
The early Miocene lake of Foieta la Sarra-A in eastern Iberian Peninsula and its relevance for the reconstruction of the Ribesalbes–Alcora Basin palaeoecology
SERGIO ÁLVAREZ-PARRA, JOAQUÍN ALBESA, SOLEDAD GOUIRIC-CAVALLI, PLINI MONTOYA, ENRIQUE PEÑALVER, JOSEP SANJUAN, and VICENTE D. CRESPO
Álvarez-Parra, S., Albesa, J., Gouiric-Cavalli, S., Montoya, P., Peñalver, E., Sanjuan, J., and Crespo, V.D. 2021. The early Miocene lake of Foieta la Sarra-A in eastern Iberian Peninsula and its relevance for the reconstruction of the Ribesalbes–Alcora Basin palaeoecology. Acta Palaeontologica Polonica 66 (Supplement to 3): 13–30.
The Ribesalbes–Alcora Basin (Castelló Province, Spain) contains two lower Miocene units that are rich in fossils. The Unit B contains oil-shale and laminated bituminous dolomicrite related to a palaeolake, whereas the Unit C is composed of sandstone and mudstone beds from distal deltaic and shallow lacustrine environments. The La Rinconada and San Chils localities from the Unit B have yielded a fossil assemblage of plants, molluscs, arthropods, and vertebrates, while the localities from the Unit C in the Campisano ravine (Araia/Mas d’Antolino outcrop) are rich in mammalian record. Here we study a new palaeolake deposit of laminated lacustrine limestone beds in the Unit C named Foieta la Sarra-A. This new locality has provided an assemblage of charophytes, terrestrial plants, molluscs, arthropods, and teleosteans. The latter represent the only known fish record from the Ribesalbes–Alcora Basin to date. Although the specimens are generally poorly preserved, the presence of soft-body preservation due to the action of microbial mats at the lake bottom allows considering the Foieta la Sarra-A locality as a Konservat-Lagerstätte. The Foieta la Sarra-A palaeolake had a different water chemistry compared to that represented in the Unit B. Its depth was about a few metres and the water level suffered periodic fluctuations. This new locality sheds light on the palaeoenvironmental dynamics of the Ribesalbes–Alcora Basin during the early Miocene and provides a new approach to the palaeoecological reconstruction of the basin.
Key words: Characeae, Poales, Gastropoda, Cladocera, Insecta, Teleostei, palaeoenvironment, taphonomy, palaeoecology, lacustrine basin, Neogene, Konservat-Lagerstätte, Spain.
Sergio Álvarez-Parra [sergio.alvarez-parra@ub.edu], Departament de Dinàmica de la Terra i de l’Oceà and Institut de Recerca de la Biodiversitat (IRBio), Facultat de Ciències de la Terra, Universitat de Barcelona, c/ Martí i Franquès s/n, 08028, Barcelona, Spain.
Joaquín Albesa [joaquin.albesa@uv.es], Departament de Botànica i Geologia, Universitat de València, Dr. Moliner 50, 46100, Burjassot, Valencia, Spain; Museu Valencià d’Història Natural, L’Hort de Feliu, P.O. Box 8460, Alginet, 46230 Valencia, Spain.
Soledad Gouiric-Cavalli [sgouiric@fcnym.unlp.edu.ar], Consejo Nacional de Investigaciones Científicas y Técnicas, División Paleontología Vertebrados, Museo de La Plata, Paseo del Bosque s/n, B1900FWA La Plata, Argentina.
Plini Montoya [p.montoya@uv.es], Departament de Botànica i Geologia, Universitat de València, Dr. Moliner 50, 46100, Burjassot, Valencia, Spain.
Enrique Peñalver [e.penalver@igme.es], Instituto Geológico y Minero de España (Museo Geominero), c/ Cirilo Amorós 42, 46004, Valencia, Spain.
Josep Sanjuan [js76@aub.edu.lb], Department of Geology, American University of Beirut-AUB, 11-0236 Beirut, Lebanon; Departament de Dinàmica de la Terra i de l’Oceà, Facultat de Ciències de la Terra, Universitat de Barcelona, c/ Martí i Franquès s/n, 08028, Barcelona, Spain.
Vicente D. Crespo [vidacres@gmail.com] (corresponding author), Consejo Nacional de Investigaciones Científicas y Técnicas, División Paleontología Vertebrados, Museo de La Plata, Paseo del Bosque s/n, B1900FWA La Plata, Argentina; Museo Paleontológico de Alpuente, Av. San Blas 17, Alpuente, 46178 Valencia, Spain; Museu Valencià d’Història Natural, L’Hort de Feliu, P.O. Box 8460, Alginet, 46230 Valencia, Spain.
Received 15 October 2020, accepted 6 April 2021, available online 19 August 2021.
Copyright © 2021 S. Álvarez-Parra et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Lakes are remarkable ecological archives due to their high sensitivity to climatic changes, a characteristic that can also be used in deep time (Cohen 2003). Indeed, the fossil content and sedimentary record of palaeolakes provide valuable information, such as palaeoclimatic, chemical, and temperature conditions (Talbot and Allen 1996). During the Miocene, a large portion of the European continent was covered by lakes (e.g., Neubauer et al. 2015a, b; Mandic et al. 2019; Vasilyan 2020). Those which developed in the Iberian Peninsula produced exceptionally well-preserved fossil assemblages with Konservat-Lagerstätte characteristics (e.g., McNamara et al. 2012; Peñalver et al. 2016). Remarkable Iberian Konservat-Lagerstätten in Miocene lacustrine deposits are the fossil localities of Rubielos de Mora (Teruel Province), Bicorb (Valencia Province), La Rinconada and San Chils (the latter two in Castelló Province). These well-preserved and diverse fossil assemblages originated under anoxic conditions at the lake bottom with presence of microbial mats (Peñalver 2002; Peñalver and Gaudant 2010).
The Ribesalbes–Alcora Basin is a Neogene complex graben covering around 150 km2 located in SE Iberian Range (Anadón et al. 1989; Fig. 1A). This basin is well-known for the mining activities leading to oil-shale extraction in the La Rinconada (Ribesalbes) and San Chils (l’Alcora) mines until the early 20th century and clay extraction in the Campisano ravine (Araia/Mas d’Antolino area) which continues nowadays (Peñalver et al. 2016; Crespo 2017). The Campisano ravine contains seven sections with mammalian fossil record that have been proposed as a Site of Geological Interest (Costa-Pérez et al. 2019), i.e., Mas dels Coixos, Mas de Torner, Araia Cantera Sud, Barranc de Campisano, Foieta la Sarra, Mas d’Antolino B and Corral de Brisca (Fig. 1A). La Rinconada is a well-known locality that has provided an outstanding record of plants, molluscs, ostracods, arachnids, insects, amphibians, bird feathers and coprolites (Peñalver et al. 2016). Furthermore, a fossil assemblage from the San Chils locality has been recently described showing a strong similarity to the assemblage of La Rinconada (Álvarez-Parra and Peñalver 2019). Both localities have been interpreted as different areas of the same meromictic palaeolake, mostly contemporaneous, that occupied the basin during the early Miocene (Álvarez-Parra and Peñalver 2019). The exceptional preservation of their fossil record is related to the presence of microbial mats at the lake bottom (Peñalver et al. 2016). Interestingly, these fossil localities lack fish record, which might be related to an endorheic origin of the lake or an inappropriate water chemistry for the species that could reach the lake (Peñalver et al. 2016).

Fig. 1. A, B. Geographic and geological location of the Ribesalbes–Alcora Basin (Castelló Province, Valencian Community, Spain) with indication of the Campisano ravine (1–7), San Chils, and La Rinconada localities. C. Synthetic stratigraphic column of the Campisano ravine (C1), and Foieta la Sarra section (C2), showing the location of the Foieta la Sarra-1 (FS1) and the studied Foieta la Sarra-A (FSA) mammalian fossiliferous localities. D. Fossiliferous laminated slabs of laminated lacustrine limestone at the lower Miocene Foieta la Sarra-A locality. E. Detail of the fine lamination of the Foieta la Sarra-A of the Foieta la Sarra-A laminated limestone rocks. Abbreviations: ACS, Araia Cantera Sud; CBR, Corral de Brisca; BC, Barranc de Campisano; MAB, Mas d’Antolino B; MCX, Mas dels Coixos; MTR, Mas de Torner. A, B modified from Crespo (2017), Crespo et al. (2019c).
Forty-five fossiliferous levels rich in mammal remains have been recently described by Crespo et al. (2019c), in addition to the ones previously studied by Agustí et al. (1988), in the seven Campisano ravine sections. Their general features, together with the geological and preliminary biostratigraphic framework, were published by Crespo et al. (2019c), who described the geology of the area and placed the sections under study a stratigraphic context. In addition, many of the small mammal groups found in these sections have already been studied, including two new species of bat and dimylid (Furió et al. 2012; Crespo 2017; Crespo et al. 2019a, b, 2020a, b, 2021a, b). Among the different studied sections from these localities, the Foieta la Sarra section includes the Foieta la Sarra-1 locality, which yields a rich record of vertebrate remains (Crespo et al. 2019c). The faunal assemblage of the Foieta la Sarra-1 locality listed by Crespo et al. (2019c) is: Amphiperatherium frequens erkertshofense (Koenigswald, 1970), Heteroxerus rubricati Crusafont, Villalta, and Truyols, 1955, Megacricetodon primitivus (Freudenthal, 1963), Democricetodon decipiens (Freudenthal and Daams, 1988), Eumyarion weinfurteri (Schaub and Zapfe, 1953), Microdyromys koenigswaldi De Bruijn, 1966, Peridyromys murinus (Pomel, 1853), Pseudodryomys ibericus De Bruijn, 1966, Simplomys julii (Daams, 1989), Glirudinus undosus Mayr, 1979, Ligerimys ellipticus Daams, 1976, Galerix symeonidisi Doukas, 1986, Talpidae sp. indet., and cf. Soricella discrepans Doben-Florin, 1964. The preliminary study of the palaeoecology of this mammalian assemblage by Crespo (2017) indicated an increase of the open forest, as well as higher humidity and temperature than the oldest localities from the Campisano ravine.
Here, we describe the fossil record and the palaeoecology of a new lower Miocene locality we name Foieta la Sarra-A, which namely consists of laminated lacustrine limestone beds located at the base of the Foieta la Sarra section. Our main goal is to infer the taphonomic and palaeoecological conditions of this outcrop. Together with the current data from La Rinconada and San Chils localities, our new findings shed light on the palaeoenvironmental dynamics of the Ribesalbes–Alcora Basin during the early Miocene.
Abbreviations.—EDX, X-ray energy microanalysis.
Geological setting
The deposits of the Ribesalbes–Alcora Basin are composed of detrital and carbonatic materials deposited in alluvial and lacustrine environments overlying a Mesozoic basement (Anadón et al. 1989) and divided into the Ribesalbes sequence (early to middle Miocene in age) and the Alcora sequence (late Miocene in age). The Ribesalbes sequence includes five depositional units (A to E from the base to the top) defined by Anadón et al. (1989). The Unit A consists of a 300 m interval of breccia of Mesozoic limestone clasts with minor interbedded red sandstone and mudstone. The Unit B is characterised by oil-shales and laminated bituminous dolomicrite of the La Rinconada and San Chils localities, which yield a rich and diverse lacustrine fossil record. The Unit C crops out in the Campisano ravine, where the mammalian fossiliferous sites are located. This unit is composed of sandstone beds ranging in thickness between 15 cm and 1.5 m, and which show cross and laminated bedding that alternates with thick massive mudstone strata interbedded with a few dolomitic and calcareous layers. These facies were interpreted by Anadón et al. (1989) as distal deltaic and shallow lacustrine environments. The Unit D represents an olistostrome of Cretaceous rocks, reflecting tectonic activity. Lastly, the Unit E is composed of thin bedded limestone strata rich in ostracods and charophytes, with interbedded mudstone horizons. On the other hand, the upper Alcora sequence consists of conglomerate beds related to alluvial deposits up to 200 m thick. This sequence displays a continuous transition without unconformities over the Ribesalbes sequence in the Campisano ravine (Anadón et al. 1989).
A synthetic stratigraphic column (Fig. 1C1) and a geological study of the Campisano ravine was presented by Crespo et al. (2019c). The stratigraphic section is 120 m thick and it is chiefly formed by mudstone and limestone beds, including some sandstone and microconglomerate layers. In the Campisano ravine, the Foieta la Sarra section is correlated to the lower part of the Mas d’Antolino B section (Fig. 1C2). The Foieta la Sarra stratigraphic section is 19 m thick and it consists of grey mudstone. A 1 m thick laminated lacustrine limestone bed, which corresponds to the Foieta la Sarra-A locality, can be distinguished at the base of the Foieta la Sarra section. The mammalian fossil site is located at the top of the Foieta la Sarra section, 16 m above the base, within grey sandy mudstone beds.
According to Crespo et al. (2019b), the presence of the genera Megacricetodon, Ligerimys, and Democricetodon in the Foieta la Sarra-1 locality is correlated with the Biozone C of the MN4 (early Aragonian, early Miocene, 16.49–15.94 Ma), based on the comparison with the Aragonian type area in the Calatayud–Montalbán Basin (García-Paredes et al. 2016). The Biozone C of the MN4 is divided in two sub-biozones, i.e., Ca and Cb, with the limit between them at approx. 16.20 Ma after Van der Meulen et al. (2012). The limit between these sub-biozones is present at the Campisano ravine (Crespo et al. 2019c). Specifically, the Foieta la Sarra-1 locality has provided the oldest record of Eumyarion weinfurteri and Ligerimys ellipticus within the biostratigraphy of the Campisano ravine localities and in the basin (Crespo et al. 2019c). Considering lower and upper stratigraphic levels, Foieta la Sarra-1 is older than the level Mas d’Antolino B-3 due to the high abundance of Megacricetodon and the scarcity of Democricetodon, which are probably correlated with the sub-biozone Ca from the Calatayud–Montalbán Basin. Foieta la Sarra-1 is younger than the level Mas d’Antolino B-0B because of the presence of both the genus Eumyarion and the species L. ellipticus, which are probably correlated with the sub-biozone Cb from the Calatayud–Montalbán Basin (Crespo et al. 2019, 2021b). Considering this information and the stratigraphic position of Foieta la Sarra-A locality studied here, its age could be approximately 16.20 Ma (early Miocene), and thus located at the base of the Cb sub-biozone.
Material and methods
The material studied herein includes compressed fossils of plants, molluscs, arthropods, and fish recorded in 45 laminated slabs excavated from the Foieta la Sarra-A locality at the Foieta la Sarra section near Araia d’Alcora (l’Alcora, Castelló Province, Spain; Fig. 1). The fossil specimens from Foieta la Sarra-A are housed at the Museu de la Universitat de València de Historia Natural (MUVHN) in Burjassot (Valencian Community, Spain). The sedimentary rock exposed at Foieta la Sarra-A does not show significant signals of weathering, although several Recent small roots and fungal mycelia were removed from the slabs. The rock slabs were observed with a Leica MS5 stereomicroscope. To enhance visibility of some microfossils, slabs were submerged in ethanol. Specimens were photographed using a Leica DMS1000 stereomicroscope with an attached digital camera, and the drawings were made using a camera lucida attached to the Leica MS5 stereomicroscope. All of these methods took place at the Department of Botany and Geology of the Universitat de València.
The micromorphology of the samples was imaged using a FEI INSPECT (5350 NE Dawson Creek Drive Hillsboro, Oregon 97124, USA) Scanning Electron Microscope (SEM) at the Museo Nacional de Ciencias Naturales (MNCN) in Madrid (Spain). The SEM microscope in low vacuum mode allows hydrated samples to be studied in their original state using the large field detector (LFD), since it is close to the sample in order to avoid electron losses. In addition, the samples were observed with the Backscattering Electron Detector (BSED), which were conductive in high vacuum mode. The SEM resolution at low vacuum was 4.0 nm at 30 kV (BSED); the accelerating voltage was 20 kV, low vacuum 0.50 torr, and 10 mm of working distance. The X-ray energy microanalysis (EDX) of the ephippial microsculpture, chironomid larval galleries and fish remains were conducted with an energy-dispersive X-ray spectrometer (INCA Energy 200 energy dispersive system, Oxford Instruments) at the MNCN (Madrid, Spain).
Anatomic terminology for fish remains follows Lagler (1947), Daniels (1996), Schultze (2015) and Bräger and Moritz (2016).
Foieta la Sarra-A fossil assemblage
The Foieta la Sarra-A locality has yielded a diverse record of plants (including charophytes and embryophytes), molluscs, arthropods (including crustaceans and insects) and vertebrates represented by fish. The botanical and faunal taxa of Foieta la Sarra-A are available in Table 1.
Table 1. Botanical and faunal record from the lower Miocene Foieta la Sarra-A locality (Ribesalbes–Alcora Basin, Castelló Province, Spain).
|
Charophyta |
|
|
Charophyceae |
|
|
Charales |
|
|
Characeae |
Chara sp. |
|
Embryophyta |
|
|
Monocotyledoneae |
|
|
Poales |
Poaceae, Cyperaceae, Sparganiaceae
|
|
Dicotyledoneae |
|
|
Incertae sedis |
Dicotylophyllum sp. |
|
Malpighiales |
|
|
Salicaceae |
Salix? sp. |
|
Mollusca |
|
|
Gastropoda |
|
|
Littorinimorpha |
Hydrobiidae? indet. |
|
Hygrophila |
Lymnaeidae indet. |
|
Planorbidae |
Ferrissia sp. Gyraulus sp. |
|
Arthropoda |
|
|
Branchiopoda |
Ostracoda indet. |
|
Cladocera |
|
|
Daphniidae |
Daphnia (Ctenodaphnia) sp. |
|
Insecta |
|
|
Coleoptera |
Coleoptera indet. |
|
Hemiptera |
Pentatomidae? indet. |
|
Diptera |
Psychodidae? indet. |
|
Vertebrata |
|
|
Actinopterygii |
|
|
Teleostei |
Teleostei indet. |
|
Cyprinodontiformes |
|
|
Cyprinodontidae |
cf. Aphanius sp. |
Plants.—Three grey-yellowish limestone intervals containing abundant charophyte remains have been distinguished in the Foieta la Sarra-A (Fig. 2A, B). These intervals are exclusively composed of compressed vegetative (thallus) fragments and poorly preserved gyrogonites (calcified fructifications of Characeae) forming a type of calcareous sedimentary rocks defined as characeite by Soulié-Märsche et al. (2010). The flattened charophyte stems studied herein consist of isostichous corticated thalli. Considering that living charophyte genera have not changed their vegetative structure since the Miocene, these charophyte thalli can be attributed to the genus Chara since among the living charophytes only this genus develops corticated stems (Soulié-Märsche et al. 2010). Complete articulated thalli would be required for a species-level attribution. Few poorly preserved gyrogonites occur among charophyte stems (Fig. 2B). The absence of complete gyrogonites and their poor preservation hinder a detailed taxonomic attribution. Several molds and casts of planorbid gastropods (Gyraulus sp.) also occur associated to these characeite intervals.

Fig. 2. Plant remains from the lower Miocene Foieta la Sarra-A locality (Ribesalbes–Alcora Basin, Castelló Province, Spain). A, B. Chara sp. corticated isostichous thalli (FS-3A) and two gyrogonites surrounded by thalli (FS-3B). C. Burned plant remains or fusinite of diverse size (arrowheads) (FS-8A-1). D. Poorly preserved Poaceae, Cyperaceae, Sparganiaceae or Juncaceae leaf remain (FS-21B-1). E, F. Dicotylophyllum sp. leaf remains (E, FS-22A-1; F, FS-24B-1). G, H. Salix? sp. leaf remains (G, FS-19B-2; H, FS-40-1). Scale bars A–C, 1 mm; D–F, 2 mm; G, H, 4 mm.
Several microlevels of Foieta la Sarra-A show accumulations of burned plant remains or fusinite (Fig. 2C) as described from La Rinconada and San Chils localities (Peñalver et al. 2016; Álvarez-Parra and Peñalver 2019), possibly resulting from wildfires near the palaeolake. Undetermined seeds with slightly bilobate apex have been found, although their preservation is poor. The Foieta la Sarra-A plant record also includes leaves, but their taxonomic affinities are challenging to determine. A large leaf remain and scarce small fragments with straight margins and parallel veins (Fig. 2D) are similar to specimens of Typha latissima A. Braun, 1851 in Heer, 1855 (Typhaceae) found in the Miocene Rubielos de Mora locality (Barrón and Diéguez 2001); this type of plant remain is also found in La Rinconada and San Chils localities. However, they are usually poorly preserved and their morphotype could correspond to Poaceae, Cyperaceae, Sparganiaceae or Juncaceae belonging to the order Poales (Peñalver et al. 2016; Álvarez-Parra and Peñalver 2019). Two slender leaves with distal areas partly preserved, 11.6 mm long and 2.9 mm maximum wide and 14.3 mm long and 2.12 mm maximum wide, respectively (Fig. 2E, F), seem to be similar to the morphotypes of Dicotylophyllum sp. present in La Rinconada (Barrón and Postigo-Mijarra 2011). Two poorly preserved leaf remains with around 6 mm maximum width have been tentatively determined as Salix? sp. (Salicaceae) based on the straight, slightly parallel margins and their similarity to the specimens of Salix lavateri A. Braun emend. Hantke, 1851 from La Rinconada (Barrón and Postigo-Mijarra 2011; Fig. 2G, H).
Molluscs.—The gastropod assemblage recorded in the studied samples is poorly preserved. Almost all specimens consist of moulds and impressions, which hinder accurate taxonomic attributions. The assemblage consists of at least four freshwater morphotypes, one of which belongs to the superfamily Truncatelloidea and the rest to the superfamily Lymnaeoidea. The Truncatelloidea superfamily is represented in Foieta la Sarra-A, with 15 recognised specimens in seven of the examined samples, by a single morphotype (Fig. 3A–C). Due to the incompleteness of the specimens and their poor preservation, they may belong to Hydrobiidae or Bithyniidae. However, the general ovate-conical morphology of the specimens, with convex spiral whorls and, above all, their small size, suggest the attribution to the family Hydrobiidae.

Fig. 3. Molluscs from the lower Miocene Foieta la Sarra-A locality (Ribesalbes–Alcora Basin, Castelló Province, Spain). A–C. Hydrobiidae? indet. specimens (A, FS-9A-1; B, FS-44C-2; C, FS-15-1). D–G. Lymnaeidae indet. specimens (D, FS-10-1; E, FS-39-1; F, FS-28B-2; G, FS-28A-2). H–L. Ferrissia sp. specimens (H, FS-16-1; I, FS-16-3; J, FS-13A-1; K, FS-31C-3; L, FS-39-2). M–R. Gyraulus sp. specimens (M, FS-42A-1; N, FS-42B-1; O, FS-35B-1; P, FS-35A-1; Q, FS-7B-1; R, FS-7A-1). Scale bars 1 mm.
Regarding the three morphotypes of the superfamily Lymnaeoidea, 120 specimens belonging to the families Lymnaeidae and Planorbidae have been studied. Lymnaeidae is represented by one morphotype, from which eight specimens have been identified (Fig. 3D–G). Seven correspond to fragments of adult specimens (Fig. 3D, E) and one to a juvenile (Fig. 3F, G). The fragments of the adults are very incomplete but show elongate morphology, a last round of rapid growth, and ornamentation of growth constrictions in the vicinity of the aperture. The juvenile specimen, with dimensions of up to 6.7 mm long and 3.7 mm wide, is compressed and deteriorated. Nevertheless, an elongate-ovate shell with three whorls is visible, its spire being short and conical, with a last whorl enlarging rapidly. The assignment at the genus level is challenging, but the characteristics observed match those of the genera Stagnicola and Radix. Vilanova y Piera (1859) cited the presence of “Lymnaeas” in several areas of the Castelló Province, including the “Balsa de Fanzara” (the classical name for the Cenozoic sediments of the Ribesalbes–Alcora Basin), although this author did not provide further information. In any case, we prefer to keep the determination of the lymnaeid morphotype at the family level.
Two species of the family Planorbidae have been identified in Foieta la Sarra-A, i.e., Ferrissia sp. (Fig. 3H–L), with 41 specimens present in 16 of the studied laminated slabs, and Gyraulus sp. (Fig. 3M–R), with 71 specimens in 21 of the slabs. The specimens of Ferrissia, the largest with 5.90 mm long and 3.89 mm wide, have a limpet-like morphology with an elliptical basal outline. The anterior shell portion is slightly broader and convex, the flanks are nearly straight or slightly convex, and the posterior part is convex. The shell apex is slightly deflected to the right, its posterior portion is slightly concave, and its anterior portion is weakly convex in lateral view. The ornamentation is poorly preserved. These specimens of Ferrissia from Foieta la Sarra-A locality differ from the middle Miocene Ferrissia illyrica (Neumayr, 1880) from the Dinaride Lake System (Neubauer et al. 2011, 2013, 2015c) by its narrower contour. The early–middle Miocene Ferrissia wittmanni (Schlickum, 1964) from Southern Germany and Austria (Harzhauser and Kowalke 2002; Kowalke and Reichenbacher 2005; Harzhauser et al. 2014) also has a narrower profile, as well as concave flanks and a cup-shaped apex. Moreover, the late Miocene Ferrissia truci Wautier, 1975, from France (Wautier 1975) differs from the Foieta la Sarra-A specimens by its more elongate contour, as well as the elongate apical region with a rounded apex located far back and strongly inclined to the right. The shape of the shell of the studied specimens is particularly similar to that of Ferrissia deperdita (Desmarest, 1814), a common species in the middle Miocene of Central Europe. However, the general degree of preservation and particularly the ornamentation prevent us from assigning our specimens to the latter species. On the other hand, Gyraulus sp. is the most abundant gastropod species in the studied samples. However, all fossils attributed to this taxon correspond to incomplete moulds and impressions, which hinders the determination to species level. However, the characteristics of this morphotype, whose shell reaches four whorls among the largest specimens, are consistent with those of Gyraulus based on the growth pattern (with whorls moderately increasing in diameter), the estimated dimensions among the studied specimens (with a maximum diameter of 5.18 mm), and the ornamentation of growth lines, i.e., prosocline in umbilical view and prosocyrt in apical view.
Arthropods.—Crustaceans and insects represent the hitherto recovered arthropod record from the Foieta la Sarra-A locality. Crustaceans include Ostracoda and Cladocera. Ostracod shells 1–2 mm long, 0.5–1 mm wide without apparent ornamentation have been found (Fig. 4A); they have been recorded both as isolated specimens and as mass records (blooms) in the laminated slabs. The poor preservation of the Foieta la Sarra-A ostracod shells prevents a proper determination. On the other hand, mass records of cladoceran ephippia occur in the Foieta la Sarra-A slabs (Fig. 4B). Specimens are around 0.8–1 mm long and 0.4–0.5 mm wide with an ovoid or sub-rectangular shape. Although the ephippia are poorly preserved, the margins of the valve (originally highly sclerotised) and the paired egg chambers can be distinguished. The dorsal margin lacks spinules. The ephippia show their typical reticular microsculpture (Fig. 5A) with high value of Ca element (Fig. 6A). The ephippial accumulations do not show uniform orientations in the slabs (see Peñalver et al. 1996). The morphotype of the Foieta la Sarra-A ephippia corresponds to the subgenus Daphnia (Ctenodaphnia) based on the sub-rectangular shape and the axes of the eggs being sub-parallel to the dorsal margin (Kotov and Taylor 2011), as in two specimens recorded from the San Chils locality (Álvarez-Parra and Peñalver 2019). Miocene cladoceran ephippia have been found in several lacustrine deposits from the Iberian Peninsula, such as Rubielos de Mora (Peñalver et al. 1996), Bellver de Cerdanya in Lleida (Martín-Closas and Delclòs 2007), and Tresjuncos in Cuenca (Bustillo et al. 2017).
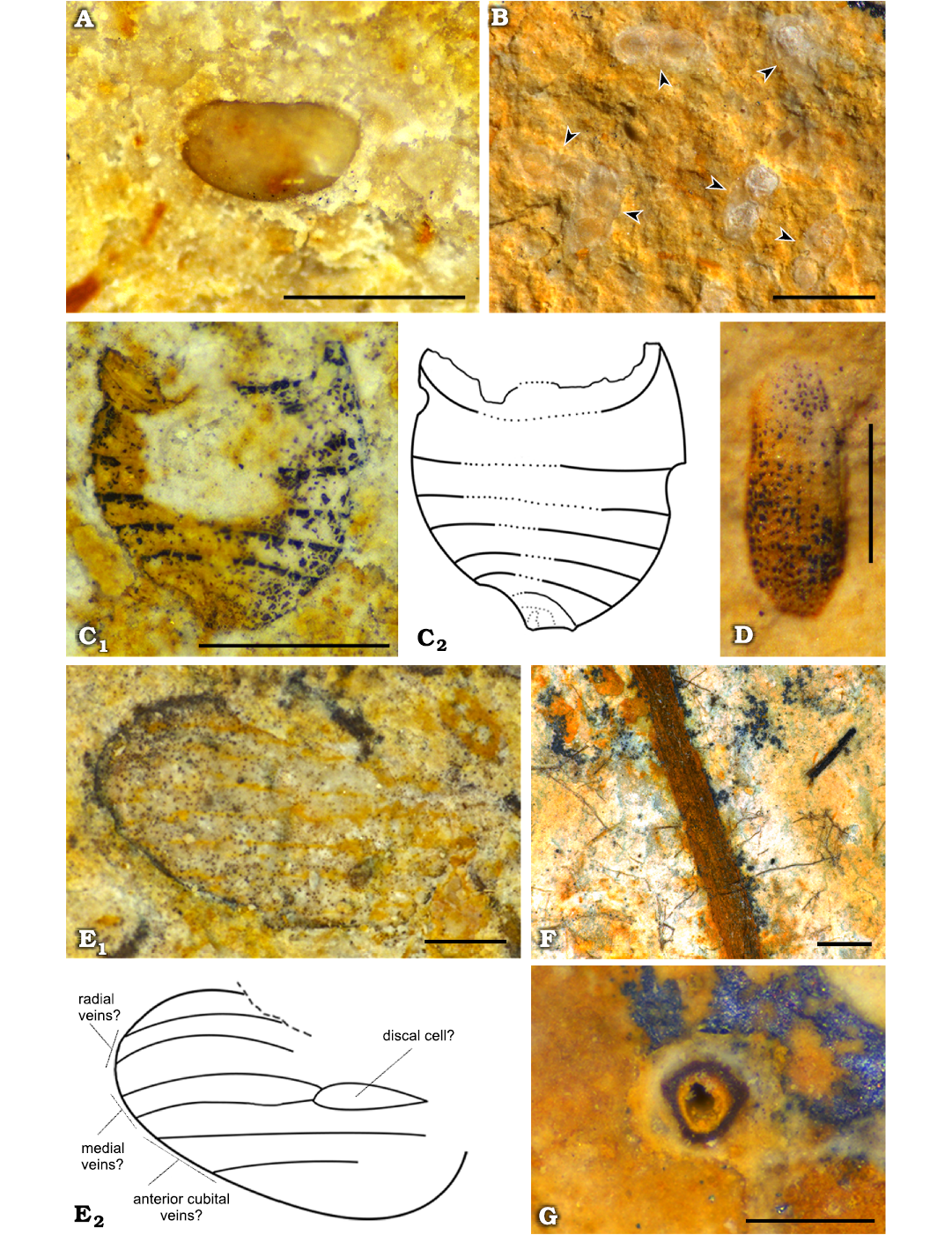
Fig. 4. Arthropods from the lower Miocene Foieta la Sarra-A locality (Ribesalbes–Alcora Basin, Castelló Province, Spain). A. Undetermined ostracod shell, FS-38A-1. B. Mass record of Daphnia (Ctenodaphnia) sp. (Cladocera) ephippia, FS-21A; each specimen is indicated with an arrowhead. C. Photograph (C1) and drawing (C2) of a Pentatomidae? (Hemiptera) abdomen, FS-41-1. D. Isolated beetle (Coleoptera) elytron, FS-19B-1. E. Photograph (E1) and drawing (E2) of a Psychodidae? (Diptera) wing, FS-44D-1. F, G. Chironomid larval galleries in longitudinal (FS-45-1) and transversal (FS-29) views, respectively. Scale bars: A, B, C, E, F, 1 mm; D, G, 0.5 mm.

Fig. 5. SEM images of arthropod remains from the lower Miocene Foieta la Sarra-A locality (Ribesalbes–Alcora Basin, Castelló Province, Spain). A. Complete Daphnia (Ctenodaphnia) sp. (Cladocera) ephippium (A1) showing an exceptionally well-preserved reticulated microsculpture (FS-43B-4); detail showing the limit between the paired egg chambers (A2), and the dorsal ridge with variation in microsculpture in the top of the image (A3). B. Chironomid larval gallery (B1) and its filling material (B2) resulting from oxidation of the original framboidal pyrite (FS-34A-1). Scale bars: A1, 0.3 mm; A2, A3, B2, 0.05 mm; B1, 0.1 mm.
Regarding insects, specimens belonging to the orders Hemiptera, Coleoptera, and Diptera have been found. First, a partly preserved abdomen divided in at least seven sclerites of 1.32 mm in length and 1.46 mm in maximum width has been found (Fig. 4C); its distal area is deformed, and the genitalia are not visible. Its morphology could correspond to a pentatomid hemipteran (Hemiptera: Pentatomidae). This group is represented in the San Chils locality by a Sciocoris? sp. specimen (Álvarez-Parra and Peñalver 2019). Moreover, beetles (Coleoptera) are represented among the Foieta la Sarra-A material by a poorly preserved isolated elytron, 0.91 mm in length with rounded apex and ornamentation constituted by ten or 11 rows of hollows (Fig. 4D). Beetles had been previously reported from the Ribesalbes–Alcora Basin (Peñalver et al. 2016; Álvarez-Parra and Peñalver 2019). Lastly, an isolated wing with a rounded apex, six longitudinal veins and a fusiform (discal?) cell likely belongs to a psychodid fly (Diptera: Psychodidae) (Quate and Vockeroth 1981). In any case, this determination should remain tentative as the wing margin is not clearly defined and the veins obscured due to preservation (Fig. 4E). This would be the first report of the family Psychodidae in the Ribesalbes–Alcora Basin. In addition, U-shaped and rather straight galleries of around 0.2–1 mm in diameter and different lengths have been found in the slabs from Foieta la Sarra-A (Fig. 4F); their sections are circular (Fig. 4G). This type of ichnofossils of around 1 mm in diameter is commonly related to bioturbation produced by chironomid (Diptera) larvae and are usually found in lacustrine deposits (Walshe 1951). Similar ichnofossils attributed to chironomid larval galleries have been found in other Cenozoic outcrops from the Iberian Peninsula (e.g., Rodríguez-Aranda and Calvo 1998). The galleries show an orange-brown colour resulted of the oxidation process of the original framboidal pyrite (Fig. 5B), as EDX microanalysis indicates (Fig. 6B).
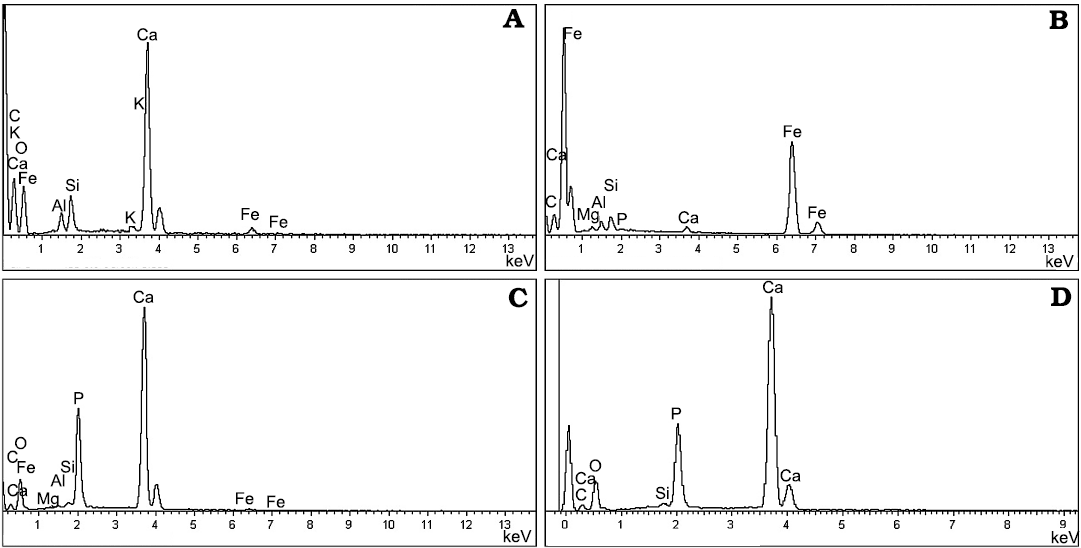
Fig. 6. EDX microanalyses of specimens from the lower Miocene Foieta la Sarra-A locality (Ribesalbes–Alcora Basin, Castelló Province, Spain). A. Ephippial microsculpture of Daphnia (Ctenodaphnia) sp. (FS-43B-4). B. Filling material of a chironomid larval gallery (FS-34A-1). C. Morphotype 2 fish scale (FS-21C-9). D. Isolated fish bone (FS-21).
Fishes.—A poorly preserved and incomplete teleostean specimen, abundant clumps of scales and bones, isolated scales, and other scattered remains comprise the fish record from Foieta la Sarra-A. The incomplete teleostean specimen, preserved as a part and a counterpart, preserved its anterior part. It is comprised by a crushed skull, scales of the abdominal region and the dorsal and pectoral fins (Fig. 7A). The skull is crushed and only the dentary bone and some bones of the opercular apparatus are recognisable, although distinctive features of the fish skull are obscured by dissolution and/or enzymatic attack, which hinder both a detailed anatomic description and an accurate taxonomic assignment. As far as it can be described, the dorsal fin appears to be triangular; no scutes, basal or fringing fulcra are observed. The dorsal fin base length is about 30 mm and dorsal pterygiophores are badly preserved. There are ca. 15 dorsal fin rays with long bases, in the preserved part of which segmentation and branching are not evident. It is unclear if only the first dorsal fin ray forms the leading margin of the fin. The distal portion of the fin is obscured by sediment. The vertebral column and its associated elements are not apparent. Although, one badly preserved vertebral centrum is observed near the base of the skull, it is not clear whether it belongs to this specimen or not. The squamation is composed by small, thin sub-squarish elasmoid scales of the cycloid type (morphotype 1, see below). Scales are imbricated, most of them completely preserved. Some isolated scales show their anterior field exposed; the posterior field, which is inserted below the epidermis, appears to be large. No lateral line scales have been recognised, nor ctenoid or spinoid scales are observed in the specimen. Circulii are present as concentric crests surrounding the focus area of the scales; the focus seems to be displaced from the central area. Circulii of the posterior and lateral fields are smooth and seems to be regularly spaced. The so-called first circulii are also smooth. The scales are composed by two layers: the superficial layer and the basal bony plate (Fig. 8B). The basal bony plate is the main part of the scale; its structure or size of collagen fibres is not evident in the available material.

Fig. 7. Fish remains from the lower Miocene Foieta la Sarra-A locality (Ribesalbes–Alcora Basin, Castelló Province, Spain). A. Poorly preserved teleostean specimen (FS-23A-1); df: dorsal fin, pf.r: pectoral fin rays; the arrow indicates the anterior region of the specimen. B, C. Clump of scales and bones (FS-38A-2); arrowheads indicate diagenetic fractures. Scale bars: A, B, 5 mm; C, 2 mm.
The clumps of scale and bones are well-sorted (Fig. 7B, C). Among the bones, some fin rays and ribs have been identified. The scales in the clumps are represented by at least the two morphotypes described below. It is unclear if these two morphotypes of scales belong to the same individual. Moreover, it is also unclear if some of these clumps represent bromalites (i.e., coprolites, cololites, and regurgitalites; see Hunt and Lucas 2012). Some fin rays and ribs (i.e., fractures perpendicular to the major axis of the bone; Fig. 7B, C), as well as most of the scales, exhibit diagenetic fractures.
Fish scales from Foieta la Sarra-A are represented by at least two morphotypes that are described below. The morphotype 1 is well-represented in the incomplete specimen covered with articulated scales (see above), as well as in the clumps of scales and bones. EDX analysis of fish scales of morphotype 2 (Fig. 6C) and an isolated bone (Fig. 6D) confirm that the scale and bone are composed namely of Ca, P, and O, indicating the prevalence of apatite.
Morphotype 1: Thin sub squarish elasmoid scales of the cycloid type. These scales measure ca. 1.5 mm long (measured antero-posteriorly) and 1.6 mm wide (measured in the widest portion) (Fig. 8A, B). They have a relatively small focus that is antero-posteriorly elongate. Circulii are well-developed. The anterior field is striated (see Bräger and Moritz 2016: fig. 3B) and bears 13 to 17 radii (= ridges). The so-called first circulii are concave to straight. Lepidonts (= small tooth-like structures anchored to circulii) are not observed. The posterior field seems to be convex and smooth. Two generations (principal and secondary) of radius (=ridges) are easily recognised. The posterior field of the scales does not show tubercles, but its absence cannot be fully ascertained due to the preservation of the available material.
This scale morphotype is commonly found in many freshwater fishes (e.g., Cyprinodontiformes and Atheriniformes), and it is most similar to the scales present in some killifish species of the genus Aphanius (e.g., Gholami et al. 2013; Teimori et al. 2017).
It is important to highlight that scales might vary in type (ctenoid and cycloid) and morphology (rounded, squarish, etc.) in the same individual relative to the different body sections of an individual. Scales are plastic structures; their general morphology is highly (intra- and interspecifically) variable; also, microstructure seems to show a high degree of intraspecific variation, at least in some cyprinodontiforms such as Aphanius (e.g., Gholami et al. 2013) and clupeiforms (e.g., Patterson et al. 2002). Because taxonomy based on isolated scales appears to be problematic (e.g., Bräger et al. 2017), we prefer to leave them unassigned.

Fig. 8. Fish scale morphotypes from the lower Miocene Foieta la Sarra-A locality (Ribesalbes–Alcora Basin, Castelló Province, Spain). A, B. General view and SEM image of the morphotype 1 (FS-21B-2 and FS-21A-10, respectively); the arrow indicates de anterior field of the scale. C. General view (C1) and SEM image (C2) of the morphotype 2 (FS-21C-9). Scale bars: A, 1 mm; B, C, D, 0.5 mm.
Morphotype 2: Elasmoid scale of the cycloid type, thin, rounded or circular in shape, diameter about 1 mm (Fig. 8C, D). The scale has a large focus located in the middle and concentric circulii. Radii and spines or ctenii are lacking. The number of circulii (seven to ten) is almost equal across the entire field. Circulii are almost equally spaced and concentric. This scale resembles those present in the freshwater gadid teleostean Lota lota Linnaeus, 1758, which is the only freshwater member of the family Gadidae and is adapted to cold waters (Lagler 1947; Oates et al. 1993; Daniels 1996). Note that some members of Salmoniformes have similar scale morphology (Patterson et al. 2002).
Discussion
Taphonomy.—The laminated limestone beds from Foieta la Sarra-A are interpreted as lacustrine deposits; the recorded taxa correspond to a typical assemblage of a perennial palaeolake, characterised by the presence of lymnaeid and planorbid molluscs, ostracods, and charophytes (Freytet and Verrecchia 2002). The Foieta la Sarra-A record fulfils the requirements to be considered a Konservat-Lagerstätte based on the preservation of soft-bodied specimens, such as the well-preserved reticulated microsculpture of the Daphnia (Ctenodaphnia) ephippia (Fig. 5A) and insect remains (Fig. 4C–E), which is explained by the presence of microbial mats in the palaeolake (Wilby et al. 1996). The incomplete fish specimen with articulated bones, fins, and scales in original position (Fig. 7A) could have reached the bottom of the lake articulated in conditions of low water energy (see below). The carcass lacks evidence of flotation, as well as of predation and/or scavenging. Aside from this singular specimen, bacterial decay and flotation appear to be evident in the remaining fish carcasses, as most of the specimens are disarticulated and forming clumps (Fig. 7B, C), perhaps due to scavengers since no clear orientation of the elements is apparent. In any case, at least some of the clumps could be bromalites; thus, their preservation resulted from rapid burial after production. The well-developed diagenetic fractures on bones and scales evidence the fossil diagenesis (Fig. 7B, C). There appears to be no directional scatter of Daphnia (Ctenodaphnia) sp. ephippia, fish scales, and/or lepidotrichia. Thus, the palaeolake was apparently quiet, lacking currents that could have affected the distribution of fish carcasses. Characeite rocks and the slabs with chironomid larval galleries correspond to the in situ record of marginal areas of the lake, while the rest of the specimens were deposited at the lake bottom covered by microbial mats.
The presence of charophytes, mass records of cladoceran ephippia, ostracods, and fish remains in Foieta la Sarra-A indicates that the water chemistry was dissimilar to that of the palaeolake of the Unit B which is apparently lacking these taxa (Peñalver et al. 2016). The laminated slabs from La Rinconada and San Chils resulted from the sedimentation of fine-grained particles on microbial mats at the anoxic bottom from a meromictic lake (Anadón et al. 1989; Peñalver et al. 2016; Álvarez-Parra and Peñalver 2019). The environmental conditions related to the presence of microbial mats in Foieta la Sarra-A could also be different. As noted above, it is unclear if the clumps of fish scales and bones resulted from scavenging or they are bromalites. Therefore, we propose two hypotheses for the presence of microbial mats at the lake bottom: (i) as finding potentially scavenged fish is incompatible with an anoxic lake bottom, the microbial mats were related to a lake bottom that was at least slightly saline (Bauld 1981; Peñalver and Gaudant 2010); this contradicts the molluscan assemblage found, which indicates freshwater conditions in the palaeolake; (ii) if the fish remains were bromalites, the lake bottom could have been anoxic. The preservation of the Foieta la Sarra-A materials is worse than other Miocene palaeolakes such as Bicorb and La Rinconada (Peñalver and Gaudant 2010; Peñalver et al. 2016), so it is plausible that the microbial mats of Foieta la Sarra-A were thin veils rather than thick mats. A detailed taphonomic study of the Foieta la Sarra-A palaeolake is beyond the aims of this contribution, and future studies will have to address that matter.
Palaeoecological reconstruction of the Foieta la Sarra-A palaeolake and its surroundings.—The botanical and faunal composition of the Foieta la Sarra-A record (Table 1) represents the community that inhabited a lacustrine environment and contiguous areas during the early Miocene.
Charophyte accumulations of Chara sp. recorded as characeite are related to the presence of charophyte meadows at the lower marginal areas of the lake, which was well-oxygenated and well-illuminated based on the finding of thalli and gyrogonites (Soulié-Märsche et al. 2010). Charophyte stems are well-preserved and somewhat fragmented, suggesting that the environment was relatively quiet, without the action of strong currents nor waves. The charophyte meadows corresponded to one of the primary producers in the palaeolake.
The assemblage of molluscs is composed of 127 poorly preserved specimens of four freshwater gastropod taxa. The 93.3% are pulmonate species (Lymnaeidae indet., Ferrissia sp., and Gyraulus sp.) which, as a whole, indicate aquatic environments that are shallow, stagnant or with weak currents and abundant aquatic vegetation (Glöer 2002). Ferrissia would also indicate the existence of probable reed-belts on the shore while Lymnaeidae and Gyraulus point out the possibility of temporary desiccation (Fetcher and Falkner 1993; Glöer 2002). Many extant individuals studied in Europe, determined as Ferrissia wautieri (Mirolli, 1960) but according to Vecchioni et al. (2017) corresponding to the allochthonous species of cosmopolitan distribution Ferrissia californica (Rowell, 1863), inhabit stagnant to lentic waters, living among the leaves of aquatic vegetation and underneath the leaves of reeds-belts on the upper littoral zone and showing little sensitivity to water quality (Glöer 2002). Some authors also indicate that the same species is thermophilous and lives in eutrophicated waters (Van der Velde 1991). Extant species of Lymnaeidae preferably inhabit shallow, permanently stagnant or weakly flowing waters, with salinities ranging from freshwater to oligohaline in the case of Stagnicola and up to mesohaline in Radix; lymnaeids typically live on macrophyte vegetation feeding on epiphytes (Adam 1960; Okland 1990; Glöer 2002). Some lymnaeid species withstand periods of desiccation and inhabit temporary water bodies, even those being located on flood plains (Fetcher and Falkner 1993). Extant species of Gyraulus live in a variety of conditions, preferably in shallow waters, including water bodies that are ephemeral, stagnant, or slowly running and abundant in vegetation (Glöer 2002; Welter-Schultes 2012). The fourth taxon (Hydrobiidae? indet.), which represents 6.7% of the specimens studied, suggests permanent waters, at least at the time of deposition. Extant species of Hydrobiidae thrive in permanently flooded freshwater to brackish water environments (Arconada and Ramos 2003).
Lymnaeids feed namely by scraping algae and diatoms from rocks or macrophytes (Pyron and Brown 2015), although they often consume filamentous algae (Kesler et al. 1986). Extant Holarctic species of Ferrissia show a preference for periphytic algae, especially diatoms (Burky 1971). Concerning the genus Gyraulus, the analysis of stomach content indicates a primarily detritivorous diet (Dudgeon and Yipp 1985), although other studies also show algal consumption (e.g., Hann et al. 2001). The species of Hydrobiidae have a widely diverse diet, including detritus, bacteria, filamentous algae, and diatoms, among others. Studies on the feeding habits of some freshwater hydrobiid species (Radea et al. 2017), as well as species from other related families such as Amnicolidae (Kesler 1981), show a diet suggesting the importance of periphyton diatom grazing. In sum, the combination of the four gastropod taxa found in Foieta la Sarra-A suggest that the palaeolake was shallow, perennial, and quiet (absence of strong currents). Moreover, this molluscan assemblage indicates that the aquatic macrophytes were abundant in the shallow areas of the lake such as the littoral zone, and the shores were also covered with helophytic vegetation. This vegetation was important in maintaining molluscan populations, as constituted the main substrate for the epiphytes on which molluscs fed on, particularly in the case of Lymnaeidae. Moreover, this vegetation would contribute to the formation of detritus which could support the Gyraulus sp. populations. These characteristics are compatible with a vegetated shore of the lake whose waters were oligohaline at the most.
Daphnia cladocerans are filter-feeding crustaceans of freshwater habitats (Ebert 2005). The specimens from Foieta la Sarra-A correspond to the subgenus Daphnia (Ctenodaphnia), which is related to shallow, temporary water bodies (Popova et al. 2016). Daphnia (Ctenodaphnia) ephippia from Foieta la Sarra-A have been found as mass records in the rock slabs, suggesting environmentally stressful conditions, such as temporary desiccation or lack of food resources, that caused the change in their reproductive cycle from parthenogenetic to ephippial females (Peñalver et al. 1996; Ebert 2005). The finding of these records is interesting, as it appears to contradict the permanent water condition inferred by charophytes, some of the molluscs and fish. Nonetheless, Daphnia (Ctenodaphnia) individuals could have occupied lake margins that desiccated during dry periods in a lake with a fluctuating extension. Predators of Daphnia (Ctenodaphnia) could have been other arthropods as well as vertebrates, although their presence did not affect the emergence of massive Daphnia populations. Although cladoceran ephippia have not been found in the extensive fossil record of La Rinconada (Peñalver et al. 2016), and they are represented in San Chils only by two specimens in a slab (Álvarez-Parra and Peñalver 2019), similar ephippial mass occurrences were found in Rubielos de Mora (Peñalver et al. 1996). Moreover, the occurrence of mass records of Daphnia (Ctenodaphnia) in Foieta la Sarra-A indicates that the water chemistry of this palaeolake was different in comparison to the lacustrine deposits from the Unit B. Furthermore, ephippia from Foieta la Sarra-A do not show a preferential orientation in the slabs, suggesting a low energy environment. Lastly, they are found associated to mass occurrences of ostracods, like the ones from Rubielos de Mora and Bellver de Cerdanya Miocene outcrops (Peñalver et al. 1996).
The absence of dipteran pupal exuviae in Foieta la Sarra-A is noteworthy, as they are abundant in La Rinconada and San Chils, where they indicate stagnant water and lack of predators (Peñalver et al. 2016; Álvarez-Parra and Peñalver 2019). The presence of fish predators matches such absence of dipteran exuviae, although the record of the latter would require a higher grade of exceptional preservation than that observed in Foieta la Sarra-A. Nonetheless, the finding of Chironomidae larval galleries occurring at the lake bottom suggests that populations of this group of flies were present in this lacustrine environment; chironomid larvae are found in ecosystems of different characteristics, although their presence in Foieta la Sarra-A could be related to shallow waters of a lentic environment and were microphagic, i.e., feeding on detritus and small plants or animals (Oliver 1981).
The teleostean fish scale morphotype 1 resembles the scales present in some killifish (Cyprinodontiformes), a large group of secondarily freshwater fishes with a wide geographical distribution. More specifically, the scale morphotype 1 resembles the scales present in species Aphanius, including small omnivorous eurythermic and eurhyhaline species that inhabit brackish and salt marshes, coastal lagoons, river mouths and freshwater environments (Gaudant 1993; Nelson et al. 2016). Therefore, Aphanius species are highly ecologically flexible fishes that tolerate a wide range of temperature and salinities, and so they may occur in a wide range of habitats (Teimori et al. 2014). Despite their omnivory, the diet of some Aphanius species has a high degree of seasonal variation and, in general terms, includes several small invertebrates and their eggs, as well as diatoms (Leonardos 2008). Also, some species of Aphanius display some differences in habitat preference related to their age (Alcaraz and García-Berthou 2007).
Although the palaeoichtyofauna can be useful to understand the palaeoenvironment, it is important to highlight that “deducing a type of environment from only a single taxon´s autoecology risks falling into a circular reasoning but this analysis is, however valuable if its incorporates a set of signals that point to the same environment” (Cavin 2017: 4). Moreover, due to the high diversity of fishes’ lifestyles and their ability to live in environments with changes in salinity, using them to determine or define types of freshwater environments is challenging.
The palaeovegetation of Foieta la Sarra-A corresponds to a typical riparian assemblage, similar to the La Rinconada and San Chils assemblages related to a lacustrine palaeoenvironment (Barrón and Postigo-Mijarra 2011; Postigo-Mijarra and Barrón 2013; Álvarez-Parra and Peñalver 2019), based on the presence of specimens of Poales, Dicotylophyllum sp., and Salix? sp. Palaeofires could be recurrent based on the accumulations of fusinite in the Foieta la Sarra-A rock. The scarcity of insects in the fossil record of Foieta la Sarra-A might be explained by the presence of vertebrates such as fishes, which fed on the former when they were washed to the lake. Abundant amphibian, crocodilian (and other reptilian), and mammalian record has been found in the other levels of the Foieta la Sarra section and Campisano ravine localities (Crespo 2017; Crespo et al. 2019c; VDC unpublished data), whereas amphibians and bird (only feathers) remains have appeared in La Rinconada and San Chils (Peñalver et al. 2016; Álvarez-Parra and Peñalver 2019). Therefore, although these vertebrates could have inhabited the riparian environment or that contiguous the palaeolake of Foieta la Sarra-A, there is a gap of such faunal record to date.
Palaeoenvironmental dynamics during the early Miocene in the Ribesalbes–Alcora Basin.—The deposits of the Ribesalbes–Alcora Basin show a dynamic environmental evolution of the water body during the early Miocene. The oil-shale and laminated bituminous dolomicrite of La Rinconada and San Chils localities, belonging to the Unit B, correspond to the record of a meromictic palaeolake (Peñalver et al. 2016). On the contrary, the stratigraphically higher fine grain facies of the Campisano ravine, belonging to the Unit C, represent the distal deposits of an alluvial fan or a muddy floodplain (Crespo et al. 2019c), indicating an environmental change during the early Miocene in the basin. The laminated slabs found in Foieta la Sarra-A hitherto represent the only record of a well-developed palaeolake in the stratigraphy of the Campisano ravine (Crespo et al. 2019c), showing that the palaeoenvironments inferred for the facies of the Unit C are more diverse than previously assumed.
The palaeolake corresponding to the deposits of the Unit B, which outcrops in La Rinconada and San Chils, had a permanent water stratification, with an anoxic lake bottom were microbial mats developed (De las Heras et al. 2003). This water stratification was probably influenced by the inferred subtropical climate that prevailed in the area (Peñalver et al. 2016). Water depth could be several tens of metres in areas away from the shore (De las Heras et al. 2003). The water was basic in character, oligo- or mesosaline and with high sulphate content (De las Heras et al. 2003). The slight salinity could have also contributed to the stratification of the water column (Peñalver et al. 2016).
The Foieta la Sarra-A palaeolake shows taphonomic and palaeoecological similarities to the lacustrine record of the Unit B, such as the fine lamination due to the presence of microbial mats at the lake bottom (which enabled the fossil preservation and is characteristic of Konservat-Lagerstätten) and the finding of plant remains and fossil insects (Peñalver et al. 2016). Interestingly, Foieta la Sarra-A shares with San Chils, but not with the widely studied La Rinconada, the presence of Daphnia (Ctenodaphnia) sp. ephippia, although only represented by two specimens in San Chils (Álvarez-Parra and Peñalver 2019), in contrast to the mass records from Foieta la Sarra-A. La Rinconada has yielded fossil record of amphibians (Peñalver et al. 2016), which have not been found in Foieta la Sarra-A. Despite that, Foieta la Sarra-A has provided charophytes and lymnaeids, not recorded in the Unit B, and the only fish remains of the Ribesalbes–Alcora Basin to date. Considering the overall data from the palaeolakes of the Unit B and Foieta la Sarra-A, the palaeoenvironment and the palaeoecology were clearly different (Fig. 9). Their palaeoenvironments differ due to a change of the water chemistry. The three main mechanisms controlling the water chemistry of lakes are atmospheric precipitation, rock dominance of the basin and evaporation-crystallisation processes (Gibbs 1970), and these three could explain the environmental dynamics of the palaeolake records of the basin. The rock dominance of the basin corresponds to the basement that crops out in the marginal areas of the basin and is eroded by the lotic waters, which arrive to the lakes formed in the central areas and set the chemistry of their waters (Gibbs 1970). The deposits of the Unit B have been tentatively dated as slightly older than the deposits of the Unit C (Peñalver et al. 2016), but it is unlikely that the rock dominance of the basin could change enough to modify the water chemistry in such a short time. Therefore, the factors that influenced a change of the water chemistry in the palaeolakes of the basin need to be further studied based on sedimentological works and geochemical analyses of the Foieta la Sarra-A rock. An increase of the humidity in the second local biozone of the Campisano ravine was proposed after the preliminary studies of the mammalian palaeoecology and isotopes in the fossiliferous localities (Ríos 2013; Crespo 2017). The faunal assemblage in the second local biozone (after Crespo et al. 2019c) apparently shows more humid preferences than in the first one (Crespo 2017; Crespo et al. 2019c). The Foieta la Sarra-A assemblage and palaeoecology confirm an increase of the humidity in the second local biozone. Indeed, there was a stable water body at least during the beginning of the second biozone, with no lacustrine sediment apparent during the first biozone. A tropical climate has been inferred for the early Miocene forest of the Campisano ravine surrounding the palaeolake (Crespo et al. 2020b).
.
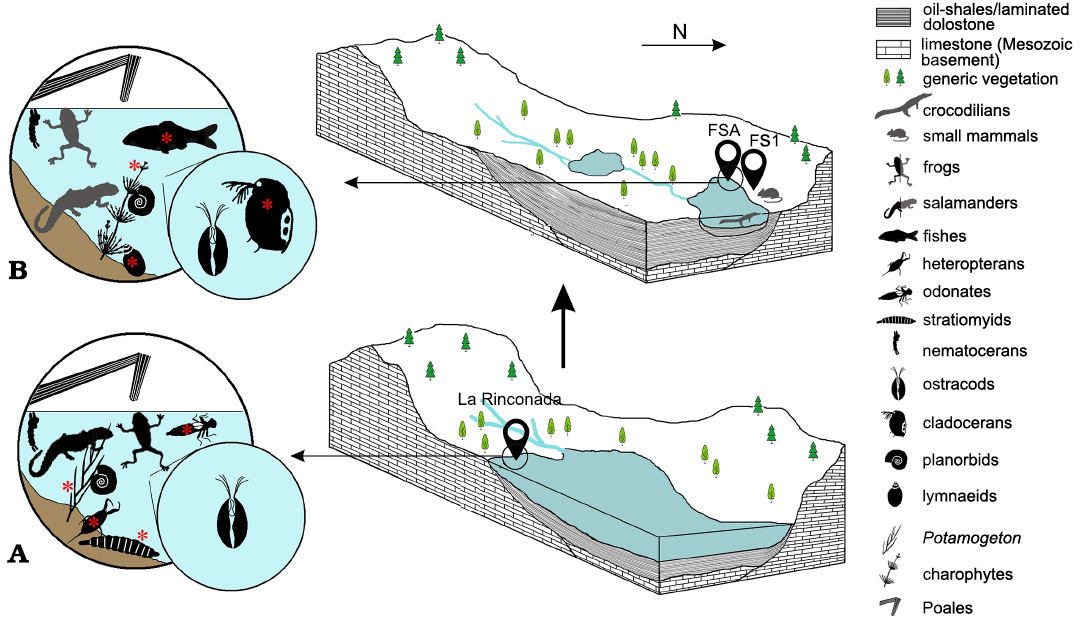
Fig. 9. Synthetic 3D representation of the aquatic environment evolution in the Ribesalbes–Alcora Basin (Castelló Province, Spain) during the early Miocene, mostly based on the La Rinconada (ca. 19 Ma, belonging to the Unit B) and Foieta la Sarra-A (ca. 16.20 Ma, belonging to the Unit C) deposits, which nowadays are separated by around 4 km. Rock deposits have been strongly simplified (e.g., breccia and sandstone are not represented). The significant differences in the aquatic biota are marked with red asterisks. Animals represented in grey correspond to the fossil record of the Foieta la Sarra-1 (FS1) mammalian locality, which are not recorded in the slabs of Foieta la Sarra-A Foieta la Sarra-A (FSA). In La Rinconada (A), heteropterans are represented by the families Nepidae and Notonectidae, odonates by the infraorders Zygoptera and Anysoptera, and nematocerans by the families Chironomidae and Chaoboridae. In Foieta la Sarra-A (B), planorbids are represented by the genera Ferrissia and Gyraulus, and nematocerans have been inferred based on chironomid larval galleries in the Foieta la Sarra-A slabs. Silhouettes not to scale.
The seven sections of the Campisano ravine, corresponding to the Unit C, include facies that represent different palaeoenvironments (Crespo 2017; Crespo et al. 2019c). Most of the deposits are mudstone beds with variegated colours, suggesting that sediments were exposed to subaerial conditions. Sandstone and microconglomerate channels are interpreted as the fluvial contribution to the distal parts of alluvial fans in a low-energy environment. These channels correspond, for instance, to specific high energy episodes related to large floods caused by storms. Thinner tabulate sandstone layers that cover a larger lateral surface, were probably deposited in the floodplains (Crespo 2017; Crespo et al. 2019c). Massive limestone beds were probably deposited in shallow lacustrine and marshy environments subjected to the fluctuation of the water level. Furthermore, several levels of the sections from the Campisano ravine indicate particular environments. These include: (i) dark lenticular beds with black-lignitiferous mudstone probably related to a palustrine environment (in the levels Mas dels Coixos-3 and Mas d’Antolino B-11), (ii) palaeosols associated with longer intervals of subaerial exposure (in the level Mas dels Coixos-4-6), (iii) fine ferruginous and red-coloured levels probably associated to low sedimentation rate episodes and combined with abundant iron supplies (in the level Araia Cantera Sud-2), (iv) evaporitic deposits in the form of gypsum layers formed on the edge of a lake (in the levels Mas d’Antolino B-0A and Mas d’Antolino B-0B), and (v) laminated limestone beds as those from Foieta la Sarra-A (Crespo 2017; Crespo et al. 2019c). Particularly, an increase of the energy in the environment is observed in the Foieta la Sarra section (Fig. 1B), as the laminated limestone beds are at the bottom of the deposit (Foieta la Sarra-A) and sandy mudstone is present at the top (Foieta la Sarra-1). Accordingly, the Campisano ravine deposits reflect the complexity of the palaeoenvironments of a palaeolake and its surroundings. The Foieta la Sarra-A locality in particular shows the environmental evolution of a palaeolake that occurred in the basin in comparison with the record of the Unit B.
Conclusions
Foieta la Sarra-A corresponds to the record of a different palaeolake to that of the Unit B, or a later phase of the same, providing evidence of the palaeoenvironmental dynamics of the Ribesalbes–Alcora Basin during the early Miocene. The Foieta la Sarra-A palaeolake had a different water chemistry in comparison to that of the Unit B. Such factor explains the presence of mass occurrences of ostracods and Daphnia (Ctenodaphnia) sp. ephippia. The occurrence of characeite beds indicates that the palaeolake depth was not significant (i.e., a few metres). Moreover, the water level underwent periodic fluctuations, favouring the mass production of Daphnia (Ctenodaphnia) sp. ephippia in the most marginal palaeolake areas under stress conditions. The scarcity of insects could be related to the abundance of fishes as predators, to preservation, and/or to sampling biases.
A new palaeontological work is planned in the near future in order to increase the known fossil assemblage from Foieta la Sarra-A. Furthermore, a sedimentological and geochemical approach would shed further light on the palaeolake environment. The search of new lacustrine limestone levels in the stratigraphy of the Campisano ravine, as well as new palaeontological excavations in La Rinconada and San Chils would provide new data on the complex palaeoecology of the Ribesalbes–Alcora Basin during the early Miocene.
Acknowledgements
We would like to thank Esther Bueno (Universitat de València, Spain), Óscar Caballero (Museo Nacional de Ciencias Naturales-Consejo Superior de Investigaciones Científicas, Madrid, Spain), Mireia Costa-Pérez (Universitat de València, Spain), María Victoria Paredes-Aliaga (Institut Cavanilles de Biodiversitat y Biologia Evolutiva, Burjassot, Spain) and Andreu Vilaplana-Climent (Universitat de València, Spain) for their participation during the prospection work. María Victoria Paredes-Aliaga provided the photographs for Fig. 1C, D. We are grateful to Eduardo Barrón (Instituto Geológico y Minero de España, Madrid, Spain) for his observations about the palaeobotanical record. We thank Thomas A. Neubauer (Justus-Liebig-Universität Gießen, Gießen, Germany) for his observations about the molluscan record, which enabled to refine the reconstructions and improved the original manuscript. SG-C wants to thank Mauricio Bigurrarena Ojeda (Universidad Nacional de La Plata, La Plata, Argentina) for valuable discussions on fish taphonomy. We are indebted to Ricardo Pérez-de la Fuente (Oxford University Museum of Natural History, UK) for the proofreading of the manuscript. SÁ-P thanks the support from the Secretary of Universities and Research of the Government of Catalonia (Spain) and the European Social Fund. PM acknowledges the Spanish Research Project PGC2018-094122-B100 (AEI/ FEDER, UE). VDC is the beneficiary of a postdoctoral fellowship from the Argentinian Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET). We are grateful to the corrections and suggestions from the editors, Nevena Andrić-Tomašević (Universität Tübingen, Germany), and an anonymous reviewer, which have significantly improved the manuscript. This study was presented as an abstract at the 2nd Palaeontological Virtual Congress, to whose organisers we are grateful for their initiative and thank for their invitation to participate in this Special Volume.
References
Adam, W. 1960. Faune de Belgique. Mollusques. Tome I. Mollusques terrestres et dulcicoles. 402 pp. Patrimoine de l’Institut royal des Sciences naturelles de Belgique, Brussels.
Agustí, J., Anadón, P., Ginsburg, L., Mein, P., and Moissenet, E. 1988. Araya et Mira: nouveaux gisements de mammifères dans le Miocène Infèrieur-Moyen des Chaînes Ibériques orientales et méditerranéennes. Conséquences stratigraphiques et structurales. Paleontologia i Evolució 22: 83–101.
Alcaraz, C. and García-Berthou, E. 2007. Food of an endangered cyprinodont (Aphanius iberus): ontogenetic diet shift and prey electivity. Environmental Biology of Fishes 78: 193–207. Crossref
Álvarez-Parra, S. and Peñalver, E. 2019. Palaeoentomological study of the lacustrine oil-shales of the lower Miocene San Chils locality (Ribesalbes–Alcora Basin, Castellón province, Spain). Spanish Journal of Palaeontology 34: 187–203. Crossref
Anadón, P., Cabrera, L., Julià, R., Roca, E., and Rosell, L. 1989. Lacustrine oil-shale basins in Tertiary grabens from NE Spain (Western European rift system). Palaeogeography, Palaeoclimatology, Palaeoecology 70: 7–28. Crossref
Arconada, B. and Ramos, M.A. 2003. The Ibero-Balearic region: one of the areas of highest Hydrobiidae (Gastropoda, Prosobranchia, Rissooidea) diversity in Europe. Graellsia 59: 91–104. Crossref
Barrón, E. and Diéguez, C. 2001. Estudio macroflorístico del Mioceno Inferior lacustre de la Cuenca de Rubielos de Mora (Teruel, España). Boletín Geológico y Minero 112: 13–56.
Barrón, E. and Postigo-Mijarra, J.M. 2011. Early Miocene fluvial-lacustrine and swamp vegetation of La Rinconada mine (Ribesalbes–Alcora Basin, Eastern Spain). Review of Palaeobotany and Palynology 165: 11–26. Crossref
Bauld, J. 1981. Occurrence of benthic microbial mats in saline lakes. Hydrobiologia 81: 87–111. Crossref
Bräger, Z. and Moritz, T. 2016. A scale atlas for common Mediterranean teleost fishes. Vertebrate Zoology 66: 275–386.
Bräger, Z., Staszny, Á., Mertzen, M., Moritz, T., and Horváth, G. 2017. Fish scale identification: from individual to species-specific shape variability. Acta Ichthyologica et Piscatoria 47: 331–338. Crossref
Burky, A. 1971. Biomass turnover, respiration, and interpopulation variation in the stream limpet Ferrissia rivularis (Say). Ecological Monographs 41: 235–251. Crossref
Bustillo, M.Á., Díaz-Molina, M., López-García, M.J., Delclòs, X., Peláez-Campomanes, P., Peñalver, E., Rodríguez-Talavera, R., and Sanchiz, B. 2017. Geology and paleontology of Tresjuncos (Cuenca, Spain), a new diatomaceous deposit with Konservat-Lagerstätte characteristics from the European late Miocene. Journal of Iberian Geology 43: 395–411. Crossref
Cavin, L. 2017. Freshwater Fishes: 250 Millon Years of Evolutionary History. 199 pp. ISTE Press, Elsevier, London.
Cohen, A.S. 2003. Paleolimnology: The History and Evolution of Lake Systems. 528 pp. Oxford University Press, New York. Crossref
Costa-Pérez, M., Álvarez-Parra, S., Paredes-Aliaga, M.V., Caballero, Ó., Bueno, E., Vilaplana-Climent, A., and Crespo, V.D. 2019. Valoración patrimonial de los yacimientos del Mioceno inferior del Barranco de Campisano de la Cuenca de Ribesalbes–Alcora (Araia d’Alcora, Castelló, España). Spanish Journal of Palaeontology 34: 125–130. Crossref
Crespo, V.D. 2017. Los mamíferos del Mioceno Inferior de la Cuenca de Ribesalbes–Alcora (Castelló, España). 695 pp. Unpublished Ph.D. Thesis, Universitat de València, Valencia.
Crespo, V.D., Fagoaga, A., Montoya, P., and Ruiz-Sánchez, F.J. 2019a. Oldtimers and newcomers: the shrews and heterosoricids from the Ribesalbes–Alcora Basin (east of Spain). Palaeontologia Electronica 22.3.64: 1–22. Crossref
Crespo, V.D., Fagoaga, A., Ruiz-Sánchez, F.J., and Montoya, P. 2021a. Diggers, gliders and runners: the squirrels from the Ribesalbes–Alcora Basin (East of Spain). Bulletin of Geosciences 96: 83–97. Cossref
Crespo V.D., Gamonal, A., Montoya, P., and Ruiz-Sánchez, F.J. 2021b. The eomyids from the Ribesalbes-Alcora Basin (early Miocene, Iberian Peninsula) and their biostratigraphic and paleoecologic implications. Rivista Italiana di Paleontologia e Stratigrafia 127: 497–514.
Crespo, V.D., Goin, F.J., Montoya, P., and Ruiz-Sánchez, F.J. 2020a. Early Miocene marsupialiforms, gymnures, and hedgehogs from Ribesalbes–Alcora Basin (Spain). Journal of Paleontology 94: 1213–1227. Crossref
Crespo, V.D., Marquina-Blasco, R., Ruiz-Sánchez, F.J., and Montoya, P. 2019b. An unusual insectivore assemblage from the early Miocene of southwestern Europe: the talpids and dimylids from the Ribesalbes–Alcora Basin (Spain). Comptes Rendus Palevol 18: 407–416. Crossref
Crespo, V.D., Sevilla, P., Montoya, P., and Ruiz-Sánchez, F.J. 2020b. A relict tropical forest bat assemblage from the early Miocene of the Ribesalbes–Alcora Basin (Castelló, Spain). Earth and Environmental Science Transactions of the Royal Society of Edinburgh 111 (4): 247–258. Crossref
Crespo, V.D., Suárez-Hernando, O., Murelaga, X., Ruiz-Sánchez, F.J., and Montoya, P. 2019c. Early Miocene mammal assemblages from the Campisano ravine in the Ribesalbes–Alcora Basin (E Spain). Journal of Iberian Geology 45: 181–194. Crossref
Daniels, R. 1996. Guide to the identification of scales of inland fishes of Northeastern North America. New York State Museum Bulletin 488: 1–105.
De las Heras, F.X.C., Anadón, P., and Cabrera, L. 2003. Biomarker record variations in lacustrine coals and oil shales: contribution from Tertiary basins in NE Spain. In: B.L. Valero-Garcés (ed.), Limnogeology in Spain: a Tribute to Kerry R. Kelts, 187–228. Consejo Superior de Investigaciones Científicas, Madrid.
Dudgeon, D. and Yipp, M. 1985. The diets of Hong Kong freshwater gastropods. In: B. Morton and D. Dudgeon (eds.), Proceedings of Second International Workshop on the Malacofauna of Hong Kong and Southern China, 491–509. Hong Kong University Press, Hong Kong.
Ebert, D. 2005. Ecology, Epidemiology and Evolution of Parasitism in Daphnia. 98 pp. National Center for Biotechnology Information, Bethesda.
Fetcher, R. and Falkner, G. 1993. Moluscos. 287 pp. Blume, Barcelona.
Freytet, P. and Verrecchia, E.P. 2002. Lacustrine and palustrine carbonate petrography: an overview. Journal of Paleolimnology 27: 221–237. Crossref
Furió, M., Ruiz-Sánchez, F.J., Crespo, V.D., Freudenthal, M., and Montoya, P. 2012. The southernmost Miocene occurrence of the last European herpetotheriid Amphiperatherium frequens (Metatheria, Mammalia). Comptes Rendus Palevol 11: 371–377. Crossref
García-Paredes, I., Álvarez-Sierra, M.Á., Van den Hoek Ostende, L.W., Hernández-Ballarín, V., Hordijk, K., López-Guerrero, P., Oliver, A., and Peláez-Campomanes, P. 2016. The Aragonian and Vallesian high-resolution micromammal succession from the Calatayud-Montalbán Basin (Aragón, Spain). Comptes Rendus Palevol 15: 781–789. Crossref
Gaudant, J. 1993. Un exemple de “régression évolutive” chez des poisons cyprinodontidae du miocène supérieur d’Espagne: Apanius illunensis nov. sp. Geobios 26: 449–454. Crossref
Gholami, Z., Teimori, A., Esmaili, H.R., Schultz-Mirbach, T., and Reichenbacher, B. 2013. Scale surface microstructure and scale size in the tooth-carp genus Aphanius (Teleostei: Cyprinodontiformes) from endorheic basins in Southwest Iran. Zootaxa 3619: 467–490. Crossref
Gibbs, R.J. 1970. Mechanisms controlling world water chemistry. Science 170: 1088–1090. Crossref
Glöer, P. 2002. Die Tierwelt Deutschlands, 73. Teil: Die Süßwassergastropoden Nord- und Mitteleuropas. Bestimmungsschlüssel, Lebensweise, Verbreitung. 327 pp. ConchBooks, Hackenheim.
Hann, B.J., Mundy, C.J., and Goldsborough, L.G. 2001. Snail-periphyton interactions in a prairie lacustrine wetland. Hydrobiologia 457: 167–175. Crossref
Harzhauser, M. and Kowalke, T. 2002. Sarmatian (late middle Miocene) gastropod assemblages of the Central Paratethys. Facies 46: 57–82. Crossref
Harzhauser, M., Neubauer, T.A., Gross, M., and Binder, H. 2014. The early middle Miocene mollusc fauna of Lake Rein (Eastern Alps, Austria). Palaeontographica, Abteilung A: Palaeozoology—Stratigraphy 302 (1–6): 1–71. Crossref
Hunt, A.P. and Lucas, S.G. 2012. Classification of vertebrate coprolites and related trace fossils. New Mexico Museum of Natural History and Science 57: 137–146.
Kesler, D.H. 1981. Periphyton grazing by Amnicola limosa: an enclosure-exclosure experiment. Journal of Freshwater Ecology 1: 51–59. Crossref
Kesler, D.H., Jokinen, E.H., and Munns, W.R. 1986. Trophic preferences and feeding morphology of two pulmonate snail species from a small New England pond, U.S.A. Canadian Journal of Zoology 64: 2570–2575. Crossref
Kotov, A.A. and Taylor, D.J. 2011. Mesozoic fossils (>145 Mya) suggest the antiquity of the subgenera of Daphnia and their coevolution with chaoborid predators. BMC Evolutionary Biology 11: 129. Crossref
Kowalke, T. and Reichenbacher, B. 2005. Early Miocene (Ottnangian) Mollusca of the Western Paratethys—ontogenetic strategies and palaeo-environments. Geobios 38: 609–635. Crossref
Lagler, K.F. 1947. Lepidological Studies 1. Scale Characters of the Families of Great Lakes Fishes. Transactions of the American Microscopical Society 66: 149–171. Crossref
Leonardos, I. 2008. The feeding ecology of Aphanius fasciatus (Valenciennes, 1821) in the lagoonal system of Messolongi (western Greece). Scientia Marina 72: 393–401. Crossref
Mandic, O., Hajek-Tadesse, V., Bakrač, K., Reichenbacher, B., Grizelj, A., and Miknić, M. 2019. Multiproxy reconstruction of the middle Miocene Požega palaeolake in the Southern Pannonian Basin (NE Croatia) prior to the Badenian transgression of the Central Paratethys Sea. Palaeogeography, Palaeoclimatology, Palaeoecology 516: 203–219. Crossref
Martín-Closas, C. and Delclòs, X. 2007. The Miocene paleolake of La Cerdanya (Eastern Pyrenees). Geo-Guías 3: 181–205.
McNamara, M.E., Orr, P J., Alcalá, L., Anadón, P., and Peñalver, E. 2012. What controls the taphonomy of exceptionally preserved taxa—environment or biology? A case study using frogs from the Miocene Libros Konservat-Lagerstätte (Teruel, Spain). Palaios 27: 63–77. Crossref
Nelson, J.S., Grande, T., and Wilson, M.V.H. 2016. Fishes of the World. Fifth edition. 707 pp. Wiley, New Jersey. Crossref
Neubauer, T.A., Mandic, O., and Harzhauser, M. 2011. Middle Miocene freshwater mollusks from Lake Sinj (Dinaride Lake System, SE Croatia; Langhian). Archiv für Molluskenkunde: International Journal of Malacology 140: 201–237. Crossref
Neubauer, T.A., Mandic, O., and Harzhauser, M. 2013. The Middle Miocene freshwater mollusk fauna of Lake Gacko (SE Bosnia and Herzegovina): taxonomic revision and paleoenvironmental analysis. Fossil Record 16: 77–96. Crossref
Neubauer, T.A., Harzhauser, M., Georgopoulou, E., Kroh, A., and Mandic, O. 2015a. Tectonics, climate, and the rise and demise of continental aquatic species richness hotspots. Proceedings of the National Academy of Sciences 112: 11478–11483. Crossref
Neubauer, T.A., Harzhauser, M., Kroh, A., Georgopoulou, E., and Mandic, O. 2015b. A gastropod-based biogeographic scheme for the European Neogene freshwater systems. Earth-Science Reviews 143: 98–116. Crossref
Neubauer, T.A., Mandic, O., and Harzhauser, M. 2015c. The freshwater mollusk fauna of the middle Miocene Lake Drniš (Dinaride Lake System, Croatia): a taxonomic and systematic revision. Austrian Journal of Earth Sciences 108: 15–67. Crossref
Oates, D.W., Krings, L.M., and Ditz, K.L. 1993. Field Manual for the Identification of Selected North American Freshwater Fish by Fillets and Scales. 181 pp. University of Nebraska, Lincoln.
Okland, J. 1990. Lakes and Snails. Environment and Gastropoda in 1500 Norwegian Lakes. 515 pp. Universal Book Service/Dr. W. Backbuys, Oegstgeest.
Oliver, D.R. 1981. Chironomidae. In: J.F. McAlpine, B.V. Peterson, G.E. Shewell, H.J. Teskey, J.R. Vockeroth, and D.M. Wood (eds.), Manual of Nearctic Diptera, Vol. 1, 27 (29), 423–458. Research Branch Agriculture Canada, Ottawa.
Patterson, R.T., Wright, C., Chang, A.S., Taylor, L.A., Lyons, P.D., Dallimore, A., and Kumar, A. 2002. Atlas of common squamatological (fish scale) material in coastal British Columbia and an assessment of the utility of various scale types in paleofisheries reconstruction. Palaeontologia Electronica 4: 1–88.
Peñalver, E. 2002. Los insectos dípteros del Mioceno del Este de la Península Ibérica; Rubielos de Mora, Ribesalbes y Bicorp. Tafonomía y sistemática. 548 pp. PhD Thesis. Universitat de València, Valencia.
Peñalver, E. and Gaudant, J. 2010. Limnic food web and salinity of the upper Miocene Bicorb palaeolake (eastern Spain). Palaeogeography, Palaeoclimatology, Palaeoecology 297: 683–696. Crossref
Peñalver, E., Barrón, E., Postigo Mijarra, J.M., García Vives, J.A., and Saura Vilar, M. 2016. El paleolago de Ribesalbes. Un ecosistema de hace 19 millones de años. 201 pp. Servicio de Publicaciones, Diputació de Castelló and Instituto Geológico y Minero de España, Ministerio de Economía y Competitividad, Castellón de la Plana.
Peñalver, E., Martínez-Delclòs, X., and De Renzi, M. 1996. Registro de pulgas de agua [Cladocera: Daphniidae: Daphnia (Ctenodaphnia)] en el Mioceno de Rubielos de Mora (Teruel, España). Comunicaciones de la II Reunión de Tafonomía y Fosilización 1: 311–317.
Popova, E.V., Petrusek, A., Kořínek, V., Mergeay, J., Bekker, E.I., Karabanov, D.P., Galimov, Y.R., Neretina, T.V., Taylor, D.J., and Kotov, A.A. 2016. Revision of the Old World Daphnia (Ctenodaphnia) similis group (Cladocera: Daphniidae). Zootaxa 4161: 1–40. Crossref
Postigo-Mijarra, J.M. and Barrón, E. 2013. Zonal plant communities of the Ribesalbes–Alcora Basin (La Rinconada mine, eastern Spain) during the early Miocene. Botanical Journal of the Linnean Society 173: 153–174. Crossref
Pyron, M. and Brown, K.M. 2015. Introduction to Mollusca and the Class Gastropoda. In: J.H. Thorp and D.C. Rogers (eds.), Thorp and Covich’s Freshwater Invertebrates, Ecology and General Biology, Fourth Edition, 18, 383–421. Academic Press, London. Crossref
Quate, L.W. and Vockeroth, J.R. 1981. Psychodidae. In: J.F. McAlpine, B.V. Peterson, G.E. Shewell, H.J. Teskey, J.R. Vockeroth, and D.M. Wood (eds.), Manual of Nearctic Diptera, Vol. 1, 27 (17), 293–300. Research Branch Agriculture Canada, Ottawa.
Radea, C., Louvrou, I., Konstantinos Bakolitsas, K., and Economou-Amilli, A. 2017. Local endemic and threatened freshwater hydrobiids of Western Greece: elucidation of anatomy and new records. Folia Malacologica 25: 3–13. Crossref
Ríos, M. 2013. Estudio multi-isotópico de la paleoecología y la paleoclimatología de la Cuenca de Ribesalbes–Alcora (Castellón, España) durante el Óptimo Climático del Mioceno. 76 pp. Unpublished MSc Thesis, Universitat de València, Valencia.
Rodríguez-Aranda, J.P. and Calvo, J.P. 1998. Trace fossils and rhizoliths as a tool for sedimentological and palaeoenvironmental analysis of ancient continental evaporite successions. Palaeogeography, Palaeoclimatology, Palaeoecology 140: 383–399. Crossref
Schultze, H.-P. 2015. Scales, enamel, cosmine, ganoine, and early osteichthyans. Comptes Rendus Palevol 15: 83–102. Crossref
Soulié-Märsche, I., Bieda, S., Lafond, R., Maley, J., Baitoudji, M., Vincent, P.M., and Faure, H. 2010. Charophytes as bio-indicators for lake level high stand at “Trou au Natron”, Tibesti, Chad, during the Late Pleistocene. Global Planetary Change 72: 334–340. Crossref
Talbot, M.R. and Allen, P.A. 1996. Lakes. In: H.G. Reading (ed.), Sedimentary Environments: Processes, Facies and Stratigraphy, 5–36. Blackwell Publishing, Oxford.
Teimori, A., Esmaeli, H.R., Erpenbeck, D., and Reichenbacher, B. 2014. A new and unique species of the genus Aphanius Nardo, 1827 (Teleostei: Cyprinodontidae) from Southern Iran: a case of regressive evolution. Zoologischer Anzeiger 253: 327–337. Crossref
Teimori, A., Motamedi, M., and Manizadeh, N. 2017. Microstructural characterization of the body key scale morphology in six Iranian endemic Aphanius species (Cyprinodontidae): their taxonomic and evolutionary significance. Journal of Ichthyology 57: 533–546. Crossref
Van der Meulen, A.J., Paredes, I.G., Sierra, M.Á., van den Hoek Ostende, L.W., and Hordijk, K. 2012. Updated Aragonian biostratigraphy: small mammal distribution and its implications for the Miocene European Chronology. Geologica Acta 10: 159–179.
Van der Velde, G. 1991. Population dynamics and population structure of Ferrissia wautieri (Mirolli, 1960) (Gastropoda, Ancylidae) in a pond near Nijmegen (The Netherlands). Hydrobiological Bulletin 24: 141–144. Crossref
Vasilyan, D. 2020. Fish, amphibian and reptilian assemblage from the middle Miocene locality Gračanica—Bugojno palaeolake, Bosnia and Herzegovina. Palaeobiodiversity and Palaeoenvironments 100: 437–455. Crossref
Vecchioni, L., Marrone, F., Aculeo, M., and Arizza, V. 2017. Are there autochthonous Ferrissia (Mollusca: Planorbidae) in the Palaearctic? Molecular evidence of a widespread North American invasion of the Old World. The European Zoological Journal 84: 411–419. Crossref
Vilanova y Piera, J. 1859. Memoria geognóstico-agrícola sobre la provincia de Castellón. Memorias de la Real Academia de Ciencias de Madrid 4: 577–803.
Walshe, B.M. 1951. The feeding habits of certain chironomid larvae (subfamily Tendipedinae). Proceedings of the Zoological Society of London 121: 63–79. Crossref
Wautier, J. 1975. Présence d’Espèces du genre Ferrissia Walker, 1903 (Gastropoda, Basommatophora) dans le Néogène du bassin rhodanien (France). Geobios 8: 423–433. Crossref
Welter-Schultes, F. 2012. European Non-marine Molluscs, A Guide For Species Identification. 760 pp. Planet Poster Editions, Göttingen.
Wilby, P.R., Briggs, D.E., Bernier, P., and Gaillard, C. 1996. Role of microbial mats in the fossilization of soft tissues. Geology 24: 787–790. Crossref
Acta Palaeontol. Pol. 66 (Supplement to 3): 13–30, 2021
https://doi.org/10.4202/app.00842.2020