
A low diversity Sinuites gastropod community from the Floian, Early Ordovician, of South Wales
JAN OVE R. EBBESTAD and JOHN C.W. COPE
Ebbestad, J.O.R. and Cope, J.C.W. 2021. A low diversity Sinuites gastropod community from the Floian, Early Ordovician, of South Wales. Acta Palaeontologica Polonica 66 (2): 319–335.
A low diversity Sinuites-dominated gastropod community is described from the Floian, Arenig Series, of the Llangynog Inlier, southwest of Carmarthen, South Wales. The abundant material comes from shallow-water siltstone and mudstone beds of the Bolahaul Member of the Ogof Hên Formation. The locality has an exceptionally diverse mollusc-dominated fauna (63.5% of the fauna), with gastropods constituting 6% and tergomyans 1% and echinoderms, arthropods and other fauna making up the rest. Except for one rare tergomyan mollusc, identified as Carcassonnella cf. vizcainoi, other tergomyans are described elsewhere. Nearly half of all gastropod specimens are represented by Sinuites ramseyensis. Three of the five taxa described are new: Mimospira llangynogensis sp. nov., Catalanispira prima sp. nov., and Ceratopea? moridunensis sp. nov. The assemblage compares best with those of contemporaneous high-latitude peri-Gondwana areas. Early Ordovician species of Carcassonnella are typically found in France, Iberia, Czech Republic, and Morocco, while species of Mimospira are found in Germany and Czech Republic, but also in Baltica where the main radiation took place later. Two of the oldest occurrences of Mimospira are from Avalonian Wales (Carmarthenshire and Anglesey). Catalanispira occurs later in Baltica and Laurentia in the late Middle and early Late Ordovician, when taxa from these areas start to appear in Wales and vice versa. The presence of Ceratopea?, a genus typical of Laurentia, is at odds with the biogeographic distribution of faunas at this time. Part of the observed distribution pattern may be explained by different latitudinal position and facies depths of Avalonia compared to Armorica, Bohemia, and Morocco. The species described herein are amongst the first Floian taxa of these groups formally described from this area and add significantly to the global Floian record.
Key words: Gastropoda, Onychochilidae, Tergomya, biogeography, Floian, Arenig, Ordovician, Avalonia, UK.
Jan Ove R. Ebbestad [jan-ove.ebbestad@em.uu.se], Museum of Evolution, Uppsala University, Norbyvägen 16, SE 752 36 Uppsala, Sweden.
John C.W. Cope [john.Cope@museumwales.ac.uk], Department of Natural Sciences, National Museum of Wales, Cathays Park, Cardiff CF10 3NP, UK.
Received 2 November 2020, accepted 10 December 2020, available online 9 June 2021.
Copyright © 2021 J.O.R. Ebbestad and J.C.W. Cope. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
The Floian record of gastropods and tergomyan molluscs is meagre and among the poorest during the Ordovician (Ebbestad et al. 2013). Records from North America and North China constitute the majority of known occurrences, and endemism was high throughout the Early Ordovician. As a result, a few key genera control the observed pattern of distribution and the biogeographical inferences that can be deduced from this. In general terms, gastropod diversity increases steadily throughout the Ordovician until it drops at the end of the period, but with different patterns of dispersal, diversity and ecological distribution among gastropod groups, palaeocontinents and facies (Novack-Gottshall and Miller 2003; Frýda and Rohr 2004; Frýda et al. 2008; Ebbestad et al. 2013), but also among members of the planktic and benthic realms (Nützel et al. 2006; Servais et al. 2010). Gastropod and tergomyan diversity and dispersal trends follow suite of other major benthic organisms during the complex and dynamic biodiversity increase seen throughout the Ordovician, constituting the so-called Great Ordovician Biodiversity Event (Servais and Harper 2018).
During the Early Ordovician, several communities and faunal provinces can be recognized, most notably among brachiopods and trilobites. Both groups show high endemism during this time (Harper et al. 2013; Adrain 2013) which in many cases enables the recognition of areas that were biogeographically close (Fortey and Cocks 2003). Among the gastropods and tergomyans the large Laurentian unit is prominent (North America, Greenland, Argentina, and North China) based on genera such as Ceratopea Ulrich, 1911, Lecanospira Butts, 1926, Lophonema Purdue and Miser, 1916, Macluritella Kirk, 1927, Malayaspira Kobayashi, 1958, Ophileta Vanuxem, 1842, Plethospira Ulrich and Scofield, 1897, Sinuopea Ulrich, 1911, Teiichispira Yochelson and Jones, 1968, and others. In the European peri-Gondwana and North Africa palaeo-areas, Ibero-Armorica, Bohemia, and Morocco are joined by high latitude area genera like Carcassonnella Horný and Peel, 1996, Lesueurilla Koken, 1898, Peelerophon Yochelson in Babin et al., 1982, Pelecyogyra Ebbestad and Lefebvre, 2015, and Thoralispira Horný and Vizcaïno, 1995 (Frýda and Rohr 1999; Ebbestad et al. 2013).
The present study reports on four gastropod taxa and one planispiral tergomyan mollusc from the Floian, Arenig Series, of the Llangynog Inlier southwest of Carmarthen in South Wales. These are amongst the first Floian taxa of these groups formally described from this area and add significantly to the global Floian record. Hicks (1873) described two species from coeval beds at Pembrokeshire to the west of the Llangynog Inlier, which are discussed in relation to the new material. Around 120 specimens were available for this study, and although variably deformed, the large sample set allows a good appreciation of their morphology. The composition of the fauna shows elements of both peri-Gondwana and Laurentian forms, and includes planispiral forms like Carcassonnella and the widespread Sinuites Koken, 1896, the rare sinistrally coiled conispiral forms Mimospira Koken in Koken and Perner, 1925, and Catalanispira Ebbestad, Kolata, and Isakar 2020a, as well as a possible Ceratopea.
Institutional abbreviations.—BGS, British Geological Survey, Nottingham, UK; NMW, National Museum of Wales, Cardiff, Wales, UK.
Nomenclatural acts.—This published work and the nomenclatural acts it contains have been registered in ZooBank: urn:lsid:zoobank.org:pub:01FF623C-11BB-4A03-8DE7-354990FBF7ED
Geological setting
The Llangynog Inlier (Cope 1982) is an area of Neoproterozoic, Cambrian and Lower Ordovician rocks that crop out to the southwest of Carmarthen, South Wales (Fig. 1). To the south the inlier is limited by the cover of Green Castle Sandstone Formation of the basal Old Red Sandstone (Cope 2018) of Late Silurian age, and is bounded to the north and west by Middle Ordovician graptolitic shale. The River Tywi (Towy) limits the inlier to the east. The rocks of the inlier were discovered by JCWC during mapping of the area begun in the late 1970s where the current Geological Survey 1967: 1:63 360 Sheet 229 (Geological Map Series, New Series, Carmarthen, https://www.bgs.ac.uk/information-hub/bgs-maps-portal/) of the area shows only Floian, Arenig Series, Lower Ordovician rocks beneath the Old Red Sandstone.
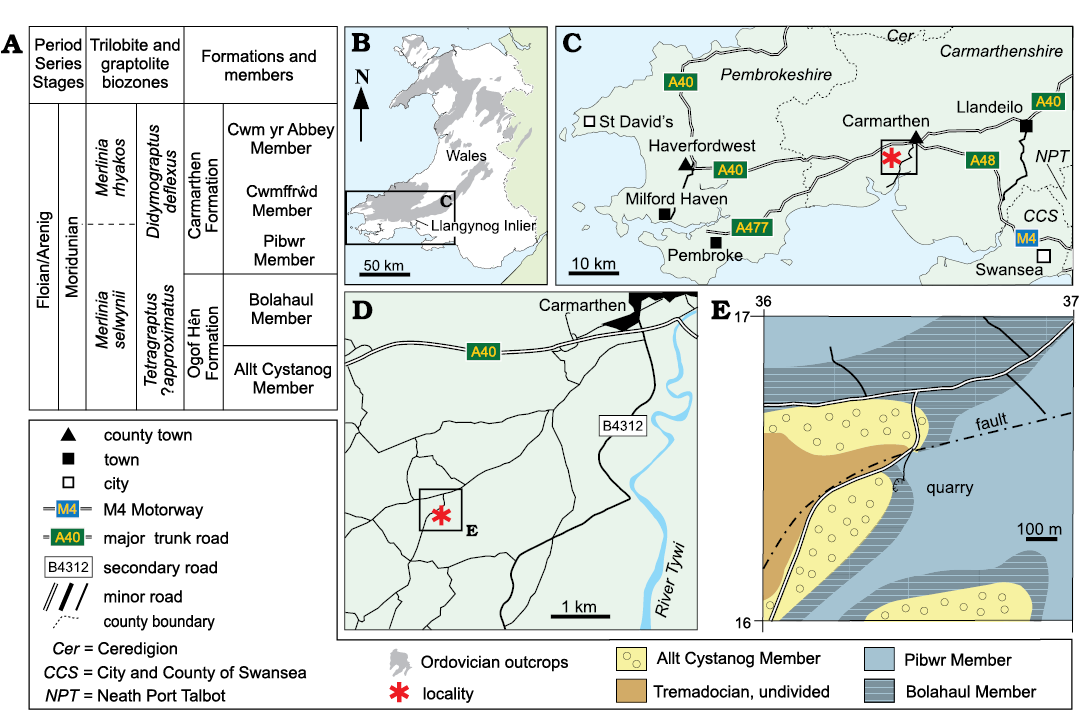
Fig. 1. A. Moridunian stratigraphy (after Fortey and Owens 1978). B–D. Simplified maps of Wales (B), the southeastern part of the country (C), and the area south of Carmarthen (D) showing the location of the study area and outcrop. E. Simplified geological map with position of the Dan-lan-y-Castell quarry (modified from Cope 1982).
The Floian rocks to the SE of Carmarthen and E of the River Tywi were investigated by Fortey and Owens (1978). They found that the succession there had been incorrectly interpreted by the Geological Survey and divided the Arenig Series there into a lower Ogof Hên Formation and an upper Carmarthen Formation; two members were recognised in the Ogof Hên Formation, a basal Allt Cystanog Member and an upper Bolahaul Member (Fig. 1). These formations and their constituent members were recognised to the W of the River Tywi in the Llangynog Inlier by Cope (1979). The fossils described herein are from a small quarry (grid reference SN 3640 1639) in the Bolahaul Member situated about 6 km southwest of Carmarthen, and known as the Dan-lan-y-Castell quarry. It lies 500 m southeast of Dan-lan-y-Castell and near to a house named Llyn-y-Llawr (locality 7 in Cocks and Popov 2019; Fig. 1). The age of the fauna is tightly constrained by the Merlinia selwynii Trilobite Zone of Fortey and Owens (1978) and of their Moridunian Stage (lower Floian, Fl1).
Although the Geological Survey Memoir (Strahan et al. 1909: 16) described this quarry they reported a very meagre faunal list that included a single unnamed bivalve. However, the quarry has yielded an exceptionally diverse fauna which is mollusc-dominated and is the earliest example of the “modern fauna” of Sepkoski (1981). Recognising that the fauna contained unusual elements for a rock of this age, bulk collection and splitting of the rock in the laboratory was carried out over a number of years. Altogether some 3 100 fossils were collected and it was found that molluscs comprised some 71.5% of the fauna (of which bivalves are 57%, nautiloids 36%, gastropods 6%, tergomyans 1%, and a single rostroconch specimen). In addition the quarry has also yielded the earliest examples of several other fossil groups including parablastoids (Paul and Cope 1982) and their later recognised holdfasts (Cope and Donovan 2005), the earliest bryozoan then-known (Taylor and Cope 1987), the earliest gorgoniids (Cope 2005) and the earliest calcified red alga (Riding et al. 1998).
Of the molluscs the bivalve fauna of 20 species belonging to 18 genera is the most diverse fauna of that age known and includes the earliest examples of several bivalve groups; that fauna and the rostroconch were described by Cope (1996) and the nautiloids by Evans (2005). Herein the gastropod fauna and a single species of tergomyan are described, the remainder of the small but varied tergomyan fauna will be described elsewhere.
Although there are several good exposures of the Bolahaul Member both E and W of the River Tywi, this one locality is unique in its rich fauna and has been designated as a Site of Special Scientific Interest (SSSI) under the auspices of the Countryside Council for Wales (CCW). The reason for the extraordinary richness of the fauna may be due to the fact that it is clearly a shallow-water close-inshore siltstone and mudstone deposit, with occasional pebbly horizons; bioturbation is frequent. The majority of the fossils are preserved as limonite coated moulds, frequently undistorted and unaffected by cleavage. The gastropods, unlike the rest of the fauna, are often distorted.
Systematic palaeontology
Phylum Mollusca Linnaeus, 1758
Class Tergomya Horný, 1965
Family Carcassonnellidae Horný, 1997b
Genus Carcassonnella Horný and Peel, 1996
Type species: Gamadiscus courtessolei Yochelson in Babin et al., 1982; by original designation of Horný and Peel (1996: 307), the Tremadocian (Tr2) to Floian (Fl3), St. Chinian–lower Landeyran formations (former horizons e–i, j?, l) in the Montagne Noire, France.
Remarks.—Shells of the planispiral Carcassonnella are readily identifiable by their distinct ornamentation of transverse lirae, shallow median sinus followed by a slit leading into a raised median dorsal crest. Hitherto Carcassonnella was known from seven species, four that are named and three that are left in open nomenclature. The type species Carcassonnella courtessolei (Yochelson in Babin et al., 1982) is extremely abundant in the Lower Ordovician (Tr3–Fl3) of Montagne Noire, France, and Fezouata, Morocco, with the more uncommon C. vizcainoi Horný and Peel, 1996, occurring in both areas in the lower Floian (Fl1) (Horný and Peel 1996; Horný1997b; Ebbestad 2016; Ebbestad et al. 2020b). A minute unnamed species also found in Morocco in the same interval as C. courtessolei (Ebbestad 2016). The Middle Ordovician (Dw2, Dw3) C. pragensis and C. sp. were described by Horný (1997a) from the Prague Basin, Czech Republic, while Carcassonnella cf. courtessolei was identified from Lower Ordovician (Tremadocian or Floian) pebbles in a Carboniferous olistostrome in Córdoba of Spain (Gutiérrez-Marco et al. 2014). The youngest species is C. multilineata (Reed, 1920) from the Upper Ordovician (Katian; Ka2–Ka4) of Girvan, Scotland, which hitherto is the only record outside peri-Gondwana (Ebbestad 2008).
Carcassonnella cf. vizcainoi Horný and Peel, 1996
Fig. 2.
Material.—Two internal moulds, NMW 2017.15G.1, NMW 2017.15G.2, with the external mould of the former preserving impression of the ornamentation. From Dan-lan-y-Castell quarry, Carmarthen, Wales, UK; Floian (Early Ordovician).
Description.—Shell planispiral, ~6.5 mm across and ~3.5 mm in width with at least 3 whorls. Whorl profile rhomboidal with periphery somewhat below mid-height of whorl. Upper and lower whorl surface straight to slightly concave. Whorl overlap just above periphery. Median dorsal carina only slightly raised above shell and bordered by thick rounded lirae. Total width of carina is about 7.5 mm (~20% of whorl width). Ornamentation consists of regularly spaced rounded ribs that are nearly straight across the upper whorl surface. Their transition to the median carina is obscured.
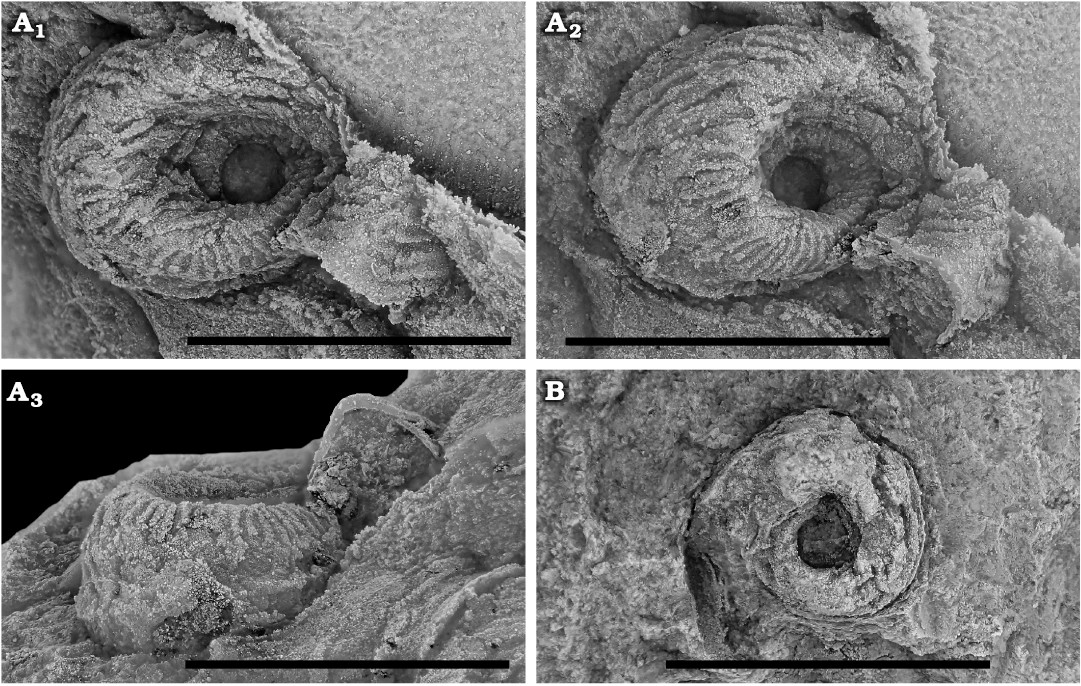
Fig. 2. Carcassonellid tergomyan Carcassonnella cf. vizcainoi Horný and Peel, 1996 from the Floian, Lower Ordovician Bolahaul Member of the Ogof Hên Formation, Dan-lan-y-Castell quarry, Wales, UK. A. NMW 2017.15G.1, silicon cast, in lateral (A1), posterior oblique (A2), and ventral (A3) views. B. NMW 2017.15G.2, internal mould in lateral view. Scale bars 5 mm.
Remarks.—The Llangynog species is recognized from two small internal moulds. A silicon cast from the external mould of specimen NMW 2017.15G.1 shows details of the ornamentation and the shape of the whorl, which allows comparison with Carcassonnella vizcainoi Horný and Peel, 1996, from Montagne Noire and Fezouata. A characteristic feature of this species is the rhomboidal whorl profile, which is also clearly seen in the Llangynog material (Fig. 2A3). The type species Carcassonnella courtessolei has a rounded whorl profile, while species like C. vizcainoi, C. multilineata (Reed, 1920), and Carcassonnella cf. C. courtessolei sensu Gutiérrez-Marco et al. (2014) have a rhomboidal whorl profile with the periphery low on the whorl (Horný and Peel 1996; Ebbestad 2008). The shape and spacing of the ornamentation in the best Llangynog specimens resembles closely that of some specimens of C. vizcainoi (see Horný and Peel 1996: pl. 6: 9–12) although this feature is quite variable in the latter (Horný and Peel 1996). The main differences between the two is the very low median carina with its thicker bordering ribs, the periphery being a bit lower and the convexity of the whorl surfaces in the Llangynog material. Horný and Peel (1996) pointed out that mode of preservation could complicate differentiation of especially juvenile specimens of this species relative to C. courtessolei. In terms of size the Llangynog specimens are smaller than the largest specimens of C. vizcainoi from the Montagne Noire, which attain a size of 13 mm (Horný and Peel 1996).
Carcassonnella vizcainoi occurs in the lowermost Floian in France and Morocco, encompassing the lowermost Taihungshania migueli Trilobite Zone in Montagne Noire and the ?Cymatograptus protobalticus Graptolite Zone in Fezouata (Horný and Peel 1996; Vizcaïno et al. 2001; Ebbestad 2016; Ebbestad et al. 2020b). The stratigraphical position of the Llangynog occurrence would be similar, within the Merlinia selwynii Trilobite Zone and the Tetragraptus approximatus Graptolite Zone.
Hicks (1873) also described Bellerophon solvensis Hicks, 1873, from a single small specimen found in contemporaneous strata at Tremanhire near Solva in Pembrokeshire (BGS A 45285). The internal mould is not readily comparable with any of the taxa described herein, but seem to constitute a planispiral form. It may be a poorly preserved species of Carcassonnella, although the whorl profile seems to be rounded and not rhomboidal.
Class Gastropoda Cuvier, 1797
Order Bellerophontida McCoy, 1851
Superfamily Bellerophontoidea McCoy, 1851
Family Sinuitidae Dall, 1913
Genus Sinuites Koken, 1896
Type species: Bellerophon bilobatus Sowerby, 1839: 643, pl. 19: 13, by subsequent designation of Bassler (1915: 1159); from the Caradoc (Sandbian, Sa2, Upper Ordovician), Horderley Sandstone Formation at Horderley, Shropshire, England, UK.
Remarks.—Sinuites is an often cited taxon from Ordovician deposits world-wide, although it likely embraces a number of sinuitid-like taxa (see discussion in Ebbestad and Yochelson 2000). Horný (1996) suggested that Sinuites could have been an active semi-infaunal predator living on a soft substrate. The genus is reported from a number of facies but it may be more common in deep subtidal siliciclastic environments (Hurst 1979; Wahlman 1992; Horný and Vonka 2002; Stewart 2012; Ebbestad et al. 2013).
Sinuites ramseyensis (Hicks, 1873)
Fig. 3.
1873 Bellerophon ramseyensis sp. nov.; Hicks 1873: 50, pl. 3: 30–32.
Material.—Fifty-five specimens, NMW 2017.15G.3–57, preserved as variably compressed internal moulds. From the Dan-lan-y-Castell quarry, Carmarthen, Wales, UK. Merlinia selwynii Trilobite Zone, Moridunian regional Stage (lower Floian Stage, Fl1).
Description.—Specimen length is 10–20 mm. The whorl profile is heart-shaped with height and width being about equal (Fig. 3D). The wide and U-shaped median sinus stretches about 1/3 of a whorl back from the aperture. The typical lateral anterior lobes are sub-angular and at the aperture the lateral lip seem to connect with a lateral-umbilical ridge. A small patch in specimen NMW 2017.15G.7 preserves ornamentation near the umbilical area about 2/3 of a whorl back from the preserved aperture. Fine, slightly wrinkled lines are expressed on the external imprint and may represent subinductural deposits (Fig. 3E) (Horný 1996).
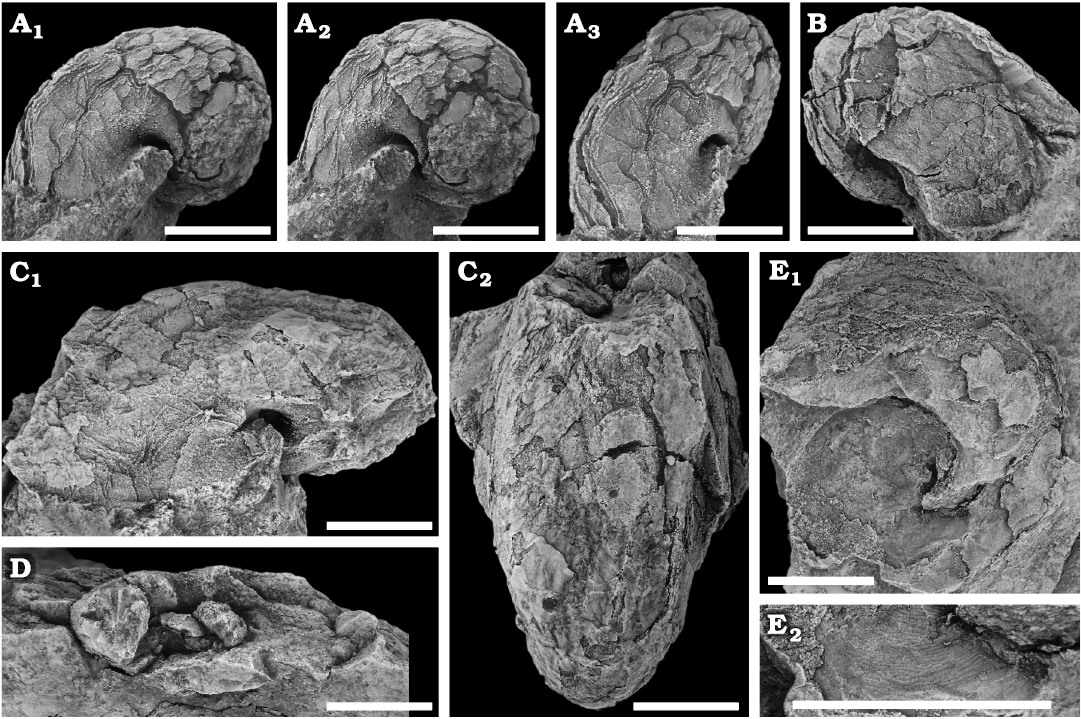
Fig. 3. Sinuitid bellerophontid Sinuites ramseyensis (Hicks, 1873) from the Floian, Lower Ordovician Bolahaul Member of the Ogof Hên Formation, Dan-lan-y-Castell quarry, Wales, UK. A. NMW 2017.15G.3, deformed shell in left lateral (A1), posterior oblique (A2), and anterior oblique (A3) views. Note the twisted and compacted median sinus seen as a line in A1 and A3. B. NMW 2017.15G.4, anterolateral shield in right lateral view. C. NMW 2017.15G.5, large, slightly dorso-ventrally flattened specimen in left lateral (C1) and dorsal (C2) views, showing median anterior sinus well. D. NMW 2017.15G.6, natural cross section in ventral view, showing the shape of the whorls. E. NMW 2017.15G.7, specimen in left lateral view (E1), showing the external mould of the initial whorls and the internal mould of the last whorl; details of the ornamentation as seen on the external mould (E2). Scale bars 5 mm.
Remarks.—Hicks (1873) described Bellerophon ramseyensis from the Ogof Hên Formation at Ramsey Island and St David’s in Pembrokeshire, south-west Wales, which is identified as a species of Sinuites (images available on the British Geological Survey homepage: http://www.3d-fossils.ac.uk/). The units in Pembrokeshire are the lateral equivalent to those of the Llangynog Inlier discussed herein (Fortey and Rushton 2000). Preservation of the Sinuites specimens in the two areas appears quite similar, and as a matter of convenience they may be regarded as constituting the same species.
With around 50 specimens Sinuites is the most abundant taxon in the present material. Deformation does not allow a full appreciation of the morphology, but there is no reason to assume that more than one species is present.
Stratigraphic and geographic range.—Ordovician; world wide.
Order Mimospirida Dzik, 1983
Family Clisospiridae Miller, 1889
Subfamily Clisospirinae Miller, 1889
Remarks.—Sinistrally coiled members of the Mimospirida Dzik, 1983, are a small but conspicuous part of Ordovician mollusc communities (total range of the group is Cambrian–Devonian (Frýda 2012)). Classification of these forms has been widely discussed, but currently they are considered as basal gastropods of uncertain systematic position (Fryda and Bouchet in Bouchet et al. 2005; Frýda 2012; Bouchet et al. 2017; Ebbestad et al. 2020a). The order contains two families within the superfamily Clisospiroidea Miller, 1889, the Clisospiridae Miller, 1889, and the Onychochilidae Koken in Koken and Perner, 1925. Members of the former are usually moderately high spired and have a wide open base (pseudoumbilicus) embraced by a frill or rib, while the members of the latter have broad and low shells with a funnel-like umbilicus (Wängberg-Eriksson 1979; Dzik 1983).
In the Llangynog fauna the Clisospiridae are represented by a species of Mimospira Koken in Koken and Perner, 1925, a genus found mainly in Ordovician strata and only two records in the Silurian (Peel 1975; Wängberg-Eriksson 1979). The Onychochilidae is represented by Catalanispira Ebbestad, Kolata, and Isakar, 2020a, as genus hitherto only known from one species in the Middle Ordovician (Dw3) of Estonia and one in the Upper Ordovician (Sa2) of the USA (Ebbestad et al. 2020a). Both new records of Mimospira and Catalanispira in Wales are amongst the oldest of their genera.
Genus Mimospira Koken in Koken and Perner, 1925
Type species: Onychochilus helmhackeri Perner, 1900: 4, 14, by subsequent designation of Knight (1937: 710), from the Arenigan (Darriwilian, Dw1, Middle Ordovician) upper pyroclastic member of the Klabava Formation near Kváň, Bohemia, Czech Republic.
Remarks.—Most species of Mimospira are small (height 5 mm or less) and many are left in open nomenclature. Ordovician Mimospira are known from at least nine species through the Ordovician of Baltoscandia and Poland (Yochelson 1962, 1963; Wängberg-Eriksson 1979; Dzik 1983; Isakar and Peel 1997; Frýda and Rohr 1999; Frisk and Ebbestad 2007; Lindskog et al. 2015), one Tremadocian (Lower Ordovician) species in the Franconian Forest (Frankenwald) of Germany (Sdzuy et al. 2001), one species from the Katian (Upper Ordovician) of the Carnic Alps in Austria (Gubanov and Bogolepova 1999), six species from the Tremadocian–Katian of the Prague Basin, Czech Republic (Frýda 1989; Frýda and Rohr 1999; Horný 1999; Kraft et al. 2013, 2015), one species from the Sandbian (Upper Ordovician) of Scotland (Longstaff 1924; Frýda and Rohr 1999; Stewart 2012), two species from the Floian (Lower Ordovician) and Dapingian (Middle Ordovician) of Wales (Bates 1963; this paper) and one Darriwilian (Middle Ordovician) species in the Precordillera of Argentina (Dzik 2020), giving a total of at least 20 species.
Rohr (1996) reported M. aff. M. cochleata Lindström, 1884, from the lower part of the Whiterockian (Dapingian, Dp1; Middle Ordovician) in the lower Antelope Valley at Meiklejohn Peak in Nevada, USA, which was compared to the Silurian species from Gotland, Sweden (Lindström 1884; Wängberg-Eriksson 1979). The two are broadly similar in having a distinct rounded angular periphery low on the whorl, which is lacking in Ordovician species. They differ in the densely spaced oblique lirae in the Silurian form that cross the periphery without a change in direction, and the much sharper angulation of the periphery in the Nevada form. The latter shows widely spaced lirae that are prosocline on the upper whorl surface sweeping strongly backwards to the sharp periphery. Just below the periphery the lirae seem to curve more strongly abaperturally before sweeping evenly backwards, although the angulation may enhance this impression. Rohr (1996) compared the whorl profile of the Nevada form with that of the Late Ordovician species of Angulospira Wängberg-Eriksson, 1979, from the Katian Boda Limestone of Sweden. The idea is followed up here, and is suggested that species of Angulospira as well as Bodaspira Wängberg-Eriksson, 1979, compare better with the Nevada form than species of Mimospira. Similarities between the Nevada form and Bodaspira undulata Wängberg-Eriksson, 1979, in the peripheral angulation, distinction of the lirae and direction of these across the periphery is compelling and the Nevada species is therefore here transferred to Bodaspira sp.
Mimospira llangynogensis sp. nov.
Fig. 4.
Zoobank LSID: urn:lsid:zoobank.org:act:E9919E4F-34D0-4B67-870E- C466C435A7B6
Etymology: After the Llangynog Inlier, where the type specimen is found.
Type material: Holotype (NMW 2017.15G.58, Fig. 4A), laterally flattened internal mould and its external mould. Paratypes (NMW 2017.15G.59–66, Fig. 4C), internal and external moulds; from the type locality. Two of the paratype specimens occur on NMW 2017.15G.60i, ii.
Type locality: Dan-lan-y-Castell quarry, Carmarthen, Wales, UK.
Type horizon: Merlinia selwynii Trilobite Zone, Moridunian regional Stage (lower Floian Stage, Fl1).
Material.—Type material only.
Diagnosis.—A large species of Mimospira with a small number of sharp ribs inclined at about 20° relative to the suture.
Description.—Shell is conispiral with 5–6 whorls, width slightly less than height, which can reach at least 12 mm. Pleural angle is about 55°. Sutures sharply incised but not deep (external view). Whorl surface steeply inclined and evenly convex between sutures. Aperture gently prosocline, simple and thin near suture, but likely getting thicker adumbilically. Ornamentation consist of regularly spaced ribs with rounded edges, separated by weakly concave interareas. The ribs are regularly spaced, with slightly increased spacing in later ontogeny. Each whorl show 5–6 ribs in lateral view, inclined at about 20° relative to the suture. Base of whorls rounded, convex, sloping gently into shallow axial area.
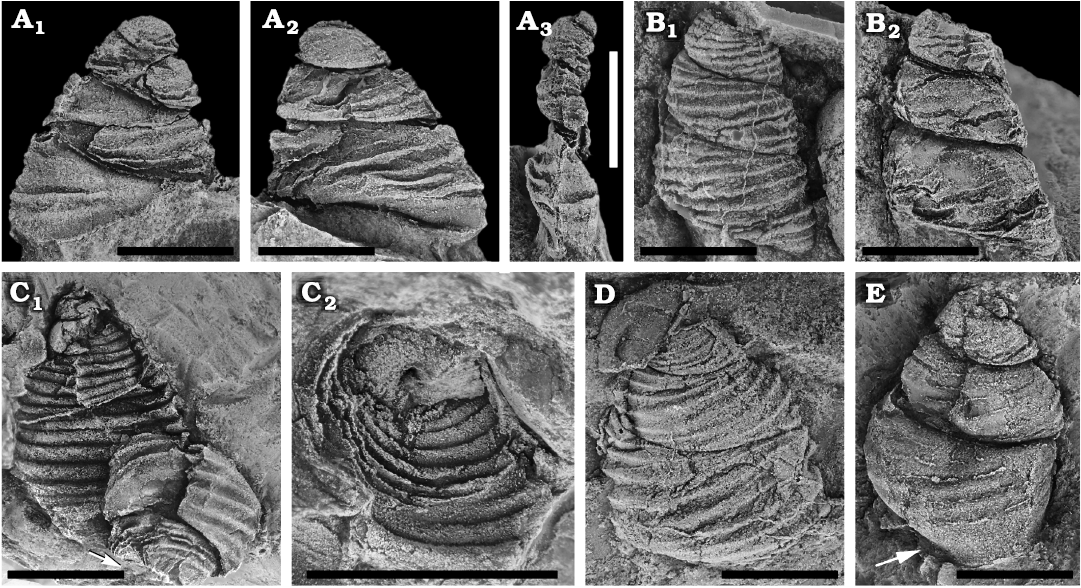
Fig. 4. Clisospirid mimospiridan Mimospira llangynogensis sp. nov. from the Floian, Lower Ordovician Bolahaul Member of the Ogof Hên Formation, Dan-lan-y-Castell quarry, Wales, UK. A. NMW 2017.15G.58, holotype, laterally flattened internal mould in apertural (A1) and lateral (A2, A3) views. B. NMW 2017.15G.59, latex cast of external (B1) and internal (B2) mould. C. NMW 2017.15G.60i, ii, two specimens partly preserved as external and internal moulds (C1); counterpart of the internal mould (arrow in C1) showing its base (C2). D. NMW 2017.15G.61, silicon cast of external mould, showing ornamentation and sutures. E. NMW 2017.15G.62, internal mould, arrow points to excavated ledge along the base. Scale bars 5 mm.
Remarks.—All specimens are laterally compressed internal or external moulds, but the large size and ornamentation allows erection of a new species. The holotype is flattened laterally, but is selected as it shows the apertural margin, which seems to be simple without any thickening (Fig. 4A). The inner lip of the aperture is not preserved, but one internal mould show an excavated ledge along the base (arrow in Fig. 4E) suggesting the presence of a thickened margin towards the umbilical area. Fig. 4C2 shows the only preserved basal view, which is an internal mould of the partial counterpart of the lower specimen in Fig. 4C1. The rounded base gives a shallow area towards the axis, but no true umbilicus. The hole seen in the base indicates the space resulting from a dissolved columella.
Only Mimospira sp. from the Tremadocian (Lower Ordovician) Vogtendorf Formation of the Franconian Forest in Germany, tentatively assigned to the genus by Sdzuy et al. (2001), attain a similar size. The single imperfect German specimen differs from M. llangynogensis sp. nov. in having more densely spaced ribs that also become much denser near the base of the whorls. Other Tremadocian species are found in the Prague Basin, with M. cf. helmhackeri in the Trenice Formation and M. aff. helmhackeri and M. sp. in the overlying Mílina Formation, although only the quite fragmentary Trenice specimen has been figured (Kraft et al. 2013, 2015). Another Tremadocian species is Mimospira atava (Moberg and Segerberg, 1906) from the Bjørkåsholmen Formation of southern Sweden, which is a small species with densely ribbed ornamentation (see further discussion in Wängberg-Eriksson 1979; Sdzuy et al. 2001).
The ornamentation in the Late Ordovician species M. kallholnensis Wängberg-Eriksson, 1979, from the Boda Limestone of Sweden is similar in their distinction and spacing compared to that of M. llangynogensis sp. nov., and the shell reaches a height of 7 mm. The two differ in the ribs being more cord-like in M. kallholnensis and its shell being proportionally narrow and slender. The type species of Mimospira, M. helmhackeri from the Darriwilian of Bohemia, reaches 8 mm in height, and the ornamentation is similar to that of M. kallholnensis, albeit more densely spaced. The base of the last whorl in the type species forms a flange-like extension not seen in other species of the genus.
Bates (1963) described Matherella? acuticostata from the Fennian (Dapingian; Middle Ordovician) Treiorwerth Formation at Trefor in the county of Anglesey, north-west Wales. The species was placed with Mimospira by Wängberg-Eriksson (1979), and consists of a tall shell reaching 8 mm in height but with much more densely spaced oblique lirae than seen in M. llangynogensis sp. nov.
Stratigraphic and geographic range.—Ordovician; Illinois (USA), Tallinn (Estonia), Carmarthen, Wales (UK).
Family Onychochilidae Koken in Koken and Perner, 1925
Subfamily Catalanispirinae Ebbestad, Kolata, and Isakar, 2020a
Genus Catalanispira Ebbestad, Kotala, and Isakar, 2020a
Type species: Catalanispira plattevillensis Ebbestad, Kotala, and Isakar, 2020a, by original designation, from the lower 2 m of the Mifflin Member of the Turinian (Sandbian, Sa2, Upper Ordovician) Platteville Formation (Diplograptus foliaceous Biozone) in northern Illinois, eastern USA.
Catalanispira prima sp. nov.
Fig. 5.
Zoobank LSID: urn:lsid:zoobank.org:act:4E4A585F-9C0C-49BF-A2B7- 467C8303733E
Etymology: From Latin primus, first; from its earliest record of the genus.
Type material: Holotype (NMW 2017.15G.67, Fig. 5B), external mould of the dorsal side, and its external mould. Paratypes (NMW 2017.15G.68–70, Fig. 5A, C, D), internal moulds from the type locality.
Type locality: Dan-lan-y-Castell quarry, Carmarthen, Wales, UK.
Type horizon: Merlinia selwynii Trilobite Zone, Moridunian regional Stage (lower Floian Stage, Fl1).
Material.—Type material only.
Diagnosis.—Species of Catalanispira with 3–3.5 whorls, strong convexity of upper whorl surface and a pleural angle of ~90°.
Description.—Shell large, up to 20 mm across, trochiform with 3–3.5 barely overlapping whorls, where last whorl expands widely. Shell height is about 2/3 of shell width, with the pleural angle being near 90°. Upper whorl surface strongly but evenly convex and sharply rounded at periphery, giving incised sutures. Aperture tangential, elongate elliptical, with a steep umbilical side leading into funnel-like umbilicus. Shell thin with simple comarginal growth lines on a slightly irregular surface.
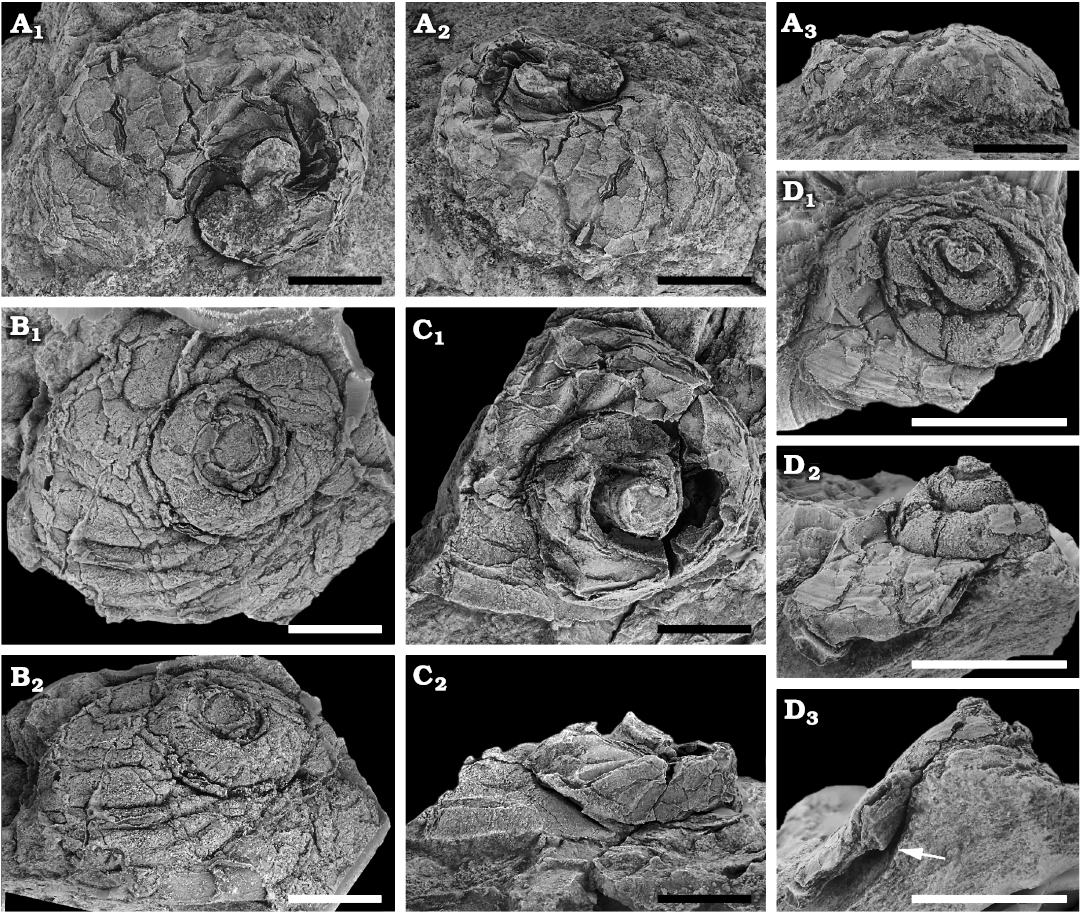
Fig. 5. Onychochilid mimospiridan Catalanispira prima sp. nov. from the Floian, Lower Ordovician, Bolahaul Member of the Ogof Hên Formation, Dan-lan-y-Castell quarry, Wales, UK. A. NMW 2017.15G.68, internal mould, in dorsal (A1), dorsal oblique (A2), and lateral (A3) views. B. NMW 2017.15G.67, silicon cast of the holotype in dorsal (B1) and dorsal oblique (B2) views. C. NMW 2017.15G.69, internal mould, in dorsal (C1) and anterior (C2) views, the latter shows the apertural plane oriented parallel to bedding plane. D. NMW 2017.15G.70, internal mould, in dorsal (D1), lateral (D2), and apertural (D3) views, the last shows original shape of the conch, the umbilical wall is preserved in the last whorl (arrowed). Scale bars 5 mm.
Remarks.—The holotype is an external mould of the dorsal side of the shell from which a silicon cast was made (Fig. 5B), and it demonstrate the large widely expanding shell, number of whorls, and trochiform shape. The convexity of the upper whorl surface is best seen in second largest specimen in Fig. 5A. None of the specimens preserve the base, but the deep funnel-like umbilicus is seen in a lateral view of specimen NMW 2017.15G.70 (Fig. 5D). Although the whorls are somewhat collapsed, most of the convexity is intact. In apertural view it is possible to see the external mould of the umbilical side of the last whorl, showing the narrow lenticular shape of the whorl and steep umbilical wall (Fig. 5D3). The tangential aperture would place the spire at a high angle of inclination, pointing only slightly abaperturally (Fig. 5C). The shell is thin and with simple growth lines (Fig. 5D1, D2). The slightly irregular upper surface is evident in the same specimen, which seems to be the least deformed exemplar. The remaining three specimens show an irregular upper surface with broad comarginal folds of variable prominence. However, this feature is interpreted as largely secondary and enhanced by deformation and compaction.
The new taxon is placed with Catalanispira recently described by Ebbestad et al. (2020a), known from the Sandbian, Late Ordovician C. plattevillensis Ebbestad, Kotala, and Isakar, 2020a, in the USA, and the Darriwilian, late Middle Ordovician C. reinwaldti (Öpik, 1930) from Estonia, owing to the large low trochiform shell, narrow elliptical aperture, deep funnel-like umbilicus and simple ornamentation. It lacks the characteristic broad axe-shaped aperture of the Tremadocian–Floian Pelecyogyra Ebbestad and Lefebvre, 2015, from Morocco and France, which is placed within the same subfamily as Catalanispira (Ebbestad et al. 2020a, b). Pelecyogyra also has fewer whorls and a much greater expansion of the last whorl.
Besides being much older than the two other species of Catalanispira, the Llangynog taxon seems to have more whorls, a more pronounced trochiform shell and stronger convexity of the upper whorl surface. In terms of size, the Estonian C. reinwaldti is larger than C. prima while C. plattevillensis is smaller. The shell shape of the latter resembles that of the Welsh species, but it has fewer whorls.
Stratigraphic and geographic range.—Lower Ordovician (Floian) of Wales, Middle Ordovician (Dw3) of Estonia, and the Upper Ordovician (Sa2) of the USA.
Order uncertain
Family Raphistomatidae Koken, 1896
Remarks.—In his clade analyses, Wagner (2002) recognized “ceratopeoids” as one of three major subclades within the “euomphalinaes” (note that these names were used informally, see Wagner 2002: 16). Wagner (2002) remarked that the “euomphalinaes”, were close to the superfamily Euomphaloidea White, 1877, as defined by Morris and Cleevely (1981, as Euomphalaceae), who included the families Ophiletidae Knight, 1956, Helicotomidae Wenz, 1938, Euomphalidae Koninck, 1881, and Omphalotrochidae Knight, 1945.
The “ceratopeoids” of Wagner (2002) encompassed two subclades, the “raphistomatids” (with the subgroups “lesueurillines”, “scalitines” and “holopeines”) and the “helicotomids” (with the subgroups “ophiletinines” and “euomphalopterines”) (see Wagner 2002: table 3). Species with the genus Ceratopea Ulrich, 1911, were included by Wagner among the early “ceratopeoids” recognized by the strong basal growth lines, a strong flange-like peripheral carina, an upper whorl surface that is rounder than the lower, and a basal carina. The sigmoidal shape of the umbilical growth lines was considered a feature of several early “ceratopeoids” encompassing species in several genera.
Traditionally, the Euomphaloidea encompassed three families in Knight et al. (1960) later increased by two in Bouchet et al. (2005, 2017), the Helicotomidae Wenz, 1938, Euomphalidae Koninck, 1881, and Omphalotrochidae Knight, 1945, respectively, and the Omphalocirridae Wenz, 1938, and Lesueurillidae Wagner, 2002. In the taxonomic revision of Wagner (2002) Ceratopea was placed with the family Raphistomatidae in the superfamily Eumophaloidea. On the other hand, Bouchet et al. (2005, 2017) placed the family with basal gastropods unassigned to a superfamily, and recognized Ceratopeidae Yochelson and Bridge, 1957, as a subjective junior synonym of the Raphistomatidae. Originally Ceratopeidae was erected to encompass taxa with a Ceratopea-like operculum (Yochelson and Bridge 1957).
Bandel and Frýda (1998) proposed the Subclass Euomphalomorpha for Palaeozoic taxa within the Euomphaloidea. The group was recognized mainly by the small egg-shaped and openly coiled protoconch, which is known primarily in Late Palaeozoic members. Morphologically the euomphalomorphid protonchs are more complicated and differ markedly from the simple protoconch morphology of the “Archaeogastropods”. Protoconchs of the early “ceratopeoids” are unknown and the placement of Ceratopea in the Raphistomatidae relies solely on the teleoconch analysis by Wagner (2002). Classification above family level is uncertain (see Wagner 2002: 21), and therefore left tentative. See also discussions in Frýda and Rohr (2004), Frýda et al. (2008), and Frýda (2012).
Genus Ceratopea Ulrich, 1911
Type species: Ceratopea keithi Ulrich, 1911: 665, by original designation, figured in Bassler (1909: 157, pl. 20: 3) as operculum of undetermined gastropod, from the Lower Ordovician Cassinian (Floian, Fl2?, Lower Ordovician) Beekmantown Group at Wytheville, Wythe County, Virginia, USA.
Remarks.—Ceratopea was originally established by Ulrich (1911) based on the horn-shaped calcareous operculum, which is often silicified (Yochelson 1979). Only three associations of partial or complete shells with an operculum in places are known, all attributed to C. unguis Yochelson and Bridge, 1957 (Yochelson and Bridge 1957; Yochelson and Wise 1972; Rohr et al. 2004), while isolated opercula are common (Yochelson 1979). Yochelson and Bridge (1957) pointed out that the question of monophyly of Ceratopea remains problematic as long as the type species is only known from the operculum. The wider scope of the genus presented by Wagner (2002) was also acknowledged as paraphyletic. Wagner (2002) suggested that taxa older than early Middle Ordovician that have been attributed to Raphistomina Ulrich and Scofield, 1897, should belong with Ceratopea.
The Middle to Late Ordovician genus Pararaphistoma Vostokova, 1955, has a somewhat Ceratopea-like adult shell, which lead Yochelson and Copeland (1974) to suggest that Pararaphistoma was a junior synonym of Ceratopea. Wagner (2002) showed that Pararaphistoma is a derived lesueurillid genus, with a marked ontogenetic change in morphology from a lesueurillid morphology in early ontogeny to a Ceratopea-like morphology in later ontogeny, mainly due to clockwise rotation of the aperture. The genus also possesses a slit at the apex of the sinus, which is lacking in Ceratopea.
Species of Pararaphistoma have been reported from the Tremadocian–Floian (Lower Ordovician) of the Montagne Noire in France (Yochelson in Babin et al. 1982) and the Canning Basin of Australia (Yü 1993), all attributed to the Darriwilian (Middle Ordovician) P. qualteriata (Schlotheim, 1820) or P. vaginata Koken in Koken and Perner, 1925, of Baltica. The latter is named from France and Australia, and the material from these two areas appears quite similar. The shells are flattened lenticular and widely phaneromphalus, with a rounded base, depressed whorls with upper whorl surface sloping adaxially and being gradate, and as such they differ from a Ceratopea-type morphology. A second species from Australia is named P. qualteriata, and is known from internal moulds (Yü 1993). The shell appears much more Ceratopea-like in the overall shape and convexity, which is of course also applicable to the shape of P. qualteriata from Baltica. In general, the internal moulds do not allow any closer comparison.
At least 40 species of Ceratopea have been recognized, ranging from the Tremadocian (Early Ordovician) to the Darriwilian (Middle Ordovician; Cullison 1944; Yochelson 1964; Yochelson and Bridge 1957; Yochelson and Copeland 1974; Yochelson and Peel 1975; Wagner 2002; Rohr et al. 2015 and references in these). It is mainly a Laurentian genus, and opercula are common in many Lower Ordovician deposits of USA, Canada, and North and East Greenland in particular but the genus is also recognized in Svalbard and Scotland (Yochelson 1964), the Darriwilian of the Farewell Terrane of Alaska (Rohr and Blodgett 1988; Wagner 2002), the Dapingian and Darriwilian (Middle Ordovician) of the Argentinian Precordillera (Bertero 2007; Ebbestad et al. 2013), and the Darriwilian of North China (Ebbestad et al. 2013; data from the Paleobiology Database compiled by Peter Wagner).
Ceratopea? moridunensis sp. nov.
Figs. 6, 7.
Zoobank LSID: urn:lsid:zoobank.org:act:564B114F-A213-458A-9423- 17585217B21F
Etymology: From Moridunum the Latin name for Carmarthen, South Wales, where the Llangynog Inlier is situated.
Type material: Holotype (NMW 2017.15G.71, Fig. 6A), large specimen with dorsal side preserved and preserving the convexity. Paratypes (45 specimens, NMW 2017.15G.72–113), internal and external moulds, all from the type locality. One slab has four specimens (Fig. 7).
Type locality: Dan-lan-y-Castell quarry, Carmarthen, Wales, UK.
Type horizon: Merlinia selwynii Trilobite Zone, Moridunian regional Stage (lower Floian Stage, Fl1).
Material.—Type material only.
Diagnosis.—A medium-sized species of Ceratopea with a prominent peripheral flange-like carina, base without circumbilical carina, and a steep inner margin without thickening.
Description.—Shell lenticular, phaneromphalus with 5–6 whorls of uniform expansion. Aperture tangential. Initial whorls probably abandoned and plugged. Largest shell close to 30 mm across. Spire low with large pleural angle, estimated to be >120°. Whorls overlap at or just below periphery, giving a slightly gradate shell with incised sutures. Upper whorl surface arched at suture, evenly convex but becoming concave near periphery which is carinated, flange-like, rounded, and prominent but narrow. Inner margin of aperture straight, seemingly without thickening, steep with an angle of about 30–35° relative to axis of coiling. Base evenly but sharply rounded, without a carina. Lower surface gently convex, sloping at about 45° relative to axis of coiling. Transition to peripheral carina abrupt. Peripheral sinus symmetrical, V-shaped and wide. Depth unknown. Transition to peripheral carina and presence/absence of notch unclear. Growth lines on upper surface gently prosocline. Growth lines on lower whorl surface gently prosocyrt with a slightly stronger abapertural curvature near base, continuing with a concave curvature on the inner margin giving an overall sigmoidal shape from the lower whorl surface to the umbilicus and an excavated shallow sinus at the innermost part of the umbilicus. Shell thin, seemingly without any thickening even at the base. Ornamentation consists of fine comarginal growth lines on upper and lower whorls, but that form slightly uneven comarginal crenulations on the lower surface.
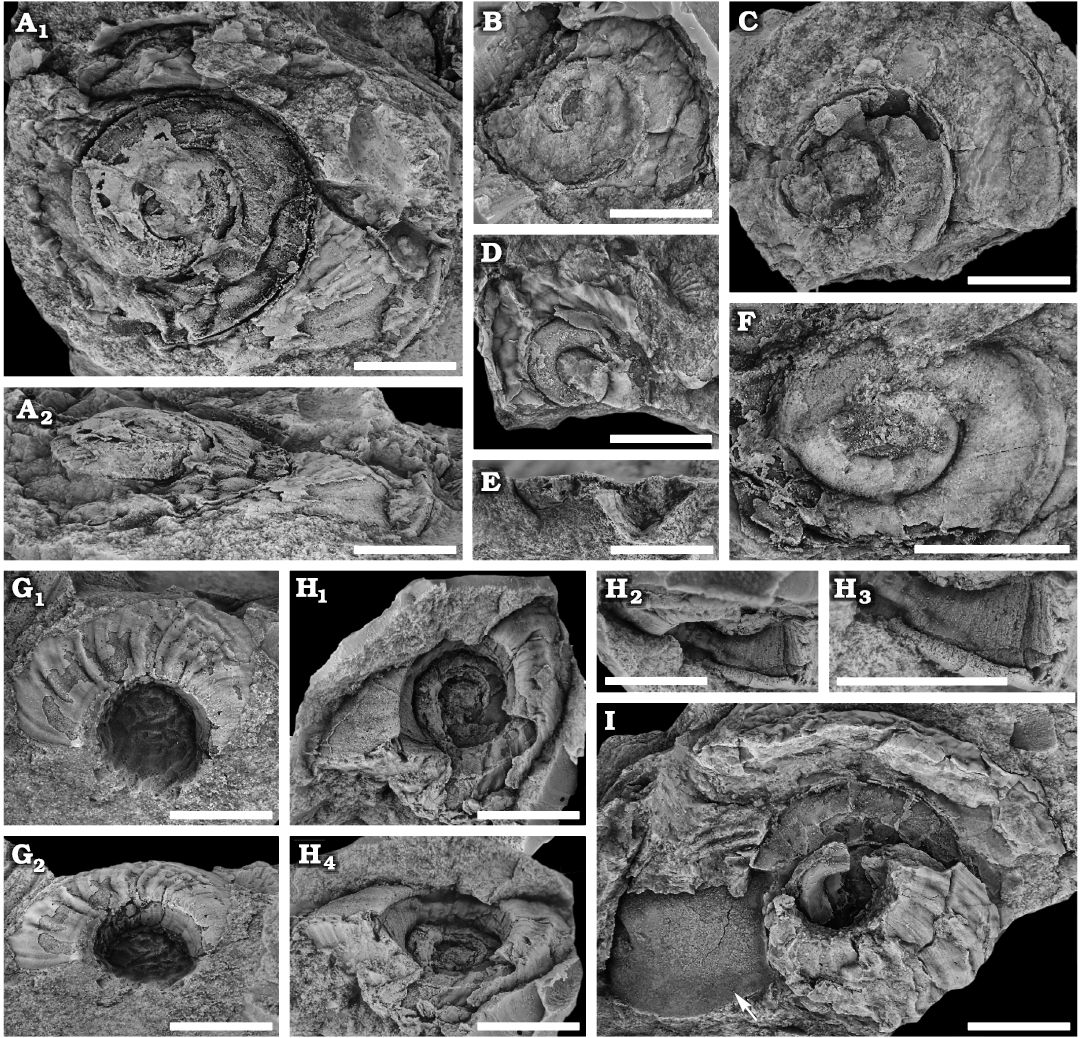
Fig. 6. Raphistomatid gastropod Ceratopea? moridunensis sp. nov. from the Floian, Lower Ordovician Bolahaul Member of the Ogof Hên Formation, Dan-lan-y-Castell quarry, Wales, UK. A. NMW 2017.15G.71, holotype in dorsal (A1) and near apertural (A2) views. B. NMW 2017.15G.72, silicon cast in dorsal view, showing the curvature of the aperture. C. NMW 2017.15G.73, internal mould in dorsal view, showing the flange-like peripheral carina in the early whorls. D. NMW 2017.15G.74, internal mould in dorsal view, with partially preserved distorted shell. E. NMW 2017.15G.76, natural cross section, showing the shape of the lower parts and base of the whorls. F. NMW 2017.15G.75, initial whorls in dorsal view, showing their convexity. G. NMW 2017.15G.77, uncompressed partial specimen in ventral (G1) and ventral oblique (G2) views, showing shape of ornamentation and the wrinkled appearance of comarginal bands. H. NMW 2017.15G.78, silicon cast of ventral side in ventral view (H1), ornamentation and details (H2, H3), umbilical morphology (H4). I. NMW 2017.15G.79, large partial specimen in ventral view, showing the wrinkled appearance of the ornamentation. The arrow points to a bivalve. Scale bars 5 mm.
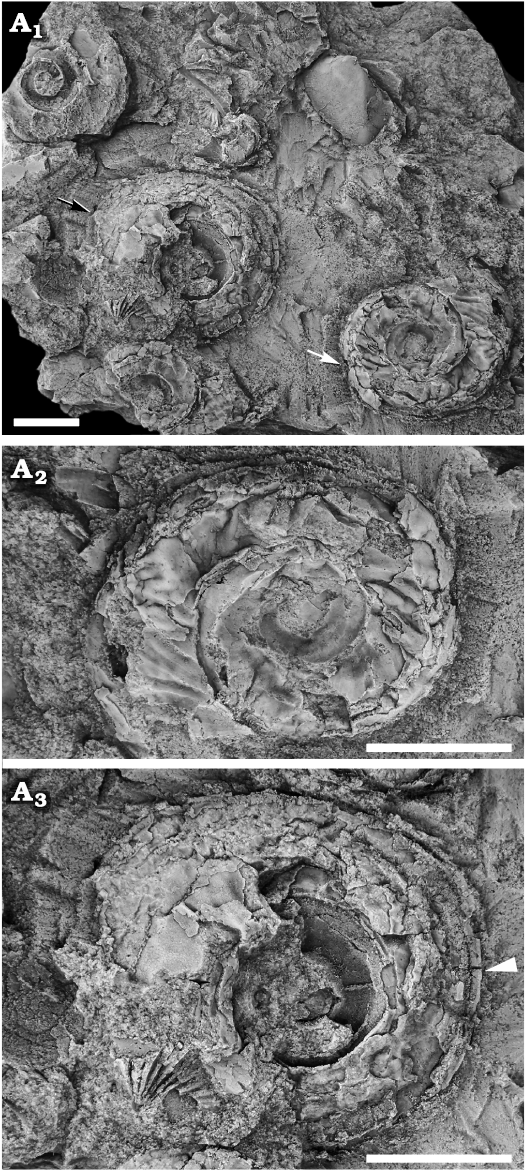
Fig. 7. Raphistomatid gastropod Ceratopea? moridunensis sp. nov. from the Floian, Lower Ordovician Bolahaul Member of the Ogof Hên Formation, Dan-lan-y-Castell quarry, Wales, UK. A. NMW 2017.15G.80; A1, overview of slab with four specimens in ventral views; A2, specimen with preserved wrinkled shell (white arrow in A1); A3, specimen (black arrow in A1) showing the flange-like peripheral carina (arrowhead). Scale bars 5 mm.
Remarks.—The apex whorls are not preserved in any specimen of Ceratopea? moridunensis sp. nov., and it is likely that these were abandoned and possibly septate or filled with (now dissolved) shell material, a condition that is found in many Palaeozoic gastropod groups (Yochelson 1971; Wagner 2002; Cook et al. 2015). Because of this condition, estimation of the number of whorls is difficult but certainly exceeds five.
Ornamentation and the shape of the comarginal growth lines are seen in several specimens (Fig. 6A, B, F–I), but the sigmoidal shape across the lower whorl surface and umbilicus is best seen in Fig. 6G2, H1. Several specimens have a markedly wrinkled surface, seen on both the dorsal and ventral sides of the shell (for instance Fig. 6D, G, I). The wrinkling of the upper surfaces is chaotic and unstructured while that of the lower surface forms prominent comarginal bands. The overall wrinkling may be partly an artefact of preservation and its effect on a very thin shell, but seems to be amplified on the lower surface by the already underlying comarginal irregularities of the ornamentation. The growth lines on the base are fine.
The rounded peripheral carina extends prominently outwards, and it seems to be present already in early ontogeny (Fig. 6C), although the initial spire is not preserved in any specimen. No growth lines are observed crossing the periphery, but it is assumed that they follow the path of the growth lines above and below the keel. There are no bordering lirae on the keel, so that a lunuate selenizone is not formed, but preservation makes it difficult to see whether there is a notch or slit present or not. Both the holotype (Fig. 6A) and NMW 2017.15G.72 (Fig. 6B) preserve the outer lip and demonstrate the gentle prosocyrt curvature, but show no indication of a notch/slit at the periphery. In the holotype the carina seems to project slightly upwards (Fig. 6A2) but owing to compaction it is unclear whether this is a preservational artefact or not. The specimen in Fig. 7A1, A3 shows a ventral view, where the peripheral carina is visible. There seem to be an abrupt transition to the carina, without any change in convexity of the lower whorl surface.
Wagner (2002) commented that Ceratopea previously was diagnosed only on the operculum morphology, although Yochelson and Copeland (1974) provided a diagnosis combining shell and operculum characters. The broader and more detailed redefinition of the genus by Wagner (2002) suggested that in addition to early “ceratopeoid” synapomorphies (strongly curved sinuses, a peripheral band that points adapically, sigmoidal innermost margin and a thick basal carina), the genus is defined by a lenticular aperture, flange-like periphery, and a rounded (convex) upper whorl surface.
The Llangynog species is best placed with Ceratopea due to the rounded upper whorl surface, the apparently deep V-shaped peripheral sinus leading into the extended peripheral flange-like carina, the steep inner margin and the sigmoidal shape of growth lines creating a shallow sinus at the innermost part of the inner margin. The lack of a basal carina is perhaps the main difference of C.? moridunensis sp. nov. relative to other species of Ceratopea. Species like C. canadensis (Billings, 1865) and C. buttsi Yochelson and Bridge, 1957, have coarser growth lines, a basal carina, less convexity of the upper whorl surface, and a less pronounced peripheral carina (see also Yochelson and Copeland 1974). C. unguis Yochelson and Bridge, 1957, have fine growth lines similar to those of C.? moridunensis sp. nov., but differs in having a basal carina, a less pronounced peripheral carina, a more gradate shell and lacking the inner margin excavation (see Yochelson and Bridge 1957; Rohr et al. 2004). C. lemonei (Flower, 1968) is similar to C.? moridunensis sp. nov. in lacking a basal carina and in the crenulation on the lower surface, but the angle of the inner margin is smaller. C. praevium (Whitfield, 1889) differs mostly from C.? moridunensis sp. nov. by having low convexity of the upper whorl surface.
Geographic and stratigraphic range.—Ordovician; Argentina?, Canada, Greenland, Scotland, and USA.
Concluding remarks
The tergomyan Carcassonnella and the gastropods described herein are a small part of the otherwise molluscan dominated fauna of the Bolahaul Member of the Ogof Hên Formation. Despite the low diversity, the molluscs herein represent an unusual assemblage of forms that in many respects are rare (the sinistral forms), and/or very early representatives of their group. Floian (Early Ordovician) was also a time of high endemism among several taxa, which makes discrete faunal elements important for biogeography. The geographical distribution of the fauna is also important as Avalonia during the Early Ordovician was in a peri-Gondwana setting with a wide Iapetus Ocean separating it from Laurentia (Cocks and Fortey 2009; Torsvik and Cocks 2013, 2017; Domeier 2016).
During the Tremadocian–Floian (Early Ordovician) Carcassonnella is only found in sediments of high latitude peri-Gondwana (Ibero-Armorica, Morocco) where it often occurs gregariously (Ebbestad 2016; Ebbestad et al. 2020b). The genus does not appear in Bohemia until the Darriwilian, and is only found outside peri-Gondwana in the Katian (Late Ordovician; Ebbestad 2008). Mimospira on the other hand is conspicuously absent in the Tremadocian–Floian of the Montagne Noire and Morocco. The genus is found in the Tremadocian peri-Gondwana settings of the Franconian Forest in Germany, Bohemia and Baltica, which also share faunal elements among the trilobites and brachiopods (Sdzuy et al. 2001; Mergl 2006; Popov et al. 2013). The genus is not seen again until the Darriwilian (Middle Ordovician) in Baltica and Bohemia. The Floian (herein) and Dapingian (Middle Ordovician) records (Bates 1963) of Mimospira in Wales are therefore amongst the first. The genus does not appear in Laurentia until the Sandbian (Stewart 2012). The Franconian Forest fauna also share brachiopod elements with Tremadocian of South America (the Puna region; Benedetto et al. 2009) as do brachiopods described from the same locality as the material presented herein (Cocks and Popov 2019).
Catalanispira cymruensis sp. nov. is the stratigraphically oldest member of this newly described genus, otherwise known from the Darriwilian of Baltica and Sandbian (Middle Ordovician) of Laurentia. Increased faunal exchange between Avalonia, Baltica and Laurentia took place from the Middle Ordovician and onwards (Cocks and Fortey 2009; Ebbestad et al. 2013; Lam et al. 2018).
Whereas all these three taxa are known from very few specimens in the collections herein, specimens of Ceratopea? moridunensis are common. This species is not associated with the characteristic operculum, but this is not unusual (Yochelson and Copeland 1974). Species of the Ceratopea are overwhelmingly Laurentian in their distribution throughout the Tremadocian–Darriwilian with rare occurrences in the Argentinian Precordillera and North China in the Darriwilian. It also occurs by then in the Farewell Terrane of Alaska, which was situated close to Siberia in the Early Palaeozoic (Rohr and Blodgett 1988; Blodgett et al. 2002). However, faunal exchange between the peri-Gondwana high latitude Avalonia and equatorial Laurentia across a wide Iapetus Ocean during the Early Ordovician must have been limited. A clear Eastern North America biogeographical entity defined by Ceratopea and several other genera of gastropods and tergomyans is clear (Ebbestad et al. 2013), and the faunal integrity of Laurentia is also documented in other well-studied groups like the rhynchonelliform brachiopods (Harper et al. 2013) and trilobites (Cocks and Fortey 2009).
In summary, it is clear that the taxonomic composition of the Llangynog assemblage described herein does not provide any strong biographical signals, other than broadly to peri-Gondwana areas. The tentative identification of Ceratopea herein is at add odds with the Laurentian dispersal pattern of the genus and the distance between Laurentia and Avalonia at this time, although Avalonia entities were separated from main Gondwana by the Early Ordovician (see discussion in Cocks and Fortey 2009). Findings of early Mimospira species in Avalonia, the Franconian Forest, Bohemia and Baltica may reflect a wide dispersal due to planktotrophy. It has been argued that the protoconch of Mimospira consist of an embryonic shell (protoconch I) and a larval shell (protoconch II) which is present in gastropods with planktotrophic larval dispersal (Frýda et al. 2008; Frýda 2012). This would allow the larvae to stay longer in the water column and facilitate a wider dispersal. Furthermore, Mergl (2006) noted that similarities in the trilobite fauna between peri-Gondwanan Bohemia and Baltic at this time is due to similar depth-related biofacies of cosmopolitan genera, rather than close biogeographic distance. The shared occurrence of Carcassonnella with the contemporaneous peri-Gondwana tergomyan and gastropod faunas of the Montagne Noire and Morocco is interesting. However, with only two specimens and lack in Wales of other components that are typical for the Montagne Noire and Morocco, like Thoralispira Horný and Vizcaïno, 1995, Pelecyogyra Ebbestad and Lefebvre, 2015, and Lesueurilla prima (Barrande in Perner, 1903) does not support any close exchange of faunas between these areas. In comparison, contemporary brachiopods from South Wales, of which 11 species were described from the same locality as the material herein, were closely tied to faunas of peri-Gondwana and Gondwana areas (Cocks and Popov 2019).
Part of the distribution pattern may be explained by different latitudinal position and facies depths. Bivalves are the most common molluscs in the Llangynog assemblage, and the bivalve fauna of the Montagne Noire (Babin in Babin et al. 1982) is in part of equivalent age. It differs in being a deeper water fauna than that of Llangynog and is preserved entirely in mudstone facies. The Montagne Noire was certainly at considerably higher latitude than South Wales in Early Ordovician times and the two factors of latitude and water depth explain a bivalve fauna dominated by heteroconchs (80.7% of the fauna, whereas pteriomorphs only account for 0.4%). At Llangynog pteriomorphians are the most important members of the fauna (47%) and outnumber heteroconchs (38.6%) (all figures from Cope and Kriz 2013).
Acknowledgements
We thank Alexander P. Gubanov (Uppsala University, Uppsala, Sweden), for comments on an early draft. Cindy Howells (National Museum of Wales, Cardiff, Wales, UK), kindly helped with specimen catalogue numbers. Sarah E. Stewart (National Museums of Scotland, Edinburgh, Scotland, UK) and David M. Rohr (Sul Ross University, Alpine, Texas, USA), are thanked for careful reviews. This is a contribution to project IGCP 653, ”The onset of the Great Ordovician Biodiversification Event.”
References
Adrain, J.M. 2013. A synopsis of Ordovician trilobite distribution and diversity. In: D.A.T. Harper and T. Servais (eds.), Early Palaeozoic Biogeography Palaeogeography. Geological Society, London, Memoirs 38: 297–336. Crossref
Babin, C., Courtessole, R., Melou, M., Pillet, J., Vizcaïno, D., and Yochelson, E.L. 1982. Brachiopodes (articulés) et mollusques (bivalves, rostroconches, monoplacophores, gastropodes) de l’Ordovicien inférieur (Trémadocien–Arenigien) de la Montagne Noire (France méridionale). Mémoire de la Société des Etudes Scientifiques de l’Aude 1982: 1–63.
Bandel, K. and Frýda, J. 1998. Position of Euomphalidae in the system of the Gastropoda. Senckenberg Lethaea 78: 103–131. Crossref
Bassler, R.S. 1909. The cement resources of Virginia west of the Blue Ridge. Virginia Geological Survey Bulletin 2-A: 1–309.
Bassler, R.S. 1915. Bibliographic index of American Ordovician and Silurian fossils. United States National Museum Bulletin 92: 1–1521. Crossref
Bates, D.E.B. 1963. A Lower Ordovician gastropod from Anglesey. Geological Magazine 100: 258–259. Crossref
Benedetto, J.L., Vaccari, N.E., Wiasfeld, B.G., Sánchez, T.M., and Foglia, R.D. 2009. Cambrian and Ordovician biogeography of the South American margin of Gondwana and accreted terranes. In: M.G. Bassett (ed.), Early Palaeozoic Peri-Gondwana Terranes: New Insights from Tectonics and Biogeography. Geological Society, London, Special Publications 325: 201–232. Crossref
Bertero, V. 2007. Moluscos gastrópodos de la formación San Juan (Ordovícico inferior). Su taxonomía y rol en las comunidades de invertebrados marinos. 59 pp. Unpublished PhD thesis, Facultad de Ciencias Exactas, Físicas y Naturales, Universidad Nacional de Córdoba, Córdoba.
Billings, E. 1865. Palaeozoic Fossils. Volume I. 426 pp. Dawson Brothers, Montreal. Crossref
Blodgett, R.B., Rohr, D.M., and Boucot, A.J. 2002. Paleozoic links among some Alaskan accreted terranes and Siberia based on megafossils. In: E.L. Miller, A. Grantz, and S.L. Klemperer (eds.), Tectonic Evolution of the Bering Shelf—Chukchi Sea—Arctic Margin and Adjacent Landmasses. Geological Society of America Special Paper 360, 273–290. Geological Society of America, Boulder. Crossref
Bouchet, P., Rocroi, J.-P., Frýda, J., Hausdorf, B., Ponder, W., Valdes, A., and Warén, A. 2005. Classification and nomenclator of gastropod families. Malacologia 47: 1–368.
Bouchet, P., Rocroi, J.-P., Hausdorf, B., Kaim, A., Kano, Y., Nützel, A., Parkhaev, P., Schrödl, M., and Strong, E.E. 2017. Revised classification, nomenclator and typification of gastropod and monoplacophoran families. Malacologia 61: 1–526. Crossref
Butts, C. 1926. The Paleozoic rocks. In: G.I. Adams, C. Butts, and L.W. Stephenson (eds.), Geology of Alabama. Geological Survey of Alabama Special Report 14: 41–230.
Cocks, L.R.M. and Fortey, R.A. 2009. Avalonia: A long-lived terrane in the lower Palaeozoic? Geological Society, London, Special Publications 325: 141–155. Crossref
Cocks, L.R.M. and Popov, L.E. 2019. Early Ordovician brachiopods from south-west Wales. Proceedings of the Geologists’ Association 130: 677–690. Crossref
Cook, A.G., Jell, P.A., Webb, G.E., Johnson, M.E., and Baarli, B.G. 2015. Septate gastropods from the Upper Devonian of the Canning Basin: implications for palaeoecology. Alcheringa 39: 519–524. Crossref
Cope, J.C.W. 1979. The early history of the southern margin of the Towy Anticline in the Carmarthen area, South Wales. In: A.L. Harris, C.H. Holland, and B.E. Leake (eds.), The Caledonides of the British Isles Reviewed. Geological Society of London Special Publication 8: 527–532. Crossref
Cope, J.C.W. 1982. The geology of the Llanstephan Peninsula. In: M.G. Bassett (ed.), Geological Excursions in Dyfed, South-West Wales, 259–269. National Museum of Wales, Cardiff.
Cope, J.C.W. 1996. Early Ordovician (Arenig) bivalves from the Llangynog Inlier, South Wales. Palaeontology 39: 979–1025.
Cope, J.C.W. 2005. Octocorallian and hydroid fossils from the Lower Ordovician of Wales. Palaeontology 48: 433–445. Crossref
Cope, J.C.W. 2018. The Green Castle Sandstone Formation: a new name for the Cwmffrwd Formation (basal Old Red Sandstone) preoccupied by an Early Ordovician Member name. Proceedings of the Geologists’ Association 129: 89–91. Crossref
Cope, J.C.W. and Donovan, S.K. 2005. Parablastoid holdfasts from the Lower Ordovician of South Wales. Geological Journal 40: 295–300. Crossref
Cope, J.C.W. and Kriz, J. 2013. The Lower Palaeozoic palaeobiogeography of bivalves. In: D.A.T. Harper and T. Servais (eds.), Early Palaeozoic Biogeography and Palaeogeography. Geological Society, London, Memoirs 38: 221–241. Crossref
Cullison, J.S. 1944. The stratigraphy of some Lower Ordovician formations of the Ozark uplift. University of Missouri School of Mines and Metallurgy Technical Series Bulletin 15: 1–112.
Cuvier, G. 1797. Tableau elementarie de l’historie naturelle des animaux. 710 pp. Baudouin, Paris. Crossref
Domeier, M. 2016. A plate tectonic scenario for the Iapetus and Rheic oceans. Gondwana Research 36: 275–295. Crossref
Dall, W.H. 1913. Class 4. Gastropoda. Snails. In: C.R. Eastmann (ed.), Textbook of Paleontology, Volume 1, 2nd edition, 514–583. MacMillan and Co., London.
Dzik, J. 1983. Larval development and relationships of Mimospira—a presumably hyperstrophic Ordovician gastropod. Geologiska Föreningens i Stockholm Förhandlingar 104: 231–239. Crossref
Dzik, J. 2020. Decline in diversity of early Palaeozoic loosely coiled gastropod protoconchs. Lethaia 53: 32–46. Crossref
Ebbestad, J.O.R. 2008. The tergomyan mollusc Carcassonnella from the Upper Ordovician of Girvan, Scotland. Palaeontology 51: 663–675. Crossref
Ebbestad, J.O.R. 2016. Gastropoda, Tergomya and Paragastropoda (Mollusca) from the Lower Ordovician Fezouata Formation, Morocco. Palaeogeography, Palaeoclimatology, Palaeoecology 460: 87–96. Crossref
Ebbestad, J.O.R. and Lefebvre, B. 2015. An unusual onychochilid mollusc from the Ordovician (Tremadocian) Fezouata Formation, Morocco. Geobios 48: 427–438. Crossref
Ebbestad, J.O.R. and Yochelson, E.L. 2000. Isostrophic molluscs (Tergomya and Gastropoda) from the Upper Ordovician of Norway. Norsk Geologisk Tidsskrift 80: 187–202. Crossref
Ebbestad, J.O.R., Frýda, J., Wagner, P., Horný, R.J., Isakar, M., Stewart, S., Percival, I.G, Bertero, V., Rohr, D.M., Peel, J.S., Blodgett, R.B., and Högström, A.E.S. 2013. Biogeography of Ordovician and Silurian gastropods, monoplacophorans and mimospirids. In: D.A.T. Harper and T. Servais (eds.), Early Palaeozoic Biogeography and Palaeogeography. Geological Society, London, Memoirs 38: 99–220. Crossref
Ebbestad, J.O.R., Kolata, D.R., and Isakar, M. 2020a. New onychochilid mollusks from the Middle and Late Ordovician of Baltica and Laurentia. Journal of Paleontology, 1–17. Crossref
Ebbestad, J.O.R., Lefebvre, B., Kundura, J.-P., and Kundura, M.-H. 2020b. Pelecyogyra (Mollusca) from the Early Ordovician of the Montagne Noire, France. Geobios 63: 23–32. Crossref
Evans, D.H. 2005. The Lower and Middle Ordovician cephalopod faunas of England and Wales. Monograph of the Palaeontographical Society, London 158: 1–81.
Flower, R.H. 1968. Some El Paso guide fossils. Part 1. New Mexico Institute of Mining and Technology Memoir 22: 1–21.
Fortey, R.A. and Cocks, L.R.M. 2003. Palaeontological evidence bearing on global Ordovician–Silurian continental reconstructions. Earth-Science Reviews 61: 245–307. Crossref
Fortey, R.A. and Owens, R.M. 1978. Early Ordovician (Arenig) stratigraphy and faunas of the Carmarthen district, south-west Wales. Bulletin of the British Museum (Natural History) Geology Series 30: 225–294.
Fortey, R.A. and Rushton, A.W.A. 2000. South Wales. In: R.A. Fortey, D.A.T. Harper, J.K. Ingham, A.W. Owen, A.W.A. Rushton, and N.H. Woodcock (eds.), A Revised Correlation of Ordovician Rocks in the British Isles. Geological Society, London, Special Reports 24: 13–17. Crossref
Frisk, Å.M. and Ebbestad, J.O.R. 2007. Paragastropoda, Tergomya, and Gastropoda (Mollusca) from the Upper Ordovician Dalby Limestone, Sweden. GFF 129: 83–99. Crossref
Frýda, J. 1989. A new species of Mimospira (Clisospiridae, Gastropoda) from the Late Ordovician of Bohemia. Věstník Ústředního ústavu geologického 64: 237–241.
Frýda, J. 2012. Phylogeny of Paleozoic gastropods inferred from their morphology. In: J. Talent (ed.), Earth and Life: Global Biodiversity, Extinction Intervals and Biogeographic Perturbations through Time, XXXII, 395–435. Springer Legacy Series, Berlin. Crossref
Frýda, J. and Rohr, D.M. 1999. Taxonomy and paleobiogeography of the Ordovician Clisospiridae and Onychochilidae (Mollusca). Acta Universitatis Carolinae. Geologica 43: 405–408.
Frýda, J. and Rohr, D.M. 2004. Gastropods. In: B.D. Webby, F. Paris, M.L. Droser, and I.G. Percival (eds.), The Great Ordovician Biodiversification Event, 184–195. Columbia University Press, New York. Crossref
Frýda, J., Nützel, A., and Wagner, P. 2008. Paleozoic gastropoda. In: W.F. Ponder and D.R. Lindberg (eds.), Phylogeny and Evolution of the Mollusca, 239–270. University of California Press, Berkeley. Crossref
Gubanov, A.P and Bogolepova, O.K. 1999. Minute fossils from the Ordovician Uggwa Limestone of the Austrian carnie Alps. Acta Universitatis Carolinae. Geologica 43: 417–420.
Gutiérrez-Marco, J.C., Sarmiento, G.N., and Rábano, I. 2014. Un olistostroma con cantos y bloques del Paleozoico Inferior en la cuenca carbonífera del guadalmellato (Córdoba). Parte II: bioestratigraphía y affinidades paleogeográficas. Revista de la Sociedad Geológica de España 27: 27–45.
Harper, D.A.T., Rasmussen, C.M.Ø., Liljeroth, M., Blodgett, R.B., Candela, Y., Jin, J., Percival, I.G., Rong, J.-Y., Villas, E., and Zhan, R.-B. 2013. Biodiversity, biogeography and phylogeography of Ordovician rhynchonelliform brachiopods. In: D.A.T. Harper and T. Servais (eds.), Early Palaeozoic Biogeography and Palaeogeography. Geological Society, London, Memoirs 38, 127–144. Crossref
Hicks, H. 1873. On the Tremadoc rocks in the neighbourhood of St. David’s, South Wales and their fossil contents. Quarterly Journal of the Geological Society, London 29: 39–52. Crossref
Horný, R.J. 1965. Cyrtolites Conrad, 1838 and its position among the Monoplacophora (Mollusca). Acta Musei Nationalis Pragae 21: 57–70.
Horný, R.J. 1996. Secondary shell deposits and presumed mode of life in Sinuites (Mollusca, Gastropoda). Acta musei Nationalis Pragae, Series B, Historia Naturalis 51: 89–103.
Horný, R.J. 1997a. New, rare, and better recognized Ordovician Tergomya and Gastropoda (Mollusca) of Bohemia. Vĕstnik Českého Geologického Ústavu 72: 223–237.
Horný, R.J. 1997b. Ordovician Tergomya and Gastropoda (Mollusca) of the Anti-Atlas (Morocco). Sbornik Národního muzea B53: 37–78.
Horný, R.J. 1999. Gastropod, paragastropod, and tergomyan biostratigraphy of the Ordovician of Bohemia. Acta Universitatis Carolinae.Geologica 43: 397–400.
Horný, R.J. and Peel, J.S. 1996. Carcassonnella, a new Lower Ordovician bellerophontiform mollusc with dorsally located retractor muscle attachments (Class Tergomya). Vĕstnik Českého Geologického Ústavu 71: 305–331.
Horný, R.J. and Vizcaïno, D. 1995. Thoralispira, a new Lower Ordovician cyrtonellid genus (Mollusca, Tergomya) from the Montagne Noire, southern France. Vĕstnik Českého Geologického Ústavu 70: 25–41.
Horný, R.J. and Vonka, V. 2002. Sinuites community in the Upper Ordovician Bohdalec Formation at Praha-Radotin (Barrandian area, Czech Republic). Časopis Národního muzea. Řada přírodovĕdná 170: 42–46.
Hurst, J.M. 1979. Evolution, succession and replacement in the type upper Caradoc (Ordovician) benthic faunas of England. Palaeogeography, Palaeoclimatology, Palaeoecology 27: 189–246. Crossref
Isakar, M. and Peel, J.S. 1997. A new species of Mimospira (Onychochiloidea, Mollusca) from the Middle Ordovician of Estonia. GFF 119: 275–277. Crossref
Kirk, E. 1927. Macluritella, a new gasteropod genus from the Lower Ordovician of Colorado. The American Journal of Science 14: 288–292. Crossref
Knight, J.B. 1937. Genotype designations and new names for invalid homonyms among Paleozoic gastropod genera. Journal of Paleontology 11: 709–714.
Knight, J.B. 1945. Some new genera of Paleozoic Gastropoda. Journal of Paleontology 19: 573–587.
Knight, J.B. 1956. New families of Gastropoda. Journal of the Washington Academy of Sciences 46: 41–42.
Knight, J.B., Cox, L.R., Keen, A.M., Batten, R.L., Yochelson, E.L., and Robertson, R. 1960. Systematic descriptions. In: R.C. Moore (ed.), Treatise on Invertebrate Paleontology, Part I, Mollusca 1, I169–I324. Geological Society of America and University of Kansas Press, Boulder, Lawrence.
Kobayashi, T. 1958. Some Ordovician Fossils from the Thailand-Malayan Borderland. Japanese Journal of Geology and Geography 29: 223–231.
Koken, E. 1896. Die Leitfossilien. 848 pp. Chr. Herm. Tauchnitz, Leipzig.
Koken, E. 1898. Ueber untersilurische Gastropoden. Neues Jahrbuch für Mineralogie, Geologie, und Palaeontologie 1: 1–25.
Koken, E. and Perner, J. 1925. Die Gastropoden den baltischen Untersilurs. Memoires de l’Academie des Sciences de Russie, Ser. 8, Classe Physicomathematique 37: i–vii + 1–326.
Koninck, L.G. de 1881. Faune du calcaire carbonifère de la Belgique, 3e partie, Gastéropodes. Musée Royale d’Historie Naturelle Belgique Annales, Série Paléontoloque 6: 1–170.
Kraft, J., Mergl, M., Hroch, T., and Kraft, P. 2013. Index of fossiliferous localities of the Třenice Formation (Lower Ordovician of the Prague Basin, Czech Republic). Folia Musei rerum naturalium Bohemiae occidentalis, Geologica et paleobiologica 47: 33–64. Crossref
Kraft, J., Mergl, M., Hroch, T., and Kraft, P. 2015. Index of fossiliferous localities of the Mílina Formation (Lower Ordovician of the Prague Basin, Czech Republic). Folia Musei rerum naturalium Bohemiae occidentalis, Geologica et paleobiologica 49: 17–50. Crossref
Lam, A.R., Stigall, A.L., and Matzke, N.J. 2018. Dispersal in the Ordovician: Speciation patterns and paleobiogeographic analyses of brachiopods and trilobites. Palaeogeography, Palaeoclimatology, Palaeoecology 489: 147–165. Crossref
Lindskog, A., Eriksson, M.E., Tell, C., Terfelt, F., Martin, E., Ahlberg, P., Schmitz, B., and Marone, F. 2015. Mollusk maxima and marine events in the Middle Ordovician of Baltoscandia. Palaeogeography, Palaeoclimatology, Palaeoecology 440: 53–65. Crossref
Lindström, G. 1884. On the Silurian Gastropoda and Pteropoda of Gotland. Kongliga Svenska Vetenskaps-Akademiens Handlingar 19: 1–250. Crossref
Linnaeus, C. 1758. Systema naturae per regna tria naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Tomus I. Editio decima, reformata. 824 pp. Salvius, Holmiae. Crossref
Longstaff, J. 1924. Descriptions of Gasteropoda chiefly in Mrs. Robert Grays’ collection, from the Ordovician and lower Silurian of Girvan. Quarterly Journal of the Geological Society, London 80: 408–446. Crossref
McCoy, F. 1851. On some new Silurian Mollusca. Annals and Magazine of Natural History, 2nd series 7: 45–63.
Mergl, M. 2006. Tremadocian trilobites of the Prague Basin, Czech Republic. Acta Musei Nationalis Pragae, Series B, Historia Naturalis 62: 1–70.
Miller, S.A. 1889. North American Geology and Palaeontology for the Use of Amateurs, Students and Scientists. 664 pp. Western Methodist Book Concern, Cincinnati. Crossref
Moberg, J.C. and Segerberg, C.O. 1906. Bidrag till kännedomen om ceratopygeregionen med särskild hänsyn till dess utveckling i Fogelsångstrakten. Lunds Universitet Årsskrift, Ny Följd, Avdelning 2 2: 1–113.
Morris, N.J. and Cleevely, R.J. 1981. Phanerotinus cristatus (Phillips) and the nature of euomphalacean gastropods, Molluscans. Bulletin of the British Museum of Natural History (Geology) 35: 195–212.
Novack-Gottshall, P.M. and Miller, A.I. 2003. Comparative geographic and environmental diversity dynamics of gastropods and bivalves during the Ordovician Radiation. Paleobiology 29: 576–604. Crossref
Nützel, A., Lehnert, O., and Frýda, J. 2006. Origin of planktotrophy evidence from early molluscs. Evolution and Development 8: 325–330. Crossref
Öpik, A. 1930. Beiträge zur kenntnis der Kukruse (C2–C3) Stufe in Eesti. Acta et Commentationes Universitatis Tartuensis. Series A, Matematica, Physica, Medica 14: 1–34.
Paul, C.R.C. and Cope, J.C.W. 1982. A parablastoid from the Arenig of South Wales. Palaeontology 25: 499–507.
Peel, J.S. 1975. New Silurian gastropods from Nova Scotia and Britain. Canadian Journal of Earth Sciences 12: 1524–1533. Crossref
Perner, J. 1900. Miscellanea Silurica Bohemiae. Příspěvky k poznáni Ċeského siluru. Díl 1. 16 pp. Česká Akademie císařte Františka Josefa pro vědy, slovesnost a umění v Praze.
Perner, J. 1903. Systéme Silurien du Centre de la Bohême par Joachim Barrande. 1ère Partie: Recherches Paléontologiques, Volume 4, Gastéropodes, Tome 1, Texte (Patellidae et Bellerophontidae) et Planches 1–89. 164 pp. C. Bellmann, Prague.
Popov, L.E., Holmer, L.E., Massett, M.G., Ghobdai Pour, M., and Percival, I.G. 2013. Biogeography of Ordovician linguliform and craniiform brachiopods. In: D.A.T. Harper and T. Servais (eds.), Early Palaeozoic Biogeography and Palaeogeography. Geological Society, London, Memoirs 38: 117–126. Crossref
Purdue A.H. and Miser H.D. 1916. Geological Atlas of the United States. Eureka Springs-Harrison folio. Arkansas-Missouri. 21 pp. United States Geological Survey, Washington.
Reed, F.R.C. 1920. A monograph of the British Ordovician and Silurian Bellerophontacea. Part I. Palaeontographical Society Monograph 72: 1–48. Crossref
Riding, R., Cope, J.C.W., and Taylor, P.D. 1998. A Coralline-like red alga from the early Ordovician of Wales. Palaeontology 41: 1069–1076.
Rohr, D.M. 1996. Ordovician (Whiterockian) Gastropods of Nevada, Part 2. Journal of Paleontology 70: 56–63. Crossref
Rohr, D.M. and Blodgett, R.B. 1988. First occurrenceof Helicotoma Salter (Gastropoda) from the Ordovician of Alaska. Journal of Paleontology 62: 304–306. Crossref
Rohr, D.M., Fix, M.F., and Darrough, G. 2004. Life association of shell and operculum of Ceratopea Ulrich, 1911 (Ordovician; Gastropoda). Journal of Paleontology 78: 218–220. Crossref
Rohr, D.M., Harper, D.A.T., Stouge, S., and Christiansen, J.L. 2015. Ordovician Gastropoda from Northeast Greenland. Bulletin of Geosciences 90: 795–805 Crossref
Schlotheim, E.F. 1820. Die Petrefactenkunde auf ihren jetzigen Standpunkte durch die Beschreibung seiner Sammlung versteinerter und fossiler Überreste des Their- und Pflanzenreichs der Vorwelt erläutert. 436 pp. Beckerschen Buchhandlung, Gotha.
Sdzuy, K., Hammann, W., and Villas, E. 2001. The Upper Tremadoc fauna from Vogtendorf and the Bavarian Ordovician of the Frankenwald (Germany). Senckenbergiana lethaea 81: 207–261. Crossref
Sepkoski, J.J. 1981. A factor analytic description of the marine fossil record. Palaeontology 24: 36–53. Crossref
Servais, T. and Harper, D.A.T. 2018. The Great Ordovician Biodiversification Event (GOBE): definition, concept and duration. Lethaia 51: 151–164. Crossref
Servais, T., Owen, A.W., Harper, D.A.T., Kröger, B., and Munnecke, A. 2010. The Great Ordovician Biodiversification Event (GOBE): The palaeoecological dimension. Palaeogeography, Palaeoclimatology, Palaeoecology 294: 99–119. Crossref
Sowerby, J.C. de 1839. Fossil shells in the lowest beds of the Old Red Sandstone and fossil shells of the Upper Ludlow Rock. In: R.I. Murchison (ed.), The Silurian System, 602–604. John Murray, London.
Stewart, S.E. 2012. Distribution and palaeoecology of Ordovician bivalves and gastropods from Girvan, SW Scotland. Earth and Environmental Science Transactions of the Royal Society of Edinburgh 102: 145–158. Crossref
Strahan, A., Cantrill, T.C., Dixon, E.E.L., and Thomas, H.H. 1909. The Geology of the South Wales Coalfield, Part 10. The Country Around Carmarthen, Being an Account of the Region Comprised in Sheet 229 of the Map. Memoir of the Geological Survey of Great Britain, England and Wales. viii + 177 pp. His Majesty’s Stationery Office, London.
Taylor, P.D. and Cope, J.C.W. 1987. A trepostome bryozoan from the Lower Arenig of South Wales: implications of the oldest described bryozoan. Geological Magazine 124: 367–371. Crossref
Torsvik, T. and Cocks, L.R.M. 2013. New global palaeogeographical reconstructions for the early Palaeozoic and their generation. In: D.A.T. Harper and T. Servais (eds.), Early Palaeozoic Biogeography and Palaeogeography. Geological Society, London, Memoirs 38: 5–24. Crossref
Torsvik, T. and Cocks, L.R.M. 2017. Earth History and Palaeogeography. 317 pp. Cambridge University Press, Cambridge. Crossref
Ulrich, E.O. 1911. Revision of the Paleozoic systems. Bulletin of the Geological Society of America 22: 281–680. Crossref
Ulrich, E.O. and Scofield, W.H. 1897. The Lower Silurian Gastropoda of Minnesota. Geology of Minnesota. Final Report of the Geological Survey of Minnesota 3: 813–1081.
Vanuxem, L. 1842. Geology of New York. Part 3. Comprising the Survey of the Third Geological District. 306 pp. New York State Geological Survey, White and Visscher, Albany.
Vizcaïno, D., Álvaro, J.J., and Lefebvre, B. 2001. The Lower Ordovician of the southern Montagne Noire. In: D. Vizcaïno and J.J. Álvaro (eds.), The Cambrian and Lower Ordovician of the Southern Montagne Noire: A Synthesis for the Beginning of the New Century. Annales de la Société Géologique du Nord (2e série) 8: 213–220.
Vostokova, V.A. 1955. Ordovician gastropods of Leningrad and Pribaltica [in Russian]. Voprosy Paleontologii 2: 82–124.
Wagner, P.J. 2002. Phylogenetic relationships of the earliest anisostrophically coiled gastropods. Smithsonian Contributions to Paleobiology 88: 1–152. Crossref
Wahlman, G.P. 1992. Middle and Upper Ordovician symmetrical univalved mollusks (Monoplacophora and Bellerophontina) of the Cincinnati Arch Region. Professional Papers of the United States Geological Survey 1066-0: 1–213. en. Sveriges Geologiska Undersökning C758: 1–33. Crossref
Wängberg-Eriksson, K. 1979. Macluritacean gastropods from the Ordovician and Silurian of Sweden. Sveriges Geologiska Undersökning C758: 1-33.
Wenz, W. 1938. Gastropoda. Teil 1: Allgemeiner Teil und Prosobranchia. In: O.H. Schindewolf (ed.), Handbuch der Paläozoologie, Band 6. XII + 480 pp. Bornträger, Berlin.
White, C.A. 1877. The invertebrate fossils collected in portions of Nevada, Utah, Colorado, New Mexico, Arizona, by parties of the Expeditions of 1871, 1872, 1873, and 1874. Chapter VI, Carboniferous age. Report upon U.S. Geographical Surveys west of the 100th Meridian (G.M. Wheeler surveys) 4: 79–161. Crossref
Whitfield, R.P. 1889. Observations on some imperfectly known fossils from the Calciferous sandrock of Lake Champlain, and descriptions of several new forms. Bulletin of the American Museum of Natural History 2: 41–63.
Yochelson, E.L. 1962. Early Ordovician gastropods from The Oslo region, Norway. Norsk Geologisk Tidsskrift 42: 239–250.
Yochelson, E.L. 1963. The Middle Ordovician of the Oslo Region, Norway. Monoplacophora and Gastropoda. Norsk Geologisk Tidsskrift 43: 133–213.
Yochelson, E.L. 1964. The Early Ordovician gastropod Ceratopea from East Greenland. Meddeleser om Grønland 164: 1–12.
Yochelson, E.L. 1971. A new late Devonian gastropod and its bearing on problems of open coiling and septation. Smithsonian Contributions to Paleobiology 3: 231–241.
Yochelson, E.L. 1979. Gastropod opercula as objects for paleobiogeographic study. In: J. Gray and A.J. Boucot (eds.), Historical Biogeography, Plate Tectonics, and the Changing Environment. Proceedings of the Annual Biology Colloquium and Selected Papers 37: 37–43.
Yochelson, E.L. and Bridge, J. 1957. The Lower Ordovician gastropod Ceratopea. U.S. Geological Survey Professional Paper 294H: 281–304. Crossref
Yochelson, E.L. and Copeland, M.J. 1974. Taphonomy and taxonomy of the Early Ordovician gastropod Ceratopea canadensis (Billings), 1865. Canadian Journal of Earth Sciences 11: 189–207. Crossref
Yochelson, E.L. and Jones, C.R. 1968, Teiichispira, a new early Ordovician gastropod genus. United States Geological Survey Professional Papers 613B: 1–15. Crossref
Yochelson, E.L. and Peel, J.S. 1975. Ceratopea and the correlation of the Wandel Valley Formation, eastern North Greenland. Rapport Grønlands Geologiske Undersøgelse 75: 28–31. Crossref
Yochelson, E.L. and Wise, O.A.J. 1972. A life association of shell and operculum in the early Ordovician gastropod Ceratopea unguis. Journal of Paleontology 46: 681–684.
Yü, W. 1993. Early Ordovician gastropods from the Canning Basin, Western Australia. Records of the Western Australian Museum 16: 437–458.
Acta Palaeontol. Pol. 66 (2): 319–335, 2021
https://doi.org/10.4202/app.00854.2020