

Two new stegosaur specimens from the Upper Jurassic Morrison Formation of Montana, USA
D. CARY WOODRUFF, DAVID TREXLER, and SUSANNAH C.R. MAIDMENT
Woodruff, D.C., Trexler, D., and Maidment, S.C.R. 2019. Two new stegosaur specimens from the Upper Jurassic Morrison Formation of Montana, USA. Acta Palaeontologica Polonica 64 (3): 461–480.
Two partial skeletons from Montana represent the northernmost occurrences of Stegosauria within North America. One of these specimens represents the northernmost dinosaur fossil ever recovered from the Morrison Formation. Consisting of fragmentary cranial and postcranial remains, these specimens are contributing to our knowledge of the record and distribution of dinosaurs within the Morrison Formation from Montana. While the stegosaurs of the Morrison Formation consist of Alcovasaurus, Hesperosaurus, and Stegosaurus, the only positively identified stegosaur from Montana thus far is Hesperosaurus. Unfortunately, neither of these new specimens exhibit diagnostic autapomorphies. Nonetheless, these specimens are important data points due to their geographic significance, and some aspects of their morphologies are striking. In one specimen, the teeth express a high degree of wear usually unobserved within this clade—potentially illuminating the progression of the chewing motion in derived stegosaurs. Other morphologies, though not histologically examined in this analysis, have the potential to be important indicators for maturational inferences. In suite with other specimens from the northern extent of the formation, these specimens contribute to the ongoing discussion that body size may be latitudinally significant for stegosaurs—an intriguing geographical hypothesis which further emphasizes that size is not an undeviating proxy for maturity in dinosaurs.
Key words: Dinosauria, Thyreophora, Stegosauria, Jurassic, Morrison Formation, USA, Montana.
D. Cary Woodruff [sauropod4@gmail.com], Great Plains Dinosaur Museum and Field Station, Malta, MT, USA; and Department of Ecology and Evolutionary Biology, University of Toronto; Royal Ontario Museum, 405 N 1st Ave E, Malta, MT 59538, Toronto, ON, Canada.
David Trexler [Dinoguy10@yahoo.com], Two Medicine Dinosaur Center, 120 2nd Ave S, Bynum, MT 59419, Bynum, MT, USA.
Susannah C. R. Maidment [susannah.maidment@nhm.ac.uk], Department of Earth Sciences, Natural History Museum, Cromwell Rd, South Kensington, London SW7 5BD, UK.
Received 18 December 2018, accepted 13 April 2019, available online 5 August 2019.
Copyright © 2019 D.C. Woodruff et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Stegosauria is a clade of ornithischian (“bird-hipped”) dinosaurs that is characterized by the possession of a series of plates and spikes that extend from the neck to the tail in two parasagittal rows. The best-known genus of the clade, Stegosaurus, is from the Upper Jurassic of the western USA, but members of the clade are known from all continents except for Antarctica and Australia, and their temporal range extends from the Middle Jurassic to the uppermost Lower Cretaceous (Maidment et al. 2008, Tumanova and Alifanov 2018). Despite the iconic status of stegosaurs among Dinosauria, they are poorly known as fossils, with most genera being known from single partial skeletons and/or fragmentary remains. Consequently, they have received relatively little attention, and much remains unknown about their paleobiology.
The Upper Jurassic Morrison Formation of the western USA comprises the deposits of rivers and floodplains that were laid down in a broad shallow basin over about 10 million years in the Kimmeridgian and Tithonian. The deposits host a diverse and well-preserved dinosaurian fauna that includes such well-known taxa as Apatosaurus, Diplodocus, and Stegosaurus. The dinosaurs of the southern part of the Morrison basin, from Colorado, Utah, and southern Wyoming, have been intensively studied for about 150 years, but those from northern outcrops in northern Wyoming and Montana are much less exploited (Foster 2007). An increasing body of evidence suggests that northern Morrison faunas were somewhat different to their southern counterparts (Maidment et al. 2018; Whitlock et al. 2018), but further exploration in this part of the basin is needed. Here, we report two stegosaurian dinosaur specimens from the most northerly outcrops of the Morrison Formation in Montana, one of which is the most northerly occurrence of a dinosaur from the Morrison Formation. The specimens, which comprise fragmentary cranial and postcranial material, are therefore important data points in our understanding of the fauna and diversity of the Morrison Formation.
Institutional abbreviations.—BYU, Brigham Young University Museum of Paleontology, Provo, USA; CM, Carnegie Museum, Pittsburgh, USA; DMN, Dinosaur National Monument, Vernal, USA; FPDM, Fukui Prefectural Dinosaur Museum, Japan; GPDM, Great Plains Dinosaur Museum and Field Station, Malta, USA; HMNH, Hayashibara Museum of Natural History, Okayama, Japan; IVPP, Institute of Vertebrate Paleontology and Paleoanthropology, Beijing, China; ML, Museum of Lourinhã, Portugal; MOR, Museum of the Rockies, Bozeman, USA; NHMUK, Natural History Museum, London, UK; USNM, United States National Museum, Washington D.C., USA; SMA (VFSMA), Sauriermuseum Aathal, Switzerland; YPM, Yale Peabody Museum of Natural History, New Haven, USA; ZDM, Zigong Dinosaur Museum, China.
Systematic palaeontology
Dinosauria (Owen, 1842)
Ornithischia (Seeley, 1887)
Stegosauria (Marsh, 1877)
Stegosauria indet.
Material.—GPDM 205, a partial postcranial skeleton from the Little Snowy Mountains, Fergus County, Montana, USA and GPDM 178, a partial skeleton including skull and partial postcrania from Giffen, Cascade County, Montana, USA (Fig. 1).
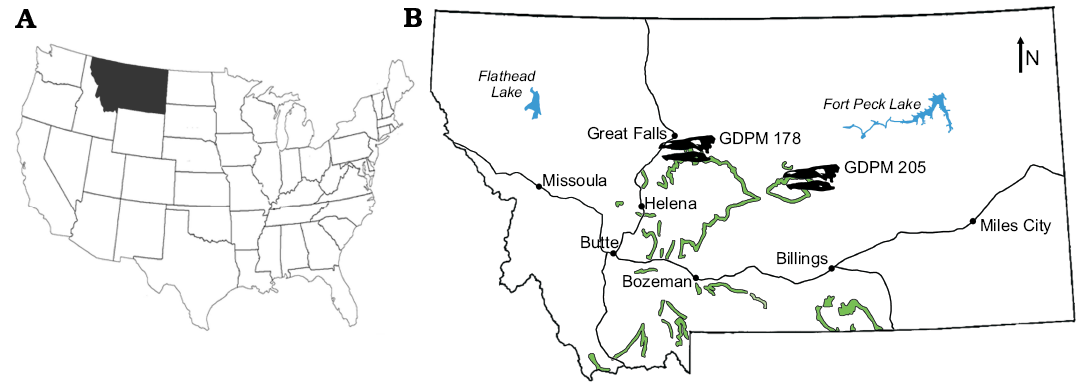
Fig. 1. A. Map of the USA with indicated the state of Montana, were the stegosaur specimens GPDM 178 and GPDM 205 were found. B. Map of Montana with the extent of the Morrison Formation in green, and the general locations of GPDM 178 and GPDM 205 indicated by the Stegosaurus skull silhouettes (modified from Maidment et al. 2018).
GPDM 205 (“Gates”) was collected in 2004 by the Judith River Dinosaur Institute from the 5E Ranch owned by the Hein family, Little Snowy Mountains, Fergus County, Montana. The specimen is now housed at the Great Plains Dinosaur Museum in partnership with the Judith River Foundation. GPDM 205 was collected within the undivided Morrison Formation, and this specimen was found in close proximity to Camarasaurus sp. GPDM 220 (Woodruff and Foster 2017). Unfortunately, neither field notebooks nor quarry maps exist for the excavation of this specimen, so we do not possess any documentation regarding taphonomy. However, SCRM visited the 5E Ranch quarry in 2005 and was told by the excavation crew that the specimen was found disarticulated, stratigraphically above the Camarasaurus remains (GPDM 220). During the 2005 field season, at the time of SCRM’s visit, a carpal and some poorly preserved dermal plates from this specimen were also recovered. Several other stegosaur specimens, referred to Hesperosaurus mjosi (= Stegosaurus mjosi) by Saitta (2015), were discovered during the 2005 field season at the 5E Ranch quarry, and were excavated over subsequent years. These specimens currently reside in a private collection, and thus are unavailable for study (sensu K. Padian and K. Carpenter in Chen 2015). At the current time, aside from a prepared humerus, ulna, and radius, only an unprepared coracoid, fragmentary scapula, humerus, and ulna have been relocated within the collections of the GPDM. Only those elements prepared to-date are described, while the skeletal reconstruction for this specimen (Fig. 2) denotes elements within the GPDM collections.
GPDM 178 (“Giffen”) was collected in 2003 and 2004 by the Judith River Dinosaur Institute near the town of Giffen, Cascade County, Montana, on land belonging to the Yurek family. Elements were initially found eroding out of a hill adjacent to the Yurek’s house. The distal end of the right tibia was the first element recovered, and it was not immediately recognized as a fossil (Linda Yurek, personal communication, 2016). For a period of time, this “black rock” (as described by Linda Yurek), served as a door stop until construction of a pole barn on the hill started, and further elements were discovered below the surface (Linda Yurek personal communication, 2016). At this time the Yurek family contacted the Judith River Dinosaur Institute, and the excavation commenced. The specimen is now housed at the Great Plains Dinosaur Museum in partnership with the Judith River Foundation Collected within the undivided Morrison Formation, GPDM 178 consists of a partial skeleton including both crania and post-crania. Several field jackets that are labeled as dorsal ribs or vertebrae await preparation. Only those elements prepared to-date are described, while the skeletal reconstruction for this specimen (Fig. 2) denotes elements collected via the quarry log.
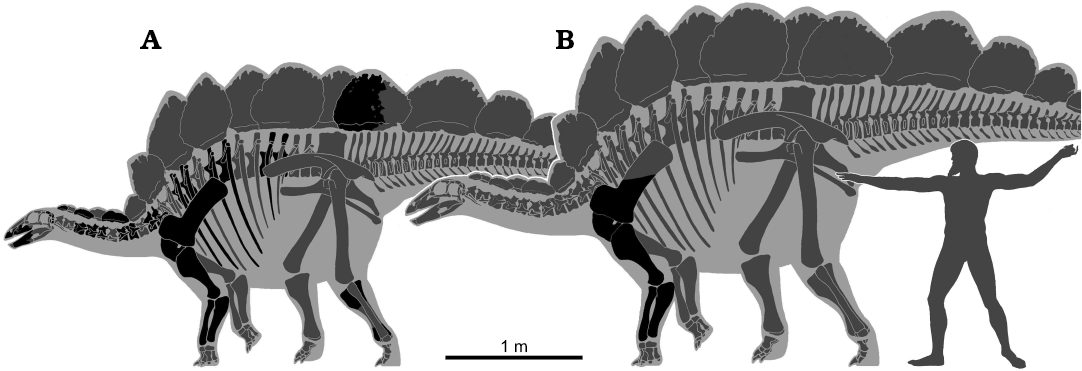
Fig. 2. Skeletal reconstruction of the stegosaurs GPDM 178 (A) and GPDM 205 (B) scaled together based on humeral lengths. Black, elements preserved in each specimen; gray, missing elements. Hesperosaurus reconstruction by Genya Masukawa (Ibaraki University, Japan). Human scale is the Artemision Bronze featuring the deity of Zeus as 1.83 m in height.
Description.—GPDM 205: For measurements of all elements, see Table 1. Humerus: The left humerus is well preserved (Fig. 3). Large cracks and the parts of the diaphysis are filled and/or restored with a form of epoxy clay. In anterior view, the humerus has a dumb bell-shaped profile, as in other stegosaurs (Maidment et al. 2015, 2018), with a stout, cylindrical diaphysis. The deltopectoral crest appears to be larger and project far more medially than in some other stegosaur specimens (e.g., Stegosaurus, NHMUK PV R36730, Maidment et al. 2015; Hesperosaurus, MOR 9728, Maidment et al. 2018; Dacentrurus, NHMUK PV OR 46013, Loricatosaurus, NHMUK PV R3167, Miragaia ML 433, Mateus et al. 2009; Kentrosaurus, NHMUK PV R37143, Henning 1925) but is similar to the condition in other specimens such as YPM 1853, YPM 1856 (Stegosaurus), MB R.4804, and MB R.4805 (Kentrosaurus). This variation appears to be intraspecific and is likely to be taphonomic or related to restoration. Transversely, the deltopectoral crest is fairly uniform in thickness throughout, only tapering at the anteromedial end. The anterolateral margin of the deltopectoral crest, particularly the anteriormost aspect, has a rugose surface texture. This rugose margin forms a “lip” along the deltopectoral crest, and in combination with the medially convex curvature of the crest, produces a prominent bowl-shaped crest. Unlike the deltopectoral crest in Stegosaurus stenops (NHMUK PV R36730; Maidment et al. 2015), which merges without a break in slope into the proximal aspect of the humerus, the deltopectoral crest of GPDM 205 has a dip, or a break in slope, along the proximal contact between the crest and diaphysis. In posterior orientation a prominent dorsomedial, posterolateral angled triceps ridge traverses along the entire length of the deltopectoral crest; this is a synapomorphy of Stegosauria (Raven and Maidment 2017). The large humeral head is very bulbous, and 68% the transverse width of the distal condyles. In lateral and medial orientations, the bulbous humeral head is offset posteriorly. Along the medial edge of the humerus, just distal to the humeral head is a symmetrical, triangular-shaped process. Though nowhere near in size, this medial triangular process opposes the deltopectoral crest.
Table 1. Greatest measurements (in mm) for GPDM 205. 1 anterior-posterior length 2 lateral width; r reconstructed.
|
Forelimb |
Humerus |
Ulna |
Radius |
|
|
Length (dorsal-ventral) |
516 |
517 |
384 |
|
|
Proximal end |
length |
121.531 |
136.83 |
110.26 |
|
width |
1512 |
1152 |
91.752 |
|
|
Distal end |
length |
115.631 |
292 |
93.18 |
|
width |
2172 |
59.04 |
92.09 r |
|
|
Deltopectoral crest length |
186.61 |
– |
– |
|
|
Diaphysis mimimum circumference |
278 |
192 |
168 |
|
The diaphysis, though largely restored, (see above), appears to be circular to sub-circular in cross section. Before sharply transversely expanding into the proximal and distal ends, this sub-circular diaphysis is uniform in thickness. The straight diaphysis, together with the slightly posteriorly oriented proximal end and the slightly anteriorly oriented distal end, gives the humerus a slightly sigmoidal lateral and medial profile. The distal portion of the humerus terminates with the transversely widened and bulbous articular condyles. The distal aspect of the humerus is 73% the width of the proximal end—contributing to the dumb bell profile. The medial epicondyle in most orientations is fairly spherical in overall morphology. The lateral epicondyle is larger than its counterpart. In anterior and posterior view, the lateral epicondyle has a sub-triangular profile that projects anterolaterally. While this condyle projects slightly anteriorly, it does not possess the triangular anterior eminence. In ventral view the lateral epicondyle is 1.3 times as transversely wide and 1.08 times anteroposteriorly elongate than the medial epicondyle.
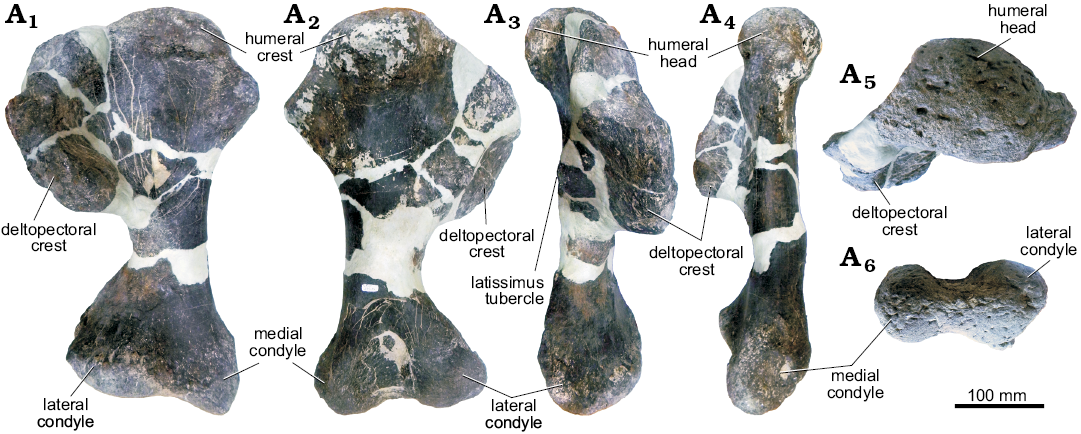
Fig. 3. Left humerus of stegosaur GPDM 205 from the 5E Ranch, Little Snowy Mountains, Fergus County, Montana, USA; in anterior (A1), posterior (A2), lateral (A3), medial (A4), dorsal (A5), and ventral (A6) views.
Ulna: The left ulna is quite well preserved; a few large cracks are filled in with epoxy clay (Fig. 4). In anterior and posterior views, the right ulna has a slightly sinusoidal profile. The olecranon process consists of a prominent triangular bony projection which extends dorsally from the diaphysis. In GPDM 205, the olecranon process is a very prominent scalene-like process, comparable in development to that of USNM 4937 (“Stegosaurus sulcatus”), YPM 1854 (Stegosauria indet.), and MB R.4800 (Kentrosaurus). In some other specimens, e.g., BYU 12290, DNM 2438, and NHMUK PV R36730 (Stegosaurus stenops), the olecranon process is less well-developed. The olecranon process of GPDM 205 is 45% the total dorsoventral length, and has a transverse thickness that is over 1.5 times that of the thinnest portion of the diaphysis. The development of the olecranon process has been related to ontogenetic stage in stegosaurs (Galton 1982a), and the well-developed olecranon process in this specimen may indicate the individual was a mature adult at time of death. This hypothesis is supported by the observation that all surfaces of the olecranon process have a very rugose bone texture. Ossification of cartilage to give a rugose bone texture in mature individuals has also been noted in other stegosaurs (Mallison 2010) and ossification of the triceps tendon to give an “olecranon horn” has been described in the stegosaurs Miragaia and Dacentrurus (Mateus et al. 2009).
In anterior view, the anterior and medial processes are more parallel and similarly oriented to one another in contrast in Stegosaurus stenops (NHMUK PV R36730; Maidment et al. 2015), in which the medial process appears nearly perpendicularly oriented to the anterior process. In Stegosaurus stenops (NHMUK PV R36730; Maidment et al. 2015), there are gently curved regions transitioning from the three proximal processes (the olecranon, anterior, and medial processes), yet in GPDM 205 the transition from each of these processes is more abrupt. For example, in GPDM 205 the medial process appears nearly flat with a sharp incline of 126° to the olecranon process. Along the entire outer length of the olecranon process is a transversely thickened margin. With this thickened outer margin and a slight curvature of this process the medial aspect is slightly concave.
In dorsal view, the olecranon process is significantly larger than the medial and anterior processes. Distal to the olecranon process, the diaphysis is fairly straight and ovoid to sub-circular in cross section, before abruptly expanding and terminating. In distal view, the distalmost region of the ulna is gently convex, and is tear dropped shaped with the tapered apex pointing anteriorly. A large, flat facet for the distal end of the radius is present, and as in Stegosaurus stenops (NHMUK PV R36730; Maidment et al. 2015), medial to this facet is a prominent dorsoventrally oriented ridge. Observable in posterior, lateral, and medial views in Stegosaurus stenops (NHMUK PV R36730; Maidment et al. 2015), the distal region of the ulna has a slight laterally oriented curvature (opposed to the largely straight rest of the ulna). Yet in GPDM 205, this region is more kinked than curved, and more similar to the corresponding region of Kentrosaurus aethiopicus (Galton 1982b) and Miragaia longicollum (ML433; Mateus et al. 2009).
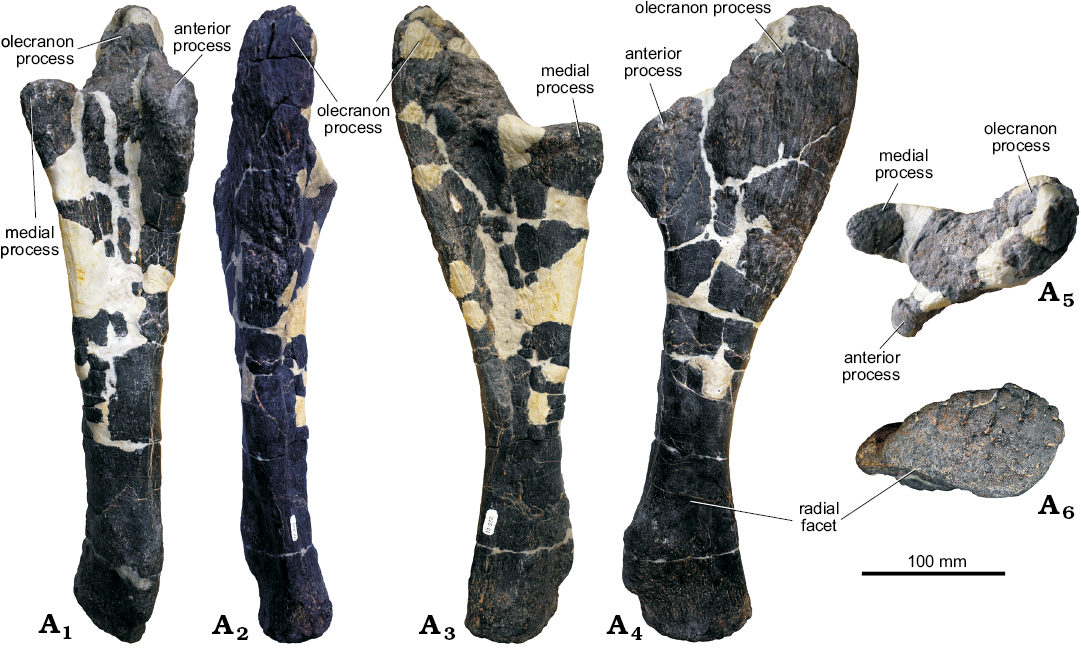
Fig. 4. Left ulna of stegosaur GPDM 205 from the 5E Ranch, Little Snowy Mountains, Fergus County, Montana, USA; in anterior (A1), posterior (A2), lateral (A3), medial (A4), dorsal (A5), and ventral (A6) views.
Radius: Like the humerus, the existing portion of the left radius appears to be well-preserved (Fig. 5). Large cracks in the diaphysis are filled in with epoxy clay, and a significant portion of the proximal region is entirely sculpted. While Stegosaurus stenops (NHMUK PV R36730; Maidment et al. 2015), has a dorsoventrally short, almost rectangular radius in lateral view, the radius of GPDM 205 is more like that of Kentrosaurus aethiopicus (Galton 1982b) in that it is very dorsoventrally elongate, with a much more transversely widened distal region.
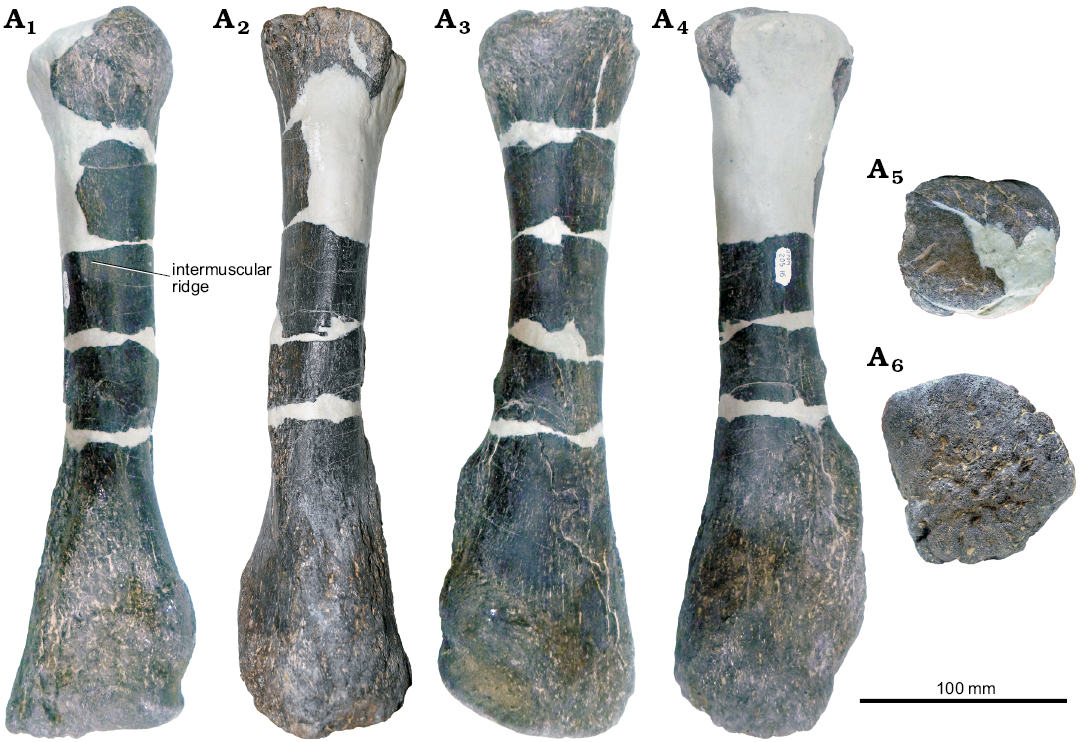
Fig. 5. Left radius of stegosaur GPDM 205 from the 5E Ranch, Little Snowy Mountains, Fergus County, Montana, USA; in lateral (A1), medial (A2), anterior (A3), posterior (A4), dorsal (A5), and ventral (A6) views.
GPDM 178: For measurements of all elements, see Table 2. Skull: The skull consist of a partial braincase, both maxillae, both dentaries, and the left premaxilla (Fig. 6).
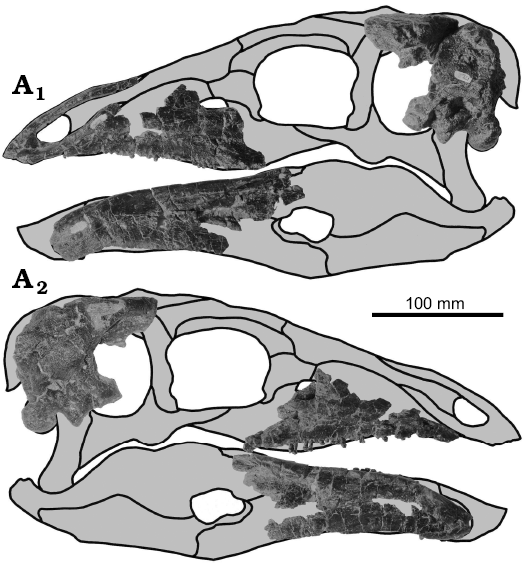
Fig. 6. Cranial elements of stegosaur GPDM 178, near Giffen, Cascade County, Montana, USA; in left (A1) and right (A2) lateral views; gray, missing elements.
Table 2. Greatest measurements (in mm) for GPDM 178. 1 anterior-posterior length; 2 lateral width; 3 anterior-posterior height; 4 length of ventral margin; d, damaged; r, reconstructed.
|
Cervical vertebra |
Centrum |
Anterior face |
Posterior face |
Neural arch |
Neural canal |
Prezyga-pophyses |
Postzyga-pophyses |
Transverse processes |
|
|
anterior view |
posterior view |
||||||||
|
Length (anterior-posterior) |
80.27 |
|
|
68.09 |
|
|
|
|
|
|
Width (lateral) |
|
78.30 |
67.94 |
|
34.11 |
45.16 |
68.67d |
52.45 |
105.99d |
|
Height (dorsal-ventral) |
|
65.33 |
68.25 |
71.43 |
34.803 |
25.743 |
|
|
|
|
Skull |
Cranium |
Maxilla |
Dentary |
Premaxilla |
Dorsal plate |
|||
|
posterior cranium |
foramen magnum |
left |
right |
left |
right |
left |
||
|
Length (anterior-posterior) |
117.97 |
|
155.52 |
170.1 |
190.8 |
204 |
123.10 |
4554 |
|
Width (lateral) |
81.26 |
21.99 |
13.03 |
17.32 |
27.30 |
26.93 |
17.76 |
86.894 |
|
Height (dorsal-ventral) |
98.78 |
26.73 |
65.99 |
65.96 |
57.77 |
72.38 |
53.72 |
550.2 |
|
Occicipital condyle height (dorsal-ventral) |
28.36 |
|
|
|
|
|
|
|
|
Tooth row length (anterior-posterior) |
|
|
116.62 |
93.81 |
124.02 |
127.59 |
|
|
|
Shoulder girdle |
Coracoid |
Scapula |
Articular connection with scapula |
|
|
Length |
anterior-posterior |
228 r |
638 r |
|
|
dorsal-ventral |
285 r |
|
130.25 |
|
|
Width (lateral) |
|
|
90.72 |
|
|
Anterior end length (proximal-distal) |
|
346 |
|
|
|
Mid-length scapular blade length (proximal-distal) |
|
146.31 |
|
|
|
Acromion (?) length (anterior-posterior) |
|
234 r |
|
|
|
Glenoid |
length (dorsal-ventral) |
|
118.54 |
|
|
width (lateral) |
|
86.16 |
|
|
|
Forelimb |
Humerus |
Ulna |
Radius |
|
|
Length (dorsal-ventral) |
454 |
374 |
364 |
|
|
Proximal end |
length |
78.511 |
165 |
116.21 |
|
width |
153 |
118.34 |
71.43 |
|
|
Distal end |
length |
114.171 |
63.55 |
109.43 |
|
width |
2042 |
83.59 |
87.98 |
|
|
Deltopectoral crest length |
125.05 |
– |
– |
|
|
Mimimum circum-ference of diaphysis |
257 |
184 |
139 |
|
Premaxilla: Given its size and delicacy, the left premaxilla is in excellent condition (Fig. 7). The premaxilla is edentulous, and as in some other stegosaurs such as Miragaia (ML 433; Mateus et al. 2009) and Stegosaurus (NHMUK PV R36730), but in contrast to the basal taxa Huayangosaurus (ZDM T7001), and Isaberrysaura (Salgado et al. 2017). The element has an anteroposteriorly elongate profile, being nearly two times the anteroposterior length than dorsoventral height. The narial opening is similarly anteroposteriorly elongate. The anteriormost region is well preserved, and possesses a crenulated and slightly rugose border (denticles; David C. Evans personal communication 2018) that would have served as an attachment for the rhamphotheca. The nasal process projects posterodorsally from the anterior margin of the bone and is triangular in cross-section with the apex pointed laterally, forming a ridge on the lateral surface of the process. It is broken at its dorsal extremity. The process is delicate and gracile, as in Stegosaurus (NHMUK PV R36730) and Miragaia (ML 433) but differs from the condition in Huayangosaurus (ZDM T7001) where it is more robust. The maxillary process forms the ventral margin of the external naris and then curves dorsally proceeding posteriorly. There is a broad, flat to slightly concave articular facet for the maxilla.

Fig. 7. Left premaxilla of stegosaur GPDM 178, near Giffen, Cascade County, Montana; in dorsal (A1), lateral (A2), medial (A3), and ventral (A4) views.
Maxilla: Neither maxilla is complete, but as preserved they are both triangular in lateral or medial view with gently upwardly convex ventral, tooth-bearing margins (Fig. 8). The anteriormost ~2–3 cm of both the left and right element is edentulous, as in Stegosaurus (USNM 4934), this diastema is transversely thin, but thickens at the tooth row. In the left maxilla 27 alveoli are present, and at least 23 in the right. There are 27 or 28 maxillary teeth present in Huayangosaurus (ZDM T7001), at least 22 are present in Stegosaurus (USNM 4934; Gilmore 1914) and 20 are present in Hesperosaurus (HMNH 001; Carpenter et al. 2001). The anterodorsal and posterodorsal margins of neither maxillae are preserved, so that relationships with other cranial elements remain unknown and the presence or absence of an antorbital fossa cannot be demonstrated.

Fig. 8. Stegosaur GPDM 178, near Giffen, Cascade County, Montana. A. Left maxilla in dorsal (A1), lateral (A2), medial (A3), and ventral (A4) views. B. Right maxilla in dorsal (B1), lateral (B2), medial (B3), and ventral (B4) views.
Dentary: Both dentaries are largely complete, except for the posteriormost parts (Fig. 9). In lateral and medial views, the dentary has a gentle sigmoidal profile. This curved profile is slight along the dorsal margin, yet more pronounced along the ventral margin, especially the anterior aspect. Overall the dentaries are transversely thin, except for the symphysis and the tooth row. The anterior is the laterally thickest portion of this element. The tooth row occupies the medial side of a medially projecting thickened ridge of bone. As in Stegosaurus (NHMUK PV R36730), but in contrast to Huayangosaurus (ZDM T7001), Gigantspinosaurus (ZDM 0019), and Jiangjunosaurus (Jia et al. 2007), the tooth row is angled slightly medially and obscured in lateral view by a thin ridge of bone.
The anterior one third of the dentary is edentulous, as in Stegosaurus (NHMUK PV R36730) and Jiangjunosaurus (Jia et al. 2007), but in contrast to the condition in Huayangosaurus (ZDM T7001), Kentrosaurus (MB.R.3806.1), and Gigantspinosaurus (ZDM 0019), where the tooth row starts immediately posterior to the predentary articular facet. The tooth row occupies nearly the entirety of the remaining length of the dentary. The tooth row is curved to gently sigmoidal, as in Huayangosaurus (ZDM T7001), Gigantspinosaurus (ZDM 0019), and Jiangjunosaurus (Jia et al. 2007) but in contrast to the condition usually seen in Stegosaurus, where it is straighter (e.g., NHMUK PV R36730). The left dentary possesses at least 24 alveoli, and the right possesses perhaps up to 26 alveoli. There are 21 dentary teeth in Huayangosaurus (ZDM T7001) and Jiangjunosaurus (Jia et al. 2007), more than 30 in Gigantspinosaurus (Ouyang 1992), and the number in Stegosaurus appears to vary from around 17 to 23 (Berman and McIntosh 1986)
Teeth: The maxillary and dentary teeth of GPDM 178 are similar to other stegosaurs, being leaf-shaped (Galton and Upchurch 2004). In lingual view the teeth are fairly symmetric with a ringed cingulum observed in all teeth. In labial view all teeth have a fairly prominent midline central denticle with several denticles of varying size and number (approximately four to eight) on either side of the midline denticle. In labial, lingual, and apical views, these denticles are strongly convex, and with the ringed cingulum, the teeth have rounded profiles.

Fig. 9. Stegosaur GPDM 178, near Giffen, Cascade County, Montana. A. Left dentary in dorsal (A1), medial (A2), lateral (A3), and ventral (A4) views. B. Right dentary in dorsal (B1), medial (B2), lateral (B3), and ventral (B4) views.
Perhaps the most interesting aspect of the teeth of GPDM 178 are the wear facets (Fig. 10). Wear facets are known in many stegosaurian teeth (Gigantspinosaurus, Hao et al. 2018; Huayangosaurus, Sereno and Dong 1992; Kentrosaurus, Hennig 1925; and Stegosaurus, Galton and Upchurch 2004), and these wear facets are largely sub-horizontal to gently inclined posteriorly and labially (Galton and Upchurch 2004; Reichel 2010). Stegosaurian tooth wear facets led Galton and Upchurch (2004) to suggest they were formed by tooth-food contact, which would indicate chewing; but the rarity and general weak degree of said facets would suggest that this was not a common occurrence. However, Reichel (2010) noted that some stegosaurian teeth have extensive wear facets, indicating prolonged chewing and/or a longer interval that the tooth was active. Interestingly, nearly all of the maxillary and dentary teeth of GPDM 178 have extensive wear facets. The majority of these wear facets are sub-horizontal, and the size of the facets vary from approximately ¼ to ½ of the crown height. The teeth of GPDM 178 have not been sectioned to document the Lines of Von Ebner correlating to tooth age in dinosaurs was studied in the Erickson (1996) analysis, so at this time we cannot say if the extensive wear facets are a direct result of the teeth being active and worn down over a prolonged period, or if they indicate a greater degree of chewing. However, contra to Hao et al. (2018), while the degree of wear may indicate that the tooth itself is more mature (i.e., it has been an erupted functional tooth for a longer period of time), as dinosaurs continually shed their teeth throughout their lives, tooth wear is not an acceptable indicator of an individual’s maturational status.
Within the wear facets of GPDM 178 are smaller features that may hint at the cause of these facets. In a majority of the sub-horizontal wear facets are numerous small-scale gouges (Fig. 10C). These gouges appear entirely restricted to the wear facets, and they uniformly consist of a series of anteroposteriorly oriented gouges with another sets of gouges cross-cutting roughly perpendicular. If the marks were infrequent or an isolated occurrence, they could be associated with taphonomic wear or even preparation damage. However, given the prevalence and uniformity of these gouges, we hypothesize that they are in fact signs of microwear.

Fig. 10. Maxillary and dentary teeth of stegosaur GPDM 178 (A–D), near Giffen, Cascade County, Montana; C2, explanatory drawing showing the wear facet and cross-cutting gouges. Not to scale, anteroposterior wear facet length of approximately 2–2.5 mm.
Posterior cranium: A co-ossified unit consists of the braincase, partial frontals, and the parietal (Fig. 11). In lateral view the frontal region slopes gently anteroventrally. The parietal is not well preserved dorsally, but is roughly square in outline, and flat, as in Stegosaurus (USNM 4934, DMNH 2818), Huayangosaurus (ZDM T7001), and Hesperosaurus (MOR 9728; Maidment et al. 2018). The frontals are flat dorsally, and incomplete anteriorly. Additionally, the sutures between the frontals and parietal is not clear, and the left and right frontals cannot be observed because the bone surface is poorly preserved in this area.
In posterior view, the supraoccipital is not well preserved, and the braincase is damaged dorsal to the foramen magnum. The paroccipital processes have broken off and are not preserved. The occipital condyle projects posteriorly, as in other stegosaurs (Hesperosaurus mjosi, Carpenter et al. 2001; Huayangosaurus taibaii, ZDM T7001; Kentrosaurus aethiopicus, Hennig 1925; and Stegosaurus stenops, NHMUK PV R 36730). In posterior view, the occipital condyle is ovoid (laterally wider than dorsoventrally tall) as in Huayangosaurus taibaii (ZDM T7001), Kentrosaurus aethiopicus (Hennig 1925), and Stegosaurus stenops (NHMUK PV R 36730). The foramen magnum is circular, and is approximately similar in dorsoventral height as the occipital condyle, but is transversely much smaller, unlike that of Hesperosaurus mjosi (Carpenter et al. 2001), but similar to Huayangosaurus taibaii (ZDM T7001), Kentrosaurus aethiopicus (Hennig 1925), and Stegosaurus stenops (Carpenter et al. 2001). In ventral view, the suture between the basisphenoid and basioccipital cannot be observed, as in other stegosaurs (Maidment et al. 2018). Anterior to the occipital condyle, the basioccipital-basisphenoid narrows transversely into a short neck, which is anteroposteriorly shorter than the occipital condyle. The fused elements then flare transversely to form the basal tubera, which are separated by a midline groove. In lateral view the basal tubera are triangular with the apex pointing ventrally, and in ventral view they are ovoid in shape, being wider transversely than they are anteroposteriorly. The basal tubera extend further ventrally than the occipital condyle, as in Stegosaurus stenops (USNM 4934), and Hesperosaurus mjosi (MOR 9728), but in contrast to the condition in the basal thyreophoran Scelidosaurus (NHMUK PV R1111). Anterior to the basal tubera, the basipterygoid processes are broken, and anterior to this area, fused to the ventral surface of the frontal, the orbitosphenoid is present as a ventral convex, hour glass-shaped element opening anteriorly. Ossified orbitosphenoids are observed in specimens of Stegosaurus (CM 106; Gilmore 1914) and are also present in the basal thyreophoran Scelidosaurus (NHMUK PV R1111).
In lateral view, the elements of the braincase are co-ossified and sutures cannot be observed. On the left side, the crista prootica of the prootic lies dorsal to a large, heart-shaped fenestra, which is the combined opening of the fenestra ovalis and fenestra pseudorotunda. The crista prootica is not as well preserved on the right side. On both sides, posterior to the fenestra ovalis, are two small foramina that likely represent the exits of the hypoglossal nerve (XII), as in the basal ornithischian Lesothosaurus (Sereno 1991). Anterior to the fenestra ovalis, a groove is present extending anteroventrally. A foramen situated in the groove at the same level as the fenestra ovalis is probably the exit for the facial nerve (VII). Ventral to this foramen, on the left side the groove is weathered and opens into a large concavity, the sella turcica. On the right side, close to the anterior margin of the braincase, the trigeminal foramen, for the exit of the maxillary and mandibular branches of the trigeminal nerve (V), is present. Dorsal to these fenestrae and foramina the lateral wall of the braincase is composed of the prootic, which is smooth and featureless. The suture between the prootic and parietal on the right side is marked by an irregular concavity that has probably been accentuated by erosion. The laterosphenoid extends anteriorly from the dorsal margin of the trigeminal foramen to contact the frontal, and appears to be roughly triangular in shape, as in the basal ornithischian Lesothosaurus, although sutures cannot be seen.

Fig. 11. Braincase of stegosaur GPDM 178, near Giffen, Cascade County, Montana; in anterior (A1), left (A2) and right (A5) lateral, dorsal (A3), posterior (A4), and ventral (A6) views.
Postcranial material: Cervical vertebra: The one fully prepared cervical vertebra is generally well-preserved (Fig. 12). By comparison with Stegosaurus stenops (NHMUK PV R36730), this vertebra appears to be an anterior cervical, perhaps cervical three or four. In lateral view, the overall vertebral profile (centrum + neural arch) is rhomboid. The anterior and posterior articular facets of the centrum are nearly flat to slightly concave. In lateral view, the centrum is roughly square; the anterior and posterior facets are slightly offset, producing a slightly kinked centrum profile. In anterior view, the centrum facet is largely circular in outline. Along the ~mid anterior centrum facet, the parapophyseal facets abut onto the rim of the centrum producing laterally projecting subtriangular profiles at this margin. In posterior view the posterior articular facet, like the anterior one, is circular in outline, except for the anterior margin with the neural canal that is rather horizontal. In lateral and ventral views, the ventral profile of the centrum is upwardly concave. This concave ventral margin does not exhibit a ventral keel or ridge. As observed to a lesser degree in Stegosaurus stenops (NHMUK PV R36730; Maidment et al. 2015), in lateral view, slightly posteroventral to the parapophyseal facets is a single, small foramen (approximately 3.8 mm anteroposteriorly long by 2.9 mm dorsoventrally tall) which is probably vascular.
While the neural arch is fully fused to the centrum, the neurocentral suture is clearly visible on all sides. The neural canal of this vertebra is very large, being 43% dorsoventrally tall and 44% laterally wide as the centrum. In anterior view, except for the curved dorsal margin, the neural canal is almost square in anterior profile, being 1.09 times dorsoventrally tall than laterally wide. In both anterodorsal and posterodorsal oblique views, the ventral margin of the neural canal is strongly curved and pervades into the midline of the centrum. In posterior view, the profile of the neural canal is triangular with the apex pointing dorsally. This triangular neural canal profile is laterally wide (approximately 1.75 times wider than tall). The left prezygapophysis is broken, and the right is damaged. The articular facet of the right prezygapophysis is strongly dorsoventrally inclined as in Stegosaurus stenops (NHMUK PV R36730; Maidment et al. 2015). The neural spine, though damaged, is slightly posteriorly inclined. The dorsal margin of the postzygapophyses are anteroposteriorly elongate and nearly horizontal. The ventral margin is gently anteriorly curved, and this curved profile seamlessly traverses ventrally to the neurocentral suture. The articular facet of the right postzygapophysis is not preserved, but the left is elongate, being 74% the length of the centrum. Anteroposteriorly elongate articular facet are observed in the Morrison stegosaurs Hesperosaurus mjosi (Carpenter et al. 2001) and Stegosaurus stenops (NHMUK PV R36730; Maidment et al. 2015).

Fig. 12. Cervical vertebra of stegosaur GPDM 178, near Giffen, Cascade County, Montana; in anterior (A1), left (A2) and right (A5) lateral, dorsal (A3), posterior (A4), and ventral (A6) views.
Scapula: The left scapula is preserved (Fig. 13). The blade is distally incomplete and the whole of the medial border has been reconstructed with epoxy clay. The dorsal surface of the scapular blade is nearly straight, and flares slightly distally. In lateral view, a ridge extends anteroposteriorly along the scapula blade, as in Stegosaurus stenops (NHMUK PV R36730). The proximal plate is square in outline with a prominent dorsoventrally oriented acromial ridge, as in Stegosaurus stenops (Maidment et al. 2015). In both dorsal and ventral view, the scapula has a gentle laterally oriented curvature. The apex of this curvature is approximately at mid length. In anterior view, the proximal plate is triangular in cross section. The ventral aspect of the plate is the transversely widest portion, and both lateral and medial borders taper proximally. The scapular portion of the glenoid fossa is transversely widened and oval in anterior view. In ventral orientation, the overall profile is triangular, the anterior region is transversely widened and strongly tapers posteriorly. The anterior aspect is over 6 times as transversely wide than the posteriormost region.
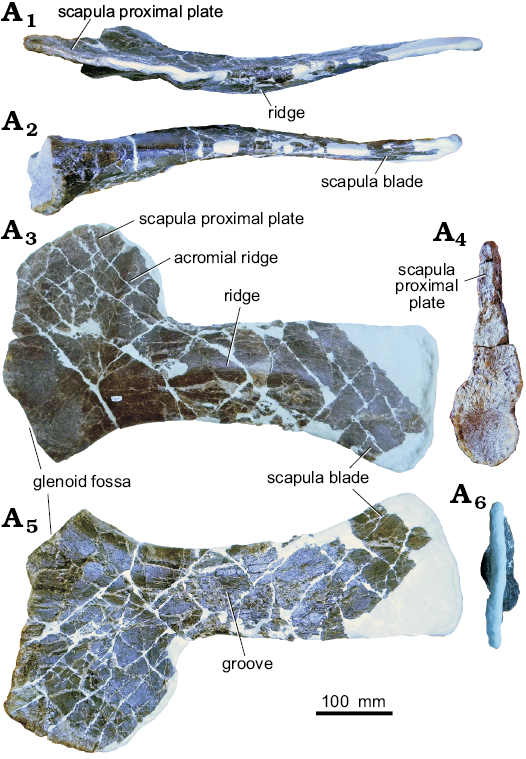
Fig. 13. Left scapula of stegosaur GPDM 178, near Giffen, Cascade County, Montana; in dorsal (A1), ventral (A2), lateral (A3), anterior (A4), medial (A5), and posterior (A6) views.
Coracoid: The left coracoid is not fused with the scapula (Fig. 14). Fusion of the scapula and coracoid in stegosaurs is intraspecifically variable and may be related to ontogenetic stage (Maidment et al. 2015). The anterior and dorsal margin of the coracoid is missing and has been reconstructed in epoxy clay. This reconstruction, likely not an accurate representation, gives the coracoid a pentagonal profile. In other stegosaurs the coracoid is sub-circular (Miragaia longicollum, ML 433, Mateus et al. 2009; Stegosaurus stenops, NHMUK PV R36730, Maidment et al. 2015) to ovoid (Kentrosaurus aethiopicus, MB R.4802 and MB R.4803, Hennig 1925). However, while the anterior and dorsal margins are incomplete, the ventral, glenoid, and posterior margins merge with distinct corners or breaks of slope, unlike that of Stegosaurus stenops (NHMUK PV R36730; Maidment et al. 2015).
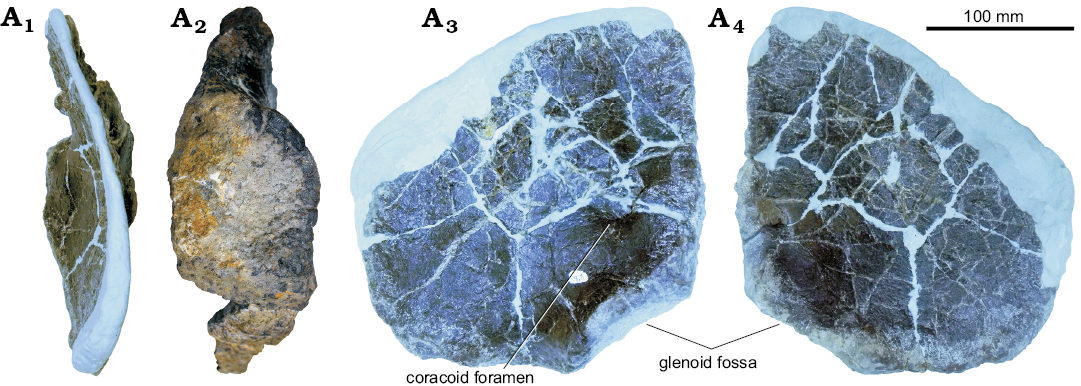
Fig. 14. Left coracoid of stegosaur GPDM 178, near Giffen, Cascade County, Montana; in anterior (A1), lateral (A2), posterior (A3), and anterior (A4) views.
Humerus: The left humerus of GPDM 178 is largely complete, except for some large cracks filled with epoxy clay and a reconstructed apex on the deltopectoral crest (Fig. 15). In anterior view, the humerus has the dumb-bell profile observed in GPDM 205 and other stegosaurs (Maidment et al. 2015, 2018). The overall morphology of deltopectoral crest of GPDM 178 is very similar to that of the Stegosaurus stenops (NHMUK PV R36730) and the Hesperosaurus mjosi (MOR 9728; Maidment et al. 2015, 2018). The deltopectoral crest occupies approximately the proximal half of the element. Transversely, the deltopectoral crest is fairly uniform in thickness, with a gentle taper along the distal aspect. The anterolateral margin of the deltopectoral crest is rugose and bears a slight rugose “lip” similar to GPDM 205; however, this rugosity and lipped region are much less pronounced in GPDM 178, and more akin to the morphologies of Stegosaurus stenops (NHMUK PV R36730) and Hesperosaurus mjosi (MOR 9728; Maidment et al. 2015, 2018). While the deltopectoral crest of GPDM 178 does have a slight anteromedial curve, it is nowhere near as curved or bowl-shaped as GPDM 205 or Miragaia longicollum (ML 433; Mateus et al. 2009). As in Stegosaurus stenops (NHMUK PV R36730) and Hesperosaurus mjosi (MOR 9728; Maidment et al. 2015, 2018), in anterior view the deltopectoral crest of GPDM 178 is somewhat proximodistally straight with the distal end oriented more medially. However, in lateral or medial view, the deltopectoral crest of GPDM 178 is like that of GPDM 205 in that there is a dip in the slope along the proximal contact between the crest and humeral body. This break in slope is weaker than that of GPDM 205, but it is not the continuous, undisrupted contact seen in the humerus of Stegosaurus stenops (NHMUK PV R36730; Maidment et al. 2015) or Miragaia longicollum (ML 433; Mateus et al. 2009). The humeral head is bulbous, rounded in outline, and offset posteriorly. The humeral head is 61% of the transverse width of the distal condyles. As in GPDM 205, distal to the humeral head on the medial side of the humerus is a symmetrical, triangular-shaped process. This triangular process (which is opposed the deltopectoral crest) projects anteromedially, and like the deltopectoral crest is proportionally smaller in GPDM 178 compared to that of GPDM 205. In lateral view the lateral dorsal process is slightly anteroposteriorly inclined with a slight anterior oriented concavity.
The nearly intact diaphysis appears to be circular to sub-circular in cross-section. The sub-rounded diaphysis transversely expands into the proximal and distal ends. This straight diaphysis, in combination with the slightly posteriorly angled proximal end and the slightly anteriorly angled distal end, gives the humerus a slightly sigmoidal profile in lateral view (as in GPDM 205, the Hesperosaurus mjosi, MOR 9728, Maidment et al. 2018, and to a much lesser degree in the Stegosaurus stenops NHMUK PV R36730, Maidment et al. 2015; versus the practically straight humeral profile of Miragaia longicollum ML433, Mateus et al. 2009). The distal portion of the humerus terminates with the transversely widened and bulbous articular condyles. In all views the medial epicondyle is ovoid in overall morphology; but as in GPDM 205, in lateral and medial views, there is a slight triangular, anteriorly oriented eminence. The lateral epicondyle is by far the larger of the two condyles. In anterior and posterior view, the lateral epicondyle has a sub-circular profile and is 1.16 times as proximodistally long as the lateral epicondyle. In ventral view, both the lateral and medial epicondyles are ovoid and have similar orientations. In ventral view the lateral epicondyle is 1.5 times as transversely wide and ~1.3 times as anteroposteriorly elongate than the medial epicondyle.
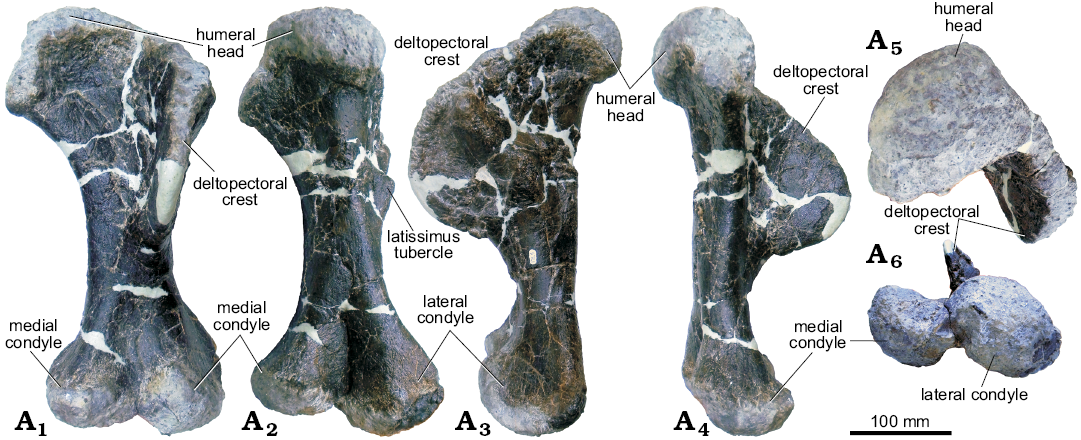
Fig. 15. Left humerus of stegosaur GPDM 178, near Giffen, Cascade County, Montana; in anterior (A1), posterior (A2), lateral (A3), medial (A4), dorsal (A5), and ventral (A6) views.
Ulna: Like the humerus, the left ulna of GPDM 178 is largely complete, and well preserved (Fig. 16). Likewise, all of the forelimb material of GPDM 178 appears to be much more “gracile” than those counterparts of GPDM 205 (and these differences are addressed in the Discussion section). In all views, the majority of the element is dorsoventrally straight, with only a slight kink in the distalmost end. The olecranon is the most prominent of the ulnar processes—being approximately twice the dorsoventral height of the medial process, and rather bulbous anteroposteriorly and lateromedially.
In anterior view, the anterior and medial processes are subparallel to one another (with the anterior process being slightly more proximal). The transition between these three proximal processes (the olecranon, anterior, and medial processes) is gentle and curved, more akin to that of Stegosaurus stenops (NHMUK PV R36730; Maidment et al. 2015) than to GPDM 205.
Distal to the proximal processes, there is a slightly flared “ring” or region that demarks these processes from the diaphysis. The diaphysis distally tapers and is sub-circular to ovoid at mid diaphysis cross section. The distal end of the ulna is slightly expanded and kinked relative to the long axis. In distal view, the distalmost aspect of the ulna is convexed and shaped like an isosceles triangle, with the apex pointing anteriorly. A flat facet for the distal end of the radius is present, but the size of this facet and the adjacent dorsoventrally oriented ridge is more reduced than these features seen in GPDM 205 and Stegosaurus stenops (NHMUK PV R36730, Maidment et al. 2015).
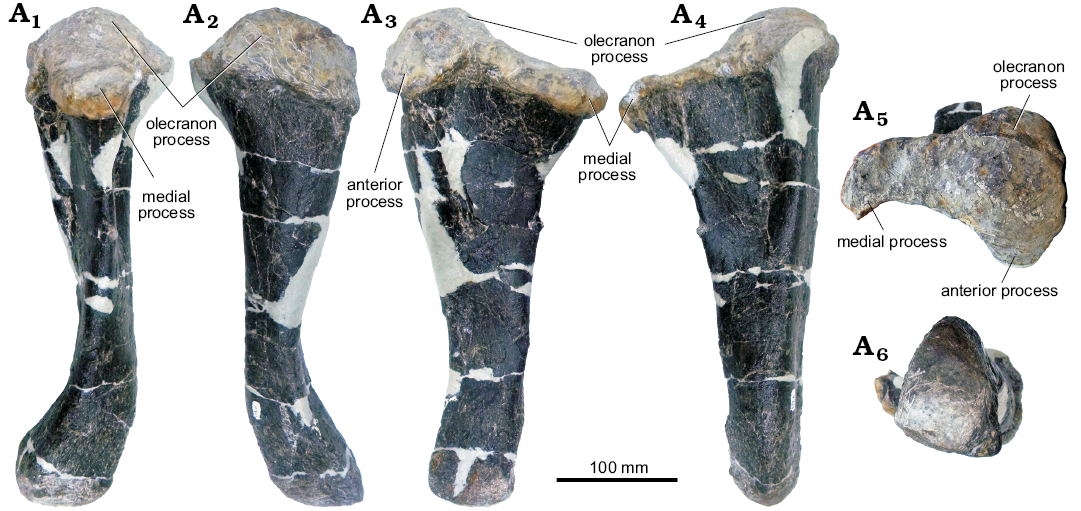
Fig. 16. Left ulna of stegosaur GPDM 178, near Giffen, Cascade County, Montana; in medial (A1), posterior (A2), lateral (A3), dorsal (A4), and ventral (A5) views.
Radius: The radius, like the other forelimb elements of GPDM 178 is well preserved (Fig. 17). Large cracks in the diaphysis are filled in with epoxy clay, but no portions of the element are sculpted or reconstructed. Like the radius of GPDM 205, the radius of GPDM 178 is dorsoventrally elongate, with a transversely widened distal region. More akin to that of Kentrosaurus aethiopicus (Galton 1982b), then the dorsoventrally short and rectangular radius of Stegosaurus stenops (NHMUK PV R36730, Maidment et al. 2015).
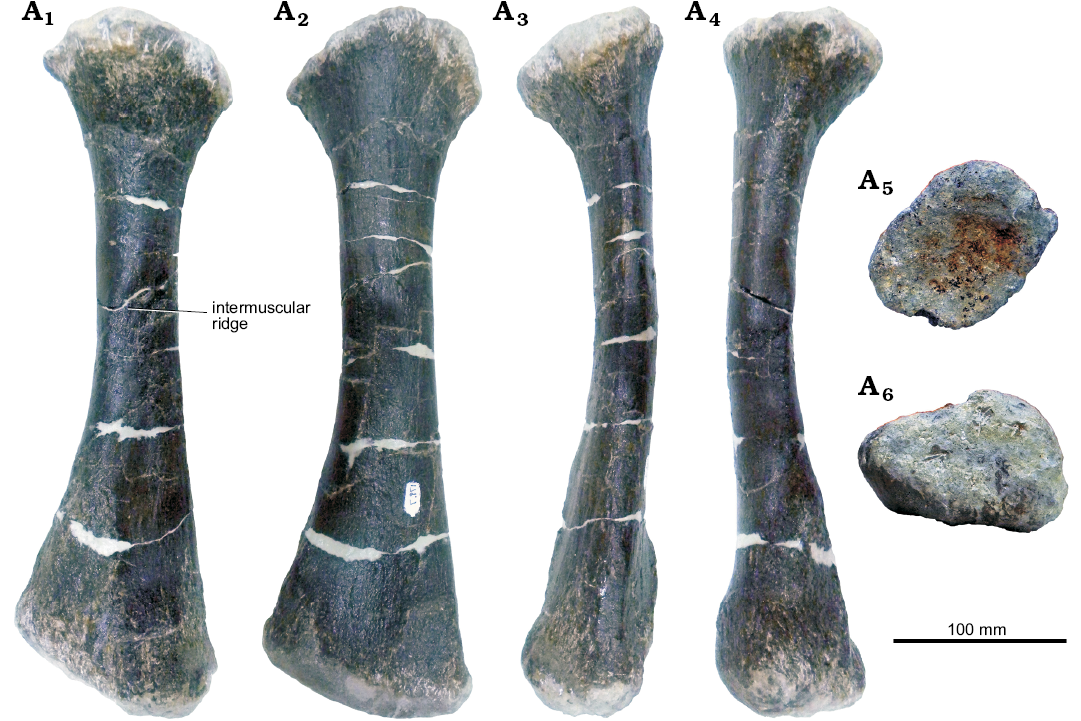
Fig. 17. Left radius of stegosaur GPDM 178, near Giffen, Cascade County, Montana; in anterior (A1), posterior (A2), medial (A3), lateral (A4), dorsal (A5), and ventral (A6) views.
Tibia: The right tibia of GPDM 178 is represented by the distal and proximal ends (Fig. 18). The proximal end was the first element recovered from the specimen (see the Material section above). While there was no diaphysis recovered, nor do we know approximately where the distal section was found in relation, given the similar states of preservation it is most parsimonious to assume these ends are parts of the same element.
In proximal view, the proximal end is broadly oval, being anteroposteriorly longer than it is transversely wide. The medial surface is convex, while the lateral surface bears a strongly laterally projecting process for articulation of the fibula. The fibula process is very well-developed and is rounded in outline in lateral and anterior view. A very well-developed fibula process is also present in Hesperosaurus mjosi (MOR 9728; Maidment et al. 2018). The cnemial crest is poorly preserved. The distal end is heart-shaped in distal view, with the apex pointing posteriorly. In anterior view, the distal end is separated into two distinct surfaces, the lateral malleolus and the medial malleolus, separated from each other by a groove. The anterior surfaces of the malleoli are flattened; the lateral malleolus is somewhat posteriorly offset by postmortem deformation. The fibula articulated with the flattened surface of the lateral malleolus. The lateral malleolus projects further ventrally than the medial malleolus, onto which the astragalus would have articulated distally. In posterior view, a vertical ridge extends dorsally along up the midline of the bone.
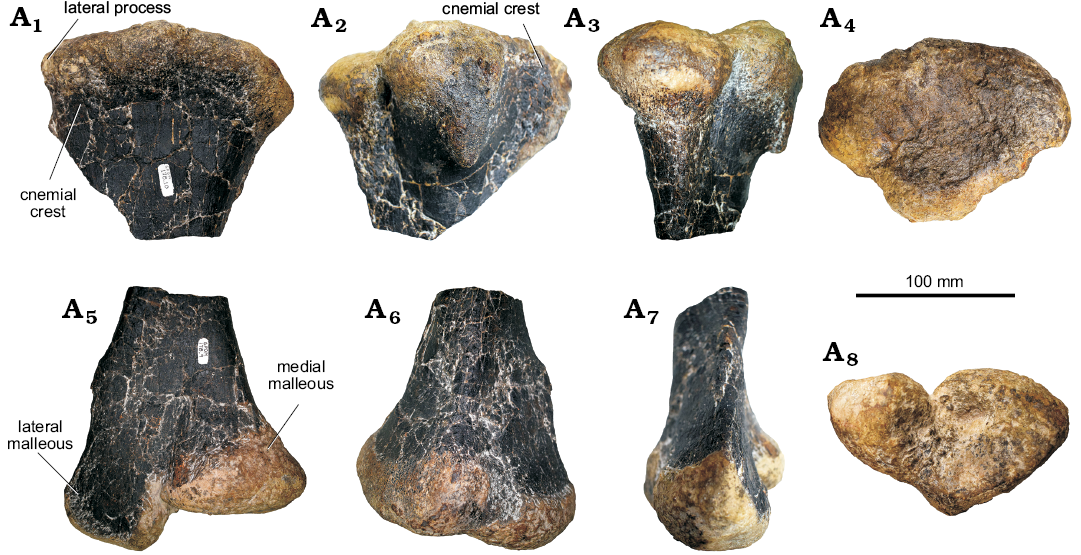
Fig. 18. Partial right tibia of stegosaur GPDM 178, near Giffen, Cascade County, Montana; in medial (A1, A6), lateral (A2, A7), anterior (A3, A5), dorsal (A4), and ventral (A8) views. A1–A4, proximal end, A5–A8, distal end.
Dermal armor: While four plates are recorded in the quarry log for GPDM 178 (three are identified as cervical plates and one as a dorsal plate), only one, a dorsal plate, is at this time fully prepared and available for study.
Dorsal plate: In notes and drawings made during excavation, this plate is located near the pelvic girdle, suggesting that it may be a large dorsal plate situated over the pelvis, by comparison with Stegosaurus stenops (NHMUK PV R36730; Maidment et al. 2015). This incomplete plate, superficially triangular in shape, has a gentle convex ventral margin (Fig. 19). The entire ventral margin of the dorsal plate of GPDM 178 is transversely widened (86.9 mm). Unlike other stegosaur plates that dorsally or anteroposteriorly taper towards the margins (as Saitta 2015 showed via CT scans), aside from the transversely widened ventral margin, the rest of the plate is a fairly uniform thickness of nearly 2 cm. This rather uniform plate thickness coupled with the widened ventral margin gives the plate more of an inverted T-shaped cross-sectional profile; again, differing from the usually observed dorsally tapering profile. The non-tapering aspect of the plate could be attributed to taphomonic processes where the thinning margins were damaged. Likewise, due to damage it is not possible to ascertain if the plate possessed the anteriormost “stepped” profile seen in plates 13–15 in Stegosaurus stenops (NHMUK PV R36730; Maidment et al. 2015). Given the plate size and the convex ventral margin, assuming similar plate homology corresponds serially within stegosaurs, we can only hypothesize that the dorsal plate of GPDM 178 corresponds to plate 13–16; potentially serially placing it more posteriorly than previously believed.

Fig. 19. Dorsal plate of stegosaur GPDM 178, near Giffen, Cascade County, Montana.
Discussion
Taxonomy.—Within the Morrison Formation there are three known stegosaur genera: Alcovasaurus, Hesperosaurus, and Stegosaurus.
According to Galton and Carpenter (2016) Alcovasaurus bears the following autapomorphies: (1) all caudal vertebrae bear transverse processes, (2) distal caudal centra short, so height greater than length, (3) femoral condylar articular surface confined almost exclusively to the distal surface, (4) two pairs of greatly elongate distal dermal tail spines (~90% of femoral length) with subequal bases and slender diaphysis, (5) posterior pair widest at ~25% of length. It is further distinguishable from other Morrison Formation stegosaurs by (6) having six pairs of sacral ribs.
Maidment et al. (2018) diagnosed Hesperosaurus using the following combination of autapomorphies (marked by an *) and plesiomorphic characters: (1) no contact between the lacrimal and prefrontal on the lateral surface of the skull, (2) ventral margin of axis upwardly concave in lateral view*, (3) postzygapophyses not elevated significantly on posterior cervical vertebrae, (4) prezygapophyses join ventrally and face dorsomedially in anterior dorsal vertebrae, (5) dorsal neural arch pedicels not elongated above the neural canal in mid-dorsal vertebrae, (6) dorsal ribs distally expanded, (7) ossified epaxial tendons present, (8) caudal neural spines not bifurcated, (9) dorsal dermal plates longer anteroposteriorly than tall dorsoventrally*, (10) 11 dorsal vertebrae*, and (11) fourth sacral vertebra not fused to the sacrum*.
Finally, Stegosaurus stenops can be diagnosed by the following autapomorphies (Maidment et al. 2015): (1) portion of the dentary anterior to the tooth row and posterior to the predentary edentulous; (2) dorsally elevated postzygapophyses of cervical vertebrae; (3) bifurcated summits of neural spines of the anterior and middle caudal vertebrae; (4) presence of dermal ossicles embedded in the skin on the underside of the cervical region.
As GPDM 205 is only represented by a prepared humerus, ulna, and radius, and no Alcovasaurus, Hesperosaurus, or Stegosaurus forelimb autapomorphies exist, we can only tentatively refer to it as Stegosauria indeterminate.
None of the autapomorphy-bearing elements of Alcovasaurus are preserved in GPDM 178, and thus we are unable to compare the specimen to that taxon. In comparison to Hesperosaurus, in GPDM 178 it is only possible to assess characters (3) and (9) of Maidment et al. (2018) based on the available material. Character (9) cannot be assessed because the prepared dorsal plate is incomplete, so its profile is unclear. Finally, regarding Stegosaurus stenops, only characters (1) and (2) can be assessed based on the available material. Character (1) is present in GPDM 178. However, it is not a robust autapomorphy of Stegosaurus because a diastema between the predentary facet and dentary tooth row is also known in Jiangjunosaurus (Jia et al. 2007). Additionally, because the dentary is unknown in Hesperosaurus (Carpenter et al. 2001; Maidment et al. 2018) and Alcovasaurus (Galton and Carpenter 2016), it is difficult to assess the importance of the feature for Morrison Formation stegosaurs. Character (3) from the Hesperosaurus diagnosis and character (2) from the Stegosaurus diagnosis serve to differentiate the two taxa. GPDM 178 has elevated postzygapophyses, an autapomorphy of Stegosaurus, but comparisons with MOR 9728 (Hesperosaurus; Maidment et al. 2018) indicate little difference in the character states between the two, suggesting this character needs reassessment in the light of new discoveries. Due to the lack of robust autapomorphies of any genus of stegosaur in GPDM 178, we consider the specimen as Stegosauria indeterminate.
It is clear that the fragmentary nature of many stegosaur specimens and the paucity of material for most genera means that the taxonomy of stegosaurs continues to be problematic, and each new discovery leads to re-diagnosis. Only by the discovery and description of new specimens will we be able to clearly understand the diversity of Stegosauria.
Ontogeny.—Though GPDM 178 and 205 have not yet been histologically sampled to document their maturational states, their relative sizes contribute to the ongoing discussion of latitudinal body size amongst Morrison Formation stegosaurs proposed by Maidment et al. (2018). In comparing humeral length, GPDM 178 and 205 appear to fall just within the upper half of humeral lengths (Fig. 20), yet, as shown by Maidment et al. (2018), humeral length does not adequately correlate to maturational age in stegosaurs. The smallest and largest Hesperosaurus mjosi humeri (MOR 9728, 390 mm; SMA0018, 545 mm) are also the histologically most mature specimens, indicating skeletal plasticity in this genus. Histologically aging GPDM 178 and 205 will help to determine if these specimens contribute to our understanding of body size versus maturity in stegosaurs (i.e., does the relative size and morphology of these specimens denote maturity, or morphologies consistent with the biomechanics of their overall size).
Additionally, Maidment et al. (2018) proposed that body size difference between Hesperosaurus and Stegosaurus might relate to ecologic partitioning where the larger Stegosaurus occupied the more arid environments, while the smaller Hesperosaurus occupied the wetter, more vegetated environments. Unfortunately, with the current available data, the relative size of these two specimens does not aide in elucidating taxomony (both GPDM 178 and 205 could be “larger” Hesperosaurus, or “mid-sized” Stegosaurus). However, it would be very interesting if these specimens, particularly GPDM 205, were Stegosaurus. Given the environmental partitioning theory of Maidment et al. (2018) and the high northern latitude of these specimens (even in the Late Jurassic), if they were Stegosaurus, this could indicate the change to a more arid climate following the northward regression of the Sundance Seaway.
Though histologic analysis of either GPDM 178 or 205 has yet to occur, some postcranial morphologies could one day substantiate generic level maturational states. In a review of immature Stegosaurus sp. remains from Utah and Wyoming, Galton (1982a) noted 14 postcranial morphologies attributable to ontogenetic development. While none of these “ontogenetic indicators” of Galton (1982a) have been histologically tested, we subscribe that many have the potential to be legitimate. Among Galton’s (1982a) “ontogenetic indicators”, some are fusion (open vertebral synostosis, lack of a fused scapula-coracoid; Galton 1982a) or proportion (slender and elongate scapulae, more gracile metacarpals; Galton 1982a) related, and four are directly related to long bone soft-tissue scarring (Galton 1982a: characters 4 and 11c) and proportion of long bone processes (Galton 1982a: characters 5, 6, 11b). Galton (1982a) noted that in immature animals, the surface of the long bones are relatively smooth, compared to the highly rugose and irregular articular end in mature animals. Likewise, in immature animals the triceps ridge and the olecranon process are much less prominent than their mature counterparts (Galton 1982a). Galton (1982a) noted that since of these traits were present within other taxa (Dacentrurus and Kentrosaurus), they were likely general stegosaur growth patterns.
In comparing GPDM 178 and 205, the deltopectoral crest on the humerus of GPDM 205 is greatly enlarged and robust compared to that of GPDM 178. An enlarged deltopectoral would correspond to the observations of Galton (1982a). In discussing the enlarged olecranon process of Miragaia longicollum, therein referred to as the “olecranon horn”, a similar consensus was reached by Mateus et al. (2009) in suggesting that the growth of this process developed throughout ontogeny. These ridges, processes, and condyles of the limb elements are all associated with connective tissues, muscles, ligaments, tendons, and cartilage. Schwarz et al. (2007) noted calcified cartilage along the distal margin of the humerus from the sauropod Cetiosauriscus greppini, while Wilson et al. (2016) noted the biomineralization of the vertebral ligaments throughout body size and ontogeny in theropods, and Cerda et al. (2015) likewise documented the occurrence of biomineralized vertebral ligaments and their possible ontogenetic development in nine sauropods. Thus, biomineralization of connective tissues, and their development throughout ontogeny, is not a dinosaurian precedent. We must reiterate that a histologic analysis is the only way to substantiate such, but in agreement with Galton (1982a), perhaps in stegosaurs, as the individuals increase in age and body mass, the cartilaginous covered ends of the ulnae (the “olecranon horn” of Mateus et al. 2009) and the ligamentous/tendinous attachment sites along the deltopectoral crests biomineralize (through metaplasia; sensu Horner et al. 2016) through ontogeny. If true, then we could hypothesize that GPDM 178 is less mature than GPDM 205. While morphology would seem to substantiate such development, currently this is a speculative hypothesis, and we await a histological analysis to test for such.
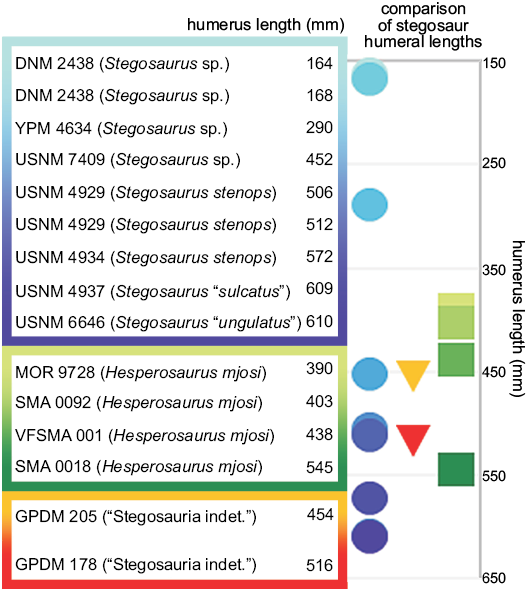
Fig. 20. Stegosaur humeral lengths (measurement from Gilmore 1914, Galton 1982a, and Maidment et al. 2018). The grading colors represent the transitions from the smallest to largest specimens. Circles, Stegosaurus; triangles, Stegosauria indeterminate; squares, Hesperosaurus.
Tooth wear.—The high prevalence of macro- and microwear on the teeth of GPDM 178 would seem to indicate that (i) this animal was possibly feeding on coarser vegetation (the possibility of a mixed vegetation diet was also proposed by Lautenschlager et al. 2016), and/or (ii) these combinations of extensive tooth wear and cross-cutting micro features may indicate a complex chewing motion. Although Lautenschlager et al. (2016), via cranial biomechanics, suggested that complex chewing in stegosaurs was unlikely, Nabavizadeh (2016) and Nabavizadeh and Weishampel (2016) discussed and reviewed stegosaur feeding motions. Possessing a cropping rhampotheca, the orthal and minor long-axis rotation of the dentaries would likely produce an initial orthal jaw motion possibly followed by a minor palinal motion (i.e., the jaws vertically come together, followed by a posterior motion while in occlusion; Barrett 2001; Galton and Upchurch 2004; Nabavizadeh 2016; Nabavizadeh and Weishampel 2016).
The extensive wear facets of GPDM 178 would seem to indicate in part, a more complex chewing motion or jaw mechanics than previously assessed. Both Barrett (2001) and Nabavizadeh (2016) note that in Stegosaurus, the equal rostrocaudal lengths in the glenoid fossa of the jaw joint and the ventral condyles of the quadrate would primarily produce an orthal jaw motion. Additionally, with a mesially focused bite force, the relative bite force (Nabavizadeh 2016) of Stegosaurus stenops (USNM 4934) is low compared to that of hadrosaurs and ceratopsians (0.6471 vs. 1.5595 and 2.7, respectively; Nabavizadeh 2016). This lower relative bite force could in part explain the “typical” lack of prominent wear facets observed in stegosaurs. While missing the portions of the jaw joint, the sub-horizontal wear facets of GPDM 178 would seem to corroborate a primarily orthal occlusion (sensu Barrett 2001 and Nabavizadeh 2016). The “flatness” could also have been formed in part by a secondary palinal motion. This is noteworthy because Nabavizadeh and Weishampel (2016) proposed that stegosaurs may have incorporated a palinal jaw motion when chewing; however, they stated that dental microwear would be needed for verification. The horizontal facets of GPDM 178 appear to verify the jaw motion hypothesis of Nabavizadeh and Weishampel (2016).
Additionally, the micro-gouges within the wear facets offers added jaw-motion insight. Following cross-cutting relationships, we hypothesize that in GPDM 178 the maxillary and dentary teeth orthally occluded, followed by a palinal occlusal motion indicated by the ~anteroposterior gouges. Finally, the cross-cutting, laterally oblique micro gouges could have been produced from the rotation of the dentaries; which we presume would have been caused by the forward rotational movement of the dentaries. Although we do not know the role of vegetation types regarding stegosaur wear facets, such prominent facets in GPDM 178 would indicate that extensive chewing was required when this animal was feeding.
Conclusions
The specimens GPDM 178 and 205 represent the northernmost stegosaurs found to-date, and GPDM 178 is the northernmost dinosaur fossil thus far recovered from the entire Morrison Formation. Interestingly, according to a Works Progress Administration report from 1938, a “Stegosaurus” was found at Sheep Creek, 25 miles north of Great Falls, MT (Syverud 1938). If true, then this specimen is the northernmost Morrison Formation dinosaur, as GPDM 178 was found near equivalent distance southeast of Great Falls, MT. Unfortunately, this report does not mention what material was collected or where it was sent. Searching for this material in Montana institutions with historic paleontological collections and the USNM has yielded no results. Therefore we are left to believe that these remains, if true, have subsequently been lost or discarded. Thus by default, GPDM 178 shall remain the northernmost record.
These two new specimens, consisting of rare cranial as well as post-cranial material, increase the number of Morrison Formation dinosaur specimens recovered from Montana. Saitta (2015) reported five specimens from the JRDI 5ES Quarry, the same locality as GPDM 205. These specimens have always resided in a private collection, and are thus unavailable to study, despite the declaration made by Saitta (2015) to the contrary. One stegosaur (unknown genus) is being collected by the Bighorn Basin Paleontological Institute in Carbon County, Montana, and another stegosaur specimen is awaiting excavation in the vicinity of Lewistown, Montana (DCW personal observations). Finally, combining MOR 9728 and two other fragmentary specimens from a single locality in Livingston, Montana, this brings the number of stegosaur specimens from Montana to 12. The Paleobiology Database (www.paleobiodb.org) lists 72 occurrences of stegosaurs from North America (67 Stegosaurus, four Hesperosaurus, and one Alcovasaurus) meaning that the Montana specimens account for 16% of the known Morrison Formation record. However, only published fossil remains are recorded in the Paleobiology Database, and it is likely that further unpublished specimens reside in museum collections across the western USA. Nonetheless, considering the relative paucity of research on the Morrison Formation in Montana in comparison to better-known areas in Colorado and Utah, the fossil record of stegosaurs in Montana indicates that these thyreophorans were a common faunal component in the north of the Morrison basin.
Acknowledgements
First and foremost, we would like to thank all of the board members of the Judith River Foundation for recognizing the significance of GPDM 178 and GPDM 205 and allowing us the opportunity to share these specimens with the scientific community. Thanks to the Hein and Yurek families of Grass Range and Great Falls, MT for donating these specimens to the Great Plains Dinosaur Museum and Field Station. ReBecca Hunt-Foster (Dinosaur National Monument, Vernal, Utah, USA) and Ali Nabavizadeh (Rowan University, Glassboro, New Jersey, USA) provided invaluable comments and suggestions that greatly improved this manuscript. Jack Wilson (Badlands Dinosaur Museum, Dickinson, North Dakota, USA) provided many of the photographs of GPDM 178 used in this manuscript. Cory Coverdell (Two Medicine Dinosaur Center, Bynum, Montana, USA), Jack Wilson, and the Two Medicine Dinosaur Center prepared many of the GPDM 178 elements examined herein. Genya Masukawa (Ibaraki University, Mito, Japan) graciously provided the skeletal reconstructions of Hesperosaurus mjosi used in Fig. 2. Thanks to the Hein family for allowing DCW to visit the GPDM 205 locality in 2016 and for personal accounts. Dixie Stordahl (Malta, Montana, USA), Sue Frary (Helena, Montana, USA), and Linda Yurek (Great Falls, Montana, USA) provided invaluable firsthand accounts on the history and excavation of GPDM 178.
References
Barrett, P.M. 2001. Tooth wear and possible jaw action of Scelidosaurus harrisonii Owen and a review of feeding mechanisms in other thyreophoran dinosaurs. In: K. Carpenter (ed.), The Armored Dinosaurs, 25–52. Indiana University Press, Bloomington.
Berman, D.S. and McIntosh, J.S. 1986. Description of the lower jaw of Stegosaurus. Annals of the Carnegie Museum 55: 29–40.
Carpenter, K., Miles, C.A., and Cloward, K. 2001. New primitive stegosaur from the Morrison Formation, Wyoming. In: K. Carpenter (ed.), The Armored Dinosaurs, 55–75. Indiana University Press, Bloomington.
Cerda, I.A., Casal, G.A., Martinez, R.D., and Ibiricu, L.M. 2015. Histological evidence for a supraspinous ligament in sauropod dinosaurs. Royal Society Open Science 2: 150369. Crossref
Chen, A. 2015. Dino “sexing” study slammed by critics. Science [published online, https://doi.org/10.1126/science.aab2534] Crossref
Erickson, G.M. 1996. Incremental lines of von Ebner in dinosaurs and the assessment of tooth replacement rates using growth line counts. Proceedings of the National Academy of Sciences 93: 14623–14627. Crossref
Foster, J.R. 2007. Jurassic West. The Dinosaurs of the Morrison Formation and Their World. 389 pp. Indiana University Press, Bloomington.
Galton, P.M. 1982a. Juveniles of the stegosaurian dinosaur Stegosaurus from the Upper Jurassic of North America. Journal of Vertebrate Paleontology 2: 47–62. Crossref
Galton, P.M. 1982b. The postcranial anatomy of stegosaurian dinosaur Kentrosaurus from the Upper Jurassic of Tanzania, East Africa. Geologica et Palaeontologica 15: 139–160.
Galton, P.M. and Carpenter, K. 2016. The plated dinosaur Stegosaurus longispinus Gilmore, 1914 (Dinosauria: Ornithischia; Upper Jurassic, western USA), type species of Alcovasaurus n. gen. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 279: 185–208. Crossref
Galton, P.M. and Upchurch, P. 2004. Stegosauria. In: P. Dodson, H. Osmólska, and D.B. Weishampel (eds.), The Dinosauria, 343–362. University of California Press, Berkeley. Crossref
Gilmore, C.W. 1914. Osteology of the armored Dinosauria in the United States National Museum, with special reference to the genus Stegosaurus. Bulletin of United States National Museum 89: 1–244. Crossref
Hao, B., Zhang, Q., Peng, G., Ye, Y., and You, H. 2018. Redescription of Gigantspinosaurus sichuanensis (Dinosauria, Stegosauria) from the Late Jurassic of Sichuan, Southwestern China. Acta Geologica Sinica, English Edition 92: 431–441. Crossref
Hennig, E. 1925. Kentrurosaurus aethiopicus; die Stegosaurierfunde vom Tendaguru, Deutsch-Ostafrika. Palaeontographica-Supplementbände (Supplement 7): 101–254.
Horner, J.R., Woodward, H.N., and Bailleul, A.M. 2016. Mineralized tissues in dinosaurs interpreted as having formed through metaplasia: A preliminary evaluation. Comptes Rendus Palevol 15: 176–196. Crossref
Jia, C.-K., Foster, C.A., Xu, X., and Clarke J.M. 2007. The first stegosaur (Dinosauria, Ornithischia) from the Upper Jurassic Shishugou Formation of Xinjiang, China. Acta Geologica Sinica 81: 351–356. Crossref
Lautenschlager, S., Brassey, C.A., Button, D.J., and Barrett, P.M. 2016. Decoupled form and function in disparate herbivorous dinosaur clades. Scientific Reports 6: 26495. Crossref
Maidment, S.C.R., Brassey, C., and Barrett, P.M. 2015. The postcranial skeleton of an exceptionally complete individual of the plated dinosaur Stegosaurus stenops (Dinosauria: Thyreophora) from the Upper Jurassic Morrison Formation of Wyoming, USA. PloS ONE 10: e0138352. Crossref
Maidment, S.C.R, Norman, D.B., Barrett, P.M., and Upchurch, P. 2008. Systematics and phylogeny of Stegosauria (Dinosauria: Ornithischia). Journal of Systematic Palaeontolgy 6: 364–407. Crossref
Maidment, S.C.R., Woodruff, D.C., and Horner, J.R. 2018. A new specimen of the ornithischian dinosaur Hesperosaurus mjosi from the Upper Jurassic Morrison Formation of Montana, USA, and implications for growth and size in Morrison stegosaurs. Journal of Vertebrate Paleontology 38: e1406366. Crossref
Mallison, H. 2010. CAD assessment of the posture and range of motion of Kentrosaurus aethiopicus Hennig 1915. Swiss Journal of Geosciences 103: 211–233. Crossref
Marsh, O.C. 1877. A new order of extinct Reptilia (Stegosauria) from the Jurassic of the Rocky Mountains. American Journal of Science 84: 513–514. Crossref
Mateus, O., Maidment, S.C.R., and Christiansen, N.A. 2009. A new long-necked “sauropod-mimic” stegosaur and the evolution of the plated dinosaurs. Proceedings of the Royal Society of London Biological Sciences 276: 1815–1821. Crossref
Nabavizadeh, A. 2016. Evolutionary trends in the jaw adductor mechanics of ornithischian dinosaurs. The Anatomical Record 299: 271–294. Crossref
Nabavizadeh, A. and Weishampel, D.B. 2016. The predentary bone and its significance in the evolution of feeding mechanisms in ornithischian dinosaurs. The Anatomical Record 299: 1358–1388. Crossref
Ouyang, H. 1992. Discovery of Gigantspinosaurus sichuanensis and its scapular spine orientation [in Chinese]. In: Youth Academic Symposium on New Discoveries and Ideas in Stratigraphic Palaeontology, Nanjing, December 1992, Abstracts and Summaries, 47–49. Chinese Science Association.
Owen, R. 1842. Report on British Fossil Reptiles. Part II. Report of the Eleventh Meeting of the British Association for the Advancement of Science; Held at Plymouth in July 1841, 60–204. John Murray, London.
Raven, T.J. and Maidment, S.C.R. 2017. A new phylogeny of Stegosauria (Dinosauria, Ornithischia). Palaeontology 60: 401–408. Crossref
Reichel, M. 2010. A model for the bite mechanics in the herbivorous dinosaur Stegosaurus (Ornithischia, Stegosauridae). Swiss Journal of Geosciences 103: 235–240. Crossref
Saitta, E.T. 2015. Evidence for sexual dimorphism in the plated dinosaur Stegosaurus mjosi (Ornithischia, Stegosauria) from the Morrison Formation (Upper Jurassic) of western USA. PloS ONE 10: e0123503. Crossref
Salgado, L., Canudo, J.I., Garrido, A.C., Moreno-Azanza, M., Martínez, L.C., Coria, R.A., and Gasca, J.M. 2017. A new primitive Neornithischian dinosaur from the Jurassic of Patagonia with gut contents. Scientific Reports 7: 42778. Crossref
Schwarz, D., Wings, O., and Meyer, C.A. 2007. Super sizing the giants: first cartilage preservation at a sauropod dinosaur limb joint. Journal of the Geological Society 164: 61–65. Crossref
Sereno, P.C. 1991. Lesothosaurus, “fabrosaurids,” and the early evolution of Ornithischia. Journal of Vertebrate Paleontology 11: 168–197. Crossref
Sereno, P.C. and Dong, Z.-H. 1992. The skull of the basal stegosaur Huayangosaurus taibaii and a cladistic diagnosis of Stegosauria. Journal of Vertebrate Paleontology 12: 318–343. Crossref
Seeley, H.G. 1887. On the classification of the fossil animals commonly named Dinosauria. Proceedings of the Royal Society of London 43: 165–171. Crossref
Syverud, E.I. 1938. Fossil Research in Eastern Montana on Pioneer Days Too (Pertinent to Recent Excavations near Fort Peck Dam). 4 pp. Federal Writers’ Project Works Progress Administration of Montana.
Tumanova, T.A. and Alifanov, V.R. 2018. First record of stegosaur (Ornithischia, Dinosauria) from the Aptian–Albian of Mongolia. Paleontological Journal 52: 1771–1779. Crossref
Whitlock, J.A., Trujillo, K.C., and Hanik, G.M. 2018. Assemblage-level structure in Morrison Formation dinosaurs, Western Interior, USA. Geology of the Intermountain West 5: 9–22. Crossref
Wilson, J.P., Woodruff, D.C., Gardner, J.D., Flora, H.M., Horner, J.R., and Organ, C.L. 2016. Vertebral adaptations to large body size in theropod dinosaurs. PloS ONE 11: e0158962.Crossref
Woodruff, D.C. and Foster, J.R. 2017. The first specimen of Camarasaurus (Dinosauria: Sauropoda) from Montana: The northernmost occurrence of the genus. PloS ONE 12: e0177423. Crossref
Acta Palaeontol. Pol. 64 (3): 461–480, 2019
https://doi.org/10.4202/app.00585.2018