

Hindlimb morphology of Palaeotis suggests palaeognathous affinities of the Geranoididae and other “crane-like” birds from the Eocene of the Northern Hemisphere
GERALD MAYR
Mayr, G. 2019. Hindlimb morphology of Palaeotis suggests palaeognathous affinities of the Geranoididae and other “crane-like” birds from the Eocene of the Northern Hemisphere. Acta Palaeontologica Polonica 64 (4): 669–678.
The early/middle Eocene Palaeotis weigelti is a flightless bird, which occurs in the fossil localities Messel and Geiseltal (Germany). The species is assigned to the Palaeognathae and some authors considered it to be a stem group representative of the Struthionidae (ostriches). Even though several partial skeletons have been found, the osteology of P. weigelti is incompletely known. In the present study, new details of the hindlimb morphology of the species are reported based on unpublished and previously described fossils from the Geiseltal. These data show that the recently described Galligeranoides boriensis from the early Eocene of southern France is another representative of the Palaeotididae and the oldest record of the taxon. It is further noted that Palaeogrus princeps from the middle Eocene of Italy, which was previously assigned to the Gruidae (cranes), may be another representative of the Palaeotididae. Galligeranoides was before assigned to the North American Geranoididae, a taxon mainly known from hindlimb elements. The Geranoididae are usually considered to be closely related to the Asian Eogruidae and both taxa are currently classified in the Gruiformes (cranes and allies). However, as detailed in the present study, derived similarities suggest close affinities between the Palaeotididae and Geranoididae. Eogruids were identified as stem group representatives of the palaeognathous Struthionidae by some earlier authors, and if close affinities between Palaeotididae and Geranoididae are corroborated in future analyses, palaeognathous affinities of the Eogruidae need to be critically revisited.
Key words: Aves, Palaeotididae, Struthionidae, evolution, systematic, Paleogene, Germany.
Gerald Mayr [Gerald.Mayr@senckenberg.de], Senckenberg Research Institute and Natural History Museum Frankfurt, Ornithological Section, Senckenberganlage 25, D-60325 Frankfurt am Main, Germany.
Received 21 June 2019, accepted 17 August 2019, available online 18 October 2019.
Copyright © 2019 G. Mayr. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Extant birds are divided into two major clades, the Palaeognathae (Tinamidae [tinamous] and the large flightless “ratites” [ostriches, emus, and allies]) and the Neognathae (all other species). In the early Paleogene of Europe and North America, palaeognathous birds are mainly represented by the volant Lithornithidae (Houde 1988; Mayr 2009), but there were also palaeognathous birds in the early Paleogene of Europe, which were identified as close relatives of some of the flightless “ratites”. The best known of these is Palaeotis weigelti from the early and middle Eocene of Germany, of which several partial skeletons were found in the fossil sites Geiseltal and Messel.
This species was initially considered to be a gruiform bird and based on hindlimb elements of two different individuals, it was actually described twice, as a bustard (Otididae; Lambrecht 1928: Palaeotis weigelti) and a crane (Gruidae; Lambrecht 1935: Palaeogrus geiseltalensis). Houde (1986) and Houde and Haubold (1987) first recognized that this long-legged bird was flightless and identified it as a stem group representative of ostriches (Struthionidae). Peters (1988) described a skeleton of P. weigelti from Messel, considered the species to be more closely related to the South American Rheidae, and assigned it to its own family-level taxon, Palaeotididae. The affinities of Palaeotis were revisited by Mayr (2015), but the analyses performed in this study did not yield congruent and well-supported results. An unconstrained analysis of morphological data resulted in a sister group relationship between Palaeotis and a clade including Struthionidae, Rheidae, and Casuariidae, which is not supported by analyses of molecular data. Analyses that were constrained to the tree topologies recovered by DNA sequence data either resulted in a sister group relationship between Palaeotis and a clade including Rheidae, Aepyornithidae, Apterygidae, and Casuariidae or recovered Struthionidae and Palaeotis in a polytomy together with two clades comprising all other palaeognathous birds.
Until recently, remains of the Palaeotididae were only found in the German fossil sites Geiseltal and Messel. However, Mayr (2015, 2016) briefly noted that leg bones from the early Eocene of southern France, which were described as Galligeranoides boriensis by Bourdon et al. (2016), closely resemble those of Palaeotis weigelti. Galligeranoides boriensis was assigned to the Geranoididae by Bourdon et al. (2016), who considered the species to be the first European record of the Geranoididae. These latter birds are classified in the Gruiformes (cranes and allies), and their fossil record was before restricted to the Eocene of North America (Cracraft 1969; Mayr 2009, 2016). The species of the Geranoididae are only known from hindlimb elements, and the exact composition of the taxon as well as the taxonomy of the included species are poorly resolved. Cracraft (1969) recognized five genus-level taxa, but Mayr (2016) synonymized Geranoides Wetmore, 1933 with Palaeophasianus Shufeldt, 1913 and hypothesized that Eogeranoides Cracraft, 1969 may be a synonym of Paragrus Lambrecht, 1933; another taxon assigned to the Geranoididae by Cracraft (1969) is Geranodornis Cracraft, 1969.
Geranoidids were considered closely related to the Eogruidae from the late Eocene of Mongolia and China (Cracraft 1973; Olson 1985). These latter birds likewise include long-legged species and are characterized by a reduced trochlea for the second toe (Kurochkin 1976, 1981; Clarke et al. 2005). Most Neogene species of the Eogruidae feature a complete loss of the second toe and, hence, a didactyl foot. These birds were previously distinguished as a separate taxon Ergilornithidae, which was synonymized with the Eogruidae by Clarke et al. (2005). Eogruids are also mainly known from hindlimb elements and it has been assumed that they were ancestral to the didactyl palaeognathous Struthionidae (Burchak-Abramovich 1951; Feduccia 1980; Olson 1985). However, most authors regarded the striking reduction of the second toe in eogruids (“ergilornithids”) and struthionids to be the result of convergence and assigned eogruids to the Gruiformes (Cracraft 1973; Clarke et al. 2005).
With Palaeotis weigelti having been identified as a palaeognathous bird and possibly even a stem lineage representative of the Struthionidae, and with geranoidids and eogruids having been considered to be stem group representatives of the Struthionidae (Olson 1985), comparisons between Palaeotis and the Geranoididae and Eogruidae suggest themselves. Unfortunately, and as noted above, Galligeranoides and the species of the Geranoididae and Eogruidae are almost exclusively based on leg bones, whereas the morphology of the hindlimb elements of Palaeotis is insufficiently known and important features have not yet been described. This is particularly true for the distal end of the tibiotarsus, which exhibits important morphological features characterizing species of the Geranoididae and palaeognathous birds, but is poorly exposed in the previously known specimens of Palaeotis.
In addition to a fossil from Messel, Houde and Haubold’s (1987) study was based on three specimens of P. weigelti from the Geiseltal, but the material from the latter locality is actually more comprehensive (Fig. 1) and some of the unpublished fossils show important features, which bear on the affinities of the species. Based on these specimens, previously unknown features of the hindlimb morphology of Palaeotis are described, which show that Galligeranoides is indeed a representative of the Palaeotididae. As detailed in the following, these data shed new light on the affinities of the Geranoididae.
Institutional abbreviations.—AMNH, American Museum of Natural History, New York, USA; GMH, Geiseltalmuseum, Martin-Luther Universität, Halle, Germany; MNHN, Muséum National d’Histoire Naturelle, Paris, France; SMF, Senckenberg Research Institute Frankfurt, Germany.
Material and methods
The present study is based on an examination of the following Palaeotis specimens from the Geiseltal: GMH 3402 (tibiotarsus lacking distal end, intratendinous ossification, and pedal phalanges; newly identified); GMH 4361 (tarsometatarsi; studied by Houde and Haubold 1987); GMH 4362 (partial skeleton; studied by Houde and Haubold 1987); GMH 4363 (pelvis; newly identified); GMH 4415, 4416, 4418 (holotype: pedal phalanx, distal end of left tarsometatarsus, and fragmentary wing phalanx; studied by Lambrecht 1928 and Mayr 2002); GMH 5882 (holotype of Palaeogrus geiseltalensis, right tibiotarsus and tarsometatarsus; studied by Lambrecht 1935 and Houde and Haubold 1987); GMH XXXVIII-6-1964 (partial skeleton; newly identified); GMH IX-566-1953 (proximal end of right tarsometatarsus; newly identified). The referral of a distal tarsometatarsus (GMH 3605) to P. weigelti by Mayr (2002) is here regarded uncertain, since the specimen shows some differences in the proportions of the trochleae. Specimens of the Geranoididae and Eogruidae were studied in the collection of AMNH, analysis of Galligeranoides boriensis is based on published data.
Results
As mentioned in the introduction, the morphology of the distal end of the tibiotarsus of Palaeotis weigelti is poorly known, and none of the previously described fossils shows the cranial surface of the bone and the shape of the condylus medialis. In GMH XXXVIII-6-1964 (Fig. 1A), the distal end of the right tibiotarsus is well exposed (Fig. 2A), even though the lateral part is broken. Most importantly, the bone exhibits a proximodistally broad pons supratendineus, which bridges a wide and medially situated sulcus extensorius. Regarding these features, the distal tibiotarsus of Palaeotis is very unlike that of Remiornis, a flightless palaeognathous birds from the Paleocene of France, which was considered to be possibly related to the Palaeotididae by Mayr (2009), and which lacks a supratendinal bridge and an extensor sulcus (Martin 1992). A supratendinal bridge and an extensor sulcus are also absent in Diogenornis (Alvarenga 1983; Agnolin 2017), a flightless palaeognathous bird from the Itaboraí fossil site in Brazil (the deposits of this locality were considered to be of late Paleocene age by earlier authors, but recent studies suggest an early Eocene age; Woodburne et al. 2014). The supratendinal bridge is likewise absent in the early Paleogene Lithornithidae, which have a narrow sulcus extensorius (Houde 1988). In crown group Palaeognathae, a supratendinal bridge is only found in the Tinamidae, some Apterygidae, and the extinct Dinornithidae; a sulcus extensorius is either absent (Struthionidae, Rheidae, Casuariidae, Aepyornithidae) or narrow (Apterygidae, Tinamidae, Dinornithidae). Because the lateral portion of the tibiotarsus of GMH XXXVIII-6-1964 is damaged, the shape of the condylus lateralis cannot be assessed. The distal rim of the condylus medialis bears a distinct notch (Fig. 2A4; contra Mayr 2016), which amongst neognathous birds is found in the Gruiformes, Podicipediformes, Phoenicopteriformes, Procellariiformes, Charadriiformes, and a few other taxa (Mayr and Clarke 2003), but which does not occur in palaeognathous birds. On the medial surface of the distal end, there is a marked ligamental scar (Fig. 2A4), which is also found in extant flightless palaeognathous birds (Bourdon et al. 2009).

Fig. 1. Fossils of the palaeotidid bird Palaeotis weigelti Lambrecht, 1928, all specimens are from the middle Eocene of the Geiseltal, Germany. A. GMH XXXVIII-6-1964, partial skeleton. B. GMH 5882, right tibiotarsus and tarsometatarsus; holotype of Palaeogrus geiseltalensis. C. GMH 3402, tibiotarsus lacking distal end, intratendinous ossification, and pedal phalanges. D. GMH 4363, pelvis. E. GMH 4361, tarsometatarsi. F. GMH 4362, partial skeleton. G. GMH 4415, 4416, 4418, holotype: distal end of left tarsometatarsus, pedal phalanx, and possible wing phalanx. H. GMH IX-566-1953, proximal end of right tarsometatarsus. Scale bars 50 mm.
The distal end of the tibiotarsus of Palaeotis weigelti corresponds well with that of Galligeranoides boriensis in its proportions and the configurations of the pons supratendineus and sulcus extensorius (compare Fig. 2A, C, D). However, Bourdon et al. (2016: 183) described a tubercle “at the distal part of the pons supratendineus” of G. boriensis that “is strongly projecting […] and is continuous with a strong crest which corresponds to the tuberositas retinaculi extensori.” In P. weigelti (GMH XXXVIII-6-1964), a well-defined tubercle is absent, but may have been broken, with the corresponding area showing some damage. Lateral of the pons supratendineus, a transverse ridge runs perpendicular to the longitudinal axis of the tibiotarsus; a fossa distal of this ridge probably encompassed the eminentia intercotylaris of the tarsometatarsus (Fig. 2A2). A cranially raised bulge proximolateral of the supratendinal bridge (Fig. 2A2) seems to correspond to the “strong crest” reported by Bourdon et al. (2016) for G. boriensis. This area is less strongly developed than in G. boriensis, in which, however, the distal tibiotarsus is either deformed (holotype) or crushed (paratype).
Bourdon et al. (2016) compared the distal tibiotarsus of Galligeranoides with that of the Geranoididae, and although these authors noted some differences, the similarities prevail. The distal tibiotarsus of P. weigelti agrees with that of geranoidids in that the distal rim of the condylus medial bears a distinct notch (the poor development of this notch in the specimen referred to “Eogeranoides”/Paragrus shown in Fig. 2F2 is here considered to be a preservational artefact). Bourdon et al. (2016: 12) considered the distal tibiotarsus of Galligeranoides to be very different from that of the Eogruidae, for which they noted, amongst others, a more centrally positioned canalis extensorius with wider openings, and a proportionally shorter pons supratendineus. However, these differences are not greater than those that exist between the Eogruidae and Geranoididae, which were considered closely related by previous authors (Cracraft 1973; Olson 1985).
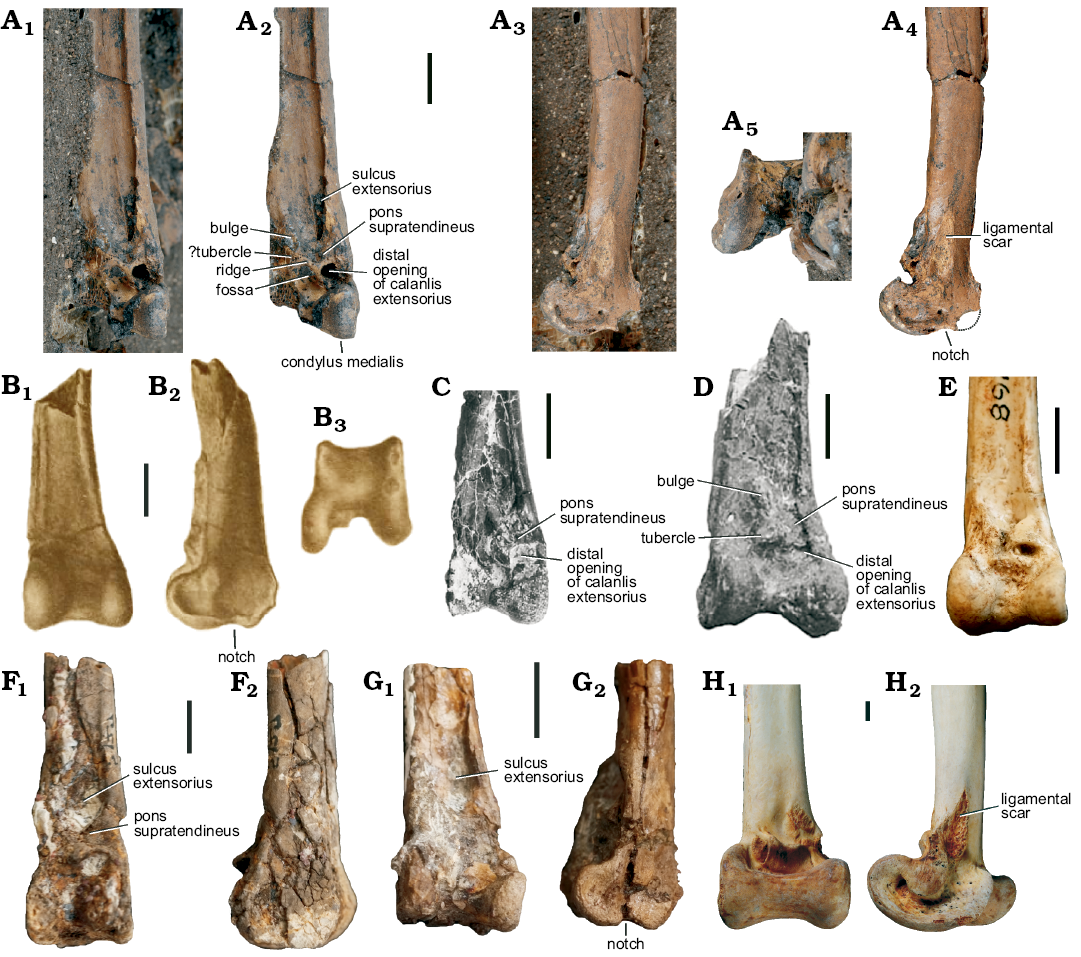
Fig. 2. Tibiotarsi of palaeotidid, geranoidid, eogruid, and struthionid birds. A. Palaeotis weigelti Lambrecht, 1928 (Palaeotididae) from the middle Eocene of the Geiseltal, Germany, GMH XXXVIII-6-1964; distal end of right tibiotarsus in cranial (A1, A2), medial (A3, A4), and distal (A5) views; in A2 and A4 surrounding matrix was digitally removed, the dotted line in A4 indicates the reconstructed outline of the missing caudal portion of the condylus medialis. B. Palaeogrus princeps Portis, 1885 (?Palaeotididae) from the middle Eocene of Italy; distal end of left tibiotarsus in cranial (B1), medial (B2), and distal (B3) views (from Portis 1885; mirrored). C. Galligeranoides boriensis Bourdon, Mourer-Chauviré, and Laurent, 2016 (Palaeotididae) from the early Eocene of southern France, paratype: MHNT.PAL.2013.16.1; distal end of right tibiotarsus in cranial view (from Bourdon et al. 2016). D. Galligeranoides boriensis Bourdon, Mourer-Chauviré, and Laurent, 2016 from the early Eocene of southern France, holotype, MHNT.PAL.2013.16.3; distal end of right tibiotarsus in cranial view (from Bourdon et al. 2016; mirrored). E. Eogrus aeola Wetmore, 1934 (Eogruidae), AMNH 2946; distal end of right tibiotarsus in cranial view (mirrored). F. cf. Eogeranoides campivagus Cracraft, 1969 from the early Eocene of Wyoming, USA, AMNH 5127; distal end of right tibiotarsus in cranial (F1) and medial (F2) views (mirrored). G. Palaeophasianus meleagroides Shufeldt, 1913 (Geranoididae) from the early Eocene of Wyoming, USA, holotype, AMNH 5128; distal end of left tibiotarsus of in cranial (G1) and medial (G2) views (mirrored). H. Extant Struthio camelus Linnaeus, 1758 (Struthionidae), SMF 2438; distal end of right tibiotarsus in cranial (H1) and medial (H2) views. Scale bars 10 mm (for B scale is based on measurements in Portis 1885).
The tarsometatarsus of Palaeotis weigelti is characterized by an unusually deep sulcus extensorius along the dorsal surface of the bone (Mayr 2002), which is nearly as deep as it is mediolaterally wide and is bordered by steeply sloping margins. Bourdon et al. (2016) noted a deep extensor sulcus for Galligeranoides, but considered it to have been exaggerated by crushing. Here it is hypothesized that the similar morphologies shared by Palaeotis and Galligeranoides reflect the actual condition of the extensor sulcus of these taxa. A deep sulcus extensorius is also present in a putative geranoidid from the North American Willwood Formation (Fig. 3E), which was tentatively assigned to Eogeranoides campivagus by Mayr (2016), who regarded it possible that this species is a junior synonym of Paragrus prentici. For Galligeranoides, Bourdon et al. (2016: 183) reported the “trace of a canalis hypotarsi, which opened on the proximal articular surface” of the tarsometatarsus. The hypotarsus of P. weigelti has not yet been described and details of it are not visible in the previously described fossils. In specimen GMH IX-566-1953 (Fig. 3B), the proximal portion of the hypotarsus is damaged, but a shallow central sulcus can be discerned in its distal section (Fig. 3B4), which is here considered homologous to a similar sulcus reported for Galligeranoides by Bourdon et al. (2016). This contrasts with the condition in extant Struthionidae, in which the hypotarsus forms only a single crest (Fig. 3J). In other palaeognathous birds, the hypotarsus is also composed of either a single long crest (Rheidae, Casuariidae) or two short crests (Apterygidae and Dinornithidae), or it is short and block-like (Lithornithidae).
Hypotarsus morphology shows some variation amongst the taxa assigned to the Geranoididae (Mayr 2016; Fig. 3G, I1). The hypotarsus of palaeotidids resembles that of a fossil, which was tentatively referred to Palaeophasianus (AMNH 5156; Fig. 3G), in which it forms a long medial crest that is laterally bordered by a shallow sulcus. The hypotarsus of Eogrus also forms a proximodistally long crest, but exhibits a large canal for the tendon of musculus flexor digitorum longus (Fig. 3F4). Owing to the fragmentary preservation of the fossil specimens it is uncertain whether there was a bony canal in the Palaeotididae, but the preserved portions of the proximal tarsometatarsi of Palaeotis and Galligeranoides rather suggest its absence.
The distal ends of the tarsometatarsi of Palaeotis and Galligeranoides closely agree in the size, proportions, and distal extent of the trochleae. The trochlea metatarsi III is very wide and its mediolateral width exceeds the dorsoplantar depth. Galligeranoides mainly differs from Palaeotis in that the trochlea metatarsi II bears a less medially directed plantar flange (compare Fig. 3C3, D3). The distal tarsometatarsus of Palaeotis and Galligeranoides differs from that of the North American Geranoididae in that the trochlea metatarsi III is mediolaterally much broader and in that the trochleae metatarsorum II and IV have an equal distal extent (in the Geranoididae, the trochlea metatarsi II does not extend as far distally); with regard to these features, palaeotidids also differ from the Eogruidae (Fig. 3F).
A distinctive feature of Palaeotis, which received little attention so far, is the presence of intratendinous ossifications along the plantar surface of the tarsometatarsus These are well visible in GMH 4362 (Figs. 1F, 3A) and GMH 5882 (Fig. 1B), and in GM 3402 (Fig. 1C) only the intratendinous ossifications but not the tarsometatarsus are preserved. Such intratendinous ossifications are unknown from extant flightless palaeognathous birds (Vanden Berge and Storer 1995). In P. weigelti, these ossifications are hypertrophied to a degree found in Gruiformes and Galliformes amongst extant birds. Intratendinous ossifications have not been reported from any fossil of the Geranoididae and Eogruidae, but the plantar surface of the tarsometatarsus of eogruids exhibits a large lateral crest, which may have guided intratendinous ossifications.

Fig. 3. Tarsometatarsi of palaeotidid, geranoidid, eogruid, and struthionid birds. A–C. Palaeotis weigelti Lambrecht, 1928 (Palaeotididae) from the middle Eocene of the Geiseltal, Germany. A. GMH 4362; right tarsometatarsus in dorsal view. B. GMH IX-566-1953; proximal end of right tarsometatarsus in dorsal (B1), plantar (B2, B4), and proximal (B3) views. C. Holotype: GMH 4416; distal end of left tarsometatarsus in dorsal (C1), plantar (C2), and distal (C3) views. D. Galligeranoides boriensis Bourdon, Mourer-Chauviré, and Laurent, 2016 (Palaeotididae) from the early Eocene of southern France, MHNT.PAL.2013.16.2; right tarsometatarsus in dorsal (D1), plantar (D2), distal (D3), and proximal (D4) views (from Bourdon et al. 2016; D1–D3 mirrored). E. cf. Eogeranoides campivagus Cracraft, 1969 (Geranoididae) from the early Eocene of Wyoming, USA, AMNH 5127; left tarsometatarsus in dorsal (E1) and dorsolateral (E2, E3) views (the fossil consists of several fragments, which were assembled for the photo). F. Eogrus aeola Wetmore, 1934 (Eogruidae) from the middle Eocene of Inner Mongolia, China, AMNH 2937; right tarsometatarsus in dorsal (F1), plantar (F2, F5), distal (F3), and proximal (F4) views (all mirrored). G. ?Palaeophasianus sp. (Geranoididae) from the early Eocene of Wyoming, USA, AMNH 5156; proximal end of right tarsometatarsus in proximal (G1) and plantar (G2) views. H. ?Palaeophasianus sp. from the early Eocene of Wyoming, USA, AMNH 5156; distal end of right tarsometatarsus in distal (H1) and plantar (H2) plantar views (H1 mirrored). I. Palaeophasianus meleagroides Shufeldt, 1913 (Geranoididae), holotype: AMNH 5128; proximal end of left tarsometatarsus in proximal view (mirrored). J. Extant Struthio camelus Linnaeus, 1758 (Struthionidae) from the early Eocene of Wyoming, USA, SMF 2438; proximal end of right tarsometatarsus in plantar view. In C3, D3, F3, H1, and H2, the tarsometatarsal trochleae are numbered. Scale bars 10 mm.
Discussion
Affinities of Palaeotis and Galligeranoides.—The tarsometatarsi of Palaeotis weigelti and Galligeranoides boriensis closely agree in their morphologies, and a distinctive shared derived feature is the unusually marked sulcus extensorius along the dorsal surface of the greatly elongated tarsometatarsus; both species furthermore agree in the wide trochlea metatarsi III, whose mediolateral width exceeds its dorsoplantar depth (usually, this trochlea is much deeper than it is wide). The new data on the morphology of the distal tibiotarsus of P. weigelti also conform with palaeotidid affinities of G. boriensis and the species is here formally transferred from the Geranoididae to the Palaeotididae. Most importantly, P. weigelti agrees with G. boriensis and differs from all known fossil and most extant palaeognathous birds in the presence of a proximodistally long supratendinal bridge and a wide sulcus extensorius of the tibiotarsus; unlike in all extant palaeognathous birds, there is a notch in the distal rim of the condylus medialis (for G. boriensis; Bourdon et al. 2016: 183 noted that “the condylus is incompletely preserved but there is an indication of the presence of a notch”). As suggested by Mayr (2015), it is likely that an unidentified ulna described by Bourdon et al. (2016) also belongs to Galligeranoides and, hence, is from a palaeotidid. Recognition of palaeotidid affinities of Galligeranoides restricts the fossil record of the Geranoididae to North America again.
Galligeranoides boriensis stems from the biostratigraphic zone MP 8–9 of the mammalian Paleogene zones (Bourdon et al. 2016). The species therefore has an absolute age of 56–51 mya (Vandenberghe et al. 2012) and is the oldest record of the Palaeotididae, predating the records of Palaeotis weigelti from Messel (MP 11) by at least 3 million years (the specimens from the Geiseltal are as old or younger than those from Messel; Mayr 2002).
The distal end of the tibiotarsus of Palaeotis weigelti is likewise similar to that of Palaeogrus princeps from the middle Eocene of Italy in those morphological features that can be discerned in the published figures of the holotype (Fig. 2B). Apart from a similar size and condylar proportions, the distal tibiotarsi of Palaeotis and Palaeogrus share the presence of a notch in the distal rim of the condylus medialis. P. princeps was originally described as a crane (Portis 1885) and is only known from the holotype, a distal tibiotarsus. Its presumed gruiform affinities have not been convincingly established, and based on morphological resemblance, geological age, and geographic provenance, it is here considered more likely that P. princeps is another representative of the Palaeotididae and not a middle Eocene crane (the earliest definitive fossil record of the Gruidae is from the early Miocene; Mayr 2017). Palaeotidid affinities of P. princeps still have to be assessed by examination of the original fossil, but if confirmed, they would reveal Lambrecht’s (1935) assignment of one Palaeotis fossil to Palaeogrus into a different light. Recognition of P. princeps as a member of the Palaeotididae may have potential taxonomic consequences, because the taxon Palaeogrus Portis, 1885 predates both Palaeotis Lambrecht, 1928 and Galligeranoides Bourdon, Mourer-Chauviré, and Laurent, 2016. Even though Bourdon et al. (2016: 186) noted that the tibiotarsus of P. princeps shows the “characteristic shape of the Gruidae, with the medial shift of the condylus medialis”, it is actually very similar to the distal tibiotarsus of the geranoidid Paragrus prentici (compare Fig. 2B3 with Cracraft 1969: fig. 2).
Bourdon et al. (2016) performed detailed comparisons between Galligeranoides and the North American Geranoididae and concluded that the similarities justify a classification of Galligeranoides into the Geranoididae. However, these authors and Mayr (2016) also noted some differences. One of these, the absence of a notch in the distal rim of the condylus medialis of the tibiotarsus, was erroneously listed by Mayr (2016) and—as shown in the present study—this notch is present in Palaeotis (Fig. 2A4). Other differences are confirmed and the most conspicuous of these is the mediolaterally wider trochlea metatarsi III of palaeotidids (compare Fig. 3C3, H1), which, by comparison with other birds, may be a synapomorphy of Palaeotis and Galligeranoides. Specialized locomotory features of Palaeotis are also indicated by marked processus supratrochleares of the ilium (Mayr 2015) and the unusually strongly developed intratendinous ossifications along the tarsometatarsus.
The most complete fossil of the Geranoididae stems from the Willwood Formation and includes a femur, tibiotarsus (Fig. 2F), and tarsometatarsus (Fig. 2E). The specimen (AMNH 5127) was tentatively assigned to Eogeranoides campivagus by Mayr (2016), who considered this species to be possibly synonymous with Paragrus prentici. AMNH 5127 agrees with Palaeotis in the presence of a very deep extensor sulcus along the dorsal surface of the tarsometatarsus (Fig. 3E3), and with regard to this feature, the fossil corresponds better with the Palaeotididae than with gruiform birds, in which the extensor sulcus is not as pronounced as in palaeotidids. Other fossils assigned to the Geranoididae are less complete and comparisons with palaeotidids are less straightforward. Geranoides, the type genus of the Geranoididae, was synonymized with Palaeophasianus by Mayr (2016), and the morphology of the fossils assigned to both taxa also conforms with an assignment to the Palaeotididae. The taxon Geranodornis is only known from a fragmentary distal tibiotarsus, which resembles the corresponding bone of palaeotidids in shape and proportions.
In summary, the Geranoididae share the following derived features with the Palaeotididae: (i) tarsometatarsus greatly elongated, (ii) sulcus extensorius along dorsal surface of tarsometatarsus very marked, (iii) hypotarsus proximodistally elongated and forming a long medial crest, (iv) tibiotarsus with distinct notch in distal rim of condylus medialis. Characters (i) and (iv) are also found in gruiform birds, from which geranoidids are distinguished in characters (ii) and (iii).
Here it is concluded that close affinities between Palaeotididae and Geranoididae are at least as well supported as the current classification of geranoidids into the Gruiformes. Close affinities between the European Palaeotididae and the North American Geranoididae would also not be surprising from a biogeographic point of view, because early and middle Eocene avifaunas of Europe and North America had a similar taxonomic composition (Mayr 2009), and the Gastornithidae even document the existence of dispersal routes for flightless birds between Europe, Asia, and North America in the early Eocene.
Higher-level affinities of Palaeotididae and Geranoididae/Eogruidae.—As noted in the introduction, the Geranoididae were considered closely related to the Eogruidae by earlier authors (e.g., Cracraft 1973; Olson 1985), some of whom hypothesized that eogruids are stem group representatives of the palaeognathous Struthionidae (Burchak-Abramovich 1951; Feduccia 1980; Olson 1985). A unique derived feature shared by eogruids and struthionids is the reduction of the tarsometatarsal trochlea for the second toe, which is completely lost in struthionids and shows a progressive diminishment in eogruids. Otherwise, the hindlimb bones of eogruids and struthionids are very different, and whereas the leg bones of eogruids resemble those of gruiform birds, the leg bones of struthionids are similar to those of other large flightless palaeognathous birds.
However, it is important to note that the tibiotarsus and tarsometatarsus of palaeotidids are equally unlike the corresponding bones of struthionids and other flightless palaeognathous birds. No osteological features of the tibiotarsus and tarsometatarsus of palaeotidids unambiguously support palaeognathous affinities of these birds, and if it were not for the fact that more bones than hindlimb elements are known from Palaeotis, palaeognathous affinities of this taxon are unlikely to have been considered. In fact, Palaeotis and Galligeranoides are clearly distinguished from extant palaeognathous birds in a number of hindlimb features, in which they resemble gruiform birds, as evidenced by previous classifications of Palaeotis (Lambrecht 1928), Galligeranoides (Bourdon et al. 2016), and, possibly, Palaeogrus (Portis 1885). In particular, the tibiotarsus of P. weigelti differs from that of other early Paleogene Palaeognathae in that a supratendinal bridge is present (an important feature that contradicts previous assumptions by Mayr 2009 that Palaeotis may form a clade together with Remiornis from the late Paleocene of France), and the bone differs from the tibiotarsus of all Palaeognathae in the presence of a notch in the distal rim of the condylus medialis. The hypotarsus of palaeotidids is distinguished from that of palaeognathous birds, in which it is either short and block-like (Lithornithidae), forms two short crests (Apterygidae and Dinornithidae), or exhibits a single long crest (Struthionidae, Rheidae, Casuariidae). Intratendinous ossifications along the plantar surface of the tarsometatarsus are not found in any of the extant flightless palaeognathous birds (Vanden Berge and Storer 1995).
The distinct differences to other palaeognathous birds may suggest that Palaeotis has erroneously been assigned to the Palaeognathae. Indeed, palaeognathous affinities of Palaeotis are mainly based on a combination of features, which are found in the flightless palaeognathous birds, the “ratites” of earlier authors. These include long and narrow “praefrontal bones” (= supraorbital processes), an unkeeled sternum, a fused scapulocoracoid, a bilaterally compressed pelvis, and the loss of a hallux (Houde and Haubold 1987: 36). Molecular studies have shown that the flightless Palaeognathae do not form a monophyletic group (Harshman et al. 2008; Philips et al. 2010; Haddrath and Baker 2012; Smith et al. 2013; Baker et al. 2014; Mitchell et al. 2014; Yonezawa et al. 2017), so that most of the above characters evolved more than once, which diminishes their phylogenetic significance. Houde and Haubold (1987) also mentioned the presence of an open frontoparietal suture in Palaeotis, but owing to the crushing of the skull, this feature cannot be clearly discerned in the skull of GMH 4362—the fossil studied by Houde and Haubold (1987)—or any other Palaeotis specimen. Critical features of the palate of Palaeotis are likewise not well exposed in the known specimens (Mayr 2015). However, an unambiguous character supporting palaeognathous affinities of Palaeotis is the presence of distinct furrows on the ventral surface of the mandibular symphysis (Houde and Haubold 1987; Mayr and Clarke 2003), and a previously unrecognized feature shared by Palaeotis and the flightless “ratites” is a distinct ligamental scar on the medial surface of the distal tibiotarsus (Fig. 2A4, H2). Houde and Haubold (1987: 31) furthermore found the bone histology of the tibiotarsus to be “consistent with what is known for ratite birds and unlike that of neognathous birds”.
If the Palaeotididae are representatives of the Palaeognathae and if these birds are closely related to the Geranoididae, palaeognathous affinities of the Eogruidae seem less far-fetched. Moreover, the Struthionidae resulted as the sister taxon of all other extant palaeognathous birds in current phylogenetic analyses based on molecular data (Harshman et al. 2008; Philips et al. 2010; Haddrath and Baker 2012; Smith et al. 2013; Baker et al. 2014; Mitchell et al. 2014; Yonezawa et al. 2017). The stem species of ostriches is therefore likely to have been similar to the stem species of neognathous birds, and there must have been a significant and previously unrecorded morphological diversity in the stem lineage of ostriches.
Eogruids are mainly known from leg bones and there exists no strong evidence for gruiform affinities of these birds, especially since some of the “gruiform” features, such as a tubercle next to the pons supratendineus (Clarke et al. 2005), have now also been reported for palaeotidids. On the other hand, however, the only derived feature supporting a sister group relationship between Eogruidae and Struthionidae is the reduction of the trochlea metatarsi II, and no shared derived characters could be identified that support a clade including Palaeotididae, Geranoididae, Eogruidae, and Struthionidae. If eogruids and struthionids form a clade, the Eogruidae would be paraphyletic, with the didactyl species traditionally assigned to the “Ergilornithidae” being more closely related to the Struthionidae than the tridactyl eogruid species (contra Clarke et al. 2005).
Differences between the Palaeotididae (Palaeotis, Galligeranoides) and taxa assigned to the Eogruidae include the broader trochlea metatarsi III of palaeotidids (compare Fig. 3C3, D3, F3) and the morphology of the hypotarsus, which exhibits a large canal in eogruids (Fig. 3F4). As noted above, the wider trochlea metatarsi III also distinguishes palaeotidids from geranoidids. The tarsometatarsus of none of the extant palaeognathous birds exhibits a hypotarsal canal, and well-developed hypotarsal sulci and canals were assumed to be an apomorphy of neognathous birds (Mayr and Clarke 2003). However, the tarsometatarsus of palaeotidids likewise shows a more complex hypotarsus than that of any of the extant Palaeognathae.
Apart from hindlimb morphology, very little is known about other aspects of the skeleton of geranoidids and eogruids. Kurochkin (1976) figured a proximal humerus of an early Oligocene eogruid, which clearly indicates flightlessness of this species, and Olson (1985: 157) mentioned an undescribed partial humerus of the Geranoididae with a reduced proximal end. A wing phalanx (phalanx proximalis digiti majoris) was tentatively assigned to Eogrus by (Wetmore 1934), who noted that by comparison with extant Gruidae, the bone “appears slighter and weaker as though it came from a wing used only moderately” (Wetmore 1934: 9). The holotype of P. weigelti also includes a fragmentary phalanx proximalis digiti majoris, which is of similar relative size to that assigned to Eogrus but is too fragmentary for meaningful comparisons and seems to exhibit a pathologic morphology with the formation of a distinct callus. A coracoid that was referred to Eogrus by Olson (1985) is also very different from the coracoid of Palaeotis, which is co-ossified with the scapula and forms a scapulocoracoid (compare Mayr 2016: fig. 3a with Mayr 2015: fig. 5a). However, assignment of this coracoid to Eogrus is likewise only tentative and the specimen was originally assigned to the Accipitriformes by Wetmore (1934).
Conclusions
In summary, it is here concluded that Palaeotis and Galligeranoides form a clade of palaeognathous birds (Palaeotididae) and that this clade is likely to be the sister taxon of the North American Geranoididae. However, whether this clade is in turn closely related to the Eogruidae and whether these birds are stem group representatives of the Struthionidae remains elusive. It is possible that long-legged flightless birds with greatly reduced wing bones evolved convergently in Europe/North America and Asia, but the surprisingly “gruiform”-like hindlimb morphology of palaeotidids (i.e., tibiotarsus with supratendinal bridge and condylus medialis with distinct notch in its distal rim, hypotarsus with plantar sulcus) potentially casts a new light on possible palaeognathous affinities of the equally “gruiform-like” Eogruidae. A critical revision of the abundant material of Oligocene and Neogene eogruids (e.g., Zelenkov and Kurochkin 2015) may shed further light on the affinities of these birds and future studies of the bone histology of geranoidids and eogruids may provide a definitive answer on the question whether these birds are representatives of the Palaeognathae.
Acknowledgements
For access to fossils, I thank Michael Stache and Oliver Wings (both GMH) and Carl Mehling (AMNH). The photos of the Palaeotis fossils were taken by Sven Tränkner (Senckenberg Research Institute, Frankfurt, Germany). Comments from Nikita Zelenkov (Borissiak Paleontological Institute of the Russian Academy of Sciences, Moscow, Russia) and two anonymous reviewers improved the manuscript.
References
Agnolin, F.L. 2017. Unexpected diversity of ratites (Aves, Palaeognathae) in the early Cenozoic of South America: palaeobiogeographical implications. Alcheringa 41: 101‒111. Crossref
Alvarenga, H.M.F. 1983. Uma ave ratitae do Paleoceno Brasileiro: bacia calcária de Itaboraí, Estado do Rio de Janeiro, Brasil. Boletim do museu nacional, Geologia 41: 1‒8.
Baker, A.J., Haddrath, O., McPherson, J.D., and Cloutier, A. 2014. Genomic support for a moa-tinamou clade and adaptive morphological convergence in flightless ratites. Molecular Biology and Evolution 31: 1686‒1696. Crossref
Bourdon, E., De Ricqlès, A., and Cubo, J. 2009. A new Transantarctic relationship: morphological evidence for a Rheidae–Dromaiidae–Casuariidae clade (Aves, Palaeognathae, Ratitae). Zoological Journal of the Linnean Society 156: 641‒663. Crossref
Bourdon, E., Mourer-Chauviré, C., and Laurent, Y. 2016. Early Eocene birds from La Borie, southern France. Acta Palaeontologica Polonica 61: 175–190.
Burchak-Abramovich, N.I. [Burčak-Abramovič, N.I.] 1951. Urmiornis (Urmiornis maraghanus Mecq.) an ostrich-like bird from the Hipparion fauna of Transcaucasia and south Ukraine [in Russian]. Izvestiâ Akademii Nauk Azerbajdžanskoj SSR 6: 83–94.
Clarke, J.A., Norell, M.A., and Dashzeveg, D. 2005. New avian remains from the Eocene of Mongolia and the phylogenetic position of the Eogruidae (Aves, Gruoidea). American Museum Novitates 3494: 1‒17. Crossref
Cracraft, J. 1969. Systematics and evolution of the Gruiformes (Class, Aves). 1. The Eocene family Geranoididae and the early history of the Gruiformes. American Museum Novitates 2388: 1‒41.
Cracraft, J. 1973. Systematics and evolution of the Gruiformes (Class Aves). 3. Phylogeny of the suborder Grues. Bulletin of the American Museum of Natural History 151: 1‒127.
Feduccia, A. 1980. The Age of Birds. 199 pp. Harvard University Press, Cambridge.
Haddrath, O. and Baker, A.J. 2012. Multiple nuclear genes and retroposons support vicariance and dispersal of the palaeognaths, and an Early Cretaceous origin of modern birds. Proceedings of the Royal Society of London, Series B 279: 4617–4625. Crossref
Harshman, J., Braun, E.L., Braun, M.J., Huddleston, C.J., Bowie, R.C.K., Chojnowski, J.L., Hackett, S.J., Han, K.-L., Kimball, R.T., Marks, B.D., Miglia, K.J., Moore, W.S., Reddy, S., Sheldon, F.H., Steadman, D.W., Steppan, S.J., Witt, C.C., and Yuri, T. 2008. Phylogenomic evidence for multiple losses of flight in ratite birds. Proceedings of the National Academy of Sciences U.S.A. 36: 13462–13467. Crossref
Houde, P. 1986. Ostrich ancestors found in the Northern Hemisphere suggest new hypothesis of ratite origin. Nature 324: 563–565. Crossref
Houde, P. 1988. Paleognathous birds from the early Tertiary of the Northern Hemisphere. Publications of the Nuttall Ornithological Club 22: 1–148.
Houde, P. and Haubold, H. 1987. Palaeotis weigelti restudied: a small Middle Eocene ostrich (Aves: Struthioniformes). Palaeovertebrata 17: 27–42.
Kurochkin, E.N. 1976. A survey of the Paleogene birds of Asia. Smithsonian Contributions to Paleobiology 27: 75‒86.
Kurochkin, E.N. [Kuročkin, E.N.] 1981. New representatives and evolution of two archaic gruiform families in Eurasia [in Russian]. Trudy Sovmestnoj Sovetsko-Mongolskoj Paleontologičeskoj Ekspedicii 15: 59‒85.
Lambrecht, K. 1928. Palaeotis Weigelti n. g. n. sp., eine fossile Trappe aus der mitteleozänen Braunkohle des Geiseltales. Jahrbuch des Halleschen Verbandes für die Erforschung mitteldeutscher Bodenschätze, Neue Folge 7: 1‒11.
Lambrecht, K. 1933. Handbuch der Palaeornithologie. 1022 pp. Gebrüder Borntraeger, Berlin.
Lambrecht, K. 1935. Drei neue Vogelformen aus dem Lutétian des Geiseltales. Nova Acta Leopoldina, Neue Folge 3: 361‒367.
Linnaeus, C. 1758. Systema naturae per regna tria naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Vol. 1: Regnum animale. Editio decima, reformata. 824 pp. Laurentii Salvii, Stockholm. Crossref
Martin, L.D. 1992. The status of the late Paleocene Birds Gastornis and Remiornis. In: K.E. Campbell (ed.), Papers in Avian Paleontology honoring Pierce Brodkorb. Natural History Museum of Los Angeles County, Science Series 36: 97‒108.
Mayr, G. 2002. Avian Remains from the Middle Eocene of the Geiseltal (Sachsen-Anhalt, Germany). In: Z. Zhou and Z. Zhang (eds.), Proceedings of the 5th Symposium of the Society of Avian Paleontology and Evolution, Beijing, 1–4 June 2000, 77–96. Science Press, Beijing.
Mayr, G. 2009. Paleogene Fossil Birds. 262 pp. Springer, Heidelberg. Crossref
Mayr, G. 2015. The middle Eocene European “ratite” Palaeotis (Aves, Palaeognathae) restudied once more. Paläontologische Zeitschrift 89: 503‒514. Crossref
Mayr, G. 2016. On the taxonomy and osteology of the Early Eocene North American Geranoididae (Aves, Gruoidea). Swiss Journal of Palaeontology 135: 315‒325. Crossref
Mayr, G. 2017. Avian Evolution: The Fossil Record of Birds and Its Paleobiological Significance. 293 pp. Wiley-Blackwell, Chichester. Crossref
Mayr, G. and Clarke, J. 2003. The deep divergences of neornithine birds: a phylogenetic analysis of morphological characters. Cladistics 19: 527‒553. Crossref
Mitchell, K.J., Llamas, B., Soubrier, J., Rawlence, N.J., Worthy, T.H., Wood, J., Lee, M.S.Y., and Cooper, A. 2014. Ancient DNA reveals elephant birds and kiwi are sister taxa and clarifies ratite bird evolution. Science 344: 898–900. Crossref
Olson, S.L. 1985. The fossil record of birds. In: D.S. Farner, J.R. King, and K.C. Parkes (eds.), Avian Biology, Volume 8, 79–238. Academic Press, New York. Crossref
Peters, D.S. 1988. Ein vollständiges Exemplar von Palaeotis weigelti (Aves, Palaeognathae). Courier Forschungsinstitut Senckenberg 107: 223–233.
Portis, A. 1885. Contribuzioni alla ornitolitologia italiana. Memorie della Reale Accademia delle Scienze di Torino 36: 361–384.
Phillips, M.J., Gibb, G.C., Crimp, E.A., and Penny, D. 2010. Tinamous and moa flock together: mitochondrial genome sequence analysis reveals independent losses of flight among ratites. Systematic Biology 59: 90–107. Crossref
Smith, J.V., Braun, E.L., and Kimball, R.T. 2013. Ratite nonmonophyly: independent evidence from 40 novel loci. Systematic Biology 62: 35–49. Crossref
Shufeldt, R.W. 1913. Further studies of fossil birds with descriptions of new and extinct species. Bulletin of the American Museum of Natural History 32: 285‒306.
Vanden Berge, J.C. and Storer, R.W. 1995. Intratendinous ossification in birds: a review. Journal of Morphology 226: 47–77. Crossref
Vandenberghe, N., Hilgen, F.J. and Speijer, R.P. with contributions by Ogg, J.G., Gradstein, F.M., Hammer, O., Hollis, C.J., and Hooker, J.J. 2012. The Paleogene period. In: F.M. Gradstein, J.G. Ogg, M. Schmitz, and G. Ogg (eds.), The Geologic Time Scale, 855–921. Elsevier, Amsterdam. Crossref
Yonezawa, T., Segawa, T., Mori, H., Campos, P. F., Hongoh, Y., Endo, H., Akiyoshi, A., Kohno, N., Nishida, S., Wu, J., Jin, H., Adachi, J., Kishino, H., Kurokawa, K., Nogi, Y., Tanabe, H., Mukoyama, H., Yoshida, K., Rasoamiaramanana, A., Yamagishi, S., Hayashi, Y., Yoshida, A., Koike, H., Akishinonomiya, F., Willerslev, E., and Jin, H. 2017. Phylogenomics and morphology of extinct paleognaths reveal the origin and evolution of the ratites. Current Biology 27: 68‒77. Crossref
Wetmore, A. 1933. Fossil bird remains from the Eocene of Wyoming. Condor 35: 115‒118. Crossref
Wetmore, A. 1934. Fossil birds from Mongolia and China. American Museum Novitates 711: 1‒16.
Woodburne, M.O., Goin, F.J., Raigemborn, M.S., Heizler, M., Gelfo, J.N., and Oliveira, E.V. 2014. Revised timing of the South American early Paleogene land mammal ages. Journal of South American Earth Sciences 54: 109‒119. Crossref
Zelenkov, N.V. and Kurochkin, E.N. 2015. Class Aves. In: E.I. Vorobyeva, E.N. Kurochkin, A.V. Lopatin, and M.A. Shishkin (eds.), Fossil Vertebrates of Russia and Adjacent Countries. Part 3, Fossil Reptiles and Birds, 86‒290. Geos, Moscow.
Acta Palaeontol. Pol. 64 (4): 669–678, 2019
https://doi.org/10.4202/app.00650.2019