
New diplodocoid sauropod dinosaur material from the Middle Jurassic of European Russia
ALEXANDER O. AVERIANOV and NIKOLAY G. ZVERKOV
Averianov, A.O. and Zverkov, N.G. 2020. New diplodocoid sauropod dinosaur material from the Middle Jurassic of European Russia. Acta Palaeontologica Polonica 65 (3): 499–509.
Two anterior caudal vertebrae from apparently a single individual from the Callovian (Middle Jurassic) marine deposits of the Podosinki Formation at Peski Quarry near Moscow, Russia, are attributed by the phylogenetic analyses of two independent datasets to the sauropod clade Diplodocoidea. Morphologically, these vertebrae differ from the anterior caudals of Diplodocidae and Rebbachisauridae, but are similar to those of Dicraeosauridae. This finding is in line with a recent discovery of the dicraeosaurid sauropod Lingwulong from the late Early–early Middle Jurassic of China, and suggests earlier diversification of the main neosauropod lineages and Asiatic origin for Dicraeosauridae. The Peski sauropod is the first Jurassic terrestrial vertebrate from the Fennoscandian landmass. Diplodocoid sauropods likely dispersed to this land massive before the Bathonian, when it was still connected with Asia.
Key words: Dinosauria, Sauropoda, Diplodocoidea, Mesozoic, Russia, Fennoscandian landmass.
Alexander O. Averianov [dzharakuduk@mail.ru], Zoological Institute of the Russian Academy of Sciences, Universitetskaya Emb. 1, 199034 Saint Petersburg, Russia.
Nikolay G. Zverkov [zverkovnik@mail.ru], Borissiak Paleontological Institute of the Russian Academy of Sciences, Profsouznaya Str. 123, 117997 Moscow, Russia; Geological Institute of the Russian Academy of Sciences, Pyzhevsky lane 7, 119017 Moscow, Russia.
Received 10 January 2020, accepted 9 March 2020, available online 13 May 2020.
Copyright © 2020 A.O. Averianov and N.G. Zverkov. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
In the Jurassic and Cretaceous, most of European Russia was covered by epicontinental seas. Therefore, the remains of continental vertebrates of this time are extremely rare in this territory and are usually found in marine deposits. The sole exception is a vertebrate locality at the Peski Quarry in Moscow Region, where vertebrate remains are found in palaeocarst cavities filled with fluvial clays which are late Bathonian in age (Fig. 1; Alekseev et al. 2001). This locality has produced remains of chimaeriformes, hybodontiformes, various actinopterygian fishes, temnospondyl and caudate amphibians, archaic aquatic turtles (Heckerochelys romani; Sukhanov 2006), choristoderes, neosuchian crocodyliforms, theropod dinosaurs, and morganucodontans (Krupina 1995; Alekseev et al. 2001; Alifanov and Sennikov 2001; Gambaryan and Averianov 2001; Bragina 2005; Sukhanov 2006; Popov and Shapovalov 2007; Pashchenko et al. 2018). The theropod teeth from this locality were hitherto the only remains of Jurassic dinosaurs from the European Russia. Here we report on the first discovery of sauropod dinosaur remains from the Middle Jurassic of European Russia. These were also found in the Peski Quarry, but they origin from a stratigraphically higher horizon, composed of the marine deposits of the Podosinki Formation, which are dated as upper Callovian to lower Oxfordian in age (Fig. 1; Tesakova 2003). The specimens were collected in 1997 by Alexander B. Vidryk from the upper part of an oolite marl bed in the upper Callovian part of the section corresponding to the Quenstedtoceras lamberti Ammonite Biozone (Fig. 1C). The specimens are two anterior caudal vertebrae which were collected in association and thus believed to be from a single individual. Both vertebrae are covered in ostreoid bivalves and serpulids (Halcobeloides sp., Serpula sp., Propomatoceros lumbricalis Schlotheim, 1820, “Filogranula” runcinata Sowerby, 1829, and Spiraserpula oligospiralis Ippolitov, 2007; all identifications were made by Alexey P. Ippolitov, Geological Institute, Russian Academy of Sciences, Moscow), which are embedded on the surface of the bone (see Figs. 2, 3), demonstrating long-term exposure above the sediment-water interface and epibiont activity. Many of these epibiont remains, as well as most of the covering rock were removed during the additional mechanical and acid preparation performed for this study by NGZ. These vertebrae were previously erroneously attributed to a gigantic pliosaurid plesiosaur (Zverkov et al. 2017: fig. 2). The Middle Jurassic sauropod vertebrae from the Peski locality belong to one of the earliest known diplodocoids and contribute significantly to our understanding of the initial radiation of Diplodocoidea.
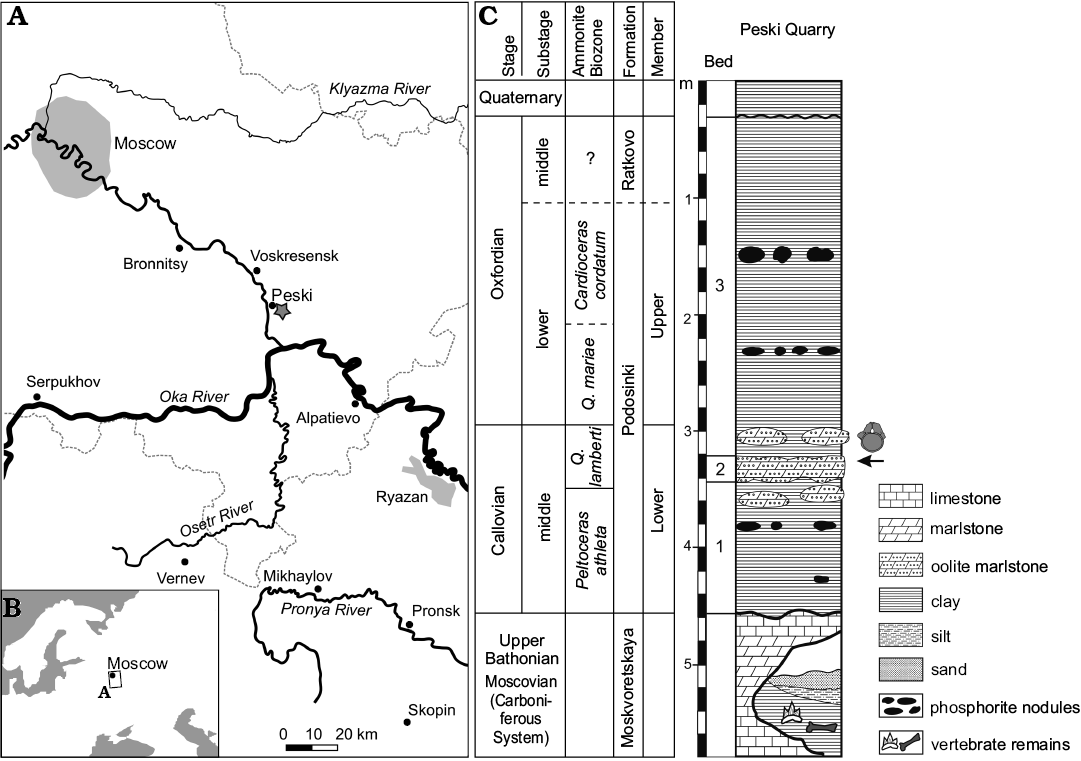
Fig. 1. A. Geographic location of the Peski locality (star). B. Map of eastern Europe with position of the studied area. C. Stratigraphic column of the Peski Quarry with position of sauropod vertebrae indicated (based on Alekseev et al. 2001 and Tesakova 2003). Abbreviation: Q., Quenstedtoceras.
Institutional abbreviation.—MCEBC, Museum of Natural History at Moscow Children’s Ecological and Biological Center, Moscow, Russia.
Other abbreviations.—ACH, anterior centrum height; ACW, anterior centrum width; CL, centrum length (with condyle); cprf, centroprezygapophyseal fossa; PCH, posterior centrum height; PCW, posterior centrum width; prdl, prezygodiapophyseal lamina; prcdf, bordering posteriorly the prezygapophyseal centrodiapophyseal fossa.
Systematic palaeontology
Sauropoda Marsh, 1878
Diplodocoidea Marsh, 1884
Diplodocoidea indet.
Figs. 2, 3.
Material.—MCEBC 1100300/216 and 1100300/215, two anterior caudal vertebrae, which likely belong to a single individual, from the upper Callovian Quenstedtoceras lamberti Ammonite Biozone of the Podosinki Formation; Peski Quarry, Moscow Region.
Measurements (in mm).—MCEBC 1100300/216 and 1100300/ 215, respectively: CL 105, 110; ACW 200, 166; ACH 171, 166; PCW 193, 156; PCH 165, 157.
Description.—The material described herein consists of two anterior caudal vertebrae, which likely belong to a single individual. MCEBC 1100300/216 and 1100300/215 (Figs. 2 and 3). MCEBC 1100300/216 is more anterior in serial position because it has wider and taller but shorter centrum. By size and relative development of the transverse process both vertebrae are close in position and likely were separated by not more than one or two vertebrae.
The centrum is weakly procoelous, with slightly concave anterior and slightly convex posterior articular surface. The centrum articular surfaces are round to heart-shaped anteriorly and round posteriorly. The dorsal margin of the articular surface is depressed under the neural canal, and this depression is more pronounced posteriorly. The centrum is anteroposteriorly short, with centrum length to posterior centrum width ratio of 0.55 (MCEBC 1100300/216) and 0.71 (MCEBC 1100300/215). In lateral view, the ventral profile of the centrum is shallowly concave. The posterior articular surface extends more ventrally compared with the anterior articular surface. The ventral centrum surface is transversely rounded, without a ventral ridge or groove. There are weakly defined lateral ridges on the centrum about one third of the centrum height above the ventral margin. The centrum surface is slightly depressed between these ridges and the ventral centrum margin. The chevron facets are developed only along the posterior margin of the centrum. These facets are very small and widely separated on the more anterior vertebra (MCEBC 1100300/216), while better developed and adjacent on the more posterior vertebra (MCEBC 1100300/215).
The vertebrae may belong to an immature individual based on the presence of suturae between the caudal rib, centrum, and neural arch (MCEBC 1100300/216; Fig. 2A2), or between the centrum and neural arch (MCEBC 1100300/215; Fig. 3A1–A3). In MCEBC 1100300/216 the caudal rib is synostosed to the dorsal third of the centrum in lateral view, with minor contribution to the neural arch. In MCEBC 1100300/215, the caudal rib is completely fused with the centrum and is placed at the dorsal margin of the centrum. In this specimen the caudal rib does not contact the neural arch, and the caudal rib is oriented laterally. In lateral view, the caudal rib is positioned closer to the anterior articular surface, especially in MCEBC 1100300/216. In this specimen, the cross-section of the caudal rib is pentagonal (better seen right laterally) and there is a ridge extending between the dorsal margin of the caudal rib and the neural arch. In MCEBC 1100300/215, the caudal rib is compressed dorsoventrally. In MCEBC 1100300/216, the caudal rib is connected to the neural arch by a tall thin plate, which is a characteristic of the first caudal in many sauropods (Upchurch 1998). This supports a serial referral for MCEBC 1100300/216 as the first caudal.
The neural arch of MCEBC 1100300/216 is incompletely preserved. The neurocentral suture is visible and located slightly dorsal to the bottom of the neural canal, which is composed by the centrum. The neural canal is completely preserved in MCEBC 1100300/216 while in MCEBC 1100300/215 its dorsal margin is missing. The neural canal is relatively small (approximately one quarter of the centrum dorsoventral height), and ovoid in outline, with a pointed dorsal apex. It is slightly larger anteriorly. In lateral view, the peduncle of the neural arch occupies the whole space between the anterior and posterior articular surfaces of the centrum and narrows dorsally. The midline of the neural arch is close to the anterior centrum articular surface. The anterior border of the neural arch is placed almost on the anterior border of the centrum. In MCEBC 1100300/215 there is a poorly defined prezygodiapophyseal lamina (prdl) bordering posteriorly the prezygapophyseal centrodiapophyseal fossa (prcdf) (Fig. 3A3, A4). In MCEBC 1100300/216, the prezygapophyses are partially preserved. The incomplete prezygapophyses are closely spaced laterally, and coalesced ventrally. The prezygapophyseal articular surfaces are convex and oriented at an angle of 28° to the vertical axis. The prezygapophyses are dorsally directed and do not project beyond the anterior end of the centrum. The articular surfaces are predominantly medially oriented. There is a prominent dorsoventrally short ventral strut between the prezygapohyses and neural canal. The height of this strut is less than the height of the neural canal. Lateral to the prezygapophysis and the neural canal there is a very shallow prezygapophyseal centrodiapophyseal fossa.
On the posterior side of MCEBC 1100300/216 the postzygapophyses are not preserved but there is a short thickened horizontal lamina which is likely the infrapostzygapophyseal lamina between the postzygapophyses. There is a prominent, posteriorly extending hyposphenial ridge along the midline between the neural canal and the infrapostzygapophyseal lamina. Its height is greater than the height of the neural canal.
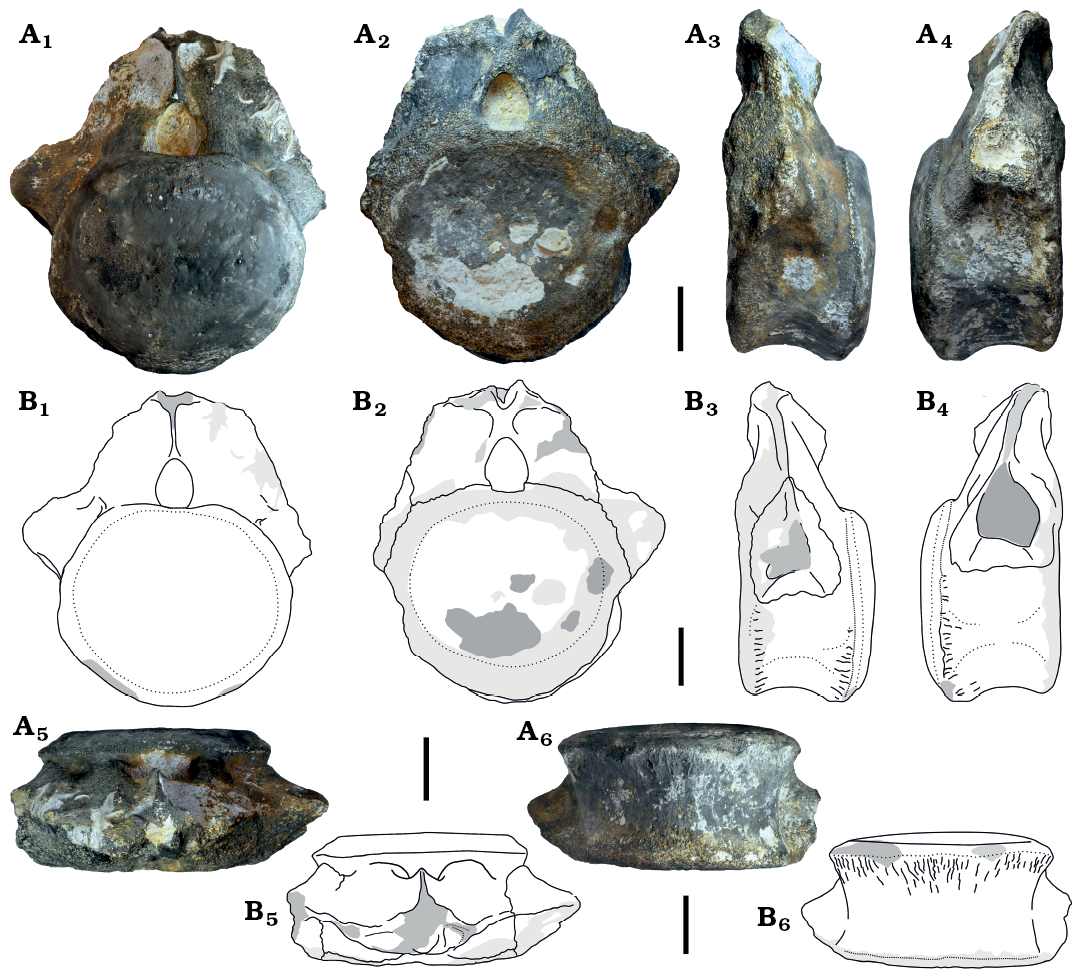
Fig. 2. Diplodocoidea indet. (MCEBC 1100300/216) from Peski, Moscow Province, Russia, Podosinki Formation, Callovian (Middle Jurassic); anterior (likely first) caudal vertebra in posterior (A1, B1), anterior (A2, B2), left lateral (A3, B3), right lateral (A4, B4), dorsal (A5, B5), and ventral (A6, B6) views. Photographs (A) and interpretative drawings (B). Dark grey, broken area; lighter grey, eroded surface, the latest lightest grey, matrix. Scale bars 50 mm.
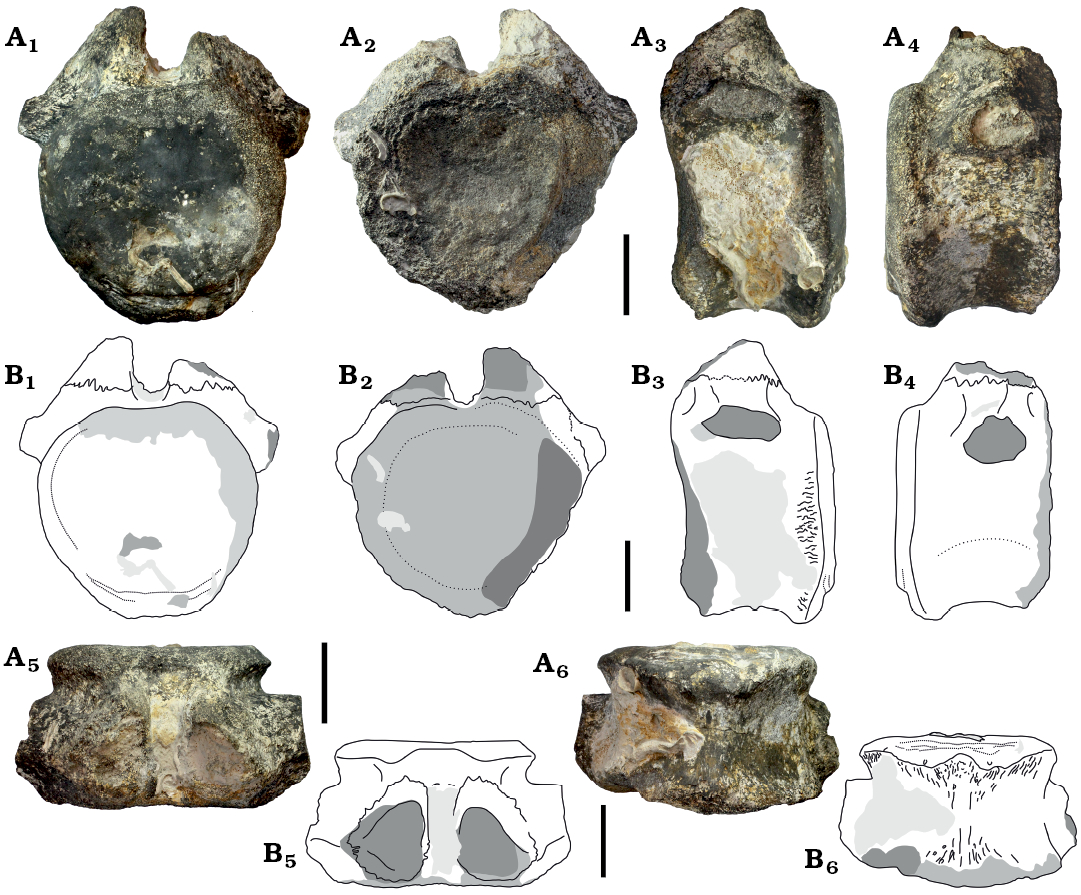
Fig. 3. Diplodocoidea indet. (MCEBC 1100300/215) from Peski, Moscow Province, Russia, Podosinki Formation, Callovian (Middle Jurassic); anterior caudal vertebra, in posterior (A1, B1), anterior (A2, B2), left lateral (A3, B3), right lateral (A4, B4), dorsal (A5, B5), and ventral (A6, B6) views. Photographs (A) and interpretative drawings (B). Dark grey, broken area; lighter grey, eroded surface, the lightest grey, matrix. Scale bars 50 mm.
Remarks.—The anterior caudals of the dicraeosaurid Lingwulong shenqi from the Toarcian–Bajocian Yanan Formation of Ningxia Hui Autonomous Region, China (Xu et al. 2018: supplementary fig. 7a–d), have moderately short centrum with deeply concave ventral margin in lateral view and anterior articulation surface projecting distinctly more ventrally compared with the posterior articulation surface. In the Peski sauropod, the ventral centrum margin is only shallowly concave in lateral view and the posterior centrum articulation surface is slightly more ventrally projecting. The anterior caudals of Lingwulong are slightly amphicoelous, while they are slightly procoelous in the Peski sauropod. In the first caudal of Lingwulong, the prezygapophyses are widely separated by a deep centroprezygapophyseal fossa (cprf), which is placed above the neural canal and between the centroprezygapophyseal laminae (Xu et al. 2018: supplementary fig. 7b). While the prezygapophyses face more dorsally than medially. However, in a more posterior anterior caudal (Xu et al. 2018: supplementary fig. 7d) the cprf is lacking and prezygapophyses are more closely spaced and face medially. The latter construction is more consistent with the condition present by MCEBC 1100300/216. The caudal rib is wing-like in the first caudal of Lingwulong, but dorsoventrally compressed and directing laterally and slightly ventrally in a more posterior anterior caudal. In the Peski sauropod, the caudal rib is wing-like in MCEBC 1100300/216 and less dorsoventrally compressed and oriented dorsolaterally in MCEBC 1100300/215.
The first caudal vertebra of the putative diplodocoid Cetiosauriscus stewarti from the Callovian Oxford Clay Formation of England (Woodward 1905: fig. 41; Charig 1980) is distinctly anteroposteriorly shorter than the first caudal of the Peski sauropod; it has a centrum length to centrum width ratio of 0.38 compared to 0.55 in MCEBC 1100300/216. It differs also with a larger neural canal, well separated prezygapophyses, anterior centrum articulation surface with overhanging dorsal margin, and anterior centrum articulation surface projecting more ventrally compared with the posterior centrum articulation surface (the opposite is true for MCEBC 1100300/216). The first caudal of Cetiosauriscus is similar with MCEBC 1100300/216 in having a convex posterior centrum articulation surface.
The first caudal vertebra of Dicraeosaurus hansemanni from the Kimmeridgian–Tithonian Tendaguru Formation of Tanzania (Janensch 1914, 1929: pl. 3) is moderately shortened (centrum length to centrum width ratio is 0.70, similar to 0.71 in MCEBC 1100300/215 but greater than in MCEBC 1100300/216). As in the Peski sauropod, the posterior centrum articulation surface is slightly convex and posterior centrum articulation surface projects more ventrally compared with the anterior centrum articulation surface. The ventral margin of the caudal rib is oriented dorsolaterally, as in MCEBC 1100300/216. The latter specimen might have had a similarly large wing-like caudal rib, as in Dicraeosaurus. The neural canal is of similar size and ovoid shape. In Dicraeosaurus, on the first caudal there is a fully developed hyposphene, with only hyposphenial ridges on the more posterior caudals. Additionally, the wing-like caudal rib is retained until the sixth caudal. If the Peski sauropod is a dicraeosaurid, then MCEBC 1100300/216, which has wing-like caudal rib and hyposphenial ridge, could serially be caudal 2 to 4, rather than the first caudal. The anterior caudals of Dicraeosaurus have more transversely compressed centra compared to the Peski sauropod.
The anterior caudal of the dicraeosaurid Suuwassea emilieae from the Tithonian Morrison Formation of Montana, USA (Harris and Dodson 2004; Harris 2006: fig. 13A–C) has a rhomboid anterior articular surface, slightly convex posterior articular surface, and a shallowly concave ventral centrum margin in lateral view. In these respects, it is very similar with MCEBC 1100300/216. It differs from the Peski sauropod by having large nutrient foramina on the ventral centrum surface.
A caudal vertebra of Pilmatueia faundezi from the Valanginian Mulichinco Formation of Argentina (Coria et al. 2019: fig. 10) was described serially as a middle caudal, although it has a distinct caudal rib. It is similar with MCEBC 1100300/ 215 in having a rhomboid outline of the centrum articular surfaces, flat posterior centrum articulation surface, relative length of the centrum, position of the caudal rib, and size of neural canal.
A more posterior anterior caudal from the Peski sauropod (MCEBC 1100300/215) is very similar to the anterior caudal (possibly C3 or C4), of the dicraeosaurid Amargatitanis macni from the Barremian La Amarga Formation of Argentina (Apesteguía 2007: fig. 6A–E; Gallina 2016: fig. 2A–D). These similarities include: a somewhat rhomboid appearance of the anterior centrum articulation surface, centrum length to centrum width ratio (0.74 in Amargatitanis; 0.71 in MCEBC 1100300/215), lack of anterior chevron facets, large posterior chevron facets with a shallow groove between them, and dorsolaterally directing caudal ribs. The posterior centrum articulation surface of this specimen was described as flat (Gallina 2016: 80), but appears to be slightly convex, as in the Peski sauropod. There is a marked hyposphenial ridge, similar to that in MCEBC 1100300/216 (not preserved in MCEBC 1100300/215). The prezyggapophyseal centrodiapophyseal fossa in MCEBC 1100300/215 is much smaller and anteriorly restricted compared with the caudal vertebra of Amargatitanis.
Diplodocids are markedly different from the Peski sauropod by “dorsalized” anterior caudal vertebrae with diapophyseal laminae and “pleurocoels” (Wilson 1999). These characters are most prominent in Diplodocus longus and D. hallorum from the Kimerridgian–Tithonian Morrison Formation of the western USA (Marsh 1878; Osborn 1899: fig. 13; Gillette 1991: figs. 4–8), while in Apatosaurus ajax from the same formation, the “pleurocoels” and diapophyseal laminae on anterior caudals are not so pronounced (Marsh 1877; Upchurch et al. 2004: pl. 5).
The first caudal of a rebbachisaurid Demandasaurus darwini from the Barremian–Aptian Castrillo la Reina Formation of Spain (Torcida Fernández-Baldor et al. 2011: fig. 10) has a slightly opisthocoelous semilunar centrum with deeply incised dorsal margin and very high neural canal. A more posterior anterior caudal has a large pneumatic depression on the anterior side of a wing-like caudal rib (Torcida Fernández-Baldor et al. 2011: fig. 11). All these characters clearly differentiate Demandasaurus and Peski sauropod.
The first caudal of the rebbachisaurid Comahuesaurus windhauseni from the Aptian–Albian Lohan Cura Formation of Argentina (Carballido et al. 2012: fig. 6) has a long slightly opisthocoelous centrum (centrum length to width ratio is 0.85), dorsoventrally compressed caudal rib, and deep fossa on the anterior side occupying part of the centrum, neural arch and caudal rib. All these features differentiate Comahuesaurus from the Peski sauropod. The only common feature between the two is a more ventrally projecting posterior centrum articulation surface.
The anterior caudal (probably C2 or C3), of the rebbachisaurid Cathartesaura anaerobica from the Turonian–Coniacian Huincul Formation of Argentina (Gallina and Apesteguía 2005: fig. 3A–C) differs from the Peski specimens by distinctly anteroposteriorly longer amphyplatian centrum, twisted caudal rib, prominent centroprezigapophyseal and centropostzygapophyseal laminae, and much larger posterior opening of the neural canal.
The preceding comparison shows that the anterior caudals of the Peski sauropod are clearly different from the anterior caudals of diplodocids, which have “pleurocoels” and diapophyseal laminae, and from the anterior caudals of rebacchisaurids, which have relatively long centrum and often slightly opisthocoelous first caudal. Morphologically, the Peski caudals are most similar to the anterior caudals of various dicraeosaurids, including Dicraeosaurus.
Phylogenetic analysis
To assess the phylogenetic position of the Peski sauropod, we used two recent taxon-character datasets for sauropods. The first matrix was developed by Gonzàlez Riga et al. (2018) and modified by Mannion et al. (2019a, b). The matrix is focused on basal somphospondylans but covers variety of other sauropod taxa. The matrix consists of 548 characters and 124 taxa. The Peski sauropod was scored for 16 characters (2.9%) from this matrix (see Appendix 1). The characters 11, 14, 15, 27, 40, 51, 104, 122, 147, 148, 195, 205, 259, 297, 426, 435, 472, and 510 were treated as ordered multistate characters and several unstable and fragmentary taxa were excluded from the analyses a priori (Astrophocaudia, Australodocus, Brontomerus, Fukuititan, Fusuisaurus, Liubangosaurus, Malarguesaurus, and Mongolosaurus). The second matrix is that of Rauhut et al. (2015) modified by Xu et al. (2018). This matrix covers all major sauropod groups. The matrix consists of 375 characters and 73 taxa. The Peski sauropod was scored for 13 characters (3.5%) from this matrix (see Appendix 1). The characters 12, 58, 95, 96, 102, 106, 108, 115, 116, 119, 120, 145, 152, 163, 213, 216, 232–235, 252, 256, 298, 299, and 301 were treated as ordered multistate characters.
We employed three analytical protocols for each matrix. In all analyses all characters were equally weighted. The first analysis was performed using NONA version 2.0 (Goloboff 1999; Goloboff and Catalano 2016) run with WinClada version 1.00.08 interface (Nixon 1999). We performed one thousand repetitions of the parsimony ratchet (island hopper) algorithm. For the second analysis we used TNT version 1.1 (Goloboff et al. 2008) New Technology Search with default settings. In the third analysis we used New Technology Search with Stabilize Consensus option, employing sectorial searches, drift and tree fusing, with the consensus stabilized five times. The produced trees were then subjected to traditional TBR branch swapping. The tree statistics, consensus tree, and distribution of characters were obtained using WinClada interface. The tree statistics for the all analyses are summarized in Table 1.
Table 1. Statistics for the most parsimonious trees obtained in the phylogenetic analyses. *overflow
|
Analysis |
Matrix 1 |
Matrix 2 |
||||||
|
Number of trees |
Tree |
Consistency index (CI) |
Retention index (RI) |
Number of trees |
Tree |
Consistency index (CI) |
Retention index (RI) |
|
|
1 |
19 |
2659 |
0.21 |
0.58 |
111 |
1142 |
0.38 |
0.72 |
|
2 |
3 |
2658 |
0.21 |
0.58 |
3 |
1142 |
0.38 |
0.72 |
|
3 |
10000* |
2655 |
0.21 |
0.58 |
952 |
1142 |
0.38 |
0.72 |
The third analysis of the Matrix 1 produced maximum number of most parsimonious trees (10 000; Table 1), and not surprisingly, the consensus tree (not shown) has little resolution. The Peski sauropod on that tree is generally collapsed with most diplodocoid sauropods. All the other analyses place the Peski sauropod within Diplodocoidea. The analyses of the Matrix 1 recovers the Peski sauropod as either a basal dicraeosaurid (NONA parsimony ratchet analysis; Fig. 4A) or basal rebbachisaurid (TNT New technology search with default settings; Fig. 4B). The analyses of Matrix 2 are more consistent in placing the Peski sauropod at the base of Diplodocidae (Fig. 4C, D), although all Diplodocidae and Dicraeosauridae are collapsed on the consensus tree from the Analysis 3 (Fig. 4E).
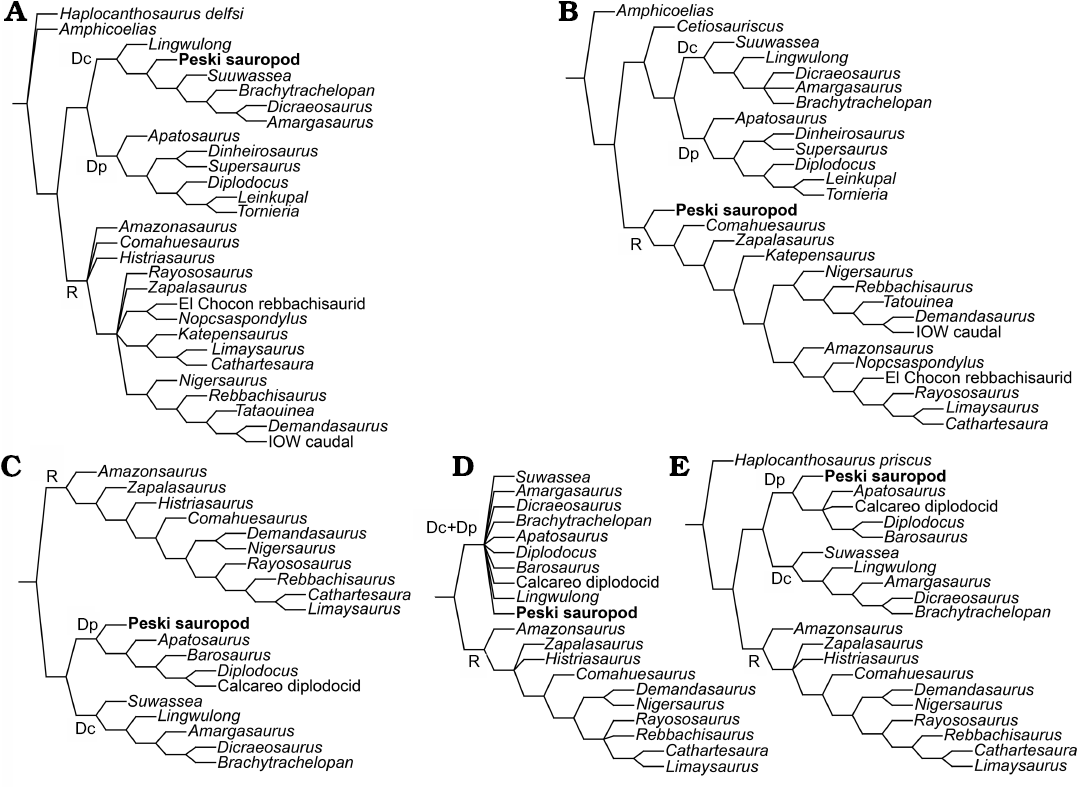
Fig. 4. Phylogenetic position of the Peski sauropod (Diplodocoidea indet.) within Diplodocoidea on the strict consensus trees recovered by phylogenetic analyses based on Matrix 1 (A, B) and Matrix 2 (C–E). A, C, NONA parsimony ratchet analysis; B, E, TNT New Technology search with defaults settings; D, TNT New Technology Search with stabilized consensus and TBR. Abbreviations: Dc, Dicraeosauridae; Dp, Diplodocidae; R, Rebbachisauridae.
In the analyses of Matrix 1, placement of the Peski sauropod within Dicraeosauridae is supported by characters 26(1) and 502(1) in Analysis 1, to Rebbachisauridae by characters 26(1), 502(1), and 505(1) in Analysis 2, to Dicraeosauridae + Diplodocidae by character 204(1) in Analysis 1, and to Diplodocoidea by characters 205(1) and 503(0) in Analysis 1 and 204(1) and 205(1) in Analysis 2. In the analyses of Matrix 2, placement of the Peski sauropod within Diplodocidae is supported by characters 190(1) and 193(2) in Analyses 1 and 2 and to Diplodocidae + Dicraeosauridae by character 198(1) in all three analyses.
Discussion
The time and place of the origin of Neosauropoda and early diversification of its main groups (Diplodocoidea and Macronaria) is one of the most controversial topics in the evolution of Sauropoda (Xu et al. 2018). The Middle Jurassic faunas of Asia are dominated by non-neosauropod eusauropods, including Shunosaurus, Datousaurus, Nebulasaurus, and diverse mamenchisaurids (Dong and Tang 1984; Zhang 1988; Chatterjee and Zheng 2002; Xing et al. 2015a, b; Averianov et al. 2019). Ferganasaurus verzilini from the Callovian Balabansai Formation of Kyrgyzstan was originally described as a neosauropod (Alifanov and Averianov 2003), but subsequent analyses place it outside of Neosauropoda (Ksepka and Norell 2010; Läng and Mahammed 2010). Bellusaurus sui from the Callovian–Oxfordian Shishugou Formation of Xinjiang Uyghur Autonomous Region, China is a basal macronarian or close relative of Neosauropoda (Dong 1990; Moore et al. 2018). Non-neosauropod eusauropods are also known from the Middle Jurassic of North Africa (Monbaron et al. 1999; Mahammed et al. 2005; Läng and Mahammed 2010). In the Middle Jurassic of Europe there are non-neosauropod eusauropods as well some putative neosauropods. One of the best-known European Jurassic non-neosauropod eusauropods is Cetiosaurus oxoniensis from the Bajocian Rutland and Bathonian Forest Marble formations of England (Phillips 1871; Upchurch and Martin 2002, 2003).
The dicraeosaurid diplodocoid Lingwulong shenqi from the Toarcian–Bajocian Yanan Formation of Ningxia Hui Autonomous Region, China, is the oldest neosauropod described to-date (Xu et al. 2018). While the brachiosaurid Vouivria damparisensis from the Oxfordian of France is the earliest known titanosauriform (Mannion et al. 2017). Additionally, there are possible brachiosaurid caudal vertebrae from the Callovian Oxford Clay Formation of England (Woodward 1905: fig. 42; Upchurch and Martin 2003: fig. 3; Noè et al. 2010: fig. 6). The phylogenetic position of Cetiosauriscus stewarti from the Oxford Clay Formation is debated (see discussion in Xu et al. 2018). This taxon was considered as a diplodocoid by a number of studies, but recent phylogenetic analyses recover it as a non-neosauropod sauropod (Rauhut et al. 2005; Mannion et al. 2012). However, in one of our cladograms (Fig. 4B) Cetiosauriscus is recovered as a sister taxon to Dicraeosauridae + Diplodocidae. The fissured chevrons of Cetiosauriscus (Woodward 1905; Charig 1980) suggest its affinities with Diplodocidae. Distal caudal vertebrae of a “whiplash tail” from the Oxford Clay Formation (Woodward 1905: fig. 45; Noè et al. 2010: fig. 11) have elongation index values lower than in known diplodocoids (Whitlock 2011); but this could be a plesiomorphic feature of Middle Jurassic diplodocoids. A possible neosauropod anterior caudal vertebra from the Oxford Clay Formation (Holwerda et al. 2019: fig. 4) is overall similar with MCEBC 1100300/216 in proportions of the centrum, more ventrally projecting posterior centrum articulation surface, and possible presence of a hyposphenial ridge. It differs by the presence of a ventral keel on the centrum (which is shown on explanatory drawings but not discernible from the photographs) and a relatively larger neural canal.
The preceding review shows that neosauropods were likely present in the Middle Jurassic of Europe and already differentiated into diplodocoids and macronarians. Discovery of a diplodocoid in the Peski locality is in line with these observations. Interestingly, the Middle Jurassic Peski sauropod is more similar to the Late Jurassic–Early Cretaceous dicraeosaurids of Africa and South America than to the Early–Middle Jurassic dicraeosaurid Lingwulong from East Asia. This may suggest that initial radiation of Dicraeosauridae took place in Western Asia or Europe in Middle Jurassic and then these more derived dicraeosaurids dispersed to Africa and the Americas. The closest land to the Peski locality and a presumable source area of sediments in the Callovian was the Fennoscandian landmass (Fig. 5), where Jurassic terrestrial vertebrates are currently unknown. In the Aalenian–early Bajocian the Fennoscandian landmass was connected to Asia and separated from North America (Ippolitov and Desai 2019: fig. 9). In the early Bathonian, it becomes disconnected from Asia by the Middle Russian Sea (Ippolitov and Desai 2019: fig. 9). Most likely diplodocoid sauropods dispersed to Fennoscandia from Asia because diplodocoids are known there in the latest Early–earliest Middle Jurassic (Lingwulong). This event should have occurred before the early Bathonian, when the land bridge between Asia and Fennoscandia disappeared. This supports pre-Bathonian diversification of the main neosauropod lineages, first suggested by the discovery of Lingwulong (Xu et al. 2018).
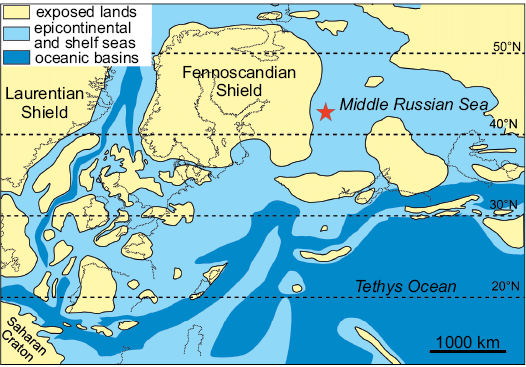
Fig. 5. Late Callovian to early Oxfordian paleogeography of Europe (after Wierzbowski and Rogov 2011; modified according to the data of Sazonova and Sazonov, 1967). The position of Peski locality is shown by a star.
Acknowledgements
We thank Maxim S. Boyko and Dmitry B. Kucher (both MCEBC) who kindly allowed the additional preparation and study of specimens under their care. Alexander B. Vidryk (Moscow, Russia) found the specimen in 1997 and granted it to the MCEBC. We thank John Whitlock (Mount Aloysius College, Cresson, USA) and Cary Woodruff (Great Plains Dinosaur Museum and Field Station, Malta, USA) for reviewing the paper and useful suggestions. Thanks to Alexey P. Ippolitov (Geological Institute, Russian Academy of Sciences, Moscow, Russia) for the taxonomic identification of epibiont serpulids covering the vertebrae and for consultations on the stratigraphy of the Peski locality. We thank the Willi Hennig Society for their sponsorship making TNT available for all researchers free of cost. Financial support to AOA was provided by the Russian Science Foundation (project 19-14-00020) and the Zoological Institute, Russian Academy of Sciences (project АААА-А19-119032590102-7). The work of NGZ was supported by the state program 0135-2019-0066 (Geological Institute of the Russian Academy of Sciences).
References
Alekseev, A.S., Agadjanian, A.K. [Agadânian, A.K.], Areshin, A.V. [Arešin, A.V.], Barskov, I.S., Gordenko, N.V., Efimov, M.B., Kabanov, P.B., Krasilov, V.A., Krasnikov, N.M., Lebedev, O.A., Rosanova, A.A., Sennikov, A.G., Smirnova, S.B., Sukhanov, V.B. [Suhanov, V.B.], Fokin, P.A., and Schmidt, A.V. 2001. Discovery of a unique locality of Middle Jurassic Fauna and Flora in Moscow Region [in Russian]. Doklady Akademii Nauk 377: 359–362.
Alifanov, V.R. and Averianov, A.O. 2003. Ferganasaurus verzilini, gen. et sp. nov., a new neosauropod (Dinosauria, Saurischia, Sauropoda) from the Middle Jurassic of Fergana Valley, Kirghizia. Journal of Vertebrate Paleontology 23: 358–372. Crossref
Alifanov, V.R. and Sennikov, A.G. 2001. About discovery of dinosaur remains in Moscow region [in Russian]. Doklady Akademii Nauk 376: 73–75.
Apesteguía, S. 2007. The sauropod diversity of the La Amarga Formation (Barremian), Neuquén (Argentina). Gondwana Research 12: 533–546. Crossref
Averianov, A.O., Krasnolutskii, S.A., Ivantsov, S.V., Skutschas, P.P., Schellhorn, R., Schultz, J.A., and Martin, T. 2019. Sauropod remains from the Middle Jurassic Itat Formation of West Siberia, Russia. Paläontologische Zeitschrift 93: 691–701. Crossref
Bragina, E.N. 2005. Hybodont shark Hybodus hauffianus Fraas, 1895 from the continental deposits of upper Bathonian in Moscow Province [in Russian]. Izvestiâ vuzov. Geologiâ i razvedka 1: 19–22.
Carballido, J.L., Salgado, L., Pol, D., Canudo, J.I., and Garrido, A. 2012. A new basal rebbachisaurid (Sauropoda, Diplodocoidea) from the Early Cretaceous of the Neuquén Basin; evolution and biogeography of the group. Historical Biology 24: 631–654. Crossref
Charig, A.J. 1980. A diplodocid sauropod from the Lower Cretaceous of England. In: L.L. Jacobs (ed.), Aspects of Vertebrate History: Essays in Honor of Edwin Harris Colbert, 231–244. Museum of Northern Arizona Press, Flagstaff.
Chatterjee, S. and Zheng, Z. 2002. Cranial anatomy of Shunosaurus, a basal sauropod dinosaur from the Middle Jurassic of China. Zoological Journal of the Linnean Society 136: 145–169. Crossref
Coria, R.A., Windholz, G.J., Ortega, F., and Currie, P.J. 2019. A new dicraeosaurid sauropod from the Lower Cretaceous (Mulichinco Formation, Valanginian, Neuquen Basin) of Argentina. Cretaceous Research 93: 33–48. Crossref
Dong, Z. 1990. On remains of the sauropods from Kelamaili region, Junggar Basin, Xinjiang, China [in Chinese with English abstract]. Vertebrata PalAsiatica 28: 43–58.
Dong, Z.-M. and Tang, Z.-L. 1984. Note on a new mid-Jurassic sauropod (Datousaurus bashanensis gen. et sp. nov.) from Sichuan Basin, China [in Chinese with English abstract]. Vertebrata PalAsiatica 22: 69–75.
Gallina, P.A. 2016. Reappraisal of the Early Cretaceous sauropod dinosaur Amargatitanis macni (Apesteguía, 2007), from northwestern Patagonia, Argentina. Cretaceous Research 64: 79–87. Crossref
Gallina, P.A. and Apesteguía, S. 2005. Cathartesaura anaerobica gen. et sp. nov., a new rebbachisaurid (Dinosauria, Sauropoda) from the Huincul Formation (Upper Cretaceous), Río Negro, Argentina. Revista del Museo Argentino de Ciencias Naturales, nueva serie 7: 153–166. Crossref
Gambaryan, P.P. and Averianov, A.O. 2001. Femur of a morganucodontid mammal from the Middle Jurassic of Central Russia. Acta Palaeontologica Polonica 46: 99–112.
Gillette, D.D. 1991. Seismosaurus halli, gen. et sp. nov., a new sauropod dinosaur from the Morrison Formation (Upper Jurassic/Lower Cretaceous) of New Mexico, USA. Journal of Vertebrate Paleontology 11: 417–433. Crossref
Goloboff, P.A. 1999. NONA (ver. 1.9). Software published by the author, Tucumán [available online at www.cladistics.org].
Goloboff, P. and Catalano, S. 2016. TNT, version 1.5, with a full implementation of phylogenetic morphometrics. Cladistics 32: 221–238. Crossref
Goloboff, P.A., Farris, J.S., and Nixon, K.C. 2008. TNT (Tree analysis using New Technology) (BETA). Software published by the authors, Tucumán.
Gonzàlez Riga, B.J., Mannion, P.D., Poropat, S.F., Ortiz David, L.D., and Coria, J.P. 2018. Osteology of the Late Cretaceous Argentinean sauropod dinosaur Mendozasaurus neguyelap: implications for basal titanosaur relationships. Zoological Journal of the Linnean Society 184: 136–181. Crossref
Harris, J.D. 2006. The axial skeleton of the dinosaur Suuwassea emilieae (Sauropoda: Flagellicaudata) from the Upper Jurassic Morrison Formation of Montana, USA. Palaeontology 49: 1091–1121. Crossref
Harris, J.D. and Dodson, P. 2004. A new diplodocoid sauropod dinosaur from the Upper Jurassic Morrison Formation of Montana, USA. Acta Palaeontologica Polonica 49: 197–210.
Holwerda, F.M., Evans, M., and Liston, J.J. 2019. Additional sauropod dinosaur material from the Callovian Oxford Clay Formation, Peterborough, UK: evidence for higher sauropod diversity. PeerJ 7: e6404. Crossref
Janensch, W. 1914. Übersicht über die Wirbeltierfauna der Tendaguru-Schichten, nebst einer kurzen Charakterisierung der neu aufgeführten Arten von Sauropoden. Archiv für Biontologie 3: 81–110.
Janensch, W. 1929. Die Wirbelsäule der Gattung Dicraeosaurus. Palaeontographica Supplementbände SVII: 35–133.
Ippolitov, A.P. 2007. Contribution to the revision of some late Callovian serpulids (Annelida, Polychaeta) of Central Russia: Part 2. Paleontological Journal 41: 429–436. Crossref
Ippolitov, A.P. and Desai, B.G. 2019. Dwarf megateuthidid belemnites from the Bathonian of Kachchh (India: Gujarat) and their significance for palaeobiogeography. Journal of Systematic Palaeontology 17: 613–634. Crossref
Krupina, N.I. 1995. First finding of Jurassic ceratodontids (Dipnoi) in Moscoe Region [in Russian]. Paleontologičeskij žurnal 1995 (2): 129–131.
Ksepka, D.T. and Norell, M.A. 2010. The illusory evidence for Asian Brachiosauridae: new material of Erketu ellisoni and a phylogenetic reappraisal of basal Titanosauriformes. American Museum Novitates 3700: 1–27. Crossref
Läng, E. and Mahammed, F. 2010. New anatomical data and phylogenetic relationships of Chebsaurus algeriensis (Dinosauria, Sauropoda) from the Middle Jurassic of Algeria. Historical Biology 22: 142–164. Crossref
Mahammed, F., Läng, E., Mami, L., Mekahli, L., Benhamou, M., Bouterfa, B., Kacemi, A., Cherief, S.-A., Chaouati, H., and Taquet, P. 2005. The “Giant of Ksour”, a Middle Jurassic sauropod dinosaur from Algeria. Comptes Rendus Palevol 4: 1–8. Crossref
Mannion, P.D., Allain, R., and Moine, O. 2017. The earliest known titanosauriform sauropod dinosaur and the evolution of Brachiosauridae. PeerJ 5: e3217. Crossref
Mannion, P.D., Upchurch, P., Jin, X., and Zheng, W. 2019a. New information on the Cretaceous sauropod dinosaurs of Zhejiang Province, China: impact on Laurasian titanosauriform phylogeny and biogeography. Royal Society Open Science 6: 191057. Crossref
Mannion, P.D., Upchurch, P., Mateus, O., Barnes, R.N., and Jones, M.E.H. 2012. New information on the anatomy and systematic position of Dinheirosaurus lourinhanensis (Sauropoda: Diplodocoidea) from the Late Jurassic of Portugal, with a review of European diplodocoids. Journal of Systematic Palaeontology 10: 521–551. Crossref
Mannion, P.D., Upchurch, P., Schwarz, D., and Wings, O. 2019b. Taxonomic affinities of the putative titanosaurs from the Late Jurassic Tendaguru Formation of Tanzania: phylogenetic and biogeographic implications for eusauropod dinosaur evolution. Zoological Journal of the Linnean Society 185: 784–909. Crossref
Marsh, O.C. 1877. Notice of some new dinosaurian reptiles from the Jurassic Formation. American Journal of Science, Series 3 14: 514–516. Crossref
Marsh, O.C. 1878. Principal characters of American Jurassic dinosaurs. Part I. American Journal of Science, Series 3 16: 411–416. Crossref
Marsh, O.C. 1884. Principal characters of American Jurassic dinosaurs. Part VII. On the Diplodocidae, a new family of the Sauropoda. American Journal of Science, Series 3 27: 160–168. Crossref
Monbaron, M., Russell, D.A., and Taquet, P. 1999. Atlasaurus imelakei n. g., n. sp., a brachiosaurid-like sauropod from the Middle Jurassic of Morocco. Comptes Rendus de l’Académie des Sciences, Series II, Fascicule A: Sciences de la Terre et des planètes 329: 519–526. Crossref
Moore, A.J., Mo, J., Clark, J.M., and Xu, X. 2018. Cranial anatomy of Bellusaurus sui (Dinosauria: Eusauropoda) from the Middle–Late Jurassic Shishugou Formation of northwest China and a review of sauropod cranial ontogeny. PeerJ 6: e4881. Crossref
Nixon, K.C. 1999. Winclada (Beta) version 0.9.9. Software published by the author, Ithaca [available online at www.cladistics.org].
Noè, L.F., Liston, J.J., and Chapman, S.D. 2010. “Old bones, dry subject”: the dinosaurs and pterosaur collected by Alfred Nicholson Leeds of Peterborough, England. In: R.T.J. Moody, E. Buffetaut, D. Nais, and D.M. Martill (eds.), Dinosaurs and Other Extinct Saurians: A Historical Perspective. Geological Society of London, Special Publications 343: 49–77. Crossref
Osborn, H.F. 1899. A skeleton of Diplodocus. Memoirs of the American Museum of Natural History 1: 191–214.
Pashchenko, D.I., Kuzmin, I.T., Sennikov, A.G., Skutschas, P.P., and Efimov, M.B. 2018. On the finding of neosuchians (Neosuchia, Crocodyliformes) in the Middle Jurassic (Bathonian) deposits of the Moscow Region. Paleontological Journal 52: 550–562. Crossref
Phillips, J. 1871. Geology of Oxford and the Valley of the Thames. 529 pp. Clarendon Press, Oxford. Crossref
Popov, E.V. and Shapovalov, K.M. [Šapovalov, K.M.] 2007. New finds of chimaeroid fishes (Holocephali, Chimaeroidei) from the Jurassic of European Russia [in Russian]. In: A.Y. Rozanov, A.V. Lopatin, and P.Y. Parhaev (eds.), Sovremennaâ paleontologiâ klassičeskie i novejšie metody, 25–44. Paleontologičeskij institut RAN, Moskva.
Rauhut, O.W.M., Carballido, J.L., and Pol, D. 2015. A diplodocid sauropod dinosaur from the Late Jurassic Cañadón Calcáreo Formation of Chubut, Argentina. Journal of Vertebrate Paleontology 35: e982798. Crossref
Rauhut, O.W.M., Remes, K., Fechner, R., Cladera, G., and Puerta, P.F. 2005. Discovery of a short-necked sauropod dinosaur from the Late Jurassic period of Patagonia. Nature 435: 670–672. Crossref
Sazonova, I.G. and Sazonov, N.T. 1967. Paleogeografiâ Russkoj platformy v ûrskoe i rannemelovoe vremâ. 260 pp. Nedra, Leningrad.
Schlotheim, E.F. von 1820. Die Petrefactenkunde auf ihrem jetzigen Standpunkte. 432 pp. Becken, Gotha.
Sowerby, J. 1829. The Mineral Conchology of Great Britain. Vol. 6. 230 pp. J. Sowerby, London.
Sukhanov, V.B. 2006. An archaic turtle, Heckerochelys romani gen. et sp. nov., from the Middle Jurassic of Moscow Region, Russia. Fossil Turtle Research 1: 112–118.
Torcida Fernández-Baldor, F., Canudo, J.I., Huerta, P., Montera, D., Pereda Suberbiola, X., and Salgado, L. 2011. Demandasaurus darwini, a new rebbachisaurid sauropod from the Early Cretaceous of the Iberian Peninsula. Acta Palaeontologica Polonica 56: 535–552. Crossref
Tesakova, E.M. 2003. Callovian and Oxfordian Ostracodes from the Central Region of the Russian Plate. Paleontological Journal 37: S107–S227.
Upchurch, P. 1998. The phylogenetic relationships of sauropod dinosaurs. Zoological Journal of the Linnean Society 124: 43–103. Crossref
Upchurch, P. and Martin, J. 2002. The Rutland Cetiosaurus: the anatomy and relationships of a Middle Jurassic British sauropod dinosaur. Palaeontology 45: 1049–1074. Crossref
Upchurch, P. and Martin, J. 2003. The anatomy and taxonomy of Cetiosaurus (Saurischia, Sauropoda) from the Middle Jurassic of England. Journal of Vertebrate Paleontology 23: 208–231. Crossref
Upchurch, P., Tomida, Y., and Barrett, P.M. 2004. A new specimen of Apatosaurus ajax (Sauropoda: Diplodocidae) from the Morrison Formation (Upper Jurassic) of Wyoming, USA. National Science Museum Monographs 26: 1–156.
Whitlock, J.A. 2011. A phylogenetic analysis of Diplodocoidea (Saurischia: Sauropoda). Zoological Journal of the Linnean Society 161: 872–915. Crossref
Wierzbowski, H. and Rogov, M. 2011. Reconstructing the palaeoenvironment of the Middle Russian Sea during the Middle–Late Jurassic transition using stable isotope ratios of cephalopod shells and variations in faunal assemblages. Palaeogeography, Palaeoclimatology, Palaeoecology 299: 250–264. Crossref
Wilson, J.A. 1999. A nomenclature for vertebral laminae in sauropods and other saurischian dinosaurs. Journal of Vertebrate Paleontology 19: 639–653. Crossref
Woodward, A.S. 1905. On parts of the skeleton of Cetiosaurus leedsi, a sauropodous dinosaur from the Oxford clay of Peterborough. Proceedings of the Zoological Society of London 1905: 232–243.
Xing, L., Miyashita, T., Currie, P.J., You, H., Zhang, J., and Dong, Z. 2015a. A new basal eusauropod from the Middle Jurassic of Yunnan, China, and faunal compositions and transitions of Asian sauropodomorph dinosaurs. Acta Palaeontologica Polonica 60: 145–154.
Xing, L., Miyashita, T., Zhang, J., Li, D., Ye, Y., Sekiya, T., Wang, F., and Currie, P.J. 2015b. A new sauropod dinosaur from the Late Jurassic of China and the diversity, distribution, and relationships of mamenchisaurids. Journal of Vertebrate Paleontology 35: e889701. Crossref
Xu, X., Upchurch, P., Mannion, P.D., Barrett, P.M., Regalado-Fernandez, O.R., Mo, J., Ma, J., and Liu, H. 2018. A new Middle Jurassic diplodocoid suggests an earlier dispersal and diversification of sauropod dinosaurs. Nature Communications 9: 2700. Crossref
Zhang, Y. 1988. The Middle Jurassic Dinosaur Fauna from Dashanpu, Zigong, Sichuan. Vol. I. Sauropod Dinosaurs (1). Shunosaurus [in Chinese]. 87 pp. Sichuan Publishing House of Science and Technology, Chengdu.
Zverkov, N.G., Shmakov, A.S. [Šmakov, A.S.], and Arkhangelsky, M.S. [Arhangelski, M.S.] 2017. Jurassic marine reptiles of Moscow and surroundings [in Russian]. Trudy Geologičeskogo Instituta Rossijskoj Akademii Nauk 615: 230–263.
Scorings of Peski sauropod for the data matrices.
Matrix 1 (Mannion et al. 2019a, b)
C25(1). Anterior caudal centra, mediolateral width to dorsoventral height (excluding chevron facets) of anterior surface ratio: 1.0 or greater [1.19 for MCEBC 1100300/216].
C26(1). Anteriormost caudal centra, lowest average Elongation Index [aEI; centrum anteroposterior length (excluding articular ball) divided by the mean average value of the anterior surface mediolateral width and dorsoventral height] value of: 0.6 or greater [0.67 for MCEBC 1100300/216].
C178(0). Anterior caudal centra, lateral pneumatic fossae or foramina: absent.
C180(0). Anterior-middle caudal centra, small, shallow vascular foramina pierce the lateral and/or ventral surfaces: absent.
C181(0). Anterior caudal centra (excluding the anteriormost caudal vertebrae), ventral longitudinal hollow: absent.
C182(0). Anterior caudal centra (excluding the anteriormost caudal vertebrae), distinct ventrolateral ridges, extending the full length of the centrum: absent.
C187(0). Anterior caudal neural arches, hyposphenal ridge: present.
C188(0). Anterior caudal neural arches, hyposphenal ridge shape: slender ridge.
C203(0). First caudal rib, expands anteroposteriorly towards its distal end, forming an “anchor” shape in dorsal view: absent.
C204(1). Anterior caudal ribs, shape in anterior view: wing-like, with a dorsolaterally oriented dorsal margin.
C205(1). Anterior caudal ribs: curve mainly laterally.
C206(0). Anterior caudal ribs: do not extend beyond posterior end of centrum (excluding posterior ball).
C501(0). Anteriormost caudal ribs (excluding the first), distal tip ventrally deflected, such that the ventral margin of the caudal rib is strongly concave in anterior/posterior view: absent.
C502(1). Anteriormost caudal ribs, orientation of ventral surface in anterior/posterior view: dorsolateral (at 30° or more to the horizontal plane, typically 40–50°).
C503(0). Anteriormost caudal ribs, anterior surface: unexcavated.
C505(1). Anterior caudal ribs, position: restricted to the neural arch and dorsal margin of centrum.
Matrix 2 (Xu et al. 2018)
C188(0). Caudal bone texture: solid.
C190(1). First caudal centrum or last sacral vertebra, articular face shape: procoelous.
C192(1). Anterior caudal vertebrae, transverse processes: ventral surface directed dorsally.
C193(2). Anterior caudal centra (excluding the first), articular face shape: slightly procoelous.
C194(0). Anterior caudal centra, pleurocoels: absent.
C195(0). Anterior caudal vertebrae, ventral surfaces: convex transversely.
C196(0). Anterior and middle caudal vertebrae, ventrolateral ridges: absent.
C198(1). Anterior caudal transverse processes shape: “winglike”, not tapering distally.
C200(1). Anterior caudal transverse processes, proximal depth: deep, extending from centrum to neural arch.
C201(0). Anterior caudal transverse processes, diapophyseal laminae (ACDl, PCDL, PRDL, PODL): absent.
C203(1). Anterior caudal vertebrae, hyposphene ridge: present.
C358(0). Anterior caudal vertebrae (mainly the first and second): ventral bulge on transverse process: absent.
C359(0). Anterior and middle caudal vertebrae, blind fossae in lateral centrum: absent.
Acta Palaeontol. Pol. 65 (3): 499–509
2020 https://doi.org/10.4202/app.00724.2020