
An additional specimen of owenettid procolophonoid from the Middle Triassic of Southern Brazil
RODRIGO T. MÜLLER
Müller, R.T. 2021. An additional specimen of owenettid procolophonoid from the Middle Triassic of Southern Brazil. Acta Palaeontologica Polonica 66 (4): 827–836.
Owenettids are small procolophonoids that survived beyond the Permo-Triassic boundary. The fossil record of owenettids range from the upper Permian to the Middle Triassic. These records come from South Africa, Madagascar, Tanzania, Germany, and Brazil. Despite its conservative anatomy, some species depict some peculiar traits, such as Candelaria barbouri. This owenettid comes from Middle Triassic of Brazil and is characterized by the presence of temporal openings, a highly unusual trait in procolophonoids. Here, a new specimen of C. barbouri is described. The specimen comes from the “Posto” site, an iconic fossiliferous locality from southern Brazil. The material is assigned to C. barbouri based on the presence of temporal openings. In addition, its owenettid affinities are supported by two phylogenetic analyses. The new specimen is the first parareptile from the “Posto” site, providing further data for biostratigraphic hypotheses. Finally, a morphological disparity analysis reveals no overlapped areas between the morphospace occupied by procolophonids and owenettids, reinforcing the existence of these two distinct groups. Furthermore, the morphospace area occupied by owenettids is far reduced in comparison to that of procolophonids, reflecting the conservative anatomy of owenettids.
Key words: Parareptilia, Procolophonia, Owenettidae, Candelaria barbouri, phylogeny, Ladinian, Santa Maria Formation, South America.
Rodrigo T. Müller [rodrigotmuller@hotmail.com], Centro de Apoio à Pesquisa Paleontológica da Quarta Colônia, Universidade Federal de Santa Maria, São João do Polêsine, RS 598, 97230-000, Brazil. Programa de Pós-Graduação em Biodiversidade Animal, Universidade Federal de Santa Maria, 97105-120, Santa Maria, Rio Grande do Sul, Brazil
Received 18 May 2021, accepted 12 July 2021, available online 8 December 2021.
Copyright © 2021 R.T. Müller. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Owenettidae (Broom 1939) is a low diversity group of small procolophonoids that range from the upper Permian (Broom 1939; Piveteau 1955) to the Middle Triassic (Cisneros et al. 2004; Martinelli et al. 2016a). The oldest owenettids are quarried from upper Permian strata of South Africa and Madagascar (Broom 1939; Piveteau 1955; Reisz and Scott 2002). Immediately after the Permo-Triassic extinction event, the group is recorded from Lower Triassic strata of South Africa and Madagascar (Reisz and Scott 2002; Modesto et al. 2003; Ketchum and Barrett 2004). Finally, Middle Triassic owenettids are recorded from Tanzania, Germany, and Brazil (Price 1947; Cisneros et al. 2004; Da-Rosa et al. 2004; Tsuji et al. 2013; Martinelli et al. 2016a).
The fossil record of owenettids from Brazil is limited to some poorly preserved specimens of Candelaria barbouri (Price 1947; Cisneros et al. 2004), one of the youngest and largest owenettid. The specimens were excavated from two distinct fossiliferous localities of southern Brazil (Da-Rosa et al. 2004), which are considered Ladinian to ?early Carnian in age (Martinelli et al. 2017; Schultz et al. 2020). In the present study, a putative new specimen of C. barbouri from the “Posto” site is described. This site is one of the most iconic fossiliferous localities of southern Brazil, yielding exceptionally preserved specimens of the giant top tier predator Prestosuchus chiniquensis (Roberto-da-Silva et al. 2020), as well as monotypic accumulations of several individuals in close association (França et al. 2011; Ugalde et al. 2020). Moreover, its faunal content precedes the dawn of the dinosaur era, providing evidences from one of the last Mesozoic terrestrial ecosystems not ruled by dinosaurs. The new specimen is the first parareptile from the “Posto” site, providing further data for biostratigraphic hypotheses. In addition, the new specimen reinforces some peculiar traits of C. barbouri (e.g., presence of temporal openings).
Institutional abbreviations.—BMNH, Natural History Museum, London, UK; BP, Evolutionary Studies Institute (formerly Bernard Price Institute for Palaeontological Research), University of the Witwatersrand, Johannesburg, South Africa; BRSUG, University of Bristol, Geological Collection, UK; CAPPA/UFSM, Centro de Apoio à Pesquisa Paleontológica da Quarta Colônia/Universidade Federal de Santa Maria, São João do Polêsine, Rio Grande do Sul, Brazil; CAMZM; Museum of Zoology, Cambridge University, Cambridge, UK; DGM, Departamento Nacional de Produção Mineral, Rio de Janeiro, Brazil; IVPP, Institute for Vertebrate Palaeontology and Palaeoanthropology, Beijing, China; NM, National Museum, Bloemfontein, South Africa; NMT, National Museum of Tanzania, Dar es Salaam, Tanzania; PIN, Palaeontological Institute, Russian Academy of Sciences, Moscow, Russia; RC, Rubidge Collection, Camdeboo Municipality, Eastern Cape, South Africa; SAM, Iziko South African Museum, Cape Town, South Africa; UFSM, Laboratório de Estratigrafia e Paleobiologia, Universidade Federal de Santa Maria, Brazil; UMZC, Museum of Zoology, Cambridge University, Cambridge, UK; YPM, Yale Peabody Museum, New Haven, USA.
Other abbrevaitions.—MPTs, most parsimonious trees; OTU, operational taxonomic unit.
Geological setting
The new specimen comes from the “Posto” (or “Posto de Gasolina”) site. This site is located in the municipality of Dona Francisca (29°37′35.60″ S; 53°22′2.86″ W), Rio Grande do Sul, Brazil (Fig. 1). Site strata belong to the Pinheiros-Chiniquá Sequence (Horn et al. 2014), which is part of the Santa Maria Suspersequence of the Paraná Basin (Zerfass et al. 2003). The new specimen was excavated from the basalmost layers of the fossiliferous site, which outcrops in ravines eroded in reddish mudstone. The presence of the cynodont Dinodontosaurus places the site in the Dinodontosaurus Assemblage Zone (Ugalde et al. 2020), which is Ladinian to early Carnian in age (Marsicano et al. 2016; Ezcurra et al. 2017; Phillip et al. 2018; Schultz et al. 2020). In addition to Dinodontosaurus, the site yielded the cynodonts Massetognathus ochagaviae Barberena, 1981, and Protheriodon estudianti Bonaparte, Soares, and Schultz, 2006, and the archosaurs Prestosuchus chiniquensis Huene, 1938, and Decuriasuchus quartacolonia França, Ferigolo, and Langer, 2011 (Bonaparte et al. 2006; França et al. 2011; Mastrantonio et al. 2013; Pavanatto et al. 2016; Martinelli et al. 2017; Roberto-da-Silva et al. 2020). The fossiliferous content varies from isolated bones to articulated and associated skeletons (e.g., França et al. 2011; Ugalde et al. 2020). A nearby outcrop named “Bortolin” site yielded the dicynodont Dinodontosaurus, as well as rauisuchian archosaurs (cf. Decuriasuchus), cynodonts (Chiniquodon and Luangwa), and a mawsoniid coelacanth (Pavanatto et al. 2020). This outcrop area is about to 1 km distant from the “Posto” site.
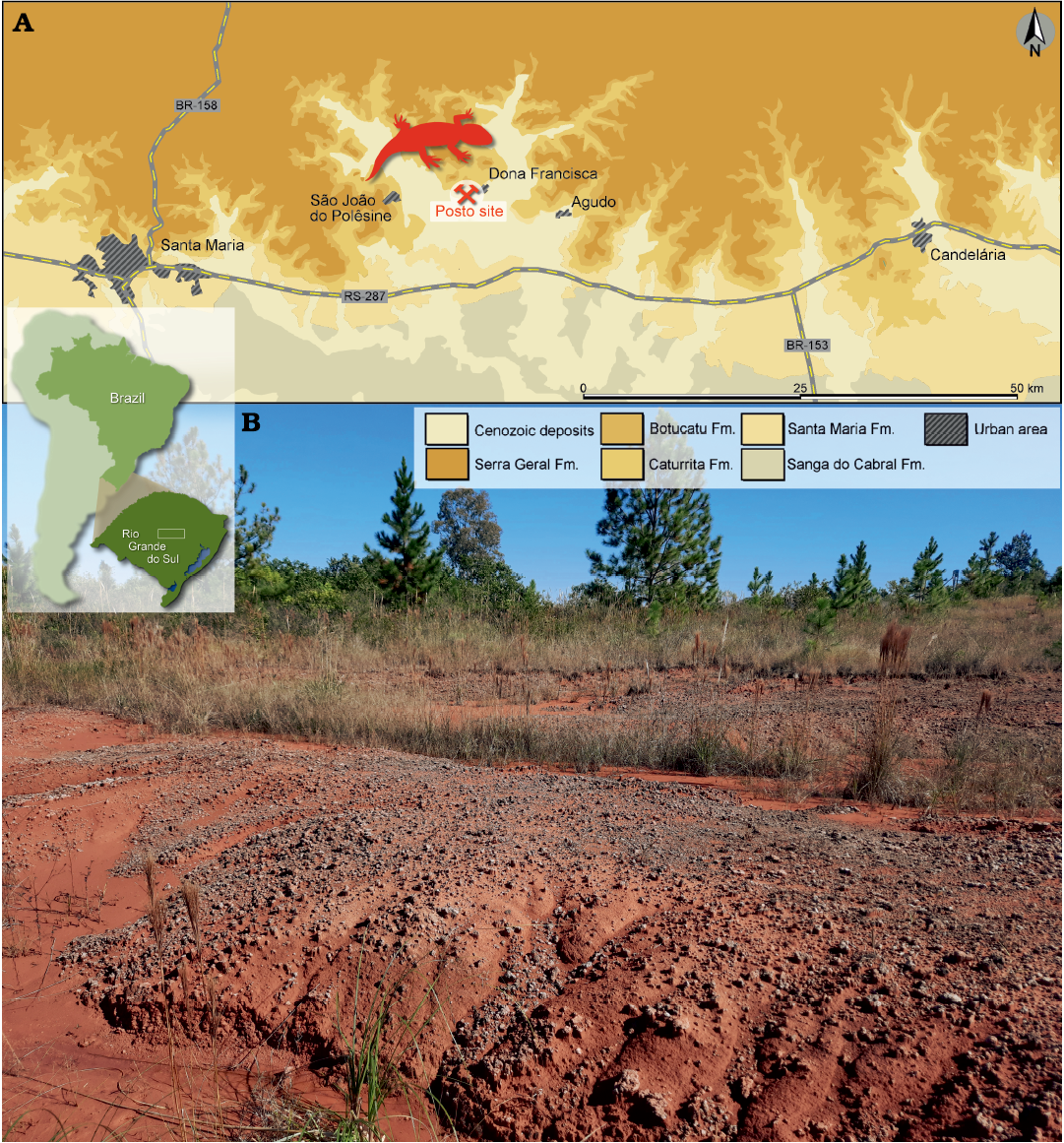
Fig. 1. A. Map of the “Posto” site and the surface distribution of the geologic units in the area (modified from Müller et al. 2020). Location of the study area within South America (inset). B. General view of the “Posto” site.
Material and methods
Material.—The specimen (Fig. 2) here described is housed at the Centro de Apoio à Pesquisa Paleontológica da Quarta Colônia/Universidade Federal de Santa Maria (CAPPA/UFSM), under the code CAPPA/UFSM 0225. It was submitted to mechanical preparation with the aid of air scribe tools, scalpels, and needles. The specimen was stabilized with Paraloid B-72 (Koob 1986).
Phylogenetic analyses.—The phylogenetic affinities of CAPPA/UFSM 0225 were investigated employing two distinct data matrices. In the first analysis, CAPPA/UFSM 0225 was scored in the data matrix of Tsuji et al. (2013), which is a modified version of the data matrices of Modesto et al. (2003) and Cisneros et al. (2004). Following the taxonomic proposal of Hamley et al. (2020), “Owenetta” kitchingorum and Saurodektes rogersorum were merged in a single operational taxonomic unit (OTU) named Saurodektes kitchingorum. The final data matrix includes 21 characters and 11 OTUs. The analysis was conducted in the software TNT v. 1.1 (Goloboff et al. 2008) following the parameters of Tsuji et al. (2013): using an exhaustive search, where all characters were weighted equally and the character 21 was treated as ordered (i.e., additive).
In the second analysis, CAPPA/UFSM 0225 was scored in the data matrix of Pinheiro et al. (2021), which combines owenettids in a single OTU named “Owenettidae”. The aim of this experimental approach is to test if the specimen nests with this OTU. Following Pinheiro et al. (2021), Phaanthosaurus was set as inactive. The final data matrix includes 59 characters and 28 active OTUs. The analysis was performed in the software TNT v. 1.1 (Goloboff et al. 2008) and employed the same parameters of Pinheiro et al. (2021), hence, the most parsimonious trees were recovered with a “Traditional search” (random addition sequence + tree bisection reconnection) with 100 replicates of Wagner trees (with random seed = 0), and using tree bisection re-connection and branch swapping (holding 100 trees save per replicate). All characters were treated as having equal weight, and the following characters were treated as ordered: 1, 8, 18, 26, 31, and 32.
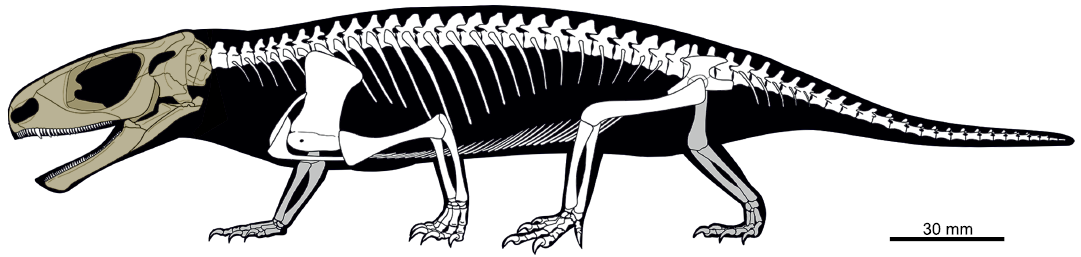
Fig. 2. Reconstructed skeleton of Candelaria barbouri depicting the preserved elements (in brown) of CAPPA/UFSM 0225 (by Maurício S. Garcia).
Decay indices (Bremer support values) and bootstrap values (1000 replicates) were obtained with TNT v. 1.1 (Goloboff et al. 2008). The strict consensus trees were generated using all trees recovered in the analysis and all OTUs. The updated data matrix of full Tsuji et al. (2013) and the full coding to CAPPA/UFSM in the second phylogenetic analysis (employing the data matrix of Pinheiro et al. 2021) are available as Supplementary Online Material.
Cluster analysis.—In order to test putative implications of the new specimen on previous biostratigraphic frameworks, the Jaccard Similarity Coefficient was used to construct a dendrogram with an updated version (incorporating the new specimen) of the dataset of Martinelli et al. (2017). The dataset of Martinelli et al. (2017) includes 11 fossiliferous localities assigned to the Dinodontosaurus Assemblage Zone, as well as 19 taxa. The analysis was carried using the software PAST (Hammer et al. 2001).
Morphospace occupation analysis.—The morphological disparity of owenettids was investigated employing the procedures of Wills (1998). First, a dataset of cranial characters was constructed (see SOM, Supplementary Online Material available at http://app.pan.pl/SOM/app66-Mueller_SOM.pdf). The dataset combines data from the phylogenetic matrices (i.e., Tsuji et al. 2013; Pinheiro et al. 2021) employed in the phylogenetic analyses performed here. The complete list of characters and the dataset are available as Supplementary Online Material. A Euclidean distance matrix (EDMA) was calculated from the dataset using the software MATRIX (Wills 1998). Then, a principal coordinate analysis (PCoA) was performed for the EDMA with the multivariate package GINKGO (Bouxin 2005). The centroid of all operational taxonomic units (OTUs) was taken as the origin of multivariate axes, and the Cailliez method of negative eigenvalue correction was adopted. Finally, a bivariate graph with axes 1 and 2 of the PCoA was constructed using the software PAST (Hammer et al. 2001).
Systematic palaeontology
Parareptilia Olson, 1947
Procolophonoidea Romer, 1956
Owenettidae Broom, 1939
Genus Candelaria Price, 1947
Type species: Candelaria barbouri Price, 1947; Candelária, Brazil, Ladinian–Carnian, Triassic.
Candelaria barbouri Price, 1947
Fig. 3.
Holotype: DGM 314Ra skull and mandible in occlusion (Price 1947).
Type locality: Sanga Pinheiros (29°46′19″ S, 52°44′54″ W), municipality of Candelária, Rio Grande do Sul, Brazil.
Type horizon: Santa Maria Supersequence, Pinheiros-Chiniquá Sequence, Paraná Basin. These strata are Ladinian to early Carnian in age (Marsicano et al. 2016; Ezcurra et al. 2017; Phillip et al. 2018; Schultz et al. 2020).
Material.—UFSM 11076 and UFSM 11131, two complete skulls with mandibles in occlusion and associated postcrania (Cisneros et al. 2004). The specimens were excavated from “Cortado” site (29°44′55″S, 53°00′06″W), municipality of Novo Cabrais, Rio Grande do Sul, Brazil (Da-Rosa et al. 2004). The outcrops pertain to the same horizon as those from which the holotype was recovered (Cisneros et al. 2004). CAPPA/UFSM 0225, an almost complete skull with mandibles in occlusion. Despite its completeness degree, the bone surface is poorly preserved. The specimen was excavated from from “Posto” site (29°37′35.60″S; 53°22′2.86″W), municipality of Dona Francisca, Rio Grande do Sul, Brazil. This site is Ladinian to early Carnian in age.
Emended diagnosis.—According to Cisneros et al. (2004), Candelaria barbouri differs from all other owenettids by its relatively large size (at least 30% larger than all other known owenettid specimens, but see Bradley and Nesbitt 2017) and by a suite of characters related to the presence of a temporal fenestra.
Description.—Whereas the skull and mandibles are almost completely preserved, bone contacts are poorly discernable (Fig. 3). Furthermore, CAPPA/UFSM 0225 lacks the right portion of the snout. The skull is 43 mm in length. The largest known specimen of Candelaria barbouri is 49 mm in length (UFSM 11076; Cisneros et al. 2004). Conversely, other owenettids are smaller in size, for instance, the length of the holotypic skull of Ruhuhuaria reiszi Tsuji, Sobral, and Müller, 2013 (CAMZM M T997) is 25 mm (Tsuji et al. 2013). However, an additional specimen (UMZC T1162) suggests that R. reiszi could have reached 40 to 50 mm in length (Bradley and Nesbitt 2017), resembling the size of Candelaria barbouri. The maximum width of the posterior half of the skull is approximately 30 mm. Therefore, the ratio between length and width of the skull is 1.43. In some procolophonids, such as Procolophon trigoniceps Owen, 1876, and Kapes bentoni Spencer and Storrs, 2002 (Carroll and Lindsay 1985; Zaher et al. 2019), it is less than 1. The height of the skull with mandibles in occlusion is about the same along its length (anterior half approximately 20.5 mm and posterior half approximately 20 mm). Nevertheless, the specimen shows some degree of dorsoventral sedimentary compression. Whereas the orbits are approximately 15 mm in length, the shape and dimensions of the external nares are uncertain.
Distinct from several parareptiles (e.g., Macroleter poezicus Tverdochlebova and Ivachnenko, 1984, PIN 4543/3; Nyctiphruretus acudens Efremov, 1938, PIN 4659/1; Tsuji 2006; Säilä 2010), the specimen lacks ornamentation (i.e., grooves and tubercles) on the dorsal surface of the skull. As typical of procolophonoids, the snout tapers anteriorly and the skull is sub-triangular in dorsal view (Fig. 3A1, A2). The anterior portion of the skull is poorly preserved. Therefore, the number of premaxillary and maxillary teeth, as well as details of the morphology of premaxilla and maxilla are uncertain. The prefrontal forms the anterior and the ventral half of the orbital margin. There is an orbital flange projecting from the anteromedial margin of the orbit (Fig. 3A1, A2), such as in other specimens of C. barbouri (e.g., UFSM 11076, UFSM 11131), where it is formed across the contact between the lacrimal and the prefrontal (Cisneros et al. 2004). In Owenetta rubidgei Broom, 1939 (RC 50) and Saurodektes kitchingorum Reisz and Scott, 2002 (BP/1/4195a; Reisz and Scott 2002) it is less developed. The orbit of CAPPA/UFSM 0225 is roofed anteriorly by the prefrontal and frontal, and posteriorly by postfrontal. The latter is large, resembling other owenettids, whereas in procolophonids it is comparatively reduced. Medially, the orbit is rimmed by a longitudinal orbitotemporal crest (Fig. 3A1, A2). This crest is usually present and well-developed in owenetiids (Cisneros et al. 2004; Tsuji et al. 2013). However, a similar crest occurs in some procolophonids (Kapes bentoni, BRSUG 29950-13; Procolophon trigoniceps, CAPPA/UFSM 0189). The frontals are narrow, resulting in a very narrow interorbital breadth, which resembles the condition of Ruhuhuaria reiszi (CAMZM T997; Tsuji et al. 2013).
The orbit of CAPPA/UFSM 0225 lacks the posterior enlargement typical of procolophonids. On the other hand, the specimen bears a putative temporal opening, such as Candelaria barbouri (Cisneros et al. 2004). The fenestra is located at the dorsal half of the posterior portion of the skull (Fig. 3A5, A6). It is longer (8 mm) than tall (2 mm). The temporal opening is separated from the orbit by the postorbital, which forms the anterior rim of the opening. The postorbital also contributes to the anterior dorsal half of the fenestra, whereas the dorsoventrally short squamosal forms the ventral margin. The lateral margin of the supratemporal bears a notch (Fig. 3A1, A2), which occurs in Saurodektes kitchingorum (BP/1/4195a; BP/1/6025; Reisz and Scott 2002; Modesto et al. 2003) and Owenetta rubidgei (SAM PK K 7582; Reisz and Scott 2002). This notch is considered absent in the other specimens of C. barbouri (Cisneros et al. 2004).
The ventral margin of the posterior portion of the jugal directs dorsally, forming an acute temporal emargination between the contact with the quadratojugal (Fig. 3A5, A6). This contact is reduced and restricted to the posterodorsal corner of the jugal, a typical trait of owenettids (Reisz and Scott 2002; Modesto et al. 2003). The posterior half of the jugal is dorsoventrally expanded, whereas in Saurodektes kitchingorum (BP/1/4195a) it is slender. The anteroventral corner of the quadratojugal projects anteriorly, forming a sigmoid anterior margin. The quadratojugal lacks any horn-like projection, which is usually present in procolophonines (e.g., Procolophon trigoniceps, NM QR3201; Teratophon spinigenis Gow, 1977, SAM PK-K10174; Cisneros 2008) and leptopleuronines (e.g., Hypsognathus fenneri Gilmore, 1928, YPM 55831; Sues et al. 2000).
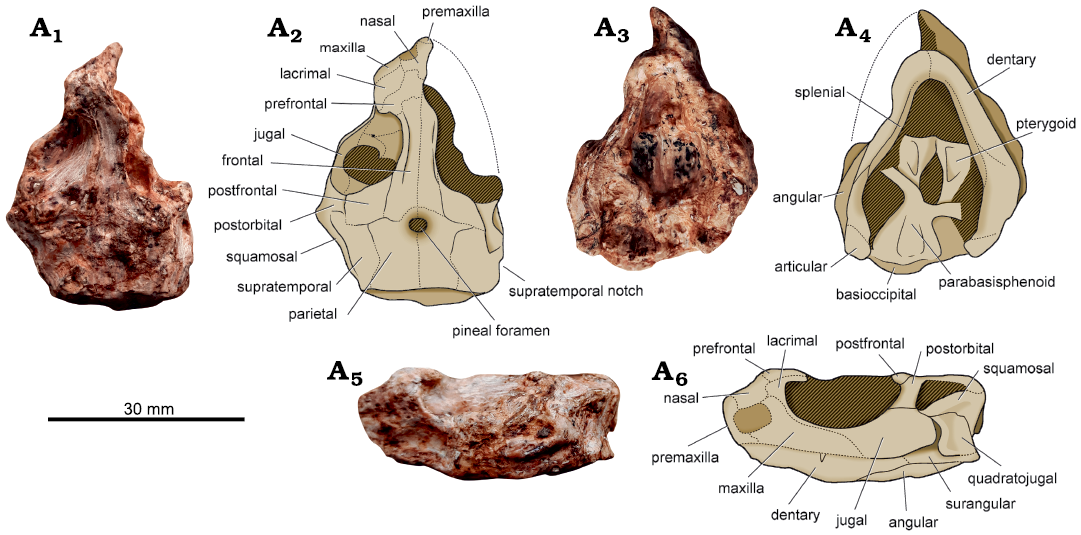
Fig. 3. Owenettid procolophonoid Candelaria barbouri (CAPPA/UFSM 0225) from the “Posto” site, Dona Francisca, Rio Grande do Sul, Brazil; Ladinian, Middle Triassic. Skull and mandible in dorsal (A1, A2), ventral (A3. A4), and left lateral (A5, A6) views. Photographs (A1, A3, A5) and explanatory drawings (A2, A4, A6).
The pineal foramen is small (2 mm in length) and circular in outline. It is located within a depressed surface (Fig. 3A1, A2). Moreover, it rests posterior to posterior margin of the orbit, such as in other owenettids (Cisneros et al. 2004). In contrast, the posterior margin of the orbitotemporal fenestra of procolophonids is anterior to or at the level of the pineal foramen (Cisneros 2008). The posterior margin of the skull is straight to concave. It differs the specimen from procolophonids like Pentaedrusaurus ordosianus Li, 1989 (IVPP V873) and Mandaphon nadra Tsuji, 2017 (NMT RB16; Tsuji 2017), which bear a posteriorly projected margin. The posterolateral corner of the skull is rounded and featureless, whereas in Procolophon trigoniceps (BMNH R1949; Carroll and Lindsay 1985) the corner forms an acute angle with a prominent spine.
The palate and ventral portion of the braincase are partially visible (Fig. 3A3, A4). The suture between the parasphenoid and basioccipital is obscured. There is a shallow depression on the ventral surface of these bones. The interpterygoid vacuity is long and narrow. The cultriform process is not preserved. The pterygoid is long and wide. On the ventral surface, there is a longitudinal fossa medially bounded by a longitudinal ridge and laterally bounded by an oblique ridge. It resembles the morphology of Saurodektes kitchingorum (BP/1/4195a), which bears rudimentary teeth in each ridge. The presence of these teeth in CAPPA/UFSM 0225 is uncertain. The transverse process of the pterygoid is anterolaterally projected. The additional palatal bones are not visible.
The mandible is triangular in ventral view (Fig. 3A3, A4), such as the skull. The posterior portion is far wider than the anterior end. The anterior portion of the mandible shows a gently transverse constriction in ventral view, resulting in a narrow symphyseal region. Conversely, Ruhuhuaria reiszi (CAMZM T997) lacks this constriction, showing a parabolic-shaped symphyseal region in ventral view. In lateral view, the dorsoventral height of each mandibular ramus is about the same along its length. Hence, the dorsal and ventral margins of the dentary run parallel. The ventral margin of the anterior end of the dentary is slightly ventrally expanded. The splenial forms the medial surface of the anterior half of the lower jaw. However, its contacts are poorly preserved. The posterior tip of the lower jaw deflects ventrally, whereas in Owenetta rubidgei (SAM PK K 7582) it is straight. Such as in other owenettids, the craniomandibular articulation occurs approximately at the same dorsoventral level of the dentary tooth line.
Phylogenetic analysis
The first phylogenetic analysis (i.e., employing the data matrix of Tsuji et al. 2013) recovered 13 most parsimonious trees (MPTs) of 33 steps each (consistency index = 0.758; retention index = 0.814). CAPPA/UFSM 0225 nests within Owenettidae in all MPTs (Fig. 4A). The node is supported by four synapomorphies: (i) presence of a notch on the lateral margin of the supratemporal [9 (0→1)]; (ii) humerus without an entepicondylar foramen [14 (0→1)]; (iii) presence of orbitotemporal crests [18 (0→1)]; and (iv) the presence of broad posteriorly expanded nostrils [20 (0→1)]. The characters 14 and 20 are encoded as uncertain (“?”) for CAPPA/UFSM 00225. The inner affinities of Owenettidae are unsolved. The clade is the sister taxa of Procolophonidae, which present the same topology recovered by Cisneros et al. (2004) and Tsuji et al. (2013).
The second phylogenetic analysis (i.e., employing the data matrix of Pinheiro et al. 2021) recovered three MPTs of 134 steps each (consistency index = 0.612; retention index = 0.787). The specimen is positioned as the sister taxon of “Owenettidae” (note: according to Cisneros 2008, characters for the “Owenettidae” have been coded on the basis of Barasaurus besairiei and Owenetta rubidgei) in all MPTs (Fig. 4B), which is supported by the postfrontal contacting frontal, parietal, postorbital and supratemporal [11 (0→1)]. The remaining topology of the strict consensus tree is equivalent to that of Pinheiro et al. (2021).
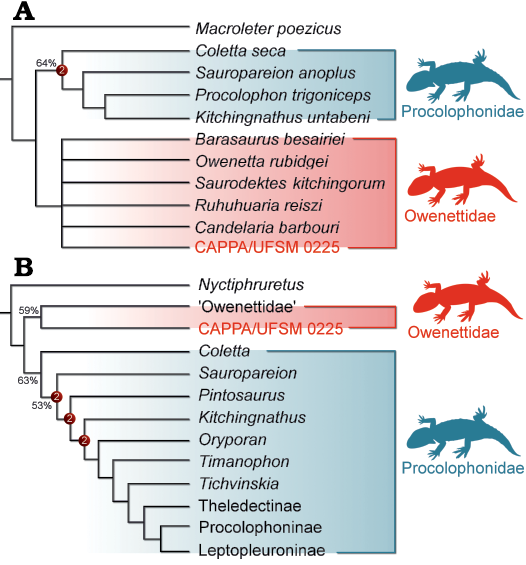
Fig. 4. Results of the phylogenetic analyses. A. Strict consensus tree of the first analysis depicting the phylogenetic position of CAPPA/UFSM 0225. B. Abbreviated strict consensus tree of the second analysis depicting the phylogenetic position of CAPPA/UFSM 0225. Numbers on nodes represent Bremer support values higher than one and Bootstrap values higher than 50%.
Discussion
Taxonomic assignation.—The inner phylogenetic affinities of parareptiles have been intensively disputed (e.g., Laurin and Piñeiro 2017; MacDougall et al. 2018; Ford and Benson 2020; Cisneros et al. 2021). Nevertheless, CAPPA/UFSM 0225 is here regarded as an owenettid procolophonoid based on the following suite of traits: (i) absence of ornamentation on the external surface of the skull, which distinguishes the specimen from non-procolophonoid procolophonians; (ii) very narrow interorbital breadth, which distinguishes the specimen from several procolophonids; (iii) presence of orbitotemporal crests, which occurs in owenettids and Nyctiphruretus (Cisneros et al. 2004); (iv) deep temporal emargination between jugal and quadratojugal, a typical trait of owenettids (Reisz and Scott 2002); (v) enlarged prefrontal that contacts the supratemporal, a synapomorphy of Owenettidae (Cisneros et al. 2004); (vi) pineal foramen set posterior to the posterior margin of the orbit, a condition shared with owenettids and non-procolophonoid procolophonians; and (vii) absence of spine-like structures or bosses on the jugal, quadratojugal and supratemporal, which distinguishes the specimen from several procolophonids. The phylogenetic analyses further support the assignation of CAPPA/UFSM 0225 to Owenettidae. Moreover, the presence of a pair of temporal fenestrae in CAPPA/UFSM 0225 reinforces the owenettid affinity. The owenettid Candelaria barbouri is the only taxon which shares with CAPPA/UFSM 0225 the aforementioned combination of traits plus a pair of temporal fenestrae. In addition, the size of the new specimen resembles that of C. barbouri and no significant differences are observable between CAPPA/UFSM 0225 and the specimens of C. barbouri. One putative difference is the presence of a notch on the lateral margin of the supratemporal in CAPPA/UFSM 0225, which seems absent in C. barbouri (Cisneros et al. 2004). This notch occurs in other owenettids (Reisz and Scott 2002; Modesto et al. 2003). Nevertheless, this portion is poorly preserved in all the specimens (including CAPPA/UFSM 0225), hence, the presence of the notch in CAPPA/UFSM 0225 and/or the absence of the notch in the other specimens is ambiguous. The assignation of CAPPA/UFSM 0225 to C. barbouri receives further support from geological data. The specimens of C. barbouri are recorded from the same geological unit (Pinheiros-Chiniquá Sequence) and Assemblage Zone (Dinodontosaurus Assemblage Zone) which yielded the new specimen. Alternatively, if CAPPA/UFSM 0225 belongs to another procolophonoid species instead of C. barbouri, this implies in a scenario where the group is more diverse than previously thought. In this scenario, the presence of a pair of temporal openings is a putative synapomorphy of CAPPA/UFSM 0225 and C. barbouri instead of an autapomorphy of the latter. This scenario is less plausible and in the present study the first hypothesis is advocated. Therefore, CAPPA/UFSM 0225 is regarded as a new specimen of C. barbouri according the following points: (i) presence of temporal openings, which is interpreted as a local autapomorphy (Cisneros et al. 2004); (ii) shared anatomy and size; (iii) and geological data.
Biostratigraphic significance.—The new specimen improves our knowledge on the faunal composition of the “Posto” site, which is dominated by large tetrapods, such as dicynodonts and archosaurs (França et al. 2011; Ugalde et al. 2020). Actually, small species are represented solely by probainognathian cynodonts (Bonaparte et al. 2006; Martinelli et al. 2016b). Moreover, CAPPA/UFSM 0225 is the first parareptile excavated from the “Posto” site, improving the taxonomic diversity of this locality. The occurrence of C. barbouri in the “Posto” site provides new information regarding the biostratigraphic framework proposed by Martinelli et al. (2017). The authors recognize the putative presence of two subzones within the Dinodontosaurus Assemblage Zone, where the older subzone can be characterized by the presence of the cynodonts Aleodon cromptoni Martinelli, Kammerer, Melo, Paes Neto, Ribeiro, Da-Rosa, Schultz, and Soares, 2017, and Luangwa sudamericana Abdala and Sá-Teixeira, 2004, and the rhynchosaur Brasinorhynchus mariantensis Schultz, Langer, and Montefeltro, 2016. Whereas the “Posto” site and the nearby “Bortolin” site lack records of A. cromptoni and B. mariantensis, the traversodontid L. sudamericana is recorded from the “Bortolin” site (Martinelli et al. 2017; Pavanatto et al. 2020). On the other hand, the “Cortado” and the “Pinheiro” region record A. cromptoni and lack records of L. sudamericana (Martinelli et al. 2017). These latter fossiliferous localities yielded C. barbouri. Therefore, the presence of C. barbouri in the “Posto” site reinforces the faunal similarity of these outcrops. The cluster analysis based on an updated version (incorporating the new record of C. barbouri for the “Posto” site) of the dataset constructed by Martinelli et al. (2017) recovered a dendrogram (Fig. 5) similar to that recovered in the former study. The cophenetic correlation coefficient of the updated version is slightly higher (0.8539) that that of the former analysis (0.8474). The arrangement of the groups follows the results of Marinelli et al. (2017), where the Dona Francisca outcrops (“Posto” plus “Bortolin”) are clustered together with the sites that yielded A. cromptoni. Martinelli et al. (2017) and Pavanatto et al. (2020) point that several records from the Dinodontosaurus Assemblage Zone lack stratigraphic control, which hampers the more accurate biostratigraphic correlations. The new specimen (CAPPA/UFSM 0225) was excavated by the author of the present study (RTM); hence, it is safely part of the fossiliferous content of the “Posto” site and stratigraphic controlled.
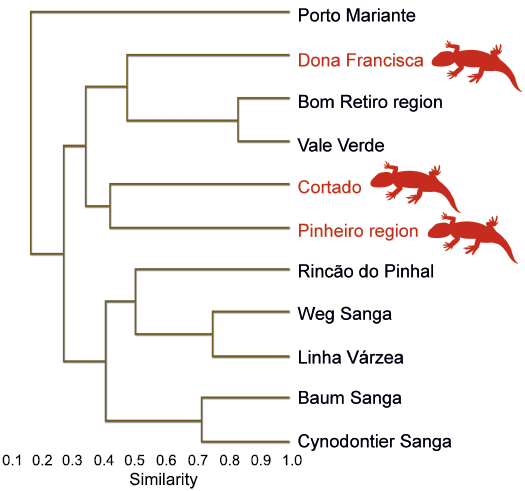
Fig. 5. Dendrogram from the updated dataset of Martinelli et al. (2017) of main localities of the state of Rio Grande do Sul yielding fossil tetrapods of the Dinodontosaurus Assemblage Zone. The silhouette represents the presence of Candelaria barbouri.
Morphological disparity.—Owenettids are far less taxonomic diverse than procolophonids, their sister group (Cisneros 2008; Tsuji et al. 2013; Hamley et al. 2020). Actually, the clade was represented by six species, which were reduced to five according the taxonomic review of Hamley et al. (2020). Conversely, there are dozens of valid species of procolophonids, which present certain morphological disparity, especially regarding dental and horns anatomy (Cisneros and Ruta 2010). As expected, the morphological disparity analysis performed here reveals a morphospace area occupied by owenettids that is far more reduced in comparison to that of procolophonids (Fig. 6). This is not a surprise given the conservative cranial anatomy of the group. Nevertheless, there are some inner specializations, such as the presence of temporal openings in Candelaria barbouri, which is an unusual trait within procolophonoids (Cisneros et al. 2004). The presence of this trait in C. barbouri is corroborated by CAPPA/UFSM 0225. Moreover, Ruhuhuaria reiszi bears peculiar enlarged anterior dentary teeth with labio-lingually expanded bases (Tsuji et al. 2013), suggesting some dietary differentiation within the clade. Despite its conservative anatomy, owenettids survive beyond the Permo-Triassic boundary (Modesto et al. 2003) and shared the terrestrial ecosystems with giant archosaurs (Roberto-da-Silva et al. 2020) and mammalian forerunners (Martinelli et al. 2016; 2017) until the Middle Triassic, when the group was extinct (Cisneros et al. 2004; Martinelli et al. 2016a).
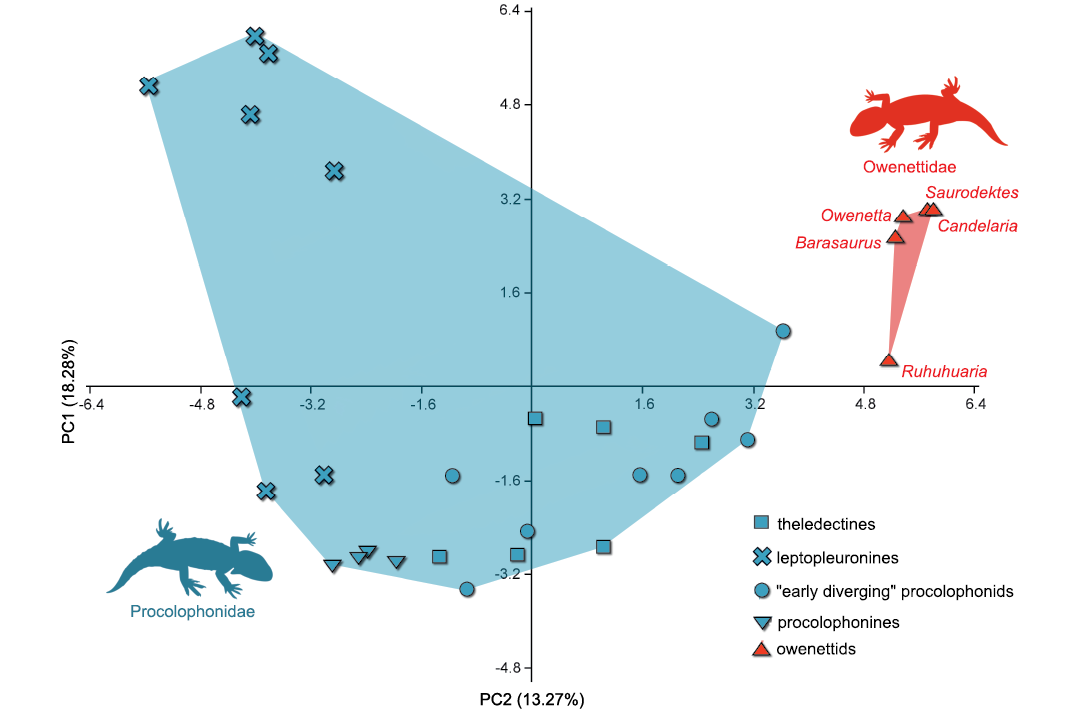
Fig. 6. Bivariate plot showing the results of the morphospace occupation analysis. Red convex hull corresponds to morphospace of owenettids and blue convex hull corresponds to morphospace of procolophonids.
Conclusions
CAPPA/UFSM 0225 is the first owenettid from “Posto” site. This assignation is supported by two phylogenetic analyses. In addition, the unusual presence of temporal openings allows the assignation of CAPPA/UFSM 0225 to Candelaria barbouri. The presence of C. barbouri in the “Posto” site provides further support for biostratigraphic correlation with two other Triassic localities from southern Brazil, reinforcing the framework proposed by Martinelli et al. (2017). Finally, a morphological disparity analysis reveals no overlapped areas between the morphospace occupied by procolophonids and owenettids, reinforcing the existence of these two distinct groups. Furthermore, the morphospace area occupied by owenettids is far reduced in comparison to that of procolophonids, which is not a surprise given the conservative cranial anatomy of the group.
Acknowledgements
I thank Eduardo Silva Neves (UFSM) and an anonymous reviewer for corrections and comments that improved this manuscript. I thank Juan Carlos Cisneros (Universidade Federal do Piauí, Brazil) for providing photographs of some specimens. I am grateful to Matthew Wills (University of Bath, UK) for the software MATRIX. I also thank the Willi Henning Society, for the gratuity of TNT software. This work was carried out with aid of the Fundação de Amparo à Pesquisa do Estado do Rio Grande do Sul (FAPERGS 21/2551-0000680-3) to RTM.
References
Abdala, F. and Sá-Teixeira, A.M.S. 2004. A traversodontid cynodont of African affinity in the South American Triassic. Palaeontologia Africana 40: 11–22.
Barberena, M.C. 1981. Uma nova espécie de Massetognathus (Massetognathus ochagaviae, sp. nov.) da Formação Santa Maria, Triássico do Rio Grande do Sul. Pesquisas 14: 181–195. Crossref
Bonaparte, J.F., Soares, M.B., and Schultz, C.L. 2006. A new non-mammalian cynodont from the Middle Triassic of Southern Brazil and its implications for the ancestry of mammals. New Mexico Museum of Natural History and Science Bulletin 37: 599–607.
Bouxin, G. 2005. Ginkgo, a multivariate analysis package. Journal of Vegetation Science 16: 355–359. Crossref
Bradley, A.B. and Nesbitt, S.J. 2017. A possible new specimen of Ruhuhuaria reiszi from the Manda Beds (? Middle Triassic) of southern Tanzania and its implications for small sauropsids in the Triassic. Journal of Vertebrate Paleontology 37: 88–95. Crossref
Broom, R. 1939. A new type of cotylosaurian, Owenetta rubidgei. Annals of the Transvaal Museum 9: 319–321.
Carroll, R.L. and Lindsay, W. 1985. Cranial anatomy of the primitive reptile Procolophon. Canadian Journal of Earth Sciences 22: 1571–1587. Crossref
Cisneros, J.C. 2008. Phylogenetic relationships of procolophonid parareptiles with remarks on their geological record. Journal of Systematic Palaeontology 6: 345–366. Crossref
Cisneros, J.C. and Ruta, M. 2010. Morphological diversity and biogeography of procolophonids (Amniota: Parareptilia). Journal of Systematic Palaeontology 8: 607–625. Crossref
Cisneros, J.C., Damiani, R., Schultz, C., Da-Rosa, A., Schwanke, C., Neto, L.W., and Aurélio, P.L. 2004. A procolophonoid reptile with temporal fenestration from the Middle Triassic of Brazil. Proceedings of the Royal Society of London. Series B: Biological Sciences 271: 1541–1546. Crossref
Cisneros, J.C., Kammerer, C.F., Angielczyk, K.D., Fröbisch, J., Marsicano, C., Smith, R.M., and Richter, M. 2021. A new reptile from the lower Permian of Brazil (Karutia fortunata gen. et sp. nov.) and the interrelationships of Parareptilia. Journal of Systematic Palaeontology 18: 1939–1959. Crossref
Da-Rosa, A.A.S., Schwanke, C., Cisneros, J.C., Neto, L.W., Aurélio, P.L.P., and Poitevin, M. 2004. Sitio Cortado, uma nova assembléia fossilífera para o Triássico Médio do sul do Brasil. Revista Brasileira de Paleontologia 7: 289–300. Crossref
Efremov, J.A. 1938. Some new Permian reptiles of the USSR. Doklady Akademiâ Nauk SSSR 19: 771–776.
Ezcurra, M.D., Fiorelli, L.E., Martinelli, A.G., Rocher, S.M., von Baczko, B., Ezpeleta, M., Taborda, J.R.A., Hechenleitner, E.M., Trotteyn, M.J., and Desojo, J.B. 2017. Deep faunistic changes preceded the raise of dinosaurs in southwestern Pangaea. Nature Ecology and Evolution 1: 1477–1483. Crossref
Ford, D.P. and Benson, R.B. 2020. The phylogeny of early amniotes and the affinities of Parareptilia and Varanopidae. Nature Ecology & Evolution 4: 57–65. Crossref
França, M.A.G., Ferigolo, J., and Langer, M.C. 2011. Associated skeletons of a new middle Triassic “Rauisuchia” from Brazil. Naturwissenschaften 379: 469–501. Crossref
Gilmore, C.W. 1928. A new fossil reptile from the Triassic of New Jersey. Proceedings of the United States National Museum 73: 1–8. Crossref
Gow, C.E. 1977. New Procolophonids from the Triassic Cynognathus Zone of South Africa. Annals of the South African Museum 72: 109–124.
Goloboff, P.A., Farris, J.S., and Nixon, K.C. 2008. TNT: a free program for phylogenetic analysis. Cladistics 24: 774–786. Crossref
Hamley, T., Cisneros, J.C., and Damiani, R. 2020. A procolophonid reptile from the Lower Triassic of Australia. Zoological Journal of the Linnean Society 192: 554–609. Crossref
Hammer, Ø., Harper, D.A., and Ryan, P.D. 2001. PAST: Paleontological statistics software package for education and data analysis. Palaeontologia Electronica 4: 1–9.
Horn, B.L.D., Melo, T.M., Schultz, C.L., Philipp, R.P., Kloss, H.P., and Goldberg, K. 2014. A new third-order sequence stratigraphic framework applied to the Triassic of the Paraná Basin, Rio Grande do Sul, Brazil, based on structural, stratigraphic and paleontological data. Journal of South American Earth Sciences 55: 123–132. Crossref
Huene, F., von 1938. Die fossilen Reptilien des südamerikanischen Gondwanalandes. Neues Jahrbuch für Mineralogie, Geologie und Paläontologie 1938: 142–151
Ketchum, H.F. and Barrett, P.M. 2004. New reptile material from the Lower Triassic of Madagascar: implications for the Permian Triassic extinction event. Canadian Journal of Earth Sciences 41: 1–8. Crossref
Koob, S.P. 1986. The use of Paraloid B-72 as an adhesive: its application for archaeological ceramics and other materials. Studies in Conservation 31: 7–14. Crossref
Laurin, M. and Piñeiro, G.H. 2017. A reassessment of the taxonomic position of mesosaurs, and a surprising phylogeny of early amniotes. Frontiers in Earth Science 5: 88. Crossref
Li, J.L. 1989. A new genus of Procolophonidae from the Lower Triassic of Shaanxi, China. Vertebrata Palasiatica 10: 248–267.
MacDougall, M.J., Modesto, S.P., Brocklehurst, N., Verrière, A., Reisz, R.R., and Fröbisch, J. 2018. Commentary: a reassessment of the taxonomic position of mesosaurs, and a surprising phylogeny of early amniotes. Frontiers in Earth Science 6: 99. Crossref
Marsicano, C.A., Irmis, R.B., Mancuso, A.C., Mundil, R., and Chemale, F. 2016. The precise temporal calibration of dinosaur origins. Proceedings of the National Academy of Sciences 113: 509–513. Crossref
Martinelli, A.G., Kammerer, C.F., Melo, T.P., Paes Neto, V.D., Ribeiro, A.M., Da-Rosa, Á.A., Schultz, C.L., and Soares, M.B. 2017. The African cynodont Aleodon (Cynodontia, Probainognathia) in the Triassic of southern Brazil and its biostratigraphic significance. PLoS One 12: e0177948. Crossref
Martinelli, A.G., Soares, M.B., and Schoch, R.R. 2016a. Owenettids and procolophonids from the lower Keuper shed new light on the diversity of parareptiles in the German Middle Triassic. Journal of Paleontology 90: 92–101. Crossref
Martinelli, A.G., Soares, M.B., and Schwanke, C. 2016b. Two new cynodonts (Therapsida) from the Middle–Early Late Triassic of Brazil and comments on South American probainognathians. PloS One 11: e0162945. Crossref
Mastrantonio, B.M., Schultz, C.L., Desojo, J.B., and Garcia, J.B. 2013. The braincase of Prestosuchus chiniquensis (Archosauria: Suchia). Geological Society, London, Special Publications 379: 425–440. Crossref
Modesto, S.P., Damiani, R.J., Neveling, J., and Yates, A.M. 2003. A new Triassic owenettid parareptile and the Mother of Mass Extinctions. Journal of Vertebrate Paleontology 23: 715–719. Crossref
Müller, R.T., von Baczko, M.B., Desojo, J.B., and Nesbitt, S.J. 2020. The first ornithosuchid from Brazil and its macroevolutionary and phylogenetic implications for Late Triassic faunas in Gondwana. Acta Palaeontologica Polonica 65: 1–10. Crossref
Olson, E.C. 1947. The family Diadectidae and its bearing on the classification of reptiles. Fieldiana Geology 11: 1–53. Crossref
Owen, R. 1876. Descriptive and Illustrated Catalogue of the Fossil Reptilia of South Africa in the Collection of the British Museum. British Museum, London.
Pavanatto, A.E.B., Da-Rosa, Á.A.S., Müller, R.T., Roberto-da-Silva, L.Ribeiro, A.M., Martinelli, A.G., and Dias-da-Silva, S. 2020. Bortolin site, a new fossiliferous locality in the Triassic (Ladinian/Carnian) of southern Brazil. Revista Brasileira de Paleontologia 23: 123–137. Crossref
Pavanatto, A.E.B., Müller, R.T., Da-Rosa, Á.A.S., and Dias-da-Silva, S. 2016. New information on the postcranial skeleton of Massetognathus ochagaviae Barberena, 1981 (Eucynodontia, Traversodontidae), from the Middle Triassic of Southern Brazil. Historical Biology 28: 978–989. Crossref
Philipp, R.P., Schultz, C.L., Kloss, H.P., Horn, B.L., Soares, M.B., and Basei, M.A. 2018. Middle Triassic SW Gondwana paleogeography and sedimentary dispersal revealed by integration of stratigraphy and U-Pb zircon analysis: the Santa Cruz Sequence, Paraná Basin, Brazil. Journal of South American Earth Sciences 88: 216–237. Crossref
Pinheiro, F.L., Silva‐Neves, E., and Da‐Rosa, Á.A.S. 2021. An early‐diverging procolophonid from the lowermost Triassic of South America and the origins of herbivory in Procolophonoidea. Papers in Palaeontology [published online http://dx.doi.org/10.1002/spp2.1355]. Crossref
Piveteau, J. 1955. Existence d’un reptile du groupe Procolophonides á Madagascar. Conséquences stratigraphiques et paléontologiques. Comptes Rendus hebdomadaires des séances de l’Académie des Sciences 241: 1325–1327.
Price, L.I. 1947. Um procolofonídeo do Triássico do Rio Grande do Sul. Boletim da Divisao de Geologia e Mineralogia 122: 7–27.
Reisz, R.R. and Scott, D. 2002. Owenetta kitchingorum, sp. nov., a small parareptile (Procolophonia: Owenettidae) from the Lower Triassic of South Africa. Journal of Vertebrate Paleontology 22: 244–256. Crossref
Roberto-da-Silva, L., Müller, R.T., França, M.A.G., Cabreira, S.F., and Dias-Da-Silva, S. 2020. An impressive skeleton of the giant top predator Prestosuchus chiniquensis (Pseudosuchia: Loricata) from the Triassic of Southern Brazil, with phylogenetic remarks. Historical Biology 32: 976–995. Crossref
Romer, A.S. 1956. Osteology of the Reptiles. 772 pp. University of Chicago Press, Chicago.
Säilä, L.K. 2010. The phylogenetic position of Nyctiphruretus acudens, a parareptile from the Permian of Russia. Journal of Iberian Geology 36: 123–143. Crossref
Schultz, C.L., Langer, M.C., and Montefeltro, F.C. 2016. A new rhynchosaur from south Brazil (Santa Maria Formation) and rhynchosaur diversity patterns across the Middle–Late Triassic boundary. Paläontologische Zeitschrift 90: 593–609. Crossref
Schultz, C.L., Martinelli, A.G., Soares, M.B., Pinheiro, F.L., Kerber, L., Horn, B.L., Pretto, F.A., Müller, R.T., and Melo, T.P. 2020. Triassic faunal successions of the Paraná Basin, southern Brazil. Journal of South American Earth Sciences 104: 102846. Crossref
Spencer, P.S. and Storrs, G.W. 2002. A Re‐evaluation of small tetrapods from the Middle Triassic Otter Sandstone Formation of Devon, England. Palaeontology 45: 447–467. Crossref
Sues, H.D., Olsen, P.E., Scott, D.M., and Spencer, P.S. 2000. Cranial osteology of Hypsognathus fenneri, a latest Triassic procolophonid reptile from the Newark Supergroup of eastern North America. Journal of Vertebrate Paleontology 20: 275–284. Crossref
Tsuji, L.A. 2006. Cranial anatomy and phylogenetic affinities of the Permian parareptile Macroleter poezicus. Journal of Vertebrate Paleontology 26: 849–865. Crossref
Tsuji, L.A. 2017. Mandaphon nadra, gen. et sp. nov., a new procolophonid from the Manda Beds of Tanzania. Journal of Vertebrate Paleontology 37: 80–87. Crossref
Tsuji, L.A., Sobral, G., and Müller, J. 2013. Ruhuhuaria reiszi, a new procolophonoid reptile from the Triassic Ruhuhu Basin of Tanzania. Comptes Rendus Palevol 12: 487–494. Crossref
Tverdokhlebova, G.I. [Tverdohlebova, G.I.] and Ivachnenko, M.F. [Ivačnenko, M.F.] 1984. Nykteroleteriids from the Upper Permian of Eastern Europe [in Russian]. Paleontologičeskij žournal 1984: 98–111.
Ugalde, G.D., Müller, R.T., de Araújo-Júnior, H.I., Dias-da-Silva, S., and Pinheiro, F.L. 2020. A peculiar bonebed reinforces gregarious behaviour for the Triassic dicynodont Dinodontosaurus. Historical Biology 32: 764–772. Crossref
Wills, M.A. 1998. Cambrian and recent disparity: the picture from priapulids. Paleobiology 24: 177–199.
Zaher, M., Coram, R.A., and Benton, M.J. 2019. The Middle Triassic procolophonid Kapes bentoni: computed tomography of the skull and skeleton. Papers in Palaeontology 5: 111–138. Crossref
Zerfass, H., Lavina, E.L., Schultz, C.L., Garcia, A.J.V., Faccini, U.F., and Chemale Jr, F. 2003. Sequence stratigraphy of continental Triassic strata of Southernmost Brazil: a contribution to Southwestern Gondwana palaeogeography and palaeoclimate. Sedimentary Geology 161: 85–105. Crossref
Acta Palaeontol. Pol. 66 (4): 827–836, 2021
https://doi.org/10.4202/app.00909.2021