

A new middle Cambrian trilobite with a specialized cephalon from Shandong Province, North China
ZHIXIN SUN, HAN ZENG, and FANGCHEN ZHAO
Sun, Z., Zeng, H., and Zhao, F. 2020. A new middle Cambrian trilobite with a specialized cephalon from Shandong Province, North China. Acta Palaeontologica Polonica 65 (4): 709–718.
Trilobites achieved their maximum generic diversity in the Cambrian, but the peak of morphological disparity of their cranidia occurred in the Middle to Late Ordovician. Early to middle Cambrian trilobites with a specialized cephalon are rare, especially among the ptychoparioids, a group of libristomates featuring the so-called “generalized” bauplan. Here we describe an unusual ptychopariid trilobite Phantaspis auritus gen. et sp. nov. from the middle Cambrian (Miaolingian, Wuliuan) Mantou Formation in the Shandong Province, North China. This new taxon is characterized by a cephalon with an extended anterior area of double-lobate shape resembling a pair of rabbit ears in later ontogenetic stages; a unique type of cephalic specialization that has not been reported from other trilobites. Such a peculiar cephalon as in Phantaspis provides new insights into the variations of cephalic morphology in middle Cambrian trilobites, and may represent a heuristic example of ecological specialization to predation or an improved discoidal enrollment.
Key words: Trilobita, Ptychopariida, ontogeny, specialization, Miaolingian, Paleozoic, Longgang, Asia.
Zhixin Sun [zxsun@nigpas.ac.cn], Han Zeng [hzeng@nigpas.ac.cn], and Fangchen Zhao [fczhao@nigpas.ac.cn] (corresponding author), State Key Laboratory of Palaeobiology and Stratigraphy, Nanjing Institute of Geology and Palaeontology and Center for Excellence in Life and Palaeoenvironment, Chinese Academy of Sciences, Nanjing 210008, China; University of Chinese Academy of Sciences, Beijing 100049, China.
Received 3 April 2020, accepted 13 July 2020, available online 26 October 2020.
Copyright © 2020 Z. Sun et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Trilobites exhibit a bottom-centered diversification history, peaking in generic diversity, origination and extinction rates, and intraspecific variation during the Cambrian Period (e.g., Foote 1988; Fortey and Owens 1990b; Webster 2007). However, unlike these diversity measures, the morphological disparity of trilobites based on cranidial outline reached the peak in post-Cambrian times (Foote 1990, 1991), during the Middle to Late Ordovician (e.g., Foote 1993; Fortey and Owens 1997). Cambrian trilobites with a specialized cephalon were rare, and despite that some species exhibited additional cephalic border spines (e.g., McNamara 1978; Knell and Fortey 2005; Ren et al. 2017), the cranidia of Cambrian trilobites were less morphologically disparate than those of post-Cambrian ones (Foote 1991, 1993). The early to middle Cambrian ptychopariids have been regarded as an ancestral group of many highly apomorphic post-Cambrian trilobites (Fortey 2001), but cases of cranidial specialization are also rare in this group.
Here we describe a ptychopariid trilobite with an unusual cephalic morphology, Phantaspis auritus gen. et sp. nov. from the middle Cambrian (Miaolingian, Wuliuan) Mantou Formation in Shandong Province, North China. Phantaspis possesses an elongated cephalon with a deep notch of the anterior area that developed in later growth stages and is unknown from other Cambrian or younger trilobites. This remarkable example of cephalic outline change helps to understand the morphological range and structural foundation of the cephalic specialization in Cambrian trilobites, and appears to provides insight into life habits or ecological adaptation of middle Cambrian ptychopariids.
Institutional abbreviations.—NIGPAS, Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences, Nanjing, China.
Other abbreviations.—L0–L4, first to fifth lateral lobes; S0–S4, first to fifth lateral glabellar furrows; sag., sagittal; exs., exsagittal; tr., transverse.
Nomenclatural acts.—This published work and the nomenclatural acts it contains have been registered in ZooBank: urn:lsid:zoobank.org:pub:51844710-D71E-40D2-9882-566300524DA8
Material and methods
All specimens described in this paper were collected from the middle Cambrian Mantou Formation in the Longgang section in Shandong Province, North China (Fig. 1), and are deposited in the Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences, Nanjing, China (NIGPAS 172104–172136). Light photographs were taken using a Nikon D810 camera fitted with a Nikon AF-S Nikkor 105 mm lens. Images were processed using Adobe Photoshop to adjust tone, contrast, and brightness. The morphological terminology employed in this article follows that of Whittington and Kelly (1997). Measurements were made parallel to and normal to the sagittal line, the directions of which are referred to as sagittal/exsagittal and transverse, respectively.

Fig. 1. A. Geographical location of studied area (star) in Shandong Province, North China. B. General geological setting of the middle Cambrian Longgang section. C. Upper part of the Honghe Member of the Cambrian (Miaolingian, Wuliuan) Mantou Formation in the Longgang section, viewed from the west. Arrow is pointed to the person for scale. D. Stratigraphic column for the Honghe and Upper Shale members and the major trilobite horizons in the Longgang section. Alternative spelling of Zhangxia is Changhia. Arrow indicates the bed yielding Phantaspis auritus gen. et sp. nov.
Geological setting
The Mantou (alternative spelling “Manto”) Formation is a lithostratigraphical unit comprising sedimentary rocks of the Longwangmiaoan (corresponding to the lower part of Cambrian Stage 4), Maozhuangian, and Hsuchuangian (corresponding to the Wuliuan) regional stages (Zhang 1996; Yuan et al. 2012). In the Shandong Province, the Mantou Formation has been subdivided into the Shidian, Lower Shale, Honghe, and Upper Shale members.
All specimens of Phantaspis auritus gen. et sp. nov. were collected from the Honghe Member in the Longgang section in Linqu County (Fig. 1) (Sun et al. 2020). In this section the Honghe Member consists mainly of alternating beds of dark purple mudstone and sandstone (Fig. 2) that were deposited in a lagoonal or subtidal environment (Zhang 1996; Chough et al. 2010). All specimens of Phantaspis are from a single mudstone bed, 10.6 m above the base of the Honghe Member. The presence of Paraszeaspis cf. quadratus Zhang and Yuan in Yuan et al., 2015 in the Longgang section indicates that the beds yielding Phantaspis correspond to the middle part of the Sunaspis laevis–Sunaspidella rara Zone (Yuan et al. 2015), which is approximately correlative to the Pentagnostus praecurrens Zone (= Ptychagnostus praecurrens Zone) in Australia, Antarctica, and Laurentia, to the Kounamkites Zone in Siberia, and to the Peronopsis ultimus Zone in Kazakhstan (Zhu et al. 2019; Geyer 2019).

Fig. 2. Alternating dark purple mudstone and sandstone yielding Phantaspis auritus gen. et sp. nov. A. Vertical face. B. Surface of bed, showing the gutter cast; not circled trilobite. The length of both hammers is 28 cm.
Systematic palaeontology
Class Trilobita Walch, 1771
Order Ptychopariida Swinnerton, 1915
Superfamily Ptychopariacea Matthew, 1887
Family uncertain
Genus Phantaspis nov.
ZooBank LSID: urn:lsid:zoobank.org:act:DBC39820-49CE-43BF-8 ACE-0144F45B56FA
Etymology: From the Ancient Greek φαντασία (phantasia), fantasy and ἀσπίς (aspis), shield; in reference to the fantastic exoskeleton and Jay Chou’s album Fantasy.
Type species: Phantaspis auritus gen. et sp. nov., by monotypy, see below.
Diagnosis.—As for the type species by monotypy.
Phantaspis auritus sp. nov.
Figs. 3–5.
ZooBank LSID: urn:lsid:zoobank.org:act:9068AC88-7FCC-47FB-AB56-EE7511C24512
Etymology: From the Latin aurītus, having ears; in reference to the cranidium having a pair of long “ears”.
Type material: Holotype: a nearly complete exoskeleton, NIGPAS 172104 (Fig. 3A). Paratypes: two complete cranidia, NIGPAS 172105 (Fig. 3B) and NIGPAS 172106 (Fig. 3C) from the type locality and horizon.
Type locality: Longgang section, Shandong Province, North China.
Type horizon: 10.6 m above the base of the Honghe Member, Mantou Formation, Cambrian Miaolingian, Wuliuan Stage (Hsuchuangian, Sunaspis laevis–Sunaspidella rara Zone).
Material.—Type material and eight complete or fairly complete dorsal exoskeletons, NIGPAS 172104, 172107, 172110, 172113, 172126–172128, 172131; 55 cranidia, 10 of which are complete, NIGPAS 172105, 172106, 172108, 172114, 172115a–172115d, 172116, 172117; nine librigenae, NIGPAS 172104, 172108, 172126, 172128, 172132–17236 and eight pygidia, NIGPAS 172107, 172110, 172115, 172126, 172127, 172129–172131. All from the type locality and horizon.
Diagnosis.—Cranidium with an extended (sag.) anterior area having a deep indentation on the sagittal line; length (sag.) and width (tr.) of the cranidium comparable; glabella elongate, moderately tapered, slightly constricted in the middle (sag.); five pairs of elliptical lateral glabellar furrows (S0–S4) not connected to the axial furrow; ocular ridges elevated and robust; palpebral area wide (tr.); anterior border narrow; librigena trapezoidal in outline, lateral margin straight, ventral doublure extended conspicuously anteriorly. Thorax composed of 13 segments having relatively long pleural spines. Pygidium semicircular, moderately large; pygidial axis moderately tapered, broad (tr.); posterior pygidial margin relatively wide.
Description.—Adults: Cranidium extremely extended, constituting nearly half of the axial length of the exoskeleton (Fig. 3A); length 4.0–15.4 mm (n = 10), width at the palpebral lobes about 80 to 100% of the cranidium length; anterior border deeply intended medially, very strongly curved adaxially and forficiform in outline; anterior area smooth, expanded, developed into a pair of lobate processes separated by a distinct rearward indentation on the sagittal line, length greater than 50–65% (n = 10) of the cephalic length (Figs. 3A–F, 4E–H). Glabella subrectangular, moderately convex, slightly tapering forward and with constriction across S2, width at the anterior end 70% of the maximum width of glabella across occipital lobe; glabellar length 35–50% (n = 10) of the cranidial length, width across S1 25–35% (n = 10) of the cranidial width across same transverse line; five pairs of lateral glabellar furrows (S0–S4) present, moderately deep, oval in outline, adjacent to the axial furrow slightly; S0 directed transversely, consisting of a par of well-developed lateral portions of the furrows and a median section consisting of a much less indented to faint furrow with s slight forward bent medially; occipital ring moderately wide (sag.), slightly convex (tr. and sag.); S1 curved obliquely rearwardly from near axial furrow, with indistinct bifurcation adaxially (sometimes emphasized by dorsoventral compaction); S2 moderately long, directed slightly oblique from near axial furrow, often with slight curvature and a faint tendency to bifurcate adaxially; S3 developed as a short oval depression with its axis directed obliquely forward and adaxially, well distant from axial furrows due to a distinctly developed abaxial lobe of the glabella; S4 developed as a short slit-like indentation oblique to axis, extending from the axial furrow at the posterior margin of the eye ridge; frontal lobe subdivided by a shallow median sulcus in large individuals, slightly curved in juveniles (Fig. 3D). Axial furrow shallow and narrow, convergent from S0 to S2, then slightly divergent anteriorly; preglabellar furrow shallow, of uniform depth, narrow and slightly curved (Figs. 3, 4), separated from the axial furrow by a couple of fossulas. Preglabellar area slightly convex, length 40–62% (n = 5) of the anterior area length (Figs. 3A–F, 4E–H), with a faint domal swelling in front of the glabella (Fig. 3A, B). Anterior border developed as a narrow, slightly convex swelling coalescent with the adjacent distant parts of the preglabellar area; anterior margin forming a spectacular double curvature due to a distinct indentation on the sagittal line, which reduces the sagittal length of the preglabellar area to ca. one-half to two-thirds; anterior border furrow obsolescent, barely recognizable in holaspids (Figs. 3A–C, 4G–H). Palpebral area of the fixigena fairly broad, with faint caeca occasionally, transversely of ca. 70% the cranidial width across S1; slightly convex in transverse profile on internal moulds and on the shell exterior, sloping gently upward from the axial furrow; poorly defined from axial furrow on the shell exterior, but well demarcated on internal moulds. Palpebral lobes crescent shaped, upturned, clearly convex in tr. profile; anterior end located about opposite L3 to S3, posterior tips located about opposite S0 or the anterior third of L0; palpebral furrow moderately wide, shallow in its central (exsag.) part, better developed along anterior and posterior thirds of the palpebral lobes. Eye ridges elevated, well developed, nearly straight, moderately posteriorly directed from near glabella, slightly less elevated and narrower than palpebral lobes, on internal moulds bilobate due to a faint furrow located slightly anterior to the center and with a progressively better impression towards the contact with the glabella, developing into slightly lobate swellings near axial furrow; dividing furrow appears to have its counterpart as a faint and short furrow near the anterolateral corners of the glabella. Anterior branches of the facial suture slightly divergent from anterior ends of the ocular suture, straight for approximately the adaxial two-thirds, reaching its maximum distance just behind the mid-length of the anterior area, then curving adaxially; posterior branches diverge strongly, nearly posteriorly directed before curving towards the posterior cranidial margin. Posterolateral projection narrow and long (tr.). Posterior border moderately convex, with a slight double curvature, expanding and flattening distally; posterior border furrow well defined, moderately broad and moderately deep (Figs. 3, 4).
Librigenae of moderate size, length (excluding the doublure) 7.5–16.5 mm (n = 9), width approximately 40% of the maximum librigenal length excluding the genal spine; lateral margin almost straight and extended anteriorly (Fig. 5H–N). Librigenal field moderately convex and wide, subtrapezoidal in outline. Lateral border narrow and slightly concave; lateral border furrow developed as a change in convexity along the lateral border; posterior border furrow short, fading in depth from posterior branch of suture. Eye socket low. Genal spine long and relatively slender. Doublure narrow and extremely long, extending anteriorly to the maximum length of the cephalon and curving inward along the margin of the cranidium (Fig. 5M, N). Surface of librigenal field occasionally with faint, centrifugally arranged caeca (Fig. 3A).
Hypostome of adult individuals unknow. An ovoid and natant hypostome known from one poorly preserved specimen in meraspid stage four (M4) (Fig. 4A).
Thorax of thirteen thoracic segments with conspicuous pleural spine. Length and width of axial rings increasing in the first five segments and then reduced progressively posteriorly (Figs. 3A, 5A, B). Thoracic pleura moderately wide, transverse width of first five axial ring about one-third total width of segments, growing in relative width to ca. 40% in subsequent segments; greatest width (tr.) at segments three to five, with slightly but increasingly backward-directed pleurae in the posterior three or four thoracic segments; posterior pleural band equal in length to the anterior pleural band. Pleural furrow moderately deep and long, form posteriorly bent and narrow lenticular furrows, extending into the pleural spines. Pleural tips extend into elongate, progressively posteriorly directed spines, length (exs.) increasing in the first five or six segments and decreasing posteriorly. Surface of thoracic pleurae smooth, axial ring bears median nodes.
Pygidium relatively small, length 2.7–5.5 mm (n = 8), width/length ratio ca. 0.7; semicircular posterior margin slightly curved terminally (Fig. 5A–G). Axis moderately tapered, mid-width 80% of the anterior axial width, mid-width 35% of the maximum pygidial width; length 65% of the pygidial length (sag.); three to four weakly convex axial rings, inter-ring furrows shallow; terminal axial piece small, smoothly triangular, with a faint, posteriorly fading postaxial ridge. Pleural regions slightly convex; with three pleural furrows of moderately width and depth, curving obliquely posteriorly; interpleural furrows indistinct. Posterior margin relatively wide (expressed on surface moulds), almost flat. Doublure of uniform width (visible impressions of its proximal rim on internal moulds) (Fig. 5E, F).
Ontogeny.—Data collected from cranidia ranging from 1 to 15.4 mm in length indicate that after the cranidial length reaches 5mm, the ratio of anterior area length (exs.) to cranidial length (sag.) and the ratio of preglabellar area length to anterior area length tend to be constant (50–65% and 40–62%, respectively). We therefore place the end of the meraspid stage (Figs. 3D, 4A–F) and the onset of the holaspid stage (Figs. 3A–C, E–I, 4G–Q) at this transition, even though we did not find any proven meraspid specimens except for one exoskeleton belonging to meraspid stage four (M4) (Fig. 4A).
The primary ontogenetic trends may be summarized as follows (Fig. 6): (i) Glabellar outline subrectagnular in meraspids, moderately tapering in holaspids, with the ratio of glabellar width (across S1) to the cranidial width across same transverse line remaining constant (approximately 0.3); frontal lobe of glabella subdivided in large individuals and slightly curved in juveniles (Fig. 3D). (ii) The anterior area increases in length, with the ratio of the length of the anterior area to the length of the cranidium (sag.) increasing from 20% in the earliest meraspid stage to approximately 57% in holaspids, and the ratio of the length of the preglabellar area to the length of the anterior area (sag.) decreasing from nearly 100% in the earliest meraspid stage to approximately 50% in holaspids. (iii) Anterior border relatively broad and upturned in M4, then narrowing during the holaspid period; border furrow relatively deep in M4 but almost disappearing in later stages. (iv) Pygidium changes from button shaped with a narrow pleural region to semicircular in shape with a broader pleural region.
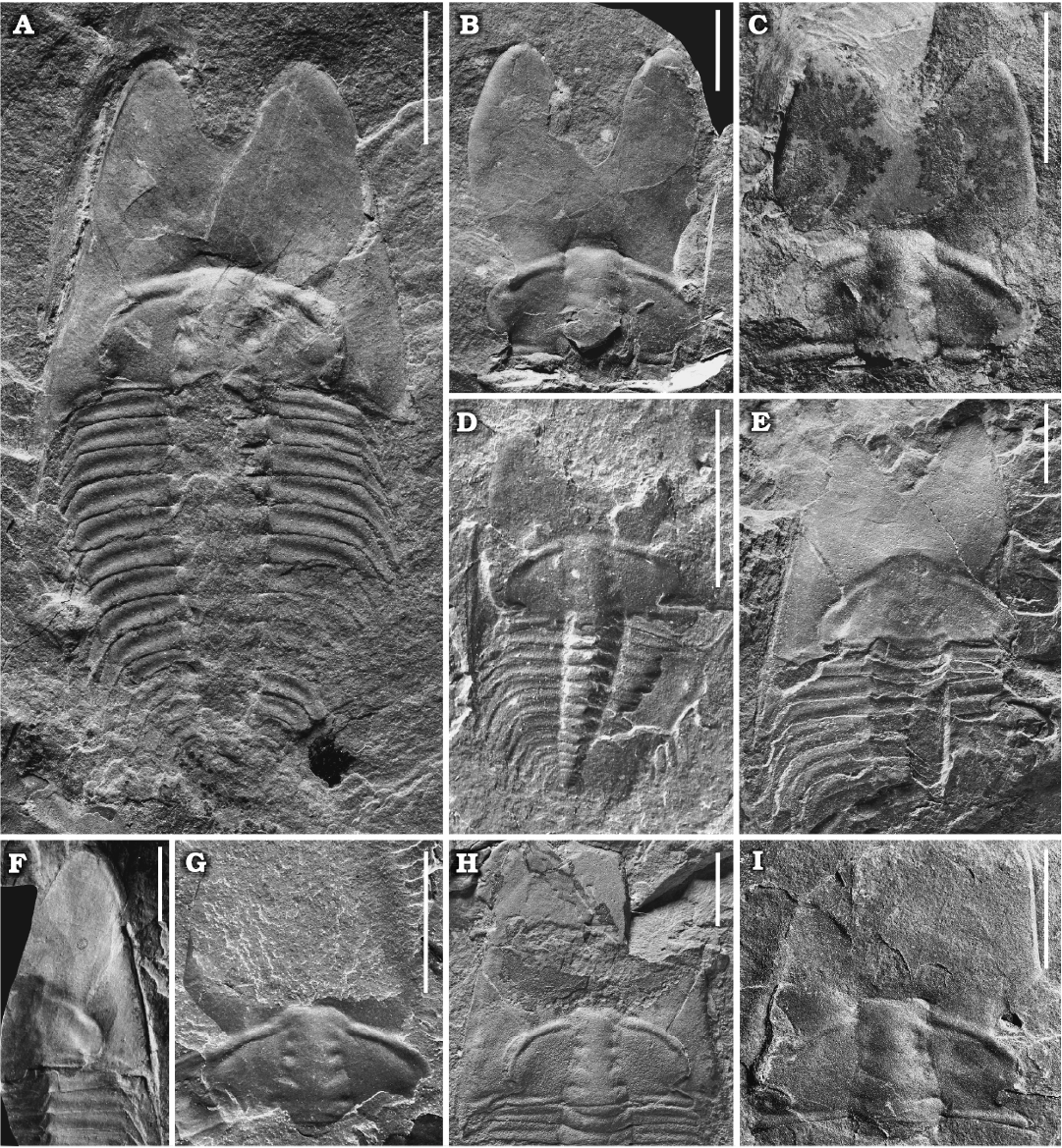
Fig. 3. A ptychoparioid trilobite Phantaspis auritus gen. et sp. nov. from the middle Cambrian (Miaolingian, Wuliuan) Mantou Formation, Longgang section, Shandong Province, North China. A. NIGPAS 172104 (holotype), nearly complete exoskeleton. B. NIGPAS 172105 (paratype), nearly complete cranidium. C. NIGPAS 172106 (paratype), external mould of a nearly complete cranidium. D. NIGPAS 172107 (paratype), external mould of a nearly complete, holaspid-stage exoskeleton. E. NIGPAS 172108 (paratype), external mould of a nearly complete cephalon. F. NIGPAS 172109 (paratype), internal mould of specimen in E. G–I. NIGPAS 172111, 172110, 172112, cranidia. Scale bars 5 mm.
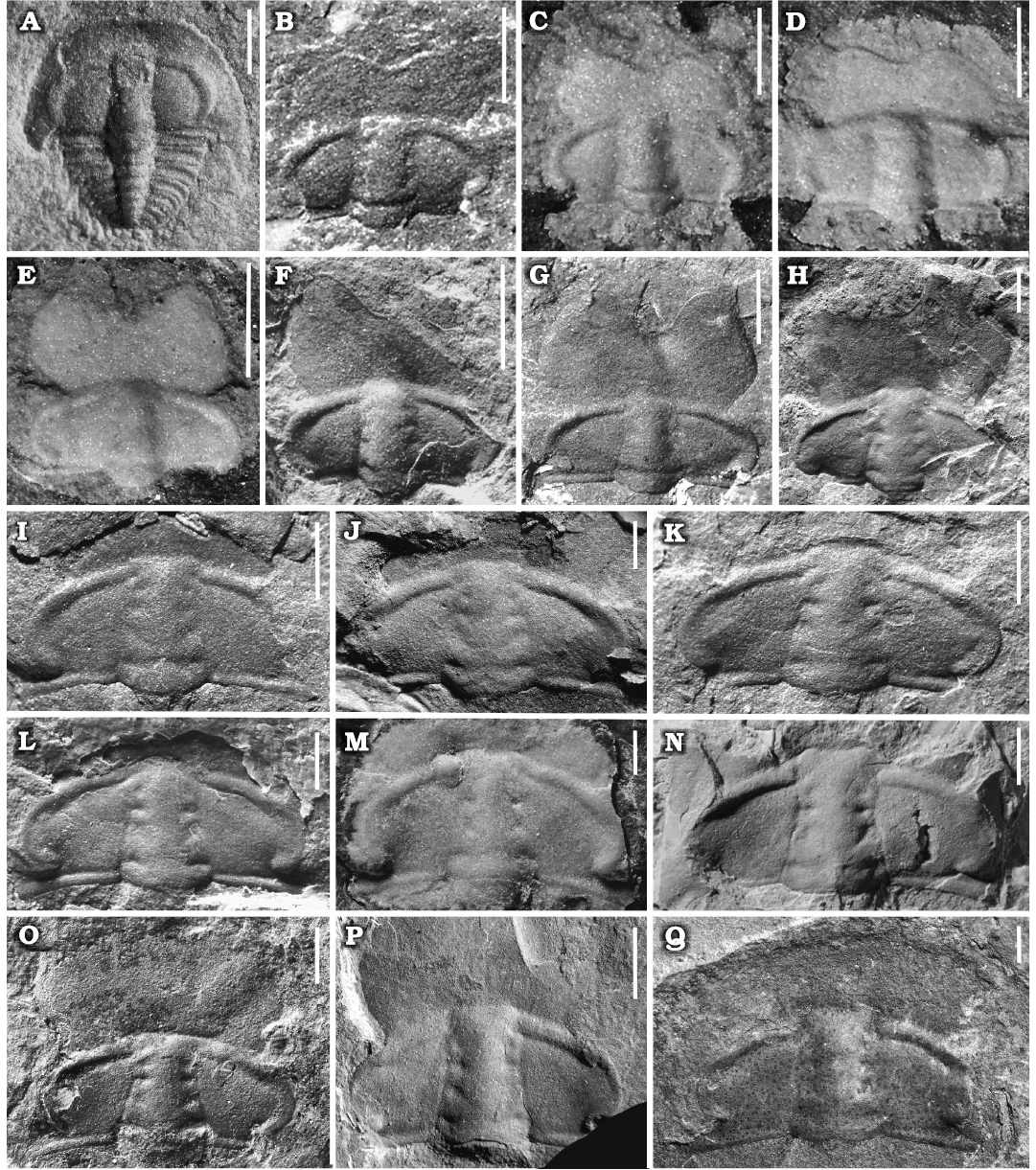
Fig. 4. Ontogenetic series of a ptychoparioid trilobite Phantaspis auritus gen. et sp. nov. from the middle Cambrian (Miaolingian, Wuliuan) Mantou Formation, Longgang section, Shandong Province, North China. A. NIGPAS 172113, complete exoskeleton in meraspid degree 4. B–F. Complete meraspid stage cranidia. B–E. NIGPAS 172114a–d, respectively. F. NIGPAS 172116. G–Q. Holaspid cranidia. G. NIGPAS 172115. H–K. NIGPAS 172117–172120, respectively. L. NIGPAS 172123. M, N. NIGPAS 172121, 172122, respectively. O. NIGPAS 172124. P. NIGPAS 172106. Q. NIGPAS 172125. Scale bars: A, 0.5 mm; B–D, I, J, 1 mm; E–H, K–Q, 2 mm.
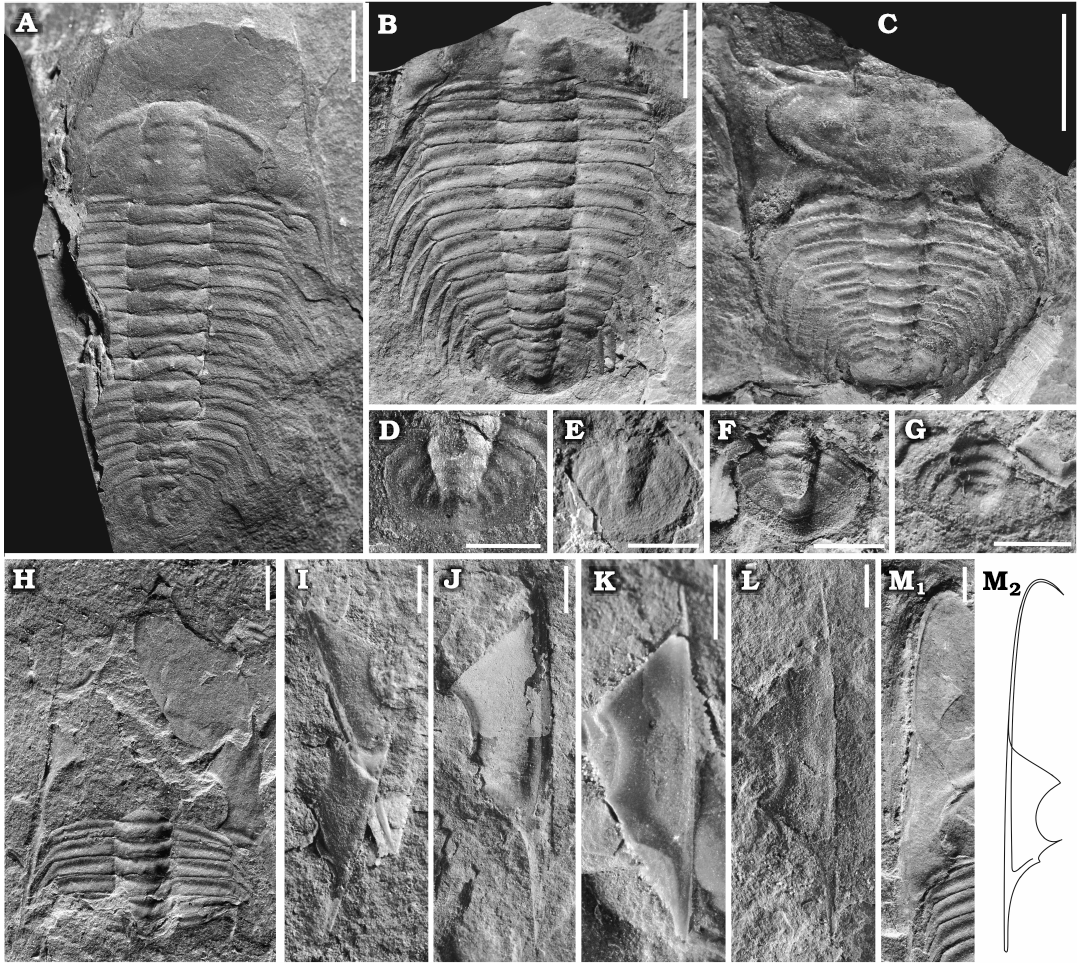
Fig. 5. A ptychoparioid trilobite Phantaspis auritus gen. et sp. nov. from the middle Cambrian (Miaolingian, Wuliuan) Mantou Formation, Longgang section, Shandong Province, North China. A. NIGPAS 172126 (paratype), nearly complete exoskeleton. B. NIGPAS 172127 (paratype), incomplete exoskeleton. C. NIGPAS 172128 (paratype), nearly complete exoskeleton showing enrollment. D–G. Pygidium, NIGPAS 172129. 172130, 172115, 172131, respectively. H. NIGPAS 172132, moult assemblage consisting of a disarticulated cranidium, librigenae, and thoracic segments. I–M. Librigenae, NIGPAS 172133, 172134, 172135, 172136, 172104, respectively; M2, reconstruction of the librigena in M1. Scale bars A–C, 5 mm; D–M, 2 mm.
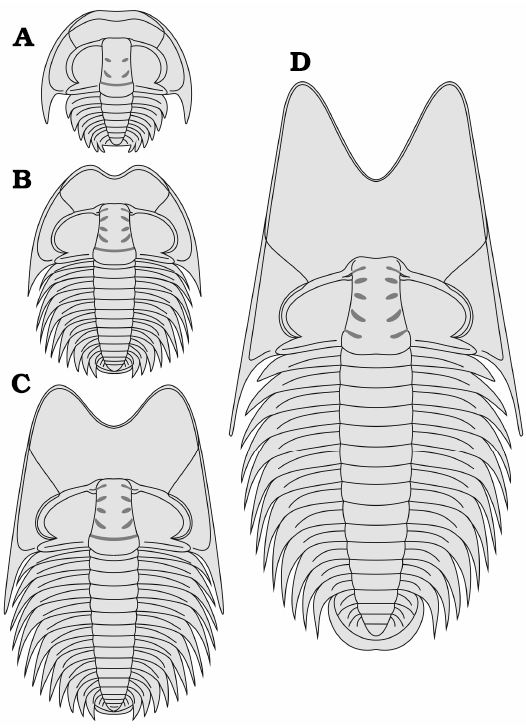
Fig. 6. Reconstructions of the dorsal exoskeleton and life cycle of Phantaspis auritus gen. et sp. nov. from the Cambrian (Miaolingian, Wuliuan) Mantou Formation, Longgang section, Shandong Province, North China. A. Meraspid degree 4. B. Late meraspid stage. C. Early holaspid stage. D. Late holaspid stage.
Remarks.—The monotypic Phantaspis gen. nov. differs from species of all other trilobite genera in having a deep sagittal indentation of the anterior area in later growth stages. Based on the combination of the tapering glabella, the outline of the preglabellar field, opisthoparian sutures and natant hypostome, this new taxon is certainly a member of the Ptychopariida. Strongly developed eye ridges, clearly convex palpebral lobes and relatively large pygidium of Phantaspis assign it to the Superfamily Ptychopariacea. Moreover, it is differentiated from roughly comparable ptychoparioids trilobites by its elliptical lateral glabellar pits, the narrow border, and the nearly straight librigenal margin.
In addition to the distinct autapomorphies, Phantaspis combines the characters known from several families in the Superfamily Ptychopariacea. In the morphology of the glabellar furrows and eye ridges, Phantaspis is somewhat similar to Tengfengia Hsiang, 1962 (Tengfengiidae Chang, 1963) from the Hsuchuangian Stage of North China, which differs from the Phantaspis in the morphology of the anterior area of the cranidium, the shape of the glabella, and the width of the palpebral area (60% of the cranidial width in Tengfengia). Lioparella Kobayashi, 1937 (Proasaphiscidae Chang, 1963) from the Hsuchuangian Stage of North China has certain features in common with Phantaspis, but there are also clear differences between these genera in the morphology of the anterior area, glabella and pygidium. Holanshania Tu in Wang et al., 1956 and Proacanthocephala (Qiu in Qiu et al., 1983) (Holanshaniidae) from the Hsuchuangian Stage also share enlarged anterior cephalic areas with Phantaspis, but the broader palpebral area and narrow anterior border in Phantaspis make them easy to distinguish. Like Phantaspis, Kochiella Poulsen, 1927 (?Ptychopariidae Matthew, 1887) from Laurentian North America has elevated eye lobes and a broad (tr.) palpebral area (Sundberg and McCollum 2002). However, Kochiella bears little resemblance to Phantaspis in other characters. Salankanaspis Egorova, 1970 (Ptychopariidae) from the Amgan Stage (corresponding mostly to the Wuliuan Stage) of the Siberian Platform is similar to Phantaspis in the morphology of the cranidium, but in all other respects there is little similarity between these two genera. Except for the extended anterior border, Salankanaspis and two other Siberian genera, Olenekina Egorova, 1970 and Regius Egorova, 1970, are similar in morphology to Kounamkites Lermontova in Chernysheva et al., 1956 (Ptychopariidae), a genus endemic to deep water areas of the Siberian plate. Except for tengfengiids, holanshaniids, proasaphiscids and ptychopariids compared above, dokimocephalids from the Furongian of Laurentia also have a specialized anterior area/anterior border (Westrop et al. 2010), but other characters indicate that these groups are not closely related to Phantaspis.
The clearly convex crescent-shaped palpebral lobes, conspicuous spinose thoracic segments and posteriorly fading postaxial ridge of Phantaspis are somewhat reminiscent to Proasaphiscidae which may include the Tengfengiidae and Holanshaniidae (Jell and Adrain 2003), but these characters just partly represent the diagnosis of proasaphiscids, and the systematic relationships of several exotic characters of Phantaspis remain uncertain. Therefore, we are unable to provide a confident family for Phantaspis in this study until further phylogenetic analysis appears.
Stratigraphic and geographic range.—Longgang section, Shandong Province, North China, Sunaspis laevis–Sunaspidella rara Zone of the Mantou Formation.
Discussion
Although trilobites had not yet achieved the peak of morphological disparity during the Cambrian (Stubblefield 1959; Whittington 1966; Foote 1990, 1991, 1993; Fortey and Owens 1991a, 1997; Webster 2007), many specialized designs already appeared in this period, including most of the common morphotypes (Fortey and Owens 1990a). Evolutionary trends such as eye loss (Álvaro and Vizcaino 2003) and effacement (see Giordanella) appeared repeatedly. Many previously known Cambrian specialized designs were repeated in post-Cambrian trilobites (Fortey and Owens 1990a). Nevertheless, P. auritus represents a form of specialization in a Cambrian trilobite that was apparently not repeated in any younger trilobites. This indicates that in addition to the morphotypes prevalent in post-Cambrian, different adaptive specializations may have been developed among the Cambrian trilobites.
Some unusual trilobites have appeared in the early to middle Cambrian such as emuellids, burlingiids, and damesellids. However, the extraordinary examples of specialized cephalic structures in trilobites tended to be such features as prominent spines, or convex areas on the anterior border (e.g., Chang 1963; Egorova 1970; McNamara 1978; Lin 1984). Phantaspis auritus, however, displays a new and unique example of cephalic specialization among Cambrian trilobites. The specialization is manifested in the anterior area rather than the border, indicating the potential of the anterior area in changing the outline of cranidia.
The extended cephalon of P. auritus is reminiscent of certain sediment feeders with a specialized cephalon, for example species of Harpina and Trinucleidae, which show a thickened anterior border and a broad cephalic doublure. However, in P. auritus the anterior border was not thickened as those of the above groups. This may explain why the anterior area of most (43 out of 55) observed cranidia are broken (Figs. 3G–I, 4H–Q, 5A). There are several examples of Ordovician olenids with narrow median cephalic doublure and nearly flat wide preglabellar areas, not underlain by doublure (and natant hypostomes), such as Hypermecaspis venulosa (Fortey, 1974) which has a nearly flat anterior like P. auritus. However, it is difficult to compare the life habits of benthic olenids, which have lived in oxygen-poor environments to the subtidal P. auritus.
Other than adaptation to a particular life habit, further possibilities should be considered. Firstly, the cephalic shape seen in P. auritus may have reduced the risk of predation by increasing the effective size of P. auritus, thus making it harder to be eaten by predators, such as other trilobites (Whittington 1950; Corbacho and López-Soriano 2012; Corbacho 2014). Alternatively, the head-to-trunk length ratio of P. auritus as well as an enrolled specimen of this species (Fig. 5C) suggest that the animal may have been able to cover the entire ventral side of the trunk with the large cephalon. Thus, P. auritus may represent another example of a complete enrollment (Bergström 1973; Ortega-Hernández et al. 2013) without caudalization. Finally, although there is no evidence of sexual dimorphism, the development and stabilization of cranidial morphology associated with sexual maturity suggests a possibility of sexual selection, similar to “beetle”-like horns known from other trilobites, which are assumed to reflect this type of selective strategy (Knell and Fortey 2005).
Conclusions
The distinctive cephalon of Phantaspis auritus gen. et sp. nov. provides valuable insights into cephalic specialization of middle Cambrian trilobites. P. auritus had an exceptionally long cephalon with a unique, strongly notched anterior area unlike that of any other known trilobite. It furthermore represents an early and very rare case of cephalic morphotypes among Cambrian trilobites. The new genus illustrates the diversity of Cambrian trilobites in morphotypes and provides an example of ptychoparioid cranidial outline variation during the middle Cambrian caused by specialisation. The unique cephalon of P. auritus may have evolved in response to sediment-feeding mode, predation, sexual selection or factors favoring a particular mode of enrollment.
Acknowledgements
We are very grateful to Gerd Geyer (University of Würzburg, Germany) and Richard Fortey (Natural History Museum, London, UK) for their constructive comments and suggestions, Heyo Van Iten (Hanover College, Hanover, USA) for improving the English of this manuscript, Jinliang Yuan (NIGPAS) for helpful suggestions and Rongrong Xing (Weifang, Shangdong, China) for assistance in the field and for collecting some of the specimens described in this study. This research was supported by grants from the Strategic Priority Research Program (B) of the Chinese Academy of Sciences (XDB26000000, XDB18000000), the National Natural Science Foundation of China (41921002, 41902014), and the State Key Laboratory of Palaeobiology and Stratigraphy (20181105).
References
Álvaro, J.J. and Vizcaïno, D. 2003. The conocoryphid biofacies, a benthic assemblage of normal-eyed and blind trilobites. Special Papers in Palaeontology 70: 127–140.
Bergström, J. 1973. Organization, life, and systematics of trilobites. Fossils and Strata 2: 1–69.
Chang, W.-T. [Zhang, W.T.] 1963. A classification of the Lower and Middle Cambrian trilobites from northern and northeastern China, with descriptions of new families and new genera [in Chinese with English summary]. Acta Palaeontologica Sinica 11: 447–487.
Chernysheva, N.E. [Černyševa, N.E.], Egorova, L.I., Ogienko, L.V., Poletaeva, O.K., and Repina, L.N. 1956. Materials on palaeontology. New families and genera. Class Trilobita [in Russian]. Trudy vsesoûznogo Navčno-Issledovatel’skogo Geologičeskogo Instituta, Novaâ seriâ 12: 145–182.
Chough, S.K., Lee, H. S., Woo, J., Chen, J.-T., Choi, D.K., Lee, S.B., Kang, I., Park, T., and Han, Z.-Z. 2010. Cambrian stratigraphy of the North China Platform: revisiting principal sections in Shandong Province, China. Geosciences Journal 14: 235–268. Crossref
Corbacho, J. 2014. Lanceaspis hammondi n. gen., n. sp.—a questionable early Asteropyginae (Trilobita) from the Pragian of the Morocco. Batalleria 20: 3–10.
Corbacho, J. and López-Soriano, F.J. 2012. A new asaphid trilobite from the Lower Ordovician (Arenig) of Morocco. Batalleria 17: 1–10.
Egorova, L.I. 1970. New Middle Cambrian trilobites from the north of the Siberian Platform [in Russian]. Paleontologičeskij žurnal 4: 72–76.
Foote, M. 1988. Survivorship analysis of Cambrian and Ordovician trilobites. Paleobiology 14: 258–271. Crossref
Foote, M. 1990. Nearest-neighbor analysis of trilobite morphospace. Systematic Zoology 39: 371–382. Crossref
Foote, M. 1991. Morphologic patterns of diversification: Examples from trilobites. Palaeontology 34: 461–485.
Foote, M. 1993. Discordance and concordance between morphological and taxonomic diversity. Paleobiology 19: 185–204. Crossref
Fortey, R.A. 1974. The Ordovician trilobites of Spitsbergen. I. Olenidae. Norsk Polarinstitutt Skrifter 160: 1–129.
Fortey, R.A. 2001. Trilobite systematics: the last 75 years. Journal of Paleontology 75: 1141–1151. Crossref
Fortey, R.A. and Owens, R.M. 1990a. Evolutionary radiations in the Trilobita. In: P.D. Taylor and G.P. Larwood (eds.), Major Evolutionary Radiations. Systematics Association Special Vol. 42, 139–164. Clarendon Press, Oxford.
Fortey, R.A. and Owens, R.M. 1990b. Trilobites. In: K.J. McNamara (ed.), Evolutionary Trends, 121–142. Belhaven Press, London.
Fortey, R.A. and Owens, R.M. 1997. Evolutionary history. In: R.L. Kaesler (ed.), Treatise on Invertebrate Paleontology. Part O. Arthropoda 1, 249–287. Geological Society of America and University of Kansas Press, Lawrence.
Geyer, G. 2019. A comprehensive Cambrian correlation chart. Episodes Journal of International Geoscience 42: 321–332. Crossref
Hsiang, L.-W. [Xiang, L.W.] 1962. Some trilobites from the Middle Cambrian of Western Honan [in Chinese with English summary]. Acta Palaeontologica Sinica 10: 394–401.
Jell, P.A. and Adrain, J.M. 2003. Available generic names for trilobites. Memoirs of the Queensland Museum 48: 331–551.
Knell, R.J. and Fortey, R.A. 2005. Trilobite spines and beetle horns: sexual selection in the Palaeozoic? Biology Letters 1: 196–199. Crossref
Kobayashi, T. 1937. Restudy of Dames’ types of the Cambrian trilobites from Liaotung. Journal of the Geological Society of Japan 44: 421–437. Crossref
Lin, T.-R. 1984. A new genus of middle Cambrian trilobite from Zaozhuang, Shandong [in Chinese with English summary]. Acta Palaeontologica Sinica 6: 738–741.
Matthew, G.F. 1887. Illustrations of the fauna of the St. John Group, 4. On the smaller-eyed trilobites of Division I., with a few remarks on the species of the higher divisions of the group. Canadian Record of Science 2: 357–363.
McNamara, K.J. 1978. Paedomorphosis in Scottish olenellid trilobites (early Cambrian). Palaeontology 21: 635–655.
Ortega-Hernández, J., Esteve, J., and Butterfield, N.J. 2013. Humble origins for a successful strategy: complete enrolment in early Cambrian olenellid trilobites. Biology Letters 9: 20130679. Crossref
Poulsen, C. 1927. The Cambrian, Ozarkian, and Canadian faunas of northwest Greenland. Meddelelser om Grønland 70: 233–343.
Qiu, H.-A., Lu, Y.-H., Zhu, Z.-L., Bi, D.-C., Lin, T.-R., Zhou, Z.-Y., Zhang, Q.-Z., Qian, Y.-Y., Ju, T.-Y., Han, N.-R., and Wei, X.-Z. 1983. Trilobita [in Chinese]. In: Nanjing Institute of Geology and Mineral Resources, Ministry of Geology and Mineral Resources (ed.), Palaeontological Atlas of East China. Vol. 1. Early Palaeozoie, 28–254. Geological Publishing House, Beijing.
Ren, G.-Y., Wei, C.-T., and Yuan, J.-L. 2017. Revision of Shantungia (Trilobita: Damesellidae) from the Kushan Formation (Guzhangian), Shandong, China. Palaeoworld 26: 423–430. Crossref
Stubblefield, C.J. 1959. Evolution in trilobites. Quarterly Journal of the Geological Society of London 115: 145–162. Crossref
Sun, Z.-X., Zeng, H., and Zhao, F.-C. 2020. First occurrence of the Cambrian arthropod Sidneyia Walcott, 1911 outside of Laurentia. Geological Magazine 157: 405–410. Crossref
Sundberg, F.A. and McCollum, L.B. 2002. Kochiella Poulsen, 1927, and Hadrocephalites new genus (Trilobita: Ptychopariida) from the early middle Cambrian of western North America. Journal of Paleontology 76: 76–94. Crossref
Swinnerton, H.H. 1915. Suggestions for a revised classification of trilobites. Geological Magazine (New Series) 6: 487–496, 538–545. Crossref
Walch, J.E.I. 1771. Die Naturgeschichte der versteinerungen zur erläuterung der Knorrischen Sammlung von Merkwürdigkeiten der Natur. Vol. 3. 235 pp. Felßecker, Nürnberg.
Wang, L.-W., Zhang, R.-S., Chang, A.-Z., Yan, E.-Z., and Wei, X.-Y. 1956. Handbook of Index fossils of China [in Chinese]. 669 pp. New Knowledge Publishing House, Shanghai.
Webster, M. 2007. A Cambrian peak in morphological variation within trilobite species. Science 317: 499–502. Crossref
Westrop, S.R., Poole, R.A.W., and Adrain, J.M. 2010. Systematics of Dokimocephalus and related trilobites from the Late Cambrian (Steptoean; Millardan and Furongian series) of Laurentian North America. Journal of Systematic Palaeontology 8: 545–606. Crossref
Whittington, H.B. 1950. A monograph of the British trilobites of the family Harpidae. Palaeontographical Society Monographs 103: 1–55.
Whittington, H.B. 1966. Phylogeny and distribution of Ordovician trilobites. Journal of Paleontology 40: 696–737.
Whittington, H.B. and Kelly, S.R.A. 1997. Morphological terms applied to Trilobita. In: R.L. Kaesler (ed.), Treatise on Invertebrate Paleontology. Part O. Arthropoda 1, Trilobita, Revised. Vol. 1: Introduction, Order Agnostida, Order Redlichiida, 313–329. Geological Society of America, Boulder, Colorado and University of Kansas Press, Lawrence.
Yuan, J.-L., Li, Y., Mu, X.-N., Lin, J., and Zhu, X.-J. 2012. Trilobite Fauna of the Changhia Formation (Cambrian Series 3) from Shandong and Adjacent Area, North China [in Chinese with English summary]. 757 pp. Science Press, Beijing.
Yuan, J.-L., Zhu, Z.-L., and Zhang, W.-T. 2015. Cambrian Stratigraphy and Trilobite Fauna in Southern and Western Marginal Parts of the Ordos Platform [in Chinese with English summary]. 534 pp. Science Press, Beijing.
Zhang, Z.-Q. 1996. Lithostratigraphy of Cambrian [in Chinese]. In: Shandong Province Bureau of Geology and Mineral Resources (ed.), Lithostratigraphy of Shandong Province, 109–154. China University of Geosciences Press, Wuhan.
Zhu, M.-Y., Yang, A.-H., Yuan, J.-L., Li, G.-X., Zhang, J.-M., Zhao, F.-C., Ahn, S.-Ye., and Miao, L.-Y. 2019. Cambrian integrative stratigraphy and timescale of China. Science China Earth Sciences 62: 25–60. Crossref
Acta Palaeontol. Pol. 65 (4): 709–718,
2020 https://doi.org/10.4202/app.00753.2020