
A new beaked whale record from the upper Miocene of Menorca, Balearic Islands, based on CT-scan analysis of limestone slabs
GIOVANNI BIANUCCI, SERGIO LLÀCER, JOSEP QUINTANA CARDONA, ALBERTO COLLARETA, and AGUSTÍ RODRÍGUEZ FLORIT
Bianucci, G., Llàcer, S., Cardona, J.Q., Collareta, A., and Florit, A.R. 2019. A new beaked whale record from the upper Miocene of Menorca, Balearic Islands, based on CT-scan analysis of limestone slabs. Acta Palaeontologica Polonica 64 (2): 291–302.
The finding of significant vertebrate remains inside commercial stone blocks is relatively rare. Here we describe a fossil cetacean skull discovered inside two slabs cut from a limestone block of Tortonian (i.e., early late Miocene) age from Menorca (Balearic Islands, Spain); this find represents the third record of a fossil cetacean from this island, as well as the best preserved. Unlike similar cases in the past, the Menorca skull was not extracted mechanically from the stone matrix, being rather imaged by means of computed tomography. The resulting 3D reconstruction of the skull allows us to refer the studied skull to the extant odontocete family Ziphiidae (beaked whales) and sheds light on the morphology of very delicate structures (e.g., the thin pterygoid hamuli and the mesorostral cartilage) that would likely have been destroyed during traditional mechanical preparation. This non-invasive investigation permits detailed osteo-anatomical comparisons between the Menorca specimen and other extinct ziphiids, leading to the referral of the former to the stem beaked whale Messapicetus cf. longirostris. We then review the geographic distribution of fossil remains of Messapicetus, which include other occurrences from Tortonian shelf deposits of southeastern Italy, southern Peru, and Maryland (eastern USA). Early branching beaked whales (including basal members of the so-called “Messapicetus clade”) likely dispersed via the Central American Seaway, which allowed a direct communication between the Pacific and Atlantic oceans (including the Mediterranean cul-de-sac) throughout the Miocene.
Key words: Mammalia, Cetacea, Ziphiidae, Messapicetus, palaeobiogeography, Neogene, Mediterranean, Spain.
Giovanni Bianucci [giovanni.bianucci@unipi.it], Dipartimento di Scienze della Terra, Università di Pisa, via Santa Maria 53, 56126, Pisa, Italy.
Sergio Llàcer [sergio.llacer@icp.cat], Institut Català de Paleontologia Miquel Crusafont, Universitat Autònoma de Barcelona, Carrer de les Columnes s/n, 08193 Cerdanyola del Vallès, Barcelona, Spain.
Josep Quintana Cardona [picoguevo@gmail.com], Institut Català de Paleontologia Miquel Crusafont, Universitat Autònoma de Barcelona, Carrer de les Columnes s/n, 08193 Cerdanyola del Vallès, Barcelona, Spain; Carrer Gustau Mas 79, 1er, 07760 Ciutadella de Menorca, Illes Balears, Spain.
Alberto Collareta [alberto.collareta@unipi.it], Dipartimento di Scienze della Terra, Università di Pisa, via Santa Maria 53, 56126, Pisa, Italy.
Agustí Rodrígez Florit [a.rodriguez@geoservei.com], carrer Himàlaia 37, 07703 Maó, Illes Balears, Spain.
Received 9 January 2019, accepted 12 February 2019, available online 23 April 2019.
Copyright © 2019 G. Bianucci et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Fossils are rather frequently encountered inside stone blocks cut for commercial use. Indeed, lithotypes such as the red ammonitic limestone or the Orthoceras limestone feature abundant fossil shells that often represent significant and appreciated decorative components of the rock itself (e.g., Robinson and Bishop 1980; Wilson 1983; Hannibal and Davis 1992; Hannibal et al. 2008). Nevertheless, the finding of significant remains of vertebrates inside commercial stone blocks is relatively rare, the few well-documented cases including the Middle Jurassic crocodylian Neptunidraco ammoniticus (Crocodylia, Metriorhynchidae) discovered in two slabs of red ammonitic limestone from northeaster Italy (Cau and Fanti 2011) and the impressive record of marine vertebrate remains found in stone blocks and slabs of the Miocene limestone “Pietra leccese” from southern Italy (e.g., Bianucci et al. 1994a, 2004; Bianucci 1996, 2001). In some cases, the study of these fossils was preceded by a long and complex procedure for preparation, in a way to free the various parts of the bones from the embedding rock before reassembling the pieces preserved within different blocks or slabs. For example, the holotype of Zygophyseter varolai (Cetacea, Physeteroidea), an almost complete skeleton, was reassembled from several large blocks of Pietra leccese after more than five years of preparation work (Bianucci and Landini 2006). Similarly, the partial skeleton of Aegyptocetus tarfa (Cetacea, Protocetidae) required preparation for one year to reassemble bone pieces from six slabs cut from a large block of middle Eocene nummulitic limestone from Egypt (Bianucci and Gingerich 2011).
Here we describe a late Miocene fossil cetacean skull discovered inside two slabs cut from a commercial limestone block extracted from the Son Salord quarry close to Ciutadella de Menorca (Menorca, Balearic Islands, Spain) and now kept in the Museu Diocesà de Menorca (Figs. 1, 2). This specimen represents the third record of a fossil cetacean from this island, as well as the best-preserved one. In fact, cetacean remains from Menorca were previously documented only on the basis of two isolated teeth: (i) one tooth referred to “Scaldicetus grandis” from the Miocene of Illa de l’Aire, at the southeastern end of Menorca (Mercadal et al. 1985); and (ii) one tooth referred to an indeterminate delphinid from the lower Tortonian of Torre d’en Penjat, at the southeastern coast of Es Castell (Mas et al. 2013).
The fossil cetacean from the Son Salord quarry was not extracted mechanically from the stone matrix; instead, it was imaged by means of computed tomography. The three-dimensional reconstruction of the skull obtained after processing of the scans proved to be extremely detailed thanks to the good density contrast between the bone and surrounding matrix. In this way, it was possible to detail the morphology of very delicate structures such as the thin pterygoid hamuli and the mesorostral cartilage, which would probably have been destroyed during traditional mechanical preparation. This kind of non-invasive investigation allowed us to refer the Menorca skull to a beaked whale, i.e., a member of the odontocete family Ziphiidae, whose fossil record is now well documented (Bianucci et al. 2016c and references therein). In particular, the skull described here is assigned to the genus Messapicetus, a stem ziphiid originally described from the Tortonian of the Pietra leccese of southern Italy (Bianucci et al. 1992, 1994b, 2016a), then tentatively recognised from the upper Miocene of the St. Marys Formation of Calvert Cliffs, Maryland, USA (Fuller and Godfrey 2007), and later recorded with several specimens from upper Miocene beds of the Pisco Formation, Peru (Bianucci et al. 2010, 2016b; Lambert et al. 2015; Di Celma et al. 2017; Ramassamy et al. 2018).
Institutional abbreviations.—ICP, Institut Català de Paleontología Miquel Crusafont, Sabadell, Barcelona, Spain; MDM, Museu Diocesà de Menorca, Ciutadella de Menorca, Spain.
Geological setting
MDM-2029 was found in an area located about 2.5 km East of Ciutadella de Menorca (Fig. 1A, B), where several limestone quarries are located. The rock that is produced in these quarries, locally known as “Mares”, is widely used for building purposes and is characterised by light colour, very porous texture and a soft consistency (Sauleau 1985). In particular, MDM-2029 comes from the still-active quarry of Son Salord (Fig. 1C), whose vertical walls display a rather massive and homogeneous appearance, with more or less differentiated and bioturbated beds. Macrofossils found at this quarry include scarce rhodolits and fragments of red calcareous algae, pectinid bivalves and irregular echinoids (belonging to the genera Clypeaster, Hypsoclypus, and Schizaster, among others). According to official geological cartography, the carbonate deposits exposed at the Son Salord quarry are Tortonian (i.e., early late Miocene) in age (Rosell et al. 1989). More recently, Pomar (2001), Pomar et al. (2002) and Mateu-Vicens et al. (2008) placed these sediments in their Lower Bar Unit, which is included in the lower Tortonian N16 zone of the planktonic foraminiferal zonation of Blow (1969). The ziphiid skull was found at a depth of 14 m below ground level, i.e., roughly at the level of the floor of the quarry as shown in Fig. 1C. It is unknown if it appeared in association with other skeletal elements belonging to the same individual. It has not been possible to locate all the blocks in which other parts of the skull were preserved, because they were sold as construction pieces before the specimen here described was recognised as a fossil vertebrate (Cristóbal Torres, personal communication 2018).
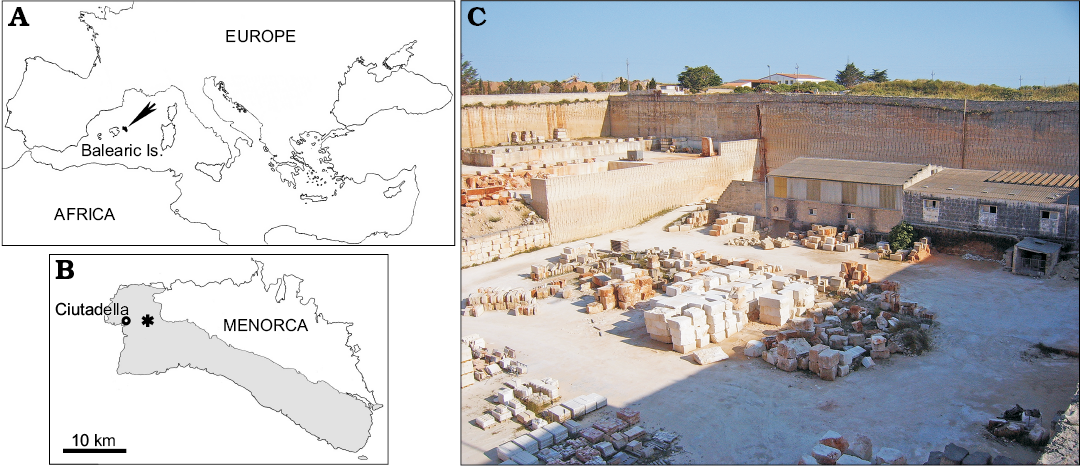
Fig. 1. A. Geographical location of Menorca in the Mediterranean Sea (black arrow). B. Close-up of Menorca showing location of the Son Salord quarry (asterisk), where the Messapicetus MDM-2029 was found. The grey area indicates the extent of the upper Miocene (Tortonian–Messinian) carbonate deposits cropping out on the island. C. General view of the Son Salord quarry. The floor of the quarry roughly corresponds to the stratigraphic level where the fossil beaked whale was found.
Material and methods
The fossil specimen studied herein is an incomplete cetacean skull preserved within two limestone blocks of similar size (ca. 400×450×140 mm) (Fig. 2). Although the two limestone blocks are contiguous, there is a hiatus of ca. 11 mm between them, corresponding to the volume of rock lost during cutting. CT segmentation of the skull was performed at Hospital Mateu Orfila (Mahón, Menorca) by using a medical scanner (Philips Brilliance 16 CT scanner). The analytical conditions of the CT-scan analysis are shown in Table 1.
Table 1. Analytical conditions of the CT-scan analysis.
| |
Slab 1 (top slab) |
Slab 2 (bottom slab) |
|
Pixel size |
0.543 mm |
0.621 mm |
|
Interslice distance |
1.5 mm |
1.5 mm |
|
Number of slices |
244 |
264 |
|
Matrix of reconstruction |
512 × 512 |
512 × 512 |
|
Slice thickness |
3 mm |
3 mm |
|
Energy (peak kilovoltage) |
120 kV |
120 kV |
|
X-ray tube current |
443 mA·s |
375 mA·s |
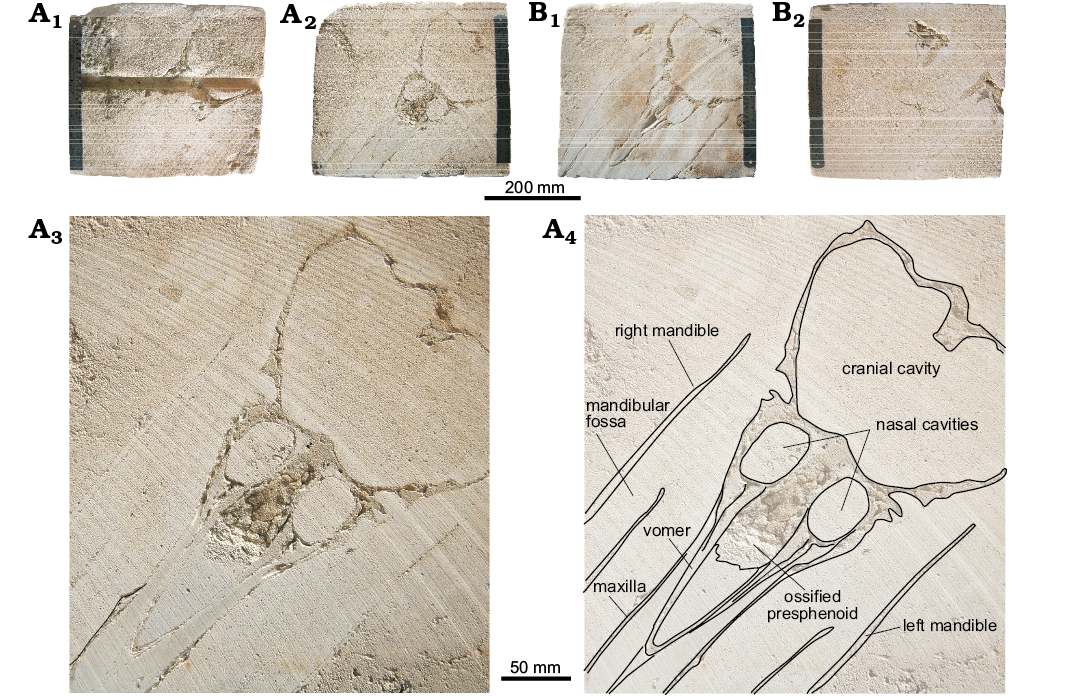
Fig. 2. The two limestone slabs showing longitudinal cross sections of the Messapicetus cf. longirostris skull (MDM-2029) from the Tortonian of Menorca (Balearic Islands, Spain). A. Surface of slab 1 (A1, top; A2, bottom), details of A2 (A3, A4, explanatory drawing). B. Surface of slab 2 (B1, top; B2, bottom).
The final processing of the images, which resulted in the 3D reconstruction of the cetacean specimen, was carried out at the ICP by using the software package AVIZO 7.1.0 (VSG, USA). The endocranial cavity, the choanae and the internal oral cavity (mainly occupied by the larynx in the living animal), all filled with sediment, were also reconstructed and associated with the 3D model of the skull (Figs. 3–8).
The skull was described on the basis of the CT-scan 3D reconstruction and CT-scan sections, integrated with direct observations on the bones exposed on the cut surfaces of the two limestone slabs.
Anatomical terminology follows Mead and Fordyce (2009), except when explicitly stated otherwise.
Systematic palaeontology
Cetacea Brisson, 1762
Odontoceti Flower, 1867
Ziphiidae Gray, 1850
“Messapicetus clade” (sensu Bianucci et al. 2016)
Genus Messapicetus Bianucci, Landini, and Varola, 1992
Type species: Messapicetus longirostris Bianucci, Landini, and Varola, 1992 from the Tortonian beds of Cisterna quarry, near Lecce (southern Italy).
Messapicetus cf. longirostris Bianucci, Landini, and Varola, 1992
Figs. 2–9.
Material.—MDM-2029, an incomplete skull (cranium and mandibles) preserved within two limestone slabs. The cranium lacks most of the rostrum, the vertex area, the posteroventral walls of the braincase, the whole right squamosal, part of the left squamosal and all the ear bones. The two incomplete mandibular rami (of which only the posteroalveolar portions are preserved) are still in anatomical position with respect to the cranium. From lower Tortonian N16 (Miocene), 2.5 km East of Ciutadella de Menorca, Spain.
Description.—Premaxilla: In dorsal view, the premaxilla appears as transversely narrow on the preserved portion of the rostrum, whereas it widens abruptly posterior to the antorbital notch (Fig. 3). The medial margins of the premaxillae do not contact each other, reaching the maximum distance of 27 mm ca. 100 mm anterior to the base of the rostrum. The transverse sections of the rostrum indicate that the premaxillae are clearly thickened and more dorsally convex than the maxillae close to the anterior end of the preserved portion of the rostrum, whereas they are thin and dorsally excavated near the base of the rostrum (Fig. 4). This proximal excavation of the rostral portion of the premaxillae, further emphasised by the lateral thickening of the maxillae (forming a prenarial basin sensu Lambert 2005), is also clearly visible in dorsal and anterodorsal views of the cranium. The premaxillary sac fossae are weakly asymmetrical, with the right fossa being slightly transversely wider than the left (ratio between the widths of the left and right premaxillary sac fossae = 0.77). At least three small foramina are visible near the anterolateral margin of the right premaxillary fossa, the anteriormost one being located 18 mm anterior to the corresponding antorbital notch. It is not clear whether one of these foramina might represent the main premaxillary foramen.
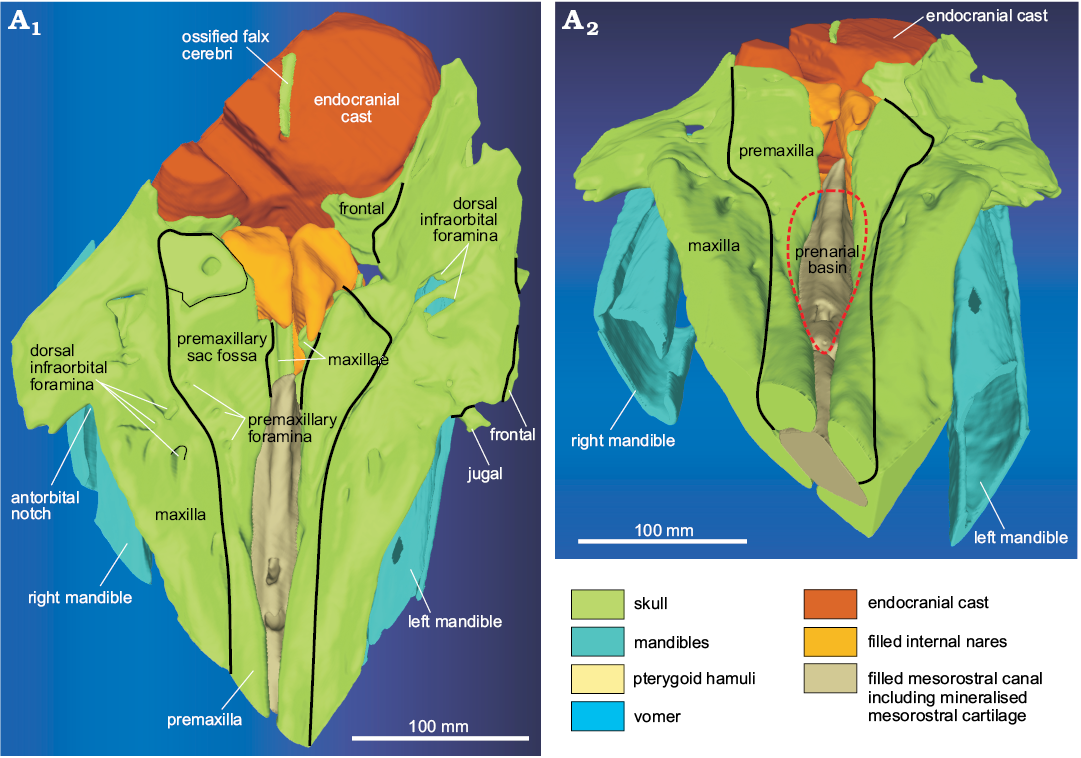
Fig. 3. 3D reconstruction resulting from the CT-scan of the ziphiid cetacean Messapicetus cf. longirostris Bianucci, Landini, and Varola, 1992 skull (MDM-2029) inside two limestone slabs from the Tortonian of Menorca (Balearic Islands, Spain), in dorsal (A1) and anterodorsal (A2) views.
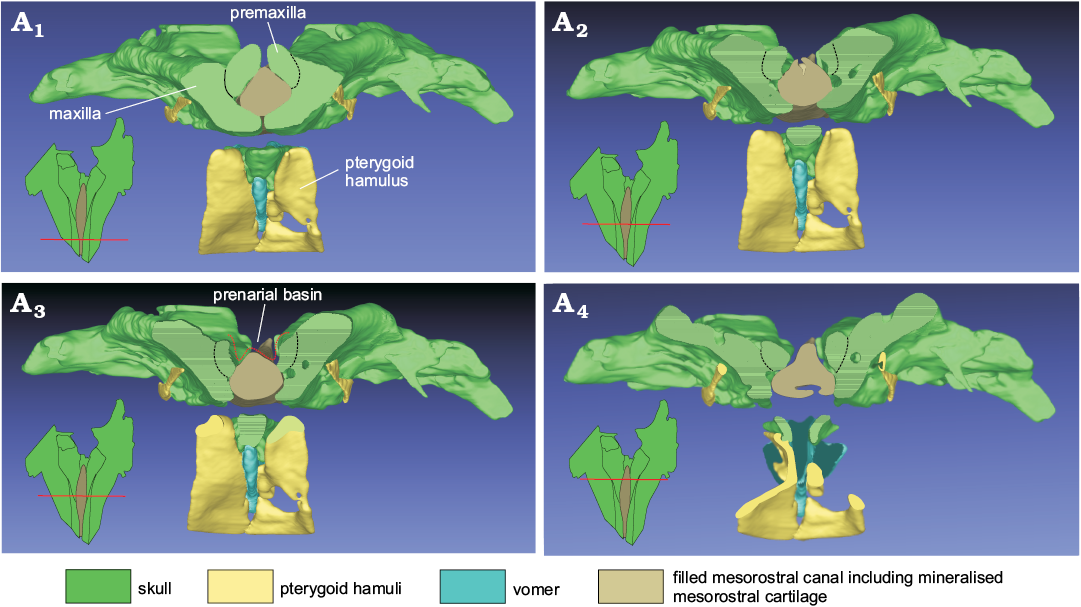
Fig. 4. 3D reconstruction resulting from the CT-scan of the ziphiid cetacean Messapicetus cf. longirostris Bianucci, Landini, and Varola, 1992 skull (MDM-2029) inside two limestone slabs from the Tortonian of Menorca (Balearic Islands, Spain); in anterior view, showing transverse cross sections of the rostrum (A1–A4) made at different distances from the rostrum base. Not to scale.
Maxilla: The maxilla is widely exposed on the dorsal surface of the rostrum, being transversely wider than the premaxilla for all the preserved portion of the rostrum (Fig. 3). The lateral margin of the preserved rostral portion of the maxilla rises progressively toward the antorbital notch, thus forming an elevated rostral maxillary crest, which is also clearly visible in the transverse sections of the rostrum (Fig. 4). This crest does not extend posterior to the antorbital notch in the supraorbital region. Clusters of large (5–9 mm wide) and small (2–3 mm) dorsal infraorbital foramina are visible on both maxillae, both anterior and posterior to the antorbital notches. Their size and position are different on the right and left maxilla. Indeed, on the right maxilla, there are three large foramina located from 5 to 22 mm anterior to the antorbital notch and at least two large foramina located from 23 to 62 mm posterior to the antorbital notch; in turn, on the left maxilla, only one of the five foramina located near the antorbital notch is large in size, whereas two large foramina are located, close to each other, 60–70 mm posterior to the antorbital notch. Posterior to the antorbital notch, the thin ascending process of the maxilla almost completely covers the frontal. No trace of a maxillary crest is observed on the lateral margin of the ascending process of the maxilla. Just anterior to the nares, a narrow strip of the maxilla is apparently exposed medial to each premaxilla. On the palatal surface of the maxillae, there is no trace of dental alveoli or an alveolar groove, indicating that no teeth were present in the posteriormost portion of the rostrum, until a level at least 160 mm anterior to the antorbital notches (Fig. 5). The ventral infraorbital foramen is visible on the posteromedial surface of the maxilla.
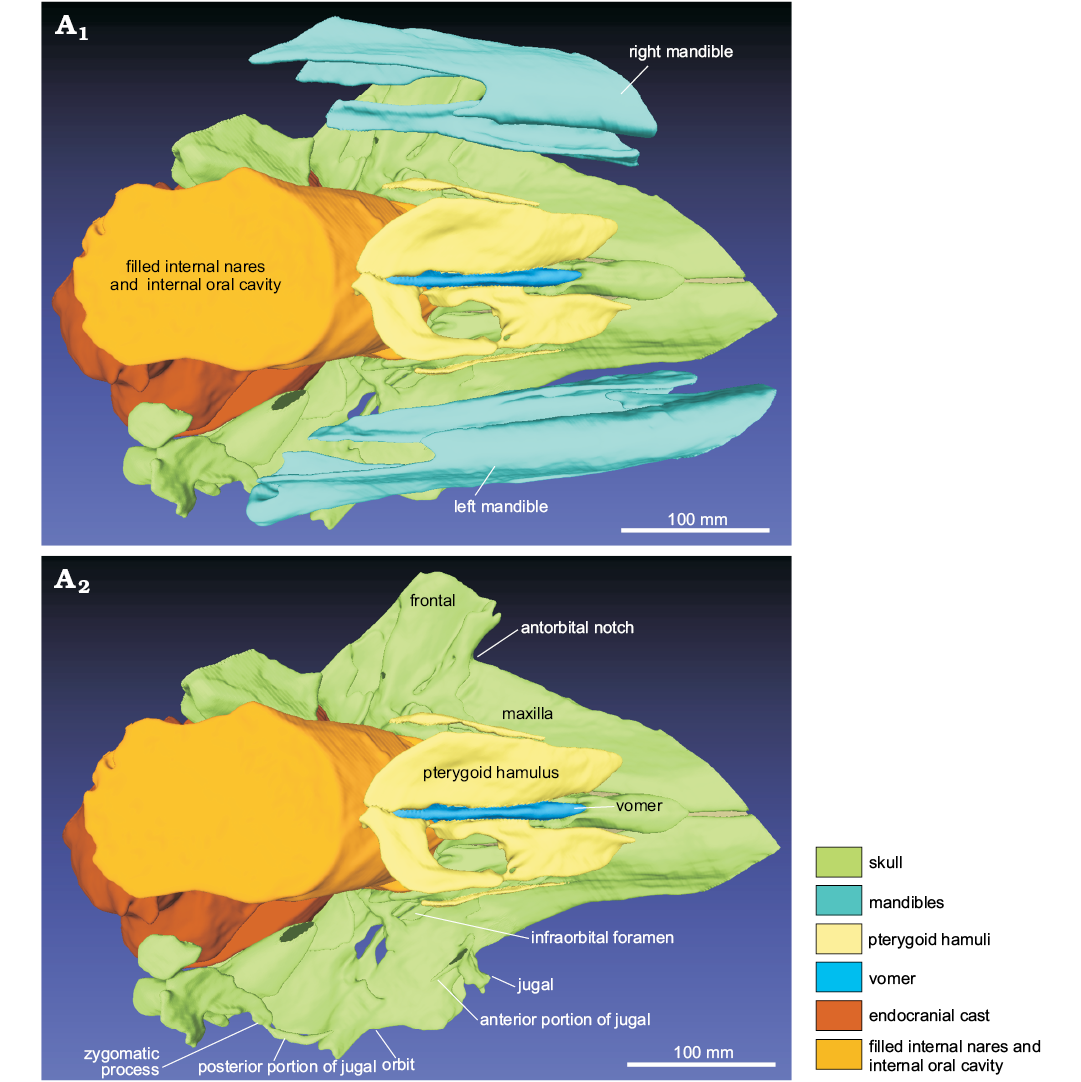
Fig. 5. 3D reconstruction resulting from the CT-scan of the ziphiid cetacean Messapicetus cf. longirostris Bianucci, Landini, and Varola, 1992 skull (MDM-2029) inside two limestone slabs from the Tortonian of Menorca (Balearic Islands, Spain); in ventral view (A1), ventral view without the mandibles (A2).
Frontal: In dorsal view, the frontal is almost totally covered by the ascending process of the maxilla, being visible for a narrow extension, located at the lateral margin of the orbit and, with a small portion, posterolateral to the nares (Fig. 3). In lateral view, the preorbital and supraorbital processes are dorsoventrally thin, whereas the postorbital process is triangular, with a wide ventral tip that points posteroventrally (Figs. 6, 7). The orbit is anteroposteriorly elongated. On the ventral surface of the skull, the frontal is widely exposed, forming the dorsal roof of the orbit and of the temporal fossa (Fig. 5).
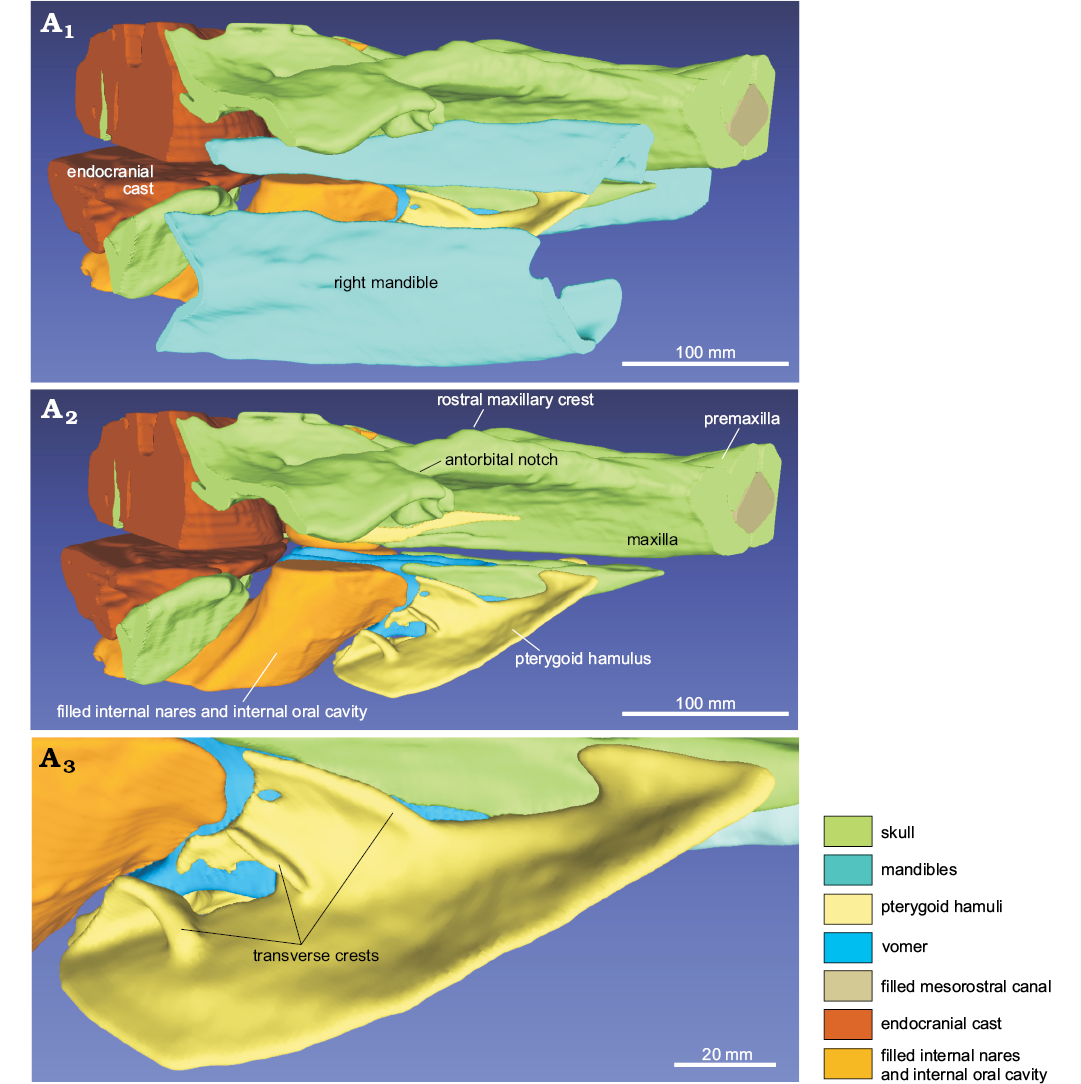
Fig. 6. 3D reconstruction resulting from the CT-scan of the ziphiid cetacean Messapicetus cf. longirostris Bianucci, Landini, and Varola, 1992 skull (MDM-2029) inside two limestone slabs from the Tortonian of Menorca (Balearic Islands, Spain); in right lateral view (A1), right lateral view without the mandibles (A2), detail of the pterygoid hamuli showing the well-preserved transverse crests (A3).
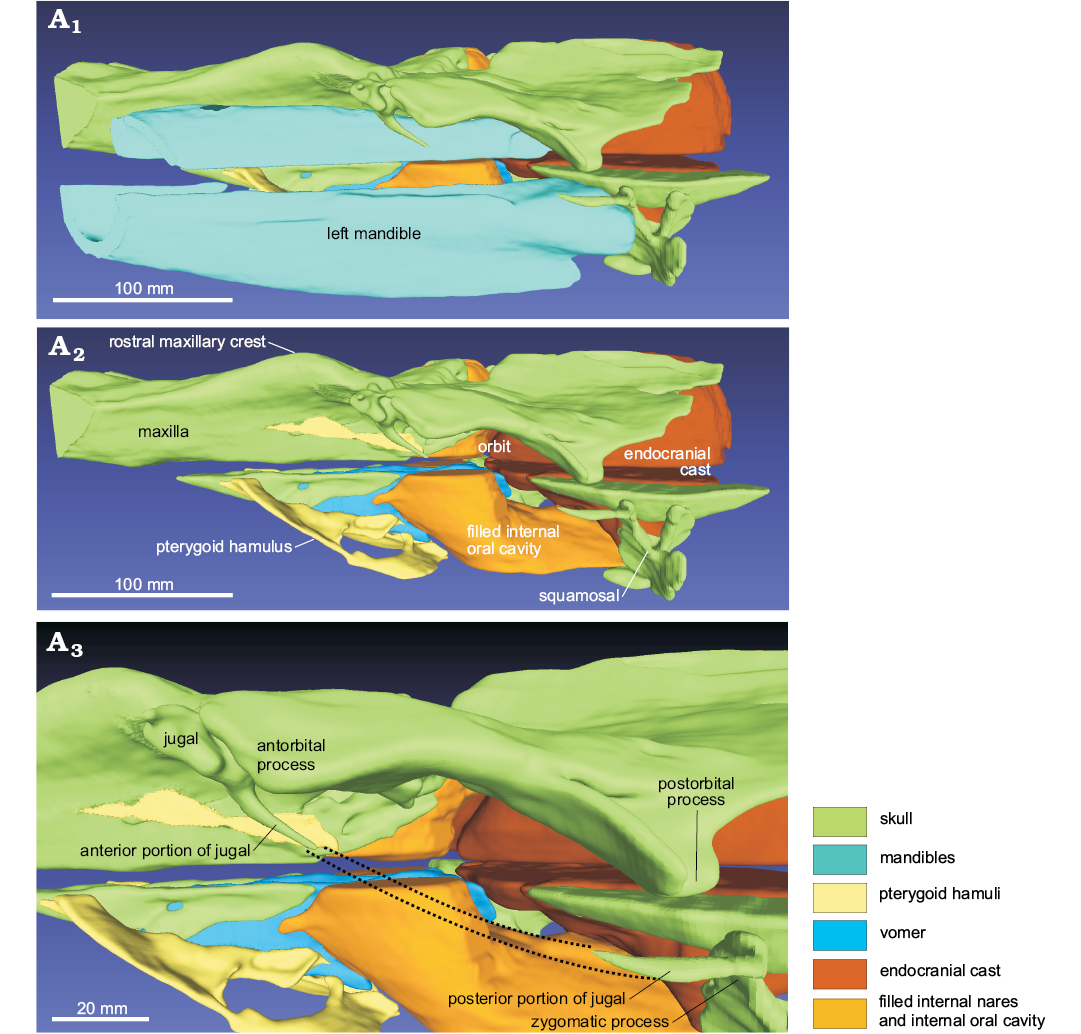
Fig. 7. 3D reconstruction resulting from the CT-scan of the ziphiid cetacean Messapicetus cf. longirostris Bianucci, Landini, and Varola, 1992 skull (MDM-2029) inside two limestone slabs from the Tortonian of Menorca (Balearic Islands, Spain); in left lateral view (A1), left lateral view without the mandibles (A2), detail of the orbital area (A3).
Presphenoid and vomer: On the cut surface of the two slabs, a section of the ossified nasal septum is observed (Fig. 2A3, A4); it is formed by the ossified presphenoid and, laterally, by the vomer. Anterior to the nasal septum, the mesorostral canal is filled internally by sediment, as also confirmed by the CT-scan, showing that this region of the slabs has density values lower than in the surrounding areas formed by dense maxillae (Fig. 8). Nevertheless, the CT-scan reconstruction of the dorsal surface of the skull (Fig. 3) and the CT cross sections of the rostrum (Fig. 4) highlights a dorsal relief at the middle of the mesorostral canal that could represent the mineralised mesorostral cartilage (i.e., cartilaginous nasal septum), perhaps in origin partially ossified. The vomer extends anterior to the nasal septum, forming a narrow and elevated keel that is visible in ventral view in a narrow fissure between the medial margins of the pterygoids (Fig. 5). The vomer is not pachyostotic and does not fill the mesorostral canal.
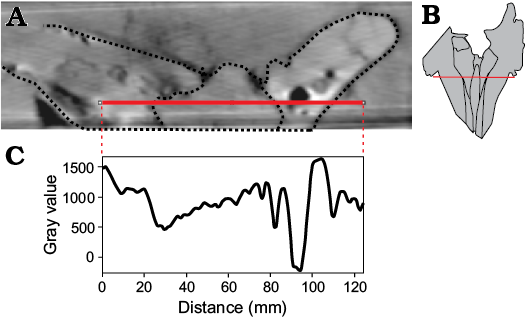
Fig. 8. CT-scan of the ziphiid cetacean Messapicetus cf. longirostris Bianucci, Landini, and Varola, 1992 skull (MDM-2029) inside two limestone slabs from the Tortonian of Menorca (Balearic Islands, Spain). A. Transverse section of the rostrum. B. Dorsal outline of the cranium showing the position of the section. C. Variation of the gray-value density along the transverse section of the rostrum.
Jugal and lacrimal: The anterior process of the jugal is dorsally exposed, forming a distinct tubercle between the antorbital notch and the antorbital process of the frontal (“lateral process” sensu Lambert et al. 2013) (Fig. 3). The preserved anterior and posterior portions of the styliform process of the left jugal are slender (Fig. 7). In ventral view, the suture between the jugal and the lacrimal is not clearly visible (Fig. 5). Compared to the anteroposterior length of the orbit, the anteroposterior extension of the lacrimojugal complex is small.
Pterygoid: The hamular process of the pterygoid is large, extending anteriorly onto the palatal surface of the rostrum for ca. 110 mm from the antorbital notch and ventrally covering the wide pterygoid sinus fossa at its anterior end (Fig. 5). Together, the posterior margins of the hamular processes form a medial point that is posteriorly directed (a plesiomorphic condition that is absent in all other ziphiids for which this feature is known). Nevertheless, it is possible that this shape is due to the fact that the posterolateral portions of the hamular processes are missing or too delicate to be imaged by means of CT-scan analyses. As in Ninoziphius platyrostris and in a few skulls of extant ziphiids (Lambert et al. 2013), the inner surface of the hamular process is crossed by clearly distinct transverse crests (Fig. 6A3).
Squamosal: The sections of both zygomatic processes of the squamosals are exposed on one of the cut slab surfaces (Fig. 2B2). The left zygomatic process, partially reconstructed using the CT-scans, appears short and directed anterodorsally. Its anterior apex articulates with the posterior preserved portion of the styliform process of the jugal (Figs. 5, 7).
Mandible: The dorsal surface of the preserved posterior portion of the right mandible (which is more complete than the left) does not exhibit hints of either distinct dental alveoli or an alveolar groove, thus suggesting that the dentigerous portion of the bone is completely missing. In lateral view, the dorsal margin is weakly concave, with an anteroposteriorly elongated coronoid crest anteriorly limited by a precoronoid crest located 21 mm anterior to the posterior end of the mandibular condyle (Fig. 9A2). Posterior to the precoronoid process, the coronoid crest slopes down progressively towards the coronoid process, which is consequently lower than the precoronoid process. The mandibular condyle is protuberant, being ventrally separated from the angular process by a deep posterior notch. The mandibular foramen extends for more than 20 cm anterior to the posterior end of the mandibular condyle (Fig. 9A4).
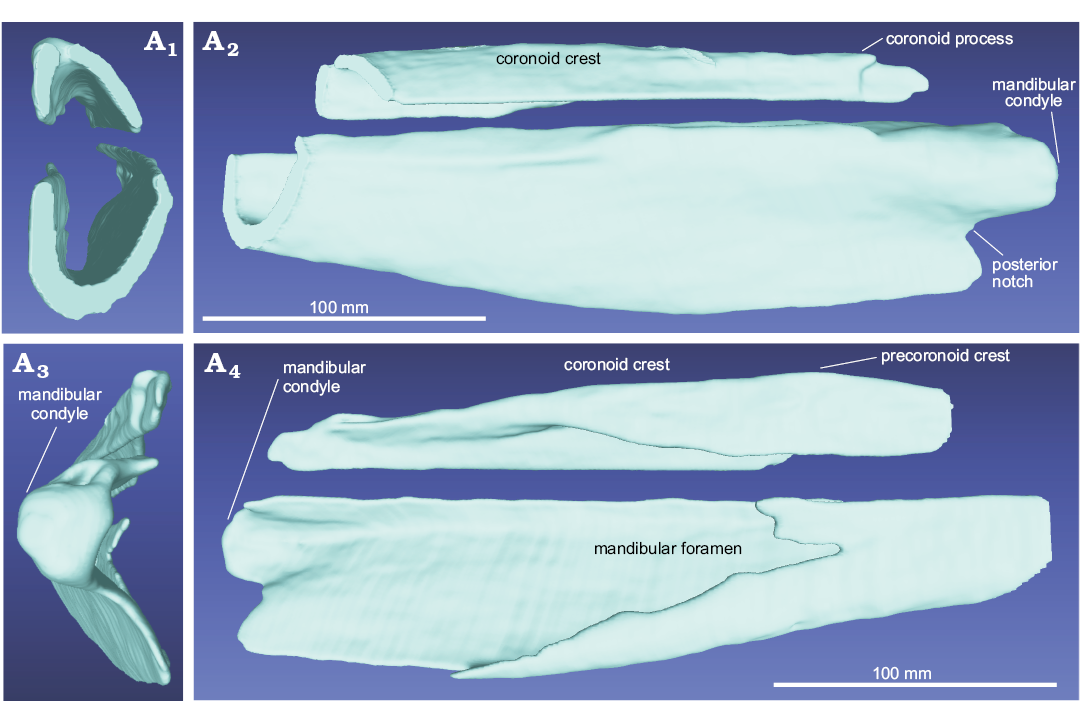
Fig. 9. CT-scan 3D reconstruction of the posterior portion of the left mandible of the ziphiid cetacean Messapicetus cf. longirostris Bianucci, Landini, and Varola, 1992 skull (MDM-2029) inside two limestone slabs from Tortonian of Menorca (Balearic Islands, Spain); in anterior (A1) lateral (A2) posterior (A3), and medial (A4) views.
Remarks.—The Menorca skull exhibits a clear ziphiid synapomorphy: the wide hamular fossa of the pterygoid sinus, extending anterior to the antorbital notch onto the palatal surface of the rostrum (Lambert et al. 2013; Bianucci et al. 2016c: character 35, state 1). Moreover, among odontocetes, transverse crests on the inner surface of the hamular process are only known for a few ziphiid species (Lambert et al. 2013). Among ziphiids, MDM-2029 shares with all the members of the “Messapicetus clade” (sensu Bianucci et al. 2016c, including also Dagonodum; Ramassamy 2016) except Notoziphius the thickening of the compact premaxillae (pachyosteosclerosis) along the rostrum (Bianucci et al. 2016c: character 30, states 1–3). Within the Messapicetus clade, MDM-2029 shares with Aporotus, Beneziphius, Dagonodum, Messapicetus, and Ziphirostrum another derived character: the presence of a prenarial basin bounded laterally by a thick stripe of the maxilla (comparison with Ziphirostrum is based on Ziphirostrum marginatum, the best-known of the three species included in this genus, as well as the only one for which a specimen with precise stratigraphic provenance is known; Lambert 2005). Aporotus and Beneziphius differ from MDM-2029 by the greater thickening of the premaxillae along the rostrum (Bianucci et al. 2016c: character 30, state 2). Aporotus further differs from MDM-2029 by having a rostral maxillary crest that extends posteriorly on the antorbital process, thus delimiting a wide and deep dorsal depression (Lambert 2005). Beneziphius further differs from MDM-2029 by its joined premaxillae that limit anteriorly the prenarial basin and by the dorsal surface of maxilla being horizontal lateral to the prenarial basin (Lambert 2005). Therefore, with respect to MDM-2029, Dagonodum, Messapicetus, and Ziphirostrum appear as the closest genera. Interestingly, all the fossils referred to Messapicetus spp., the holotype and only known specimen of Dagonodum mojnum, and the only Ziphirostrum marginatum skull for which precise information about the stratigraphical provenance is known come from Tortonian strata, as does the Menorca skull.
Although comparisons between MDM-2029 and the sole known specimen Dagonodum are complicated by the fragmentary status of both skulls, clear differences reside in the shape of the posterior portion of the rostrum, as Dagonodum exhibits a well-developed maxillary tubercle and associated prominential notch that is absent in MDM-2029. Messapicetus and Ziphirostrum marginatum are very similar to each other, differing essentially for the degree of elongation of the rostrum—a feature that is not useful for comparisons with the Menorca skull, which only preserves the posterior portion of the rostrum. Nevertheless, two characters, possibly related to the degree of elongation of the rostrum, attribute MDM-2029 to Messapicetus rather than to Z. marginatum: (i) the length of the dorsally open posterior portion of the mesorostral groove is at least 180 mm in the Menorca skull, close to Messapicetus longirostris (206–230 mm) and in the range of Messapicetus gregarius (163–210 mm), but consistently larger than in Z. marginatum (roughly 100 mm); (ii) the posteroalveolar portion of the rostrum is at least 160 mm long in MDM-2029, close to M. longirostris (180 mm) and M. gregarius (150–255 mm), but consistently larger than in Z. marginatum (roughly 100 mm) (Fig. 10).
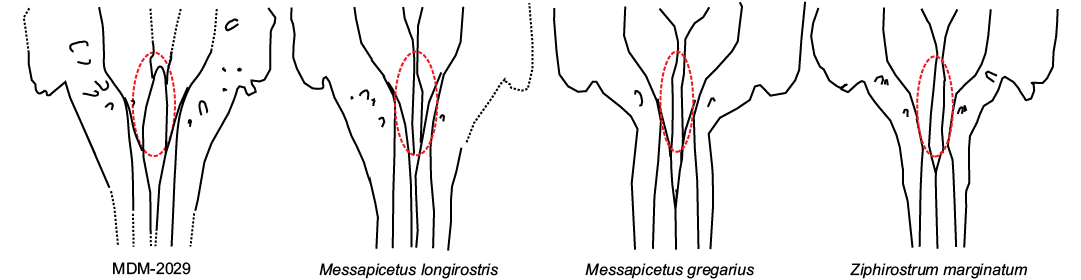
Fig. 10. Comparison of the dorsal view of the preserved cranium of Messapicetus cf. longirostris (MDM-2029) with some stem beaked whales. The dotted ovals indicate the extent of the prenarial basin.
Finally, the aforementioned absence of a distinct maxillary tubercle and of a prominential notch, coupled with the presence on the right maxilla of a cluster of two-three dorsal infraorbital foramina (contra only one foramen in M. gregarius; Bianucci et al. 2016a), also facilitates attribution of MDM-2029 to M. longirostris rather than to M. gregarius. Consequently, MDM-2029 is most closely related to Messapicetus longirostris, both the former and the latter being found in Tortonian deposits of the Mediterranean basin. Indeed, there is a good overlap in size and shape between MDM-2029 and M. longirostris; moreover, several preserved portions of the Menorca skull appear to be very similar to the corresponding features in M. longirostris (e.g., the shape of the rostral maxillary crest, the slender antorbital process, the thin supraorbital process, the anteroposteriorly elongated coronoid crest of the mandible and the protuberant mandibular condyle). The only substantial differences between the Menorca skull and Messapicetus longirostris (and also M. gregarius) is the more anteroposteriorly elongate orbit (Table 2) and the greater dorsoventral thickening of the lateral margin of the preserved rostral portion of the maxilla (Figs. 3A2, 4). The first difference could partly be due to some inaccuracy in the CT-scan 3D reconstruction, whereas the different degrees of thickening of the maxilla could be due to intraspecific ontogenetic variation. Nevertheless, considering also the incompleteness of MDM-2029, we prefer to refer this specimen to Messapicetus cf. longirostris, pending the discovery of more complete material.
Table 2. Cranial measurements (in mm) of Messapicetus cf. longirostris from the Tortonian of Spain (MDM-2029), Messapicetus longirostris from Italy, and Messapicetus gregarious from Peru. Asterisk indicates doubling of measurement from one side.
| |
MDM-2029 |
Messapicetus longirostris |
Messapicetus gregarius |
|
Width of rostrum base at level antorbital notch |
203 |
190* |
203–217 |
|
Width of premaxillae at level antorbital notch |
105 |
|
80–109 |
|
Preorbital width of skull |
300 |
|
296–313 |
|
Postorbital width of skull |
346* |
|
323–352 |
|
Length of orbit |
113 |
88 |
80–88 |
|
Total width of premaxillary sac fossae |
140 |
|
136–142 |
|
Maximum width of right premaxillary sac fossa |
60 |
|
59–67 |
|
Maximum width of left premaxillary sac fossa |
46 |
|
47–58 |
|
Length of left pterygoid hamulus |
170 |
|
195 |
|
Condylar width |
+106 |
|
110–120 |
|
Bizygomatic width |
+335? |
|
310–313 |
Palaeobiogeography
Among extant toothed whales, beaked whales are likely the most enigmatic family, due to their deep-sea habits and elusive behaviour. Scanty and fragmentary until a few decades ago, the fossil record of Ziphiidae is now rich in taxa and well documented, thanks to recent research on specimens from phosphorite layers cropping out on the bottom of deep oceanic areas and from inland deposits, notably the Pisco Formation of Peru (Lambert et al. 2009, 2010, 2013, 2018; Bianucci et al. 2010, 2016a, c; Ichishima et al. 2017). A recent phylogenetic and biogeographical analysis (Bianucci et al. 2016b) recognised two major clades among Ziphiidae characterised by a high degree of diversity and disparity during the late Miocene: (i) the so-called Messapicetus clade, which dominated the North Atlantic and went extinct during the Pliocene; (ii) crown Ziphiidae, most of which inhabited the Southern Ocean and subsequently dispersed globally. Both clades apparently followed similar evolutionary trends, including an increase in the compactness and thickness of the rostrum, the loss of functional teeth, and an increase of body size, possibly related to a convergent ecological shift from ancestral epipelagic and raptorial trophic habits to suction feeding in deep waters (Lambert et al. 2015; Bianucci et al. 2016c). In this evolutionary and biostratigraphic scenario, the geographic distribution of Messapicetus, the basal most member of the Messapicetus clade, is remarkable. In fact, this genus was recorded from the Pisco Formation of Peru (Bianucci et al. 2010, 2016b; Lambert et al. 2015; Di Celma et al. 2017; Ramassamy et al. 2018); it was tentatively recognised from the St. Marys Formation of Maryland, USA (Fuller and Godfrey 2007); it is here reported for the first time in Menorca, Spain; and it was first described from the Pietra leccese of southern Italy (Bianucci et al. 1992, 1994b, 2016a); all these occurrences (Fig. 11) take place in shelf deposits of Tortonian (i.e., early late Miocene) age. Interestingly, Messapicetus and the two earliest branching stem ziphiids Chavinziphius and Ninoziphius are known from the Pisco Formation, suggesting a dispersal route running between the southeastern Pacific and the eastern Mediterranean for early beaked whales. Such a distributional pattern was possible since the Central American Seaway allowed a direct communication between the Pacific and Atlantic oceans in late Miocene times (Jacobs et al. 2004; Bianucci et al. 2010, 2016a).
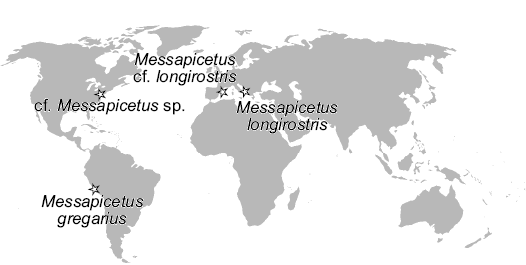
Fig. 11. Geographic distribution of fossil remains of Messapicetus from upper Miocene deposits worldwide (see text for data sources).
Conclusions
We describe a beaked whale skull discovered inside two slabs cut from a commercial block of upper Miocene limestone from Menorca (Balearic Islands, Spain). This specimen was not freed mechanically from the entombing stone, but rather was imaged by means of computed tomography. The resulting three-dimensional reconstruction of the skull proved extremely detailed and pivotal for resolving the morphology of very delicate structures (probably including the only partly ossified mesorostral cartilage) that would probably have been destroyed during traditional mechanical preparation. Messapicetus shows a geographic distribution from Tortonian shelf deposits of Italy, Peru, and the Eastern USA. Early branching beaked whales (including basal members of the so-called “Messapicetus clade”) likely dispersed via the Central American Seaway, which allowed for faunal interchange between the southeastern Pacific and the northern Atlantic/Mediterranean realms until latest Miocene times.
Acknowledgements
The authors want to express their gratitude to the following people and institutions, whose help made this study possible. Thanks are especially due to owner of the Son Salord quarry, Cristobal Torres (Ciutadella de Menorca, Spain), who discovered and brought to our attention the skull here described, for facilitating its study at all times. Thanks also to the management of the Hospital Mateu Orfila (Maó, Menorca) for allowing the use of their CT service, which provided the images on which this work is based. The technical assistance of Marina Febrer Febrer (specialist doctor in radiodiagnosis) and Pedro Delgado Cantero (technician in radiodiagnosis), both at the Hospital Mateu Orfila, allowed us to obtain these images. This project was carried out, to a large extent, thanks to the institutional support of Salvador Moyà-Solà, former director of the ICP. Logistical support from Miquel Fernández (Maó, Spain) and Josep Lluis Florit (Consell Insular de Menorca, Spain) was essential for moving the specimen to the Hospital Mateu Orfila. Antoni Obrador (Universitat Autonoma de Barcelona, Spain) collaborated in the interpretation of the geological context in which the specimen was found. We are sincerely indebted to Stephen J. Godfrey (Calvert Marine Museum, Solomons, USA), Pavel Gol’din (I.I. Schmalhausen Institute of Zoology, National Academy of Sciences of Ukraine, Kyiv, Ukraine), and Olivier Lambert (Institut royal des Sciences naturelles de Belgique, Brussels, Belgium), whose thorough reviews and constructive criticisms substantially contributed to improve an early draft of this paper.
References
Bianucci, G. 1996. I cetacei fossili del Museo di Storia Naturale dell’Università di Pisa. Atti della Società Toscana di Scienze Naturali, Memorie, Serie A 103: 63–69.
Bianucci, G. 2001. A new genus of kentriodontid (Cetacea: Odontoceti) from the Miocene of South Italy. Journal of Vertebrate Paleontology 21: 573–577. Crossref
Bianucci, G. and Gingerich, P.D. 2011. Aegyptocetus tarfa, n. gen. et sp. (Mammalia, Cetacea), from the middle Eocene of Egypt: clinorhynchy, olfaction, and hearing in a protocetid whale. Journal of Vertebrate Paleontology 31: 1173–1188. Crossref
Bianucci, G. and Landini, W. 2006. Killer sperm whale: a new basal physeteroid (Mammalia, Cetacea) from the Late Miocene of Italy. Zoological Jounal of the Linnean Society 148: 103–131. Crossref
Bianucci, G., Collareta, A., Post, K., Varola, A., and Lambert, O. 2016a. A new record of Messapicetus from the Pietra leccese (Late Miocene, Southern Italy): antitropical distribution in a fossil beaked whale (Cetacea, Ziphiidae). Rivista Italiana di Paleontologia e Stratigrafia 122: 63–74.
Bianucci, G., Di Celma, C., Landini, W., Post, K., Tinelli, C., de Muizon, C., Gariboldi, K., Malinverno, E., Cantalamessa, G., Gioncada, A., Collareta, A., Salas-Gismondi, R., Varas-Malca, R., Urbina, M., and Lambert, O. 2016b. Distribution of fossil marine vertebrates in Cerro Colorado, the type locality of the giant raptorial sperm whale Livyatan melvillei (Miocene, Pisco Formation, Peru). Journal of Maps 12: 543–557. Crossref
Bianucci, G., Di Celma, C., Urbina, M., and Lambert, O. 2016c. New beaked whales from the late Miocene of Peru and evidence for convergent evolution in stem and crown Ziphiidae (Cetacea, Odontoceti). PeerJ 4: e2479. Crossref
Bianucci, G., Lambert, O., and Post, K. 2010. High concentration of long-snouted beaked whales (genus Messapicetus) from the Miocene of Peru. Palaeontology 53: 1077–1098. Crossref
Bianucci, G., Landini, W., and Varola, A. 1992. Messapicetus longirostris a new genus and species of Ziphiidae (Cetacea) from the Late Miocene of “Pietra leccese” (Apulia; Italy). Bollettino della Società Paleontologica Italiana 31: 261–264.
Bianucci, G., Landini, W., and Varola, A. 1994a. New remains of Cetacea Odontoceti from the “Pietra leccese” (Apulia, Italy). Bollettino della Società Paleontologica Italiana 33: 215–230.
Bianucci, G., Landini, W., and Varola, A. 1994b. Relationships of Messapicetus longirostris (Cetacea, Ziphiidae) from the Miocene of South Italy. Bollettino della Società Paleontologica Italiana 33: 231–241.
Bianucci, G., Landini, W., and Varola, A. 2004. First discovery of the Miocene northern Atlantic sperm whale Orycterocetus in the Mediterranean. Geobios 37: 569–573. Crossref
Blow, W.H. 1969. Late Middle Eocene to Recent planktonic foraminiferal biostratigraphy. In: R. Bronnimann and H.H. Renz (eds.), Proceedings of the First International Conference on Planktonic Microfossils, Volume I, 199–421. Brill, Leiden.
Cau, A. and Fanti, F. 2011. The oldest known metriorhynchid crocodylian from the Middle Jurassic of North-eastern Italy: Neptunidraco ammoniticus gen. et sp. nov. Gondwana Research 19: 550–565. Crossref
Di Celma, C., Malinverno, E., Bosio, G., Collareta, A., Gariboldi, K., Gioncada, A., Molli, G., Basso, D., Varas-Malca, R.M., Pierantoni, P.P., Villa I.M., Lambert, O., Landini, W., Sarti, G., Cantalamessa, G., Urbina, M., and Bianucci, G. 2017. Sequence stratigraphy and paleontology of the upper Miocene Pisco Formation along the western side of the lower Ica valley (Ica Desert, Peru). Rivista Italiana di Paleontologia e Stratigrafia 123: 255–274.
Fuller, A.J. and. Godfrey, S.J. 2007. A Late Miocene ziphiid (Messapicetus sp.: Odontoceti: Cetacea) from the St. Marys Formation of Calvert Cliffs, Maryland. Journal of Vertebrate Paleontology 27: 535–540. Crossref
Hannibal, J.T. and Davis, R.A. 1992. Guide to the building stones of downtown Cincinnati: A walking tour. Ohio Division of Geological Survey Guidebook 7: 1–44.
Hannibal, J.T., Thomas, S.F., and Straw, W.T. 2008. Building stones and cultural geology of Evansville, Indiana, USA. GSA Field Guides 12: 81–103. Crossref
Ichishima, H., Augustin, A.H., Toyofuku, T., and Kitazato, H. 2017. A new species of Africanacetus (Odontoceti: Ziphiidae) found on the deep ocean floor off the coast of Brazil. Deep Sea Research Part II: Topical Studies in Oceanography 146: 68–81. Crossref
Jacobs, D.K., Haney, T.A., and Louie, K.D. 2004. Genes, diversity, and geological process on the Pacific coast. Annual Review of Earth and Planetary Sciences 32: 601–652. Crossref
Lambert, O. 2005. Systematics and phylogeny of the fossil beaked whales Ziphirostrum du Bus, 1868 and Choneziphius Duvernoy, 1851 (Mammalia, Cetacea, Odontoceti), from the Neogene of Antwerp (North of Belgium). Geodiversitas 27: 443–497.
Lambert, O., Bianucci, G., and Post, K. 2010. Tusk bearing beaked whales from the Miocene of Peru: Sexual dimorphism in fossil ziphiids? Journal of Mammalogy 91: 19–26. Crossref
Lambert, O., Collareta, A., Landini, W., Post, K., Ramassamy, B., Di Celma, C., Urbina, M., and Bianucci, G. 2015. No deep diving: evidence of predation on epipelagic fish for a stem beaked whale from the Late Miocene of Peru. Proceedings of the Royal Society B 282: 20151530. Crossref
Lambert, O., de Muizon, C., and Bianucci, G. 2013. The most basal beaked whale Ninoziphius platyrostris Muizon, 1983: clues on the evolutionary history of the family Ziphiidae (Cetacea: Odontoceti). Zoological Journal of the Linnean Society 167: 569–598. Crossref
Lambert, O., de Muizon, C., Duhamel, G., and Plicht, J.V.D. 2018. Neogene and Quaternary fossil remains of beaked whales (Cetacea, Odontoceti, Ziphiidae) from deep-sea deposits off Crozet and Kerguelen islands, Southern Ocean. Geodiversitas 40: 135–140. Crossref
Mas, G., Escalante, F., and Quintana, J. 2013. Primera cita de un Delphinidae en el Neógeno de las Islas Baleares. Batalleria 18: 45–51.
Mateu-Vicens, G., Hallock, P., and Brandano, M. 2008. A depositional model and paleoecological reconstruction of the lower Tortonian distally steepened ramp of Menorca (Balearic Islands, Spain). Palaios 23: 465–481. Crossref
Mead, J.G. and Fordyce, R.E. 2009. The therian skull: a lexicon with emphasis on the odontocetes. Smithsonian Contributions to Zoology 627: 1–261. Crossref
Mercadal, B., Pilleri, G., and Casinos, A. 1985. A tooth of Scaldicetus grandis (Du Bus, 1872) (Physeteridae) from Aire Island (Menorca, Spain). Investigations on Cetacea 17: 31–33.
Pomar, L. 2001. Ecological control of sedimentary accommodation: evolution from a carbonate ramp to rimmed shelf, Upper Miocene, Balearic Islands. Palaeogeography, Palaeoclimatology, Palaeoecology 175: 249–272. Crossref
Pomar, L., Obrador, A., and Westphal, H. 2002. Sub-wavebase cross-bedded grainstones of a distally steepened carbonate ramp, Upper Miocene, Menorca, Spain. Sedimentology 49: 139–169. Crossref
Ramassamy, B. 2016. Description of a new long-snouted beaked whale from the Late Miocene of Denmark: evolution of suction feeding and sexual dimorphism in the Ziphiidae (Cetacea: Odontoceti). Zoological Journal of the Linnean Society 178: 381–409. Crossref
Ramassamy, B., Lambert, O., Collareta, A., Urbina, M., and Bianucci, G. 2018. Description of the skeleton of the fossil beaked whale Messapicetus gregarius: searching potential proxies for deep-diving abilities. Fossil Record 21: 11–32. Crossref
Robinson, E. and Bishop, C. 1980. Geological walks around St. Paul’s. Proceedings of the Geologists’ Association 91: 241–260. Crossref
Rosell, J., Gómez-Gras, D., and Elizaga, E. 1989. Mapa Geológico de España escala 1:25.000. Hoja 617bis-II, 618-III (Ciutadella). Instituto Tecnológico Geominaro de España, Madrid.
Sauleau, L. 1985. Pedreres i trencadors. Quaderns de Folklore 22–23: 86 pp.
Wilson, E. 1983. Swedish limestone paving in 17th- and 18th-century English buildings. Post-Medieval Archaeology 17: 95–109. Crossref
Acta Palaeontol. Pol. 64 (2): 291–302, 2019
https://doi.org/10.4202/app.00593.2019