
Priabonian, late Eocene chronostratigraphy, depositional environment, and paleosol-trace fossil associations, Pipestone Springs, southwest Montana, USA
DEBRA L. HANNEMAN, DONALD LOFGREN, STEPHEN T. HASIOTIS, and WILLIAM C. MCINTOSH
Hanneman, D.L., Lofgren, D., Hasiotis, S.T., and McIntosh, W.C. 2022. Priabonian, late Eocene chronostratigraphy, depositional environment, and paleosol-trace fossil associations, Pipestone Springs, southwest Montana, USA. Acta Palaeontologica Polonica 67 (1): 5–20.
Sanidine 40Ar/39Ar ages of lapilli tuffs and the mammalian fauna of Pipestone Springs strata provide a high-resolution chronostratigraphy for upper Eocene (Priabonian) rock units in southwestern Montana. Two felsic lapilli tuffs with weighted-mean 40Ar/39Ar single crystal sanidine ages of 37.50±0.02 Ma and 36.00±0.20 Ma both fall within the Priabonian, late Eocene. These tuffs occur within the basal to upper part of the 55 m of exposed Pipestone Springs strata. The uppermost 15 m yield a diverse and abundant assemblage of mostly small-bodied middle Chadronian (Priabonian, late Eocene) mammals. The older lapilli tuff is an ashfall tuff, whereas the younger lapilli tuff exhibits minor aeolian reworking. The new 40Ar/39Ar age constraints significantly increase the age range of Pipestone Springs strata to include uppermost Duchesnean–lowermost Chadronian (Priabonian, upper Eocene) deposits in addition to its well-known middle Chadronian vertebrate assemblage. These new 40Ar/39Ar ages combined with its mammalian fauna further support Pipestone Springs strata as age-correlative to the Flagstaff Rim section in central Wyoming, and provide a basis for better determining late Eocene mammalian paleogeography and regional paleolandscapes in the United States Rocky Mountain to Great Plains areas. Loessites intercalated with paleosols dominate Pipestone Springs deposits. The recognition of loessites comprising these strata is a new depositional interpretation of Pipestone Springs strata, making these loessites some of the oldest known aeolian Eocene strata in the Great Plains–Rocky Mountains region. Pipestone Springs paleosols developed on lapilli tuffs are vertisols. Alfisols and inceptisols, developed from a parent material of volcanic glass mixed with non-volcanic grains, are the remaining paleosols within the loessite strata. Additionally, a new and important discovery in this project is the recognition that all paleosols are extensively bioturbated, containing trace fossils similar to Rebuffoichnus and newly identified trace fossils resembling Feoichnus, Eatonichnus, Fictovichnus, and Coprinisphaera.
Key words: Trace fossils, Coprinisphaera, Eatonichnus, Feoichnus, Fictovichnus, Rebuffoichnus, continental strata, paleosol, vertisol, alfisol, inceptisol, loessite, Chadronian, Priabonian, Renova Formation, Montana.
Debra L. Hanneman [whgeol@gmail.com], Whitehall GeoGroup Inc., 107 Whitetail Road, Whitehall, Montana 59759, USA.
Donald Lofgren [dlofgren@webb.org], Raymond M. Alf Museum of Paleontology, 1175 Base Line Road, Claremont, California 91711, USA.
Stephen T. Hasiotis [hasiotis@ku.edu], Department of Geology, University of Kansas, 1475 Jayhawk Boulevard, Lawrence, Kansas 66045, USA.
William C. McIntosh [william.mcintosh@nmt.edu], New Mexico Bureau of Mines and Mineral Resources, 801 Leroy Place, Socorro, New Mexico 87801, USA.
Received 2 May 2021, accepted 7 September 2021, available online 30 March 2022.
Copyright © 2022 D.L. Hanneman et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Pipestone Springs lies at the northwestern edge of the Jefferson Valley, southwestern Montana (Fig. 1), where Eocene continental strata and pyroclastic units unconformably overlie Upper Cretaceous batholithic rocks. The Pipestone Springs Main Pocket (PSMP), a middle Chadronian (Priabonian, late Eocene) vertebrate locality that is world-renowned for an abundant and diverse assemblage of small bodied mammals, is contained within these basin-margin strata. The PSMP is one of the best studied mammal localities within Cenozoic (Paleogene and Neogene) continental deposits of southwestern Montana (Tabrum et al. 1996, 2001; Tabrum and Metais 2007; Lofgren et al. 2017). As noted by Tabrum et al. (1996: 285), “if the number of type specimens collected from a locality is a measure of its significance, then the Main Pocket at Pipestone Springs is arguably the single most important locality of Chadronian age known.” Extensive collections from PSMP in the early 1900s resulted in the description of many new mammalian taxa (Douglass 1901, 1903, 1905; Matthew 1903). Reports of new records of mammals from the PSMP continued for decades (Tabrum and Fields 1980; Emry and Hunt 1980; Lillegraven et al. 1981; Lillegraven and Tabrum 1983; Ostrander 1983; Garcia 1992; Korth 1994; Emry and Korth 1996). Most recently, Lofgren et al. (2017) provided a review of PSMP carnivores, as well as an analysis of the coprolites commonly recovered from the locality.
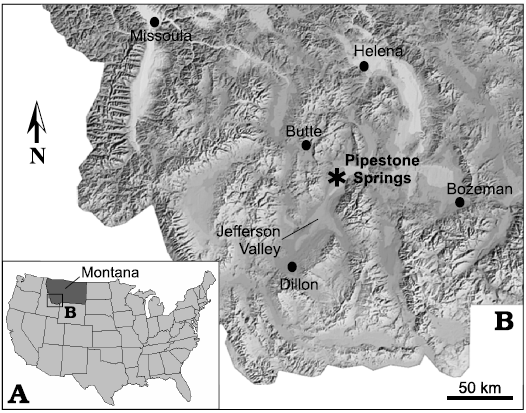
Fig. 1. Study area location maps. A. Location of studied area within Montana (USA). B. Digital relief image of southwest Montana depicting main towns and the Jefferson Valley which contains the study locality of Pipestone Springs (sourced from Montana State Library, Natural Resource Information System, 2001 data bank).
Although vertebrate fossils from Pipestone Springs strata have been extensively studied, other aspects of these upper Eocene strata have not been accorded the same level of inquiry. In this study, we document newly acquired 40Ar/39Ar single crystal sanidine ages for Pipestone Springs strata. These are the first radioisotopic age constraints for Pipestone Springs strata. They also provide a basis for detailed correlation with the central Wyoming Flagstaff Rim section, thus enabling a more regional foundation for understanding Priabonian (late Eocene) mammalian and landscape paleogeography.
We also describe ichnofossils that occur within several paleosol horizons of Pipestone Springs strata and give 40Ar/39Ar age constraints for ichnofossil horizons and for the base of the PSMP. Additionally, we present a new interpretation for the depositional environment of Pipestone Springs rock units containing the ichnofossil horizons and PSMP as loessites, making these strata the oldest documented Eocene aeolian strata in the Great Plains–Rocky Mountain area of the United States.
Institutional abbreviations.—CM, Carnegie Museum of Natural History, Pittsburgh, Pennsylvania, USA; ICAL, Montana State University Image and Chemical Laboratory, Bozeman, Montana, USA; MV, University of Montana vertebrate locality; RAM, Raymond M. Alf Museum of Paleontology, Claremont, California, USA; UMVC, University of Montana Vertebrate Collections, Missoula, Montana USA.
Other abbreviations.—EDS, energy dispersive spectrometry; GPTS, geomagnetic polarity time scale; MSWD, mean squared weighted deviation; PS, Pipestone Springs; PSMP, Pipestone Springs Main Pocket.
Geological setting
General geology.—Priabonian (upper Eocene) clastic continental strata and pyroclastic rocks at Pipestone Springs, Montana (USA), occur on the west flank of the local, informally named Whitehall depositional basin (Hanneman 1989) that is present within the northern part of the Jefferson Valley, and north of the local Waterloo Basin, the main depositional basin in the Jefferson Valley (Fig. 2). Similar to other depositional basins in western Montana’s intermontane valleys, both the Whitehall and the Waterloo depositional basins formed during the Cenozoic, with the bulk of deposition occurring during late Paleocene–Eocene extensional tectonism. Minor middle Miocene and late Miocene–Recent extension has modified these basins (Hanneman 1989; Hanneman and Wideman 1991, 2006, 2013, 2016; Constenius 1996; Constenius et al. 2003; Hanneman et al. 2003; Fan et al. 2017; Schwartz et al. 2019). Cenozoic basin-fill in the Whitehall and Waterloo basins is approximately 760 m to 3000 m thick, respectively; stratal thicknesses contained within other southwestern Montana Cenozoic basins can exceed 4500 m (Fields et al. 1985; Hanneman 1989; Hanneman and Wideman 1991). Continental clastic and carbonate strata as well as minor amounts of volcanic and hydrothermal rocks comprise the fill of these southwestern Montana basins (Fields et al. 1985; Hanneman and Wideman 1991; Hanneman et al. 2003; Barnosky et al. 2007; Lielke et al. 2012; Schwartz and Graham 2017). Similar Cenozoic deposits also occur in a few higher elevation areas in southwestern Montana (Hanneman and Lofgren 2017; Fan et al. 2017; Li et al. 2017; Lofgren et al. 2020).
Age constraints for southwestern Montana Cenozoic strata are based largely on vertebrate assemblages. Douglass (1899, 1901, 1903, 1905) initially interpreted ages for his collection of southwest Montana Cenozoic vertebrates; further documentation of Cenozoic vertebrate assemblages in southwestern Montana provided additional age control for basin deposits (Matthew 1903; Hough 1955, 1958, 1961; Kay et al. 1958; Fields et al. 1985; a comprehensive list of other works is provided by Tabrum et al. 1996, 2001), and limited age control is available for scattered volcanic units interbedded with these strata (Chadwick 1978, 1985; Hanneman and Wideman 1991; Hanneman et al. 2003; Fritz et al. 2007; Kellogg and Harlan 2007; Dudás et al. 2010).
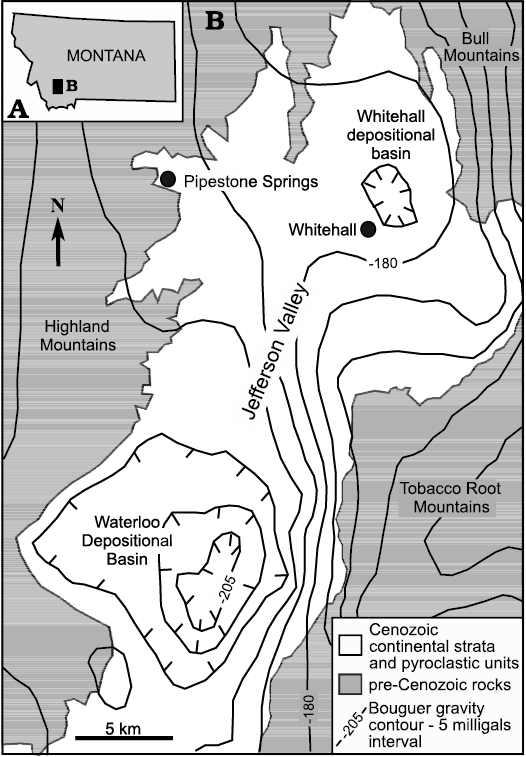
Fig. 2. Bouguer gravity map of the Jefferson Valley and surrounding area, southwest Montana. A. The state of Montana with the black rectangle showing location of the Jefferson River Valley. B. Bouguer gravity map of the Jefferson River Valley showing both the Whitehall and Waterloo depositional basins. Gravity contour interval of 5 milligals (modified from Hanneman 1989; Hanneman and Wideman 1991; data archived in McCafferty et al. 1998). Note that Pipestone Springs lies in a basin-margin position on the northwestern edge of the Jefferson Valley.
Pipestone Springs and PSMP vertebrate locality geology.— Priabonian (upper Eocene) strata of the Pipestone Springs area and PSMP locality (both located in section 29, Township 2 North, Range 5 West) are part of the informal Sequence 2 of Hanneman and Wideman (1991, 2006) and a lithostratigraphic proxy, the Renova Formation, of Kuenzi and Fields (1971). Post-depositional erosion has dissected the Eocene strata in this area to the extent that a deeply entrenched, 100 m wide gully separates two main Eocene stratal accumulations (Fig. 3). Because of this physical separation, Pipestone Springs area strata and their contained vertebrate fossil localities have historically been labeled as two separate stratigraphic sections and named variously as Main Pocket, South Pocket, north section, and south section (Kay et al. 1958; Garcia 1992; Tabrum et al. 1996; Lofgren et al. 2017). In keeping with the stratal separation, vertebrate fossils from the two sections have been curated separately (Tabrum et al. 1996; Lofgren et al. 2017).

Fig. 3. Overview of the Pipestone Springs area, southwest Montana (from Google Earth 7.3.3.7786, July 21, 2020, Pipestone Springs, Montana, 45°53’42.79”N, 112°15’42.00”W, Eye altitude 5264 feet. Digital Globe 2012. http://www.earth.google.com [September 19. 2020]). Pipestone Springs (PS) strata occur in both a south and north area, separated by a 100 m-wide gully. Because of this geographic separation, vertebrate fossils from the two areas have been curated separately. Abbreviations: MV, University of Montana vertebrate locality; CM, Carnegie Museum of Natural History locality; RAM, Raymond M. Alf Museum of Paleontology locality.
Measured sections through the north and south areas of Pipestone Springs are illustrated in Fig. 4. The sections were measured from basal Tertiary strata exposed at either side of the entrenched gully shown in Fig. 3 through the strata’s contained vertebrate fossil localities. The measured sections range from approximately 55 m in thickness for the northern area to 33 m in thickness for the southern area, with section bases lying unconformably on Cretaceous granodiorite and the top of the southern section being truncated by a normal fault. Both sections contain massive silty mudstone units. A lapilli tuff bed occurs at about the 10 m level in the north section and at about 7 m in the south section; a reworked lapilli tuff is found at the 40 m level in the north section and at the 23 m level in the south section. Multiple ichnofossil horizons and associated paleosols occur from approximately 10 m to 42 m in the northern section and from about 7 m to 26 m in the southern section (Fig. 4).
We confirm that the two physically separated sections of Pipestone Springs Priabonian (upper Eocene) strata represent a common set of depositional and non-depositional events, based on stratigraphic section measurements and their detailed description. Because the northern section is the most complete of the two, we designate it as the reference section for Pipestone Springs Priabonian (upper Eocene) strata (Fig. 4). The differences between the two sections can be attributed to basin margin variations in deposition and to fault truncation of the south section.
The majority of vertebrate fossils have been collected from a discrete horizon in Pipestone Springs strata that occurs in the upper part of both sections (Lofgren et al. 2017), and the name of PSMP is herein restricted to that horizon (Fig. 4). The PSMP is, thus, equivalent to several collected localities, including UMVC locality MV 5811, CM locality 3575, and RAM locality V200301 in the northern Pipestone Springs reference section, and UMVC locality MV 5810, CM locality 3576, and RAM locality V200322 in the Pipestone Springs south section (Fig. 3; stratigraphic placement of localities shown in Fig. 4). Fossil mammal vertebrates from the PSMP are middle Chadronian (Priabonian, late Eocene) in age (Tabrum et al. 1996; Tabrum et al. 2001). UMVC MV 5902 occurs below the PSMP (Fig. 4) and yields scant material of a general Chadronian (Priabonian, late Eocene) age.
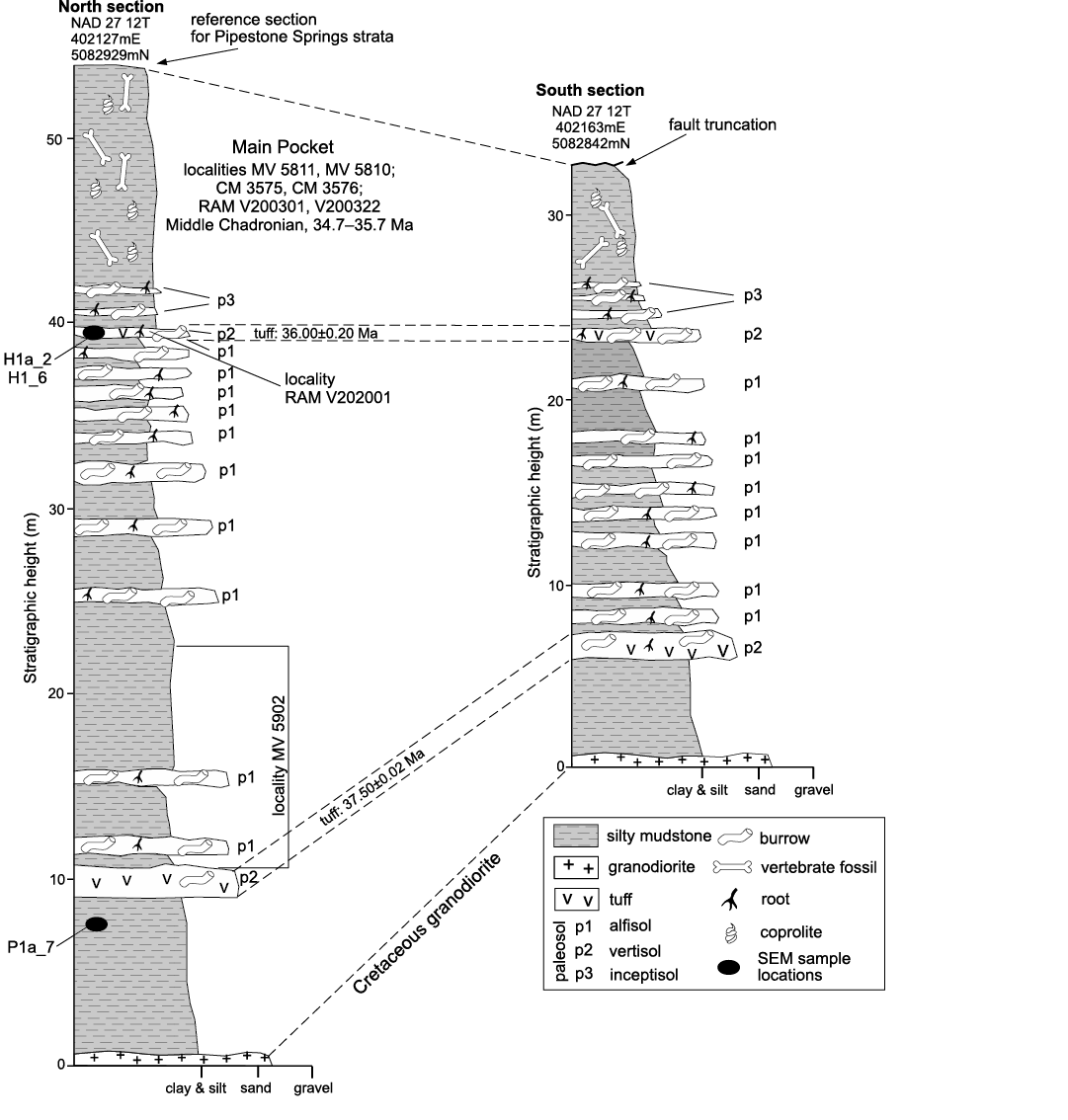
Fig. 4. Pipestone Springs north and south stratigraphic sections showing paleosols and SEM sample locations, with the north section specified as the reference section.
Material and methods
Section measurement was done using a 1.5-m-long Jacob’s staff. Sedimentary units were described using standard features noted by Tucker (2011) that include unit thickness, lithology, sediment texture, color, sedimentary structures, pedogenic features, and fossil and ichnofossil content. Paleosols were categorized using soil taxonomy developed by the Soil Survey Staff (1999). Paleosol profiles were delineated using horizons designated as A (uppermost; zone of eluviation), B (intermediate; zone of illuviation), and C (lowermost; parent material) (Soil Survey Staff 1951; for applications to paleosols e.g., Retallack 2001; Hasiostis et al. 2007; Fischer and Hasiotis 2018). Other pedogenic features of the paleosols were described in the field by noting dry matrix color, soil structure, and ichnofossil and rhizolith presence. Matrix color was determined using a Munsell soil color chart (Munsell Color 1975).
Ichnofossils were collected from all burrow horizons (Fig. 4). The ichnofossils were described according to their architectural and surficial morphologies and internal fill pattern (Hasiotis and Mitchell 1993; Bromley 1996) and assigned to an ichnotaxon based on distinguishing characteristics. Ichnofossils were assigned to one or more categories of burrowing behavior that indicate their spatial position and interpreted moisture zone within the paleosol profile (Hasiotis 2000, 2004, 2007, 2008; Hasiotis et al. 2007). Ichnofossils interpreted as epiterraphilic behavior were constructed on the paleosurface. Ichnofossils interpreted as terraphilic behavior were constructed in the upper vadose zone above the water table near the surface of the soil-water profile where soils are well drained. Ichnofossils interpreted as hygrophilic behavior were constructed above the water table in the upper, intermediate, and lower vadose zone where soil moisture is better retained.
Two lapilli tuff units (Fig. 4) were sampled for 40Ar/39Ar age analysis, with one lapilli tuff at the 7 m level in the south section (sample location NAD 27 12T 402183 m East 5082888 m North) and the other at approximately the 40 m section level in the Pipestone Springs reference section (sample location NAD 27 12T 402185 m East 5082978 m North). Matrix samples were taken from the basal 15 cm of each unit. The sample analyses were done by New Mexico Geochronology Laboratory (Socorro, New Mexico) personnel for single crystal weighted-mean 40Ar/39Ar ages. Minerals were separated by crushing and sieving, followed by etching in 7% HF (sanidine only) or 7% HCl and rinsing in an ultrasonic bath, then concentrated using a Franz magnetic separator and/or density liquids, followed by hand picking. Samples and Fish Canyon Tuff sanidine monitors in machined aluminum trays were irradiated in-vacuo at the Denver USGS Triga reactor. Samples were heated using a defocused CO2 laser for 30 seconds. Reactive gases were removed during analysis by reaction with a SAES NP-10 getter. Sample gas analyses were performed using Thermo Argus VI and/or Helix mass spectrometers on line with automated all-metal extraction system. J-factors were determined by CO2 laser-fusion of 4 to 6 single crystals from multiple radial positions around the irradiation tray. Calculations were performed using Pychron software, v. 18.2. Mean age errors are quoted herein as ±2σ, with all other errors ±1σ. Weighted-mean and error are inverse-variance-weighted mean and error Taylor (1982), with error multiplied by root MSWD where MSWD >1. Decay constants are after Min et al. (2000), λ total = 5.464e – 10/a; isotopic abundances are after Steiger and Jäger (1977). Ages are calculated relative to FC-2 Fish Canyon Tuff sanidine interlaboratory standard at 28.201 Ma after Kuiper et al. (2008).
SEM of sediment samples from Pipestone Springs strata was performed at Montana State University’s ICAL, Bozeman, Montana. A JEOL JSM-6100 SEM, with a magnification range between 90×–3500×, a 20 KeV accelerating voltage, and a working distance of 39 mm was used for this work. Sample grains were washed with distilled water, disaggregated for three minutes using a Crest Ultrasonic cleaner, then dried on a hotplate. Grains were directly mounted onto carbon tape, then tapped lightly to eliminate excessive material and to produce a fine layer on the tape. Sediment grains were sputter-coated with gold in a vacuum evaporator to prevent static build-up. EDS was done on most grains during the project’s SEM work.
Results
Pipestone Springs strata 40Ar/39Ar single crystal sanidine age constraints.—The basal lapilli tuff unit, located at approximately 10 m in the Pipestone Springs reference section and at about 7 m in the south section (Fig. 4), has a weighted-mean 40Ar/39Ar single crystal sanidine age of 37.50±0.02 Ma. The slightly reworked lapilli tuff that occurs at approximately the 40 m level of the Pipestone Springs reference section and at about the 23 m in the south section (Fig. 4), has a weighted-mean 40Ar/39Ar single crystal sanidine age of 36.00±0.20 Ma.
Pipestone Springs strata description and interpretation.—Pipestone Springs strata are comprised primarily of massive silty mudstone units 8 m to 15 m thick, and thinner (0.5 m to 3 m in thickness), structureless beds, with the entirety resting nonconformably on Upper Cretaceous granodiorite of the Boulder Batholith (Fig. 4). The thinner beds are separated from each other by pedogenically altered zones that contain burrows, rhizoliths, and minor pedogenic features. Within the reference section, paleosol-silty mudstone intervals comprise about 60% of the section, occurring from about 10 m to about the 40 m level (Figs. 4, 5A). These strata range from white, light gray, to pale brown/light yellowish brown (10YR 8/1–2, 7/2, and 6/3–4) in color on dry bedrock surfaces. A massive, white (10YR 8/1), 15 m thick silty mudstone unit constitutes the remaining part of the reference section and is the locus of PSMP fossil vertebrate collections (Fig. 5B).
Pipestone Springs strata are generally tabular and maintain a constant thickness for a short distance, but because they occupy a basin-margin position with an underlying surface of moderate relief, overall section thickness changes over even a few hundred meters. With the exception of a weathered zone developed atop the Upper Cretaceous granodiorite, these strata are well consolidated.

Fig. 5. Field and SEM appearance of Pipestone Spring section strata and sediments. A. Pipestone Springs reference section’s lower strata, including the 37.50±0.02 Ma basal lapilli tuff and overlying paleosol horizons. The basal lapilli tuff is 1.6 m thick for scale. B. A massive, white, 15 m-thick silty mudstone unit constitutes the upper-most part of the Pipestone Springs reference section and the Pipestone Springs south section. This massive unit is the locus of PSMP fossil vertebrate collections, including the localities MV 5811, CM 3575, and RAM V200301 occurring in the Pipestone Springs reference section, and locality MV 5810, CM 3576, and RAM V200322 located in the Pipestone Springs south section. Students, with an average height, are surface collecting fossil vertebrates from the PSMP strata. C. Backscattered SEM image showing fresh to altered glass and detrital grains that display grain morphology ranging between rounded and angular. Sample location is H1a_2 as shown in the stratigraphic column of Fig. 4, sampled from the Pipestone Springs reference section’s 36.00±0.20 Ma reworked tuff. The coexistence of these grains and mixed grain morphology indicates sediment mixing. D. SEM micrograph of glass shard heavily coated by smectite. Many grain samples from Pipestone Springs strata are heavily coated by smectite which is the dominant clay mineral in these rock units. Sample location is H1_6, shown in the stratigraphic column of Fig. 4, sampled from the Pipestone Springs reference section’s 36.00±0.20 Ma reworked tuff.
As noted previously, two ashfall lapilli tuffs are present in these deposits. One tuff occurs at about the 10 m level in the Pipestone Springs reference section and at about 7 m in the nearby southern exposure (Fig. 4). A younger tuff exhibits minor reworking and crops out at the 40 m level in the Pipestone Springs reference section and at about 23 m in the nearby southern exposure (Fig. 4).
An average clay:silt:sand ratio for Pipestone Springs strata is 47:46:7 (Kuenzi 1966), with silt- to sand-sized angular to subangular grains typically floating within the mudstone matrix. The coarser-grain fraction (coarse silt to medium sand), in order of abundance, is predominantly comprised of volcanic glass, quartz, plagioclase, potassium feldspar, and granitic fragments (Kuenzi 1966; Hanneman 1989). In some samples, fresh to altered glass and detrital grains coexist, with overall grain morphology displaying a range between rounded and angular, indicating sediment mixing (Fig. 4: sample H1a_2; Fig. 5C) as in the sections’ 36.00±0.20 Ma upper lapilli tuff unit (Fig. 4). Many grain samples show extensive smectite overgrowths (Fig. 4: sample H1_6; Fig. 5D), with smectite being the dominant clay mineral in Pipestone Springs strata (Kuenzi 1966; Hanneman 1989). That smectite is the dominant clay mineral in these strata is also indicated by the popcorn texture developed on many weathered surfaces.
The depositional environment of Pipestone Springs strata has historically been interpreted as a fluvial-floodplain environment, with minor debris-flow and ashfall deposits (Kuenzi 1966; Kuenzi and Fields 1971; Axelrod 1984; Garcia 1992). These strata, however, exhibit no fluvial or debris-flow features. Instead, the Pipestone Springs strata represent direct ashfall, slightly reworked ashfall, and loessite deposits. That these strata result from aeolian deposition is supported by their overall fine-grained make-up that is typical of loess (Zarate and Blasi 1991; Rowley and Fan 2016; Raigemborn et al. 2018). This depositional interpretation is further corroborated by stratal sedimentary features that include: (i) massive stratification caused by protracted suspension or fallout, (ii) angular to subangular shape of fresher vitric fragments indicating low abrasion, (iii) mantle bedding for relatively very short distances, and (iv) numerous paleosols interspersed within the section that indicate stable land surfaces formed between aeolian events. These sedimentary features are very similar to those noted by Bellosi (2010) for massive pyroclastic mudstone interpreted as subaerial distal ashfall deposits or tephric loessites of the middle Eocene to Miocene Sarmiento Formation at Gran Barranca, in central Patagonia. Analogous sedimentary deposits occur in loess-paleosol sequences such as those found in the northern European Loess Belt (Lemkuhl et al. 2016) and the China Loess Plateau (Maher 2016).
Pipestone Springs paleosol pedogenesis.—Pedogenic features including horizonation, color change, structure, pedogenic clay occurrence, rhizoliths, and burrows define paleosols that occur at various intervals throughout the majority of Pipestone Springs strata (Fig. 4). Overall, these paleosols are weakly to strongly developed on silty mudstone parent material that is either predominantly volcanic glass (i.e., lapilli tuff) or contains volcanic glass reworked with a significant component of non-volcanic grains. As noted previously, Pipestone Springs strata are overall light in color, with high value and low chroma, thus enhancing pedogenic color change that occurs within the strata’s contained paleosols. Additionally, extensively developed ichnofossil zones are associated with most Pipestone Springs paleosols, and these combine to form indurated ledges within the Pipestone Springs sections (Fig. 6A).
The presence of an A horizon is interpreted from limited granular structure, lack of depositional features, and abundant burrows and rhizoliths present both on indurated surfaces and often within 15–40 cm of the indurated surface, indicating a near-surface location for the uppermost part of a pedogenically-altered zone (Hasiotis 2002, 2007; Fischer and Hasiotis 2018). There is no evidence for the possible erosion of the pre-existing A horizon, such as scours, fluvial deposits, or debris-flow deposits. Rather, features typically associated with a modern A horizon were masked and/or removed by burial compaction by overlying sediments and the oxidation of organic matter as suggested by McDonald and Busacca (1992) for paleosols present in late Quaternary loess of Washington state. Similar trends of organic matter oxidation in well-drained paleosol horizons have been observed in other Mesozoic and early to mid-Cenozoic paleosols (e.g., Bown and Kraus 1987; Kraus 1987; Hasiotis 2004, 2008; Hasiotis et al. 2007; Hembree and Hasiotis 2007, 2008; Fischer and Hasiotis 2018).

Fig. 6. Paleosol features and grain morphology of the Pipestone Springs strata. A. Alfisols occur as rubbly zones that weather to become resistant units from approximately 12–40 m in the Pipestone Springs reference section. These paleosols are composite and are developed upon extensively reworked volcanic glass and non-volcanic grains. A vertisol occurs atop the 36.00±0.20 Ma upper lapilli tuff at Pipestone Springs. Backpack is 50 cm in length for scale. B. The Pipestone Springs basal lapilli tuff, with a 40Ar/39Ar age of 37.50±0.02 Ma, is a direct ashfall tuff and is partially overprinted by a vertisol paleosol. The vertisol contains pseudo-anticlines and micro-highs and lows. Scale for paleosol features is the circled field notebook at 19 cm in length and the shovel at 54 cm in length. C. Alfisols are the most numerous paleosols within Pipestone Springs strata. These paleosols have composite A–Bt1–Bt2–Bt3 profiles. Field notebook is 19 cm in length for scale. D. Alfisol A–Bt horizons are delineated primarily by the occurrence of abundant trace fossils. E. Backscattered electron microscopy image of sample P1a_7, taken from near the base of the Pipestone Springs reference section (sample location shown in the stratigraphic column of Fig. 4), shows volcanic grains (a glass grain indicated by arrow) mixed with detrital non-volcanic components (magnification ×160). A conspicuous rounded and pitted quartz grain is indicated by the white arrow. Overall grain morphology is rounded to subangular, with substantial smectitic clay overgrowth on most grains (indicated by arrow for one grain).
Pipestone Springs vertisols.—Vertisol paleosols comprise some Pipestone Springs paleosols. These paleosols occur atop the 37.50±0.02 Ma basal ashfall lapilli tuff (Fig. 6B) and on the 36.00±0.20 Ma upper, reworked lapilli tuff in both the Pipestone Springs reference section and in the southern Pipestone Springs section (see Fig. 4 for stratigraphic locations). The vertisols have A–Bss–C and A-Bss1–Bss2 profiles. The A horizon has abundant burrows and rhizoliths concentrated within 5–15 cm of the horizon’s surface. An underlying smectite-rich (Bss) horizon, 24–45 cm thick, is characterized by pseudo-anticlines containing angular blocky, wedge-shaped, and small prismatic peds, some exhibiting pressure faces. The pseudo-anticlines form micro-high and micro-low areas in vertical cross section. The pseudo-anticlines typically rest upon a C horizon of lapilli tuff. In other places the Bss horizon is welded on a pre-existing Bss horizon. Scattered burrows and rhizoliths persist into the Bss horizon. Paleosol matrix color darkens slightly from the C horizon to the A–Bss horizons, with Munsell color being white (10YR 8/2) for the lapilli tuff C horizon and light gray to very pale brown (10YR 7/2–7/4) for the A-Bss horizons.
Pipestone Springs alfisols.—Alfisol paleosols are the most numerous pedogenically-altered zones in Pipestone Springs strata (see Fig. 4 for stratigraphic locations of alfisol paleosols). These paleosols have A–Bt1–Bt2–Bt3 and Bt1–Bt2 profiles (Fig. 6C). The A horizon is granular, with abundant burrows and rhizoliths; thickness varies 10–40 cm (Fig. 6D). Rhizoliths from hairline size to 0.5 cm in diameter are common in these horizons; some rhizoliths and burrows extend from the A into the B horizons. Argillic (Bt) horizons range 25–40 cm thick. Scattered burrows and rhizoliths persist into the Bt horizon. The Bt horizons have a blocky, subangular blocky to prismatic structure and colors of 10YR 7/2 (very pale brown) to 10YR 6/3–6/4 (pale brown to light yellowish brown). Alfisol paleosols are developed upon parent material comprised of reworked volcanic glass and non-volcanic grains as typified by sample P1a_7 (stratigraphic location shown in Fig. 4) and shown in the backscattered electron microscopy image of Fig. 6E.
Pipestone Springs inceptisols.—The remaining Pipestone Springs paleosols occur in the uppermost part of the Pipestone Springs section and are best categorized as inceptisols with A–Bw–C profiles developed on parent material composed of reworked volcanic glass and non-volcanic grains (stratigraphic locations shown in Fig. 4). The A horizon ranges 15–40 cm thick and is delineated by granular structure and ichnofossils. Rhizoliths from hairline size to 0.5 cm in diameter are common in these horizons; some rhizoliths and burrows extend from the A into the Bw horizons. The underlying Bw horizon has a granular to massive structure and a darkening of color from the underlying parent-material strata of 10YR 8/1 (white) to 10YR 7/3 (very pale brown). All horizons contain volcanic glass, but they also include some non-volcanic grains.
Pipestone Springs ichnofossil zones.—All paleosols exhibit extensive bioturbation (Fig. 7A), containing an abundance of newly identified ichnofossils likely constructed by dung beetles (Coleoptera) based on comparisons to modern and ancient traces attributed to this group. Close examination shows that the tracemakers built complex reproductive structures using dung that, depending on completeness, larval ontogeny, and preservation, appear as: (i) elongate ellipsoids with meniscate backfills within an irregular ellipsoid structure or with massive fill (Fig. 7B, C), (ii) weakly organized helical patterns of masses constructed from inside-outward by the addition of pelletized sediment pressed together to form meniscate backfill from the base upward (Fig. 7D), (iii) spheres and hemispheres of material that are partly hollowed out (Fig. 7E), and (iv) the addition of material to an initial cavity from outside-inward in a rosette pattern that is subsequently hollowed out into a hemisphere (Fig. 7F). These architectural morphologies contain single or multiple elements found in ichnofossils attributed to dung beetle reproduction structures, such as Feoichnus, Eatonichnus, Rebuffoichnus, Fictovichnus, and Coprinisphaera (e.g., Roselli 1939, 1987; Halffter and Edmonds 1982; Halffter et al. 1985; Howden and Peck 1987; Johnston et al. 1996; Hasiotis 2003; Laza 2006). These preserved forms likely represent a continuum of reproductive structures in various stages of completion by adults and usage by larvae and pupae, and final preservation in these paleosols. The architectural differences encountered in these ichnofossils indicate that extensive future work is necessary to determine their ichnotaxonomic affinities and their taphonomy.

Fig. 7. Ichnofossils in the Pipestone Springs strata. A. Example of ichnofossils in the A horizon. B. Elongate ellipsoidal trace similar in morphology to Rebuffoichnus. C. Cylindrical to elliptical structure, partly walled, with meniscate backfill, similar to dung sausages of modern dung beetle nests. D. Helical constructed structure composed of pelletized sediment pressed together to form a wall with meniscate backfill or massive backfill, similar to Eatonichnus. E. Constructed spheres with an opening to a concave hemispherical impression, similar in morphology to Coprinisphaera and Feoichnus. F. Structures composed of meniscate backfills in a rosette pattern that is subsequently hollowed out into a hemisphere with architectural elements of Coprinisphaera, Eatonichnus, Feoichnus, and Rebuffoichnus.
The occurrence of the ichnofossils and the association of other pedogenic features of the paleosol horizons indicate terraphilic behavior (Hasiotis 2000, 2002, 2003, 2004, 2007, 2008) of the tracemakers. The traces were most likely constructed in the upper vadose zone of the soil-water profile and near the surface, well above the phreatic zone. These combined pedogenic features suggest that the paleosols had abundant groundcover, were well rooted, and well drained.
Discussion
The newly acquired 40Ar/39Ar single crystal sanidine ages are the first radioisotopic age constraints for Pipestone Springs strata. They provide a basis for detailed stratal chronostratigraphy, depositional environment interpretation, ichnofossil identification, and correlation with the Flagstaff Rim section located in central Wyoming, about 42 km southwest of Casper, Wyoming (Fig. 8A). They also enable a more regional foundation for understanding late Eocene mammalian and landscape paleogeography.
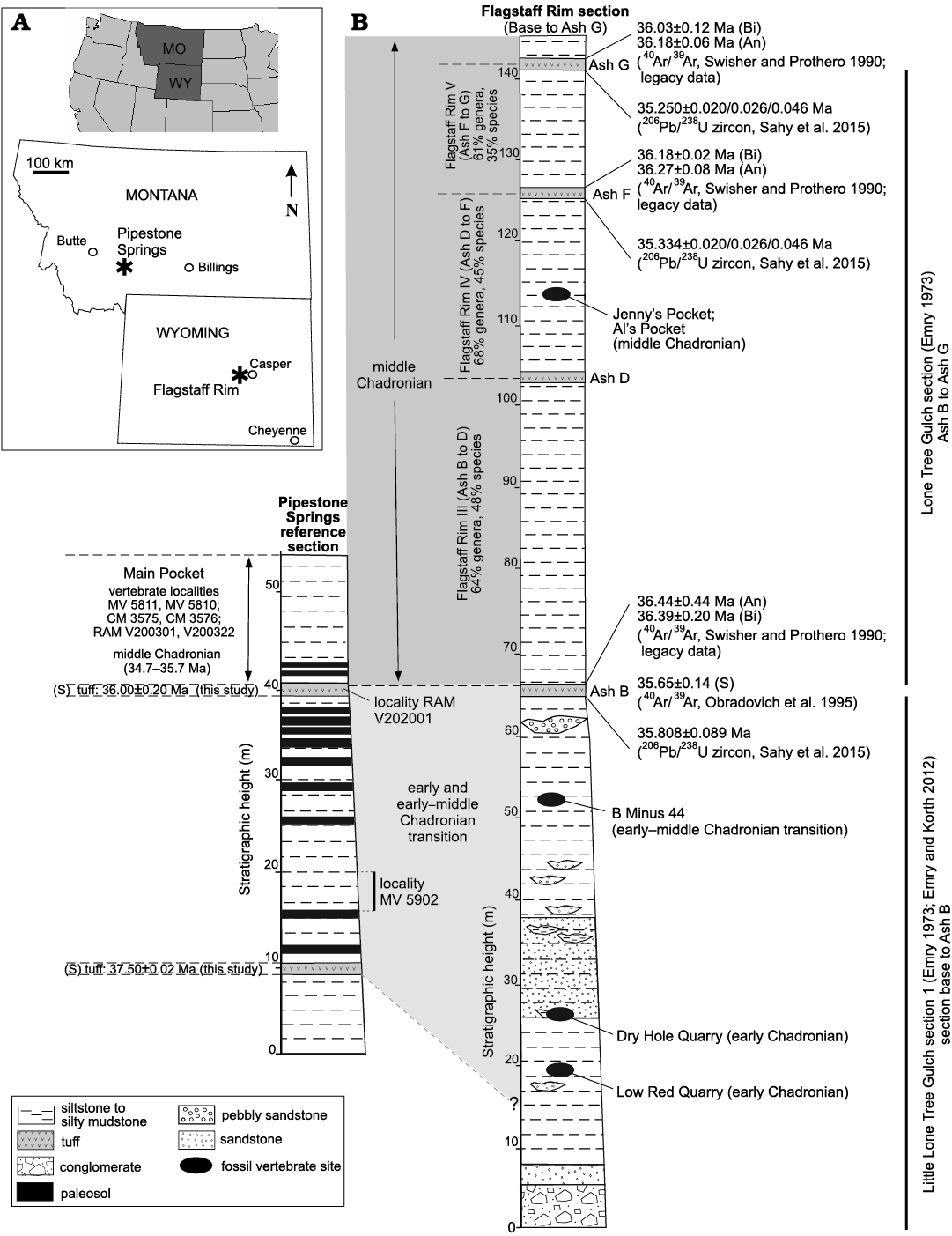
Fig. 8. Correlation of Pipestone Springs and Flagstaff Rim upper Eocene lithostratigraphy, and stratigraphic position of tuffs and vertebrate assemblages. A. Location of Flagstaff Rim (Wyoming) and Pipestone Springs (Montana). B. Pipestone Springs reference section and Flagstaff Rim section showing its lithostratigraphy, and stratigraphic position of tuffs and vertebrate assemblages. Sections are correlated based upon Pipestone Spring’s 40Ar/39Ar age of 36.00±0.20 Ma tuff and Flagstaff Rim’s Ash B, and on middle Chadronian vertebrate assemblages in the PSMP and between Flagstaff Rim’s Ash B to Ash G. 40Ar/39Ar age controls for Pipestone Springs strata are from this study; Flagstaff Rim tuff ages are from Swisher and Prothero (1990), Obradovich et al. (1995), and Sahy et al. (2015) Abbreviations: An, anorthoclase; Bi, biotite; S, sanidine (denote minerals used for single crystal 40Ar/39Ar age analyses of tuffs).
The combination of new 40Ar/39Ar single crystal sanidine ages for two lapilli tuffs with the known PSMP middle Chadronian vertebrate assemblage for the Pipestone Springs reference section not only makes this a high-resolution age late Eocene (Priabonian) reference section, but it also significantly extends the section’s age. These strata have previously been labeled as only middle Chadronian (approximately 35.7–34.7 Ma) (Tabrum et al. 1996, 2001; Tabrum and Metais 2007; Lofgren et al. 2017). Instead, given the newly acquired 40Ar/39Ar single crystal sanidine ages of 37.50±0.02 Ma and 36.00±0.20 Ma (Priabonian, late Eocene) for lapilli tuffs located near the base and immediately below the PSMP, Pipestone Springs strata represent a longer time duration (~2 Ma) and are significantly older than previously thought. The longer time duration for Pipestone Springs strata extends them into the latest Duchesnean based upon the 37 Ma Duchesnean–Chadronian numerical boundary estimated by Prothero and Swisher (1992), Prothero and Emry (1996), and retained by Robinson et al. (2004).
The longer time duration for these strata requires reevaluation of earlier interpretations of how they correlate to the GPTS. Based on paleomagnetic analysis of Prothero (1984) and its contained middle Chadronian vertebrate assemblage, the Pipestone Springs section was most recently correlated to chrons C15r–C15n (Tabrum et al. 1996; Prothero and Emry 2004). However, with strata now known to be as old as 37.50±0.02 Ma, and the acknowledged presence of numerous paleosols that represent hiatuses in the basin-margin depositional record of the Pipestone Springs sections (Fig. 4), the C15r–C15n GPTS correlation is most likely untenable. Thus, based on 40Ar/39Ar ages now available in this study for Pipestone Spring strata, the sections appear to at best correlate with chrons C17n.1n through C15n.
The depositional environment for Pipestone Springs strata is aeolian, which is a much different interpretation than those previously published for these strata (Kuenzi 1966; Kuenzi and Fields 1971; Axelrod 1984; Garcia 1992). Our aeolian interpretation is based upon these rock units being fine-grained deposits and on stratal sedimentary features such as lack of fluvial, paludal, or debris flow structures, massive stratification, low abrasion of fresher vitric grains, and mantle bedding. The numerous paleosols intercalated within the Pipestone Springs section represent a loess-paleosol sequence, similar to those loess-paleosol sequences found in the northern European Loess Belt (Lemkuhl et al. 2016) and the China Loess Plateau (Maher 2016). Thus, the Pipestone Springs paleosols indicate a much-repeated record of stable land surfaces existing between aeolian events that encompass both the deposition of lapilli tuffs and of other re-worked volcaniclastic material. Speculation on a source(s) of volcanic material has previously been proposed to be northwestern United States or the Nevada–Utah Great Basin early Cenozoic volcanic centers (Fields et al. 1985; Larson and Evanoff 1998; Sahy et al. 2015; Rowley and Fan 2016). However, our on-going studies of southwestern Montana Cenozoic tuffs suggest a more local source such as the Eocene–early Oligocene Helena Volcanic Field, located in west-central Montana near Helena, Montana. This inference is based on the presence of similar age lapilli tuff in several southwestern Montana locations, and similar tuff chemistry of these tuffs with those in the Helena Volcanic Field (Chadwick 1985; Schmidt et al. 1994; O’Neill et al. 2005).
An interesting aspect of the aeolian depositional interpretation for Pipestone Springs strata is that their maximum age, at 37.50±0.02 Ma, makes them the oldest aeolian strata identified in Cenozoic deposits of the central Rockies and Great Plains. Evanoff et al. (1992), Evanoff (2015), and Benton et al. (2015) have reported younger, early Oligocene aeolian deposits in the Wyoming region. Rowley and Fan (2016) do suggest that aeolian strata of ca. 36 Ma in age could be present in a small area of central Wyoming. However, these authors were unaware of the Pipestone Springs loessites and thus they concluded that widespread aeolian deposition did not occur in the central Rockies and Great Plains until about 30 Ma.
In addition to the Pipestone Springs loessites, similar age strata found in other areas of southwestern Montana represent aeolian as well as a variety of other continental depositional environments (see Hanneman and Wideman 1991 for a summary of these studies). These co-existing depositional environments suggest a dynamic Eocene landscape rather than one with a predominance of a single depositional environment. Given that western Montana was undergoing extensional tectonism from at least middle Eocene to early Oligocene and that regional uplift coincided with a global cooling trend (Hanneman and Wideman 1991; Constenius 1996; Fan et al. 2017; Schwartz et al. 2019), a range of associated depositional environments is expected.
The ichnofossil-paleosol patterns exhibited in the Pipestone Springs vertisol, alfisol, and inceptisol paleosols record periods of landscape stability within a loessic depositional environment (e.g., Muhs and Bettis 2003; Mason et al. 2008) where pedogenesis was the most prevalent process that occurred between sedimentation episodes. Collectively, the Pipestone Springs paleosols are tentatively interpreted as including compound, composite, and cumulative paleosols because of their stacking pattern within the pedogenic profiles with respect to the parent material, formation on pre-existing pedogenic horizons, and/or overthickening of horizons (e.g., Morrison 1967, 1978; Kraus 1999; Hasiotis and Platt 2012; Fischer and Hasiotis 2018). Vertisol paleosols exhibit both compound and composite profiles depending on whether the profile reflects rapid sedimentation and insignificant erosion (compound paleosol) or the rate of pedogenesis was greater than sedimentation (composite paleosol). The alfisol paleosols typically developed composite profiles because the rate of pedogenesis was greater than that of sedimentation. Nonsteady and steady sedimentation produced inceptisols with reworked volcanic and non-volcanic parent material, thus generating both compound and cumulative paleosols.
During intervals of soil formation, the Pipestone Springs landscape was covered with ample vegetation that fed dung-producing herbivores. This dung was most likely used by adult dung beetles to construct shallow nests in which their eggs were laid. These eggs later hatched into larvae that fed on the provisions (e.g., Halffter and Edmond 1982; Halffter et al. 1985). The combination of biotic and abiotic features in the vertisols, alfisols, and inceptisols indicate that they all represent well-drained paleosol profiles in which vadose zone conditions dominated (e.g., Hasiotis 2002, 2004, 2007, 2008; Hasiotis et al. 2007; Hasiotis and Platt 2012). The ichnofossil-bearing soils were each subsequently buried when the accumulation of loess resumed.
Interestingly, the Pipestone Springs stratigraphic sections have strong similarities with the Flagstaff Rim stratigraphic section of central Wyoming (Emry 1973; Emry 1992; Emry and Korth 2012; Sahy et al. 2015) in terms of age and vertebrate assemblages (Fig. 8). The Pipestone Springs’ upper lapilli tuff, with a sanidine 40Ar/39Ar age of 36.00±0.20 Ma, suggests an age correlation with the Ash B tuff at Flagstaff Rim, based on the 35.808±0.089 Ma 206Pb/238U (zircon) age published by Sahy et al. (2015). These same authors recalibrated a sanidine 40Ar/39Ar age for Ash B previously published by Obradovich et al. (1995). Their recalculated age of 35.65±0.14 Ma for Ash B was done relative to a Fish Canyon sanidine age of 28.201 Ma using the intercalibration factor of Renne et al. (1998). 40Ar/39Ar ages using biotite and anorthoclase for Ash B were done earlier by Swisher and Prothero (1990) for Ash B, but because of problems with these legacy data (Sahy et al. 2015), these 40Ar/39Ar ages were not used for comparison purposes in our study and are noted in Fig. 8 as legacy data. Possible equivalents of Flagstaff Rim tuffs D through G (Emry 1973; Evanoff et al. 1992; Obradovich et al. 1995; Larson and Evanoff 1998; Sahy et al. 2015, 2020) are not found at Pipestone Springs, but as expected, some differences in deposition between these two sections are probably due to the non-continuous sedimentation of basin-margin sites such as that of Pipestone Springs (Fig. 8).
The similarities between the two sections are most apparent based on the relatively high percentage of shared mammalian taxa between the PSMP and middle Chadronian assemblages known from sites stratigraphically between Ash B and Ash G at Flagstaff Rim. The middle Chadronian is typified by Flagstaff Rim collections from ca. 15 m below Ash B to ca. 15 m above Ash G (Prothero and Emry 2004) (Fig. 8), based on the section and observed ranges of taxa provided by Emry (1992: figs. 5.1, 5.2). A principal correlative is the mammalian assemblage from the PSMP (Tabrum et al. 1996, 2001; Prothero and Emry 2004; Lofgren et al. 2017) (Fig. 8). In spite of their geographic separation of 570 km, the PSMP mammalian assemblage is very similar to three assemblages from the Flagstaff Rim section: the Flagstaff Rim III based on sites between Ash B and Ash D, the Flagstaff Rim IV based on sites between Ash D and Ash F (compilation of collections from Jenny’s Pocket, Al’s Pocket, and sites about 16 m below Ash F in the Middle Fork of Lone Tree Gulch) (Korth and Emry 1997), and the Flagstaff Rim V based on sites between Ash F and Ash G in the South Fork of Lone Tree Gulch. When comparing the 45 mammalian genera known from PSMP (Lofgren et al. 2017: table 1), to mammalian genera identified from the III, IV, and V faunal levels at Flagstaff Rim (III, 16962; IV, 16963: V, 16964; source: https://paleobiodb.org/classic/displayCollResults?collection_no), about 64% of Flagstaff Rim III, 68% of Flagstaff Rim IV, and 61% of Flagstaff Rim V genera are shared with PSMP. Similarly, with respect to shared mammalian species, 48% of Flagstaff Rim III, 45% of Flagstaff Rim IV, and 35% of Flagstaff Rim V are shared with PSMP, with most shared species for each comparison representing rodents and lipotyphlans.
The basal lapilli tuff of 37.50±0.02 Ma at Pipestone Springs does not have a correlative tuff at Flagstaff Rim, but vertebrate assemblages from about 20–24 m above the Flagstaff Rim section’s base are identified as earliest to early Chadronian (Emry and Korth 2012), so are approximately age equivalent. Pipestone Springs’ localities MV 5902 and RAM V202001 are present between the basal lapilli tuff and the 36.00±0.20 Ma lapilli tuff (Fig. 4), but the age ranges of mammals from these localities are too broad to be useful for faunal correlations.
Conclusions
Priabonian (upper Eocene) tuffs and middle Chadronian mammalian fauna provide a high-resolution chronostratigraphy for strata in the Pipestone Springs area of southwestern Montana. Two lapilli tuffs within the basal to upper portions of exposed strata have 40Ar/39Ar single crystal sanidine ages of 37.50±0.02 Ma and 36.00±0.20 sanidine 40Ar/39Ar ages, respectively. Uppermost Pipestone Springs strata contain a diverse middle Chadronian vertebrate fauna. The new 40Ar/39Ar ages extend the age of the Pipestone Springs strata to include deposits as old as latest Duchesnean–early Chadronian (Priabonian). In combination with its mammalian fauna, the new 40Ar/39Ar ages confirm that Pipestone Springs strata are correlative to part of the Flagstaff Rim section in central Wyoming, and now provide a basis for better determining late Eocene mammalian paleogeograpy and regional paleolandscapes for the Rocky Mountain–Great Plains region.
A loessite–paleosol sequence comprises Pipestone Springs deposits and consequently our interpretation indicating that the Pipestone Springs strata are aeolian requires new insight into depositional environments of southwestern Montana basin-margin strata during the late Eocene. Paleosols at Pipestone Springs include vertisols, alfisols, and inceptisols that contain newly identified ichnofossils likely constructed by dung beetle (Coleoptera) adults and modified by larvae and pupae based on comparisons to modern and ancient traces attributed to this group. The ichnofossil–paleosol patterns exhibited by the Pipestone Springs paleosols reflect periods of landscape stability within a loessic depositional environment where pedogenesis was the most prevalent process that occurred between the infrequent sedimentation episodes.
Acknowledgements
We are thankful to be able to honor Rich Cifelli as he has been an inspiration to every geoscientist and vertebrate paleontologist who has worked on Mesozoic and Cenozoic strata over the past 40 or more years, and highly value Rich’s field crews and camps that have always been great places for fun as well as science. We also thank the Museum of the Rockies (Bozeman, Montana, USA) and the Raymond M. Alf Museum of Paleontology (Claremont, California, USA) and the David B. Jones Foundation for funding the 40Ar/39Ar analyses of Pipestone Springs tuffs. The assistance of David Mogk (ICAL) for SEM and EDS analyses is gratefully acknowledged and much appreciated. Whitehall Geogroup, Inc., Whitehall, Montana, USA provided funds for the ICAL analyses. We greatly appreciate manuscript reviews by Barry Allbright (University of North Florida, Jacksonville, Florida, USA), Emmett Evanoff (University of Northern Colorado, Greeley, Colorado, USA), and Donald Prothero (La Crescenta-Montrose, California, USA). We thank Daniel Hirmas (University of California, Riverside, California, USA) for detailed discussions on soil processes. Our appreciation is also extended to all who have worked at the Pipestone Springs locality and thus helped provide a biostratigraphic and geochronologic framework for our current study.
References
Axelrod, R.B. 1984. Tertiary Sedimentary Facies, Depositional Environments, and Structure, Jefferson Basin, Southwestern Montana. 64 pp. M.Sc. Thesis, University of Montana, Missoula.
Barnosky, A.D., Bibi, F., Hopkins, S.S.B., and Nichols, R. 2007. Biostratigraphy and magnetostratigraphy of the mid-Miocene Railroad Canyon sequence, Montana and Idaho, and age of the mid-Tertiary unconformity west of the continental divide. Journal of Vertebrate Paleontology 27: 204–224. Crossref
Bellosi, E.S. 2010. Loessic and fluvial sedimentation in Sarmiento Formation pyroclastics, middle Cenozoic of central Patagonia. In: R.H. Madden, A.A. Carlini, M.G. Vucetich, and R.F. Kay (eds.), The Paleontology of Gran Barranca: Evolution and Environmental Change through the Middle Cenozoic of Patagonia, 278–292. Cambridge University Press, Cambridge.
Benton, R.C., Terry, D.O. Jr., Evanoff, E., and McDonald, H.G. 2015. The White River Badlands: Geology and Paleontology. 222 pp. Indiana University Press, Bloomington.
Bown, T.M. and Kraus, M.J. 1987. Integration of channel and floodplain suites. Developmental sequence and lateral relations of alluvial paleosols. Journal of Sedimentary Petrology 57: 587–601. Crossref
Bromley, R.G. 1996. Trace Fossils: Biology, Taphonomy, and Applications. 2nd Edition. 361 pp. Chapman and Hall, London.
Chadwick, R.A. 1978. Chronology of post-Eocene rhyolitic and basaltic volcanism in southwestern Montana. Isochron/West 22: 25–28.
Chadwick, R.A. 1985. Overview of Cenozoic volcanism in the west central United States. In: R.M. Flores and S.S. Kaplan (eds.), Cenozoic Paleogeography of the West-Central United States, 359–382. Society of Economic Paleontologists and Mineralogists, Tulsa.
Constenius, K.N. 1996. Late Paleogene extensional collapse of the Cordilleran foreland fold and thrust belt. Geological Society of America Bulletin 108: 20–39. Crossref
Constenius, K.N., Esser, R.P., and Layer, P.W. 2003. Extensional collapse of the Charleston-Nebo salient and its relationship to space-time variations in Cordilleran orogenic belt tectonism and continental stratigraphy. In: R.G. Raynolds and R.M. Flores (eds.), Cenozoic Systems of the Western United States, 303–353. Society of Economic Paleontologists and Mineralogists, Denver.
Douglass, E. 1899. The Neocene Lake Beds of Western Montana. 27 pp. Unpublished M.Sc. Thesis, University of Montana, Missoula.
Douglass, E. 1901. Fossil Mammalia of the White River Beds of Montana. Transactions of the American Philosophical Society 20: 237–279. Crossref
Douglass, E. 1903. New vertebrates from the Montana Tertiary. Annals of Carnegie Museum 2: 145–199.
Douglass, E. 1905. The Tertiary of Montana. Memoirs of the Carnegie Museum 2: 203–224. Crossref
Dudás, F.Ö., Ispolatov, V.O., Harlan, S.S., and Snee, L.W. 2010. 40Ar/39Ar geochronology and geochemical reconnaissance of the Eocene Lowland Creek Volcanic Field, West‐Central Montana. The Journal of Geology 118: 295–304. Crossref
Emry, R.J. 1973. Stratigraphy and preliminary biostratigraphy of the Flagstaff Rim area, Natrona County, Wyoming. Smithsonian Contributions to Paleobiology 18: 1–48. Crossref
Emry, R.J. 1992. Mammalian range zones in the Chadronian White River Formation at Flagstaff Rim, Wyoming. In: D.R. Prothero and W.A. Berggren (eds.), Eocene–Oligocene Climatic and Biotic Evolution, 106–115. Princeton University Press, Princeton. Crossref
Emry, R.J. and Hunt, R.M. Jr. 1980. Maxillary dentition and new records of Daphoenictis, an Oligocene amphicyonid carnivore. Journal of Mammalogy 61: 720–723. Crossref
Emry, R.J. and Korth, W.W. 1996. The Chadronian squirrel “Sciurus” jeffersoni Douglass, 1901: a new generic name, new material, and its bearing on the early evolution of Sciuridae (Rodentia). Journal of Vertebrate Paleontology 16: 775–780. Crossref
Emry, R.J. and Korth, W.W. 2012. Early Chadronian (late Eocene) rodents from the Flagstaff Rim area, central Wyoming. Journal of Vertebrate Paleontology 32: 419–432. Crossref
Evanoff, E. 2015. Cenozoic geology and geomorphology of the Laramide Mountains,Wyoming. In: E. Evanoff (ed.), Colorado Scientific Society Field Trip Guide. 46 pp. Colorado Scientific Society, Denver.
Evanoff, E., Prothero, D.R., and Lander, R.H. 1992. Eocene–Oligocene change in North America: The White River Formation near Douglas, east-central Wyoming. In: D.R. Prothero and W.A. Berggren (eds.), Princeton Series in Geology and Paleontology. Eocene–Oligocene Climatic and Biotic Evolution, 116–130. Princeton University Press, Princeton. Crossref
Fan, M., Constenius, K.N., and Dettman, D.L. 2017. Prolonged high relief in the northern Cordilleran orogenic front during middle and late Eocene extension based on stable isotope paleoaltimetry. Earth and Planetary Science Letters 457: 376–384. Crossref
Fields, R.W., Rasmussen, D.L., Tabrum, A.R., and Nichols, R. 1985. Cenozoic rocks of the intermontane basins of western Montana and eastern Idaho. In: R.M. Flores and S.S. Kaplan (eds.), Cenozoic Paleogeography of the West Central United States, 9–36. Society of Economic Paleontologists and Mineralogists, Tulsa.
Fischer, S.A. and Hasiotis, S.T. 2018. Stratigraphic changes in ichnopedofacies of the Upper Triassic Chinle Formation, Utah: implications to paleohydrology and paleoclimate. Annales Societatis Geologorum Poloniae 88: 127–162.
Fritz, W.J., Sears, J.W., and Wampler, J.M. 2007. Cenozoic volcanic rocks of southwestern Montana. Northwest Geology 36: 91–110.
Garcia, D. 1992. Fossil Mammalia from the Pipestone Creeks Region, late Eocene and Oligocene (Chadronian and Orellan), Jefferson County, Montana. 215 pp. Unpublished Ph.D. Thesis, University of California, Berkeley.
Halffter, G. and Edmonds, W.D. 1982. The Nesting Behavior of Dung Beetles (Scarabaeinae): an Ecological and Evolutive Approach. 176 pp. Publicación del Instituto de Ecología, Mexico, D.F.
Halffter, V., Lopez-Guerrero, Y., and Halffter, G. 1985. Nesting and ovarian development in Geotrupes cavicollis Bates (Coleoptera: Scarabaeidae). Acta Zoologica Mexico 7: 1–28.
Hanneman, D.L. 1989. Cenozoic Basin Evolution in a Part of Southwestern Montana. 347 pp. Ph.D. Thesis, University of Montana, Missoula.
Hanneman, D.L. and Wideman, C.J. 1991. Sequence stratigraphy of Cenozoic continental rocks. Geological Society of America Bulletin 103: 1335–1345. Crossref
Hanneman, D.L. and Wideman, C.J. 2006. Calcic pedocomplexes—regional sequence boundary indicators. In: A.M. Alonso-Zarza and L.H. Tanner (eds.), Paleoenvironmental Record and Applications of Calcretes and Palustrine Carbonates. Geological Society of America Special Paper 416: 1–15.
Hanneman, D.L. and Wideman, C.J. 2013. Cenozoic tectonic sequences and basin evolution in western Montana. Geological Society of America. Abstracts with Programs 45 (7): 134. [published online, https://gsa.confex.com/gsa/2013AM/webprogram/Paper226039.html]
Hanneman, D.L. and Wideman, C.J. 2016. Cenozoic stratigraphy in western Montana: current status and a way forward. Geological Society of America. Abstracts with Programs 48 (7): 456. [published online, https://gsa.confex.com/gsa/2016AM/webprogram/Paper279878.html] Crossref
Hanneman, D.L., Cheney, E., and Wideman, C.J. 2003. Cenozoic sequence stratigraphy of northwestern USA. In: R.G. Raynolds and R.M. Flores (eds.), Cenozoic Systems of the Western United States, 135–156. Society of Economic Paleontologists and Mineralogists, Denver.
Hanneman, D.L. and Lofgren, D. 2017. Vertebrate paleontology and geology of high elevation Tertiary deposits in the Gravelly Range, southwestern Montana. Geological Society of America. Abstracts with Programs 49 (5) [published online, https://gsa.confex.com/gsa/2017RM/webprogram/Paper293156.html]. Crossref
Hasiotis, S.T. 2000. The invertebrate invasion and evolution of Mesozoic soil ecosystems: the ichnofossil record of ecological innovations. In: R.A. Gastaldo and W.A. Dimichele (eds.), Phanerozoic Terrestrial Ecosystems. Paleontological Society Papers 6: 141–170. Crossref
Hasiotis, S.T. 2002. Continental Trace Fossils. Society for Sedimentary Geology Short Course Notes Number 51. 132 pp. SEPM Society for Sedimentary Geology, Tulsa.
Hasiotis, S.T. 2003. Complex ichnofossils of solitary to social soil organisms: understanding their evolution and roles in terrestrial paleoecosystems. Palaeogeography, Palaeoclimatology, Palaeoecology 192: 259–320. Crossref
Hasiotis, S.T. 2004. Reconnaissance of Upper Jurassic Morrison Formation ichnofossils, Rocky Mountain Region, USA: paleoenvironmental, stratigraphic, and paleoclimatic significance of terrestrial and freshwater ichnocoenoses. Sedimentary Geology 167: 177–268. Crossref
Hasiotis, S.T. 2007. Continental ichnology: Fundamental processes and controls on trace fossil distribution. In: W. Miller, III. (ed.), Trace Fossils—Concepts, Problems, Prospects, 268–284. Elsevier Press, Cambridge. Crossref
Hasiotis, S.T. 2008. Reply to the Comments by Bromley et al. of the paper “Reconnaissance of the Upper Jurassic Morrison Formation ichnofossils, Rocky Mountain Region, USA: Paleoenvironmental, stratigraphic, and paleoclimatic significance of terrestrial and freshwater ichnocoenoses. Sedimentary Geology 208: 61–68. Crossref
Hasiotis, S.T. and Mitchell, C.E. 1993. A comparison of crayfish burrow morphologies: Triassic and Holocene fossil, paleo- and neo-ichnological evidence, and the identification of their burrowing signatures. Ichnos 2: 291–314. Crossref
Hasiotis, S.T. and Platt, B.F. 2012. Exploring the sedimentary, pedogenic, and hydrologic factors that control the occurrence and role of bioturbation in soil formation and horizonation in continental deposits: An integrative approach. The Sedimentary Record 10: 4–9. Crossref
Hasiotis, S.T., Kraus, M.J., and Demko, T.M. 2007. Climatic controls on continental trace fossils. In: W. Miller, III (ed.), Trace Fossils—Concepts, Problems, Prospects, 172–195. Elsevier, Amsterdam. Crossref
Hembree, D.I. and Hasiotis, S.T. 2007. Paleosols and ichnofossils of the White River Formation of Colorado: Insight into soil ecosystems of the North American midcontinent during the Eocene–Oligocene transition. Palaios 22: 123–142. Crossref
Hembree, D.I. and Hasiotis, S.T. 2008. Miocene vertebrate and invertebrate burrows defining compound paleosols in the Pawnee Creek Formation, Colorado, USA. Palaeogeography, Palaeoclimatology, Palaeoecology 270: 349–365. Crossref
Hough, J. 1955. An Upper Eocene fauna from the Sage Creek area, Beaverhead County, Montana. Journal of Paleontology 29: 22–36.
Hough, J. 1958. Tertiary beds of the Sage Creek area, Beaverhead County, Montana. In: R.W. Fields (ed.), Eighth Field Conference, Society of Vertebrate Paleontology, Western Montana Guidebook, 41–45. Montana State University, Missoula.
Hough, J. 1961. Review of Oligocene didelphid marsupials. Journal of Paleontology 35: 218–228.
Howden, H.F. and Peck, S.B. 1987. Adult habits, larval morphology, and phylogenetic placement of Taurocerastes patagonicus Philippi (Scarabaeidae: Geotrupinae). Canadian Journal of Zoology 65: 329–332. Crossref
Johnston, P.A., Eberth, D.A., Anderson, P.K. 1996, Alleged vertebrate eggs from Upper Cretaceous redbeds, Gobi Desert, are fossil insect (Coleoptera) pupal chambers: Fictovichnus new ichnogenus. Canadian Journal of Earth Sciences 33: 511–525. Crossref
Kay, J.L., Fields, R.W., and Orr, J. 1958. Faunal lists of Tertiary vertebrates from western and southwestern Montana. In: R.W. Fields (ed.), Eighth Field Conference, Society of Vertebrate Paleontology, Western Montana Guidebook, 33–39. Montana State University, Missoula.
Kellogg, K.S. and Harlan, S.S. 2007. New 40Ar/39Ar age determinations and paleomagnetic results bearing on the tectonic and magmatic history of the northern Madison Range and Madison Valley region, southwestern Montana, USA. Rocky Mountain Geology 42: 157–174. Crossref
Korth, W.W. 1994. Middle Tertiary marsupials (Mammalia) from North America. Journal of Paleontology 68: 376–397. Crossref
Korth, W.W. and Emry, R.J. 1997. A new species of Aulolithomys (Rodentia, Eomyidae) from the Chadronian (late Eocene) of Wyoming. Paludicola 1: 112–116.
Kraus, M.J. 1987. Integration of channel and floodplain suites. Vertical relations of alluvial paleosols. Journal of Sedimentary Petrology 57: 602–612. Crossref
Kraus, M.J. 1999. Paleosols in clastic sedimentary rocks: their geologic applications. Earth Science Reviews 47: 41–70. Crossref
Kuenzi, W.D. 1966. Tertiary Stratigraphy in the Jefferson River Basin, Montana. 293 pp. Ph.D. Thesis, University of Montana, Missoula.
Kuenzi, W.D. and Fields, R.W. 1971. Tertiary stratigraphy, structure and geologic history, Jefferson Basin, Montana. Geological Society of America Bulletin 82: 3374–3394. Crossref
Kuiper, K.F., Deino, A., Hilgen, F.J., Krijgsman, W., Renne, P.R., and Wijbrans, J.R. 2008. Synchronizing rock clocks of Earth history. Science 320: 500–504. Crossref
Larson, E.E. and Evanoff, E. 1998. Tephrostratigraphy and source of the tuffs of the White River sequence. In: D.O. Terry, H.E. LaGarry, and R.M. Hunt (eds.), Depositional Environments, Lithostratigraphy, and Biostratigraphy of the White River and Arikaree Groups (late Eocene to early Miocene, North America). Geological Society of America Special Paper 325: 1–14. Crossref
Laza, J.H. 2006. Dung-beetle fossil brood balls: The ichnogenera Coprinisphaera Sauer and Quirogaichnus (Coprinisphaeridae). Ichnos 13: 217–235. Crossref
Lehmkuhl, F., Zens, J., Krauss, L., Schulte, P., and Kels, H. 2016. Loess-paleosol sequences at the northern European loess belt in Germany: Distribution, geomorphology and stratigraphy. Quaternary Science Reviews 153: 11–30. Crossref
Li, L., Fan, M., and Hanneman, D.L. 2017. Stable isotope evidence (δ18O and δD) for late Eocene–late Miocene topography and climate in the northern Rocky Mountains, southwestern Montana. In: Geological Society of America. Abstracts with Programs 49 (6) [published online, https://gsa.confex.com/gsa/2017AM/meetingapp.cgi/Paper/299842]. Crossref
Lielke, K., Manchester, S., and Meyer, H. 2012. Reconstructing the environment of the Northern Rocky Mountains during the Eocene/Oligocene transition: constraints from the palaeobotany and geology of south-western Montana, USA. Acta Palaeobotanica 52: 317–358.
Lillegraven, J.A. and Tabrum, A.R. 1983. A new species of Centetodon (Mammalia, Geolabididae) from southwestern Montana and its biogeographical implications. Contributions to Geology, University of Wyoming 22: 57–73.
Lillegraven, J.A., McKenna, M.C., and Krishtalka, L. 1981. Evolutionary relationships of middle Eocene and younger species of Centetodon (Mammalia, Insectivora, Geolabididae) with a description of the dentition of Ankylodon (Adapisocricidae). University of Wyoming Publications 45: 1–115.
Lofgren, D., Hanneman, D.L., Bibbens, J., Gerken. L., Hu, F., Runkel, A., Kong, I., Tarakji, A., Helgeson, A., Gerard, I., Li, R., Li, S., and Ji, Z. 2020. Eocene and Oligocene mammals from the Gravelly Range of southwest Montana. Paludicola 12: 263–297.
Lofgren, D.L., Shen, C.Y., Buday, N.N., Ylagan, C.A., Lofgren, K.K., Lai, R.L., Santana-Grace, D.D., and Tabrum, A.R. 2017. Coprolites and mammalian carnivores from Pipestone Springs, Montana and their paleoecological significance. Annals of Carnegie Museum 84: 265–285. Crossref
Maher, B. 2016. Palaeoclimatic records of the loess/palaeosol sequences of the Chinese Loess Plateau. Quaternary Science Reviews 154: 23–84. Crossref
Mason, J.A., Miao, X.D., Hanson, P.R., Johnson, W.C., Jacobs, P.M., and Goble, R.J. 2008. Loess record of the Pleistocene–Holocene transition on the northern and central Great Plains, USA. Quaternary Science Reviews 27: 1772–1783. Crossref
Matthew, W.D. 1903. The fauna of the Titanotherium beds at Pipestone Springs, Montana. Bulletin of the American Museum of Natural History 19: 197–226.
McCafferty, A., Bankey, V., and Brenner, K.C. 1998. Merged Aeromagnetic and Gravity Data for Montana, a Web Site for Distribution of Gridded Data and Plot Files. Open-File Report 98-333. 20 pp. United States Geological Survey, Denver.
McDonald, E.V. and Busacca, A.J. 1992. Late Quaternary stratigraphy of loess in the channeled scabland and Palouse of Washington State. Quaternary Research 38: 141–156. Crossref
Min, K.W., Mundil, R., Renne, P.R., and Ludwig, K.R. 2000. A test for systematic errors in 40Ar/39Ar geochronology through comparison with U/Pb analysis of a 1.1-Ga rhyolite. Geochimica et Cosmochimica Acta 64: 73–98. Crossref
Morrison, R.B. 1967. Principles of Quaternary soil stratigraphy. In: R.B. Morrison and H.E. Wright (eds.), Means of Correlation of Quaternary Successions. INQUA, VII Congress, 1–69. International Association of Quaternary Research, Denver.
Morrison, R.B. 1978. Quaternary soil stratigraphy—concepts, methods, and problems. In: W.C. Mahaney (ed.), Quaternary Soils, 77–108. Geo Abstracts, Norwich.
Muhs, D.R and Bettis, E.A. III. 2003. Quaternary loess-paleosol sequences as examples of climate-driven sedimentary extremes. In: M.A. Chan and A.W. Archer (eds.), Extreme Depositional Environments: Mega End Members in Geologic Time. Geological Society of America Special Paper 370: 53–74. Crossref
Munsell Color 1975. Munsell Soil Color Charts. Natural Resources Conservation Service, United States Department of Agriculture, Baltimore.
Obradovich, J.D., Evanoff, E., and Larson, E.E. 1995. Revised single crystal laser fusion 40Ar/39Ar ages of Chadronian tuffs in the White River Formation of Wyoming. Geological Society of America. Abstracts with Programs 27: 77–78.
O’Neill, J.M., Lund, K., Van Gosen, B.S., Smith, B.D., Sole, T.C., and DeWitt, E.H. 2005. Geologic framework [Boulder and Basin watersheds, Montana]. In: D.A. Nimick, S.E. Church, and S.E. Finger (eds.), Integrated Investigations of Environmental Effects of Historical Mining in the Basin and Boulder Mining Districts, Boulder River Watershed, Jefferson County, Montana. U.S. Geological Survey Professional Paper 1652: 53–88.
Ostrander, G.E. 1983. A new genus of eomyid (Mammalia, Rodentia) from the early Oligocene (Chadronian), Pipestone Springs, Montana. Journal of Paleontology 57: 140–144.
Prothero, D.R. 1984. Magnetostratigraphy of the early Oligocene Pipestone Springs locality, Jefferson County, Montana. Contributions to Geology, The University of Wyoming 23: 33–36.
Prothero, D.R. and Swisher, C.C. III. 1992. Magnetostratigraphy and geochronology of the terrestrial Eocene–Oligocene transition in North America. In: D.R. Prothero and W.A. Berggren (eds.), Eocene–Oligocene Climatic And Biotic Evolution, 46–73. Princeton University Press, Princeton. Crossref
Prothero, D.R. and Emry, R.J. 1996. Summary. In: D.R. Prothero and R.J. Emry (eds.) The Terrestrial Eocene–Oligocene Transition in North America, 664–683. Cambridge University Press, New York. Crossref
Prothero, D.R. and Emry, R.J. 2004. The Chadronian, Orellan, and Whitneyan North American Land Mammal Ages. In: M.O. Woodburne (ed.), Late Cretaceous and Cenozoic Mammals of North America, 56–168. Columbia University Press, New York. Crossref
Raigemborn, M.S., Beilinson, E., Krause, J.M., Varela, A.N., Bellosi, E., Matheos, S.D., and Sosa, N. 2018. Paleolandscape reconstruction and interplay of controlling factors of an Eocene pedogenically-modified distal volcaniclastic succession in Patagonia. Journal of South American Earth Sciences 86: 475–496. Crossref
Renne, P.R., Swisher, C.C., Deino, A.L., Karner, D.B., Owens, T.L., and DePaolo, D.J. 1998. Intercalibration of standards, absolute ages and uncertainties in 40Ar/39Ar dating. Chemical Geology 145: 117–152. Crossref
Retallack, G.G. 2001. Soils of the Past. An Introduction to Paleopedology. 2nd Edition. 404 pp. Blackwell Science, Oxford.
Robinson, P., Gunnell, G.F., Walsh, S.L., Clyde, W.C., Storer, J.E., Stucky, R.K., Froehlich, D.J., Ferrusquia-Villafranca, I., and McKenna, M.C. 2004. Wasatchian through Duchesnean Biochronology. In: M.O. Woodburne (ed.), Late Cretaceous and Cenozoic Mammals of North America, 106–155. Columbia University Press, New York. Crossref
Roselli, F.L. 1939. Apuntes de geología y paleontología Uruguaya. Sobre insectos del Cretacico del Uruguay o descubrimiento de admirables instintos constructivos de esaepoca. Boletín de la Sociedad Amigos de las Ciencias Naturales “Kraglievich-Fontana” 1: 72–102.
Roselli, F.L. 1987. Paleoicnología: Nidos de insectos f´osiles de la cubertura Mesozoica del Uruguay. Publicaciones del Museo Municipal de Nueva Palmira 1: 1–56.
Rowley, J. and Fan, M. 2016, Middle Cenozoic diachronous shift to eolian deposition in the central Rocky Mountains: Timing, provenance, and significance for paleoclimate, tectonics, and paleogeography. Geosphere 12: 1795–1812. Crossref
Sahy, D., Condon, D.J., Terry, D.O., Fischer, A.U., and Kuiper, K.F. 2015. Synchronizing terrestrial and marine records of environmental change across the Eocene–Oligocene transition. Earth and Planetary Science Letters 427: 171–182. Crossref
Sahy, D., Hiess, J, Fischer, A.U., Condon, D.J., Terry, D.O. Jr, Abels, H.A., Hüsing, S.K., and Kuiper, K.F. 2020. Accuracy and precision of the late Eocene–early Oligocene geomagnetic polarity time scale. Geological Society of America Bulletin 132: 373–388. Crossref
Schmidt, R.G., Loen, J.S., Wallace, C.A., and Mehnert, H.H. 1994. Geology of the Elliston region, Powell and Lewis and Clark counties, Montana. U.S. Geological Survey Bulletin 2045: i–iv + 1–25.
Schwartz, T.M. and Graham, S.A. 2017. Depositional history and provenance of Paleogene strata in the Sage Creek basin, southwestern Montana. Geosphere 13: 1–25. Crossref
Schwartz, T.M., Methner, K., Mulch, A., Graham, S.A., and Chamberlain, C.P. 2019. Paleogene topographic and climatic evolution of the Northern Rocky Mountains from integrated sedimentary and isotopic data. Geological Society of America Bulletin 131: 1203–1223. Crossref
Soil Survey Staff 1951. Soil Survey Manual. U.S. Department Agricultural Handbook 18. 230 pp. Government Printing Office, Washington, D.C.
Soil Survey Staff 1999. Soil Taxonomy: A Basic System of Soil Classification for Making and Interpreting Soil Surveys. 2nd Edition. 886 pp. United States Department of Agriculture, Washington DC.
Steiger, R.H. and Jäger, E. 1977. Subcommission on geochronology: convention on the use of decay constants in geo-and cosmochronology. Earth and Planetary Science Letters 36: 359–362. Crossref
Swisher, C.C. and Prothero, D.R. 1990. Single-crystal 40Ar/39Ar dating of the Eocene–Oligocene transition in North America. Science 249: 760–762. Crossref
Tabrum, A.R. and Fields, R.W. 1980. Revised mammalian faunal list for the Pipestone Springs local fauna (Chadronian, early Oligocene), Jefferson County, Montana. Northwest Geology 9: 45–51.
Tabrum, A.R. and Métais, G. 2007. Pipestoneia douglassi, a new genus and species of selenodont artiodactyl from the Pipestone Springs area, Jefferson County, Montana. Bulletin of Carnegie Museum of Natural History 39: 83–96. Crossref
Tabrum, A.R., Nichols, R., and Barnosky, A.D. 2001. Tertiary paleontology of southwestern Montana and adjacent Idaho. In: C.L. Hill (ed.), Mesozoic and Cenozoic Paleontology in the Western Plains and Rocky Mountains. Museum of the Rockies Occasional Paper No. 3, 3–112. Museum of the Rockies, Bozeman.
Tabrum, A.R., Prothero, D.R., and Garcia, D. 1996. Magnetostratigraphy and biostratigraphy of the Eocene–Oligocene transition, southwestern Montana. In: D.R. Prothero and R.J. Emory (eds.), The Terrestrial Eocene–Oligocene Transition in North America, 287–311. Cambridge University Press, Cambridge. Crossref
Taylor, J.R. 1982. An Introduction to Error Analysis: The Study of Uncertainties in Physical Measurements. 270 pp. University Science Books, Mill Valley.
Tucker, M.E. 2011. Sedimentary Rocks in the Field: A Practical Guide. 4th Edition. 288 pp. John Willey and Sons Ltd., Hoboken.
Zarate, M. and Blasi, A.M. 1991. Late Pleistocene and Holocene loess deposits of the southeastern Buenos Aires Province, Argentina. GeoJournal 24: 211–220. Crossref
Acta Palaeontol. Pol. 67 (1): 5–20, 2022
https://doi.org/10.4202/app.00901.2021