
Palaeoecology of tropical marine invertebrate assemblages from the Late Triassic of Misurina, Dolomites, Italy
IMELDA M. HAUSMANN, ALEXANDER NÜTZEL, VANESSA JULIE RODEN, and MIKE REICH
Hausmann, I.M., Nützel, A., Roden, V.J., and Reich, M. 2021. Palaeoecology of tropical marine invertebrate assemblages from the Late Triassic of Misurina, Dolomites, Italy. Acta Palaeontologica Polonica 66 (1): 143–192.
Two marine invertebrate fossil assemblages from the Late Triassic Cassian Formation (Dolomites, northern Italy) were examined to assess their diversity and palaeoecology. Surface and bulk samples from the localities Misurina Landslide and Lago Antorno were taken and analysed separately. Both benthic assemblages are relatively similar in taxonomic composition. Gastropods form the most abundant and diverse group, followed by bivalves. Disarticulated echinoderm ossicles are also common in the bulk sample from Misurina Landslide, but they are rare at Lago Antorno. The Misurina Landslide outcrop has yielded two echinoderm Palaeozoic holdovers, the ophiocistioid Linguaserra triassica and plates of putative proterocidarids, supporting the earlier hypothesis that such basins acted as refugia. The gastropod species Coelostylina conica, Prostylifer paludinaris, and Ampezzopleura hybridopsis are characteristic elements of both assemblages. The gastropod Jurilda elongata, however, is the most abundant species at Misurina Landslide, whereas juveniles of the gastropod species Dentineritaria neritina dominate the assemblage from Lago Antorno. Newly described gastropod taxa are Angulatella bizzarinii Nützel and Hausmann gen. et sp. nov., Bandellina compacta Nützel and Hausmann sp. nov., and Ampezzogyra angulata Nützel and Hausmann sp. nov. Fifty-seven invertebrate species were found in the bulk sample from Misurina Landslide and 26 species were recovered from the bulk sample from Lago Antorno. However, sample size from Lago Antorno was much smaller than that from Misurina. Diversity indices (Shannon, Simpson, Berger-Parker) show similar moderate diversities in both assemblages. Rarefaction curves and rank-abundance distributions also point to very similar diversities and ecological structures of the fossil assemblages. Both assemblages are autochthonous or parautochthonous, stemming from basinal, soft-bottom habitats. Their taxonomic composition differs significantly from that of other faunas known from the Cassian Formation. The tropical marine Cassian palaeoecosystem was highly complex and its diversity is still far from being fully explored.
Key words: Mollusca, Echinodermata, diversity, small body size, Ladinian, Carnian, Cassian Formation, Alps.
Imelda M. Hausmann [hausmann.i@snsb.de], SNSB, Bavarian State Collection of Palaeontology and Geology; and Department of Earth and Environmental Sciences, Palaeontology & Geobiology, Ludwig-Maximilians-Universität München, Richard-Wagner-Str. 10, 80333 München, Germany.
Alexander Nützel [nuetzel@snsb.de], SNSB, Bavarian State Collection of Palaeontology and Geology; Department of Earth and Environmental Sciences, Palaeontology & Geobiology; and GeoBio-Center LMU, Richard-Wagner-Str. 10, 80333 München, Germany.
Vanessa Julie Roden [vanessa.roden@fau.de], GeoZentrum Nordbayern, Section Paleobiology, University Erlangen-Nürnberg, Loewenichstr. 28, 91054 Erlangen, Germany; and SNSB – Staatliche Naturwissenschaftliche Sammlungen Bayerns, Menzinger Str. 67, 80638 München, Germany.
Mike Reich [reich@snsb.de], SNSB, Bavarian State Collection of Palaeontology and Geology; Department of Earth and Environmental Sciences, Palaeontology & Geobiology, Ludwig-Maximilians-Universität München; and GeoBio-Center LMU, Richard-Wagner-Str. 10, 80333 München, Germany.
Received 28 July 2019, accepted 12 August 2020, available online 19 January 2021.
Copyright © 2021 I.M. Hausmann et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
The Cassian Formation sensu lato (late Ladinian–middle Carnian) has yielded by far the most diverse marine invertebrate fauna known from the early Mesozoic. 1421 nominate invertebrate species have been described at present, with Mollusca (950 species) being the most diverse phylum (Roden et al. 2020a). The main reason for this very high diversity is a high primary alpha and beta diversity (Roden et al. 2018, 2020a) and the fact that the poorly lithified marls qualify the Cassian Formation as a liberation lagerstätte sensu Roden et al. (2020a). This lagerstätten type can be defined by an exceptionally good preservation plus a high abundance of fossils which can be easily extracted from the source sediment that is poorly lithified (Roden et al. 2020a). The vast majority of the literature on Cassian Formation fossils addresses taxonomic or stratigraphic questions, and in most cases refers to particular fossil groups. In contrast, only few quantitative datasets covering entire invertebrate assemblages have been provided including relative abundances (Fürsich and Wendt 1977; Nützel and Kaim 2014; Hausmann and Nützel 2015). However, abundance data are needed for sample standardisation methods used in biodiversity estimations and palaeoecological considerations (e.g., Magurran 2004; Kowalewski and Novack-Gottshall 2010). It is therefore desirable to provide such data sets of high quality, i.e., taxonomic descriptions, count data and discussion of the taxa present in a sample or fauna. The taxa present in the assemblages should be illustrated and described so that identifications are easily falsifiable. Moreover, it should be stated how many specimens were collected in the field by surface sampling and how many were obtained by bulk sampling and sieving as these methods produce different results (Hausmann and Nützel 2015; Forcino and Stafford 2020).
A considerable variety of fossil assemblages has been found in the Cassian Formation, ranging from highly diverse shallow-water reef dweller assemblages to various low-diversity soft-bottom assemblages. In addition to differences in diversity, rank-abundance distributions and taxonomic composition differ considerably between the studied assemblages (Fürsich and Wendt 1977; Nützel and Kaim 2014; Hausmann and Nützel 2015; Roden et al. 2020a). Previous quantitative datasets suggest that the entire complex tropical marine ecosystem of the Cassian Formation is far from being fully explored (Fürsich and Wendt 1977; Nützel and Kaim 2014; Hausmann and Nützel 2015). The present study quantitatively and systematically investigated the two hitherto unknown invertebrate fossil assemblages from the vicinity of Lake Misurina (Dolomites, northern Italy; Fig. 1).

Fig. 1. Maps of northern Italy (A, B) showing the sampling localities Misurina Landslide and Lago Antorno (marked with stars). C. Upper part of Misurina Landslide locality where the samples were taken. D. Detailed view of the sediments at the Misurina Landslide locality.
The studied assemblages are strongly dominated by gastropods as in the other assemblages from the Cassian Formation (e.g., Nützel and Kaim 2014; Hausmann and Nützel 2015; Roden et al. 2020a). The taxonomy of the gastropods is intricate, due to a complex research history. Therefore, we also focus on systematic description and illustration of the gastropod fauna from our samples. The study of the exceptionally preserved Cassian biota helps to understand the evolution of diversity at a large scale, i.e., addressing the question as to whether early Mesozoic tropical marine ecosystems were as diverse as modern ones.
Institutional abbreviations.—NHMW, Natural History Museum, Vienna, Austria; NHMUK, Natural History Museum, London, UK; PZO, Naturmuseum Südtirol, Bolzano, Italy; SNSB-BSPG, Staatliche Naturwissenschaftliche Sammlungen Bayerns-Bayerische Staatssammlung für Paläontologie und Geologie, Munich, Germany.
Nomenclatural acts.—This published work and the nomenclatural acts it contains have been registered in ZooBank urn:lsid:zoobank.org:pub:52C87838-856E-468B-9215-1065205FA02A.
Geological setting
Surface and bulk samples were taken near Lago Antorno at a landslide scar (GPS 46°35’39.8”N 12°15’39.6”E). In addition, several samples were taken at another landslide scar near the locality Misurina (named here Misurina Landslide; GPS 46°35’41.6”N 12°15’34.6”E) (Fig. 1). The latter sampling site corresponds to outcrop number 4 of Bizzarini and Laghi (2005). The section exposed in the scar is in situ and consists largely of marl and claystone with a few marly limestone beds (Fig. 1D). In the upper part of this landslide area, a relatively thick marly limestone bed is exposed, situated a few metres above our sampling spot (Fig. 1C).
Both sites, Lago Antorno and Misurina Landslide, expose greyish to brownish claystone, which is poorly lithified. They belong to the Cassian Formation sensu lato (late Ladinian–middle Carnian) as is indicated by several typical invertebrate species (see below; Carnian: Julian, ?Austrotrachyceras austriacum Ammonite Biozone; see Reich et al. 2018: fig. 1; Roden et al. 2020a: table 1). The Cassian Formation sensu lato also includes those fossil sites which belong to the A. austriacum and Tropites dilleri ammonite biozones, which are part of the Heiligkreuz Formation that overlies the Cassian Formation (Keim et al. 2006; Urlichs 2017; Nose et al. 2018; Reich et al. 2018; Roden et al. 2020a). We do not share the view that our locations as well as others such as Misurina Skilift, Alpe di Specie etc. represent the Heiligkreuz Formation because at its type location, the Heiligkreuz Formation differs considerably from the deposits studied herein regarding lithology and fauna. The Heiligkreuz Formation at its type locality consists of a limestone marl alternation with superabundant ostracods and with the gastropod Ptychostoma sanctaecrucis and the bivalve Unionites muensteri dominating the macrofauna. It is interpreted as a brackish water deposit (Koken 1913; Keim et al. 2006: “Member A”). The material studied herein and other exposures such as Misurina Skilift and Alpe di Specie are assigned to the Upper Cassian Formation as was also done by others (e.g., Bizzarini and Laghi 2005; Urlichs 2017; Nose et al. 2018; Reich et al. 2018; Roden et al. 2020a).
During the Late Triassic, the Cassian Formation was positioned in the northern tropics (at approximately 16°N) in the western Tethys region (Broglio Loriga et al. 1999; Keim et al. 2006). It constitutes marine basin sediments, which were situated between carbonate platforms (e.g., Fürsich and Wendt 1977; Bosellini 1998; Hausmann and Nützel 2015). Sea surface temperatures were comparable to modern tropical seas (e.g., Korte et al. 2005; Nützel et al. 2010a) and it has been shown for the nearby locality Misurina Skilift that seasonal fluctuations were pronounced (Nützel et al. 2010a).
Material and methods
Samples from the landslide scar near Lago Antorno stem from sampling campaigns conducted in 2008, 2010, and 2016. Samples from Misurina Landslide were collected in the upper part of the landslide scar in June 2016. A marly limestone bed is located near this sampling locality. Samples were taken from unconsolidated marls. One bulk and three surface samples from Lago Antorno and two bulk and four surface samples from Misurina Landslide were taken. Regarding surface sampling, all fossils were removed by hand from the sediment surface of both outcrops. The three surface collections from Lago Antorno were sampled in three different years. Since the upper part of the landslide scar at Misurina Landslide was too large for a complete surface sampling, areas of several square metres within this site were sampled. For the bulk samples, sediment was collected from two nearby lying spots in each outcrop down to a maximum depth of 20 cm, comprising 5–10 kg per bulk sample.
The marly bulk samples were treated with a 7% H2O2 solution and wet-sieved over mesh sizes of 5, 0.5, and 0.11 mm. For the Lago Antorno bulk sample, a 0.4 mm instead of a 0.5 mm mesh was used. All fossil remains were picked from the residues. The finest fraction, 0.11–0.5/0.4 mm, was only analysed qualitatively and was therefore excluded from the statistical analyses, since the larger size fractions were analysed quantitatively. Nevertheless, the fossil content from this size fraction is briefly discussed herein. The fossil material from bulk (>0.5/0.4 mm) and surface samples was quantified and identified to species level (although in open nomenclature if preservation was insufficient).
To account for the best possible estimation of the abundance of fossils with more than one shell part (i.e., bivalves, brachiopods, ostracods, echinoderms), we calculated the minimum number of individuals as follows: in bivalves, brachiopods, and ostracods, the estimated number of specimens was calculated as articulated fossils plus higher number of left/right or ventral/dorsal valves. Regarding echinoderms, each species was counted as being represented by one specimen as long as the number of hard parts did not exceed the number present in one specimen (see also Fürsich and Wendt 1977; Nützel and Kaim 2014; Hausmann and Nützel 2015).
To investigate evidence of predation, all shells were additionally investigated for predatory drill holes irrespective of complete or incomplete drilling. Drill holes were defined as circular bore holes similar to Recent naticid or muricid predation traces. Other traces of predatory origin, such as bite marks, were not analysed in this study.
Alpha diversity was estimated using the diversity indices Shannon, Simpson, and Berger-Parker, which were calculated in PAST version 3.20 (Hammer et al. 2001). All three indices were calculated because every diversity index has its own advantages and weaknesses. For instance, the Shannon index is more strongly influenced by the rare portion of species whereas the Simpson index is influenced more by the common species in an assemblage. The Shannon index calculates the degree of uncertainty, i.e., it estimates how uncertain it is to predict what species will be collected next. The Simpson index calculates how likely it is that two randomly chosen specimens represent two different species (e.g., Harper 1999). The Berger-Parker index is a simple indicator of dominance, calculated by the specimen number of the dominant species relative to the whole number of specimens in the analysed sample (Hammer 2019). Rarefaction curves were performed with PAST and R version 3.5.3 (R Core Team 2019) using the package “vegan” (Oksanen et al. 2019). Best model fit for rank-abundance distributions, using AIC values, was calculated with the package “vegan” (Oksanen et al. 2019). Rank-abundance distributions are helpful, because they easily illustrate evenness and ecological structure of an assemblage (e.g., Bastow 1991). The R package “sads” (Prado et al. 2018) was used for plotting rank-abundance distributions. Beta diversity was calculated based on dissimilarities in community composition among samples from this study and previous works (Nützel and Kaim 2014; Hausmann and Nützel 2015) using identified species which were assigned to species names, but excluding species left in open nomenclature. Beta diversity is expressed as pairwise proportional dissimilarity (PPD), which is relatively insensitive to unequal sample sizes (Krebs 1989) and is calculated as djk = 1 – ∑min(xij, xik), with xij and xik being the proportions of species abundance in each sample. Beta diversity calculations were performed using R version 3.5.0 (R Core Team 2016) and the package “vegan” (Oksanen et al. 2016).
Results
Taxonomic composition.—57 species were found in the bulk sample from Misurina Landslide and 26 species in that from Lago Antorno. Fossil assemblages at both sites are dominated by gastropods (Tables 1 and 2, Fig. 2). The collection from Misurina Landslide consists of 567 specimens of gastropods from 20 species. In comparison, 230 specimens representing 17 species of gastropods were found at Lago Antorno. Among these, vetigastropods are represented by only one species and one specimen found at Misurina Landslide. Bivalves make up the second-most abundant and species-rich group. Scaphopods and ostracods are rare. A single cephalopod (ammonite) was found at Misurina Landslide and a single brachiopod at Lago Antorno. Twenty-one echinoderm species were retrieved from Misurina Landslide, with echinoids and holothurians being the most diverse groups. The Lago Antorno assemblage yields a much less diverse echinoderm fauna as only two species were found in the bulk sample. Many skeletal elements, most of them belonging to echinoids, were recovered from the bulk samples from Misurina Landslide, whereas only 5 elements could be retrieved from Lago Antorno. Additionally, a single fragment of a potential hydrozoan was found in the Lago Antorno sample.
In the surface samples, gastropods again make up the most species-rich and abundant group, followed by bivalves. Fourteen and 11 gastropod species were found at Misurina Landslide and Lago Antorno, respectively, comprising 89 and 136 shells (Table 1). Eight species and 24 specimens of bivalves were found in the surface samples from Misurina Landslide, in comparison to 5 species and 10 specimens from Lago Antorno. Scaphopods are also rare and cephalopods, brachiopods, and ostracods are absent. No echinoderms were found in the surface samples from both localities, except for a single skeletal element of a cidaroid echinoid, which was found at Misurina Landslide. In addition, one small sponge fragment was found at Misurina Landslide.
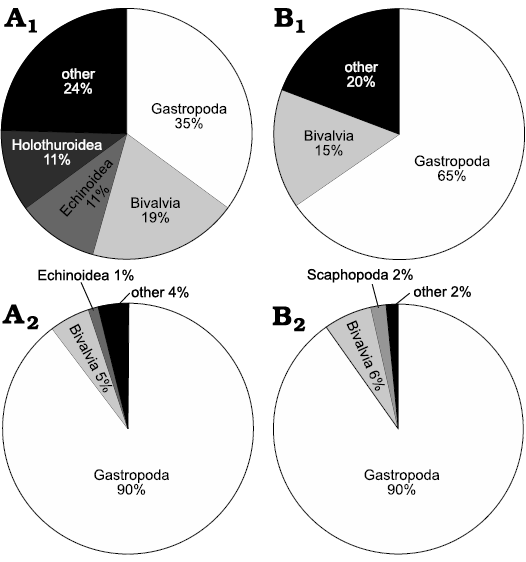
Fig. 2. Species (A1, B1) and specimen (A2, B2) proportions within higher taxa of the bulk samples from Misurina Landslide (A) and Lago Antorno (B).
Table 1. Species and abundances lists of the bulk and surface collections from Misurina Landslide and Lago Antorno. * not included in statistical analyses because of allochthonous origin and therefore not being part of the original palaeocommunity; ** not included in statistical analyses because size fraction was <0.5 mm.
|
Taxa |
Misurina Landslide bulk |
Misurina Landslide surface |
Lago Antorno bulk |
Lago Antorno surface |
|
Porifera* |
||||
|
Porifera indet.* |
|
1 |
|
|
|
Cnidaria* |
||||
|
Hydrozoa? indet.* |
|
|
1 |
|
|
Gastropoda |
||||
|
Eunema badioticum Kittl, 1891 |
|
|
|
1 |
|
“Turbo” sp. |
|
1 |
|
|
|
Hologyra? expansa (Laube, 1869) |
|
|
|
1 |
|
Neritaria mandelslohi (Klipstein, 1843) |
5 |
|
29 |
|
|
Dentineritaria neritina (Münster, 1841) |
38 |
|
68 |
|
|
Coelostylina conica (Münster, 1841) |
53 |
19 |
29 |
26 |
|
Coelostylina sp. 1 |
1 |
|
|
|
|
Coelostylina sp. 2 |
|
1 |
|
|
|
Coelochrysalis pupaeformis (Münster, 1841) |
|
2 |
|
|
|
Caenogastropoda indet. |
|
|
|
2 |
|
Helenostylina convexa Nützel and Kaim, 2014 |
1 |
2 |
6 |
4 |
|
Prostylifer paludinaris (Münster, 1841) |
128 |
50 |
24 |
81 |
|
Angulatella bizzarinii Nützel and Hausmann sp. nov. |
3 |
|
5 |
|
|
Atorcula sp. |
|
|
|
3 |
|
Atorcula canalifera (Münster, 1841) |
1 |
4 |
1 |
6 |
|
Ampezzopleura hybridopsis Nützel, 1998 |
95 |
1 |
29 |
9 |
|
Ampezzopleura bandeli Nützel, 1998 |
21 |
3 |
|
|
|
Ampezzopleurinae indet. |
1 |
|
|
|
|
Zygopleura sp. |
1 |
|
1 |
|
|
Kittliconcha aonis (Kittl, 1894) comb. nov. |
|
2 |
|
2 |
|
Tyrsoecus zeuschneri (Klipstein, 1843) |
|
|
|
1 |
|
Flemingella bistriata (Münster, 1841) |
|
1 |
|
|
|
Jurilda elongata (Leonardi and Fiscon, 1959) comb. nov. |
166 |
1 |
5 |
|
|
Promathildia cf. milierensis Zardini, 1980 |
19 |
1 |
|
|
|
Promathildia cf. decorata (Klipstein, 1843) |
2 |
|
|
|
|
Promathildia subnodosa (Münster, 1841) |
|
1 |
|
|
|
Camponaxis lateplicata (Klipstein, 1843) |
1 |
|
|
|
|
Bandellina compacta Nützel and Hausmann sp. nov. |
|
|
2 |
|
|
Alexogyra marshalli Bandel, 1996 |
|
|
7 |
|
|
Ampezzogyra angulata Nützel and Hausmann sp. nov. |
14 |
|
9 |
|
|
Sinuarbullina sp. 1 |
14 |
|
10 |
|
|
Sinuarbullina sp. 2 |
2 |
|
2 |
|
|
Sinuarbullina sp. 3 |
|
|
2 |
|
|
Gastropoda indet. 1 |
1 |
|
|
|
|
Gastropoda indet. 2 |
|
|
1 |
|
|
Bivalvia |
||||
|
Palaeonucula strigilata (Goldfuss, 1837) |
10 |
8 |
2 |
|
|
Nuculana sulcellata (Wissmann, 1841) |
3 |
5 |
|
2 |
|
Palaeoneilo distincta (Bittner, 1895) |
2 |
|
2 |
|
|
Palaeoneilo elliptica (Goldfuss, 1837) |
|
2 |
|
1 |
|
Modiolus paronai Bittner, 1895 |
1 |
|
|
|
|
Gervillia (Cultriopsis) sp. |
3 |
1 |
|
|
|
Cassianella beyrichi Bittner, 1895 |
1 |
3 |
|
3 |
|
Cassianella sp. 1 |
|
1 |
|
|
|
Cassianella sp. 2 |
|
|
|
2 |
|
Antijanira sp. |
1 |
|
|
|
|
Leptochondria sp. |
1 |
|
1 |
|
|
Schafhaeutlia cf. astartiformis (Münster, 1841) |
|
1 |
|
|
|
Septocardia pichleri (Bittner, 1895) |
2 |
3 |
|
2 |
|
Bivalvia indet. 1 |
8 |
|
11 |
|
|
Bivalvia indet. 2 |
1 |
|
|
|
|
Scaphopoda |
||||
|
Dentalium klipsteini Kittl, 1891 |
2 |
1 |
5 |
6 |
|
Plagioglypta undulata (Münster, 1841) |
1 |
1 |
|
|
|
Cephalopoda |
||||
|
Trachyceratidae indet. |
1 |
|
|
|
|
Ammonoidea indet. |
4 |
|
|
|
|
Brachiopoda |
||||
|
Brachiopoda indet. |
|
|
1 |
|
|
Ostracoda |
||||
|
Ostracoda indet. 1 |
1 |
|
|
|
|
Ostracoda indet. 2 |
|
|
1 |
|
|
Echinodermata |
||||
|
Echinodermata indet. |
1 |
|
1 |
|
|
Crinoidea |
||||
|
Isocrinus? sp. |
1 |
|
|
|
|
Crinoidea indet. |
1 |
|
|
|
|
Asteroidea |
||||
|
Asteroidea indet. |
1 |
|
|
|
|
Ophiuroidea |
||||
|
Ophiuroidea indet. 1 |
1 |
|
|
|
|
Ophiuroidea indet. 2 |
1 |
|
|
|
|
Ophiuroidea indet. 3 |
1 |
|
|
|
|
Ophiuroidea indet. 4 |
1 |
|
|
|
|
Other Ophiuroidea indet. |
2 |
|
|
|
|
Echinoidea |
||||
|
“Cidaris” cf. decoratissima Wöhrmann, 1889 |
1 |
|
|
|
|
“Cidaris” cf. flexuosa Münster, 1841 |
1 |
|
|
|
|
Cidaroida indet. 1 |
1 |
|
|
|
|
Pronechinus? sp. |
2 |
|
|
|
|
stem Cidaroida/Cidaroida indet. |
1 |
1 |
1 |
|
|
Echinoidea indet. |
2 |
|
|
|
|
Ophiocistioidea** |
||||
|
Linguaserra triassica Reich in Reich et al., 2018 ** |
1 |
|
|
|
|
Holothuroidea |
||||
|
Apodida gen. et. sp. nov. 1 |
1 |
|
|
|
|
Apodida gen. et. sp. nov. 2 |
1 |
|
|
|
|
Jumaraina sp. |
1 |
|
|
|
|
Tetravirga cf. perforata Mostler, 1968 |
1 |
|
|
|
|
Tetravirga sp. 1 |
1 |
|
|
|
|
Theelia cf. multiplex Speckmann, 1968 |
1 |
|
|
|
The most abundant species in the bulk sample from Misurina Landslide is the gastropod Jurilda elongata (Leonardi and Fiscon, 1959), which is rather rare in the collection from Lago Antorno. In contrast, the most abundant species in the Lago Antorno bulk sample is the gastropod Dentineritaria neritina (Münster, 1841) sensu Bandel (2007), which is also abundant in the bulk sample from Misurina Landslide but less frequent than at Lago Antorno (Table 1). The gastropod Neritaria mandelslohi (Klipstein, 1843) sensu Bandel (2007) is also very abundant at Lago Antorno, but rare at Misurina Landslide. The caenogastropods Coelostylina conica (Münster, 1841), Prostylifer paludinaris (Münster, 1841), and Ampezzopleura hybridopsis Nützel, 1998, are characteristic faunal components at both locations. Regardless of minor differences in species abundances, both fossil assemblages from Lago Antorno and Misurina Landslide are similar in composition, indicating that they derive from similar palaeoenvironments.
Diversity.—All three calculated diversity indices, i.e., Simpson, Shannon, and Berger-Parker, indicate that diversity is approximately at the same level in the bulk collections of Lago Antorno and Misurina Landslide (Table 3). Simpson and Shannon indices suggest a moderate diversity for both fossil assemblages. The dominance is not very high, as indicated by the Berger-Parker index. Diversity indices of the surface collections show that diversity is lower and dominance is much higher in comparison to the bulk samples. The Lago Antorno surface collection is less diverse than the surface collection from Misurina Landslide.
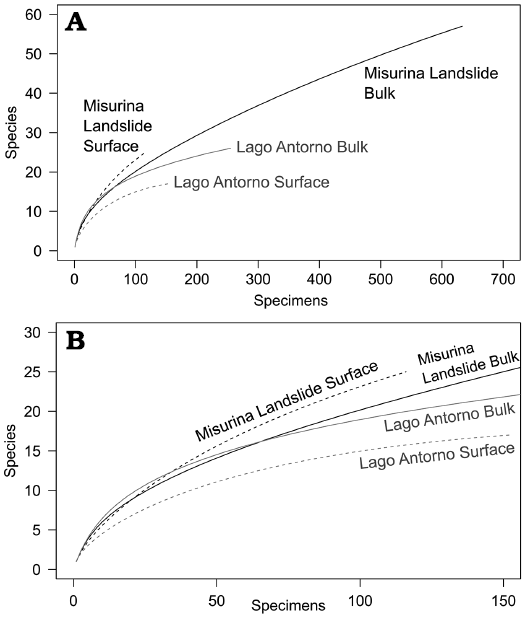
Fig. 3. Rarefaction curves of surface and bulk samples from Misurina Landslide and Lago Antorno. A. Complete curves. B. Detailed view.
Rarefaction curves suggest that the bulk collections from both Cassian localities are similar in diversity (Fig. 3). In addition, the surface samples show high similarities with the corresponding bulk samples. All rarefaction curves are not yet saturated. Therefore, it is expected that additional species would be found in larger samples.
Rank-abundance distributions of Lago Antorno and Misurina Landslide bulk samples best fit the Zipf-Mandelbrot model (Table 4, Fig. 4). This indicates that both assemblages have the same ecological complexity, i.e., being influenced by similar ecological processes (Frontier 1985; Magurran 2004) and characterised by a similar evenness, since they follow the same distribution.
Table 2. Number of species and specimens per higher taxon in the Misurina Landslide and Lago Antorno bulk samples.
|
Taxa |
Misurina Landslide |
Lago Antorno |
||
|
species |
specimens |
species |
specimens |
|
|
Gastropoda |
20 |
567 |
17 |
230 |
|
Bivalvia |
11 |
33 |
4 |
16 |
|
Scaphopoda |
2 |
3 |
1 |
5 |
|
Cephalopoda |
2 |
5 |
|
|
|
Brachiopoda |
|
|
1 |
1 |
|
Ostracoda |
1 |
1 |
1 |
1 |
|
Echinodermata |
1 |
1 |
1 |
1 |
|
Crinoidea |
2 |
2 |
|
|
|
Asteroidea |
1 |
1 |
|
|
|
Ophiuroidea |
5 |
6 |
|
|
|
Echinoidea |
6 |
8 |
1 |
1 |
|
Holothuroidea |
6 |
6 |
|
|
Table 3. Diversity indices of bulk and surface samples from Misurina Landslide and Lago Antorno.
| |
Misurina Landslide bulk |
Misurina Landslide surface |
Lago Antorno bulk |
Lago Antorno surface |
|
Simpson |
0.85 |
0.78 |
0.87 |
0.68 |
|
Shannon |
2.47 |
2.19 |
2.48 |
1.73 |
|
Berger-Parker |
0.26 |
0.43 |
0.27 |
0.53 |
Drilling predation.—Some molluscan shells from Misurina Landslide and Lago Antorno showed drill holes similar to Recent naticid borings (see for instance Fig. 10). In most cases, gastropods were drilled, for instance Coelostylina conica (Münster, 1841), Ampezzopleura hybridopsis Nützel, 1998, Ampezzogyra angulata Nützel and Hausmann sp. nov., and Angulatella bizzarinii Nützel and Hausmann sp. nov. However, only a small portion of all present species and specimens was drilled. Only 15 species out of a total of 54 molluscan species show drill holes which suggests a certain prey preference. In addition, only a comparatively small number of shells was drilled in total (c. 60 shells). All drilled species that were found simultaneously in both bulk and surface samples were considerably more frequently drilled in the corresponding surface samples, regardless of the fossil locality. In several cases, shells display more than one drill hole (see also Klompmaker et al. 2016).
Fossil size.—The fossils from Lago Antorno and Misurina Landslide are small, often only a few millimetres in size. The marly limestone bed situated near the sampling spot at Misurina Landslide (Fig. 1C) also contained fossils, mostly gastropods of the genus Coelostylina Kittl, 1894, and the bivalves Nuculana Link, 1807, and Cassianella Beyrich, 1862. The largest fossil found there was a specimen of Cassianella with a height of 32 mm and a width of 27 mm. We assume that the fossil assemblage embedded in the marly limestone bed is the same assemblage type as those in the unconsolidated sediments from Lago Antorno and Misurina, because their taxonomic compositions do not differ significantly. However, fossils of the size of the Cassianella specimen were not found in the unconsolidated marls. In contrast, they were much smaller in these samples. It is unclear whether this size difference is primary or was caused by diagenesis, which favoured preservation of larger specimens.
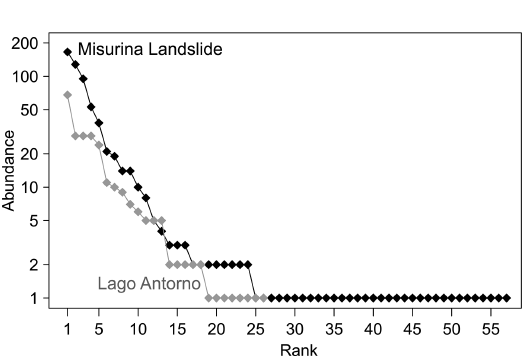
Fig. 4. Rank-abundance distributions of the bulk samples from Misurina Landslide and Lago Antorno.
Table 4. Best model fit for rank-abundance distributions of the Misurina Landslide and Lago Antorno bulk samples. Best fit is shown in bold.
|
Models |
AIC values |
|
|
Misurina Landslide |
Lago Antorno |
|
|
Brokenstick |
861.32 |
145.82 |
|
Preemption |
428.68 |
108.87 |
|
Log-normal |
285.5 |
107.08 |
|
Zipf |
270.89 |
122.81 |
|
Zipf-Mandelbrot |
203.45 |
103.98 |
Systematic palaeontology
Foraminifera
Foraminifera are abundant in the bulk samples from Misurina Landslide, with the most abundant being a species of Glomospira Rzehak, 1888, and encrusting forms of unknown identity. The fine fraction (<0.5 mm) from both localities was not analysed quantitatively for this study but yields an abundant foraminiferal assemblage. Several specimens of the foraminifera Variostoma exile Kristan-Tollmann, 1960, were found in the bulk sample from Lago Antorno.
Porifera and Hydrozoa
One small calcareous sponge fragment was found in the surface collection from Misurina Landslide (PZO 12680). A small fragment of a potential hydrozoan was recovered from the bulk sample from Lago Antorno (PZO 12795).
Mollusca
Class Gastropoda Cuvier, 1795
Subclass uncertain
Family Trochonematidae Zittel, 1895
Genus Eunema Salter, 1859
Type species: Pleurotomaria umbilicata Hall, 1847; original designation, USA, Ordovician.
Eunema badioticum Kittl, 1891
Fig. 5.
1891 Eunema badiotica n.f.; Kittl 1891: 246, pl. 2: 35.
1926 Eunema badioticum Kittl; Diener 1926: 65.
1978 Worthenia canalifera (Münster); Zardini 1978: pl. 5: 2.
Material.—One specimen (PZO 12796), from Lago Antorno surface sample, Italy, Carnian, Triassic.
Description.—Shell trochoid, comprising five whorls (apical whorls missing), 5.2 mm high, 4.4 mm wide; whorl face angulated at about mid-whorl with selenizone situated on angulation; whorl-face above angulation forming oblique, somewhat concave ramp; whorl-face below angulation approximately parallel to shell axis, concave; base convex joining whorl face at distinct angular edge at suture; whorl-face and base covered with numerous spiral threads and enhanced growth lines; at least six spiral threads on whorl face each, above and below median angulation; base with one or two weak angulations; whorls adpressed with subsutural bulge; this bulge, the median and the basal angulation form three prominent spiral ribs; growth lines opisthocyrt between adapical suture and median angulation and rather straight, almost opisthocline below median angulation; growth lines curve back abruptly and sharply at median angulation; minute pits present on median and basal angulations; aperture elongated, oblique.

Fig. 5. Trochonematid gastropod Eunema badioticum Kittl, 1891, PZO 12796, from Lago Antorno, northern Italy, Cassian Formation, Carnian, Upper Triassic. In apertural (A1) and lateral (A2, A3) views; A4, detail of last whorl showing fine ornamentation including pits on upper angulation, in lateral view.
Remarks.—The present specimen agrees well with the type specimen of Eunema badioticum (AN personal observation, NHMW 1899-0005-0184). Zardini (1978: pl. 5: 2a, b) illustrated this species as Wortheniella canalifera (Münster, 1841) (Pleurotomariida) but E. badioticum lacks a selenizone. The small pits on the spiral angulations of E. badioticum are remarkable and are shown here for the first time; they suggest the presence of periostracal hair. Eunema badioticum probably does not belong in the genus Eunema which has an Ordovician type species nor does it belong to Trochonematidae. This species might represent a new vetigastropod or caenogastropod genus but that remains open until specimens with protoconchs can be studied. At present, we leave E. badioticum in the systematic position that was suggested by Kittl (1891).
Stratigraphic and geographic range.—Carnian, Upper Triassic; northern Italy (Diener 1926).
Subclass Vetigastropoda Salvini-Plawén, 1980
?Family Turbinidae Rafinesque, 1815
“Turbo” sp.
Fig. 6.
Material.—One specimen (PZO 12740) from the Misurina Landslide surface sample, Italy, Carnian, Triassic.
Description.—Shell turbiniform, comprising ca. 6 whorls 5.4 mm high, ca. 4.4 mm wide (specimen slightly compressed, appearing wider); whorl embracing at mid-whorl periphery; last whorl distinctly higher than spire; whorl face moderately convex with shoulder giving spira gradate appearance; whorls ornamented with strengthened prosocline and slightly prosocyrt growth-lines; base evenly convex, seemingly anomphalous.

Fig. 6. Turbinid? gastropod “Turbo” sp., PZO 12740, from Misurina Landslide, northern Italy, Cassian Formation, Carnian, Upper Triassic. In apertural (A1) and lateral (A2), and apical (A3) views; A4, early whorls in lateral view.
Remarks.—Turbiniform shape and prosocline ornament of strengthened growth lines suggest that this specimen represents a turbinid vetigastropod species. The specimen from Misurina that was figured by Zardini (1978: pl. 12: 7 and 1985: pl. 10: 2) as Solarioconulus nudus (Münster, 1841) is probably conspecific with the present specimen. However, S. nudus is much broader with a greater apical angle according to the figures provided by Münster (1841) and Kittl (1892). Moreover, it lacks the strengthened growth lines and is smooth according to Kittl (1892). Trochus lissochilus Kittl, 1892, is similar but lacks a shoulder and the transition to the base is rounded angular. Turbo? vixcarinatus Münster, 1841, resembles the present specimen but lacks a shoulder and according to Kittl (1892) the growth lines are straight. We think that the present specimen represents an undescribed species but more material is necessary to characterize it sufficiently.
Subclass Neritimorpha Koken, 1896
Family Naticopsidae Waagen, 1880
Genus Hologyra Koken, 1892
Type species: Hologyra alpina Koken, 1892, subsequent designation by Kittl (1899); northern Italy, Late Triassic.
Hologyra? expansa (Laube, 1869)
Fig. 7.
1869 Naticopsis expansa; Laube 1869: 11, pl. 22: 5.
1892 Naticopsis expansa Laube sp.; Kittl 1892: 82, pl. 7: 22–24.
1978 Hologyra expansa (Laube); Zardini 1978: 35, pl. 19: 6–12, 23, 24.
Material.—One specimen (PZO 12802) from Lago Antorno surface sample, Italy, Carnian, Triassic.
Description.—Shell bulbous, consisting of about four rapidly expanding whorls, 3.6 mm wide, 3.9 mm high; spire slightly elevated; whorls markedly convex, with broad subsutural shelf; whorl embracing somewhat above periphery; whorls smooth except for distinct, prosocline, prosocyrt growth lines; aperture with outer lip broken off, with pronounced callus on parietal lip.

Fig. 7. Naticopsid gastropod Hologyra? expansa (Laube, 1869), PZO 12802, from Lago Antorno, northern Italy, Cassian Formation, Carnian, Upper Triassic; apertural view showing callus on inner and parietal lip.
Remarks.—This specimen differs from the other neritimorphs in the studied samples, Neritaria mandelslohi and Dentineritaria neritina, by having a low but nevertheless distinctly elevated spire. It is much larger than the specimens representing these species in our samples that are only present with small juveniles. It agrees well with the illustrations of H. expansa given by Kittl (1892) who also figured Laube’s (1869) type specimens. It is also close to the specimens that were illustrated by Zardini (1978) as H. expansa. Species identity is not entirely beyond doubt because this specimen is not fully grown and large parts of the aperture are broken off. The placement of this species in Hologyra is doubtful because the type species of Hologyra has a blunt and not an elevated spire.
Stratigraphic and geographic range.—Only known from northern Italy (Cassian Formation), Carnian, Upper Triassic.
Family Neritariidae Wenz, 1938
Genus Neritaria Koken, 1892
Type species: Natica mandelslohi Klipstein, 1843; Italy, Late Triassic.
Neritaria mandelslohi (Klipstein, 1843)
Fig. 8.
2007 Neritaria mandelslohi (Klipstein, 1843); Bandel 2007: 261, fig. 11N–P, fig. 12A–H.
Material.—Thirty four specimens, all from bulk samples; 29 from Lago Antorno (PZO 12798, 12799, 2 figured specimens; PZO 12797, 27 specimens); 5 from Misurina Landslide (PZO 12681, 5 specimens); Italy, Carnian, Triassic.
Description.—Shell egg-shaped, low-spired with rapidly increasing whorls; illustrated specimen with about three whorls, 0.7 mm wide and high; initial whorl corroded but seemingly convolute, smooth; larval shell largely smooth, only with faint reticulate ornament on adapical portion of whorls; reticulate ornament consists of somewhat strengthened growth lines and spiral lirae which diverge somewhat in an apertural direction; base smooth, evenly rounded; larval shell with diameter of about 0.5 mm, ends at simple, faint suture; only small part of teleoconch present, seemingly smooth.
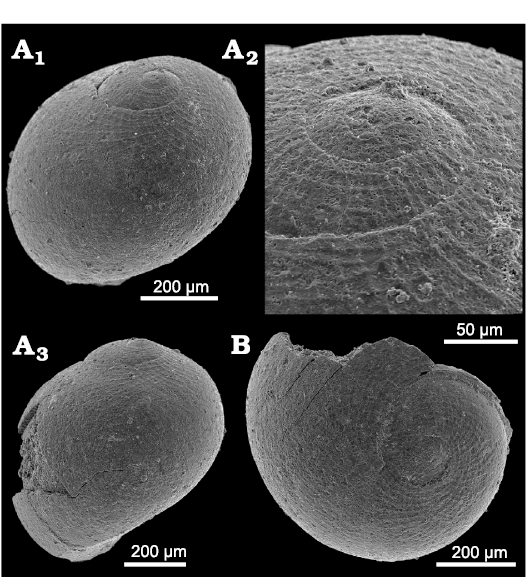
Fig. 8. Neritariid gastropods Neritaria mandelslohi (Klipstein, 1843) from Lago Antorno, northern Italy, Cassian Formation, Carnian, Upper Triassic. A. PZO 12798, in oblique lateral (A1) and lateral view (A3), and oblique lateral view of protoconch (A2). B. PZO 12799 in apical view.
Remarks.—Only isolated larval shells or juvenile specimens with a small teleoconch portion are at hand. They do not exceed a size of 1 mm. However, this species reaches a size of several millimetres at other locations (Kittl 1892; Bandel 2007). This could indicate that the studied specimens represent mortal larval fall (failure to undergo metamorphosis) or that specimens died at or shortly after metamorphosis. The larval shell is rather characteristic and resembles that of Neritaria mandelslohi as reported by Bandel (2007: fig. 11N–P).
Stratigraphic and geographic range.—Ladinian–Carnian, Triassic; Muschelkalk in Poland; northern Italy (Cassian Formation); Hungary (Diener 1926; Kutassy 1940).
Genus Dentineritaria Bandel, 2007
Type species: Natica neritina Münster, 1841; Italy, Late Triassic.
Dentineritaria neritina (Münster, 1841)
Fig. 9.
2007 Dentineritaria neritina (Münster); Bandel 2007: 265, fig. 13A–H.
Material.—One hundred and six specimens, all from bulk samples; 68 from Lago Antorno (PZO 12801, figured specimen; PZO 12800, 67 specimens); 38 from Misurina Landslide (PZO 12683, 12684, 2 figured specimens; PZO 12682, 36 specimens); Italy, Carnian, Triassic
Description.—Shell egg-shaped, low-spired with rapidly increasing whorls; illustrated specimen (Fig. 9A) with about three whorls, 0.9 mm wide, 0.8 mm high; protoconch consists of about three smooth whorls, 0.43 mm wide; initial whorls corroded but seemingly convolute; last larval whorl with distinct adapical suture; larval shell ends at simple, faint suture; somewhat less than one teleoconch whorl preserved; early teleoconch with subsutural bulge and initially with weak axial ribs (strengthened growth lines) which are restricted to adapical portion of whorls; axial ribs fade after half whorl so that remaining teleoconch is entirely smooth; base smooth, evenly rounded; aperture higher than wide with evenly arched outer lip; inner lip reflexed, forming columellar callus; first illustrated specimen from Misurina Landslide (Fig. 9B) is 0.8 mm wide and 0.7 mm high; consists of about three whorls; second specimen (Fig. 9C) 0.7 mm wide, with about two whorls; both illustrated specimens from Misurina Landslide less well preserved than illustrated shell from Lago Antorno.
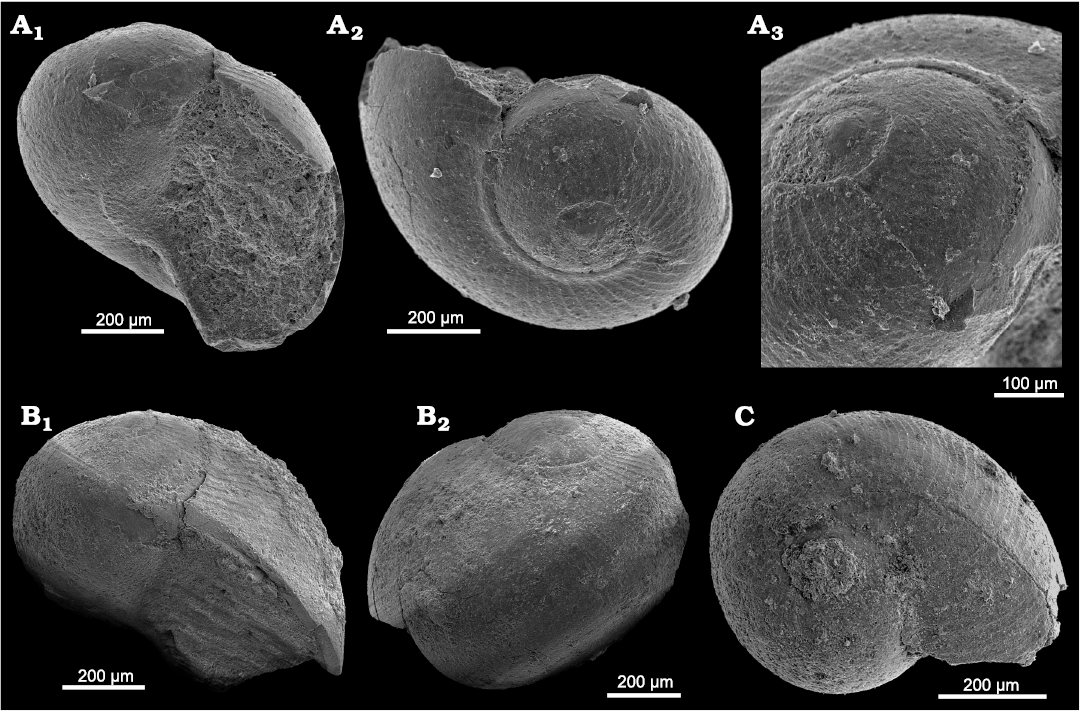
Fig. 9. Neritariid gastropods Dentineritaria neritina (Münster, 1841) from Lago Antorno (A) and Misurina Landslide (B, C), northern Italy, Cassian Formation, Carnian, Upper Triassic. A. PZO 12801, in apertural (A1) and apical (A2) views; A3, end of smooth larval shell well visible in oblique lateral view. B. PZO 12683, in apertural (B1) and abapertural (B2) views. C. PZO 12684 in apical view.
Remarks.—As in Neritaria mandelslohi, only isolated larval shell or specimens with a small teleoconch portion are at hand. Thus even this neritimorph species did not find suitable living conditions in the sampled area. The present specimens resemble early growth stages of Dentinerita neritina as reported by Bandel (2007). Bandel (2007) interpreted the axial ribbed shell as a part of the larval shell which according to him has a diameter of 0.8 mm. However, the present material suggests that the larval shell is much smaller (0.4 mm wide) and that the axially ribbed shell represents the early teleoconch.
Stratigraphic and geographic range.—Ladinian–Carnian, Triassic; Italy (southern Alps, Cassian Formation, Esino Limestone, Marmolata Limestone; eastern Alps, Coquina of Ghegna); South America (Diener 1926; Kutassy 1940).
Subclass Caenogastropoda Cox in Knight et al., 1960
Superfamily unassigned
Family Coelostylinidae Cossmann, 1909
Genus Coelostylina Kittl, 1894
Type species: Melania conica Münster, 1841; Italy, Late Triassic.
Emended diagnosis.—Shell acute conical, medium-sized; last whorl higher than spire; whorls moderately convex with narrow shoulder; sutures distinctly impressed; whorls smooth or with very faint spiral ornament including spirally arranged micro-pits and furrows; growth lines slightly opisthocyrt to orthocline; base convex, not demarcated from whorl face; base minutely phaneromphalous; aperture higher than wide, with straight columellar lip and convex outer lip; protoconch conical, orthostrophic, smooth; protoconch whorls convex, without shoulder, demarcated from teleoconch by a distinct sinusigera.
Remarks.—The re-investigation of Münster’s (1841) type specimen and the well-preserved material at hand allow a sharper diagnosis of the genus Coelostylina which has been used as a dustbin taxon for many years.
Coelostylina conica (Münster, 1841)
Figs. 10, 11.
1841 Melania conica n. sp.; Münster 1841: 28, pl. 23: 20.
1894 Coelostylina conica (Münster, 1841); Kittl 1894: 181, pl. 14: 1–7.
1959 Coelostylina conica (Münster, 1841); Leonardi and Fiscon 1959: 54, pl. 5: 20.
1978 Coelostylina conica (Münster, 1841); Zardini 1978: 44–45, pl. 28: 12–14, pl. 29: 1–3.
pars 1992 Coelostylina conica (Münster, 1841); Bandel 1992: 55, pl. 6: 6, pl. 7: 1, 2, 6, non 5
non 2006, pl. 12: 10–12.
1996 Carboninia valvatiformis n. sp.; Bandel 1996: 55, figs. 17a–h, 18a.
Lectotype: SNSB-BSPG AS VII 504, the specimen illustrated by Münster (1841: pl. 23: 20).
Type locality: Area near St. Cassian, northern Italy.
Type horizon: Cassian Formation, Late Triassic; Early Carnian.
Material.—One hundred and twenty seven specimens, 82 from bulk samples, 45 from surface samples; 55 from Lago Antorno (PZO 12803–12810, 8 figured specimens; PZO 12803, 47 specimens); 72 from Misurina Landslide (PZO 12686–12690, 5 figured specimens; PZO 12685, 67 specimens); Italy, Carnian, Triassic.
Description.—Shell acutely conical; apical angle about 45°; lectotype comprises about eight whorls, 13.4 mm high, 6.7 mm wide (Fig. 10A); last whorl higher than spire; whorls moderately convex with narrow shoulder; sutures distinctly impressed; whorls smooth or with rows of minute pits (seen on lectotype) or very faint spiral grooves which are only visible in oblique light; growth lines slightly opisthocyrt to orthocline; base convex, not demarcated from whorls face; base minutely phaneromphalous; aperture higher than wide, with straight columellar lip and convex outer lip; protoconch conical, more low-spired than teleoconch, apical angle 70–80°; protoconch orthostrophic, smooth, consisting of about 3.5 whorls 0.35–0.45 mm high, 0.38–0.50 mm wide; protoconch whorls convex, without shoulder (in contrast to teleoconch); protoconch demarcated from teleoconch by opisthocyrt ledge; initial whorl flattened, almost planispiral to immersed, with diameter of 0.11–0.13 mm.

Fig. 10. Coelostylinid gastropod Coelostylina conica (Münster, 1841) from Lago Antorno (A–D) and Misurina Landslide (E, F), northern Italy, Cassian Formation, Carnian, Late Triassic. A. PZO 12803, in apertural (A1) and abapertural (A2) views; A3, spire whorls in lateral view; A4, close up of spire whorl showing faint spirally arranged micro-pits; A5, close up of apical whorls in lateral view. B. PZO 12804 in apertural view. C. PZO 12805, with drill hole, in apertural view (C1); C2, early whorls including protoconch, in lateral view. D. PZO 12806, in lateral view (D1); D2, early whorls including protoconch, in lateral view; D3, whorl detail with boring. E. PZO 12686, in apertural view. F. PZO 12687, with drill hole in apertural view (F1); F2, detail, in apertural view.

Fig. 11. Coelostylinid gastropods Coelostylina conica (Münster, 1841) from Lago Antorno (A–D) and Misurina Landslide (E–G), northern Italy, Cassian Formation, Carnian, Upper Triassic. A. PZO 12807, A1, teleoconch shows faint spiral furrows in oblique apical view; A2, apertural view; A3, protoconch in lateral view with sinusigera. B. PZO 12808, in apertural view, with drill hole. C. PZO 12809, C1, juvenile whorls including protoconch ending at sinusigera, in oblique lateral view; C2, juvenile whorls including protoconch ending at sinusigera, in apertural view. D. PZO 12810, teleoconch shows faint spiral furrows, end of smooth larval shell well visible, in oblique apical view. E. PZO 12688, in lateral view. F. PZO 12689, F1, juvenile whorls including protoconch in apical view; F2, juvenile whorls including protoconch, in lateral view; F3, juvenile whorls including protoconch, in lateral view; F4, juvenile whorls including protoconch with dipping initial whorl, in lateral view. G. PZO 12690, juvenile whorls including protoconch, in lateral view.
Remarks.—The present material from Lago Antorno and Misurina Landslide closely resembles Münster’s (1841) type specimen (lectotype) from the Stuores Wiesen and is obviously conspecific with it. The specimens illustrated by Bandel (1992: pl. 6: 6, pl. 7: 1, 2, 6) also agree well with the present material. However, the protoconchs illustrated by Bandel (1992: pl. 7: 5; 2006: pl. 12: 10–12) and assigned by him to C. conica differ significantly from the protoconchs reported here. According to Bandel’s (1992) illustration and description, the protoconch of Coelostylina conica is about 0.25 mm high and wide, consists of about 2.5 whorls, and has an ornament of minute tubercles. In contrast, the protoconchs reported here consist of 3.5 whorls, are much larger (twice the size) and lack any ornament. The juvenile specimens identified by Bandel (2006: pl. 12: 10–12) have a larval shell with numerous spirally arranges tubercles. This specimen which has only one teleoconch whorl preserved does not represent C. conica but another species and genus. Carboninia valvatiformis Bandel, 1996, type species of Carboninia Bandel, 1996, is conspecific with our material and hence represents a synonym of Coelostylina conica and therefore Carboninia is a synonym of Coelostylina. Bandel (1996) interpreted the immersed initial whorl to represent larval heterostrophy with a protoconch of 1.8 whorls but our better preserved material shows that this species has a caenogastropod larval shell of 3.5 whorls which can also be seen in Bandel’s (1996: fig. 17e) illustration of Carboninia valvatiformis.
Stratigraphic and geographic range.—Middle–Upper Triassic, Anisian–Norian/Rhaetian; Italy (southern Alps, Cassian Formation, Raibler Schichten, Marmolata Limestone, Esino Limestone; Latemar Limestone near Forno, Italy, Apennines, Sicily); Italy (eastern and northern Alps, Hauptdolomit); Bulgaria; Hungary; Poland (Silesia, Upper Wellenkalk); Romania (western Carpathians, Siebenbürgen); Iran (Nayband Formation) (Diener 1926; Kutassy 1940; Sachariewa-Kowatschewa 1962; Kollarova-Andrusova and Kochanova 1973; Nützel and Senowbari-Daryan 1999; Nützel et al. 2012).
Coelostylina sp. 1
Fig. 12B.
Material.—One specimen (PZO 12691) from Misurina Landslide bulk sample, Italy, Carnian, Triassic.
Description.—A single small shell is present with a height of about 1 mm. It consists of about two teleoconch whorls which are smooth and bulbous and a poorly preserved mammilated larval shell.
Remarks.—Species identification is impossible, but the specimen resembles the genus Coelostylina or Prostylifer to some degree.
Coelostylina sp. 2
Fig. 12C.
Material.—One specimen (PZO 12692) from the Misurina Landslide surface sample, Italy, Carnian, Triassic.
Description.—This fusiform shell consists of about four whorls (apex missing), has largely smooth, convex whorls with almost straight orthocline growth lines and a faint spiral micro-striation.
Remarks.—This form resembles Coelostylina species, for instance Coelostylina (Pseudochrysalis) stotteri (Klipstein, 1843) (see Zardini 1978: pl. 30: 6), to some degree but an umbilical chink seems to be absent.

Fig. 12. Caenogastropods from Lago Antorno (A) and Misurina Landslide (B–F), northern Italy, Cassian Formation, Carnian, Upper Triassic. A. Unidentified caenogastropod, PZO 12829, in lateral view. B. Coelostylina sp. 1, PZO 12691, in apertural view. C. Coelostylina sp. 2, PZO 12692, in lateral view. D. Flemingella bistriata (Münster, 1841), PZO 12713, in apertural view. E. Coelochrysalis pupaeformis (Münster, 1841), PZO 12714, in lateral view, with constriction of last whorl. F. Coelochrysalis pupaeformis (Münster, 1841), PZO 12715, deformed specimen in lateral view, showing constriction of last whorl.
Genus Coelochrysalis Kittl, 1894
Type species: Melania pupaeformis Münster, 1841; Italy, Late Triassic.
Coelochrysalis pupaeformis (Münster, 1841)
Fig. 12E, F.
1894 Euchrysalis (Coelochrysalis) pupaeformis; Kittl 1894: 225, pl. 6: 15–20.
Material.—Two specimens (PZO 12714, 12715) from the Misurina Landslide surface samples, Italy, Carnian, Triassic.
Description.—Two specimens show the characteristic pupoid shape of this species that is produced by the constriction of the body whorl. The whorls are low; the larger specimen is 18 mm high. Both specimens lack the early whorls and are compressed.
Remarks.—Fürsich and Wendt (1977) reported a single specimen from their Ampullina association.
Unidentified caenogastropod
Fig. 12A.
Material.—Two specimens (PZO 12829, 12828) from Lago Antorno surface samples, Italy, Carnian, Triassic.
Description.—Two small fragments of a smooth shelled high-spired gastropod species that may represent Caenogastropoda.
Genus Helenostylina
Kaim, Jurkovšek,
and Kolar-Jurkovšek, 2006
Type species: Helenostylina mezicaensis Kaim, Jurkovšek, and Kolar-Jurkovšek, 2006; Slovenia, Late Triassic.
Helenostylina convexa Nützel and Kaim, 2014
Fig. 13.
1992 Ptychostoma sanctaecrucis (Laube, 1868); Bandel 1992: 52, pl. 6: 4, 5.
1994 Ptychostoma sanctaecrucis (Laube, 1868); Bandel 1994: pl. 4: 8.
2014 Helenostylina convexa n. sp.; Nützel and Kaim 2014: 418, fig. 7e–h.
Material.—Thirteen specimens, 7 from bulk samples, 6 from surface samples; 10 from Lago Antorno (PZO 12812–12814, 3 figured specimens; PZO 12811, 7 specimens); 3 from Misurina Landslide and Misurina Skilift (PZO 12694, 12695, 2 figured specimens; PZO 12693, 1 specimen); Italy, Carnian, Triassic.
Description.—Shell turbiniform, conical; apical angle about 70°; illustrated specimen from Misurina Landslide (Fig. 13E) with about seven whorls, 4 mm high, 2.8 mm wide; last whorl higher than spire; teleoconch whorls smooth, convex, without shoulder; base flatly convex with two spiral ribs and furrows surmounting the centre of the base; aperture as high as wide; protoconch comprising about 3.5–4 whorls, 0.4–0.5 mm high and wide, with about same apical angle as teleoconch (70–80°); initial whorl smooth with a diameter of about 0.14 mm; larval whorls convex, somewhat angulated at lower third; larval whorls with subsutural row of tubercles, followed by a smooth zone below extending to the whorl angulation; lower zone between angulation and abapical suture ornamented with 5–6 rows of fine tubercles; larval shell ends abruptly.

Fig. 13. Coelostylinid gastropods Helenostylina convexa (Nützel and Kaim, 2014) from Lago Antorno (A, B, C), Misurina Skilift (D), and Misurina Landslide (E), northern Italy, Cassian Formation, Carnian, Upper Triassic. A. PZO 12812, relatively large specimen with well-preserved base with spiral ribs, in apertural view (A1); A2, early whorls in lateral view; A3, basal view showing spiral ribs; A4, oblique basal view showing spiral ribs on base. B. PZO 12813, relatively large specimen with spiral ribs on base and protoconch with tubercles, in apertural view (B1); B2, close up of base showing spiral ribs on base; B3, early whorls with tubercles on larval shell, in lateral view. C. PZO 12814, juvenile shell with tubercles on larval shell, in lateral view. D. PZO 12694, juvenile shell with tubercles on larval shell and healed shell fracture, in apertural (D1) and lateral (D2, D4) views; details showing end of larval shell (arrows), in lateral (D3) and apical (D5) views. E. PZO 12695, relatively large specimen with well-preserved base with spiral ribs and protoconch with tubercles, in apertural view (E1); E2, oblique apertural view to show base with spiral ribs; E3, protoconch in lateral view.
Remarks.—This species has a very characteristic larval shell of rows of small tubercles below the suture and on the lower third of the larval whorls separated by a wide smooth zone. Our material resembles the more juvenile type specimens as reported by Nützel and Kaim from the Settsass Scharte—one of their figured specimens (Nützel and Kaim 2014: fig. 7e) shows the tubercular ornament on the larval shell although it was not mentioned in the description. The same species and larval shell was reported by Bandel (1992, 1994) who assigned it to Ptychostoma sanctaecrucis (Wissmann, 1841) [erroneously cited as Ptychostoma sanctaecrucis (Laube, 1868) by Bandel (1992, 1994)]. However, Bandel’s (1992, 1994) illustrated specimen from the locality Campo near Cortina d’Ampezzo does not represent P. sanctaecrucis. Ptychostoma sanctaecrucis was first described from the type location of the Heiligkreuz Formation which overlies the Cassian Formation. This species lacks the spiral ornament on the base and is larger. Moreover, Ptychostoma sanctaecrucis has a smaller larval shell which is ornamented by spiral crests, the same type that is present in Prostylifer (own observation on material from the type locality).
Helenostylina mezicaensis Kaim, Jurkovšek, and Kolar-Jurkovšek, 2006, the type species of Helenostylina resembles H. convexa in shape and in having a spiral ornament around the umbilicus. However, this species has a subsutural ramp and the protoconch is seemingly smooth.
Omphaloptycha muensteri (Wissmann, 1841) is similar. However the type specimen (SNSB-BSPG AS VII 1676) lacks circumumbilical spiral ribs and furrows and its whorls are higher. The type is larger (height 7 mm) than the specimens assigned to Helenostylina convexa by Nützel and Kaim (2014) and herein. The possibility that Omphaloptycha muensteri has basal spiral ribs and furrows only in the middle whorls but not on a more mature growth stage as present in the type specimen cannot be ruled out. If this is the case, Helenostylina convexa could represent a synonym of Omphaloptycha muensteri but this is seen as unlikely.
Stratigraphic and geographic range.—Only known from the Carnian, Upper Triassic; northern Italy (Cassian Formation).
Family Prostyliferidae Bandel, 1992
Genus Prostylifer Koken, 1889
Type species: Melania paludinaris Münster, 1841; Late Triassic, Italy.
Prostylifer paludinaris (Münster, 1841)
Fig. 14.
1841 Melania paludinaris n. sp.; Münster 1841: 97, pl. 9: 50.
1992 Prostylifer paludinaris (Münster, 1841); Bandel 1992: 50, pl. 5: 3–6.
1993 Prostylifer paludinaris (von Münster, 1841); Bandel 1993: 49, pl. 6: 1.
1994 Prostylifer paludinaris (Münster, 1841); Bandel 1994: 143, pl. 4: 10.
Material.—Two hundred eighty three specimens, 152 from bulk samples, 131 from surface samples; 105 from Lago Antorno (PZO 12823, 12824, 13680, 3 figured specimens; PZO 12821, 102 specimens); 178 from Misurina Landslide (PZO 12699, figured specimen; PZO 12698, 177 specimens); Italy, Carnian, Triassic.
Description.—Shell littoriniform to turbiniform, bulbous; spire conical, acute; shell with 8 whorls (including protoconch), 5.0 mm high, 3.8 mm wide (Fig. 14A); last whorl much higher than spire; whorls strongly convex; sutures distinct; teleoconch whorls smooth; base rounded, evenly convex, not demarcated from whorl face, cryptomphalous; aperture oblique teardrop-shaped; inner lip oblique, evenly arched; columellar lip strengthened, somewhat reflexed; protoconch orthostrophic, mammilate, in general more slender than teleoconch; protoconch consisting of about 3.1–3.2 whorls, 0.33–0.40 mm high, 0.42–0.45 mm wide; initial whorl smooth, with diameter of 0.14–0.15 mm; protoconch whorls convex, ornamented with up to six sharp, narrow spiral crests; spiral crests evenly spaced, separated by wide concave interspaces; spiral crests crenulated with fine tubercles; abapical crest emerges at lower suture; additional crests present on convex base of the larval whorls; larval shell terminates abruptly with a distinct sinusigera; larval projection largely covered by succeeding teleoconch whorls; illustrated specimen from Misurina Landslide (Fig. 14D) comprising about seven whorls; last whorl broken; shell 2.1 mm high, 1.7 mm wide; protoconch 0.43 mm high and 0.4 mm wide.

Fig. 14. Prostyliferid gastropods Prostylifer paludinaris (Münster, 1841) from Lago Antorno (A–C) and Misurina Landslide (D), northern Italy, Cassian Formation, Carnian, Upper Triassic. A. PZO 13680, relatively large specimen with preserved protoconch and inner lip, in apertural view (A1); A2, close up showing umbilical chink and arched inner lip; A3, apex with mammilated protoconch having spiral crests, in lateral view. B. PZO 12823, juvenile specimen with protoconch having spiral crests, in apertural view. C. PZO 12824, juvenile specimen with protoconch having spiral crests; C1, larval shell ending at sinusigera, in apertural view; C2, close up of whorl of larval shell. D. PZO 12699, juvenile specimen with mammilated protoconch having spiral crests, in lateral view (D1); D2, early whorls, larval shell ending at sinusigera, in lateral view.
Remarks.—This material from Lago Antorno and Misurina Landslide agrees well with the specimens illustrated by Bandel (1992: pl. 5: 3–6) which were reported from the localities Misurina, Alpe di Specie, and Costalaresc by this author. It also agrees with Münster’s (1841) type specimens at the Bavarian State Collection. One of the specimens illustrated by Bandel (1992: pl. 5: 5) has shouldered teleoconch whorls; Bandel (1992) also mentioned a great variation in the course of the growth line from slightly opisthocyrt to specimens which have an apertural sinus in the upper part of the whorls. Moreover, he mentioned the presence of a fine spiral striation on some specimens. This character was not observed in the present material. The larval shells documented by Bandel (1992) agree well with those in the present material.
Stratigraphic and geographic range.—Carnian, Triassic; northern Italy (Cassian Formation); Germany (eastern Alps, Raibler Schichten; northern Alps, Cardita Schichten); China, Yunnan; Timor; Bulgaria (Diener 1926; Kutassy 1940; Sachariewa-Kowatschewa 1962).
Genus Angulatella Nützel and Hausmann nov.
ZooBank LSID: urn:lsid:zoobank.org:act:A90DB9CA-6986-4819-BA BB-ECBE164CFDA2
Type species: Angulatella bizzarinii Nützel and Hausmann sp. nov., see below.
Species included: Type species only.
Diagnosis.—High-spired shell, spiral keels on teleoconch; upper keel angulates whorl profile in early teleoconch whorls, with a steep ramp between adapical suture and keel; whorl profile concave, parallel to shell axis between upper and lower keels; later whorl more or less convex with weaker angulations; base convex, anomphalous; protoconch orthostrophic; larval whorls ornamented with two strong spiral keels; third spiral keel present on third protoconch whorls; spiral keels angulate larval whorls; spiral keels ornamented with minute tubercles; protoconch ends abruptly at sinusigera; sinus strengthened with a varix.
Remarks.—The carinate, small larval shell of Angulatella closely resembles larval shells of many Recent and fossil Cerithioidea. The teleoconch resembles that of some Mathildoidea. However, Mathildoidea have a heterostrophic protoconch. The finding of Angulatella is further evidence for an early radiation of Cerithioidea. The small bicarinated larval shell resembles that of Permocerithium Nützel, 2012 (Nützel and Nakazawa 2012) from the Middle Permian of Japan. However, the teleoconch of this genus is not angulated. The Triassic Camposcala has the same type of larval shell but differs from Angulatella in having a strong cancellate teleoconch ornament. Small bicarinated larval shells of the same type have been reported from numerous Mesozoic Cerithoidea (e.g., Bandel 1992; Schröder 1995; Kaim 2004; Guzhov 2004).
Stratigraphic and geographic range.—Only known from the Carnian, Upper Triassic; northern Italy (Cassian Formation).
Angulatella bizzarinii Nützel and Hausmann sp. nov.
Fig. 15.
1993 Protuba winkleri (Klipstein, 1894); Bandel 1993: pl. 4: 3.
ZooBank LSID: urn:lsid:zoobank.org:act:BEA466D7-1E46-4E41-B4 EA-FE393C27B0D8
Etymology: After the palaeontologist Fabrizio Bizzarini for his excellent work on the fauna of the Cassian Formation.
Type material: Holotype PZO 12701, specimen with larval shell and 4 teleoconch whorls from Misurina Landslide. Paratypes: PZO 12826, specimen with larval shell and 4 teleoconch whorls; PZO 12827, relatively large specimen with 4 teleoconch whorls but lacking protoconch, from Lago Antorno.
Type locality: Misurina Landslide, near Cortina d’Ampezzo, northern Italy.
Type horizon: Cassian Formation; lower Carnian, Upper Triassic.
Material.—Eight specimens, all from bulk samples; 5 from Lago Antorno (PZO 12826, 12827, 2 paratypes; PZO 12825, 3 specimens); 3 from Misurina Landslide (PZO 12701, holotype; PZO 12700, 2 specimens); Italy, Carnian, Triassic.
Diagnosis.—High-spired, conical shell; teleoconch with two spiral keels on whorl face; upper keel at about mid-whorl; lower keel emerging at abapical suture; upper keel angulates whorl profile with a steep ramp between adapical suture and keel; shell axis of orthostrophic protoconch somewhat oblique to teleoconch axis; larval whorls with spiral keels which angulate larval whorls; spiral keels ornamented with minute tubercles.
Description.—Shell high-spired, conical; apical angle about 45°; shell with 6.5 whorls about 1.4 mm high, 0.8 mm wide (Fig. 15A); teleoconch with two spiral keels on whorl face of earliest teleoconch whorls; upper keel at about mid-whorl; lower keel emerging at abapical suture; upper keel angulates whorl profile with a steep ramp between adapical suture and keel; ramp slightly convex to straight; whorl profile concave, parallel to shell axis between upper and lower keels; in later whorl the abapical spiral keel moves in an adapical direction; the latest preserved whorl has a rounded convex appearance with only slight angulations (Fig. 15B); at least early teleoconch whorls covered with micro-ornament of tubercles (Fig. 15A5), base convex, anomphalous ornamented with several weak spiral ribs; protoconch orthostrophic, consisting of about three whorls, 0.25 mm high, 0.22 mm wide; shell axis of protoconch somewhat oblique to teleoconch axis; initial whorl without visible ornament, with diameter of 0.12 mm; first larval whorl ornamented with two strong spiral keels; third spiral keel present on third protoconch whorls; spiral keels angulate larval whorls; spiral keels ornamented with minute tubercles; protoconch ends abruptly at sinusigera; sinus strengthened with a varix.

Fig. 15. Caenogastropods Angulatella bizzarinii Nützel and Hausmann sp. nov. from Lago Antorno (A, B) and Misurina Landslide (C), northern Italy, Cassian Formation, Carnian, Upper Triassic. A. Paratype PZO 12826, specimen with preserved protoconch having bicarinate shell, in apertural view (A1); protoconch (A2–A5): A2, early whorls in lateral view inclucing bicarinate larval shell with axis oblique to teleoconch shell axis; A3, early whorls in oblique lateral view showing larval shell terminating at sinusigera; A4, close up of larval whorls with carinae ornamented with minute tubercles, in lateral view; A5, close up to show tuberculate micro-ornament on teleoconch whorl, upper portion shows sinusigera where larval shell end B. Paratype PZO 12827, relatively large specimen lacking protoconch, in lateral view. C. Holotype, PZO 12701, specimen with preserved protoconch having bicarinate shell, in apertural (C1) and apical (C3) views; C2, early whorls including bicarinate larval shell, in lateral view.
Remarks.—Bandel (1993: pl. 4: 3) illustrated a protoconch obviously representing Angulatella bizzarinii Nützel and Hausmann sp. nov. and identified it as “Protuba winkleri (Klipstein, 1843)”. However, Protuba Cossmann, 1912, is a monotypic genus holding P. intermittens Kittl, 1894, from the Cassian Formation. Bandel (1993) probably meant Promathildia winkleri Klipstein, 1894. Promathildia winkleri is known from relatively large teleoconch fragments, its protoconch is unknown. It has a much stronger median spiral angulation and the whorls do not become more or less rounded convex in mature whorls so that species identity with our material can be excluded.
Stratigraphic and geographic range.—Only known from the Carnian, Upper Triassic; northern Italy (Cassian Formation).
Family Protorculidae Bandel, 1991
Genus Atorcula Nützel, 1998
Type species: Melania canalifera Münster, 1841; Late Triassic, Italy.
Atorcula sp.
Fig. 16A, B.
Material.—Three teleoconch fragments (PZO 12815, 12816, 12817) from Lago Antorno surface samples, Italy, Carnian, Triassic.
Description.—Shell high-spired; one fragment (apex missing) comprises five whorls, is 13.4 mm high and 7.0 mm wide (Fig. 16A); whorls moderately convex; sutures distinct; shell smooth; growth lines parasigmoidal with shallow opisthocyrt sinus below suture and opisthocline on remaining part of whorl face; base moderately convex, smooth with growth lines only; transition from whorl face to base a rounded edge; protoconch unknown; two illustrated specimens (Fig. 16A, B) show circular borings: one specimen with two borings, one unsuccessful; other specimen with a single boring.

Fig. 16. Protorculid gastropods from Lago Antorno (A–D) and Misurina Landslide (E), northern Italy, Cassian Formation, Carnian, Upper Triassic. A. Atorcula sp., PZO 12816, in lateral view with drill holes (A1), A2, close up showing incomplete drill hole (right) and complete one (left). B. Atorcula sp., PZO 12817, in lateral view (B1), B2, close up showing drill hole. C. Atorcula canalifera (Münster, 1841), PZO 12819, in apertural view. D. Atorcula canalifera (Münster, 1841), PZO 12888, in apertural view. E. Atorcula canalifera (Münster, 1841), PZO 12697, axially ribbed larval shell and first smooth teleoconch whorls, in lateral view.
Remarks.—The high-spired shape, the smooth shell and the parasigmoidal course of the growth lines on the base suggest that the present specimens represent the genus Atorcula. However, Atorcula is also characterized by an axially ribbed larval shell but the protoconch of the present material is unknown. Moreover, Atorcula has spiral furrows on the base whereas the base of the present specimens lacks spiral furrows. Atorcula canalifera also has a less convex, almost straight whorl face. This species was obviously a preferred target for a boring predator. At least some of the specimens that were illustrated by Zardini (1978: pl. 27: 6–13) as Anoptychia canalifera (Münster, 1841) are probably conspecific with the present specimens. These specimens are mainly from the location Costalaresc. As in the present specimens, several of Zardini’s (1978) specimens have drill holes.
Atorcula canalifera (Münster, 1841)
Fig. 16C–E.
1998 Atorcula canalifera (Münster, 1841 sensu Kittl 1894); Nützel 1998: 161–162, pl. 26: F(?), G–I. (additional synonymy therein)
Material.—Twelve specimens, 2 from bulk samples, 10 from surface samples; 7 from Lago Antorno (PZO 12819, 12888, 2 figured specimens; PZO 12818, 5 specimens); 5 from Misurina Landslide (PZO 12697, figured specimen; PZO 12696, 4 specimens); Italy, Carnian, Triassic.
Description.—Shell high-spired slender, acutely conical; teleoconch fragment of 6 whorls 14.9 mm high, 6.3 mm wide; whorl face straight to very slightly convex; whorls smooth or with faint spiral striation; suture distinct; base flatly convex with several spiral furrows. Juvenile specimen (Fig. 16E) with relatively large larval shell with distinct axial ribs and a single smooth teleoconch whorl is preserved.
Remarks.—The material at hand is very close to Kittl’s (1894: pl. 4: 45, 46) type material of Atorcula canalifera (see Nützel 1998).
Stratigraphic and geographic range.—Carnian, Upper Triassic; northern Italy (Cassian Formation).
Family Zygopleuridae Wenz, 1938
Subfamily Ampezzopleurinae Nützel, 1998
Genus Ampezzopleura Bandel, 1991
Type species: Ampezzopleura tenuis Nützel, 1998; Late Triassic, northern Italy.
Ampezzopleura hybridopsis Nützel, 1998
Fig. 17A–G.
Material.—One hundred thirty four specimens, 124 from bulk samples, 10 from surface samples; 38 from Lago Antorno (PZO 12831–12833, 3 figured specimens; PZO 12830, 35 specimens); 96 from Misurina Landslide (PZO 12705–12708, 4 figured specimens; PZO 12704, 92 specimens); Italy, Carnian, Triassic.
Description.—Shell high-spired, slender; largest specimen from Lago Antorno comprises 8–9 whorls, 1.9 mm high, 0.8 mm wide; protoconch 4.5–5 whorls, 0.94 mm high, 0.54 mm wide; protoconch whorls somewhat convex; initial whorls almost flat; remaining larval shell high-spired; first two whorls smooth; remaining larval whorls with strong somewhat opisthocyrt axial ribs numbering 15–20 per whorl; axial ribs rather sharp, much narrower than interspaces between them; larval axial ribs curve strongly forward just above lower suture, becoming thinner at the same time and fusing to a sharp, suprasutural spiral thread which forms angular edge between base and whorl face of larval whorls; base of larval whorls flat to slightly convex; larval ribs do not continue onto base; axial ribs reduced to subsutural row of nodules in last half whorl of larval shell; larval shell ends at wide opisthocyrt arc which is poorly demarcated and not strengthened by a varix; teleoconch whorls slightly convex with broad, wave-like axial ribs numbering about ten per whorl; sutures distinct; base rather flat to slightly convex, joining whorl face at an angle; ribs do not continue onto base; base flat to slightly convex; base and whorls face meet at rounded edge.

Fig. 17. Zygopleurid gastropods from Lago Antorno (A–C) and Misurina Landslide (D–H), northern Italy, Cassian Formation, Carnian, Upper Triassic. A. Ampezzopleura hybridopsis Nützel, 1998, PZO 12831, axially ribbed larval shell and 3 teleoconch whorls, in lateral view. B. Ampezzopleura hybridopsis Nützel, 1998, PZO 12832, axially ribbed larval shell and 1 teleoconch whorl, in lateral view (B1), B2, last whorl axial ribs reduced to subsutural nodes, in lateral view; B3, earliest whorls smooth, then strong axial ribs, in lateral view. C. Ampezzopleura hybridopsis Nützel, 1998, PZO 12833, axially ribbed larval shell. D. Ampezzopleura hybridopsis Nützel, 1998, PZO 12705, axially ribbed larval shell and 2 teleoconch whorls, in apertural (D1), lateral (D2), and apical (D3) views. E. Ampezzopleura hybridopsis Nützel, 1998, PZO 12706, teleoconch. F. Ampezzopleura hybridopsis Nützel, 1998, PZO 12707, axially ribbed larval shell and 3 teleoconch whorls, in lateral view (F1), F2, protoconch in lateral view, axial ribs of larval shell reduced to subsutural nodes on last whorl; F3, protoconch in lateral view, axial ribs of larval shell reduced to subsutural nodes and teleoconch ribs begin to appaer on last whorl. G. Ampezzopleura hybridopsis Nützel, 1998, PZO 12708, axially ribbed larval shell and 4 teleoconch whorls, in lateral view. H. Ampezzopleura bandeli Nützel, 1998, PZO 12703, protoconch and ca. 4 teleoconch whorls.
Remarks.—The specimens at hand are very close to the type material from Campo near Cortina (type locality) and Alpe di Specie as illustrated and described in Nützel (1998). The reduction of the larval ribs on the last larval whorl to a subsutural row of nodules was interpreted by Nützel (1998) as evidence for a close phylogenetic relationship between Ampezzopleurinae (with axial ribs on larval shell) and Zygopleuridae with a subsutural row of nodules or riblets on larval whorls.
Stratigraphic and geographic range.—Carnian, Upper Triassic; northern Italy (Cassian Formation).
Ampezzopleura bandeli Nützel, 1998
Fig. 17H.
Material.—Twenty four specimens, 21 from bulk samples, 3 from surface samples; all from Misurina Landslide (PZO 12703, 1 figured specimen; PZO 12702, 23 specimens); Italy, Carnian, Triassic.
Remarks.—The illustrated specimen (Fig. 17H) from Misurina Landslide is deformed over its entire shell length (shell 1.6 mm high; protoconch about 0.3 mm high). It has preserved its small, bulbous larval shell that is typical of that species (Nützel 1998).
Ampezzopleurinae indet.
Material.—One specimen (PZO 12712) from Misurina Landslide bulk sample; Italy, Carnian, Triassic.
Remarks.—An isolated globular protoconch with strong axial ribs, about 0.5 mm; such larval shells are found in the genus Striazyga Nützel, 1998 or in Ampezzopleura bandeli.
Genus Zygopleura Koken, 1892
Type species: Turritella hybrida Münster, 1841; Late Triassic, northern Italy.
Zygopleura sp.
Fig. 18.
Material.—Two larval shells, both from bulk samples; 1 from Lago Antorno (PZO 12711); 1 from Misurina Landslide (PZO 12836); Italy, Carnian, Triassic.
Description.—Broadly conical larval shells with immersed initial whorls; whorls smooth except for sub- and suprasutural rows of minute nodules; specimen from Lago Antorno (Fig. 18A) broken; 0.67 mm high, 0.57 mm wide; specimen from Misurina Landslide (Fig. 18B) 1.02 mm high, 0.85 mm wide.

Fig. 18. Zygopleurid larval shells of Zygopleura sp. from Lago Antorno (A) and Misurina Landslide (B), northern Italy, Cassian Formation, Carnian, Upper Triassic. PZO 12836 (A) and PZO 12711 (B), in lateral view.
Remarks.—The larval shells at hand are typical of zygopleurid protoconchs (Bandel 1991; Nützel 1998).
Family Settsassiidae Bandel, 1992
Genus Kittliconcha Bonarelli, 1927
Kittliconcha aonis (Kittl, 1894) comb. nov.
Fig. 19A–C.
1894 Pseudomelania? aonis n. sp.; Kittl 1894: 194, pl. 6: 32–34, pl. 8: 19.
1978 Pseudomelania? aonis; Zardini 1978: 46, pl. 41: 3.
Material.—Four specimens, all from surface samples; 2 from Lago Antorno (PZO 12834, 12835, 2 figured specimens); 2 from Misurina Landslide (PZO 12710, 1 figured specimen; PZO 12709, 1 specimen); Italy, Carnian, Triassic.
Description.—High-spired shell with very low, convex whorls that are ornamented with orthocline, broad irregular axial ribs; ribs are as wide as interspaces, irregular in strength, number and spacing; growth lines orthocline, distinct, irregular; axial ribs tending to be reduced on last preserved whorls; suture distinct; base convex with traces of spiral lirae and umbilical chink.
Remarks.—The present specimens closely resemble Kittl’s (1894) type specimens and those illustrated by Zardini (1978: pl. 41: 3). Kittl (1894) placed this species tentatively in the genus Pseudomelania. However, Pseudomelania is based on Cretaceous steinkerns and is therefore problematic. We place Pseudomelania? aonis in Kittliconcha, which has a similar gross morphology and axial ribbing (see Nützel 2010). The type species Kittliconcha obliquecostata has higher whorls and oblique axial ribs. The Palaeozoic genus Palaeostylus Mansuy, 1914, is also similar. The species Kittliconcha aonis is insufficiently known i.e., its protoconch is unknown and its aperture is insufficiently known. Therefore, its systematic placement may change in the future.
Stratigraphic and geographic range.—Carnian, Upper Triassic; northern Italy (Cassian Formation).
Genus Tyrsoecus Kittl, 1892
Type species: Tyrsoecus cassiani Cox, 1960; Late Triassic, northern Italy.
Remarks.—The genus Tyrsoecus is problematic because it was based on Turritella compressa Münster, 1841 (Tyrsoecus cassiani Cox, 1960, is a replacement name) which was based on a poorly illustrated teleoconch fragment. According to Kittl (1894), the type specimen is lost.
Tyrsoecus zeuschneri (Klipstein, 1843)
Fig. 19D, E.
1843 Turritella zeuschneri n. sp.; Klipstein 1843: 178, pl. 11: 24.
1849 Loxonema zeuschneri (Klipstein); Orbigny 1849: 187.
1894 Coronaria? zeuschneri (Klipstein); Kittl 1894: 167.
?1894 Coronaria subcompressa n. sp.; Kittl 1894: 166, pl. 4: 31, 32.
1909 Tyrsoecus zeuschneri (Klipstein); Cossmann 1909: 35.
1926 Stephanocosmia? zeuschneri (Klipstein); Diener 1926: 199.
1940 Stephanocosmia (Tyrsoecus) zeuschneri (Klipstein); Kutassy 1940: 369.
non 1978 cfr. Coronaria zeuschneri (Klipstein); Zardini 1978: 41, pl. 26: 25.
Material.—One shell (PZO 12837) from Lago Antorno surface sample, Italy, Carnian, Triassic. Klipstein’s (1843) illustrated type specimen (NHMUK PI OR 35421(1)) from unspecified locality, “St. Cassian” Formation.
Description.—Shell high-spired, present specimen from Lago Antorno is a teleoconch fragment of four whorls, 8.9 mm high, 4.6 mm wide specimen (flattened by compaction; 5.3 and 3.9 mm wide); the holotype from the Klipstein collection comprises c. six whorls, is 16.2 mm high, 7.0 mm wide; whorls low, stout, slightly adpressed; sutures distinct; whorls convex with slight median angulation which bears a faint spiral rib; additional faint spiral present; whorl with relatively weak axial ribs (12–14 per whorl) which do not reach sutures and are strongest at mid-whorl; whorl with faint, mostly obscured micro-striation; base flatly convex, without prominent ornament.

Fig. 19. Caenogastropods from Lago Antorno (A, B, D), Misurina Landslide (C) and Klipstein’s (1843) collection (E), northern Italy, Cassian Formation, Carnian, Upper Triassic. A. Kittliconcha aonis (Kittl, 1894), PZO 12834, in lateral view. B. Kittliconcha aonis (Kittl, 1894), PZO 12835, in lateral view. C. Kittliconcha aonis (Kittl, 1894); PZO 12710, in lateral view. D. Tyrsoecus zeuschneri (Klipstein, 1843); PZO 12837, in lateral view. E. Tyrsoecus zeuschneri (Klipstein, 1843), Klipstein’s (1843) illustrated type specimen (NHMUK PI OR 35421(1)), in lateral (E1) and apertural (E2) views.
Remarks.—Tyrsoecus zeuschneri seems to be a rare species. Since its initial description, only a single specimen was assigned to this species as “cfr. Coronaria zeuschneri” from Alpe di Specie (Zardini 1978: 41–42, pl. 26: fig. 1; written as “cfr. Coronaria subcompressa” in the plate caption). However, Bandel (1995) listed Zardini’s (1978) specimen in the synonymy list of the mathildoid species Camponaxis lateplicata (Klipstein, 1843). Indeed, Zardini’s (1978) specimen is not very close to Klipstein’s (1843) illustration and type specimen. This illustration is inaccurate; it overemphasizes the axial ribs and does not show the slight median angulation. The type specimen housed in the Natural History Museum in London (NHMUK PI OR 35421(1)) agrees well with the specimen from the present collection. Tyrsoecus subcompressus (Kittl, 1894) is close to T. zeuschneri and could represent a synonym.
Stratigraphic and geographic range.—Carnian, Upper Triassic; northern Italy (Cassian Formation).
Family Ladinulidae Bandel, 1992
Genus Flemingella Knight, 1936
Type species: Trochella prisca McCoy, 1844; Millicent, Clare, Ireland, Carboniferous.
Flemingella bistriata (Münster, 1841)
Fig. 12D.
1841 Trochus bistriatus n. sp.; Münster 1841: 108, pl. 11: 16.
1891 Flemmingia bistriata; Kittl 1891: 253, pl. 7: 14–16.
1992 Flemmingia bistriata; Bandel 1992: 47, pl. 4: 4, 5.
Material.—One shell (PZO 12713) from the Misurina Landslide surface sample, Italy, Carnian, Triassic.
Remarks.—This shell that is about 8 mm high, closely resembles the specimens illustrated by Kittl (1891) and Bandel (1992). There are several other Cassian species that have been assigned to Flemmingia. Flemmingia bicarinata (Klipstein, 1843) as reported by Kittl (1891) is very similar to our specimen. However, this species has been considered a synonym of Flemmingia bistriata by Bandel (1992).
Stratigraphic and geographic range.—Only known from the Carnian, Upper Triassic; northern Italy (Cassian Formation).
Subclass Heterobranchia
Family Mathildidae Dall, 1889
Genus Jurilda Gründel, 1973
Type species: Mathilda (Jurilda) crasova Gründel, 1973; OD; Middle Jurassic, Poland.
Remarks.—The type species of Tirolthilda Bandel, 1995, Tirolthilda seelandica Bandel, 1995, closely resembles the type species of Jurilda and therefore we consider Tirolthilda to represent a junior synonym of Jurilda. Mathilda (Jurilda) crasova Gründel, 1973 according to Gründel (1997a) is a synonym of Promathildia (Teretrina) concava Walther, 1951 (material from Germany).
Jurilda elongata (Leonardi and Fiscon, 1959) comb. nov.
Fig. 20.
1959 Promathildia (?) peracuta n. var. elongata; Leonardi and Fiscon 1959: pl. 9: 14, 15, 20.
1978 Promathildia peracuta f. elongata (Leonardi-Fiscon); Zardini 1978: pl. 35: 24, 25, non 23.
1995 Tirolthilda seelandica n. sp.; Bandel 1995: 13, pl. 5: 6, 8–10.
Material.—One hundred and seventy specimens, 171 from bulk samples, 1 from surface samples; 5 from Lago Antorno (PZO 12838, 5 specimens); 167 from Misurina Landslide (PZO 12716–12720, 5 figured specimens; PZO 12635, 162 specimens; Italy, Carnian, Triassic.
Description.—Shell high-spired, slender, largest illustrated specimen 3.7 mm high, 1.4 mm wide (apex broken off); two prominent spiral cords present immediately after protoconch; spiral cords relatively close to each other on early whorls and widely separated on later whorl; whorl angulated at spiral cord so that whorl face is bevelled towards sutures; whorl straight to slightly concave between spiral cords; whorl angulated with fine axial threads which are much weaker than spiral cords; axial threads straight between spiral cords, prosocline above adapical spiral cord and opisthocline below abapical spiral cord; number of axial threads increases to up to more than 30 per mature teleoconch whorls; bordering spiral cord emerges at suture and is fully exposed on base.

Fig. 20. Mathildid gastropod Jurilda elongata (Leonardi and Fiscon, 1959) from Misurina Landslide, northern Italy, Cassian Formation, Carnian, Upper Triassic. A. PZO 12716, in lateral view (A1), A2, early whorls including heterostrophic protoconch, in lateral view. B. PZO 12717, in apical (B1) and lateral (B2) views; B3, early whorls including heterostrophic protoconch, in lateral view. C. PZO 12718, in apertural (C1) and oblique basal (C2) views. D. PZO 12719, in lateral (D1) and oblique basal (D2) views. E. PZO 12720, detail teleoconch whorl, in lateral view.
Remarks.—Leonardi and Fiscon (1959) described this species from Costalaresc as variety of Flemingia bicarinata Kittl, 1891 (placed tentatively in Promathildia by them), a species from the Middle Triassic Esino Limestone. Since this variety was published before 1961 it has subspecific rank (ICZN Article 45.6.4.). Leonardi and Fiscon (1959) discussed several characters separating Flemingia bicarinata from their new variety. Therefore, they clearly had the intention to describe a new taxon and not an infrasubspecific name as becomes obvious from the synonymy list provided by them. Bandel (1995) was aware that his material is conspecific with the variety described by Leonardi and Fiscon (1959) but nevertheless described the species as new (Tirolthilda seelandica). We raise Promathildia (?) peracuta n. var. elongata to species rank, there is no doubt that the morphological differences mentioned by Leonardi and Fiscon (1959) (ribbing, shape) are sufficient to raise it from subspecies rank. Tirolthilda seelandica Bandel, 1995, falls in synonymy of Jurilda elongata.
The Middle Jurassic type species of Jurilda as reported by Walther (1951), Gründel (1973, 1997a) and Kaim (2004) closely resembles Jurilda elongata from the Cassian Formation. The only major difference is that the apical carination is stronger and thus the whorl face above it is oblique instead of parallel to the shell axis. Gründel and Nützel (2013) gave an emendated diagnosis of Jurilda which fits the present material well. Especially the ornament with two primary spiral cords close to the sutures which are crossed by fine axial threads is similar. Thus placement of Jurilda seelandica in Jurilda and the synonymisation of Jurilda and Tirolthilda are justified. It is the first report of this genus from the Triassic.
Bandel (1995) reported about 40 specimens from the type locality Alpe di Specie (Seelandalpe) and three specimens from Misurina. Zardini (1978) reported that it is very abundant at Misurina, abundant at Alpe di Species and rare at Costalaresc and Campo. Here it is one of the most abundant species at Misurina Landslide but very rare at Lago Antorno.
Stratigraphic and geographic range.—Only known from the Carnian, Upper Triassic; northern Italy (Cassian Formation).
Genus Promathildia Andreae, 1887
Type species: Mathilda janeti Cossmann, 1885; subsequent designation by Gründel and Nützel (2013); Europe, Jurassic.
Promathildia cf. milierensis Zardini, 1980
Fig. 21C–H.
Material.—Twenty specimens, 19 from bulk samples, 1 from surface sample; all from Misurina Landslide (PZO 12722–12727, 6 figured specimens; PZO 12721, 14 specimens); Italy, Carnian, Triassic.
Description.—Shell high-spired, slender; whorl face distinctly angulated low on the whorls; whorl ornamented with two spiral cords and ca. 10 axial ribs per whorl; adapical spiral cord weak to absent, in subsutural position; abapical spiral cord strong, forming angulation and periphery, somewhat above abapical suture but higher on whorl face in earlier whorls; nodes at intersections of spiral cords and axial ribs, strong on abapical spiral cord, weak on adapical cord; whorl face with fine spiral threads; protoconch consisting of somewhat more than one whorl with a diameter of the first whorl 0.24 mm; protoconch smooth except a very fine spiral striation; first teleoconch whorls angulated but without distinct ornament; teleoconch ornament develops from the third teleoconch whorl onward; base flat with spiral ornament and a spiral cord at angulation at border to whorl face.

Fig. 21. Heterobranch gastropods from Misurina Landslide, northern Italy, Cassian Formation, Carnian, Upper Triassic. A. Promathildia cf. decorata (Klipstein, 1843), PZO 12729, in lateral (A1) and apical (A3) views; A2, close up to show heterostrophic protoconch, in lateral view. B. Promathildia subnodosa (Münster, 1841), PZO 12730, in apertural view. C. Promathildia cf. milierensis Zardini, 1980; PZO 12722, in apertural view (C1); C2, close up to show heterostrophic protoconch, in lateral view; C3, close up to show heterostrophic protoconch, transition to teleoconch well visible, in lateral view. D. Promathildia cf. milierensis Zardini, 1980, PZO 12725, in lateral view. E. Promathildia cf. milierensis Zardini, 1980, PZO 12723, in lateral view. F. Promathildia cf. milierensis Zardini, 1980, PZO 12726, in lateral view. G. Camponaxis lateplicata (Klipstein, 1843), PZO 12731, in lateral view. H. Promathildia cf. milierensis Zardini, 1980, PZO 12727, in lateral view. I. Promathildia cf. milierensis Zardini, 1980; PZO 12724, in lateral view.
Remarks.—The main characteristic of this species is the pronounced carina with strong, almost spiny nodules low on the whorls and a much weaker subsutural spiral cord. The protoconch with only about one whorl and a relatively large diameter suggest that this species had non-planktotrophic larval development. The specimens are more or less encrusted or fragmented making an identification difficult. They resemble Promathildia milierensis as illustrated by Zardini (1980: pl. 5: 12, 14, 15, but not 13) and Hausmann and Nützel (2015: fig. 8I) but these specimens have the knobby angulation higher on the whorls. Tofanella cancellata Bandel, 1995, is similar but has more spiral cords and it lacks strong to almost spine-like nodes on the lower spirals. Promathildia colon (Münster, 1841) and P. pygmaea (Münster, 1841) as illustrated by Kittl (1894) and Zardini (1978) are similar but examination of Münster’s type specimens of these species showed that they are probably not conspecific with our specimens.
Promathildia cf. decorata (Klipstein, 1843)
Fig. 21A.
Material.—Two shells (PZO 12729, 12728) from Misurina Landslide bulk samples, Italy, Carnian, Triassic.
Description.—Two juvenile specimens with preserved protoconch show strongly angulated whorls; spiral carina below mid-whorl, additional spiral cord present below suture; additional spiral cords on base; protoconch coaxial heterostrophic with radial fold at the end of larval shell; axial ornament either weak or absent, obscured due to preservation.
Remarks.—These two juvenile specimens resemble the early whorls of Promathildia decorata (Klipstein, 1843) as reported by Bandel (1995) who also reported the same type of coaxial protoconch with folds. In addition, Bandel’s (1995) specimens have axial threads as teleoconch ornament which might be obscured due to preservation in our specimens.
Promathildia subnodosa (Münster, 1841)
Fig. 21B.
1841 Fusus subnodosus n. sp.; Münster 1841: 124, pl. 13: 51.
1894 Promathildia subnodosa; Kittl 1894: 243, pl. 9: 36–45.
1978 Promathildia subnodosa; Zardini 1978: 50–51, pl. 35: 4–9.
?1995 Promathildia subnodosa; Bandel 1995: 8, pl. 2: 5, 7, pl. 3: 1–4, 6, 7.
2015 Promathildia subnodosa; Hausmann and Nützel 2015: fig. 8H.
Material.—One teleoconch fragment (PZO 12730) from Misurina Landslide surface sample, Italy, Carnian, Triassic.
Description.—A single teleoconch fragment consisting of about 5 whorls, 12 mm high, 4.7 mm wide; whorls convex with median angulation; growth lines distinct, sometimes thread-like, orthocline below suture and then curving forward so that they become markedly opisthocline in the two thirds of whorls; two prominent spiral cords, one at mid-whorl angulation, the other half way between mid-whorl and abapical suture; both main spiral cords undulating to nodular at intersections with opisthocline axial ribs, numbering 10–14 per whorl; a third faint spiral cord somewhat below suture; base convex, with two distinct spiral cords and additional spiral lirae.
Remarks.—This is a rather variable species; the specimen at hand is close to some of the specimens illustrated by Kittl (1894) and Zardini (1978) as P. subnodosa. The identity of the specimens illustrated by Bandel (1995) is doubtful because they represent early juvenile shell portions only.
Stratigraphic and geographic range.—Carnian, Upper Triassic; northern Italy (southern Alps, Cassian Formation; Apulia, Raibl Formation) (Diener 1926; Kutassy 1940).
Family Tofanellidae Bandel, 1995
Genus Camponaxis Bandel, 1995
Type species: Cerithium(?) lateplicatum Klipstein, 1843; northern Italy, Late Triassic.
Camponaxis lateplicata (Klipstein, 1843)
Fig. 21G.
Material.—One shell (PZO 12731) from Misurina Landslide bulk sample, Italy, Carnian, Triassic.
Description.—Shell high-spired; whorls convex with slight angulation high on the whorls; whorl ornamented with axial ribs and spiral cords of about same strength with intersection not or only slightly nodular; axial ribs, straight orthocline, rounded, separated by wide interspaces, numbering up to 20 per whorl; spiral cords of variable strength and irregularly species; two adapical spiral cords particularly weak.
Remarks.—This specimen resembles Bandel’s (1995) illustrations of Camponaxis lateplicata (Bandel 1995: pl. 13: 9; pl. 14: 1–5) and is probably conspecific. However, it is questionable whether Bandel’s (1995) material is conspecific with Klipstein’s (1843) type specimen which AN has examined at the Museum of Natural History in London. This type specimen is relatively large, fusiform and has fewer but stronger axial ribs. It is therefore likely that Camponaxis has a misidentified type species.
Family Cornirostridae Ponder, 1990
Genus Bandellina Schröder, 1995
Type species: Bandellina laevissima Schröder, 1995; Poland, Early Cretaceous.
Remarks.—The genus was discussed by Bandel (1996) and Kaim (2004) who pointed out its resemblance to the modern sunken wood and hot vent species Hyalogyra Marshall, 1988. Kaim (2004) did not follow Bandel’s (1996) assignment to the family Cornirostridae but left the family assignment open.
Bandellina compacta Nützel and Hausmann sp. nov.
Fig. 22.
ZooBank LSID: urn:lsid:zoobank.org:act:6B8D87DF-6B27-402E-B0 A3-EB2908DB78DF
Etymology: In reference to the compact shell form.
Holotype: PZO 12840, specimen with larval shell and 2 teleoconch whorls.
Type locality: Lago Antorno, near Cortina d’Ampezzo, northern Italy.
Type horizon: Cassian Formation; lower Carnian, Upper Triassic.
Material.—Two specimens (PZO 12839, 12840) from Lago Antorno bulk samples, Italy, Carnian, Triassic.
Description.—Shell low-spired, turbiniform; specimen comprises 3.6 whorls (two larval), 0.9 mm wide, 0.7 mm high; protoconch blunt, comprising about two whorls; initial whorl sinistral, sunken, smooth; second protoconch whorl dextral, round, convex with axial subsutural rib character of strengthened growth lines; protoconch clearly demarcated from teleoconch by a suture; teleoconch smooth, with evenly convex whorls embracing somewhat above periphery; base evenly convex; minutely phaneromphalous, covered with numerous fine spiral ribs.
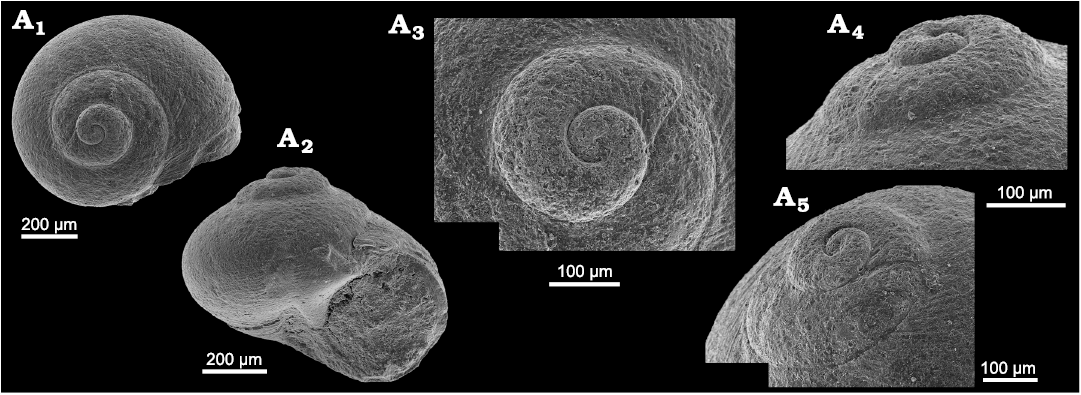
Fig. 22. Cornirostrid gastropod Bandellina compacta Nützel and Hausmann sp. nov. from Lago Antorno, northern Italy, Cassian Formation, Carnian, Upper Triassic. Holotype PZO 12840, in apical (A1) and apertural (A2) views; heterostrophic protoconch, in apical (A3), lateral (A4), and oblique lateral (A5) views.
Remarks.—Bandellina compacta differs from B. cassiana and B. laevissima in being more high-spired and in having a narrower umbilicus. Moreover, B. cassiana and B. laevissima lack spiral ribs on the base and axial riblets on the last larval whorl.
Stratigraphic and geographic range.—Only known from the Carnian, Upper Triassic; northern Italy (Cassian Formation).
Family Hyalogyrinidae Warén and Bouchet, 1993
Genus Alexogyra Bandel, 1996
Type species: Alexogyra marshalli Bandel, 1996; Italy, Late Triassic.
Remarks.—Kaim (2004) considered Alexogyra to represent a synonym of Bandellina Schröder, 1995. However, Alexogyra is almost planispiral whereas Bandellina is distinctly helicoidal and more massive. Therefore, a possible synonymy is considered here as unlikely although both genera are probably closely related.
Alexogyra marshalli Bandel, 1996
Fig. 23.
Material.—Seven specimens (PZO 12842–12845, 4 figured specimens; PZO 12841, 3 specimens), all from Lago Antorno bulk samples, Italy, Carnian, Triassic.
Description.—Shell almost planispiral with very slightly elevated spire; teleoconch flatly dextral; with about three whorls, 1.0 mm wide, 0.5 mm high; protoconch flat, comprising about 1.5 whorls, diameter 0.26 mm; initial whorls dipping, sinistral; remaining protoconch flatly dextral; protoconch smooth, with convex whorls; protoconch demarcated from teleoconch by a distinct suture and a bulge; teleoconch whorls smooth, convex, circular in transverse section; suture distinct; base distinctly phaneromphalous.
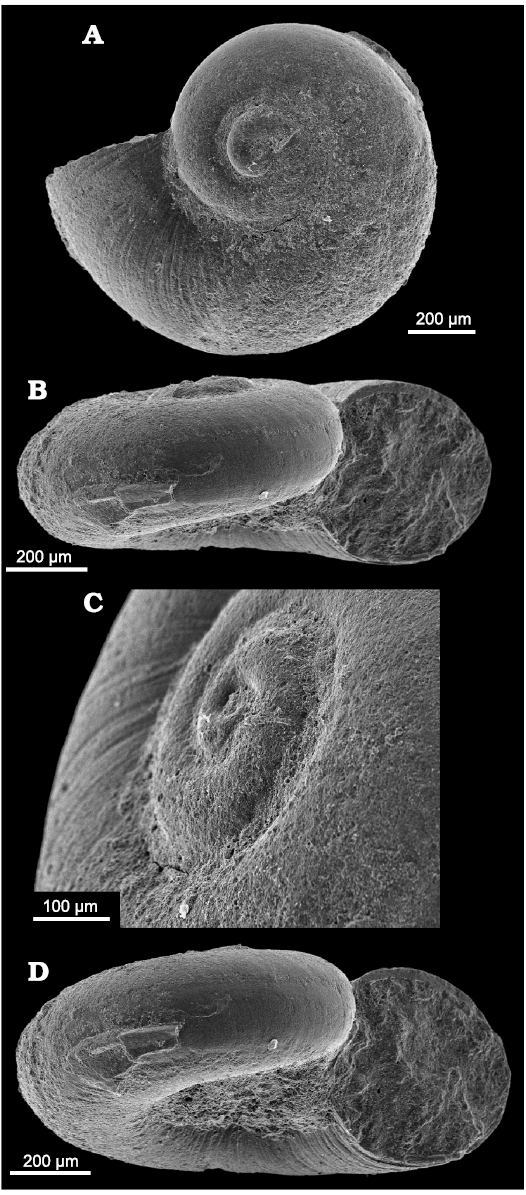
Fig. 23. Hyalogyrinid gastropod Alexogyra marshalli Bandel, 1996 from Lago Antorno, northern Italy, Cassian Formation, Carnian, Upper Triassic. A. PZO 12842, in apical view. B. PZO 12843, in apertural view. C. PZO 12844, protoconch in oblique lateral view. D. PZO 12845, in oblique apertural view to show base.
Remarks.—The present specimens agree well with Bandel’s (1996) type specimens from the type locality Misurina Skilift.
Stratigraphic and geographic range.—Only known from the Carnian, Upper Triassic; northern Italy (Cassian Formation).
Family Stuoraxidae Bandel, 1996
Genus Ampezzogyra Bandel, 1996
Type species: Ampezzogyra schroederi Bandel, 1996; Italy, Late Triassic.
Ampezzogyra angulata Nützel and Hausmann sp. nov.
Fig. 24.
ZooBank LSID: urn:lsid:zoobank.org:act:3DC97C77-0812-4E8B-91E6-AF5F4445F06D
Etymology: In reference to the angulated whorl profile.
Type material: Holotype (PZO 12847), paratypes (PZO 12733, 12734, 12848); four well-preserved specimens with preserved protoconchs. Holotype and paratype PZO 12848 from Lago Antorno, other paratypes from Misurina Landslide.
Type locality: Lago Antorno, near Cortina d’Ampezzo, northern Italy.
Type horizon: Cassian Formation; lower Carnian, Upper Triassic.
Material.—Twenty three specimens, all from bulk samples; 9 from Lago Antorno (PZO 12847, holotype; PZO 12848, paratype; PZO 12846, 7 specimens); 14 from Misurina Landslide (PZO 12733, 12734, paratypes; PZO 12732, 12 specimens); Italy, Carnian, Triassic.
Description.—Shell planispiral with somewhat depressed upper side (assuming dextral teleoconch) and somewhat deeper, widely phaneromphalous umbilicus; shell with about 3.4 whorls, 0.9 mm wide, 0.3 mm high; protoconch flat, comprising about 1.5 whorls, diameter 0.16–0.18 mm; initial whorls dipping, sinistral; protoconch smooth, with convex whorls; protoconch demarcated from teleoconch by a distinct suture and a bulge; teleoconch whorls convex at periphery, ornamented with seven to eight distinct spiral ribs which angulate the whorls somewhat especially at transition from periphery to upper side; axial ribs having character of strengthened growth lines; intersections of weak axial ribs and upper spiral rib somewhat pronounced, node-like, especially in early portions of teleoconch; teleoconch covered with micro-ornament of fine tubercles; shell structure crossed lamellar.
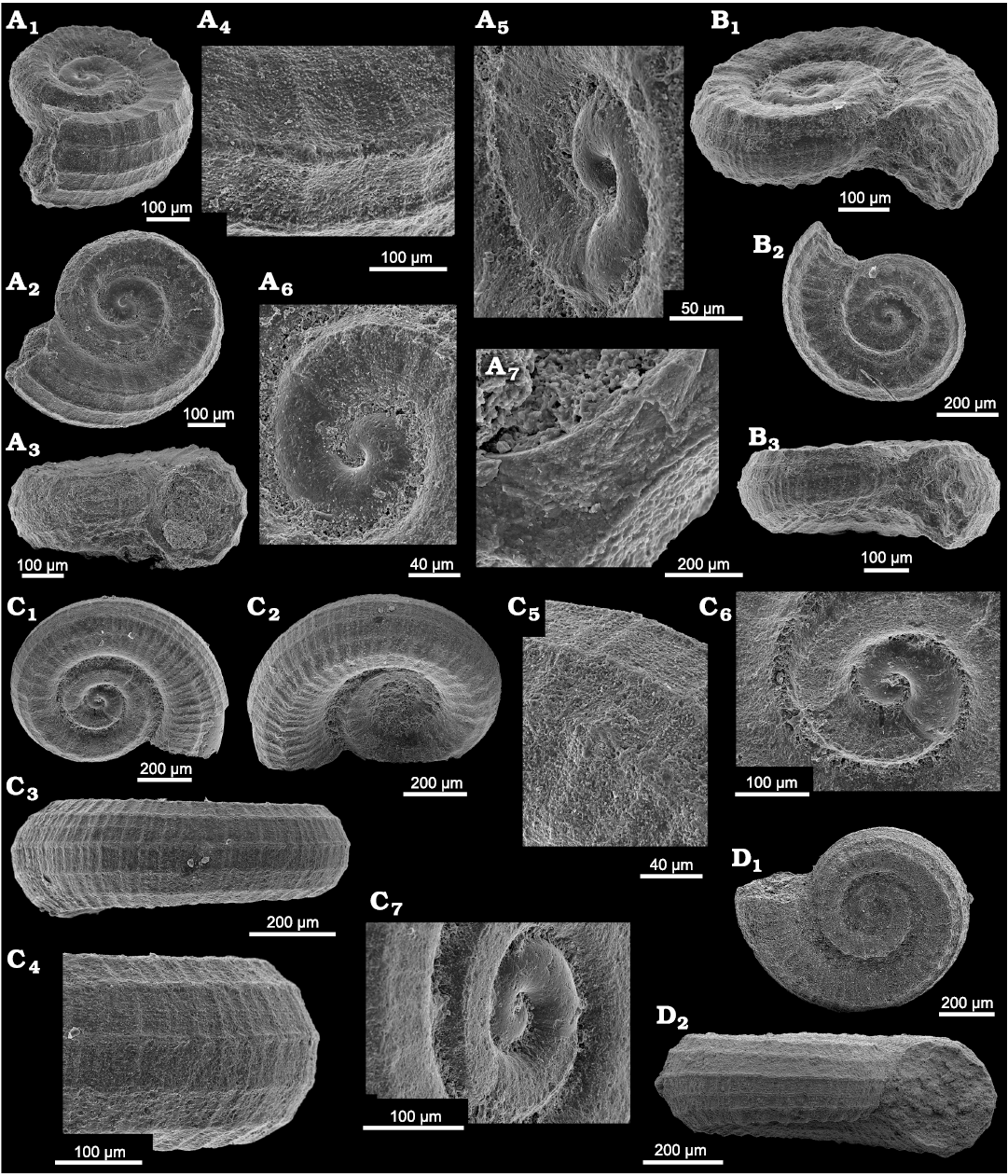
Fig. 24. Stuoraxid gastropod Ampezzogyra angulata Nützel and Hausmann sp. nov. from Lago Antorno (A, C) and Misurina Landslide (B, D), Cassian Formation, northern Italy, Late Triassic. A. Paratype PZO 12848, in oblique lateral (A1), apical (A2), and apertural (A3) views; A4, detail teleoconch at adapical whorl angulation with micro-ornament of fine tubercles; A5, protoconch in oblique lateral view; A6, protoconch in apical view; A7, crossed lamellar shell structure. B. Paratype PZO 12733, in oblique lateral (B1), apical (B2), and apertural (B3) views. C. Holotype PZO 12847, in apical (C1), basal (C2), and apertural (C3) views; C4, detail of C3; C5, detail teleoconch with micro-ornament of fine tubercles; C6, protoconch in apical view; C7, protoconch in oblique view. D. Paratype PZO 12734, in apical (D1) and apertural (D2) views.
Remarks.—Ampezzogyra angulata sp. nov. resembles the type species A. schroederi Bandel, 1996. However, A. schroederi has strong axial ribs and only subordinate, weak numerous spiral threads at which the whorls are not angulated so that the whorls are circular in transverse section. Moreover, the larval whorls have axial folds in A. schroederi while those of A. angulata are entirely smooth. Sloeudaronia karavankensis Kaim, Jurkovšek, and Kolar-Jurkovšek, 2006, from the Carnian of Slovenia is similar but has widely spaced distinct axial ribs and fewer, more distinct spiral ribs on the whorls. Kaim et al. (2006) placed Sloeudaronia in Vetigastropoda based on the poorly preserved protoconch which they interpreted consisting of one whorl and as being of the vetigastropod type. However, if this protoconch turns out to be heterostrophic, Sloeudaronia and Ampezzogyra could be considered synonyms. The Rinaldoconchus Bandel, 1996, from the Cassian Formation (R. bieleri and R. ampezzanus) is similar but has an elevated spire.
Stratigraphic and geographic range.—Only known from the Carnian, Upper Triassic; northern Italy (Cassian Formation).
Family Tubiferidae Cossmann, 1895
Genus Sinuarbullina Gründel, 1997b
Type species: Sinuarbullina ansorgi Gründel, 1997b; Poland, Middle Jurassic.
Sinuarbullina sp. 1
Fig. 25A, D.
Material.—Twenty four specimens, all from bulk samples; 10 from Lago Antorno (PZO 12850, 1 figured specimen; PZO 12849, 9 specimens); 14 from Misurina Landslide (PZO 12736, figured specimen; PZO 12735, 13 specimens); Italy, Carnian, Triassic.
Description.—Shell slender, fusiform with last whorl larger than spire, embracing above mid-whorl; illustrated specimens with about 2.5 teleoconch whorls; whorls smooth, evenly convex with narrow subsutural ramp which is demarcated from outer whorl face by an angular edge and a weak spiral furrow; aperture elongated, much higher than wide, narrow posteriorly, wide and rounded anteriorly; protoconch heterostrophic sinistral, transaxial, consisting of about two smooth whorls, 0.28 mm wide (Fig. 25A); illustrated specimen from Lago Antorno (Fig. 25A) 0.7 mm wide, 1.2 mm high; illustrated specimen from Misurina Landslide (Fig. 25D) 0.5 mm wide, 0.9 mm high.

Fig. 25. Heterobranch gastropods from Lago Antorno (A–C) and from Misurina Landslide (D, E), northern Italy, Cassian Formation, Carnian, Upper Triassic. A. Sinuarbullina sp. 1, PZO 12850, in apertural view. B. Sinuarbullina sp. 3, PZO 12852, in lateral view. C. Sinuarbullina sp. 3, PZO 12853, in lateral view. D. Sinuarbullina sp. 1, PZO 12736, in apertural view. E. Sinuarbullina sp. 2, PZO 12738, in lateral view.
Remarks.—This is a typical representative of the informal group of “shelled opisthobranchs” which occurs in the Early Triassic (Batten and Stokes 1986; Nützel 2005; Nützel and Schulbert 2005; Foster et al. 2017) and ranges into the Cretaceous (Kaim 2004).
Sinuarbullina sp. 2
Fig. 25E.
Material.—Four specimens, all from bulk samples; 2 from Lago Antorno (PZO 12851, 2 specimens); 2 from Misurina Landslide (PZO 12738, 1 figured specimen; PZO 12737, 1 specimen); Italy, Carnian, Triassic.
Description.—Illustrated specimen PZO 12738 from Misurina Landslide, 0.5 mm wide, 0.9 mm high.
Remarks.—Sinuarbullina sp. 2 differs from Sinuarbullina sp. 1 by having a lower spire. The protoconch size of both species is more or less the same.
Sinuarbullina sp. 3
Fig. 25B, C.
Material.—Two shells (PZO 12852, 12853) from Lago Antorno bulk samples, Italy, Carnian, Triassic.
Description.—Shell slender, 0.4 mm wide, 1.1 mm high (Fig. 25B).
Remarks.—This species is distinct from the two Sinuarbullina species in being more slender and having higher whorls and more oblique sutures. Moreover, the protoconch is smaller than in the other Sinuarbullina species.
Gastropoda indet. 1
Material.—One shell (PZO 12739) from Misurina Landslide bulk sample, Italy, Carnian, Triassic.
Remarks.—The specimen is poorly preserved. The shell is heavily encrusted and compressed. It resembles Camposcala biserta (Münster, 1841) to some degree, but the encrustation does not allow a clear assignment.
Gastropoda indet. 2
Material.—One shell (PZO 12854) from Lago Antorno bulk sample, Italy, Carnian, Triassic.
Remarks.—The shell is smooth, high-spired with low whorls; the apex is missing. Although distinct in this collection, this specimen cannot be identified.
Class Bivalvia Linnaeus, 1758
Order Nuculida Dall, 1889
Superfamily Nuculoidea Gray, 1824
Family Nuculidae Gray, 1824
Genus Palaeonucula Quenstedt, 1930
Type species: Nucula hammeri Defrance, 1825; France, Jurassic.
Palaeonucula strigilata (Goldfuss, 1837)
Fig. 26C, D.
1837 Nucula strigilata; Goldfuss 1837: 153–154, pl. 124: 18a–c.
1841 Nucula strigilata Goldf.; Münster 1841: 83, pl. 8: 10a, b.
1865 Nucula strigilata Goldf.; Laube 1865: 65, pl. 19: 2.
1895 Nucula strigilata Goldf.; Bittner 1895: 137–138, pl. 17: 1–17.
1977 Palaeonucula strigilata (Goldfuss); Fürsich and Wendt 1977: fig. 11.
1981 Palaeonucula strigilata (Goldfuss); Zardini 1981: pl. 1: 1–14, pl. 39: 11.
1988 Palaeonucula strigilata Goldfuss; Zardini 1988: pl. 1: 3, 4.
2012 Palaeonucula strigilata (Goldfuss, 1837); Urlichs 2012: 5, pl. 1: 3.1–3.3, 4, 8.
2014 Palaeonucula strigilata (Goldfuss, 1837); Nützel and Kaim 2014: 423, figs. 10a, d.
Material.—Twenty specimens, 12 from bulk samples, 8 from surface samples; 2 from Lago Antorno (PZO 12855, 2 specimens); 18 from Misurina Landslide (PZO 12742, 12743, 2 figured specimens; PZO 12741, 16 specimens); Italy, Carnian, Triassic.
Remarks.—All specimens are double-valved and the shells are often diagenetically compressed. In many cases, the co-marginal ribs are concealed by encrustation. Palaeonucula strigilata (Goldfuss, 1837) is a very common bivalve in Cassian deposits and was therefore also found at many other Cassian localities, including Costalaresc, Piz Stuores, Pralongia, Rio Pocol, Settsass Scharte and Stuores Wiesen (Fürsich and Wendt 1977; Urlichs 2012; Nützel and Kaim 2014). This species constitutes the second most abundant taxon in the Rhaphistomella radians/Palaeonucula strigilata association and even the most abundant species of the Palaeonucula strigilata/Dentalium undulatum association described by Fürsich and Wendt (1977).
Stratigraphic and geographic range.—Carnian, Upper Triassic; northern Italy (Cassian Formation), southern Italy (Sicily, Raibl Formation); Hungary (Bakony); Spain (Boyar section); Turkey (Anatolia); Switzerland (Röti Formation). Upper Triassic, Norian, Russia (Primorskiy Kray). Middle Triassic, Anisian–Ladinian, Vietnam. Triassic: Oman; Libya (Tripolis); Indonesia. Middle to Upper Triassic, China (Liyantang, Kuahongdong, Qiaotou). Jurassic, upper Bajocian–Bathonian, Poland (Polish lowland) (Diener 1923; Kutassy 1931; Paleobiology Database, https://paleobiodb.org).
Order Nuculanida Carter, Campbell, and Campbell, 2000
Superfamily Nuculanoidea Adams and Adams, 1858
Family Nuculanidae Adams and Adams, 1858
Genus Nuculana Link, 1807
Type species: Arca rostrata Bruguière, 1789; Europe, Recent.
Nuculana sulcellata (Wissmann, 1841)
Fig. 26A, E.
1841 Nucula sulcellata Wissm.; Münster 1841: 85, pl. 8: 15.
1843 Nucula sulcellata Wissm.; Klipstein 1843: 263, pl 17: 19.
1857 Nucula sulcellata Wissm.; Hauer 1857: 558, pl. 2: 11, 12.
1865 Leda sulcellata Wissmann sp.; Laube 1865: 68–69, pl. 19: 5.
1895 Leda sulcellata Wissm. sp. (emend.); Bittner 1895: 147–148, pl. 18: 10.
1981 Naculana [sic!] sulcellata (Muenster); Zardini 1981: pl. 3: 4, 6–13.
1988 Nuculana sulcellata Muenster; Zardini 1988: pl. 4: 34.
Material.—Ten specimens, 3 from bulk samples, 7 from surface samples; 2 from Lago Antorno (PZO 12857, 1 figured specimen; PZO 12856, 1 specimen); 8 from Misurina Landslide (PZO 12745, figured specimen; PZO 12744, 7 specimens); Italy, Carnian, Triassic.
Remarks.—Single- and double-valved shells occur in the samples. The characteristic long rostrum at the posterior end is broken in all specimens. Nevertheless, the shells fit best to Bittner’s (1895), Laube’s (1865) and Zardini’s (1981) illustrated specimens of this species. This bivalve species is relatively common in the Cassian Formation and was also found at the localities Costalaresc, Rumerlo, Alpe di Specie and Stuores Wiesen (see Roden et al. 2020a and references therein). Bittner (1895) mentioned that Münster’s (1841) type specimen of this species does not fit to Münster’s (1841) respective description and illustrations and assumed that it is not the real holotype. He therefore described it as Leda wissmanniana sp. nov. and emphasised that there are doubts if the type specimen really comes from the Cassian Formation.
Stratigraphic and geographic range.—Carnian, Upper Triassic; northern Italy (Cassian Formation). Norian, Russia; Lacian–Alaunian, Peru (Suta Formation) (Diener 1923; Paleobiology Database, https://paleobiodb.org).

Fig. 26. Nuculid bivalves from Lago Antorno (A, B) and Misurina Landslide (C–E), northern Italy, Cassian Formation, Carnian, Upper Triassic. A. Nuculana sulcellata (Wissmann, 1841), PZO 12857, in lateral view. B. Palaeoneilo elliptica (Goldfuss, 1837), PZO 12859, in lateral (B1) and dorsal (B2) views. C. Palaeonucula strigilata (Goldfuss, 1837), PZO 12742, in lateral (C1) and dorsal (C2) views. D. Palaeonucula strigilata (Goldfuss, 1837), PZO 12743, in lateral view. E. Nuculana sulcellata (Wissmann, 1841), PZO 12745, in lateral (E1) and dorsal (E2) views.
Family Malletiidae Adams and Adams, 1858
Genus Palaeoneilo Hall and Whitfield, 1869
Type species: Nuculites constricta Conrad, 1842; USA, Devonian.
Palaeoneilo distincta (Bittner, 1895)
1895 Leda (?) distincta n. sp.; Bittner 1895: 150–152, pl. 16: 38, 39.
1981 Palaeoneilo distincta f. laubei (Bittner); Zardini 1981: pl. 3: 14, 15.
Material.—Four specimens, all from bulk samples; 2 from Lago Antorno (PZO 12858, 2 specimens); 2 from Misurina Landslide (PZO 12746, 2 specimens); Italy, Carnian, Triassic.
Remarks.—All specimens have the same outline and show only a weak sculpture of slightly strengthened growth lines similar to Bittner’s (1895) and Zardini’s (1981) illustrations of Palaeoneilo distincta. The umbo is central, the shell outline circular to oval-shaped. The specimens studied herein also resemble Bittner’s (1895) Leda (?) zelima d’Orbigny, 1849, to some degree, however, this species is more oval than our shells. All four specimens are double-valved.
Stratigraphic and geographic range.—Carnian, Upper Triassic; northern Italy (Cassian Formation); Hungary (Bakony) (Diener 1923; Paleobiology Database, https://paleobiodb.org).
Palaeoneilo elliptica (Goldfuss, 1837)
Fig. 26B.
1837 Nucula elliptica sp. nov.; Goldfuss 1837: 153, pl. 124: 16 (pars).
1841 Nucula elliptica Goldf.; Münster 1841: 83, pl. 8: 8.
1843 Nucula tenuis ?; Klipstein 1843: 263, pl. 17: 17.
1848 Leda elliptica; Deshayes 1848: 278.
1849 Leda elliptica; d’Orbigny 1849: 197.
1852 Nucula Münsteri; Giebel 1852: 389 (pars.).
1864 Nucula elliptica; Alberti 1864: 102.
1864 Leda elliptica; Laube 1864: 407.
1865 Leda elliptica Goldf. sp.; Laube 1865: 67–68, pl. 19: 6.
1895 Palaeoneilo elliptica Goldf. spec.; Bittner 1895: 142–143, pl. 16: 26–31.
1981 Palaeoneilo elliptica (Goldfuss); Zardini 1981: pl. 2: 12–14, pl. 3: 1, 2.
Material.—Three specimens, all from surface samples, 1 from Lago Antorno (PZO 12859, figured specimen); 2 from Misurina Landslide (PZO 12747, 2 specimens); Italy, Carnian, Triassic.
Remarks.—All specimens from Lago Antorno and Misurina Landslide are articulated. They fit best to Zardini’s (1981) illustrated shells of Palaeoneilo elliptica (Goldfuss, 1837). Also Bittner’s (1895) illustrated specimens are similar in shape to our specimens.
Stratigraphic and geographic range.—Carnian, Upper Triassic; northern Italy (Cassian Formation); southern Italy (Sicily); Germany (Muschelkalk, Lettenkohle); Switzerland; Hungary (Bakony); Libya (Tripolis). Rhaetian, Argentina (Mendoza, Arroyo Malo Formation). Anisian, Bosnia and Herzegovina. Triassic: China; Japan; Russia. Sinemurian, Mexico (Sonora, Sierra de Santa Rosa Formation). Jurassic, Portugal. Hettangian, United Kingdom (Blue Lias Formation) (Kutassy 1931; Paleobiology Database, https://paleobiodb.org).
Order Mytilida Férussac, 1822
Superfamily Mytiloidea Rafinesque, 1815
Family Mytilidae Rafinesque, 1815
Genus Modiolus Lamarck, 1799
Type species: Mytilus modiolus Linnaeus, 1758; Mediterranean, Recent.
Modiolus paronai Bittner, 1895
1895 Modiola Paronai nov. spec.; Bittner 1895: 48, pl. 5: 19, 20.
1981 Modiolus paronai (Bittner); Zardini 1981: pl. 6: 16, 17, pl. 7: 1, 2.
2015 Modiolus paronai; Hausmann and Nützel 2015: fig. 9G.
Material.—One valve (PZO 12748) from Misurina Landslide bulk sample, Italy, Carnian, Triassic.
Remarks.—The single-valved specimen is heavily encrusted and, with a length of ca. 1 mm, very small. However, it closely resembles Zardini’s (1981) illustrated specimens of Modiolus paronai Bittner, 1895. M. paronai Bittner, 1895 was also found at the Cassian locality Stuores Wiesen, where it was represented by 15 specimens (Hausmann and Nützel 2015).
Stratigraphic and geographic range.—Carnian, Upper Triassic; northern Italy (Cassian, Gorno, and Lombardische formations); Libya (Tripolis); Argentina(?), Middle Triassic; Jordan (Wadi Hesban); Slovenia (Julian); China (Leidapo Member) (Diener 1923; Kutassy 1931; Paleobiology Database, https://paleobiodb.org).
Order Ostreida Férussac, 1822
Superfamily Pterioidea Gray, 1847 (1820)
Family Bakevelliidae King, 1850
Genus Gervillia Defrance, 1820
Subgenus Cultriopsis Cossmann, 1904
Type species: Gervilleia (Cultriopsis) falciformis Cossmann, 1904; France, Jurassic.
Gervillia (Cultriopsis) sp.
Material.—Four specimens (PZO 12749); 3 from Misurina Landslide bulk samples; 1 from Misurina Landslide surface sample; Italy, Carnian, Triassic.
Remarks.—Only fragments were found in the samples from Misurina Landslide. Four of them were single-valved (to account for an estimation of the real number of bivalve individuals, this number was divided by two), two were double-valved. Our specimens were compared with Gervillia (Cultriopsis) species from Bittner (1895) (who spelled this genus Gervilleia throughout his monograph), Zardini (1981) and Hautmann (2001). Because of their fragmented nature, a species identification was not possible. Nevertheless, the characteristic form of the large byssal notch assign the Misurina Landslide specimens to this genus.
Family Cassianellidae Ichikawa, 1958
Genus Cassianella Beyrich, 1862
Type species: Avicula gryphaeata Münster, 1836; Austria, Triassic.
Remarks.—Cassianella is very diverse in the Cassian Formation. Bittner (1895) listed 10 species from the Cassian beds. The genus ranges from the Middle to the Upper Triassic.
Cassianella beyrichi Bittner, 1895
Fig. 27B, C.
pars 1838 Avicula gryphaeata Münst.; Goldfuss 1838: 127, pl. 106: 10e (non fig. 10 a–d).
pars 1841 Avicula gryphaeata; Münster 1841: 75–76, pl. 7: 7a–c.
pars 1865 Cassianella gryphaeata Münst. sp.; Laube 1865: 46–47, pl. 17: 1a–h.
1895 Cassianella Beyrichii nov. spec.; Bittner 1895: 54–55, pl. 6: 16–21.
1977 Cassianella beyrichii; Fürsich and Wendt 1977: 288, fig. 13.6.
1981 Cassianella beyrichi (Bittner); Zardini 1981: pl. 13: 3, 4.
Material.—Seven specimens, 1 from bulk samples, 6 from surface samples; 3 from Lago Antorno (PZO 12861, 12862, 2 figured specimens; PZO 12860, 1 specimen); 4 from Misurina Landslide (PZO 12751, 4 specimens); Italy, Carnian, Triassic.
Remarks.—Only the characteristic left valves are preserved. They were identified according to Zardini (1981). This Cassianella species, together with C. ampezzana Bittner, 1895, belongs to the trophic nucleus of Fürsich and Wendt’s (1977) Ampullina association, which is very similar in taxonomic composition to the Misurina Landslide and Lago Antorno assemblages studied herein.
Stratigraphic and geographic range.—Carnian, Upper Triassic; northern Italy (Cassian Formation), Sicily, Apennines; Austria (northern Alps). Upper Triassic; China, Yunnan (Diener 1923; Kutassy 1931; Paleobiology Database, https://paleobiodb.org).
Cassianella sp. 1
Material.—One left valve (PZO 12752) from Misurina Landslide surface sample, Italy, Carnian, Triassic.
Remarks.—The specimen is diagenetically compressed. The smooth surface distinguishes this species from both other Cassianella species found at Lago Antorno and Misurina Landslide. A species comparable to Cassianella sp. 1 was not found in the literature. Therefore, using open nomenclature was the best solution in the case of this single specimen.
Cassianella sp. 2
Material.—Two left valves (PZO 12863) from Lago Antorno surface samples, Italy, Carnian, Triassic.
Description.—This species is ornamented with distinct, densely packed co-marginal ribs, between which radially arranged densely spaced threads are visible.

Fig. 27. Cassianellid bivalves from Lago Antorno, northern Italy, Cassian Formation, Carnian, Upper Triassic. A. Cassianella spp., PZO 12886, in lateral view (A1); A2, lateral view showing prodissoconch; A3, internal view with partly preserved hinge. B. Cassianella beyrichi Bittner, 1895, PZO 12861, in lateral view. C. Cassianella beyrichi Bittner, 1895, PZO 12862, in internal (C1) and lateral (C2) views.
Order Pectinida Gray, 1854
Superfamily Heteropectinoidea Beurlen, 1954
Family Antijaniridae Hautmann in Carter et al., 2011
Genus Antijanira Bittner, 1901
Type species: Pecten hungaricus Bittner, 1901; Hungary, Triassic.
Antijanira sp.
Material.—One specimen (PZO 12750) from Misurina Landslide bulk sample, Italy, Carnian, Triassic.
Remarks.—Only the anterior auricle is preserved. The specimen is assigned to the genus Antijanira Bittner, 1901, because it resembles the shells of Antijanira auristriata (Münster, 1841) illustrated in Zardini (1981). Unfortunately, the auricle region was not depicted distinctly enough in Bittner (1895) to differentiate between species shown in his monograph.
Family Hunanopectinidae Yin, 1985
Genus Leptochondria Bittner, 1891
Type species: Pecten aeolicus Bittner, 1891; Turkey, Triassic.
Leptochondria sp.
Material.—Two specimens, both from bulk samples, 1 from Lago Antorno (PZO 12753); 1 from Misurina Landslide (PZO 12864); Italy, Carnian, Triassic.
Remarks.—Only fragments were found. They showed the characteristic ornamentation of Leptochondria monilifera (Braun in Münster, 1841) depicted in Zardini (1981), but the fragments did not show enough shell features to justify an unambiguous assignment to this species.
Order Lucinida Gray, 1854
Superfamily Lucinoidea Fleming, 1828
Family Lucinidae Fleming, 1828
Genus Schafhaeutlia Cossmann, 1897
Type species: Gonodon ovatum Schafhäutl, 1863; Austria, Triassic.
Schafhaeutlia cf. astartiformis (Münster, 1841)
Material.—One specimen (PZO 12754) from Misurina Landslide surface sample, Italy, Carnian, Triassic.
Remarks.—The single specimen is articulated and partly fragmented. It is the same species as depicted in Hausmann and Nützel (2015). However, the illustrated specimen in Hausmann and Nützel (2015) is heavily encrusted whereas the individual studied herein shows no incrustations.
Order Cardiida Férussac, 1822
Superfamily Cardioidea Lamarck, 1809
Family Cardiidae Lamarck, 1809
Genus Septocardia Hall and Whitfield, 1877
Type species: Septocardia typica Hall and Whitfield, 1877; USA, Jurassic.
Septocardia pichleri (Bittner, 1895)
Fig. 28.
1895 Cardita Pichleri nov. spec.; Bittner 1895: 38–39, pl. 24: 8–10.
1895 Cardita cf. Pichleri nov. spec.; Bittner 1895: 38–39, pl. 4: 17.
1981 Palaeocardita pichleri (Bittner); Zardini 1981: pl. 31: 3–11.
2001 Septocardia pichleri; Schneider and Carter 2001: fig. 3, 4.
Material.—Seven specimens, 2 from bulk samples, 5 from surface samples, 2 from Lago Antorno (PZO 12866, figured specimen, PZO 12865, 1 specimen); 5 from Misurina Landslide (PZO 12755, 5 specimens); Italy, Carnian, Triassic.
Description.—Septocardia pichleri (Bittner, 1895) is subovate-shaped with a subtruncate posterior margin; length and width of the illustrated specimen (Fig. 28) are 3.8 mm and 3.5 mm; shell highly inflated; externally ornamented primarily with strongly developed radial costae; fine concentric lirae are present, which are most obvious in the interspaces between the radial costae; tubercles and cross bars are occasionally present on the radial costae with stronger developed cross bars at the anterior ventral side; disarticulated and articulated specimens occur.

Fig. 28. Cardiid bivalve Septocardia pichleri (Bittner, 1895) from Lago Antorno, northern Italy, Cassian Formation, Carnian, Upper Triassic. PZO 12866, in ventral (A1), dorsal (A2), and lateral (A3, A4) views.
Remarks.—The present material agrees well with the type specimens illustrated by Bittner (1895) from the localities Issberg-Gehänge and Bergangerl near Hall in Northern Tyrol, Austria (northern Alps). Bittner (1895: pl. 4: 17) also illustrated an articulated specimen from the Cassian Formation of the Seelandalpe. Bittner stated that he could not study the hinge of the material from the southern Alps but that it shows no external morphological differences to the type material from the northern Alps. The present specimens are relatively small. The specimen from Alpe di Specie illustrated by Schneider and Carter (2001) closely resembles the present specimens and is of approximately the same size.
Stratigraphic and geographic range.—Carnian, Upper Triassic; northern Italy (Cassian Formation, northern Alps); Hungary (Bakony); Russia (Volzeia Beds Formation) (Diener 1923; Paleobiology Database, https://paleobiodb.org).
Bivalvia indet. 1
Material.—Nineteen specimens, all from bulk samples; 11 specimens (PZO 12867) from Lago Antorno; 8 specimens (PZO 12756) from Misurina Landslide; Italy, Carnian, Triassic.
Description.—The thin-shelled specimens are fragmented and in most cases smaller than two millimetres. Inflation is minimal and a weak concentric ornamentation is visible in some specimens.
Remarks.—Most specimens are disarticulated. The fossils are not determinable because of their fragmentary preservation.
Bivalvia indet. 2
Material.—One valve (PZO 12757) from Misurina Landslide bulk sample, Italy, Carnian, Triassic.
Remarks.—The single specimen consists of only one valve and is relatively thin-shelled. The fossil could not be determined properly, but an affiliation to the family Entoliidae is possible (Michael Hautmann, personal communication 2017).
Class Scaphopoda Bronn, 1862
Family Dentaliidae Children, 1834
Genus Dentalium Linnaeus, 1758
Type species: Dentalium elephantinum Linnaeus, 1758; Indo-Pacific, Recent.
Dentalium klipsteini Kittl, 1891
Fig. 29.
1891 Dentalium Klipsteini Kittl; Kittl 1891: 172, pl. 1: 3.
1978 Dentalim klipsteini (Kittl) [sic!]; Zardini 1978: 56, pl. 39: 14–17, 20, pl. 40: 7.
1978 Dentalium klipstein (Kittl) n. f. tenuistriatum [sic!]; Zardini 1978: 56, pl. 39: 19, 22, 23, pl. 40: 13.
Material.—Fourteen specimens, 7 from bulk samples, 7 from surface samples; 11 from Lago Antorno (PZO 12869–12871, 3 figured specimens; PZO 12868, 8 specimens); 3 specimens (PZO 12758) from Misurina Landslide; Italy, Carnian, Triassic.
Remarks.—Only broken specimens were found. Characteristic of this species are the more or less oblique annulations in combination with variably strongly pronounced longitudinal ribs.
Stratigraphic and geographic range.—Carnian, Upper Triassic; northern Italy (Cassian Formation) (Paleobiology Database, https://paleobiodb.org).

Fig. 29. Dentaliid scaphopods Dentalium klipsteini Kittl, 1891, from Lago Antorno, Cassian Formation, northern Italy, Late Triassic. PZO 12869 (A), PZO 12870 (B), and PZO 12871 (C), in lateral view.
Genus Plagioglypta Pilsbry in Pilsbry and Sharp, 1898
Type species: Dentalium undulatum Münster, 1844; Cassian Formation, northern Italy, Triassic.
Plagioglypta undulata (Münster, 1841)
1841 Dentalium undulatum; Münster 1841: 91, pl. 9: 6.
1844 Dentalium undulatum Münster; Goldfuss 1844: 3, pl. 166: 8.
1891 Dentalium undulatum Münster; Kittl 1891: pl. 1: 1.
1977 Dentalium undulatum Münster; Fürsich and Wendt 1977: 282, 284, 287, 292, 301, fig. 12.
1991 Pseudobactrites subundatus (Münster); Bizzarini and Gnoli 1991: 114, pl. 1: 5a–b.
2012 Plagioglypta undulata (Münster, 1841); Urlichs 2012: 8, figs. 3.11, 3.12, 8.
2014 Plagioglypta undulata (zu Münster 1841); Nützel and Kaim 2014: 424–425, fig. 11e, f.
2015 Plagioglypta undulata; Hausmann and Nützel 2015: 246, fig. 10K.
Material.—Two specimens (PZO 12759), 1 from bulk and 1 from surface samples; from Misurina Landslide, Italy, Carnian, Triassic.
Remarks.—This taxon was the second most abundant species in Fürsich and Wendt’s (1977) Palaeonucula strigilata/Dentalium undulatum association, which was found at Costalaresc. It is also a major component of the basinal soft-bottom fauna described from the Settsass Scharte (Nützel and Kaim 2014). In the allochthonous assemblage from Stuores Wiesen, P. undulata was only moderately represented (Hausmann and Nützel 2015).
Stratigraphic and geographic range.—Carnian, Upper Triassic; northern Italy (Cassian Formation); Germany (Bavarian Alps). Upper Triassic, Indonesia. Upper Triassic, Ladinian, Carnian, Peru. Jurassic, Bathonian, Poland (Paleobiology Database, https://paleobiodb.org).
Cephalopoda
A single fragment of a trachyceratid ammonite (PZO 12760) and four fragments of unidentified ammonites (PZO 12761) are present in the Misurina Landslide bulk sample, Italy, Carnian, Triassic.
Brachiopoda
A single, presumably juvenile (very small specimen, only 1 mm in size) unidentified brachiopod (PZO 12872) was found in the Lago Antorno bulk sample, Italy, Carnian, Triassic.
Arthropoda
Crustacean arthropods are represented in our samples by two ostracod specimens of two species. One of them (PZO 12762) was found in the bulk collection from Misurina Landslide, the other (PZO 12873) in the bulk sample from Lago Antorno, Italy, Carnian, Triassic. Both species were not determined. Ostracods are much more abundant and diverse quantitatively in the finer fraction (<0.5 mm) which was not analysed.
Echinodermata
Crinoidea
One poorly preserved columnal (PZO 12765) of isocrinid affinities (Fig. 30A) from Misurina Landslide bulk sample, Italy, Carnian, Triassic.
Asteroidea and Ophiuroidea
“Asterozoan” echinoderms from bulk samples collected at Misurina Landslide, Italy, Carnian, Triassic are mostly represented by a few ophiuroid species (around 20 ossicles and partly articulated arm fragments, including PZO 12878–12881), based on lateral arm plates, vertebrae, and spines whose detailed taxonomic affinities have yet to be determined (Fig. 30B–D, H).
Ophiocistioidea
Class Ophiocistioidea Sollas, 1899
Order unnamed
Family Linguaserridae Reich and Haude, 2004
Genus Linguaserra Langer, 1991
Type species: Linguaserra ligula Langer, 1991; Germany, Devonian.
Linguaserra triassica Reich in Reich et al., 2018
Fig. 30F.
Material.—One goniodont (PZO 11827) from Misurina Landslide bulk sample, Italy, Carnian, Triassic.
Description.—See Reich et al. (2018: 805–806).
Remarks.—This small linguaserrid goniodont (Fig. 30F) was described recently by Reich et al. (2018) as a new species, representing the stratigraphically youngest record of ophiocistioid teeth worldwide. Our Misurina material is important because it provides the only evidence of this “Palaeozoic” echinoderm group known from Mesozoic strata. Ophiocistioids, which are closely related to holothurians and echinoids (Reich 2010a; Smith and Reich 2013; Rahman et al. 2019), are exclusively benthic and were presumably active predators using their lantern and teeth to capture small prey (Haude 2004; Smith 2004a).
Stratigraphic and geographic range.—Carnian, Upper Triassic; northern Italy (southern Alps, Cassian Formation).
Class Echinoidea Leske, 1778
Remarks.—Sea urchin plates, spines and other ossicles are most common in our material. As the Cassian Formation contains the largest and most important Triassic echinoid fauna worldwide (cf. Kroh 2011), it is not suprising that even very rare or unexpected forms, hitherto unknown as articulated findings, can be found.
Subclass Perischoechinoidea McCoy, 1849
Order Palaechinoidea Smith, 1984
Suborder Palaeodiscoida Smith, 1984
Family Proterocidaridae Smith, 1984
Genus Pronechinus Kier, 1965
Type species: Pronechinus anatoliensis Kier, 1965; Turkey, Permian.
Pronechinus? sp.
Fig. 30G, J, L, M.
?2014 Unknown echinoderm plate; Nützel and Kaim 2014: fig. 11p, q.
?2017 Dissociated ambulacral test plate of a proterocidaroid, with peripodial rim indicated; Thuy et al. 2017a: fig. 1B.
Material.—Total more than 60 ambulacral and interambulacral plates (including PZO 12777–12780) from Misurina Landslide bulk samples, Italy, Carnian, Triassic.
Description.—This species is known only from disarticulated plates. One type of ambulacral plates oblongate polygonal in shape, with a marginal pore pair (without distinct peripodial rim) and an imperforate primary tubercle accompanied by a minute secondary tubercle (Fig. 30G). Another, and more common second type of ambulacral plates almost subcircular to circular in shape, with a central pore pair surrounded by a slightly distinct peripodial rim, and also bearing adjacent smaller secondary tubercles (Fig. 30J, L, M). Interambulacral plates rectangular to polygonal in shape, with up to 10 minute secondary tubercles.
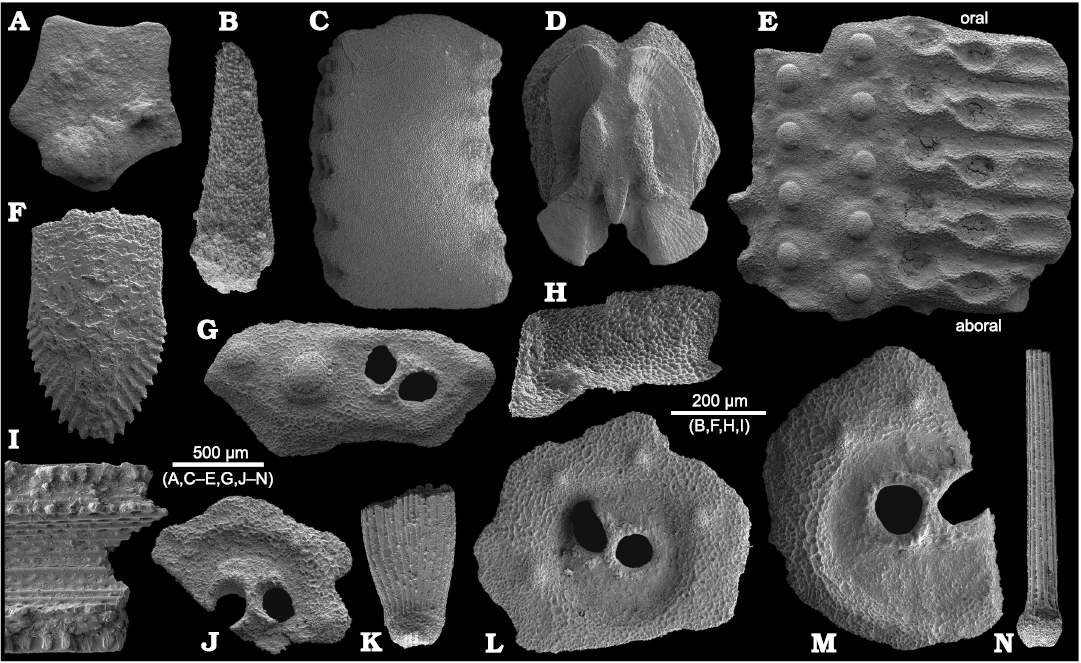
Fig. 30. Echinoderm plates, ossicles, and elements from Misurina Landslide, northern Italy, Cassian Formation; Carnian, Upper Triassic. A. PZO 12765, isocrinid? (sediment encrusted) columnal. B. PZO 12878, asteroid?/asterozoan spine. C, D, H. Ophiuroids. C. PZO 12879, lateral arm plate, external view, showing the articulation for the attachment of spines. D. PZO 12880, arm vertebra, distal view, showing the muscle attachment area. E. PZO 12885, part of ambulacrum of unknown cidaroid? species. H. PZO 12881, distal lateral arm plate, in external view. F. PZO 11827, ophiocistioid goniodont Linguaserra triassica Reich in Reich et al., 2018, abaxial face, in outer view. G. Pronechinus? sp., PZO 12777, ambulacral plate. I. PZO 12884, broken hollow cidaroid? spine fragment, showing the microstructure at the inner side of the cylinder. J, L, M. Ambulacral plates of Pronechinus? sp. with slightly developed peripodial rim surrounding pore pairs (J, PZO 12780; L, PZO 12778; M, PZO 12779). K, N. Cidaroid? spines (partially broken) of unknown species (K, PZO 12882; N, PZO 12883).
Remarks.—This putative proterocidarid material is remarkable because it provides evidence for persistence of this “Palaeozoic” echinoid stem group for at least 16 Ma after the Permian/Triassic boundary. Misurina Landslide is the same locality from which a representative of another “Palaeozoic-type” echinoderm group, the Ophiocistioidea, was recently described (Reich et al. 2018). These findings are in accordance with other Triassic stem group echinoids recently reported from France (Hagdorn 2018), China (Thuy et al. 2017a; Thompson et al. 2018), and potentially Italy (Thompson et al. 2019).
Subclass Cidaroidea Smith, 1984
Order Cidaroida Claus, 1880
Isolated spines and partly articulated test fragments (gen. et spp. indet.)
Fig. 30E, I, K, N.
Material.—More than 90 spines, ambulacral plates, and other ossicles (teeth, rotulae, etc.) (including PZO 12777–12780) from Misurina Landslide bulk samples and few other remains from Lago Antorno bulk samples, Italy, Carnian, Triassic.
Remarks.—In addition to the stem group echinoid taxon, we have found well preserved and partly articulated test fragments (Fig. 30E) and spines (a few types; e.g., Fig. 30I, K, N) of cidaroid echinoids (including “Cidaris” cf. decoratissima Wöhrmann, 1889, and “Cidaris” cf. flexuosa Münster, 1841) at Misurina Landslide, which represent the majority of sea urchin material from the Ladinian/Carnian Cassian Formation (see also Kier 1984; Vadet 1999a, b; Kroh 2011). However, as more than 40% of the worldwide reported Triassic echinoid “species” are spine-based taxa (Kroh 2011), disarticulated material remain often indeterminate to species or genus level. Spines (and other ossicles) can be distinct and recognizable, but more studies combining isolated spines and test material, like ambulacral plates, are needed to validate different single species.
Regular echinoids, like in our material, are vagile benthos, moving by means of their spines over the sea floor and using their tube-feet to climb and grip hard substrata (Smith 2004b). The proterocidarid species described and figured here is presumably a deposit feeder, using its enlarged oral tube-feet for detritus gathering (cf. Thompson and Bottjer 2019).
Holothuroidea
Sea cucumbers are represented by calcareous ring (CR) elements and body-wall ossicles of Apodida (Fig. 31A–C, E, F) and Synallactida (Fig. 31D, G, H) only.
Class Holothuroidea Blainville, 1834
Subclass Paractinopoda Ludwig, 1891
Order Apodida Brandt, 1835
“Stem group chiridotids”
Gen. et sp. indet. 1
Fig. 31A, B.
Material.—Four calcareous ring elements (PZO 12784, 12785) from Misurina Landslide bulk samples, Italy, Carnian, Triassic.
Description and remarks.—Radial (Fig. 31A) and interradial (Fig. 31B) calcareous ring elements of a medium-sized apodid sea cucumber, probably with ?15 tentacles. Radial and interradial nearly squarish in shape and with a deep posterior indent, radial with two unequal medium long anterior processes, the shorter one notched for the passage of the radial nerve (Fig. 31A1, A2, A4; here filled with sediment). The additional anterior process is longer and pointed like that of the interradial (Fig. 31B). On the external face of radial a tongue-shaped insertion area for the tentacle ampullae. The stereom of radial resembles that of the interradial. Interradial with a single well-developed pointed anterior process and associated frontal excavations.
Both types of elements are closely comparable with the calcareous ring elements found in modern chiridotids (and myriotrochids), but presenting apparently a stem group chiridotid, with respect to the stout morphology in combination with medium-sized anterior processes.
Definitely associated body-wall ossicles missing, but probably represented by Theelia cf. multiplex (Fig. 31E).
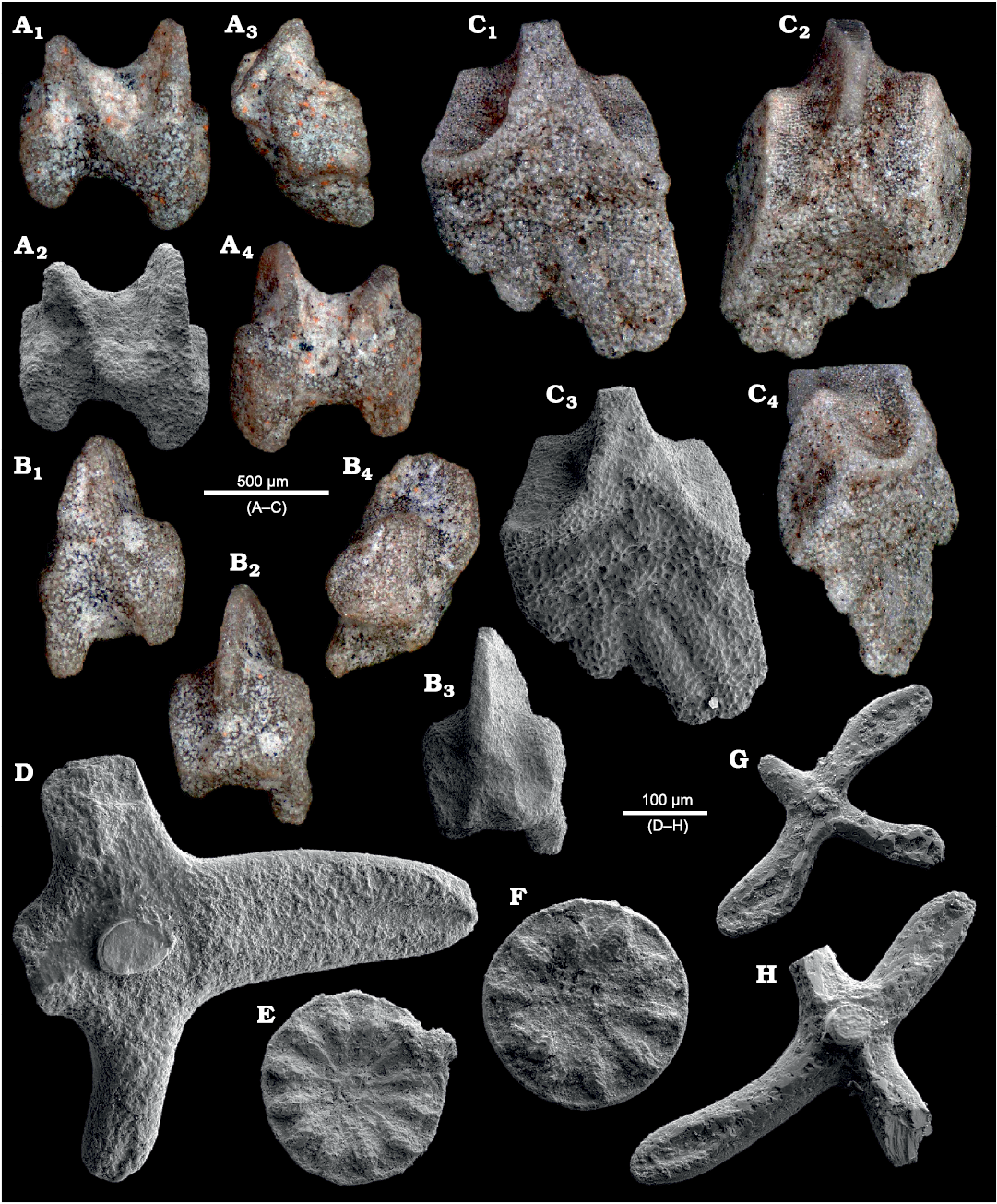
Fig. 31. Holothurian calcareous ring elements
and body-wall ossicles from Misurina Landslide, northern Italy,
Cassian Formation; Carnian, Upper Triassic. A.
Apodida gen. et sp. indet. 1, PZO 12784, radial calcareous ring
element in outer view (A1,
A2), lateral
oblique view from right (A3),
inner view (A4).
B. Apodida gen. et sp. indet.
1, PZO 12785, interradial calcareous ring element in outer view (B1), lateral oblique view
from left (B4),
inner view (B2,
B3). C. Apodida gen. et sp. indet. 2, PZO
12787, interradial calcareous ring element in outer view (C1, C3)
inner view (C2),
lateral oblique view from right (C4).
D. Tetravirga
sp., PZO 12792, cross, upper side, two arms and the central solid
spire broken, perforated arm areas filled with sediment. E. Theelia
cf. multiplex Speckmann,
1968, PZO 12794, wheel, lower side, interspoke areas filled with
sediment. F. Jumaraina
sp., PZO 12789, wheel, lower side (interspoke areas filled with
sediment). G. Tetravirga cf. perforata
Mostler, 1968, PZO 12790, cross, upper side, one arm and the central
solid spire broken, perforated arm areas filled with sediment. H. Tetravirga
cf. perforata Mostler,
1968, PZO 12791, cross, upper side, two arms and the central solid
spire broken, perforated arm areas filled with sediment. SEM (A2, B3,
C3, D–H) and
LM (A1, A3, A4,
B1, B2, B4,
C1, C2, C4)
images.
Incertae familiae
Gen. et sp. indet. 2
Fig. 31C.
Material.—Two calcareous ring elements (PZO 12787) from Misurina Landslide bulk sample, Italy, Carnian, Triassic.
Description.—Large and robust interradial (Fig. 31C) calcareous ring elements, with prominent anterior insertion areas for the tentacle ampullae, and padded-shaped medium long posterior processes (Fig. 31C1, C3). The single pointed anterior process is short in comparison to the whole length. Different stereom types visible.
Remarks.—Due to the general shape and morphology, this type of calcareous ring elements is comparable with apodid calcareous rings. However, due to the posterior processes, a taxonomic position within molpadid sea cucumbers (Actinopoda: Molpadida) is also possible.
“Stem group chiridotids”
or Chiridotidae Östergren, 1898
Genus Theelia Schlumberger, 1890, sensu Gilliland 1992
Type species: Chirodota undulata Schlumberger, 1888; France, Eocene.
Theelia cf. multiplex Speckmann, 1968
Fig. 31E.
Material.—One single wheel body-wall ossicle (PZO 12794) from Misurina Landslide bulk sample, Italy, Carnian, Triassic.
Description.—Rim slightly inclined to the plane of the wheel and subtle serrated; hub slightly convex at upper side, but ring-shaped at lower side; with 12 spokes. All interspoke areas filled with sediment, why an specific designation is not possible.
Genus Jumaraina Soodan, 1973
Type species: Jumaraina indica Soodan, 1973; India, Jurassic.
Jumaraina sp.
Fig. 31E.
Material.—One single wheel body wall ossicle (PZO 12789) from Misurina Landslide bulk sample, Italy, Carnian, Triassic.
Description.—Rim slightly inclined to the plane of the wheel, unserrated; hub convex at upper side, and indifferent flat at lower side; with 10 spokes. All interspoke areas filled with sediment, why a specific designation is not possible.
Subclass Actinopoda Ludwig, 1891
Order Synallactida Miller, Kerr, Paulay, Reich, Wilson, Carvajal, and Rouse, 2017
Family Synallactidae Ludwig, 1894
or “Stem group synallactids”
Genus Tetravirga Frizzell and Exline, 1956
Type species: Tetravirga imperforata Frizzell and Exline, 1956; USA, Pennsylvanian.
Tetravirga cf. perforata Mostler, 1968
Fig. 31G, H.
2005 Tetravirga sp.; Bizzarini and Laghi 2005: pl. 2: 5.
Material.—Two body-wall ossicles (PZO 12790, 12791) from Misurina Landslide bulk sample, Italy, Carnian, Triassic.
Description and remarks.—Ossicles in form of a cross, arms placed at 70–110° angles; slender arms distally expanded and grooved. Grooved area is restricted to the distal third of each arm. These trenchlike impressions should be perforated, but are filled with sediment in our specimens. A solid spire (circular in shape) in the centre of the ossicle (outer/upper side) is broken (e.g., Fig. 31H). After Reich (2010b), this type of ossicle resembles synallactid taxa with respect to general shape and morphology, spire, and especially the distally grooved and perforated arms.
Tetravirga sp.
Fig. 31D.
non 2014 Tetravirga sp.; Nützel and Kaim 2014: fig. 11n, o. [= calcareous sponge spicules].
Material.—One body-wall ossicle (PZO 12792) from Misurina Landslide bulk sample, Italy, Carnian, Triassic.
Description.—Large ossicle in form of a cross, arms placed at 100° angles; wide arms distally decreasing and grooved. Grooved area is restricted to around 40% of each arm. These impressions should be perforated, but are filled with sediment in our specimen. A solid spire (oval in shape) in the centre of the ossicle (outer/upper side) is broken (e.g., Fig. 31D). This type of ossicle is closely related to T. cf. perforata and therefore resembles representatives of the Synallactidae.
Remarks.—Apodid sea cucumbers were mainly infaunal deposit feeders, whereas synallactids were epibenthic deposit feeders (e.g., rake-feeder or sweeper) and capable of active swimming. Representatives of both reported groups are therefore active and important bioturbators of the sediment floor.
Problematicum
Abundant unknown trace fossils resembling crustacean coprolites (including PZO 12822, 12876, 12877) were found in the Misurina Landslide bulk samples, Italy, Carnian, Triassic (Fig. 32). They are oval in tranverse section and have fine, irregular longitudinal ridges on their lateral sides. Bizzarini and Laghi (2005: pl. 2: 9) illustrated similar specimens and identified them as Bactryllium sp. However, these specimens are not as laterally compressed and have straight, pronounced longitudinal furrows. The identification of the present problematicum and its producer needs further study.
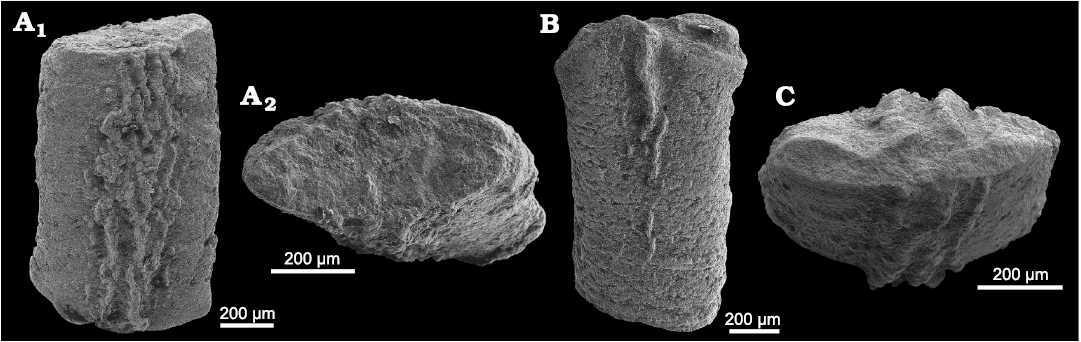
Fig. 32. Trace fossils (taxonomic assignment unknown) from Misurina Landslide, northern Italy, Cassian Formation, Carnian, Upper Triassic. A. PZO 12822. B. PZO 12876. C. PZO 12877.
Discussion
Faunal composition.—Even though the overall faunal content is similar at Lago Antorno and Misurina Landslide, there are differences in species composition and abundances (Table 1). The majority of the more abundant species is shared between the Lago Antorno and the Misurina Landslide localities (e.g., the gastropods Prostylifer paludinaris, Coelostylina conica, Ampezzopleura hybridopsis, and Jurilda elongata). However, Jurilda elongata is the most abundant species at Misurina Landslide but has never been reported as a dominant species from elsewhere (Fürsich and Wendt 1977; Nützel and Kaim 2014; Hausmann and Nützel 2015; Roden et al. 2020b) and is not abundant at Lago Antorno. The rare species are commonly not shared between the studied sites, but this is probably due to random effects, the likelihood that rare species are found at various sites is lower than finding common shared species.
Although there is some similarity to the Ampullina assemblage sensu Fürsich and Wendt (1977), the assemblages from Misurina Landslide and Lago Antorno differ considerably in taxonomic composition from all other Cassian invertebrate assemblages described quantitatively so far (Fürsich and Wendt 1977; Nützel and Kaim 2014; Hausmann and Nützel 2015). The most abundant species from Settsass Scharte (Goniospira armata, Schartia carinata, Helenostylina convexa, Plagioglypta undulata) are either absent or rare in the samples from Lago Antorno and Misurina Landslide. In contrast, all abundant species from the samples studied herein are absent at Settsass Scharte (except Coelostylina conica, which is, however, not very abundant there) (Nützel and Kaim 2014).
The Rhaphistomella radians/Palaeonucula strigilata association (largely composed of the name-giving species as well as the bivalve Prosoleptus lineatus), reported by Fürsich and Wendt (1977) from Stuores Wiesen and Pralongia, likewise represents an autochthonous basin association. The bivalve P. strigilata is rarer in the Lago Antorno and Misurina Landslide samples, and the gastropod R. radians is absent. Fürsich and Wendt (1977) based their analysis on surface collections and therefore did not consider the microgastropod fauna, which may partly explain the differences in faunal composition, but not the absence of the index species.
Alpha and beta diversity.—Both fossil assemblages from Lago Antorno and Misurina Landslide are characterised by a moderate to low diversity when compared to other Cassian faunas studied so far (Fürsich and Wendt 1977; Hausmann and Nützel 2015). A comparison of Misurina Landslide and Lago Antorno with the Cassian assemblage from Stuores Wiesen shows that, in addition to a difference in taxonomic composition, diversity is also profoundly different. The Stuores Wiesen diversity is very high (Shannon index of 4.01; Simpson index of 0.96), and is supported by rarefaction analyses (Fig. 33). The fossil assemblage found at this locality is thought to stem from a shallow water lagoonal palaeohabitat (Hausmann and Nützel 2015).
In contrast, Misurina Landslide and Lago Antorno have a similar moderate to low diversity as the basinal soft-bottom assemblage reported by Nützel and Kaim (2014) from the Settsass Scharte (Shannon index of 2.24; Simpson index of 0.85; Fig. 33) but also differ strongly in composition.
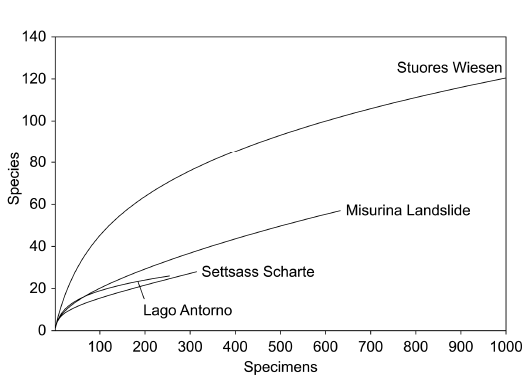
Fig. 33. Rarefaction curves of Cassian assemblages from Misurina Landslide, Lago Antorno (assemblages studied herein), Settsass Scharte (Nützel and Kaim 2014) and Stuores Wiesen (Hausmann and Nützel 2015). Only bulk samples are used.
A comparison of beta diversity based on dissimilarities across the quantitatively studied Cassian assemblages (Nützel and Kaim 2014; Hausmann and Nützel 2015) shows that all assemblage pairs are highly dissimilar (values >0.9), except for Misurina Landslide and Lago Antorno (Table 5); a value of 0.52 indicates that both assemblages are quite similar to each other, confirming our previous results and those of Roden et al. (2020b). Nevertheless, beta diversity is very high among the studied Cassian assemblages (Misurina Landslide, Lago Antorno, Settsass Scharte, Nützel and Kaim 2014; Stuores Wiesen, Hausmann and Nützel 2015), with a mean dissimilarity value of 0.88. High beta diversity in the Cassian Formation is probably due to high environmental heterogeneity, including a broad range of water depths. Water depth itself, although often found to be a driver of beta diversity (e.g., Ellingsen and Gray 2002; Harborne et al. 2006), has not been shown to be the main driver for the Cassian Formation (Roden et al. 2020b). However, data in Roden et al. (2020b) are too limited to allow definite conclusions on the effect of water depth on beta diversity.
Table 5. Dissimilarity (using mean proportional dissimilarity) between Cassian assemblages from Misurina Landslide, Lago Antorno, Stuores Wiesen (Hausmann and Nützel 2015) and Settsass Scharte (Nützel and Kaim 2014). Only bulk samples were used.
| |
Stuores Wiesen |
Misurina Landslide |
Lago Antorno |
Settsass Scharte |
|
Stuores Wiesen |
|
0.96 |
0.96 |
0.93 |
|
Misurina Landslide |
0.96 |
|
0.52 |
0.95 |
|
Lago Antorno |
0.96 |
0.52 |
|
0.96 |
|
Settsass Scharte |
0.93 |
0.95 |
0.96 |
|
Palaeoenvironment.—Several factors indicate that the assemblages stem from basinal soft-bottom settings. Nuculids are characteristic representatives of the bivalve assemblages and are considered to be soft-bottom dwellers, living infaunally as deposit-feeders (e.g., Fürsich and Wendt 1977; Aberhan 1994; Fürsich et al. 2004). They almost always occur with conjoined valves in the investigated sections. Ooids and oncoids, which are typical components of transported shallow-water assemblages from the Cassian Formation (Hausmann and Nützel 2015), are lacking. In addition, the marly to clayey sediments yielding the fauna point to soft-bottom conditions. The large number of double-valved bivalve shells indicates a predominantely autochthonous origin of both fossil assemblages. However, it is also possible that nuculids occur with co-joined valves in assemblages transported to a small degree because their taxodont hinge type allows a relatively strong teeth interlocking (e.g., Schenck 1934; Oliver 1992). Nevertheless, we assume that the great majority of the assemblage was not transported and only a few allochthonous elements from other palaeoenvironments are present. For example, a single fragmentary calcareous sponge was found in the surface sample from Misurina Landslide and a small fragment of a potential hydrozoan was found in the bulk sample from Lago Antorno; both taxa may have grown in reefal environments. The presence of echinoderms, nuculid bivalves and clearly marine gastropods that had planktotrophic larval development (Nützel 2014) indicates fully marine conditions.
Fürsich and Wendt (1977) described a variety of faunal assemblages from the Cassian Formation stemming from different palaeoenvironments, including, for example, patch reef faunas of partly allochthonous origin and several autochthonous basin associations. The fossil assemblages studied herein resemble the Ampullina association of Fürsich and Wendt (1977) from the Cortina Basin, which includes the region of Misurina. This basinal, autochthonous, soft-bottom association is characterised mainly by the smooth-shelled, bulbous caenogastropod species Helenostylina convexa and Prostylifer paludinaris, which were identified as Ampullina sanctaecrucis and A. paludinaris in Fürsich and Wendt (1977) and Bizzarini and Laghi (2005). It is unclear which species the authors were referring to with Ampullina sanctaecrucis (= Ptychostoma sanctaecrucis) because no photographs were provided. Ptychostoma sanctaecrucis is a relatively large gastropod that resembles P. paludinaris. Ptychostoma sanctaecrucis is abundant at its type locality which is also the type locality of the Heiligkreuz Formation and is interpreted as a brackish-water habitat (Koken 1913; Bizzarini and Rottonara 1997); this species is not present in the assemblages studied herein. Prostylifer paludinaris is the most abundant species in our bulk samples and is by far the most abundant species in the surface samples, corresponding to the observations made by Fürsich and Wendt (1977) that this species is highly abundant in the Ampullina association found in marls in the vicinity of Cortina d’Ampezzo (Rudavoi, Costalaresc, Misurina). Smooth, bulbous gastropods such as Helenostylina convexa, “Ampullina” sanctaecrucis and P. paludinaris look very similar to each other and can only be identified clearly when the protoconch is present and the base is well exposed. Since this is most often not the case, species identification is difficult and we assume that the two most abundant gastropods of Fürsich and Wendt’s (1977) Ampullina association (“A. paludinaris” and “A. sanctaecrucis”) represent Prostylifer paludinaris, and more rarely Helenostylina convexa. Fürsich and Wendt’s (1977) Ampullina association and our studied assemblages share the presence of palaeonuculid, cassianellid and cardiid bivalves. These bivalves are more abundant in Fürsich and Wendt’s (1977) dataset. This is probably due to the fact that these authors based their study on surface samples and not on washed bulk samples. Since bivalve species are commonly larger than gastropod species, their proportion in surface samples is expected to be higher. Many gastropods in our assemblages are juveniles, but not isolated larval shells (although these are also present). Therefore, the size difference between gastropods and bivalves, at least in our samples, is not due to a significantly larger portion of larvae in gastropods compared to bivalves.
The water depths of the palaeosettings, in which the faunal assemblages from Misurina Landslide and Lago Antorno lived, cannot be determined with certainty. According to Fürsich and Wendt (1977), the autochthonous soft-bottom faunas they described lived in shallow basins, whereas almost no autochthonous benthic assemblages are found in deep basins. Wendt and Fürsich (1980) suggested that the benthic soft-bottom assemblages lived in the upper subtidal or at least within the photic zone. However, Urlichs (2012) concluded that they stem from greater water depths, reaching 100 m and more. Roden et al. (2020b) suggested an intermediate water depth for Misurina Landslide and a more shallow depth for Lago Antorno based on various criteria. In washed residues from lower parts of the Misurina Landslide section not analysed here, stems of dasycladacean algae were found, indicating deposition in the photic zone (AN unpublished observation). However, in the residues studied herein, algae are absent. This could suggest that the Misurina Landslide outcrop exposes a shallowing upward sequence.
Our calculated diversity indices show that alpha diversity is approximately the same across bulk samples from both studied Cassian localities (Table 3). Similar evenness and ecological complexity as well as a relatively similar fossil content suggest an origin from similar palaeoenvironments, supported by their spatial proximity.
Body size.—The fossil assemblages from Lago Antorno and Misurina Landslide are strongly dominated by molluscs, with gastropods being the most diverse and abundant group. Most of the fossils are very small. The size of most fossils does not exceed a few millimetres, which is a common feature of Cassian assemblages, regardless of the palaeoenvironment from which they originally derive (e.g., Fürsich and Wendt 1977; Nützel and Kaim 2014; Hausmann and Nützel 2015). The majority of Recent gastropods from marine tropical settings are also characterised by small to very small sizes (e.g., Bouchet et al. 2002; Bouchet 2009; Albano et al. 2011). Therefore the small sizes of the gastropod-dominated fossil assemblages from Misurina Landslide, Lago Antorno, and other Cassian locations (e.g., Fürsich and Wendt 1977; Nützel and Kaim 2014; Hausmann and Nützel 2015) are less remarkable than previously thought. Stunting due to unfavourable conditions has been proposed to explain the allegedly exceptionally small size of the Cassian fossils (e.g., Richthofen 1860; Leonardi and Fiscon 1959). However, as pointed out by Fuchs (1871) and Nützel et al. (2010b), the high overall diversity of the Cassian Formation argues against pronounced environmental stress, because such stress produces communities of low diversity and high dominance. Stunting has been reported for some species at Stuores Wiesen and surrounding areas near San Cassiano (Urlichs 2012) but cannot be proposed for the Cassian fauna as a whole. On the other hand, the question arises why only a very small fraction or a complete lack of larger fossils characterises samples from the Cassian Formation. In allochthonous fossil assemblages, size sorting during transport could, at least in part, be responsible for the exclusively small sizes (Hausmann and Nützel 2015). However, since the samples from Misurina Landslide and Lago Antorno are of predominantely autochthonous origin, transport-induced size sorting cannot be a reason for the lack of fossils larger than 10 mm. An explanation concerning surface samples could be the selective removal of large specimens by private collectors (e.g., Fürsich and Wendt 1977; Urlichs 2004, 2012). However, historical collections from the Cassian beds made during the last 180 years consist of a great majority of specimens smaller than 15 mm. Regarding bulk samples, a comparison with modern (sub-) tropical marine sediment samples may help to explain the scarcity of large fossils, since modern benthic sediments typically consist of only a small number of large specimens (e.g., Bouchet et al. 2002; Bouchet 2009; Hausmann et al. 2018). Some larger Cassian fossils have been found, but in most cases they are fragmented or disarticulated (e.g., Hausmann and Nützel 2015; AN unpublished data). Molluscs, for example, can reach sizes of 40 mm and even more; however, as in Recent (sub-) tropical marine sediment samples, members of these size classes are scarce (e.g., Bouchet et al. 2002; Bouchet 2009; AN unpublished data).
As mentioned before, many of the gastropods from the studied samples are juveniles or fragments, as is typical for gastropods from bulk samples. Most gastropods are clearly post-larval and only a small number of larvae has been found, so that these assemblages do not represent larval fall communities as has been reported, for instance, from the Carboniferous of Arkansas by Nützel and Mapes (2001). Some of the species are fully grown but still small, for instance Bandellina compacta Nützel and Hausmann sp. nov., Alexogyra marshalli Bandel, 1996, Ampezzogyra angulata Nützel and Hausmann sp. nov. and species of the genus Sinuarbullina, which never grew to large sizes. A decision to exclude juvenile specimens from analyses would be most arbitrary, because even juveniles are ecologically active actors and it is unclear for most species at which point they have reached their maximum adult size. Also the degree of fragmentation is unknown. High spired species such as Jurilda elongata consist largely of fragments. The high number of juvenile specimens may also point to a high mortality due to environmental conditions. This is especially true for the neritimorphs Neritaria mandelslohi and Dentineritaria neritina, which are abundant but only present as isolated larval shells or juvenile specimens with a small teleoconch portion. Obviously, the larvae of these species failed to settle in a suitable habitat.
Additionally, unfavourable environmental conditions may have also led to a smaller body size in adult specimens. It is possible that fluctuating oxygen concentrations and the soft substrate limited body size in this case, as has also been proposed for other soft-bottom communities (e.g., from the Early Jurassic; Nützel and Kiessling 1997; Nützel and Gründel 2015). Framboidal pyrite was observed in the problematic trace fossils from Misurina Landslide (Fig. 32), and framboids were occasionally detected at Lago Antorno (Fig. 34). Many studies demonstrate that framboidal pyrite is characteristic of oxygen-deficient conditions in benthic habitats and that the size of framboidal structures indicate the severity of oxygen deficiency (e.g., Wilkin et al. 1996, 1997; Wignall and Newton 1998; Bond and Wignall 2010; Shen et al. 2007; Wignall et al. 2005). Lower oxygen content in the bottom waters of the palaeohabitats from the two sampling sites, Lago Antorno and Misurina Landslide, could argue for the relatively low biodiversity, but stunting at these localities has not been investigated so far.

Fig. 34. Pyrite framboid from Lago Antorno, northern Italy, Cassian Formation, Carnian, Upper Triassic. PZO 12887.
Echinoderms.—Several invertebrate groups, including echinoderms, were apparently severely decimated during the Permian–Triassic mass extinction interval (e.g. Sepkoski 1981; Erwin 1989; Benton and Twitchett 2003; Twitchett and Oji 2005; Pietsch et al. 2019). However, existing paradigms that most of the echinoderm lineages are inferred to have gone extinct at the end of the Palaeozoic shifted recently due to the presence of “hangovers” or “holdovers” in Triassic strata (e.g., Thuy et al. 2017a–c; Thuy 2017; Blake 2017; Hunter and McNamara 2017; Salamon and Gorzelak 2017; Hagdorn 2018; Reich et al. 2018; Thompson et al. 2018, 2019). Due to the rapid post-mortem disarticulation of echinoderms, resulting in a relatively sparse articulated fossil record, the use of the disarticulated fossil record seems rather more favourable (e.g., Kutscher and Reich 2004; Reich and Smith 2009; Thompson and Denayer 2017; Thompson et al. 2019).
Liberation lagerstätten (Roden et al. 2020a), like the Cassian Formation, yield highly diverse echinoderm assemblages (e.g., Zardini 1973, 1988) with a large proportion of disarticulated material from Crinoidea, Asteroidea, Ophiuroidea, Echinoidea, and Holothuroidea (e.g., Gümbel 1869; Deflandre-Rigaud 1963; Kristan-Tollmann 1970; Janofske 1992; Bizzarini 1993, 1996; Broglio Loriga et al. 1999; Bizzarini and Laghi 2005; Gale 2011; Hagdorn 2011; Kroh 2011; Nützel and Kaim 2014; Hausmann and Nützel 2015).
Until now, Cassian stem group echinoids (i.e., putative proterocidarids) have been found solely in samples investigated herein and from the Settsass Scharte locality (Nützel and Kaim 2014), all of which represent deeper water soft-bottom habitats. In the bulk collection from Misurina Landslide, the abundance of isolated proterocidarid plates was high (74 ossicles). Hagdorn (2018) stated that it is unclear whether stem group echinoids from the Triassic were inhabitants of shallow or deep water habitats (cf. also Godbold et al. 2017; Thompson et al. 2019). However, results from Cassian assemblages studied so far suggest that they preferred deeper water settings (Nützel and Kaim 2014; Hausmann and Nützel 2015). The 0.11–0.5 mm size fraction from Misurina Landslide was additionally analysed, but in contrast to the other size fraction only qualitatively. One skeletal element (goniodont) was found, which belongs to the ophiocistioid Linguaserra triassica. Ophiocistioids were previously only found in Palaeozoic strata and were therefore thought to have gone extinct at the end of the Palaeozoic. However, they in fact survived much longer, at least up to the Late Triassic (Reich et al. 2018). The fact that putative proterocidarids and the ophiocistioid L. triassica from the Cassian Formation were until now solely found from basinal palaeohabitats (Nützel and Kaim 2014; Reich et al. 2018) suggests that the Late Triassic basinal settings in this area may have acted as refugia for such hangover taxa, as also suggested by Thuy et al. (2017c).
Predation.—Drill holes are not uncommon in autochthonous soft-sediment assemblages from the Cassian Formation (Klompmaker et al. 2016). In addition to the samples studied herein, drilling was also reported from a surface sample from the Stuores Wiesen, which is representative of the basinal Raphistomella radians/Palaeonucula strigilata association sensu Fürsich and Wendt (1977) (Klompmaker et al. 2016). However, drilling predation was not found in the basinal soft-bottom assemblage described from the Settsass Scharte by Nützel and Kaim (2014). It is unknown which organisms produced the drill holes found in the shells from the Late Triassic Cassian Formation (Klompmaker et al. 2016). The shape of the drill holes resembles naticid borings. However, these predatory gastropods must be excluded as potential drill hole producers since they originated much later in the Cretaceous (Bandel 1999; Kase and Ishikawa 2003; Klompmaker et al. 2016; Kaim et al. 2017).
Preservation.—Low-grade lithification is a remarkable aspect of the Cassian Formation and points to the potential of these sediments to preserve fossil assemblages, which are taphonomically highly unbiased by lithification (Kowalewski et al. 2006; Sessa et al. 2009). Kowalewski et al. (2006) found that bulk samples from the Cassian Formation are more similar to unlithified late Cenozoic samples than lithified Jurassic samples in several respects, including diversity and higher taxonomic composition. Interestingly, gastropods are a major component of the late Cenozoic assemblages, whereas they play only a minor role in lithified Jurassic assemblages (Kowalewski et al. 2006). Kowalewski et al. (2006) and Sessa et al. (2009) suggest that differences in diversity and size distributions are at least partly due to diagenetic effects resulting from lithification and aragonite loss.
Diversification in the Triassic.—The new data from Misurina Landslide and Lago Antorno support Hautmann’s (2014) model of diversity partitioning. This model explains how diversification takes place after mass extinctions. It was found that the main recovery phase following the end-Permian mass extinction took place in the Middle Triassic (Hautmann 2007; Friesenbichler et al. 2019), which may have allowed early Late Triassic faunas from the Cassian Formation to further diversify with respect to alpha and beta diversity. Hautmann’s (2014) model recognises three phases during the full recovery process: (i) niche overlap phase, (ii) habitat contraction phase, and (iii) niche differentiation phase. During the first phase after a mass extinction, mainly alpha diversity rises since no or only low competition takes place. Therefore, the habitat width of individual species is not yet limited by competition across species. In the second phase, competition leads to a narrowing of the species’ habitat width and, consequently, to a stagnation of alpha diversity but to a rise in beta diversity. In phase three, alpha diversity may again be the main driver of diversity increase because of adaptive divergence (see Hofmann et al. 2013a, b, 2014 and Hautmann 2014 for further details). The high degree of variation in diversity and taxonomic composition between invertebrate assemblages in combination with a high diversity found in specific assemblages from the Cassian Formation (Fürsich and Wendt 1977; Nützel and Kaim 2014; Hausmann and Nützel 2015) indicates that faunal assemblages from the Late Triassic already reached the transition to the second phase of Hautmann’s (2014) model.
Conclusions
Basinal soft-bottom invertebrate assemblages from the Cassian Formation can vary in their taxonomic composition; however, previous data (Nützel and Kaim 2014) together with our dataset suggest that they have similarly moderate alpha diversity. The small size of Cassian invertebrate fossils is not as exceptional as previously thought but is very similar to Recent tropical marine benthic assemblages (e.g., Bouchet et al. 2002; Bouchet 2009; Albano et al. 2011). Soft-bottom basin habitats may have acted as refugia for echinoderm Lazarus (“hangover”) taxa after the end-Permian mass extinction in the Cassian Formation (Nützel and Kaim 2014; Reich et al. 2018), together with at least rarely present suitable habitats (both, in deeper but also shallow water) in other parts of the Tethys (Godbold et al. 2017; Thuy et al. 2017c; Thompson et al. 2019). This study confirms the need to further investigate Cassian assemblages to get a full picture of the complex Cassian palaeoecosystem.
Acknowledgements
We would like to thank Michael Hautmann (Universität Zürich, Switzerland) and Baran Karapunar (Ludwig‐Maximilians‐Universität München, Germany) for their help with species identifications. William Foster (University College Dublin, Ireland), Simon Schneider (CASP, Cambridge, UK) and Jeffrey R. Thompson (University College London, UK) are acknowledged for their helpful reviews. Special thanks go to Evelyn Kustatscher (Naturmuseum Südtirol, Bozen, Italy) and the Naturmuseum Südtirol for supporting this study. Markus Moser (Bayerische Staatssammlung für Paläontologie und Geologie, Munich, Germany) is acknowledged for his helpful comments on the manuscript. The research of IMH, VJR and AN was supported by the “Deutsche Forschungsgemeinschaft” (IMH and AN: project number NU 96/13-1; VJR: project number KI 806/14-1). MR acknowledges support from the „Bayerischer Pakt für Forschung und Innovation (BayPFI)“ of the „Bayerisches Staatsministerium für Wissenschaft und Kunst“ (StMWK).
References
Aberhan, M. 1994. Guild-structure and evolution of Mesozoic benthic shelf communities. Palaios 9: 516–545. Crossref
Adams, H. and Adams, A. 1858. The Genera of Recent Mollusca; Arranged According to Their Organisation. 660 pp. Van Voorst, London. Crossref
Albano, P.G., Sabelli, B., and Bouchet, P. 2011. The challenge of small and rare species in marine biodiversity surveys: microgastropod diversity in a complex tropical coastal environment. Biodiversity and Conservation 20: 3223–3237. Crossref
Alberti, F. von 1864. Überblick über die Trias. 353 pp. Cotta, Stuttgart.
Andreae, A. 1887. Die Glossophoren des Terrain à chailles der Pfirt. Abhandlungen der geologischen Specialkarte von Elsaß-Lothringen 4: 1–45.
Bandel, K. 1991. Über triassische „Loxonematoidea“ und ihre Beziehungen zu rezenten und paläozoischen Schnecken. Paläontologische Zeitschrift 65: 239–268. Crossref
Bandel, K. 1992. Über Caenogastropoda der Cassianer Schichten (Obertrias) der Dolomiten (Italien) und ihre taxonomische Bewertung. Mitteilungen aus dem Geologisch-Paläontologischen Institut der Universität Hamburg 73: 37–97.
Bandel, K. 1993. Caenogastropoda during Mesozoic times. Scripta Geologica, Special Issue 2: 7–56.
Bandel, K. 1994. Triassic Euthyneura (Gastropoda) from St. Cassian Formation (Italian Alps) with a discussion on the evolution of the Heterostropha. Freiberger Forschungsheft C 452: 79–100.
Bandel, K. 1995. Mathildoidea (Gastropoda, Heterostropha) from the Late Triassic St Cassian Formation. Scripta Geologica 111: 1–83.
Bandel, K. 1996. Some heterostrophic gastropods from Triassic St. Cassian Formation with a discussion on the classification of the Allogastropoda. Paläontologische Zeitschrift 70: 325–365. Crossref
Bandel, K. 1999. On the origin of the carnivorous gastropod group Naticoidea (Mollusca) in the Cretaceous with description of some convergent but unrelated groups. Greifswalder Geowissenschaftliche Beiträge 6: 143–175.
Bandel, K. 2006. Families of the Cerithioidea and related superfamilies (Palaeo-Caenogastropoda; Mollusca) from the Triassic to the Recent characterized by protoconch morphology—including the description of new taxa. Freiberger Forschungshefte C 511: 59–138.
Bandel, K. 2007. Description and classification of Late Triassic Neritimorpha (Gastropoda, Mollusca) from the St Cassian Formation, Italian Alps. Bulletin of Geosciences 82: 215–274. Crossref
Bastow, W.J. 1991. Methods for fitting dominance/diversity curves. Journal of Vegetation Science 2: 35–46. Crossref
Batten, R.L. and Stokes, WM.L. 1986. Early Triassic gastropods from the Sinbad Member of the Moenkopi Formation, San Rafael Swell, Utah. American Museum Novitates 2864: 1–33.
Benton, M.J. and Twitchett, R.J. 2003. How to kill (almost) all life: The end-Permian extinction event. Trends in Ecology & Evolution 18: 358–365. Crossref
Beurlen, K. 1954. As faunas de lamellibrânquios do sistema Gonduânico no Paraná. In: F.W. Lange (ed.), Paleontologia do Paraná, 107–136. Comissão de Comemorações do Centenário do Paraná, Curitiba.
Beyrich, E. 1862. Zwei aus dem deutschen Muschelkalk noch nicht bekannte Avicula-artige Muscheln. Zeitschrift der Deutschen Geologischen Gesellschaft 14: 9–10.
Bittner, A. 1891. Triaspetrefakten von Balia in Kleinasien. Jahrbuch der Kaiserlich-Königlichen Geologischen Reichsanstalt 41: 97–116.
Bittner, A. 1895. Lamellibranchiaten der alpinen Trias. 1. Theil: Revision der Lamellibranchiaten von St. Cassian. Abhandlungen der Kaiserlich-Königlichen Geologischen Reichsanstalt 18: 1–235.
Bittner, A. 1901. Lamellibranchiaten aus der Trias des Bakonyer Waldes. Resultate der wissenschaftlichen Erforschung des Balatonsees. I. Band: Physische Geographie des Balatonsees und seiner Umgebung. I. Theil: Geographische Beschreibung der Balatonsee-Umgebung, samt deren Orographie und Geologie. Anhang: Palaeontologie der Umgebung des Balatonsees, II. Band, III. Abhandlung. 107 pp. Hölzel, Vienna.
Bizzarini, F. 1993. Microfossili cassiani al limite Ladinico-Carnico nella successione dei Prati di Stuores (Dolomiti orientali). Annali dei Musei Civici di Rovereto (Sez.: Archeologia, Storia e Scienze Naturali) 8: 141–168.
Bizzarini, F. 1996. Stratigraphic distribution of the microfossils in St. Cassian Formation (Upper Triassic between Badia and Ampezzo valley (Dolomites, Italy). In: G. Braga, F. Finotti, and G. Piccoli (eds.), Reports of Shallow Tethys 4. International Symposium, Albrechtsberg, Austria, 8–11 September 1994. Annali dei Musei Civici di Rovereto (Sez.: Archeologia, Storia e Scienze Naturali), Supplement 11: 207–213.
Bizzarini, F. and Gnoli, M. 1991. Trematoceras elegans (Münster) and other Late Triassic cephalopods from the San Cassiano Formation, Eastern Dolomites (Italy). Bolletino della Societa Paleontologica Italiana 30: 109–116.
Bizzarini, F. and Laghi, G.F. 2005. La successione “Cassiana” nell’area a nord di Misurina (Trias, Dolomiti). Lavori—Società Veneziana di Scienze Naturali 30: 127–143.
Bizzarini, F. and Rottonara, E. 1997. La Tanatocenosi a Vertebrati delle Heiligenkreuzschichten in Alta Val Badia, Dolomiti. Bollettino del Museo Civico di Storia Naturale di Venezia 47: 307–316.
Blainville, H.M.D. de 1834. Manuel d’Actinologie et de Zoophytologie. 2 volumes. viii + 694 pp. Levrault, Paris. Crossref
Blake, D.B. 2017. Paleozoic echinoderm hangovers: Waking up in the Triassic: Comment. Geology 45: e417. Crossref
Bonarelli, G. 1927. Fósiles de la Formación Petrolífera o “Sistema de Salta”. Boletín de la Academia Nacional de Ciencas de la república Argentina 30: 51–115.
Bond, D.P.G. and Wignall, P.B. 2010. Pyrite framboid study of marine Permian/Triassic boundary sections: A complex anoxic event and its relationship to contemporaneous mass extinction. GSA Bulletin 122: 1265–1279. Crossref
Bosellini, A. 1998. Geologie der Dolomiten. 192 pp. Athesia, Bolzano.
Bouchet, P. 2009. From specimens to data, and from seashells to molluscs: The Panglao Marine Biodiversity Project. Vita Malacologica 8: 1–8.
Bouchet, P., Lozouet, P., Maestrati, P., and Heros,
V. 2002. Assessing the magnitude of species richness in tropical marine
environments: exceptionally high numbers of molluscs at a New
Caledonia site. Biological Journal of the
Linnean Society 75: 421–436. Crossref
Bouchet, P., Rocroi, J.-P., Hausdorf, B., Kaim, A., Kano, Y, Nützel, A., Parkhaev, P., Schrödl, M., and Strong, E.E. 2017. Revised classification, nomenclator and typification of gastropod and monoplacophoran families. Malacologia 61: 1–526. Crossref
Brandt, J.F. 1835. Prodromus descriptionis animalium ab H. Mertensio in orbis terrarum circumnavigatione observatorum. In Animalia Mertensii. Fascic[ulus] 1. 75 pp. Graeff, Petropoli [= Saint Petersburg] and L. Voss, Lipsiae [= Leipzig].
Broglio Loriga, C.B., Cirilli, S., De Zanche, V., Di Bari, D., Gianolla, P., Laghi, G.F., Lowrie, W., Manfrin, S., Mastandrea, A., Mietto, P., Muttoni, G., Neri, C., Posenato, R., Rechichi, M., Rettori, R., and Roghi, G. 1999. The Prati di Stuores/Stuores Wiesen section (Dolomites, Italy): A candidate global stratotype section and point for the base of the Carnian Stage. Rivista Italiana di Paleontologia e Stratigrafia 105: 37–78.
Bronn, H.G. 1862 [in 1862–1866]. Klassen und Ordnungen des Thier-Reichs, wissenschaftlich dargestellt in Wort und Bild. Dritter Band, Malacozoa. Erste Abtheilung. 1500 pp. 136 pls [1862: 523–650, pls 45–49]. C.F. Winter’sche Verlagshandlung, Leipzig. Crossref
Bruguière, J.G. 1789–1816. Encyclopédic méthodique ou par ordre de matières. Histoire naturelle de Vers, des Mollusques. 758 pp. Charles-Joseph Panckoucke, Paris. Crossref
Carter, J.G., Altaba, C.R., Anderson, L.C., Araujo, R., Biakov, A.S., Bogan, A.E., Campbell, D.C., Campbell, M., Jin-hua, C., Cope, J.C.W., Delvene, G., Dijkstra, H.H., Zong-jie, F., Gardner, R.N., Gavrilova, V.A., Goncharova, I.A., Harries, P.J., Hartman, J.H., Hautmann, M., Hoeh, W.R., Hylleberg, J., Bao-yu, J., Johnston, P., Kirkendale, L., Kleemann, K., Koppka, J., Kříž, J., Machado, D., Malchus, N., Márquez-Aliaga, A., Masse, J.-P., McRoberts, C.A., Middelfart, P.U., Mitchell, S., Nevesskaja, L.A., Özer, S., Pojeta, J.Jr., Polubotko, I.V., Pons, J.M., Popov, S., Sánchez, T., Sartori, A.F., Scott, R.W., Sey, I.I., Signorelli, J.H., Silantiev, V.V., Skelton, P.W., Steuber, T., Waterhouse, J.B., Wingard, G.L., and Yancey, T. 2011. A synoptical classification of the Bivalvia (Mollusca). Paleontological Contributions 4: 1–47.
Carter, J.G., Campbell, D.C., and Campbell, M.R. 2000. Cladistic perspectives on early bivalve evolution. In: E.M. Harper, J.D. Taylor, and J.A. Crame (eds.), The Evolutionary Biology of the Bivalvia. Geological Society London Special Publications 177: 47–79. Crossref
Children, J.G. 1834. Synopsis of the Contents of the British Museum, 28th Edition. iv + 240 pp. British Museum, London.
Claus, C.F.W. 1880. Grundzüge der Zoologie, 4th ed., Vol. 1. vii + 821 pp. N.G. Elwert, Marburg.
Conrad, T.A. 1842. Descriptions of new species of organic remains belonging to the Silurian, Devonian and Carboniferous systems of the United States. Journal of the Academy of Natural Sciences of Philadelphia 8: 183–190.
Cossmann, M. 1885. Contribution à l’étude de la faune de l’étage Bathonien en France (Gastropodes). Mémoirs de la Société géologique de France (3) 3: 1–374.
Cossmann, M. 1895–1918. Essais de Paléoconchologie comparée. Livraison 1 (1895): 159 pp., 2 (1896): 179 pp., 6 (1904): 151 pp., 8 (1909): 348 pp., 9 (1912): 215 pp., 11 (1918): 388 pp. Cossmann, Paris.
Cossmann, M. 1897. Paléoconchologie et ouvrages généraux. Terrains paléozoiques et secondaires. Revue Critique de Paléozoologie 1: 46–59.
Cossmann, M. 1904. Note sur l’Infralias de la Vendée et des Deux-Sèvres. II. Pélécypodes. Bulletin de la Société géologique de France (4) 3: 497–537.
Cox, L.R. 1960. Thoughts on the classification of the Gastropoda. Proceedings of the Malacological Society of London 33: 239–261.
Cuvier, G. 1795. Second Mémoire sur l’organisation et les rapports des animaux à sang blanc, dans lequel on traite de la structure des Mollusques et de leur division en ordre, lu à la société d’Histoire naturelle de Paris, le 11 prairial, an troisième. Magazin Encyclopédique, ou Journal des Sciences, des Lettres et des Arts 2: 433–449.
Dall, W.H. 1889. On the hinge of pelecypods and its development, with an attempt toward a better subdivision of the group. The American Journal of Science and Arts, Series 3 38: 445–462. Crossref
Deflandre-Rigaud, M. 1963. État actuel de nos connaissances sur la représentation, au Trias, de quelques groupes de micro-organites. Vestiges microscopiques d’Invertébrés. Memoires du Bureau de Recherches Géologiques et Minières 15: 562–563.
Defrance, M.J.L. 1820. Gervillie, Gervillia. Dictionaire des Sciences Naturelles 18: 502–503.
Defrance, M.J.L. 1825. Nucule (Fossiles). Dictionnaire des Sciences Naturelles 35: 216–219.
Deshayes, G.P. 1839–1853. Traité élémentaire de conchiologie, avec les applications de cette sciences à la geologie. 1588 pp. V. Masson, Paris. Crossref
Diener, C. 1923. Lamellibranchiata triadica I. In: C. Diener (ed.), Fossilium Catalogus, I Animalia, 19. Gustav Feller, Neubrandenburg.
Diener, C. 1926. Glossophora triadica. In: C. Diener (ed.), Fossilium Catalogus, I Animalia, 34. W. Junk, Berlin.
Ellingsen, K. and Gray, J.S. 2002. Spatial patterns of benthic diversity: is there a latitudinal gradient along the Norwegian continental shelf? Journal of Animal Ecology 71: 373–389. Crossref
Erwin, D.H. 1989. The end-Permian mass extinction: what really happened and did it matter? Trends in Ecology & Evolution 4: 225–229. Crossref
Férussac, A.E.J. d’Audebard de 1822. Tableaux systématiques des animaux mollusques suivis d’un Prodrome général pour tous les mollusques terrestres ou fluviatiles vivants ou fossiles. Première partie, Tableaux systématiques généraux. 47 pp. Arthus-Bertrand, Paris.
Fleming, J. 1828. A History of British Animals, Exhibiting the Descriptive Characters and Systematical Arrangement of the Genera and Species of Quadrupeds, Birds, Reptiles, Fishes, Mollusca, and Radiata of the United Kingdom; Including the Indigenous, Extirpated, and Extinct Kinds, Together with Periodical and Occasional Visitants. xxii + 565 pp. + corrigenda. Bell & Bradfute, Edinburgh and James Duncan, London. Crossref
Forcino, F.L. and Stafford, E.S. 2020. The influence of collection method on paleoecological datasets: In-place versus surface-collected fossil samples in the Pennsylvanian Finis Shale, Texas, USA. PLoS ONE 15 (2): e0228944. Crossref
Foster, W.J., Danise, S., and Twitchett, R.J. 2017. A silicified Early Triassic marine assemblage from Svalbard. Journal of Systematic Palaeontology 15: 851–877. Crossref
Friesenbichler, E., Hautmann, M., Nützel, A., Urlichs, M., and Bucher, H. 2019. Palaeoecology of Late Ladinian (Middle Triassic) benthic faunas from the Schlern/Sciliar and Seiser Alm/Alpe di Siusi area (South Tyrol, Italy). Paläontologische Zeitschrift 93: 1–29. Crossref
Frizzell, D.L. and Exline, H. 1956. Monograph of Fossil Holothurian Sclerites. Bulletin of School of Mines and Metallurgy (Technical Series) 89 (for 1955): 1–204. Crossref
Frontier, S. 1985. Diversity and structure in aquatic ecosystems. In: M. Barnes (ed.), Oceanography and Marine Biology—An Annual Review 23, 253–312. Aberdeen University Press, Aberdeen.
Fuchs, T. 1871. Über die lokale Anhäufung kleiner Organismen und insbesondere über die Fauna von St. Cassian. Verhandlungen der Kaiserlich-Königlichen Geologischen Reichsanstalt 1871: 204–206.
Fürsich, F.T. and Wendt, J. 1977. Biostratinomy and palaeoecology of the Cassian Formation (Triassic) of the Southern Alps. Palaeogeography, Palaeoclimatology, Palaeoecology 22: 257–323. Crossref
Fürsich, F.T., Oschmann, W., Pandey, D.K., Jaitly, A.K., Singh, I.B., and Liu. C. 2004. Palaeoecology of Middle to Lower Upper Jurassic macrofaunas of the Kachchh Basin, Western India: An overview. Journal of the Palaeontological Society of India 49: 1–26.
Gale, A. 2011. Phylogeny of the Neoasteroidea (post-Palaeozoic Asteroidea, Echinodermata). Special Papers in Palaeontology 85: 1–112.
Giebel, C.G. 1852. Deutschlands Petrefacten – Ein systematisches Verzeichniss aller in Deutschland und den angrenzenden Ländern vorkommenden Petrefacten nebst Angabe der Synonymen und Fundorte. 706 pp. Ambrosius Abel, Leipzig.
Gilliland, P.M. 1992. Holothurians in the Blue Lias of southern Britain. Palaeontology 35: 159–210.
Godbold, A, Schoepfer, S., Shen, S., and Henderson, C.M. 2017. Precarious ephemeral refugia during the earliest Triassic. Geology 45: 607–610. Crossref
Goldfuss, G.A. 1834–44. Petrefacta Germaniae, II: 1–312, pls. 72–165, III. 7. Lieferung (1841): 1–28, pls. 166–171; 8.–9. Lieferung (1844): 29–128, pls. 172–200. Arnz, Düsseldorf.
Gray, J.E. 1824. A Supplement to the Appendix of Captain Perry’s Voyage for the Discovery of a North West Passage in the Years 1819–1820, Containing an Account of the Subjects of Natural History. Appendix X. Natural History, Shells, 240–246. J. Murray, London.
Gray, J.E. 1847. A list of genera of Recent Mollusca, their synonyma and types. Proceedings of the Zoological Society of London 15: 129–219.
Gray, J.E. 1854. A revision of the arrangement of the families of bivalve shells (Conchifera). The Annals and Magazine of Natural History, Series 2 13: 408–418. Crossref
Gründel, J. 1973. Zur Gastropodenfauna aus dem Dogger. I. Die Gattungen Mathilda und Eucycloidea. Zeitschrift für Geologische Wissenschaften 1: 947–965.
Gründel, J. 1997a. Heterostropha (Gastropoda) aus dem Dogger Norddeutschlands und Nordpolens. I. Mathildoidea (Mathildidae). Berliner Geowissenschaftliche Abhandlungen, Reihe E 25: 131–175.
Gründel, J. 1997b. Heterostropha (Gastropoda) aus dem Dogger Norddeutschlands und Nordpolens. III. Opisthobranchia. Berliner Geowissenschaftliche Abhandlungen, Reihe E 25: 177–223.
Gründel, J. and Nützel, A. 2013. Evolution and classification of Mesozoic mathildoid gastropods. Acta Palaeontologica Polonica 58: 803–826. Crossref
Gümbel, C.W. 1869. Ueber Foraminiferen, Ostracoden und mikroskopische Thier-Ueberreste in den St. Cassianer und Raibler Schichten. Jahrbuch der Kaiserlich-Königlichen Geologischen Reichsanstalt 19: 175–186.
Guzhov, A.V. 2004. Jurassic gastropods of European Russia (orders Cerithiiformes, Bucciniformes, and Epitoniiformes). Paleontological Journal 36, Supplement 5: S457–S592.
Hagdorn, H. 2011. Benthic crinoids from the Triassic Cassian Formation of the Dolomites. Geo.Alp 8: 128–135.
Hagdorn, H. 2018. Slipped through the bottleneck: Lazarechinus mirabeti gen. et. sp. nov., a Paleozoic-like echinoid from the Triassic Muschelkalk (late Anisian) of East France. Paläontologische Zeitschrift 92: 267–282. Crossref
Hall, J. 1847. Palaeontology of New York Vol. I. 338 pp. Van Benthuysen, Albany. Crossref
Hall, J. and Whitfield, R.P. 1869. Preliminary Notice of the Lamellibranchiate Shells of the Upper Helderberg, Hamilton and Chemung Groups, With Others from the Waverly Sandstones, Pt. 2. 97 pp. New York State Museum, Albany.
Hall, J. and Whitfield, R.P. 1877. Paleontology, v. 4. U.S. Geological Survey Exploration of the 40th Parallel. U.S. Geological Survey Report 2: 197–302.
Hammer, Ø. 2019. PAST: PAleontological STatistics, Version 3.25, Reference Manual, 1999–2019. 275 pp. Natural History Museum, University of Oslo.
Hammer, Ø., Harper, D.A.T., and Ryan, P.D. 2001. PAST: Paleontological statistics software package for education and data analysis. Palaeontologia Electronica 4: 1–9.
Harborne, A.R., Mumby, P.J., Żychaluk, K., Hedley, J.D., and Blackwell, P.G. 2006. Modeling the beta diversity of coral reefs. Ecology 87: 2871–2881. Crossref
Harper, D.A.T. 1999. Numerical Palaeobiology: Computer-Based Modelling and Analysis of Fossils and their Distributions. 468 pp. John Wiley & Sons Ltd, Chichester.
Haude, R. 2004. Mode of life of ophiocistioids (Echinozoa) according to plated and “naked” forms in the Rhenish Devonian. In: T. Heinzeller and J.H. Nebelsick (eds.), Echinoderms: München, 409–416. Taylor & Francis Group, London. Crossref
Hauer, F. Ritter von 1857. Ein Beitrag zur Kenntniss der Raibler Schichten. Sitzungsberichte der Kaiserlichen Akademie der Wissenschaften, Mathematisch-Naturwissenschaftliche Classe 24: 537–566.
Hausmann, I.M. and Nützel, A. 2015. Diversity and palaeoecology of a highly diverse Late Triassic marine biota from the Cassian Formation of north Italy. Lethaia 48: 235–255. Crossref
Hausmann, I.M., Domanski, H., and Zuschin, M. 2018. Influence of setting, sieve size, and sediment depth on multivariate and univariate assemblage attributes of coral reef-associated mollusc death assemblages from the Gulf of Aqaba. Facies 64: 20. Crossref
Hautmann, M. 2001. Die Muschelfauna der Nayband-Formation (Obertrias, Nor-Rhät) des östlichen Zentraliran. Beringeria 29: 3–181.
Hautmann, M. 2007. Steuerungsfaktoren globaler Biodiversifizierungsmuster am Beispiel frühmesozoischer Muscheln. Beringeria 37: 61–74.
Hautmann, M. 2014. Diversification and diversity partitioning. Paleobiology 40: 162–176. Crossref
Hofmann, R., Hautmann, M., and Bucher, H. 2013a. A new paleoecological look at the Dinwoody Formation (Lower Triassic, western USA): intrinsic versus extrinsic controls on ecosystem recovery after the end-Permian mass extinction. Journal of Paleontology 87: 854–880. Crossref
Hofmann, R., Hautmann, M., Brayard, A., Nützel, A., Bylund, K.G., Jenks, J.F., Vennin, E., Olivier, N., and Bucher, H. 2014. Recovery of benthic marine communities from the end-Permian mass extinction at the low latitudes of eastern Panthalassa. Palaeontology 57: 547–589. Crossref
Hofmann, R., Hautmann, M., Wasmer, M., and Bucher, H. 2013b. Palaeoecology of the Spathian Virgin Formation (Utah, USA) and its implications for the Early Triassic recovery. Acta Palaeontologica Polonica 58: 149–173.
Hunter, A.W. and McNamara, K.J. 2017. Paleozoic echinoderm hangovers: Waking up in the Triassic: Comment. Geology 45: e431. Crossref
Ichikawa, K. 1958. Zur Taxonomie und Phylogenie der triadischen „Pteriidae“ (Lamellibranch.) mit besonderer Berücksichtigung der Gattungen Claraia, Eumorphotis, Oxytoma und Monotis. Palaeontographica A 111: 131–212.
Janofske, D. 1992. Kalkiges Nannoplankton, insbesondere Kalkige Dinoflagellaten-Zysten der alpinen Ober-Trias: Taxonomie, Biostratigraphie und Bedeutung für die Phylogenie der Peridiniales. Berliner geowissenschaftliche Abhandlungen, Reihe E: Paläobiologie 4: 1–53.
Kaim, A. 2004. The evolution of conch ontogeny in Mesozoic open sea gastropods. Palaeontologia Polonica 62: 3–183.
Kaim, A., Hryniewicz, K., Little, C.T.S., and Nakrem, H.A. 2017. Gastropods from the Late Jurassic–Early Cretaceous seep deposits in Spitsbergen, Svalbard. Zootaxa 4329: 351–374. Crossref
Kaim, A., Jurkovšek, B., and Kolar-Jurkovšek, T. 2006. New associations of Carnian gastropods from Mežica region in the Karavanke Mountains of Slovenia. Facies 52: 469–482. Crossref
Kase, T. and Ishikawa, M. 2003. Mystery of naticid predation history solved: Evidence from a “living fossil” species. Geology 31: 403–406. Crossref
Keim, L., Spötl, C., and Brandner, R. 2006. The aftermath of the Carnian carbonate platform demise: a basinal perspective (Dolomites, Southern Alps). Sedimentology 53: 361–386. Crossref
Kier, P.M. 1965. Evolutionary trends in Paleozoic echinoids. Journal of Paleontology 39: 436–465.
Kier, P.M. 1984. Echinoids from the Triassic (St. Cassian) of Italy, their lantern supports, and a revised phylogeny of Triassic echinoids. Smithsonian Contributions to Paleobiology 56: i–iii + 1–41. Crossref
King, W. 1850. A monograph of the Permian fossils of England. Palaeontographical Society Monograph, London 30: i–xxxviii + 1–258. Crossref
Kittl, E. 1891. Die Gastropoden der Schichten von St. Cassian der südalpinen Trias. I. Theil. Annalen des Kaiserlich-Königlichen Naturhistorischen Hofmuseums 6: 166–262. Crossref
Kittl, E. 1892. Die Gastropoden der Schichten von St. Cassian der südalpinen Trias. II. Theil. Annalen des Kaiserlich-Königlichen Naturhistorischen Hofmuseums 7: 35–97. Crossref
Kittl, E. 1894. Die Gastropoden der Schichten von St. Cassian der südalpinen Trias. III. Theil. Annalen des Kaiserlich-Königlichen Naturhistorischen Hofmuseums 9: 143–277.
Kittl, E. 1899. Die Gastropoden der Esinokalke, nebst einer Revision der Gastropoden der Marmolatakalke. Annalen des Naturhistorischen Museums in Wien 14: 1–238.
Klipstein, A. von 1843–1845. Mittheilungen aus dem Gebiete der Geologie und Palaeontologie. 1. Lieferung (1843), 1–144, pls. 1–9, 2. Lieferung (1844), 145–240, pls. 10–15, 3. Lieferung (1845), 241–311, pls. 16–20. G.F. Heyer’s Verlag, Gießen.
Klompmaker, A.A., Nützel, A., and Kaim, A. 2016. Drill hole convergence and a quantitative analysis of drill holes in mollusks and brachiopods from the Triassic of Italy and Poland. Palaeogeography, Palaeoclimatology, Palaeoecology 457: 342–359. Crossref
Knight, J.B. 1936. Notes on Paleozoic Gastropoda. Journal of Paleontology 10: 520–534.
Koken, E. 1889. Ueber die Entwicklung der Gastropoden vom Cambrium bis zur Trias. Neues Jahrbuch für Mineralogie, Geologie und Palaeontologie, Beilage-Band 6: 440–463.
Koken, E. 1892. Ueber die Gastropoden der rothen Schlernschichten nebst Bemerkungen über Verbreitung und Herkunft einiger triassischer Gattungen. Neues Jahrbuch für Mineralogie, Geologie und Palaeontologie 1892: 25–36.
Koken, E. 1896. Die Gastropoden der Trias um Hallstatt. Jahrbuch der Kaiserlich-Königlichen Geologischen Reichsanstalt 46: 37–126.
Koken, E. 1913. Beiträge zur Kenntnis der Schichten von Heiligenkreuz (Abteital, Südtirol). Abhandlungen der Kaiserlich-Königlichen Geologischen Reichssanstalt 16: 1–43.
Kollarova-Andrusova, V. and Kochanova, M. 1973. Die Molluskenfauna des Bleskovy pramen bei Drnava (Nor, Westkarpaten). 215 pp. Slovenska Akademia Vied, Bratislava.
Korte, C., Kozur, H.W., and Veizer, J. 2005. δ13C and δ18O values of Triassic brachiopods and carbonate rocks as proxies for coeval seawater and palaeotemperature. Palaeogeography, Palaeoclimatology, Palaeoecology 226: 287–306. Crossref
Kowalewski, M. and Novack-Gottshall, P. 2010. Resampling methods in paleontology. In: J. Alroy and G. Hunt (eds.), Quantitative Methods in Paleobiology, Paleontological Society Short Course, October 30th, 2010. The Paleontological Society Papers 16: 19–54. Crossref
Kowalewski, M., Kiessling, W., Aberhan, M., Fürsich, F.T., Scarponi, D., Barbour Wood, S.L., and Hoffmeister, A.P. 2006. Ecological, taxonomic, and taphonomic components of the post-Paleozoic increase in sample-level species diversity of marine benthos. Paleobiology 32: 533–561. Crossref
Krebs, C.J. 1989. Ecological Methodology. 654 pp. Harper & Row, New York.
Kristan-Tollmann, E. 1960. Rotaliidea (Foraminifera) aus der Trias der Ostalpen. Jahrbuch der Geologischen Bundesanstalt, Sonderband 5: 47–78.
Kristan-Tollmann, E. 1970. Die Osteocrinusfazies, ein Leithorizont von Schwebcrinoiden im Oberladin-Unterkarn der Tethys. Erdöl und Kohle, Erdgas, Petrochemie, vereinigt mit Brennstoff-Chemie 23: 781–789.
Kroh, A. 2011. Echinoids from the Triassic of St. Cassian—a review. Geo.Alp 8: 136–140.
Kutassy, A. 1931. Lamellibranchiata triadica II. In: W. Quenstedt (ed.), Fossilium Catalogus, I Animalia, 51, iii + iv + 261–475. W. Junk, Berlin.
Kutassy, A. 1940. Fossilium Catalogus, I Animalia, 81, Glossophora triadica II. 477 pp. W. Junk, Berlin.
Kutscher, M. and Reich, M. 2004. Archaeocidarid and bothriocidarid Echinozoa from the Silurian of Gotland. In: T. Heinzeller and J.H. Nebelsick (eds.), Echinoderms: München, 457–458. Taylor and Francis Group, London. Crossref
Lamarck, J.B. 1799. Prodrome d’une nouvelle classification des coquilles. Mémoires de la Société d’Histoire Naturelle de Paris 1: 63–91.
Lamarck, J.B. 1809. Philosophie Zoologique: Tome 1. xxv + 428 pp. Dentu, Paris.
Langer, W. 1991. Beiträge zur Mikropaläontologie des Devons im Rheinischen Schiefergebirge. Geologisches Jahrbuch A 128: 35–65.
Laube, G.C. 1864. Bemerkungen über die Münster’schen Arten von St. Cassian in der Münchener paläontologischen Sammlung. Jahrbuch der Kaiserlich-Königlichen Geologischen Reichsanstalt 14: 402–412.
Laube, G.C. 1865. Fauna der Schichten von St. Cassian. Ein Beitrag zur Paläontologie der Alpinen Trias. II. Abtheilung: Brachiopoden und Bivalven. Denkschriften der Kaiserlichen Akademie der Wissenschaften, Mathematisch-Naturwissenschaftliche Classe 25 (2): 1–76.
Laube, G.C. 1868. Fauna der Schichten von St. Cassian. Ein Beitrag zur Paläontologie der Alpinen Trias. III. Abtheilung: Gastropoden. I. Hälfte. Denkschriften der Kaiserlichen Akademie der Wissenschaften, Mathematisch-Naturwissenschaftliche Classe 28 (2): 29–94.
Laube, G.C. 1869. Die Fauna der Schichten von St. Cassian. IV. Abtheilung. Gastropoden. II. Hälfte. Denkschriften der Kaiserlichen Akademie der Wissenschaften, Mathematisch-Naturwissenschaftliche Classe 30: 1–48.
Leonardi, P. and Fiscon, F. 1959. La fauna Cassiana di Cortina d’Ampezzo. Parte III Gasteropodi. Ricerce Geo-Paleontologiche nelle Dolomiti. Memorie degli Istituti di Geologia e Mineralogia dell’Università di Padova 21: 1–103.
Leske, N.G. 1778. Jacobi Theodori Klein naturalis dispositio echinodermatum. Accesserunt lucubratiuncula de aculeis echinorum marinorum et spicilegium de belemnitis. xx + 278 pp. Gleditsch, Lipsiae. Crossref
Link, H.F. 1807. Beschreibung der Naturalien-Sammlung der Universität zu Rostock. Abtheilung 3, 101–165. Adler, Rostock. Crossref
Linnaeus, C. 1758. Systema naturae per regna tria naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Editio decima, reformata. 824 pp. Laurentius Salvius, Stockholm. Crossref
Ludwig, H. 1891. Ankyroderma musculus (Risso), eine Molpadiide des Mittelmeeres, nebst Bemerkungen zur Phylogenie und Systematik der Holothurien. Zeitschrift für wissenschaftliche Zoologie 51: 569–612.
Ludwig, H. 1894. 12. Holothurioidea. In: Reports on an exploration off the west coasts of Mexico, Central and South America, and off the Galapagos Islands, in charge of Alexander Agassiz, by the U. S. Fish Commission Steamer “Albatross”, during 1891, Lieut. Commander Z.L. Tanner, U.S.N., commanding. Memoirs of the Museum of Comparative Zoölogy at Harvard College 17: 1–183.
Magurran, A.E. 2004. Measuring Biological Diversity. 256 pp. Blackwell Publishing, Malden.
Mansuy, H. 1914. Faune des calcaires à Productus de l’Indo-Chine. Mémoires du Service Géologiques de l’Indochine 3: 1–59.
Marshall, B.A. 1988. Skeneidae, Vitrinellidae and Orbitestellidae (Mollusca: Gastropoda) associated with biogenic substrata from bathyal depths off New Zealand and New South Wales. Journal of Natural History 22: 949–1004. Crossref
M’Coy [= McCoy], F. 1844. A Synopsis of the Characters of the Carboniferous Limestone Fossil of Ireland. 207 pp. University Press by M.H. Gill, Dublin. Crossref
M’Coy [= McCoy], F. 1849. On some new Palæozoic Echinodermata. Annals and Magazine of Natural History Series 2 3: 244–254. Crossref
Miller, A.K., Kerr, A.M., Paulay, G., Reich, M., Wilson, N.G., Carvajal, J.I., and Rouse, G.W. 2017. Molecular phylogeny of extant Holothuroidea (Echinodermata). Molecular Phylogenetics and Evolution 111: 110–131. Crossref
Mostler, H. 1968. Holothurien-Sklerite aus oberanisischen Hallstätterkalken (Ostalpen, Bosnien, Türkei). Alpenkundliche Studien (Universität Innsbruck) 2: 44 pp.
Münster, G. zu 1836: Mitteilungen an Professor Bronn gerichtet. Neues Jahrbuch für Mineralogie, Geognosie, Geologie und Petrefaktenkunde, Stuttgart 1836: 580–583.
Münster, G. zu 1841. Beschreibung und Abbildung der in den Kalkmergelschichten von St. Cassian gefundenen Versteinerungen. In: H.L. Wissmann and G. Graf zu Münster (eds.), Beiträge zur Geognosie und Petrefacten-Kunde des südöstlichen Tirol’s vorzüglich der Schichten von St. Cassian. Beiträge zur Petrefacten-Kunde 4: 25–152.
Nose, M., Schlagintweit, F., and Nützel, A. 2018. New record of halimedacean algae from the Upper Triassic of the Southern Alps (Dolomites, Italy). Rivista Italiana di Paleontologia e Stratigrafia 124: 421–431.
Nützel, A. 1998. Über die Stammesgeschichte der Ptenoglossa (Gastropoda). Berliner Geowissenschaftliche Abhandlungen, Reihe E 26: 1–229.
Nützel, A. 2005. Recovery of gastropods in the Early Triassic. Comptes Rendus Palevol 4: 501–515. Crossref
Nützel, A. 2010. A review of the Triassic gastropod genus Kittliconcha Bonarelli, 1927—implications for the phylogeny of Caenogastropoda. Zitteliana A 50: 9–24.
Nützel, A. 2014. Larval ecology and morphology in fossil gastropods. Palaeontology 57: 479–503. Crossref
Nützel, A. and Gründel, J. 2015. Early Jurassic (Pliensbachian) gastropods from Franconia, Southern Germany. Palaeontographica A 305 (1–3): 1–89. Crossref
Nützel, A. and Kaim, A. 2014. Diversity, palaeoecology and systematics of a marine fossil assemblage from the Late Triassic Cassian Formation at Settsass Scharte, N Italy. Paläontologische Zeitschrift 88: 405–431. Crossref
Nützel, A. and Kiessling, W. 1997. Gastropoden aus dem Amaltheenton (oberes Pliensbachium) Frankens. Geologische Blätter für Nordost-Bayern 47: 381–414.
Nützel, A. and Mapes, R.H. 2001. Larval and juvenile gastropods from a Carboniferous black shale: palaeoecology and implications for the evolution of the Gastropoda. Lethaia 34:143–162. Crossref
Nützel, A. and Nakazawa, K. 2012. Permian (Capitanian) gastropods from the Akasaka Limestone (Gifu Prefecture, Japan). Journal of Systematic Palaeontology 10: 103–169. Crossref
Nützel, A. and Schulbert, C. 2005. Facies of two important Early Triassic gastropod lagerstätten: implications for diversity patterns in the aftermath of the end-Permian mass extinction. Facies 51: 480–500. Crossref
Nützel, A. and Senowbari-Daryan, B. 1999. Gastropods from the Late Triassic (Norian–Rhaetian) Nayband Formation of central Iran. Beringeria 23: 93–132.
Nützel, A., Aghababalou, B., and Senowbari-Daryan, B. 2012. Gastropods from the Norian (Late Triassic) Nayband Formation near Natanz (Iran). Bulletin of Geosciences 87: 53–65. Crossref
Nützel, A., Joachimski, M., and López Correa, M. 2010a. Seasonal climatic fluctuations in the Late Triassic tropics—high-resolution oxygen isotope records from aragonitic bivalve shells (Cassian Formation, Northern Italy). Palaeogeography, Palaeoclimatology, Palaeoecology 285: 194–204. Crossref
Nützel, A., Mannani, M., Senowbari-Daryan, B., and Yazdi, M. 2010b. Gastropods from the Late Triassic Nayband Formation (Iran), their relationships to other Tethyan faunas and remarks on the Triassic gastropod body size problem. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 256: 213–228. Crossref
Östergren, H.J. 1898. Das System der Synaptiden (Vorläufige Mitteilung). Öfversigt af Kongliga Vetenskaps-Akademiens Förhandlingar 55: 111–120.
Oksanen, J., Blanchet, F.G., Friendly, M., Kindt, R., Legendre, P., McGlinn, D., Minchin, P.R., O’Hara, R.B., Simpson, G.L., Solymos, P., Stevens, M.H.H., Szoecs, E., and Wagner, H. 2016. vegan: Community Ecology Package. R package version 2.4-1. https://CRAN.R-project.org/package=vegan.
Oksanen, J., Blanchet, F.G., Friendly, M., Kindt, R., Legendre, P., McGlinn, D., Minchin, P.R., O‘Hara, R.B., Simpson, G.L., Solymos, P., Stevens, M.H.H., Szoecs, E., and Wagner, H. 2019. vegan: Community Ecology Package. R package version 2.5-4. https://CRAN.R-project.org/package=vegan.
Oliver, P.G. 1992. Bivalved Seashells of the Red Sea. 330 pp. Christa Hemmen, Wiesbaden.
Orbigny, A. d’ 1849. Prodrome de paléontologie stratigraphique universelle des animaux mollusques et rayonnés. Premier volume, 394 pp. V. Masson, Paris.
Pietsch, C., Ritterbush, K.A., Thompson, J.R., Petsios, E., and Bottjer, D.J. 2019. Evolutionary models in the Early Triassic marine realm. Palaeogeography, Palaeoclimatology, Palaeoecology 513: 65–85. Crossref
Pilsbry, H.A. and Sharp, B. 1897–1898. Class Scaphopoda. In: G.W. Tryon (ed.), Manual of Conchology: Structural and Systematic. With Illustrations of the Species. Vol. XVII, 1–144, pls. 1–26 (1897), 145–280, pls. 27–39 (1898). Conchological Section, Academy of Natural Sciences, Philadelphia.
Ponder, W.F. 1990. The anatomy and relationships of the Orbistellidae (Gastropoda: Heterobranchia). Journal of Molluscan Studies 56: 515–532. Crossref
Prado, P.I., Dantas Miranda, M., and Chalom, A. (2018). sads: Maximum Likelihood Models for Species Abundance Distributions. R package version 0.4.2. https://CRAN.R-project.org/package=sads.
Quenstedt, W. 1930. Die Anpassung an die grabende Lebensweise in der Geschichte der Solenomyiden und Nuculaceen. Geologische und Paläontologische Abhandlungen 18: 1–120.
Rafinesque, C.S. 1815. Analyse de la Nature ou Tableau de l’univers et des corps organisés. 224 pp. The author, Palermo. Crossref
Rahman, I.A., Thompson, J.R., Briggs, D.E.G., Siveter, D.J., Siveter, D.J. and Sutton, M.D. 2019. A new ophiocistioid with soft-tissue preservation from the Silurian Herefordshire Lagerstätte, and the evolution of the holothurian body plan. Proceedings of the Royal Society B: Biological Sciences 286: 20182792. Crossref
R Core Team 2016. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna. https://www.R-project.org.
R Core Team 2019. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna. https://www.R-project.org.
Reich, M. 2010a. Evolution and diversification of ophiocistioids (Echinodermata: Echinozoa). In: L.G. Harris, S.A. Boettger, C.W. Walker, and M.P. Lesser (eds.), Echinoderms: Durham: Proceedings of the 12th International Echinoderm Conference, 51–54. Taylor & Francis Group, London. Crossref
Reich, M. 2010b. The oldest synallactid sea cucumber. Paläontologische Zeitschrift 84: 541–546. Crossref
Reich, M. and Haude, R. 2004. Ophiocistioidea (fossil Echinodermata): an overview. In: T. Heinzeller and J.H. Nebelsick (eds.), Echinoderms: München: Proceedings of the 11th International Echinoderm Conference, 489–494. A.A. Balkema, Rotterdam. Crossref
Reich, M. and Smith, A.B. 2009. Origins and biomechanical evolution of teeth in echinoids and their relatives. Palaeontology 52: 1149–1168. Crossref
Reich, M., Stegemann, T.R., Hausmann, I.M., Roden, V.J., and Nützel, A. 2018. The youngest ophiocistioid: A first Palaeozoic-type echinoderm group representative from the Mesozoic. Palaeontology 61: 803–811. Crossref
Richthofen, F.F. von 1860. Geognostische Beschreibung der Umgegend von Predazzo, Sanct Cassian und der Seisser Alpe in Süd-Tyrol. 327 pp. Perthes, Gotha.
Roden, V.J., Hausmann, I.M., Nützel, A., Seuss, B., Reich, M., Urlichs, M., Hagdorn, H., and Kiessling, W. 2020a. Fossil liberation: a model to explain high biodiversity in the Triassic Cassian Formation. Palaeontology 63: 85–102. Crossref
Roden, V.J., Kocsis, Á.T., Zuschin, M., and Kiessling, W. 2018. Reliable estimates of beta diversity with incomplete sampling. Ecology 99: 1051–1062. Crossref
Roden, V.J., Zuschin, M., Nützel, A., Hausmann, I.M., and Kiessling, W. 2020b. Drivers of beta diversity in modern and ancient reef-associated soft-bottom environments. PeerJ 8, e9139. Crossref
Rzehak, A. 1888. Die Foraminiferen der Nummuliten-Schichten des Waschberges und Michelsberges bei Stockerau in Nieder-Österreich. Verhandlungen der Geologischen Bundesanstalt 1888: 226–229.
Sachariewa-Kowatschewa, K. 1962. Die Trias von Kotel (Ost-Balkan). II. Teil. Scaphopoden und Gastropoden. Annuaire de l’Université de Sofia, Faculté de Biologie, Géologie et Géographie, Livre 2, Géologie 55: 91–140.
Salamon, M.A. and Gorzelak, P. 2017. Paleozoic echinoderm hangovers: Waking up in the Triassic: Comment. Geology 45 (7): e419. Crossref
Salter, J.W. 1859. Figures and Descriptions of Canadian Organic Remains, Decade 1. vi + 47 pp., 10 pl. Geological Survey of Canada, Montreal. Crossref
Salvini-Plawén, L. von 1980. A reconsideration of systematics in the Mollusca (phylogeny and higher classification). Malacologia 19: 249–278.
Schafhäutl, K.F.E. von 1863. Süd-Bayern’s Lethea Geognostica. Der Kressenberg und die südlich von ihm gelegenen Hochalpen geognostisch betrachtet in ihren Petrafakten. Vol. 1 Atlas zu Süd-Bayern’s Lethea Geognostica. Einhundert Tafeln enthaltend 1758 Original Abbildungen und 2 Karten. Vol. 1: xvi + 487 pp.; Vol. 2: iv–viii + 86 pls. + 2 maps. Leopold Voss, Leipzig.
Schenck, H.G. 1934. Classification of nuculid pelecypods. Bulletin du Musée royal d’histoire naturelle de Belgique 10 (20): 1–78.
Schlumberger, C. 1888. Note sur les holothuridées du Calcaire Grossier. Bulletin de la Société géologique de France (Série 3) 16: 437–441.
Schlumberger, C. 1890. Second note sur les holothuridées du Calcaire Grossier. Bulletin de la Société géologique de France (Série 3) 18: 191–206.
Schneider, J.A. and Carter, J.G. 2001. Evolution and phylogenetic significance of Cardioidean shell microstructure (Mollusca, Bivalvia). Journal of Paleontology 75: 607–643. Crossref
Schröder, M. 1995. Frühontogenetische Schalen jurassischer und unterkretazischer Gastropoden aus Norddeutschland und Polen. Palaeontographica, Abteilung A 238: 1–95.
Sepkoski, J.J., Jr. 1981. A factor analytic description of the Phanerozoic marine fossil record. Paleobiology 7: 36–53. Crossref
Sessa, J.A., Patzkowsky, M.E., and Bralower, T.J. 2009. The impact of lithification on the diversity, size distribution, and recovery dynamics of marine invertebrate assemblages. Geology 37: 115–118. Crossref
Shen, W., Lin, Y., Xu, L., Li, J., Wu, Y., and Sun, Y. 2007. Pyrite framboids in the Permian–Triassic boundary section at Meishan, China: Evidence for dysoxic deposition. Palaeogeography, Palaeoclimatology, Palaeoecology 253: 323–331. Crossref
Smith, A.B. 1984. Echinoid Palaeobiology. Special Topics in Palaeontology 1. xii + 190 pp. Allen and Unwin, London.
Smith, A.B. 2004a. Echinoderms (other than echinoids). In: R. Seeley and R. Cocks (eds.), Encyclopaedia of Geology, 334–341. Academic Press/Elsevier, Cambridge. Crossref
Smith, A.B. 2004b. Echinoids. In: R. Seeley and R. Cocks (eds.), Encyclopaedia of Geology, 350–356. Academic Press/Elsevier, Cambridge. Crossref
Smith, A.B. and Reich, M. 2013. Tracing the evolution of the holothurian body plan through stem-group fossils. Biological Journal of the Linnean Society 109: 670–681. Crossref
Sollas, W.J. 1899. Fossils in the University Museum. Oxford. I. On Silurian Echinoidea and Ophiuroidea. Quarterly Journal of the Geological Society of London 55: 692–715. Crossref
Soodan, K.S. 1973. Fossil Holothuroidea from Kutch, India—Part II. Geophytology 3: 81–84.
Speckmann, P. 1968. Holothurien-Sklerite aus der Mittel-Trias der Ostalpen. Mitteilungen der Bayerischen Staatssammlung für Paläontologie und historische Geologie 8: 197–218.
Stiller, F. 2001. Fossilvergesellschaftungen, Paläoökologie und paläosynökologische Entwicklung im Oberen Anisium (Mittlere Trias) von Qingyan, insbesondere Bangtoupo, Provinz Guizhou, Südwestchina. Münstersche Forschungen zur Geologie und Paläontologie 92: 1–523.
Thompson, J.R. and Bottjer, D.J. 2019. Quantitative analysis of substrate preference in Carboniferous stem group echinoids. Palaeogeography, Palaeoclimatology, Palaeoecology 513: 35–41. Crossref
Thompson, J.R. and Denayer, J. 2017. Revision of echinoids from the Tournaisian (Mississippian) of Belgium and the importance of disarticulated material in assessing palaeodiversity. Geological Journal 52: 529–538. Crossref
Thompson, J.R., Hu, S., Zhang, Q., Petsios, E., Cotton, L.J., Huang, J., Zhou, C., Wen, W., and Bottjer, D.J. 2018. A new stem group echinoid from the Triassic of China leads to a revised macroevolutionary history of echinoids during the end-Permian mass extinction. Royal Society Open Science 5: 171–548. Crossref
Thompson, J.R., Posenato, R., Bottjer, D.J., and Petsios, E. 2019. Echinoids from the Tesero member (Werfen Formation) of the Dolomites (Italy): implications for extinction and survival of echinoids in the aftermath of the end-Permian mass extinction. PeerJ 7: e7361. Crossref
Thuy, B. 2017. Paleozoic echinoderm hangovers: Waking up in the Triassic: Reply. Geology 45: e420. Crossref
Thuy, B., Hagdorn, H., and Gale, A. 2017a. Paleozoic echinoderm hangovers: Waking up in the Triassic. Geology 45: 531–542. Crossref
Thuy, B., Hagdorn, H., and Gale, A. 2017b. Paleozoic echinoderm hangovers: Waking up in the Triassic: Reply. Geology 45: e418. Crossref
Thuy, B., Hagdorn, H., and Gale, A. 2017c. Paleozoic echinoderm hangovers: Waking up in the Triassic: Reply. Geology 45: e432. Crossref
Twitchett, R.J. and Oji, T. 2005. Early Triassic recovery of echinoderms. Comptes Rendus Palevol 4: 531–542. Crossref
Urlichs, M. 2004. Kümmerwuchs bei Lobites Mojsisovics, 1902 (Ammonoidea) aus dem Unter-Karnium der Dolomiten (Ober-Trias, Italien) mit Revision der unterkarnischen Arten. Stuttgarter Beiträge zur Naturkunde, Serie B (Geologie und Paläontologie) 344: 1–37.
Urlichs, M. 2012. Stunting in some invertebrates from the Cassian Formation (Late Triassic, Carnian) of the Dolomites (Italy). Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 265: 1–25. Crossref
Urlichs, M. 2017. Revision of some stratigraphically relevant ammonoids from the Cassian Formation (latest Ladinian–early Carnian, Triassic) of St. Cassian (Dolomites, Italy). Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 283: 173–204. Crossref
Vadet, A. 1999a. Révision des Echinides de Saint Cassian et évolution des échinides post-carbonifères. I partie. Mémoires de la Société Académique du Boulonnais (Série Histoire Naturelle) 20: 1–60.
Vadet, A. 1999b. Révision des Echinides de Saint Cassian et évolution des échinides post-carbonifères. II partie. Mémoires de la Société Académique du Boulonnais (Série Histoire Naturelle) 20: 61–116.
Waagen, W. 1880. Salt-Range fossils. I, Productus-limestone fossils. Part 2, Pisces-Cephalopoda: supplement. Gasteropoda. Memoirs of the Geological Survey of India. Palaeontologia Indica series 13: 73–183.
Walther, H. 1951. Jurassische Mikrofossilien, insbesondere Gastropoden, am Südrand des Hils. Paläontologische Zeitschrift 25: 35–106. Crossref
Warén, A. and Bouchet, P. 1993. New records, species, genera and a new family of gastropods from hydrothermal vents and hydrocarbon seeps. Zoologica Scripta 22: 1–90. Crossref
Wendt, J. and Fürsich, F.T. 1980. Facies analysis and palaeogeography of the Cassian Formation, Triassic, Southern Alps. Rivista Italiana di Paleontologia e Stratigrafia 85: 1003–1028.
Wenz, W. 1938–1944. Gastropoda, Teil I. In: O.H. Schindewolf (ed.), Handbuch der Paläozoologie, Vol. 6, 1639. Borntraeger, Berlin.
Wignall, P.B. and Newton, R. 1998. Pyrite framboid diameter as a measure of oxygen deficiency in ancient mudrocks. American Journal of Science 298: 537–552. Crossref
Wignall, P.B., Newton, R., and Brookfield, M.E. 2005. Pyrite framboid evidence for oxygen-poor deposition during the Permian–Triassic crisis in Kashmir. Palaeogeography, Palaeoclimatology, Palaeoecology 216: 183–188. Crossref
Wilkin, R.T., Arthur, M.A., and Dean, W.E. 1997. History of water-column anoxia in the Black Sea indicated by pyrite framboid size distributions. Earth and Planetary Science Letters 148: 517–525. Crossref
Wilkin, R.T., Barnes, H.L., and Brantley, S.L. 1996. The size distribution of framboidal pyrite in modern sediments: An indicator of redox conditions. Geochimica et Cosmochimica Acta 60: 3897–3912. Crossref
Wissmann, H.L. 1841. Beiträge zur Geognosie und Petrefactenkunde des südöstlichen Tirols. In: H.L. Wissmann and G. Graf zu Münster (eds.), Beiträge zur Geognosie und Petrefacten-Kunde des südöstlichen Tirol’s vorzüglich der Schichten von St. Cassian. Beiträge zur Petrefacten-Kunde 4: 1–25.
Wöhrmann, S. von 1889. Die Fauna der sogenannten Cardita- und Raibler-Schichten in den Nordtiroler und bayerischen Alpen. Jahrbuch der Kaiserlich-Königlichen Geologischen Reichsanstalt 39: 181–258.
Yin, H.-F. 1985. On Hunanopecten [in Chinese]. Acta Palaeontologica Sinica 24: 635–639.
Zardini, R. 1973. [Fossili di Cortina 1] Atlante degli echinodermi cassiani (Trias medio-superiore) della regione dolomitica attorno a Cortina d’Ampezzo. 29 pp., pls. 22. Foto Ghedina, Cortina d’Ampezzo. [reprint 1976]
Zardini, R. 1978. Fossili Cassiani (Trias Medio-Superiore). Atlante dei Gasteropodi Della Formazione di S. Cassiano Raccolti Nella Regione Dolomitica Attorno a Cortina D’Ampezzo. 58 pp. Edizione Ghedina, Cortina d’Ampezzo.
Zardini, R. 1980. Fossili Cassiani (Trias Medio-Superiore). Primo aggiornamento all’ Atlante dei Gasteropodi della Formazione di S. Cassiano raccolti nella regione dolomitica attorno a Cortina D’ Ampezzo. 14 pp. Edizione Ghedina, Cortina d’Ampezzo.
Zardini, R. 1981. Fossili Cassiani (Trias Medio-Superiore) Atlante dei Bivalvi Della Formazione di S. Cassiano Raccolti Nella Regione Dolomitica Attorno a Cortina D’Ampezzo. 14 pp. Edizione Ghedina, Cortina d’Ampezzo.
Zardini, R. 1985. Fossili Cassiani (Trias Medio-Superiore). Primo aggiornamento all’Atlante dei Bivalvi e secondo aggiornamento all’Atlante dei Gasteropodi della Formazione di S. Cassiano raccolti nella regione dolomitica attorno a Cortina D’Ampezzo. 17 pp. Edizione Ghedina, Cortina d’Ampezzo.
Zardini, R. 1988. Geologia e Fossili delle Dolomiti di Cortina e dintorni. Nuova edizione a cura di Massimo Spampani. 45 pp. Edizioni Dolomiti, San Vito di Cadore.
Zittel, K.A. von 1895. Grundzüge der Paläontologie (Paläozoologie), Abhandlungen I, Invertebrata. 971 pp. Oldenburg, München.
Acta Palaeontol. Pol. 66 (1): 143–192, 2021
https://doi.org/10.4202/app.00659.2019