
Sauropod diversity in the Upper Cretaceous Nemegt Formation of Mongolia—a possible new specimen of Nemegtosaurus
ALEXANDER O. AVERIANOV and ALEXEY V. LOPATIN
Averianov, A.O. and Lopatin A.V. 2019. Sauropod diversity in the Upper Cretaceous Nemegt Formation of Mongolia— a possible new specimen of Nemegtosaurus. Acta Palaeontologica Polonica 64 (2): 313–321.
Currently, there are two sauropod taxa known from the Upper Cretaceous (Maastrichtian) Nemegt Formation of Gobi Desert, Mongolia: Nemegtosaurus from the Nemegt locality and Opisthocoelicaudia from the Altan Uul IV locality. Both taxa are represented by not overlapping elements (skull and partial postcranial skeleton respectively), which arises question on their possible synonymy. Five articulated sauropod dorsal vertebrae (PIN 3837/P821, dorsals 6–10) were found in 1949 by the Mongolian Expedition of the Academy of Sciences of the USSR at the Nemegt locality. This specimen is similar to Opisthocoelicaudia in having a strong ventral ridge on dorsal centra, a low neural arch which is anteroposteriorly narrowest at the junction with the centrum and widens dorsally, and lack of hyposphene–hypantrum articulations. PIN 3837/P821 differs from Opisthocoelicaudia by having the less dorsoventrally flattened dorsal centra, a shallow ventral concavity of dorsal centra in lateral view, a vertical posterior centrodiapophyseal lamina (pcdl) in dorsals 8 and 9, a postzygodiapophyseal lamina (podl) that roofs the centrodiapophyseal fossa (pocdf), and strongly developed accessory laminae within the parapophyseal centrodiapophyseal fossa (pacdf). The sauropod femora from Nemegt Formation differ from the femur of Opisthocoelicaudia by the medial condyle extending more distally compared with the lateral condyle. Most likely these femora and PIN 3837/P821 belong to Nemegtosaurus, which would make this taxon distinct from Opisthocoelicaudia by discussed characters of dorsal vertebrae and femur.
Key words: Dinosauria, Sauropoda, Nemegtosaurus, Upper Cretaceous, Nemegt Formation, Mongolia.
Alexander O. Averianov [dzharakuduk@mail.ru], Zoological Institute of the Russian Academy of Sciences, Universitetskaya nab. 1, 199034 Saint Petersburg, Russia; Institute of Earth Sciences, Saint Petersburg State University, 16 Liniya VO 29, 199178 Saint Petersburg, Russia; Institute of Geology and Petroleum Technology, Kazan Federal University, Kremlevskaya str. 4/5, 420008 Kazan, Russia.
Alexey V. Lopatin [alopat@paleo.ru], Borissiak Paleontological Institute, Russian Academy of Sciences, Profsouznaya ul. 123, 117997 Moscow, Russia.
Received 10 January 2019, accepted 11 February 2019, available online 24 April 2019.
Copyright © 2019 A.O. Averianov and A.V. Lopatin. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
In spite of the intensive dinosaur records in the Cretaceous of Mongolia, sauropods remain there the least diverse and poorly known among the main dinosaurian groups. Until now there are five named sauropod taxa from the Cretaceous of Mongolia: Asiatosaurus mongoliensis (currently nomen dubium) from the Oshih locality, Lower Cretaceous (Osborn 1924), Erketu ellisoni from the Bor Guvé locality, ?Bayan Shireh Formation, lower Upper Cretaceous (Ksepka and Norell 2006, 2010), Quaesitosaurus orientalis from the Shar Tsav locality, Barun Goyot Formation, Campanian (Kurzanov and Bannikov 1983), Nemegtosaurus mongoliensis from the Nemegt locality, Nemegt Formation, Maastrichtian (Nowinski 1971; Wilson 2005), and Opisthocoelicaudia skarzynskii from the Altan Uul IV locality, Nemegt Formation, Maastrichtian (Borsuk-Białynicka 1977). Nemegtosaurus was based on an isolated skull while Opisthocoelicaudia on a skeleton lacking the skull and cervical vertebrae. Because the known skeletal parts of these two taxa do not overlap and because both of them come from the same stratigraphic units (Nemegt Formation) and localities separated by about 50 km, they were considered as possible synonyms (Currie et al. 2018). This idea was supported by discovery of some postcranial elements in the Nemegtosaurus quarry, possibly belonging to the holotype individual (Currie et al. 2018).
Abundant sauropod remains were mentioned in the narratives and preliminary reports of the Mongolian Paleontological Expedition carried out by the Moscow Paleontological Institute in 1946–1948 under the leadership of Ivan A. Efremov. These findings (Fig. 1) are reviewed in the next section. In particular, Efremov (1954a: 10) cited remains of a new genus of sauropod from an unspecified locality. Later in that paper (Efremov 1954a: 18) he mentioned new forms of sauropod with shortened hindlimbs from the Nemegt locality. These materials were never described and were not mentioned in the reviews of Mongolian sauropods (Maryańska 2000; Alifanov 2012). One of these specimens, the five articulated dorsal vertebrae collected at the Nemegt locality, is on display in the Orlov Paleontological Museum of the Borissiak Paleontological Institute in Moscow. This specimen was found few km aside from the Nemegtosaurus quarry and may belong to the same taxon. This specimen is described herein and its implications for the sauropod taxonomic diversity in the Nemegt Formation are discussed.
The nomenclature of the vertebral fossae and laminae follows Wilson (1999, 2012) and Wilson et al. (2011).
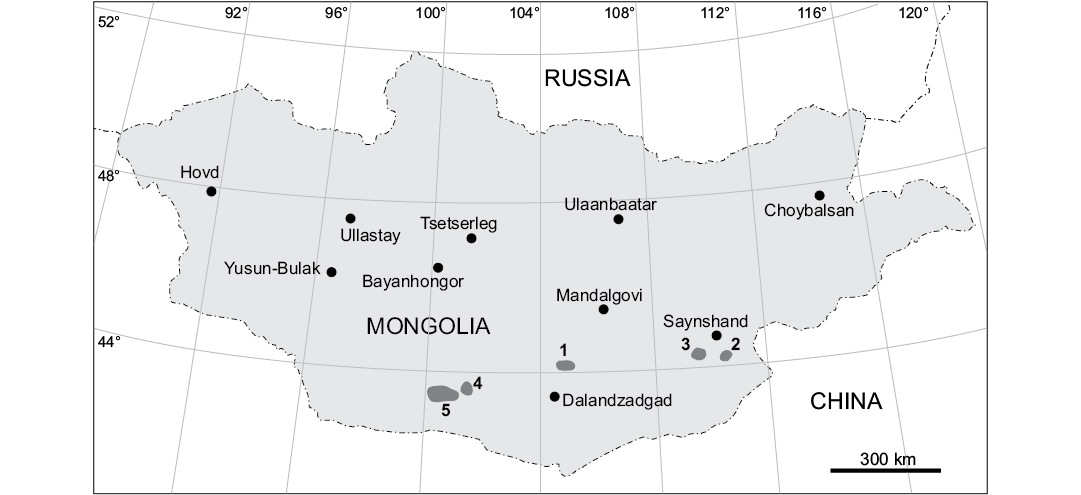
Fig. 1. Map of Mongolia showing position of the prospecting and excavation areas of the Mongolian Paleontological Expedition of the Academy of Sciences of the USSR in 1946–1949 that produced sauropod remains (1, Olgoy Ulan Tsav; 2, Khamarin Khural; 3, Khara Khutul; 4, Sheeregeen Gashoon; 5, Nemegt and Tsagan Khushuu). Modified after Efremov (1949) and Rozhdestvensky (1952).
Institutional abbreviations.—MPC-D, Institute of Paleontology and Geology of the Mongolian Academy of Sciences, Ulaanbaatar, Mongolia; PIN, Borissiak Paleontological Institute, Russian Academy of Sciences, Moscow, Russia; ZPAL, Institute of Paleobiology, Polish Academy of Sciences, Warsaw, Poland.
Other abbreviations.—acpl, anterior centroparapophyseal lamina; cpaf, centroparapophyseal fossa; cpol, centropostzygapophyseal lamina; cprf, centroprezygapophyseal fossa; cprl, centroprezygapophyseal lamina; pacdf, parapophyseal centrodiapophyseal fossa; pacprf, parapophyseal centroprezygapophyseal fossa; pcdl, posterior centrodiapophyseal; pcpl, posterior centroparapophyseal lamina; pocdf, postzygapophyseal centrodiapophyseal fossa; podl, postzygodiapophyseal lamina; ppdl, paradiapophyseal lamina; prdl, prezygodiapophyseal lamina; prpl, prezygoparapophyseal lamina; tprl, intraprezygapophyseal lamina.
Review of sauropod remains discovered by the Mongolian Paleontological Expedition of the Academy of Sciences of the USSR in 1946–1948
Olgoy Ulan Tsav.—Northern Gobi, 160 km north to Dalanzadgad. Barunbayan Formation, Lower Cretaceous (Aptian–Albian) (Shuvalov 2000). An isolated hill of Cretaceous red-beds within the metamorphosed pre-Mesozoic rocks. Well preserved sauropod bones, sometimes in associations, were found in September 1946 in a sandstone 30–35 m below the top of the hill (Efremov 1949: 7). Worse preserved sauropod bones come from the clay below and above this fossiliferous level. A partial sauropod skeleton, including sacrum, pelvis, and caudal vertebrae, was preliminarily excavated in a hard sandstone on the northern side of the hill (Efremov 1948a: 51; 1949: pl. 1: 2). Only two huge anterior caudal vertebrae have been collected from this specimen. Abundant sauropod remains were cited for this locality (Efremov 1954b: 130; 1957: 90).
Khamarin Khural.—Eastern Gobi, 50 km SSW of Sainshand. Khukhtyk Formation, Lower Cretaceous (Aptian–Albian). Associated sauropod bones were discovered by Soviet geologists in red and yellow sand and sandstone (Efremov 1949: 15).
Khara Khutul.—Eastern Gobi, 70 km NNW to Bayn Shireh. Lower part of Bayan Shireh Formation (Upper Cretaceous, Cenomanian–Turonian). Sauropod bones were found in October 1946 and April 1948 in gray and yellow sand above the basalt layer (Efremov 1949: 18; Rozhdestvensky 1952: 12). A sauropod pelvis was found in April 1948 in greenish-gray sandstone below the basalt layer (Rozhdestvensky 1969: 53). Abundant sauropod have been mentioned (Efremov 1954a: 10; 1954b: 130; 1957: 89).
Sheeregeen Gashoon.—Trans-Altai Gobi, 350 km west to Dalanzadgad. “Sheeregeen Gashoon beds”, a lateral equivalent of the upper part of Bayan Shireh Formation (Upper Cretaceous, Turonian–Santonian). A series of low hills at the bottom of Sheeregeen Gashoon Depression. Abundant large bones of sauropods (Efremov 1949: 14) or sauropod remains (Maleev 1952: 893; 1956: 89) have been noted.
Nemegt.—350 km WSW to Dalanzadgad. Nemegt Formation (Upper Cretaceous, Maastrichtian). Scattered sauropod limb bones, including a gigantic femur, were found in September–October 1946 (Efremov 1948b; 1949: 12, pl. 3: 1). The sauropod vertebra labelled “caudal vertebra” and figured in Efremov (1948b) is a partial dorsal vertebra. Humeri, femora, ribs, and fragments of vertebral column of sauropods were mentioned by Efremov (1955: 797). Unspecified sauropod bones were noted by Rozhdestvensky (1969: 80). Some bones attributed previously to Therizinosaurus (Nemegt, Quarry V) belong to sauropods (Rozhdestvensky 1969: 82). Part of a vertebral column of a sauropod was found between Nemegt and Altan Ula (= Altan Uul) in May 1948 (Rozhdestvensky 1969: 87). Rare and mostly disarticulated sauropod remains were mentioned from the Nemegt locality (Efremov 1954a: 18; 1954b: 130). The femur from Nemegt has a prominent lateral bulge that is generally similar to that of Opisthocoelicaudia (Borsuk-Białynicka 1977), but differs in having the medial (tibial) condyle extending more distally compared with the lateral (fibular) condyle (the opposite is characteristic for the femur in Opisthocoelicaudia) (Efremov 1949: pl. 3: 1). The specimen described in this paper, PIN 3837/P821 was found in 1949 in a close proximity to the above mentioned femur in the excavation field near the Central Camp of the Mongolian Paleontological Expedition of the Academy of Sciences of the USSR. These were indicated as items 5 and 6 on the excavation map published by Efremov (1955: fig. 2). Currently the Nemegt locality represents a vast area, approximately 5 × 5 km, where the dozens of dinosaur skeletal remains are concentrated (Gradziński et al. 1968: fig. 2; Gradziński and Jerzykiewicz 1972: fig. 1; Funston et al. 2018: fig. 1).
Tsaagan Khushuu (= Tsaagan Uul).—Several km west to Nemegt locality. Sauropod remains found more frequently than in Nemegt locality were cited for the Tsaagan Khushuu locality (Rozhdestvensky 1952: 18; Efremov 1955: 801).
Description of PIN 3837/P821
The specimen PIN 3837/P821 consists of five articulated middle-posterior dorsal vertebrae from the Nemegt locality. By comparison with dorsal series in Diplodocus carnegii, Euhelopus, and Opisthocoelicaudia (Hatcher 1901; Borsuk-Białynicka 1977; Wilson and Upchurch 2009), the preserved vertebrae could be dorsals 6 to 10. In sauropods the cervicodorsal transition occurs between the last three cervical vertebrae and first four dorsal vertebrae (Wilson 1999). In particular, in the first four dorsal vertebrae the parapophysis is placed on the junction of centrum and neural arch. In dorsal 5 it is placed on the neural arch but closer to the centrum than to the diapophysis. The first preserved vertebrae in PIN 3837/P821 can be identified as dorsal 6 because of a relatively large parapophysis which is placed adjacent to the prezygapophysis.
The dorsals 6–9 preserve partially the neural arch. Only parapophysis and acpl are preserved from the neural arch of the dorsal 10. The diapophysis of dorsals 9 and the posterior articular surface of the centrum of dorsal 10, seen in a historical photograph (Fig. 2A1), are now missing.
The centra are of similar length, while the posterior centrum height increases from dorsal 6 to 10 (Table 1). The centra are opisthocoelous, with a deep ball-like anterior articular surface and concave posterior articular surface. The centra are spool shaped, deeply constricted laterally. The posterior articular surface is transversely wider than the anterior articular surface. The dorsoventral flattening of the centrum is maximal in dorsal 6 (PCH/PCW = 0.80; Table 1). This flattening decreases in more posterior dorsals (Table 1). The ventral profile of the centrum is shallowly concave. There is a distinct ridge along the midline of the centrum ventral surface. This ridge is flanked laterally by a depressed surface, bordered laterally by a ridge extending for the most of the centrum length. These lateral depressions and ridges are better developed on dorsals 6–8 compared with the dorsals 9–10 (Fig. 2A2, A3).
Table 1. Measurements (in cm) of dorsal vertebrae in PIN 3837/P821. Abbreviations: ACH, anterior height of centrum; ACW, anterior width of centrum; CL, centrum length (without ball); PCH, posterior height of centrum; PCW, posterior width of centrum.
| |
CL |
ACH |
ACW |
PCH |
PCW |
PCH/PCW |
|
dorsal 6 |
21.9 |
17.3 |
21.5 |
18.6 |
23.4 |
0.80 |
|
dorsal 7 |
23.4 |
17.2 |
21.5 |
22.3 |
23.0 |
0.97 |
|
dorsal 8 |
22.1 |
20.1 |
21.3 |
22.6 |
22.6 |
1.00 |
|
dorsal 9 |
21.9 |
20.7 |
20.8 |
23.5 |
22.3 |
1.05 |
|
dorsal 10 |
– |
21.2 |
21.2 |
– |
– |
– |
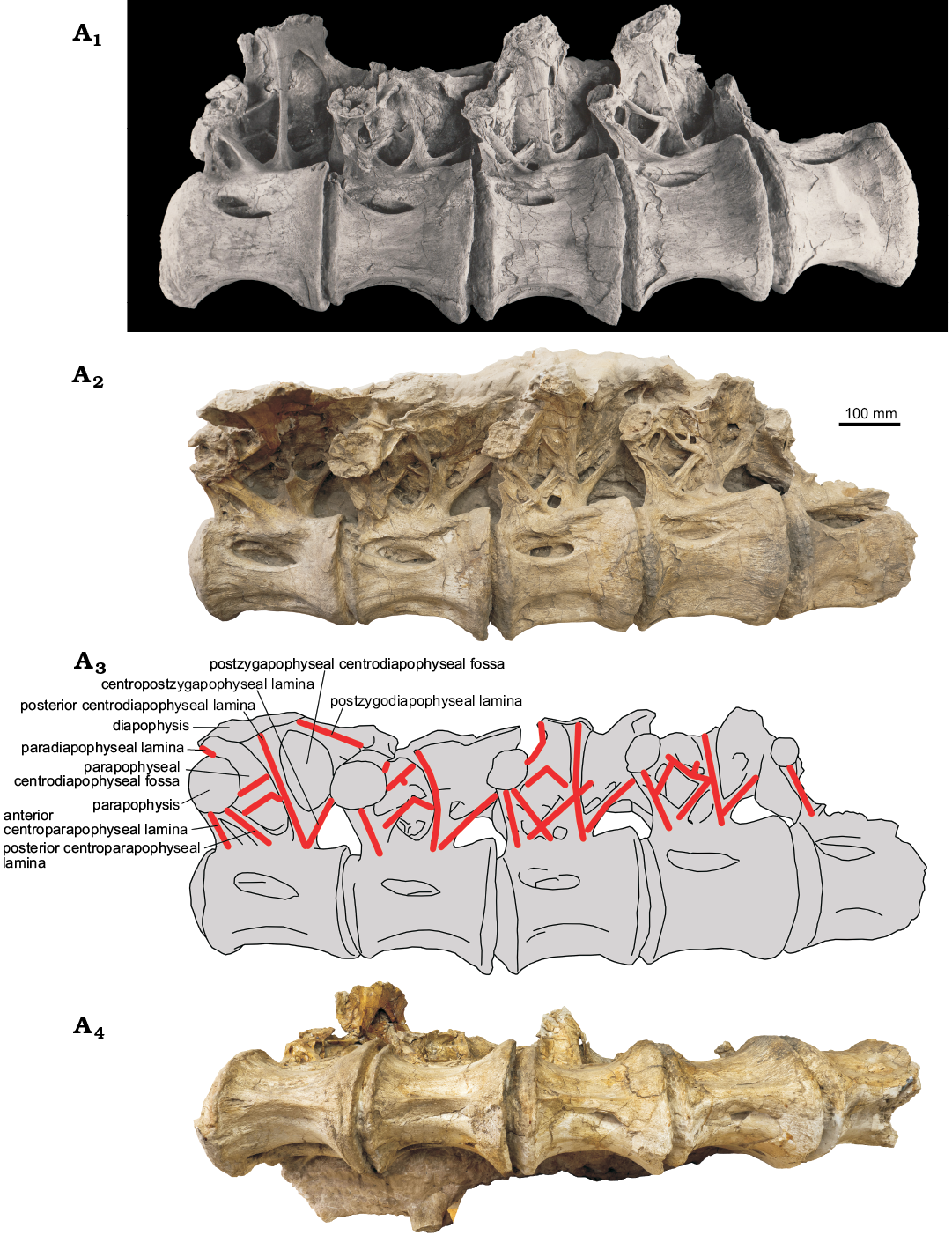
Fig. 2. Sauropod PIN 3837/P821 from Nemegt, Gobi Desert, Mongolia; Nemegt Formation, Maastrichtian (Upper Cretaceous); dorsal vertebrae 6–10 in lateral view (A1, photo from PIN archive, A2, the actual photo, A3, explanatory drawing) and in ventral view (A4).
A large spindle-shaped pleurocoel occupies about half of the centrum length. The pleurocoel is round anteriorly and pointed posteriorly in dorsals 6 and 7, round on both ends in dorsal 8, and pointed on both ends in dorsals 9 and 10. The pleurocoel is subdivided by a variable developed longitudinal lamina. The anterior articular surface of the centrum of the dorsal 6 is wider than high, with slightly concave dorsal margin (Fig. 3).
The neural arch occupies about two thirds of the centrum length in lateral view (Fig. 2). Its anterior margin is almost level with the centrum anterior margin, excluding the ball. In lateral view the neural arch is anteroposteriorly shortest at the neurocentral suture and widens in dorsal direction. This contrasts with the dorsal neural arches of many sauropods which have the narrowest neural arch some distance dorsal to the neurocentral suture (Janensch 1950: figs. 56, 62; Powell 1992: fig. 15; Curry Rogers 2009: fig. 21E; Coria et al. 2013: fig. 4A). The height of the neural arch at the diapopyseal articular surface is only slightly greater than the height of the centrum.
The prezygapophyses are best exposed on the dorsal 6 (Fig. 3). The prezygapophyseal articular surface is mediolaterally wide and slightly convex. This surface is oriented to approximately 30° to the horizontal line. The medial ends of both prezygapophyses are widely separated. The postzygapophysis is preserved only on dorsal 6 where it is articulating with the prezygapophysis of dorsal 7 (Fig. 2). It seems that the entire prezygapophyseal articular surface is placed beyond the posterior articular surface of the centrum.
The parapophysis is adjacent to the prezygapophysis. The dorsal margins of their articulated surfaces are nearly contacting. The parapophyseal articular surface is round to oval shaped. It is large on dorsals 6 and 7, distinctly smaller on dorsals 8 and 9, and very small on dorsal 10. The diapophyseal articular surface is anteroposteriorly elongate and dorsoventrally narrow.
The pcdl is the longest and most robust lamina on the neural arch. It extends between the diapophyseal articular surface (partially preserved on dorsal 8) and posteroventral corner of the neural arch. It is oriented anterodorsolaterally on the dorsals 6 and 7 and dorsolaterally on the dorsals 8 and 9. The ventral end of pcdl is not divided or bifurcating and is confluent with that of the cpol. On dorsal 7 there is a small oval depression at the base of pcdl.
There is no prdl because the prezygapophysis is almost completely (dorsal 7) or completely (remaining vertebrae) overlapped laterally by the parapophsyis, leaving no space for this lamina.
The podl extends between the diapophysis and postzygapophysis. It is preserved only on the dorsal 6 and better seen on the historical photograph (Fig. 2A1).
The ppdl joins the diapophysis and parapophysis. It is partially preserved (by its medial part) on dorsals 6–8 (Fig. 2A2, A3). In dorsal 6 this lamina is nearly horizontal (extended mediolaterally) because the diapophysis and parapophysis are placed on a similar vertical level. In dorsal 8 it is more vertical because the diapophysis is placed distinctly higher than the parapophysis.
The acpl joins the parapophysis and the anterior point of the neurocentral suture. It extends along the anterior margin of the neural arch. Dorsally this lamina is confluent with the pcpl, which connects the parapophysis with the middle of the neurocentral suture. On dorsals 6 and 8 both acpl and pcpl are similarly robust, with a deep cleft-like cpaf between them (Fig. 2A2, A3). On dorsals 7 and 9 acpl and pcpl are confluent along their entire length.
The prpl is a short strut between the parapophysis and prezygopophysis seen on the anterior side of dorsal 6 (Fig. 3). This lamina closes dorsally the pacprf.
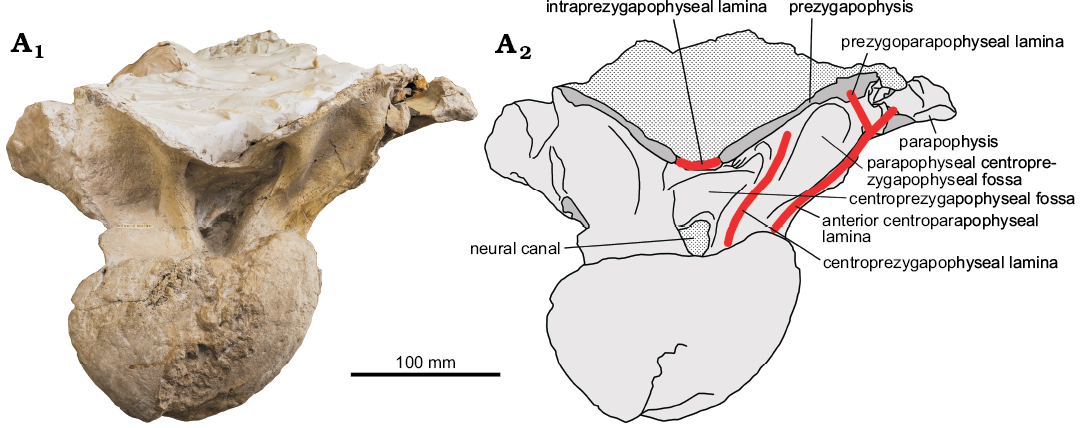
Fig. 3. Sauropod PIN 3837/P821 from Nemegt, Gobi Desert, Mongolia; Nemegt Formation, Maastrichtian (Upper Cretaceous); dorsal vertebra 6 in anterior view (A1, photo; A2, explanatory drawing).
The cprl is a robust strut between the ventrolateral aspect of the prezygapophysis and base of the neural arch. The cprl is directed dorsolaterally so the paired cprls form a V-shaped figure. Within the triangle formed by the cprls and tprl there is a deep cprf dorsally and a small opening of the neural canal ventrally. The lateral wall of the neural canal is formed by the cprl. Ventrally the cprl is almost confluent with the acpl. Both laminae diverge dorsally, with a distinct pacprf between them.
Between the widely separated medial ends of the prezygapophyses there is a tprl, which anterior margin is damaged. The apparent lack of a hypantral facet indicates that hyposphene-hypantrum articulation was absent in this region.
PIN 3837/P821 is peculiar in having accessory laminae within the pacdf. The configuration of these laminae differs between the vertebrae. On dorsal 6 these accessory laminae are best seen in the historical photograph providing a lateroventral view of pacdf (Fig. 2A1). There are two accessory laminae directed posterodorsally. The dorsal lamina connects the parapophysis with the middle of pcdl. The shorter ventral accessory lamina starts at the middle of pcpl and parallels the dorsal accessory lamina. It meets at a right angle a shorter lamina extending anterodorsally from a ventral part of pcdl. A similar pattern of accessory laminae is present on dorsal 7 (Fig. 2A2, A3). However, in this vertebra, the space between the two accessory laminae is very shallow, with two distinct depressions at the anterior and posterior ends of the dorsal accessory lamina. The internal margins of these depressions can be considered as two additional poorly developed accessory laminae (Fig. 2A2, A3). In dorsal 8 the pattern of the accessory lamina is opposed to that of dorsal 6. The dorsal accessory lamina meets at the right angle a short lamina emanating from pcdl, while the ventral accessory lamina directly connects to the pcdl (Fig. 2A2, A3). The ventral accessory lamina is longer than in other vertebrae. It intersects pcpl and pcdl and enters the cpaf and pocdf (Fig. 2A2, A3). The pattern of the accessory laminae in dorsal 9 is completely different from that in the previous vertebrae. The two accessory laminae, starting from the parapophysis (dorsal lamina) and middle of the pcpl (ventral lamina), are not parallel each other but meet at a right angle in a ventral part of the pcdl (Fig. 2A2, A3). These two accessory laminae and pcdl form a “K” laminae pattern, described previously for Euhelopus (see discussion). There are also two smaller other accessory laminae. The longer of these two is between the pcpl and the dorsal accessory lamina, close to the parapophysis and subparallel to the ventral accessory lamina (Fig. 2A3). A short accessory lamina is emanating from the dorsal accessory lamina, near its junction with the pcdl, and is directed towards the pcdl but does not reach it (Fig. 2A2, A3).
The broken posterior part of dorsal 10 reveals a camellate bone texture with the cells of 1–3 cm size.
Comparison with Opisthocoelicaudia
PIN 3837/P821 is similar with Opisthocoelicaudia in size, in having a relatively low neural arch in middle-posterior dorsal vertebrae, small neural canal, and apparent lack of hyposphene–hypantrum articulations. The low neural arch of PIN 3837/P821 and Opisthocoelicaudia is narrowest anteroposteriorly at the junction with the centrum and widens dorsally, while in many other sauropods the higher neural arch has a narrowest point at the midheight. Wilson (2002) considered presence of the pcpl in middle and posterior dorsal neural arches as an autapomorphy of Opisthocoelicaudia. This lamina is well developed at least in dorsal 6 (Borsuk-Białynicka 1977: fig. 3A6). In PIN 3837/P821 the pcpl is present on dorsals 6 and 8 and it is confluent with acpl on dorsals 7 and 9. D’Emic (2012) listed divided pcpl on middle dorsal vertebra as an autapomorhy for Opisthocoelicaudia, but we see no evidence for this.
In Opisthocoelicaudia the dorsoventral flattening of the centra increases in the posterior part of dorsal series due to the broadening of the centra in that region and constant centrum height (Borsuk-Białynicka 1977: 9). In dorsals 6–8 the centrum height to centrum width ratio is 0.60–0.64. This ratio is 0.47 in dorsal 9 and 0.52 in dorsal 10. In PIN 3837/P821 the posterior centrum width is constant while the posterior centrum height increases from dorsal 6 to dorsal 9 (Table 1). This result in decreasing of the dorsoventral flattening from dorsal 6 to dorsal 9 (PCH/PCW changes from 0.80 to 1.05; Table 1).
In Opisthocoelicaudia the ventral concavity of the centrum in dorsal vertebrae is delimited by two ventrolateral crests and divided by a midline ridge (Borsuk-Białynicka 1977: 9). This concavity becomes increasingly deeper towards the posterior dorsals. In PIN 3837/P821, the ventral profile of the centrum is much less concave, with a strong ventral ridge flanked by ventrolateral ridges and depressions between them. Thus, the construction of the ventral centrum surface in dorsals of Opisthocoelicaudia and PIN 3837/P821 is quite similar. The difference is that the ventral concavity is emphasized in Opisthocoelicaudia and ventral ridge is emphasized in PIN 3837/P821. This results in a much less concave ventral profile of the centrum in PIN 3837/P821. A concave ventral surface that bears a ventral midline keel in dorsal and sacral vertebrae has been considered a diagnostic character of Opisthocoelicaudia (Upchurch et al. 2004; D’Emic 2012).
In Opisthocoelicaudia the pcdl is anterodorsally oriented in all dorsals. In PIN 3837/P821 this is true for the dorsal 6 and 7 while in the dorsals 8 and 9 pcdl is almost vertical.
In Opisthocoelicaudia the podl is absent and pocdf is open dorsally. The absence of podl has been considered an autapomorphy of Opisthocoelicaudia (Wilson 2002). In PIN 3837/P821 this lamina is well developed at least in dorsal 6 (this region is damaged in other dorsals), roofing the pocdf.
In PIN 3837/P821 the pacprf on the anterior aspect of the neural arch, bordered by cprl, acpl, and prpl, is undivided (this side can be seen only in dorsal 6). This is true for the dorsal vertebrae in Opisthocoelicaudia except the dorsal 6, where pacprf is divided by a middle accessory lamina which is perpendicular to the prezygapophysis and acpl (Borsuk-Białynicka 1977: fig. 3A3).
The accessory laminae within the pacdf, well pronounced in PIN 3837/P821, are poorly developed in Opisthcoelicaudia. The ventral accessory lamina can be seen only on the left side of cervical 7 (Borsuk-Białynicka 1977: pl. 3: 1). The other accessory laminae are absent.
Although PIN 3837/P821 shares some common features with Opisthocoelicaudia, it clearly belongs to a different genus which is characterized by less dorsoventrally flattened dorsal centra, shallow ventral concavity of dorsal centra in lateral view, vertical pcdl in dorsals 8 and 9, presence of podl that closes pocdf dorsally, and strongly developed accessory laminae within the pacdf.
Diagnostic characters of PIN 3837/P821
Centrum articulation.—The posterior dorsal centra are generally amphicoelous to platycoelous in non-macronarian sauropods; all presacral vertebrae are strongly opisthocoelous in Macronaria (Wilson and Sereno 1998). In PIN 3837/P821 the centra are strongly opisthocoelous, supporting its attribution to the Macronaria.
Shape of pleurocoels.—In PIN 3837/P821 the pleurocoels are relatively low, with acuminate posterior, or both ends. These spindle-shaped, or eye-shaped pleurocoels of dorsal vertebrae have been considered a synapomorphy of Titanosauria (Salgado et al. 1997). In non-titanosaurian neosauropods the pleurocoels are oval to subcircular in shape.
Hyposphene-hypantrum articulations.—Hyposphene–hypantrum articulation is present in the middle and posterior dorsal vertebrae of most sauropods, except Lithostrotia (Salgado et al. 1997; Upchurch et al. 2004; Apesteguía 2005). In PIN 3837/P821 the hyposphene is lacking on the dorsal 6 and most likely is absent on more posterior dorsals, which supports its attribution to the Lithostrotia.
Posterior centroparapophyseal lamina.—Presence of this lamina has been considered a synapomorphy for Titanosauria (Salgado et al. 1997), or Titanosauriformes (Wilson 1999, 2002). The pcpl was independently acquired in Diplodocidae (Wilson 2002). At the same time Wilson (2002) considered presence of pcpl as an autapomorphy for Opisthocoelicaudia (see the previous chapter). The pcpl is present on dorsals 6 and 8 of PIN 3837/P821, which may support its attribution to the Titanosauriformes or Titanosauria.
Accessory laminae.—PIN 3837/P821 has a rare pattern of accessory laminae within the pacdf. The homology of these laminae is difficult to interpret. The dorsal 9 has “K” laminae cross pattern found in dorsal 5 and more posterior dorsal of Euhelopus that was considered a diagnostic feature for that taxon (Wilson and Upchurch 2009). Wilson and Upchurch (2009) proposed homology of these laminae based on laminae intersection: pcdl (vertical portion of “K”), pcpl (upper arm of “K”), and acdl (lower arm of “K”). While interpretation of pcdl is beyond the question in PIN 3837/P821, interpretation of the two other laminae of “K” is less certain. The dorsal accessory lamina cannot be considered the pcpl. In more anterior dorsals (6–8) it starts from the parpophysis and directed posterodorsally, while pcpl is directed posteroventrally towards the posterolateral aspect of the centrum. The ventral accessory lamina in dorsals 6–8 is very short and does not connect the diapophysis; it can be homologized with the acdl. Moreover, acdl does not exist on dorsal vertebrae as it is interrupted by the parapophsis and separates into two distinct laminae, acpl and ppdl (Wilson 1999). Thus these laminae are simple referred here as accessory laminae. In Diplodocus a similar “K” laminae patter is present on dorsal 13, while on dorsals 10–12 there is only ventral accessory laminae (Osborn 1899: fig.7). On dorsal 13 of this specimen there is an additional accessory lamina that intersects the ventral accessory lamina. The “K” laminae pattern is also developed in dorsals 8 and 9 of Neuquensaurus (Salgado et al. 2005: fig. 4B, C). The laminae pattern in dorsal 9 of that taxon is especially similar with that in dorsal 9 of PIN 3837/P821 in having an additional lamina between acpl + pcpl and upper arm of “K”.
Internal bone structure.—In PIN 3837/P821 the bone structure is composed of large chambers, about 1–3 cm in size, separated by thin bony plates. Such camellate bony texture is a synapomorphy for the clade Somphospondyli (Wilson and Sereno 1998; Upchurch et al. 2004).
Sauropod diversity in the Nemegt Formation
Currently there are two sauropod taxa described from the Upper Cretaceous (Maastrichtian) Nemegt Formation in Gobi Desert, Mongolia: Nemegtosaurus mongoliensis, based on a skull found in Central Sayr at the Nemegt locality (Nowiński 1971) and Opisthocoelicaudia skarzynskii based on a skeleton lacking the skull and cervical vertebrae from Altan Uul IV locality (Borsuk-Białynicka 1977). Currie et al. (2018) rediscovered the original quarry of Nemegtosaurus and found some postcranial elements which may belong to the holotype individual (caudal or sacral centrum, femur, tibia, fibula, astragalus, and pedal ungual phalanx). These authors also reported seven additional sites within the Nemegt locality producing sauropod remains. Currie et al. (2018) concluded that a single sauropod taxon is present inthe Nemegt Formation and Opisthocoelicaudia skarzynskii is a junior subjective synonym of Nemegtosaurus mongoliensis. However, this conclusion cannot be followed because there are some problems with interpretation of the referred postcranial elements. The centrum was interpreted as “clearly opisthocoelous” but this specimen does not present any features indicative of its anterior-posterior orientation. Interpretation of this centrum as procoelous is equally possible. Currie et al. (2018: fig. 7) compared the femora of Nemegtosaurus (MPC-D100/413) and Opisthocoelicaudia (MPC-D100/404). The latter specimen was cited as the holotype of O. skarzynskii, originally published under the number ZPAL MgD-I/48 (Borsuk-Białynicka 1977). However, this specimen is markedly different from the femur of Opisthocoelicaudia published by Borsuk-Białynicka (1977: fig. 15, pl. 13: 1) and certainly does not belong to the holotype. It differs from the holotype femur by much shorter femoral neck and the opposite relative size of the distal condyles. In MPC-D100/404 and in MPC-D100/413 (Nemegtosaurus) the medial condyle protrudes more distally compared with the lateral condyle while the opposite is characteristic for the femur of Opisthocoelicaudia. Moreover, the femur MPC-D100/413 differs from that in Opisthocoelicaudia by a more dorsal position of the fourth trochanter (Borsuk-Białynicka 1977: fig. 15A; Currie et al. 2018: fig. 7B). A femur similar to MPC-D100/413 with the medial condyle extending more distally compared with the lateral condyle, was found at the Nemegt locality by the Mongolian Paleontological Expedition of the Academy of Sciences of the USSR (Efremov 1949: pl. 3: 1; Fig. 4B). This morphology of femur, distinct from that of Opistocoelicaudia, might be diagnostic for Nemegtosaurus. The part of vertebral column from the Nemegt locality described herein (PIN 3837/P821) is distinct from Opisthocoelicaudia. This unequivocally indicates the presence of two sauropod taxa in Nemegt Formation, likely Nemegtosaurus at the Nemegt locality and Opisthocoelicaudia at the Altan Uul IV locality.
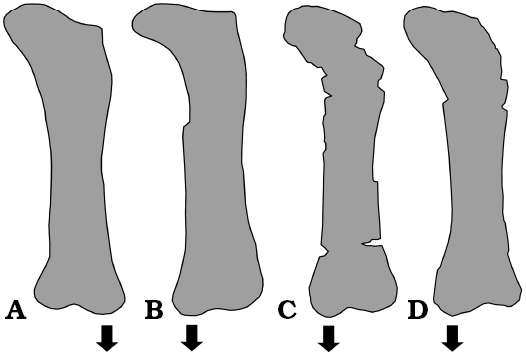
Fig. 4. Outlines of sauropod femora from the Upper Cretaceous (Maastrichtian) Nemegt Formation of Gobi Desert, Mongolia. A. ZPAL MgD-I/48, right femur in posterior view, the holotype of Opisthocoelicaudia skarzynskii Borsuk-Białynicka, 1977, modified from Borsuk-Białynicka (1977: fig. 15). B. Specimen collected by the Mongolian Paleontological Expedition of the Academy of Sciences of the USSR at Nemegt locality (current deposition unknown), right femur in anterior view (reversed), modified from Efremov (1949: pl. 3: 1). C. MPC-D100/413 attributed to Nemegtosaurus, right femur in posterior view, modified from Currie et al. (2018: fig. 7B). D. MPC-D100/404 (provenance unknown), left femur in anterior view, modified from Currie et al. (2018: fig. 7C). Arrows indicate the condyle protruding distally: lateral condyle in A and medial condyle in B–D. Not to scale.
Acknowledgements
We are grateful to Dmitry V. Grigoriev (Russian State Geological Prospecting University, Moscow, Russia) for photographing the vertebrae, to Konstantin K. Tarasenko (Borissiak Paleontological Institute, Moscow, Russia) for the assistance. We thank Philip Mannion (Department of Earth Science and Engineering, Imperial College London, London, UK) and Verónica Díez Díaz (Museum für Naturkunde, Leibniz Institute for Evolution and Biodiversity Science, Berlin, Germany) for reviewing the paper and providing useful comments. The laboratory research by AA received support from the Russian Scientific Fund (19-14-00020) and from the Zoological Institute, Russian Academy of Sciences (project АААА-А17-117022810195-3).
References
Alifanov, V.R. 2012. Superorder Dinosauria [in Russian]. In: E.N. Kuročkin and A.V. Lopatin (eds.), Iskopaemye pozvonočnye Rossii i sopredel’nyh stran. Iskopaemye reptilii i pticy. Čast 2, 153–309. GEOS, Moskva.
Apesteguía, S. 2005. Evolution of the hyposphene–hypantrum complex within Sauropoda. In: V. Tidwell and K. Carpenter (eds.), Thunder-Lizards. The Sauropodomorph Dinosaurs, 248–267. Indiana University Press, Bloomington.
Borsuk-Białynicka, M. 1977. A new camarasaurid sauropod Opisthocoelicaudia skarzynskii, gen. n., sp. n. from the Upper Cretaceous of Mongolia. Palaeontologia Polonica 37: 1–64.
Coria, R.A., Filippi, L.S., Chiappe, L.M., García, R.A., and Arcucci, A.B. 2013. Overosaurus paradasorum gen. et sp. nov., a new sauropod dinosaur (Titanosauria: Lithostrotia) from the Late Cretaceous of Neuquén, Patagonia, Argentina. Zootaxa 3683: 357–376. Crossref
Currie, P.J., Wilson, J.A., Fanti, F., Mainbayar, B., and Tsogtbaatar, K. 2018. Rediscovery of the type localities of the Late Cretaceous Mongolian sauropods Nemegtosaurus mongoliensis and Opisthocoelicaudia skarzynskii: Stratigraphic and taxonomic implications. Palaeogeography, Palaeoclimatology, Palaeoecology 494: 5–13. Crossref
Curry Rogers, K.A. 2009. The postcranial osteology of Rapetosaurus krausei (Sauropoda: Titanosauria) from the Late Cretaceous of Madagascar. Journal of Vertebrate Paleontology 29: 1046–1086. Crossref
D’Emic, M.D. 2012. The early evolution of titanosauriform sauropod dinosaurs. Zoological Journal of the Linnean Society 166: 624–671. Crossref
Efremov, I.A. 1948a. First Mongolian paleontological expedition of the Academy of Sciences of the USSR [in Russian]. Vestnik Akademii Nauk SSSR 1: 47–58.
Efremov, I.A. 1948b. In the footsteps of giant reptiles [in Russian]. Vokrug Sveta 1: 28–34.
Efremov, I.A. 1949. Preliminary results of the works of the I Mongolian paleontological expedition of the Academy of Sciences of the USSR of 1946 year [in Russian]. Trudy Mongol’skoj Komissii 38: 5–28.
Efremov, I.A. 1954a. Paleontological studies in the Mongolian People’s Republic (preliminary results of expeditions in 1946, 1948, and 1949) [in Russian]. Trudy Mongol’skoj Komissii 59: 3–32.
Efremov, I.A. 1954b. Some remarks on the questions of historical development of dinosaurs [in Russian]. Trudy Paleontologičeskogo Instituta AN SSSR 48: 125–141.
Efremov, I.A. 1955. Dinosaur burial in Nemegt (South Gobi, MPR) [in Russian]. In: N.S. Šackij (ed.), Voprosy geologii Azji. Tom 2, 789–809. Izdatel’stvo AN SSSR, Moskva.
Efremov, I.A. 1957. To the taphonomy of fossil faunas of terrestrial vertebrates of Mongolia [in Russian]. Vertebrata PalAsiatica 1 (2): 83–102.
Funston, G.F., Mendonca, S.E., Currie, P.J., and Barsbold, R. 2018. A dinosaur community composition dataset for the Late Cretaceous Nemegt Basin of Mongolia. Data in Brief 16: 660–666. Crossref
Gradziński, R. and Jerzykiewicz, T. 1972. Additional geographical and geological data from the Polish-Mongolian palaeontological expeditions. Palaeontologia Polonica 27: 17–30.
Gradziński, R., Kaźmierczak, J., and Lefeld, J. 1968. Geographical and geological data from the Polish-Mongolian palaeontological expeditions. Palaeontologia Polonica 19: 33–82.
Hatcher, J.B. 1901. Diplodocus (Marsh): Its osteology, taxonomy, and probable habits, with a restoration of the skeleton. Memoirs of the Carnegie Museum 1: 1–63. Crossref
Janensch, W. 1950. Die Wirbelsäule von Brachiosaurus brancai. Palaeontographica, Abteilung A: Paläozoologie, Stratigraphie 3 (Supplement 7 Reihe 1): 27–93.
Ksepka, D.T. and Norell, M.A. 2006. Erketu ellisoni, a long-necked sauropod from Bor Guvé (Dornogov Aimag, Mongolia). American Museum Novitates 3508: 1–16. Crossref
Ksepka, D.T. and Norell, M.A. 2010. The illusory evidence for Asian Brachiosauridae: new material of Erketu ellisoni and a phylogenetic reappraisal of basal Titanosauriformes. American Museum Novitates 3700: 1–27. Crossref
Kurzanov, S.M. and Bannikov, A.F. 1983. A new sauropod from the Upper Cretaceous of Mongolia [in Russian]. Paleontologičeskij žurnal 1983 (2): 90–96.
Maleev, E.A. 1952. Some comments on the geologic age and stratigraphic distribution of the armoured dinosaurs of Mongolia [in Russian]. Doklady Akademii Nauk SSSR 85: 893–896.
Maleev, E.A. 1956. Armored dinosaurs of the Upper Cretaceous of Mongolia [in Russian]. Trudy Paleontologičeskogo Instituta AN SSSR 62: 51–91.
Maryańska, T. 2000. Sauropods from Mongolia and the former Soviet Union. In: M.J. Benton, M.A. Shishkin, D.M. Unwin, and E.N. Kurochkin (eds.), The Age of Dinosaurs in Russia and Mongolia, 456–461. Cambridge University Press, Cambridge.
Nowiński, A. 1971. Nemegtosaurus mongoliensis n. gen., n. sp. (Sauropoda) from the uppermost Cretaceous of Mongolia. Palaeontologia Polonica 25: 57–81.
Osborn, H.F. 1899. A skeleton of Diplodocus. Memoirs of the American Museum of Natural History 1: 191–214.
Osborn, H.F. 1924. Sauropoda and Theropoda of the Lower Cretaceous of Mongolia. American Museum Novitates 128: 1–7.
Powell, J.E. 1992. Osteologia de Saltasaurus loricatus (Sauropoda–Titanosauroda) del Cretacico Superior del Noreostre Argentino. In: J.L. Sanz and A.D. Buscalioni (eds.), Los Dinosaurios y Su Entorno Biotico: Actas del Segundo Curso de Paleontologia in Cuenca, 165–230. Instituto “Juan de Valdes” Excmo, Cuenca.
Rozhdestvensky, A.K. [Roždestvenskij, A.K.] 1952. Issledovaniâ sovetskih paleontologov v Tsentral’noj Azji (Mongol’skaâ paleontologičeskaâ ekspediciâ Akademii nauk SSSR). 31 pp. Znanie, Moskva.
Rozhdestvensky, A.K. [Roždestvenskij, A.K.] 1969. Na poiski dinozavrov v Gobi. 293 pp. Nauka, Moskva.
Salgado, L., Apesteguia, S., and Heredia, S.E. 2005. A new specimen of Neuquensaurus australis, a Late Cretaceous saltasaurine titanosaur from North Patagonia. Journal of Vertebrate Paleontology 25: 623–634. Crossref
Salgado, L., Coria, R.A., and Calvo, J.O. 1997. Evolution of the titanosaurid sauropods I: Phylogenetic analysis based on the postcranial evidence. Ameghiniana 34: 3–32.
Shuvalov, V.F. 2000. The Cretaceous stratigraphy and palaeobiogeography of Mongolia. In: M.J. Benton, M.A. Shishkin, D.M. Unwin, and E.N. Kurochkin (eds.), The Age of Dinosaurs in Russia and Mongolia, 256–278. Cambridge University Press, Cambridge.
Upchurch, P., Barrett, P.M., and Dodson, P. 2004. Sauropoda. In: D.B. Weishampel, P. Dodson, and H. Osmólska (eds.), The Dinosauria. Second Edition, 259–322. University of California Press, Berkeley. Crossref
Wilson, J.A. 1999. A nomenclature for vertebral laminae in sauropods and other saurischian dinosaurs. Journal of Vertebrate Paleontology 19: 639–653. Crossref
Wilson, J.A. 2002. Sauropod dinosaur phylogeny: critique and cladistic analysis. Zoological Journal of the Linnean Society 136: 217–276. Crossref
Wilson, J.A. 2005. Redescription of the Mongolian sauropod Nemegtosaurus mongoliensis Nowinski (Dinosauria: Saurischia) and comments on Late Cretaceous sauropod diversity. Journal of Systematic Palaeontology 3: 283–318. Crossref
Wilson, J.A. 2012. New vertebral laminae and patterns of serial variation in vertebral laminae of sauropod dinosaurs. Contributions from the Museum of Paleontology, The University of Michigan 32 (7): 91–110.
Wilson, J.A. and Sereno, P.C. 1998. Early evolution and higher-level phylogeny of sauropod dinosaurs. Journal of Vertebrate Paleontology 18 (Supplement to 2): 1–72. Crossref
Wilson, J.A. and Upchurch, P. 2009. Redescription and reassessment of the phylogenetic affinities of Euhelopus zdanskyi (Dinosauria: Sauropoda) from the Early Cretaceous of China. Journal of Systematic Palaeontology 7: 199–239. Crossref
Wilson, J.A., D’Emic, M.D., Ikejiri, T., Moacdieh, E.M., and Whitlock, J.A. 2011. A nomenclature for vertebral fossae in sauropods and other saurischian dinosaurs. PLoS One 6 (2): e17114. Crossref
Acta Palaeontol. Pol. 64 (2): 313–321, 2019
https://doi.org/10.4202/app.00596.2019