

Ordovician enigmatic sclerite-type elements from western Argentina: possible oldest axial components of alcyonacean octocorals
MARCELO G. CARRERA, GUSTAVO G. VOLDMAN, MATIAS J. MANGO, and GALINA P. NESTELL
Carrera, M.G., Voldman G.G., Mango, M.J., and Nestell, G.P. 2021. Ordovician enigmatic sclerite-type elements from western Argentina: possible oldest axial components of alcyonacean octocorals. Acta Palaeontologica Polonica 66 (3): 535–544.
The unusual findings of Lower and Middle Ordovician tiny sclerite-type elements in the San Juan Formation of the Argentine Precordillera are described and analysed. The well-preserved silicified and phosphatized association consists of spindle-shaped morphotypes, ornamented with small regular granules or tubercles with some elements connected by their tips in a linear manner. Some morphologic characteristics of these mostly fused or connected sclerite-type elements are present in the alcyonacean octocorals. Their affinities with sponge spicules are also discussed. The oldest records of alcyonacean sclerites have been dated as late Llandovery to late Wenlock (Silurian). The discovery of possible octocoral skeletal elements in the Lower Ordovician of western Argentina may represent the oldest occurrence of such fundamental skeletal elements in the evolutionary history of the octocorals. Although a proper group assignment is still speculative at this state of knowledge, there is an implication that these early forms could be axial skeletal elements comparable to those seen in the Scleraxonia or Calcaxonia octocorals. One new genus Catenatus and new species Catenatus argentinus are described.
Key words: Octocoralia, sclerite-type elements, Ordovician, Palaeozoic, Precordillera, Argentina.
Marcelo G. Carrera [mcarrera@unc.edu.ar], Gustavo G. Voldman [gvoldman@unc.edu.ar], and Matías J. Mango [matiasjmango@gmail.com], CICTERRA (CONICET-Universidad Nacional de Córdoba), Facultad de Ciencias Exactas, Físicas y Naturales, Av. Vélez Sarsfield 1699, X5016GCA, Córdoba, Argentina.
Galina P. Nestell [gnestell@uta.edu], Department of Earth and Environmental Sciences, University of Texas at Arlington, Arlington, Texas 76019, USA.
Received 19 December 2020, accepted 18 February 2021, available online 20 August 2021.
Copyright © 2021 M.G. Carrera et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
The finding of enigmatic sclerite-type or spicule-type skeletal elements in Ordovician limestone of western Argentina encourages the review and analysis of modern and fossil sclerite- or spicule-type records to assess their taxonomic assignment. According to the characteristic morphology observed, the potential affinities fall into the octocoral sclerites, which are small skeletal elements that are found among the main octocoral groups, and are particularly common in alcyonacean octocorals. However, a critical feature such as the longitudinal connection of such skeletal elements has not been seen in fossil or Recent octocoral forms.
Similar skeletal elements have been previously reported from the Ordovician limestone beds in Argentina and were described as foraminifers (Nestell et al. 2009). However, characteristic feature such as a “bumpy” surface in the sense of Nestell et al. (2009) is not compatible with this group as noted by Georgescu (2018), who proposed that these “foraminifers” probably belong to alcyonacean octocorals.
There are some reports of the presence of octocorals in the Ordovician (Lindström 1978; Reich 2002, 2009; Cope 2005; Fernández-Martínez et al. 2019). These records include mostly gorgonian and pennatulacean-like forms, but do not include alcyonaceans (suborder Alcyoniina), whose oldest undoubtable record comes from the Silurian Visby Beds and a rare occurrence in the lowest part of the Högklint Beds of the Swedish island of Gotland (Bengtson 1981). In general, representatives of the Alcyoniina contain tiny calcareous sclerites of different shapes within their soft tissue (coenenchyme). In the fossil record, the presence of octocorals is commonly known based on the findings of their disarticulated sclerites. Many sclerites have been misinterpreted as echinoid spines (Regnéll 1956; Bengtson 1981, and references therein), calcareous sponge spicules (Bengtson 1979; Finks et al. 2004: 171), and monothalamous foraminifers (Nestell et al. 2009, 2011; Mestre et al. 2013). In the Silurian strata of Gotland, the presence of soft corals has been confirmed based on the findings of calcareous spindle-shaped sclerites (or spicules in the sense of Bengtson 1981) of the genus Atractosella Hinde, 1888.
The aim of this contribution is to report and describe the exceptional findings of Early and Middle Ordovician sclerite-type elements, that could be related to alcyonacean octocorals. These Argentinean forms would be the earliest report of alcyonacean octocorals in the geological record.
Institutional abbreviations.—CEGH-UNC, Centro de investigaciones en Ciencias de la Tierra, Consejo Nacional de Investigaciones Científicas y Técnicas, Universidad Nacional de Córdoba (CICTERRA, CONICET-UNC), Argentina.
Nomenclatural acts.—This published work and the nomenclatural acts it contains have been registered in ZooBank: urn:lsid:zoobank.org:pub:3789F943-B5A7-4B61-9EB5-316185763E2F
Geological setting
The San Juan Formation (Kobayashi 1937; Keller et al. 1994) of the Argentine Precordillera is well known for its excellent outcrops of this early Palaeozoic carbonate unit in the foothills of the Andes (Fig. 1). The unit contains a rich fauna of brachiopods, trilobites, crinoids, gastropods, nautiloids, ostracods, sponges, calcareous algae, and problematic microorganisms. The limestone beds of the San Juan Formation, up to 300 m thick, consist mainly of skeletal and intraclastic wackestone and packstone deposited between the fair-weather and storm-wave base in a mid-ramp setting (Cañas 1995, 1999; Carrera et al. 2013).
The age of the San Juan Formation is well constrained by conodonts, ranging from the upper part of the Paltodus deltifer–Macerodus dianae zones (middle Tremadocian, Early Ordovician) (Mango and Albanesi 2020) up to the Yangtzeplacognathus crassus Zone (early–middle Darriwilian, Middle Ordovician) (reviewed by Albanesi and Ortega 2016). The end of the carbonate factory is attributed to a relative rise of sea level that led to the drowning of the platform below the photic zone. Subsequently, sedimentation took place in more restricted environments, with a facies change toward the graptolitic black shales of the Gualcamayo and Los Azules formations (Darriwilian, Middle Ordovician). Tabulate corals have previously been reported from younger, Sandbian (Late Ordovician) units (Fernández-Martínez et al. 2004).

Fig. 1. The geographical and geological context of the Ordovician alcyonacean octocoral Catenatus argentinus gen. et sp. nov. from the San Juan Formation, Argentina. A. Location map of the studied localities with Ordovician alcyonacean sclerite-type elements from the Argentine Precordillera, San Juan Province, Argentina (1, Oculta Creek, Sierra de Los Cauquenes; 2, Los Gatos Creek, Cerro Viejo; 3, Amarilla Creek, Cerro Viejo; 4, Punta Negra Anticline; 5, Sierra de Rinconada). B. Simplified stratigraphic column of the San Juan Formation in the Argentine Precordillera, with location of the productive samples.
Material and methods
The discovery of tiny sclerite-type elements in acid-insoluble residues of conodont samples from lower Palaezoic strata of the Argentine Precordillera encouraged the systematic search for these elements in our repository collections. Only five carbonate samples (total rock weight: 10.5 kg) obtained from different sections of the San Juan Formation were productive, yielding 202 specimens. All of the rock samples were processed in 10% acetic acid or 7% buffered acetic acid following the standard laboratory procedures for recovering conodonts (Jeppsson et al. 1985; Stone 1987). The number of sclerite-type elements and weight of each dissolved limestone sample is presented in Appendix 1.
In this study, the sclerite-type elements were found in three separate regions of the Argentine Precordillera in outcrops of the San Juan Formation (Fig. 1). The oldest assemblage (sample U+20) is from the Punta Negra Anticline, where discontinuous exposures of the San Juan Formation are included in the subvolcanic bodies present in the Ullum Department of the San Juan Province, which obscures the original stratigraphic relationships (Alonso et al. 2014). The associated conodont fauna, preliminarily studied by Voldman et al. (2011), is characterised by the species of Juanognathus variabilis, Periodon flabellum, Reutterodus andinus, Scolopodus krummi, and Tropodus sweeti, and the index species of the Oepikodus evae Zone, precisely indicating a middle–late Floian age (Early Ordovician) for the limestone outcrop.
A second set of samples containing sclerite-type elements is from the extensively studied, top stratum of the San Juan Formation exposed in the Huaco area. In the Los Gatos Creek section, located on the western flank of the Cerro Viejo, sclerite-type elements (sample LG42) are associated with conodonts indicative for the Paroistodus horridus Subzone of the Lenodus variabilis Zone (lower Darriwilian; Mango and Albanesi 2018). One kilometre to the south-west, in the Amarilla Creek section, sclerite-type elements (sample TFSJ QA) occur together with conodonts referred to the overlying Yangtzeplacognathus crassus Zone (lower–middle Darriwilian; Mango et al. 2019). The diachronous nature of the top of the San Juan Formation is further supported by graptolite records, which are indicative of the Levisograptus dentatus Subzone of the L. dentatus Zone in the Los Gatos Creek section, and of the Arienigraptus angulatus Subzone of the L. dentatus Zone in the Amarilla Creek section (Mango et al. 2019). In the Oculta Creek section at the Sierra de Los Cauquenes, 4 km to the west, correlative stratigraphic levels with sclerite-type elements (sample FSJ 3º hardground) are also associated with conodonts of the Y. crassus Zone (Voldman et al. 2013).
Thirdly, sclerite-type elements were recovered from a dm-scale limestone block (sample RIN34), most likely derived from the San Juan Formation, which is embedded in the Rinconada mélange at the eastern margin of the Argentine Precordillera (Voldman et al. 2018). The accompanying conodont fauna is also characteristic of the lower–middle Darriwilian Yangtzeplacognathus crassus Zone.
The sclerite-type elements previously assigned to the foraminiferal genera Lavella? and Amphitremoida (Nestell et al. 2009; Mestre et al. 2013) were recovered from the upper part of the San Juan Formation (Darriwilian, Middle Ordovician) in the Cerro Viejo area (San Juan Province). Nestell et al. (2011) also described such forms as the foraminiferal species Amphitremoida longa Nestell and Tolmacheva, 2004, from the top of the San Juan Formation (Floian, Lower Ordovician) in the Salagasta region, Mendoza Province.
In the case of the top stratum of the San Juan Formation, selective silicification of the possibly originally calcareous sclerite-type elements of alleged octocorals (indicated by Energy-Dispersive X-ray spectroscopy analysis, Fig. 2A) was apparently controlled by the transgressive sequence boundary that marks its top (Carrera et al. 2013), and a hiatus-related silica remobilisation process. Other secondarily silicified microfossils, such as ostracods and trilobites, are also commonly found at these levels along with chert nodules. Some sclerite-type elements show a high siliceous composition, with small impurities mostly represented by aluminum, calcium, iron, sodium and magnesium, possibly related to clay alteration or diagenetic mineral overgrowths (Fig. 2A). Most likely, bacterial sulphate reduction and pyrite formation created low-pH conditions favourable to calcite dissolution and silica precipitation deep in the sediment column (e.g., Muscente et al. 2015).
Additionally, some sclerite-type elements were preserved through phosphatization, possibly as calcium phosphate minerals, such as fluorapatite (Fig. 2B). Despite the fact that phosphatization and silicification of microfossils occurs by similar mineralization processes, they form under notably different conditions. For instance, the decaying soft tissues of the organism itself may create a phosphatizing microenvironment (Briggs and Kear 1993). Replacement of the originally calcareous skeletal elements by phosphate was possibly facilitated in the outer carbonate shelf by upwelling of deep, nutrient-rich seawaters and relatively low rates of sedimentation (e.g., Glenn et al. 1994) during the Darriwilian transgression. The absence of reworked conodont specimens in the studied assemblage attests against phosphatization of the sclerite-type elements in shallower waters, and its subsequent concentration through reworking and winnowing prior to final burial, as documented in other basins (e.g., Glenn et al. 1994; Muscente et al. 2015).
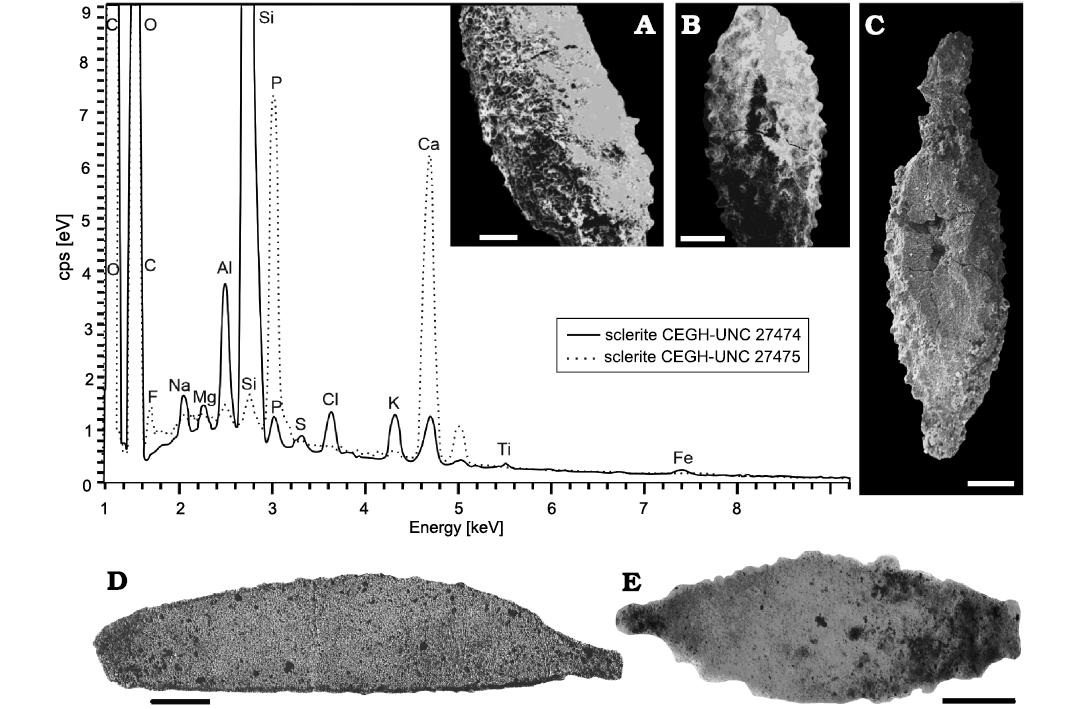
Fig 2. Alcyonacean octocoral Catenatus argentinus gen. et sp. nov. sclerite-type elements from the Middle Ordovician (Darriwilian), Lenodus variabilis– Yangtzeplacognathus crassus zones, San Juan Formation, San Juan Province, Argentina and its chemical composition by surface EDX analysis. A. Siliceous sclerite-type element, CEGH-UNC 27474, sample LG42 (unpolished sample). B. Phosphatic sclerite-type, CEGH-UNC 27475, sample RIN34. C. Partially broken element showing a solid inner structure, CEGH-UNC 27471, sample LG42. D, E. Thin sections of isolated elements of Catenatus argentinus gen. et sp. nov., showing the internal homogeneous structure, CEGH-UNC 27472, sample LG42 (D), and CEGH-UNC 27473, sample FSJ 3º hardground (E). Scale bars 100 μm.
Thin sections of sclerite-type elements were made by embedding an isolated element in Canada balsam and then following the usual procedure for preparing a thin section. The SEM images of sclerite-type elements were taken by JEOL JSM-T300 microscope in the Department of Earth and Environmental Sciences of the University of Texas at Arlington, Arlington, Texas, USA. The Energy-Dispersive X-ray spectroscopy (EDX) of sclerite-type elements was done at the Nano Technology Research Center at the University of Texas at Arlington. The studied specimens are housed under the prefix CEGH-UNC in the Centro de Investigaciones en Ciencias de la Tierra (CICTERRA), Universidad Nacional de Córdoba, Argentina.
Systematic palaeontology
Class Anthozoa Ehrenberg, 1834
Subclass Octocorallia Haeckel, 1866
Order Alcyonacea? Lamouroux, 1812
Genus Catenatus nov.
ZooBank LSID: urn:lsid:zoobank.org:act:71A0EAE5-6756-4BBD-B4 D2-5F3305E70F0A
Etymology: From Latin catenatus, chained; in reference to the longitudinally connected spindle-shaped elements.
Type species: Catenatus argentinus sp. nov; monotypic, see below.
Diagnosis.—Sclerite-type element association of connected spindle-shaped elements or less frequent isolated elements, ornamented with regularly distributed granules or tubercles. The elements show a solid homogeneous structure.
Stratigraphic and geographic range.—Lower to Middle Ordovician (Floian–Darriwilian), San Juan Formation, San Juan Province, Argentina.
Catenatus argentinus sp. nov.
Fig. 3.
2009 Lavella? sp.; Nestell et al. 2009: 332, pl. 1: 6, pl. 4: 2; misidentified as a foraminifer.
2009 Amphitremoida sp. 2; Nestell et al. 2009: 332, pl. 1: 7; misidentified as a foraminifer.
2011 Amphitremoida longa Nestell and Tolmacheva; Nestell et al. 2011: 604, fig. 3 (3); misidentified as a foraminifer.
2013 Lavella? sp.; Mestre et al. 2013: fig. 2 (3); misidentified as a foraminifer.
ZooBank LSID: urn:lsid:zoobank.org:act:CAE9314D-7B79-42F8-AC 1A-ADB671A33FF9
Etymology: After Argentina, the country where the type material was collected.
Type material: Holotype: two connected, spindle shaped sclerite-type elements CEGH-UNC 27465. Paratypes: a set of 17 loose or connected spindle shape sclerites-type elements, CEGH-UNC 27454–27464, CEGH-UNC 27466–27472 from the type ocality and horizon.
Type locality: Los Gatos creek section, western flank of the Cerro Viejo, Huaco area, San Juan Province, Argentina.
Type horizon: Top of the San Juan Formation at the Los Gatos Creek section (Paroistodus horridus Subzone of the Lenodus variabilis Zone (lower Darriwilian; Mango and Albanesi 2018)
Material.—202 sclerite-type elements of which 37 were photographed in SEM and 17 illustrated (CEGH-UNC 27454–27495). 32 elements are from sample FSJ 3o hardground, 19 from sample U+20, 41 from sample RIN 34, 85 from sample LG42, and 25 from sample TFSJ QA.
Description.—The sclerite-type element association consists of a possibly connected association of spindle-shaped elements, ornamented with regularly distributed granular or tubercles (Fig. 3). In our collection, few connected forms were found (Fig. 3I, K1, L, M1). The spindles range from slightly fusiform elongated, rod-like forms to oval or strongly ellipsoidal. The elongated forms range 1–1.8 mm long and 0.2 mm wide, whereas the more inflated spindle forms are 1 mm long to 0.3 mm wide.
The surface of most spindle elements is covered or ornamented by small granules or rounded to oval tubercles (Fig. 3A2, O2, P2), some of them terminating with a tip (Fig. 3H1, M1, M2). Some broken elements show a solid inner structure, which appears porous and homogeneous in thin section (Fig. 2C–E).
Some elements have rounded terminations whereas others show small expansions of the tips. These expansions are frequently found connecting the elements (“neck” connection), forming a linear continuous structure. Small longitudinally aligned corrugations or ridges can be seen at neck or tip junctions (Fig. 3I, K, L, M).
Some sclerite-type elements have two articulation tips (thus they possibly occur in the middle of some linear construction) and some only at one tip, the other one being rounded, suggesting that it was the final (apical) element in the linear structure. There is one form that apparently shows two rounded terminations (Fig. 3H), and if it is the case (excluding physical abrasion of the tips) it can be considered as a typical isolated sclerite commonly found in alcyonacean octocorals.
Remarks.—The studied forms, based on their shape, resemble the Silurian genus Atractosella from the island of Gotland (Bengtson 1981), and they could be included in the same spindle morphotype. However, different morphologies in Atractosella, such as bifurcated or branched spindle forms, are absent in our material, and in contrast, the commonly occurring fused sclerite-type elements in the Argentinean forms are absent in the Silurian Atractosella.
Reich (2002) also reported simple sclerites, which are referred to Atractosella, from glacial erratic Silurian age boulders of Northern Germany. These forms are simple elongated spindles ornamented with crowded small granules, the same type of fine ornamentation as observed in Atractosella and in our sclerite-type elements. However, the Silurian sclerites represent only one of the morphotypes found in Atractosella, whereas the bifurcated or branched forms are absent. The connecting neck, commonly found in the Argentinean material, is also absent among the sclerites of Atractosella described by Reich (2002).
The Silurian Termieralcyon Fernández-Martínez, Coronado, Rodríguez, Tourneur, and Badpa, 2019, an encrusting form attached to a syringoporoid coral described by Coronado et al. (2015) and Fernández-Martínez et al. (2019), has sclerites with slightly pointed and serrated ends that are clearly different and not comparable with the Argentinean forms.
Stratigraphic and geographic range.—Lower to Middle Ordovician (Floian–Darriwilian), San Juan Formation, San Juan Province, Argentina.

Fig. 3. Alcyonacean octocoral Catenatus argentinus gen. et sp. nov. sclerite-type elements. CEGH-UNC 27465 (holotype) and CEGH-UNC 27454–27464, 27466–27472 (paratypes); from Darriwilian, Middle Ordovician (Lenodus variabilis–Yangtzeplacognathus crassus zones (except P, Floian, Lower Ordovician, Oepikodus evae Zone); San Juan Formation, San Juan Province, Argentina. A–C, F, N, P, Q. Spindle shaped, ellipsoidal sclerite-type morphotypes (in the sense of Fernández-Martínez et al. 2019). A. CEGH-UNC 27454 (sample FSJ 3o hardground), general view (A1), enlarged view of the surface of the specimen showing granular sculpture (A2). B. CEGH-UNC 27455 (sample FSJ 3o hardground). C. CEGH-UNC 27456 (sample FSJ 3o hardground). F. CEGH-UNC 27457 (sample TFSJ QA). N. CEGH-UNC 27458 (sample LG42). P. CEGH-UNC 27459 (sample U+20), general view (P1), enlarged view of the surface of the specimen (P2). Q. CEGH-UNC 27460 (sample RIN 34). D, G. Inflated spindle morphotypes. D. CEGH-UNC 27461 (sample FSJ 3o hardground). G. CEGH-UNC 27462 (sample TFSJ QA). K–M. Spindle elements connected by a thin and fragile neck. K. CEGH-UNC 27463 (sample LG42), general view (K1), enlarged view of the neck of the specimen (K2). L. CEGH-UNC 27464, sample LG42. M. Holotype CEGH-UNC 27465 (sample LG42), two connected sclerite-type elements (M1), detailed view of the neck of the specimen, and tips or spine-like protuberances (M2). E, H–J. Elongate spindle morphotypes (in the sense of Fernández-Martínez et al. 2019). E. CEGH-UNC 27466 (sample TFSJ QA), element with possible broken neck. H. CEGH-UNC 27467 (sample TFSJ QA), general outline (H1), note small spine-like protuberances at the top edge, enlarged view of the surface of the specimen showing porous structure (H2). I. CEGH-UNC 27468 (sample TFSJ QA), two elongate spindle elements connected by a thin and fragile neck. J. CEGH-UNC 27469 (sample LG42), elongate spindle morphotype with two polar necks. O. CEGH-UNC 27470 (sample RIN 34), elongate spindle morphotype, slightly curved sclerite-type element (O1), enlarged view of the surface of the specimen showing granular sculpture (O2). Scale bars 100 µm (A–G, H1, I–N, O1) and 10 µm (H2, O2).
Discussion
In the alcyonacean octocorals, the skeleton is made up of sclerites loosely embedded in a soft tissue or connected by organic tissue (Bayer 1956, 1981). The Scleraxonia, a group of gorgonian octocorals, are recognized by the presence of an axis composed of fused or unfused sclerites (see Bayer 1956, 1981; Daly et al. 2007; Reich and Kutscher 2011; Lau et al. 2019 for Recent and fossil examples). Many taxa have an axial skeleton that consists of a dense brown calcified, horn-like substance (gorgonin), or of sclerites that are densely packed together (Lau et al. 2019, and references therein).
Considering Catenatus argentinus gen. et sp. nov. as separated sclerite-type element (as for example the element in Fig. 3H), it shows morphologic characteristics of the sclerites found in fossil and Recent alcyonacean octocorals. Particularly, their shape, dimensions, ornamentation and solid structure are in accordance with those from previously described Palaeozoic octocoral sclerites (Bengtson 1981; Reich 2002, 2009; Fernández-Martínez et al. 2019, and references therein).
We also explored the possibility that these elements were in fact sponge spicules. Externally, octocoral sclerites could resemble some spicules of sponges. In our case, the sclerite-type elements resemble some longitudinally inflated spicules (see for example inflated hexactinellid-base spicules, Carrera and Maletz 2014) or more specifically, the hexactinellid-base spicules described as Rigbyella Mostler and Mosleh-Yazdi, 1976 (Elicki 2011) or thick oxeas (monaxons) of particular sponges (Kozur et al. 1996). However, sponge spicules are clearly differentiated (even in the absence of a preserved spicule axial canals) for their characteristic geometry, and particularly in Rigbyella, a vertical ray perpendicular to a four orthogonal rays at the base of the pinnular spicule, which is clearly different from the structure seen in octocoral sclerites. No orthogonal rays or other spicule-type geometry are seen at the end of each skeletal element from the studied material, especially at the neck connections. Some inflated monaxons, as seen among sponges mainly in microscleres (Mostler 1990; Kozur et al. 1996), could be comparable in shape to the octocoral sclerites; however, internally they are clearly different and they do not show neck type or other kind of connections at their tips. Monaxons are commonly loose in the sponge body and only in some sponge groups they could be laterally welded together.
Catenatus argentinus gen. et sp. nov. displays a porous internal structure in thin sections, which obscures its original structure and the mineralogical composition (Fig. 2D, E). The structure of Recent octocoral sclerites is variable, from simple straight or gently curved layers or homogenously distributed granules to complex multiple divergent fibrous-like alignments (Deflandre‐Rigaud, 1957; Vargas et al. 2010; Fernández-Martínez et al. 2019; Lau et al. 2019 and reference therein).
As we stated above, the general shape and sculpture of the Ordovician forms from Argentina are similar to the Silurian Atractosella sclerites, but they are much more regular in shape. However, the fact that they were articulated forming a linear structure shows an important feature against their interpretation as isolated sclerites. In this sense, the octocoral axial structures in Recent forms shows a variety of skeletal structures from simple calcareous rods (alcyonacean Calcaxonia) to an aggregation of fused sclerites (alcyonacean Scleraxonia). Although unique and not seen among fossil and Recent octocorals, the longitudinally fused structure observed in the Argentinean forms can be related to this octocoral axial structure. Functionally, they are different structures (and probably not homologous), because real sclerites form loose armour in the ectosome/mesenchyme, whereas those described herein look like elements of some linear structure. However, the axial structure in the Scleraxonia is commonly composed of laterally attached, medular branched rods surrounding by extra-medular sclerites (see Daly et al. 2007; Reich and Kutscher 2011; Cairns and Wirshing 2015) and in this sense, it could be considered comparable to the described here “axial” structure.
Conclusions
The association of sclerite-type elements from Argentina are comparable with the octocoral sclerites seen in the representatives of the suborders Alcyoniina, and Scleraxonia. The association is unique because it contains elements connected by an expanded neck termination. This particular structure is not seen among recent octocorals, where the axial structure, for example in the Scleraxonia and Calcaxonia, is composed of laterally attached, medular branched rods or sclerites.
The oldest records of alcyonacean sclerites have been dated as late Llandovery to late Wenlock (Silurian). The discovery of possible octocoral skeletal elements in the Lower Ordovician of western Argentina could represent the oldest occurrence of such skeletal elements in the evolutionary history of the octocorals.
Although a proper group assignment is still speculative at this state of knowledge, there is an implication that these early forms could be axial skeletal elements comparable to those seen in the scleraxonian or calcaxonian octocorals. Based on the morphological differences between the Atractosella Hinde, 1888, and the Argentinean forms, a new genus Catenatus and new species Catenatus argentinus are described.
Acknowledgements
The authors thank Nader Hozhabri and Huan Nguyen (both Shimadzu Institute, Nano Technology Research Center, University of Texas at Arlington, USA) for aiding with geochemical analyses of samples. The authors also thank Merlynd Nestell (Department of Earth and Environmental Sciences, University of Texas at Arlington, USA) for checking English. We thank Mike Reich (University of Göttingen, Germany), Stefan Bengtson (Swedish Museum of Natural History, Stockholm, Sweden), and an anonymous reviewer for their constructive comments and suggestions on earlier versions of the manuscript. This article is a contribution to the International Geoscience Programme (IGCP) Project 653 “The onset of the Great Ordovician Biodiversification Event” and IGCP Project 735 “Rocks and the Rise of Ordovician Life”. This study was supported by CONICET (Consejo Nacional de Investigaciones Científicas y Técnicas): PUE 2016 (CICTERRA-CONICET). MGC acknowledge support from FONCyTGrant PICT 2016 (ANPCYT 285/2017-0588).
References
Albanesi, G.L. and Ortega, G. 2016. Conodont and graptolite biostratigraphy of the Ordovician system of Argentina. In: M. Montenari (ed.), Stratigraphy and Timescales, Vol. 1, 61–121. Elsevier. Crossref
Alonso, J.L., Gallastegui, J., Rodríguez Fernández, L.R., and García-Sansegundo, J. 2014. Stratigraphy and structure of the Punta Negra Anticline. Implications on the structural evolution of the Argentine Precordillera. Journal of Iberian Geology 40: 283–292. Crossref
Bayer, F.M. 1956. Octocorallia. In: R.C. Moore (ed.), Treatise on Invertebrate Paleontology, Part F, Coelenterata, F166–F231. University of Kansas Press, Lawrence.
Bayer, F.M. 1981. On some genera of stoloniferous Octocorals (Coelenterata: Anthozoa) with descriptions of new taxa. Proceedings of the Biological Society of Washington 94: 878–901.
Bengtson, S. 1979. Sponges. In: V. Jaanusson, S. Laufeld, and R. Skoglund (eds.), Lower Wenlock Faunal and Floral Dynamics—Vattenfallet Section, Gotland. Sveriges Geologiska Undersökning C 762: 61–62.
Bengtson, S. 1981. Atractosella, a Silurian alcyonacean octocoral. Journal of Palaeontology 55: 281–294.
Briggs, D.E.G. and Kear, A.J.1993. Fossilization of soft tissue in the laboratory. Science 259: 1439–1442. Crossref
Cairns, S.D. and Wirshing, H.H. 2015. Phylogenetic reconstruction of scleraxonian octocorals supports the resurrection of the family Spongiodermidae (Cnidaria: Alcyonacea). Invertebrate Systematics 29: 345–368. Crossref
Cañas, F.L. 1995. Estratigrafía y evolución paleoambiental de las sucesiones carbonáticas del Cámbrico tardío y Ordovícico temprano de la Precordillera Septentrional, República Argentina. 216 pp. Unpublished Ph.D. Thesis, Facultad de Ciencias Exactas, Físicas y Naturales, Universidad Nacional de Córdoba.
Cañas, F.L. 1999. Facies and sequences of the late Cambrian–Early Ordovician carbonates of the Argentine Precordillera: a stratigraphic comparison with Laurentian platforms. Geological Society of America Special Paper 336: 43–62. Crossref
Carrera, M.G. and Maletz, J. 2014. Ordovician sponge spicules from Spitsbergen, Nevada and Newfoundland: new evidence for hexactinellid and demosponge early diversification. Journal of Systematic Palaeontology 12: 961–981.
Carrera, M.G., Fenoglio, F., Albanesi, G.L., and Voldman, G.G. 2013. Conodonts, sequence stratigraphy and the drowning of the San Juan carbonate platform in the Ordovician of the Argentine Precordillera. In: G.L. Albanesi and G. Ortega (eds.), Conodonts from the Andes. Proceedings of the 3rd International Conodont Symposium and IGCP591 Regional Field Meeting. Asociación Paleontológica Argentina Publicación Especial 13, 5–12. Asociación Paleontológica Argentina, Mendoza.
Cope, J.C.W. 2005. Octocorallian and hydroid fossils from the Lower Ordovician of Wales. Palaeontology 48: 433–445. Crossref
Coronado, I., Fernández-Martínez, E., Rodríguez, S., and Tourneur, F. 2015. Reconstructing a Carboniferous inferred coral-alcyonarian association using a biomineralogical approach. Geobiology 13: 340–356. Crossref
Daly, M., Brugler, M.R., Cartwright, P., Collins, A.G., Dawson, M.N., Fautin, D.G., France, S.C., McFadden, C.S., Opresko, D.M., Rodríguez, E., Romano, S.L., and Stake, J.L. 2007. The phylum Cnidaria: A review of phylogenetic patterns and diversity 300 years after Linnaeus. Zootaxa 1668: 127–182. Crossref
Deflandre‐Rigaud, M. 1957. A classification of fossil alcyonarian sclerites. Micropaleontology 3: 357–366. Crossref
Elicki, O. 2011. First skeletal microfauna from the Cambrian Series 3 of the Jordan Rift Valley (Middle East). Memoirs of the Association of Australasian Palaeontologists 42: 153–173.
Ehrenberg, C.G. 1834. Beiträge zur physiologischen Kenntniss der Corallenthiere im allgemeinen, und besonders des Rothen Meeres, nebst einem Versuche zur physiologischen Systematik derselben. Abhandlungen der Königlichen Akademie der Wissenschaften 1: 225–380.
Fernández-Martínez, E., Carrera, M.G., and Rodríguez, S. 2004. Corales tabulados del Ordovícico (Caradoc) de la Precordillera Argentina. Revista Española de Paleontología 19: 47–59. Crossref
Fernández-Martínez, E., Coronado, I., Rodríguez, S., Tourneur, F., and Badpa, M. 2019. Alcyonacea awakens: palaeobiology and palaeoecology of Palaeozoic octocorals known from their sclerites. Geological Journal 54: 3593–3618. Crossref
Finks, R.M., Reid, R.E., and Rigby, J.K. 2004. Porifera (Demospongea, Hexactinellida, Heteractinida, Calcarea). In: R.L. Kaesler (ed.), Treatise on Invertebrate Paleontology, Part E (Revised), Porifera, Vol. 3, 1–872. The Geological Society of America, Inc. and The University of Kansas, Boulder and Lawrence.
Georgescu, M.D. 2018. Microfossils Through Time: An Introduction First Steps in Micropaleontology. 400 pp. Schweizerbart Science Publishers, Stuttgart.
Glenn, C.R., Föllmi, K.B., Riggs, S.R., Baturin, G.N., Grimm, K.A., Trappe, J., Abed, A.M., Galli-Olivier, C., Garrison, R.E., Ilyin, A.V., Jehl, C., Rohrlich, V., Sadaqah, R.M.Y., Schidlowski, M., Sheldon, R.E., and Siegmund, H. 1994. Phosphorus and phosphorites: Sedimentology and environments of formation. Eclogae Geologicae Helvetiae 87: 747–788.
Haeckel, E. 1866. Generelle Morphologie der Organismen, 2, Anthozoa. 1036 pp. Georg Reimer, Berlin. Crossref
Hinde, J.J. 1888. A monograph of the British fossil sponges. Part II. Palaeontographical Society 40: 93–188. Crossref
Jeppsson, L., Fredholm, S., and Mattiasson, B. 1985. Acetic acid and phosphatic fossils—a warning. Journal of Paleontology 59: 952–956.
Keller, M., Cañas, F., Lehnert, O., and Vaccari, N.E. 1994. The upper Cambrian and Lower Ordovician of the Precordillera (Western Argentina): some stratigraphic reconsiderations. Newsletters on Stratigraphy 31: 115–132. Crossref
Kobayashi, T. 1937. The Cambro–Ordovician shelly faunas of South America. Faculty of Science Journal, Imperial University of Tokyo, Section II, Geology, Mineralogy, Geography, Seismology 4: 369–522.
Kozur, H.W., Mostler, H., and Repetski, J.E. 1996. “Modern” siliceous sponges from the lowermost Ordovician (early Ibexian–early Tremadocian) Windfall Formation of the Antelope Range, Eureka County, Nevada, U.S.A. Geologisch-Paläontologische Mitteilungen Innsbruck 21: 201–221.
Lamouroux, M. 1812. Extrait d’un mémoire sur la classification des Polypiers coralligènes non entierement pierreux. Nouveau Bulletin des Science, Société Philomatique III 63: 181–188.
Lau, Y.W., Poliseno, A., Kushida, Y., and Quéré, G. 2019. The classification, diversity and ecology of shallow water octocorals. In: M.L. Goldstein and D.A. DellaSala (eds.), Encyclopedia of the World’s Biomes, 597–611. Elsevier. Crossref
Lindström, M. 1978. An octocoral from the lower Ordovician of Sweden, with Appendix by Pietzner, H.: Geochemische Untersuchungen an einem apatitischen Problematicum aus dem Ordovicium von Schweden. Geologica et Palaeontologica 12: 41–52.
Mango, M.J. and Albanesi, G.L. 2018. Bioestratigrafía y provincialismo de conodontes del tramo medio-superior de la Formación San Juan en el cerro Viejo de Huaco, Precordillera, Argentina. Andean Geology 45: 274–299. Crossref
Mango, M.J. and Albanesi, G.L. 2020. Conodont biostratigraphy and correlation of the San Juan Formation at the Cerro La Silla section, middle Tremadocian–lower Dapingian, Central Precordillera, Argentina. Andean Geology 47: 577–598. Crossref
Mango, M.J., Ortega, G., and Albanesi, G.L. 2019. Conodont and graptolite biostratigraphy of the lower–middle Darriwilian (Middle Ordovician), Cerro Viejo of Huaco, Argentine Precordillera. Geological Journal 54: 3349–3361. Crossref
Mestre, A., Beresi, M., Heredia, S., and Nestell, G.P. 2013. Microfossils of the Yangtzeplacognathus crassus zone in the middle Darriwilian of the Argentine Precordillera. In: G.L. Albanesi and G. Ortega (eds.), Conodonts from the Andes. Proceedings of the 3rd International Conodont Symposium and IGCP591 Regional Field Meeting. Asociación Paleontológica Argentina, Publicación Especial 13, 79–83. Asociación Paleontológica Argentina, Mendoza.
Mostler H, 1990. Mikroscleren von Demospongien (Porifera) aus dem Basalen Jura der Nordlichen Klakalpen. Geologisch-Paläontologische Mitteilungen Innsbruck 17: 119–142.
Mostler, H. and Mosleh-Yazdi, A. 1976. Neue Poriferen aus oberkambrischen Gesteinen der Milaformation im Elbruzgebirge (Iran). Geologisch-Paläontologische Mitteilungen Innsbruck 5: 1–36.
Muscente, A.D., Hawkins, A.D., and Xiao, S. 2015. Fossil preservation through phosphatization and silicification in the Ediacaran Doushantuo Formation (South China): a comparative synthesis. Palaeogeography, Palaeoclimatology, Palaeoecology 434: 46–62. Crossref
Nestell, G.P. and Tolmacheva, T.Y. 2004. Early Ordovician foraminifers from the Lava Section, northwestern Russia. Micropaleontology 50: 253–280. Crossref
Nestell, G.P., Mestre, I., and Heredia, S. 2009. First Ordovician foraminifera from South America: a Darriwilian (Middle Ordovician) fauna from the San Juan Formation, Argentina. Micropaleontology 55: 329–344.
Nestell, G.P., Heredia, S., Mestre I., Beresi, M., and González, M. 2011. The oldest Ordovician foraminifers (Oepikodus evae Conodont Zone, Floian) from South America. Geobios 44: 601–608. Crossref
Regnéll, G. 1956. Silurian Echinoids from Gotland. Arkiv for Mineralogi och Geologi 2: 155–178.
Reich, M. 2002. Skleren von Alcyonacea (Anthozoa: Octocorallia) aus einem Silur-Geschiebe Norddeutschlands. Neues Jahrbuch für Geologie und Paläontologie Monatshefte 2002 (9): 551–561. Crossref
Reich, M. 2009. A critical review of octocorallian fossil record (Cnidaria: Anthozoa). In: M.R. Smith, L.J. O’Brien, and J.-B. Caron (eds.), International Conference on the Cambrian Explosion, Banff, Alberta, Canada, Abstracts, 85. The Burgess Shale Consortium, Toronto.
Reich, M. and Kutscher, M. 2011. Sea pens (Octocorallia: Pennatulacea) from the Late Cretaceous of northern Germany. Journal of Paleontology 85: 1042–1051. Crossref
Stone, J. 1987. Review of investigative techniques used in the study of conodonts. In: R.L. Austin (ed.), Conodonts: Investigative Techniques and Applications, 17–34. Ellis Horwood Limited, Chichester.
Vargas, S., Breedy, F., Siles, O., and Guzman, H.M. 2010. How many kinds of sclerite? Towards a morphometric classification of gorgoniid microskeletal components. Micron 41: 158–164.
Voldman, G.G., Albanesi, G.L., and Ortega, G. 2013. Middle Ordovician conodonts and graptolites at Los Cauquenes Range, Central Precordillera of San Juan, Argentina. In: G.L. Albanesi and G. Ortega (eds.), Conodonts from the Andes. Proceedings of the 3rd International Conodont Symposium and IGCP591 Regional Field Meeting, Asociación Paleontológica Argentina, Publicación Especial 13, 117–121. Mendoza, Argentina.
Voldman, G.G., Albanesi, G.L., Banchig, A.L., Ramos, V.A., and Ortega, G. 2011. Conodontes metamorfizados del Ordovícico Inferior en la Formación San Juan expuesta en Ullum, Precordillera Central de San Juan, Argentina. Reunión Anual de comunicaciones de la Asociación Paleontológica Argentina, Luján. Ameghiniana 48 (4, Supplement): R28.
Voldman, G.G., Alonso, J.L., Fernández, L.P., Ortega, G., Albanesi, G.L., Banchig, A.L., and Cardó, R. 2018. Tips on the SW-Gondwana margin: Ordovician conodont-graptolite biostratigraphy of allochthonous blocks in the Rinconada mélange, Argentine Precordillera. Andean Geology 45: 399–409. Crossref
Acta Palaeontol. Pol. 66 (3): 535–544, 2021
https://doi.org/10.4202/app.00869.2020
Lower and Middle Ordovician conodont samples from the Argentine Precordillera with octocoral sclerite-type elements.
|
Sample |
Section |
Stratigraphic unit |
Level |
Biozone (age) |
Number of alcyonacean sclerites |
Reference |
|
U+20 |
Punta Negra Anticline, Ullum Department |
San Juan Formation |
limestone block |
Oepikodus
evae |
19 |
|
|
RIN 34 |
Sierra de Rinconada |
Rinconada mélange |
limestone block |
Yangtzeplacognathus crassus (Darriwilian) |
41 |
|
|
FSJ (3o hardground) (2150 g) |
Oculta Creek, Sierra de Los Cauquenes |
San Juan Formation |
uppermost levels |
Yangtzeplacognathus crassus (Darriwilian) |
32 |
|
|
LG42 |
Los Gatos Creek, Cerro Viejo of Huaco |
San Juan Formation |
top stratum |
Lenodus
variabilis |
85 |
|
|
TFSJ QA |
Amarilla Creek, Cerro Viejo of Huaco |
San Juan Formation |
top stratum |
Yangtzeplacognathus crassus (Darriwilian) |
25 |