
Mississippian colonial tabulate and rugose corals from the Flett Formation, Liard Basin, northwest Canada
JERZY FEDOROWSKI, E. WAYNE BAMBER, and BARRY C. RICHARDS
Fedorowski, J., Bamber, E.W., and Richards, B.C. Mississippian colonial tabulate and rugose corals from the Flett Formation, Liard Basin, northwest Canada. Acta Palaeontologica Polonica 66 (3): 679–704.
Colonial Rugosa and Tabulata from the lower and middle Viséan Flett Formation of the Liard Basin (Canada) are described. Michelinia expansa is the first tabulate coral described from the Flett Formation. The new generic name Cordilleria is introduced for the North American Lithostrotionidae and its differences from the European Siphonodendron are documented. Extremely wide variability in all characters and the small size of the studied collection allowed description of numerous Liard Basin specimens in open nomenclature only. Discussion of the most important characteristics of Cordilleria gen. nov. forms an introduction to the comprehensive revision of the North American Lithostrotionidae. The state of preservation of a colony identified as Stelechophyllum cf. banffense emphasizes the necessity of recognizing the diagenetic alterations of specimens prior to their taxonomic identification.
Key words: Rugosa, Tabulata, Favositina, Lithostrotionina, taxonomy, Carboniferous, Canada.
Jerzy Fedorowski [jerzy@amu.edu.pl], Institute of Geology, Adam Mickiewicz University, Bogumiła Krygowskiego 12, 61-680 Poznań, Poland.
E. Wayne Bamber [wayne.bamber@canada.ca] and Barry C. Richards [barry.richards@canada.ca], Geological Survey of Canada (Calgary) Natural Resources Canada, Government of Canada, 3303-33rd Street Northwest, Calgary, Alberta T2L 2A7, Canada.
Received 15 September 2020, accepted 17 August 2021, available online 14 September 2021.
Copyright © 2021 J. Fedorowski et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
This paper is devoted to Mississippian (Viséan) colonial Rugosa and Tabulata collected from the Flett Formation at its type section by Jackfish Gap on Yohin Ridge in the southwest corner of the Northwest Territories (NWT), northwestern Canada and in the Flett Formation at several other localities in that region (Figs. 1–6; Table 1). It is supplemented by the paper of Fedorowski et al. (2019) devoted to the Mattson Formation rugose corals (Bashkirian) from the Liard Basin. The Flett Formation is correlated to the lower and middle Viséan, i.e., Moliniacian and Livian in the Franco-Belgian scale or upper Chadian, Arundian and Holkerian in the British scale. The Flett Formation, deposited in the eastern Liard Basin (Fig. 1), consists of the Tlogotsho, Jackfish Gap and Meilleur members, in ascending order. The eastern Cordillera in the southwest corner of the NWT has been extensively mapped and its lithostratigraphy is well established (see Geological setting for details). Prior to this study, Tabulata had not been described from the Flett Formation. Four colonies, collected in 2018 for the purpose of this paper and described below, are the first tabulate taxa described from the Flett Formation. Rugose corals are rather numerous in the Tournaisian and lower to middle Viséan strata of the northeastern Liard Basin and adjacent areas. However, the paper by Sutherland (1958) is the only detailed study of rugose corals from that area and its strata, although they are not from the Flett Formation sections described and illustrated by Richards et al. (1989), i.e., from the area sampled for the purpose of this paper. Only two fragmentary specimens of colonial rugose corals were collected by Sutherland (1958) from separate localities within the Liard Basin, but neither of them was derived from the type Flett Formation at Jackfish Gap, measured and sampled for corals by BCR in 1975 (Richards et al. 1989) and sampled for corals by Julita Biernacka and Edward Chwieduk (both UAM) in 2018 for this paper (Fig. 6).
An introduction by Sutherland (1958) of the European genus and species names for the North American Lithostrotionidae d’Orbigny, 1852, and his application of the so-called genomorph concept of Smith and Lang (1930) required revision. That concept has already been criticized and rejected by Wilson (1963), a position fully accepted and followed here. Fedorowski and Bamber (2007) suggested the necessity for the introduction of new generic names for North American Lithostrotionidae. One of such names (Cordilleria) is introduced here.
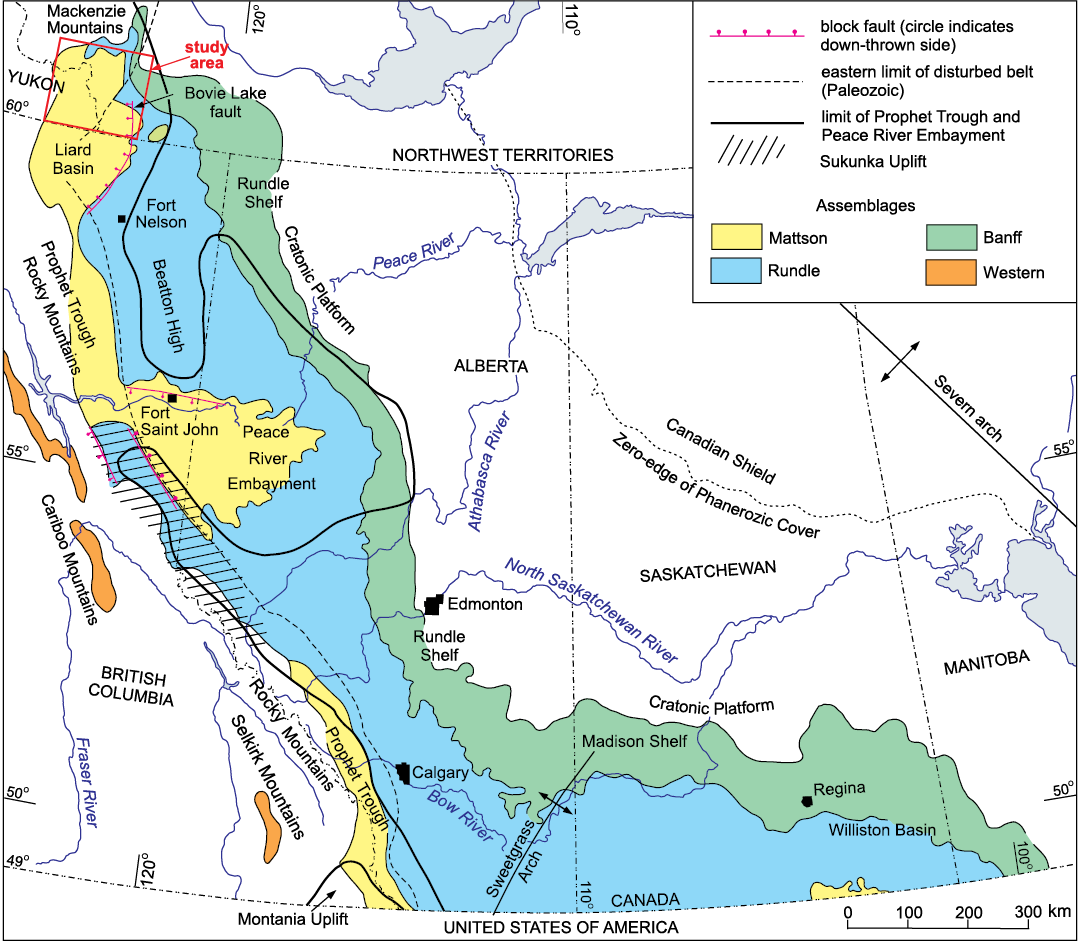
Fig. 1. Map showing location of study area, Carboniferous lithofacies assemblages subcropping beneath Permian and Mesozoic deposits in the Western Canada Sedimentary Basin, principal sub-basins, uplifts, and shelves. In the Liard Basin western occurrences of the Rundle and Matson lithofacies assemblages overlie and pass westwards into the Besa River lithofacies assemblage. See Fig. 3 for the formational composition of the Banff, Rundle, and Mattson lithofacies assemblages. Modified from Richards et al. (1994).
Specimens here identified as Cordilleria gen. nov. demonstrate extremely large variability in all skeletal structures. This resulted in misunderstandings or misinterpretations of morphological characters, making identifications of particular species doubtful. This, and a small number of specimens studied resulted in the unconventional descriptions of species. The holotypes of “Diphyphyllum mutabile Kelly, 1942”, and “Lithostrotion (Siphonodendron) oculinum Sando, 1963”, and only them, were characterized first on the basis of their earlier descriptions and illustrations, whereas the materials described herein were identified only as affinis. Such an approach allows: (i) identification of characters of a species free from a subjectivity, always possible when other samples are described as paratypes without a careful study of all characters, (ii) to establish well documented frames of the intraspecific variability when a collections larger than available for the present paper are studied.
Institutional abbreviations.—GSC (= C-), Geological Survey of Canada, Calgary, Canada; IG (= UAM-Tc.Can.), Institute of Geology, Adam Mickiewicz University, Poznań, Poland.
Other abbreviations.—d, corallite diameter (in mm); JG, Jackfish Gap; n, number of major septa; NAD27, North American Datum 1927 map projection; NATMAP, Geological Survey of Canada program name; NTS, National Topographic System; RAH, Geological Survey of Canada officer code for BCR; Tc, Tetracoralla.
Nomenclatural acts.—This published work and the nomenclatural act it contains have been registered in ZooBank urn:lsid:zoobank.org:pub:F4B631A5-CE04-4B92-B0C3-277DBDD4B386
Geological setting
Deposition of the Flett Formation occurred in the Liard Basin (Fig. 1) of Leckie et al. (1991), equivalent to the Liard Trough of Douglas et al. (1970). The Liard Basin, a sub-basin of the Prophet Trough and composite Western Canada Sedimentary Basin, occupies a prominent structural re-entrant in the eastern Cordilleran deformation front near 60°N. The composite Western Canada Sedimentary Basin, as defined by Wright et al. (1994), comprises Phanerozoic sedimentary rocks that overlie the crystalline Precambrian basement of the stable interior of the ancestral North American craton north of the United States border at 49°N and extends westwards into the eastern Cordillera, which includes the Rocky Mountains, Mackenzie Fold Belt and Franklin Mountains (Figs. 1, 2). On the east, the Bovie Lake normal fault (MacLean 2002; MacLean and Morrow 2004), with the hanging wall to the west, defines the eastern margin of the Liard Basin. Uplifted lower Paleozoic strata in the Cordillera define the western margin of the Liard Basin. The Bovie Lake fault, active from the Famennian to the Cretaceous, separates thick upper Paleozoic and Cretaceous successions in the Liard Basin from substantially thinner packages of strata to the east (Leckie et al. 1991; Richards et al. 1994; Henderson et al. 1994; Ferri et al. 2015).
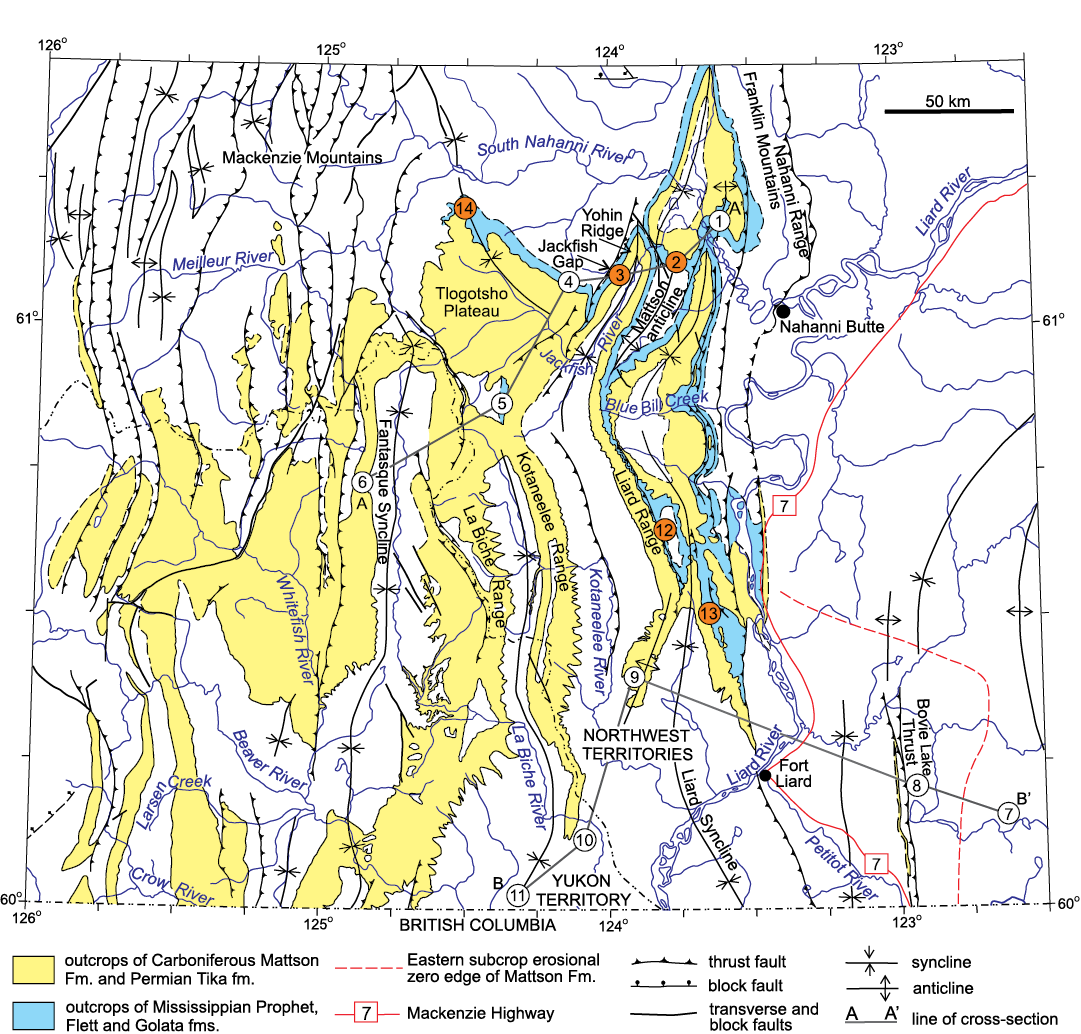
Fig. 2. Simplified geological map of study region showing distribution of outcrops of Carboniferous Prophet, Flett, and Mattson formations in southwest corner of Northwest Territories and southeast Yukon Territory and lines of cross-section A–A’ and B–B’ (see Figs. 4, 5 for details). Surface and subsurface sections (coral localities are in orange): 1, Twisted Mountain; 2, north end Mattson anticline; 3, Jackfish Gap (main coral locality); 4, Tlogotsho Plateau; 5, Etanda Lakes; 6, Tika Creek; 7, Pan American Home Signal C.S.P. Celibeta Number 7; 8, Texaco N.F.A. Bovie Lake J-72; 9, Pan American A-1 Pointed Mountain P-53; 10, Canada Southern et al. North Beaver River YT 1-27; 11, Pan American Beaver YT G-01; 12, West Flett anticline; 13, Southern Liard Range; 14, Sheaf Cree. Abbreviations: Fm., Formation (formal unit); fm., formation (informal unit); fms., formations; S., south; W., west; YT, Yukon Territory; C.S.P., Canada Southern Petroleum. Modified from Harker (1963) and an unpublished map by Andrew Okulitch from 2005 (reproduced with permission).

Fig. 3. A. Partly schematic, palinspastic stratigraphic cross-section A–A’ showing stratigraphic relationships of Flett Formation in eastern Cordillera of northern part of project area in southwest corner of Northwest Territories and southeast Yukon. B. Environments of deposition. See Fig. 2 for line of cross-section A–A’. Abbreviations: Fm., Formation (formal unit); fm., formation (informal unit); Mbr., Member (formal unit). Modified from Richards et al. (1989).
Deposition of the Flett Formation, defined by Harker (1961, 1963), occurred during the latest Tournaisian to latest middle Viséan (Ivorian to Livian in Franco-Belgian nomenclature). In this report, the revised concept of the Flett Formation as used by Richards et al. (1989) is utilized. Richards et al. (1989) restricted the definition of the Flett Formation of Harker (1961, 1963) and excluded the occurrences of the Golata and Prophet formations that Harker (1961, 1963) incorporated into the Flett Formation type section. The Flett Formation, which is separated by a strictly nomenclatural boundary at 60°N from the largely coeval and lithologically identical Debolt Formation of northeast British Columbia, comprises mainly limestone and occurs in the southwest corner of the Northwest Territories (Fig. 2). The southwestward (basinward) thinning Flett Formation is generally between 120 and 350 m thick. The type section of the Flett Formation comprises 347.3 m of strata and crops out on the west side of Yohin Ridge (Fig. 2, locality 3) north of Jackfish Gap (Richards et al. 1989). The section is 32 km west of the hamlet of Nahanni Butte in the Twisted Mountain map area (NTS 95F/4) at latitude 61°05’54”N, and longitude 123°59’26”W. Douglas and Norris mapped the Yohin Ridge (1976b, c) and the Liard Range (1976a) areas at a scale of 1:250 000. More recently, Lane (2006) and Lane and Hynes (2005) mapped the Flett Formation in the Liard Range at a scale of 1:50 000.
In most of the study area, the Flett Formation conformably overlies the Prophet Formation. Southwestern occurrences of the Flett Formation grade basinward into the Prophet Formation, with the upper Flett Formation extending farthest (Figs. 3, 4). The Flett Formation and Prophet Formation constitute a progradational succession overall representing carbonate ramps (Figs. 3, 4) that developed into poorly differentiated carbonate platforms characterized by shelf-margin sand belts. The Flett Formation is gradationally overlain by the shale-dominated Golata Formation in the eastern Cordillera and on the eastern Interior Platform. East of the subcrop edge of the Golata Formation, Cretaceous and Permian strata unconformably overlie the Flett Formation (Figs. 3, 4).

Fig. 4. A. Partly schematic, palinspastic stratigraphic cross-section showing stratigraphic relationships of Flett Formation in southern part of project area. B. Environments of deposition. See Fig. 2 for line of cross-section B–B’. Abbreviations: Fm., Formation (formal unit); fm., formation (informal unit); Mbr., Member (formal unit); mbr., member (informal unit); YT, Yukon Territory; C.S.P., Canada Southern Petroleum. Modified from Richards et al. (1994).
Richards et al. (1989) divided the Flett Formation into the Tlogotsho, Jackfish Gap and Meilleur members, in ascending order (Fig. 5). The cliff-forming, limestone-dominated Tlogotsho and Meilleur members have almost the same distribution as the Flett. In contrast, the recessive Jackfish Gap Member, which comprises siltstone, sandstone, shale, mudstone, and limestone, is confined to the Cordillera north of 60°30’N. A mixed-skeletal lime-packstone lithofacies and a lime grainstone lithofacies constitute most of the Flett Formation. A lithofacies comprising calcareous to non-calcareous shale and mudstone is intercalated with the other two lithofacies.
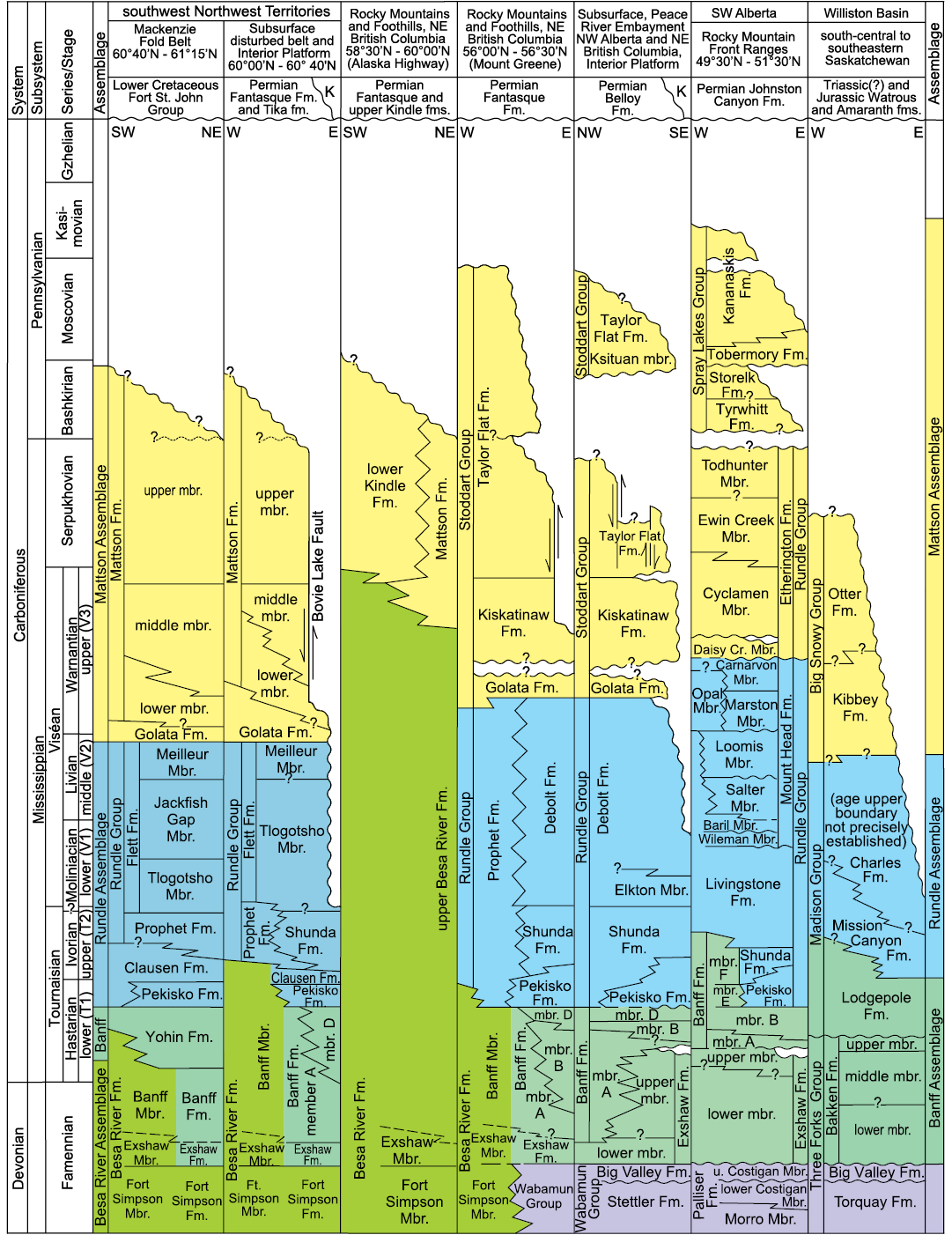
Fig. 5. Correlation of Carboniferous lithostratigraphic units in southwest corner of Northwest Territories to the Wilson Basin in southern Saskatchewan. Relationship of Viséan Flett Formation to other carbonate-dominated Mississippian formations in the West Canada Sedimentary Basin is shown and formational composition of assemblages mapped on Fig. 1; colours correlate with those on Fig. 1. Abbreviations: Cr., Creek; Fm., Formation (formal unit); fm., formation (informal unit); K, Cretaceous; Mbr., Member (formal unit); mbr., member (informal unit); u., upper. Modified from Richards et al. (1994).
Cyclic deposits of the mixed skeletal-lime packstone lithofacies constitute most of the Tlogotsho Member, the lower and western occurrences of the Meilleur Member and the limestone beds in the Jackfish Gap Member. Skeletal-lime packstone and wackestone, rhythmically intercalated with marlstone predominate in this lithofacies. Most of the limestone beds are sharp-based, massive, have great lateral continuity, and resemble turbidites but debris-flow deposits also occur. Large-scale submarine paleochannels and overlying channel fills are common in the Flett Formation. The mixed skeletal-lime packstone lithofacies grades southwestward into lower- and middle-slope deposits of the Prophet Formation. Upward and eastward, the skeletal-lime packstone lithofacies grades into the skeletal-lime grainstone lithofacies of the Flett Formation. Richards et al. (1989) interpreted most of the mixed skeletal-lime packstone lithofacies as middle-slope deposits. Most of the corals described in this report were collected from the mixed skeletal-lime packstone lithofacies.
The lime-grainstone lithofacies mainly occurs in the upper Meilleur Member but is also present in the eastern Tlogotsho Member south of 60°45’N. Bryozoan–pelmatozoan lime-grainstone and -packstone predominate. Medium- to large-scale crossbedding and hummocky cross-bedded storm deposits are moderately common. Large-scale submarine paleochannels filled with this lithofacies truncate the bedding at some localities. Richards et al. (1989) interpreted the lime-grainstone lithofacies to be of upper-slope and shelf-margin origin.
The siltstone and sandstone lithofacies occurs in the Jackfish Gap Member forming prominent coarsening-upward sequences underlain and overlain by the shale and mudstone lithofacies. At some localities, abundant trace fossils of the Cruziana ichnofacies (Seilacher 1967; Frey and Pemberton 1985) and wave- and current-formed crossbedding occur in the sandstone and siltstone. This lithofacies records deposition in slope to shallow-neritic settings (Richards et al. 1989).
Material and methods
The coral specimens discussed herein from the Flett Formation at Jackfish Gap were collected by BCR in July and August, 1975. Most of the corals described and illustrated in this paper were collected from the Flett Formation type section (Fig. 6) but several specimens are from collections BCR obtained in 1975 (Table 1) from Flett Formation sections at the northwest corner of the Tlogotsho Plateau, the Liard Range, and at the north end of the Mattson anticline (Fig. 2; localities 2, 12, 13, 14). The coral specimens that BCR collected have locality numbers with prefix “C” and their museum sample numbers with prefix “GSC”. In the main body of the text and in the figure captions herein both those numbers are mentioned (e.g., C-7449 = GSC 142468). That part of the collection is housed in the Geological Survey of Canada, Calgary, Alberta. In July and August, 2018, Julita Biernacka and Edward Chwieduk (both UAM) collected additional coral specimens from the Flett Formation with locality numbers with prefix “IG” and museum samples with prefix “UAM-Tc.Can” (e.g., IG-82 = UAM-Tc.Can./8).
Table 1. Locality data for Geological Survey of Canada-Calgary coral collection localities. Abbreviations: FGSC, Geological Survey of Canada; MTA-BCR, Geological Survey of Canada officer code for David W. Morrow and Barry C. Richards; NTS; National Topographic System; NWT, Northwest Territories.
|
GSC field collection number |
Section name |
Section location |
Stratigraphic level |
Lithostratigraphic unit |
Age |
|
C-52124 = GSC-142471 |
Jackfish Gap, |
latitude 61°05’54”N, longitude 123°59’26”W; map NTS 95-G/4, NWT; locality 3; Fig. 2 |
220.4 m above base Flett Formation |
Meilleur Member of Flett Formation |
middle Viséan |
|
C-52546 = GSC-142470 |
Jackfish Gap, |
latitude 61°05’54”N, longitude 123°59’26”W; map NTS 95-G/4, NWT; locality 3; Fig. 2 |
252.9–253.4 m above base Flett Formation |
Meilleur Member of Flett Formation |
middle Viséan |
|
C-47930 = GSC-142469 |
north end Mattson anticline, |
latitude 61°06’38”N, longitude 123°45’54”W; map NTS 95-G/4, NWT; locality 2; Fig. 2 |
176.7 m above base Flett Formation |
Jackfish Gap Member of Flett Formation |
early to middle Viséan |
|
C-52205 = GSC-142472 |
north end Mattson anticline, |
latitude 61°06’38”N, longitude 123°45’54”W; map NTS 95-G/4, NWT; locality 2; Fig. 2 |
135.0 m above base Prophet Formation; 16.5 m below base of Flett Formation |
Upper Prophet Formation |
late Tournaisian to early Viséan |
|
C-52268 = GSC-142473 |
north end Mattson anticline, |
latitude 61°06’38”N, longitude 123°45’54”W; map NTS 95-G/4, NWT; locality 2; Fig. 2 |
307.0–309.5 m above base Flett Formation |
Meilleur Member of Flett Formation |
middle Viséan |
|
C-52434 = GSC-142476 |
West Flett anticline, 75MTA-BCR10 |
latitude 60°33’50”N, longitude 123°48’20”W; map NTS 95-B/12, NWT; locality 12; Fig. 2 |
213 m above base Flett Formation |
Meilleur Member of Flett Formation |
middle Viséan |
|
C-52361 = GSC-142474 |
Southern Liard Range, |
latitude 60O29’46”N, longitude 123°38’53”W; map NTS 95-B/5, NWT; locality 13; Fig. 2 |
193.3 m above base Flett Formation |
Meilleur Member of Flett Formation |
middle Viséan |
|
C-47942 = GSC-142475 |
Sheaf Creek, |
latitude 61°12’23”N, longitude 124°33’32”W; map NTS 95-F/2, NWT; locality 14; Fig. 2 |
68.5–69.5 m above base Flett Formation |
Flett Formation undivided |
middle Viséan |
|
C-07349 = GSC-142468 |
8.05 km NW of Bone Mountain |
latitude 54°42’59.90”N, longitude 120°54’05.13”W; map NTS 93-I/10, east-central British Columbia |
384.4 m above base of section |
Rundle Group undivided |
Viséan |
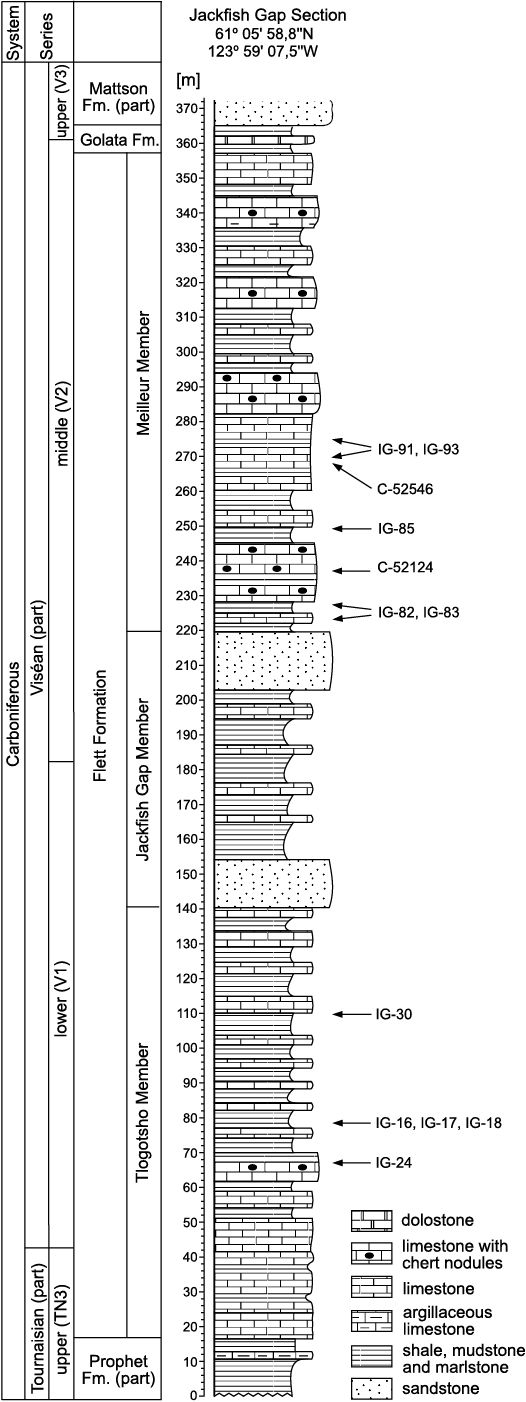
Fig. 6. Stratigraphic column of type section of Viséan Flett Formation at Jackfish Gap on Yohin Ridge, eastern Cordillera, southwest corner of Northwest Territories. Shows lithostratigraphic positions of coral collections discussed in this paper. Abbreviations: Fm., Formation. Stratigraphic column modified from Richards et al. (1989: fig. 24).
The 2018 coral collections are housed in the Institute of Geology, Adam Mickiewicz University, in Poznań, Poland. All the tabulate specimens were collected in situ in 2018 but not necessarily in growth position. The tabulates do not show traces of erosion but are corroded and recrystallized. Like the tabulates, the rugose coral colonies collected from the Flett Formation were not in growth position, being worn and apparently redeposited from their original neritic setting. The internal deformation of the corallites in the specimens varies. In most specimens, compression during compaction and diagenesis crushed only a few corallites, whereas silicification and recrystallization from deep burial diagenesis are common. Such preservation excludes effective use of the acetate peeling method for the study of blastogeny. Therefore, because we could not make usable series of peels, we described the blastogeny using random transverse and longitudinal thin sections of offsetting corallites. We slabbed and made thin sections from all colonies collected in 1975 and 2018.
Systematic palaeontology
Class Anthozoa Ehrenberg, 1834
Subclass Tabulata Milne Edwards and Haime, 1855
Order Favositina Wedekind, 1937
Suborder Favositina Wedekind, 1937
Family Michelinidae Waagen and Wentzel, 1886
Subfamily Michelininae Waagen and Wentzel, 1886
Genus Michelinia de Koninck, 1841
Diagnosis.—See Hill (1981: F561).
Remarks.—Colonies included here in the Michelinia de Konnick, 1841, belong to a morphologically complex group of species developing both tabulae and abundant dissepiments. That morphological complexity led Stumm (1948) to introduce the new genus Tabellaephyllum Stumm, 1948, based on the new species T. peculiare Stumm, 1948, and to include it in the Rugosa. Oliver and Sando (1977) rejected both the Devonian age of T. peculiare and its position within the Rugosa, identifying that species as belonging to the tabulate genus Michelinia. Also, they suggested the synonymy of T. peculiare with Michelinia expansa White, 1883, “a common and widespread element of the Early Mississippian … coral fauna…” as mentioned by Oliver and Sando (1977: 422). However, Lafuste and Plusquellec (1990) cast doubt on the identification of “M. expansa” as Michelinia by placing that generic name in inverted comas as far as the topotypes of that species are concerned. They identified Michelinia expansa of Sando (1969a) as Turnecipora sp. and in the case of Tabellaephyllum peculiare concluded: “The beautiful wording of Oliver and Sando suits us perfectly ‘Tabellaephyllum Stumm is a Michelinia’ however, silicification of the wall precludes precise attribution to a genus.” (“La belle formule de Oliver et Sando (1977) nous agrée parfaitement ‘Tabellaephyllum Stumm is a Michelinia’ mais la silicification de la muraille empêche toute attribution générique precise”) (Lafuste and Plusquellec 1990: 23). The morphology of the intercorallite walls, observed in fragments of two colonies (Figs. 7A2, 8C3), suggest they are related to Turnecipora Lafuste and Plusquellec, 1985, whereas the microstructure of the walls in the remaining parts of those colonies and in the other two colonies were destroyed by recrystallization with only the darker middle line recognizable. Thus, our suggestion is not supported firmly, precluding a further discussion of that matter and our genus and species identifications rely on the general morphology of the coral skeletons.
Michelinia expansa White, 1883
Figs. 7, 8.
Material.—Four almost complete colonies (Fig. 6) from the Flett Formation, Rundle Group, Liard Basin, Northwest Territories, Canada: IG-17 = UAM-Tc.Can./1, Tlogotsho Member (70 m above base), lower Viséan; IG-91-1 = UAM-Tc.Can/2, IG-91-2 = UAM-Tc.Can/3, IG-93 = UAM-Tc.Can/4, Meilleur Member (270 m above base and 50 m above base of member), middle Viséan. Peripheral parts of colonies, i.e., holotheca and fragments of external corallites corroded. Corallite skeletons recrystallized, partly dolomitized and/or replaced by silica. Microstructure of wall destroyed in most. Septal spines extremely rare and may have been dissolved. Corallite calices preserved in fragments of two colonies.
Description.—Colonies hemispherical, cerioid as documented by tripartite composition of inter-corallite walls (Fig. 8C), i.e., partition of Fedorowski and Jull (1976). Corallites within colonies differentiated in size into a few large corallites and many smaller corallites, surrounding large ones (Figs. 7A1, 8B2). Diameters of largest corallites: 6.5 mm in specimens UAM-Tc.Can./1, 3 and 7.5 mm in colonies UAM-Tc.Can./2, 4. Pattern described suggests either differentiated preferences in offsetting with only some polyps within colony being able to offset, or ability to offset appearing late in polyps growth. Offsetting lateral (Fig. 8A2, B1, C1, C2, C4). Offsets separated by partitions early in their growth, but a communication duct or pore connecting parental polyp and its offset recognizable at very beginning of offsetting when longitudinal section centric (e.g., Fig. 8A2, arrows; B1, grey arrows). Such communication apparently absent when section eccentric (Fig. 8B1, black arrow) and can be missing from transverse section (Fig. 8C1). Pores of early growth stage of one offset open towards two mature corallites (Fig. 8C4) recognized as well. Also, pores connecting offset with adjacent corallites common (Fig. 8A2, arrows; B1, grey arrows). Septal spines extremely rare, very small when present (Fig. 7A3). Pores connecting mature corallites rare, but both types (P1 and P2) documented; sometimes both types occurring close to one another (Fig. 7A4). Dissepiments absent from young corallites (Figs. 7A1, 8C1, C2, C4), growing in number from one row in small corallites to 2–4 rows in large corallites. Long, steeply down sloping dissepiments may occupy up to 1/2 corallite lumen. Only complete tabulae, expanding through entire corallite lumen, present during several millimeters of earliest corallite growth. In mature corallites tabulae vary from horizontal or slightly sagging to convex; all observed in same colony or even same corallite (Figs. 7A1, upper, 8A2, B1).
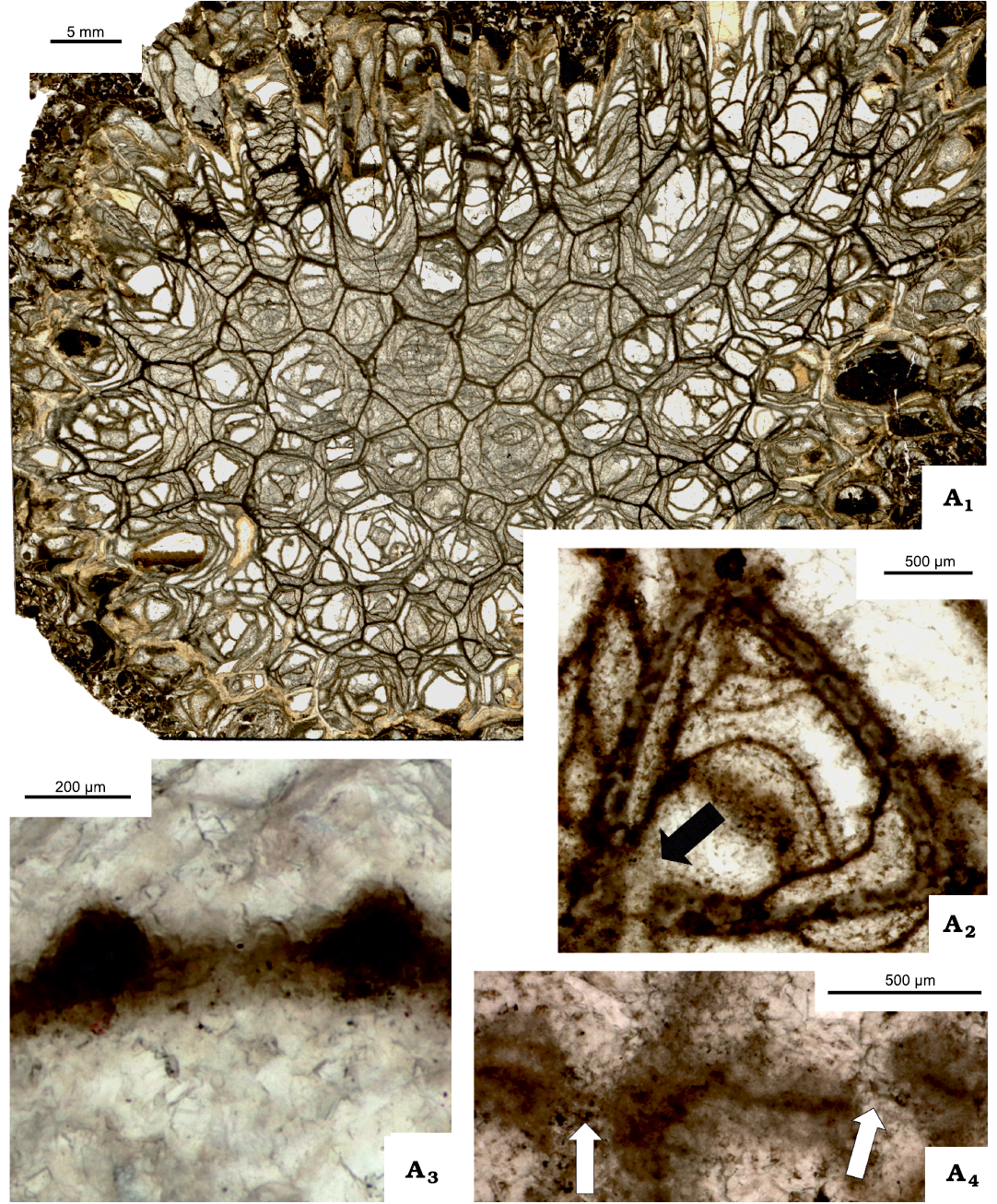
Fig. 7. Tabulate coral Michelinia expansa White, 1883. IG-91-1 = UAM-Tc.Can./2, from locality 3 (Fig. 2), Meilleur Member, Fleet Formation (middle Viséan, Mississipian), Rundle Group, Liard Basin, Northwest Territories, Canada. A1, thin section of colony, transverse in most part and longitudinal at colony edge; A2, immature corallite, note remnants of wall microstructure, arrow points to P1 pore; A3, septal spines attached to tabula; A4, pores P1 and P2 situated close to each other (arrows).
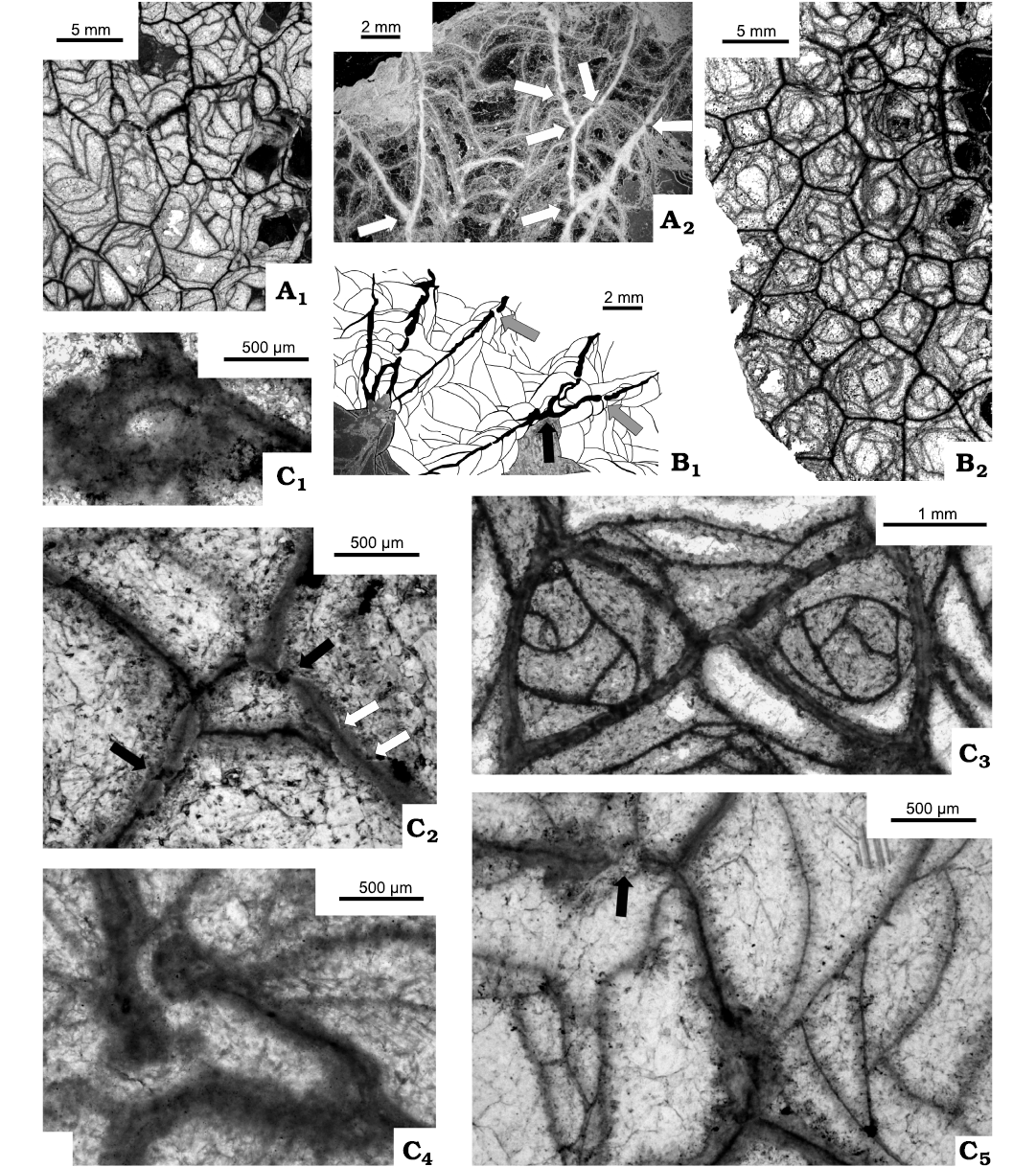
Fig. 8. Tabulate corals Michelinia expansa White, 1883, from locality 3 (Fig. 2), Meilleur Member, Flett Formation (Viséan, Mississipian), Rundle Group, Liard Basin, Northwest Territories, Canada. Transverse thin sections unless stated otherwise. A. IG-91-2 = UAM-Tc.Can./3; A1, fragment of colony; A2, longitudinal section (peel) of offsetting corallite; showing pores (arrows). B. IG-17 = UAM-Tc.Can./1; B1, drawing on peel image of longitudinal section of corallites in different growth stage; showing pores (grey arrows) and apparent absence of pore resulting from eccentric section (black arrow); B2, fragment of a colony. C. IG-91-1 = UAM-Tc.Can./2, different growth stages; C1, very early growth stage of offset surrounded by thick, solid wall; C2, very early growth stage of offset; showing P2 pores (black arrows) and overgrown pores? (white arrows); note differentiated thickness of partitions; C3, two offsets adjacent to one another with first dissepiments developed; C4, very early growth stage of offset with two P1 pores open to two adjacent mature corallites; C5, partitions differentiated in thickness, showing P2 pores (arrow).
Remarks.—Doubts expressed above in the remarks on the genus resulted in both, removal from the synonymy and identification of species based on the macro-morphology. Also, a wide intraspecific variability is here accepted rather than attempting to identify the colonies as different species. Such characters as “thinner walls and fewer and generally more convex tabulae” listed by Sando (1969a: 311) as the only characters distinguishing M. leptosphragma Armstrong, 1962, from M. expansa are here considered inadequate for distinction between species. Thickness of walls may vary (Fig. 8C2, C5). Thus, the synonymy of those two seems very probable. All colonies described here resemble both “Tabellaephyllum peculiaris Stumm, 1948”, (Stumm 1948: pl. 12: 1, 2, 9, 11; Lafuste and Plusquellec 1990: fig. 6) and the type and topotype specimens of Michelinia expansa White, 1883 (White 1883: pl. 39: 2a, b; Sando 1969a: pl. 39: 1–3; Lafuste and Plusquellec 1990: fig. 7a, b). They differ clearly from Michelinia meekiana Girty, 1910 (Girty 1910: 189 only; Sando 1969b: pl. 2: 6–9; Plusquellec and Sando 1987) in possessing short tabulae but dissepiments common, long and steeply sloping down.
Stratigraphic and geographic range.—Viséan (Mississipian, Carboniferous), Flett Formation, Rundle Group, Liard Basin, Northwest Territories, Canada; known also from Shunda Formation, Rundle Group, Liard Basin, Northwest Territories, Canada; Chouteau Limestone, Missouri; Redwall Limestone, Arizona; Lodgepole Formation, Utah; all upper Tournaisian (Mississipian, Carboniferous).
Subclass Rugosa Milne Edwards and Haime, 1850
Order Stauriida Verrill, 1865
Suborder Lithostrotionina Spasskiy and Kachanov, 1971
Family Lithostrotionidae d’Orbigny, 1852
Subfamily Lithostrotioninae d’Orbigny, 1852
Genus Cordilleria nov.
ZooBank LSID: urn:lsid:zoobank.org:act:DA56472A-7C79-4EB9-A8 E7-8CC9B68FFE56
Etymology: Named after the occurrence of the type species in the North American Cordillera.
Type species: Diphyphyllum mutabile Kelly, 1942: 358, pl. 51: 7, 8; “southwest slope of the mountain northwest of the junction of the McLeod River with Whitehorse Creek” (Kelly 1942: 359), lower Rundle Group, upper Tournaisian. Here designated.
Species included: Diphyphyllum mutabile Kelly, 1942; Lithostrotion flexuosum Warren, 1927; Lithostrotion (Siphonodendron) warreni Nelson, 1960; Lithostrotion (Siphonodendron) oculinum Sando, 1963.
Diagnosis.—Fasciculate colonial Lithostrotioninae. Major septa amplexoid, elongated on surfaces of tabulae to various extent. Protosepta indistinguishable from remaining major septa. Cardinal fossula absent. Pseudocolumella formed on upper surfaces of tabulae independently from major septa; may be continuous, or underdeveloped to various extent, up to atrophy. Increase lateral with peripheral parts of parents’ septa inherited by offsets (atavosepta). Acolumellate growth of offsets long-lasting, may be permanent up to maturity. Tabularium normal; down sloping of peripheral parts of tabulae differently accentuated, rarely so strong as to form short-lasting columnotheca. Microstructure of septa finely trabecular.
Remarks.—The holotype of Diphyphyllum mutabile Kelly, 1942, has been the most thoroughly studied and best illustrated of all the North American lithostrotionid specimens and the species itself has been formerly included to three different genera: Diphyphyllum Lonsdale, 1845, Lithostrotion Fleming, 1828, and Siphonodendron McCoy, 1849 (e.g., White 1875; Warren 1927; Kelly 1942; Nelson 1960; Bamber 1966; Sando and Bamber 1985; Fedorowski and Bamber 2007). Diphyphyllum mutabile is designated here as the type species for Cordilleria gen. nov. Most of the summary by Fedorowski and Bamber (2007), based mainly on the holotype of D. mutabile, is followed here, allowing a considerable reduction of the remarks that follow. Only the idea of the ancestral position of the North American species included in European genus Dorlodotia Salée, 1910, by Sando (1969a), Sando and Bamber (1985) and Fedorowski and Bamber (2007), is not followed here. All those North American Dorlodotia-like species are here considered as endemics, belonging to a new, yet unnamed genus, closely related to Cordilleria gen. nov. Fedorowski and Bamber (2007) postulated a position of that unnamed new genus as ancestral for the North American Siphonodendron-like corals. We follow that idea for two reasons: (i) the early growth stages of both “D. mutabile Kelly, 1942”, and “Dorlodotia sp.” (Fedorowski and Bamber 2007: pl. 2: 3 and 7, respectively) are nearly identical; (ii) the occurrence of the oldest “Dorlodotia sp.”-like species preceded the appearance of the oldest Cordilleria gen. nov., i.e., “D. mutabile”.
The collection from the Flett Formation in the Liard Basin is too geographically and stratigraphically restricted for a more comprehensive discussion on the content and the inter-relationships of the species possibly belonging to Cordilleria gen. nov. A large collection derived from more than 200 localities in western Canada and studied by the authors simultaneously, documents very large morphological variability in all characters of colonies potentially belonging to that genus. Limits of the intraspecific variability and emended diagnoses of these species are impossible to establish using only the Liard Basin colonies. They will be discussed in detail and introduced in the study of the large collection mentioned above. Thus, the total species content of the genus Cordilleria gen. nov. is not established here and descriptions of species that follow are reduced to characters typical for the specimens from the Liard Basin collection. For the reasons mentioned above, all species from the Liard Basin are here temporarily identified as affinis to the nominative species. Such an approach is safer than the application of names when the limits of intraspecific variability of those species have not been established. Brief characteristics of the most important features of the holotypes of species present in the Liard Basin are introduced for comparison. Those characteristics are based on the papers by Kelly (1942), Nelson (1960), Sando (1963), Bamber (1966), Sando and Bamber (1985), and Fedorowski and Bamber (2007). A brief discussion below on some morphological characters of corals identified here as Cordilleria gen. nov. is introduced to illustrate their variability and bearing on species identifications.
(i) The characteristics of major septa. All specimens considered by us as belonging to Cordilleria gen. nov. have amplexoid major septa. That characteristic, not named in descriptions published so far, is differently accentuated in particular species. Its bearing on the length of major septa has been recognized by several authors dealing with those corals (e.g., Sutherland 1958; Nelson 1960; Sando 1963; Bamber 1966). To that question Bamber (1966: 8) devoted a good deal of his remarks on “Lithostrotion (Siphonodendron) sinuosum (Kelly, 1942)”, i.e., Cordilleria flexuosa (Warren, 1927) in the name accepted here. The amplexoid character of the major septa and the consistent length of the non-amplexoid minor septa, led to wide limits being accepted for the major/minor septa relationship in all detailed descriptions of the species in the papers listed above. We consider that mutual relationship as diagnostically important, but with the restriction that minor septa are compared to the parts of major septa consistent in their length. Only parts of the amplexoid major septa, i.e., those located between external walls or dissepimentarium on the one side and steep parts of tabulae on the other side, fulfill that criterion. Parts of major septa lengthened to various extent on surfaces of inner parts of tabulae, cannot be used for comparison since their length depends on the advancement of their amplexoid character. They are only slightly elongated when weakly amplexoid as exemplified by C. warreni (Nelson, 1960) or are elongated along surfaces of tabulae to approach the pseudocolumella when strongly amplexoid as in C. flexuosum. That difference should be omitted when the major/minor septal relationships are compared. Thus, we suggest considering only the non-lengthened sectors of major septa as their referring length.
(ii) The morphology in longitudinal sections and the shape of tabulae in particular, has been accepted by Sando (1963: 1076) as the most important feature distinguishing his new species “Lithostrotion (Siphonodendron) oculinum” from the most similar species “L. (S.) mutabile” and “L. (S.) warreni”. Also, Bamber (1966) considered the morphology in longitudinal section as one of the most important characters allowing the distinction between extremely variable species traditionally included by the North American authors in Lithostrotion (Siphonodendron). Unfortunately, the strong variability in the development of pseudocolumellae within corallites of particular colonies, varying from almost continuous to non-existent, strongly reflects on the shapes of tabulae, making them similar in different species when the pseudocolumella is absent. Diagenetic alterations, in turn, camouflage the distinction of steep, peripheral parts of tabulae from longitudinal sections of peripheral major septa that can imitate steep parts of tabulae resting upon underlying tabulae. All those taxonomically inconvenient features can be eliminated by thorough investigation of perfectly centric longitudinal sections. Nevertheless, they increase the difficulty in applying the characters of longitudinal sections as diagnostic. It is enough to compare the longitudinal sections of the holotype of “L. (S.) oculinum” (Sando 1963: pl. 145: 4, 5), or the longitudinal sections illustrated by Bamber (1966: pl. 1: 4c–i) to demonstrate those difficulties. The illustrated specimens (Figs. 9A2, A3, B1, B2, 11A2, A3, B1, 13D) may serve as additional examples. Nevertheless, the morphology in longitudinal section is diagnostically important, but it should be treated in a way less rigid than suggested by Nelson (1960), Sando (1963), and Bamber (1966). This means that the prevailing morphology of several characters rather than a strong single feature should be accepted as diagnostic for a given species.
(iii) The presence or absence of a pseudocolumella is one of the most important characteristics for distinguishing between the European Diphyphyllum Lonsdale, 1845, and Siphonodendron McCoy, 1849. Following that European distinction, Kelly (1942) followed by Sutherland (1958) and Sando and Bamber (1985) applied the name Diphyphyllum to the lithostrotionids from North America that lack a pseudocolumella or have it extensively reduced. Fedorowski and Bamber (2007) rejected that name by informally using the name Siphonodendron for “Diphyphyllum mutabile Kelly, 1942”. However, the question arises what to do with the North American colonies that totally lack pseudocolumella, but expose all other characters resembling particular species of Cordilleria gen. nov. We now provisionally accept such colonies as belonging to the latter genus.
(iv) The dissepimentarium in most North American lithostrotionids described so far can be characterized as consistent for species. Two main groups can be distinguished: those with a single row of interseptal and mostly regular dissepiments and others developing two or more rows of dissepiments with or without lonsdaleoid dissepiments developed in addition to interseptal ones. However, two or even three rows of dissepiments may for a very short period of growth appear in specimens characterized otherwise by a single, incomplete row of interseptal dissepiments (Fig. 9A1). We provisionally consider such a multiplied development as a diagnostically unimportant and environmentally controlled event and include its bearers within species overwhelmingly developing only a single complete or incomplete row of interseptal dissepiments.
The name Lithostrotion flexuosum, introduced by Warren (1927) for a species included by him in the genus Lithostrotion Fleming, 1828, was renamed Lithostrotion sinuousum by Kelly (1942) due to the earlier preoccupation of the former by Lithostrotion flexuosum Trautschold, 1879. However, that pre-occupation failed when specimens of flexuosum–sinuosum were included here in Cordilleria gen. nov. Thus, the name L. flexuosum Warren, 1927, is here re-introduced as valid and is applied in this paper instead of L. sinuousum Kelly, 1942.
Cordilleria mutabile (Kelly, 1942)
1942 Diphyphyllum mutabile Kelly, 1942: 358, pl. 51: 7, 8.
1966 Lithostrotion mutabile Kelly, 1942; Bamber 1966: 4, pl. 1: 1a–e.
2007 “Diphyphyllum” mutabile Kelly, 1942; Fedorowski and Bamber 2007: pl. 2: 1–6, 10–12; pl. 7: 1.
Holotype: GSC 9642.
Type locality: “Southwest slope of the mountain northwest of the junction of the McLeod River with Whitehorse Creek” (Kelly 1942: 359).
Type horizon: Lower Rundle Group, upper Tournaisian.
Emended diagnosis.—Fasciculate colonial coral with n:d value of 20:3.9 mm to 25:6.0 mm (Fig. 10). Major septa weakly amplexoid, reach up to 1/2 corallite radius on tabulae surfaces, reduced to 1/3–1/4 corallite radius when sectioned between tabulae, i.e., terminated on steep, peripheral parts of tabulae. Protosepta indistinguishable. Cardinal fossula absent. Minor septa almost as long as major septa when latter not elongated. Pseudocolumella short, simple, slightly thickened or thin, rarely underdeveloped. Tabulae complete, slope peripherally under 10–20° when pseudocolumella present, elevated slightly upward next to pseudocolumella. Tabulae horizontal, slightly convex or concave when pseudocolumella absent; both kinds strongly deflected at periphery, rare inner parts of tabulae rest upon underlying tabulae, most meet dissepimentarium or external wall under a variable angle. In immature specimens, tabulae span entire corallite lumen, slightly convex or horizontal with weakly marked peripheral deflection, or straight. Dissepimentarium in one incomplete row of regular, interseptal dissepiments. Microstructure of septa finely trabecular.
Remarks.—The synonymy is restricted to the papers discussing with the holotype. The emended diagnosis, comprising all main characteristics of skeleton of the holotype, is based on description and illustrations by Kelly (1942: pl. 51: 7, 8), restudy of the holotype by Bamber (1966: pl. 1: 1a–e) and on the further restudy of the holotype by Fedorowski and Bamber (2007: pl. 1: 1–11, pl. 2: 1–6, 10–12).
Stratigraphic and geographic range.—Type horizon and locality only.
Cordilleria aff. mutabile (Kelly, 1942)
Figs. 9–11.
Material.—Three fragments of colonies (Fig. 6): C-47930 = GSC 142469, C-52546 = GSC 142470, and C-52124 = GSC 142471 from the Flett Formation, Rundle Group, Viséan, Liard Basin, Northwest Territories, Canada and colony C-7349 = GSC 142468 from Rundle Group, Viséan, Rocky Mountains, Bone Mountain, east-central British Columbia, Canada, all comparable in age (Table 1), included in this description as bearing morphological features important for the discussion on this species and the new genus Cordilleria. All colonies recrystallized.
Description.—N:d values of all colonies vary, some comparable to those of holotype (Fig. 10). However, increasing number of septa does not always follow increase of corallite diameter in all colonies, holotype included. Corallites in GSC 142469 display smallest number of septa; largest of them equal in septal number to smallest corallites of holotype. In contrast, corallites in GSC 142470 display number of septa comparable to holotype, but diameters larger. Thus, n:d values of those two colonies differ from that in holotype and remaining specimens (Fig. 10). Also, they differ from one another in morphology (Figs. 9A3, B4 and 11A1, B2, respectively) suggesting limits of intraspecific variability provisionally accepted here for C. mutabile are too broad.
Amplexoid character of major septa differentiated. Most advanced in GSC 142469 (Fig. 9B4), where some major septa reach more than 2/3 corallite radius. In GSC 142471 and GSC 142470 (Figs. 9A3 and 11B2, respectively), amplexoid character of major septa hardly recognizable. Longest major septa in corallites of those colonies reach only slightly more than 1/3 corallite radius. In all specimens studied, major septa thin and protosepta indistinguishable irrespective of presence or absence of pseudocolumella. Minor septa longest in GSC 142469 (Fig. 9B4), closely comparable to those in holotype. In remaining specimens, minor septa vary in length both between corallites and during growth of individual corallites; longest reach 2/3 length of major septa or slightly more. Those reaching 1/2–2/3 length of major septa prevail, but slightly shorter ones occur. Occurrence of pseudocolumellae strongly differentiated; present as a slightly thickened body in all corallites observed in transverse section of colony GSC 142469, but not in fragments of longitudinal sections of several corallites within that colony (Fig. 9B4 and B1–B3, respectively). Pseudocolumella absent from all corallites in two other colonies studied from the Flett Formation (Figs. 9A, 11B2). Differences in longitudinal section depend mainly on development of pseudocolumella. Closely resembling each other, including holotype (Fedorowski and Bamber 2007: pl. 2: 5, 6, 10), when pseudocolumella not developed (Figs. 9A1, A2, B1 (white arrow), B2, B3, 11B1). In columellate part of longitudinal section of GSC 142469 (Fig. 9B1, black arrow), inner tabulae elevated steeply towards thickened pseudocolumella. Many of them rest upon underlying tabulae to form short lasting columnotheca. Columnotheca rarely occurs in holotype. Also, inner parts of its tabulae elevated much less steeply than tabulae in the illustrated GSC 142469. Colony GSC 142468 additionally included in this paper, (Fig. 11A) displays morphology intermediate between GSC 142469 and holotype. This especially concerns the development vs. absence of pseudocolumella. Pseudocolumella in colony GSC 142468 strongly interrupted, making acolumellate colonies from the Liard Basin an extreme variant of a sequence starting with colonies with pseudocolumella almost invariably continuous in all corallites to colonies composed of acolumellate corallites. Also, it documents secretion of pseudocolumella by epidermis of polyp’s floor (Fig. 11A2, A4), discussed in more detail in general remarks. Dissepimentarium mostly in one incomplete row of rectangular, interseptal dissepiments, i.e., like in holotype, but in colony GSC 142471 may occasionally increase to 2–3 rows in small parts of corallite and during short period of the corallite growth (Fig. 9A1). Offsetting (Fig. 11C, D) documented by offsets in various growth stages appearing by chance in transverse thin sections. Their morphology follows particular growth stages illustrated by Fedorowski and Bamber (2007: pl. 1: 1–11) in holotype.
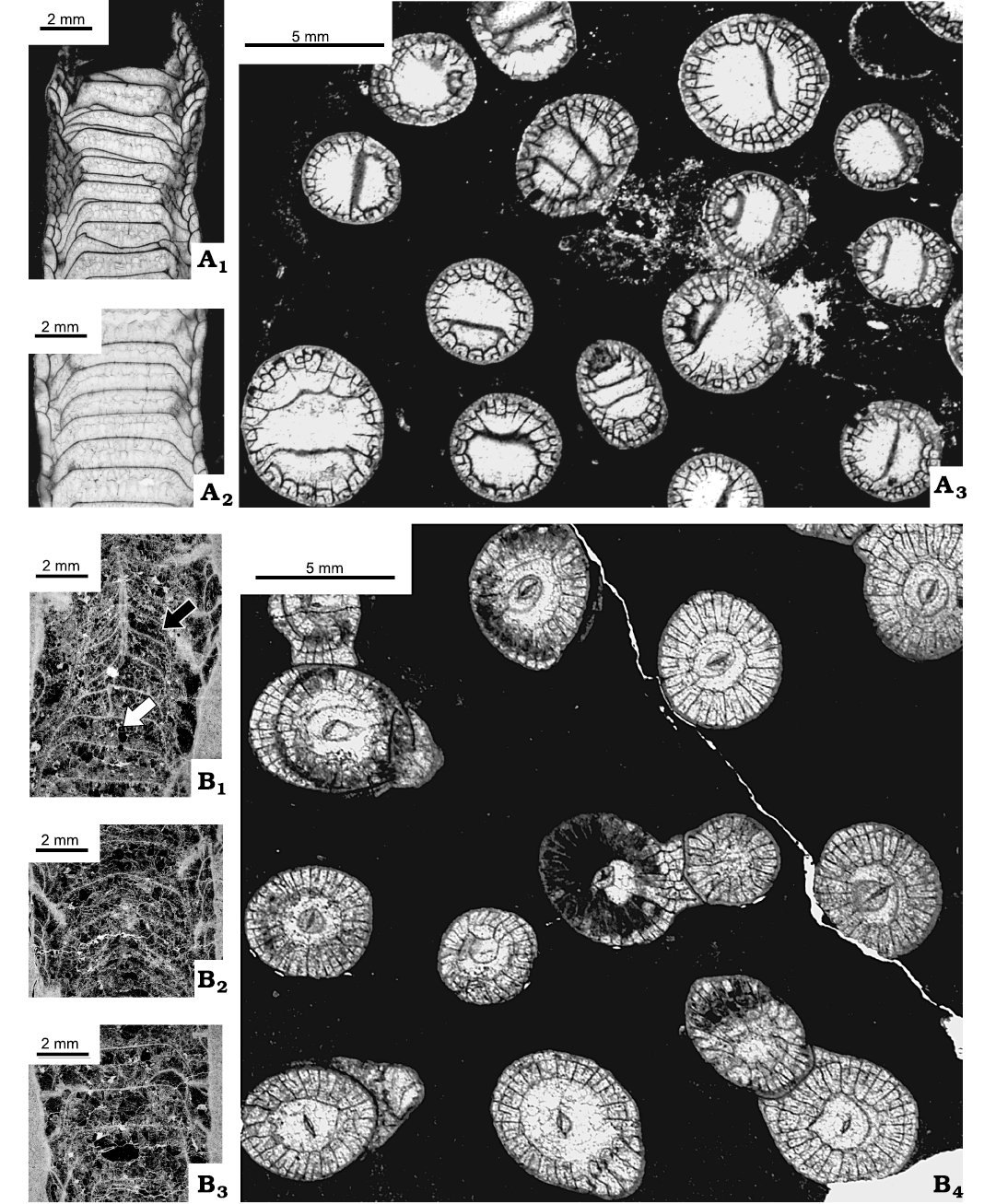
Fig. 9. Rugose corals Cordilleria aff. mutabile (Kelly, 1942), from Flett Formation (Mississipian), Rundle Group, Liard Basin, Northwest Territories, Canada. A. C-52124 = GSC142471, locality 3 (Fig. 2); longitudinal thin sections (A1, A2), transverse thin section (A3). B. C-47930 = GSC-142469, locality 14 (Fig. 2), Flett Formation undivided; longitudinal sections, peels (B1–B3), transverse thin section (B4). Note in B1 steeply arranged tabulae when pseudocolumella present (black arrow), flat tabulae when pseudocolumella absent (white arrow).
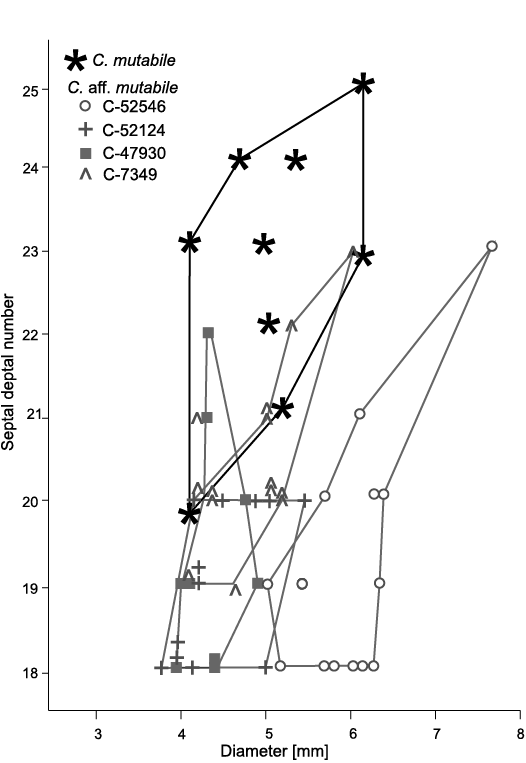
Fig. 10. Variation in septal number and corallite diameters of rugose coral Cordilleria aff. mutabile (Kelly, 1942) and the holotype colony of Cordilleria mutabile (Kelly, 1942) added for comparison. Large differences in that character between colonies comparable to that noted by Sando (1963: fig. 1) in “Lithostrotion (Siphonodendron) oculinum” (Sando, 1963) = Cordilleria oculina (Sando, 1963) (see Fig. 12).
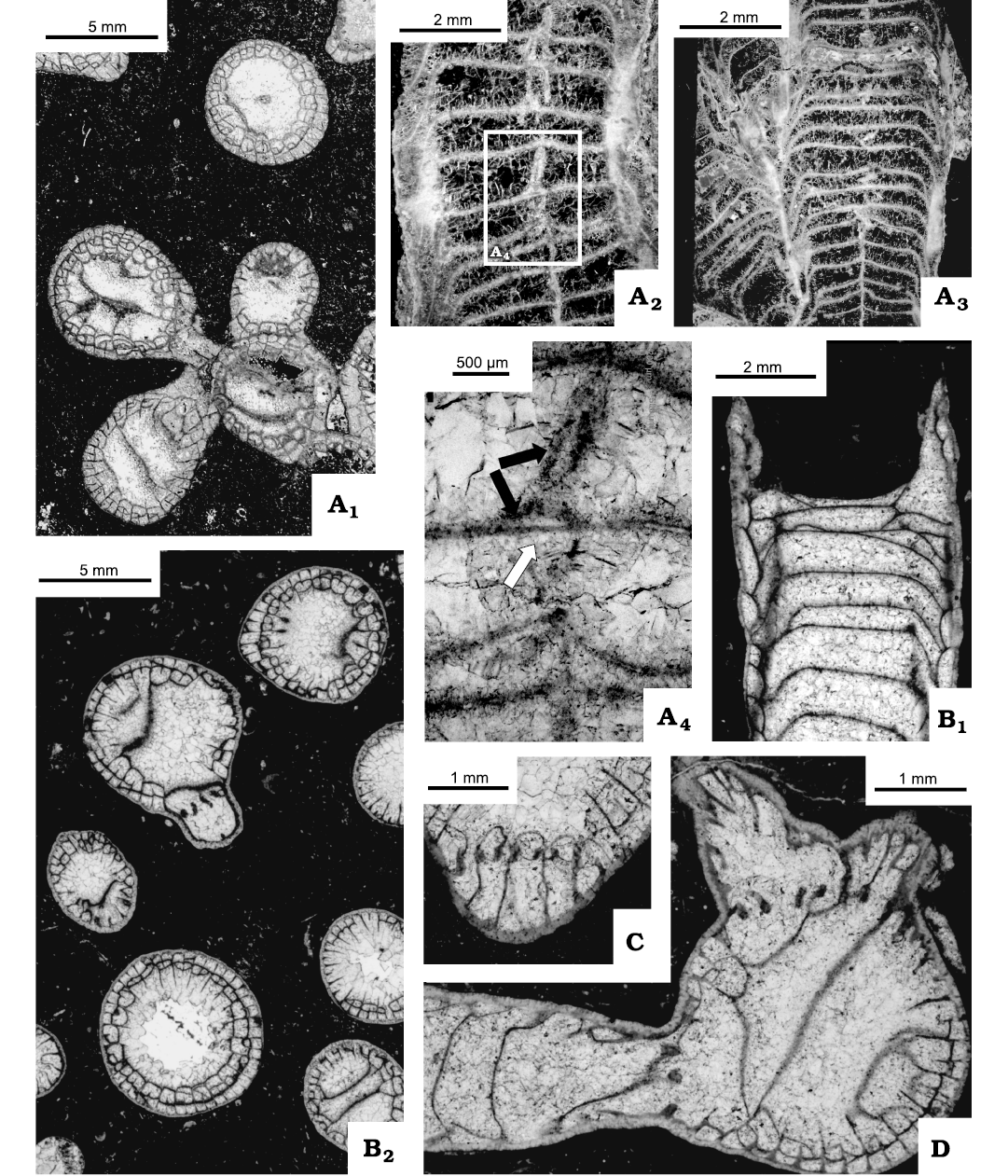
Fig. 11. Rugose corals Cordilleria aff. mutabile (Kelly, 1942), from Rundle Group undivided, east-central British Columbia, Canada (A) and Meilleur Member, Flett Formation (Mississipian), Rundle Group, Liard Basin, Northwest Territories, Canada (B–D). A. C-07349 = GSC 142468, 8.05 km NW of Bone Mountain, east-central British Columbia; transverse thin section (A1), longitudinal thin sections (A2, A3), pseudocolumella (A4, black arrows) created on tabula surface (white arrow) and tabulae/pseudocolumella relationships in lower part of picture. B. C-52546 = GSC 142470, locality 3 (Fig. 2); longitudinal thin section (B1), transverse thin section (B2). C. C-47930 = GSC 142469, locality 2 (Fig. 2); transverse thin section; very early growth stage of offset. D. C-52124 = GSC 142471, locality 3 (Fig. 2); thin section of three offsets in different growth stage; one cut longitudinally, two transversally.
Remarks.—The morphological variability in almost all characters of specimens described from the Flett Formation extends beyond the parameters established above for the holotype. The small n:d value of GSC 142469 and its rather strongly amplexoid major septa resemble those in C. flexuosum (Warren, 1927), whereas its very long minor septa and its morphology in longitudinal section distinguishes it clearly from that species. The morphology in transverse sections of the other colonies and the slightly reduced length of some of their minor septa resemble C. oculinum (Sando, 1963), whereas the great length of most of the minor septa, the weakly marked amplexoid character of the major septa and the morphology in longitudinal sections form a set of characters distinguishing them from the latter species. All those characters, the long minor septa and the morphology in the longitudinal sections allow all the colonies discussed to be distinguished from C. warreni (Nelson, 1960).
Cordilleria oculina (Sando, 1963)
1963 Lithostrotion (Siphonodendron) oculinum sp. nov.: Sando 1963: 1075, pl. 145: 2–5 only.
1969 Lithostrotion (Siphonodendron) oculinum Sando; Sando 1969a: 309, pl. 35: 2–5.
1985 Lithostrotion (Siphonodendron) oculinum Sando; Sando and Bamber 1985: pl. 5: 8, 9.
Type material: USNM 143243.
Type locality: Redwall between Bridge Canyon and the Peach Springs-Nelson area, south of the Grans Canyon (Sando 1963: 1074).
Type horizon: Redwall Limestone, C Member, Kinderhookian Series, upper Tournaisian. Mississipian, Carboniferous.
Emended diagnosis.—Fasciculate colonial coral with maximum n:d value of 23–25:5.6 mm and most common n:d value of 20:4.5–5.2 mm and 21:4.5–4.8 mm (Fig. 12). Major septa amplexoid, on tabula surfaces reach up to 2/3 corallite radius, reduced to 1/3–1/2 corallite radius when sectioned between tabulae, i.e., terminated at steep, peripheral parts of tabulae. Minor septa up to 1/2 length of majors when latter not elongated, commonly enter peripheral part of tabularium. Pseudocolumella short, thin, commonly interrupted. Tabulae slope from pseudocolumella at angle of 20°–40° in long inner parts, trapezoid to slightly convex when pseudocolumella absent, sharply deflected downward at periphery; most horizontal near dissepimentarium; some rest upon underlying tabulae. Dissepimentarium in one, incomplete row of interseptal dissepiments
Remarks.—The synonymy is restricted to the papers dealing with the holotype. The emended diagnosis, comprising all main characteristics of skeleton of the holotype, is based on Sando 1963: 1075, 1076, pl. 145: 2–5.
Stratigraphic and geographic range.—Type locality and horizon only.
Cordilleria aff. oculinum (Sando, 1963)
Figs. 12, 13.
Material.—Five fragments of colonies from Flett Formation, Rundle Group, Liard Basin, Northwest Territories, Canada: IG-16 = UAM-Tc./Can.5, IG-30 = UAM-Tc./Can.7, Tlogotsho Member, lower Viséan; IG-85 = UAM-Tc./Can.6, Meilleur Member, middle Viséan (Fig. 6); C-52268 = GSC 142473, C-52361 = GSC 142474, Meilleur Member, middle Visean; and C-52205 = GSC 142472 from Prophet Formation, Rundle Group, Liard Basin, Northwest Territories, Canada. All colonies diagenetically altered by recrystallization and dolomitization that destroyed microstructure of septa in all corallites.
Description.—Intra- and inter-colony variation advanced in both n:d values (Fig. 12) and in morphological characteristics (Fig. 13). Amplexoid character of major septa weakly accentuated, resulting in their comparatively small length, not exceeding 1/2 corallite radius, commonly less. All major septa thin, equal in length. Cardinal fossula absent. Minor septa differentiated in length from hardly recognizable projections on surface of external wall to well developed, penetrating outermost tabularium. Pseudocolumellae underdeveloped to various extent, short and thin when present; very rare in most colonies (Fig. 13A1, E, F), probably absent from some corallites (Fig. 13A2). In acolumellate parts of corallites tabulae complete, with flat sectors extending through major part of corallite diameter, sloping steeply down in their short peripheral sectors to meet external walls or dissepimentarium (Fig. 13A2, B1, D1, white arrow). In columellate corallite growth intervals, tabulae elevated towards pseudocolumella under various angles, but not steeply (Fig. 13C, D1, black arrow). Dissepiments in one row, may be absent from great distances of corallite growth (Fig. 13A2).
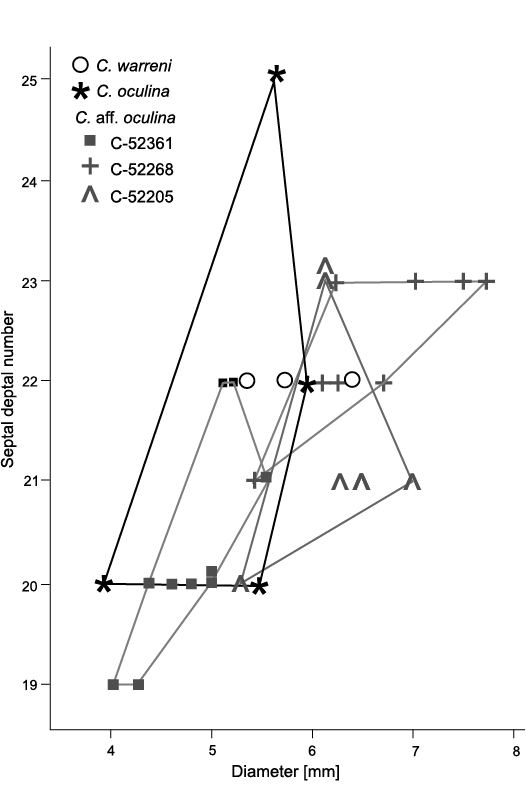
Fig. 12. Variation in septal number and corallite diameters in colonies of rugose coral Cordilleria aff. oculinum (Sando, 1963) and holotypes of Cordilleria oculina (Sando, 1963) and Cordilleria warreni (Nelson, 1960) added for comparison. Data for C. oculina after Sando (1963: part of fig. 1); for C. warreni after Nelson (1960: pl. 24: 11).
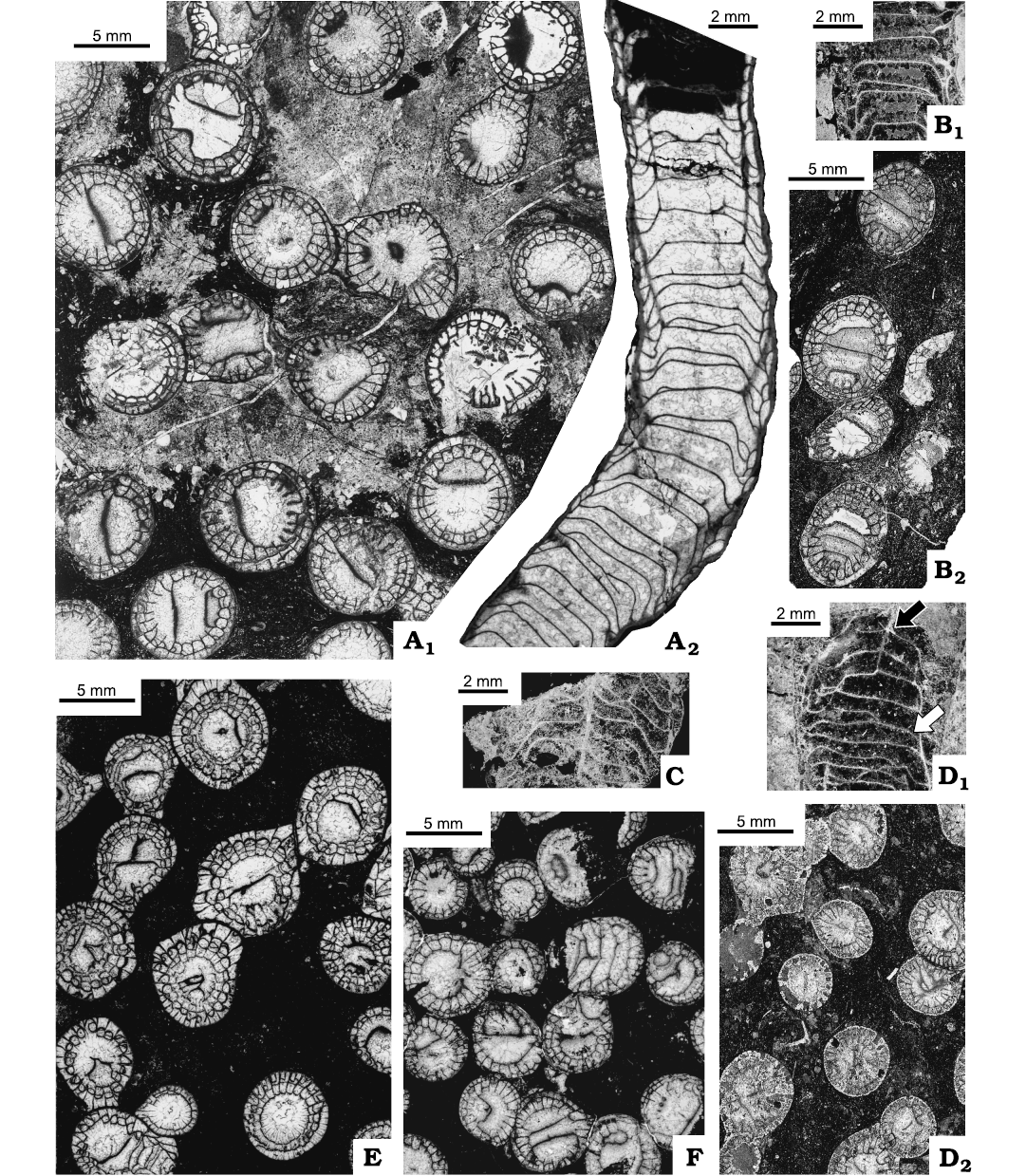
Fig. 13. Rugose coral Cordilleria aff. oculinum (Sando, 1963), from Meilleur Member, Flett Formation (Mississipian), Rundle Group, Liard Basin, Northwest Territories, Canada (A, C, F), Tlogotsho Member, Tlett Formation (Mississippian), Rundle Group, Liard Basin, Northwest Territories, Canada (B, D), and upper Prophet Formation (Mississippian), Roundle Group, Northwest Territories, Canada (E). A. C-52268 = GSC-142473, locality 2 (Fig. 2); transverse thin section (A1), longitudinal thin section (A2). B. IG-16 = UAM-Tc.Can./5 B-D, locality 3 (Fig. 2); longitudinal section, peel (B1), transverse thin section (B2). C. IG-85 = UAM-Tc.Can./6, locality 3 (Fig. 2); longitudinal section (peel). D. IG-30 = UAM-Tc.Can./7, locality 3 (Fig. 2); longitudinal section, peel (D1), transverse thin section (D2). E. C-52205 = GSC-142472, locality 2 (Fig. 2); transverse thin section. F. C-52361 = GSC-142474, locality 13 (Fig. 2); transverse thin section.
Remarks.—Sando (1963: fig. 1) suggested wide intraspecific variability in n:d value of this species. However, a great majority of his specimens are located within narrow limits similar to the intra-colony variation in the holotype. Small specimens are immature (omitted here from Fig. 12). Large corallites are very rare. Most specimens here identified as C. aff. oculinum resemble the type collection in the number of septa present, but differ in larger corallite diameter, i.e., their n:d value differ from that in both the holotype and the type collection. The amplexoid character of the major septa, weakly accentuated in the specimens described here, differs from that in the type collection. That amplexoid weakness results in the absence of short projections of major septa on surfaces of tabulae, apparently characteristic for the holotype. Very short minor septa in some colonies are characteristic for C. warreni rather than C. oculinum, whereas the morphology in longitudinal section in those colonies points towards the latter.
Cordilleria sp.
Figs. 14, 15.
Material.—Four fragments of colonies from Flett Formation, Rundle Group, Liard Basin, Northwest Territories, Canada: IG-18 = UAM-Tc.Can./10, middle part of the Tlogotsho Member, lower Viséan; IG-82 = UAM-Tc.Can./8, IG-83 = UAM-Tc.Can./9, lower part of the Meilleur Member, middle Viséan (Fig. 6); C-47942 = GSC 142475, Flett Formation undivided, Viséan. All specimens recrystallized and/or dolomitized; some strongly (Fig. 15A1, B). Thin sections and peels available for the study.
Description.—Diameters and numbers of septa in all colonies similar in most corallites measured with only rare corallites of colony UAM-Tc.Can./8 larger (Fig. 14). Major septa thin, vary in length in particular colonies; shortest and weakly amplexoid in colony UAM-Tc.Can./10, reach 2/3 corallite radius at most (Fig. 15C3), longest and almost non-amplexoid in colony GSC 142475 (Fig. 15B1, B2), almost approach corallite axis in some corallites. Length of major septa in remaining two colonies (Fig. 15A1, D) intermediate between those two. In all colonies, minor septa penetrate tabularium; in colony GSC 142475, longest, extending up to 4/5 length of major septa. Dissepimentarium complete in most corallites, comprises 2–4 rows of interseptal, rectangular dissepiments. Only in colony UAM-Tc.Can./10 dissepimentarium incomplete in some fragments of corallites, but locally up to 3 rows of dissepiments. Pseudocolumella thin and short, when present; in colony UAM-Tc.Can./9 (Fig. 15D) not traced. In remaining colonies, interrupted to various extent, absent from long interval of corallite growth in some (Fig. 15A2). Morphology in longitudinal section depends only in part on presence/absence of pseudocolumella. Shape of tabulae most irregular in colony UAM-Tc.Can./10 irrespective of presence/absence of pseudocolumella (Fig. 15C1, C2). Also, down sloping of peripheral parts of tabulae in that corallite very weakly accentuated. This character most distinctly accentuated in acolumellate part of corallites of colony UAM-Tc.Can./8 (Fig. 15A2, A3, arrow), but weak in its columellate part. Same corallite demonstrates differentiated deflection of tabulae from pseudocolumella and their variable density in parts of growth. Deflection of tabulae steepest in colony GSC 142475 (Fig. 15B3).
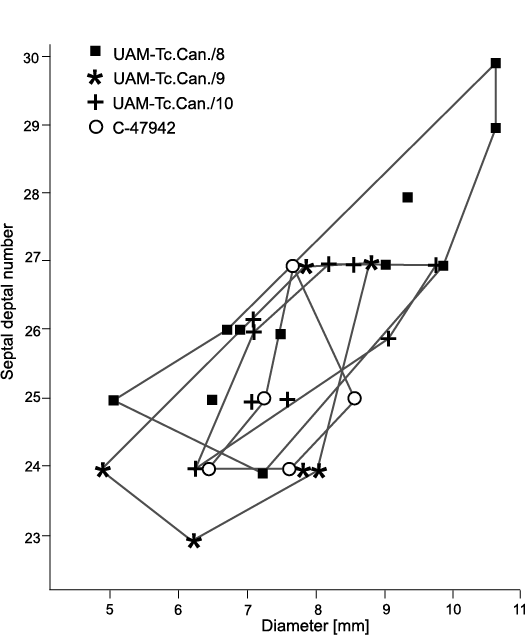
Fig. 14. Variation in septal number and corallite diameters of rugose coral Cordilleria sp.
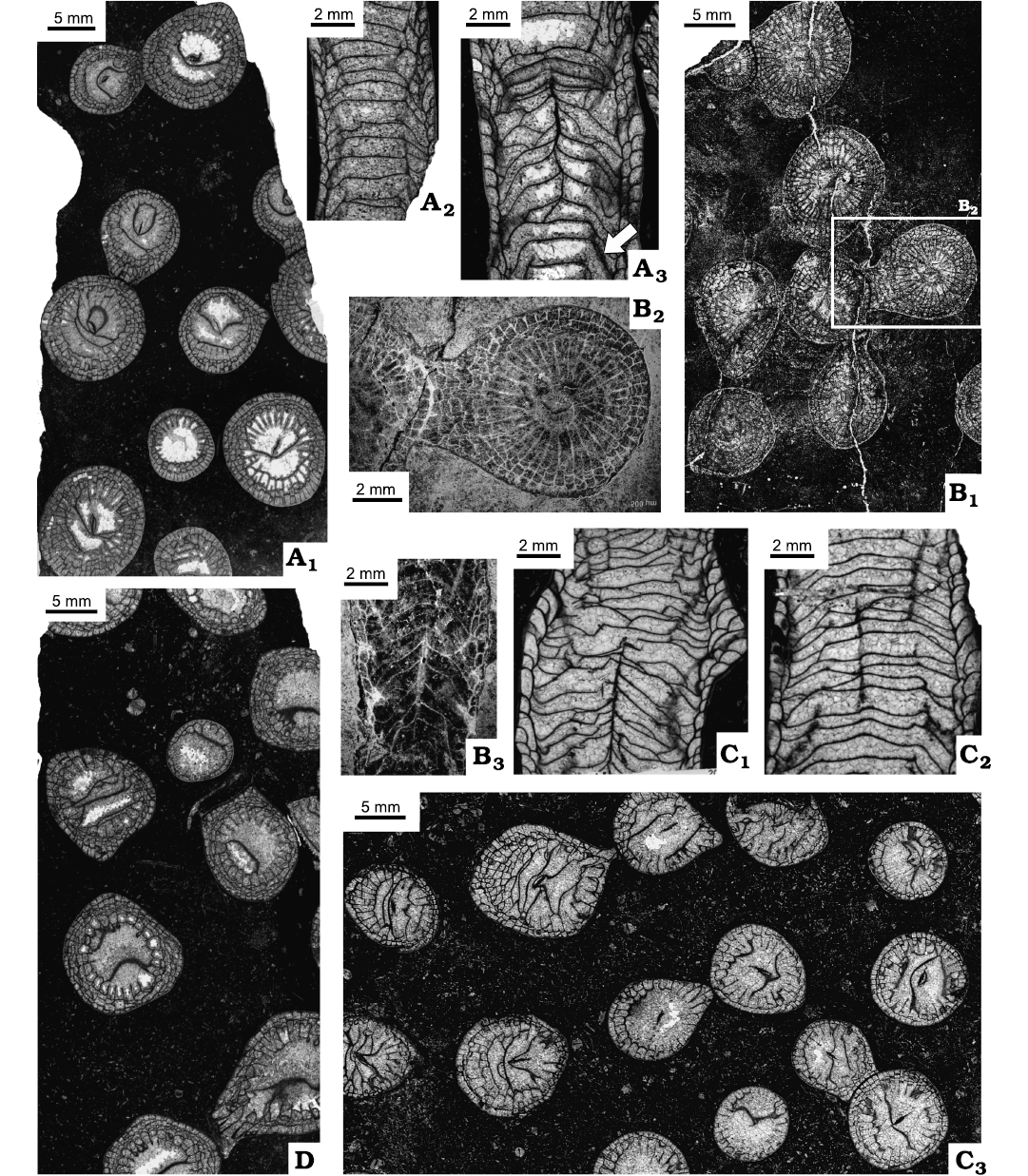
Fig. 15. Rugose corals Cordilleria sp. A. IG-82 = UAM-Tc.Can./8, locality 3 (Fig. 2), from Meilleur Member, Flett Formation (Mississipian), Rundle Group, Liard Basin, Northwest Territories, Northwest Territories, Canada; transverse thin section (A1); longitudinal thin sections (A2, A3), weakly accentuated down sloping of peripheral part of tabulae (arrow). B. C-47942 = GSC-142475, locality 14 (Fig. 2), from Flett Formation undivided, Rundle Group, Liard Basin, Northwest Territories, British Columbia, Canada; transverse section (B1, B2), longitudinal section, peel (B3). C. IG-18 = UAM-Tc.Can./10, locality 3 (Fig. 2), from Tlogotsho Member, Flett Formation (Mississipian), Rundle Group, Liard Basin, Northwest Territories, British Columbia, Canada; longitudinal thin sections (C1, C2), transverse thin section (C3). D. IG-83 = UAM-Tc.Can./9, locality 3 (Fig. 2), from Meilleur Member, Flett Formation (Mississipian), Rundle Group, Liard Basin, Northwest Territories, Canada; transverse thin section.
Remarks.—Colonies described as Cordilleria sp. differ from all the other specimens of Cordilleria gen. nov. described above, i.e., C. mutabile, C. aff. mutabile, C.oculinum and C. aff. oculinum by having larger numbers of septa and corallite diameters. Three of those colonies (Fig. 15A1, B, D) differ from them in possessing wider dissepimentaria, comprising 2–4 rows of interseptal dissepiments. The dissepimentarium in corallites of colony UAM-Tc.Can./10, comprising 1–3 rows of dissepiments in most, absent from some part of corallites, is intermediate between three colonies described here and the other species of Cordilleria gen. nov. described above. Other characters, except for those in some longitudinal sections (e.g., Fig. 15B3, C1, C2) are comparable. All four colonies resemble specimens of “Lithostrotion whitneyi Meek, 1877” of Nelson (1960: pl. 25: 1–4), who pointed out that “a complete redescription of the primary types or topotypes is desirable” (Nelson 1960: 124).
Sando (1965) made such a revision starting from a discussion on the authorship of “Lithostrotion whitneyi”. According to him, White’s authorship has priority over the paper by Meek (1877) due to White’s (1875) preprint of his report, published eventually in White (1877). We accept Sando’s reasoning in that matter, but we do not follow Sando’s (1965: E15) identification of that species as belonging to Orygmophyllum Fomichev, 1953. The latter genus has nothing in common with the colony of “Lithostrotion whitneyi”, catalogue number USNM 8480, selected by Sando (1965: E15, pl. 5: 1–9) as the lectotype of that species. First of all, the Ukrainian genus has a solitary growth form. Secondly, its morphology (Fomichev 1953: pl. 18: 2–14; pl. 19: 1, 2), discussed recently by Fedorowski (2019), differs considerably from that in “L. whitneyi”, pointing towards the family Neokoninckophyllidae Fomichev, 1953, as Fedorowski (2019) documented. Sando (1965) did not study the offsetting and the septal microstructure in the lectotype and we did not restudy that colony. However, its morphology closely resembles that of Heintzella Fedorowski, 1967, into which it should probably be transferred. With all the mistakes and doubts, the colonies resembling those identified as Cordilleria sp. require a new species name supported by a thoroughly studied and well documented holotype. None of the colonies studied here can serve as such a type. Besides, the restricted number of specimens are inadequate for establishing the intraspecific variation of such a new species. It may be considerable as suggested by the variability of the colonies described here. Thus, the introduction of a new species is not proposed.
Genus Stelechophyllum Tolmachev, 1933
Remarks.—The diagnosis of this genus emended by Sando (1983: 9) is too detailed in such characters as two orders of septa, common for the entire suborder and incomplete in such an important character as the microstructure of septa. We accepted it here on the condition of being temporary. The same is true for the geographical distribution and the species content of that genus suggested by Sando (1983). Unfortunately, the single colony available for this study cannot form the basis for a comprehensive discussion.
Stelechophyllum cf. banffense (Warren, 1927)
Fig. 16.
Material.—Fragment of cerioid colony C-52434 = GSC 142476, ca. 90×70×80 mm in size from Flett Formation undivided, Viséan, Rundle Group, Liard Basin, Northwest Territories, Canada. Some corallites strongly re-crystallized (see below).
Description.—Maximum number of septa measured—18 with n:d value 18:7.8×6.1 mm. Most common number of septa is 17 with corallite diameters 7.1×6.1 mm, 9.0×6.0 mm. Morphology of corallites varies greatly as a result of diagenetic alteration. Most altered corallites form a belt surrounding less altered part of the colony (Fig. 16, arrows). Boundary between those two fragments crosses several corallites as indicated by their morphology. Parts of corallites adjacent to less altered sectors of colony closely resemble corallites from that sector, whereas their opposite parts look like corallites from the diagenetically altered sector. In parts of colony showing least diagenetic alteration, major septa thin, long, approaching and some meeting thin pseudocolumella, commonly connected to both protosepta. Minor septa thin, in full number, all enter tabularium. Dissepiments mostly interseptal, regular. Lonsdaleoid dissepiments small when present. In corallites most altered diagenetically, either all, or most major septa restricted to tabularium. Minor septa very restricted in number. Few of them remain as thin, spike-like bodies attached to external walls, mostly absent. Dissepimentarium consists solely or mostly of large lonsdaleoid dissepiments, some surrounding more than 1/4 corallite diameter. Pseudocolumellae either absent from diagenetically most advanced corallites or free when surviving. Dissepiments in longitudinal sections either in one vertical row (Fig. 16A5) or up to three, gently dipping rows (Fig. 16A2). Tabulae complete or incomplete, deflected gently from thin pseudocolumella. Some tabellae rest upon underlying tabulae. Increase lateral (Fig. 16A3, A4, A6, A7). Acolumellate early growth stage of offsets continue up to their total separation from parental polyp (Fig. 16A6). Derivation of pseudocolumella from axial septum very probable, but diagenetic alterations precludes firm recognition of that process (Fig. 16A7).
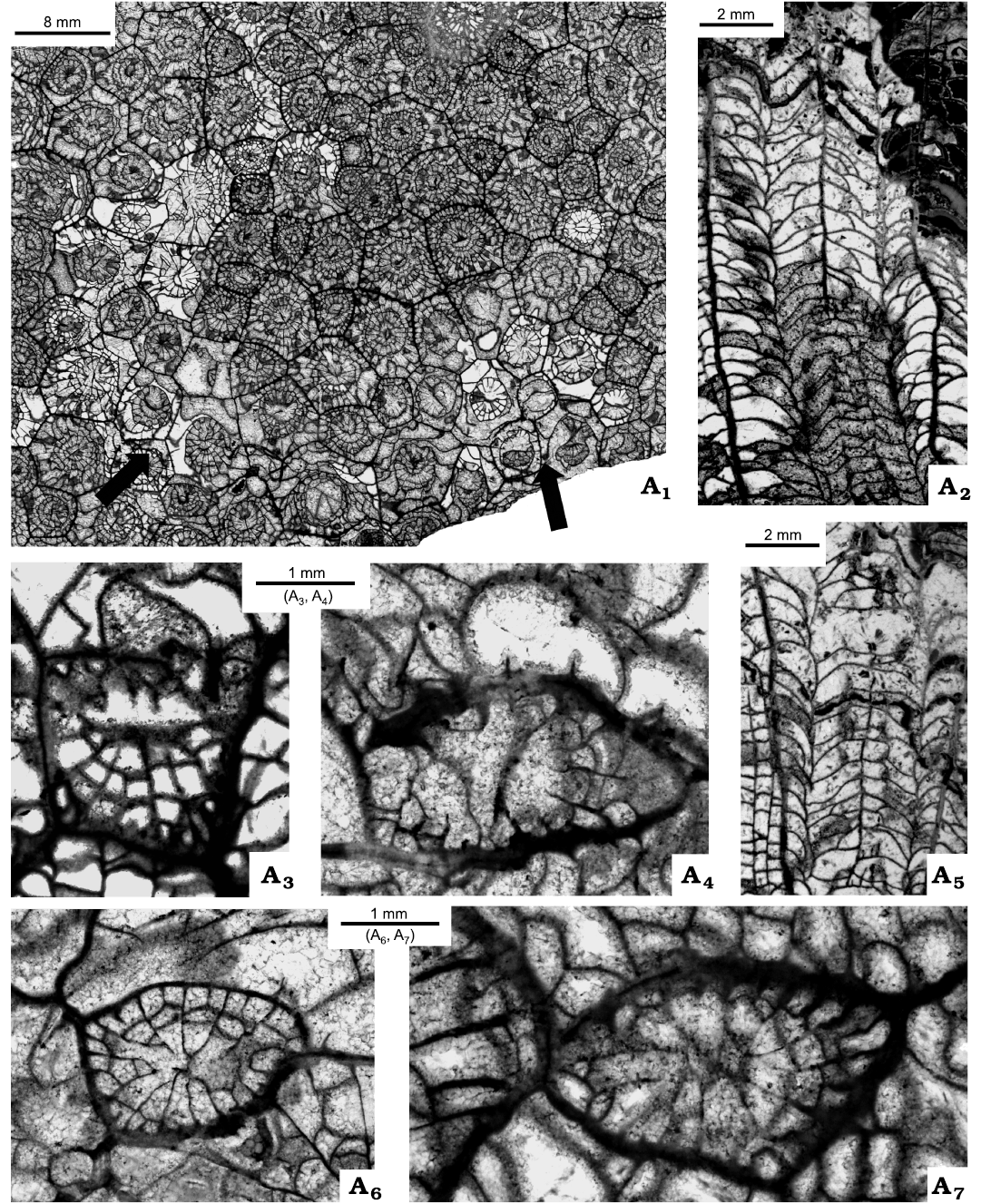
Fig. 16. Rugose coral Stelechophyllum cf. banffense (Warren, 1927), C-52434 = GSC-142476, locality 12 (Fig. 2), from Meilleur Member, Flett Formation (Mississipian), Rundle Group, Liard Basin, Northwest Territories, Canada. Thin sections. A1, transverse section of fragment of colony, note different state of preservation in middle of picture and area around it (arrows); A2, A3, longitudinal sections; A4–A7, offsets in ascending growth stages.
Remarks.—The morphology of the colony under discussion differs from the lectotype (Nelson 1960: pl. 23: 4, 5; Bamber 1966: pl. 3: 5) in its n:d value, in having stronger minor septa in its less altered part and in having a weaker pseudocolumella. It resembles small and simple variants of S. banffense (Warren, 1927), characterized by Sando (1983: 13) as “highly variable species”. Differences in the morphology illustrated by Sando (1963: pls. 8, 9) may have resulted from a differentiated intensity in diagenetic alterations. His colonies with large lonsdaleoid dissepiments and reduced minor septa may correspond to more altered parts of the colony described above, whereas those with the minor septa well developed and the lonsdaleoid dissepiments are rare and may correspond to the less altered part of that colony (Sando 1983: pl. 8: 1, 3 and pl. 9: 1, respectively). The longitudinal section illustrated by Sando (1983: pl. 4: 2c) resembles that illustrated here (Fig. 16A2, A5). It is probable that those two corallites are conspecific. The question is whether they can be included within the limits of intraspecific variability of S. banffense or should be described as new. Having such restricted material, taking in mind the large variability of the specimens included in S. banffense so far, and considering the influence of diagenesis on the morphology of the corals documented above, the conformity of the species described here to S. banffense is the safest solution.
Discussion
Three items require additional comments: (i) the distinction between the European and North American species of Siphonodendron: the American species were either included to Siphonodendron as a subgenus of Lithostrotion, to Lithostrotion treated broadly, or to Diphyphyllum. That question has already been raised by Fedorowski and Bamber (2007), but the discovery that pseudocolumella is formed on the tabulae and independently from the major septa (see description of Cordilleria aff. mutabile) verifies that suggestion, i.e., the independent taxonomic position of the North American Lithostrotionidae, included here in Cordilleria gen. nov.; (ii) the interspecific variability of the taxa here included in Cordilleria gen. nov., resulting in the indefinite identification of species; and (iii), the influence of diagenetic alteration on the taxonomy of rugose corals.
The longitudinal sections investigated in this paper (Fig. 11A2, A4) leave no doubt as to the derivation of the pseudocolumella as suggested above. It is invariably deleted in North American lithostrotionids, sometimes up to the mature growth stage inclusively and is invariably free from any major septum from its beginning. We interpret the image illustrated in Fig. 11A2, A4, as showing a continuous tabula, thin beneath the pseudocolumella (Fig. 11A4, white arrow) and thickened both sides of it as follows: the epidermis of the polyp’s floor leaves the axial part of the tabula to form an epidermal pocket wide at the bottom, but narrowing upwards to form a body typical for septa creating pockets (Fig. 11A4, black arrows). The septum-like skeleton of the pseudocolumella is secreted in that pocket. The pseudocolumella created that way may either stop growing after a short period of its existence, i.e., as in the case discussed, or may continue to grow for some time as demonstrated in the lower part of the same picture. The disconnection of the polyp’s epidermis from both the tabula and the pseudocolumella and its upwards shifting may be strong and complete, as demonstrated by the interruption of the pseudocolumella, or local when only peripheral parts of the polyp’s epidermis are disconnected from the tabula and shifted upwards to form the next tabulae, whereas the pseudocolumella’s pocket remains stable and secretion of the pseudocolumella is continuous for various periods of the corallite growth. The pseudocolumella in the European Lithostrotionidae is also absent from the very early growth of the offset (e.g., Khoa 1977: figs. 10–13, 15, 16), but in contrast to the North American Lithostrotionidae, its derivation is invariably connected to protosepta. Fedorowski and Bamber (2007) accepted that fundamental difference as adequate for the addition of the new genus to North American Lithostrotionidae, an idea accepted and followed here by the introduction of Cordilleria gen. nov.
The North American Lithostrotionidae have served as types for some biostratigraphic zones (e.g., Nelson 1960; Sando and Bamber 1985). Indeed, the holotypes of most species used for those zones differ to the extent permitted for accepting such an approach. However, the analysis of specimens belonging to a small collection such as we deal with in this paper creates many difficulties in the identification of particular colonies. Figures 9, 12, and 14 demonstrate variety in the n:d values, a character traditionally and correctly treated as an important taxonomic criterion within the Rugosa. The morphology of all characters, such as the number of rows of dissepiments, the length of the minor septa, and the morphology in longitudinal sections, considered important in the case of the holotypes of the lithostrotionid species discussed, becomes much less rigid when other specimens are studied. Only a large collection of specimens, derived from a wide area will allow establishing of limits of the intra- vs. inter-specific variability of the North American Lithostrotionidae. Such a collection, derived from more than 200 localities, is being studied with the target of establishing a new approach to those corals. The data published here and its discussion are treated by us as an introduction study, suggesting that new students should take a careful approach to them.
The role of diagenetic alteration is commonly omitted from descriptions of taxa showing changes created by recrystallization, replacement and other factors not treated as responsible for differences in morphology. We purposely avoid pointing to literature examples other than that of Sando (1983), although a comparison of the colony illustrated here (Fig. 16A1) can easily find its counterparts in the literature dealing with the North American Lithostrotionidae. We consider drawing attention to that question important. It would be enough to treat the weakly and strongly altered parts of colony GSC 142476 as separate fragments in order to place them into separate species. Occurrence vs absence of minor septa, confinement of major septa to the tabularium vs their common attachment to the external wall and small, uncommon lonsdaleoid dissepiments hidden between interseptal ones vs large lonsdaleoid dissepiments, occupying an entire dissepimentarium create a set of characters adequate for creation of such different species.
Conclusions
This is the first time that the tabulate coral, identified with some uncertainty as Michelinia expansa White, 1883, is reported, discussed and illustrated on the basis of a specimen from the Liard Basin.
The reconstruction of the formation process of pseudocolumella in North American lithostrotionid rugose corals identified so far as Diphyphyllum, Lithostrotion, Siphonodendron, and/or Dorlodotia (e.g., Warren 1927; Kelly 1942; Sutherland 1958; Nelson 1960; Sando 1963, 1969a; Bamber 1966; Sando and Bamber 1985) shows pseudocolumella originating from the floor epidermis of the polyp. This contrasts with the formation of pseudocolumella from protosepta in European lithostrotionids and warrants the introduction of a new genus Cordilleria to the family Lithostrotionidae.
The large variability in all skeletal structures of the North American lithostrotionid species of the genera listed above and the differences in the interpretation of certain characters by authors requires revision of all those species. Such a revision should rely on a large collection sourced from numerous localities, whereas the collection available for this study is restricted in number and geography. Thus only the main characteristics of two holotypes were compiled from the literature as the reference characteristics of those species and the specimens from the Liard Basin, probably belonging to these species, are described as affinis.
A variety of diagenetic alterations to skeletons within a colony of Stelechophyllum cf. banffense resulted in localized changes to species-defining characters. This precluded a closer identification of that colony and suggests that this phenomenon should be factored into any study of rugose corals.
Acknowledgements
We are indebted to Mikołaj Zapalski (Warsaw University, Poland) for his kind remarks on some tabulate corals genera. We want to thank the critical reviewers, Sergio Rodriguez (Complutense University, Madrid, Spain) and Calvin H. Stevens (San Jose University, California, USA) for their valuable remarks. We are also thankful to Marta Bartkowiak (Adam Mickiewicz University, Poznań, Poland) for her excellent technical assistance. This research is funded by the National Research Center, grant UMO-2016/21/B/ST10/0186.
References
Armstrong, A.K. 1962. Stratigraphy and paleontology of the Mississippian System in southwestern New Mexico and adjacent southeastern Arizona. New Mexico Bureau of Mines and Mineral Resources Memoir 8: 1–99.
Bamber, E.W. 1966. Type lithostrotionid corals from the Mississippian of western Canada. Geological Survey of Canada Bulletin 135: i–iv + 1–28. Crossref
Douglas, R.J.W and Norris, D.K. 1976a. Geology, Fort Liard, District of Mackenzie. Geological Survey of Canada, Map 1379A, scale 1:250,000 [published online, https://doi.org/10.4095/109154]. Crossref
Douglas, R.J.W. and Norris, D.K. 1976b. Geology, Sibbeston Lake, District of Mackenzie. Geological Survey of Canada, Map 1377A, scale 1:250,000 [published online, https://doi.org/10.4095/109051]. Crossref
Douglas, R.J.W. and Norris, D.K. 1976c. Geology, Virginia Falls, District of Mackenzie. Geological Survey of Canada, Map 1378A, scale 1:250,000 [published online, https://doi.org/10.4095/109052]. Crossref
Douglas, R.J.W., Gabrielse, H., Wheeler, J.O., Stott, D.F., and Belyea, H.R. 1970. Geology of western Canada. In: R.J.W. Douglas (ed.), Geology and Economic Minerals of Canada. Geological Survey of Canada, Economic Geology Report 1: 366–488. Crossref
Ehrenberg, C.G. 1834. Beiträge zur physiologischen Kenntniss der Corallenthiere im allgemeinen, und besonders des rothen Meeres, nebst einem Versuche zur physiologischen Systematik derselben. Abhandlungen der Königlichen Akademie der Wissenschaften, Berlin 1: 225–380.
Fedorowski, J. 1967. The Lower Permian Tetracoralla and Tabulata from Treskelodden,Vestspitsbergen. Norsk Polarinstitut Skrifter 142: 11–42.
Fedorowski, J. 2019. Bashkirian Rugosa (Anthozoa) from the Donets Basin (Ukraine). Part 7. The family Neokoninckophyllidae Fomichev, 1953, with preliminary revision of Moscovian taxa. Acta Geologica Polonica 69: 59–87.
Fedorowski, J. and Bamber, E.W. 2007. Remarks on lithostrotionid phylogeny in eastern North America and western Europe. Österreichische Akademie der Wissenschaften. Schriftenreihe der Erdwissenschaftlichen Kommissionen 17: 251–273.
Fedorowski, J. and Jull, R.K. 1976. Review of blastogeny in Palaeozoic corals and description of lateral increase in some Upper Ordovician rugose corals. Acta Palaeontologica Polonica 21: 37–78.
Fedorowski, J., Bamber, E.W., and Richards, B.C. 2019. Bashkirian rugose corals from the Carboniferous Mattson Formation in the Liard Basin, northwest Canada—stratigraphic and paleobiogeographic implications. Acta Palaeontologica Polonica 64: 851–870. Crossref
Ferri, F., McMechan, M., and Creaser, R. 2015. Oil and Gas Geoscience Reports 2015, B.C. The Besa River Formation in Liard Basin, British Columbia. 27 pp. Ministry of Natural Gas Resources, Ottawa.
Fleming, J. 1828. A history of British animals. xxiii + 265 pp. Bell and Bredfute, Edinburgh.
Fomichev, V.D. [Fomičev, V.D.] 1953. Korally Rugosa i stratigrafiâ sredne i verhnekamennougol’nyh i permskih otloženij Donetskogo bassejna. Text 622 pp. + Atlas 70 pp. Trudy Vsesoûznogo Naučno-Issledovatel’skogo Geologičnego Instituta (VSEGEI), Moskva.
Frey, R.W. and Pemberton, S.G. 1985. Biogenic structures in outcrop and cores. I. Approaches to ichnology. Bulletin of Canadian Petroleum Geology 33: 72–115.
Girty, G.H. 1910. New genera and species of Carboniferous fossils from the Fayetteville Shale of Arkansas. New York Academy of Sciences Annals 20: 189–238. Crossref
Harker, P. 1961. Summary of Carboniferous and Permian formations, southwestern District of Mackenzie. Geological Survey of Canada, Paper 61-1: 1–9. Crossref
Harker, P. 1963. Carboniferous and Permian rocks, southwestern District of Mackenzie. Geological Survey of Canada, Bulletin 95: 1–102. Crossref
Henderson, C.M., Richards, B.C., and Barclay, J.E. 1994. Permian strata of the Western Canada Sedimentary Basin. In: G.D. Mossop, J. Dixon, J. Kramers, M. Madanicky, H. Blackie, D. Kent, D. Smith, and J. Monro (eds.), Geological Atlas of the Western Canada Sedimentary Basin, 251–258. Canadian Society of Petroleum Geologists and Alberta Research Council, Calgary.
Hill, D. 1981. Coelenterata, Supplement 1. In: C. Teichert (ed.), Treatise on Invertebrate Paleontology, Part F: Rugosa and Tabulata, 1–762. Geological Society of America, Boulder and University of Kansas Press, Lawrence.
Kelly, W.H. 1942. Lithostrotionidae in the Rocky Mountains. Journal of Paleontology 16: 351–361.
Khoa, N.D. 1977. Carboniferous Rugosa and Heterocorallia from boreholes in the Lublin Region (Poland). Acta Palaeontologica Polonica 22: 301–404.
Koninck, L.G de 1841. Description des animaux fossils qui se trouvent dans le terrain Carbonifére de Belgique. iv + 178 pp. H. Dessain, Liège.
Lafuste, J. and Plusquellec, Y. 1985. Structure et microstructure de quelques Michelinidae et Michelinimorphes (Tabulata, paleozoiques). Bulletin du Muséumnational d’histoire naturelle, Serie 4 7: 13–63.
Lafuste, J. and Plusquellec, Y. 1990. Les genres Utaratuia, Tabellaephyllum, Michelinia et ladistinction Tabulata-Rugosa. Annales de Paléontologie 76: 13–28.
Lane, L.S. 2006. Geology Mount Flett, District of Mackenzie, Northwest Territories (95B/12). Geological Survey of Canada, Open File map 5236, scale 1:50000, doi:10.4095/221970. Crossref
Lane, L.S. and Hynes, G.F. 2005. Geology Sawmill Mountain (95B/13), District of Mackenzie, Northwest Territories. Geological Survey of Canada, Open File 4940, scale 1:50,000 [published online, https://doi:10.4095/220620. Crossref
Leckie, D.A., Potocki, D.J. and Visser, K. 1991. The Lower Cretaceous Chinkeh Formation: A frontier-type play in the Liard Basin of Western Canada. American Association of Petroleum Geologists Bulletin 75: 1324–1352. Crossref
Lonsdale, W. 1845. Description of some characteristic Palaeozoic corals from Russia. In: R.I. Murchison, E. de Verneuil and A. von Keyserling (eds.), The Geology of Russia in Europe and Ural Mountains. Volume 1, 591–634. John Murray, London.
MacLean, B.C. 2002. Bovie structure: A decapitated thrust? Geological Survey of Canada Open File 4274: https://doi.org/10.4095/213391. Crossref
MacLean, B. C. and Morrow, D.W. 2004. Bovie Structure: its evolution and regional context. Bulletin of Canadian Petroleum Geology 52: 302–324. Crossref
McCoy, F. 1849. On some new genera and species of Palaeozoic corals and Foraminifera. The Annals and Magazine of Natural History. Series two 3 (13): 1–20, 119–136. Crossref
Meek, F.B. 1877. Paleontology. U.S. Geological Exploration of the Fortieth Parallel (King) 4: 1–17.
Milne Edwards, H. and Haime, J. 1850. A Monograph of the British Fossil Corals. i–lxxxv + 1–72. The Palaeontographical Society of London, London.
Nelson, S.J. 1960. Mississippian lithostrotionid zones of the southeastern Canadian Rocky Mountains. Journal of Paleontology 34: 107–126.
Oliver, W.A., Jr. and Sando, W.J. 1977. Tabellaephyllum Stumm, 1948 is a Michelinia (Carboniferous, Tabulata). Journal of Paleontology 51: 422.
d’Orbigny, A. 1852. Cours élémentaire de paleontologie et géologie stratigraphique.Volume 2 (1). 328 pp. Victor Masson, Paris.
Plusquellec, Y. and Sando, W.J. 1987. The microstructure of Michelinia meekiana Girty, 1910. Journal of Paleontology 61: 10–13. Crossref
Richards, B.C., Barclay, J.E., Bryan, D., Hartling, A., Henderson, C.M., and Hinds, R.C. 1994. Chapter 14. Carboniferous strata of the Western Canada Sedimentary Basin. In: G.D. Mossop, J. Dixon, J. Kramers, M. Madanicky, H. Blackie, D. Kent, D. Smith, and J. Monro (eds.), Geological Atlas of the Western Canada Sedimentary Basin, 221–250. Canadian Society of Petroleum Geologists and Alberta Research Council, Calgary.
Richards, B.C., Mamet, B.L., Bamber, E.W., Higgins, A.C., McGregor, D.C., and Utting, J. 1989. Uppermost Devonian and lower Carboniferous stratigraphy, sedimentation, and diagenesis, southwestern District of Mackenzie and southeastern Yukon Territory. Geological Survey of Canada, Bulletin 390: 1–135. Crossref
Salée, A. 1910. Sur un genre nouveau de Tétracoralliaires (Dorlodotia) et la valeur stratigraphique des Lithostrotion. Annales de la Société Scientifique de Bruxelles 39: 145–154.
Sando, W.J. 1963. New species of colonial rugose corals from the Mississippian of southern Arizona. Journal of Paleontology 37: 1074–1079.
Sando, W.J. 1965. Revision of some Paleozoic coral species from the western United States.U.S. Geological Survey Professional Paper 503-E: E1–E38. Crossref
Sando, W.J. 1969a. Corals. In: E.D. McKee and R.C. Gutschick (eds.), History of the Redwall Limestone of northern Arizona. Geological Society of America Memoir 114: 257–342.
Sando, W.J. 1969b. Revision of some of Girty’s invertebrate fossils from the Fayetteville Shale (Mississippian) of Arkansas and Oklahoma—corals. Geological Survey Professional Paper 606B: 9–14.
Sando, W.J. 1983. Revision of Lithostrotionella (Coelenterata, Rugosa) from the Carboniferous and Permian. Geological Survey Professional Paper, 1247:1–52. Crossref
Sando, W.J. and Bamber, E.W. 1985. Coral zonation of the Mississippian system in the Western Interior Province of North America. U.S. Geological Survey Professional Paper 1334: i–iii + 1–61. Crossref
Seilacher, A. 1967. Bathymetry of trace fossils. Marine Geology 5: 413–428. Crossref
Smith, S. and Lang, W.D. 1930. Descriptions of the type specimens of some Carboniferous corals of the genera “Diphyphyllum”, “Stylastraea”, Aulophyllum and Chaetetes. The Annals and Magazine of Natural History. Series 10 5: 177–194. Crossref
Spasskiy, N.Yu. [Spasskij, N.Û.] and Kachanov, E.I. 1971. New primitive early Carboniferous corals of the Altay and Urals [in Russian]. Zapiski Leningradskogo Gornogo Instituta 59: 48–64.
Stumm, E.C. 1948. Upper Devonian compound tetracorals from the Martin Limestone. Journal of Paleontology 22: 40–47.
Sutherland, P.K. 1958. Carboniferous stratigraphy and rugose coral faunas of northeastern British Columbia. Geological Survey of Canada Memoir 295: i–vi + 1–177. Crossref
Tolmachev, I.P. 1933. New names for two genera of Carboniferous corals. Geological Magazine 70: 287. Crossref
Trautschold, H. 1879. Die Kalkbrüche von Myatschkowa. Eine Monographie des Oberen Bergkalks. Société Impériale des Naturalistes de Moscou, Mémoir 14, 1–82.
Verrill, A.E. 1865. Classification of polyps (Extract condensed from a synopsis of the polypi of the North Pacific Exploring Expedition, under captains Ringgold and Rogers, U.S.N.). Proceedings of the Essex Institute 4: 145–149. Crossref
Waagen, W.H. and Wentzel, J. 1886. Salt Range fossils. Volume 1. Productus limestone fossils; 6. Coelenterata. Palaeontologia Indica 13: 835–924.
Warren, P.S. 1927. Banff area, Alberta. Canada Geological Survey Memoir 153: 1–94. Crossref
Wedekind, R. 1937. Einführung in die Grundlagen der historischen Geologie. II. Band. Die Korallen- und Foraminiferenzeit. 136 pp. Ferdinand Enke, Stuttgart.
White, C.A. 1875. Report upon the invertebrate fossils collected in portions of Nevada, Utah, Colorado, New Mexico and Arizona by parties of the expeditions of 1871, 1872, 1873 and 1874. U.S. Geographical and Geological Explorations and Surveys West of the One Hundredth Meridian (Wheeler) Report 4: 1–219 (preprint).
White, C.A. 1877. Report upon the invertebrate fossils collected in portions of Nevada, Utah, Colorado, New Mexico and Arizona by parties of the expeditions of 1871, 1872, 1873 and 1874. U.S. Geographical and Geological Explorations and Surveys West One Hundredth Meridian (Wheeler) Report 4: 1–219. Crossref
White, C.A. 1883. Contributions to invertebrate paleontology. No. 8 – fossils from Carboniferous rocks of the Interior States. U.S. Geological and Geographical Survey of the Territories (Haiden), Annual Report 12 (for 1878): 155–171.
Wilson, E.C. 1963. An evaluation of the genomorph concept in rugose corals. Systematic Zoology 12: 83–90. Crossref
Wright, G.N., McMechan, M.E., Potter, D.E.G, and Holter, M.E. 1994. Structure and Architecture of the Western Canada Sedimentary Basin. In: G.D. Mossop, J. Dixon, J. Kramers, M. Madanicky, H. Blackie, D. Kent, D. Smith and J. Monro (eds.), Geological Atlas of the Western Canada Sedimentary Basin, 25–40. Canadian Society of Petroleum Geologists and Alberta Research Council, Calgary.
Acta Palaeontol. Pol. 66 (3): 679–704, 2021
https://doi.org/10.4202/app.00817.2020