises the posterodorsal process of the prefrontal, which forms the anterodorsal edge of the orbital rim. The third fragment is a simple slight concave surface that would have formed the dorsal portion of the prefrontal and would have articulated with the nasal in life. When reconstructed, the prefrontal is large and semi-lunate in shape. However, we note that the damaged prefrontal of Clevosaurus cambrica might have been deeper and more robust than reconstructed here, meaning the skull overall might have been broader.
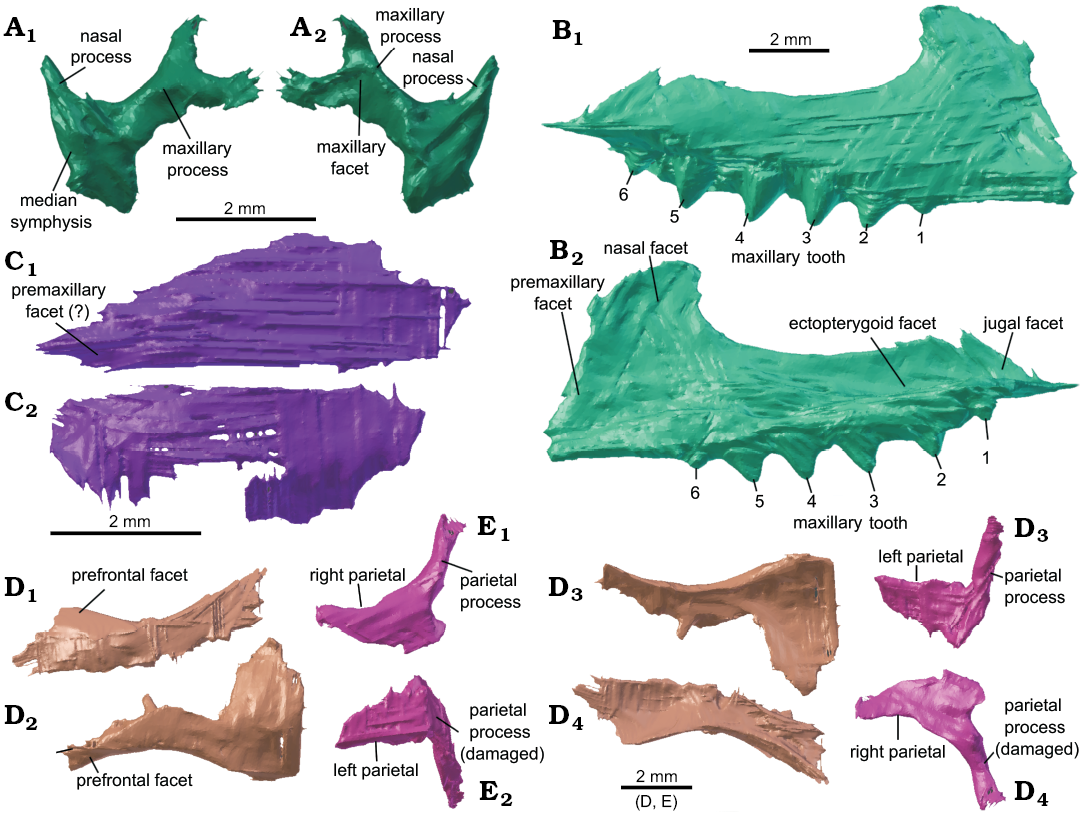
Fig. 6. Cranial bones of lepidosaur rynchocephalian Clevosaurus cambrica Keeble, Whiteside, and Benton, 2018 (NHMUK PV R37014) from Late Triassic, Pant-y-ffynnon, Wales, UK. A. The right premaxilla in medial (A1) and lateral ( A2) views. B. The right maxilla in lateral (B1) and medial (B2) views. C. The right (C1) and left (C2) nasal in dorsal view. D. The right (D1, D4) and left (D2, D3) frontals in dorsal (D1, D2) and ventral (D3, D4) views. E. The right (E1, E4) and left (E2, E3) parietals in dorsal (E1, E2) and ventral (E3, E4) views.
Postfrontal: The left postfrontal (Fig. 7B) is a small triradiate bone with a slightly recurved medial surface and it is closely associated with the left frontal and is nearly articulated with the dorsal process of the postorbital.
Postorbital: The left postorbital (Fig. 7C) is a large triradiate bone with a broad and elongated ventral jugal process that extends nearly as far as the maxilla and bears a broad jugal facet on its medial surface (Fig. 7C2). It was found to have broken into at least two pieces, with the dorsal portion still articulated with the postfrontal but having been displaced ventrally beneath the lower portion of the postorbital, and the dorsal portion appears to have been broken a second time so that the postfrontal process is at an angle to the rest of the element, which was corrected by digitally rebreaking the bone at this point and rotating it dorsally. Unlike C. brasiliensis (Hsiou et al. 2015: 8), C. cambrica has both a long ventral and dorsal process, which encompasses almost the entire posterior orbital rim.
Squamosal: The preserved left squamosal lacks the anterior flange that would have articulated medially with the postorbital (Fig. 7D), but the tapering ventral quadrate process, which would have extended approximately half the height of the quadrate, as in C. brasiliensis and C. wangi (Bonaparte and Sues 2006; Jones 2006), and the medially recurved posterior supratemporal process are both well preserved.
Supratemporal: Both supratemporals appear to be preserved. The left supratemporal is still articulated with the left parietal, and it is a thin medially convex plate-like bone (Fig. 7E). As in Clevosaurus hudsoni (Fraser 1988: 134), it is broadest dorsally and narrows ventrally, and this broad extension would have articulated with the squamosal in life. The possible right supratemporal is part of a narrow, flattened process that appears to extend from the anterior surface of the braincase (see Fig. 4A2). Its identity is inferred from its position relative to the braincase, but it could be a fragment of bone from elsewhere in the skeleton. Another possible candidate for a right supratemporal is a fragment of bone overlying the right opisthotic (see Fig. 4A1). The shape of the supratemporal differs from that of C. brasiliensis, which is elongate (Hsiou et al. 2015) but similarly tapers anteriorly to contact the parietal process.
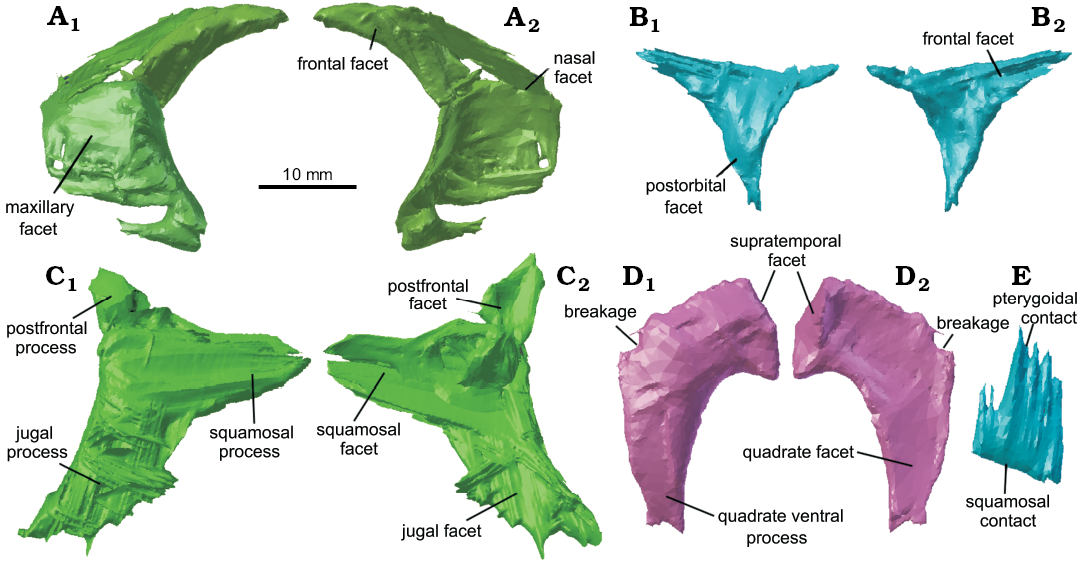
Fig. 7. Cranial bones of lepidosaur rynchocephalian Clevosaurus cambrica Keeble, Whiteside, and Benton, 2018 (NHMUK PV R37014) from Late Triassic, Pant-y-ffynnon, Wales, UK. A. The reassembled left prefrontal. B. The left postfrontal. C. The reassembled left postorbital. D. The left squamosal. E. The left supratemporal. In lateral (A1–D1, E) and medial (A2–D2) views.
Vomer: Despite their rare preservation in clevosaurs because of their fragility (Fraser 1988: 136), remarkably both vomers are present and intact in C. cambrica (Fig. 8B). The two vomers are in close association and located close to where they would have been in life. They are thin convex structures with robust medial and anterior margins. Vomerine teeth are hard to discern at the resolution of the scans, but at least two appear along the posteromedial edge, in the same position as in C. hudsoni (Fraser 1988: fig. 14) and C. brasiliensis, which latter species bears three teeth in this medial position (Hsiou et al. 2015: 10).
Palatine: Both palatines are present in Clevosaurus cambrica, with the right palatine well preserved and with the whole maxillary process intact and barely disarticulated (Fig. 8A), forming much of the boundary of the choanae. The palatines have a robust lateral raised ridge bearing six conical teeth, as in the C. hudsoni specimens described by Fraser (1988: 137), while medially the palatine forms a thin flattened flange that widens anteriorly and becomes more concave, bearing a single medio-anteriorly placed tooth. The palatine is distinct from that of C. minor as it possesses a raised lateral tooth ridge and concave vomerine facet (Fraser 1988: 159–160) not found in that species. The maxillary process is robust and bifurcates at the maxilla to form an extensive but thin surface of contact. The six laterally positioned teeth in C. cambrica, are the same as in C. hudsoni, C. brasiliensis, and C. sectumsemper (Fraser 1988; Hsiou et al. 2015; Klein et al. 2015), but greater in number than the five in C. convallis (Säilä 2005) or the four in C. minor (Fraser 1988), and fewer than the nine in C. hudsoni NHMUK PV R36832 or the seven in C. wangi (Wu 1994; Jones 2006). The lateral tooth row is also much straighter than in the Chinese clevosaurs C. wangi and C. petilus (Wu 1994; Jones 2006).
Epipterygoid: Both epipterygoids may be present. The left epipterygoid (Fig. 8C) is a tall, columnar and slender element that does not appear to have the medio-lateral flattening seen in the Clevosaurus hudsoni of O’Brien et al. (2018). It is positively identified as it is still articulated with the left pterygoid, albeit it has collapsed antero-posteriorly so that it lies parallel to the long axis of the skull, and the cephalic region appears to have been broken. There is a possible left epipterygoid but at the current resolution this cannot be confirmed, and its identity is debatable (see Fig. 4). It is a flattened rod-like bone. The shape of the left epipterygoid is much narrower and more columnar than in Sphenodon. The epipterygoid of C. bairdii is similar to that of C. cambrica in being “rod-like dorsally” and flattening ventrally towards its attachment above the basipterygoid joint (Sues et al. 1994); the epipterygoid has not be described in any other species of Clevosaurus.
Ectopterygoid: Both ectopterygoids are present and intact, and the left ectopterygoid is still articulated with the left pterygoid, palatine and a fragment of the maxilla. The ectopterygoid (Fig. 8D) is formed of a posteromedial robust rod-like bone that bifurcates at either end, with a broad lateral process that is more elongate anteriorly and would have had an extensive contact with the medial surfaces of maxilla and the jugal, as in Clevosaurus brasiliensis (Bonaparte and Sues 2006) and C. bairdii (Sues et al. 1994). The medial process by contrast is longer posteriorly, and this section forms a triangular facet that overlaps and articulates with the anteroventrally facing facet on the lateral posterior pterygoid flange, while anteriorly it forms a flattened short surface where the palatine would have articulated. Overall, the ectopterygoids appear very similar to the C. hudsoni specimens described by Fraser (1988).
Pterygoid: Pterygoids are present on both sides of the skull, though the right appears to be better preserved (Fig. 8D). Pterygoid teeth cannot be definitively distinguished at the resolution of the scans, but several small conical bumps are present on the flattened palatal shelf that appear to form two parallel rows, which differs from the three rows found in Clevosaurus brasiliensis (Hsiou et al. 2015). The transverse process that runs between the palatal shelf and the posterior quadrate flange is short, as in C. hudsoni (Fraser 1988: 137–138; NHMUK PV R36832). The quadrate flange is still articulated with the pterygoid flange of the left pterygoid. The anteriormost portion of the left pterygoid has broken off but remains articulated anteriorly with the right pterygoid. The pterygoids bear robust transverse ectopterygoid processes with triangular facets that articulate with the medial surface of the ectopterygoids. While the tooth number cannot be discerned, the overall shape of the pterygoids is as described for C. hudsoni by Fraser (1988) with no medial “neck” between the ectopterygoid processes and the basipterygoid facet as in most other Clevosaurus but differing in C. wangi where this region is described as elongate (Wu 1994; Jones 2006).
Quadrate: The left quadrate is preserved and appears to be complete (Fig. 8E), though at the CT scan resolution some detail is unclear. The medially placed vertical strut is recurved medially, and thickens towards both the cephalic and articular condyles, but notably more so for the articular condyle where it has a pronounced ventrally convex lateral keel that attaches to the lateral plate-like structure of the quadrate. The condyle is very similar to that described for the Clevosaurus hudsoni specimens of Fraser (1988: 136), with the anterior-posterior length much shorter than the width, and the cotyle being medio-laterally deeply concave. The medial edge of the condyle projects strongly ventrally much as in C. brasiliensis (Hsiou et al. 2015), but differing from C. hudsoni (Fraser 1988: 13a, 17a; NHMUK PV R36832). The thin blade-like pterygoid-quadrate process is preserved, extending medio-anteriorly from the vertical strut, and as in C. hudsoni, is reinforced by a more robust ventral edge, as also seen in C. hudsoni (NHMUK PV R36832; Fig. 2D, E). The medially recurved lateral side of the quadrate is very thin and poorly resolved here, but a quadratojugal foramen appears to be present. Relatively the quadrate of C. cambrica is much shorter than in C. hudsoni (NHMUK PV R36832) but is only moderately shorter than that of the C. hudsoni of Fraser (1988). The quadratojugal cannot be discerned from the quadrate, and we assume that the bones are fused.
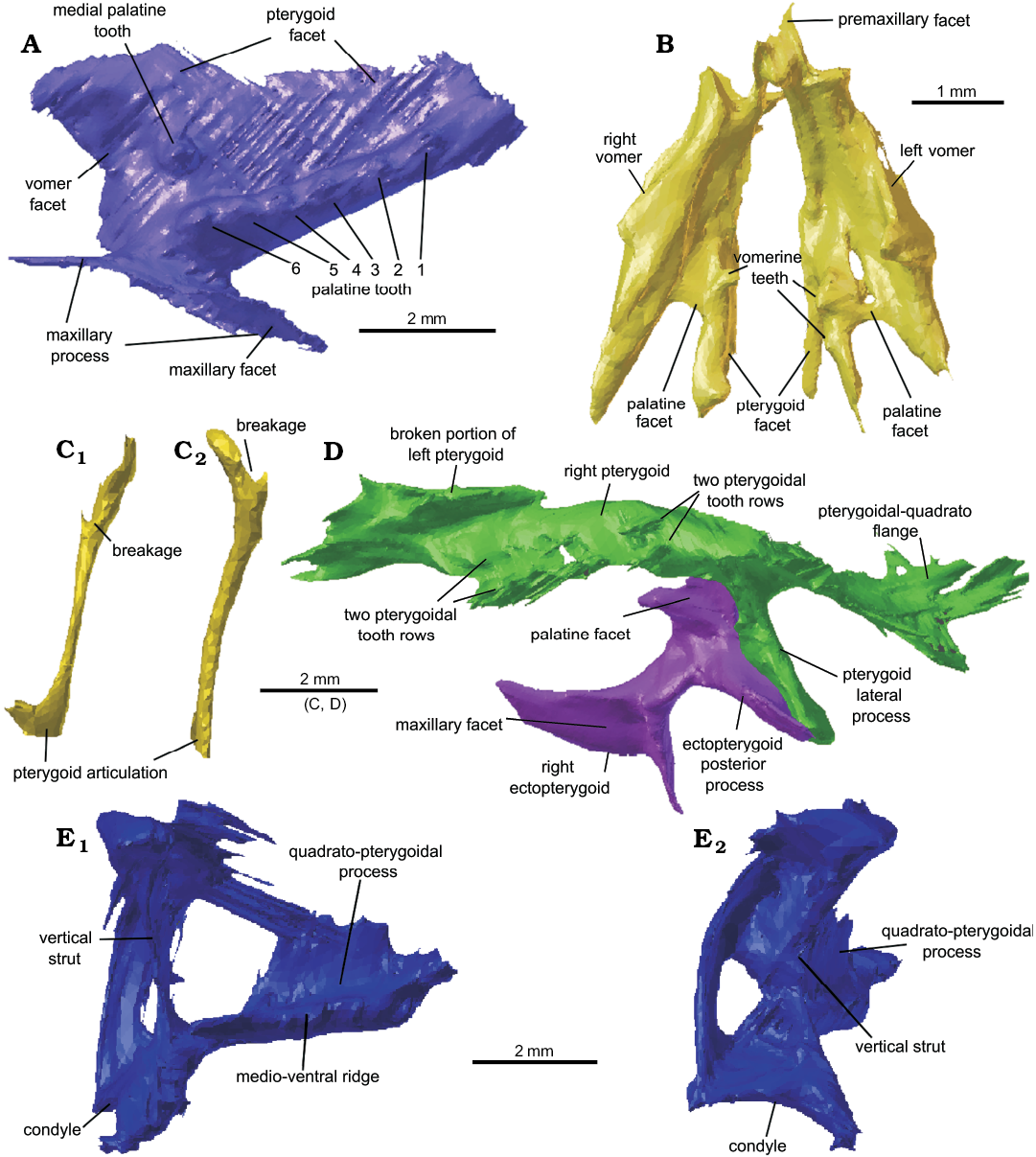
Fig. 8. Cranial bones present in lepidosaur rynchocephalian Clevosaurus cambrica Keeble, Whiteside, and Benton, 2018 (NHMUK PV R37014) from Late Triassic, Pant-y-ffynnon, Wales, UK. A. The right palatine in ventral view. B. The left and right vomers in ventral view. C. The left epipterygoid in lateral (C1) and posterior (C2) views. D. The left pterygoid and ectopterygoid in ventral view. E. The left quadrate in medial (E1) and posterior (E2) views.
Basioccipital and exoccipitals: The basioccipital remains articulated with the basisphenoid and parabasisphenoid (Fig. 9). The shape appears similar to that of Clevosaurus sectumsemper and C. hudsoni (Fraser 1988; Klein et al. 2015), with a lunate occipital condyle, and two robust and rounded basal tubera that project postero-laterally. The left and right exoccipitals, in close association with the basioccipital, appear to have sustained some damage and have broken apart asymmetrically; together they would have formed the lateral edges of the foramen magnum.
Parabasisphenoid: Aside from the basioccipital, the floor of the braincase is primarily formed by the fusion of the parasphenoid and basisphenoid (Fraser 1988: fig. 18; Hsiou et al. 2015: 12; figs. 9C, D) and is relatively much broader in Clevosaurus cambrica than in C. hudsoni (Fraser 1988; O’Brien et al. 2018) but similar to that of C. brasiliensis (Hsiou et al. 2015: fig. 2). Anteriorly, the basisphenoid forms the paired basipterygoid processes, but in C. cambrica (NHMUK PV R37014) these two processes have been clearly broken off (Fig. 9A3). We located what we infer to be the left basipterygoid process located ventral to the basipterygoid facet on the left pterygoid. It appears as a small but robust bone that is roughly tear-shaped. Like the basipterygoid processes, the cultriform process also appears to have been broken off (Fig. 9A3), though it might be preserved, but as it is a very delicate thin structure (Fraser 1988: fig. 17b; NHMUK PV R36832), it is not visible at the resolution of the CT scans used here.
Supraoccipital, opisthotics, and prootics: The supraoccipital in Clevosaurus cambrica is still fused to both left and right prootics, which are in turn both fused to their respective opisthotics (Fig. 9). As has been observed in the C. hudsoni specimens of Fraser (1988: 140), these elements are often fused. Posteriorly the supraoccipital forms the dorsal margin of the foramen magnum, but the bone is so thin here (as in C. hudsoni; NHMUK PV R36832), that if it is not damaged then it is not visible at this resolution. The dorsal surface is convex and broad with no sign of a dorsal crest, but the area is poorly resolved and may have been damaged. The paroccipital processes of the opisthotics are robust and short, with a concave ventral surface, and they project posterolaterally. Suture lines between these bones cannot be distinguished, and though there clearly has been some displacement and breakage, digital reconstruction is limited. The left pila antotica, formed from the prootic (Fraser 1988: 141), is preserved and appears similar to that of C. hudsoni (NHMUK PV R36832), though relatively dorsoventrally shorter, and it appears to have been broken off the main body of the prootic. These elements have not been well preserved in other species of Clevosaurus and cannot therefore be compared here.
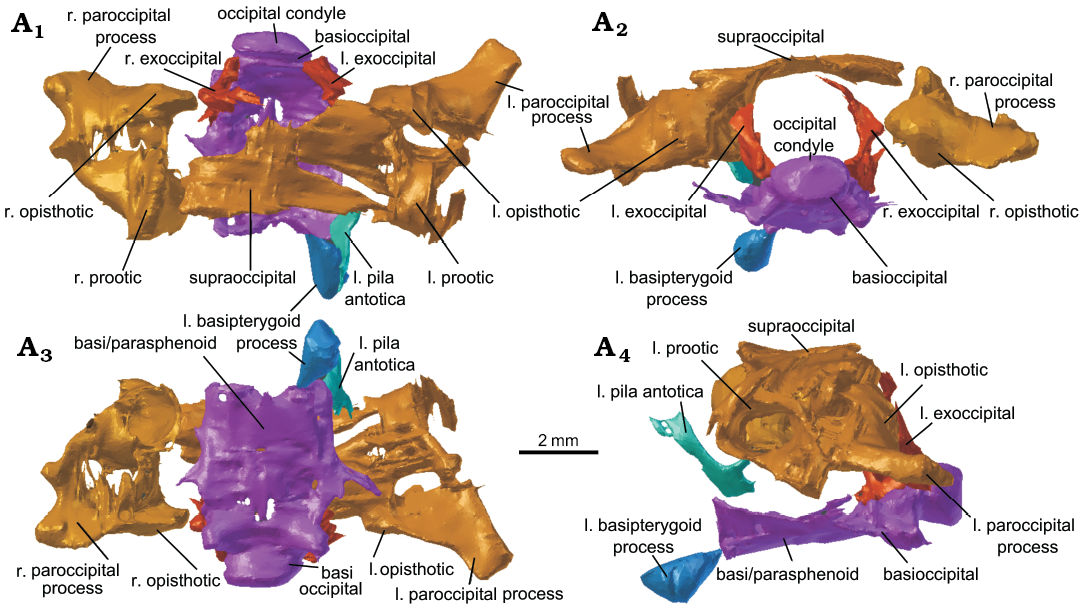
Fig. 9. The reassembled braincase of lepidosaur rynchocephalian Clevosaurus cambrica Keeble, Whiteside, and Benton, 2018 (NHMUK PV R37014) from Late Triassic, Pant-y-ffynnon, Wales, UK. In dorsal (A1), posterior (A2), ventral (A3), and lateral (A4) views. Abbreviations: l., left; r., right.
Dentary: Both dentaries have been partially described (Keeble et al. 2018: 104–105). In the amended scans, both dentaries were found to be complete, although the left dentary appears to have sustained some damage along its lateral and ventral margins, and we identified the anterior fragment which in the original description had been tentatively suggested to be part of the hyoid. The right dentary is intact, while the left dentary has a break in the anteriormost third of its length. The anterior third is still articulated with the right dentary in the symphyseal region, while the posterior two-thirds of the left dentary have been displaced laterally. The anterior fragment bears no teeth and forms a dorso-anteriorly concave structure that inclines anteriorly very much like that of Clevosaurus hudsoni (Fraser 1988: fig. 19). The symphyseal contact is ovoid, gracile and positioned at a shallow angle, which is like the condition in C. sectumsemper (Klein et al. 2018: fig. 4), but differs from the oval near-vertical symphyseal contact of C. brasiliensis (Hsiou et al. 2015) or the stout symphyseal contact of C. convallis (Säilä 2005). There are six acrodont teeth on the right dentary, with pronounced wear facets, which are further described by Keeble et al. (2018), and below them is a pronounced lateral ridge visible indicating the development of secondary bone (Fig. 10A1). The penultimate tooth is longest mesio-distally, but roughly the same height as the ultimate tooth. The Meckelian groove is present but not pronounced, expanding posteriorly below the tooth row. The region of dentary immediately behind the posteriormost tooth tapers into a narrow upwardly slanted neck, which then expands dorsally to form the coronoid process, where it resumes running parallel to the anterior portion of the dentary. Both left and right dentaries are still articulated with the surangular, angular, articular and prearticular, as well as both being in close association with their respective coronoids, so a complete articulated mandible was easy to reconstruct (Fig. 10A). The dentary of C. cambrica is slighter than those of C. convallis (Säilä 2005), C. brasiliensis (Bonaparte and Sues 2006) and C. bairdii (Sues et al. 1994). It is most similar in morphology to those of C. hudsoni and C. sectumsemper (Fraser 1988; Klein et al. 2015) which, although they have only four additional teeth, are more gracile and possess teeth with prominent anterolateral flanges. The C. cambrica dentary is more slender and has a lower coronoid process than the C. hudsoni specimen NHMUK PV R36832 (Fig. 10B).
Coronoid: Previously undocumented in any species of Clevosaurus from the British Isles, though observed in North American and Chinese clevosaurs (Sues et al. 1994; Wu 1994; Jones 2006), both left and right coronoids are present and complete, with only slight disarticulation. Both bear clear facets for the surangular on their posterior surfaces in the form of a corresponding convex structure (Fig. 10A2). The coronoid in life would have articulated with the antero-medial surface of the coronoid process. The shape is similar to that of C. hudsoni (NHMUK PV R36832) that we describe here, with both bearing a smoother and rounder lateral profile than in Sphenodon. Similar to C. bairdii (Sues et al. 1994: 334), the coronoid bone projects slightly above the coronoid process. The coronoid is roughly an inverted tear-drop in shape, with a rounded, broader and wider dorsal margin that corresponds with the medio-dorsal edge of the coronoid process of the dentary. Ventrally the coronoid tapers and flattens.
Articular, prearticular, surangular and angular: The posterior bones of the mandible are present on both sides of the skull of Clevosaurus cambrica. As mentioned by Keeble et al. (2018: 109), the angular cannot be separated from the prearticular, which in turn is fused to the articular complex, but it can be inferred as it extends beyond the anterior edge of coronoid eminence and reaches as far as the penultimate tooth. The articular and surangular are fused to form the articular complex, as in C. hudsoni (Fraser 1988) and C. convallis (Säilä 2005). Both prearticular and surangular are latero-medially compressed and are fused posteriorly to the angular, forming the ventral and dorsal edges respectively of the adductor fossa (Fig. 10A). The prearticular runs parallel to the length of the dentary and is anteriorly fused to the angular. The surangular runs at a shallow angle towards the apex of the coronoid process, forming its posterodorsal edge, and anteriorly articulating with the coronoid. The articular is robust, forming the articular condyle before rapidly tapering into an elongate medially recurved retroarticular process which remains at a constant width along its length (Fig. 10A). The mandibular foramen cannot be discerned. The mandible terminates posteriorly in an elongated retroarticular that is similar in relative length to that of C. hudsoni (NHMUK PV R36832; Fig. 10B) and C. brasiliensis (Bonaparte and Sues 2006; Hsiou et al. 2015).
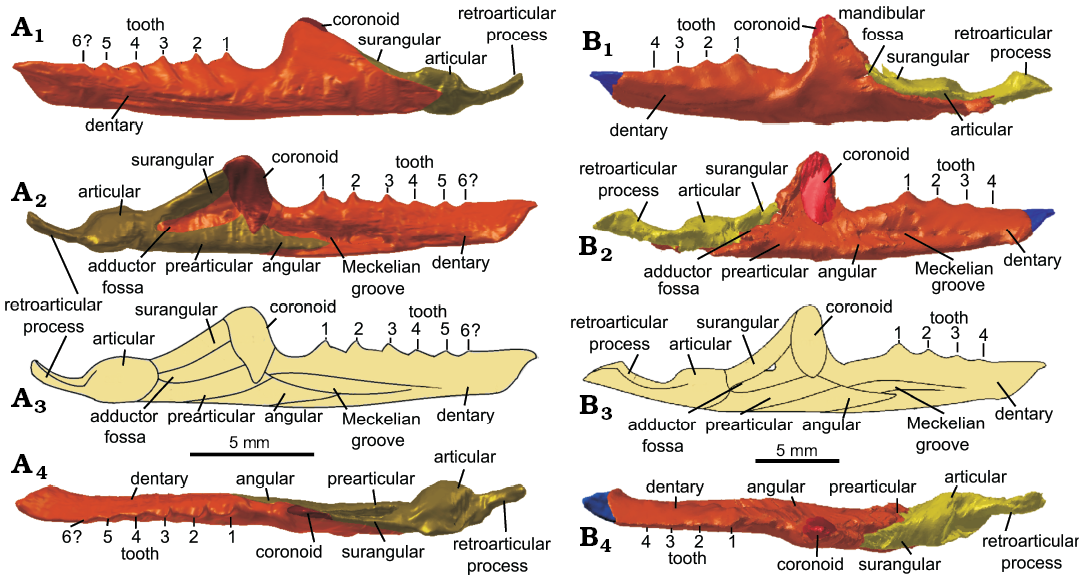
Fig. 10. The reconstructed left mandible of lepidosaur rynchocephalians. A. Clevosaurus cambrica Keeble, Whiteside, and Benton, 2018 (NHMUK PV R37014) from Late Triassic, Pant-y-ffynnon, Wales, UK. B. Clevosaurus hudsoni Swinton, 1939 (NHMUK PV R36832) from Rhaetian, Cromhall Quarry, England, UK. Note: the anterior tip of the dentary is highlighted in blue to indicate where bone is taken from C. cambrica. In lateral (A1, B1), medial (A2, B2), and dorsal (A4, B4) views, simplified reconstruction in medial view (A3, B3).
Teeth: Maxillary and dentary teeth in Clevosaurus cambrica were already well described by Keeble et al. (2018), and the main amendments we make is that there is an anterior small sixth tooth (probably not flanged) on the right maxilla and that the dentary teeth appear to be bounded ventrally by secondary bone or dentine with wear facets of individual maxillary teeth typical of Clevosaurus. We also add that the trailing flanges that recurve posteromedially on the teeth have large prominent medially placed escape structures on the additional teeth of the dentary. The additional dentary and maxillary teeth of C. cambrica bear prominent flanges, and “saddle-shaped” additional teeth on the dentary with prominent anteromedially placed escape structures; this morphology also resembles that in C. hudsoni as described by Fraser (1988) and in C. sectumsemper (Klein et al. 2015). This latter species has gaps with no overlap between the teeth on the dentary, a feature which C. cambrica appears to share (Keeble et al. 2018: fig. 5C; Fig. 6B), but the dentary of C. cambrica differs markedly from both in having 5–6 teeth rather than 4 (including C. minor; Fraser 1988). Furthermore, unlike C. hudsoni and C. minor, it is not the ultimate but the penultimate dentary (or the antepenultimate) tooth that is the largest in C. cambrica, similar to C. convallis (Säilä 2005). However, the dentition of C. cambrica differs from that of C. convallis in that hatchling teeth do not remain visible in adult specimens, and the tooth number is between 5–6, rather than 6–10 (maximum 8 on the dentary and 10 on the maxilla; Säilä 2005: 820). C. convallis also lacks the pronounced escape structures or anterolateral flanges on its dentary teeth which often overlap and lack any gap between them.
Atlas/axis complex: The atlas (in NHMUK PV R37013) has undergone some fragmentation, and the antero-posteriorly short neural arch has broken in half and has disarticulated from the atlas centrum (Fig. 11A) and shares a similar morphology to that of Clevosaurus sectumsemper (Klein et al. 2015: fig. 5A, B). The centrum of the atlas itself is small, rounded and appears to be fused to the front of the axis centrum, forming the odontoid complex. There are no visible intercentra, but many small fragments of bone in the region could be parts of atlantal intercentra. The neural spine of the axis may be partially incomplete, as the expected axial neural spine (seen in NHMUK PV R36832, O’Brien et al. 2018) is seemingly absent, damaged or too thin to be visible at the resolution of the CT scans, and instead we see a medially placed notably high pointed spike at the antero-dorsal surface of the neural arch that rises and then appears to drop sharply (Fig. 11A2). The zygosphene-zygantrum articulation is preserved between the axis and all successive cervical vertebra. No proatlas was identified, but it might be present, as the resolution of the scans is poor in this region.
Cervical vertebrae: Excluding the atlas and axis, there are three articulated cervical vertebrae in the scan (Fig. 11B), making five cervical vertebrae altogether (Fig. 11D) in NHMUK PV R37013. These have shorter neural spines than the axis. They appear typical of sphenodontians, with an hourglass-shaped centrum, which is both notochordal and amphicoelous. The neural arches appear to be complete and are all fused to their respective centra. Intercentra for these vertebrae have either been lost from this specimen or at not visible at the resolution of the CT scans. These cervical vertebrae appear typical of Clevosaurus (Fraser 1988), with shorter spines than in Sphenodon.
Posterior, cervical, and dorsal vertebrae: As noted by Keeble et al. (2018), we found two isolated but articulated vertebrae close to the left scapula (in NHMUK PV R37013), but we add a further four, making a total of six closely associated vertebrae running beneath the forelimb (Fig. 11E). Their close association with the shoulder elements, and the number of cervical vertebrae already counted (five), suggests these are dorsal, or at least some are dorsal, vertebrae, as in C. hudsoni which had eight cervical vertebrae (Fraser 1988; O’Brien et al. 2018). Not much detail can be discerned from the scans, and only the two caudalmost of the six vertebrae can be seen in sufficient detail to be compared with the cervical vertebrae. The dorsal vertebrae both bear amphicoelous notochordal centra, and as in C. hudsoni (O’Brien et al. 2018) they are wider than the cervical vertebrae (Fig. 11C). In close association with the six articulated vertebrae we found three fragments of ribs.
Scapulae and coracoids: Here we identified in addition to the left scapulacoracoid (re-identified here as the left scapula; Fig. 11F1), the right scapula (Fig. 11F2) and both coracoids (Fig. 11G), displaced from their original positions in NHMUK PV R37013. Both scapula and coracoids are found separate, suggesting that they were not fused (cf. the Clevosaurus hudsoni of Fraser 1988; O’Brien et al. 2018) as seen in C. sectumsemper, indicating that the animal may have been a sub-adult at the time of death (Klein et al. 2015). The left scapula is disarticulated but closely associated with the proximal head of the humerus, while the right scapula is found displaced anteriorly of the distal head of the humerus, and Keeble et al. (2018) suggested it might be part of the sternum. The scapula blade here is found to be much shorter than previously described, and more like that of C. hudsoni, with the start of the blade clearly marked by a sharp transverse flexure in the bone. The glenoid protuberance is preserved in close association with the distal head of the humerus. Not much further detail can be discerned from the scans. The two coracoids are found displaced either side of the vertebral column. One coracoid is positioned posteriorly to the distal head of the humerus and had been segmented by Keeble et al. (2018) but was left out of the description. The second coracoid, found on the opposite side of the vertebral column, was segmented here for the first time. The coracoids are both damaged, but their identities can be inferred by their close association with the other shoulder elements and their positions either side of the vertebral column. Further, the robust protuberance found on each is likely to have formed the ventral half of the glenoid fossa.
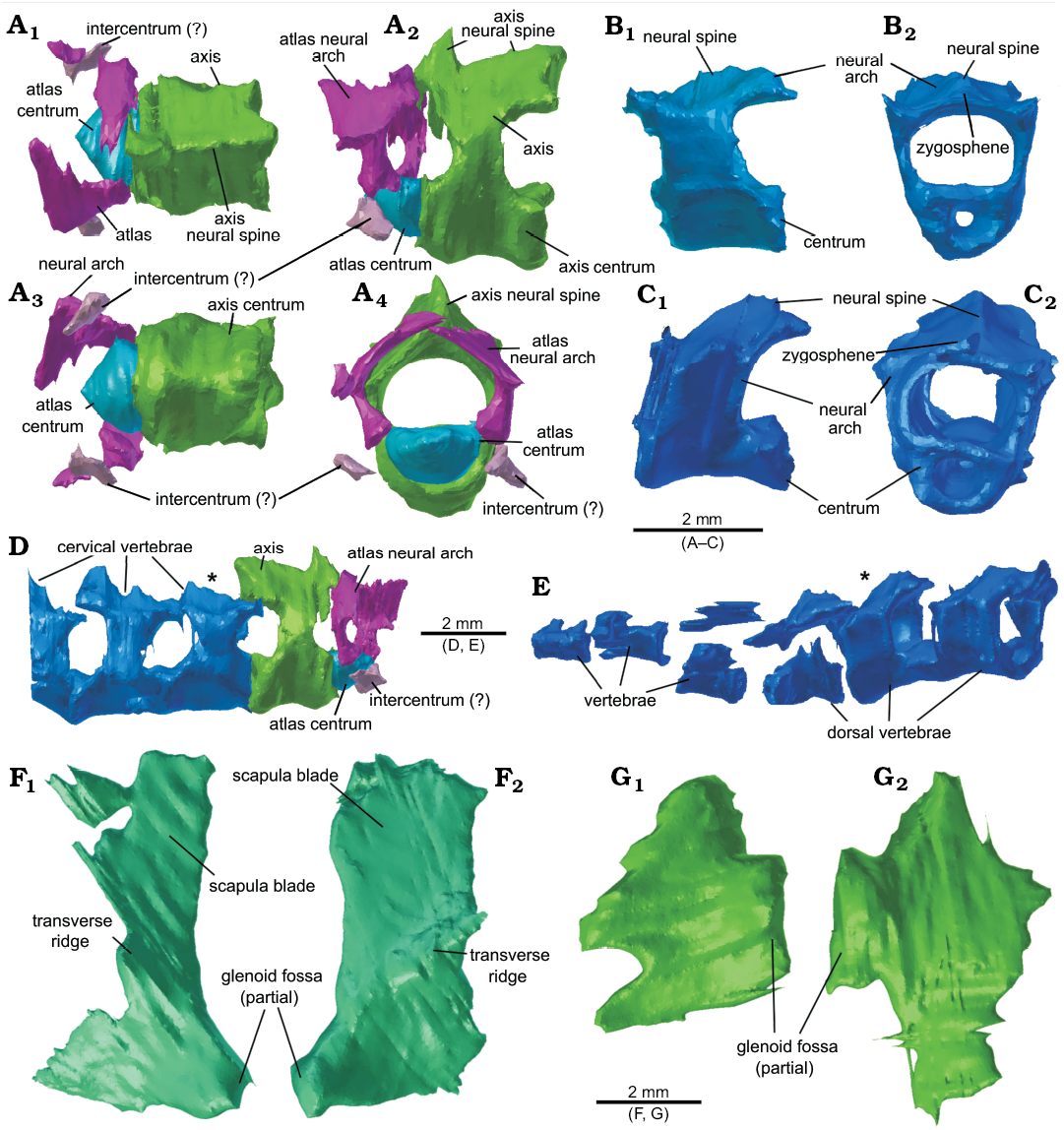
Fig. 11. The vertebral column and pectoral girdle elements of lepidosaur rynchocephalian Clevosaurus cambrica Keeble, Whiteside, and Benton, 2018 (NHMUK PV R37013 and NHMUK PV R37014) from Late Triassic, Pant-y-ffynnon, Wales, UK. A. The axial-atlas complex in dorsal (A1), lateral (A2), ventral (A3) and anterior (A4) views. B. The third cervical vertebra in right lateral (B1) and anterior (B2) views. C. A dorsal vertebra in right lateral (C1) and anterior (C2) views. D. The five cervical vertebrae present in the scans taken of NHMUK PV R37014, in lateral view, with asterisk indicating the cervical vertebra seen in B; note the posterior-most vertebra terminates prematurely due to the extent of the CT scans. E. The partial vertebral column of NHMUK PV R37013 in lateral view, with asterisk indicating the dorsal vertebra seen in C. F. The left (F1) and right (F2) scapula in lateral view. G. The left (G1) and right (G2) coracoids in lateral view.
Humerus: The left humerus (in NHMUK PV R37013) was described by Keeble et al. (2018), and we have refined the scan data to give a smoother surface and remove segmentation artefacts. Some alterations were made to the distal head of the humerus. The length of the humerus is nearly twice the mean length for C. sectumsemper but is around 80% the mean length for C. hudsoni (Fraser 1988, Klein et al. 2015).
Radius and ulna: These elements have already been described, but here they were segmented separately, allowing us to re-measure them individually, and we find them to be slightly shorter than they had been previously described, at 11 mm each (Table 1). The distal end of the radius appears to have a recurved head, and while in position for a carpal, no gaps could be observed to indicate it was a separate bone. The length ratio between the length of the ulna to the length of the humerus (in NHMUK PV R37013) falls closest to C. hudsoni measured by Fraser (1988: 157, tables 2, 3; Table 2), rather than C. hudsoni (NHMUK PV R36832), but this may not be a reliable comparison as they are the only two articulated specimens reported. However, the ulna/ humerus length ratios of the two articulated C. hudsoni specimens differ greatly and it is possible that these two specimens belong to two morphotypes. Skull length and forelimb proportions show some variations between the species of Clevosaurus (Table 3), as Fraser (1988) also noted.
Stratigraphic and geographic range.—Upper Triassic (Rhaetian) fissure infill within Lower Carboniferous Limestone, Pant-y-ffynnon Quarry, near Bonvilston, Vale of Glamorgan, South Wales, UK (NGR ST 046742).
Table 1. Measurements (in mm) of Clevosaurus hudsoni (NHMUK PV R36832), Clevosaurus cambrica (NHMUK PV R37014 and NHMUK PV R37013), and Sphenodon punctatus (from Maisano 2001).
| |
C. hudsoni |
C. cambrica |
S. punctatus |
|
Skull height |
14.0 |
9.5 |
32.7 |
|
Skull width |
19.4 |
14.0 |
47.7 |
|
Skull length |
24.0 |
19.0 |
55.0 |
|
Mandible length |
28.7 |
21.5 |
57.9 |
|
Maxilla length |
16.7 |
12.0 |
27.7 |
|
Coronoid height |
6.8 |
4.2 |
13.3 |
|
Radius length |
12.0 |
11.0 |
– |
|
Ulna length |
12.2 |
11.0 |
– |
|
Humerus length |
20.0 |
14.0 |
– |
Table 2. Measurements (in mm) of Clevosaurus cambrica (NHMUK PV R37013), Clevosaurus hudsoni (NHMUK PV R36832, UMZC T1271, * specimen measured by Fraser 1988: 157, table 2, 3); note that Fraser (1988) used isolated bones without a noted registration number to compile the mean given here.
| |
Humerus length |
Ulna |
Ratio
ulna/humerus |
|
|
C. cambrica |
NHMUK PV R37013 |
14.0 |
11.0 |
0.786 |
|
C. hudsoni |
NHMUK PV R36832 |
20.0 |
12.2 |
0.610 |
|
UMZC T1271 |
21.0 |
18.0 |
0.857 |
|
|
mean* |
20.4 |
18.5 |
0.907 |
|
Table 3. Measurements of skull lengths (in mm) and forelimb bone ratios of Clevosaurus cambrica (NHMUK PV R37013 and R37014) and Clevosaurus hudsoni (NHMUK PV R36832, UMZC T1271).
| |
Skull |
Radius |
Humerus |
Ulna |
Radius |
Ratio
|
|
|
C. cambrica |
NHMUK PV R37013 and NHMUK PV R37014 |
19 |
1.000 |
0.737 |
0.579 |
0.579 |
1.31 |
|
C. hudsoni |
NHMUK PV R36832 |
24 |
0.983 |
0.833 |
0.508 |
0.500 |
1.63 |
|
UMZC T1271 |
35 |
0.944 |
0.600 |
0.514 |
0.486 |
1.11 |
|
Results
Reconstruction of the skulls of Clevosaurus cambrica and C. hudsoni.—The two skulls were reassembled prior to reconstruction, in which broken or partial bones have been restored, allowing for a full reconstruction of the skulls of both specimens (Fig. 12). From among the other seven species of Clevosaurus, articulated skulls are available only for two from which reliable skull reconstructions have been made, namely C. bairdii (Sues et al. 1994: fig. 5) and C. brasiliensis (Bonaparte and Sues 2006). We regard the reconstructions for the three Chinese Clevosaurus by Wu (1994) as unreliable because they are based on fragmentary material, and two have been considered to be synonymous for the same reason (Jones 2006). Both C. bairdii and C. brasiliensis, with estimated adult skull lengths of 19 mm (Sues et al. 1994) and 17–23 mm (Bonaparte and Sues 2006) respectively, are more similar in size to the two skulls reconstructed here, than the estimated size of 35–40 mm given by Fraser (1988). Estimated skull lengths for the Chinese clevosaurs (Jones 2006) were 21–34 mm.
In dorsal view, the skull of C. hudsoni (NHMUK PV R36832) is broad and roughly triangular, with a robust rounded antorbital region (Fig. 12A1). The large orbits occupy approximately one third of the skull’s length, and half of the skull’s height (Fig. 12A4). The upper and lower temporal fenestrae also occupy approximately one-third of the skull’s length. The skull shape is like Fraser’s reconstruction of C. hudsoni (1988: fig. 3), with a similar skull height, but relatively broader (Table 4). It is more like that of C. bairdii or C. cambrica (see below) but much narrower than C. brasiliensis, and the antorbital region in particular is more robust and with more prominent prefrontals than in C. brasiliensis, C. bairdii, and C. cambrica. The orbits appear more squared and angular than the smooth and rounded orbits illustrated by Fraser (1988: fig. 3). Other interesting features that differ include a higher coronoid process (a feature shared with C. bairdii) and elongated retroarticular which has an enlarged recurved tip (a feature shared with C. brasiliensis and C. cambrica). The anteriormost portion of the skull had broken off and is not present in the scans, but there is enough of the maxilla and dentary margin in this region to say that not much is missing here, especially from the dentary whose anteriormost portion curves medially. Other species of Clevosaurus similarly have a very short antorbital region that is approximately one quarter of the length of the skull, or less in C. brasiliensis. The skull of both C. hudsoni (NHMUK PV R36832) and C. cambrica are noticeably less robust than that of C. bairdii, which has very deep maxillae and dentaries. C. hudsoni (NHMUK PV R36832) has a wider intertemporal than interorbital gap, as with the other clevosaurs examined here, but differing from the reconstruction of C. hudsoni by Fraser (1988).
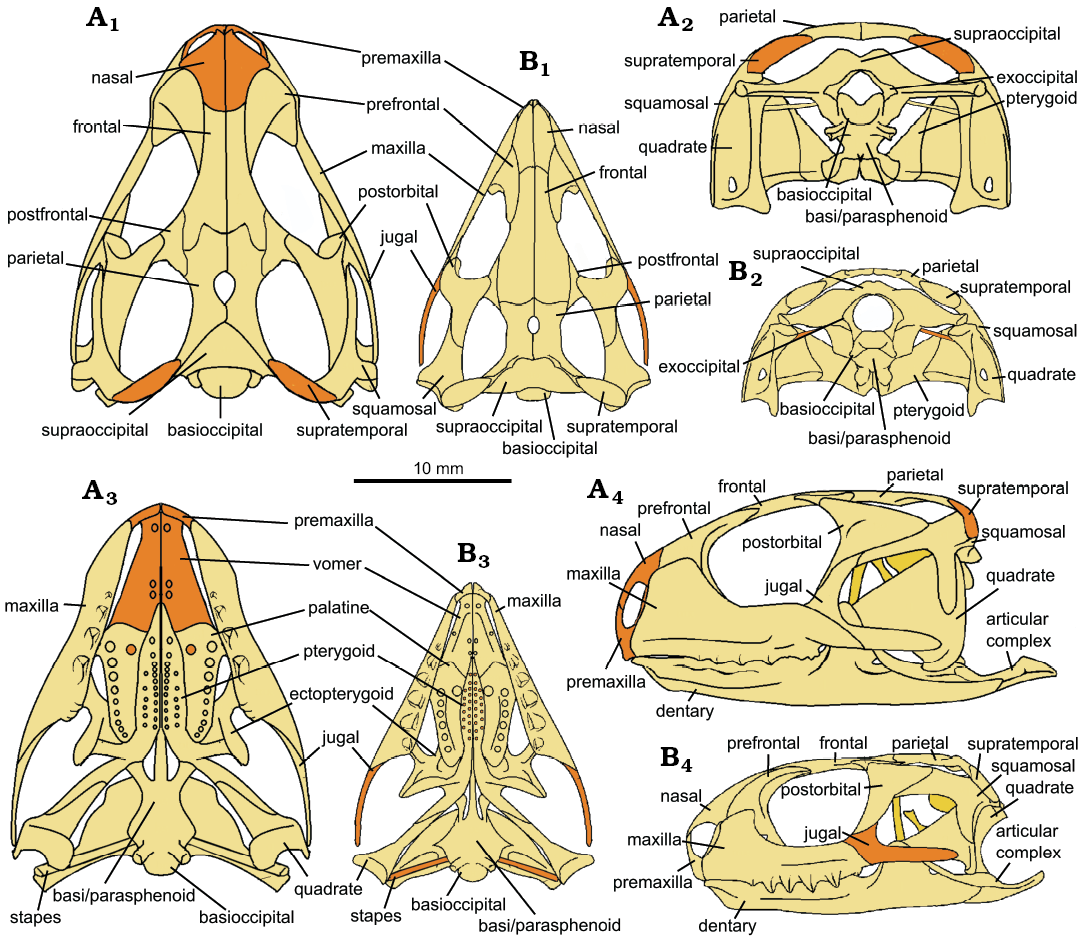
Fig. 12. Reconstruction of the skulls of lepidosaur rynchocephalians. A. Clevosaurus hudsoni Swinton, 1939 (based on NHMUK PV R36832). B. Clevosaurus cambrica Keeble, Whiteside, and Benton, 2018 (based on NHMUK PV R37014). In dorsal (A1, B1), posterior (A2, B2), ventral (A3, B3), lateral (A4, B4) views. Bones types that are absent in their respective datasets are highlighted in orange. Note that the pterygoids of C. cambrica bear two rows of multiple teeth, but the exact number of teeth at this time is unknown.
In dorsal view, the skull of C. cambrica is triangular (Fig. 12B1), with a pointed antorbital region. As with C. hudsoni (NHMUK PV R36832), orbital and temporal fenestrae each form about one-third of the skull’s length. Skull width is around three quarters of skull length, while skull height is slightly over half of skull length (Table 4). Skull width is like that of NHMUK PV R36832 but relatively shallower (Table 4). The antorbital region of C. cambrica appears to be much more delicate and narrower than NHMUK PV R36832 and the other clevosaurs compared here.
Table 4. Skull dimensional ratios for Clevosaurus and Sphenodon, with specimens shown in sequence of values for each. Clevosaurus hudsoni, NHMUK PV R36832 (Fig. 12A); C. hudsoni* (from Fraser 1988: fig. 3); C. cambrica, NHMUK PV R37014 (Fig. 12B); C. bairdii (from Sues et al. 1994: fig. 5); C. brasiliensis (from Bonaparte and Sues 2006: fig. 2); Sphenodon punctatus (from Maisano 2001). Skull width is the highest value measured across the jugals, skull height is the highest point of the skull around the region of the coronoid, and skull length is measured from the premaxillae to the squamosals.
|
Species |
Height / Width |
Species |
Height / Length |
Species |
Width / Length |
|
C. hudsoni* |
0.82 |
C. hudsoni |
0.60 |
C. brasiliensis |
0.90 |
|
C. hudsoni |
0.77 |
C. hudsoni* |
0.57 |
S. punctatus |
0.87 |
|
C. bairdii |
0.75 |
C. bairdii |
0.57 |
C. hudsoni |
0.77 |
|
S. punctatus |
0.69 |
S. punctatus |
0.55 |
C. bairdii |
0.76 |
|
C. cambrica |
0.68 |
C. brasiliensis |
0.52 |
C. cambrica |
0.75 |
|
C. brasiliensis |
0.58 |
C. cambrica |
0.51 |
C. hudsoni* |
0.69 |
The braincases of C hudsoni and C. cambrica were digitally reassembled and reconstructed (Figs. 13A, B, 14A, B), but the low resolution of the scans of the C. cambrica specimen mean its braincase could not be as confidently reconstructed. The braincase of Sphenodon (Maisano 2001) was used for comparison (Figs. 13C, 14C) as braincases have not been reported or reconstructed for any other species of Clevosaurus. The most striking difference is in the paroccipital processes, which are short and robust in C. cambrica, long and slender in C. hudsoni, and intermediate in Sphenodon. Proportionally, the supraoccipitals, and the fused parabasisphenoid, are much broader in C. cambrica than C. hudsoni or Sphenodon, indicating a larger space for the brain in life. In other respects, the braincases of both clevosaurs appear relatively similar, differing from Sphenodon in possessing flatter supraoccipitals and longer, more slender, basipterygoid processes and pila antotica. NHMUK PV R36832 differs from Fraser’s (1988) C. hudsoni in possessing a pila antotica that is notably larger and taller relative to the rest of the braincase.
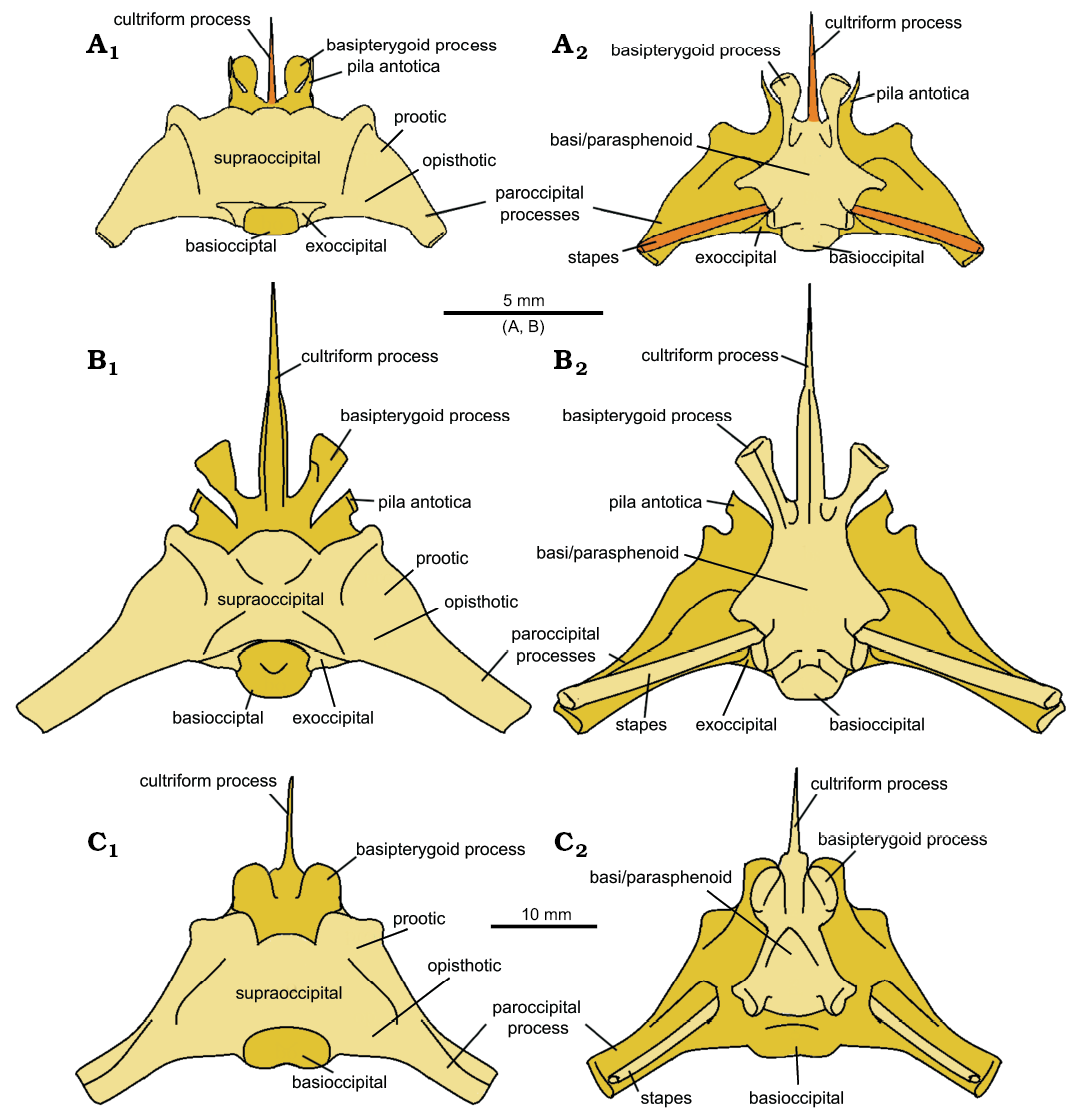
Fig. 13. The reconstructed braincases of lepidosaur rynchocephalians. A. Clevosaurus cambrica Keeble, Whiteside, and Benton, 2018. B. Clevosaurus hudsoni Swinton, 1939. C. Sphenodon punctatus (from Maisano 2001). In dorsal (A1–C1) and ventral (A2–C2) views. Bones types that are absent in their respective datasets are highlighted in orange.
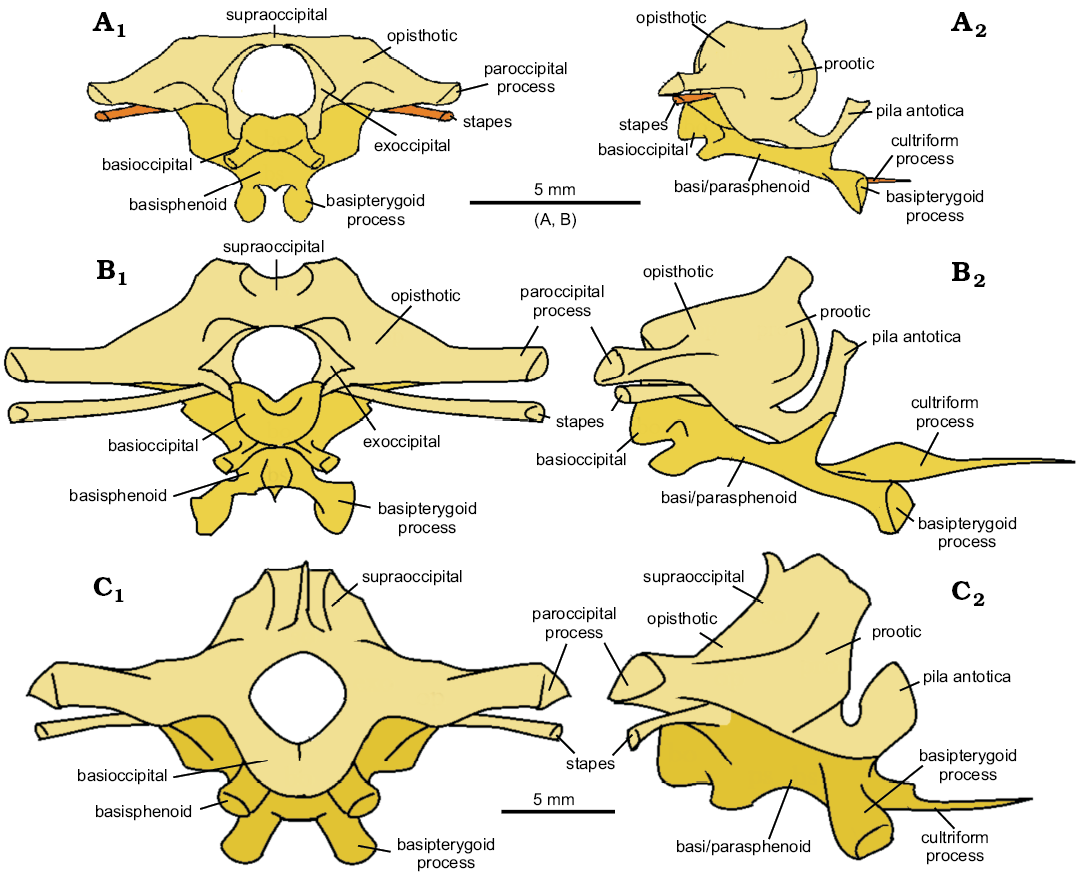
Fig. 14. The reconstructed braincases of lepidosaur rynchocephalians. A. Clevosaurus cambrica Keeble, Whiteside, and Benton, 2018. B. Clevosaurus hudsoni Swinton, 1939. C. Sphenodon punctatus (from Maisano 2001). In posterior (A1–C1) and right lateral (A2–C2) views. Bones types that are absent in their respective datasets are highlighted in orange.
Morphology of the ultimate dentary teeth in Clevosaurus cambrica, C. hudsoni, and Sphenodon.—The ultimate and largest tooth of Clevosaurus hudsoni (NHMUK PV R36832) has a height-to-length ratio of 0.32, similar to that of C. cambrica (NHMUK PV R37014) at 0.45, though this changes to 0.35 for the penultimate tooth, which is the largest tooth in this species. In Sphenodon, the ultimate tooth height-to-length ratio is 0.76, nearly twice the value in the clevosaurs considered here, and reflecting their more conical tooth shape.
Overall, dentary tooth height in C. hudsoni decreases anteriorly uniformly, but in C. cambrica the most posterior three teeth are of approximately equal height, and the trend of decreasing height applies only to those teeth anterior to that. Tooth length decreases anteriorly almost linearly in C. hudsoni, but in C. cambrica increases in the penultimate tooth before decreasing nearly linearly anteriorly. In both species, tooth width decreases anteriorly nearly linearly.
Tooth cross-sectional areas from tip to gum line increase most rapidly in C. cambrica. In C. hudsoni the expansion is gradual and linear, and more closely resembles the pattern in Sphenodon.
The teeth of C. cambrica have a different morphology from the blade-shaped teeth of C. hudsoni and the conical teeth of Sphenodon, showing a 90° twist from the tip, which is perpendicular to the jaw line, to the base, which is parallel to the jaw line. The dentary teeth of C. cambrica have large and prominent antero-lingually placed escape structures, while those of C. hudsoni (NHMUK PV R36832) are small or absent.
These variations in tooth shape are reflected in their labiolingual to mesiodistal width ratio. The ratio for the ultimate blade-shaped dentary tooth of C. hudsoni never exceeds a value of 0.7, meaning that labiolingual width is always less than mesiodistal width, showing a simple blade-like shape. The teeth of Sphenodon are more conical, which is reflected by having ratios closer to 1.0. For C. cambrica ratios exceed a value of 1.0 for the top half of the tooth, before dropping below 1.0, reflecting the spiral structure of their teeth caused by the presence of a large antero-lingually placed escape structure (see SOM 6).
While it is beyond the scope of this study, from observation, the additional dentary teeth of C. sectumsemper (Klein et al. 2015) are very similar in morphology to those of C. cambrica, with large antero-medially placed escape structures and prominent antero-lateral flanges, which we predict would produce a very similar ratio profile to C. cambrica. C. convallis also has elongated antero-lateral flanges, but these are less pronounced than those of C. cambrica, C. minor or Fraser (1988)’s reconstruction of C. hudsoni and would likely fall closer to the values recorded for NHMUK PV R36832 on the graph (SOM 6). Comparisons with other clevosaurs are difficult: the Chinese clevosaurs are too fragmentary, in C. bairdii the dentary teeth are obscured by matrix, and in C. brasiliensis the crowns of the additional dentary teeth wear into a near-single surface in mature specimens (Bonaparte and Sues 2006; Hsiou et al. 2015).
Discussion
Tooth morphology.—The diet of Clevosaurus has been interpreted as herbivory (Fraser and Walkden 1983; Fraser 1988), omnivory (Jones 2009), or faunivory (Jones 2008; Meloro and Jones 2012; Whiteside et al. 2017), but most generally it is thought to have been adapted to consuming arthropod prey. The clearest difference between C. hudsoni and C. cambrica is in the number of teeth, but tooth morphology also differs. The cusp of the ultimate tooth of C. cambrica is oriented nearly perpendicular to the long axis of the mandible, unlike that of C. hudsoni, which is orientated parallel. The direction of this blade in C. cambrica rotates down its height to be nearly parallel to the long axis of the dentary at its base. Further, the tooth broadens more rapidly from tip to gum line than in C. hudsoni, and this would have resulted in greater damage after initial tooth penetration of prey, at the cost a more reduced relative pressure, as in the molariform teeth of crocodilians (Erickson et al. 2012). Struggling insect prey could have been impaled on these teeth and then further broken apart by the “cork-screw”-like morphology of the teeth, which could have effectively pulled the chitin apart in multiple directions during biting. The lenticular teeth of C. hudsoni would have likely been more effective in slicing.
In the arthropod exoskeleton, chitin is often laid down in parallel layers, and long blade-like teeth like those of C. hudsoni are ideal for cuticle shredding, as observed in insectivorous mammals (Lucas and Peters 2007). The positioning of the tooth row differs between the two clevosaurs. The type C. hudsoni and our specimen NHMUK PV R36832 has all four tooth bases parallel to the long axis of the dentary, forming a jagged continuous shearing structure, whereas those of C. cambrica are positioned slightly “en echelon” so that the teeth slightly overlap mesio-distally. Another notable difference is the presence of large distinct escape structures on the antero-lingual edge of the mandibular teeth of C. cambrica, which appear to be absent or unpronounced in our specimen of C. hudsoni. These large escape structures would have helped C. cambrica clear food fragments away from its cutting surface and move them towards the palatine teeth, and thereby improve cutting and food reduction efficiency (Evans and Sanson 2003), possibly indicating that C. cambrica processed food more rapidly than C. hudsoni.
Cranial differences (Fig. 15): Aside from tooth morphology, the cranial morphology of Clevosaurus cambrica differs strikingly from C. hudsoni (NHMUK PV R36832) in the relative size of the braincase. The increased volume of the braincase of C. cambrica (see SOM 7 and 8) would have reduced the space for adductor musculature, the volume of which may have been further decreased by its relatively shallower skull. Further, the more gracile skull of C. cambrica (Fig. 15B) probably gave it a weaker bite than C. hudsoni (Fig. 15C). Therefore, C. cambrica might have preyed on smaller arthropod prey than C. hudsoni, which concentrated on tougher prey, such as beetles and small vertebrates.
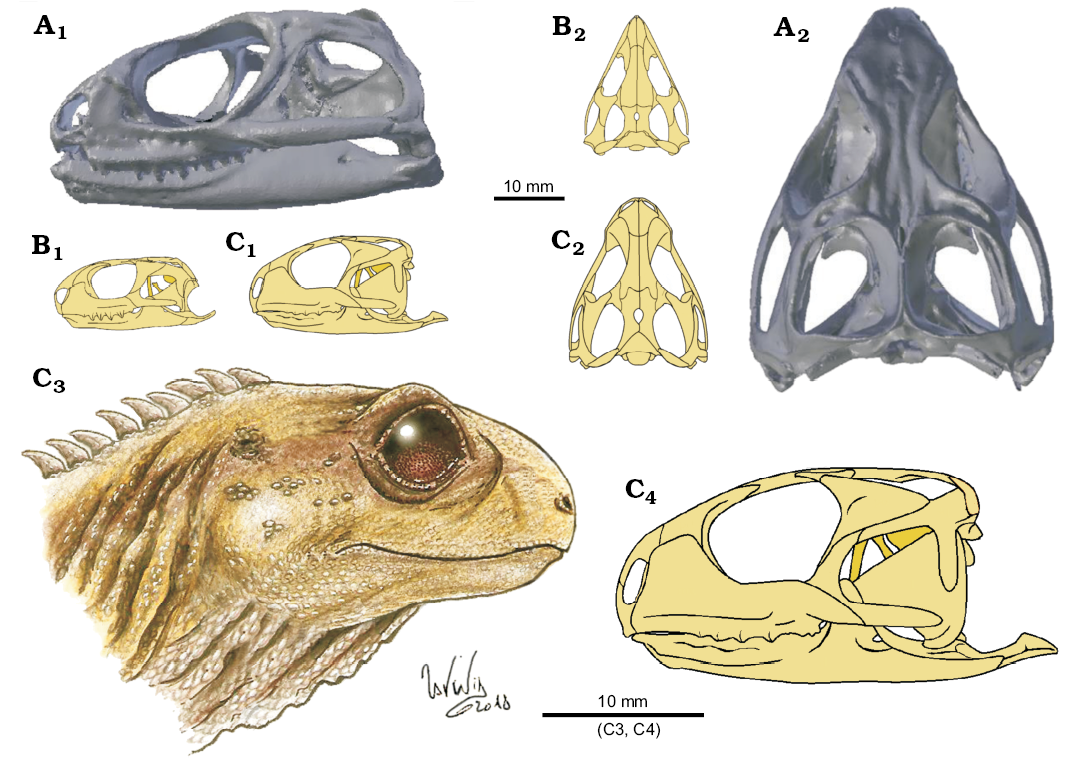
Fig. 15. A size scale comparison of the skulls of lepidosaur rynchocephalians. A. Sphenodon punctatus (from Maisano 2001?). B. Clevosaurus cambrica Keeble, Whiteside, and Benton, 2018. C. Clevosaurus hudsoni Swinton, 1939; C3, based on NHMUK PV R36832 (adapted from illustrations by Lavinia Gandolfi). In lateral (A1–C1, C4) and dorsal (A2–C2) views.
Life stage of Clevosaurus cambrica.—Clevosaurus cambrica (NHMUK PV R37014) is interpreted to be a sub-adult because its scapulae and coracoids are both found separately, rather than being fused to form a scapulacoracoid as in C. hudsoni (Fraser 1988: 149; O’Brien et al. 2018: 184). In juvenile Sphenodon, for example, these shoulder elements only fuse on maturity (Howes and Swinnerton 1901: 58, pl. 6), and this might also have been the case in C. sectumsemper (Klein et al. 2015). The maxilla of NHMUK PV R37014 is 12 mm long, whereas the holotype presented in Keeble et al. (2018) is 15 mm long. In addition, the specimen has a relatively enlarged braincase compared to C. hudsoni (Fig. 13), suggesting further that this specimen might be a sub-adult. However, the dentaries and maxillae already bear pronounced wear facets, and no trace of definite hatchling teeth. It should be noted too that the C. hudsoni specimen NHMUK PV R36832 has also been interpreted as a young adult from its small size (O’Brien et al. 2018: 191), but, unlike C. cambrica NHMUK PV R37014, its scapulae and coracoids are fused. The account of C. hudsoni given by Robinson (1973: fig. 4) suggests a smaller skull of length of 26 mm than Fraser’s (1988: 155) estimate of 40 mm. Overall, our reconstruction more closely resembles that of Robinson (1973) than Fraser’s (1988), with a “prominent lateral bulge” in the region of the prefrontals, and in the skull being broader with a steeper antorbital region. Fraser (1988) suggests this may reflect phenotypic variation, and we suggest so again that there may be at least two morphotypes within the species, and that the smaller size of NHMUK PV R36832 may not indicate that this individual was sub-adult. Rather it may well be a young adult, as suggested by O’Brien et al. (2018).
The significance of the coronoid bone in Clevosaurus.—No species of Clevosaurus from the British Isles had yet been found to have a separate coronoid bone, including C. hudsoni, where it was instead thought to have fused to the dentary to form the coronoid process (O’Brien et al. 2018). We have discovered separate coronoid bones in the scan data of both C. hudsoni and C. cambrica. We suggest that this bone has not been observed previously in C. hudsoni as it may be fused in fully mature adults, and this specimen may be a young adult (O’Brien et al. 2018: 191) or it may simply be that the coronoid has been missed in all previous descriptions.
Limitations of the reconstructions.—In the reconstruction of the skull of C. hudsoni (NHMUK PV R36832), skull bilateral symmetry was assumed. Approximately half of the skull is missing, which means that the skull had to be reconstructed using a mosaic of mirrored elements from both sides. There is noticeable asymmetry between the paired frontals and parietals in NHMUK PV R36832, which caused problems of spatial placement of the additional cranial bones. To compensate for this, the frontals and parietals were divided down the middle and mirrored from the left side of the skull to the less complete right side. Missing or damaged bones, such as the anterior end of the left dentary and maxilla, were replaced or repaired with data from C. cambrica and from illustrations in Fraser (1988) and Robinson (1973). An educated guess had to be made in defining the anterior extent of the rostral-most portion of the skull, as the premaxillae were missing, and the anterior edge of the maxilla was broken off. In piecing the skull back together, some facets were found to be well defined, such as the prefrontal-frontal facet. However, the extents of some facets were not well defined, such as the connection between the pterygoid flange and quadrate flange, and the articular complex and dentary, and previous reconstructions (Robinson 1973; Fraser 1988) were used as a guide.
The skull of C. cambrica was reconstructed using the same methodology, including an assumption of bilateral symmetry. Lower resolution scans meant that few facets were clearly defined. An additional problem in C. cambrica, but not in C. hudsoni, was the fragmentation of bones such as the prefrontal and postorbital.
The C. hudsoni skull (NHMUK PV R36832) shows some morphological features that differ from Fraser’s (1988) reconstruction, including: a coronoid region that, even with the correction of the coronoid bone, remains quite high and prominent; an unusually long retroarticular process (though not unusual when compared to C. cambrica here); and the nine palatine teeth, compared to five or six in Fraser (1988). We cannot certainly conclude whether these are errors, poor preservation, or evidence of intraspecific variation in C. hudsoni. There is scope for future work to investigate morphological variation within the species, to evaluate as much material referred to this species as possible, in similar manner to the study of dentaries of C. brasiliensis by Martínez and Soares (2015). The examination of the C. hudsoni dentaries at the NHMUK revealed two possible morphotypes, and such morphological variation within the species might explain some of the discrepancies. The two morphotypes might reflect geographic or temporal variation, sexual dimorphism, or species differences. Importantly, our skull is of the same morphotype as the type specimens described by Swinton (1939).
Conclusions
The skull, cervical vertebrae and forelimb of C. cambrica were investigated using a combination of CT scans, digital segmentation and reassembly, and the skull of C. hudsoni was also reconstructed. This approach allowed, for the first time, for 3-D digital reassembly of skulls and braincases of two species of Clevosaurus, as well as the first record of coronoid bones in clevosaurs from the British Isles. There were some limitations in the resolution of the C. cambrica scans, which had been previously thought not to contain much more than some mandibular and maxillary bones; in fact, we discovered the whole skull of C. cambrica. We also provide new information on the anatomy of C. cambrica such as the epipterygoid and demonstrate interspecific cranial variability among Clevosaurus species, where key differences in dental morphology suggest different dietary habits between the two species.
Acknowledgements
We are grateful to Aileen O’Brien (University of Southampton, UK) and Emily Keeble (University of Bristol, UK) for access to the CT scans and segmented surfaces used in their respective papers on Clevosaurus hudsoni and C. cambrica. We particularly thank Sandra Chapman (NHMUK) who has continually supported and encouraged this work and also Mike Day (NHMUK) who registered some of the specimens at short notice. We thank Mark Mavrogordato and Kathryn Rankin (both µ-VIS X-Ray Imaging Centre, Faculty of Engineering and the Environment, University of Southampton, UK) for carrying out the CT scans of C. hudsoni, and Tom Davies and Benjamin Moon (both University of Bristol, UK) for producing the scans of C. cambrica on the Palaeontology Research Group CT scanner in the Life Sciences Building. We also thank Annie Hsiou (Universidade de São Paulo, Ribeirão Preto, Brazil) for her comments on the manuscript which have significantly enhanced the quality of this paper.
References
Bonaparte, J.F. and Sues, H.-D. 2006. A new species of Clevosaurus (Lepidosauria: Rhynchocephalia) from the Upper Triassic of Rio Grande do Sul, Brazil. Palaeontology 49: 917–923. Crossref
Erickson, G.M., Gignag, P.M., Steppan, S.J., Lappin, A.K., Vliet, K.A., Brueggen, J.D., Inouye, B.D, Kledzik, D., and Webb, G.J.W. 2012. Insights into the ecology and evolutionary succes of crocodilians revealed through bite-force and toot-pressure experimentation. PLoS ONE 7 (3): e31781 Crossref
Evans, A.R. and Sanson, G.D. 2003. The tooth of perfection: functional and spatial constraints on mammalian tooth shape. Biological Journal of the Linnean Society 78: 173–191. Crossref
Fraser, N.C. 1988. The osteology and relationships of Clevosaurus (Reptilia: Sphenodontida). Philosophical Transactions of the Royal Society of London B 321: 125–178. Crossref
Fraser, N.C. 1993. A new sphenodontian from the early Mesozoic of England and North America: Implications for correlating early Mesozoic continental deposits. New Mexico Museum of Natural History and Science Bulletin 3: 135–139.
Fraser, N.C. and Walkden, G. M. 1983. The ecology of a late Triassic reptile assemblage from the Gloucestershire, England. Palaeogeography, Palaeoclimatology, Palaeoecology 42: 341–365. Crossref
Gignac, P.M. and Erickson, G.M. 2015. Ontogenetic changes in dental form and tooth pressures facilitate developmental niche shifts in American alligators. Journal of Zoology 295: 132–142. Crossref
Gill, P.G., Purnell, M.A., Crumpton, N., Brown, K.R., Gostling, N.J., Stampanoni, S., and Rayfield, E.J. 2014. Dietary specializations and diversity in feeding ecology of the earliest stem mammals. Nature 512: 303–305. Crossref
Herrera-Flores, J., Stubbs, T.L., and Benton, M.J. 2017. Macroevolutionary patterns in Rhynchocephalia: is the tuatara (Sphenodon punctatus) a living fossil? Palaeontology 60: 319–328. Crossref
Herrera-Flores, J., Stubbs, T.L., Elsler, J.A., and Benton, M. J. 2018. Taxonomic reassessment of Clevosaurus latidens Fraser, 1993 (Lepidosauria, Rhynchocephalia) and rhynchocephalian phylogeny based on parsimony and Bayesian inference. Journal of Paleontology 94: 734–742. Crossref
Howes, G.B. and Swinnerton, H.H. 1901. On the development of the skeleton of the Tuatara, Sphenodon punctatus; with remarks on the egg, on the hatchling, and on the hatched young. Transactions of the Zoological Society of London 16: 1–86. Crossref
Hsiou, A.S., Gallo De Franca, M.A., and Ferigolo, J. 2015. New data on the Clevosaurus (Sphenodontia: Clevosauridae) from the Upper Triassic of Southern Brazil. PLoS ONE 10 (9): e0137523. Crossref
Jones, M.E.H. 2006. The Early Jurassic clevosaurs from China. (Diapsida: Lepidosauria). New Mexico Museum of Natural History, Science Bulletin 37: 548–562.
Jones, M.E.H. 2008. Skull shape and feeding strategy in Sphenodon and other Rhynchocephalia (Diapsida: Lepidosauria). Journal of Morphology 269: 945–966. Crossref
Jones, M.E.H. 2009. Dentary tooth shape in Sphenodon and its fossil relatives (Diapsida: Lepidosauria: Rhynchocephalia). Frontiers of Oral Biology 13: 9–15. Crossref
Jones, M.E.H., Anderson, C.L., Hipsley, C.A., Müller, J., Evans, S.E., and Schoch, R.R. 2013. Integration of molecules and new fossils supports a Triassic origin for Lepidosauria (lizards, snakes, and tuatara). BMC Evolutionary Biology 13: 208. Crossref
Keeble, E., Whiteside, D.I., and Benton, M.J. 2018. The terrestrial fauna of the Late Triassic Pant-y-ffynnon Quarry fissures, South Wales, UK and a new species of Clevosaurus (Lepidosauria: Rhynchocephalia). Proceedings of the Geologists’ Association 129: 99–119.
Klein, C.G., Whiteside, D.I., Selles de Lucas, V., Viegas P.A., and Benton, M.J. 2015. A distinctive Late Triassic microvertebrate fissure fauna and a new species of Clevosaurus (Lepidosauria: Rhynchocephalia) from Woodleaze Quarry, Gloucestershire, UK. Proceedings of the Geologists’ Association 126: 402–416. Crossref
Lautenschlager, S. 2016. Reconstructing the past: methods and techniques for the digital restoration of fossils. Royal Society Open Sciences 3: 160342. Crossref
Lucas, P.W. and Peters, C.R. 2007. Function of postcanine tooth crown shape in mammals. In: M.F. Teaford, M. Smith, and M.W.J. Ferguson (eds.), Development, Function, and Evolution of Teeth, 282-289. Cambridge University Press, New York.
Maisano, J. 2001. “Sphenodon punctatus”, Digital Morphology. Accessed April 22, 2018 at http://digimorph.org/specimens/Sphenodon_punctatus/adult/.
Martínez R.N., Apaldetti, C., Colombi, C.E., Praderi, O.A., Fernandez, E., Malnis, P.S., Correa, G.A., Abelin, D., and Alcober, O. 2013. A new sphenodontian (Lepidosauria: Rhynchocephalia) from the Late Triassic of Argentina and the early origin of the herbivore opisthodontians. Proceedings of the Royal Society B 280: 20132057. Crossref
Meloro, C. and Jones, M.E.H. 2012. Tooth and cranial disparity in the fossil relatives of Sphenodon (Rhynchocephalia) dispute the persistent “living fossil” label. Journal of Evolutionary Biology 25: 2194–2209. Crossref
O’Brien, A., Whiteside, D.I., and Marshall, J.E.A. 2018. Anatomical study of two previously undescribed specimens of Clevosaurus hudsoni (Lepidosauria: Rhynchocephalia) from Cromhall Quarry, UK, aided by computed tomography, yields additional information on the skeleton and hitherto undescribed bones. Zoological Journal of the Linnean Society 183: 163–195. Crossref
Robinson, P.L. 1973. A problematic reptile from the British Upper Trias. Journal of the Geological Society of London 29: 457–479. Crossref
Säilä, L.K. 2005. A new species of the sphenodontian reptile Clevosaurus from the Lower Jurassic of South Wales. Palaeontology 48: 817–832. Crossref
Sues, H.-D., Shubin, N.H., and Olsen, P.E. 1994. A new sphenodontian (Lepidosauria: Rhynchocephalia) from the McCoy Brook Formation (Lower Jurassic) of Nova Scotia, Canada. Journal of Vertebrate Paleontology 14: 327–340. Crossref
Swinton, W.E. 1939. A new Triassic rhynchocephalian from Gloucestershire. Journal of Natural History 4: 591–594.
Whiteside, D.I. and Duffin, C.J. 2017. Late Triassic terrestrial microvertebrates from Charles Moore’s “Microlestes” quarry, Holwell, Somerset, UK. Zoological Journal of the Linnean Society 179: 677–705. Crossref
Whiteside, D.I., Duffin, C.J., and Furrer, H. 2017. The Late Triassic lepidosaur fauna from Hallau, North-Eastern and a new “basal” rhynchocephalian Deltadectes elvetica gen et. sp. nov. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 285: 53–74. Crossref
Whiteside, D.I., Duffin, C.J., Gill, P.G., Marshall, J.E.A., and Benton, M.J. 2016. The Late Triassic and Early Jurassic fissure faunas from Bristol and South Wales: Stratigraphy and setting. Palaeontologia Polonica 67: 257–287.
Wu, X.C. 1994. Late Triassic–Early Jurassic sphenodontians from China and the phylogeny of the Sphenodontia. In: N.C. Fraser and H.D. Sues (eds.), In the Shadow of the Dinosaurs: Early Mesozoic Tetrapods. 38–69. Cambridge University Press, Cambridge.
Acta Palaeontol. Pol. 64 (1): 41–64, 2019
https://doi.org/10.4202/app.00569.2018