
New data on the distal tarsals in Ornithomimidae
RACHEL E. NOTTRODT and ANDREW A. FARKE
Nottrodt, R.E. and Farke, A.A. 2021. New data on the distal tarsals in Ornithomimidae. Acta Palaeontologica Polonica 66 (4): 789–796.
The ankle in non-avian theropod dinosaurs consists of the astragalus and calcaneum proximally and a distal series of tarsal bones capping the metatarsals. Nearly all theropods have only two distal tarsals, identified as distal tarsal 3 and distal tarsal 4. Historically, the morphology and anatomical relationships of these distal tarsals is uncertain in ornithomimosaurs due to loss and/or disarticulation; even in articulated specimens, the bones can be difficult to access. A previously undescribed ornithomimid fossil from the Kaiparowits Formation (upper Campanian) of southern Utah, USA, provides unique views of the distal tarsals in articulation with their surrounding elements, allowing the most complete assessment yet of this region in an ornithomimid from North America. Distal tarsal 3 contacts both metatarsals II and III, whereas distal tarsal 4 contacts only metatarsal IV. Distal tarsal 4 also shows a tab-like process that projects laterally. Comparison of the new fossil with other ornithomimosaurs shows that distal tarsals in Ornithomimosauria can be generalized as: (i) paired as distal tarsals 3 and 4; (ii) not fused to one another or to the proximal metatarsus; and (iii) proximo-distally compressed. The distal tarsals of ornithomimosaurs vary in the antero-posterior positioning and extent to which they cover the proximal metatarsal surface.
Key words: Theropoda, Ornithomimosauria, Ornithomimidae, tarsals, ankle.
Rachel E. Nottrodt [rachel.nottrodt@ucalgary.ca], University of Calgary, 2500 University Drive NW, Calgary, Alberta, T2N 1N4 Canada.
Andrew A. Farke [afarke@webb.org], Raymond M. Alf Museum of Paleontology at The Webb Schools, 1175 Base Line Rd. Claremont, California, 91711 USA.
Received 25 February 2021, accepted 17 May 2021, available online 26 October 2021.
Copyright © 2021 R.E. Nottrodt and A.A. Farke. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Despite their crucial function in the mesotarsal ankle (Martin et al. 1980), distal tarsals are rarely discussed in the literature describing theropod dinosaurs. These small and thin bones facilitate the articulation between the astragalus and calcaneum on one side of the joint and the metatarsus on the other, yet the morphology and functional significance of the distal tarsals are often less studied in comparison to the larger, more anatomically complex, and more commonly preserved proximal tarsals (astragalus and calcaneum; e.g., Holtz 1995).
Plesiomorphically, dinosaurs have two distal tarsals (Langer 2003; Ossa-Fuentes et al. 2020), either referred to as medial and lateral distal tarsals or numbered as distal tarsals 3 (or III) and 4 (or IV), based upon the metatarsal with which they are most closely associated. This general pattern is hypothesized to result from a change in the ancestral ossification pattern during development that would eventually change further to co-ossify the distal tarsals and the metatarsals into the tarsometatarsus during the dinosaur-bird transition (Ossa-Fuentes et al. 2020). Within non-avian theropods, verifiable reports of a third tarsal (distal tarsal 2) are quite rare, and observations in tyrannosaurids and compsognathids may result from the persistence of three cartilaginous tarsals in early embryonic development (Ossa-Fuentes et al. 2020). Overall, two distal tarsals occur in the vast majority of theropods for which the relevant anatomy is preserved (e.g., Allosaurus, Ostrom 1976; Deinonychus, Ostrom 1969; Sinraptor, Currie and Zhao 1993; oviraptorids: Currie et al. 2016; Funston and Currie 2021; and in ornithomimids: Osmólska et al. 1972).
Throughout Ornithomimosauria, few specimens preserve the distal tarsals completely, due to the small size and delayed ossification of these bones (Currie and Zhao 1993; Padian 2017). In specimens that do preserve the distal tarsals in articulation, their position within the ankle and the “death pose” of many well-preserved specimens often make it difficult, if not impossible, to access the distal tarsals for study. Further, the distal surfaces of the distal tarsals, i.e., the surfaces that articulate with the proximal metatarsus, are rarely accessible. Thus, the tarsals are only partially visible in even the best circumstances.
To date, distal tarsals have been described previously in the basal ornithomimosaur, Nqwebasaurus thwazi (De Klerk et al. 2000; Choiniere et al. 2012), as well as the Asian ornithomimosaurs Sinornithomimus dongi (Kobayashi and Lü 2003), Archaeornithomimus asiaticus (Gilmore 1933; Russell 1972; Smith and Galton 1990), Gallimimus bullatus (Osmólska et al. 1972), Harpymimus okladnikovi (Barsbold and Perle 1984; Kobayashi and Barsbold 2005b), Garudimimus brevipes (Barsbold 1981; Kobayashi and Barsbold 2005a), Aepyornithomimus tugrikinensis (Chinzorig et al. 2017), and the Bissekty ornithomimid (Sues and Averianov 2016). Among North American ornithomimids, distal tarsals have been noted and described for Ornithomimus velox (Marsh 1890), Struthiomimus altus (Lambe 1902; Osborn 1916; Russell 1972), and Rativates evadens (McFeeters et al. 2016). Most of these descriptions are very brief, generally without detailed figures, and often rely on disarticulated specimens or specimens with incomplete exposure of the distal tarsals. Thus, the distal tarsal anatomy remains incompletely characterized for most ornithomimids, which can in turn limit functional and developmental inferences as well as anatomical comparisons.
The first depictions of ornithomimosaur distal tarsals were for the Ornithomimus velox holotype, YPM 542 (Marsh 1890), and the Struthiomimus altus holotype, CMN 930 (Lambe 1902), in which three tarsals were identified as “t2”, “t3”, and “t4”. Gilmore (1933) then described the distal tarsals of the Archaeornithomimus asiaticus holotype, AMNH 6565, noting that this taxon has two distal tarsals, designated as “t2” and “t3”. It was not until the description of Gallimimus bullatus (Osmólska et al. 1972), that the two distal tarsals were designated as “dtIII” and “dtIV” to match the naming convention used in other theropods. In the same year, Russell (1972) found that the description of the Struthiomimus altus holotype (Lambe 1902) was incorrect due to a reconstruction that overestimated both the number and the shape of distal tarsals present. Referencing a nearly complete specimen, AMNH 5339, Russell (1972) concluded that Struthiomimus altus has only two distal tarsals, resembling those of the Asian taxon, Archaeornithomimus asiaticus. However, this observation, that Asian and North American ornithomimosaur taxa share similar distal tarsal morphology, has variable uptake in subsequent literature that follows the incorrect historical descriptions of Ornithomimus velox (Marsh 1890) and Struthiomimus altus (Lambe 1902) for comparative purposes (e.g., Sues and Averianov 2016). This discrepancy highlights the need for a more thorough review of ornithomimosaur distal tarsal morphology. Fortunately, some previously undescribed North American ornithomimid specimens preserve the distal tarsals, thereby serving as excellent comparisons with the Asian ornithomimosaur taxa.
The current study was spurred by recognition of the unique views for the distal tarsals offered by a partially articulated ornithomimid hind limb from the Kaiparowits Formation of southern Utah, USA. As the first ornithomimid specimen from North America to reveal both the proximal and distal surfaces of both distal tarsal bones (Fig. 1), and one of the very few ornithomimids for which tarsal morphology can be seen in articulation from multiple views, this fossil adds new information for ornithomimosaur distal tarsal morphology in particular and theropod tarsal anatomy in general.
Institutional abbreviations.—AMNH, American Museum of Natural History, New York City, USA; CMN, Canadian Museum of Nature, Ottawa, Canada; GIN, Paleontological Center of Mongolia, Ulaan Bataar, Mongolia; IGM, Mongolian Academy of Sciences, Ulaan Baatar, Mongolia; MNA, Museum of Northern Arizona, Flagstaff, USA; RAM, Raymond M. Alf Museum of Paleontology at The Webb Schools, Claremont, USA; TMP, Royal Tyrrell Museum of Palaeontology, Drumheller, Canada; YPM, Yale Peabody Museum, New Haven, USA; ZPAL, Institute of Paleobiology, Polish Academy of Sciences, Warsaw, Poland.
Other abbreviations: dt, distal tarsal.
Systematic palaeontology
Dinosauria Owen, 1842
Theropoda Marsh, 1881
Coelurosauria von Huene, 1914
Maniraptoriformes Holtz, 1996
Ornithomimosauria Barsbold, 1976
Ornithomimidae Marsh, 1890
Ornithomimidae indet.
Material.—RAM 6794, an associated and partially articulated partial postcranial skeleton, including pelvis, sacrum, some anterior caudal vertebrae, and both hind limbs. RAM 6794 was collected at locality RAM V1998008, within the middle unit of the upper Campanian, Upper Cretaceous Kaiparowits Formation, Grand Staircase-Escalante National Monument, Garfield County, Utah, USA. Detailed locality data are on file at the Raymond M. Alf Museum.
Description.—RAM 6794 represents a relatively small individual, as compared to other ornithomimids from the upper Campanian of North America. The femur measures 327 mm in maximum length, approximately 20% smaller than the 411 mm long femur in an Ornithomimus edmontonicus (Sternberg, 1933) with a body length of 3.6 m (Zelenitsky et al. 2012; TMP 1995.110.1). The most complete metatarsal, right MT IV, is 236 mm long. This is approximately 35% smaller than the Kaiparowits Formation MNA Pl. 1762A, in which the equivalent element is 365 mm long (Decourten and Russell 1985).
The remainder of the description here focuses nearly exclusively on the tarsal complex in RAM 6794, which is most relevant to the questions under consideration. The astragalus and calcaneum are fused with each other, with a faint recessed area indicating the point of contact (Fig. 2A2, A3, A6). This complex is firmly attached to the tibia, but the contact with the tibia remains open. The fibula is not fused with the astragalus-calcaneum complex, either.
During collection and preparation, the left distal tarsals were kept in place against their corresponding astragalus and calcaneum, leaving the distal surface visible (Fig. 1A), whereas the right distal tarsals were retained atop the corresponding metatarsals, leaving the proximal surface exposed (Fig. 1B). Thus, the following description is a composite of observations from both the left and right distal tarsals, which provide distal and proximal views of the distal tarsals, respectively.

Fig. 1. Tarsal region of Ornithomimidae indet., RAM 6794 from upper Campanian Kaiparowits Formation, Utah, USA, as preserved in articulation. A. Left tarsus, tibia, proximal tarsal region, and distal tarsals in lateral view (A1); distal tarsals in distal view and tibia, astragalus, and calcaneum in oblique anterior view (A2). B. Right tarsus, metatarsals and distal tarsals in proximal (B1), anterior (B2), medial (B3), lateral (B4), and posterior (B5), views.
Distal tarsal 3 underlies the medial third of the astragalus and overlies the posterior (plantar) surfaces of the proximal ends of metatarsals II and III, with a small portion of the lateral edge also overlying metatarsal IV (Figs. 1B1, 2A1). The tarsal bone has a somewhat pear-shaped outline in proximal and distal views, with the narrower end directed medially (lies along the medial border), and the broader, lateral end only narrowly separated from distal tarsal 4 (Fig. 1A2, B1). The distal articular surface of distal tarsal 3 is relatively flattened, divided into medial and lateral halves by a transverse ridge that trends antero-posteriorly (Fig. 1A2). This ridge accommodates the proximal surface of metatarsals II and III. The proximal articular surface is strongly convex, imparting a lenticular cross-sectional profile to the bone (Figs. 1B3, 2A4). Visually, distal tarsal 3 appears taller than distal tarsal 4; however, the bones are roughly similar in thickness when measured. In the part that remains in position relative to the astragalus, a modest gap separates the two bones, indicating the distal tarsals are not fused to each other (Fig. 1A2).
Distal tarsal 4 underlies the middle portion of the astragalus, does not articulate with the calcaneum, and overlies the proximal posterior (plantar) surface of metatarsal 4 alone (Figs. 1B1, B5, 2A1, A2, A5). The position of distal tarsal 4 leaves visible the anterior and very small portions of the posterior surface of the proximal end of metatarsal IV (Figs. 1B1, 2A1). This tarsal is divided into two parts, with a medial portion that forms most of the element and a narrower, laterally directed flange. The distal surface of the overall tarsal is relatively flat, with a slight bit of concavity towards the ventral edge of this surface and a slight convexity towards the dorsal edge (this morphology is somewhat obscured by matrix in Fig. 1A2). Like distal tarsal 3, distal tarsal 4 has a prominent lenticular cross-section, with the proximal surface strongly rounded (Figs. 1B4, 2A6). In combined articulation, distal tarsals 3 and 4 together produce a continuous, rounded proximal surface, with the peak of the rounding positioned slightly ventral, relative to the main bodies of the tarsals. The combined effect is a subtly saddle-shaped articular surface between proximal and distal tarsals, with the long axis of the “saddle” directed mediolaterally.
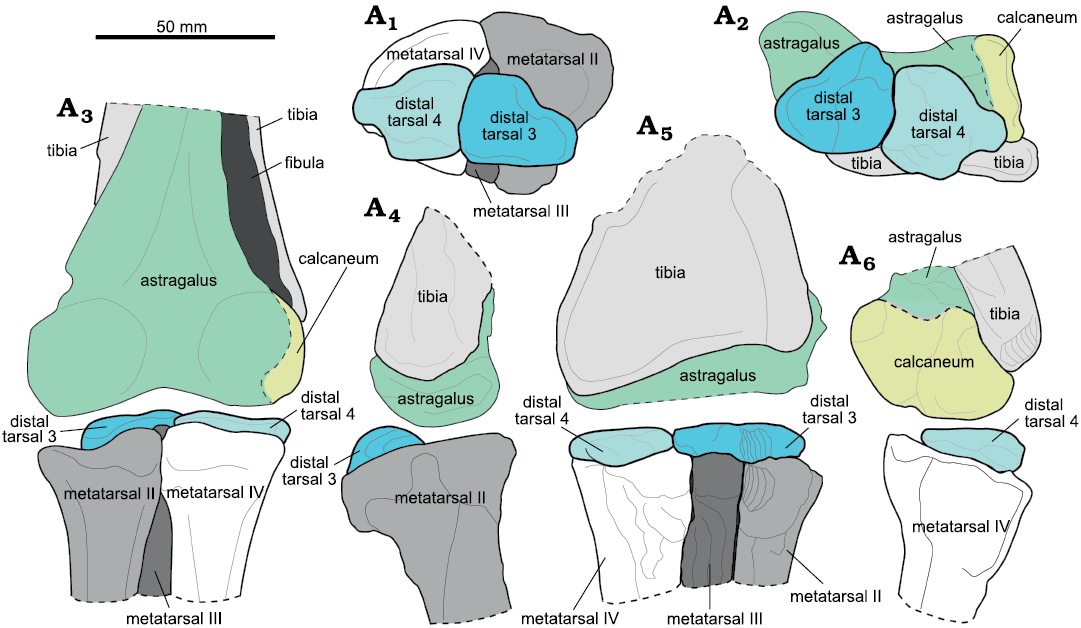
Fig. 2. Reconstruction of left tarsal region of Ornithomimidae indet., RAM 6794 from upper Campanian Kaiparowits Formation, Utah, USA. Proximal view within tarsal joint (A1), distal view within tarsal joint (A2), anterior (A3), medial (A4), posterior (A5), and lateral (A6) views. Artwork by Hannah Caisse (2020).
Remarks.—RAM 6794 can be assigned to Ornithomimidae on the basis of the arctometatarsalian pes (Figs. 1 and 2), in which the proximal end of metatarsal III is constricted and is not visible anteriorly where metatarsals II and IV articulate (Holtz 1995), because this is the sole unambiguous synapomorphy of the family (Kobayashi and Lü 2003). Additionally, the medial expansion of distal tarsal 3 and straight pedal unguals in RAM 6794 also support an assignment to Ornithomimidae. The distal tarsals will be referred to as distal tarsal 3 (dt3) and distal tarsal 4 (dt4) following the naming convention of Ossa-Fuentes et al. (2020). The ornithomimids from the Kaiparowits Formation have not yet been fully described, and their relationship with other taxa in North America at the time is still unexplored (Zanno et al. 2013). Claessens and Loewen (2016) suggested they may be referable to Ornithomimus sp., but this has not yet been tested in detail.
Discussion
Observations of RAM 6794 permit a comprehensive interpretation of distal tarsal anatomy in ornithomimids, clarifying and providing context to some previously reported features. Based on the anatomy observed in RAM 6794 and the morphology reported for Sinornithomimus dongi (Kobayashi and Lü 2003), Archaeornithomimus asiaticus (Gilmore 1933; Russell 1972; Smith and Galton 1990), Gallimimus bullatus (Osmólska et al. 1972), Harpymimus okladnikovi (Barsbold and Perle 1984; Kobayashi and Barsbold 2005b), Garudimimus brevipes (Barsbold 1981; Kobayashi and Barsbold 2005a), and Struthiomimus altus (Lambe 1902; Osborn 1916; Russell 1972), the distal tarsals of ornithomimosaurs can be generalized as: (i) paired, representing distal tarsals 3 and 4; (ii) not fused to each other or to the proximal metatarsals; and (iii) proximo-distally compressed. The intact association of the distal tarsals with the metatarsals in RAM 6794 makes it an important specimen for comparison with both ornithomimosaurs and other theropods to better understand the gross anatomy and function of the theropod ankle.
Anatomical comparisons.—Tarsal morphology and position vary across Ornithomimosauria in: (i) shape of the posterior surface of distal tarsal 3; (ii) antero-posterior position of the distal tarsals relative to the proximal ends of the metatarsals; (iii) extent of distal tarsal coverage of the proximal metatarsal surfaces; and (iv) presence of a lateral flange on distal tarsal 4 (Fig. 3). In many cases, the distal tarsal 3 of ornithomimosaurs is smaller than the distal tarsal 4 (Kobayashi and Lü 2003) and covers the posterior proximal surfaces of metatarsals II and III; distal tarsal 4 is primarily in contact with the proximal surface of metatarsal IV, and distal tarsal 4 is thinner than distal tarsal 3 (Smith and Galton 1990). The distal tarsals also share the general characteristic of being convex proximally and concave distally to articulate with the astragalus and calcaneum and the metatarsus, respectively (Smith and Galton 1990, as reported for Archaeornithomimus asiaticus). The transverse ridge on the distal surface of left distal tarsal 3 observed in RAM 6794 (Fig. 1A2) has also been described in Archaeornithomimus asiaticus (Gilmore 1933; Smith and Galton 1990). Only an incompletely preserved distal tarsal 3 with a unique pair of concavities on the posterior surface is known from Aepyornithomimus tugrikinensis (Chinzorig et al. 2017: fig. 2g, h). This same posterior surface is concave in Harpymimus okladnikovi (Kobayashi and Barsbold 2005b), almost straight to convex in Garudimimus brevipes, and convex in Gallimimus bullatus, Archaeornithomimus asiaticus, Struthiomimus altus, RAM 6794, and the Bissekty ornithomimid (Sues and Averianov 2016: fig. 20F, G; Chinzorig et al. 2017; Fig. 3).
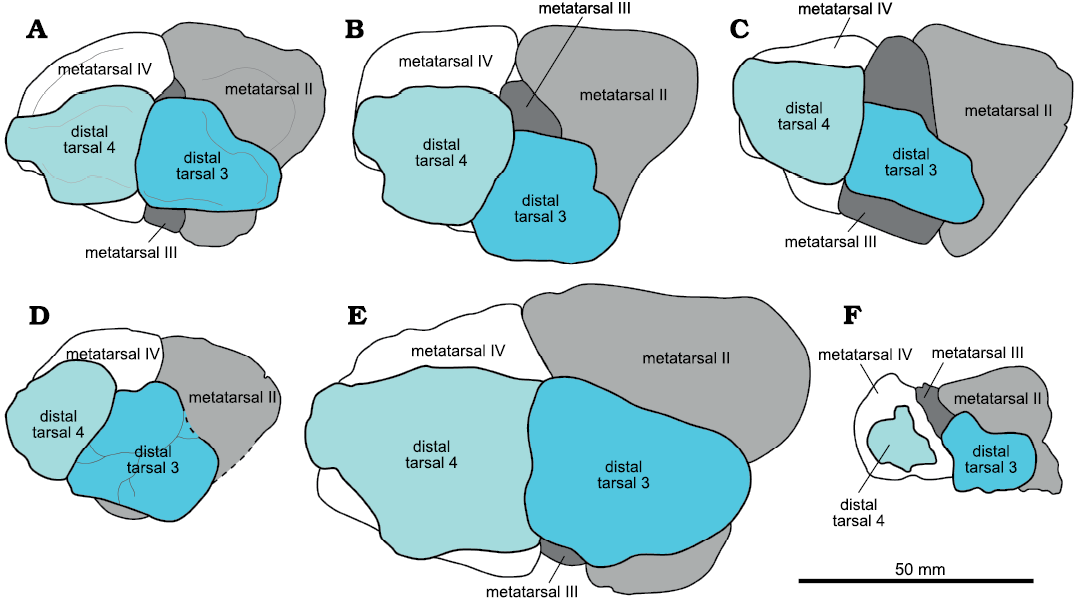
Fig. 3. Comparisons of ornithomimosaur metatarsals with distal tarsals 3 and 4 in proximal view. All are left elements as drawn here, with anterior to the top and lateral to the left of the image, respectively. A. Ornithomimidae indet., RAM 6794. B. Archaeornithomimus asiaticus Russell, 1972, AMNH 6565, redrawn (rotated/reversed) from Smith and Galton (1990). C. Garudimimus brevipes Barsbold, 1981, GIN 100/13, from Kobayashi and Barsbold (2005b). D. Struthiomimus altus Lambe, 1902, CMN 930 from Brad McFeeters personal photos. E. Gallimimus bullatus Osmólska, Roniewicz, and Barsbold, 1972, ZPAL MgD-I/8, redrawn (reversed) from Osmólska et al. (1972). F. Harpymimus okladnikovi Barsbold and Perle, 1984, IGM 100/29, redrawn (rotated) and relabeled from Kobayashi and Barsbold (2005a). Artwork by Hannah Caisse (2021).
The position of the distal tarsals atop the metatarsus is similar in RAM 6794 and Gallimimus bullatus (ZPAL MgD-I/8, Osmólska et al. 1972: fig. 16A1–A3), in that the very posterior edge of all three metatarsals remains visible in proximal view (Figs. 2A1, 3). However, the two differ in that the distal tarsal 3 of RAM 6794 does not overhang the medial edge, leaving more of the anterior proximal surface of metatarsal II visible, yet not as much as in Harpymimus okladnikovi (Barsbold 1981; Kobayashi and Barsbold 2005a: fig. 6.10D, E; Fig. 3). It is unclear if these differences in the antero-posterior positioning could be influenced by some degree of taphonomic displacement, because there would presumably be a number of tendons that act to tightly bind the distal tarsals within the ankle region. Alternatively, it may represent differences in ossification patterns relative to the original cartilaginous precursors.
RAM 6794, Gallimimus bullatus (ZPAL MgD-I/8), Archaeornithomimus asiaticus, and some Struthiomimus altus (AMNH 5339 and UCZM VP 1980.1; REN personal observation; this does not occur in CMN 930 shown in Fig. 3) share a distal tarsal 4 with a narrow, anterolaterally projecting flange on the lateral surface of the tarsal (Figs. 1A2, 2A1, 3). This flange is absent in Garudimimus brevipes (Fig. 3), which differs from these three in that it is a non-arctometatarsalian ornithomimosaur. Harpymimus okladnikovi is also non-arctometatarsalian, but the state of the flange cannot be evaluated in this and the remaining ornithomimosaur taxa because distal tarsal 4 is either incomplete or not preserved. In RAM 6794, Gallimimus bullatus, and Struthiomimus altus, distal tarsal 3 is oriented such that the posterior proximal surface of metatarsal III is visible, whereas the anterior proximal surface of metatarsal III is exposed in Archaeornithomimus asiaticus (Smith and Galton 1990: fig. 4H), Sinornithomimus dongi (Kobayashi and Lü 2003: fig. 23C), Garudimimus brevipes (Kobayashi and Barsbold 2005b: fig 15F), and Harpymimus okladnikovi (Fig. 3). In Archaeornithomimus asiaticus, Sinornithomimus dongi, and Harpymimus okladnikovi, the distal tarsals are situated across the posterior surface of the proximal metatarsus such that one or both distal tarsals overhang posterior metatarsal edges. Ostrom (1969) proposed that in the mesotarsal joint the exposed concave proximal metatarsus in combination with the gently convex anterior portion of the proximal distal tarsals acts as the weight-bearing surface, with the final phases of flexion during each stride provided by the remaining strongly convex posterior aspect of the proximal distal tarsals. Given this relationship, the variation in distal tarsal orientations relative to the metatarsus throughout Ornithomimosauria could be indicative of taxon-specific differences in the degree of typical tarsus flexion and extension.
Marsh (1890) originally depicted the Ornithomimus velox holotype, YPM 542, with a series of three round bones atop the metatarsals that were interpreted as the distal tarsals at the time. Yet, the distal tarsals were largely missing in the specimen when it was redescribed (Claessens and Loewen 2016). Two small fragments of bone remain atop the metatarsus of Ornithomimus velox, situated on the posterior proximal surfaces of metatarsals II and IV and leaving metatarsal III completely uncovered (REN personal observation). Thus, it is difficult to ascertain the distal tarsal morphology. The distal tarsals of Rativates evadens (McFeeters et al. 2016) and MNA Pl. 1762A (Decourten and Russell 1985) are also described as being poorly preserved. In MNA Pl. 1762A, a partial ornithomimid specimen from the Kaiparowits Formation in southern Utah, two fragmentary bones cover the posterior portion of the proximal surfaces of both metatarsals II and III (REN personal observation). In medial view, the bone fragment atop the proximal surface of metatarsal II resembles that of distal tarsal 3 in RAM 6794 and Ornithomimus sp. (TMP 1995.110.1). In all these examples above, only the posterior portions of the distal tarsals are preserved as fragments, which could be a result of more ligaments in this region associated with the tight capping of the metatarsals.
The only ornithomimosaur specimen with published distal tarsal measurements for comparison with RAM 6794 is Archaeornithomimus asiaticus, AMNH 6565 (dt3 length = 33.9 mm, dt3 min. width = 29.4, dt4 length = 32.6, dt4 min. width = 28.5) (Smith and Galton 1990; see Table 1 for comparative measurements of RAM 6794). AMNH 6565 has a length of 259 mm for metatarsal IV, only slightly larger (110%) than the equivalent in RAM 6794 (236 mm). Based on maximum dimensions of the distal tarsals (115% for dt3; 110% for dt4) as well as the reconstructions presented by Smith and Galton (1990: fig. 4H), the tarsals in AMNH 6565 are proportionately the same size as in RAM 6794.
Overall, RAM 6794 is significant in showing unambiguous positioning of distal tarsals versus the metatarsals from multiple angles, and in highlighting similarities and differences between specimens and species as outlined above.
Table 1. Measurements (in mm) of tarsals in RAM 6794. Abbreviations: L, left; R, right.
| |
Maximum medio-lateral width |
Maximum dorso-ventral width |
Maximum proximo-distal thickness |
|
Astragalus+calcaneum (L/R) |
59.3/64.4 |
– |
– |
|
Distal tarsal 3 (L/R) |
29.4/28.4 |
22.7/27.8 |
7.9/7.9 |
|
Distal tarsal 4 (L/R) |
29.7/25.5 |
27.7/24.6 |
–/10.4 |
|
Metatarsal II, proximal articular surface (L/R) |
28.8/30.0 |
44.5/48.1 |
– |
|
Metatarsal III, proximal articular surface (L/R) |
11.3/12.7 |
29.2/29.1 |
– |
|
Metatarsal IV, proximal articular surface (L/R) |
29.0/32.5 |
37.9/37.1 |
– |
Distal tarsals in other theropods.—RAM 6794 is similar in tarsal count to conditions observed in most tyrannosaurids, caenagnathids, oviraptorids, and dromaeosaurids (Currie et al. 2016), although there are differences in aspects of overall shape and degree of fusion. The two distal tarsals of tyrannosaurids are subequal in size and disk-like, exhibiting a narrow and elongate morphology and positioned along the posterior edge of the proximal metatarsus (Holtz 2004). This differs from the more oblong shapes seen in RAM 6794. Brochu (2003) noted that the topography of the distal surface of the distal tarsals varies throughout Tyrannosauridae, suggesting that these differences are created as the distal tarsals ossify and grow into the crevices atop and between the metatarsals. Within Caenagnathidae, the distal tarsals are unfused in caenagnathines and fused to each other and the proximal metatarsus in elmisaurines (Currie et al. 2016; Funston et al. 2016). Elmisaurus rarus and Leptorhynchos elegans uniquely possess a posterodorsal hook on distal tarsal 4 among theropods, which projects vertically to create an ossified arch with metatarsal V (Osmólska 1981; Currie et al. 2016; Funston et al. 2016). By studying a series of Elmisaurus rarus specimens, Currie et al. (2016) found that fusion of the distal tarsals to the proximal metatarsus occurred in a posterior to anterior direction. Distal tarsal fusion in the oviraptorid oviraptorosaur, Oksoko avarsan, is ontogenetically linked, with fusion between distal tarsal 3 and metatarsal III in only the mature individual (Funston et al. 2020). In the Dromaeosauridae, the two flat distal tarsals cap the metatarsals and may exhibit partial fusion to one another and the metatarsus in Velociraptor (Norell and Makovicky 2004), contrasting with the tightly bound but unfused distal tarsals in Deinonychus antirrophus (Ostrom 1969). The shape of distal tarsal 3 is comparable in dromaeosaurs and RAM 6794, but distal tarsal 4 has a more rounded overall shape in dromaeosaurs, with a concave posterolateral corner and no lateral flange. The distal tarsals are unfused with the metatarsals in RAM 6794, and no fusion is noted in any published descriptions for other ornithomimosaurs, although given the variability in fusion in other clades, fusion within ornithomimosaurs and other clades is conceivably possible.
A recent embryological study of extant birds and crocodilians has found that developing avian theropod (bird) ankles consisted of three embryonic distal tarsal cartilages referred to as dt2, dt3, and dt4 that would ossify to become the distal tarsal bones (Ossa-Fuentes et al. 2020). The loss of an ossification centre in the distal tarsal 2 embryonic cartilage would explain the decrease from three to two distal tarsal bones observed in Archosauria. This was accompanied by increasing distal tarsal fusion in maniraptorans (Norell and Makovicky 1997, 1999; Rauhut et al. 2018), with complete fusion of the distal tarsals and metatarsals to form the tarsometatarsus in Avialae (Baumel and Witmer 1993; Ossa-Fuentes et al. 2020). In Maniraptoriformes (which includes Ornithomimosaurs such as RAM 6794), distal tarsal 3 contacts both metatarsals II and III, and this is hypothesized to be the result of the fusion between the ossification centre of the distal tarsal 3 embryonic cartilage and the distal tarsal 2 embryonic cartilage to form a broader distal tarsal 3 element (Ossa-Fuentes et al. 2020: fig. S2). The unambiguously articulated distal tarsal complex preserved in RAM 6794 allows clear confirmation of the tarsal/metatarsal contacts, and thus the specimen is an important comparative model of ornithomimid distal tarsal morphology that can be used to understand the anatomical changes in the ankle that led up to the dinosaur-bird transition.
Across Ornithomimosauria, the greatest variation in the distal tarsals appears to be the degree to which they cover the proximal metatarsus and how they are oriented anteroposteriorly on this surface. One important point of consideration is that this variation could lie in evolutionary divergence between these taxa (in part correlated with temporal separation), as Harpymimus okladnikovi and Garudimimus brevipes belong to Deinocheiridae, and Archaeornithomimus asiaticus, Sinornithomimus dongi, Gallimimus bullatus, and Struthiomimus altus (plus RAM 6794 based on the arctometatarsalian pes) belong to the clade Ornithomimidae (Serrano-Brañas et al. 2020). The explanation for this variation remains unclear and requires further data, but could be related to body size, potential range of flexion and extension at the ankle joint, differences in patterns of ossification, the relationship to the soft tissue components of the ankle (e.g., meniscus, cartilage, ligaments, and tendons), or even post-mortem displacement. These ideas can be explored in greater detail with additional comparative material in the future, allowing better documentation of the variation that exists in distal tarsal morphology.
If tarsal positions are compared with and used to infer embryological patterns, it is critical that future descriptions of this region explicitly state if bones are observed in articulation or if their position is inferred. The unique exposure of the articulated and complete distal tarsals in RAM 6794 makes it the first ornithomimid specimen from North America for which the distal tarsals can be described in detail; however, the paucity of intact ornithomimosaur distal tarsal material limits comparisons. As more specimens become available it will be possible to investigate ontogenetic changes, functional and biomechanical implications of morphological differences, and the broader evolutionary developmental context of the distal tarsals in theropod ankle evolution.
Conclusions
RAM 6794 is the first North American ornithomimid specimen that not only preserves both distal tarsals completely and in articulation, but also allows for the description of the proximal and distal surfaces in the same articulated individual. This specimen highlights some of the variation across species and specimens, while also supporting the conclusion that both North American and Asian ornithomimosaurs have two distal tarsal bones designated distal tarsal 3 and distal tarsal 4, which remain unfused to the metatarsals. New insights into the mechanisms of embryonic distal tarsal ossification emphasize the importance of specimens, like RAM 6794, that preserve the unaltered distal tarsals in articulation, in order to understand developmental and osteological changes leading up to and across the dinosaur-bird transition.
Acknowledgements
We thank the following persons for their assistance during collections visits: Carl Mehling and Mark Norell (AMNH); Margaret Currie, Jordan Mallon, and Kieran Shepherd (all CMN); Tom Courtenay, Heather Feeney, Rhian Russell, Rebecca Sanchez, and Brandon Strilisky (all TMP); Arminty Clarke and Warren Fitch (both University of Calgary Zoological Museum, Calgary, Canada); and Dan Brinkman (YPM). We thank Hannah Caisse (RAM) for detailed illustrations of RAM 6794 and other ornithomimosaur taxa, Janet Gillette (MNA) for photographs of MNA Pl. 1762A, Brad McFeeters (Carleton University, Ottawa, Canada) for photographs of CMN 930, Bailey Jorgensen and Gabriel-Philip Santos (both RAM) for collection and conservation assistance at RAM, and Jared Heuck (RAM) for additional preparation on the specimen. The manuscript was improved by insightful comments from Rebecca Hunt-Foster (Dinosaur National Monument, Jensen, USA), an anonymous reviewer, and the editor Danny Barta (Oklahoma State University, Tahlequah, USA). Fieldwork to collect RAM 6794 was carried out under Paleontological Resources Use Permit UT-S-98-005, issued through the Utah State Office of the Bureau of Land Management, United States Department of the Interior. The discovery was made by Don Lofgren (RAM), with fieldwork assistance from The Webb Schools students, faculty, and volunteers, and initial preparation by Duncan Everhart (Alum Bank, Pennsylvania, USA). This project was funded in part through financial support from the Augustyn Family.
References
Barsbold, R. 1976. On the evolution and systematics of the late Mesozoic dinosaurs [in Russian with English summary]. Sovmestnaâ Sovetsko-Mongol’skaâ Paleontologičeskaâ Ekspediciâ Trudy 3: 68–75.
Barsbold, R. 1981. Toothless carnivorous dinosaurs of Mongolia [in Russian]. Joint Soviet-Mongolian Palaeontological Expedition Transactions 15: 28–39.
Barsbold, R. and Perle, A. 1984. On first new find of a primitive ornithomimosaur from the Cretaceous of the MPR [in Russian]. Paleontologičeskij žurnal 1984 (2): 121–123.
Baumel, J.J. and Witmer, L.M. 1993. Osteologia. In: J.J. Baumel, A.S. King, J.E. Breazile, H.E. Evans, and J.C. Vanden Berge (eds.), Handbook of Avian Anatomy: Nomina Anatomica Avium, 45–132. Nuttal Ornithological Club, Cambridge University Press, Cambridge.
Brochu, C.A. 2003. Osteology of Tyrannosaurus rex: Insights from a nearly complete skeleton and high-resolution computed tomographic analysis of the skull. Journal of Vertebrate Paleontology 22 (Supplement 4): 1–138. Crossref
Chinzorig, T., Kobayashi, Y., Tsogtbaatar, K., Currie, P.J., Watabe, M., and Barsbold, R. 2017. First ornithomimid (Theropoda, Ornithomimosauria) from the Upper Cretaceous Djadokhta Formation of Tögrögiin Shiree, Mongolia. Scientific Reports 7: 1–14. Crossref
Choiniere, J.N., Forster, C.A., and de Klerk, W.J. 2012. New information on Nqwebasaurus thwazi, a coelurosaurian theropod from the Early Cretaceous Kirkwood Formation in South Africa. Journal of African Earth Sciences 71–72: 1–17. Crossref
Claessens, L.P. and Loewen, M.A. 2016. A redescription of Ornithomimus velox Marsh, 1890 (Dinosauria, Theropoda). Journal of Vertebrate Paleontology 36 (1): e1034593. Crossref
Currie, P.J. and Zhao, X.-J. 1993. A new carnosaur (Dinosauria, Theropoda) from the Jurassic of Xinjiang, People’s Republic of China. Canadian Journal of Earth Sciences 30: 2037–2081. Crossref
Currie, P., Funston, G., and Osmólska, H. 2016. New specimens of the crested theropod dinosaur Elmisaurus rarus from Mongolia. Acta Palaeontologica Polonica 61: 143–157. Crossref
De Klerk, W.J., Forster, C.A., Sampson, S.D., Chinsamy, A., and Ross, C.F. 2000. A new coelurosaurian dinosaur from the Early Cretaceous of South Africa. Journal of Vertebrate Paleontology 20: 324–332. Crossref
Decourten, F.L. and Russell, D.A. 1985. A specimen of Ornithomimus velox (Theropoda, Ornithomimidae) from the terminal Cretaceous Kaiparowits Formation of southern Utah. Journal of Paleontology 59: 1091–1099.
Funston, G.F. and Currie, P.J. 2021. New material of Chirostenotes pergracilis (Theropoda, Oviraptorosauria) from the Campanian Dinosaur Park Formation of Alberta, Canada. Historical Biology 33: 1671–1685. Crossref
Funston, G.F., Chinzorig, T., Tsogtbaatar, K., Kobayashi, Y., Sullivan, C., and Currie, P.J. 2020. A new two-fingered dinosaur sheds light on the radiation of Oviraptorosauria. Royal Society Open Science 7 (10): 201184. Crossref
Funston, G., Currie, P., and Burns, M. 2016. New elmisaurine specimens from North America and their relationship to the Mongolian Elmisaurus rarus. Acta Palaeontologica Polonica 61: 159–173. Crossref
Gilmore, C.W. 1933. On the dinosaurian fauna of the Iren Dabasu Formation. Bulletin of the American Museum of Natural History 47 (2): 23–78.
Holtz, T.R. 1995. The arctometatarsalian pes, an unusual structure of the metatarsus of Cretaceous Theropoda (Dinosauria: Saurischia). Journal of Vertebrate Paleontology 14: 480–519. Crossref
Holtz, T.R. 1996. Phylogenetic taxonomy of the Coelurosauria (Dinosauria: Theropoda). Journal of Paleontology 70: 536–538. Crossref
Holtz, T.R. 2004. Tyrannosauroidea. In: D.B. Weishampel, P. Dodson, and H. Osmólska (eds.), Dinosauria, 227–275. University of California Press, Berkeley.
Kobayashi, Y. and Barsbold, R. 2005a. Anatomy of Harpymimus okladnikovi Barsbold and Perle 1984 (Dinosauria; Theropoda) of Mongolia. In: K. Carpenter (ed.), The Carnivorous Dinosaurs, 97–126. Indiana University Press, Bloomington.
Kobayashi, Y. and Barsbold, R. 2005b. Reexamination of a primitive ornithomimosaur, Garudimimus brevipes Barsbold, 1981 (Dinosauria: Theropoda), from the Late Cretaceous of Mongolia. Canadian Journal of Earth Sciences 42: 1501–1521. Crossref
Kobayashi, Y. and Lü, J.-C. 2003. A new ornithomimid dinosaur with gregarious habits from the Late Cretaceous of China. Acta Palaeontologica Polonica 48: 235–259.
Lambe, L.M. 1902. New genera and species from the Belly River Series (mid-Cretaceous). Contributions to Canadian Palaeontology 3 (2): 25–81.
Langer, M.C. 2003. The pelvic and hind limb anatomy of the stem-sauropodomorph Saturnalia tupiniquim (Late Triassic, Brazil). PaleoBios 23 (2): 1–30.
Marsh, O.C. 1881. Classification of the Dinosauria. American Journal of Science (series 3) 23: 81–86. Crossref
Marsh, O.C. 1890. Description of new dinosaurian reptiles. American Journal of Science 229: 81–86. Crossref
Martin, L.D., Stewart, J.D., and Whetstone, K.N. 1980. The origin of birds: structure of the tarsus and teeth. The Auk 97: 86–93.
McFeeters, B., Ryan, M.J., Schröder-Adams, C., and Cullen, T.M. 2016. A new ornithomimid theropod from the Dinosaur Park Formation of Alberta, Canada. Journal of Vertebrate Paleontology 36 (6): e1221415. Crossref
Norell, M.A. and Makovicky, P.J. 1997. Important features of the dromaeosaur skeleton: information from a new specimen. American Museum Novitates 3215: 28.
Norell, M.A. and Makovicky, P.J. 1999. Important features of the dromaeosaurid skeleton: information from newly collected specimens of Velociraptor mongoliensis. American Museum Novitates 3282: 48.
Norell, M.A. and Makovicky, P.J. 2004. Dromaeosauridae. In: D.B. Weishampel, P. Dodson, and H. Osmólska (eds.), Dinosauria, 386–409. University of California Press, Berkeley. Crossref
Osborn, H.F. 1916. Skeletal adaptations of Ornitholestes, Struthiomimus, Tyrannosaurus. Bulletin of the American Museum of Natural History 35: 733–771.
Osmólska, H. 1981. Coossified tarsometatarsi in theropod dinosaurs and their bearing on the problem of bird origins. Palaeontologia Polonica 42: 79–95.
Osmólska, H., Roniewicz, E., and Barsbold, R. 1972. A new dinosaur, Gallimimus bullatus n. gen., n. sp. (Ornithomimidae) from the Upper Cretaceous of Mongolia. Palaeontologia Polonica 27: 103–143.
Ossa-Fuentes, L., Soto-Acuña, S., Bona, P., Sallaberry, M., and Vargas, A.O. 2020. Developmental evolution of the distal ankle in the dinosaur-bird transition. Journal of Experimental Zoology Part B: Molecular and Developmental Evolution [published online, https://doi.org/10.1002/jez.b. 23022]. Crossref
Ostrom, J.H. 1969. Osteology of Deinonychus antirrhopus, an unusual theropod from the Lower Cretaceous of Montana. Bulletin of the Peabody Museum of Natural History 30: 1–165.
Ostrom, J.H. 1976. Archaeopteryx and the origin of birds. Biological Journal of the Linnean Society 8: 91–182. Crossref
Owen, R. 1842. Report on British fossil reptiles. Part II. Report of the British Association for the Advancement of Science for 1841: 60–204.
Padian, K. 2017. Structure and evolution of the ankle bones in pterosaurs and other ornithodirans. Journal of Vertebrate Paleontology 37 (5): e1364651. Crossref
Rauhut, O.W.M., Foth, C., and Tischlinger, H. 2018. The oldest Archaeopteryx (Theropoda: Avialae): a new specimen from the Kimmeridgian/Tithonian boundary of Schamhaupten, Bavaria. PeerJ 6: e4191. Crossref
Russell, D.A. 1972. Ostrich dinosaurs from the Late Cretaceous of western Canada. Canadian Journal of Earth Sciences 9: 375–402. Crossref
Serrano-Brañas, C.I., Espinosa-Chávez, B., Maccracken, S.A., Gutiérrez-Blando, C., de León-Dávila, C., and Ventura, J.F. 2020. Paraxenisaurus normalensis, a large deinocheirid ornithomimosaur from the Cerro del Pueblo Formation (Upper Cretaceous), Coahuila, Mexico. Journal of South American Earth Sciences 101: 102610. Crossref
Smith, D. and Galton, P. 1990. Osteology of Archaeornithomimus asiaticus (Upper Cretaceous, Iren Dabasu Formation, People’s Republic of China). Journal of Vertebrate Paleontology 10: 255–265. Crossref
Sternberg, C.M. 1933. A new Ornithomimus with complete abdominal cuirass. Canadian Field Naturalist 57: 79–83.
Sues, H.-D. and Averianov, A. 2016. Ornithomimidae (Dinosauria: Theropoda) from the Bissekty Formation (Upper Cretaceous: Turonian) of Uzbekistan. Cretaceous Research 57: 90–110. Crossref
Von Huene, F. 1914. Das natürliche System der Saurischia. Centralblatt für Mineralogie, Geologie und Paläontologie 1914: 154–158.
Zanno, L.E., Loewen, M.A., Farke, A.A., Kim, G.-S., Claessens, L.P.A.M., and McGarrity, C.T. 2013. Late Cretaceous theropod dinosaurs of Southern Utah. In: A.L. Titus and M.A. Loewen (eds.), At the Top of Grand Staircase: The Late Cretaceous of Southern Utah, 504–525. Indiana University Press, Bloomington.
Zelenitsky, D.K., Therrien, F., Erickson, G.M., DeBuhr, C.L., Kobayashi, Y., Eberth, D.A., and Hadfield, F. 2012. Feathered non-avian dinosaurs from North America provide insight into wing origins. Science 338: 510–514. Crossref
Acta Palaeontol. Pol. 66 (4): 789–796, 2021
https://doi.org/10.4202/app.00884.2021