
The first Triassic vertebrate fossils from Myanmar: Pachypleurosaurs in a marine limestone
KHAING KHAING SAN, NICHOLAS C. FRASER, DAVIDE FOFFA, OLIVIER RIEPPEL, and STEPHEN L. BRUSATTE
As ecosystems recovered from the end-Permian extinction, many new animal groups proliferated in the ensuing Triassic. Among these were the sauropterygians, reptiles that evolved from terrestrial ancestors and transitioned to a marine environment. The first sauropterygians were small, marine-adapted taxa such as pachypleurosaurs, which are known from Middle–Late Triassic deposits, particularly in the Tethyan realm of Europe, and more recently from Lagerstätten in southwestern China. Here we report two pachypleurosaurs from Myanmar, the first Triassic vertebrate fossils from the country. These specimens demonstrate that their entombing rocks in northern Shan State, which have received less study than terrestrial sediments in southern Shan State and whose ages have long been uncertain, are Triassic. The specimens may be among the oldest pachypleurosaurs globally, potentially corroborating biogeographic scenarios that posit an eastern Tethyan origin for pachypleurosaurs, and raise the potential for future discoveries of well-preserved Triassic reptiles in Myanmar.
Institutional abbreviations.—CMLV, Cultural Museum, Lashio, Myanmar; YDBGLV, Geology Museum, Yadanabon University, Myanmar.
Geological and geographical setting
Myanmar is divided into four tectonic provinces. From east to west, these are the (i) Shan-Tanintharyi Block; (ii) Central Cenozoic Belt; (iii) Western Fold Belt; and (iv) Rakhine Coastal Belt (Chhibber 1934; Win Swe 1972; Maung Thein 1973). The pachypleurosaur fossils were collected immediately west of the town of Lashio, in northern Shan State, Myanmar (Fig. 1). They were found in rocks of the Lashio Basin, in the northern part of the Shan Massif, which is part of the Shan-Tanintharyi Block. The Lashio area is composed mainly of sedimentary rocks that date from the Middle Devonian to the Jurassic.
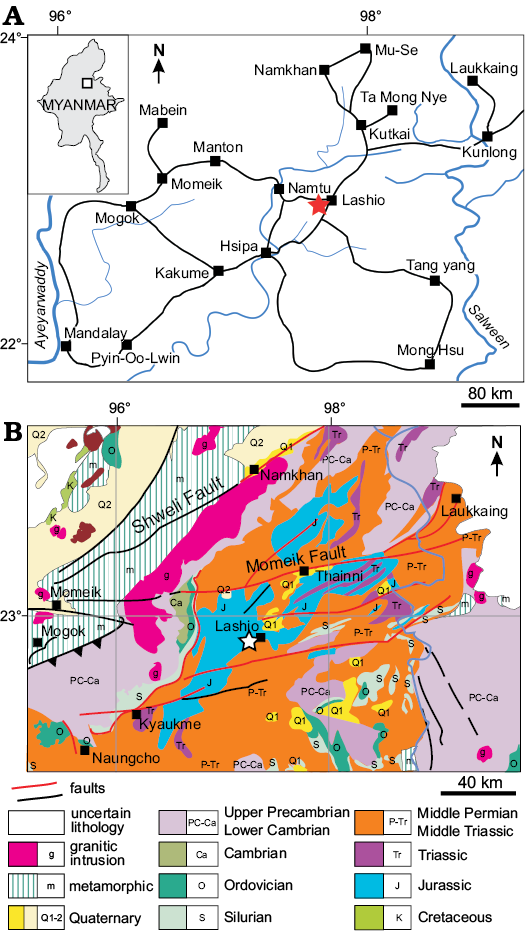
Fig 1. Geographical (A) and geological setting (B) for the Triassic pachypleurosaurs from Myanmar. Study area indicated by a star. B based on the 2014 geological map of Myanmar from the Myanmar Geoscience Society.
Both specimens were found in a ferruginous, micritic limestone, associated with indeterminate fish bones and scales. The limestone crops out in a mountain range, locally called the Yebawhaung Kyauk-taung (UTM map 2297-9; 22°56′04″ N, 97°42′49″ E). The fossil-bearing limestone is sandwiched between thin- to medium-bedded, light to dark grey, hard and compact limestone and red to purple, uniformly thin-bedded, siltstone and mudstone. Dark grey chert stringers or nodules are intercalated within the fossil-bearing limestone, and minor amounts of ore (lead-zinc) mineralization can be observed in some parts of the bed. This limestone unit apparently grades into the surrounding dolomites.
The fossil-bearing limestone was previously mapped as belonging to the Nwabangyi Dolomite Formation (Garson et al. 1976), considered Late Permian to Middle Triassic in age based on the occurrence of the foraminiferan Glomospirella irregularis in the Kyaukme-Longtawkno area of northern Shan State (Brönnimann et al. 1975). This limestone has been correlated with the Thigaungtaung Limestone (Early–Middle Triassic, Induan–Anisian) of southern Shan State (Amos 1975; Whittaker in Brönnimann et al. 1975; see also the recent review of the “Plateau Limestone” of southern Shan State by Win et al. 2015). Furthermore, Sahni (1936) compiled other records of lower-most Triassic units (Scythian, Induan–Olenekian) at Namhkam (north of Lashio and west of Hsenwi), to which he gave the informal name of the “Na-hkan Beds”. The detailed stratigraphy, correlations, and ages of these various units are in need of revision. Vertebrate fossils may help better constrain their ages (see below).
Description
Pachypleurosaur morphology is best known from complete specimens of Anarosaurus and Dactylosaurus from the lower Muschelkalk (lower Anisian) of the Germanic basin (Sues and Carroll 1985; Rieppel and Lin 1995; Klein 2009, 2012), the late Anisian Serpianosaurus mirigiolensis from the southern Alps (Rieppel 1989), the early and middle Ladinian Neusticosaurus (three species) again from the southern Alps (Carroll and Gaskill 1985; Sander 1989), and the Ladinian (possibly early Carnian) Keichousaurus from southwestern China (Lin and Rieppel 1998; Cheng et al. 2009; Cheng et al. 2004; Holmes et al. 2008). Dianopachysaurus dingi is another well-preserved pachypleurosaur from the Anisian of southwestern China, which has played an important role in pachypleurosaur phylogeny reconstruction (Liu et al. 2011). In this section, we compare the Myanmar specimens to these taxa, especially Keichousaurus and Dianopachysaurus given the geographical proximity of their occurrences (see also Rieppel 2000).
Specimen CMLV_1 (Fig. 2) includes a partial skull, much of the neck and pectoral girdles, and a substantial portion of the trunk. Few morphological details are observable on the skull, but the retroarticular process is clearly distinct, as in other pachypleurosaurs. The humerus is evenly curved, more distinctly so than in other pachypleurosaurs, except perhaps in immature individuals. There appears to be a large number of ossifications in the left carpus, although some of these may represent broken bones instead of separate carpals. The dorsal ribs are not pachyostotic and the thin gastral ribs number at least two per vertebral segment in the posterior trunk, as in other pachypleurosaurs.
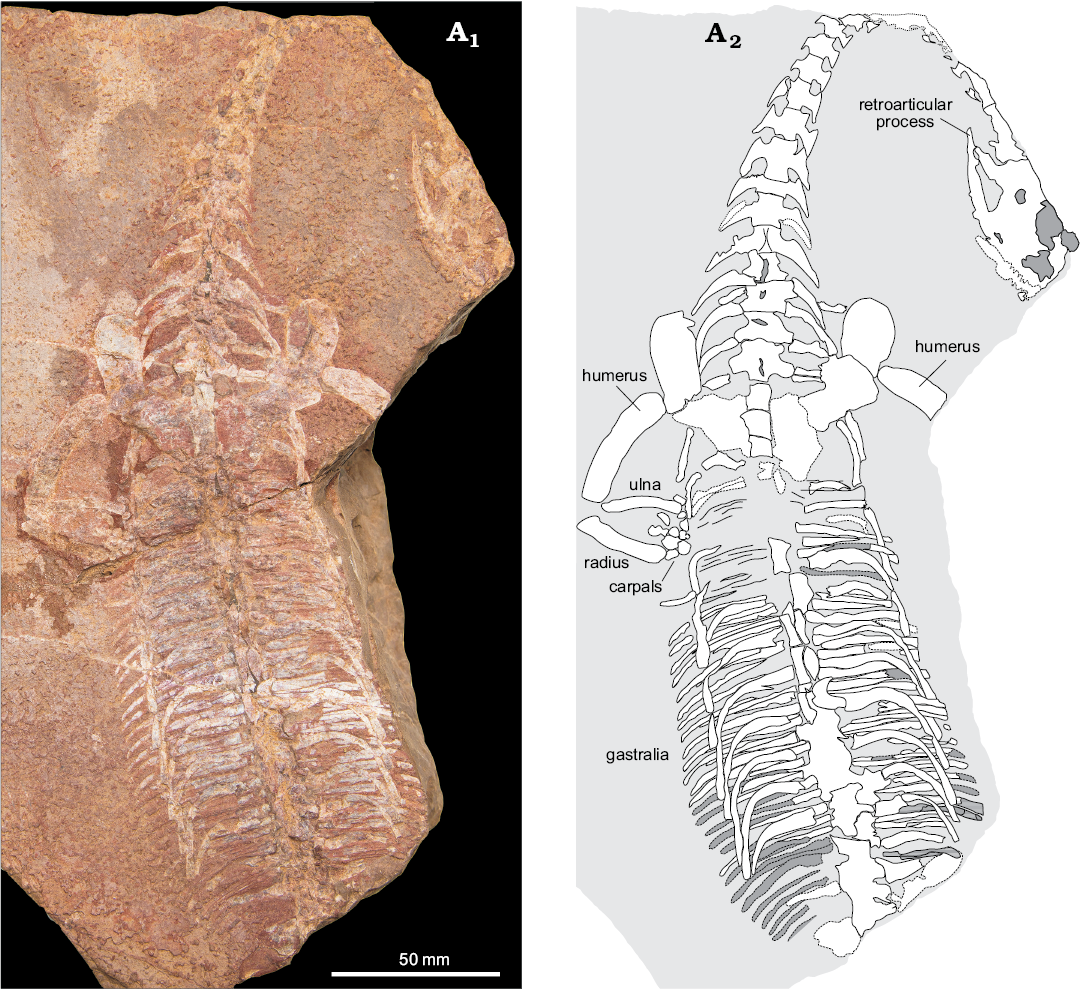
Fig. 2. Unidentified pachypleurosaur (CMLV_1) from the Triassic of Myanmar, in dorsal view. Much of the neck, pectoral girdles, and trunk are preserved, along with part of the skull and forelimbs. The pelvis, hindlimbs, and tail are not preserved. Photograph (A1) and explanatory drawing (A2). For measurements see Table 1.
Table 1. Key measurements (in mm) of unidentified pachypleurosaurs from Myanmar (after Sander 1989). Standard length is defined as the length of the four posterior dorsal centra; na, not available (as the specimen is incomplete).
| |
CMLV_1 |
YDBGLV_4 |
|
skull |
51 |
50 |
|
neck length |
151 |
127 |
|
neck length |
165 |
112 |
|
standard length |
na |
27 |
|
incomplete tail |
na |
42 |
|
humerus |
46 |
37 |
|
radius |
2.7 |
22 |
|
ulna |
29 |
23 |
|
femur |
na |
38 |
|
fibula |
na |
22 |
|
tibia |
na |
23 |
|
snout-vent length |
– |
315 |
Specimen YDBGLV_4 (Fig. 3) preserves much of the skeleton, and it has the lizard-like habitus, with a short-snouted skull and overall body proportions, that are typical for pachypleurosaurs. Also characteristic of pachypleurosaurs is the homodont dentition comprising numerous small, peg-like teeth. The snout is not constricted and the orbits are large compared to the postorbital region of the skull, as in other pachypleurosaurs. As in Anarosaurus, Dactylosaurus, and Keichousaurus, the cheek is deeply embayed. There are no clear supratemporal fenestrae, and it appears they have been very much reduced. Possible fragmentary traces of these openings occur in the posterolateral corners of the parietal table, but there are no clear margins, and they could even be completely absent. Scleral ossicles appear to be absent in the better-preserved right orbit; these are variably present in other pachypleurosaurs. The pineal foramen is located in the centre of the parietal skull table, as is typical for pachypleurosaurs.
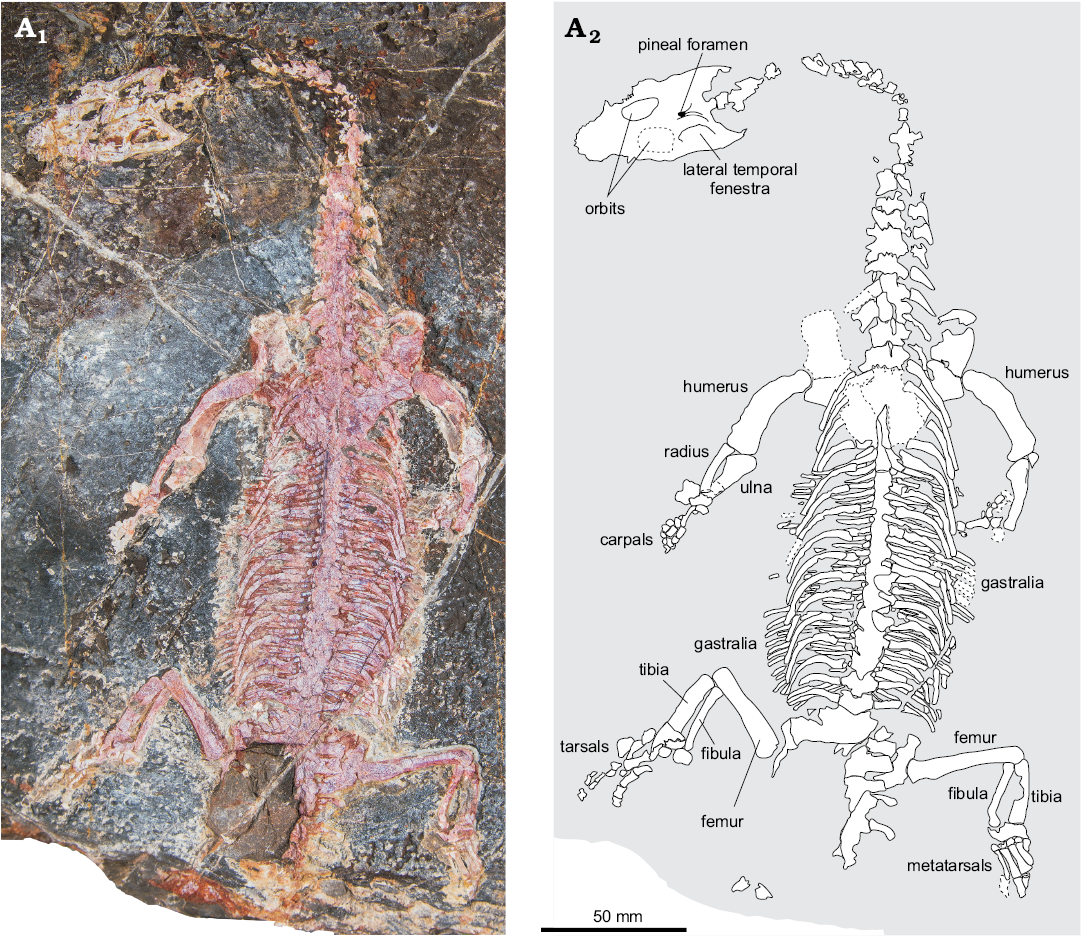
Fig. 3. Unidentified pachypleurosaur (YDBGLV_4) from the Triassic of Myanmar, in dorsal view. Most of the skeleton is preserved, except for the tail. Photograph (A1) and explanatory drawing (A2). For measurements see Table 1.
In the axial skeleton CMLV_1 exhibits the individual cervical vertebrae more clearly than YDBGLV_4, but unfortunately the series runs along the edge of the block and only traces of each of the anterior cervicals remain. In YDBGLV_4 the outline of the neck is more intact but there is a significant gap between the posterior margin (occiput) of the skull and the mid cervical series, which renders it difficult to obtain more than an approximate cervical count. Moreover, in both specimens it is difficult to demarcate the cervical series from the first dorsal vertebra, but we consider there to be at least 20 cervical vertebrae, and perhaps as many as 24. These numbers are within the range of variation known in other pachypleurosaurs, although tending towards the higher end (e.g., 25–26 Keichousaurus; 20 Dianopachysaurus; 17 Dactylosaurus; 15–18 Serpianosaurus; 17 Neusticosaurus edwardsii; 18–20 Neusticosaurus pusillus; Carroll and Gaskill 1985; Rieppel and Lin 1995; Sues and Carroll 1985; Sander 1989; Lin and Rieppel 1998; Liu et al. 2011).
There are at least 16 left dorsal ribs. This would correspond to an unusually low number of dorsal vertebrae for a pachypleurosaur, although not too dissimilar to Keichousaurus (18–19) and Dianopachysaurus (19). Other members of the group have more dorsal vertebrae, with, for example, 22–24 in Neusticosaurus pusillus and as many as 25–26 in Anarosaurus (Sander 1989; Rieppel and Lin 1995; Lin and Rieppel 1998). It is possible that two vertebrae in the cervical/dorsal transitional zone are not cervicals but dorsals, meaning that there would be approximately 22 cervicals and approximatley 18 dorsals in YDBGLV_4. There are three sacral vertebrae, as in other pachypleurosaurs except for Keichousaurus, which has 2 or 3 (Lin and Rieppel 1998). The neural arches of the dorsal vertebrae are compressed, and apparently pachyostotic. Pachyostosis is conspicuously absent, however, in the dorsal ribs. The delicate gastral ribs are numerous in number, and there are at least two per vertebral segment in the posterior trunk region.
In the appendicular skeleton, the coracoid is elongate, plate-like, and biconcave, as is standard for pachypleurosaurs. The scapula is difficult to discern in detail, but appears unremarkable for a pachypleurosaur, and the interclavicle is indistinct. The most salient feature of the appendicular skeleton is the distinctly and evenly curved humerus, as in CMLV_1. This differs from other (mature) pachypleurosaurs, but is a feature the Myanmar pachypleurosaur shares with Dianopachysaurus (Liu et al. 2011). The entepicondylar foramen seems to be absent, an unusual character state otherwise known only in Keichousaurus amongst pachypleurosaurs. The ulna is distinctly broadened proximally, more so than in most other pachypleurosaurs, but not overall as broad as in Keichousaurus. The number and morphology of the carpal ossifications and phalanges in the manus are difficult to observe. The pelvis is also difficult to describe, but the femur is slender, weakly sigmoidally curved, and longer than the humerus. This latter feature is again shared with Dianopachysaurus (Liu et al. 2011), and also with Anarosaurus, where the femur length exceeds humerus length to an even greater degree (Rieppel and Lin 1995; Klein 2012). In other pachypleurosaurs known from a larger number of specimens (Neusticosaurus, Serpianosaurus, Carroll and Gaskill 1985; Rieppel 1989; Sander 1989), the ratio of humerus to femur length is subject to ontogenetic variation as well as sexual dimorphism (Rieppel 1993: fig. 8). The metatarsals of the right foot are well preserved; the first metatarsal is much shorter than the others, as is standard for pachypleurosaurs.
Concluding remarks
The two new specimens from Myanmar, CMLV_1 and YDBGLV_4, can both be assigned to Pachypleurosauria because they possess several diagnostic features of the clade, including: preorbital region of the skull longer or subequal in length to postorbital region; upper temporal fenestra much smaller than the orbit; pre- and postzygapophyses pachyostotic; sacral ribs not expanded distally. The two specimens most likely belong to the same lower-level taxon, due to their shared presence of an evenly and distinctly curved humerus, which is unknown in other pachypleurosaurs except Dianopachysaurus (Liu et al. 2011), and apparently similar cervical and dorsal vertebral numbers. Obliteration of the upper temporal fenestrae would be an important autapomorphy of a new taxon, but the specimens are too poorly preserved to allow an unequivocal assessment of this trait. We also are unable to confidently include the Myanmar specimens in a phylogenetic analysis at this time, as we feel we do not yet have enough morphological information from the limited fossils to render such an analysis informative. Preferring a phylogenetic approach to species delimitation, we therefore refrain from diagnosing a new species, due to the incompleteness of the specimens and the potential for finding new fossils with additional fieldwork. There has recently been a flourishing of phylogenetic studies of basal eosauropterygian relationships, some of which have questioned the monophyly of Pachypleurosauria and other traditional clades (e.g., Rieppel et al. 2002; Holmes et al. 2008; Liu et al. 2011; Shang et al. 2011, 2017; Neenan et al. 2013; Jiang et al. 2014; Sato et al. 2014; Shang and Li 2015; Cheng et al. 2016). Addressing this debate is outside the scope of this paper, but additional material of the Myanmar pachypleurosaur, analysed in a phylogenetic context, may contribute important new insights in the future.
The presence of pachypleurosaurs in Myanmar is important for several reasons. First, these reptiles are known only from the Triassic, so they are a key piece of evidence that the Nwabangyi Dolomite Formation and coeval rocks in northern Shan State, whose ages have long been the subject of uncertainty, are Triassic in age. Second, there is a possibility that the Myanmar specimens may be among the oldest pachypleurosaurs in the world, which might give insight into the origins of the group and potentially help untangle the thorny phylogenetic relationships of basal eosauropterygians. The current oldest unequivocal pachypleurosaurs are Anisian (Liu et al. 2011), but if the Nwabangyi Dolomite Formation is indeed correlated with the Thigaungtaung Limestone of southern Shan State, then it is possible that the Myanmar taxon is anywhere from Induan to Anisian in age. Third, if the Myanmar taxon is among the oldest pachypleurosaurs, it might corroborate the often-discussed hypothesis of an eastern Tethyan origin for the group (e.g., Rieppel 1999, 2000; Rieppel and Hagdorn 1997), or alternatively, pending its inclusion in a phylogenetic analysis, hint that the fossil record is currently too poor to strongly support any biogeographic scenario.
Finally, the discovery of articulated reptilian material in the Triassic of Myanmar, although currently limited to these two specimens, suggests that fine preservation of aquatic and nearshore reptiles may be common in particular units, but has gone unrecognized due to limited collecting. The fossil-bearing Burmese rocks are not particularly far geographically from the Lagerstätten (conservation deposits) of Guizhou and Yunnan in southwestern China, which over the past two decades have yielded a wealth of spectacularly preserved Triassic reptiles (e.g., Wang et al. 2008; Zhang et al. 2008). There may be potential for similar Lagerstätten in Myanmar.
Acknowledgements.—We thank Maung Maung Naing (Rector of Yadanabon University, Amarapura, Myanmar) for his kind help, encouragement, and suggestions. We also thank James Neenan (University of Oxford, UK) and an anonymous referee for their helpful reviews. DF and SLB were funded by a Marie Curie Career Integration Grant (CIG 630652).
References
Amos, B.J. 1975. Stratigraphy of some of the Upper Paleozoic and Mesozoic carbonate rocks of the Eastern Highlands, Burma. Newsletter on Stratigraphy 4: 49–70. Crossref
Brönnimann, P., Whittaker, J.E., and Zennetti, L. 1975. Triassic foraminiferal biostratigraphy of the Kyaukme-Lonhtawkno area, Northern Shan State, Burma. Rivista Italiana di Paleontologia e Stratigrafia 81: 1–30.
Carroll, R.L., and Gaskill, P. 1985. The nothosaur Pachypleurosaurus and the origin of plesiosaurs. Philosophical Transactions of the Royal Society of London B 309: 343–393. Crossref
Cheng, Y.-N., Holmes, R., Wu, X.-C., and Alfonso, N. 2009. Sexual dimorphism and life history of Keichousaurus hui (Reptilia: Sauropterygia). Journal of Vertebrate Paleontology 29: 401–408. Crossref
Cheng, Y.-N., Wu, X.-C., and Ji, Q. 2004. Triassic marine reptile gave birth to live young. Nature 432: 383–386. Crossref
Cheng, Y.N., Wu, X.-C., Sato, T., and Shan, H.Y. 2016. Dawazisaurus brevis, a new eosauropterygian from the Middle Triassic of Yunnan, China. Acta Geologica Sinica 90: 401–424. Crossref
Chhibber, H.L. 1934. Geology of Burma. Macmillan, London.
Garson, M.S., Amos, B.J., and Mitchell, A.H.G. 1976. The geology of the area around Neyaungga and Yengan, southern Shan State, Burma. Overseas Memoir Institute of Geological Sciences 2: 19–30.
Holmes, R., Cheng, Y.-N., and Wu, X.-C. 2008. New information on the skull of Keichosaurus hui (Reptilia, Sauropterygia) with comments on sauropterygian interrelationships. Journal of Vertebrate Paleontology 28: 76–84. Crossref
Jiang, D.Y., Motani, R., Tintori, A., Rieppel, O., Chen, G.B., Huang, J.D., Zhang, R., Sun, Z.Y., and Ji, C. 2014. The Early Triassic eosauropterygian Majiashanosaurus discocoracoidis, gen. et sp. nov. (Reptilia, Sauropterygia), from Chaohu, Anhui Province, People’s Republic of China. Journal of Vertebrate Paleontology 34: 1044–1052. Crossref
Klein, N. 2009. Skull morphology of Anarosaurus heterodontus (Reptilia: Sauropterygia: Pachypleurosauria) from the lower Muschelkalk of the Germanic Basin (Winterswijk, the Netherlands). Journal of Vertebrate Paleontology 29: 665–676. Crossref
Klein, N. 2012. Postcranial morphology and growth of the pachypleurosaur Anarosaurus heterodontus (Sauropterygia) from the lower Muschelkalk of Winterswiik, The Netherlands. Paläontologische Zeitschrift 86: 389–408. Crossref
Lin, K. and Rieppel, O. 1998. Functional morphology and ontogeny of Keichousaurus hui (Reptilia, Sauropterygia). Fieldiana (Geology) N.S. 39: 1–35. Crossref
Liu, J., Rieppel, O., Jiang, D.Y., Aitchison, J.C., Motani, R., Zhang, Q.Y., Zhou, C.Y., and Sun, Y.Y. 2011. A new pachypleurosaur (Reptilia: Sauropterygia) from the lower Middle Triassic of southwestern China and the phylogenetic relationships of Chinese pachypleurosaurs. Journal of Vertebrate Paleontology 31: 291–302. Crossref
Maung Thein 1973. A preliminary synthesis of the geological evolution of Burma with reference to the tectonic development of Southeast Asia. Geological Society of Malaysia Bulletin 6: 87–116.
Neenan, J.M., Klein, N., and Scheyer, T.M. 2013. European origin of placodont marine reptiles and the evolution of crushing dentition in Placodontia. Nature Communications 4: 1621. Crossref
Rieppel, O. 1989. A new pachypleurosaur (Reptilia: Sauropterygia) from the Middle Triassic of Monte San Giorgio, Switzerland. Philosophical Transactions of the Royal Society of London B 323: 1–73. Crossref
Rieppel, O. 1993. Status of the pachypleurosauroid Psilotrachelosaurus toeplitschi Nopcsa (Reptilia, Sauropterygia), from the Middle Triassic of Austria. Fieldiana (Geology) N.S. 27: 1–17. Crossref
Rieppel, O. 1999. Phylogeny and paleobiogeography of Triassic Sauropterygia: problems resolved and unresolved. Palaeogeography, Palaeoclimatology, Palaeoecology 153: 1–15. Crossref
Rieppel, O. 2000. Sauropterygia I: Placodontia, Pachypleurosauria, Nothosauroidea, Pistosauroidea. Encyclopedia of Paleoherpetology, Part 12A. 134 pp. Friedrich Pfeil, Munich.
Rieppel, O. and Hagdorn, H. 1997. Paleobiogeography of Middle Triassic Sauropterygia in Central and Western Europe. In: J.M. Callaway and E.L.Nicholls (eds.), Sea Reptiles of the Past, 121–144. Academic Press, San Diego. Crossref
Rieppel, O. and Lin, K. 1995. Pachypleurosaurs (Reptilia: Sauropterygia) from the Lower Muschelkalk, and a review of the Pachypleurosauroidea. Fieldiana (Geology) N.S. 32: 1–44. Crossref
Rieppel, O., Sander, P.M., and Storrs, G.W. 2002. The skull of the pistosaur Augustasaurus from the Middle Triassic of northwestern Nevada. Journal of Vertebrate Paleontology 22: 577–592. Crossref
Sahni, M.R. 1936. On the geological age of the Namyau, Liu-Wun and Napeng Beds and the certain other formations in Indo China. Records of the Geological Survey of India 71: 217–230.
Sander, P.M. 1989. The pachypleurosaurids (Reptilia: Nothosauria) from the Middle Triassic of Monte San Giorgio, (Switzerland), with the description of a new species. Philosophical Transactions of the Royal Society of London B 325: 561–670. Crossref
Sato, T., Cheng, Y.-N., Wu, X.-C., and Shan, H.-Y. 2014. Diadongosaurus acutidentatus Shang, Wu & Li, 2011 (Diapsida: Sauropterygia) and the relationships of Chinese eosauropterygians. Geological Magazine 151: 121–133. Crossref
Shang, Q.H. and Li, C. 2015. A new small-sized eosauropterygian (Diapsida: Sauropterygia) from the Middle Triassic of Luoping, Yunnan, southwestern China. Vertebrata PalAsiatica 53: 265–280.
Shang, Q.H., Li, C., and Wu, X.C. 2017. New information on Dianmeisaurus gracilis Shang & Li, 2015. Vertebrata PalAsiatica 55: 145–161.
Shang, Q.H., Wu, X.C., and Li, C. 2011. A new eosauropterygian from the Middle Triassic of eastern Yunnan Province, southwestern China. Vertebrata PalAsiatica 49: 155–173.
Sues, H.-D. and Carroll, R.L. 1985. The pachypleurosaurid Dactylosaurus schroederi (Diapsdida: Sauropterygia). Canadian Journal of Earth Sciences 22: 1602–1608. Crossref
Wang, X.F., Bachmann, G.H., Hagdorn, H.D., Sander, P.M., Cuny, G., Chen, X.H., Wang, C., Lide, C., Long, C., Meng, F.S., and Xu, G. 2008. The Late Triassic black shales of the Guanling area, Guizhou Province, south-west China: a unique marine reptile and pelagic crinoid fossil Lagerstätte. Palaeontology 51: 27–61. Crossref
Win Swe 1972. Strike-slip faulting in Central Belt of Burma. In: N.S. Haile (ed.), Regional Conference on the Geology of South-East Asia, Kuala Lumpur. Geological Society of Malaysia Newsletter (Annex) 34: 59.
Win, Z., Shwe, K.K., and Yin, O.S. 2015. Sedimentary facies and biotic associations in the Permian–Triassic limestones on the Shan Plateau, Myanmar. Geological Society of London, Memoirs 48: 343–363. Crossref
Zhang, Q.Y., Zhou, C.Y., Lü, T., Xie, T., Lou, X.Y., Liu, W., Sun, Y.Y., and Jiang, X.S. 2008. Discovery and significance of the Middle Triassic Anisian Biota from Luoping, Yunnan Province. Geological Review 54: 523–526.
Khaing Khaing San [khaingkhaingsan2014@gmail.com], Geology Department, Yadanabon University, Amarapura, Myanmar.
Nicholas C. Fraser [nick.fraser@nms.ac.uk], Davide Foffa [davide.foffa@ed.ac.uk], Stephen L. Brusatte [stephen.brusatte@ed.ac.uk], School of GeoSciences, University of Edinburgh, Grant Institute, James Hutton Road, Edinburgh EH9 3FE, UK; National Museums of Scotland, Chambers Street, Edinburgh EH1 1JF, UK.
Olivier Rieppel [orieppel@fieldmuseum.org], Center of Integrative Research, The Field Museum, Chicago, Illinois 60605-2496, USA.
Received 9 January 2019, accepted 8 February 2019, available online 26 April 2019.
Copyright © 2019 Khaing Khaing San et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Acta Palaeontol. Pol. 64 (2): 357–362, 2019
https://doi.org/10.4202/app.00594.2019