
First known gigantic sea turtle from the Maastrichtian deposits in Egypt
GEBELY ABDEL-MAKSOUD ABU EL-KHEIR, MOHAMED KORANY ABDELGAWAD, and WALID GAMAL KASSAB
Abu El-Kheir, G.A.-M., AbdelGawad, M.K., and Kassab, W.G. 2021. First known gigantic sea turtle from the Maastrichtian deposits in Egypt. Acta Palaeontologica Polonica 66 (2): 349–355.
The Maastrichtian deposits in Egypt and generally in Africa contain very few records of chelonioids. This scarcity hinders understanding the evolution and extinction of African chelonioids during this age. The Dakhla Basin in Egypt presents excellent Maastrichtian outcrops that consist of highly fossiliferous, calcareous siltstone and sandstone layers rich in ammonites, bivalves (Exogyra overwagi, Pycnodonta vesicularis, pectinids), gastropods, echinoids, corals, fossilized fruits of mangrove plants (Nypa) and vertebrate remains. The well-preserved humerus of giant turtle and other vertebrate remains have been discovered in the early late Maastrichtian Ammonite Hill Member of the Dakhla Formation in the Abu Minqar area, Southern Western Desert, Egypt. The Ammonite Hill Member was deposited in very shallow coastal, inner neritic to littoral environments. The well-preserved giant humerus represents the first record of gigantic Mesozoic chelonioids in Africa and the second record of chelonioids in the Maastrichtian deposits in Egypt. The present specimen contributes to fill the missing gaps in the history of Egyptian turtles from the Cretaceous through the Cenozoic.
Key words: Testudines, Panchelonioidea, Cheloniidae, humerus, Cretaceous, Maastrichtian, Dakhla, Abu Minqar, Egypt.
Gebely Abdel-Maksoud Abu El-Kheir [gebely2006@sci.nvu.edu.eg; ORCID: https://orcid.org/0000-0001-5905-8790], Department of Geology, Faculty of Sciences, New Valley University, New Valley, Kharga, Egypt.
Mohamed Korany AbdelGawad [mkabdelgawad@gmail.com; mkabdelgawad@sci.cu.edu.eg; ORCID: https://orcid.org/0000-0002-3229-3800] and Walid Gamal Kassab [wkassab@sci.cu.edu.eg; ORCID: https://orcid.org/0000-0001-9882-9607] (corresponding author), Department of Geology, Faculty of Science, Cairo University, Giza, Egypt.
Received 23 October 2020, accepted 23 January 2021, available online 9 June 2021.
Copyright © 2021 G. Abdel-Maksoud Abu El-Kheir et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
The southern Western Desert of Egypt exhibits good exposures of successive Cretaceous and early Paleocene deposits. These deposits were formed in fluvial, fluvio-marine, lacustrine and marine environments. The early to middle Maastrichtian to early Paleocene deposits are represented by approximately 200-m-thick alternating shale and sandstone units that formed in inner neritic to lagoonal environments (Tantawy et al. 2001). The Ammonite Hill Member at the base of the Dakhla Formation in the village of Abu Minqar, Farafra (Zittel 1883; Lapparent de Broin and Werner 1998), yielded well-preserved turtle shell fragments, fish bones, Mosasaurus sp. and shark teeth (Lapparent de Broin and Werner 1998). Six turtle taxa were discovered by Project 69 “Geoscientific problems in arid and semiarid areas”, Technical University of Berlin, in the calcareous sandstone layers of the Ammonite Hill Member near the Bir Abu Minqar area, in particular the bottremydids Zolhafah bella, Taphrosphys cf. sulcatus, cf. Taphrosphys sp., Arenila krebsi, and cf. Arenila krebsi, and the chelonioid Tasbacka sp. (Lapparent de Broin and Werner 1998). These discoveries are the first records of reptiles in the Maastrichtian deposits of Egypt.
Our present work describes the further discovery of a marine turtle (Panchelonioidea) in the Maastrichtian deposits in the southern Western Desert of Egypt, which is represented by a well-preserved large humerus collected from the Ammonite Hill Member, Dakhla Formation north of Bir Abu Minqar behind Qaret Selmi (Fig. 1).
Institutional abbreviations.—NVP, New Valley Vertebrate Paleontology Center, New Valley University, Kharga, Egypt.
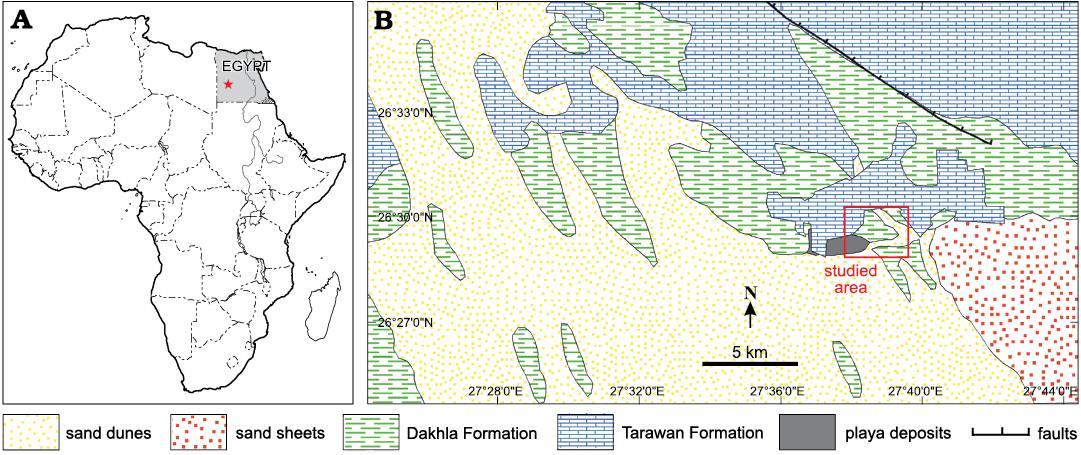
Fig. 1. A. Location map of the south Western Desert of Egypt (star). B. Geologic map of the studied area at Abu Minqar.
Geological setting
The Bir Abu Minqar section (26º31ʹ05ʺN, 27º39ʹ43ʺE) is located approximately 260 km northwest of Mut Dakhla, at the curvature of the Dakhla-Farafra road, and north of Abu Minqar Village in the southern Western Desert (Fig. 1). The sediments in this section are composed of two rock units (Fig. 2): the Dakhla Formation (Maastrichtian) and the Tarawan Formation (early Paleocene). These two rock units are cut by many sedimentary discontinuity surfaces.
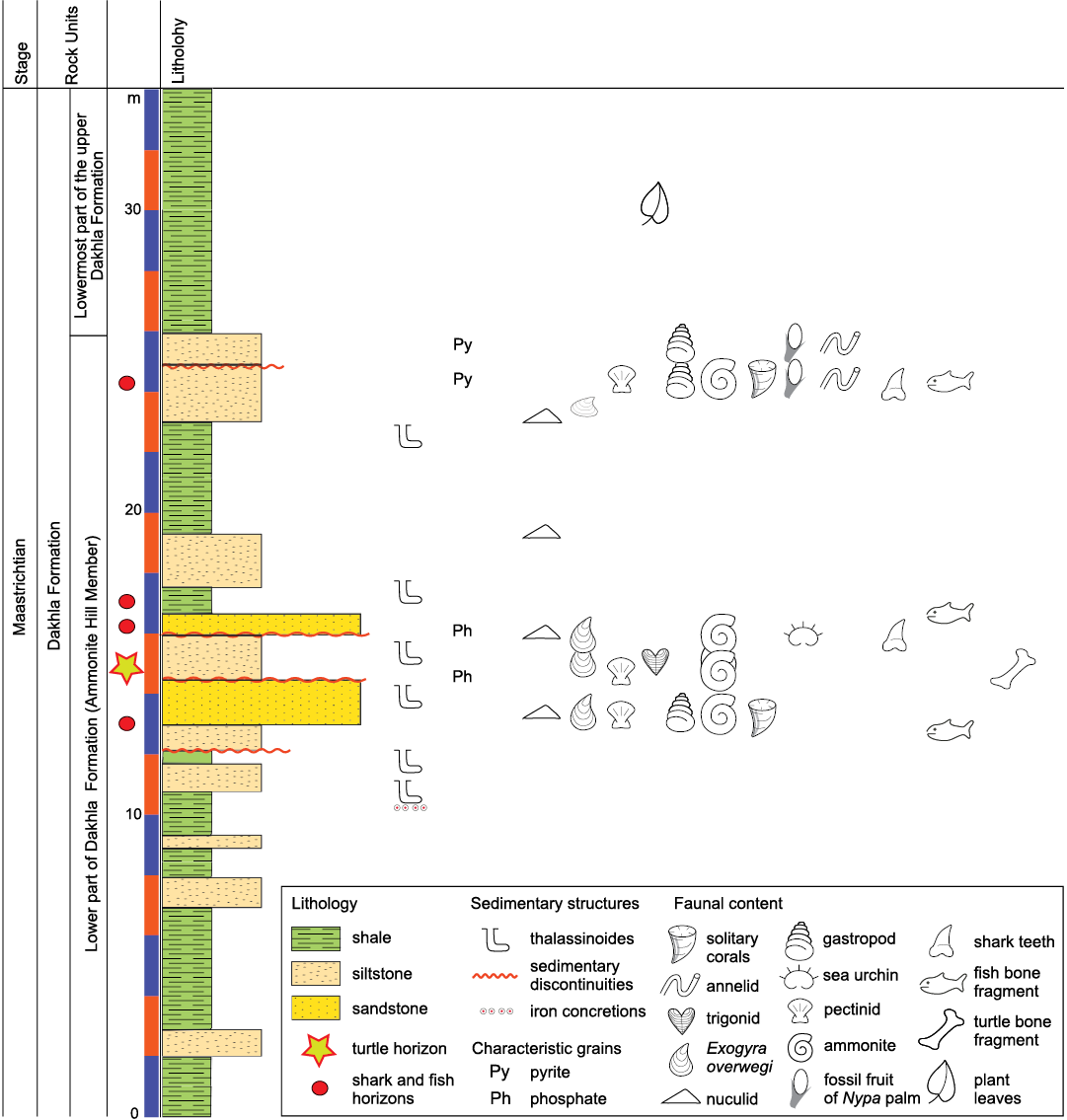
Fig. 2. Stratigraphic section of Qaret Selmi, north Abu Miqar village, showing the Maastrichtian–Paleocene successions of Dakhla Formation.
The Dakhla Formation (Said 1961) outcrops not only in the Dakhla Oasis but also in vast areas all over southern Egypt. In the Bir Abu Minqar section, the Dahkla Formation is subdivided into lower and upper parts (Barthel and Herrmann-Degen 1981; Fig. 3).
The lower part of the Dakhla Formation is chiefly composed of dark shale interbedded with highly fossiliferous calcareous siltstone and sandstone (named the Ammonite Hill Member, introduced by Barthel and Herrmann-Degen 1981). The fossiliferous siltstone and sandstone layers show either intensive bioturbation or enormous concentrations of ammonites (e.g., Baculites sp.), bivalves (e.g., Exogyra overwagi, Pycnodonta vesicularis, pectinids), gastropods, echinoids, corals, fossilized fruits of mangrove plants (Nypa) and vertebrate remains (Fig. 3). El Hedeny et al. (2020) introduce two types of borings Teredolites clavatus and Apectoichnus longissimus in the disconnected aggregates of allochthonous fossil Nypa fruits. The diversity, as well as the abundance of this faunal assemblage, reflects deposition in well-oxygenated shallow waters in a nearshore or lagoonal environment during sea-level lowstand periods (Tantawy et al. 2001). In the Bir Abu Minqar area, all vertebrate remains, including those of marine reptiles (turtles), bony fishes, and shark teeth, were found in the lower part of the Dakhla Formation. Westward of Abu Minqar, the occurrence of vertebrate remains gradually increases, reflecting increasing proximity to the paleo-shoreline and increasing terrestrial input (Lapparent de Broin and Werner 1998). The abundance of Nypa fruits indicates that the paleoenvironment involved brackish water (lagoons or estuaries) and that the paleoclimatic conditions were tropical (El-Soughier et al. 2011). The lower part of the Dakhla Formation is dated to the Maastrichtian based on the occurrence of the ammonite Libycoceras ismaelis and a large number of Exogyra overwegi. More recently, Tantawy et al. (2001) assigned an early Maastrichtian age to the lower part of the Dakhla Formation due to its equivalency to the lower Maastrichtian planktonic foraminiferal zone CF7 and the calcareous nannofossil subzone CC25a.

Fig. 3. A. General view of Qaret Selmi, north Abu Minqar, showing the Dakhla and Tarawan formations. B. The fossiliferous siltstone and sandstone layer, showing bioturbations or enormous concentrations of ammonites (e.g., Baculites sp.), bivalves (e.g., Exogyra overwagi, Pycnodonta vesicularis, pectinids), gastropods, echinoids, corals, fossilized fruits of mangrove palm (Nypa) and vertebrate remains.
The upper part of the Dakhla Formation consists mainly of green shale that is poor in fossils, except for the presence of plant remains, indicating that sedimentation during sea-level highstands in inner neritic to lagoonal environments was characterized by dysaerobic or low-oxygen conditions. Tantawy et al. (2001) assigned a early late Maastrichtian age for the upper part of the Dakhla Formation where this part can be correlated with the early upper Maastrichtian planktonic foraminiferal zone CF6 and the calcareous nannofossil subzone CC25b. The clastic sediment source of the Dakhla Formation was derived primarily from the tectonic activity of the Gilf El Kebir spur to the southwest of Dakhla and the Bahariya arch, according to Tantawy et al. (2001). Uplift in the region during the Maastrichtian–Paleocene resulted in major erosive sedimentary unconformity surfaces.
The Tarawan Formation (Awad and Ghobrial 1966) forms the topmost part of the Abu Minqar section, unconformably overlying the Dakhla Formation. This unconformity is indicated by the presence of an irregular surface with bioturbated marl to silty clay with limestone nodules and ferruginous sandstone at the top of the Dakhla Formation. It is composed mainly of chalk and chalky limestone with few marl and shale intercalations. The chalk is snow white to yellowish white, hard, thickly bedded, highly enriched with dwarfed solitary corals and wall forming. This formation is correlated to the lower Palaeocene planktonic foraminiferal zone P1d and the calcareous nannofossil zone NP4 (Tantawy et al. 2001).
Material and methods
The NVP010 is a giant turtle humerus and was collected by the authors in the spring of 2018 from the Maastrichtian layers of the Dakhla Formation at Abu Minqar Village, approximately 420 km west of the Kharga oasis in the southern Western Desert of Egypt. It was almost completely embedded in sediments except for the ventro-proximal end, which was affected by the erosive factors, causing complete loss of the lateral process and material between the medial process and the head of the humerus. The specimen has two separate fractures, one in the middle of the shaft and another one behind the head of the humerus. The specimen exhibits a very fine ventral surface of the cortical bone coating the cancellous bone of the humerus, except for some parts that have been abraded.
Systematic palaeontology
Testudines Batsch, 1788
Cryptodira Cope, 1868
Panchelonioidea Joyce, Parham, and Gauthier, 2004 (sensu Joyce et al. 2021)
Gen. et sp. indet.
Fig. 4.
Material.—Humerus (NVP010) from Maastrichtian layers of the Dakhla Formation at Abu Minqar Village, approximately 420 km west of the Kharga oasis in the southern Western Desert of Egypt.
Description.—The left humerus is virtually complete, but the ventral side of the proximal margin and the lateral process are broken off (Fig. 4). The specimen is slightly curved and has a length of 65 cm (measured from the ventral side). The maximum width of the shaft is 25 cm (measured at the widest point of the distal region). The minimum width of the shaft is 12.6 cm at the narrow waist. Cortical bone mostly covers the cancellous bones in the well-preserved parts of the ventral side of the humerus, while it is almost removed from the distal side of the specimen. The specimen has some erosional fractures in one-third of the distal margin, but this area is still connected to the shaft. Another fracture separates the proximal margin from the shaft with loss of some parts of the ventral side, and the surfaces connected to the shaft on the dorsal side are complete. The specimen is characterized by a massive humeral head, long curved shaft and large medial process, which is nearly perpendicular to the head of the humerus.
The dorsoventral medial process extends more medially than proximally. It has smooth surfaces and is dorso-ventrally flattened. The maximum length of the medial process is 14.3 cm (as measured from the proximal-most margins of the head), and its length is 12.2 cm (as measured from the external margin). The tip of the medial process is thick and slightly straight, while its base has a thinner concave surface with thinly flattened attachment to the shaft. The medial process extends from the head at an angle of approximately 96°, which means that it is almost perpendicular to the head of the humerus. The medial process does not expand proximally beyond the plane of the head of the humerus. The external most margin of the medial process is lost.
The lateral process is lost due to erosion, except for part of its basal margin situated on the ventral side of the shaft, distally and close to the humeral head.
The head of the humerus is a massive, sub-spherical surface with greater dorsal enclosure. The width of the surface is 12 cm, and its dorso-ventral length is approximately 4.4 cm. The head of the humerus is offset approximately 40° from the shaft. It is situated above the plane of the medial process. The angle between the axis of the head and the shaft is nearly 115° (α angle; Zangerl 1953), and the longitudinal axis of the shaft makes an angle of approximately 40° with the plane of movement (β angle; Zangerl 1953).
The shaft is slightly curved from the proximal to the distal end in the ventral plane. It is relatively slender at the midlength, forming a prominent slightly thinner waist. It expands distally into the epicondyles. The distal region is wider than the epicondyle region and is dorso-ventrally flattened. The inclined angle of the shaft from the proximal to the distal end is 34°. The shaft has a concave outline from the lateral and medial faces and distally has a convex surface. The ectepicondylar foramen is situated dorsally in the distal extremity border. The foramen is small, oval and slightly deep, and it extends dorso-ventrally but is not open on the ventral side.
The distal epiphysis is characterized by well-developed articulated surfaces (ulnar epicondyle, trochlea, capitellum and radial epicondyle) situated in the ventral surface of the shaft. The outline of the distal margin forms a trapezoidal shape. The distal end extends in apparent elevation to the ventral side. The trochlea is more extending and less dipping to the ventro-dorsal side, with a smooth articular surface. The ulnar epicondyle represents the wider point of the distal margin of the shaft. It extends more medially of the shaft. The coronoid fossa is a shallow wide groove, situated medially on the ventral side of the shaft, in the front of the trochlea. The capitellum is a massive oval body, slightly pointed to the medio-dorsal surface. It extends more distally behind the trochlea. The radial epicondyle is situated in the ventral surface of the shaft and extends more dorsally than the capitellum. The radial epicondyle represents the apex of the distal end of the shaft and is separated from the capitellum by a V-shaped dorso-ventrally extending groove. The outlines of the distal margin form a trapezoidal shape.
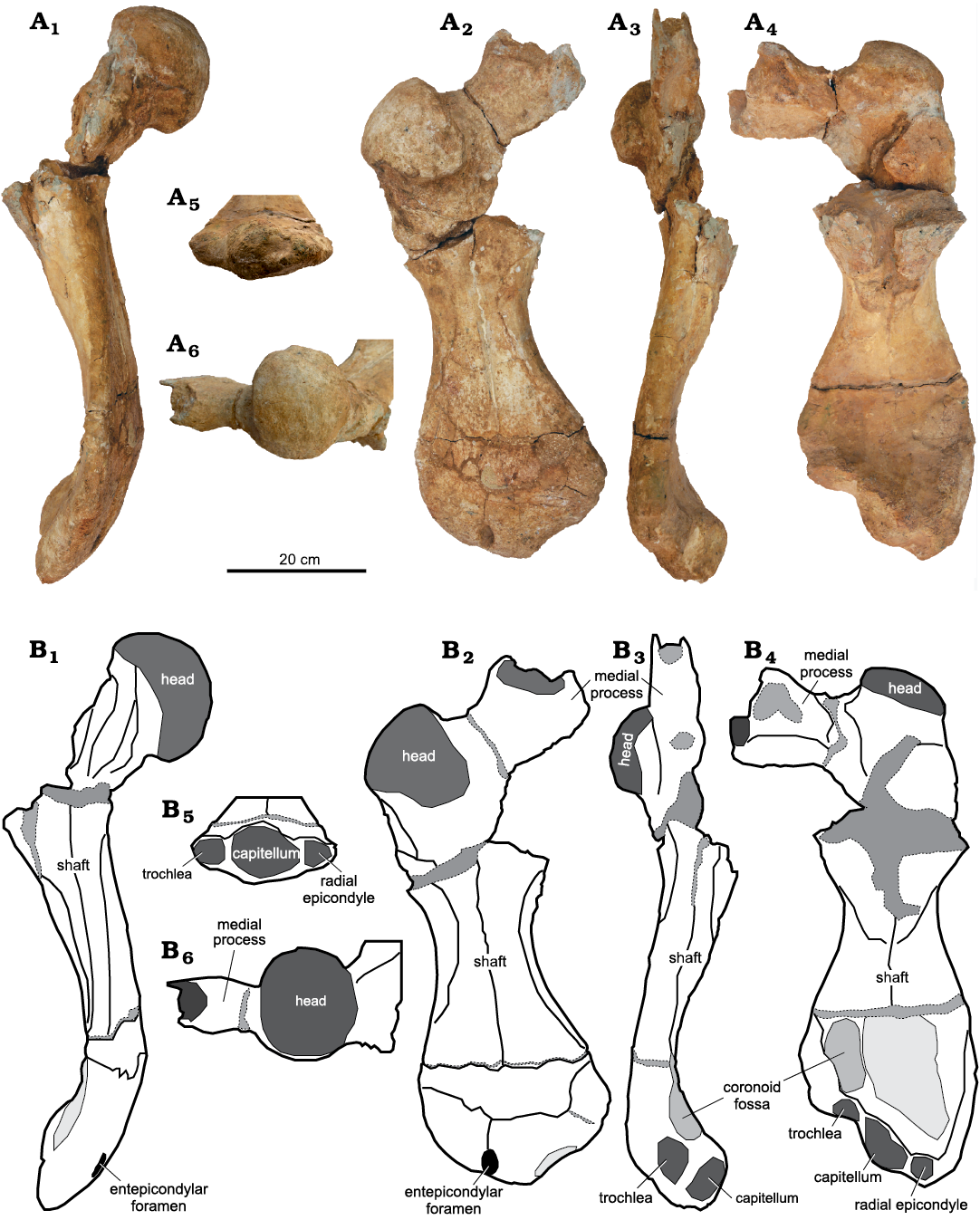
Fig. 4. Left humerus (NVP010) of panchelonioidean turtle, from Maastrichtian, Abu Minqar, southern Western Desert of Egypt. Photograps (A) and explanatory drawings (B), in anterior (A1, B1), dorsal (A2, B2), posterior (A3, B3), ventral (A4, B4), distal (A5, B5), and proximal (A6, B6) views.
Discussion
NVP010 has diagnostic features of Panchelonioidea such as a flattened humeral shaft, a lateral humeral process that is situated distal to the head of the humerus (Hirayama 1994), and a distal end that is laterally prolonged and possesses an ectepicondylar groove (Hirayama 1992).
Current phylogenies suggest the presence of up to three speciose clades within Panchelonioidea: Pancheloniidae, Dermochelyidae, and Protostegidae, the latter either being situated within Dermochelyidae, or at the stem of crown Chelonioidea (see Joyce et al. 2021 for recent summary).
NVP010 has prominent morphological features that are more similar to those of Pancheloniidae than Protostegidae and Dermochelyidae (Zangerl and Sloan 1960; Hirayama 1994). These features are summarized as follows: (i) the humerus of NVP010 is flattened and curved, whereas in Dermochelyidae it is cylindrical in cross section and slightly straight (e.g., Mesodermochelys undulatus, see Hirayama and Chitoku 1996; and Hirayama and Hikida 1998); (ii) the head of the humerus of NVP010 is not well separated from the adjoining process, comparing with the head of the humerus of Protostegidae which is well separated from the adjoining processes (Hirayama 1997); (iii) the remnant of the distal margin of the lateral process in NVP010 is distally situated close to the head of the humerus but in dermochelyids, it is further distally situated on the shaft of the humerus (Hirayama 1992; Hirayama and Hikida 1998; Kaddumi 2006); (iv) NVP010 has a prominent coronoid fossa and other well-articulated surfaces of the distal end, but dermochelyids lack the coronoid and the other articular surfaces are nearly massive (Mulder 2003; Evers et al. 2019); (v) the ectepicondylar foramen of NVP010 is represented as closed, shallow groove, but in Protostegidae it is an open groove in the distal end of the humerus (Cadena and Parham 2015); (vi) the head of NVP010 humerus is situated proximally above the medial process, which is different from Dermochelyidae and Protostegidae, where the medial process has a high level than the head of humerus (Hirayama 1994; Hirayama and Chitoku 1996); (vii) the medial process of NVP010 is laterally wider and dorso-ventrally thicker in the proximal end, but in Protostegidae it is almost straight (Kaddumi 2006; Evers et al. 2019); (viii) The medial process is separated from the head of the humerus by deep and wide notch (nearly perpendicular), but in protostegids it is separated from the head of the humerus by shallow notch (Evers et al. 2019).
NVP010 also has diagnostic features that differentiate it from large bodied, coeval chelonioids: massive humeral size; the humerus is more curved than in Gigantatypus salahi Kaddumi, 2006, Allopleuron hoffmani (Gray, 1831), Atlantochelys mortoni Agassiz, 1849; the medial process extends medially, but in Gigantatypus salahi, it extends proximally; the medial process lacks the concavity between the head and the medial process; it is not elevated above the plane of the proximal extension of the head of the humerus; the distal margin is strongly ventrally curved, containing slightly deep and elongated coronoid fossa proximal to the epicondyle; the radial capitellum is situated on the ventral side, but in the other chelonioids, it is situated on the dorsal side (Kaddumi 2006). Incidentally, the shaft of the humerus is similar to the shaft of toxochelyidae and the distal joints of the surface have radial elevation (Zangerl 1953). The medial process of toxochelyids lies at the level of the humerus head (Zangerl 1953), but in Pancheloniidae, it lies beyond the head of the humerus, as in the present sample.
Unfortunately, large bodied turtles that are known worldwide from coeval marine sediments that lack humeri, in particular the purported Moroccan dermochelyids Ocepechelon bouyai (Bardet, Jalil, Lapparent de Broin, Germain, and Lambert, 2013) and Alienochelys selloumi (Lapparent de Broin, Bardet, Amaghzaz, and Meslouh, 2014). To avoid adding to parataxonomy that plagues marine turtle systematics, we therefore here refrain from naming a new species. In addition, as the relationship of Mesozoic marine turtles is still under dispute, we carefully refer our specimen to Panchelonioidea indet., even if similarities with described pancheloniids are apparent.
Conclusions
The bone-bearing horizon containing the studied specimen is located in the Ammonite Hill Member in the lower Dakhla Formation, South Western Desert, Egypt. This horizon is composed of sandstone and siltstone and contains abundant and diverse of Maastrichtian-age fossils, such as ammonites (Libycoceras ismaelis and Baculites sp.), bivalves (Exogyra overwegi, Pycnpdonta vesicularis), gastropods, echinoids, corals, fossilized fruits of mangrove palm Nypa and vertebrate remains. These faunal and floral associations reflect deposition in well-oxygenated shallow waters in a nearshore or lagoonal environment during sea-level lowstand periods.
The recorded turtle bone in the Ammonite Hill Member could reflect terrestrial influence within a shallow marine environment. In other words, the turtle may have lived along the coasts in salt water or in lagoons or estuaries in brackish waters or may have moved from salty to brackish environments.
The massive and widened humerus of NVP010 reflects a huge body size and its ability to swim long distances.
The loss of the complete skeleton of NVP010 and the discovery of only separated elements, such as a humerus, may have been due to (i) the predation by other vertebrate animals, such as sharks or mosasaurs, which occur in the same horizon as the specimen, or (ii) the redeposition of the skeleton after death by currents along the coastal area.
The present specimen represents the first record of gigantic Mesozoic chelonioids in Africa and the second record of chelonioids in the Maastrichtian deposits in Egypt. It contributes to fill the missing gaps in the history of Egyptian turtles from the Cretaceous through the Cenozoic.
Acknowledgements
The authors deeply thank Salem Ahmed Hamed and his sons (Abu Minqar, Egypt), as well as other Abu Minqar residents, for their hospitality and help during the field camping. Additionally, the authors would like to thank Anne Schulp (Naturalis Biodiversity Center, Darwinweg, The Netherlands) for his help with the manuscript. A great thanks is extended to Cairo University and New Valley University for their support. Many thanks to Mohamed Kamal Mosa (New Valley Vertebrate Paleontology Center, Kharga, New Valley, Egypt) who helped us with the camera set up. This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors. The quality of the manuscript was significantly enhanced thanks to comments by the editor Daniel Barta and the two reviewers Walter Joyce (Department of Geosciences, University of Fribourg, Fribourg, Switzerland) and Ren Hirayama (School of International Liberal Studies, Waseda University, Tokyo, Japan).
References
Agassiz, L. 1849. Remarks on crocodiles of the green sand of New Jersey and on Atlantochelys. Proceedings of the Academy of Natural Sciences of Philadelphia 4: 169.
Awad, G.H. and Ghobrial, M.G. 1966. Zonal stratigraphy of the Kharga Oasis. Ministry of Industry, General Egyptian Organisation for Geological Research and Mining, Geological Survey, Paper 34: 1–77.
Bardet, N., Jalil, N.E., Lapparent de Broin, F., Germain, D., and Lambert, O. 2013. A giant chelonioid turtle from the Late Cretaceous of Morocco with a suction feeding apparatus unique among tetrapods. PLoS ONE 8 (7): e63586. Crossref
Barthel, K.W. and Herrmann-Degen, W. 1981. Late Cretaceous and early tertiary stratigraphy in the Great Sand Sea and its SE margins (Farafra and Dakhla Oases, SW Desert, Egypt). Mitteilungen Bayerische Staatssammlung Paläontologie Historische Geologie 21: 141–182.
Cadena, E.A. and Parham, J.F. 2015. Oldest known marine turtle? A new protostegid from the Lower Cretaceous of Colombia. PaleoBios 32: 1–42. Crossref
El Hedeny, M., Kassab, W., Rashwan, M., Abu El-Kheir, G., and Abdel Gawad, M. 2020. Bivalve borings in Maastrichtian fossil Nypa fruits: Dakhla Formation, Bir Abu Minqar, South Western Desert, Egypt. Ichnos 28 (1): 24–33. Crossref
El-Soughier, M.I., Mehrotra, R., Zhou, Z.Y., and Shi, G. 2011. Nypa fruits and seeds from the Maastrichtian–Danian sediments of Bir Abu Minqar, South Western Desert, Egypt. Palaeoworld 20: 75–83. Crossref
Evers, W.S., Barett, M.P., and Benson, B.J. 2019. Anatomy of Rhinochelys pulchriceps (Protostegidae) and marine adaptation during the early evolution of cheloniids. PeerJ 7 [published online, http://dx.doi.org/10.7717/peerj.6811] Crossref
Gray, J.E. 1831. Synopsis Reptilium; or Short Descriptions of the Species of Reptiles. Part 1. Catapharcata, Tortoises, Crocodiles and Enaliosaurians. 84 pp. Treuttel, Wurtz & Co., London. Crossref
Hirayama, R. 1992. Humeral morphology of Chelonioid sea turtles; its functional analysis and phylogenetic implications. Bulletin of the Hobetsu Museum 8: 17–57.
Hirayama, R. 1994. Pylogentic systematics of Chelonioids sea turtles. The Island Arc 3: 270–284. Crossref
Hirayama, R. 1997. Distribution and diversity of Cretaceous chelonioids. In: J.M. Callaway and E.L. Nicholls (eds.), Ancient Marine Reptiles, 225–241. Academic Press, San Diego. Crossref
Hirayama, R. and Chitoku, T. 1996. Family Dermochelyidae (super family Chelonioidea) from the upper Cretaceous of North Japan. Transactions and Proceedings of the Paleontological Society of Japan. New series 184: 597–622.
Hirayama, R. and Hikida, Y. 1998. Mesdermochelys (Testudines, Chelonioidea; Dermochelyidae) from the Late Cretaceous of North Nakagawa-Cho, Hokkaido, North Japan. Bulletin of the Nakagawa Museum of Natural History 1: 69–76.
Joyce, W.G., Anquetin, J., Cadena, E.-A., Claude, J., Danilov, I.G., Evers, S.W., Ferreira, G.S., Gentry, A.D., Georgalis, G.L., Lyson, T.R., Pérez-García, A., Rabi, M., Sterli, J., Vitek, N., and Parham, J.F. 2021. A nomenclature for fossil and living turtles using phylogenetically defined clade names. Swiss Journal of Paleontology 140: article 5. [published online, https://doi.org/10.1186/s13358-020-00211-x] Crossref
Joyce, W.G., Parham, J.F., and Gauthier, J.A. 2004. Developing a protocol for the conversion of rank-based taxon names to phylogenetically defined clade names, as exemplified by turtles. Journal of Paleontology 78: 989–1013. Crossref
Kaddumi, H.F. 2006. A new genus and species of gigantic marine turtles (Cheonioidea: Cheloniidae) from Maastrichtian of the Harrana Fauna-Jordan. Vertebrate Paleontology 3 (1): 1–14.
Lapparent de Broin, F. and Werner, C. 1998. New late Cretaceous turtle from the Western Desert, Egypt. Annales de Paléontologie 84: 131–214. Crossref
Lapparent de Broin, F., Bardet, N., Amaghzaz, M., and Meslouh, S. 2014. A strange new chelonioid turtle from the Latest Cretaceous Phosphates of Morocco. Comptes Rendus Palevol 13: 87–95. Crossref
Mulder, E.W.A. 2003. Comparative osteology, palaeoecology and systematics of the Late Cretaceous turtle Allopleuron hofmanni (Gray, 1831) from the Maastrichtian type area. In: E.W.A. Mulder (ed.), On the Latest Cretaceous Tetrapods from the Maastrichtian Type Area. Publicaties van het Natuurhistorisch Genootschap in Limburg 44: 23–92.
Said, R. 1961. Tectonic framework of Egypt and its influence on distribution of Foraminifera. American Association of Petroleum Geologists Bulletin 45: 198–218. Crossref
Said, R. 1961. Tectonic framework of Egypt and its influence on distribution of foraminifera. AAPG Bulletin 45 (2): 198–218. Crossref
Staesche, K. 1929. Schildkroten aus der oberen Kreide Patagnoiens. Paleontographica 72: 103–123.
Tantawy, A.A., Keller, G., Addatte, T., Stinnesbeck,W., Kassab, A., and Schulte, P. 2001. Maastrichtian to Paleocene depositional environment of the Dakhla Formation, Western Desert, Egypt: sedimentology, mineralogy, and integrated micro-and macrofossil biostratigraphies. Cretaceous Research 22: 795–827. Crossref
Zangerl, R. 1953. The vertebrate fauna of the Selma Formation of Alabama, part IV the turtle of family Toxochelyidae. Fieldiana: Geology Memoirs 3: 153–277. Crossref
Zangerl, R. and Sloan, R.E. 1960. A new specimen of Desmatochelys lowii Williston. A primitive cheloniid sea turtle from the Cretaceous of South Dakota. Fieldiana, Geology 14 (2): 7–70. Crossref
Zittel, K.A. 1883. Beitrtige , wr Geologie und Palaontologie der Libyschen Wiiste und der angrewenden Gebiete Agyptens. Palaeontographica 30 (1): l–147.
Acta Palaeontol. Pol. 66 (2): 349–355, 2021
https://doi.org/10.4202/app.00849.2020