

A revaluation of rhipidocystid echinoderms based on a new flattened blastozoan from the Upper Ordovician of Maryland, USA
SAMUEL ZAMORA, JAMES SPRINKLE, and COLIN D. SUMRALL
Zamora, S., Sprinkle, J. and Sumrall, C.D. 2020. A revaluation of rhipidocystid echinoderms based on a new flattened blastozoan from the Upper Ordovician of Maryland, USA. Acta Palaeontologica Polonica 65 (3): 455–465.
A new rhipidocystid echinoderm from the Upper Ordovician Chambersburg Formation in western Maryland (USA) is here described based on four exquisitely preserved specimens. Specimens of Durhamicystis americana gen. et sp. nov. preserve both thecal sides and the oral area, with identification of major apertures including peristome, periproct, gonopore, and hydropore. This allows orientation of this species and proper comparison across all rhipidocystids. Durhamicystis americana has a large theca composed of ten marginal plates with only two basals on the posterior side, two ambulacra and seven large oral plates with brachioles attached either on orals or smaller flooring plates. Rhipidocystids include taxa with lateral flattening along the BC-DE axis (i.e., Rhipidocystis) and others along the anterior-posterior A-CD axis (i.e., Durhamicystis, Neorhipidocystis, and Petalocystites) suggesting rhipidocystids may be paraphyletic.
Key words: Echinodermata, Blastozoa, Rhipidocystidae, Paleozoic, Chambersburg Formation, North America.
Samuel Zamora [s.zamora@igme.es], Instituto Geológico y Minero de España (IGME), C/Manuel Lasala, 44, 9ºB, 50006, Zaragoza, Spain and Grupo Aragosaurus-IUCA, Área de Paleontología, Facultad de Ciencias, Universidad de Zaragoza, Zaragoza, Spain.
James Sprinkle [echino@jsg.utexas.edu], Department of Geological Sciences, Jackson School of Geosciences, University of Texas, 1 University Station C1100, Austin TX, 78712-0254, USA.
Colin D. Sumrall [csumrall@utk.edu], Department of Earth and Planetary Sciences, University of Tennessee, Knoxville, TN 37996-1410, USA.
Received 18 December 2019, accepted 12 February 2020, available online 5 June 2020.
Copyright © 2020 S. Zamora et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Rhipidocystids (i.e., Rhipidocystis, Batherocystis, and Mandalacystis) are a small group of Paleozoic echinoderms, exclusively found in the Ordovician, which had a flattened theca, long brachioles, and reduced stems (Jaekel 1901; Hecker 1940; Bassler 1950; Ubaghs 1968; Sprinkle 1973; Bockelie 1981; Lewis et al. 1987; Rozhnov 1989). The family Rhipidocystidae was erected by Jaekel (1901) to accommodate Rhipidocystis, then considered a member of the Carpoidea because it bore a flattened theca. A second genus of rhipidocystids, Batherocystis was described by Bassler (1950) and included within the new family Batherocystidae, as part of Eocrinoidea and was later synonymized with Rhipidocystidae by Ubaghs (1968). Sprinkle (1973) erected the genus Petalocystites that was also included in the family Rhipidocystidae. Mandalacystis Lewis, Sprinkle, Bailey, Moffit, and Parsley, 1987 is one of the better-known taxa within the family, including complete specimens that preserve the stem, brachioles, and the oral area. Some specimens of Rhipidocystis baltica described by Rozhnov (1989) preserved the oral area and show a pentaradial pattern with a BC-DE flattening direction. Rozhnov (1989) transferred some material originally described as Rhipidocystis (Hecker 1938; Bockelie 1981) to the new genus Neorhipidocystis, which included bilateral forms with only the two shared ambulacra. Ambulacral homologies in flattened forms with two ambulacra remain unresolved (Sumrall et al. 2001), and this results in part, from a lack of information from the oral area, which is critical for the understanding of homologies in early pentaradial echinoderms (Sumrall 2010; Kammer et al. 2013; Zamora and Rahman 2014).
The aim of this paper is the description of a historic collection of one of the best-preserved rhipidocystids known. The collection includes four exquisitely preserved specimens showing both sides of the theca documenting that, unlike other rhipidocystids, this species only bears two basals. The anal pyramid opens between the marginal plates on the upper right edge of the theca. Brachioles are long and slender, with a very flexible proximal part and stiffer distal portion. The oral area is well preserved, showing only two shared ambulacra. This feature combined with the position of main apertures, provides a better understanding of rhipidocystids that are here reevaluated.
Institutional abbreviations.—PMO, Paleontologisk museum, Oslo, Norway; USNM, National Museum of Natural History (Smithsonian Institution), Washington DC, USA.
Nomenclatural acts.—This published work and the nomenclatural acts it contains, have been registered in ZooBank: urn:lsid:zoobank.org:pub:2D997956-215B-4290-B5DE-88D6D6CA3652
Geological setting, material, and methods
This paper describes four specimens from a historic collection housed at the Smithsonian Institution (National Museum of Natural History), Washington DC, USA. These specimens are fully articulated in a dark grey highly lithified limestone. The specimens were apparently hand prepared by the original collectors. They are preserved in calcite that appears to be softer than the surrounding matrix.
Information on the original labels indicates that the material was “brought in by amateurs” and shown to John Wyatt Durham, University of California, Berkeley, who was visiting the Smithsonian, on June 6, 1966. This small collection of rhipidocystid echinoderms was collected from the “Edinburg Formation”, as originally defined (see discussion below), in western Maryland (USA). Specimens were collected in an old quarry, west of Conococheague Road, east of Wilson, MD. The first record of these specimens being deposited at the Smithsonian is from June 24, 1969, indicating that at least “two specimens of Rhipidocystis sp. and one specimen of disarticulated rhipidocystid plates” were received by Thomas F. Phelan, who was Curator Porter Kier’s Assistant at that time, who placed them in the Springer Room Echinoderm Collection. Since then, nothing has been done with these specimens, with the exception of JS who, while visiting Durham at the Smithsonian in August, 1967, made a drawing of one of the rhipidocystid brachioles to use in his dissertation (Sprinkle 1973: fig. 5C). While reviewing the collections at the National Museum of Natural History in 2017, one of us (SZ) rediscovered these specimens in a museum drawer, but there is no historical record about what happened with these fossils between 1966 and 2017.
The original label indicates that specimens derive from what was formerly called “Edinburg Formation”, and were collected from the Schemeompf’s old Quarry, located at the top of a hill, west of Conococheague Creek, about 2.1 miles (3.4 km) NNE and 0.5 miles (0.8 km) east of Wilson, Washington County, Maryland. Coordinates recorded on the label place the quarry at 77º between 49’ + 50’’ W; 39º approximate 41’ N. Based on Google Earth©, there is now a mostly filled-in quarry north of Cresspond Road, North Wilson, NW of Hagerstown, MD, that almost certainly is the Schemeompf’s old Quarry.
The Edinburg Formation is now only used in the Shenandoah Valley in western and northern Virginia. The equivalent unit in western Maryland (Hagerstown Valley) and southern Pennsylvania is now called the Chambersburg Formation. Brezinski et al. (2012) indicates that the Chambersburg Formation belongs to the Amorphognathus tvaerensis Conodont Zone, in the lower and middle Turinian (Laurentian) Stage, belonging to the lower Mohawkian Series, Sandbian (Upper Ordovician). The Chambersburg Formation in Maryland is about 150 m thick and was deposited in a shallow outer ramp with a small amount of deep ramp deposits (sensu Brezinski et al. 2012). These authors also document that the nodular-bedded outer-ramp facies of the lower Chambersburg Formation contains Echinosphaerites beds, showing that the Chambersburg Formation is fossiliferous and favorable for echinoderm preservation, such as the rhipidocystids described herein.
Systematic palaeontology
The classification follows Sprinkle (1973). Morphologic terminology follows Ubaghs (1968) and Sprinkle (1973). Elongate U- or V-shaped plates from the marginal framework are called marginals, and those contributing to the integument, central plates (sensu Sprinkle 1973). Numbering of marginal plates follows Lewis et al. (1987), but this does not necessarily imply homology of named plates among taxa. Dorsal and ventral are used with respect to the position of the substrate, with the marginal plates bearing large projections on the ventral side that contribute to the central part of the theca (see discussion below). Anterior and posterior are used with respect to the position of the oral plates and ambulacra (see below). A–E are used to designate the five ambulacra in pentaradially symmetric echinoderms, with the anterior part indicated by ambulacrum A and the posterior part by interambulacrum CD. Orientation of flattening echinoderms with ambulacral reduction necessitates recognition of oral plates and ambulacra and potential homologies with five-fold echinoderms (see Sumrall 1997; Sumrall et al. 2001; Sumrall and Wray 2007).
Phylum Echinodermata Bruguière, 1791 (ex Klein, 1734)
Subphylum Blastozoa Sprinkle, 1973
Class Eocrinoidea Jaekel, 1918
Order Digitata Hecker, 1938
Family Rhipidocystidae Jaekel, 1901
Genus Durhamicystis nov.
Zoobank LSID: urn:lsid:zoobank.org:act:D0CB18AA-6423-4615-B331-18A8A3C28B36
Type species: Durhamicystis americana sp. nov.; by monotypy, see below.
Etymology: In honour of the paleontological specialist on echinoids and Cambrian echinoderms, John Wyatt Durham (University of California, Berkeley) who examined these specimens while visiting the Smithsonian in 1966–1967, and arranged for their eventual donation.
Diagnosis.—As for the type species by monotypy.
Remarks.—Durhamicystis is assigned to the family Rhipidocystidae based on the elongate and flattened theca, brachioles on both sides of the oral area, and marginal plates. The original description and illustration of Rhipidocystis baltica (type species of the genus) by Jaekel (1901) was very limited including a single drawing that shows an echinoderm with large marginal plates, centrals, and stem facet. The genus was reviewed by Ubaghs (1968) for his chapter on eocrinoids for the Treatise, who figured additional material assigned to Rhipidocystis that was bilaterally symmetrical and showed only two ambulacra with flattening on the A-CD plane. Rozhnov (1989) was first to reconstruct Rhipidocystis in detail showing an animal with a flattened theca and flattening along the BC-DE plane. He also moved the material showing bilateral symmetry described by Ubaghs (1968) to the genus Neorhipidocystis, that also included R. norvegica (Bockelie, 1981) described from a different part of Baltica. The new material described herein is very similar to Neorhipidocystis in the shape of the theca, oral area, split marginal above the periproct, and flattening pattern. If previous observations are correct, then Rhipidocystis includes forms with a very different flattening pattern, and thus the family status should be reconsidered (see discussion below).
Durhamicystis americana sp. nov.
Figs. 1–3, 6D.
Zoobank LSID: urn:lsid:zoobank.org:act:E7B7523F-14C8-4112-946F-A37EC172B26C
Etymology: In reference to North America, the continent where the type species was found.
Type material: Holotype: USNM 642510, a complete specimen preserving the theca, brachioles, and proximal stem (Fig. 1A). Paratypes: USNM 642512, another complete specimen (Fig. 1B), USNM 642511, a small slab containing two well preserved overlapping specimens (Fig. 2); all from type locality and horizon.
Type locality: Schemeompf’s old Quarry, top of hill, west of Conococheague Creek, about 2.1 miles (3.4 km) NNE and 0.5 miles (0.8 km) east of Wilson, Washington Co., Maryland.
Type horizon: Chambersburg Formation, Sandbian, lower Upper Ordovician.
Material.—USNM 642513–642515, fragments and isolated plates; all from type locality and horizon.
Diagnosis.—Rhipidocystid with large theca composed of ten marginal plates with only two basals on the posterior side. Two ambulacra corresponding to BC and DE unsplit rays. Seven large oral plates with brachioles attached either on orals or smaller flooring plates. Brachioles between 22 and 26, biserial, long, slender, and showing a cryptic suture between paired brachiolars. Anal pyramid lateral aligned with the BC ray, hydropore between O1–O7, and gonopore within O1.
Description.—Rhipidocystid with large theca (Fig. 1A1, B1) composed of ten marginal plates and orals closing the marginal framework (see below). Three apertures are recognized that allow orientation of the material. Peristome appears in the central anterior part, bounded by seven oral plates (Fig. 1A2, B1). Two elongate ambulacra corresponding with BC and DE rays. Anal pyramid (Fig. 1B2) covering the periproctal opening (Figs. 1A3, 2A4) appears in the left anterior side of the marginal ring. This is a large aperture of ca. 2 mm (Fig. 1A3) that is covered by a domed anal pyramid composed of about eight?, thin, subtriangular plates (Fig. 1B2). A single small pore (ca. 300 µm) within the plate O1 probably corresponds to the gonopore (Fig. 3B). The suture between O1 and O7 has a slit corresponding with the hydropore (Fig. 3B).
Nine U-shaped marginal plates contribute to the marginal framework on the “ventral” surface closing it (Fig. 1A1), and an extra small plate with a vertical suture lies above the periproct on the “dorsal surface” (Fig. 1A3); thus, most of the marginal framework is singly plated. This framework is closed posteriorly with two large marginal plates (M1, M2), usually considered as basals (Fig. 1A1). Those plates are very large and abut with large sutures closing the marginal frame. They are both trapezoidal in shape and articulate anteriorly with plates M3 and M4, and posteriorly form the stem facet. They both have large, thin projections on the ventral side that contribute to the central part of the theca. Marginal plates M3–M5, and M6 are longer than wide and also have large, thin projections contributing to the central theca. Marginals M4 and M6 are the longest marginals (Fig. 3A2). Marginal plate M7 articulates posteriorly with M5, forms the posterior edge of the periproct, and is overlain by M9 and M10 separated by a vertical suture (Fig. 1A3) at the left anterior corner of the theca, followed anteriorly by oral plates. M8–M10 contribute to the peristomial opening; M8 contributes to the marginal framework in both dorsal and ventral aspects, while M10 and M9 are only visible in dorsal and ventral views, respectively. M9 also articulates anteriorly with an ambulacral flooring plate.
Ventral central plates are large, polygonal, and very thin in cross section (Fig. 3). They are probably four in number and form a tessellate pavement that covers the ventral surface of the theca. Dorsal central plates are more disrupted and only visible in one specimen (Figs. 2A3, 3B); they are apparently smaller than ventral centrals, polygonal, and also very thin in section. They form a tessellate integument that covers the dorsal surface and connects with marginal plates and anterior oral plates.
At least seven oral plates are recognized in the anterior part of the theca (Figs. 1A2, 2A2, 3). Four of them are large plates that articulate aborally with marginal plates M8–M10 and have facets for feeding appendages. O1 and O6 are on the dorsal side of the oral area and have an indented aboral margin to accommodate a small pentagonal O7 plate (Figs. 2A2, 3B2). O3 and O4 are large, relatively thick, subpentagonal elements that appear on the anterior side of the oral area. O2? and O5? are questionably recognized (aborally from the central orals) at the ends of the ambulacra (where branches BC and DE would normally split ontogenetically). They are smaller elements bearing facets for feeding appendages and are visible on both sides of the theca.
Flooring plates appear aborally from the large central oral plates and also have facets for feeding appendages (Figs. 1A2, 2A2, 3). They are difficult to distinguish in size and morphology from oral plates O2? and O5?, except that they do not wrap around the ends of the ambulacra.
Brachioles range from 22 and 26 in number, are biserial, long, and slender (Fig. 2A2). Brachiolar plates appear to be adjacent (not alternating), and have cryptic sutures (Fig. 2A5). They attach either in groups of two (rarely three) on both oral plates and flooring plates. Proximally, the three or four lowest segments of the feeding appendages are wider than long and have deep facets composed of a transverse prominent ridge and three fossa (Fig. 2A4, A5), a large posterior one and two small anterior ones. These create wedge-shaped spaces for ligament attachment, and indicate some mobility. Higher segments are longer than high, tightly sutured, and have a thin, vertical suture visible on the aboral side accentuated by weathering (Fig. 2A4, A5). The adjacent brachiolars here may be partly fused, but still show different crystallographic orientations (see Sprinkle 1973: fig. 5C).
The stem is incomplete in all specimens. Proximally, there is a single large, tapering, subtriangular columnal with a distally positioned facet for one or more additional columnals (Fig. 1B1). Only one? columnal is present in the best preserved specimens. Some disarticulated material preserved in slab USNM 642514 suggests the presence of a normal stem with cylindrical columnals showing crenulae with symplexy articulations in the columnal facets, but this columnal material is not closely associated with any of the complete rhipidocystid specimens.
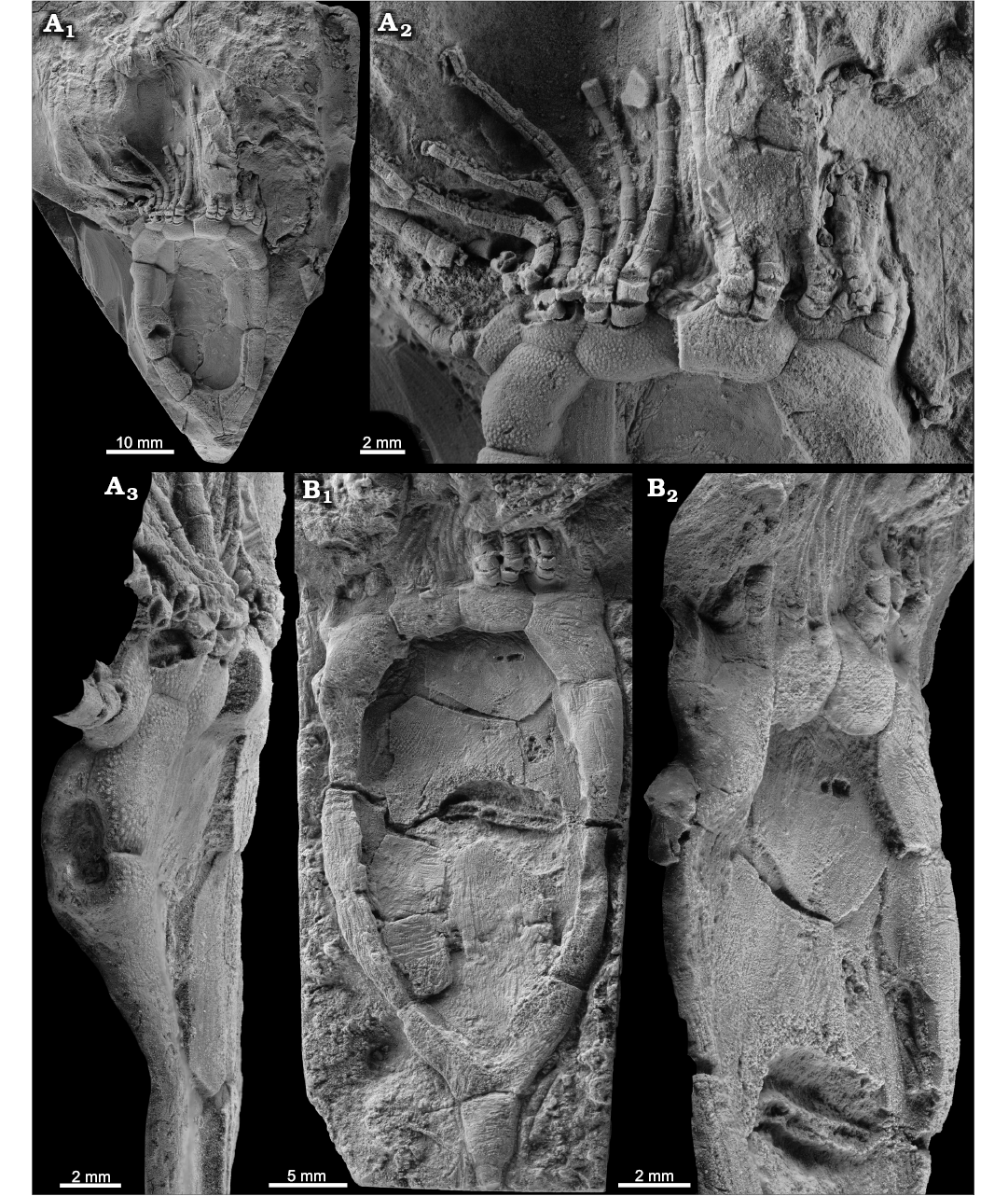
Fig. 1. Rhipidocystid echinoderm Durhamicystis americana gen. et sp. nov. from the Upper Ordovician Chambersburg Formation, Maryland, USA. A. Holotype USNM 642510 showing the theca, proximal stem, and brachioles; general view of the complete specimen in ventral view (A1), detail of oral area and proximal part of the brachioles (A2), specimen in lateral view showing the periproct surrounded by three plates (A3). B. Paratype USNM 642513; general view of the complete specimen in ventral view (B1), note central plates in thecal interior, and single columnal attached to basals; detail of the anal pyramid on left edge (B2). Specimens whitened with ammonium chloride sublimate.
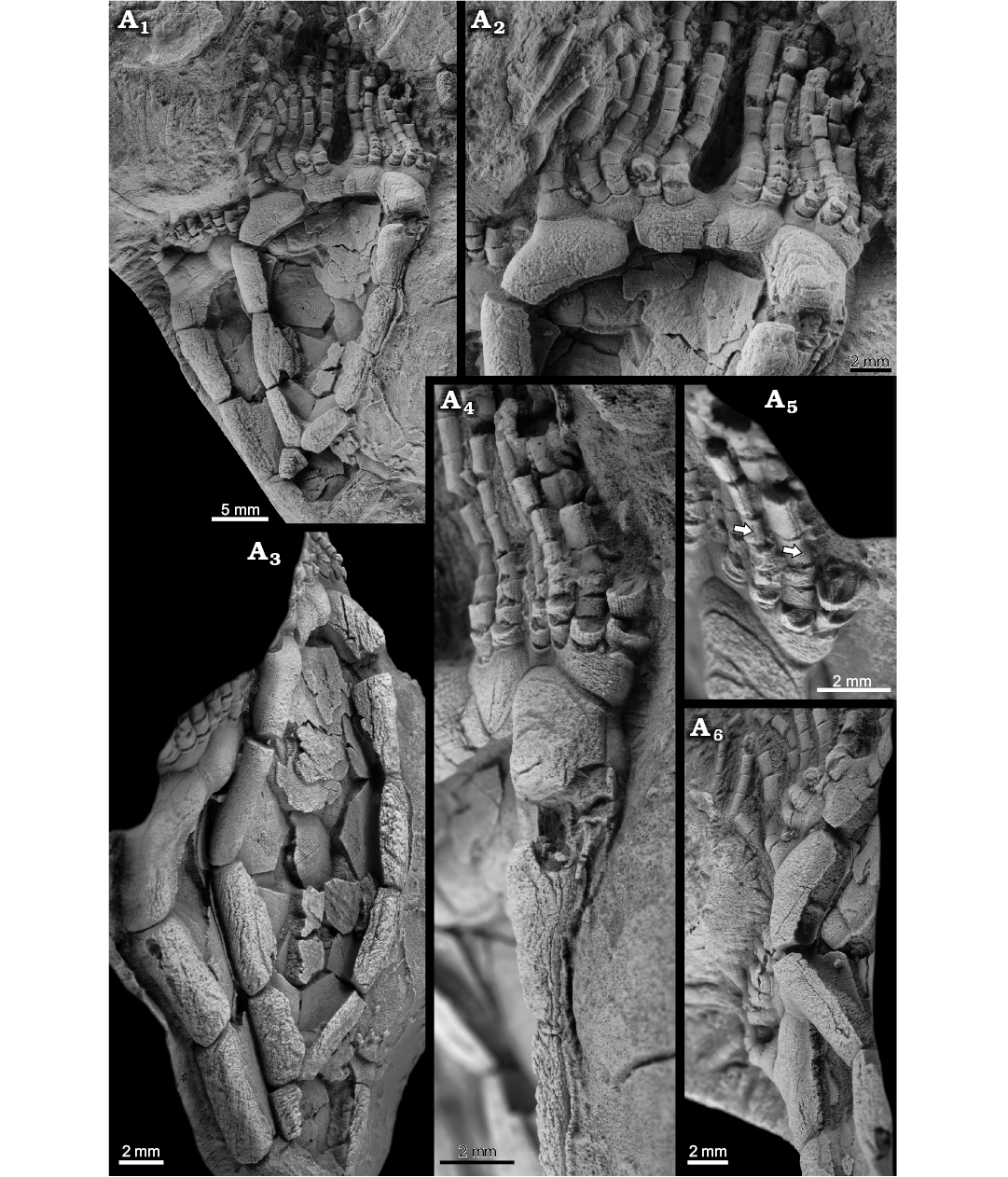
Fig. 2. Rhipidocystid echinoderm Durhamicystis americana gen. et sp. nov. from the Upper Ordovician Chambersburg Formation, Maryland, USA. A. Paratype USNM 642511a, b; general view showing two nearly complete superimposed specimens (A1); detail of the oral area, proximal brachioles attached to marginal, oral, and floor plates, and three apertures (periproct, hydropore slit, and tiny gonopore) (A2); specimen in left lateral view showing the thin central plates from both integuments (A3); specimen in right lateral view showing the position of the periproct (A4); detail of facets between brachiolar plates and cryptic vertical sutures indicated by arrows (A5); detail showing thin internal projections of weathered, U-shaped, marginal plates (A6). Specimens whitened with ammonium chloride sublimate.
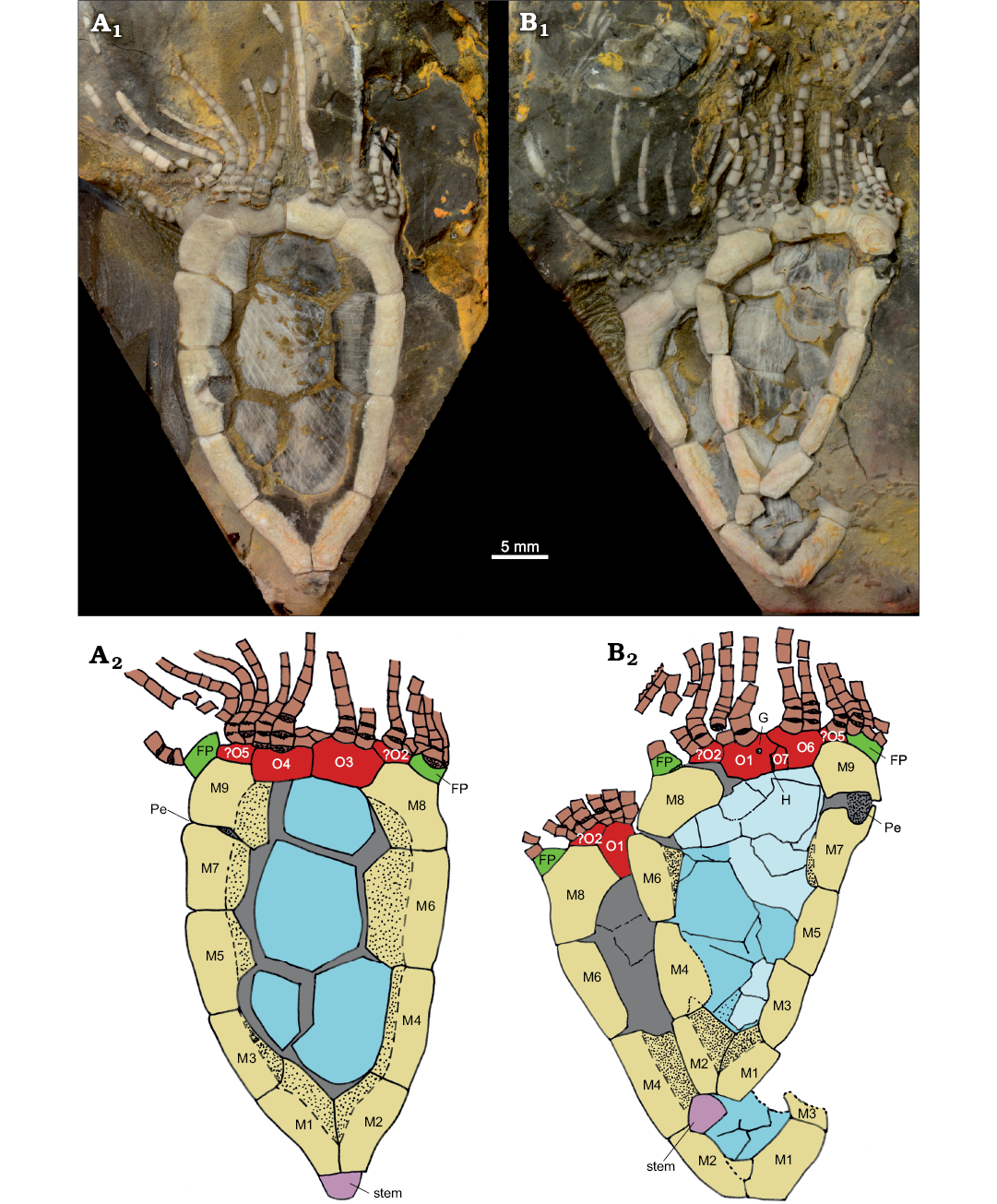
Fig. 3. Rhipidocystid echinoderm Durhamicystis americana gen. et sp. nov. from the Upper Ordovician Chambersburg Formation, Maryland, USA. A. USNM 642510. B. USNM 642511a, b. Photographs of specimens submerged in water to increase contrast of plates and plate boundaries (A1, B1). Camera lucida drawings indicating plate arrangements (A2, B2). Abbreviations: FP, flooring plate (green); G, gonopore; H, hydropore; M1–M9, marginal plates (yellow); O1–O7, oral plates (red); Pe, periproct. Central plates colored in blue, stem in purple, and brachioles in brown.
Remarks.—Durhamicystis americana shows many features that are similar to other members of the family, except for: (i) having only two basals. None of these specimens show M0, or it does not extend out to the margin to form part of the stem facet. This is a difference from the other rhipidocystid genera, with only Neorhipidocystis and few specimens of R. baltica (Sergei Rozhnov, personal communication 2020) also having two basals; (ii) the lack of an elongate M4, only documented in two other genera (Petalocystites and Mandalacystis); (iii) orals (that otherwise look like adoral marginals) supporting the brachioles (usually two of each); (iv) a relatively small stem facet, possibly with normal, thin columnals attached, unlike Batherocystis, which has only a few, elongate, inflated columnals; (v) about 22–26 brachioles present on two elongate ambulacra, probably combined BC and DE; this number of brachioles is intermediate between Mandalacystis (38–44 brachioles) and Petalocystis (16–18 brachioles); the 4 other rhipidocystids all have fewer brachioles; and (vi) 10 marginals total, including a split marginal above the periproct, instead of all marginals being thick, with a U-shaped cross section, and extending onto both sides; Neorhipidocystis is the only other genus that shows a split marginal like this. Also, the brachioles appear to be uniserial, with wedges near their attachment, and with a faint suture line down the back side, indicating that the brachiolars are composites with different crystallography (noted in Sprinkle 1973: fig. 5C). Both the thecal plates and brachioles also show faint to moderate pustules, another typical rhipidocystid feature.
O2? and O5? are questionably recognized in the current material. These elements seem to show on both sides of the theca, but this is difficult to prove based on the current material. Favoring such an interpretation is that aboral (distal) elements from O2? and O5? bearing facets for feeding appendages are single elements on both sides.
Stratigraphic and geographic range.—Chambersburg Formation, Sandbian, lower Upper Ordovician; Maryland, USA.
Discussion
Understanding the orientation of echinoderms that have undergone ambulacral reduction (Sumrall and Wray 2007) is sometimes problematic and rhipidocystids are a clear example. This is exacerbated because flattening in eocrinoids is polyplyletic (Sumrall et al. 2001), and any attempt to recognize possible orientation requires recognition of oral plates, ambulacra, and in some cases thecal openings. Many clades of echinoderms have independently developed flattened theca in response to living in close proximity to the substrate (Sumrall et al. 2001) or as adaptations to high energy environments with oscillatory currents (Nardin 2007a). Flattening in echinoderms is recognized to occur in three planes: lateral where the sides of the theca area are compressed along the BC-DE axis, anterior-posterior where the sides of the theca are compressed along the A-CD axis, and dorsal-ventral where the theca is compressed along the polar axis (Sumrall 1997).
Thecae showing lateral flattening have the BC and DE sides of the theca compressed parallel to the polar axis of the theca. This results in a lens-shaped to discoidal theca that is flattened parallel to the A-CD and polar axes, and perpendicular to the BC-DE axis. This occurs only in Rhipidocystis (sensu Sumrall et al. 2001). Rozhnov (1994) interpreted Rhipidocystis as flattened in the plane of symmetry A-CD, but this plane is unlikely because his drawings (Rozhnov 1994: fig. 2) indicate flattening in the perpendicular orientation (sensu Sumrall 1997). Anterior-posterior flattening occurs where flattening is along the A-CD axis. This results in a lens-shaped to discoidal theca that is flattened parallel to the BC-DE and polar axes and perpendicular to the A-CD axis. This is seen in pleurocystitid glyptocystitoids, the eocrinids Lingulocystis, Cardiocystites, and Haimacystis, and many rhipidocystids such as Neorhipidocystis, Petalocystites, and Durhamicystis. Dorsal-ventral flattening occurs where flattening is along to the polar axis of the theca (Sumrall et al. 2001; Nardin 2007a, b). This is seen in many echinoderms such as asteroids, some echinoids, edrioasteroids, and cyclocystoids (Sumrall 1997).
As with other rhipidocystids, Durhamicystis has a flattened theca and large U-shaped marginal plates. The position of inferred oral plates and openings allows comparison with other pentaradial echinoderms (Kammer et al. 2013), suggesting that ambulacra correspond to the shared BC and DE rays. Two other rhipidocystids have a similar ambulacral pattern; one corresponds to Neorhipidocystis oepiki (Hecker, 1938), that was re-described by Rozhnov (1989) to accommodate material included by Hecker (1938) within Rhipidocystis. A second species of Neorhipidocystis was described by Bockelie (1981) and originally included in R. norvegica (Figs. 4, 5), later transferred to Neorhipidocystis by Rozhnov (1989). Both Bockelie (1981) and Rozhnov (1989) suggested that the periproct in Neorhipidocystis was located on the left edge of the theca (in the DE interray based on our interpretation), opening through the marginal plate (Fig. 6C), but this position is unlikely based on the observations in Durhamicystis (Fig. 6D) in which the periproct opens in the opposite position, between plates M7 and M9, likely coinciding with the BC interray. This suggests the periproct migrated from its original position within the CD interlay to the BC. Mandalacystis (Fig. 6A)—one of the best known rhipidocystids, was described from abundant material (Lewis et al. 1987). Four ambulacra in an elongate, moderately tilted array were proposed: possibly B (short), C (long and recurved), D (medium), and E (medium), with nearly all brachioles attached on the aboral side of each ambulacrum. Unfortunately, the oral area in Mandalacystis cannot be resolved because extensive brachioles cover the underlying ambulacra, mouth, and surrounding oral plates, and thus comparison to Durhamicystis is difficult. Material of Rhipidocystis described by Rozhnov (1989) preserves the oral area and main apertures. The raised, circular, oral area is composed of five short ambulacra with the anal pyramid located between orals O1 and O7 on one edge of the theca. If correct, this orientation corresponds to a lateral flattening pattern through the BC-DE plane, and therefore, at a right angle to what is observed in Durhamicystis and Neorhipidocystis. Contrary to our interpretation (see above), Rozhnov (1994) suggested the theca of Rhipidocystis is flattened in the plane of symmetry A-CD, but this situation is inconsistent with the position of periproct, gonopore, and hydropore, that are situated in the CD interray in most early pentaradial echinoderms, and coincides with their position in Rhipidocystis (sensu Rozhnov 1994: fig. 2).
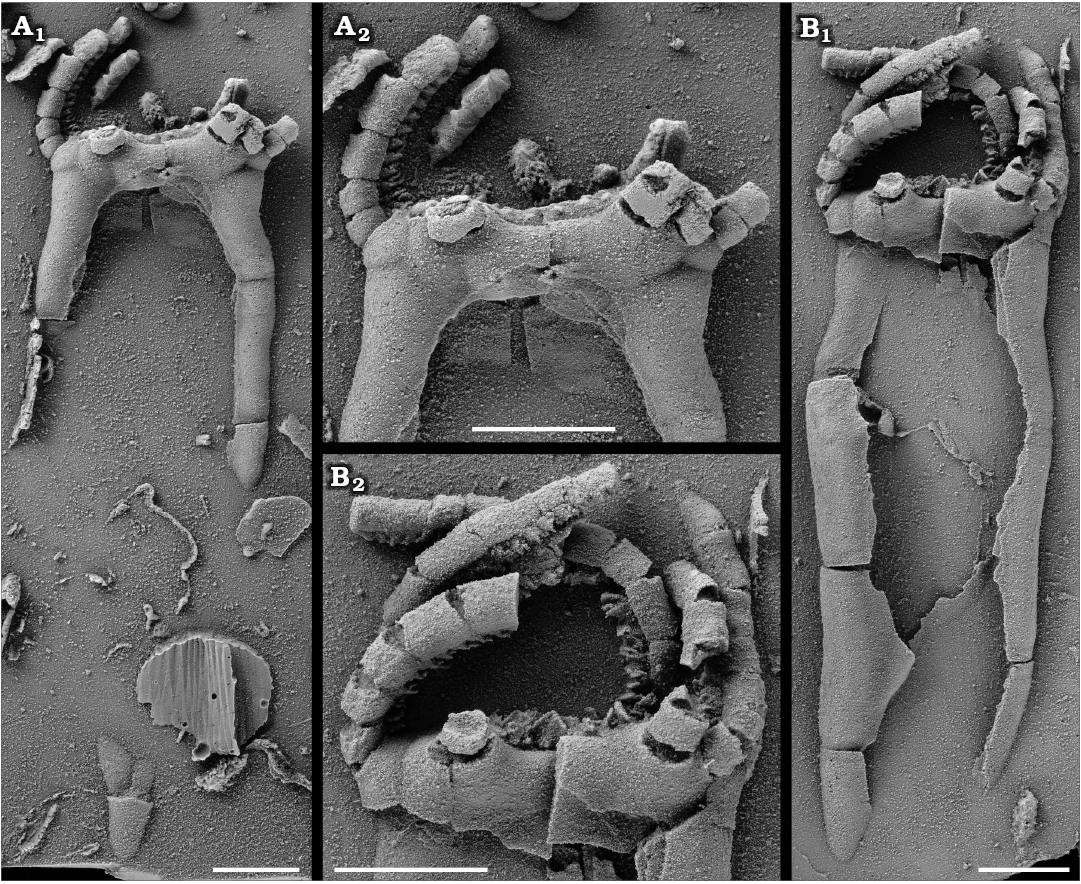
Fig. 4. Rhipidocystid echinoderm Neorhipidocystis norvegica (Bockelie, 1981) from the Late Ordovician of the Oslo Region, Norway. A. PMO 101.143, half complete specimen; detail of the oral area (A2). B. PMO 101.144 (holotype), almost complete specimen lacking the lower portion; detail of the oral area (B2), note V-shaped notches and cover plates on uniserial brachioles. Specimens are latex casts whitened with ammonium chloride sublimate. Scale bars 5 mm.
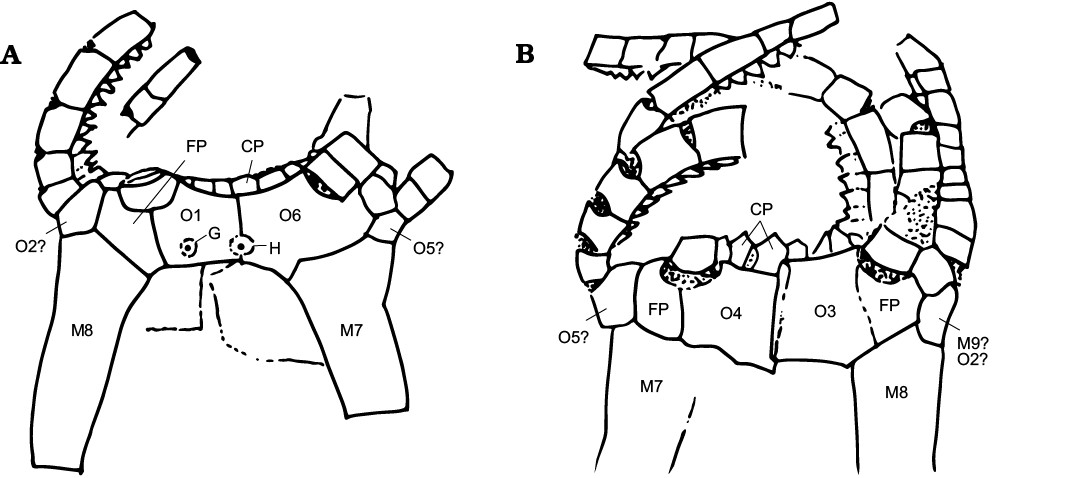
Fig. 5. Camera lucida drawings and plate interpretations of the oral area in Neorhipidocystis norvegica (Bockelie, 1981). A. PMO 101.143. B. PMO 101.144, holotype. Abbreviations: CP, cover plates over ambulacra and brachioles; FP, flooring plates; G, gonopore; H, hydropore; M, marginal plates; O1–O6, oral plates.
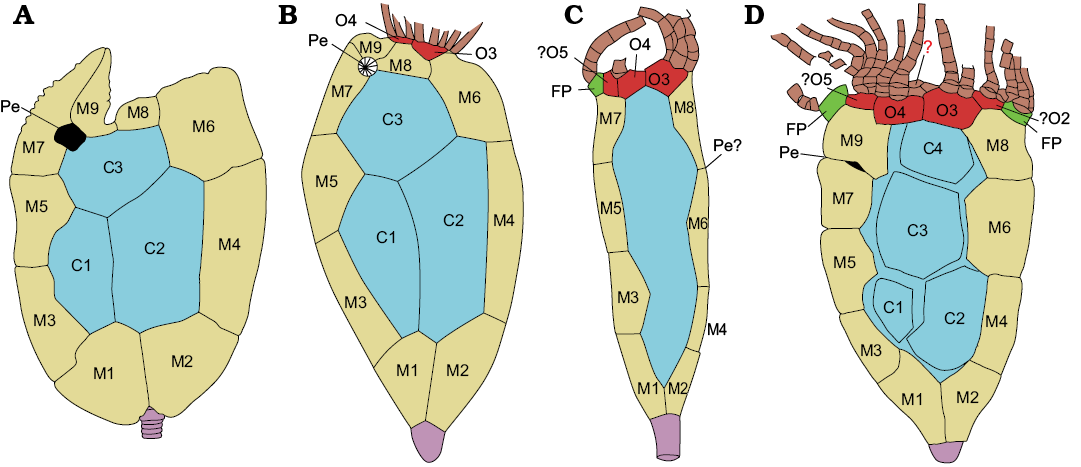
Fig. 6. Comparisson between the rhipidocystids Mandalacystis dockery Lewis, Sprinkle, Bailey, Moffit, and Parsley, 1987 (A), Petalocystites ikecanensis (Sprinkle, 1973) (B), Neorhipidocystis norvegica (Bockelie, 1981) (C), and Durhamicystis americana gen. et sp. nov. (D). Abbreviations: FP, flooring plate (green); M1–M9, marginal plates (yellow); O1–O7, oral plates (red); central plates (blue); stem (purple), brachioles (brown); Pe, periproct. Based on Lewis et al. 1987 (A), Sprinkle 1973 (B), and Bockelie 1981 (C).
Other echinoderms experimented with the reduction of ambulacra to only the BC-DE rays associated with anterior-posterior thecal flattening. The earliest eocrinoid showing such a reduction is Ubaghsicystis segurae from the Cambrian Series of Spain (Gil Cid and Domínguez Alonso 2002). This species shows two large feeding appendages extending from O1 and O6, and the anal pyramid is positioned on the right side. Hydropore-gonopore is interpreted to be between the O1 and O6. Lingulocystids (i.e., Lingulocystis) also have the shared ambulacra BC and DE, with the anal pyramid in the upper third of the theca, opening through the integument just inside the marginals (Ubaghs 1968: fig. 320). Some of the marginal plates along the adoral side also have facets for feeding appendages (Ubaghs 1968; Nardin 2007b), similar to what is observed in Durhamicystis, and might be interpreted as oral plates. Pleurocystitids are derived glyptocystitid rhombiferans with shared BC-DE ambulacra (Sumrall and Sprinkle 1995). In this group stout brachioles arise from the shared ambulacra with anterior-posterior flattening of the theca, and the bulk of the ventral surface is occupied by a greatly enlarged periproctal membrane. In Haimacystis the ambulacra are in the form of a curved C-shaped filled groove around a centrally positioned peristome. The periproct is centered below in the CD interray but the hydropore and gonopore have not been identified. Brachioles arise along the anterior margin of the food groove, but the details are not presently clear (Sumrall et al. 2001). In Cardiocystites the theca is flattened but the orientation of the flattening is doubtful as there is no evidence for the hydropore and gonopore. Five, short, erect ambulacra bearing brachioles arise from the oral area (Dean and Smith 1998). The anal pyramid is located at the junction between the most adoral marginal plate and the adoral mound, suggesting anterior-posterior flattening (Nardin 2007a).
Rhipidocystids have been argued to be related to paracrinoids based on similar peristomial plate arrangement, three distinctive basal plates, position of main thecal apertures (gonopore, hydropore, and periproct), and proximal stem morphology (Sumrall 1997; Sprinkle and Rushlau 2016; Sprinkle and Guensburg 2017). Most highly flattened rhipidocystids with U-shaped marginals, and most lens-shaped paracrinoids with 2–3 ambulacra, appear to be flattened into the anterior-posterior plane (through the A-CD), although there is considerable variability in where the periproct is located and what ambulacra are retained (usually BC and DE). At least three taxa, Petalocystites, Neorhipidocystis, and Durhamicystis agree with this configuration, and thecal plates can be homologized across the three taxa (Fig. 6). Rhipidocystis, that gives the name to the family Rhipidocystidae, is apparently flattened in the opposite direction (see discussion above). If confirmed this probably responds to different flattening planes in Rhipidocystidae, and thus the family, as presently described, should be reconsidered.
Conclusions
Rhipidocystids include some of the most enigmatic echinoderm taxa from the lower Palaeozoic. Their fattened body and aberrant morphology, with little morphological information from the oral area, limited in part the understanding of homologies across different taxa. The discovery and description of Durhamicystis americana gen. et sp. nov. from the Upper Ordovician Chambersburg Formation in western Maryland (USA), preserving both thecal sides and the oral area, and major apertures including peristome, periproct, gonopore, and hydropore, allows orientation of the flattening this species and proper comparison across all rhipidocystids. This questioned previous hypothesis about homology within rhipidocystids, and suggests that two different groups, one including Rhipidocystis and the other comprising Petalocystites, Neorhipidocystis, and Durhamicystis, show flattening in opposite directions.
Acknowledgments
We thank Mark Florence (USNM) for help searching historical information about specimens, and Jessica K. Nakano and Kathy Hollis (both USNM), for arranging the loan of specimens. Franz-Josef Lindemann (Paleontologisk museum, Oslo, Norway) arranged the loan of latex casts of Neorhipidocystis norvegica. Sergey Rozhnov (Paleontological Institute, Moscow, Russia), Martina Nohejlová (Czech Geological Survey, Prague, Czech Republic), and Ronald Parsley (Tulane University, Louisiana, USA), reviewed and improved an earlier version of this manuscript. This work was supported by the Spanish Ministry of Science, Innovation and Universities (CGL2017-87631) and co-financed by the European Regional Development Fund. SZ was funded by project “Aragosaurus: recursos geológicos y paleoambientales” (ref: E18_17R ) funded by the Government of Aragón. The JS visits to the Smithsonian Institution in 1966 and 1967 were funded by an NSF Graduate Fellowship through Harvard University.
References
Bassler, R.S. 1950. New genera of Middle Ordovician “Cystidae”. Journal of the Washington Academy of Sciences 40: 273–277.
Bockelie, J.F. 1981. The Middle Ordovician of the Oslo region, Norway. The eocrinoid genera Cryptocrinites, Rhipidocystis and Bockia. Norsk Geologisk Tidsskrift 61: 123–147.
Brezinski, D.K., Taylor, J.F., and Repetski, J.E. 2012. Sequential development of platform to off-platform facies of the Great American Carbonate Bank in the central Appalachians. In: Derby, J.R., Fritz, R.D., Longacre, S.A., Morgan, W.A., and Sternbach, C.A. (eds.), The Great American Carbonate Bank: The Geology and Economic Resources of the Cambrian–Ordovician Sauk Megasequence of Laurentian. AAPG Memoir 98: 383–420.
Bruguière, J.G. 1791. Tableau Encyclopédique et Méthodique des Trois Règnes de la Nature, contenant l’Helminthologie, ou les Vers Infusoires, les Vers Intestins, les Vers Mollusques, etc. Volume 7. 180 pp. Panckoucke, Paris. Crossref
Dean, J. and Smith, A.B. 1998. Palaeobiology of the primitive Ordovician pelmatozoan echinoderm Cardiocystites. Palaeontology 41: 1183–1194.
Gil Cid, M.D. and Domínguez Alonso, P. 2002. Ubaghsicystis segurae nov. gen. y sp., nuevo Eocrinoide (Echinodermata) del Cámbrico Medio del Norte de España. Coloquios de Paleontología 53: 21–32
Hecker, R.F. 1938. New data on Rhipidocystis Jkl. (order Digitata n.o., class Carpoidea Jkl.) and on a new genus Bockia (subclass Eocrinoidea Jkl., class Crinoidea Mill.) from the Ordovician of Leningrad Province, U.S.S.R., and Estonia [in Russian]. Doklady Akademii Nauk SSSR 19 (5): 421–425.
Hecker, R.F. [Gekker, R.F.] 1940. Carpoidea, Eocrinoidea and Ophiocistia from the Ordovician of the Leningrad region and Estonia [in Russian with German summary]. Trudy Paleontologičeskogo Instituta Akademii Nauk SSSR 9: 5–82.
Jaekel, O. 1901. Uber Carpoideen, eine neue Klasse von Pelmatozoen. Deutsche Geologische Gesellschaft, Zeitschrift 52: 661–677.
Jaekel, O. 1918. Phylogenie und System der Pelmatozoen. Paläontologische Zeitschrift 3: 1–128. Crossref
Kammer, T., Sumrall, C.D., Zamora, S., Ausich, W.A., and Deline, B. 2013. Oral region homologies in Paleozoic crinoids and other plesiomorphic pentaradial echinoderms. PLoSOne 8 (11): e77989. Crossref
Lewis, R.D., Sprinkle, J., Bailey, J.B., Moffit, J., and Parsley, R.L. 1987. Mandalacystis, a new rhipidocystid eocrinoid from the Whiterockian Stage (Ordovician) in Oklahoma and Nevada. Journal of Paleontology 61: 1222–1235. Crossref
Nardin, E. 2007a. New occurrence of the Ordovician eocrinoid Cardiocystites: Palaeogeographical and palaeoecological implications. Acta Palaeontologica Polonica 52: 17–26.
Nardin, E. 2007b. New specimens of Lingulocystis Thoral, 1935 (Eocrinoidea, Blastozoa) from the Arenig (Lower Ordovician) of Montagne noire (Southern France): Intraspecific morphological variability, stratigraphic, and palaeoecological implications. Annales de Paléontologie 93: 199–214. Crossref
Rozhnov, S.V. [Rožnov, S.V.] 1989. New data on rhipidocystids (Eocrinoidea) [in Russian]. In: D.L. Kal’o (ed.), Problemy izučâ iskopaemyh i sovremennyh iglokožih, 38–57. Academy of Sciences of the Estonian SSR Institute of Geology, Tallinn.
Rozhnov, S.V. 1994. Comparative morphology of Rhipidocystis Jaekel, 1900 and Cryptocrinites von Buch, 1840 (Eocrinoidea, Ordovician). In: B. David, A. Guille, J.-P. Féral, and M. Roux (eds.), Echinoderms through Time: Proceedings of 8th International Echinoderm Conference Dijon France, 173–178. Balkema, Rotterdam.
Sprinkle, J. 1973. Morphology and evolution of blastozoan echinoderms. Harvard University Museum of Comparative Zoology, Special Publication. 283 pp. Cambridge. Crossref
Sprinkle, J. and Guensburg, T.E. 2017. The convoluted history of paracrinoids and rhipidocystids (blastozoan echinoderms) during the Ordovician Radiation. Geological Society of America Abstracts with Programs 49(6) [published online https://doi.org/10.1130/abs2017AM-302126]. Crossref
Sprinkle, J. and Rushlau, W. 2016. New flattened, asymmetrical, rhipidocystid echinoderm from the Bromide Formation (Sandbian) of southern Oklahoma, and a revised classification of eocrinoids, rhipidocystids, and paracrinoids. Geological Society of America Abstracts with Programs 48 (7) [published online, https://doi.org/10.1130/abs/2016AM-283419]. Crossref
Sumrall, C.D. 1997. The role of fossils in the phylogenetic reconstruction of Echinodermata. In: J.A. Waters and C.G. Maples (eds.), Paleontological Society Papers Volume 3, Geobiology of Echinoderms, 267–288. The Paleontological Society, Pittsburgh. Crossref
Sumrall, C.D. 2010. A model for elemental homology for the peristome and ambulacra in blastozoan echinoderms. In: L.G. Harris, S.A. Böttger, C.W. Walker, and M.P. Lesser (eds.), Proceedings of the 12th International Echinoderms Conference Echinoderms:Durham, 269–276. CRC Publications, Taylor & Francis Group, London. Crossref
Sumrall, C.D. and Sprinkle, J. 1995. Plating and pectinirhombs of the Ordovician rhombiferan Plethoschisma. Journal of Paleontology 69: 772–778. Crossref
Sumrall, C.D. and Wray, G.A. 2007. Ontogeny in the fossil record: diversification of body plans and the evolution of “aberrant” symmetry in Paleozoic echinoderms. Paleobiology 33: 149–163. Crossref
Sumrall, C.D., Sprinkle, J., and Guensburg, T.E. 2001. Comparison of flattened blastozoan echinoderms: insights from the new Early Ordovician eocrinoid Haimacystis rozhnovi. Journal of Paleontology 75: 985–992. Crossref
Ubaghs, G. 1968. Eocrinoidea. In: R.C. Moore (ed.), Treatise on Invertebrate Paleontology. Part S. Echinodermata 1 (2), S455–S495. Geological Society of America, New York and University of Kansas, Lawrence.
Zamora, S. and Rahman, I. 2014. Deciphering the early evolution of echinoderms with Cambrian fossils. Palaeontology 57: 1105–1119. Crossref
Acta Palaeontol. Pol. 65 (3): 455–465,
2020 https://doi.org/10.4202/app.00718.2019