
The last erythrosuchid—a revision of Chalishevia cothurnata from the late Middle Triassic of European Russia
RICHARD J. BUTLER, ANDREY G. SENNIKOV, MARTÍN D. EZCURRA, and DAVID J. GOWER
Butler, R.J., Sennikov, A.G., Ezcurra, M.D., and Gower, D.J. 2019. The last erythrosuchid—a revision of Chalishevia cothurnata from the late Middle Triassic of European Russia. Acta Palaeontologica Polonica 64 (4): 757–774.
Erythrosuchidae is a clade of early archosauriform reptiles that were large-bodied, hypercarnivorous, possibly apex predators in late Early and Middle Triassic ecosystems following the Permo-Triassic mass extinction. Chalishevia cothurnata from the late Middle Triassic (Ladinian) of Russia, is the stratigraphically youngest known erythrosuchid species, but the holotype and referred material of this taxon has received little study. Here, we provide the first detailed anatomical description of C. cothurnata, including comparisons to other erythrosuchids. Although known from relatively fragmentary material, the anatomy of C. cothurnata is distinctive, including an autapomorphic strongly slanted ventral border of the antorbital fossa. The presence of a large accessory opening (the “accessory antorbital fenestra”) in the skull between the premaxilla, nasal and maxilla, together with the inferred presence of a narrow postnarial process of the premaxilla that articulated with a slot on the nasal, provides strong evidence for a sister taxon relationship between C. cothurnata and the erythrosuchid Shansisuchus shansisuchus from the early Middle Triassic (Anisian) of China. The inferred basal skull length of C. cothurnata was approximately 80 cm, making it one of the largest erythrosuchids known.
Key words: Archosauriformes, Erythrosuchidae, anatomy, Triassic, Ladinian, Russia, Bukobay Gorizont.
Richard J. Butler [r.butler.1@bham.ac.uk], School of Geography, Earth and Environmental Sciences, University of Birmingham, Edgbaston, Birmingham B15 2TT, UK.
Andrey G. Sennikov [sennikov@paleo.ru], Borissiak Paleontological Institute RAS, Profsoyuznaya 123, Moscow 117647, Russia; Kazan Federal University, Institute of Geology and Petroleum Technologies, Kremlyovskaya str. 4, Kazan 420008, Russia.
Martín D. Ezcurra [martindezcurra@yahoo.com.ar], School of Geography, Earth and Environmental Sciences, University of Birmingham, Edgbaston, Birmingham B15 2TT, UK; Sección Paleontología de Vertebrados, CONICET-Museo Argentino de Ciencias Naturales “Bernardino Rivadavia”, Ángel Gallardo 470 (C1405DJR), Buenos Aires, Argentina.
David J. Gower [d.gower@nhm.ac.uk], Department of Life Sciences, The Natural History Museum, London SW7 5BD, UK.
Received 4 June 2019, accepted 10 July 2019, available online 28 October 2019.
Copyright © 2019 R.J. Butler et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Erythrosuchids are a clade of early archosauriform reptiles, the fossil remains of which have been recovered from Triassic rock sequences in South Africa, Russia, China, and India (Gower 2003; Ezcurra et al. 2013, 2019; Gower et al. 2014; Ezcurra 2016). They were hypercarnivorous, possibly apex predators in late Early and Middle Triassic terrestrial ecosystems, and reached large sizes, with the skull of the South African species Erythrosuchus africanus reaching more than one metre in length (Gower 2003). The clade apparently went extinct prior to the end of the Middle Triassic, with the putatively geologically youngest erythrosuchid being Chalishevia cothurnata Ochev, 1980, from the late Middle Triassic (Ladinian) of European Russia (Ochev 1980; Shishkin et al. 1995; Sennikov 1995, 2008; Gower and Sennikov 2000; Tverdokhlebov et al. 2003; Ezcurra et al. 2013; Ezcurra 2016).
Ochev (1980) described Chalishevia and its only species C. cothurnata on the basis of fragmentary cranial and possibly referable postcranial remains from several localities in the Bukobay Gorizont of Orenburg Province. He referred Chalishevia to Erythrosuchidae, and noted the characteristic presence of an additional opening in the skull in front of the antorbital fenestra. Sennikov (1995, 2008) referred additional fragmentary remains from the same stratigraphic horizon to C. cothurnata. Gower and Sennikov (2000) provided some brief additional anatomical notes on C. cothurnata, and hypothesized a close relationship between this taxon and the Chinese erythrosuchid Shansisuchus shansisuchus (Young 1964). Most recently, Ezcurra (2016) provided a revised diagnosis for C. cothurnata and incorporated it for the first time into a numerical phylogenetic analysis, recovering it within Erythrosuchidae as the sister taxon of S. shansisuchus (see also Butler et al. 2019; Ezcurra et al. 2019).
With these few exceptions, Chalishevia cothurnata has been largely ignored, despite its importance as potentially the stratigraphically youngest erythrosuchid and its large size and unusual morphological features. Here, we provide a full redescription of the type and referred material of C. cothurnata, and discuss its phylogenetic relationships.
Institutional abbreviations.—BP, Evolutionary Studies Institute, University of the Witwatersrand, Johannesburg, South Africa; BSPG, Bayerische Staatssammlung für Paläontologie und Geologie, Munich, Germany; GHG, Geological Survey, Pretoria, South Africa; IVPP, Institute of Vertebrate Paleontology and Paleoanthropology, Beijing, China; NHMUK, Natural History Museum, London, UK; PIN, Borissiak Paleontological Institute of the Russian Academy of Sciences, Moscow, Russia; RC, Rubidge Collection, Wellwood, Graaf-Reinet, South Africa; SAM, Iziko South African Museum, Cape Town, South Africa; UTGD, School of Earth Sciences, University of Tasmania, Hobart, Australia.
Systematic palaeontology
Archosauromorpha Huene, 1946 sensu Dilkes 1998
Archosauriformes Gauthier, Kluge, and Rowe, 1988 sensu Gauthier et al. 1988
Erythrosuchidae Watson, 1917 sensu Ezcurra et al. 2010
Genus Chalishevia Ochev, 1980
Type species: Chalishevia cothurnata Ochev, 1980; see below.
Diagnosis.—As for type and only species.
Chalishevia cothurnata Ochev, 1980
Type material: Holotype: PIN 4366/1 (formerly SGU 104/3852, wrongly cited as PIN 4356/1 by Gower and Sennikov 2000), left maxilla and both nasals (Ochev 1980: fig. 2a−c; Gower and Sennikov 2000: fig. 8.6). Paratypes: PIN 4366/2 (formerly SGU 104/3853), an incomplete right quadrate, and PIN 4366/3 (formerly SGU 104/3854), a tooth, both from the same locality and bed as the holotype.
Type locality: Bukobay VII locality (Shishkin et al. 1995; Sennikov 1995, 2008; Gower and Sennikov 2000; wrongly cited as Bukobay VI by Ochev 1980), Sol-Iletsk distrinct, Orenburg Province, Russia (Fig. 1). Tverdokhlebov et al. (2003) referred to this locality as Belyaevsky II (their locality 31).
Type horizon: Bukobay Gorizont, Ladinian, late Middle Triassic (Shishkin et al. 1995, 2000).
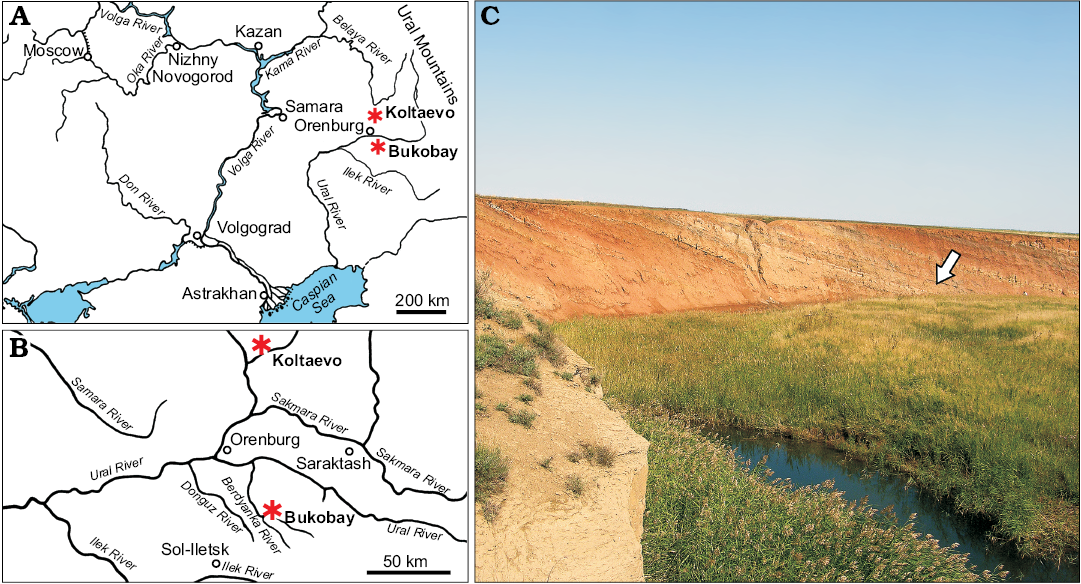
Fig. 1. Location of the Bukobay and Koltaevo localities (asterisks) in Orenburg Province and the Republic of Bashkortostan, Russia (A, B). Photograph of the Bukobay VII locality (C). Arrow indicates the point at which the holotype specimen of Chalishevia cothurnata was collected. Photograph by AGS.
Referred material.—PIN 4366/4, 4366/5, 4366/7, partial left pterygoid, originally preserved in three separately numbered pieces, and PIN 4366/8, a tooth. Both specimens come from the same locality and bed as the holotype (Sennikov 1995, 2008). PIN 2867/18 (formerly SGU 104/3862), fragment of right nasal, from Koltaevo III, Kuyurgazinskiy district, Republic of Bashkortostan, Russia (Ochev 1980). PIN 2867/7, partial right nasal from Koltaevo III (Sennikov 1995, 2008). Tverdokhlebov et al. (2003) referred to this locality as Starokoltaevo I (their locality 8). All from the Bukobay Gorizont, Ladinian, late Middle Triassic (Shishkin et al. 1995, 2000).
Emended diagnosis.—Chalishevia cothurnata is a large erythrosuchid that differs from other archosauromorphs in the following combination of features (autapomorphy indicated with an asterisk): large, subcircular accessory antorbital fenestra and fossa; maxillo-nasal tuberosity; maxilla with two rows of large foramina on the lateral surface; and maxilla with an edentulous anterior tip, mainly vertical ascending process, and oblique, anteroventrally-to-posterodorsally oriented ventral border of the antorbital fossa on the horizontal process* (modified from Ezcurra 2016).
Description.—The holotype consists of the left maxilla, partial left nasal and partial right nasal, all belonging to a single individual (Ochev 1980). All bones articulate well with one another. The paratypic quadrate and tooth and referred pterygoid and tooth from the same locality (Bukobay VII) and bed were recovered close to the holotype remains and probably also represent the same individual based on similar size, preservation, and phylogenetic consistency of morphology.
Maxilla: The left maxilla of the holotype (PIN 4366/1) is nearly complete (Figs. 2, 3), but is damaged at the lateral extremity of the posterolateral end of the main body, below the posterior margin of the antorbital fenestra, at the posterodorsal tip of the ascending process, and anterodorsally, below the anterior part of the accessory antorbital fenestra. Medially, the palatal process is damaged at its base and is missing its distal tip (Fig. 2A2, 3), and the medial surface of the maxilla is missing at the posterior end of the tooth row. The external bone surface is generally very well preserved. The maximum anteroposterior length of the maxilla as preserved is approximately 310 mm, and the maximum dorsoventral height as preserved is approximately 180 mm.
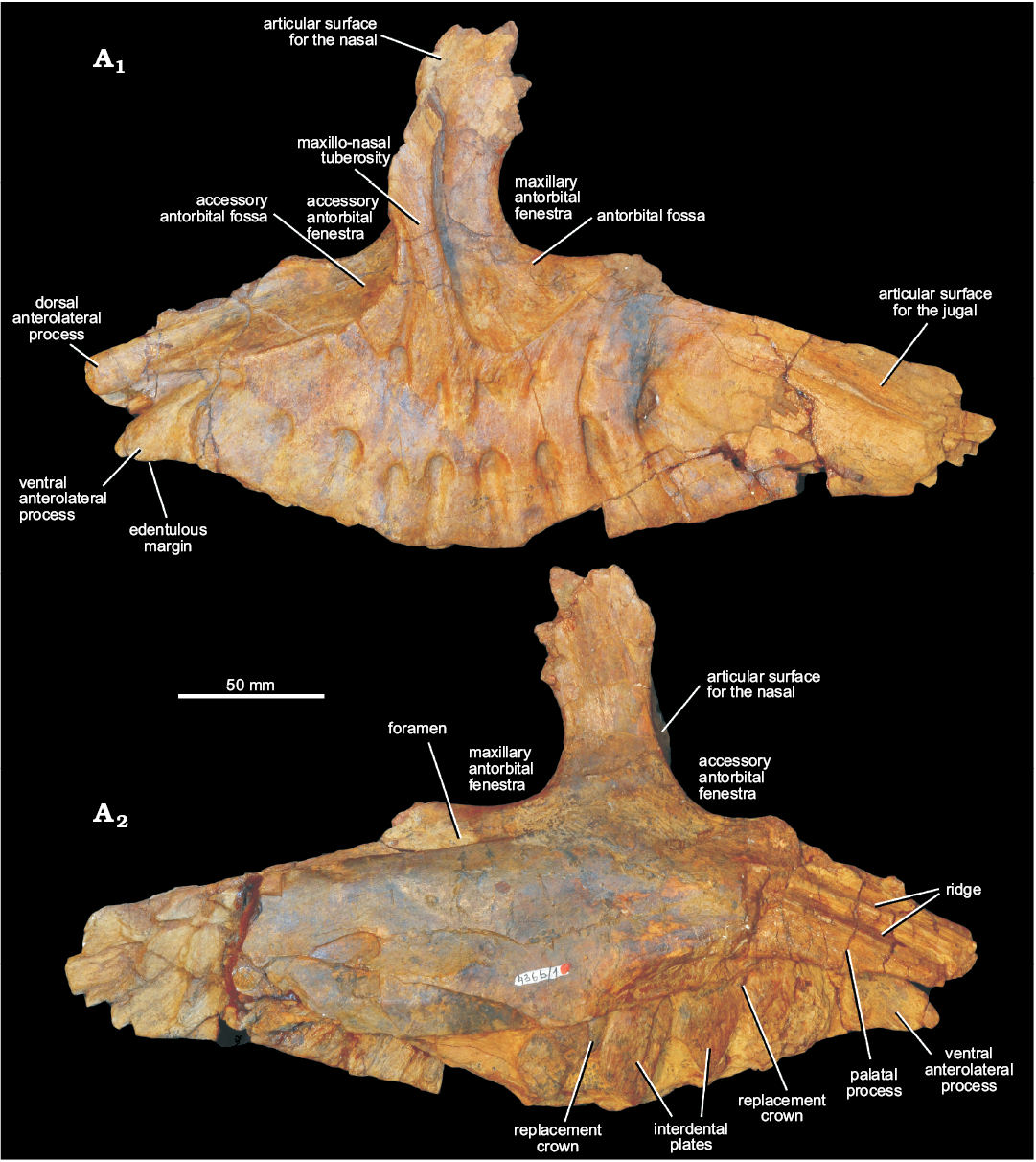
Fig. 2. Left maxilla of erythrosuchid archosauriform Chalishevia cothurnata Ochev, 1980 (holotype, PIN 4366/1) from the Bukobay VII locality, Sol-Iletsk district, Orenburg Province, Russia; Bukobay Gorizont, Ladinian, late Middle Triassic; in lateral (A1) and medial (A2) views.
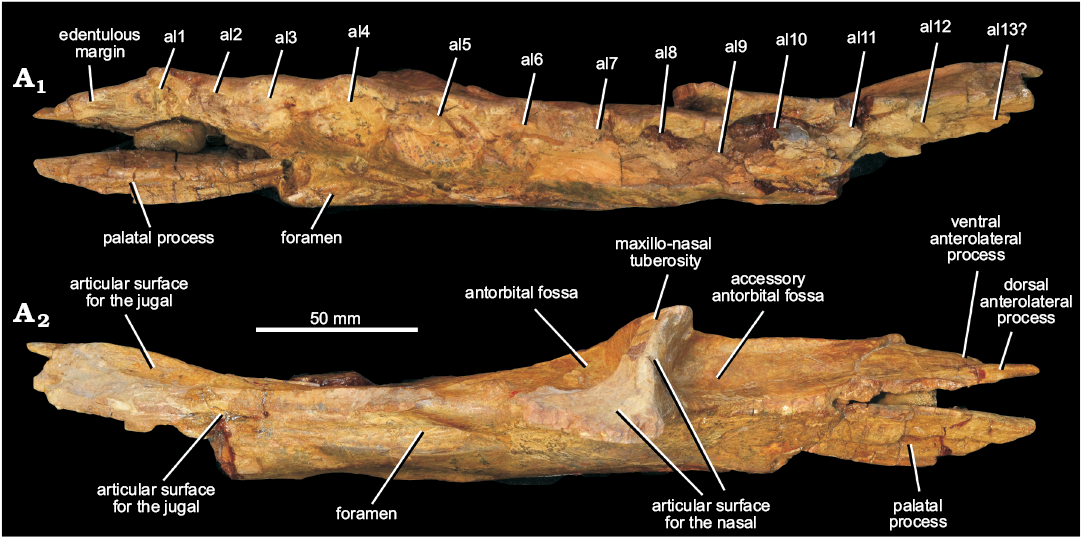
Fig. 3. Left maxilla of erythrosuchid archosauriform Chalishevia cothurnata Ochev, 1980 (holotype, PIN 4366/1) from the Bukobay VII locality, Sol-Iletsk district, Orenburg Province, Russia; Bukobay Gorizont, Ladinian, late Middle Triassic; in ventral (A1) and dorsal (A2) views. Abbreviations: al1–13, alveoli 1–13.
The main body of the maxilla is anteroposteriorly elongated relative to its dorsoventral height, and tapers at its anterior and posterior ends in lateral view (Fig. 2A1). This condition differs from the mostly horizontal dorsal margin of the anterior process of the maxilla of Garjainia prima, whereas the condition in Erythrosuchus africanus and Shansisuchus shansisuchus seems to be intraspecifically variable (Young 1964; Wang et al. 2013; Ezcurra et al. 2019). The alveolar margin is strongly convex centrally, becoming gently concave anteriorly (towards the contact with the premaxilla) and posteriorly, resembling the condition in a referred specimen of Shansisuchus shansisuchus (Wang et al. 2013). By contrast, the alveolar margin along the anterior process is straight in lateral view in Erythrosuchus africanus and Garjainia prima (Gower 2003; Ezcurra et al. 2019). In dorsal or ventral view (Fig. 3) the lateral surface of the maxilla is flat to gently convex anteriorly, but becomes gently concave posteriorly, towards the jugal contact.
The ascending process is strongly offset posterior to the anterior margin of the maxilla, with the base of the ascending process lying approximately one third of the distance along the anteroposterior length of the element, closely resembling the condition in Garjainia prima (Ezcurra et al. 2019) and Shansisuchus shansisuchus (IVPP V2501, V2505, V2508; Wang et al. 2013). The ascending process rises nearly vertically, and is excavated anteriorly (visible most clearly in anterior or dorsal views) by a fossa that bordered the accessory antorbital fenestra (Figs. 2A1, 3A2: accessory antorbital fossa). This fossa is termed here the accessory antorbital fossa and is also present in Shansisuchus shansisuchus (IVPP V2501, V2505, V2508). Posteriorly the ascending process is excavated by the maxillary antorbital fossa. Between these two excavations there is a very strongly raised bar or ridge of bone (the “maxillo-nasal tuberosity”; Ezcurra 2016) on the lateral surface of the ascending process (Figs. 2A1, 3A2: maxillo-nasal tuberosity), which has been found as a synapomorphy of the clade that includes all erythrosuchids with the exception of Fugusuchus hejiapanensis and Guchengosuchus shiguaiensis (Ezcurra 2016; Ezcurra et al. 2019; Butler et al. 2019). At its base, this ridge is anteroposteriorly expanded and smoothly merges with the lateral surface of the maxilla. Dorsally, this ridge curves first gently anteriorly and then gently posteriorly. Dorsal to the lower third of the contact of this ridge with the ventral process of the nasal (see below), the ridge becomes gradually reduced in transverse width, merging smoothly with the preserved dorsal tip of the ascending process. The lateral surface of this ridge is flattened and laterally facing along its ventral half, but it twists to face posterolaterally as it diminishes in transverse width towards its dorsal apex.
In lateral view the tapered anterior end of the maxilla has two distinct prong-like processes separated by a posterodorsally-tapering groove (Fig. 2A1), a condition that resembles that of Garjainia prima, though this species has shorter prong-like processes (Ezcurra et al. 2019). The more ventral of these processes tapers to a bluntly rounded and slightly downturned tip (Fig. 2A1: ventral anterolateral process). Based on comparison with the holotype of Garjainia prima (PIN 2394/5; Ezcurra et al. 2019), this process did not articulate with the premaxilla, but instead projected into a subnarial notch along the tooth margin between the premaxilla and maxilla. The ventral surface of this process is edentulous (Figs. 2A1, 3A1: edentulous margin), meaning that there was a short gap between maxillary and premaxillary tooth rows, as occurs in Garjainia prima (Ezcurra et al. 2019) and Shansisuchus shansisuchus (Young 1964). The process projects slightly medially as preserved, but this likely reflects damage given that there is a substantial, dorsoventrally extending break at the base of the process.
The more dorsal of the two anterolateral processes is also slightly downturned towards its anterior end, and is bluntly squared off at its tip (Fig. 2A1: dorsal anterolateral process). The tip is slightly bevelled, presumably forming a small, loose contact with the premaxilla. The groove between the two anterolateral processes is open at its anterior end and backed by bone more posteriorly; it tapers and connects to a foramen posteriorly.
The lateral surface of the more dorsal of the two anterolateral processes is flattened, and merges posteriorly with the accessory antorbital fossa (Figs. 2A1, 3A2: accessory antorbital fossa). This fossa is delimited ventrally and posteriorly by a sharp ridge that forms the anterior margin of the ascending process in lateral view. The fossa excavates the anterior surface of the ascending process, and is deepest immediately anterior to the base of the process, as occurs in Shansisuchus shansisuchus (IVPP V2508). The dorsal part of the maxilla anterior to the ascending process and the medial part of the anterior margin of the ascending process form the smoothly curved ventral and posterior margins of the accessory antorbital fenestra. Within the accessory antorbital fossa, medial to the ridge of the ascending process (at the point at which it begins to curve posteriorly and diminish in thickness), there is an articular surface for the ventral process of the nasal that is gently concave mediolaterally (Fig. 3B: articular surface for the nasal).
The ventral and anterior rims of the antorbital fenestra (Fig. 2A1: maxillary antorbital fenestra) are preserved, and are smoothly curved. The ventral margin of the antorbital fenestra is almost exactly level with the ventral margin of the accessory antorbital fenestra. The antorbital fenestra is surrounded ventrally, anteriorly and anterodorsally by an antorbital fossa (Figs. 2A1, 3A2: antorbital fossa) that extends onto the dorsolateral surface of the main body of the maxilla and excavates the posterolateral part of its ascending process, as is the case in Erythrosuchus africanus (Gower 2003) and Shansisuchus shansisuchus (Young 1964). By contrast, Fugusuchus hejiapanensis, Garjainia prima (Ezcurra et al. 2019) and Guchengosuchus shiguaiensis (Butler et al. 2019) lack an antorbital fossa on the horizontal process of the maxilla. The antorbital fossa of Chalishevia cothurnata is bounded by a sharp ridge on the ascending process, and a well-defined but low ridge on the main body. This latter ridge extends from anteroventral to posterodorsal, and its orientation is very similar to that of the boundary of the antorbital fenestra in Garjainia prima (PIN 2394/5; Ezcurra et al. 2019). The orientation of the ventral margin of the antorbital fossa contrasts with the horizontal margin present in Erythrosuchus africanus (Gower 2003). The maxillary antorbital fossa of Shansisuchus shansisuchus becomes shallower posteriorly on the horizontal process and lacks a clearly defined posteroventral margin (IVPP V2505, V2508). Within the fossa of the holotype of Chalishevia cothurnata, two small inflated areas that form volcano-like features are present. These are unusual, and may represent some kind of pathology, although this cannot be determined without histological or computed tomographic analysis.
There are numerous large nutrient foramina on the lateral surface of the maxilla (Fig. 2A1), and these are connected with broad and deeply incised grooves. Nine of these foramina are placed in a ventral convex row that extends parallel to the alveolar margin, with each foramen being positioned approximately above one of the alveoli. The anteriormost of this row of foramina is continuous with the anterior groove between the anterolateral processes described above. These foramina open towards the alveolar margin; broad, well-defined, and deeply impressed grooves extend from the foramina to the alveolar margin, as occurs in some other erythrosuchids and loricatans (Ezcurra 2016). The foramina have oval to subcircular outlines. The posteriormost of the foramina is larger than those positioned more anteriorly.
Dorsal to this first row of foramina there are six foramina in a second row, an unusual condition among early archosauromorphs. This second row of foramina could be autapomorphic for Chalishevia cothurnata, but a similar feature may be present in at least one specimen of Shansisuchus shansisuchus (Young 1964: fig. 10a), and the condition in this latter taxon requires further investigation. The anteriormost of these foramina is small, and positioned ventral to the accessory antorbital fenestra, and is not connected to a well-defined groove. The second foramen in this row is positioned below the anterior margin of the ascending process, opens ventrally, and communicates with three grooves. The deepest of these grooves extends dorsally onto the ascending process, terminating adjacent to the anterior margin of the process immediately ventral to the articulation with the nasal. A second, weakly expressed groove extends ventrally towards the tooth row but fades out before reaching the more ventral row of foramina. A third, narrow groove extends anterodorsally, reaching the boundary of the accessory antorbital fossa. The third foramen is small, ventrally opening, and positioned below the ascending process. A shallow groove extends ventrally from this opening. The fourth is positioned below the posterior margin of the ascending process. It also opens ventrally and is partially obscured by a small thickening of the surface of the maxilla posterodorsal to it. There is no clearly developed groove extending ventrally from this opening, but a groove does extend dorsally along the posterior margin of the ascending process, fading out level with the dorsal margin of the volcano-like projections. This groove appears to subdivide close to its base, with a short groove extending to reach the anteroventral corner of the antorbital fossa. The fifth and sixth of this second row of foramina open posteroventrally. Well-developed grooves extend posterodorsally from these foramina, curving along their lengths. A clear groove also extends ventrally from the sixth foramen, connecting to the eighth foramen of the lower row.
There is a well-developed facet for the reception of the jugal laterally at the posterior end of the maxilla (Figs. 2A1, 3A2: articular surface for the jugal). A slot for articulation with the jugal also appears to be present on the posterodorsal margin of the maxilla (Fig. 3A2). Anterodorsally, at the tip of the ascending process, a second facet for articulation with the nasal is present (Fig. 3A2: articular surface for the nasal). Due to damage to the posterodorsal margin of the ascending process it cannot be determined if a contact surface was present for the lacrimal, a contact that occurs in Shansisuchus shansisuchus (Wang et al. 2013).
Medially the palatal process is broken at its tip, and has three longitudinal grooves separated by two ridges on its medial surface (Fig. 2A2: palatal process). It has been broken at its base and displaced slightly laterally. There is a ventrally opening large foramen posteroventral to the base of the palatal process (Fig. 3A1: foramen). Another very deep and large foramen is positioned below the antorbital fenestra (Figs. 2A2, 3A2: foramen), posterior to the ascending process, opening into a groove that extends along the dorsal surface of the main body of the maxilla. The medial surface of the main body is convex dorsoventrally. A sharp ridge separates the main body from the alveolar region. Interdental plates are present and contact one another at their bases (Fig. 2A2: interdental plates). The interdental plates extend ventrally to the same level as the ventrolateral margin of the bone, indicating a thecodont tooth implantation (sensu Bertin et al. 2018). Small foramina are present between them, and small parts of replacement crowns are visible in these foramina dorsal to the third and fifth alveoli (Fig. 2A2: replacement crown). Almost no details of these crowns are visible, although the one above the third alveolus has several denticles preserved along the distal margin of the tip (the mesial margin is not preserved).
At least twelve (possibly 13) alveoli appear to be present (Fig. 3A1), rather than 10 as identified by Ochev (1980). Two alveoli appear to be present in the space identified by Ochev (1980) as for the first alveolus. It seems likely that, although preservation is poor, two alveoli (6 and 7) were present in the gap identified as for alveolus 5 by Ochev (1980). At the very posterior end of the tooth row it is unclear whether or not a thirteenth alveolus was present. The condition in Chalishevia cothurnata resembles the relatively low maxillary tooth counts of Guchengosuchus shiguaiensis (IVPP V8808: maxillary tooth count 14 or 15), Garjainia prima (PIN 2394/5: maxillary tooth count 13 or 14), Erythrosuchus africanus (BP/1/5207: maxillary tooth count 11), Shansisuchus shansisuchus (Young [1964] described 9 or possibly 10 tooth positions and Wang et al. [2013] probably 13 maxillary teeth), and Euparkeria capensis (Ewer, 1965; maxillary tooth count 13). By contrast, higher maxillary tooth counts are present in the less crownward archosauromorphs Tasmaniosaurus triassicus (UTGD 54655; >21), Proterosuchus fergusi (BP/1/3993, BSPG 1934 VIII 514, GHG 231, RC 59, 96, SAM-PK-11208, K140, K10603; maxillary tooth count 20–31, depending on ontogenetic stage; Ezcurra and Butler 2015), “Chasmatosaurus” yuani (>23 in IVPP V90002 and ≥29 in IVPP V2719) and Prolacerta broomi (BP/1/471, Modesto and Sues 2004; tooth count 24–25). There is no evidence of ankylosis between the base of the crowns and the alveolar margin of the bone.
Nasal: A substantial portion of the left nasal is preserved (Fig. 4), and articulates with the ascending process of the maxilla via a ventral process. The left nasal is broken and damaged at its anterior end, and posteriorly is broken posterior to the ventral process. A major crack extends from posterodorsal to anteroventral through the contact between the main body of the nasal and the ventral process (Fig. 4: cr), and this means that the orientation of these two parts of the element to one another (and thus the orientation of the anterior part of the nasal to the maxilla when in articulation) is not accurately preserved.
Medially, the left nasal is dorsoventrally thickened with a convex dorsal surface at its midline contact with the right nasal. The medial surface of the element is grooved for articulation with a ridge of the right nasal (Fig. 4A2: articular surface for the nasal); below this groove there is a ridge that fits into a corresponding groove on the right nasal. At its anterior end there is a concavity for articulation with the prenarial process of the premaxilla (Fig. 4A2: articular surface for the premaxilla). The external surface of the nasal is arched, facing dorsally at the midline and dorsolaterally towards the contact with the maxilla and the border of the accessory antorbital fenestra. It is covered with rugose ornamentation towards the posterior end of the preserved element.
Anteriorly, parts of the dorsal border of the naris are preserved (Fig. 4A1: border of the external naris). Behind the naris, there is an elongate, narrow, posterodorsally extending and tapering slot (Fig. 4A1: articular surface for the premaxilla), that would have accommodated a narrow, tapering postnarial process of the premaxilla, similar to that present in Shansisuchus shansisuchus (Young 1964). Dorsal to and adjacent to this slot, the external surface of the nasal is anteroposteriorly grooved (Fig. 4A1: groove).
Posteroventral to the slot for the postnarial process of the premaxilla, the nasal forms the dorsal and posterodorsal margins of the accessory antorbital fenestra (Fig. 4A1: accessory antorbital fenestra), as occurs in Shansisuchus shansisuchus (Young 1964; Wang 2013). Several small foramina are present on the external surface of the nasal adjacent to the fenestra (Fig. 4A1: foramen), and are associated with short anteroventrally extending grooves.
The ventral process of the nasal tapers towards its tip. An extensive facet for the ascending process of the maxilla is present on its posteromedial surface (Fig. 4A3: articular surface for the maxilla). In medial and anterior views, a very deeply recessed fossa (= “sulcus” of Ochev 1980) is present adjacent to the dorsal margin of the accessory antorbital fenestra (Fig. 4A2, A3: accessory antorbital fossa).
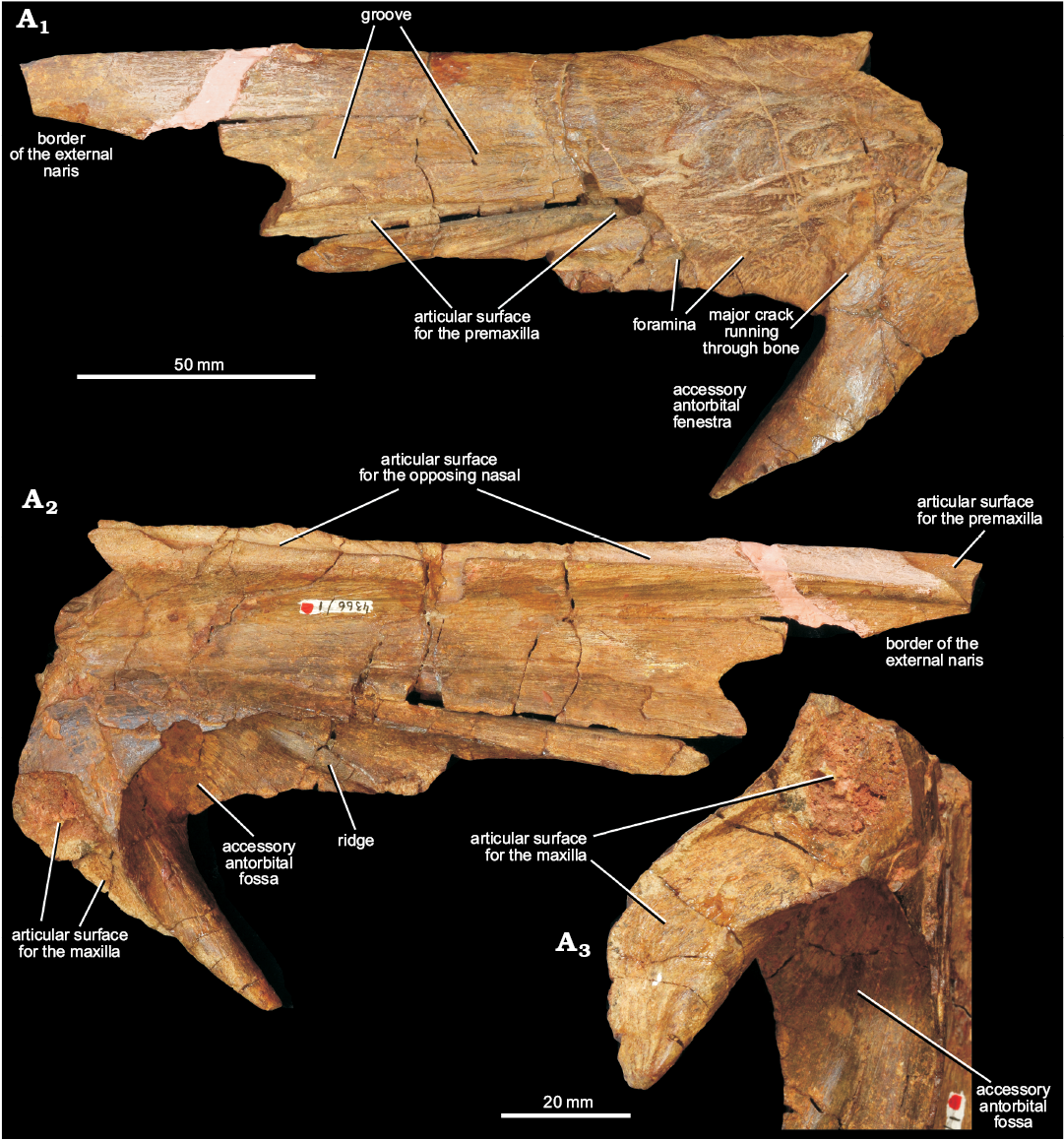
Fig. 4. Left nasal of erythrosuchid archosauriform Chalishevia cothurnata Ochev, 1980 (holotype, PIN 4366/1) from the Bukobay VII locality, Sol-Iletsk district, Orenburg Province, Russia; Bukobay Gorizont, Ladinian, late Middle Triassic; in lateral (A1) and medial (A2) views, and in oblique posteroventromedial view showing the articular surface for the ascending process of the maxilla (A3).
The right nasal is less complete than the left, and is broken anteriorly, just anterior to the beginning of the dorsal margin of the external naris, ventrolaterally, and posteriorly (Fig. 5). The dorsal margin of the accessory antorbital fenestra is almost entirely missing, and the ventral process of the bone has broken away. It does not add anatomical information not present in the left nasal.

Fig. 5. Right nasal of erythrosuchid archosauriform Chalishevia cothurnata Ochev, 1980 (holotype, PIN 4366/1) from the Bukobay VII locality, Sol-Iletsk district, Orenburg Province, Russia; Bukobay Gorizont, Ladinian, late Middle Triassic; in lateral (A1) and medial (A2) views.
Quadrate: A right quadrate (PIN 4366/2), broken into two pieces, was referred to Chalishevia cothurnata by Ochev (1980), and was identified as from the same individual as the holotype but was not described (Fig. 6). The two pieces of the quadrate do not fit together, and a section of the midshaft of the bone is missing. The maximum dorsoventral height of the bone would have been at least 220 mm. The pterygoid wing of the quadrate is missing, as is much of the lateral margin of the bone. Ventrally, the condyles are preserved along with part of the lateral expansion for articulation with the quadratojugal (Fig. 6: articular surface for the quadratojugal). The maximum transverse expansion at the condyles is 66.5 mm. The condyles are strongly compressed anteroposteriorly and expanded transversely (Fig. 6A5), with the medial condyle being broader anteroposteriorly. The medial condyle is slightly anteriorly projected, resembling the condition in Erythrosuchus africanus (NHMUK PV R3592) and Shansisuchus shansisuchus (Young 1964). By contrast, the medial condyle is distinctly more anteriorly projected in Garjainia madiba (BP/1/6232f) and, particularly, Garjainia prima, in which the quadrate acquires an L-shaped profile in ventral view (Ezcurra et al. 2019). Both condyles have strongly convex articular surfaces, although the medial condyle is more strongly convex, resembling the condition in Garjainia prima and other erythrosuchids (Ezcurra et al. 2019). The ventral condyles of the quadrate of Chalishevia cothrunata are separated from each other by a shallow, anteroposteriorly oriented notch, closely resembling the condition in Erythrosuchus africanus (NHMUK PV R3592; Gower 2003) and Shansisuchus shansisuchus (Young 1964). By contrast, the separation between the ventral condyles is much deeper in posterior or anterior view in Garjainia prima (Ezcurra et al. 2019) and an intermediate condition occurs in Garjainia madiba (Gower et al. 2014). Above the condyles, the posterior surface of the quadrate is flattened, and tapers in transverse width toward the midshaft. The anterior surface of the shaft is transversely concave above the condyles. Ventrolaterally the quadrate is expanded into a depressed articular surface for the quadratojugal. This surface tapers dorsally, and the shaft reaches a minimum transverse width just above the dorsal termination of the quadratojugal flange. The borders of the quadrate foramen (if present) are not preserved. The blind, laterally open pit present on the quadrate of other erythrosuchids (e.g., Gower 2003) may have been present immediately dorsal to the articular surface for the quadratojugal, but this area is poorly preserved.
The pterygoid wing is broken, but clearly would have originated some distance above the quadrate condyles, because the broken base of the wing is separated from the condyles on the medial surface by at least 30 mm (Fig. 6A4). Dorsally, the head has a triangular cross section (Fig. 6A5), with a flattened anterior surface (lacking the anterior projection that is present in Garjainia prima: PIN 951/57), a short flat to gently convex lateral surface (broken at its anterior margin) and a broad, gently concave posteromedial surface. The head is convex in posterior and lateral views.
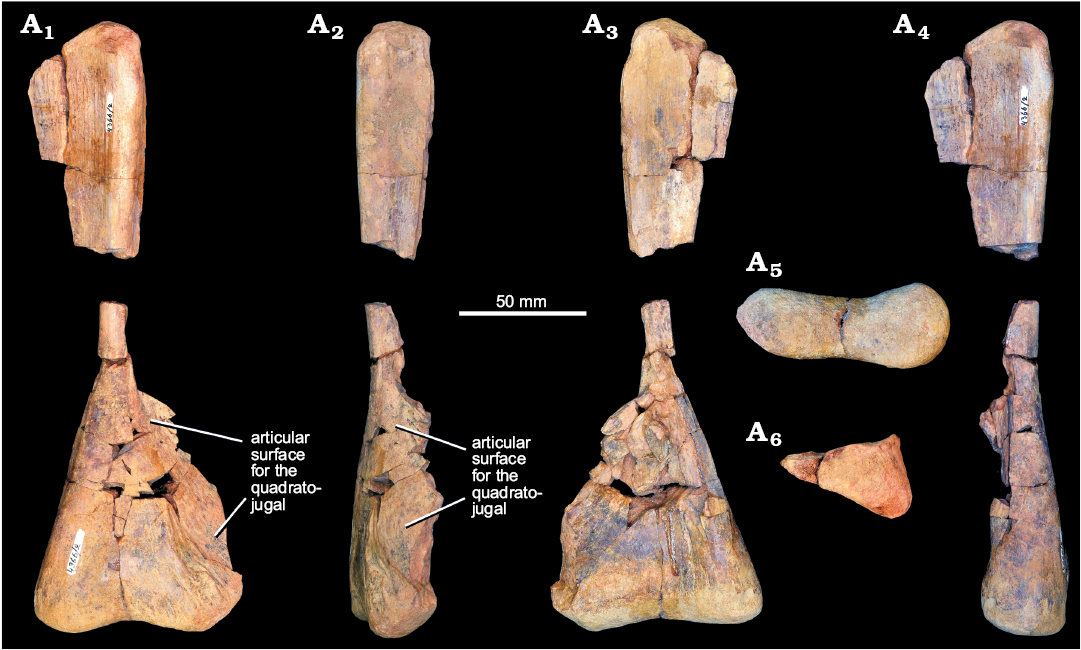
Fig. 6. Right quadrate of erythrosuchid archosauriform Chalishevia cothurnata Ochev, 1980 (paratype, PIN 4366/2) from the Bukobay VII locality, Sol-Iletsk district, Orenburg Province, Russia; Bukobay Gorizont, Ladinian, late Middle Triassic; in posterior (A1), lateral (A2), anterior (A3), medial (A4), ventral (A5), and dorsal (A6) views.
Pterygoid: A partial left pterygoid was originally preserved in three pieces: the posterolateral process (PIN 4366/4), the central part of the element, with broken anterior process, quadrate flange and basipterygoid articulation (PIN 4366/5), and part of the anterior process (PIN 4366/7). The three pieces have been reassembled (Fig. 7), and show a pterygoid that is extremely similar in morphology and slightly larger in size than the holotype specimen of the erythrosuchid Uralosaurus magnus (PIN 2973/70). No palatal teeth are present on any preserved part of the pterygoid, resembling the condition in Erythrosuchus africanus (Gower 2003), Uralosaurus magnus (PIN 2973/70), Shansisuchus shansisuchus (Young 1964: fig. 15a) and the holotype of Garjainia prima (Ezcurra et al. 2019). The posterolateral process is elongate and projects strongly posteriorly and ventrally, and slightly laterally (Fig. 7: posterolateral process), resembling the condition in Erythrosuchus africanus (NHMUK PV R3592), Shansisuchus shansisuchus (Young, 1964: fig. 15a), Uralosaurus magnus (PIN 2973/70) and Sarmatosuchus otschevi (PIN 2865/68). It tapers strongly distally to a point and is slightly curved anteriorly at its distal end. The process is broken along its anterior margin, except for the distal third, which is complete. The dorsal surface of the process is convex anteroposteriorly, with the apex of this convexity offset posteriorly. Ventrally, there is a deeply inset facet along the anterior margin for articulation with the ectopterygoid along the entire preserved length of the process (Fig. 7A3: articular surface for the ectopterygoid). This facet is offset by a distinct step from the rest of the ventral surface, and is flattened medially and slightly convex with low ridges within it laterally. Due to poor preservation, it is unclear if the ectopterygoid also articulated with the dorsal surface of the posterolateral process. The ventral surface of the posterolateral process posterior to this facet is anteroposteriorly convex.
At its base, where the posterolateral process merges into the ventral margin of the quadrate flange, the posterior surface of the posterolateral process bears a dorsoventrally extending and posteromedially opening fossa that continues onto the base of the quadrate flange (Fig. 7A1: fossa on the posterior surface of the posterolateral process), as occurs in Erythrosuchus africanus (Gower 2003: fig. 14b), Shansisuchus shansisuchus (Young 1964: fig. 15a) and Uralosaurus magnus (PIN 2973/70). This fossa is delimited by ridges medially and laterally in these species, but it is absent in Sarmatosuchus otschevi (PIN 2865/68-2). The anterior process of the pterygoid is very incomplete, but a deep ventromedially opening fossa is present on the ventral surface of the pterygoid at the point at which the anterior and posterolateral processes join (Fig. 7A1, A3: fossa on the ventral surface of the anterior process), and expands anteriorly onto the ventral surface of the anterior process, a condition widespread among early archosauriforms (e.g., Erythrosuchus africanus: Gower 2003: fig. 14; Shansisuchus shansisuchus: Young 1964: fig. 15a; Uralosaurus magnus: PIN 2973/70; Sarmatosuchus otschevi: PIN 2865/68-2). The medial margin of the anterior process is drawn out dorsally into a transversely compressed flange, and the lateral surface of the anterior process is dorsoventrally concave. Posterodorsally, only the transversely expanded base of the quadrate wing is preserved, and a medial part of the dorsally expanding anterior wall of the fossa for the basipterygoid process is preserved, although this fossa itself is infilled with matrix and its lateral wall is completely broken away.

Fig. 7. Left pterygoid of erythrosuchid archosauriform Chalishevia cothurnata Ochev, 1980 (PIN 4366/4, PIN 4366/5, PIN 4366/7) from the Bukobay VII locality, Sol-Iletsk district, Orenburg Province, Russia; Bukobay Gorizont, Ladinian, late Middle Triassic; in ventromedial (A1), dorsolateral (A2), and ventrolateral (A3) views.
Dentition: PIN 4366/3 is an isolated tooth including the crown and root (Fig. 8A) that was collected from the type locality and referred to Chalishevia cothurnata by Ochev (1980). It is generally well preserved but damaged at the tip and broken basally, with some damage to labial and lingual surfaces. The entire tooth is 92 mm high and has a mesiodistal length of 25 mm at the base of the crown. The crown is gently recurved in labial and lingual views. One surface of the crown (interpreted as labial) is more strongly convex than the other (lingual). Chisel-like, fine serrations orthogonal to the carina are present along the mesial and distal margins, extending from the base of the crown to the preserved apex. They vary in size along the carinae, being small basally, reaching a maximum size at the dorsoventral midpoint of the crown, and then decreasing slightly apically. There are no blood grooves. Along the mesial margin of the lingual surface, faint crenulations (enamel wrinkles) are present, curving basally away from the carina (Fig. 8A5). No other enamel ornamentation or wear facets are present in the preserved portion of crown. There is no constriction or expansion at the base of the crown with respect to the root.
PIN 4366/8 is a second tooth including the complete crown and nearly the entire root (Fig. 8B). The tooth is 123 mm long as preserved, and the crown makes up approximately the apical 38 mm. The root is oval in cross section, being slightly longer mesiodistally than labiolingually. It tapers strongly towards its base. The crown is recurved, with the apex extending distal to the distal margin. The mesial and distal margins possess finely serrated carinae, with serrations extending from the very base of the crown right to the apex. Variation in denticle size is similar to the other crown. Labial and lingual crown surfaces are equally convex mesiodistally. Faint, basally curved wrinkles in the enamel surface are present (Fig. 8J), but only on one surface and more extensively developed adjacent to the distal carina.

Fig. 8. Teeth of erythrosuchid archosauriform Chalishevia cothurnata Ochev, 1980 (PIN 4366/3, A; PIN 4366/8, B) from the Bukobay VII locality, Sol-Iletsk district, Orenburg Province, Russia; Bukobay Gorizont, Ladinian, late Middle Triassic; in labial (A1), lingual (A2), mesial (A3, B3), distal (A4, B4), and labial/lingual (B1, B2) views, and with close-ups of enamel wrinkles and serrations (A5, B5).
Two fragments of right nasal from the Koltaevo III locality are referred to Chalishevia cothurnata (Fig. 9). One fragment (PIN 2867/7) corresponds to the main body (Fig. 9A3, B1), 99 mm in length with a maximum width of 43 mm as preserved. It is broken posteriorly before the beginning of the ventral process and anteriorly before the margin of the external naris. The size of this specimen, particularly the transverse distance between the midline and the facet for the premaxilla (20 mm) compares well with the right nasal of the holotype (17.5 mm). By contrast, a second fragment (PIN 2867/18), also of a right nasal but consisting of the ventral process and base of anterior ramus of main body (Fig. 9A1, A2), is smaller than the holotype specimen, probably juvenile, and represents a second individual from this locality.
In PIN 2867/7 the posterior end of the narrow and posterodorsally tapering sutural surface for the postnarial process of the premaxilla is visible (Fig. 9B1: articular surface for the premaxilla). Posterior and lateral to this the preserved lateral margin appears to have been broken and worn, such that it superficially resembles a finished edge. Thus, the dorsal border of the accessory antorbital fenestra does not appear to be preserved. Medial to the premaxillary facet, the external surface of the nasal is transversely convex, and the longitudinal depression extending from the posterior end of the external naris parallel to the premaxillary facet that is present in the holotype of Chalishevia cothurnata is absent. Medially, close to the midline, there is a deep and sharply delimited groove that extends anteroposteriorly (Fig. 9B1: groove); this groove is truncated anteriorly by the breakage of the anterior end of the bone, and it is not clear whether a foramen was present at this point. The external surface of the posterior part of the bone is rugose and ornamented.
Ventrally, there is a well-developed, anteroposteriorly extending ridge that runs parallel to the medial margin of the bone (Fig. 9B2: ridge), separated from it by approximately 12 mm anteriorly, but diverging from the midline slightly and fading out towards the posterior end of the bone. This ridge is not present in the holotype of Chalishevia cothurnata. At the lateral margin the bone appears to be broken, but the base of the deep fossa that surrounds the accessory antorbital fenestra in the holotype of Chalishevia cothurnata is preserved (Fig. 9B2: accessory antorbital fossa).
The bone differs from the holotype specimen of Chalishevia cothurnata in the ventral ridge, the absence of an external groove adjacent to the facet for the postnarial process of the premaxilla, and the presence of a more medially placed groove on the external surface. It is strongly similar to Chalishevia cothurnata in possessing the fossa surrounding the accessory antorbital fenestra, and the very strongly tapered postnarial process of the premaxilla, although these features are also present in Shansisuchus shansisuchus (Young 1964).
The second nasal fragment (PIN 2867/18) includes a more posterior part of the main body and the almost complete ventral process (the tip is missing) for articulation with the dorsal process of the maxilla (Fig. 9A1, A2). The external surface of the bone is convex and rugose, with several foramina present, and is very similar to the same region in the holotype. The ventral process is triangular in lateral view (Fig. 9A1: ventral process). In anterior view, the anterodorsal surface of the ventral process and the ventral surface of the main body are deeply excavated to form an accessory antorbital fossa as in the holotype of Chalishevia cothurnata (Fig. 9A2: accessory antorbital fossa). The anterior border of the ventral process and the preserved ventral margin of the main body would have formed the posterodorsal margins of an accessory antorbital fenestra, as in the holotype of Chalishevia cothurnata and Shansisuchus shansisuchus. Medially, the main body is transversely concave, and at its medial margin forms an articular surface for the opposite nasal with two grooves and a median ridge. The posteromedial surface of the ventral process is bevelled ventrally for the ascending process of the maxilla.
Based on the strong similarities of these nasal fragments to those in the holotype, and their occurrence within the same stratigraphic horizon, we retain their referral to Chalishevia cothurnata. Minor differences between PIN 2867/7 and the nasals of Chalishevia cothurnata are interpreted as individual intraspecific variation.
Stratigraphic and geographic range.—Bukobay Gorizont, Ladinian, late Middle Triassic; Sol-Iletsk district, Orenburg Province and Kuyurgazinskiy district, Republic of Bashkortostan, Russia.
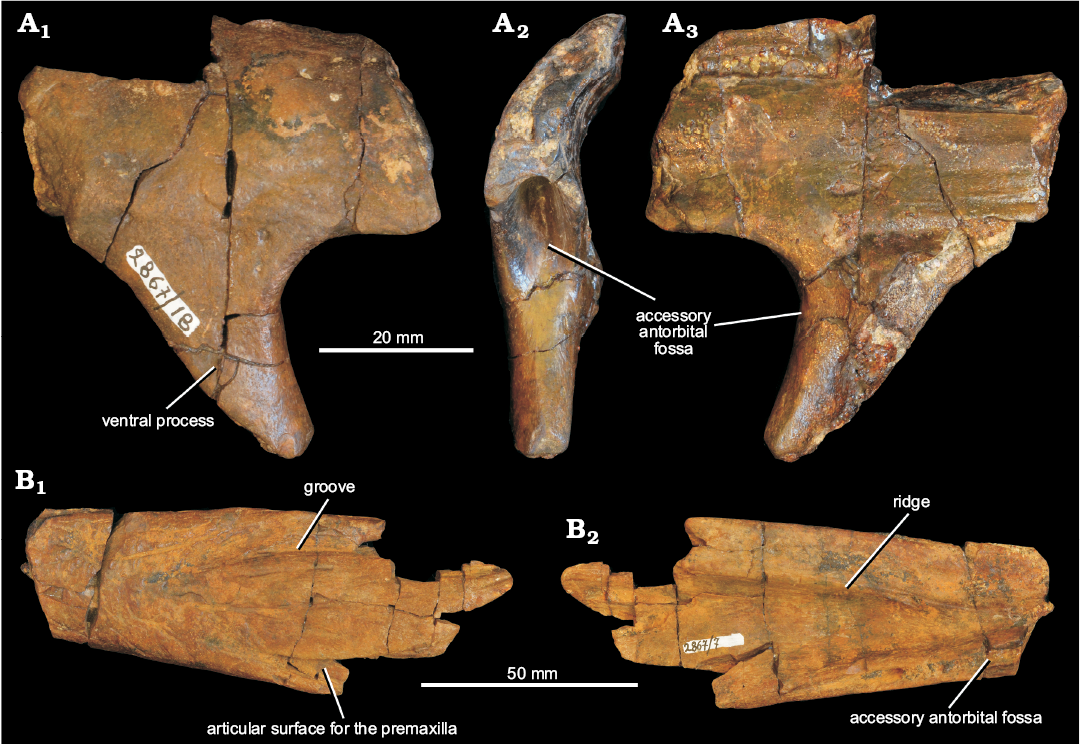
Fig. 9. Partial right nasals referred by Ochev (1980) and Sennikov (1995, 2008) to erythrosuchid archosauriform Chalishevia cothurnata Ochev, 1980 (PIN 2867/18, A; PIN 2867/7, B) from the Koltaevo III locality, Kuyurgazinskiy district, Republic of Bashkortostan, Russia; Bukobay Gorizont, Ladinian, late Middle Triassic; in lateral (A1, B1), anterior (A2), and medial (A3, B2) views.
Erythrosuchidae indet.
Figs. 10, 11.
Material.—PIN 525/30, partial left surangular, from Koltaevo III, Kuyurgazinskiy district, Republic of Bashkortostan (Sennikov 1995, 2008). PIN 4165/18 (formerly SGU 104/3859), two cervical vertebrae, from Bukobay I, Sol-Iletsk district, Orenburg Province, Russia (Ochev 1980). PIN 4188/97 (formerly SGU 104/3861), cervical centrum, from Bukobay V, Sol-Iletsk district, Orenburg Province, Russia (Ochev 1980). PIN 4188/98 (formerly SGU 104/3861), dorsal vertebra, from Bukobay V (Ochev 1980). The Bukobay I and V and Koltaevo III localities are in the Bukobay Gorizont (Ladinian, late Middle Triassic: Shishkin et al. 1995, 2000). Tverdokhlebov et al. (2003) referred to the Bukobay V locality as Belyaevsky I (their locality 30).
Description.—Surangular: A fragmentary posterior half of left surangular (PIN 525/30; Fig. 10) includes part of the dorsal margin of the element and the ventromedial expansion (preglenoid process) that would have articulated with the anterior surface of the articular and formed the anterior margin of the glenoid fossa. The surangular is broken anteriorly, ventrally, and posteriorly. The preserved portion of the surangular is 148 mm long, with the dorsal margin reaching a maximum width of 27 mm and the element being approximately 42 mm wide at the level of the preglenoid process. The dorsal margin is broadly expanded dorsally. The dorsal surface is slightly thickened into low ridges at its medial and lateral margins (Fig. 10A1: ridge), between which the dorsal surface is gently concave transversely, with this concavity interrupted by a low anteroposteriorly extending ridge that shifts slightly laterally and becomes more prominent towards its anterior end. An almost identical transverse expansion of the dorsal margin that is gently concave with lateral, medial and median ridges is also present in surangulars of Garjainia prima (e.g., PIN 951/46), and the surangular is generally very similar to those of other erythrosuchids (e.g., Erythrosuchus africanus; Gower 2003). The transverse expansion of the dorsal margin overhangs the (mostly missing) ventral, transversely compressed, and sheet like flange of the surangular (Fig. 10A2: broken base of ventral flange), with the overhang being greater medially than laterally. The lateral expansion diminishes and the medial expansion increases in width anteriorly. The dorsal transverse expansion is dorsoventrally thickest medially and tapers laterally. There is a groove on its medial surface at the anterior preserved end (Fig. 10A4: groove). An anteroventrally opening fossa is present on the ventral surface of the dorsal expansion adjacent to the broken anterior surface (Fig. 10A2: fossa). At the posterior end of the dorsal expansion a slightly raised transverse ridge delimits the anterior margin of the glenoid region (Fig. 10A1: ridge defining anterior border of glenoid region). Medial to this, the surangular is expanded ventromedially into the flange-like preglenoid process (Fig. 10A1: preglenoid process), the anterior surface of which is twisted to face anterodorsally and is transversely concave. Posteriorly and posteroventrally there are two articular surfaces for the articular.

Fig. 10. Partial left surangular referred by Sennikov (1995, 2008) to erythrosuchid archosauriform Chalishevia cothurnata Ochev, 1980 (PIN 525/30) but here considered as Erythrosuchidae indet., from the Koltaevo III locality, Kuyurgazinskiy district, Republic of Bashkortostan, Russia; Bukobay Gorizont, Ladinian, late Middle Triassic; in dorsal (A1), ventral (A2), lateral (A3), and medial (A4) views.
Vertebrae: From the Bukobay I locality there are two articulated vertebrae that probably represent posterior cervicals (PIN 4165/18; Fig. 11A). These vertebrae are very erythrosuchid-like in appearance, with the very short and deep centra and large size consistent with being referable to the clade (e.g., Ezcurra 2016). The two vertebrae are poorly preserved, and the more posterior of the two is missing most of its centrum. Neurocentral sutures are closed but visible, indicating that these vertebrae do not belong to an ontogenetically mature individual. It is unclear whether or not intercentra would have been present. The centrum of the first (more anterior) vertebra is anteroposteriorly short, dorsoventrally deep and transversely broad (maximum length 33.9 mm; maximum depth of anterior margin 51.5 mm; maximum width of anterior margin 54 mm). The articular faces are gently concave, with sub-circular outlines, although the ventral margin of the anterior articular face of the first vertebra is slightly damaged, and the posterior articular face is incomplete ventrally. In ventral view, the lateral surfaces are arched strongly inwards giving the centrum a spool-like appearance. The parapophysis is placed very close to the anteroventral corner of the centrum, and has an oval outline, with the long axis of the oval aligned dorsoventrally. Posterodorsal to the parapophysis there is a deep blind fossa on the lateral surface of the centrum immediately below the neurocentral suture—a small nutrient foramen is present within this fossa on the left side. The centrum has a low median ridge on its ventral surface, resembling the condition in Garjainia madiba (Gower et al. 2014) and Erythrosuchus africanus (Gower 2003). By contrast, Sarmatosuchus otschevi lacks a ventral keel on its posterior cervical and cervico-dorsal centra (PIN 2865/68).
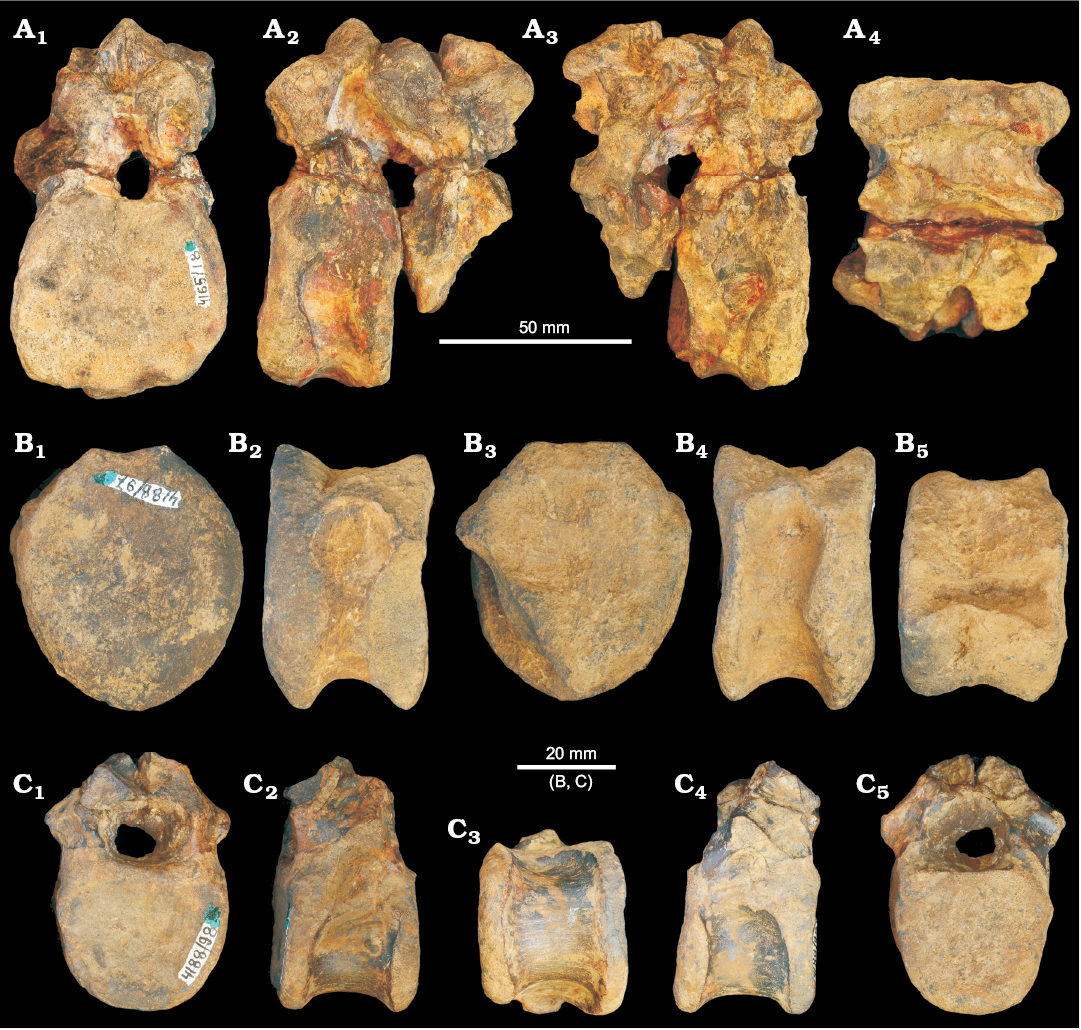
Fig. 11. Vertebrae referred by Ochev (1980) to erythrosuchid archosauriform Chalishevia cothurnata Ochev, 1980, but here considered as Erythrosuchidae indet., from Sol-Iletsk district, Orenburg Province, Russia; Bukobay Gorizont, Ladinian, late Middle Triassic. A. Two cervico-dorsal vertebrae (PIN 4165/18) from the Bukobay I locality in anterior (A1), left lateral (A2), right lateral (A3), and ventral (A4) views. B. Centrum of cervical vertebra (PIN 4188/97) from the Bukobay V locality in anterior (B1), left lateral (B2), posterior (B3), right lateral (B4), and dorsal (B5) views. C. Dorsal vertebra (PIN 4188/98) from the Bukobay V locality in anterior (C1), left lateral (C2), ventral (C3), right lateral (C4), and posterior (C5) views.
The neural arch is poorly preserved in both vertebrae and the neural spines are broken away. The neural canal is dorsoventrally deep with an oval outline. The base of the transverse process is partially preserved only on the right side of the second vertebra and its base is placed lateral to the dorsolateral corner of the neural canal and at about mid length of the neural arch, resembling the condition in the cervico-dorsal vertebrae of Garjainia prima (Ezcurra et al. 2019) and Sarmatosuchus otschevi (Gower and Sennikov 1997). The transverse process is strongly downturned but is eroded at its distal end such that the diapophyseal articulation is not preserved. Ventral to the transverse process, an anteroposteriorly narrow and dorsoventrally deep fossa is present (clearly visible on the right side of the second vertebra) and is bounded by a sharp and narrow anterior centrodiapophyseal lamina, and a thick posterior centrodiapophyseal lamina. A deep and broad fossa is also present anterior to the transverse process and is bounded by the anterior centrodiapophyseal lamina and a very broad ridge in an equivalent position to the prezygodiapophyseal lamina. Posterior to the transverse process is a poorly developed third fossa, delimited by the posterior centrodiapophyseal and postzygodiapophyseal laminae. The presence of the set of four laminae originating from the diapophysis also occurs in other erythrosuchids (Ezcurra 2016). The pre- and postzygapophyses are large but project only short distances beyond the anterior and posterior articular surfaces. There are deep pre-and postspinal fossae. There is no development of a fossa on the neural arch lateral to the base of the neural spine. The base of the neural spine is subrectangular in cross-section, being approximately 1.2 times anteroposteriorly longer than transversely wide, and located approximately at mid-length on the neural arch.
Two isolated partial vertebrae from the Bukobay V locality have also been previously assigned to Chalishevia cothurnata. PIN 4188/97 is a relatively deep and short (superficially erythrosuchid-like) cervical centrum (Fig. 11B), missing the neural arch and damaged at its posterolateral corner. The ventral parts of the anterior and posterior articular faces are slightly bevelled, but it is unclear if these represent articular surfaces for intercentra. The anterior and posterior articular faces have shield shaped outlines and concave surfaces. The centrum is anteroposteriorly compressed and the lateral surfaces are gently concave in ventral view with shallow blind fossae immediately ventral to the neurocentral suture. A low median ridge is present on the ventral margin, as is the case in Sarmatosuchus otschevi (PIN 2865/68), Guchengosuchus shiguaiensis (Butler et al. 2019), Garjainia prima (Ezcurra et al. 2019) and Erythrosuchus africanus (Gower 2003). The parapophysis is placed very low on the anterior margin of the centrum. The centrum has a maximum dorsoventral height of the anterior face of 52.7 mm; the maximum width of the anterior face is 45 mm; and the anteroposterior length of centrum is 32 mm.
PIN 4188/98 is a probable dorsal vertebra that includes the neural arch, but lacking the neural spine, pre- and postzygapophyses, and diapophyses (Fig. 11C). It generally resembles an early archosauriform, but little more can be said about its taxonomic identity. The neurocentral suture is visible and unfused. The vertebra is probably too small to belong to the same individual as PIN 4188/97 from the same locality. The centrum has a maximum dorsoventral height of the anterior face of 30 mm; the maximum width of the anterior face is 34 mm; and the anteroposterior length of centrum is 28 mm. The articular surfaces of the centrum are subcircular with flattened dorsal margins and the anterior face is gently concave whereas the posterior face is flattened. In lateral view, the parapophysis is not clearly recognizable, but may be an eroded, thickening at the dorsal margin of the anterior end of the centrum. There is no ventral keel or ridge on the centrum and the lateral surfaces are gently concave, as occurs in the cervico-dorsal and dorsal centra of other erythrosuchids (Ezcurra et al. 2019). The transverse process would have been at least slightly downturned (although only the base is preserved), and the bases of anterior centrodiapophyseal (or possibly paradiapophyseal, if the parapophysis was indeed on the centrum), posterior centrodiapophyseal, and pre- and postzygodiapophyseal laminae are preserved, delimiting fossae. There is no fossa on the neural arch dorsal to the transverse process.
Concluding remarks
Age of the Bukobay Gorizont and the stratigraphic distribution of Erythrosuchidae.—The faunal assemblage of the Bukobay Gorizont, which is widespread in the south Cis-Ural, north Cis-Ural and Caspian regions, is relatively poorly known. In the south Cis-Ural region this assemblage includes the temnospondyls “Mastodonsaurus” torvus, Bukobaja enigmatica, Cyclotosauridae gen. indet. and Plagiorophus danilovi (Shishkin et al. 1995), the archosauromorph Malutinisuchus gratus (Ochev 1986), the archosauriforms Chalishevia cothurnata, Jushatyria vjushkovi and Energosuchus garjainovi (Kalandadze and Sennikov 1985; Ochev 1986; Sennikov 1995, 2008; Gower and Sennikov 2000), indeterminate small therapsids (theriodonts), and the dicynodonts Elephantosaurus jachimovitchi and Elatosaurus facetus (Shishkin et al. 1995; Battail and Surkov 2000). In the north Cis-Ural region the Bukobay assemblage includes the temnospondyl Bukobaja (?) sp., the chroniosuchian Synesuchus muravjevi (Novikov and Shishkin 2000), the sauropterygian Pistosaurus (?) sp. (Sennikov 2008), the archosauromorph Malutinisuchus gratus (Sennikov 2008), the archosauriform Energosuchus garjainovi (Sennikov 2008) and the dicynodont Planitorostris pechoriensis (Surkov 1999). In the Caspian region this assemblage comprises the temnospondyls “Mastodonsaurus” torvus, Bukobaja sp., and Plagioscutum caspiense (Shishkin et al. 2000), indeterminate procolophonid remains (Sennikov personal observation), indeterminate archosauriforms and interminate dicynodonts (AGS personal observation). The Bukobay Gorizont has been dated to the Ladinian and usually correlated with the Lettenkeuper (Erfurt Formation) of Germany based on some similarities in the temnospondyl faunas between the two areas, the presence of a plant assemblage in the Bukobay belonging to the Scytophyllum Flora, of Ladinian–Carnian age, and the presence in the Bukobay of a miospore assemblage similar to that of the Ladinian of Germany (Shishkin et al. 1995, 2000).
If a Ladinian age for the Bukobay Gorizont is correct, then Chalishevia cothurnata may represent the stratigraphically youngest known erythrosuchid. The stratigraphically oldest occurrences of the clade are known from the late Early Triassic (Olenekian) of South Africa (Garjainia madiba) and Russia (Garjainia prima) (Gower et al. 2014; Ezcurra et al. 2019), and the latest Early Triassic to earliest Middle Triassic of China (Fugusuchus hejiapanensis; Ezcurra et al. 2019). The other known erythrosuchids, Guchengosuchus shiguaiensis, Erythrosuchus africanus, Uralosaurus magnus, and Shansisuchus shansisuchus, are all dated to the Anisian (Ezcurra et al. 2013; Ezcurra 2016; Butler et al. 2019), although the range of the genus Shansisuchus may extend into the early Ladinian (Liu and Sullivan 2017). Cuyosuchus huenei, from the Late Triassic of Argentina, has been referred to Erythrosuchidae (Charig and Reig 1970), but this has not been supported by recent analyses (Desojo et al. 2002; Ezcurra 2016). Thus, the currently known stratigraphic range of Erythrosuchidae extends from the late Olenekian to the Ladinian.
Reconstruction of the skull of Chalishevia cothurnata.—Ochev (1980) reconstructed the anterior part of the skull of Chalishevia cothurnata in lateral view, showing strongly downturned nasals, and inferred a comparatively small premaxilla, and an accessory antorbital fenestra considerably smaller in relative size than that of Shansisuchus shansisuchus (Young 1964; Wang et al. 2013). Gower and Sennikov (2000) disputed this reconstruction, arguing that the left nasal of the holotype had been crushed during preservation, leading to an artificial shortening and tapering of the snout. We agree with the arguments of Gower and Sennikov (2000). As discussed above, a major crack extends from posterodorsal to anteroventral through the contact between the main body of the nasal and the ventral process. The effect of this is to cause the nasal to appear more strongly anteroventrally downturned in lateral view (and the posterior end of the accessory antorbital fenestra more narrowly rounded) than would have originally been the case (Fig. 12A). The exact original orientation of the nasal relative to the maxilla cannot be determined with certainty, but it would certainly have been directed less strongly anteroventrally (Fig. 12B), and the accessory antorbital fenestra would have large, and similar in relative size to that of Shansisuchus shansisuchus.
In Garjainia prima, Erythrosuchus africanus, and Shansisuchus shansisuchus, the anteroposterior length of the maxilla along the alveolar margin, from the anterior contact with the premaxilla to the posterior contact with the jugal, is slightly less than 40% of the basal skull length (Gower 2003: fig. 1; Wang et al. 2013: fig. 2; Ezcurra et al. 2019: fig. 2). Assuming similar skull proportions in Chalishevia cothurnata, this suggests that given a preserved length of the maxilla of 31 cm, total basal skull length in the latter taxon would have been approximately 80 cm (a similar skull length of 80−90 cm was estimated originally by Ochev 1980), making it one of the largest erythrosuchid specimens known to date, and consistent with a general pattern of larger body sizes in stratigraphically younger erythrosuchids.
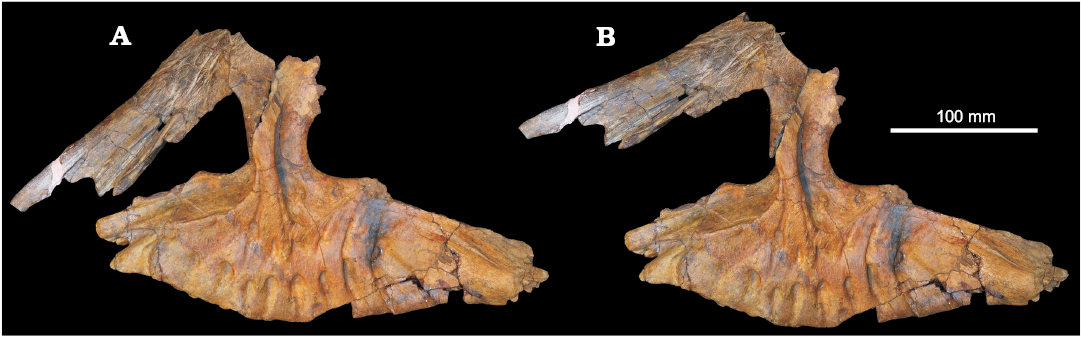
Fig. 12. Reconstruction of maxilla and left nasal of erythrosuchid archosauriform Chalishevia cothurnata Ochev, 1980, in articulation as preserved (A), and close to the articulation reconstructed by Ochev (1980). This reconstruction produces a strongly downturned nasal, but is incorrect due to damage to the nasal between the main body and the ventral process. An alternative reconstruction (B) shows a more anteriorly directed nasal, providing space to accommodate a Shansisuchus-like premaxilla and a larger accessory antorbital fenestra.
Phylogenetic relationships and comparisons to Shansisuchus.—As noted by Gower and Sennikov (2000), there are strong similarities between the anatomy of Chalishevia cothurnata and that of the erythrosuchid Shansisuchus spp. from the Anisian–Ladinian of China (Young 1964; Wang et al. 2013; Liu and Sullivan 2017). Most notably, the nasal of Shansisuchus shares with that of Chalishevia cothurnata the presence of an elongate, narrow notch for articulation with a slender postnarial process of the premaxilla, and the presence of a large accessory antorbital fenestra (= subnarial fenestra) bordered by the maxilla, nasal, and premaxilla (Young 1964; Wang et al. 2013; Liu and Sullivan 2017). These features are absent in other erythrosuchids, were incorporated into a revised differential diagnosis of Shansisuchus shansisuchus by Wang et al. (2013), and provide strong support for a sister taxon relationship between the two species (Gower and Sennikov 2000; Ezcurra 2016). This relationship is also consistent with the relatively late stratigraphic position of the two species (see above). Assessment of differences between Chalishevia cothurnata and Shansisuchus shansisuchus (and thus a detailed consideration of whether the former might be a junior subjective synonym of the latter) is complicated by the relatively brief anatomical descriptions available for the latter taxon, the unresolved species level taxonomy of the latter taxon, and the limited material available for the former taxon. However, the oblique, anteroventrally-to-posterodorsally oriented ventral border of the antorbital fossa that is present in Chalishevia cothurnata appears to be absent in Shansisuchus shansisuchus (Wang et al. 2013: fig. 2). Ezcurra (2016) identified this character state as an autapomorphy of Chalishevia cothurnata.
Acknowledgements
We thank Luke Meade (University of Birmingham, UK) for taking photographs of the pterygoid. DJG thanks Mike Benton (University of Bristol, UK) for supervising his NERC scholarship-funded Ph.D. research that first allowed him to examine Chalishevia. Liu Jun (Institute of Vertebrate Paleontology and Paleoanthropology, Beijing, China) and David Dilkes (University of Wisconsin, Oshkosh, USA) provided helpful reviews, and we also thank the editor Steve Brusatte for handling the manuscript. This research was supported by an International Exchange grant co-funded by the Royal Society (IEC\R2\170064 to RJB) and the Russian Foundation for Basic Research (RFBR № 17-54-10013 to AGS). AGS was also funded by the Russian Foundation for Basic Research through the research project No. 17-04-00410, and by a subsidy of the Russian Government to support the Program of “Competitive Growth of Kazan Federal University among World’s Leading Academic Centers”.
References
Battail, B. and Surkov, M.V. 2000. Mammal-like reptiles from Russia. In: M.J. Benton, E.N. Kurochkin, M.A. Shishkin, and D.M. Unwin (eds.), The Age of Dinosaurs in Russia and Mongolia, 86–119. Cambridge University Press, Cambridge.
Bertin, T.J., Thivichon-Prince, B., LeBlanc, A.R., Caldwell, M.W., and Viriot, L. 2018. Current perspectives on tooth implantation, attachment, and replacement in Amniota. Frontiers in Physiology 9: 1630. Crossref
Butler, R.J., Ezcurra, M.D., Liu, J., Sookias, R.B., and Sullivan, C. 2019. The anatomy and phylogenetic position of the erythrosuchid archosauriform Guchengosuchus shiguaiensis from the earliest Middle Triassic of China. PeerJ 7: e6435. Crossref
Charig, A.J. and Reig, O.A. 1970. The classification of the Proterosuchia. Biological Journal of the Linnean Society 2: 125–171. Crossref
Desojo, J.B., Arcucci, A.B., and Marsicano, C.A. 2002. Reassessment of Cuyosuchus huenei, a Middle–Late Triassic archosauriform from the Cuyo Basin, west-central Argentina. New Mexico Museum of Natural History and Science Bulletin 21: 143–148.
Dilkes, D.W. 1998. The Early Triassic rhynchosaur Mesosuchus browni and the interrelationships of basal archosauromorph reptiles. Philosophical Transactions of the Royal Society of London B 353: 501–541. Crossref
Ezcurra, M.D. 2016. The phylogenetic relationships of basal archosauromorphs, with an emphasis on the systematics of proterosuchian archosauriforms. PeerJ 4: e1778. Crossref
Ezcurra, M.D. and Butler, R.J. 2015. Post-hatchling cranial ontogeny in the Early Triassic diapsid reptile Proterosuchus fergusi. Journal of Anatomy 226: 387–402. Crossref
Ezcurra, M.D., Butler, R.J., and Gower, D.J. 2013. “Proterosuchia”: the origin and early history of Archosauriformes. In: S.J. Nesbitt, J.B. Desojo, and R.B. Irmis (eds.), Anatomy, Phylogeny and Palaeobiology of Early Archosaurs and Their Kin. Geological Society of London, Special Publications 379: 9–33. Crossref
Ezcurra, M.D., Gower, D.J, Sennikov, A.G., and Butler, R.J. 2019. The osteology of the holotype of the early erythrosuchid Garjainia prima Ochev, 1958 (Diapsida: Archosauromorpha) from the upper Lower Triassic of European Russia. Zoological Journal of the Linnean Society 185: 717–783. Crossref
Ezcurra, M.D., Lecuona, A., and Martinelli, A. 2010. A new basal archosauriform diapsid from the Lower Triassic of Argentina. Journal of Vertebrate Paleontology 30: 1433–1450. Crossref
Gauthier, J.A., Kluge, A.G., and Rowe, T. 1988. Amniote phylogeny and the importance of fossils. Cladistics 4: 105–209. Crossref
Gower, D.J. 2003. Osteology of the early archosaurian reptile Erythrosuchus africanus Broom. Annals of the South African Museum 110: 1–84.
Gower, D.J. and Sennikov, A.G. 1997. Sarmatosuchus and the early history of the Archosauria. Journal of Vertebrate Paleontology 17: 60–73. Crossref
Gower, D.J. and Sennikov, A.G. 2000. Early archosaurs from Russia. In: M.J. Benton, E.N. Kurochkin, M.A. Shishkin, and D.M. Unwin (eds.), The Age of Dinosaurs in Russia and Mongolia, 140–159. Cambridge University Press, Cambridge.
Gower, D.J., Hancox, P.J., Botha-Brink, J., Sennikov, A.G., and Butler, R.J. 2014. A new species of Garjainia Ochev, 1958 (Diapsida: Archosauriformes: Erythrosuchidae) from the Early Triassic of South Africa. PLoS ONE 9: e111154. Crossref
Huene, F. von 1946. Die grossen Stämme der Tetrapoden in den geologischen Zeiten. Biologisches Zentralblatt 65: 268–275.
Kalandadze, N.N. and Sennikov, A.G. 1985. New reptiles from the Middle Triassic of the southern Cis-Urals [in Russian]. Paleontologičeskij žurnal 1985 (2): 77–84.
Liu, J. and Sullivan, C. 2017. New discoveries from the Sinokannemeyeria–Shansisuchus Assemblage Zone: 3. Archosauriformes from Linxian, Shanxi, China. Vertebrata PalAsiatica 55: 110–128.
Modesto, S.P. and Sues, H.-D. 2004. The skull of the Early Triassic archosauromorph reptile Prolacerta broomi and its phylogenetic significance. Zoological Journal of the Linnean Society 140: 335–351. Crossref
Novikov, I.V. and Shishkin, M.A. 2000. Triassic chroniosuchians (Amphibia, Anthracosauromorpha) and the evolution of the trunk dermal ossifications in the bystrowianids. Paleontological Journal 34 (Supplement): S165–S178.
Ochev, V.G. [Očev, V.G.] 1980. New archosaurs from the Middle Triassic of the southern Cis-Urals [in Russian]. Paleontologičeskij žurnal 1980 (2): 101–107.
Ochev, V.G. [Očev, V.G.] 1986. On Middle Triassic reptiles of the southern Cis-Urals [in Russian]. Ežegodnik Vsesoûznogo Paleontologisčeskogo Obŝestva 29: 171–180.
Sennikov, A.G. 1995. Early thecodonts of Eastern Europe [in Russian]. Trudy Paleontologičeskogo Instituta RAN 263: 1–141.
Sennikov, A.G. 2008. Subclass Archosauromorpha [in Russian]. In: M.F. Ivahnenko and E.N. Kuročkin (eds.), Iskopaemye pozvonočnye Rossii i sopredel'nyh stran: Iskopaemye reptilii i pticy. Čast' 1, 266–318. GEOS, Moskva.
Shishkin, M.A. [Šiškin, M.A.], Ochev, V.G. [Očev, V.G.], Tverdokhlebov, V.P. [Tverdohlebov, V.P.], Vergay, I.F. [Vergaj, I.F.], Gomankov, A.V., Kalandadze, N.N., Leonova, E.M., Lopato, A.Y., Makarova, I.S., Minikh, M.G. [Minih, M.G.], Molostovsky, E.M. [Molostovskij, E.M.], Novikov I.V., and Sennikov, A.G. 1995. Biostratigrafiâ kontinental'nogo triasa Ûžnogo Priuralâ. 206 pp. Nauka, Moskva.
Shishkin, M.A., Ochev, V.G., Lozovskii, V.R., and Novikov, I.V. 2000. Tetrapod biostratigraphy of the Triassic of eastern Europe. In: M.J. Benton, E.N. Kurochkin, M.A. Shishkin, and D.M. Unwin (eds.), The Age of Dinosaurs in Russia and Mongolia, 120–139. Cambridge University Press, Cambridge.
Surkov, M.V. 1999. A New Middle Triassic Kannemeyeriin from the Pechora District [in Russian]. Paleontologičeskij žurnal 1999 (4): 83–84.
Tverdokhlebov, V.P., Tverdokhlebova, G.I., Surkov, M.V., and Benton, M.J. 2003. Tetrapod localities from the Triassic of the SE of European Russia. Earth Science Reviews 60: 1–66. Crossref
Wang, R., Xu, S., Wu, X., Li, C., and Wang, S. 2013. A new specimen of Shansisuchus shansisuchus Young, 1964 (Diapsida: Archosauriformes) from the Triassic of Shanxi, China. Acta Geologica Sinica 87: 1185–1197. Crossref
Watson, D.M.S. 1917. A sketch classification of the Pre-Jurassic tetrapod vertebrates. Proceedings of the Zoological Society of London 1917: 167–186. Crossref
Young, C.-C. 1964. The pseudosuchians in China. Palaeontologia Sincia new series C 151: 1–205.
Acta Palaeontol. Pol. 64 (4): 757–774, 2019
https://doi.org/10.4202/app.00648.2019