
A new late-surviving early diverging Ibero-Armorican duck-billed dinosaur and the role of the Late Cretaceous European Archipelago in hadrosauroid biogeography
ALBERT PRIETO-MÁRQUEZ and MIGUEL ÁNGEL CARRERA FARIAS
Prieto-Márquez, A. and Carrera Farias, M.Á. 2021. A new late-surviving early diverging Ibero-Armorican duck-billed dinosaur and the role of the Late Cretaceous European Archipelago in hadrosauroid biogeography. Acta Palaeontologica Polonica 66 (2): 425–435.
The anatomy and phylogenetic systematics of a well preserved hadrosauroid dentary collected in the 1990s from uppermost Maastrichtian strata (within chron C29r) of the Figuerola Formation, in the Àger syncline of the southern Pyrenees in northeastern Spain are revised. The specimen represents a new genus and species of basally branching hadrosauroid dinosaur, Fylax thyrakolasus gen. et sp. nov. This taxon is the third hadrosaurid outgroup species erected in Europe so far and the stratigraphically youngest non-hadrosaurid hadrosauroid known to date. It is diagnosed on the basis of a unique combination of characters such as a dorsal region of the coronoid process that is at least as wide anteroposteriorly as 30% of dental battery length, a coronoid process inclined anteriorly less than 80º relative to the alveolar margin of the dentary, steeply inclined and flat occlusal surface of the dental battery, and dentary tooth crowns 2.8–3.3 times taller than wide without marginal denticles and with a pair of major long ridges on the enameled lingual surface. The parsimony analysis confirmed that F. thyrakolasus gen. et sp. nov. is a close outgroup to Hadrosauridae. However, we also found that it is sister to the hadrosauroid Tethyshadros insularis form the late Campanian–early Maastrichtian Adriatic-Dinaric Carbonate Platform (present-day Italy). Maximum likelihood reconstruction of ancestral areas on the time calibrated phylogeny revealed that the ancestry of the Fylax–Tethyshadros clade has Appalachian roots. This study supports an Asian origin for hadrosauroids and subsequent dispersal to Appalachia, where Hadrosauridae likely originated. In this scenario, the Late Cretaceous European Archipelago could have facilitated the dispersal of hadrosaurid outgroups from Asia to Appalachia.
Key words: Dinosauria, Hadrosauroidea, anatomy, systematic, end-Cretaceous, Spain, Pyrenees.
Albert Prieto-Márquez [albert.prieto@icp.cat], Institut Català de Paleontologia Miquel Crusafont, Universitat Autònoma de Barcelona, c/Escola Industrial 23, 08201 Sabadell, Barcelona, Spain.
Miguel Angel Carrera Farias [miguelangel.carrera@e-campus.uab.cat], Universitat Autònoma de Barcelona, Facultat de Ciències i Biociències, Entrada Sud Edifici de Ciències, Carrer de la Vall Moronta, 08193 Bellaterra (Cerdanyola del Vallès), Barcelona.
Received 18 September 2020, accepted 6 November 2020, available online 2 June 2021.
Copyright © 2021 A. Prieto-Márquez and M.A.Carrera Farias. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
The Late Cretaceous European Archipelago, an extensive array of islands that existed between the Atlantic and Tethys oceans, supported a diverse assemblage of vertebrate clades (Csiki-Sava et al. 2015). Among the various dinosaurian groups present in these islands, hadrosauroid ornithopods were a major component of the Maastrichtian terrestrial ecosystems (Prieto-Márquez et al. 2013; Dalla Vecchia 2014; Dalla Vecchia et al. 2014). These herbivores were iguanodontian ornithopods that evolved dental batteries (Erickson et al. 2012) and, in the more morphologically derived hadrosaurids, hypertrophied nasal passages and a wide variety of cranial crests (Ostrom 1962; Hopson 1975). Here, we follow Sereno (1998) in regarding Hadrosauroidea as all dinosaurs more closely related to Parasaurolophus than Iguanodon.
Most of the European hadrosauroid record correspond to lambeosaurine hadrosaurids from Maastrichtian strata of the Pyrenean region of northeastern Spain (Casanovas-Cladellas et al. 1993; Pereda-Suberbiola et al. 2009a, b; Riera et al. 2009; Cruzado-Caballero et al. 2010; Dalla Vecchia et al. 2014; Prieto-Márquez et al. 2019) and southern France (Laurent et al. 2002; Billotte et al. 2010; Prieto-Márquez et al. 2013), both formerly part of the large Ibero-Armorican Island at the southwestern end of the archipelago. Remains of earlier diverging hadrosauroids have been less forthcoming, with the notable exception of the relatively abundant materials of Telmatosaurus transsylvanicus, from the Maastrichtian Sânpetru Formation of Romania (Weishampel et al. 1993; Dalla Vecchia 2006), and the articulated skeletons of Tethyshadros insularis, from the upper Campanian–lower Maastrichtian Liburnian Formation of Italy (Dalla Vecchia 2009, 2020). Additional, yet fragmentary remains of indeterminate hadrosauroids have also been recovered from Upper Cretaceous strata of Ukraine, Bulgaria, Slovenia, Germany, Belgium, the Netherlands, Spain, and France (Dalla Vecchia 2014).
Aside from the richer Romanian and Italian record, the next best-preserved material of a European hadrosaurid outgroup corresponds to a dentary (IPS-36338) collected in the early 90s from uppermost Maastrichtian strata of the Figuerola Formation cropping out at the Àger Syncline (Lleida province, northeastern Spain; Fig. 1). Casanovas et al. (1999) originally described IPS-36338 and their phylogenetic analysis positioned the specimen as sister to Saurolophidae (Lambeosaurinae + Saurolophinae sensu Prieto-Márquez 2010b). In a review of the Iberian hadrosauroid record, Pereda-Suberbiola et al. (2009a) provided a cursory redescription of IPS-36338 and confirmed its position as a hadrosaurid outgroup species. More recently, Blanco et al. (2015) undertook a series of quantitative analyses of morphological characters in a large sample of Ibero-Armorican dentaries and recognized IPS-36338 as representing a distinct morphotype from all other hadrosauroid taxa in the region.
The last decade has seen much progress on the taxonomy and phylogenetic systematics of hadrosauroid dinosaurs. These studies more than doubled both the diversity of species and the number of phylogenetically informative characters of the clade since the time of the discovery and early work on IPS-36338. The phylogenetic analysis of Casanovas et al. (1999) included only eight taxa, in addition to IPS-36338, and only 17 dental and mandibular characters. In the current context, that represents only about 10% of the skeletal phylogenetically informative characters available for hadrosauroids and less than 10% of the known diversity of the clade. Pereda-Suberbiola et al. (2009a) performed several phylogenetic analyses, but these focused on Lambeosaurinae and it is not clear whether IPS-36338 was included (the resulting topologies were not figured in the article and are nowhere to be found in their supplementary material).
The present study addresses these shortcomings by revising the comparative anatomy of IPS-36338 and its phylogenetic position in the light of the state of the art of the diversity and evolution of hadrosauroid dinosaurs. The unique morphology of IPS-36338 recognized by Blanco et al. (2015) is documented in detail and shown to be sufficiently distinct from that of other hadrosauroids to warrant the erection of a new genus and species. Finally, possible functional implications of the dental morphology of IPS-36338, as well as insights into its biogeographic history, are discussed.
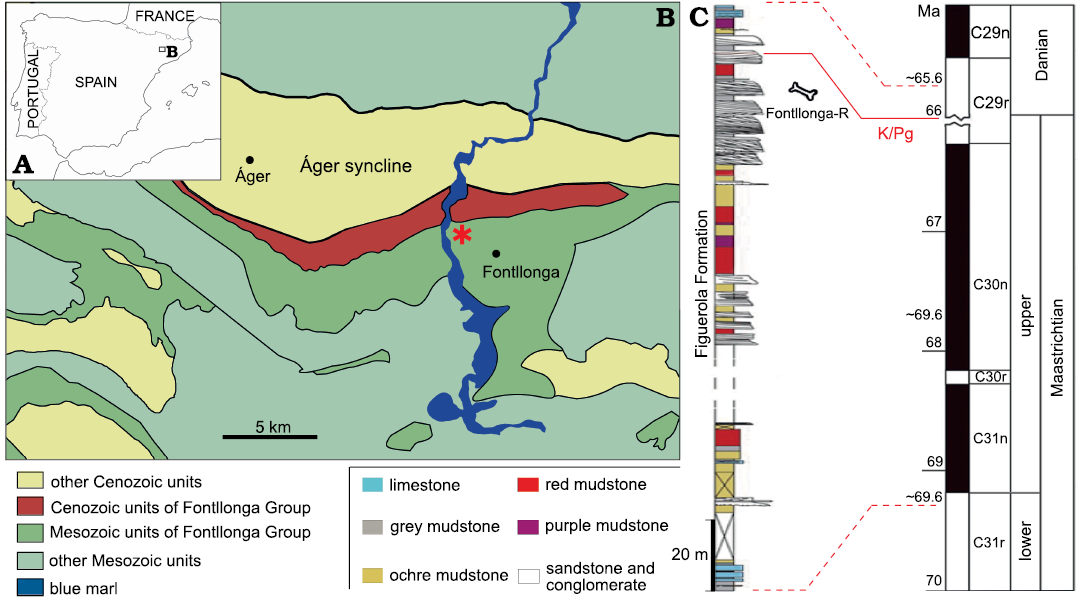
Fig. 1. Geographic location and stratigraphic position of Fontllonga-R, the type locality of Fylax thyrakolasus gen. et sp. nov. A. Simplified map showing the location of the Àger syncline in northeastern Spain. B. Geological map of the Àger syncline and adjacent areas displaying the location of the Fontllonga-R locality (asterisk), near the eponymous town. C. Stratigraphic section of the Fontllonga Formation cropping out at the Fontllonga-R locality, showing the position of the holotype dentary IPS-36338 (modified from Fondevilla et al. 2019).
Institutional abbreviations.—AMNH FARB, American Museum of Natural History, Fossil Amphibian, Reptile, and Bird Collection, New York, USA; IPS, Museu de l’Institut Català de Paleontologia Miquel Crusafont (formerly Institut de Paleontologia de Sabadell), Sabadell, Spain; MGUV, Museo de Geología, Universidad de Valencia, Spain; MOR, Museum of the Rockies, Bozeman, USA; MVP, Museo Paleontológico de Valencia, Spain; NMMNH, New Mexico Museum of Natural History and Science, Albuquerque, USA; ROM, Royal Ontario Museum, Toronto, Canada; TMP, Royal Tyrrell Museum of Paleontology, Drumheller, Canada; YPM, Yale Peabody Museum of Paleontology, New Haven, USA.
Nomenclatural acts.—This published work and the nomenclatural acts it contains have been registered in ZooBank:urn:lsid:zoobank.org:pub:DBBAD226-62A9-4588-B8A2-E35492DDC382.
Systematic palaeontology
Dinosauria Owen, 1842
Ornithischia Seeley, 1887
Ornithopoda Marsh, 1881
Iguanodontia Dollo, 1888
Hadrosauroidea Sereno, 1986 (sensu Sereno 1998)
Genus Fylax nov.
Zoobank LSID: urn:lsid:zoobank.org:act:CD63DB79-1031-4664-B6 D3-80786EF580AA
Type species: Fylax thyrakolasus gen. et sp. nov., monotypic, see below.
Etymology: From the Greek fýlax, keeper; for rationale see below.
Diagnosis.—As for the type species by monotypy.
Fylax thyrakolasus sp. nov.
Fig. 2.
Zoobank LSID: urn:lsid:zoobank.org:act:97E63552-0A0B-4208-8F25-9161CA2731EF
1999 Euadrosauria indeterminate; Casanovas et al. 1999: fig. 2.
2009 Hadrosauroidea indeterminate; Pereda-Suberbiola et al. 2009: fig. 1G.
Etymology: From the Greek thýra, door or gate and kólasi, hell; meaning together with the generic name “keeper of the gates of hell”, in reference to the extreme temporal proximity of this taxon to the Late Cretaceous mass extinction event.
Holotype: IPS-36338, a nearly complete left dentary with teeth.
Type locality: Fontllonga-R, a locality near the eponymous village, next to the c-13 road that lies between the municipalities of Camarasa and La Baronia de Sant Oisme, Noguera county, in the southern reaches of the Montsec mountain range of Lleida province, in the southern limb of the Àger syncline, northeastern Spain (Colombo and Cuevas 1993; López-Martínez et al. 1999; Fondevilla et al. 2019). Fontllonga-R is the same locality that Casanovas et al. (1999) designated as simply Fontllonga (see also Sellés and Vila 2015: table 1).
Type horizon: Rocks corresponding to chron C29r within the Figuerola Formation, a few meters from the K/Pg boundary (Galbrun et al. 1993; Caus et al. 2016; Oms et al. 2016; Fondevilla et al. 2019: figs. 9, 11). The Figuerola Formation spans chrons C31n through C29n and consists of alternating brown, ochre, and reddish marl with sandstone that have been interpreted as fluvial deposits (Cuevas 1992; Rosell et al. 2001; Riera et al. 2009). This locality is one of the few in the world preserving a continental geological record across the K/Pg boundary (López-Martínez et al. 1998).
Material.—Holotype only.
Diagnosis.—Non-hadrosaurid hadrosauroid dinosaur characterized by the following unique combination of characters: dorsal region of coronoid process at least as wide anteroposteriorly as 30% of dental battery length (convergent in the lambeosaurine Parasaurolophus tubicen); coronoid process of dentary lacking ridge on posteromedial surface and inclined anteriorly less than 80° relative to alveolar margin; steeply inclined (i.e., angled less than 45° with coronoid process) and flat occlusal surface of dental battery; dentary tooth crowns 2.8–3.3 times taller than wide, lacking marginal denticles and with two major long ridges on enameled lingual surface.
Description.—The dentary of Fylax thyrakolasus gen. et sp. nov. is nearly completely preserved, except for the missing symphyseal process (Fig. 2). It measures 276 mm in length and 180 mm from the dorsal margin of the coronoid process to ventral margin of dentary, perpendicular to the dental battery. The mandibular ramus is relatively deep, being only 2.7 times longer than tall at its deepest point. The ventral margin is nearly straight in lateral view, displaying only a very gentle sinuous profile (Fig. 2A5).
The most salient attribute of the dentary of F. thyrakolasus gen. et sp. nov. is its disproportionately massive coronoid process. In particular, the dorsal region of the coronoid process is greatly enlarged, being 31% of the length of the dental battery. Among hadrosauroids, only the lambeosaurine Parasaurolophus tubicen displays a similar ratio (e.g., 30% in NMMNH P-25100; Fig. 3). All other examined hadrosauroid species have lower ratios, the majority of them ranging 18–25% (Fig. 3). However, when compared to the minimum width of the ascending ramus of the coronoid process, the dorsal region of this process in F. thyrakolasus gen. et sp. nov. is less expanded than in hadrosaurids. Thus, in Hadrosauridae and Claosaurus agilis (e.g., YPM 1190), the dorsal region of the coronoid process is at least 1.5 times wider than the ascending ramus, whereas in F. thyrakolasus gen. et sp. nov. and other early diverging hadrosauroids the dorsal region is less than 1.5 times wider than the ramus (SOM 1, Supplementary Online Material available at http://app.pan.pl/SOM/app66-PrietoMarquez_CarreraFarias_SOM.pdf). The medial surface of the dorsal region of the coronoid process in F. thyrakolasus gen. et sp. nov. is paddle-shaped, slightly deeper than wide. Most of what is seen of the coronoid process above the lingual side of the dental battery consists of this medial surface of the dorsal region. A short acute apex is present along the posterodorsal margin of the coronoid process. The anterior and posterior margins of the ascending ramus of the coronoid process are parallel. As in the vast majority of hadrosaurids (SOM 2) and Tethyshadros insularis (Dalla Vecchia 2009), the coronoid process of F. thyrakolasus gen. et sp. nov. is angled anteriorly less than 80° relative to the dorsal margin of the alveolar sulci of the dental battery (74° in the case of IPS-36338) (contra Pereda-Suberbiola et al. 2009a). Unlike in many early diverging hadrosauroids such as Sirindhorna khoratensis (Shibata et al. 2015) or Protohadros byrdi (Head 1998), but as in hadrosaurids (Prieto-Márquez 2010b) and a few hadrosaurid outgroups like Penelopognathus weishampeli (Godefroit et al. 2005) and Telmatosaurus transsylvanicus (Weishampel et al. 1993), there is no prominent longitudinal ridge on the medial surface of the ascending ramus of the coronoid process of F. thyrakolasus gen. et sp. nov.
The lateral surface of the dentary near the base of the coronoid process is substantially expanded laterally in IPS-36338, so that the angle between the lateral surface of the dentary and that of the region posteroventral to the coronoid process is 150°. This angle is also less than 165° in hadrosaurids (Prieto-Márquez 2010b) and hadrosauroids Penelopognathus weishampeli (Godefroit et al. 2005) and Telmatosaurus transsylvanicus (Weishampel et al. 1993). Other early diverging hadrosauroids like Probactrosaurus gobiensis (Norman 2002) or Eolambia caroljonesa (McDonald et al. 2012) feature greater angles indicative of less expanded lateral surfaces near the base of the coronoid process. The coronoid process is well offset laterally from the lateral surface of the mandibular ramus, so that there is a well-developed concave platform separating the dental battery from the base of the coronoid process, as in hadrosaurids (Prieto-Márquez 2010b) and some hadrosaurid outgroups like Bactrosaurus johnsoni (Prieto-Márquez 2011) or Jeyawati rugoculus (McDonald et al. 2010). A deep and large adductor fossa exists between the medial surface of the ascending ramus of the coronoid process and the posterior extent of the dental battery. The base of this fossa connects medioventrally with a deep sulcus the extends adjacent to the ventral margin of the dentary. This sulcus wedges gradually anteriorly until disappearing into the ventral margin of the dentary past mid-length of the dental battery (Fig. 2A5).

Fig. 2. Dentary of the hadrosauroid dinosaur Fylax
thyrakolasus gen. et sp. nov. (IPS-36338, holotype) from the
uppermost Maastrichtian Fontllonga-R locality; in posterior (A1),
medial (A2), dorsal (A4), anterior (A5),
lateral (A6), and ventral
(A7) views. A detailed
lingual view of the tooth crowns appears in A3.
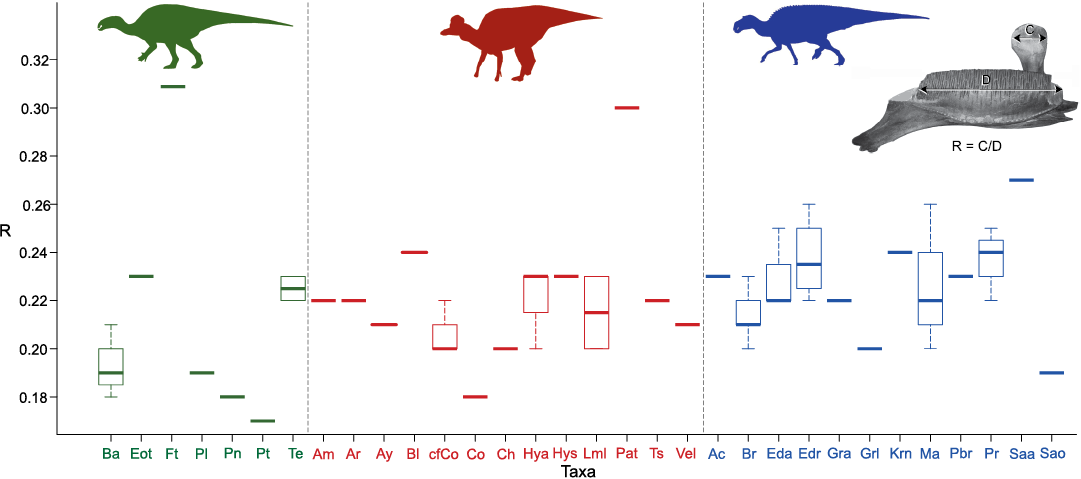
Fig. 3. Distribution of the ratio between the maximum width of the dorsal region of the coronoid process (C) and the length of the dental battery (D) in a sample of hadrosauroid dinosaurs. Taxon abbreviations: Ac, Acristavus gagslarsoni; Am, Amurosaurus riabinini; Ar, Aralosaurus tuberiferus; Ay, Arenysaurus ardevoli; Ba, Bactrosaurus johnsoni; Bl, Blasisaurus canudoi; Br, Brachylophosaurus canadensis; cfCo, cf. Corythosaurus sp.; Ch, Charonosaurus jiayinensis; Co, Corythosaurus sp.; Eda, Edmontosaurus annectens; Edr, Edmontosaurus regalis; Eot, Eotrachodon orientalis; Ft, Fylax thyrakolasus; Gra, ?Gryposaurus alsatei; Grl, Gryposaurus latidens; Hya, Hypacrosaurus altispinus; Hys, H. stebingeri; Krn, Kritosaurus navajovius; Lml, Lambeosaurus lambei; Ma, Maiasaura peeblesorum; Pat, Parasaurolophus tubicen; Pbr, Probrachylophosaurus bergei; Pl, Plesiohadros djadokhtaensis; Pn, Penelopognathus weishampeli; Pr, Prosaurolophus maximus; Pt, Protohadros byrdi; Saa, Saurolophus angustirostris; Sao, Saurolophus osborni; Te, Telmatosaurus transsylvanicus; Ts, Tsintaosaurus spinorhinus; Vel, Velafrons coahuilensis. Silhouettes were downloaded from http://phylopic.org and drawn by Pete Buchholz (https://creativecommons.org/licenses/by-sa/3.0/), Scott Hartman (https://creativecommons.org/licenses/by-nc-sa/3.0/) and Craig Dylke (https://creativecommons.org/publicdomain/zero/1.0/).
The incompletely preserved anterior end of the dentary is ventrally deflected, forming 13.5° with the long axis of the dental battery in medial view. The point of inflexion between the relatively straight ventral margin of the mandibular ramus and the start of the deflection occurs anteriorly past the mid-length of the dentary, precisely at 72% of the length of the dental battery. Because the symphyseal process is not preserved, it is uncertain whether this region of the dentary was further curved ventrally. Anterior to the first alveolar sulcus, the dorsal margin of the dentary abruptly slopes ventrally, indicating a relatively short (less than 20% of the length of the dental battery) proximal edentulous margin preceding the predentary, as in most early diverging hadrosauroids like Bactrosaurus johnsoni (Prieto-Márquez 2011), Gilmoreosaurus mongoliensis (Prieto-Márquez and Norell 2010) or Tethyshadros insularis (Dalla Vecchia 2009).
The dental battery of F. thyrakolasus gen. et sp. nov. contains 29 alveolar positions, of which 17 preserve teeth. There are up to three replacement teeth per alveolar sulcus at the deepest point of the dental battery. The occlusal surface displays a maximum of two functional teeth, unlike the three teeth present in hadrosaurids (Prieto-Márquez 2010b). The longitudinal axis of the dorsal margin of the dental battery is obliquely oriented, converging anteriorly with the lateral surface of the mandibular ramus (Fig. 2A5), as in most non-hadrosaurid hadrosauroids (Prieto-Márquez 2010b; Prieto-Márquez et al. 2019), except Plesiohadros djadokhtaensis (Tsogbataar et al. 2014). This is unlike the condition in Hadrosauridae, where the dorsal margin of the dental battery is parallel to the lateral surface of the mandibular ramus (Prieto-Márquez 2010b). Also unlike in hadrosaurids, but as in some early diverging hadrosauroids like Sirindhorna khoratensis (Shibata et al. 2015) and Penelopognathus weishampeli (Godefroit et al. 2005), the dental battery ends anterior to the posterior margin of the coronoid process (Fig. 2A2) (contra Casanovas et al. 1999).
Tooth crowns exhibit relatively tall diamond-shaped enameled lingual faces. The height/width ratio of the crowns ranges 2.8–3.2. No marginal denticles are observed; instead, the margins of these crowns are smooth. As in many early diverging hadrosauroids like Sirindhorna khoratensis (Shibata et al. 2015), Probactrosaurus gobiensis (Norman 2002), Bactrosaurus johnsoni (Prieto-Márquez 2011), Plesiohadros djadokhtaensis (Tsogbataar et al. 2014), or Tethyshadros insularis (Dalla Vecchia 2009), the enameled lingual surfaces of IPS-36338 feature two major ridges, one of them slightly more prominent than the other. This pair of long ridges extend apicobasally and are accompanied by a few fainter, shorter accessory ridges. The primary most prominent ridge is posteriorly offset from the midline of the crown. The occlusal surface is steeply inclined (forming less than 45° with the ascending process of the coronoid process) and flat, as in Bactrosaurus johnsoni (Prieto-Márquez 2011). This condition is reminiscent of the steep and planar occlusal surfaces of Prosaurolophus maximus and Saurolophus spp. (Erickson et al. 2012).
Remarks.—Pereda-Suberbiola et al. (2009a) noted that in the dental battery of Fylax thyrakolasus gen. et sp. nov. the distal tooth crowns are not more erupted and worn than the ones near the center of the battery. It is certainly common among hadrosauroids to display progressively more worn teeth (i.e., shallower as seen rising in labial view above the alveolar margin of the dentary) towards the posterior region of the dental battery, particularly nearing the level of the coronoid process (e.g., Bactrosaurus johnsoni, AMNH FARB 6553; Prosaurolophus maximus, MOR 447-8-20-87; Lambeosaurus lambei, TMP 81-37-1; Hypacrosaurus stebingeri, MOR 549; or Corythosaurus casuarius, ROM 858). However, the teeth of the anterior two fifths of the dental battery in IPS-36338 are missing, so it is not possible to ascertain whether the extent of eruption and wear would remain more or less constant throughout the dental battery, or gradually more worn posteriorly as in other hadrosauroids.
Stratigraphic and geographic range.—Type locality and horizon only.
Phylogenetic relationships of Fylax thyrakolasus
The phylogenetic position of the Fylax thyrakolasus gen. et sp. nov. was inferred using parsimony. The character/taxon matrix has been updated from that of Prieto-Márquez et al. (2019) and consisted of 283 equally weighted discrete morphological characters (201 cranial and 82 postcranial; see SOM 3 and 4). Multistate characters containing states that are not mutually exclusive, following a natural morphocline, were ordered. The taxonomic sample consisted of 68 species, including 23 lambeosaurine and 23 saurolophine hadrosaurids. The updated matrix differs from that of Prieto-Márquez et al. (2019) in containing a greater number of hadrosauroid outgroups given the focus of the present study on members of that evolutionary grade. The current matrix sees also the addition and revision of some of the dental and dentary characters used by Prieto-Márquez et al. (2019). In particular, characters 5 (maximum number of ridges on the enameled lingual side of dentary tooth crowns), 30 (angle between the ascending ramus of the coronoid process and the dorsal margin of the alveolar sulci of the dental battery; SOM 2), and 31 (expansion of the dorsal region of the coronoid process relative to the minimum width of the ascending ramus; SOM 1) have been revised. Added characters include the orientation (character 10) and morphology (character 11) of the occlusal surface of the dentary dental battery, based on the work by Erickson et al. (2012) and Erickson and Zelenitsky (2014), and character 40 (ratio between the anteroposterior width of the dorsal region of the coronoid process and the length of the dental battery; Fig. 3).
The tree search was conducted in TNT version 1.5 (Goloboff and Catalano 2016). A heuristic search of 10 000 replicates using random additional sequences was performed, followed by branch swapping by tree-bisection-reconnection holding ten trees per replicate. Bremer support (Bremer 1988) was assessed by computing decay indices (Donoghue et al. 1992) using TNT. Bootstrap proportions (Felsenstein 1985) were also calculated using TNT, setting the analysis for 5000 replicates using heuristic searches, in which each search was conducted via random additional sequences with branch-swapping by subtree pruning and regrafting and 25 replicates.
The analysis resulted in 79 most parsimonious trees of 1128 steps each (C.I: 0.42, R.I: 0.77). The best score was hit 1204 times out of the 10 000 replicates. In agreement with previous studies (Casanovas et al. 1999; Pereda-Suberbiola et al. 2009a; Blanco et al. 2015), F. thyrakolasus gen. et sp. nov. is positioned as a close outgroup to Hadrosauridae (Fig. 4, SOM 5). The hadrosauroid from Fontllonga-R is excluded from hadrosaurids by lacking more than 30 tooth positions in the dentary, having an occlusal surface of the dentary dental battery consisting of less than three teeth per position at the maximum (middle) breadth of the battery, possessing more than a single long major ridge on the enameled side of dentary tooth crowns (with the primary ridge not occupying a median position), lacking a sufficiently expanded dorsal region of the coronoid process (i.e., at least 1.5 times wider than the minimum width of the ascending ramus of the process), the lack of a longer proximal edentulous slope of the dentary (20% or more of the length of the dental battery), and displaying a dental battery that does not reach the posterior margin of the coronoid process.
Notably, F. thyrakolasus gen. et sp. nov. forms a clade with the hadrosauroid Tethyshadros insularis from the late Campanian–early Maastrichtian Adriatic-Dinaric Carbonate Platform (Fig. 4). Their sister relationship is unambiguously supported by the possession of a coronoid process inclined anteriorly less than 80° (independently evolved in Saurolophidae) and marginal denticles that are at least extremely reduced (also independently acquired in Saurolophidae, with reversals to papillae in Gryposaurus latidens, Acristavus gagslarsoni, and various lambeosaurines).
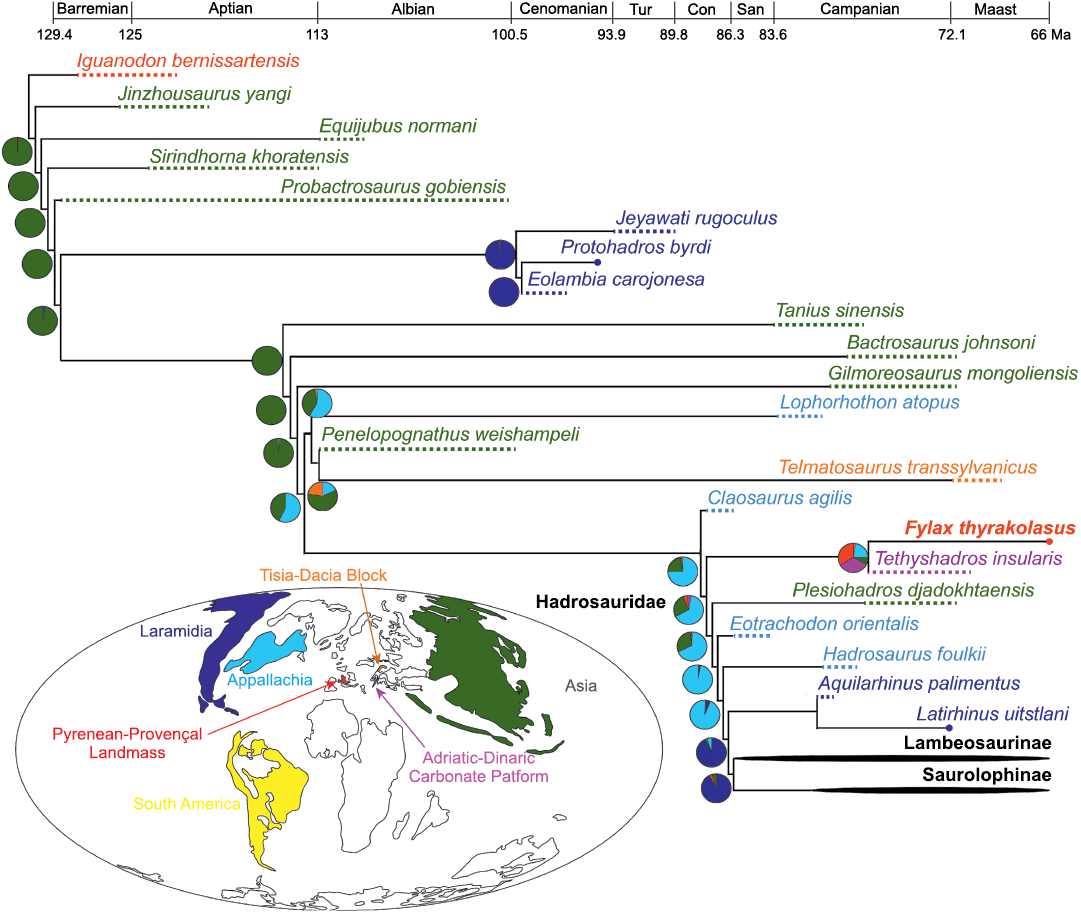
Fig. 4. Time calibrated cladogram based on the strict consensus tree (unresolved branches in the consensus are within saurolophine and lambeosaurine hadrosaurids, only shown in the SOM 5) resulting from the parsimony analysis of 67 hadrosauroid taxa, showing the position of IPS-36338. Ancestral areas are reconstructed for each clade of the phylogram, with maximum likelihood proportions indicated by the pie charts. Colors represent the various areas considered in the maximum likelihood ancestral state reconstruction analysis. The global paleogeographic map corresponds to the late Campanian (75–73.8 Ma) and is based on Scotese (2014), except the paleogeographic configuration of the European Archipelago, which is based on the greater detailed offered by Csiki et al. (2015: fig. 3). Geochronological ages are from Walker et al. (2018). The Pyrenean-Provençal Landmass presented in the figure will become part of the Ibero-Armorican Island in the Maastrichtian. Abbreviations: Con, Coniacian; Maast, Maastrichtian; San, Santonian; Tur, Turonian.
Historical biogeography of Fylax thyrakolasus
Ancestral areas for each node of the resulting most parsimonious trees were reconstructed using the maximum likelihood ancestral state reconstruction algorithm in Mesquite version 3.51 (Maddison and Maddison 2018). The areas under consideration were those that harbored the 67 species included in the phylogenetic analysis. These areas correspond to a base paleomap integrating the 73.8 Ma late Campanian global paleogeography of Scotese (2014) (Asia, Appalachia, Laramidia, and South America) and the ca. 75 Ma late Campanian paleogeography of the European Archipelago shown in Csiki et al. (2015) (the Pyrenean-Provençal Landmass, that in the Maastrichtian would become the northern region of the Ibero-Armorican Island, the Adriatic-Dinaric Carbonate Platform, and the Tisia-Dacia Block). A step matrix was created to account for the distance among those seven areas. The number of steps between two areas indicated the minimum number of landmasses (according to the paleogeographic configuration in Fig. 4) that a species would need to cross to travel from one area to another.
The sister relationship between Fylax thyrakolasus gen. et sp. nov. and Tethyshadros insularis indicates the existence of a clade of basally branching hadrosauroids during the late Campanian through the Maastrichtian of the European Archipelago. The split between these two species took place no later than the late Campanian. It is equally likely that the ancestral area of the clade was either the Pyrenean-Provençal Landmass (fated to become the northern Ibero-Armorican island during the Maastrichtian) or the Adriatic-Dinaric Carbonate Platform (both scenarios with a 33% likelihood proportion; Fig. 4, SOM 6). In the first scenario, F. thyrakolasus gen. et sp. nov. would have become the last or one of the last survivors of this clade in the Ibero-Armorican island; in the second scenario, the species from Fontllonga-R would have dispersed from the Adriatic-Dinaric Carbonate Platform into the Pyrenean-Provençal Landmass no later than the late Campanian–early Maastrichtian (Fig. 4). A less likely, yet still possible scenario is that the ancestral area of this clade would have been located in Appalachia (22% likelihood proportion; Fig. 4, SOM 6), suggesting a dispersal event from this continental landmass into the European Archipelago no later than the late Campanian.
At a deeper, more inclusive level in the phylogeny, the ancestry of these European hadrosauroids can be traced back to North America, specifically Appalachia. The most recent common ancestor of the Fylax–Tethyshadros clade and the clade including Hadrosauridae and its most closely related outgroups is likely to have lived in Appalachia (with a likelihood proportion in our results of 67%; Fig. 4, SOM 6).
Discussion
A late surviving non-hadrosaurid hadrosauroid.—Fylax thyrakolasus gen. et sp. nov. is the stratigraphically youngest non-hadrosaurid hadrosauroid species known to date, having survived to the end of the Cretaceous. All other known hadrosaurid outgroups are older, spanning taxa from the Early Cretaceous such as Probactrosaurus gobiensis (Norman 2002), Equijubus normani (McDonald et al. 2014), Sirindhorna khoratensi (Shibata et al. 2015), and also species from the early Maastrichtian like Telmatosaurus transsylvanicus (Bojar et al. 2011), and possibly Tethyshadros insularis (Dalla Vecchia 2020) (Fig. 4). Other latest Maastrichtian hadrosauroid species recorded in Europe (Fondevilla et al. 2019: fig. 11) are morphologically derived members of Hadrosauridae, such as lambeosaurines Pararhabdodon isonensis from the Talarn Formation of the eastern Tremp syncline (Lleida Province, northeastern Spain; Prieto-Márquez et al. 2019) and Canardia garonnensis from the Marnes d’Auzas Formation of the Aurignac anticline (southwest of Toulouse, southern France; Prieto-Márquez et al. 2013), as well as the indeterminate hadrosaurid from the lower and middle part of the Tremp Formation cropping out in the northern flank of the Tremp Syncline (Huesca Province, northeastern Spain; Company et al. 2015) and the partial dentary (MPZ2010/952) from an indeterminate hadrosaurid described by Cruzado-Caballero et al. (2014) from the Blasi 3.4 locality of the upper Maastrichtian La Posa Formation (Huesca Province, northeastern Spain).
Company et al. (1998) and Pereda-Suberbiola et al. (2009a) reported on dentary and dental remains of what they regarded as a basal hadrosaurid from upper Maastrichtian strata of the Sierra Perenchiza Formation cropping out at the locality of La Solana, Chera Basin, eastern Spain. This unnamed taxon shares with F. thyrakolasus gen. et sp. nov. dentary teeth with a pair of apicobasally long ridges on the enameled surface and crowns with height/width ratios within the 2.8–3.2 range, as well as a coronoid process of the dentary lacking a ridge on the posteromedial surface. However, the form from La Solana differs from F. thyrakolasus gen. et sp. nov. in that at least one of the dentaries (MPV 181) possesses 36–37 tooth positions (Pereda-Suberbiola et al. 2009a), the dental battery extends posterior to the level of the coronoid process (Company et al. 1998), the coronoid process is subvertically oriented and has a dorsal region that is less than 30% of the length of the dental battery (e.g., MGUV 2200), and dentary teeth display marginal denticles (Company et al. 1998). The relatively high number of tooth positions (over 30) and the dental battery extending posterior to the coronoid process support the referral of this species to Hadrosauridae. In the vast majority of hadrosaurids the coronoid process is oriented anteriorly (Prieto-Márquez 2010b) but it has occasionally been reported to be subvertically to vertically oriented (e.g., in the saurolophine Kritosaurus navajovius, according to Prieto-Márquez 2014). The presence of two major pair of long ridges on the enameled surface of dentary teeth is a condition absent in all hadrosaurids, except the lambeosaurine from the lower Maastrichtian Basturs Poble locality of the southern Pyrenees of northeastern Spain (Fondevilla et al. 2018). Thus, the presence of this pair of ridges in itself does not necessarily preclude inclusion of the species from La Solana within Hadrosauridae. Overall, the anatomical evidence supports Company et al. (1998) and Pereda-Suberbiola et al. (2009a) in regarding the form from La Solana as an indeterminate hadrosaurid, leaving F. thyrakolasus gen. et sp. nov. as the only known late Maastrichtian non-hadrosaurid hadrosauroid recorded so far.
The role of the European Archipelago in historical biogeography.—Although it is not the goal of this study to delve deeply into hadrosauroid biogeography, there are some ancillary results of our analysis that are worth discussing. At a broader level, our inference of ancestral areas on the temporally calibrated cladogram (Fig. 4) supports previous studies that postulated an Asian origin for hadrosauroids (Wiman 1929; Rozhdestvensky 1967; Brett-Surman 1979; Milner and Norman 1984; You et al. 2003; Godefroit et al. 2008). After originating in Asia during the Early Cretaceous, hadrosauroids are reconstructed here to have diversified in this continent for most of their pre-hadrosaurid evolution, with the exception of the dispersal to Laramidia no later than the early Cenomanian leading to the Eolambia–Jeyawati clade (Fig. 4). The present analysis also supports the hypothesis that subsequently, during the Coniacian–early Campanian, much of the branching sequence of cladistic events leading to Hadrosauridae appears to have occurred in Appalachia (Fig. 4), as hypothesized by Prieto-Márquez (2010a, 2016). In this context, the European Archipelago may have played an important role in providing a pathway for the dispersal of hadrosauroids from Asia to Appalachia. European species Telmatosaurus transsylvanicus, Tethyshadros insularis, and Fylax thyrakolasus gen. et sp. nov. might be representatives of that grade of hadrosaurid outgroups that colonized Appalachia.
Conclusions
The dentary IPS-36338, from the Fontllonga Formation of the Àger syncline in the southern Pyrenees of northeastern Spain, represents a new genus and species of non-hadrosaurid hadrosauroid, Fylax thyrakolasus gen. et sp. nov. This species is diagnosed on the basis of a combination of dental and dentary characters, the most remarkable of which is the great development of the dorsal region of the coronoid process and the flat high-angled occlusal surface of the dental battery. This steep and flat occlusal surface likely indicates a slicing mode of food processing.
Phylogenetically, F. thyrakolasus gen. et sp. nov. is positioned as sister to the Italian hadrosauroid Tethyshadros insularis. The ancestral area of this clade is ambiguous, most likely originating either in the Pyrenean-Provençal Landmass or the Adriatic-Dinaric Carbonate Platform of the Late Cretaceous European Archipelago, no later than the late Campanian. Ancestrally, the Fylax–Tethyshadros clade might have had deeper Appalachian roots. This study supports an Appalachian origin for hadrosaurids and the possibility that the Late Cretaceous European Archipelago served as a pathway for dispersals from Asia to Appalachia of hadrosaurid outgroups.
Acknowledgements
We thank Àngel Galobart (IPS) for informing on the occurrence of IPS-36338 and Fabio Dalla Vecchia (IPS) for providing details on the dentition of Tethyshadros insularis. We are also grateful for the comments and advice provided by Andrew McDonald (Western Science Center, Hemet, USA) and José Ignacio Canudo (Universidad de Zaragoza, Zaragoza, Spain), which improved the quality of the manuscript. This work was supported by the Ministry of Economy, Industry and Competitivity of the Government of Spain via the Ramón y Cajal Program [RyC-2015-17388] and grant [CGL2016-73230-P]. Additional support was provided by the CERCA Programme of the Generalitat de Catalunya, as well as grant CLT009/18/00067 (Paleoecological Networks of the Dinosaur Localities from the Catalan Cretaceous) from the Departament de Cultura of the Generalitat de Catalunya, and EDUCAFIN (Talentos de Exportación) from the Government of the State of Guanajuato, Mexico.
References
Blanco, A., Prieto-Márquez, A., and De Esteban-Trivigno, S. 2015. Diversity of hadrosauroid dinosaurs from the Late Cretaceous Ibero-Armorican Island (European Archipelago) assessed from dentary morphology. Cretaceous Research 56: 447–457. Crossref
Bilotte, M., Laurent, Y., and Téodori, D. 2010. Restes d’Hadrosaure dans le Crétacé terminal marin de Larcan (Petites Pyrénées, Haute-Garonne, France). Carnets de Géologie 2010/02: CG2010_L02. Crossref
Bojar, A.V., Halas, S., Bojar, H.-P., Grigorescu, D., and Vasile, Ş. 2011. Upper Cretaceous volcanoclastic deposits from the Haţeg Basin, South Carpathians (Romania): K-Ar ages and intrabasinal correlation. Geochronometria 38: 182–188. Crossref
Bremer, K. 1988. The limits of amino acid sequence data in angiosperm phylogenetic reconstruction. Evolution 42: 795–803. Crossref
Brett-Surman, M.K. 1979. Phylogeny and palaeobiogeography of hadrosaurian dinosaurs. Nature 277: 560–562. Crossref
Caus, E., Frijia, G., Parente, M., Robles-Salcedo, R., and Villalonga, R. 2016. Constraining the age of the last marine sediments in the Late Cretaceous of central South Pyrenees (NE Spain): Insights from larger benthic foraminife ra and strontium isotope stratigraphy. Cretaceous Research 57: 402–413. Crossref
Casanovas-Cladellas, M.L., Santafé-Llopis, J.V., and Isidro-Llorens, A. 1993. Pararhabdodon isonense n. gen. n. sp. (Dinosauria). Estudio morfológico, radiotomográfico y consideraciones biomecánicas. Paleontologia i Evolució 26–27: 121–131.
Casanovas, M.L., Pereda Suberbiola, X.P., Santafé, J.V., and Weishampel, D.B. 1999. A primitive euhadrosaurian dinosaur from the uppermost Cretaceous of the Ager syncline (southern Pyrenees, Catalonia). Geologie En Mijnbouw 78: 345–356. Crossref
Colombo, F. and Cuevas, J.L. 1993. Características estratigráficas y sedimentológicas del “Garumniense” en el sector de Ager (Pre-Pirineo, Lleida). Acta Geologica Hispanica 28: 15–32.
Company, J., Cruzado Caballero, P., and Canudo, J.I. 2015. Presence of diminutive hadrosaurids (Dinosauria: Ornithopoda) in the Maastrichtian of the south-central Pyrenees (Spain). Journal of Iberian Geology 41: 71–81. Crossref
Company, J., Galobart, A., and Gaete, R. 1998. First data on the hadrosaurid dinosaurs (Ornithischia, Dinosauria) from the Upper Cretaceous of Valencia, Spain. Oryctos 1: 121–126.
Cruzado-Caballero, P., Pereda-Suberbiola, X., and Ruiz-Omeñaca, J.I. 2010. Blasisaurus canudoi gen. et sp. nov., a new lambeosaurine dinosaur Hadrosauridae from the latest Cretaceous of Arén Huesca, Spain. Canadian Journal of Earth Sciences 4712: 1507–1517. Crossref
Cruzado-Caballero, P., Ruiz-Omeñaca, J.I., Gaete, R., Riera, V., Oms, O., and Canudo, J.I. 2014. A new hadrosaurid dentary from the latest Maastrichtian of the Pyrenees (north Spain) and the high diversity of the duck-billed dinosaurs of the Ibero-Armorican Realm at the very end of the Cretaceous. Historical Biology 26: 619–630. Crossref
Csiki-Sava, Z., Buffetaut, E., Ösi, A., Pereda-Suberbiola, A., and Brusatte, S.L. 2015. Island life in the Cretaceous—faunal composition, biogeography, evolution, and extinction of land-living vertebrates on the Late Cretaceous European archipelago. ZooKeys 469: 1–161. Crossref
Cuevas, J.L. 1992. Estratigrafía del” Garumniense” de la Conca de Tremp. Prepirineo de Lérida. Acta Geológica Hispánica 27: 95–108.
Dalla Vecchia, F.M. 2006. Telmatosaurus and the other hadrosaurids of the Cretaceous European Archipelago. An overview. Natura Nascosta 32: 1–55.
Dalla Vecchia, F.M. 2009. Tethyshadros insularis, a new hadrosauroid dinosaur (Ornithischia) from the Upper Cretaceous of Italy. Journal of Vertebrate Paleontology 29: 1100–1116. Crossref
Dalla Vecchia, F.M. 2014. An overview of the latest Cretaceous hadrosauroid record in Europe. In: D.A. Eberth and D.C. Evans (eds.), Hadrosaurs, 268–297. Indiana University Press, Bloomington.
Dalla Vecchia, F.M. 2020. The unusual tail of Tethyshadros insularis (Dinosauria, Hadrosauroidea) from the Adriatic Island of the European Archipelago. Rivista Italiana di Paleontologia e Stratigrafia 126: 583–628.
Dalla Vecchia, F.M., Gaete, R., Riera, V., Oms, O., Prieto-Márquez, A., Vila, B., Sellés, A.G., and Galobart, A. 2014. The hadrosauroid record in the Maastrichtian of the eastern Tremp Syncline (northern Spain). In: D.A. Eberth and D.C. Evans (eds.), Hadrosaurs, 298–314. Indiana University Press, Bloomington.
Dollo, L. 1888. Iguanodontidae et Camptosauridae. Comptes Rendus de l’Académie des Sciences 106: 775–777.
Donoghue, M.J., Olmstead, R.G., Smith, J.F., and Palmer, J.D. 1992. Phylogenetic relationships of Dipsacales based on rbcL sequences. Annals of the Missouri Botanical Garden 79: 672–685. Crossref
Erickson, G.M. and Zelenitsky, D.K. 2014. Osteohistology and occlusal morphology of Hypacrosaurus stebingeri teeth throughout ontogeny with comments on wear-induced form and function. In: D.A. Eberth and D.C. Evans (eds.), Hadrosaurs, 422–432. Indiana University Press, Bloomington.
Erickson, G.M., Krick, B.A., Hamilton, M., Bourne, G.R., Norell, M.A., Lilleodden, E., and Sawyer, W.G. 2012. Complex dental structure and wear biomechanics in hadrosaurid dinosaurs. Science 338: 98–101. Crossref
Felsenstein, J., 1985. Confidence limits on phylogenies: an approach using the bootstrap. Evolution 39: 783–791. Crossref
Fondevilla, V., Dalla Vecchia, F.M., Gaete, R., Galobart, À., Moncunill-Solé, B., and Köhler, M. 2018. Ontogeny and taxonomy of the hadrosaur (Dinosauria, Ornithopoda) remains from Basturs Poble bonebed (late early Maastrichtian, Tremp Syncline, Spain). PLoS ONE 12: e0206287. Crossref
Fondevilla, V., Riera, V., Vila, B., Sellés, A.G., Dinarès-Turell, J., Vicens, E., Gaete, R., Oms, O., and Galobart, À. 2019. Chronostratigraphic synthesis of the latest Cretaceous dinosaur turnover in south-western Europe. Earth-Science Reviews 191: 168–189. Crossref
Galbrun, B., Feist, M., Colombo, F., Rocchia, R., and Tambareau, Y. 1993. Magnetostratigraphy and biostratigraphy of Cretaceous–Tertiary continental deposits, Ager basin, province of Lerida, Spain. Palaeogeography, Palaeoclimatology, Palaeoecology 102: 41–52. Crossref
Godefroit, P., Li, H., and Shang, C.-Y. 2005. A new primitive hadrosauroid dinosaur from the Early Cretaceous of Inner Mongolia (P.R. China). Comptes Rendus Palevol 4: 697–705. Crossref
Godefroit, P., Shulin, H., Tinxiang, Y., and Lauters, P. 2008. New hadrosaurid dinosaurs from the uppermost Cretaceous of northeastern China. Acta Palaeontologica Polonica 53: 47–74. Crossref
Goloboff, P.A. and Catalano, S.A. 2016. TNT version 1.5, including a full implementation of phylogenetic morphometrics. Cladistics 3: 221–238. Crossref
Head, J.J. 1998. A new species of basal hadrosaurid (Dinosauria, Ornithischia) from the Cenomanian of Texas. Journal of Vertebrate Paleontology 18: 718–738. Crossref
Hopson, J.A. 1975. The evolution of cranial display structures in hadrosaurian dinosaurs. Paleobiology 1: 21–43. Crossref
Laurent, Y., Bilotte, M., and Le Loeuff, J., 2002. Late Maastrichtian continental vertebrates from southwestern France: correlation with marine fauna. Palaeogeography, Palaeoclimatology, Palaeoecology 187: 121–135. Crossref
López-Martinez, N., Ardevol, L., Arribas, M.E., Civis, J., and Gonzalez-Delgado, A. 1998. The geological record in non-marine environments around the K/T boundary (Tremp Formation, Spain). Bulletin de la Société géologique de France 169: 11–20.
López-Martínez, N., Fernández-Marron, M.T., and Valle M.F. 1999. The succession of vertebrates and plants across the Cretaceous–Tertiary boundary in the Tremp Formation, Ager valley (South-central Pyrenees Spain). Geobios 32: 617–627. Crossref
Maddison, W. P. and Maddison, D. R. 2018. Mesquite: a Modular System for Evolutionary Analysis. Version 3.51. http://www.mesquiteproject.org.
Marsh, O.C. 1881. Principal characters of American Jurassic dinosaurs. Part IV. American Journal of Science 21: 417–423. Crossref
McDonald, A.T., Bird, J., Kirkland, J.I., and Dodson, P. 2012. Osteology of the basal hadrosauroid Eolambia caroljonesa (Dinosauria: Ornithopoda) from the Cedar Mountain Formation of Utah. PLoS ONE 7(10): e45712.
McDonald, A.T., Maidment, S.C.R., Barrett, P.M., You, H., and Dodson, P. 2014. Osteology of the basal hadrosauroid Equijubus normani (Dinosauria, Ornithopoda) from the Early Cretaceous of China. In: D.A. Eberth and D.C. Evans (eds.), Hadrosaurs, 44–72. Indiana University Press, Bloomington.
McDonald, A.T., Wolfe, D.G., and Kirkland, J.I. 2010. A new basal hadrosauroid (Dinosauria: Ornithopoda) from the Turonian of New Mexico. Journal of Vertebrate Paleontology 30: 799–812. Crossref
Milner, A.R. and Norman, D.B. 1984. The biogeography of advanced ornithopods dinosaurs (Archosauria: Ornithischia)—a cladistic-vicariance model. In: W.E. Reif and F. Westphal (eds.), Third Symposium on Mesozoic Terrestrial Ecosystems, Short Papers, 145–150. Attempto Verlag, Tubingen.
Norman, D.B. 2002. On Asian ornithopods (Dinosauria: Ornithischia). Probactrosaurus Rozhdestvensky, 1966. Zoological Journal of the Linnean Society 136: 113–144. Crossref
Oms, O., Fondevilla, V., Riera, V., Marmi, J., Vicens, E., Estrada, R., Anadón, P., Vila, B., and Galobart, À. 2016. Transitional environments of the lower Maastrichtian South-Pyrenean Basin (Catalonia, Spain): the Fumanya Member tidal flat. Cretaceous Research 57: 428–442. Crossref
Ostrom, J.H. 1962. The cranial crests of hadrosaurian dinosaurs. Postilla 62: 1–29.
Owen, R. 1842. Report on British fossil reptiles. Part 2. Report of the British Association for the Advancement of Science 11: 60–204.
Pereda-Suberbiola, X., Canudo, J.I., Company, J., Cruzado-Caballero, P., and Ruiz-Omeñaca, J.I. 2009a. Hadrosauroid dinosaurs from the latest Cretaceous of the Iberian Peninsula. Journal of Vertebrate Paleontology 29: 946–951. Crossref
Pereda-Suberbiola, X., Canudo, J.I., Cruzado-Caballero, P., Barco, J.L., López-Martínez, N., Oms, O., and Ruiz-Omeñaca, J. I. 2009b. The last hadrosaurid dinosaurs of Europe: a new lambeosaurine from the uppermost Cretaceous of Arén (Huesca, Spain). Comptes Rendus Palevol 8: 559–572. Crossref
Prieto-Márquez, A. 2010a. Global historical biogeography of hadrosaurid dinosaurs. Zoological Journal of the Linnean Society 159: 503–525. Crossref
Prieto-Márquez, A. 2010b. Global phylogeny of Hadrosauridae (Dinosauria: Ornithopoda) using parsimony and Bayesian methods. Zoological Journal of the Linnean Society 159: 435–502. Crossref
Prieto-Márquez, A. 2011. Cranial and appendicular ontogeny of Bactrosaurus johnsoni, a hadrosauroid dinosaur from the Late Cretaceous of northern China. Palaeontology 54: 773–792. Crossref
Prieto-Márquez, A. 2016. A primitive hadrosaurid from southeastern North America and the origin and early evolution of “duck-billed” dinosaurs. Journal of Vertebrate Paleontology 36 (2): e1054495. Crossref
Prieto-Márquez, A. and Norell, M.A. 2010.
Anatomy and relationships of Gilmoreosaurus
mongoliensis (Dinosauria: Hadrosauroidea) from the Late
Cretaceous of Central Asia. American
Museum Novitates 3694: 1–49. Crossref
Prieto-Márquez, A., Dalla Vecchia, F.M., Gaete, R., and Galobart, A. 2013. Diversity, relationships, and biogeography of the lambeosaurine dinosaurs from the European Archipelago, with description of the new aralosaurin Canardia garonnensis. PLoS ONE 8 (7): e69835. Crossref
Prieto-Márquez, A., Fondevilla, V., Sellés, A.G., Wagner, J.R., and Galobart, À. 2019. Adynomosaurus arcanus, a new lambeosaurine dinosaur from the Late Cretaceous Ibero-Armorican Island of the European archipelago. Cretaceous Research 96: 19–37. Crossref
Riera, V., Oms, O., Gaete, R., and Galobart, À. 2009. The end-Cretaceous dinosaur succession in Europe: the Tremp Basin record (Spain). Palaeogeography, Palaeoclimatology, Palaeoecology 283: 160–171. Crossref
Rosell, J., Linares, R., and Llompart, C. 2001. El “Garumniense” prepirenaico. Revista de la Sociedad Geológica de España 14: 47–56.
Rozhdestvensky, A.K. 1967. New iguanodonts from central Asia. International Geology Review 9: 556–566. Crossref
Scotese, C.R. 2014. Atlas of Late Cretaceous Maps, PALEOMAP Atlas for ArcGIS, Volume 2, The Cretaceous, Maps 16–22. Mollweide Projection, PALEOMAP Project, Evanston, IL.
Seeley, H.G. 1887. On the classification of the fossil animals commonly named Dinosauria. Proceedings of the Royal Society of London 43: 165–171. Crossref
Sellés, A.G. and Vila, B. 2015. Re-evaluation of the age of some dinosaur localities from the southern Pyrenees by means of megaloolithid oospecies. Journal of Iberian Geology 41: 125–139. Crossref
Sereno, P.C. 1986. Phylogeny of the bird-hipped dinosaurs (Order Ornithischia). National Geographic Research 2: 234–256.
Sereno, P.C. 1998. A rationale for phylogenetic definitions, with application to higher-level taxonomy of Dinosauria. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 210: 41–83. Crossref
Shibata, M., Jintasakul, P., Azuma, Y., and You, H.-L. 2015. A new basal hadrosauroid dinosaur from the Lower Cretaceous Khok Kruat Formation in Nakhon Ratchasima Province, northeastern Thailand. PLoS ONE 10 (12): e0145904. Crossref
Tsogtbaatar, K., Weishampel, D.B., Evans, D.C., and Watabe, M. 2014. A new hadrosauroid (Plesiohadros djadokhtaensis) from the Late Cretaceous Djadokhtan Fauna of southern Mongolia. In: D.A. Eberth and D.C. Evans (eds.), Hadrosaurs, 108–135. Indiana University Press, Bloomington.
Walker, J.D., Geissman, J.W., Bowring, S.A., and Babcock, L.E. 2018. Geologic Time Scale v. 5.0. Geological Society of America, https://doi.org/10.1130/2018.CTS005R3C. Crossref
Weishampel, D.B., Norman, D.B., and Grigorescu, D. 1993. Telmatosaurus transsylvanicus from the Late Cretaceous of Romania: the most basal hadrosaurid dinosaur. Palaeontology 36: 361–385.
Wiman, C. 1929. Die Kreide. Dinosaurier aus Shantung. Palaeontologia Sinica, Series C 6: 1–67.
You, H.-L., Luo, Z., Shubin, N.H., Witmer, L.M., Tang, Z.-L., and Tang, F. 2003. The earliest-known duck-billed dinosaur from deposits of late Early Cretaceous age in northwestern China and hadrosaur evolution. Cretaceous Research 24: 347–355. Crossref
Acta Palaeontol. Pol. 66 (2): 425–435, 2021
https://doi.org/10.4202/app.00821.2020