
New protoceratopsid specimens improve the age correlation of the Upper Cretaceous Gobi Desert strata
ŁUKASZ CZEPIŃSKI
Czepiński, Ł. 2020. New protoceratopsid specimens improve the age correlation of the Upper Cretaceous Gobi Desert strata. Acta Palaeontologica Polonica 65 (3): 481–497.
New protoceratopsid (Dinosauria: Ceratopsia) specimens from two Late Cretaceous Mongolian localities with problematic stratigraphy are described. A specimen of Protoceratops andrewsi from the Zamyn Khond locality enables its correlation with other sites of the Djadokhta Formation. P. andrewsi is also abundant in the Üüden Sair locality, variously assigned to the Djadokhta or Baruungoyot formations. However, one new specimen from that site exhibits a fused nasal horn and a sharp buccal crest of the dentary. With these apomorphic features, it resembles Bagaceratops rozhdestvenskyi, known from the Baruungoyot and Bayan Mandahu formations. It may be an evidence for: sympatric evolution of B. rozhdestvenskyi and P. andrewsi; a dispersal of Bagaceratops to Üüden Sair; hybridization between the two parapatric taxa; or the anagenetic evolutionary transition from P. andrewsi to Bagaceratops. The anagenetic explanation appears to be most strongly supported by given data. This new record advocates for the age of the sediments from the Üüden Sair locality being intermediate between the Djadokhta and Baruungoyot formations. The observed changes in the frequency of the apomorphic features within protoceratopsid samples from various Late Cretaceous sites of the Gobi Desert potentially enable their correlations and chronological ordering.
Key words: Dinosauria, Ceratopsia, Protoceratopsidae, biostratigraphy, gradual evolution, anagenesis, Mesozoic, Mongolia.
Łukasz Czepiński [lczepinski@biol.uw.edu.pl, ORCID: https://orcid.org/0000-0002-8621-3888], Department of Palaeobiology and Evolution, Faculty of Biology, Biological and Chemical Research Centre, University of Warsaw, Żwirki i Wigury 101, 02-089 Warszawa, Poland.
Received 10 November 2019, accepted 18 February 2020, available online 8 May 2020.
Copyright © 2020 Ł. Czepiński. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
The Upper Cretaceous sandstone deposits of the Gobi Desert yield one of the most productive and diverse Late Cretaceous vertebrate assemblages in the world (Gradziński et al. 1977; Osmólska 1980; Gao and Norell 2000; Dashzeveg et al. 2005; Dingus et al. 2008; Currie 2016). Despite the long interest of paleontologists and geologists in studies of the Late Cretaceous ecosystems of the Gobi Basin, stratigraphy of this region is still a subject of disagreement (Jerzykiewicz and Russell 1991; Jerzykiewicz 2000; Shuvalov 2000; Kielan-Jaworowska et al. 2003; Dashzeveg et al. 2005; Watabe et al. 2010). The classic stratigraphic methods (comparison with the global marine biostratigraphy, radiometric dating, palynological studies) are hardly applicable to the Gobi Basin due to the lack of marine and volcanic sediments within the fossiliferous sections and limited palynological data.
The main dating method for the Gobi Basin is the comparison of the local faunas with better calibrated localities around the world, especially with the North American nonmarine units. Such a biostratigraphic approach was performed on several groups of vertebrates: dinosaurs, turtles and mammals (Gradziński et al. 1977; Shuvalov and Chkhikvadze 1979; Osmólska 1980; Barsbold 1983; Lillegraven and McKenna 1986; Makulbekov and Kurzanov 1986). However, the method does not allow for a high resolution, it is affected by the high level of endemism of the Gobi faunas, and it is biased by the subjective approach of taxonomists working on particular groups of organisms (Gradziński et al. 1977; Lucas and Estep 1998; Jerzykiewicz 2000; Averianov and Sues 2012). Moreover, even the relationships between most of the Cretaceous formations and localities of the Gobi Desert are unclear, because many exposures lack a good record of their geological succession (Jerzykiewicz 2000).
Among dinosaurs, protoceratopsid material is especially abundant in the sandstone beds of the Djadokhta and Baruungoyot formations (Fig. 1). The formations differ in faunal composition and in some environmental aspects, with sediments of the Djadokhta Formation having been deposited under more arid conditions (Jerzykiewicz and Russell 1991; Jerzykiewicz et al. 1993). Based on examination of the protoceratopsid material, I previously proposed that differences observed between samples of Protoceratopsidae from different localities and formations may reflect anagenetic evolution and suggest temporal differences in sediment deposition between distinct sites (Czepiński 2019). Here I supplement this claim by data on previously undescribed protoceratopsid specimens found at the Üüden Sair and Zamyn Khond localities. Additionally, I present a more detailed discussion on the distribution of protoceratopsid taxa, offering new perspectives for correlation and temporal ordering of vertebrate assemblages from each of the Late Cretaceous Gobi localities.
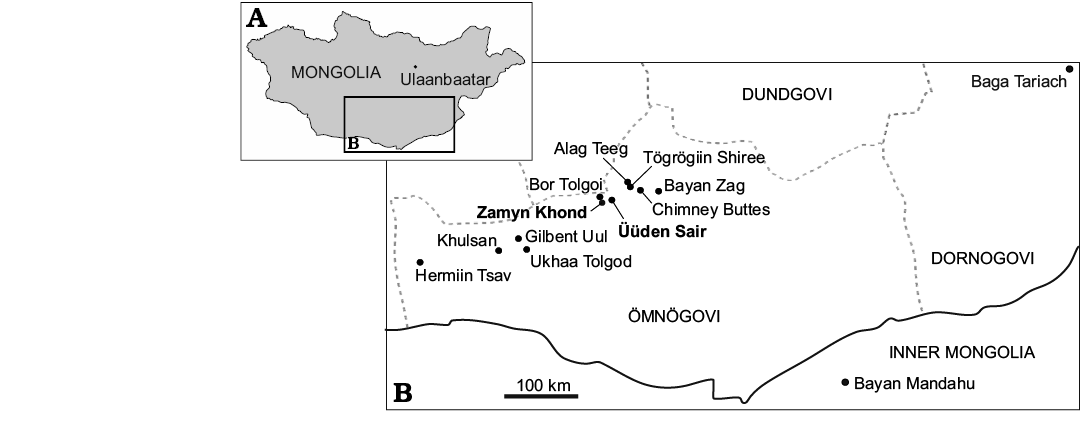
Fig. 1. Map of Mongolia (A) with the Late Cretaceous sites yielding remains of protoceratopsid dinosaurs (B). Localities that yielded the specimens described in this paper are in bold. Geographic data from: Jerzykiewicz and Russell 1991; Norell and Makovicky 1999; Dingus et al. 2008; Watabe et al. 2010; Czepiński 2019. Transcriptions of the locality names follow Benton 2000.
Institutional abbreviations.—AMNH, American Museum of Natural History, New York, USA; GMV, National Geological Museum of China, Beijing, China; HMNS, Hayashibara Museum of Natural Sciences, Okayama, Japan; IGM, Institute of Geology, Ulaanbaatar, Mongolia; IMM, Inner Mongolia Museum, Hohhot, China; IVPP, Institute of Vertebrate Paleontology and Paleoanthropology of the Chinese Academy of Sciences, Beijing, China, MPC, Mongolian Paleontological Center (Institute of Paleontology and Geology) of the Mongolian Academy of Sciences, Ulaanbaatar, Mongolia; PIN, Paleontological Institute of the Russian Academy of Sciences, Moscow, Russia; ZPAL, Institute of Paleobiology, Polish Academy of Sciences, Warsaw, Poland.
Systematic palaeontology
Dinosauria Owen, 1842
Ornithischia Seeley, 1887
Ceratopsia Marsh, 1890
Neoceratopsia Sereno, 1986
Coronosauria Sereno, 1986
Protoceratopsidae Granger and Gregory, 1923
Genus Protoceratops Granger and Gregory, 1923
Type species: Protoceratops andrewsi Granger and Gregory, 1923 Bayan Zag, Djadokhta Formation, Ömnögovi Province, Mongolia, Upper Cretaceous.
Protoceratops andrewsi Granger and Gregory, 1923
Figs. 2, 3, 4B.
Material.—MPC-D 100/505, a weathered skull of a large individual (Fig. 2) from the Zamyn Khond locality, Ömnögovi Province, Gobi Desert, Mongolia, Djadokhta Formation (?middle Campanian; Jerzykiewicz and Russell 1991), and MPC-D 100/551, a nearly complete articulated skeleton with skull and mandible of a medium-sized individual (Fig. 3) from the eolian sandstone beds of the Zhinst Tolgoi site, Üüden Sair area, Ömnögovi Province, Gobi Desert, Mongolia, Djadokhta or Baruungoyot Formation (?middle–late Campanian; Jerzykiewicz and Russell 1991; Watabe et al. 2010; Kielan-Jaworowska et al. 2003).
Description.—MPC-D 100/505 (Fig. 2) has a short and high rostral bone. At least one premaxillary tooth is preserved in the left premaxilla. Maxillary diastema seems to be relatively long (plausibly longer than 30% of the tooth row). However, its length cannot be established with certainty because not all teeth are preserved. As seen on the right side, premaxillo-maxillary suture is not pierced by any fenestra. The nasals are paired and form low, bowed horns. The frontonasal suture is U-shaped posteriorly. The frontals are relatively narrow and short. The frontoparietal depression is restricted to the small area of the posterior portion of frontals and is separated medially by the sagittal crest. The frontoparietal suture is W-shaped. The sagittal crest is slightly bifurcated at the contact with frontals. The skull in anterior view is folded in the postorbital region, with the postorbitals distinctly facing dorsally. The lateral buccal crest of the dentary is U-shaped in cross section. The frill expands posteriorly for nearly a half of the basal skull length. The symmetrical parietal fenestrae of the frill project laterally, nearly reaching the squamosals. The posterior margin of the frill is marked with a low, delicate ridge. Basal skull length (sensu Dodson 1976; measured from the anterior margin of the rostral to the posterior margin of the quadrate) of the specimen equals 223 mm.
Presence of paired nasals, the premaxillary dentition, the absence of the accessory antorbital fenestration and the U-shaped buccal crest of the dentary are diagnostic features of Protoceratops andrewsi (Makovicky 2002; Morschhauser 2012; Czepiński 2019). MPC-D 100/505 has a folded dorsal surface of the postorbitals, similar to large specimens of P. hellenikorhinus from Bayan Mandahu (e.g., IMM 95BM1/1). The development of the ridge on the posterior margin of the frill varies among P. andrewsi (Handa et al. 2012). The lateral expansion of the parietosquamosal frill is typical for individuals of advanced ontogenetic stages (Hone et al. 2016).
MPC-D 100/551 (Fig. 3) is a nearly complete skeleton measuring ~570 mm in total preserved length (measured from the tip of the rostral to the posterior extremity of the ischium). The sagittal skull length equals 216 mm, the height equals 85 mm. Basal skull length equals 120 mm, and the partial basal skull length (sensu Czepiński 2019; measured from the anterior margin of the maxilla to the posterior margin of the quadrate) is 91 mm.
The rostral is nearly complete; however, it is rather small and restricted to the anteroventral portion of the snout. Dorsally it reaches only slightly beyond the level of the ventral margin of external nares. Although the anterior keel is preserved, the lateral portion of the rostral barely shows any rugosities.
The premaxilla is rather short. Two premaxillary teeth are preserved on the left side. They are directed slightly laterally, protruding closely outside the predentary bone, when the jaws are closed, as observed by Varriale (2011). With this condition, it is similar to some other specimens of Protoceratops andrewsi (e.g., AMNH 6430 from Bayan Zag, Djadokhta Formation).
The maxilla is well preserved. The premaxillo-maxillary suture is seen at the right side of the specimen and it shows no evidence for the “true” accessory antorbital fenestration (sensu Czepiński 2019). However, the anterior margin of the maxilla that forms the suture is slightly concave at its midpoint, resembling the natural border of the bone, suggesting that some “loosening” (where the bones did not overlap along the entire premaxillo-maxillary suture, but had some loose contact at mid-height of the suture, however, without the formation of the “true” accessory antorbital fenestration with well-defined margins and a size similar to that of the external nares; Czepiński 2019: fig. 20) was present. However, due to the poor preservation of the area, it remains ambiguous. Nine maxillary teeth are preserved. The maxillary diastema is rather long, reaching 30.8% of the tooth row length.
The lacrimal forms up to two-thirds of the anterior orbit margin. The prefrontal is shallow, placed at the dorsal tip of the orbit. Preserved on the right prefrontal the descending process projects slightly below the midpoint of the orbit.
The nasals are not fused and form paired nasal horns, as can be noticed from the morphology of the preserved left nasal. It is covered by rugosities and slightly arched along the longitudinal axis of the bone.
The frontals are paired and covered by rugosities. They meet the postorbitals along slightly oblique sutures. A small and shallow frontoparietal depression is present. Frontals contribute extensively in the formation of the anterior margin of the supratemporal fenestra.
The jugals are complete. Although epijugals are not preserved, scars for the epijugal attachment occur on both sides of the specimen. The postorbitals are covered by rugosities. A small tubercle is visible beyond the posterodorsal corner of the orbit, especially on the right postorbital. The posterior process of the postorbital is short.
The squamosals form a wide contact with the parietal bone, as seen in dorsal view. In lateral view, the posteriormost portion of the squamosal is low and elongated. The posteromedial process of the squamosal is expanding anteriorly, and contributes to the lateral margin of the parietal fenestra. A low ridge occurs along the posterior margin of the parietal, as seen in other specimens of P. andrewsi from the same locality (Handa et al. 2012). The posterior margin of the parietal is slightly concave, resulting in a heart-shaped morphology in dorsal view. The sagittal crest is low and straight in lateral view.
The tip of the predentary does not project anteriorly, being placed behind the anterior tip of the premaxilla. The dentary is low, lacking a distinct buccal crest (Fig. 4B). A posteroventral process of the bone is present, as in other specimens of P. andrewsi and P. hellenikorhinus.
Presence of the premaxillary dentition, paired nasals, and a short maxillary diastema, and the lack of the distinct dentary buccal crest and accessory antorbital fenestration indicate that MPC-D 100/551 belongs to Protoceratops andrewsi.
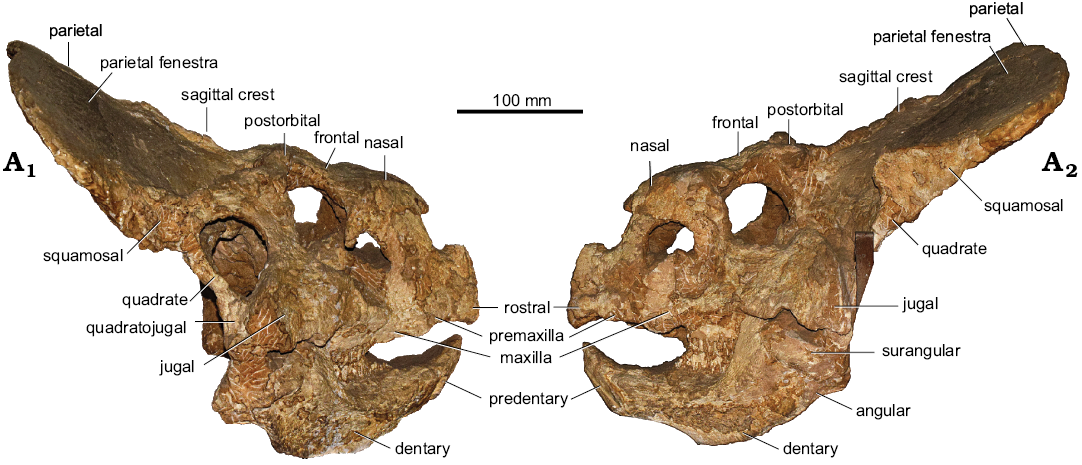
Fig. 2. Ceratopsian dinosaur Protoceratops andrewsi Granger and Gregory, 1923 (MPC-D 100/505), from the Late Cretaceous, Zamyn Khond, Ömnögovi, Mongolia. Skull in right (A1) and left (A2) lateral views.
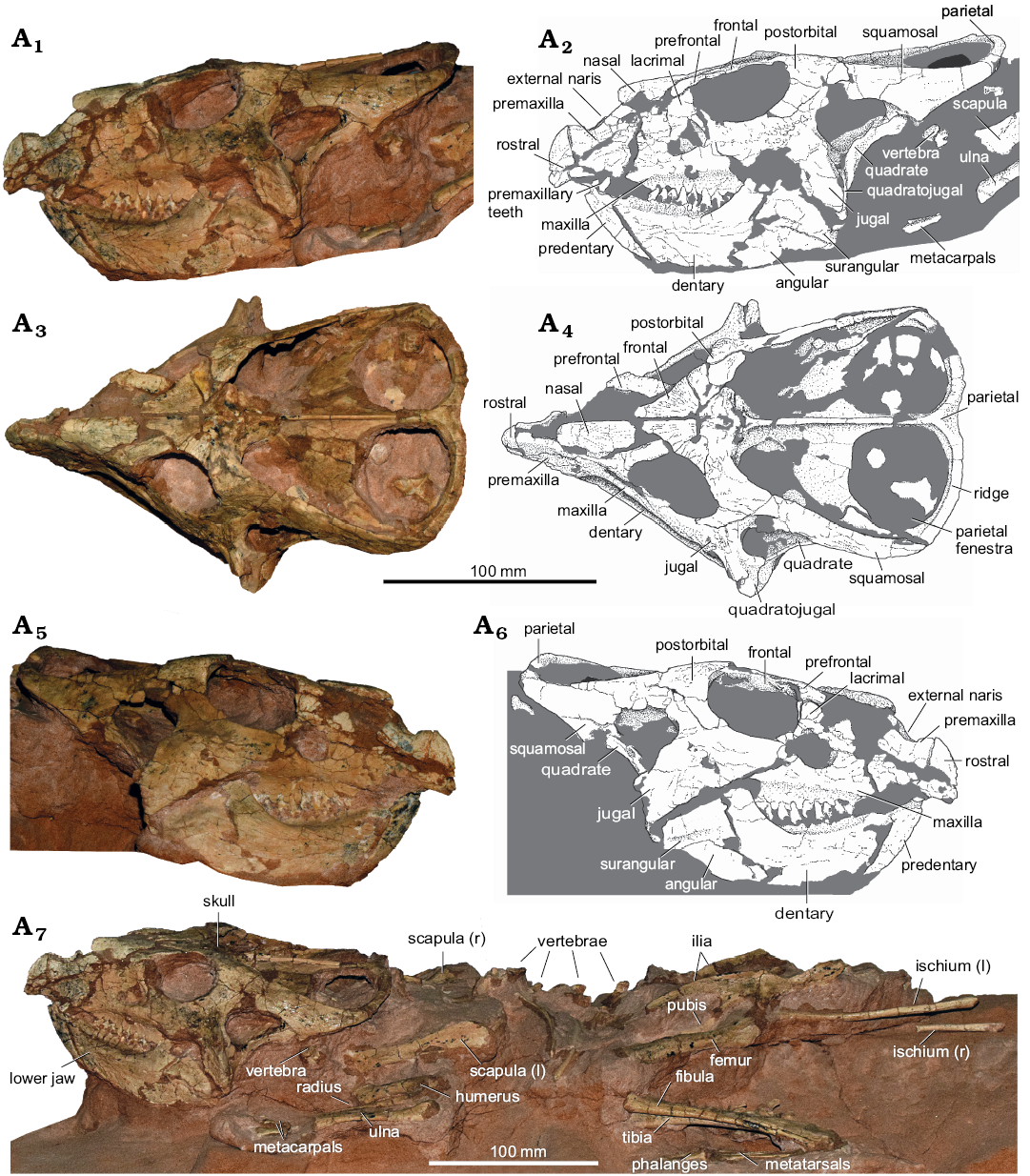
Fig. 3. Ceratopsian dinosaur Protoceratops andrewsi Granger and Gregory, 1923 (MPC-D 100/551), from the Late Cretaceous, Üüden Sair, Ömnögovi, Mongolia. Skull in left lateral (A1, A2), dorsal (A3, A4), and right lateral (A5, A6) views. Slab with the skeleton in left laterodorsal view (A7). Abbreviations: l, left; r, right.
Remarks.—MPC-D 100/505 was collected in 1977 by MPC (Ulziitseren Sanjaadash, personal communication 2016); MPC-D 100/551 was collected by the HMNS-MPC Joined Paleontological Expedition on July 20, 2000, with a field number 000720 US-Zhinst (Watabe and Tsogtbaatar 2004). The latter specimen was listed as Protoceratops in a report of the HMNS-MPC expedition by Watabe and Tsogtbaatar (2004). It is the same specimen described as Bagaceratops GI SPS 100/588 by Varriale (2011) and GI SPS 100/528 by Nabavizadeh and Weishampel (2016). Only recently has it been referred to as Protoceratops andrewsi (Czepiński 2019).
Stratigraphic and geographic range.—Upper Cretaceous, Gobi Desert, Mongolia.
Genus Bagaceratops Maryańska and Osmólska, 1975
Type species: Bagaceratops rozhdestvenskyi Maryańska and Osmólska, 1975, Hermiin Tsav, Baruungoyot Formation, Ömnögovi Province, Mongolia, Upper Cretaceous.
cf. Bagaceratops sp.
Figs. 4A, 5.
Material.—MPC-D 100/551B (field number 00019US FJM Baga), a skull and mandible with a partial articulated skeleton, containing both scapulae, humeri, radii, ulnae, right manus, and ribs (Fig. 5), from the Cretaceous (?middle–late Campanian), Djadokhta or Baruungoyot Formation (Jerzykiewicz and Russell 1991; Watabe et al. 2010; Kielan-Jaworowska et al. 2003), eolian sandstone beds of the Üüden Sair locality, Ömnögovi Province, Gobi Desert, Mongolia (Fig. 1).
Description.—Sagittal skull length is 230 mm. Skull width between the jugals is 146 mm. Basal skull length is 127 mm, and partial basal skull length is 97 mm.
The rostral was disarticulated from the specimen, revealing the morphology of the anteriormost portion of premaxilla. The rostral bone is rather complete, with a straight ventral margin. The ratio between height and length equals 1.55. In lateral view, the rostral originally reached nearly the level of the dorsal margin of the external nares.
The premaxilla is quite elongated, with the rostral covering only one third of the bone. The ventral margin of both premaxillae is weathered; however, the snout is distinctly narrower than the predentary width. Together with flat lateral sides of the bone this condition suggests a lack of premaxillary teeth. Indeed, in specimens with a premaxillary dentition the premaxilla is usually thicker and bulbous in this region. An edentulous premaxilla is a diagnostic feature of Bagaceratops rozhdestvenskyi and Protoceratops hellenikorhinus (Maryańska and Osmólska 1975; Lambert et al. 2001). However, a few specimens of Protoceratops andrewsi from Üüden Sair (MPC-D 100/539) and Bayan Zag (AMNH 6431) also lack teeth on the premaxilla (Handa et al. 2012; Czepiński 2019). The premaxilla contributes to the ventral and, together with the ventral projection of nasal, to the anterior and posterior margins of the elongated external nares.
The maxilla is poorly preserved on both sides of the specimen. The premaxillo-maxillary suture is weathered, and the bone is extremely thin in this area. In the anterior portion of the left maxilla, a part of the possible natural border of the bone is preserved. Due to the poor preservation of the premaxilla’s posterior portion, it cannot be concluded if the “loosening” of the premaxillo-maxillary suture was present, or if the premaxilla simply overlapped the maxilla. However, presence of the “true” accessory antorbital fenestration, with distinct margins and a size similar to that of the external nares (as in B. rozhdestvenskyi), is not likely, because the eroded area of the bone is limited, being much smaller than the nares. A few maxillary teeth are poorly preserved, and the length of the tooth row and of the maxillary diastema cannot be measured.
The lacrimal contributes to most of the posterodorsal margin of the antorbital fossa. Its contribution to the anterior margin of the orbit is restricted to one third of its height, due to the development of the ventral process of the prefrontal and of the anterodorsal process of the jugal.
The prefrontal is deep, and its descending process reaches the level below the mid-height of the orbit. In dorsal view, the prefrontal is wider rostrally. In anterior view, the prefrontals lean toward the longitudinal axis of the skull.
The “horn-like projection” of the nasals was described in the field report on the specimen (Watabe and Tsogtbaatar 2004). That part of the bone was no longer attached to the specimen during my visit in MPC, Ulaanbaatar (September 2016). However, its morphology can be reconstructed thanks to the close examination of the preserved part of the bones, and the study of the archival pictures from the field report (Watabe et al. 2004: pl. 3: 5) and those taken in MPC in the past. Only the midline and right portion of the fused nasals were preserved, forming a low, narrow, elongated horncore. Its dorsal tip nearly reached the level of the dorsal margin orbit. The fused nasals are an autapomorphy of Bagaceratops rozhdestvenskyi (Maryańska and Osmólska 1975; Makovicky 2002; Morschhauser 2012; Czepiński 2019), and are seen also in Ceratopsidae; however, in the latter the nasal fusion occurs during the ontogeny (McDonald 2011).
The paired frontals are roughly triangular in dorsal view. Anteriorly, proximal to the nasal horncore, the frontals are elevated medially, and the nasofrontal suture is V-shaped. The frontals meet with the postorbitals at a nearly straight suture. The interfrontal suture is slightly convex along most of its longitudinal length, with the posterior portion of the suture being concave. A distinct frontoparietal depression occurs on both sides of the posterior portion of the interfrontal suture. The frontals form a small contribution on the anterior margin of the supratemporal fenestra.
The left jugal is better preserved than the right one. However, the bone is deformed in the region of the contact with the maxilla. The bone contributes to the whole ventral margin of the orbit, and the short ascending anterior process forms the anteroventral corner of the margin. Its quadratojugal portion is deformed. Posteriorly, the jugal contributes to the formation of the anterior margin of the infratemporal fenestra, excluding the postorbital. The epijugal is not preserved and no trace of its attachment is preserved on the jugal.
The postorbitals are elongated, reaching half of the parietosquamosal frill length. Small rugosities occur on the bone at the posterodorsal margin of the orbit. A small tubercle occurs on the suture between the postorbital and frontal. Such a tubercle was also observed in some specimens of B. rozhdestvenskyi (e.g., ZPAL MgD-I/125, 129, IVPP V12513; Czepiński 2019).
The squamosals are widely separated from the margin of the parietal fenestra. In lateral view, the squamosals lean posteroventrally, with a high posteriormost margin. The posterior tip of the squamosals is covered with longitudinal rugosities. The parietal fenestrae are small and asymmetrical. The left fenestra is nearly circular, whereas the right is elongated mediolaterally. The parietal fenestration is small, even when compared with smaller specimens of P. andrewsi (i.e., the parietal fenestrae area of MPC-D 100/551 is more than twice as large as in the cf. Bagaceratops sp. specimen, although the basal skull length of the latter is 7% greater). It is similar to the known material of B. rozhdestvenskyi, where the fenestration is relatively smaller than in Protoceratops spp. (Czepiński 2019). The ridge at the posterior margin of the parietal frill is subtle, and the sagittal crest is somewhat bifurcated at its contact with the frontal, resembling the holotype specimen of B. rozhdestvenskyi (ZPAL MgD-I/126).
The predentary is elongated, but its lateral surface is poorly preserved. It is longer than in P. andrewsi (MPC-D 100/551), extending anterior to the premaxilla. The dentary bears a buccal crest, nearly V-shaped in cross-section. The crest is distinct; however, it does not expand laterally to the same degree as in some specimens of B. rozhdestvenskyi (e.g., ZPAL MgD-I/144). There is a posteroventral (angular) projection of the dentary, similar to the condition seen in Protoceratops spp. (Czepiński 2019).
Given the presence of the fused nasals and V-shaped buccal crest of dentary, the specimen is similar to Bagaceratops. However, it lacks the well-developed premaxillo-maxillary fenestration (accessory antorbital fenestra) and the angular is not projecting anteroventrally. The mosaic of plesiomorphic and apomorphic features makes the morphology of the specimen intermediate between P. andrewsi and B. rozhdestvenskyi (Fig. 4).
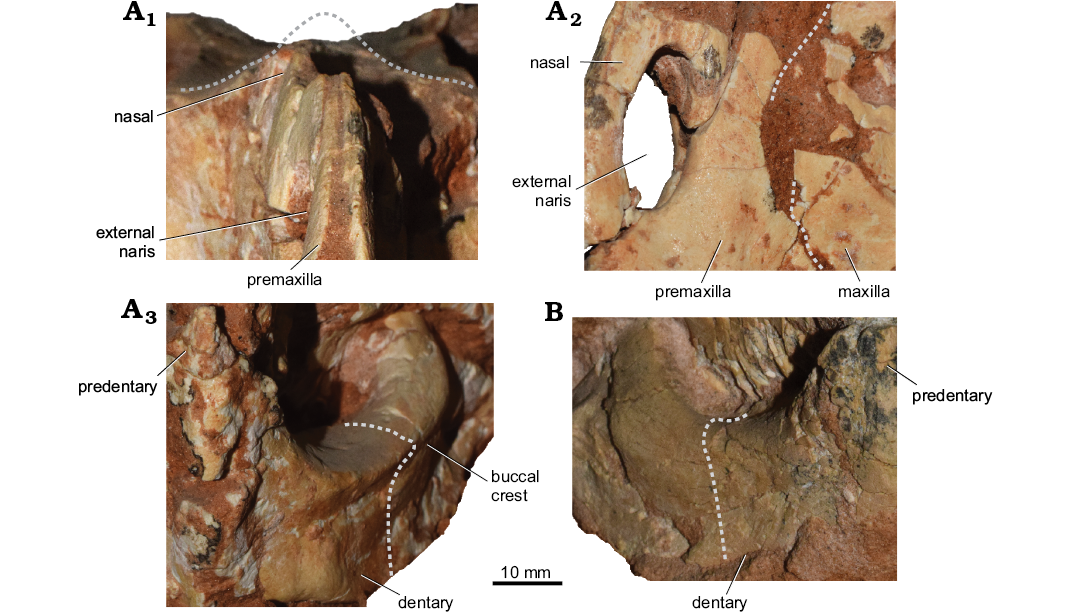
Fig. 4. Comparison of two ceratopsian dinosaurs cf. Bagaceratops sp. (A) and Protoceratops andrewsi (B) from the Late Cretaceous, Üüden Sair, Ömnögovi, Mongolia. A. Plesiomorphic and apomorphic features of MPC-D 100/551B (field number 000719 US FJMBaga). Skull in anterior view (A1), showing the folded nature of the preserved nasals, suggesting the presence of a single midline nasal horn (the dashed line shows its reconstruction following archival photographs). Premaxillo-maxillary suture (the dashed line) in left anterolateral view (A2), showing no evidence for the “true” accessory antorbital fenestra. Right mandible in anterior view (A3), showing the sharp buccal crest of the dentary. B. Plesiomorphic condition of the left mandible of MPC-D 100/551 in anterior view (the dashed lines show the lateral outline at the mid-length of the mandible).
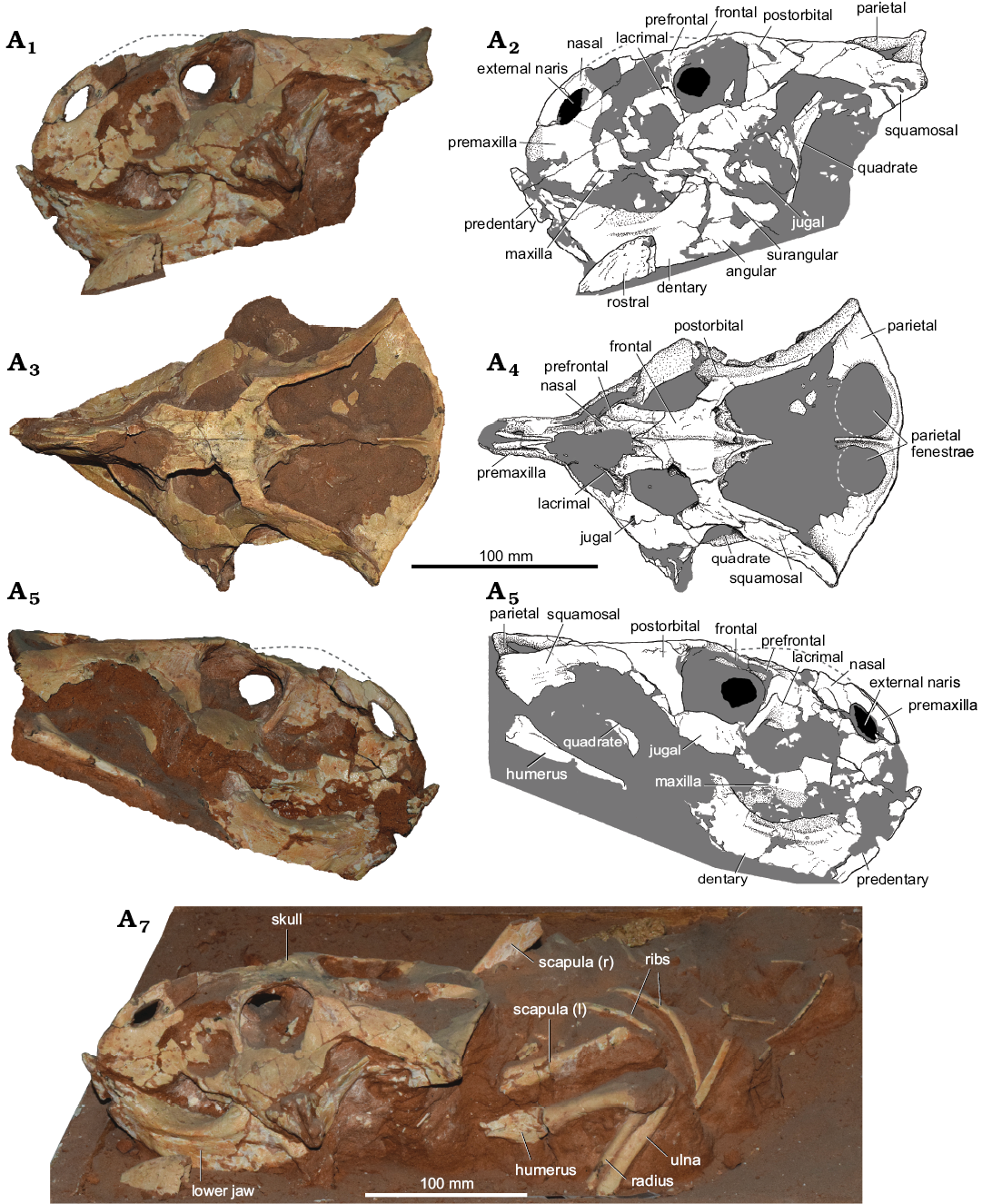
Fig. 5. Ceratopsian dinosaur cf. Bagaceratops sp. (MPC-D 100/551B, field number 000719 US FJMBaga), from the Late Cretaceous, Üüden Sair, Ömnögovi, Mongolia. Skull in left lateral (A1, A2), dorsal (A3, A4), and right lateral (A5, A6) views. Slab with the skeleton in left laterodorsal view (A7). Abbreviations: l, left; r, right.
Remarks.—The specimen was collected on July 19–20, 2000 at Üüden Sair by the HMNS-MPC expedition to the Gobi Desert. It was photographed in the report of the HMNS expedition with the field number 000719 US FJMBaga (Watabe and Tsogtbaatar 2004). It was, probably erroneously, described with the field number 00720 US by Tsogtbaatar (2004).
Discussion
Intermediate nature of the cf. Bagaceratops sp. specimen.—The presence of Bagaceratops in the Üüden Sair locality can be explained in four different ways.
(i) The sympatric evolution of Bagaceratops and P. andrewsi in one locality. Although possible, the sympatric speciation would require evidence, i.e., a niche partitioning, that would lead to the reproductive isolation within the population (Smith 1966). So far there is no evidence for a different ecology between the two taxa (i.e., no differences in the morphology of teeth; Czepiński 2019). Differences in the feeding apparatus are quite subtle, and include an elongated maxillary diastema and the lack of a premaxillary dentition in B. rozhdestvenskyi. However, premaxillary teeth are also lacking in some specimens referred to P. andrewsi (AMNH 6431, MPC-D 100/539), and its contribution in known sample of this species is higher in the Üüden Sair locality (16%) than in any other (Fig. 6), suggesting that gradual changes were occurring in a lineage of P. andrewsi. Some subtle differences in the morphology of a given region of the skull (i.e., aspect of horns and frills) were suggested as an evidence for species recognition among sympatric taxa (Padian and Horner 2011); however, this was criticized recently, among others, on the basis of the Gobi protoceratopsids (Hone and Naish 2013; Knapp et al. 2018).
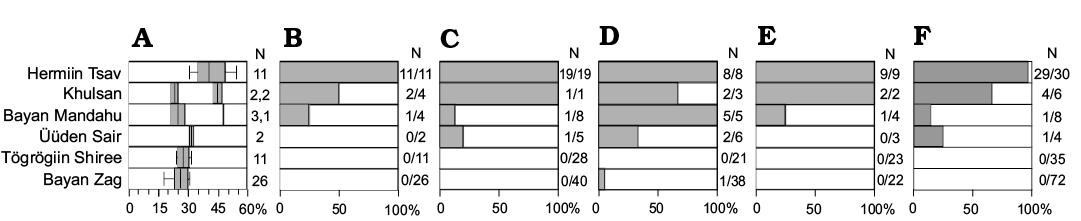
Fig. 6. Distribution of the apomorphic features within the protoceratopsid material from different Late Cretaceous localities of the Gobi Desert. A. Ratio of maxillary diastema to tooth row length (after Czepiński 2019). B. Long diastema (>35% of tooth row). C. Fused nasal horn. D. No premaxillary dentition. E. Accessory antorbital fenestra. F. Sharp buccal crest of dentary. Note: A, number of specimens for each sample, with sympatric taxa separated by a comma (Bagaceratops rozhdestvenskyi, Breviceratops kozlowskii in Khulsan, and B. rozhdestvenskyi, Protoceratops hellenikorhinus in Bayan Mandahu); B–F, number of specimens showing the apomorphic features (grey-shaded area) and the total number of specimens in which the condition state can be recognized. Detailed information in SOM.
(ii) The dispersal of Bagaceratops. It would be possible that B. rozhdestvenskyi emerged in a different region and subsequently migrated to Üüden Sair. However, the specimen of cf. Bagaceratops sp. shows only a few of the autapomorphic features of B. rozhdestvenskyi (there is no “true” accessory fenestra and no anterior projection of the angular bone). No Bagaceratops specimens are known from the “typical” Djadokhta localities, and the material from the Inner Mongolia Bayan Mandahu site (IVPP V12513) is already indistinguishable from B. rozhdestvenskyi from the Baruungoyot Formation (Czepiński 2019).
(iii) Hybridization event. Üüden Sair is placed between the southwestern Baruungoyot localities with the record of B. rozhdestvenskyi (Hermiin Tsav, Khulsan) and the northeastern typical Djadokhta sites with P. andrewsi (Bayan Zag, Tögrögiin Shiree; Fig. 1). It is possible that the two species, although morphologically different, did not establish full reproductive isolation from each other, and Üüden Sair was a hybrid zone. Hence, the intermediate phenotype of the cf. Bagaceratops sp. specimen may be an evidence for hybridization between the parapatric populations of P. andrewsi and B. rozhdestvenskyi.
However, it does not explain why in the adjacent Zamyn Khond locality, placed even closer to the Baruungoyot sites than Üüden Sair, only the typical P. andrewsi is recorded. In addition, one would expect the presence of both parentals and intermediate phenotypes in the hybrid zone, as it is observed in extant animals (e.g., Lawson and Lieb 1990; Szymura 1993). This is not the case here: except for the intermediate specimen of cf. Bagaceratops sp. described here, most of the protoceratopsid material can be referred to P. andrewsi (MPC-D 100/537, 538, 551; Handa et al. 2012) and no definitive B. rozhdestvenskyi is known in Üüden Sair. Moreover, in Bayan Mandahu B. rozhdestvenskyi co-occurs with P. hellenikorhinus, the latter being closely related to P. andrewsi, without the record of the intermediate individuals suggesting hybridization. As stated earlier, no single locality records the co-occurrence of both P. andrewsi and B. rozhdestvenskyi.
(iv) Anagenetic evolutionary transition. The specimen from Üüden Sair may be an intermediate stage of an anagenetic evolution within the lineage leading from P. andrewsi to B. rozhdestvenskyi. Although anagenesis is rarely considered in the evolution of dinosaurs, and whereas most of the research suggests that it is of little importance in evolutionary processes (Hunt and Rabosky 2014), that may actually be an artifact due to the nature of the fossil record (Dzik 2005). Some recent studies have provided evidence of anagenesis in dinosaur phylogeny (Horner et al. 1992; Scanella et al. 2014; Carr et al. 2017; Wilson et al. 2020). The best support for such an interpretation requires dense and precise stratigraphic sampling, temporally successive specimens of close phylogenetic relationship, and a large sample size that includes growth series (Carr et al. 2017). Although the number of protoceratopsid specimens currently known is one of the highest among dinosaurs, postulating an anagenetic evolution without precise stratigraphic data remains challenging.
Nevertheless, the presence of both plesiomorphic and apomorphic features in the cf. Bagaceratops sp. specimen hampers the precise assignation of that material. Given the faunistic and sedimentological evidence (suggesting that Üüden Sair was intermediate in environment and/or time between Djadokhta and Baruungoyot formations; Table 1), and the accumulation of diagnostic features for B. rozhdestvenskyi in protoceratopsid material from distinct localities (suggesting an increase in the frequency of the apomorphic Bagaceratops-phenotype in the protoceratopsid populations; Fig. 6), it would be reasonable to consider the specimen as intermediate between P. andrewsi and B. rozhdestvenskyi. Hence, the sediments of Üüden Sair would record the population where Bagaceratops-type traits were evolving (Fig. 7). It is plausibly the most reasonable explanation for the mosaic morphology of the specimen MPC-D 100/551B (field number 000719 US FJMBaga).
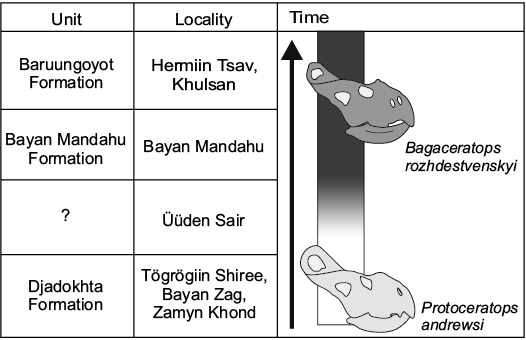
Fig. 7. Hypothesized chronology of the Late Cretaceous Gobi Desert strata with a schematic representation of the accumulation of apomorphic Bagaceratops-phenotype features in the Protoceratops andrewsi–Bagaceratops rozhdestvenskyi lineage recorded in each of localities.
The samples from different Mongolian localities can be regarded as quasi-populations, representing a time-averaged record of the protoceratopsid populations that lived in the same place. Differences between samples from distinct sites cannot be explained by isolation only, due to their geographic proximity and the lack of known barriers that would hamper the interbreeding of the parapatric populations. Distances between even relatively remote locations (Bayan Mandahu is placed less than 500 kilometers from the main Gobi fossil sites; Fig. 1) are not beyond the mobility abilities of medium-sized animals as protoceratopsids (Godefroit et al. 2008; Longrich et al. 2010).
The suggestion that the different Gobi localities were deposited at different times was raised in some earlier studies (Kielan-Jaworowska et al. 2003; Dashzeveg et al. 2005; Makovicky 2008; Longrich et al. 2010). Despite the lack of detailed stratigraphic data for the collected specimens, future work, with the record of the precise stratigraphic position data, is crucial. It would lead to the confirmation or rejection of the proposed anagenetic hypothesis and an evolutionary connection between the two species. More data on the less known taxa and closely related taxa, Protoceratops hellenikorhinus and Breviceratops kozlowskii, would increase our knowledge of the evolutionary history and relationships of protoceratopsid dinosaurs.
Correlation between the Bayan Zag and Tögrögiin Shiree localities.—The Bayan Zag (= Flaming Cliffs, Shabarkh Usu) locality belongs to the Djadokhta Formation, plausibly Campanian in age (Gradziński et al. 1977; Jerzykiewicz et al. 1993; Dashzeveg et al. 2005). It was the first locality to yield specimens of Protoceratops andrewsi (Granger and Gregory 1923; Brown and Schlaikjer 1940). So far, at least 130 protoceratopsid specimens were collected from there. From the Tögrögiin Shiree (= Tugrugyin Shireh = Tugrik = Toogreeg) locality, also assigned to the Djadokhta Formation (Jerzykiewicz and Russell 1991), more than 60 protoceratopsid specimens were collected.
There is some degree of intraspecific variation in cranial material of P. andrewsi from both sites. Several specimens have a maxillary diastema that is longer than 30% of the tooth row (AMNH 6433, 6637, ZPAL MgD-II/2, BMNH R10060, UALVP 49397 from Bayan Zag and MPC-D 100/522, 534a, and PIN 3143/1 from Tögrögiin Shiree; Fig. 6), the loosening of the premaxillo-maxillary suture (e.g., AMNH 6251), the anteroventral projection of the angular (AMNH 6416, 6428), the lack of the premaxillary dentition (e.g., AMNH 6431), or lack of the well-developed frontoparietal depression in adult individuals (e.g., the large skull AMNH 6439, although variable presence of the depression was considered ontogeny-dependent by Farke 2010). When combined, these features are diagnostic for Bagaceratops rozhdestvenskyi (Czepiński 2019). However, the firsthand examination of 95 cranial specimens collected from Bayan Zag and 35 from Tögrögiin Shiree suggests that P. andrewsi is the only ceratopsian species in these two localities (SOM, Supplementary Online Material available at http://app.pan.pl/SOM/app65-Czepinski_SOM.pdf).
A recent study on vertebral morphology hypothesized the presence of four morphotypes within the P. andrewsi sample, suggesting that more than one protoceratopsid taxon was present in the Djadokhta Formation (Tereschenko 2018). However, observed differences are subtle and better explained by the relatively wide intraspecific variation in Protoceratopsidae (Makovicky and Norell 2006; Czepiński 2019). The recently described skeleton of B. rozhdestvenskyi from Hermiin Tsav suggests no distinct differences in the postcranial anatomy between B. rozhdestvenskyi and P. andrewsi (Kim et al. 2019). Given this, postcranial specimens PIN 614/29, 34 and 53 from Bayan Zag and PIN 3143/11 from Tögrögiin Shiree, previously referred to Bagaceratops sp. (Tereschenko 2007) most likely fall into the postcranial variation within P. andrewsi. No cranial material with the apomorphic Bagaceratops features (e.g., fused nasals) was found in these two localities; therefore, there is no evidence for the presence of B. rozhdestvenskyi in the Djadokhta Formation. Because all diagnostic features focus on the cranium, the isolated postcranial material should better be regarded as Protoceratopsidae indet. (Czepiński 2019).
It was reported recently that specimens of P. andrewsi from Bayan Zag and Tögrögiin Shiree differ in the length of the maxillary diastema and the number of maxillary teeth (Fig. 6; Czepiński 2019: figs. 12, 14). It is possible that the observed differences are caused by differences in the geological age of the sediments where the specimens were found or regional radiation. This is congruent with some earlier attempts to elucidate the temporal sequence of the Late Cretaceous Mongolian paleobiotas (Makovicky 2008). Also, geological observations have suggested the division of the Djadokhta Formation into the Bayan Zag Member and the overlying Tögrögiin Shiree Member (Dashzeveg et al. 2005). However, these observations were questioned more recently (Hasegawa et al. 2009). Given all the evidence currently available, it is most likely that the Djadokhta sediments of Bayan Zag and Tögrögiin Shiree are nearly of equal age (Jerzykiewicz et al. 1993; Hasegawa et al. 2009), although some of the observations mentioned above indicated that the Tögrögiin Shiree deposits are slightly younger (Makovicky 2008).
Age of the Zamyn Khond locality.—The Zamyn Khond (= Dzamyn Khondt = Dzamin Khond) locality is placed 9 km southwest of Üüden Sair (Fig. 1). It has been assigned to the Upper Cretaceous Djadokhta Formation (Barsbold 1988; Jerzykiewicz and Russell 1991; Tsuihiji et al. 2014). Its lithology (eolian sandstone) resembles that of the Üüden Sair and Bor Tolgoi localities (Watabe et al. 2010).
Zamyn Khond has yielded material of the oviraptorosaur Citipati sp. (Barsbold 1983; Clark et al. 2002; Longrich et al. 2010), the troodontid Gobivenator mongoliensis (Tsuihiji et al. 2014), the nanhsiungchelyid turtle Zangerlia dzamynchondi (Danilov et al. 2013), remains of ankylosaurids (Watabe and Suzuki 2000; Carpenter et al. 2011; Arbour et al. 2014; Burns et al. 2015), protoceratopsids (Watabe and Suzuki 2000), Gobipteryx-type eggs, and remains of mammals and lizards (Watabe and Suzuki 2000; Tsogtbaatar et al. 2014). So far, there were no overlapping species between Zamyn Khond and any other locality of the Djadokhta or Bayan Mandahu Formation. Thus, it was thought that the locality cannot be referred to the “typical” Djadokhta Formation (Pei et al. 2017).
The oviraptorosaur Citipati osmolskae is known from the sediments of the Ukhaa Tolgod locality, possibly younger than other Djadokhta Formation localities (Makovicky 2008). However, material of Citipati sp. from Zamyn Khond most plausibly represents a distinct species (Clark et al. 2002; Longrich et al. 2010). Different species of the nanhsiungchelyid genus Zangerlia are present in the Ukhaa Tolgod, Nemegt Formation and Bayan Mandahu (the later currently known as Jiangxichelys neimongolensis; Tong et al. 2016). The purported presence of the ankylosaurs Saichania chulsanensis (Carpenter et al. 2011) and Pinacosaurus sp. (Watabe and Suzuki 2000) in Zamyn Khond is not confirmed (Arbour and Currie 2013).
Among protoceratopsids, material of Protoceratops was collected in Zamyn Khond in 1977 (MPC-D 100/505) and in 1998 (Watabe et al. 2000). In addition to the specimen MPC-D 100/505 described above (Fig. 2), a second specimen housed in MPC could not be examined in the curse of this work. This specimen (MPC-D 100/518) is an articulated skeleton of a medium-sized individual, with a skull lacking the roof and the parietosquamosal frill. The presence of the premaxillary dentition can be noticed on archival pictures of the specimen. There is no accessory antorbital fenestration, and the maxillary diastema is relatively short. The nasals are paired and do not form a prominent horn, possibly because of the young ontogenetic age of the individual. The labial dentary crest is U-shaped in cross-section.
The presence of the premaxillary dentition and of paired nasals suggests that both specimens (MPC-D 100/505 and 518) belong to P. andrewsi. Thus, this species is the only one that is recognized in both the Zamyn Khond locality and other localities of the “typical” Djadokhta Formation (Table 2). The rather long maxillary diastema of MPC-D 100/505 may provide an evidence of a relatively derived condition of the individual; however, this feature displays quite a wide intraspecific variation in P. andrewsi (Fig. 6). With the shared presence of P. andrewsi, similarity in age of Zamyn Khond with other Djadokhta localities (i.e., Bayan Zag and Tögrögiin Shiree) is very plausible.
Table 2. The Djadokhta Formation dinosaur taxa from the Bayan Zag, Tögrögiin Shiree, and Zamyn Khond localities. Data from: Chinzorig et al. 2017, Czepiński 2019, Elżanowski and Wellnhofer 1992, Longrich et al. 2010.
| |
Bayan Zag |
Tögrögiin Shiree |
Zamyn Khond |
|
Protoceratopsidae |
Protoceratops andrewsi |
Protoceratops andrewsi |
Protoceratops andrewsi |
|
Leptoceratopsidae |
Bainoceratops efremovi |
|
|
|
Ankylosauria |
Pinacosaurus grangeri |
Pinacosaurus sp. |
|
|
Alvarezsauria |
Alvarezsauridae indet. |
|
|
|
|
?Parvicursor sp. |
|
|
|
Oviraptorosauria |
Oviraptor philoceratops |
|
Zamyn Khond taxon |
|
Dromaeosauridae |
Velociraptor mongoliensis |
Velociraptor mongoliensis |
|
|
|
Mahakala omnogovae |
|
|
|
Troodontidae |
Archaeornithoides deinosauriscus |
|
Gobivenator mongoliensis |
|
Saurornithoides mongoliensis |
|
|
|
|
Ornithomimosauria |
|
Aepyornithomimus tugrikinensis |
|
Age of the Üüden Sair locality.—The Üüden Sair (= Udyn Sayr = Udan Sayr) locality was discovered by the Soviet-Mongolian Paleontological Expeditions in the 1980s (Ivakhnenko and Kurzanov 1988). The area is located about 65 kilometers west of Bayan Zag (Fig. 1). The site was also explored by AMNH-MPC and HMNS-MPC expeditions; the latter focused on the southern area called Zhinst Tolgoi (Watabe et al. 2010; Rougier et al. 2016). In the literature, it was variously referred to as the Djadokhta Formation (Ivakhnenko and Kurzanov 1988; Watabe et al. 2010; Handa et al. 2012; Funston et al. 2019) or the Baruungoyot Formation (Jerzykiewicz and Russell 1991; Kielan-Jaworowska et al. 2003; Rougier et al. 2016).
The presence of Protoceratops andrewsi in the Üüden Sair, along with Pinacosaurus sp., makes it similar to the Djadokhta localities, Ukhaa Tolgod and Bayan Mandahu (Table 1). The anguimorph Gobiderma pulchra found there is also known from Tögrögiin Shiree (Djadokhta Formation), Ukhaa Tolgod, Hermiin Tsav and Khulsan (Baruungoyot Formation; Gao and Norell 2000).
Table 1. Distribution of the overlapping vertebrate taxa within the Upper Cretaceous formations of the Gobi Desert yielding protoceratopsid remains. Data from: Alifanov 2012, Currie et al. 2011, Czepiński 2019, Dong et al. 2018, Gao and Hou 1996, Gao and Norell 2000, Kielan-Jaworowska et al. 2003, Norell and Makovicky 1999, Turner et al. 2012, Watabe et al. 2010. Abbreviations: AT, Alag Teeg; BM, Bayan Mandahu; BT, Bor Tolgoi; BZ, Bayan Zag; CB, Chimney Buttes; HT, Hermiin Tsav; Kh, Khulsan; TS, Tögrögiin Shiree; US, Üüden Sair (HMNS-MPC collection); US-PIN, Üüden Sair (PIN collection); UT, Ukhaa Tolgod; YK, Yagaan Khovil; ZK, Zamyn Khond; Fm., Formation.
| |
Taxa |
AT Fm. |
Djadokhta Fm. |
? |
BM Fm. |
Baruungoyot Fm. |
|||||||||
|
AT |
BZ |
TS |
ZK |
CB |
US |
US-PIN |
YK |
BT |
UT |
BM |
HT |
Kh |
|||
|
Dinosauria |
Ceratopsia |
Bagaceratops rozhdestvenskyi |
|
|
|
|
|
? |
|
|
|
|
× |
× |
× |
|
Breviceratops kozlowskii |
|
|
|
|
|
|
|
|
|
|
? |
? |
× |
||
|
Protoceratops andrewsi |
? |
× |
× |
× |
× |
× |
|
|
|
|
|
|
|
||
|
Protoceratops hellenikorhinus |
|
|
|
|
|
|
|
|
|
|
× |
|
|
||
|
Ukhaa Tolgod protoceratopsid |
|
|
|
|
|
|
|
|
|
× |
|
|
|
||
|
Bainoceratops efremovi |
|
× |
|
|
|
|
|
|
|
|
|
|
|
||
|
Udanoceratops tschizhovi |
|
|
|
|
|
|
× |
|
? |
|
? |
|
|
||
|
Ankylosauria |
Pinacosaurus grangeri |
? |
× |
? |
|
? |
? |
|
|
× |
× |
× |
|
|
|
|
Pinacosaurus mephistocephalus |
|
|
|
|
|
|
|
|
|
|
× |
|
|
||
|
Saichania chulsanensis |
|
|
|
|
|
|
|
|
|
|
|
× |
× |
||
|
Theropoda |
Avimimus portentosus |
|
|
|
|
|
|
× |
× |
|
|
|
|
|
|
|
Conchoraptor gracilis |
|
|
|
|
|
|
|
|
|
|
|
× |
× |
||
|
Shuvuuia deserti |
|
|
? |
|
|
|
|
|
|
× |
|
|
|
||
|
Tsaagan mangas |
|
|
|
|
|
|
|
|
|
× |
? |
|
|
||
|
Velociraptor mongoliensis |
|
× |
× |
|
|
|
|
|
|
|
|
|
|
||
|
Velociraptor osmolskae |
|
|
|
|
|
|
|
|
|
|
× |
|
|
||
|
Velociraptor sp. |
|
|
|
|
|
|
|
|
|
|
|
× |
× |
||
|
Gobipteryx minuta |
|
|
|
|
|
|
|
|
|
× |
|
× |
× |
||
|
Croco- dyliformes |
Gobiosuchus kielanae |
|
× |
|
|
|
|
? |
|
|
|
|
|
|
|
|
Gobiosuchus sp. |
|
|
× |
|
|
|
|
|
|
|
|
|
|
||
|
Shamosuchus djadochtaensis |
|
× |
|
|
|
|
|
|
|
× |
|
|
|
||
|
Squamata |
Iguania |
Isodontosaurus gracilis |
|
× |
× |
|
|
|
|
|
|
× |
× |
|
|
|
Mimeosaurus crassus |
|
× |
? |
|
|
|
|
|
|
× |
× |
|
|
||
|
Phrynosomimus asper |
|
|
|
|
|
|
|
|
|
× |
|
× |
× |
||
|
Priscagama gobiensis |
|
× |
|
|
|
|
|
|
|
× |
× |
× |
× |
||
|
Pleurodontagama aenigmatoides |
|
|
|
|
|
|
|
|
|
|
× |
× |
|
||
|
Temujinia ellisoni |
|
|
× |
|
|
|
|
|
|
× |
|
|
|
||
|
Anguimorpha |
Cherminotus longifrons |
|
|
× |
|
|
|
|
|
|
? |
|
× |
|
|
|
Carusia intermedia |
|
× |
|
|
|
|
|
|
|
× |
× |
× |
× |
||
|
Gobiderma pulchra |
|
|
× |
|
|
× |
|
|
|
× |
? |
× |
× |
||
|
Estesia mongoliensis |
|
× |
|
|
|
|
|
|
|
× |
|
× |
× |
||
|
Paravaranus angustifrons |
|
|
|
|
|
|
|
|
|
|
|
× |
× |
||
|
Parviderma inexacta |
|
|
|
|
|
|
|
|
|
|
× |
|
× |
||
|
Teiidae |
Adamisaurus magnidentatus |
|
× |
× |
|
|
|
|
|
|
× |
× |
× |
× |
|
|
Cherminsaurus kozlowskii |
|
|
|
|
|
|
|
|
|
|
|
× |
|
||
|
Cherminsaurus tereschenkoi |
|
|
|
|
|
|
|
|
|
|
|
|
× |
||
|
Conicodontosaurus djadochtaensis |
|
× |
|
|
|
|
|
|
|
|
× |
? |
|
||
|
Gobileon emersoculatus |
|
|
|
|
|
|
|
|
|
|
|
× |
× |
||
|
Gobinatus arenosus |
|
|
|
|
|
|
|
|
|
? |
? |
× |
× |
||
|
Gilmoreteius ferrugenous |
|
× |
|
|
|
|
|
|
|
|
|
|
|
||
|
Gilmoreteius chulsanensis |
|
|
|
|
|
|
|
|
|
|
|
× |
× |
||
|
Gilmoreteius gilmorei |
|
|
|
|
|
|
|
|
|
|
|
× |
× |
||
|
Gilmoreteius sulimskii |
|
|
|
|
|
|
|
|
|
|
|
× |
|
||
|
Gilmoreteius sp. |
|
|
|
|
|
|
|
|
|
× |
|
|
|
||
|
Pyramicephalosaurus cherminicus |
|
|
|
|
|
|
|
|
|
|
|
× |
× |
||
|
Tchingisaurus multivagus |
|
|
|
|
|
|
|
|
|
× |
? |
× |
|
||
|
other |
Eoxanta lacertifrons |
|
|
|
|
|
|
|
|
|
× |
|
× |
× |
|
|
Globaura venusta |
|
× |
|
|
|
|
|
|
|
× |
|
× |
× |
||
|
Parmeosaurus scutatus |
|
? |
|
|
|
|
|
|
|
× |
× |
|
|
||
|
Slavoia darevskii |
|
|
|
|
|
|
|
|
|
× |
|
× |
× |
||
| |
Taxa |
AT Fm. |
Djadokhta Fm. |
? |
BM Fm. |
Baruungoyot Fm. |
|||||||||
|
AT |
BZ |
TS |
ZK |
CB |
US |
US-PIN |
YK |
BT |
UT |
BM |
HT |
Kh |
|||
|
Mammalia |
Allotheria |
Bulganbaatar nemegtbaataroides |
|
× |
|
|
|
|
|
|
|
? |
|
|
|
|
Catopsbaatar catapsaloides |
|
|
|
|
|
|
|
|
|
? |
|
× |
× |
||
|
Chulsanbaatar vulgaris |
|
|
|
|
|
|
|
|
|
× |
|
× |
× |
||
|
Djadochtatherium matthewi |
|
× |
× |
|
|
|
|
|
|
× |
× |
|
|
||
|
Kamptobaatar kuczynskii |
|
× |
|
|
|
|
|
|
|
? |
|
|
|
||
|
Kryptobaatar dashzevegi |
|
× |
× |
|
|
|
|
|
|
× |
× |
× |
|
||
|
Nemegtbaatar gobiensis |
|
? |
|
|
|
|
|
|
|
? |
|
× |
× |
||
|
Sloanbaatar mirabilis |
|
× |
|
|
|
|
|
|
|
? |
|
|
|
||
|
Tombaatar sabuli |
|
|
|
|
|
|
|
|
|
× |
? |
|
|
||
|
Eutheria |
Asioryctes nemegtensis |
|
|
|
|
|
|
|
|
|
|
|
× |
× |
|
|
Barunlestes butleri |
|
|
|
|
|
|
|
|
|
|
|
× |
× |
||
|
Kennalestes gobiensis |
|
× |
× |
|
|
|
|
|
|
? |
? |
|
|
||
|
Zalambdalestes grangeri |
|
× |
|
|
|
|
|
|
|
|
|
|
|
||
|
Zalambdalestes lechei |
|
× |
× |
|
|
|
|
|
|
× |
|
|
|
||
|
Zalambdalestes sp. |
|
× |
× |
|
|
|
|
|
|
|
× |
|
|
||
|
Meta- theria |
Deltatheridium pretrituberculare |
|
× |
|
|
? |
? |
|
|
|
× |
? |
× |
? |
|
|
Deltatheroides cretacicus |
|
× |
|
|
|
|
|
|
|
× |
|
|
|
||
|
Hyotheridium dobsoni |
|
× |
|
|
|
|
|
|
|
? |
|
|
|
||
Ivakhnenko and Kurzanov (1988) reported the collection from Üüden Sair of at least eight specimens of the oviraptorosaur Avimimus portentosus (including the holotype) by the Soviet-Mongolian Expeditions. They come from the layers below and above the lens with the holotype of the ceratopsian Udanoceratops tschizhovi (according to Kurzanov 1992, it comes from the Bed 2; Ivakhnenko and Kurzanov 1988: fig. 2). The presence of Avimimus was considered typical for the Nemegt-age deposits of the Gobi Desert (Watabe et al. 2010; Funston et al. 2018). Hence, it was suggested that the specimens collected by the Soviet-Mongolian Palaeontological Expeditions in Üüden Sair in fact come from the nearby locality known as the Yagaan Khovil, where the sediments of the Nemegt Formation are exposed (Watabe et al. 2010). The Üüden Sair section lithology (sensu Ivakhnenko and Kurzanov 1988) is similar, but not identical to that of Yagaan Khovil as described by Watabe et al. (2010). The co-occurrence of Udanoceratops with Avimimus may suggest that the former is the only ceratopsian taxon known from the Nemegt Formation.
On the other hand, the material of Avimimus from the typical Nemegt Formation localities (Nemegt, Bügiin Tsav) was recognized recently by Funston et al. (2018) as a separate species, A. nemegtensis. Funston et al. (2019) suggested that Üüden Sair, with the record of A. portentosus, may instead belong to the Djadokhta Formation. In addition, one fragmentary specimen of a gobiosuchid crocodyliform, collected from there by the Soviet-Mongolian Expeditions and described as Gobiosuchus parvus, may be conspecific with Gobiosuchus kielanae, known from the Djadokhta Formation (Storrs and Efimov 2000). This may indicate that the place discovered by the Soviet-Mongolian Expeditions is really the same as the more recently explored Üüden Sair area. Future investigation of the Üüden Sair and Yagaan Khovil localities, and particularly the confirmation of the purported co-occurrence in there of A. portentosus, Udanoceratops and P. andrewsi, will solve this issue.
As mentioned, the number of protoceratopsid specimens displaying the apomorphic features (especially the length of the maxillary diastema and the absence of the premaxillary dentition) varies in samples collected from different sites (Fig. 6). With the presence of the cf. Bagaceratops sp. specimen described here, this variation may support an interpretation that observed changes reflect the evolution of a single lineage within a restricted area through time and related changing environment (Fig. 7). Hence, deposits of Üüden Sair may be younger than those from Bayan Zag and Tögrögiin Shiree.
Other protoceratopsid localities in the Ömnögovi Province, Mongolia.—Protoceratopsid remains from localities of the Baruungoyot Formation were redescribed recently (Czepiński 2019). From the Hermiin Tsav locality, nearly 60 protoceratopsid specimens, mostly medium-sized, were collected (see SOM). Nearly all diagnostic specimens belong to Bagaceratops rozhdestvenskyi. Only one fragmentary dentary (ZPAL MgD-I/139a) with the plesiomorphic condition of a smooth buccal crest may represent another protoceratopsid (Protoceratops sp., or Breviceratops, the latter known from the sediments of the Baruungoyot Formation; Czepiński 2019). From the Khulsan locality, four specimens of Bagaceratops rozhdestvenskyi and two specimens of Breviceratops kozlowskii are currently known (Makovicky 2002; Czepiński 2019).
The Ukhaa Tolgod locality is the richest Late Cretaceous site of the Gobi Desert so far. It was variously assigned to either the Djadokhta Formation (Dingus et al. 2008) or thought to be even younger than the Baruungoyot Formation (Makovicky 2008). The faunal composition is similar to both the Djadokhta and Baruungoyot formations (Table 1). An abundant collection of protoceratopsid material belongs to a plausibly new taxon (Peter Makovicky and Mark Norell, personal communication 2017).
From Gilbent Uul (Gilbentu), two protoceratopsid specimens consisting of vertebrae were described as Protoceratops sp. (PIN 4550/1; Tereschenko 1996) and Bagaceratops sp. (PIN 4550/3; Tereschenko 2007). However, no cranial material was reported from there, hence these identifications are dubious.
From the Bor Tolgoi locality (?Djadokhta Formation), west of Üüden Sair, one specimen of protoceratopsid, MPC-D 100/550, was illustrated by Tsogtbaatar and Chinzorig (2010). It is the skull of a large individual with paired nasals, which can be assigned to Protoceratops sp.
The Chimney Buttes locality is a small exposure close to Tögrögiin Shiree. It was suggested that it lies stratigraphically between the older Bayan Zag and younger Tögrögiin Shiree sediments (Norell and Makovicky 1999). A specimen of Velociraptor mongoliensis was collected here, in addition to protoceratopsids, ankylosaurids, lizards, and mammals (Gao and Norell 2000). The protoceratopsid remains were neither described nor illustrated, and were tentatively referred to P. andrewsi by Norell and Makovicky (1999). The small skull IGM 100/1078 was discovered in the same locality in 1993 (Carl Mehling, personal communication 2018). The presence of a premaxillary dentition is uncertain, given the poor preservation of the specimen. However, the paired nasal bones suggest that most likely the specimen belongs to Protoceratops sp.
The Alag Teeg locality was discovered by the Soviet-Mongolian Paleontological Expeditions (Currie et al. 2011). It was originally assigned to the Djadokhta Formation (Tverdochlebov and Tsybin 1974; Watabe et al. 2010; Tsogtbaatar et al. 2014), although a new stratigraphic unit, the Alagteeg Formation was recently established, underlying the Djadokhta Formation, (Hasegawa et al. 2009; Burns et al. 2015). The most common dinosaur remains belong to the ankylosaur Pinacosaurus grangeri (Currie et al. 2011; Burns et al. 2015); however, it might represent a distinct species (Averianov and Lopatin 2020). The hadrosauroid Plesiohadros djadokhtaensis (Tsogtbaatar et al. 2014) was also described from that locality. Although present, protoceratopsid remains, (referred to P. andrewsi by Currie et al. 2011) are rare; however, no material was described nor illustrated.
Protoceratopsid material from the Dornogovi Province, Mongolia.—The Baga Tariach (= Baga Tariachi) locality has yielded several remains of ceratopsians. The sediments exposed there were preliminarily assigned to the Baruungoyot Formation (Watabe and Suzuki 2000). PIN 4046/11, a ceratopsian tarsal, was assigned by Tereschenko (2008) to Udanoceratops sp. Fragmentary skulls of protoceratopsid dinosaurs were collected there by the HMNS-MPC Expedition in 1994 (Watabe and Suzuki 2000); however, they were neither illustrated nor described.
In 1977, a medium-sized skull (MPC-D 100/517) was collected from the Shurg Uul (Shurguuli = Shurguul’) locality of the Dornogovi Province. The detailed coordinates or the locality are unknown, and there is no record of the site in the scientific literature. The specimen is housed at the exhibition in Hunnu Mall, Ulaanbaatar, Mongolia, and was provisionally assigned to Protoceratops andrewsi. The plesiomorphic condition of the paired nasals and smooth buccal crest of the dentary make it similar to Protoceratops spp. However, no premaxillary teeth can be traced at the ventral margin of the premaxilla, and the area of the premaxillo-maxillary suture is eroded on both sides, preventing determination of the presence of accessory antorbital fenestration (personal observation). MPC-D 100/517 is determined here as Protoceratops sp.
Protoceratopsid material from Inner Mongolia, China.— The Bayan Mandahu locality was considered as representing deposits equivalent to the Djadokhta Formation (Jerzykiewicz et al. 1993), although these deposits are sometimes assigned to a distinct unit, the Bayan Mandahu (= Wulansuhai) Formation (Godefroit et al. 2008; Longrich et al. 2010). The faunal composition of Bayan Mandahu is quite different from that of the Djadokhta Formation, with the presence of a few species of squamates (Carusia intermedia and Pleurodontagama aenigmatoides) typical for the Baruungoyot Formation (Gao and Hou 1996), and with protoceratopsid taxa with a more derived anatomy than in the Djadokhta Formation (Table 1). The ankylosaurian Pinacosaurus grangeri is also known from the Djadokhta Formation (Burns et al. 2011). However, material from Bayan Mandahu was described as a distinct species, Pinacosaurus mephistocephalus (Godefroit et al. 1999). It is possible that the age of Bayan Mandahu strata is in fact younger than the Djadokhta Formation (Fig. 7; Longrich et al. 2010).
Most of the diagnostic ceratopsian material from Bayan Mandahu belongs to Protoceratops hellenikorhinus (Lambert et al. 2001). Only one specimen (IVPP V12513) can be referred to Bagaceratops rozhdestvenskyi (Czepiński 2019). No specimen of Protoceratops andrewsi could be unambiguously identified from Bayan Mandahu.
However, distinguishing P. andrewsi and P. hellenikorhinus may be challenging. Features suggested as diagnostic for the latter, including the development of the nasal horn and the parietosquamosal frill (Lambert et al. 2001), are subtle and most likely related to ontogeny (Morschhauser 2012). Moreover, the alleged single autapomorphy of P. andrewsi, the small process projecting posteriorly on the anterior portion of the parietal fenestrae (Makovicky 2002; Morschhauser 2012), can also be observed in some undescribed protoceratopsid specimens from Bayan Mandahu (own unpublished data). Nevertheless, the very large holotype of P. hellenikorhinus has proportionally shorter exoccipitals and fewer maxillary teeth than the relatively smaller individuals of P. andrewsi (Lambert et al. 2001; Czepiński 2019). Several specimens of P. andrewsi from Bayan Zag and Üüden Sair lack the premaxillary dentition, the state observed in all material assigned to P. hellenikorhinus. A plausible explanation is that P. hellenikorhinus emerged from a population of P. andrewsi, where the phenotype of the toothless premaxilla was established. For a better understanding of the relationship between P. andrewsi and P. hellenikorhinus, more material from Bayan Mandahu, with further studies on ontogeny and intraspecific variation, is required.
Although the presence of the leptoceratopsid Udanoceratops in Bayan Mandahu was reported (Jerzykiewicz et al. 1993), no single described specimen can be assigned to that taxon. It is possible that the very large specimens of P. hellenikorhinus were erroneously interpreted as Udanoceratops, because of the similar size to the holotype of the latter (Kurzanov 1992).
The Ulan Tsonch locality is placed 25 km east of Bayan Mandahu. It was assigned to the Minhe Formation, Lower Cretaceous in age (Lucas and Estep 1998). Bohlin (1953) described fragmentary material (two dentaries and isolated teeth) from that locality, which he assigned to Protoceratops andrewsi. Personal observations of the casts housed in New York (AMNH 2034–2042) confirmed the undiagnostic nature of this fragmentary material, as suggested by Lambert et al. (2001). With the mandible anatomy being plesiomorphic for neoceratopsian dinosaurs, I suggest considering the material described by Bohlin as a Neoceratopsia indet.
Protoceratopsid material from Alxa, Inner Mongolia, was reported recently (Ji et al. 2017). It was found with material of Pinacosaurus sp. that suggests similarities with the Djadokhta Formation and the Bayan Mandahu locality. Protoceratopsids from Alxa have no premaxillary dentition (Ji Shu’an, personal communication 2018), a condition similar to P. hellenikorhinus from Bayan Mandahu.
Chen (2017) assigned a fragmentary skull lacking the snout, roof and frill (GMV V02651) from the Early Cretaceous of the Ordos Basin, Inner Mongolia, to Protoceratops sp. It is relatively large in size (25 cm in length of the preserved portion); however, due to its fragmentary nature, the lack of diagnostic features, and the reported geological age (found together with remains of Psittacosaurus neimongoliensis), it should probably be considered as an indetermined neoceratopsian.
Conclusions
New material of protoceratopsid dinosaurs from Mongolia helps comparing several Upper Cretaceous sites of the Gobi Desert with a poorly constrained stratigraphic context. Specimens of Protoceratops andrewsi from Zamyn Khond enable correlation with the typical Djadokhta Formation localities in Bayan Zag and Tögrögiin Shire.
One protoceratopsid specimen collected from the Üüden Sair locality is intermediate in morphology between P. andrewsi and Bagaceratops rozhdestvenskyi. It possesses a fused nasal horn, a sharp buccal crest of the dentary, and lacks the premaxillary dentition (apomorphic features of B. rozhdestvenskyi). At the same time, it has no accessory antorbital fenestration at the premaxillo-maxillary suture, a plesiomorphic condition seen in P. andrewsi. The specimen, referred here to cf. Bagaceratops sp., supports the hypothesis of an anagenetic transition from P. andrewsi to B. rozhdestvenskyi. It seems reasonable to propose that the samples of protoceratopsid specimens from the adjacent Djadokhta Formation localities may represent a single lineage. Differences observed in the anatomy of P. andrewsi from different sites may reflect the gradual change of these features (with an increasing contribution of the Bagaceratops phenotype) within the lineage. It probably reflects the evolution of the population through time.
The age of the sediments in Üüden Sair is plausibly intermediate between the Djadokhta and Baruungoyot formations. However, the remaining question is whether the site examined by the Soviet-Mongolian Paleontological Expeditions is the same as the recently explored Üüden Sair area, or whether it is a distinct location with sediments of the Nemegt Formation. In the affirmative, does the co-occurrence of Udanoceratops with Avimimus material constitute evidence for the record of ceratopsians in the Nemegt Formation.
The wide variation observed within the abundant protoceratopsid material in the Late Cretaceous of the Gobi Desert reflects a major issue in palaeontology, namely the definition and distinction of species in the fossil record. On the other hand, the rich vertebrate material from the Late Cretaceous of the Gobi Desert can enable research at a nearly-populational level through deep time.
The hypothesis on the temporal ordering of the Late Cretaceous Gobi paleobiotas, based on dinosaur assemblages as presented here, could be tested through the examination of intraspecific variation and potential lineage evolution within the other well-represented vertebrate taxa from these deposits, for example squamates and mammals.
Acknowledgements
I thank Khishigjav Tsogtbaatar and Ulziitseren Sanjaadash (both Institute of Paleobiology, Mongolian Academy of Sciences, Ulaanbaatar, Mongolia) for help during my visit in Ulaanbaatar. I am grateful to Andrew Farke (Raymond M. Alf Museum of Paleontology, Claremont, USA), Mark Loewen (Natural History Museum of Utah, Salt Lake City, USA), Olivier Lambert (Royal Belgian Institute of Natural Sciences, Brussels, Belgium), Mateusz Tałanda and Jerzy Dzik (both University of Warsaw, Poland) for useful suggestions that improved the manuscript. Lukas Panzarin (Museo Paleontologico Cittadino, Monfalcone, Italy) provided the archival photographs of the specimens from the MPC. Frank Varriale (King’s College London, UK) and Andrew Knapp (Queen Mary University of London, UK) helped to obtain the historical details on the MPC specimens. Dave Hone (Queen Mary University of London, UK), Philip Currie (University of Alberta, Edmonton,, Canada), and Ji Shu’an (Chinese Academy of Geological Sciences, Beijing, China) provided details on the material collected from Inner Mongolia. This work was supported by the foundation Universitatis Varsoviensis (grant “Diversity of the horned dinosaurs from the Central Asia”).
References
Alifanov, V.R. 2012. Order Lacertilia [in Russian]. In: E.N. Kuročkin and A.V. Lopatin (eds.), Iskopaemye pozwonočnye Rossii i sopredel’nyh stran. Iskopaemye reptilii i pticy. Čast’ 2, 7–136, GEOS, Moskva.
Arbour, V.M. and Currie, P.J. 2013. The taxonomic identity of a nearly complete ankylosaurid dinosaur skeleton from the Gobi Desert of Mongolia. Cretaceous Research 46: 24–30. Crossref
Arbour, V.M., Currie, P.J., and Badamgarav, D. 2014. The ankylosaurid dinosaurs of the Upper Cretaceous Baruungoyot and Nemegt formations of Mongolia. Zoological Journal of the Linnean Society 172: 631–652. Crossref
Averianov, A.O. and Lopatin, A.V. 2020. An unusual new sauropod dinosaur from the Late Cretaceous of Mongolia. Journal of Systematic Palaeontology [published online http://doi.org/10.1080/14772019.2020.1716402]. Crossref
Averianov, A.O. and Sues, H.D. 2012. Correlation of Late Cretaceous continental vertebrate assemblages in Middle and Central Asia. Journal of Stratigraphy 36: 462–485.
Barsbold, R. 1983. Carnivorous dinosaurs from the Cretaceous of Mongolia [in Russian]. Sovmestnaâ Sovetsko-Mongol’skaâ Paleontologičeskaâ Ekspeditciâ, Trudy 19: 1–117.
Barsbold, R. I988. About the bone crest and the horn cover of the carnivorous-oviraptors dinosaurs [in Russian]. Sovmestnaâ Sovetsko-Mongol’skaâ Paleontologičeskaâ Ekspeditciâ, Trudy 34: 77–80.
Benton, J.M. 2000. Mongolian place names and stratigraphic terms. In: M.J. Benton, M.A. Shishkin, D.M. Unwin, and E.N. Kurochkin (eds.), The Age of Dinosaurs in Russia and Mongolia, xxii–xxviii. Cambridge University Press, Cambridge.
Bohlin, B. 1953. Fossil reptiles from Mongolia and Kansu. The Sino-Swedish Expedition, Publication 37: 1–105.
Brown, D.B. and Schlaikjer, D.E.M. 1940. The structure and relationships of Protoceratops. Transactions of the New York Academy of Sciences 2: 99–100. Crossref
Burns, M.E., Currie, P.J., Sissons, R.L., and Arbour, V.M. 2011. Juvenile specimens of Pinacosaurus grangeri Gilmore, 1933 (Ornithischia: Ankylosauria) from the Late Cretaceous of China, with comments on the specific taxonomy of Pinacosaurus. Cretaceous Research 32: 174–186. Crossref
Burns, M.E., Tumanova, T.A., and Currie, P.J. 2015. Postcrania of juvenile Pinacosaurus grangeri (Ornithischia: Ankylosauria) from the Upper Cretaceous Alagteeg Formation, Alag Teeg, Mongolia: implications for ontogenetic allometry in ankylosaurs. Journal of Paleontology 89: 168–182.Crossref
Carpenter, K., Hayashi, S., Kobayashi, Y., Maryańska, T., Barsbold, R., Sato, K., and Obata, I. 2011. Saichania chulsanensis (Ornithischia, Ankylosauridae) from the Upper Cretaceous of Mongolia. Palaeontographica Abteilung A 294: 1–61. Crossref
Carr, T.D., Varricchio, D.J., Sedlmayr, J.C., Roberts, E.M., and Moore, J.R. 2017. A new tyrannosaur with evidence for anagenesis and crocodile-like facial sensory system. Scientific Reports 7: 44942. Crossref
Chen, X. 2017. The discovery and significance of the protoceratopsid dinosaur from the Lower Cretaceous of Ordos Region, Inner Mongolia. Acta Geoscientica Sinica 38: 154–158.
Chinzorig, T., Kobayashi, Y., Tsogtbaatar, K., Currie, P.J., Watabe, M., and Barsbold, R. 2017. First ornithomimid (Theropoda, Ornithomimosauria) from the Upper Cretaceous Djadokhta Formation of Tögrögiin Shiree, Mongolia. Scientific Reports 7: 5835. Crossref
Clark, J.M., Norell, M.A., and Rowe, T. 2002. Cranial anatomy of Citipati osmolskae (Theropoda, Oviraptorosauria), and a reinterpretation of the holotype of Oviraptor philoceratops. American Museum Novitates 2002: 1–24. Crossref
Currie, P.J. 2016. Dinosaurs of the Gobi: Following in the footsteps of the Polish-Mongolian Expeditions. Palaeontologica Polonica 67: 83–100.
Currie, P.J., Badamgarav, D., Koppelhus, E.B., Sissons, R., and Vickaryous, M.K. 2011. Hands, feet, and behaviour in Pinacosaurus (Dinosauria: Ankylosauridae). Acta Palaeontologica Polonica 56: 489–505. Crossref
Czepiński, Ł. 2019. Ontogeny and variation of a protoceratopsid dinosaur Bagaceratops rozhdestvenskyi from the Late Cretaceous of the Gobi Desert. Historical Biology [published online http://doi.org/10.1080/08912963.2019.1593404]. Crossref
Danilov, I.G., Sukhanov, V.B., and Syromyatnikova, E.V. 2013. Redescription of Zangerlia dzamynchondi (Testudines: Nanhsiungchelyidae) from the Late Cretaceous of Mongolia, with a reassessment of the phylogenetic position and relationships of Zangerlia. In: D.B. Brinkman, P.A. Holroyd, and J.D. Gardner (eds.), Morphology and Evolution of Turtles, 407–417. Springer, Dordrecht. Crossref
Dashzeveg, D., Dingus, L., Loope, D.B., Swisher, C.C., Dulam, T., and Sweeney, M.R. 2005. New stratigraphic subdivision, depositional environment, and age estimate for the Upper Cretaceous Djadokhta Formation, southern Ulan Nur Basin, Mongolia. American Museum Novitates 2005: 1–31. Crossref
Dingus, L., Loope, D.B., Dashzeveg, D., Swisher, C.C., Minjin, C., Novacek, M.J., and Norell, M.A. 2008. The geology of Ukhaa Tolgod (Djadokhta Formation, Upper Cretaceous, Nemegt Basin, Mongolia). American Museum Novitates 2008: 1–40. Crossref
Dodson, P. 1976. Quantitative aspects of relative growth and sexual dimorphism in Protoceratops. Journal of Paleontology, 50: 929–940.
Dong, L., Xu, X., Wang, Y., and Evans, S.E. 2018. The lizard genera Bainguis and Parmeosaurus from the Upper Cretaceous of China and Mongolia. Cretaceous Research 85: 95–108. Crossref
Dzik, J. 2005. The chronophyletic approach: stratophenetics facing an incomplete fossil record. Special Papers in Palaeontology 73: 159–183.
Elżanowski, A. and Wellnhofer, P. 1992. A new link between theropods and birds from the Cretaceous of Mongolia. Nature 359: 821–823. Crossref
Farke, A.A. 2010. Evolution, homology, and function of the supracranial sinuses in ceratopsian dinosaurs. Journal of Vertebrate Paleontology 30: 1486–1500. Crossref
Funston, G.F., Currie, P.J., Ryan, M.J., and Dong, Z.M. 2019. Birdlike growth and mixed-age flocks in avimimids (Theropoda, Oviraptorosauria). Scientific Reports 9: 18816. Crossref
Funston, G.F., Mendonca, S.E., Currie, P.J., and Barsbold, R. 2018. Oviraptorosaur anatomy, diversity and ecology in the Nemegt Basin. Palaeogeography, Palaeoclimatology, Palaeoecology 494: 101–120. Crossref
Gao, K. and Hou, L. 1996. Systematics and taxonomic diversity of squamates from the Upper Cretaceous Djadochta Formation, Bayan Mandahu, Gobi Desert, People’s Republic of China. Canadian Journal of Earth Sciences 33: 578–598. Crossref
Gao, K. and Norell, M.A. 2000. Taxonomic composition and systematics of Late Cretaceous lizard assemblages from Ukhaa Tolgod and adjacent localities, Mongolian Gobi Desert. Bulletin of the American Museum of Natural History 2000: 1–118. Crossref
Godefroit, P., Currie, P.J., Hong, L., Yong, S.C., and Dong, Z. 2008. A new species of Velociraptor (Dinosauria: Dromaeosauridae) from the Upper Cretaceous of northern China. Journal of Vertebrate Paleontology 28: 432–438. Crossref
Godefroit, P., Suberbiola, X.P., Li, H., and Dong, Z. 1999, A new species of the ankylosaurid dinosaur Pinacosaurus from the late cretaceous of inner Mongolia (P.R. China): Bulletin de l’Institut Royal des Sciences Naturelles de Belgique, Sciences de la terre 69: 17–36.
Gradziński, R., Kielan-Jaworowska, Z., and Maryańska, T. 1977. Upper Cretaceous Djadokhta, Barun Goyot and Nemegt formations of Mongolia, including remarks on previous subdivisions. Acta Geologica Polonica 27: 281–318.
Granger, W. and Gregory W.K. 1923. Protoceratops andrewsi, a pre-ceratopsian dinosaur from Mongolia. American Museum Novitates 72: 1–7.
Handa, N., Watabe, M., and Tsogtbaatar, K. 2012. New specimens of Protoceratops (Dinosauria: Neoceratopsia) from the Upper Cretaceous in Udyn Sayr, southern Gobi area, Mongolia. Paleontological Research 16: 179–199. Crossref
Hasegawa, H., Tada, R., Ichinnorov, N., and Minjin, C. 2009. Lithostratigraphy and depositional environments of the Upper Cretaceous Djadokhta Formation, Ulan Nuur basin, southern Mongolia, and its paleoclimatic implication. Journal of Asian Earth Sciences 35: 13–26. Crossref
Hone, D.W. and Naish, D. 2013. The “species recognition hypothesis” does not explain the presence and evolution of exaggerated structures in non-avialan dinosaurs. Journal of Zoology, 290: 172–180. Crossref
Hone, D.W., Wood, D., and Knell, R.J. 2016. Positive allometry for exaggerated structures in the ceratopsian dinosaur Protoceratops andrewsi supports socio-sexual signaling. Palaeontologia Electronica 19: 1–13. Crossref
Horner, J.R., Varricchio, D.J., and Goodwin, M.B. 1992. Marine transgressions and the evolution of Cretaceous dinosaurs. Nature 358: 59–61. Crossref
Hunt, G. and Rabosky, D.L. 2014. Phenotypic evolution in fossil species: pattern and process. Annual Review of Earth and Planetary Sciences 42: 421–441. Crossref
Ivakhnenko, M.F. [Ivahnenko, M.F.] and Kurzanov, S.M. 1988. Geological structure and age of localities Udan Sayr and Shara Tsav [in Russian]. Sovmestnaâ Sovetsko-Mongol’skaâ Paleontologičeskaâ Ekspeditciâ, Trudy 34: 100–105.
Jerzykiewicz, T. 2000. Lithostratigraphy and sedimentary settings of the Cretaceous dinosaur beds of Mongolia. In: M.J. Benton, M.A. Shishkin, and D.M. Unwin (eds.), The Age of Dinosaurs in Russia and Mongolia, 279–296, Cambridge University Press, Cambridge.
Jerzykiewicz, T., and Russell, D.A. 1991. Late Mesozoic stratigraphy and vertebrates of the Gobi Basin. Cretaceous Research 12: 345–377. Crossref
Jerzykiewicz, T., Currie, P.J., Eberth, D.A., Johnston, P.A., Koster, E.H., and Zheng, J.J. 1993. Djadokhta Formation correlative strata in Chinese Inner Mongolia: an overview of the stratigraphy, sedimentary geology, and paleontology and comparisons with the type locality in the pre-Altai Gobi. Canadian Journal of Earth Sciences 30: 2180–2195. Crossref
Ji, S.A., Zhang, L., Lu, L., and Hao, J. 2017. The first discovery of the Late Cretaceous protoceratopsid fauna from Alxa, Inner Mongolia, China. Acta Geologica Sinica 91: 1908–1909. Crossref
Kielan-Jaworowska, Z., Hurum, J.H., and Badamgarav, D. 2003. An extended range of the multituberculate Kryptobaatar and distribution of mammals in the Upper Cretaceous of the Gobi Desert. Acta Palaeontologica Polonica 48: 273–278.
Kim, B., Yun, H., and Lee, Y-N. 2019. The postcranial skeleton of Bagaceratops (Ornithischia: Neoceratopsia) from the Baruungoyot Formation (Upper Cretaceous) in Hermiin Tsav of southwestern Gobi, Mongolia [in Korean]. Journal of the Geological Society of Korea 55: 179–190. Crossref
Knapp, A., Knell, R.J., Farke, A.A., Loewen, M.A., and Hone, D.W. 2018. Patterns of divergence in the morphology of ceratopsian dinosaurs: sympatry is not a driver of ornament evolution. Proceedings of the Royal Society B: Biological Sciences 285:20180312. Crossref
Kurzanov, S.M. 1992. A gigantic protoceratopsid from the Upper Cretaceous of Mongolia. Paleontological Journal 26: 103–116.
Lambert, O., Godefroit, P., Li, H., Shang, C.Y., and Dong, Z.M. 2001. A new species of Protoceratops (Dinosauria, Neoceratopsia) from the Late Cretaceous of Inner Mongolia (P.R. China). Bulletin de l’Institut royal des Sciences naturelles de Belgique, Sciences de la terre 71: 5–28.
Lawson, R. and Lieb, C.S. 1990. Variation and hybridization in Elaphe bairdi (Serpentes: Colubridae). Journal of Herpetology 24: 280–292. Crossref
Lillegraven, J.A. and McKenna, M.C. 1986. Fossil mammals from the “Mesaverde” Formation (Late Cretaceous, Judithian) of the Bighorn and Wind River basins, Wyoming, with definitions of Late Cretaceous North American land-mammal “ages”. American Museum Novitates 2840: 1–68.
Longrich, N.R., Currie, P.J., and Dong, Z. 2010. A new oviraptorid (Dinosauria: Theropoda) from the Upper Cretaceous of Bayan Mandahu, Inner Mongolia. Palaeontology 53: 945–960. Crossref
Lucas, S.G. and Estep, J.W. 1998. Vertebrate biostratigraphy and biochronology of the Cretaceous of China. New Mexico Museum of Natural History and Science Bulletin 14: 1–20.
Makovicky, P.J. 2002. Taxonomic Revision and Phylogenetic Relationships of Basal Neoceratopsia (Dinosauria: Ornithischia). 288 pp. Ph.D. Thesis, Columbia University, New York, NY.
Makovicky, P.J. 2008. Telling time from fossils: a phylogeny-based approach to chronological ordering of paleobiotas. Cladistics 24: 350–371. Crossref
Makovicky, P.J. and Norell, M.A. 2006. Yamaceratops dorngobiensis, a new primitive ceratopsian (Dinosauria: ornithischia) from the Cretaceous of Mongolia. American Museum Novitates 3530:1–42. Crossref
Makulbekov, N.M. and Kurzanov, S.M. 1986. The biogeographical relationships of the Late Cretaceous biota of Mongolia [in Russian]. Sovmestnaâ Sovetsko-Mongol’skaâ Paleontologičeskaâ Ekspeditciâ, Trudy 29: 106–112.
Maryańska, T. and Osmólska, H. 1975. Protoceratopsidae (Dinosauria) of Asia. Palaeontologia Polonica 33: 133–181.
McDonald, A.T. 2011. A subadult specimen of Rubeosaurus ovatus (Dinosauria: Ceratopsidae), with observations on other ceratopsids from the Two Medicine Formation. PloS one 6: e22710. Crossref
Morschhauser, E.M. 2012. The Anatomy and Phylogeny of Auroraceratops (Ornithischia: Ceratopsia) from the Yujingzi Basin of Gansu Province, China. 629 pp. PhD Thesis, University of Pennsylvania, Philadelphia.
Nabavizadeh, A. and Weishampel, D.B. 2016. The predentary bone and its significance in the evolution of feeding mechanisms in ornithischian dinosaurs. The Anatomical Record 299: 1358–1388. Crossref
Norell, M. and Makovicky, P.J. 1999. Important Features of the Dromaeosaurid Skeleton II: Information from Newly Collected Specimens of Velociraptor mongoliensis. American Museum Novitates 3282: 1–45.
Osmólska, H. 1980. The Late Cretaceous vertebrate assemblages of the Gobi Desert, Mongolia. Memoirs de la Societé Geologique de France 139: 145–150.
Padian, K. and Horner, J.R. 2011. The evolution of “bizarre structures” in dinosaurs: biomechanics, sexual selection, social selection or species recognition? Journal of Zoology 283: 3–17. Crossref
Pei, R., Norell, M.A., Barta, D.E., Bever, G.S., Pittman, M., and Xu, X. 2017. Osteology of a new Late Cretaceous troodontid specimen from Ukhaa Tolgod, Ömnögovi Aimag, Mongolia. American Museum Novitates 2017: 1–48. Crossref
Rougier, G.W., Sheth, A.S., Spurlin, B.K., Bolortsetseg, M., and Novacek, M.J. 2016. Craniodental anatomy of a new Late Cretaceous multituberculate mammal from Udan Sayr, Mongolia. Palaeontologia Polonica 67: 197–248.
Scannella, J.B., Fowler, D.W., Goodwin, M.B., and Horner, J.R. 2014. Evolutionary trends in Triceratops from the Hell Creek formation, Montana. Proceedings of the National Academy of Sciences 111: 10245–10250. Crossref
Shuvalov, V.F. 2000. The Cretaceous stratigraphy and palaeobiogeography of Mongolia. In: M.J. Benton, M.A. Shishkin, and D.M. Unwin (eds.), The Age of Dinosaurs in Russia and Mongolia, 256–278. Cambridge University Press, Cambridge.
Shuvalov, V.F. [Šuvalov, V.F.], and Chkhikvadze, V.M. [Čhikvadze, V.M.] 1979. On stratigraphical and systematical position of some freshwater turtles from new Cretaceous localities in Mongolia [in Russian]. Sovmestnaâ Sovetsko-Mongol’skaâ Paleontologičeskaâ Ekspeditciâ, Trudy 8: 58–76.
Smith, J.M. 1966. Sympatric speciation. The American Naturalist 100: 637–650. Crossref
Storrs, G.W. and Efimov, M.B. 2000. Mesozoic crocodyliforms of north-central Eurasia. In: M.J. Benton, M.A., Shishkin, D.M. Unwin, and E.N. Kurochkin (eds.), The Age of Dinosaurs in Russia and Mongolia, 402–419. Cambridge University Press, Cambridge.
Szymura, J.M. 1993. Analysis of hybrid zones with Bombina. In: R.G. Harrison and R.G. Harrison (eds.), Hybrid Zones and the Evolutionary Process, 261–289. Oxford University Press, Oxford.
Tereschenko, V.S. 1996. A reconstruction of the locomotion of Protoceratops. Paleontological Journal 30: 232–245.
Tereschenko, V.S. 2007. Key to Protoceratopoid vertebrae (Ceratopsia, Dinosauria) from Mongolia. Paleontological Journal 41: 175–188. Crossref
Tereschenko, V.S. 2008. Adaptive features of protoceratopoids (Ornithischia: Neoceratopsia). Paleontological Journal 42: 273–286. Crossref
Tereschenko, V.S. 2018. On Polymorphism of Protoceratops andrewsi Granger et Gregory, 1923 (Protoceratopidae, Neoceratopsia). Paleontological Journal 52: 429–444. Crossref
Tong, H., Li, L., Jie, C., and Yi, L. 2016. New material of Jiangxichelys ganzhouensis Tong & Mo, 2010 (Testudines: Cryptodira: Nanhsiungchelyidae) and its phylogenetic and palaeogeographical implications. Geological Magazine 154: 456–464. Crossref
Tsogtbaatar, K. 2004. Fossil specimens prepared in Mongolian Paleontological center 1993–2001. Hayashibara Museum of Sciences Research Bulletin 2: 123–128.
Tsogtbaatar, K. and Chinzorig, T. 2010. Fossil specimens prepared in Mongolian Paleontological Center: 2002–2008 Hayashibara Museum of Sciences Research Bulletin 3: 155–166.
Tsogtbaatar, K., Weishampel, D.B., Evans, D.C., and Watabe, M. 2014. A new hadrosauroid (Plesiohadros djadokhtaensis) from the Late Cretaceous Djadokhtan Fauna of southern Mongolia. In: D.A. Eberth and D.C. Evans (eds.), Hadrosaurs, 108–135. Indiana University Press, Bloomington.
Tsuihiji, T., Barsbold, R., Watabe, M., Tsogtbaatar, K., Chinzorig, T., Fujiyama, Y., and Suzuki, S. 2014. An exquisitely preserved troodontid theropod with new information on the palatal structure from the Upper Cretaceous of Mongolia. Naturwissenschaften 101: 131–142. Crossref
Turner, A.H., Makovicky, P.J., and Norell, M.A. 2012. A review of dromaeosaurid systematics and paravian phylogeny. Bulletin of the American Museum of Natural History 371: 1–206. Crossref
Tverdochlebov, V.P. [Tverdohlebov, V.P.] and Zybin, J.I. 1974. Genesis of the Upper Cretaceous sediments with dinosaur remains at Tugrikin-us and Alag-Taag localities [in Russian]. Sovmestnaâ Sovetsko-Mongol’skaâ Paleontologičeskaâ Ekspeditciâ, Trudy 1: 314–319.
Varriale, F.J. 2011. Dental Microwear and the Evolution of Mastication in Ceratopsian Dinosaurs. 470 pp. Ph.D. Thesis, The Johns Hopkins University, Baltimore.
Watabe, M. and Suzuki, S. 2000. Report on the Japan-Mongolia Joint Paleontological expedition to the Gobi Desert, 1996. Hayashibara Museum of Natural Sciences Research Bulletin 1: 30–44.
Watabe, M. and Tsogtbaatar, K. 2004. Report on the Japan-Mongolia Joint Paleontological expedition to the Gobi Desert, 2000. Hayashibara Museum of Natural Sciences Research Bulletin 2: 45–67.
Watabe, M., Tsogtbaatar, K., Suzuki, S., and Saneyoshi, M. 2010. Geology of dinosaur-fossil-bearing localities (Jurassic and Cretaceous: Mesozoic) in the Gobi Desert: Results of the HMNS-MPC Joint Paleontological Expedition. Hayashibara Museum of Natural Sciences Research Bulletin 3: 41–118.
Wilson, J.P., Ryan, M.J., and Evans, D.C. 2020. A new, transitional centrosaurine ceratopsid from the Upper Cretaceous Two Medicine Formation of Montana and the evolution of the “Styracosaurus-line” dinosaurs. Royal Society Open Science 7: 200284. Crossref
Acta Palaeontol. Pol. 65 (3): 481–497,
2020 https://doi.org/10.4202/app.00701.2019