
First spalacotheriid and dryolestid mammals from the Cretaceous of Germany
THOMAS MARTIN, ALEXANDER O. AVERIANOV, JULIA A. SCHULTZ, RICO SCHELLHORN, and ACHIM H. SCHWERMANN
Martin, T., Averianov, A.O., Schultz, J.A., Schellhorn, R., and Schwermann, A.H. 2022. First spalacotheriid and dryolestid mammals from the Cretaceous of Germany. Acta Palaeontologica Polonica 67 (1): 155–175.
The new spalacotheriid “symmetrodontan” Cifellitherium suderlandicum gen. et sp. nov. from the Barremian–Aptian of the Balve locality in northwestern Germany is the first record of spalacotheriids in Central Europe. The new taxon is based on one lower and two upper molars. Cifellitherium is similar to Spalacotherium, but differs by smaller stylocone and larger paraststyle on the upper molars, and a labially interrupted cingulid on the lower molar. The new dryolestid Minutolestes submersus gen. et sp. nov. is based on one lower and two upper molars of small size. Phylogenetic analysis revealed it as sister taxon of the clade comprising Laolestes, Krebsotherium, Dryolestes, and Guimarotodus. A dryolestid mandible with unevenly rooted molars and extremely worn down teeth cannot be attributed to Minutolestes submersus gen. et sp. nov. due to possessing molars twice as large in size. The mandible is assigned to a new dryolestid taxon, Beckumia sinemeckelia gen. et sp. nov., and has the dental formula 3i, 1c, 4p, 8m and a fully reduced Meckel’s groove. The mandible lacks any trace of a coronoid or splenial. In the phylogenetic analysis, it appears as sister taxon of a clade comprising Achyrodon, Phascolestes, Crusafontia, and Hercynodon. The new mammals are the stratigraphically youngest European representatives of their clades. The late survival of Beckumia sinemeckelia gen. et sp. nov. and Minutolestes submersus gen. et sp. nov. is possibly the result of isolated evolution in an Early Cretaceous island environment. This finding of new spalacotheriid and dryolestid mammals from Central Europe adds to an emerging paleobiogeographic pattern that Europe was distinct from Asia in the constituents of mammalian faunas during the Barremian–Aptian.
Key words: Mammalia, Spalacotheriidae, Cifellitherium, Crusafontia, Dryolestidae, Beckumia, Minutolestes, Balve, Cretaceous.
Thomas Martin [tmartin@uni-bonn.de], Julia A. Schultz [jaschultz@uni-bonn.de], and Rico Schellhorn [rico.schellhorn@uni-bonn.de], Institute of Geosciences, Section Paleontology, Rheinische Friedrich-Wilhelms-Universität Bonn, Nussallee 8, 53115 Bonn, Germany.
Alexander O. Averianov [dzahrakuduk@mail.ru], Zoological Institute of the Russian Academy of Sciences, Universitetskaya Emb. 1, Saint Petersburg, 199034, Russia.
Achim H. Schwermann [achim.schwermann@lwl.org], LWL-Museum of Natural History, Westphalian State Museum and Planetarium, Sentruper Straße 285, 48161 Münster, Germany.
Received 31 May 2021, accepted 27 September 2021, available online 30 March 2022.
Copyright © 2022 T. Martin et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
The record of spalacotheriid “symmetrodontans” and dryolestids in Europe has, until now, been restricted to the western part of the continent: the Iberian Peninsula, France, and the British Isles. Here we report the first spalacotheriid “symmetrodontan” from Central Europe, along with two new dryolestids, which are geologically the youngest representatives of their clades in Europe. Spalacotheriids are a monophyletic group within the paraphyletic “symmetrodontans” and are characterized by an acute angulation of the molar main cusps, in contrast to the paraphyletic assemblage of “obtuse angular symmetrodontans”, comprising the Late Triassic to Early Jurassic Kuehneotherium (Kuehneotheriidae) and the Late Jurassic to earliest Cretaceous Tinodon, among other less well-known taxa (Gill 2004b; Kielan-Jaworowska et al. 2004; Martin 2018).
Spalacotheriids are known from the Tithonian to the Campanian of the Northern Hemisphere, from North America (Patterson 1955, 1956; Fox 1976; Cifelli 1990; Cifelli and Gordon 1999; Cifelli and Madsen 1999; Cifelli et al. 2014), Europe (references see following paragraph), Uzbekistan (Archibald and Averianov 2005), China (Hu et al. 2005; Li and Luo 2006; Han and Meng 2016), and Japan (Tsubamoto et al. 2004).
Spalacotherium tricuspidens from the Early Cretaceous (Berriasian–Valanginian) of the Purbeck Limestone Group is the best-known European spalacotheriid “symmetrodontan” and the first one described (Owen 1854; Simpson 1928). Clemens and Lees (1971) announced a new species Spalacotherium taylori based on a lower molar from the late Valanginian Paddockhurst bone bed of the Wealden Supergroup in Great Britain. Ensom and Sigogneau-Russell (2000) described a fragmentary mandible from the Berriasian of the Sunnydown Farm Quarry in the Lulworth Formation of the Purbeck Limestone Group as Spalacotherium evansae, which also has been reported for the Berriasian of Cherves-de-Cognac (Charente, France) by Pouech et al. (2006). An additional species, Spalacotherium hookeri from the Berriasian of the Lulworth Formation at Durlston Bay, is based on a partial dentary with five posterior molars (Gill 2004a). Krebs (1985) reported Spalacotherium from the Iberian Peninsula with a large new species, S. henkeli, found in the Barremian of the Spanish locality Galve. Cuenca-Bescós et al. (2004) described Aliaga molinensis from the Hauterivian–Barremian of Molina Alto 1 in northeastern central Spain. Slightly younger (Barremian) is Yaverlestes gassoni from the Isle of Wight (Sweetman 2008). Krusat (1989) reported a lower molariform from the Tithonian–Berriasian of Porto Pinheiro (or Dinheiro) in Portugal and assigned it to ?Tinodon. Unfortunately, the tooth was destroyed while being mounted for SEM (Krusat 1989: 279). Based on Krusat’s (1989) description, detailed drawings, and conventional photographs, Sweetman (2008) suggested that the tooth should rather be placed within Spalacotheriidae.
In Europe, dryolestids are known mainly from the Middle Jurassic to the earliest Cretaceous of the Iberian Peninsula and of Great Britain, beginning with molars assigned to ?Dryolestidae from the Bathonian Oxfordshire in England (Simpson 1928; Freeman 1979; Martin 1999, 2000). Recently the first dryolestid from Central Europe, Hercynodon germanicus, was described from the Kimmeridgian of the Langenberg Quarry in the Harz Mountains in northern Germany (Martin et al. 2021). Cretaceous dryolestids have been reported from the Berriasian of the Purbeck Limestone Group: Amblotherium pusillum, Achyrodon nanus, and Phascolestes mustelulus. Isolated dryolestid molars have been reported from the Berriasian of Cherves-de-Cognac, but have not yet been described in detail (Mazin et al. 2006).
The youngest representative of dryolestids in Europe was hitherto Crusafontia cuencana from the Barremian of Uña and Galve localities in Spain (Henkel and Krebs 1969; Krebs 1971, 1985, 1993). The species Crusafontia amoae, based on two upper molars from the Hauterivian–Barremian of Galve (Cuenca-Bescós et al. 2011), was recently synonymized with C. cuencana (Martin et al. 2021). The mandible of C. cuencana is characterized by derived characters such as the full reduction of the coronoid and splenial as well as a very short Meckel’s groove (Henkel and Krebs 1969; Krebs 1971). The youngest record of a possible dryolestid in the Northern Hemisphere is a fragmentary lower molar from the Late Cretaceous (Campanian) of North America (Lillegraven and McKenna 1986). Groebertherium stipanici from the Campanian–Maastrichtian of Argentine Patagonia indicates an even longer survival in the Southern Hemisphere (Rougier et al. 2009).
Institutional abbreviations.—MfN, Museum für Naturkunde Berlin, Germany; WMNM, LWL-Museum of Natural History, Westphalian State Museum and Planetarium, Münster, Germany; Galve Th, Galve collection, housed at the Institute of Geosciences, section Paleontology, Universität Bonn, Germany.
Other abbreviations.—We follow standard convention in abbreviating tooth families as I, C, P, and M, with upper and lower case letters referring to upper and lower teeth, respectively.
Nomenclatural acts.—This published work and the nomenclatural acts it contains have been registered in ZooBank: urn:lsid:zoobank.org:pub:915DAF63-442C-4404-A3C9-F2F0A9490E54
Geological setting
The Cretaceous fossil locality of Balve is a part of the paleo karst structure in the northern Rhenish Massif (Fig. 1). Multiple such karst structures are ranging across its east-west axis in Sauerland. Comparable structures are also observed in the western extensions of the Rhenish Massif, beyond Wuppertal to Wülfrath and west of the Lower Rhine Embayment, in the northern Eifel Mountains. The discovery of the Rohdenhaus Cave near Wülfrath (Drozdzewski et al. 1998) initiated a new interpretation of the genesis of the cavities and sinkholes in this and other sites in the northern Rhenish Massif (Drozdzewski et al. 2017).
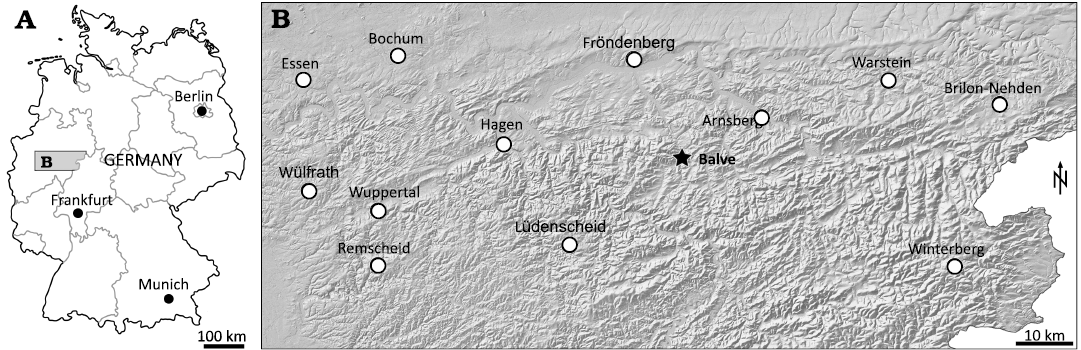
Fig. 1. Location of the Busche quarry near Balve in North Rhine-Westphalia (northwestern Germany). A. Map showing location of the northern Sauerland region of North Rhine-Westphalia in Germany. B. Map of the Sauerland region with topography, cities, and the Balve locality (asterisk).
The Rohdenhaus Cave lies more than 200 m below today’s ground surface. It is 100–200 m wide and up to 20 m high, and is assumed to be more than one kilometer long. According to the new interpretation, such a large cave is most probably not of exokarst origin, but rather of hydrothermal endokarst origin, which is similar as a geological process to the formation of other deep karstic structures in the region (Drozdzewski et al. 2017). They are all located in Devonian and Carboniferous limestone in the northern Rhenish Massif. Drozdzewski et al. (2017) attribute the hydrothermal corrosion to underlying Devonian black shale, which acted as a CO2 source and gave the rising water an acidic character. The exposure of these endokarst cavities to the surface occurred from the Early Cretaceous onward. Thus, Early Cretaceous or younger terrestrial infillings are found at various sites in the northern Sauerland (Wirth 1970; Huckriede 1982; Schudack 1987; Drozdzewski et al. 1998; Lanser and Heimhofer 2013; Richter et al. 2014). The vertebrate fossil-bearing sediments are Cretaceous infill of the endokarst cavities.
The Busche quarry is located near Balve, in the area of the Remscheid-Altena anticline. The fossil-bearing site belongs to a NW-SE striking fault, which is exposed within the quarry. The sedimentary infill of the sinkhole has been opened up by quarry operations at 300 m and 290 m above sea level, respectively, about 15 to 25 m below the original ground level before limestone quarrying began. Three horizontal outcrops of refilled karst cavities can be observed at the 290 m level. The largest has a nearly circular shape of about 30 × 35 m (Lanser and Heimhofer 2013).
In 2000, the first fossils, dinosaur teeth and bone fragments, were found in the sediments of this cavity. The infill consists mainly of gray clay and silt, intercalated with coarser boulders (Lanser and Heimhofer 2013). In the course of the excavations, a complex depositional pattern of primarily fine-grained sediments was revealed, mainly clayey silts which regularly contain small charcoal particles. The vertebrate fossils are usually isolated bone fragments showing varying degree of rolling due to transportation. In addition, well-rounded allochthonous clasts and sharp-edged fragments of lydite, quartz, and sandstone occur, beside more or less weathered larger blocks of autochthonous Devonian limestone containing calcite crystals. Calcite also occurs as small sharp-edged fragments and calcite arenite, but also in the form of isolated marine fossils (e.g., crinoid stalk elements, brachiopods) originating from the Devonian limestone. The sediments are layered, indicating successive accumulation of the infill. The sediment body exhibits distinct subsidence and sliding structures. In some cases, the sediments have subsided to such an extent that layered structures are parallel to the vertical sidewall of the cavity.
The sediments of the cavity infilling in Balve have been dated to be late Barremian to Aptian age based on pollen analysis by Lanser and Heimhofer (2013). In the Early Cretaceous, the area belonged to the Central European continental range, which includes the London-Brabant Massif, the Rhenish Massif, and the Bohemian Massif (Fig. 2).
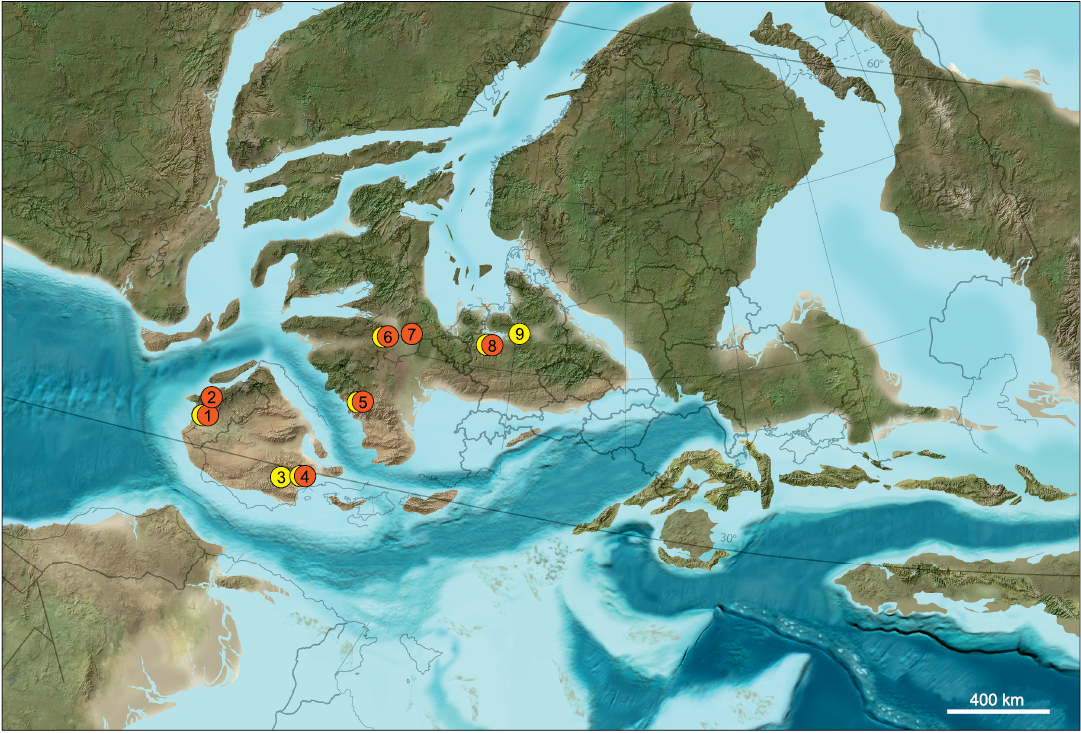
Fig. 2. Paleogeographic map of Europe in the Early Cretaceous, about 125 mya. Marked are Late Jurassic and Early Cretaceous localities that have yielded “symmetrodontans” (red) and dryolestids (yellow). 1, Guimarota Coal Mine (Portugal), Kimmerdigian; 2, Porto Pinheiro (or Dinheiro) (Portugal), Jurassic/Cretaceous boundary (Krusat 1989); 3, Uña (Spain), Barremian; 4, Galve (Spain), Hauterivian–Barremian; 5, Cherves-de-Cognac (France), Berriasian; 6, Purbeck Limestone Group (southern England), Berriasian–Valanginian; 7, Wealden Supergroup (southern England), Berriasian–Valanginian; 8, Balve (Germany), Barremian–Aptian; 9, Langenberg Quarry (Germany), Kimmeridgian. Map by Ron Blakey, Colorado Plateau Geosystems, Phoenix, Arizona.
Material and methods
While the macroscopically recognizable fossils were recovered directly, the excavated sediments were treated by screen washing procedures. The dried sediment was softened in water without the addition of chemicals and then screen washed. The minimum mesh size was 0.5 mm. About half of the teeth presented here derive from four tons of dried sediment (collected in 2020) that was broken into the following fractions: 263 kg of grain sizes greater than 6.7 mm, 76 kg of grain sizes between 2.0 and 6.7 mm, 38 kg of grain sizes between 1.0 and 2.0 mm, and 52 kg of grain sizes between 0.5 and 1.0 mm.
The fossils from the Balve excavation are curated into the paleontological collection of WMNM.
The dryolestid mandible WMNM P82301 was scanned using a 240 kV x-ray tube on the v|tome|x s μCT device (GE Sensing & Inspection Technologies GmbH phoenix|x-ray) in the Institute of Geosciences, University of Bonn, Germany, generating data with an isotropic voxel size of 13.7 µm. The tube settings were 100 kV and 120 µA, using a shutter speed of 333 ms per capture. Avizo 8 (Thermo Fisher Scientific) was used for the segmentation of teeth within their associated alveoli and creating the surface models of the jaw parts.
The isolated teeth were scanned using the 180 kV x-ray tube of the same µCT device (v|tome|x s, GE Sensing & Inspection Technologies GmbH phoenix|x-ray). Specimens WMNM P82304–P82308 were scanned at a resolution of 1.72 µm. Specimen WMNM P82302 was scanned at a resolution of 2.06 µm, with tube settings of 105 kV and 130 µA with a shutter speed of 500 ms. The tube settings of specimen WMNM P82304 were 100 kV and 100 μA with a shutter speed of 500 ms per capture. The tube settings of specimen WMNM P82305 were 110 kV and 140 μA with a shutter speed of 333 ms per capture. The tube settings of specimens WMNM P82306 and WMNM P82307 were 110 kV and 130 μA with a shutter speed of 333 ms per capture. The tube settings of specimen WMNM P82308 were 100 kV and 140 μA with a shutter speed of 500 ms per capture. VG Studio Max 3 was used for creating polygonal surface models and Polyworks 2014 (innovmetric) for smoothing the surface with the Gouraud filter function (at correction levels 0.1 and 0.3) as needed, and for reduction of data sizes (deleting doubled and floating polygons, reducing number of triangles for 3D PDF production). The polygonal STL models were converted to OBJ using Polyworks. OBJs were converted to U3D file format using DAZ Studio version 4.5 in order to create 3D PDF files which are available as SOM (Supplementary Online Material available at http://app.pan.pl/SOM/app67-Martin_etal_SOM.pdf). For light microscopy a Zeiss AxioZoom.V16 stereomicroscope with AxioCam HRC camera was used.
For the phylogenetic analysis of the new dryolestid taxa we used the data matrix from Martin et al. (2021) with the same settings for the analysis, applying a heuristic parsimony tree-search algorithm that included 1000 random addition sequences (RAS), followed by tree bisection and reconnection (TBR) branch swapping and keeping 10 trees in each replication traditional search option in TNT version 1.1 (Goloboff et al. 2008; Goloboff and Catalano 2016). The analysis produced a single most parsimonious tree with a tree length of 70 steps, a consistency index of 0.78, and a retention index of 0.86. Data matrix and 3D PDF files are provided in SOM.
For the homology of “symmetrodontan” cusps we follow Rougier et al. (2003b).
Systematic palaeontology
Mammalia Linnaeus, 1758
Trechnotheria McKenna, 1975
Spalacotheriidae Marsh, 1887
Genus Cifellitherium nov.
Zoobank LSID: urn: lsid:zoobank.org:act:C4591BC4-B0BF-4AE3-9C 4B-22476E353BE4
Etymology: After Richard L. Cifelli, in acknowledgement of his outstanding contributions to the knowledge of Mesozoic mammals, and therium, latinized version of the Greek θηρίον, a wild animal.
Type species: Cifellitherium suderlandicum sp. nov.; see below.
Diagnosis.—As for the type and only known species.
Stratigraphic and geographic range.—Barremian–Aptian (Lower Cretaceous), northwestern Germany (Europe).
Cifellitherium suderlandicum sp. nov.
Figs. 3–5.
Zoobank LSID: LSID urn:lsid:zoobank.org:act:4195BDBD-1C75-4D 0D-899D-252B1D2B7266
Etymology: From the medieval name Suderlande for the Sauerland region of Germany where Cifellitherium was found.
Holotype: WMNM P82305, right lower molar.
Type locality: Fissure filling within the Busche quarry near Balve, Sauerland, North Rhine-Westphalia, Germany.
Type horizon: Barremian–Aptian (Lower Cretaceous).
Material.—Holotype and left upper molars (WMNM P82308, P82302) from the type locality and horizon.
Diagnosis.—Medium to large sized spalacotheriid. Referred to Spalacotheriidae based on the combination of the following characters: acute angulation of principal cusps of upper (ranging from 40° to 54° on different teeth) and lower (55°) molars (derived); postvallum/prevallid shearing surfaces complete, extending on entire respective aspects on the upper and lower molars (derived); distinctive cusp B and well developed hook-like parastyle present (plesiomorphic); pronounced lingual cingulum; lower molar asymmetrically high crowned (labial side higher than lingual) with well developed proto-, para-, and metaconid; single distal talonid cusp d small and without heel, sitting at the lingual side of trigonid (derived); distal cuspule d overlapping labially with mesial cuspule e of the succeeding molar (derived); only one small mesial cingular cusp e present, distinct and in lingual position (derived); prominent lingual and labial cingulid, the latter interrupted above the interradical notch (derived).
Differs from Spalacotheridium Cifelli, 1990, primary trigon by the retention of three distinctive cusps (paracone, cusp B, and cusp C), and a less strongly compressed primary trigon on the upper molars, and by the interruption of the labial cingulid on the lower molars. Differs from Spalacolestes Cifelli and Madsen, 1999, and Symmetrodontoides Fox, 1976, by the retention of three distinctive cusps (paracone, cusp B, and cusp C), and a less strongly compressed primary trigon on the upper molars, and by the alignment of the paraconid with the metaconid as well as the interruption of the labial cingulid on the lower molars. Differs from Anebodon Bi, Zheng, Meng, Wang, Robinson, and Davis, 2016, and Kiyatherium Maschenko, Lopatin, and Voronkevich, 2002, by more acute angulation of upper molars. Differs from Maotherium Rougier, Ji, and Novacek, 2003a, by more acute triangulation of upper molars and a smaller stylocone and a larger parastyle. Differs from Zhangheotherium Hu, Luo, Li, and Wang, 1997, by a smaller stylocone and a larger parastyle and presence of a complete lingual cingulum on the upper molars (Hu et al. 1997; Rougier et al. 2003a; Lopatin et al. 2010; Bi et al. 2016; Plogschties and Martin 2019). Differs from Spalacotherium Owen, 1854, by a smaller stylocone, a larger parastyle, and a complete lingual cingulum on the upper molars and the interruption of the labial cingulid on the lower molars (labial cingulid interrupted in S. evansae). Differs from Spalacotheroides Patterson, 1955, by the presence of a complete lingual cingulum on the upper molars (Cifelli and Madsen 1999). Differs from Yaverlestes Sweetman, 2008, by double size and presence of a cingulum on upper molars. Differs from Heishanlestes Hu, Fox, Wang, and Li, 2005, Lactodens Han and Meng, 2016 (25% smaller), Aliaga Cuenca-Bescós, Canudo, Gasca, Moreno-Leanza, and Cifelli 2004, and Symmetrolestes Tsubamoto and Rougier in Tsubamoto et al., 2004 (half the size) by the interruption of the labial cingulid on lower molars. Differs from Infernolestes Cifelli, Davis, and Sames, 2014, by a dorsally flexed lingual cingulid and in having the paraconid/metaconid not closely appressed to the protoconid.
Measurements (in mm).—WMNM P82308, upper molar: length 1.45; width: 1.53. WMNM P82302, upper molar: length: 1.60; width: 1.45. WMNM P82305, lower molar (holotype): length: 1.53; width: 1.00.
Description.—Upper molars: In occlusal view, the primary trigon forms an isosceles triangle with the lingual angle varying from 40° (WMNM P82308) to 54° (WMNM P82302) (Figs. 3A1 and 4A1). This variable angle is dependent on the position of the upper molar along the tooth row, which is known for other spalacotherioids (Rougier et al. 2003a; Li and Luo 2006; Chen and Luo 2008; Han and Meng 2016). The upper molars have a well developed ectoflexus on the labial side (Figs. 3A1 and 4A1). The crowns of both upper molars are unilaterally hypsodont, being lingually higher than labially as in Akidolestes Li and Luo, 2006, and are moderately worn. By far the highest cusp is the paracone, separated from the second highest cusp B by a sharp narrow notch on the paracrista (Fig. 3A3). Stylocone and cusp C are smaller than cusp B; cusp C is separated from the paracone by a narrow v-shaped notch (Figs. 3A5 and 4A6). The metastylar area is damaged in both upper molars. The paracone is triangular in cross section (occlusal view) with a convex lingual and a flat labial side bearing a low vertical bulge. In WMNM P82302, this bulge extends into the primary trigon basin where it soon fades. Within the trigon basin of WMNM P82302, there is a small, linguo-labially elongated, isolated enamel rising; in WMNM P82308 the primary trigon basin is almost flat and bears only two tiny enamel knots. The paracone is oriented vertically and its apex is worn in both specimens. The small stylocone at the labial end of the paracrista is vertically oriented and bears a small, mesio-distally extended wear facet (Figs. 3A1, A4, 4A1). Distally from the deepest point of the ectoflexus is a median cusp which is one third of the size of the stylocone and also bears a small, mesio-distally extended wear facet. The paracrista and metacrista are robust and considerably worn. The parastyle is well developed and hook-like; at the mesial flank it is indented for accommodation of the metastyle of the preceding molariform (upper molar interlock) (Figs. 3A1, A3, 4A1, A4). The parastyle is worn, and the wear facet along the paracrista is confluent with the wear facet of the parastyle before reaching the stylocone. Like the paracrista, the metacrista is considerably worn and interrupted at the base of the paracone by a narrow v-shaped notch that separates cusp C from the paracone. On the mesial, lingual, and distal sides the crown is surrounded by a sharp, partially crenelated cingulum (Figs. 3A1, A5, A6, 4A1, A2, A6). Although not preserved completely in any of the specimens, the cingulum is assumed to be uninterrupted based on the almost complementary preservation in both specimens. In WMNM P82302 the mesial part of the cingulum is largely preserved (Fig. 3A1, A3); it rises to the stylocone and bears a groove-like linguo-labially oriented wear facet. The distal flank of the paracone and the distal side of the more labial part of the crown bear shiny wear facets with faint striations oriented about 45° labio-cervically in relation to the occlusal plane. WMNM P82302 shows an additional small wear facet linguocervically of the cingulum (less clear in the holotype specimen). On the mesial flank of the paracone and the mesial sides of the more labial parts of the crown, there are also wear facets detectable with striations oriented 45° labio-cervically, but less clear than on the distal side. The upper molars have two roots, of which in both specimens only the distal root is preserved (Figs. 3A2, 4A3). The distal root is mesio-distally compressed and straight; a mesio-distal compression can also be assumed for the mesial root according to its preserved base. The lesser degree of mesio-distal compression of the distal root and the longer crown suggest that WMNM P82302 derives from a more anterior position in the tooth row than WMNM P82308.
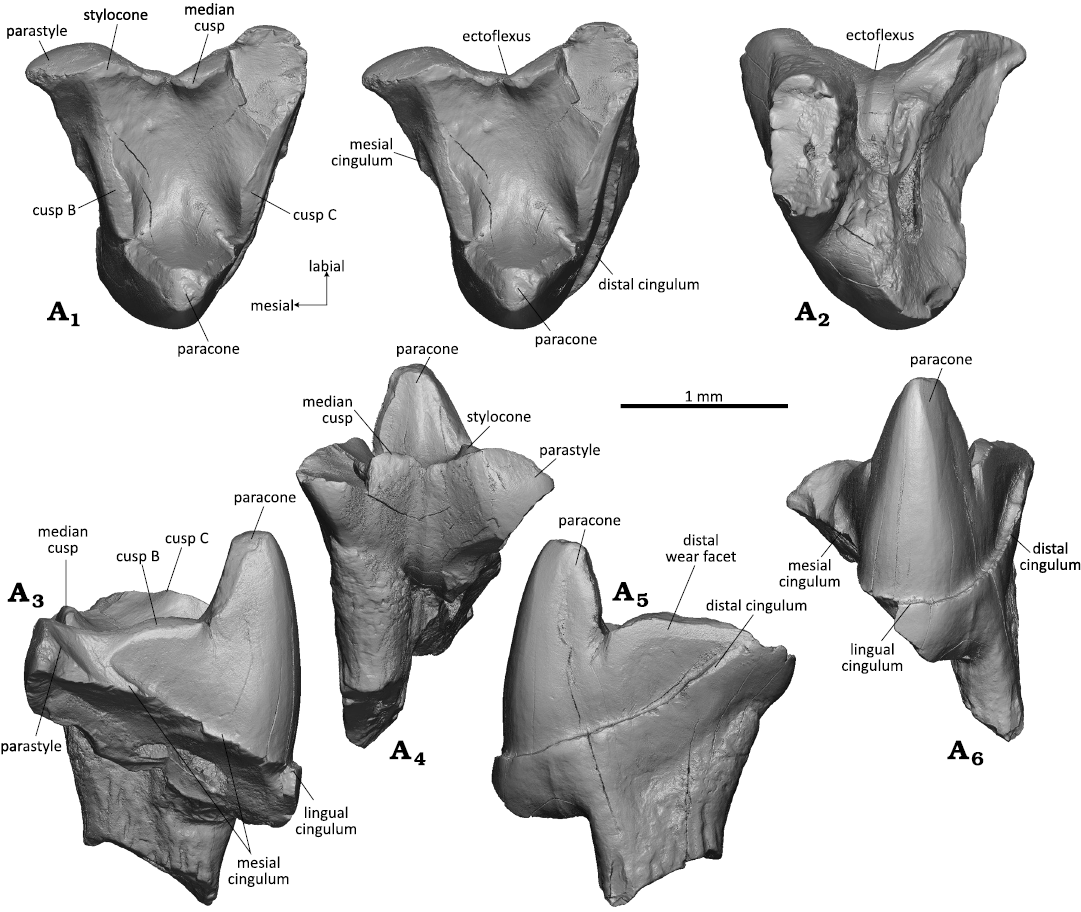
Fig. 3. Spalacotheriid mammal Cifellitherium suderlandicum gen. et sp. nov., WMNM P82308 from Busche Quarry near Balve, North Rhine-Westphalia, Germany, Barremian–Aptian (Lower Cretaceous). Right upper molar in occlusal (stereopair, A1), dorsal (A2), mesial (A3), labial (A4), distal (A5), and lingual (A6) views.
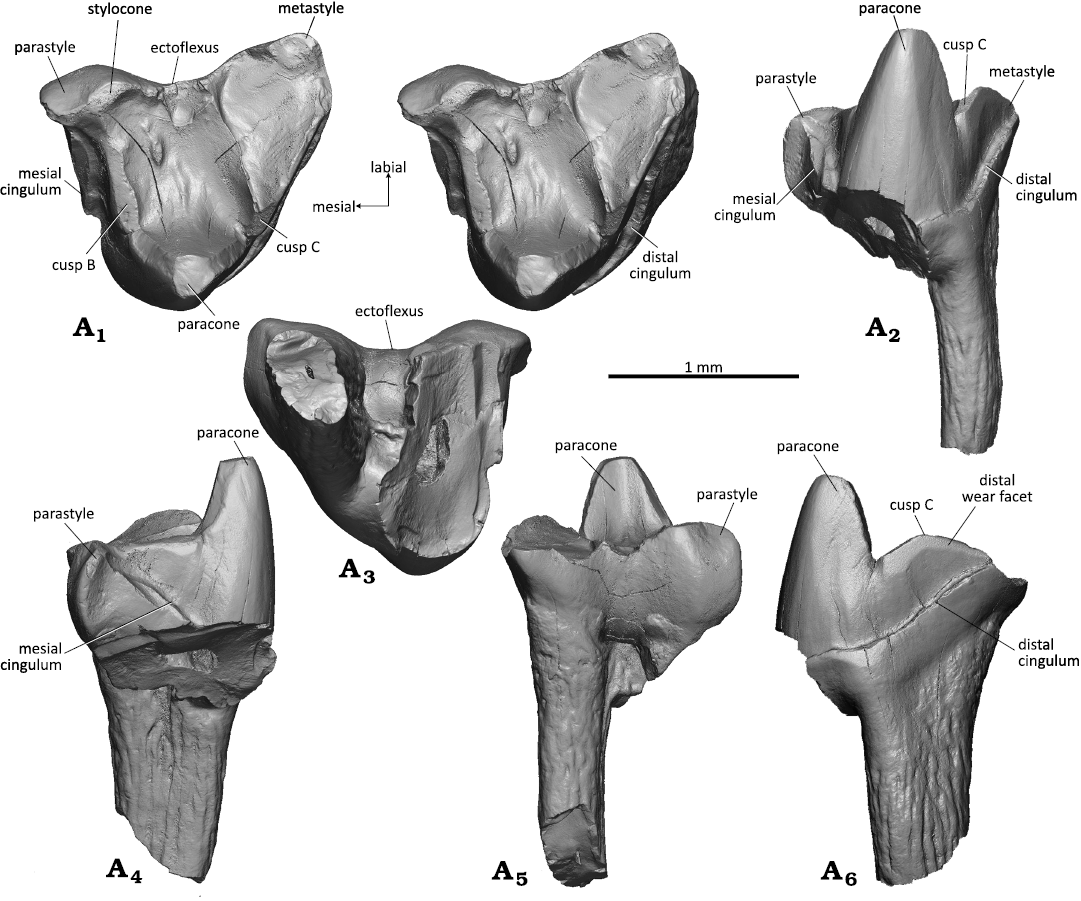
Fig. 4. Spalacotheriid mammal Cifellitherium suderlandicum gen. et sp. nov., WMNM P82302 from Busche Quarry near Balve, North Rhine-Westphalia, Germany, Barremian–Aptian (Lower Cretaceous). Right upper molar in occlusal (stereopair, A1), lingual (A2), dorsal (A3), mesial (A4), labial (A5), distal (A6) views.
Right lower molar: The crown in WMNM P82305 (holotype) has the shape of an isosceles triangle and the main cusps of the trigonid are arranged in an angle of about 55° (acute angulation); the lingual side (base line of the triangle) is straight (Fig. 5A1). The lower molar crown is moderately high, somewhat in between Zhangheotherium/Maotherium (Rougier et al. 2003a; Ji et al. 2009; Plogschties and Martin 2019) and Spalacotherium (Cifelli and Madsen 1999). Highest cusp is the protoconid which is slightly bent distally (Fig. 5A3, A5). The protoconid has a triangular cross section with slightly convex mesiolabial and distolabial flanks and a slightly concave lingual flank with a shallow convex bulge running down from the apex into the trigonid basin. Second highest cusp is the metaconid that reaches two thirds of the protoconid height. It has a flat distal flank whereas the lingual, and mesiolingual sides are rounded. It is curved distally and separated from the protoconid by a sharp notch (Fig. 5A1, A3, A4). The apex of the metaconid bears a round wear facet that continues along the metacristid. The paraconid is about half as high as the protoconid and curved mesially (Fig. 5A1, A3, A6). It has a conical shape without pre- and postparacristid. Apically the paraconid bears a drop-shaped wear facet that continues onto the paracristid. The notch between paraconid and protoconid is as deep as that between protoconid and metaconid; it appears less pronounced because the paraconid is more worn than the metaconid. The bases of paraconid and metaconid are confluent with that of the protoconid; the paraconid and the metaconid are clearly separated from each other, resulting in a small, lingually open trigonid basin with lingually sloping floor. The trigonid is surrounded by a marked cingulid which is interrupted above the interradical notch on the labial side (Fig. 5A1, A5). There is a well developed cusp d in the distolingual corner of the tooth crown (Fig. 5A1, A3, A5). It is triangular in shape with flat lingual and rounded mesial side. Its distolabial side is concave for accommodation of cusp e of the succeeding molariform (lower molar interlock). The prominent distal edge of cusp d is crest-like and separates the flat lingual side from the concave accommodation concavity. Its tip bears a small oval wear facet and continues into the crest-like distal portion of the cingulid. The lingual portion of the cingulid is less prominent and merges into the base of cusp d. Mesial cusp e (Fig. 5A1, A3, A5, A6) is about half the size of cusp d and is integrated into the mesial cingulid as a swelling at its lingual end. Its tip bears a drop-shaped wear facet that continues onto the mesial cingulid. The mesial and lingual cingulids are strongly developed and are crenelated. They slope in labial direction and taper out before reaching the interradical notch on the labial side. The lingual cingulid (Fig. 5A3) is somewhat less prominent and also crenelated; it is uninterrupted and rises slightly at the notch between paraconid and metaconid. The roots are slightly curved lingually and the mesial root is somewhat stronger than the distal one (Fig. 5A2–A6). The cross section is triangular for the mesial root and square for the distal root; in the latter the apex is missing. Both roots are concave in the upper part of their inner sides, the mesial root on the distal side and the distal root on the mesial side.
Stratigraphic and geographic range.—Type locality and horizon only.
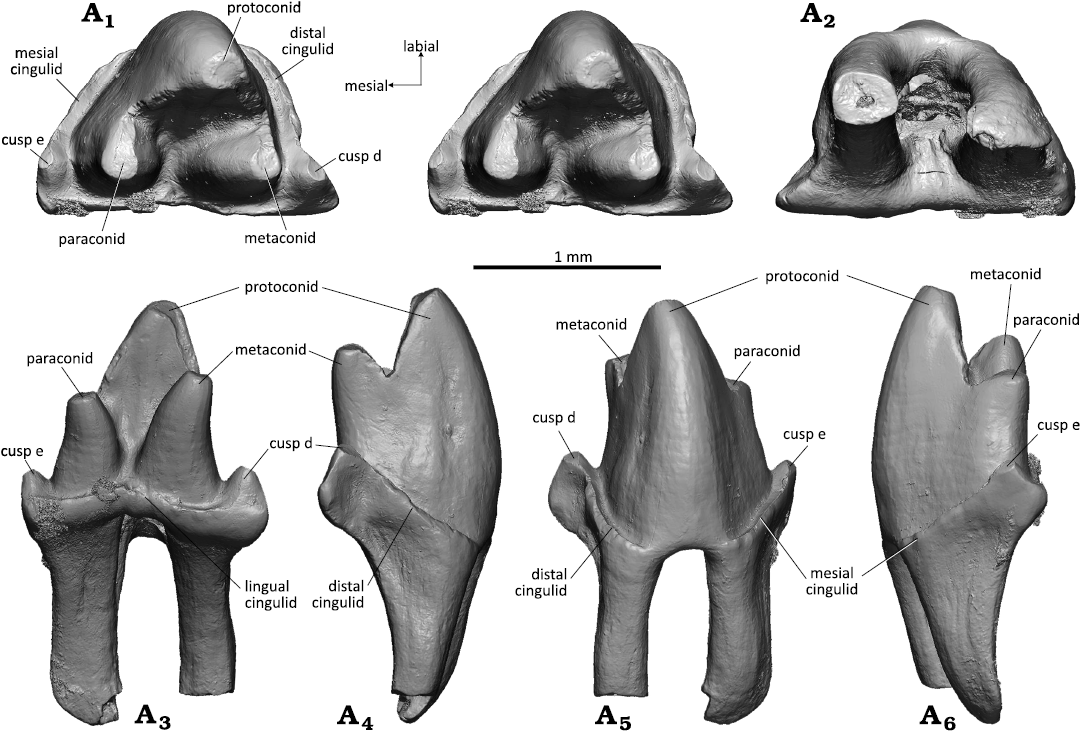
Fig. 5. Spalacotheriid mammal Cifellitherium suderlandicum gen. et sp. nov., WMNM P82305 (holotype) from Busche Quarry near Balve, North Rhine-Westphalia, Germany, Barremian–Aptian (Lower Cretaceous). Right lower molar in occlusal (stereopair, A1), ventral (A2), lingual (A3), distal (A4), labial (A5), and mesial (A6) views.
Cladotheria McKenna, 1975
Dryolestida Prothero, 1981
Dryolestidae Marsh, 1879
Genus Minutolestes nov.
Zoobank LSID: urn:lsid:zoobank.org:act:240D4DE4-0EF3-4959-922F- BF2C8B4E6F61
Etymology: From Latin minutus, tiny, referring to the small size and Greek λησήτς, robber.
Type species: Minutolestes submersus sp. nov.; see below.
Diagnosis.—As for the type and only known species.
Stratigraphic and geographic range.—Barremian–Aptian (Lower Cretaceous), northwestern Germany (Europe).
Minutolestes submersus sp. nov.
Figs. 6–8.
Zoobank LSID: urn:lsid:zoobank.org:act:7DD409B2-0C58-4AAA-875 C-7F3F881AAD64
Etymology: From Latin submersus, sunken, referring to the origin of the fossils from a sinkhole.
Holotype: WMNM P82306, left upper molar.
Type locality: Fissure filling within the Busche quarry near Balve, Sauerland, North Rhine-Westphalia, Germany.
Type horizon: Barremian–Aptian (Lower Cretaceous).
Material.—Holotype and left upper (WMNM P82304) and lower (WMNM P82307) molars from the type locality and horizon.
Diagnosis.—Tiny dryolestid with small stylocone and flat primary trigon basin. Referred to Dryolestidae based on the following derived characters: upper molars much wider than long, stylocone large, median cusp present, metaconid more than half of protoconid height, and mesial root much larger than distal root on lower molars.
Differs from Dryolestes Marsh 1878, Laolestes Simpson 1927, Krebsotherium Martin 1999, and Portopinheirodon Martin 1999, by smaller size and flat primary trigon basin without median ridge between stylocone and paracone (Laolestes, Portopinheirodon, Lakotalestes Cifelli, Davis, and Sames 2014, Groebertherium Bonaparte, 1986) or bulge between metacone and stylocone (Dryolestes, Krebsotherium) on upper molars. Differs from Amblotherium Owen, 1871 by less prominent (not hook-like) parastyle on the upper molars and blunt metaconid. Differs from Anthracolestes Averianov, Martin, and Lopatin, 2014, by a blunt metaconid on the lower molars. Differs from Hercynodon Martin, Averianov, Schultz, Schwermann, and Wings, 2021, by presence of a metacone. Differs from Achyrodon Owen, 1871, by the presence of cups along the metacrista on the upper molars. Similar to Crusafontia Henkel and Krebs, 1969 by the following characters: small size, shallow ectoflexus on upper molars present, metacone reduced (derived), labial cingulid on lower molars (derived). Differs from Crusafontia by a weaker ectoflexus and a stronger metacone on upper molars and a blunt metaconid on the lower molars.
Measurements (in mm).—WMNM P82305, upper molar (holotype): length: 0.75; width: 0.86. WMNM P82304, upper molar: length: 0.65; width: 0.97. WMNM P82307, lower molar: length: 0.60 (without talonid); width: 0.61.
Description.—Upper molars: WMNM P82306 (holotype) and attributed specimen WMNM P82304 (Figs. 6, 7) are unilaterally hypsodont with the crown being higher lingually than labially. In occlusal view, the crown forms an asymmetrical triangle with the distal border longer than the mesial border in the holotype and vice versa in WMNM P82304 (Figs. 6A1, 7A1). In the holotype, the labial border is slightly shorter than the distal border, and in WMNM P82304 it has only half the length of the distal border. It is almost straight in the holotype and exhibits a slight labial bulging in the stylocone area in WMNM P82304. The tooth crown is compressed mesio-distally and the angulation of the principal cusps is 43° in the holotype specimen and only 25° in WMNM P82304. Judging by the degree of compression, the holotype specimen derives from an anterior position and WMNM P82304 from a middle position in the tooth row.
Of the three primary trigon main cusps, the paracone is the highest. Its outline is triangular in cross section with flat mesial and distal flanks, a rounded lingual angle, and a slightly concave labial flank. The paracone bears an oval apical wear facet with exposed dentine that extends along the metacrista (Fig. 6A1, A4). The stylocone is broken on the holotype but was likely the second largest cusp. In WMNM P82304, the stylocone is present and small (Fig. 7A1, A3–A6). It is worn (likely by corrosion) apically and on the mesial and labial sides; it is the second-highest cusp of the primary trigon. The metacone, third-largest cusp, is a prominent swelling on the metacrista that bulges towards the primary trigon basin (Figs. 6A1, A4, 7A1) and is separated from the paracone by a sharply indented notch. The metacone bears a large linguo-labially extended wear facet (Figs. 6A1, A6, 7A1). Labially of the metacone, cusp C is represented as a slight elongate swelling on the metacrista (present in both upper molars) (Figs. 6A1, A3, A4, A6, 7A1). The parastyle is triangular in shape, bulging mesio-labially, and clearly separated from the primary trigon by a marked lingual notch (Fig. 6A1, A3, A4, A5). It bears an oval apical wear facet in the holotype specimen and is corroded in WMNM P82304. The metastyle (Fig. 6A1, A3, A4, A6) is much smaller and bears a large oval wear facet in the holotype and is strongly corroded in WMNM P82304. In the middle of the labial rim there is a tiny median cusp (Fig. 6A1) with an apical wear facet; from the median cusp extends a small worn crest in distal direction in the holotype. In WMNM P82304 the median cusp and the area distally from it including the metastyle are corroded. Paracrista (so far preserved) and metacrista are worn with exposure of dentin. The mesial and distal flanks of the crown bear fine striations that are oriented about 45° in labio-cervical direction. The primary trigon basin is flat without any crest or bulge.
There are three roots, of which two are broken off in the holotype. According to the broken base with opened pulp cavity, largest root was the one supporting the paracone, whereas the roots supporting the para- and metastyle were much smaller (Fig. 6A2). The root supporting the parastyle is largely intact and only its tip is missing (Fig. 6A2–A6). The root is slightly bent disto-labially and has an oval cross section with a slight mesio-distal compression. In WMNM P82304 all three roots are partially preserved. The larger root supporting the paracone is almost complete. It is curved mesio-labially (Fig. 7A3–A6) and has a sub-quadrangular outline in cross section (Fig. 7A2). Of both smaller roots supporting the para- and metastyle only the proximal confluent portion is preserved (Fig. 7A2–A6); the distal parts are broken off. The incompletely separated roots have oval cross sections with mesio-distal compression. A slight curvature of the remaining portion of the roots indicates a former labial bending.
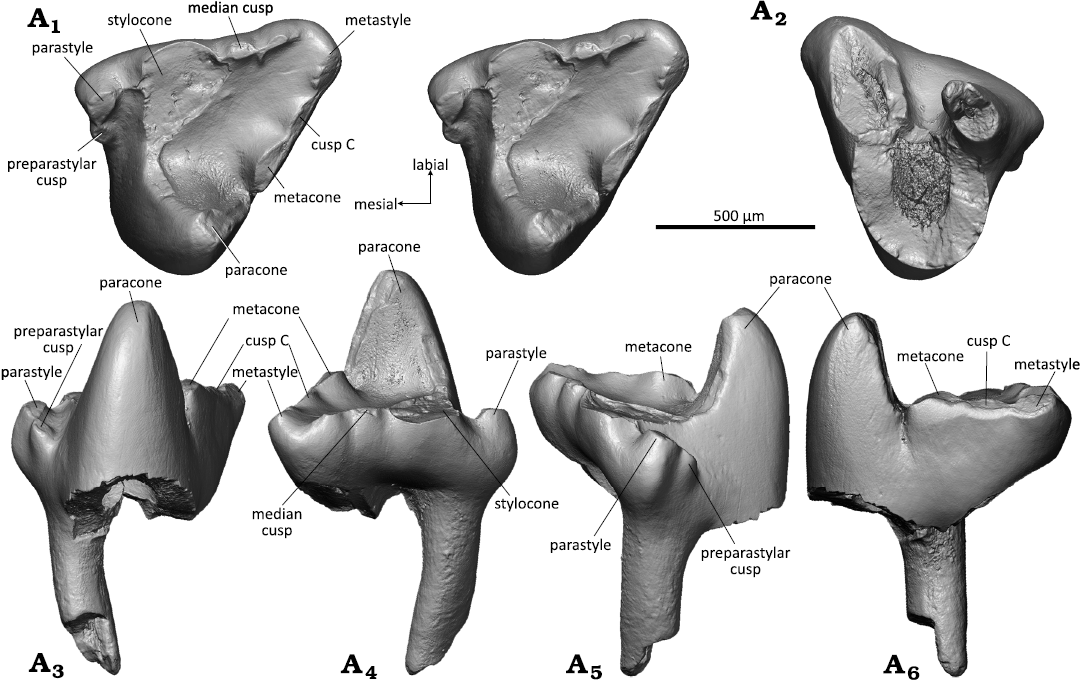
Fig. 6. Dryolestid mammal Minutolestes submersus gen. et sp. nov., WMNM P82306 (holotype) from Busche Quarry near Balve, North Rhine-Westphalia, Germany, Barremian–Aptian (Lower Cretaceous). Left upper molar in occlusal (stereopair, A1), dorsal (A2), lingual (A3), labial (A4), mesial (A5), and distal (A6) views.
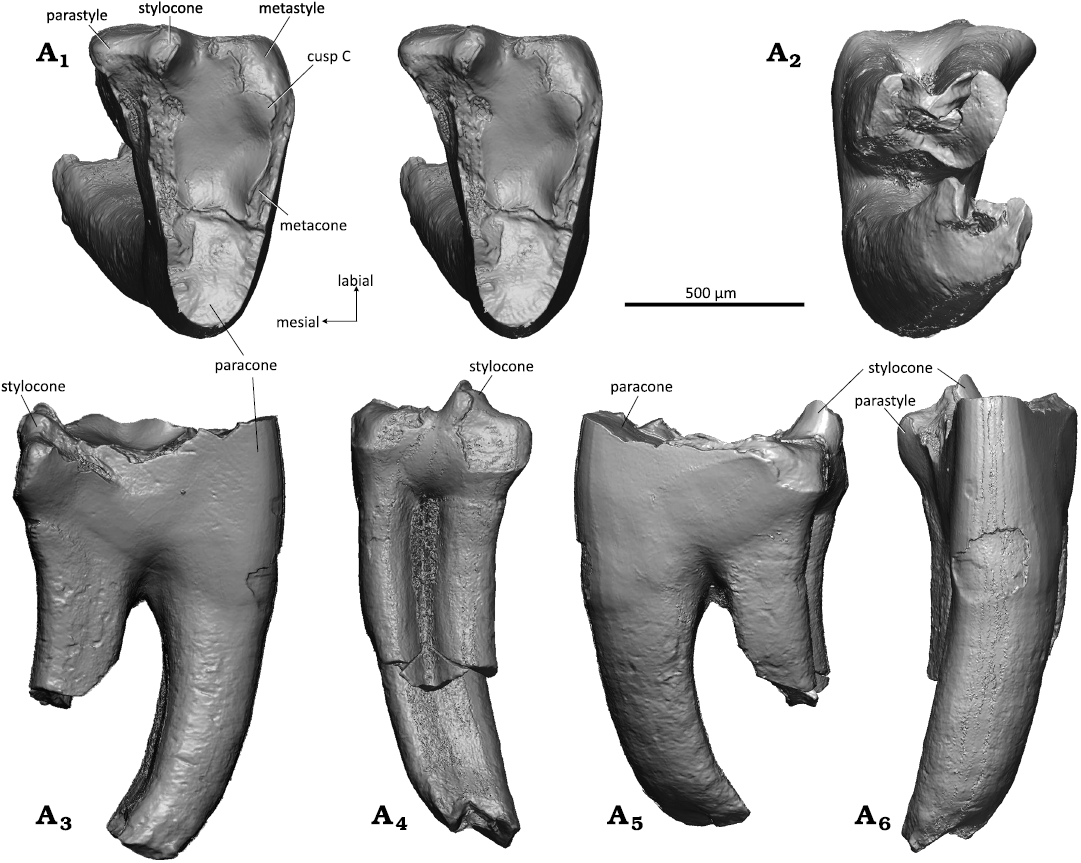
Fig. 7. Dryolestid mammal Minutolestes submersus gen. et sp. nov., WMNM P82304 from Busche Quarry near Balve, North Rhine-Westphalia, Germany, Barremian–Aptian (Lower Cretaceous). Left upper molar in occlusal (stereopair, A1), dorsal (A2), mesial (A3), labial (A4), distal (A5), and lingual (A6) views.
Left lower molar: The left lower molar WMNM P82307 (Fig. 8) is unilaterally hypsodont with the labial part higher than the lingual. The part of the tooth where a talonid would be expected is broken off. On the crown, the three main cusps, proto-, para-, and metaconid are preserved. The protoconid (Fig. 8A1, A3–A6) is the highest cusp and has a triangular cross section; it has a flat mesiolingual and slightly concave (in the lower part) distal side and a slightly convex mesiolabial side. The distal side of the protoconid shows faint striations that are oriented at about 45° in labio-cervical direction in relation to the occlusal plane (Fig. 8A6, inset). The protoconid is separated from the para- and metaconid by sharp notches. Second largest cusp is the metaconid (Fig. 8A1, A3–A6), with a cross section that is quadrangular in outline; the lingual and distal sides are flat, the labial side is slightly concave, and the anterior side slightly convex. At the distal side of the metaconid, a small portion of the distal metacristid is preserved (Fig. 8A6). The upper third of the metaconid is broken. The base of the metaconid sits close to the base of the protoconid; both cusps are separated by a sharp notch in the metacristid. The paraconid (Fig. 8A1, A3–A5) is crest-like, compressed mesio-distally and tapering labially. In lingual view, the paraconid appears pointed and inclined mesially; it attains about half the height of the protoconid. The trigonid is moderately compressed mesio-distally and has an angle of 38°. Due to the close approximation of protoconid and metaconid, the basin is largely confined to the mesial portion of the trigonid. The talonid is broken off. On the mesial side of the paraconid, close to its base, there is a short mesial cingulid (Fig. 8A3, A4) which is inclined cervically and exhibits slight wear in its middle part. The cingulid becomes weaker in labial direction, but continues along the preserved part of the labial side; the distal side is broken. The paracristid is moderately worn along the paraconid, and the lingual portion of the metacristid is broken off including the tip of the metaconid. The protoconid bears a small oval wear facet that continues along the preserved part of the metacristid. Of the mesial, much larger root, the anterior portion is preserved (Fig. 8A2, A4, A6) whereas its distal part and the posterior small root are broken off together with the talonid and the disto-cervical part of the protoconid, exposing the large pulp cavity Fig. 8A2, A3, A6).
Stratigraphic and geographic range.—Type locality and horizon only.
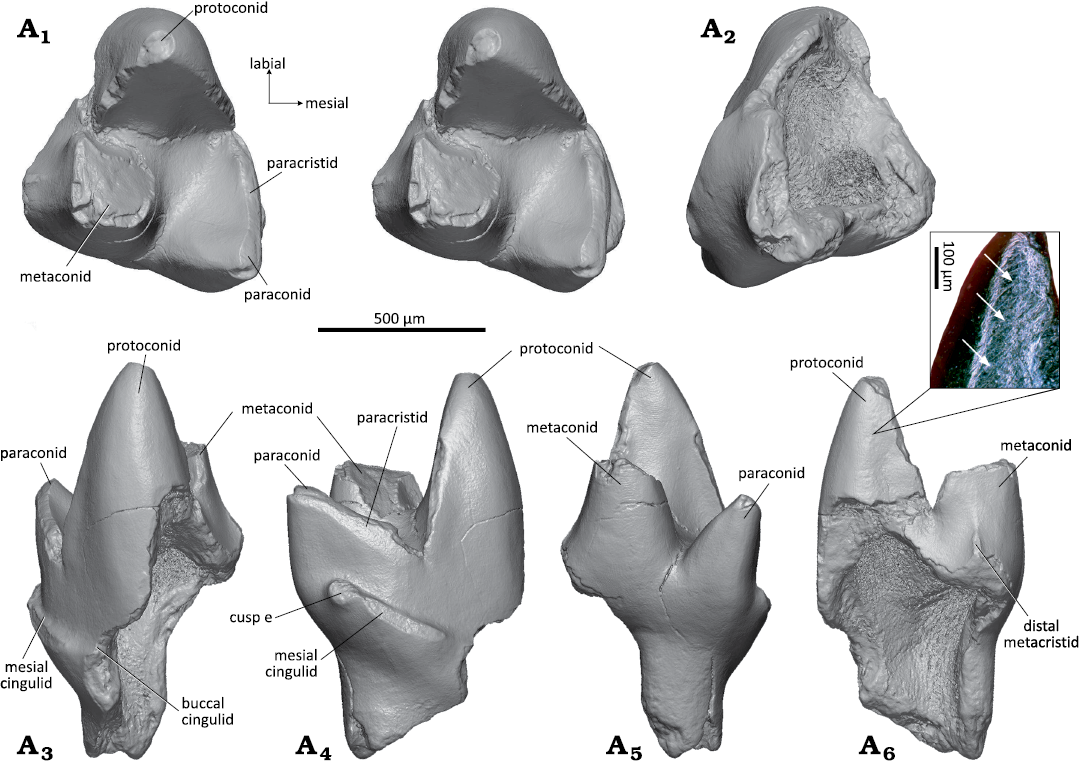
Fig. 8. Dryolestid mammal Minutolestes submersus gen. et sp. nov., WMNM P82307 from Busche Quarry near Balve, North Rhine-Westphalia, Germany, Barremian–Aptian (Lower Cretaceous). Left lower molar in occlusal (stereopair, A1). ventral (A2), labial (A3), mesial (A4), lingual (A5), and distal (A6) views. Enlarged inset of protoconid (stereo microscopic image under sided light) showing striations (arrows).
Genus Beckumia nov.
Zoobank LSID: urn:lsid:zoobank.org:act:C93A0BA6-7DA9-475D-8D 45-11FECE0E6049
Etymology: After Beckum, urban district of the town of Balve, next to the Busche Quarry where Beckumia gen. nov. was found.
Type species: Beckumia sinemeckelia sp. nov.; see below.
Diagnosis.—As for the type and only known species.
Stratigraphic and geographic range.—Barremian–Aptian (Lower Cretaceous), northwestern Germany (Europe).
Beckumia sinemeckelia sp. nov.
Figs. 9, 10.
Zoobank LSID: urn:lsid:zoobank.org:act:738E1658-6C43-4516-B3C3-2D23D15856B2
Etymology: From Latin sine, without, and Meckel, referring to the absence of Meckel’s groove.
Holotype: WMNM P82301, left mandible with strongly worn m2–m5 present, other teeth fallen out or broken off postmortem.
Type locality: Fissure filling within the Busche quarry near Balve, Sauerland, North Rhine-Westphalia, Germany.
Type horizon: Barremian–Aptian (Lower Cretaceous).
Material.—Holotype and left lower premolar (WMNM P82310) from the type locality and horizon.
Diagnosis.—Referred to Dryolestidae based on the following derived characters: roots of lower molars extremely disparate with mesial root much larger and stronger than distal root; mandibular border medially higher than laterally, molars unilaterally (labially) hypsodont; lower molars strongly compressed mesio-distally with small unicuspid talonid cusp; mandibular body deep, with s-shaped curvature in dorsal view (Fig. 9). Dental formula 4i, 1c, 4p, 8m; the first lower incisor (i1) implanted almost horizontally in alveolus. Incisors single-rooted, canine, premolars and molars double-rooted, ultimate molar (m8) single-rooted; coronoid process rising almost vertically; angular process well developed and pointing backwards; well developed masseteric and pterygoid fossae; two mental foramina below p1 and m1; no trace of coronoid or splenial detectable.
Differs from all other known dryolestids by complete reduction of Meckel’s groove (derived). Differs further from Crusafontia cuencana Henkel and Krebs, 1969, by 50% larger molars and absence of a labial cingulid as evident on m5.
Measurements (in mm).—Preserved length of mandible WMNM P82301: 23.0; height of corpus below ultimate molar: 3.5; m2, length*: 1.0, width*: 1.1; m3*, length*: 1.0, width*: 1.3; m4, length*: 1.2, width*: 1.5. WMNM P82310, lower p4: length: 0.72; width: 0.3 (* as preserved).
Description.—WMNM P82301 is a left mandible (Fig. 9) with the dorsal part of the coronoid process, the condylar process, and the distal tip of the angular process broken off. Most of the teeth have fallen out postmortem. According to the present alveoli, the dental formula is 4i, 1c, 4p, and 8m (Fig. 9A5). The single roots of the incisors have an oval shape in cross section (long axis of cross section oriented linguo-labially). Judging by the alveoli, i1 was inserted almost horizontally, i2 at 45°, and i3 and i4 at 70° in the mandible. The canine was double-rooted with both roots of equal size. All four premolars had two roots with a round cross section. In p1 and p2, the anterior root was slightly larger than the posterior; in p3 the posterior root was slightly larger; and in p4 both roots were of equal size. The sizes of the alveoli suggest that p4 was the largest, with p3, p1, and p2 being successively smaller. Except for the ultimate, the molars are double-rooted with dryolestid-typical large anterior and tiny posterior root; m8 was single-rooted with only the anterior root present. The m1 and m6–m8 are not present (posterior root of m1 still sitting in the alveolus). The heavily worn m2–m5 are still in place (m5 partly broken), their crowns are almost entirely worn away with the pulp cavities open. The molars are more strongly worn on the labial side, resulting in an oblique occlusal surface. On the lingual side, parts of the enamel-covered crowns are still present. The paraconid (best preserved on m4) is erect and separated from the metaconid by a narrow v-shaped notch. The paraconid and the metaconid taper apically and it can be inferred that they were pointed, although their tips are removed by the oblique wear. The single-cusped talonids are very short. On m5, a small cervical portion of the enamel-covered labial part of the crown is preserved which lacks any trace of a labial cingulid. The extremely worn down molars with extensive transversely oriented striations suggest a senile individual. As typical for dryolestids, the alveolar border is much higher on the lingual than on the labial side which is correlated with a unilateral (labial) hypsodonty of the molars.
The mandibular body is slender and has a slightly curved ventral border. There are two mental foramina, one at the position of the posterior root of p2 and one slightly smaller at the position of the posterior root of m1; both foramina are oval in longitudinal direction and sitting just below the alveolar border (Fig. 9A2). The external border of the mandibular body is convex with foveal bone surface (especially in the anterior part in front of the molars). The masseteric fossa is well developed with a strong anterior crest and shelf-like ventral border (Fig. 9A2). The coronoid process rises steeply (about 80° to the alveolar line) but its dorsal half (or more) is missing (Fig. 9A2). There is no trace of a coronoid facet. The angular process is well developed and pointing posteriorly (distal end is broken off) (Fig. 9A2, A4).
The lingual (internal) surface of the mandibular body is flat (Fig. 9A4). A large symphysis extends from the tip of the mandible to the posterior root of p2 with a strongly pitted surface. Above the symphysis, the mandibular surface is slightly concave and bears two tiny foramina below the canine. From the posterior end of the symphysis to the position of m3 a ventral edge extends along the mandibular body. The pterygoid fossa (Fig. 9A4) is well developed with a pronounced ventral edge. It bears a high oval mandibular foramen. Meckel’s groove is completely reduced and no traces of a splenial bone is detectable.
In dorsal view (Fig. 9A1), the mandibular body is somewhat bent with a slight labial curvature in the region of the anterior-most molars and the premolars which is exaggerated by a crack in that area.
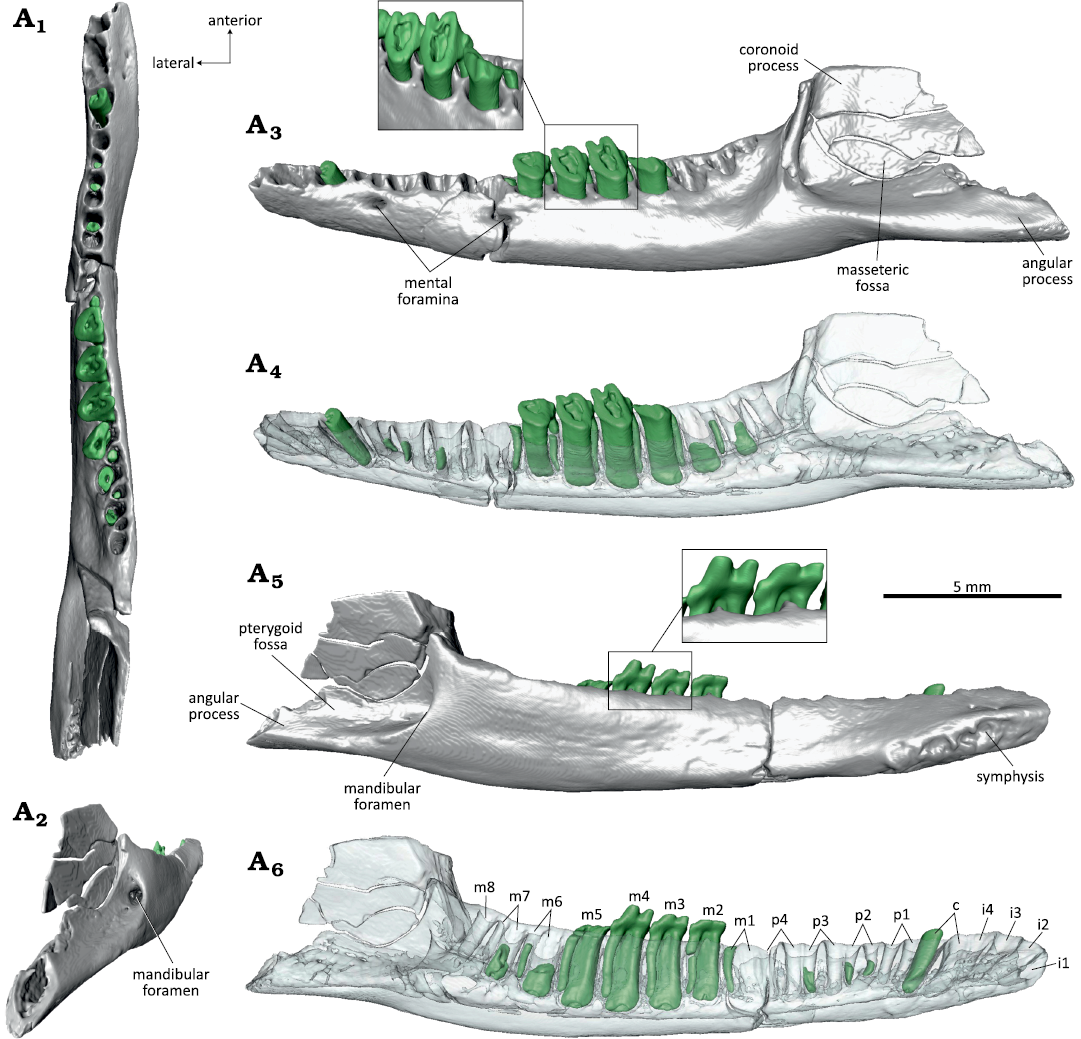
Fig. 9. Dryolestid mammal Beckumia sinemeckelia gen. et sp. nov., WMNM P82301 (holotype) from Busche Quarry near Balve, North Rhine-Westphalia, Germany, Barremian–Aptian (Lower Cretaceous). Left mandible in dorsal (A1), linguoposterior (A2), lateral (A3, A4), and medial (A5, A6) views. Inset in A3 showing enlarged m3–m5 in distolabial view, inset in A5 showing m3 and m4 in lingual view.
Left p4: WMNM P82309 (Fig. 10) has been identified as a lower p4 based on similarity to the p4 of Crusafontia cuencana (Henkel and Krebs 1969: fig. 1c–e; Martin 1998: fig. 4; Figs. 11 and 12). The crown of p4 is formed by a large main cusp (protoconid) and a mesial and distal cuspule. The main cusp is compressed labio-lingually and slightly bent lingually, and bears an apical wear facet with exposed dentin that continues on the lingual side (Fig. 10A1). The protoconid has a convex mesial side; the lingual side is slightly concave; and the distal side is straight. On the lingual side, the enamel is corroded centrally near the crown base. The mesial cuspule (Fig. 10A1, A3–A5) sits high on the mesial side of the protoconid, at about half of the main cusp’s height. It is separated from the protoconid by a pronounced notch and bears a round apical wear facet. The distal cuspule (Fig. 10A1, A3, A5, A6) sits much lower at the base of the protoconid. It is somewhat larger than the mesial cuspule and bears an oval, slightly lingually inclined apical wear facet. The distal cuspule is separated from the protoconid by a rounded notch. The premolar has two roots of which the distal root is slightly larger than the mesial root. Both roots are broken off, exposing the pulp cavities; the mesial root has a round and the distal root an oval (slightly labio-lingually compressed) cross section (Fig. 10A2).
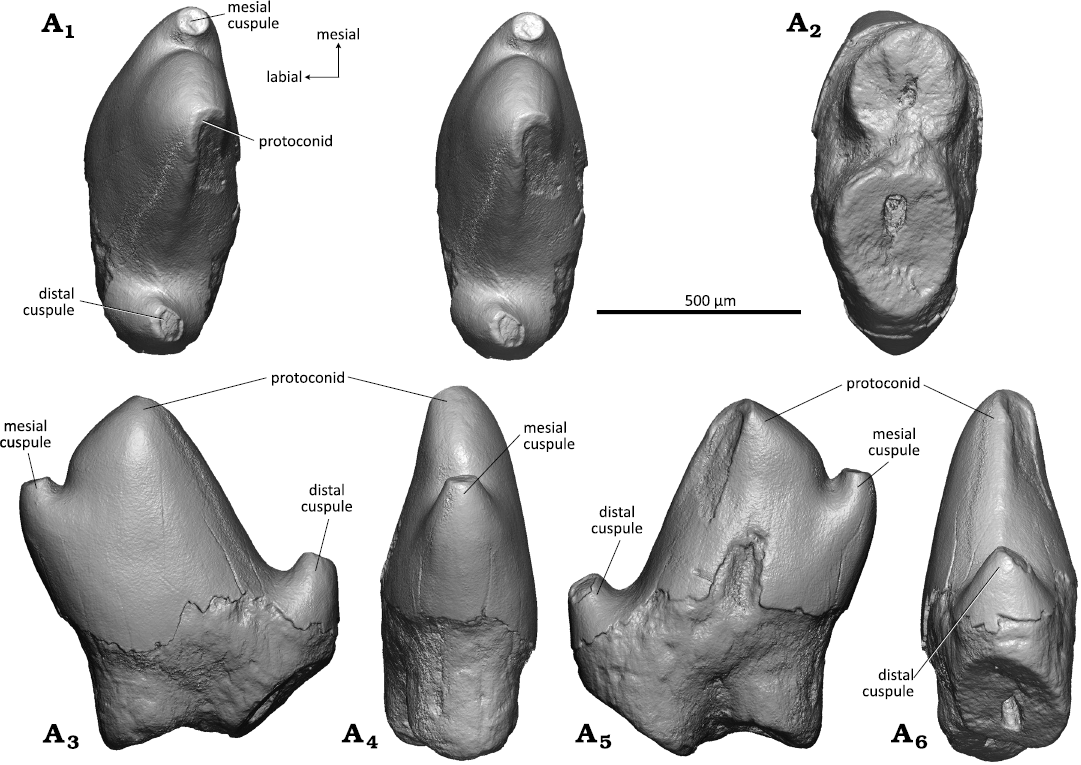
Fig. 10. Dryolestid mammal Beckumia sinemeckelia gen. et sp. nov., WMNM P82310 from Busche Quarry near Balve, North Rhine-Westphalia, Germany, Barremian–Aptian (Lower Cretaceous). Left lower p4 in occlusal (stereopair, A1), ventral (A2), labial (A3), mesial (A4), lingual (A5), and distal (A6) views.
Remarks.—The attribution of WMNM P82301 to Dryolestidae is unquestionable because the molar trigonids are strongly compressed mesio-distally, have a small lingually positioned talonid, and possess extremely uneven roots, which all are derived dryolestid characters. In lingual view, the molars are reminiscent to those of Crusafontia cuencana, showing a slender and pointed paraconid. They differ from the molars of C. cuencana by the absence of a buccal cingulid (evident on m5 of WMNM P82301; the respective area is worn off in the other molars) and a 50% larger size. The molars of WMNM P82301 are also twice in size of the molars of Minutolestes submersus gen. et sp. nov. Dryolestids have a diphyodont mode of tooth replacement, and permanent molars are morphologically clearly distinct from deciduous premolars (Martin 1997). Therefore, it can be ruled out that the small molariform teeth attributed to Minutolestes submersus gen. et sp. nov. derive from juvenile Beckumia individuals. The complete reduction of Meckel’s groove in the mandible from Balve is unique among Dryolestidae; in C. cuencana Meckel’s groove is short and reduced to the distal portion, but still clearly discernible (Henkel and Krebs 1969; Figs. 12 and 13). In MfN Uña 2 (paratype specimen), a crack is present in that area which artificially exaggerates Meckel’s groove, but in MfN Uña 1 (holotype specimen) it is preserved undisturbed. In both C. cuencana specimens (MfN Uña 1 and MfN Uña 2), the medial side of the mandibular body bears a longitudinal depression which is rather a postmortem phenomenon (diagenetic compression of the bone) than a further remnant of Meckel’s groove. Minutolestes submersus gen. et sp. nov. and C. cuencana lack a rough surface area that can be interpreted to be for the coronoid bone and any trace for accommodation of a splenial on the inner side of the mandible. By comparison, the coronoid facet and a possible “splenial” facet are still present in Late Jurassic dryolestids (e.g., Krebs 1969, 1971; Martin 1995, 1999). Like the mandible of C. cuencana, WMNM P82301 is rather slender for a dryolestid mandible. Size-wise the mandible of WMNM P82301 (preserved length 23 mm; corpus height below last molar 3.5 mm) corresponds to the adult paratype mandible of C. cuencana from Uña (preserved length 19.5 mm, portion anterior of posterior alveolus of canine missing; corpus height below last molar 3.7 mm (Henkel and Krebs 1969). The adult paratype mandible (MfN Uña 2) is about 50% larger than the juvenile holotype mandible (MfN Uña 1), but the teeth are of comparable dimensions. The holotype and paratype specimens of C. cuencana have three mental foramina (Henkel and Krebs 1969; Krebs 1971; Figs. 11A3 and 12A2) whereas there are only two in WMNM P82301.
Premolar WMNM P82310 closely resembles a left p4 from Galve attributed to Crusafontia cuencana (Galve Th 21) (Martin 1998: fig. 4), both in morphology and size (Galve Th 21, length: 0.73 mm, width 0.37 mm), and fits very well the dimensions of mandible WMNM P82301. In dryolestids, wear on the premolars occurs on the disto-lingual side rather than on the labial side as in most other mammals. This wear pattern regularly occurs in connection with pronounced apical wear which is typical for dryolestids (Martin 1999; Schultz and Martin 2011).
We offer a new interpretation for the premolar and molar count in the holotype specimen of Crusafontia cuencana (MfN Uña 1) based on µCT analysis (Fig. 11). In the original description, Henkel and Krebs (1969) interpreted the dental formula as 4i, 1c, 4p, 7m, with single-rooted p1 and p2 (only represented by alveoli). According to our re-interpretation the dental formula is 4i, 1c, 4p, 6m, with dP4 still in place (“m1” of Henkel and Krebs 1969) (Fig. 11A6). This tooth differs from the molars by a more elongated crown (145% of width compared to 133–126% in m1–m5), which is a typical trait of dryolestid deciduous premolars (Martin 1997). The shape of the two alveoli immediately behind the canine indicates that they belong to a double-rooted premolar (p1) and not to two single-rooted teeth as suggested by Henkel and Krebs (1969). Judging by the size of the alveoli, p1 was larger than p2, which is the smallest of the premolars. This is in accordance with the observation that p2 generally is the smallest premolar in dryolestids (e.g., Martin 1999). The stage of eruption of the premolars, p2 in process of eruption, p3 fully erupted, and dp4 still in place, is also in congruence with the earlier observed alternating mode of tooth replacement in dryolestids with p4 being the last premolar to erupt (Martin 1997). This new interpretation of premolar positions in the juvenile C. cuencana holotype specimen resolves the discrepancy to the premolar count of the adult paratype specimen (MfN Uña 2, Fig. 12) noticed by Henkel and Krebs (1969). According to Henkel and Krebs (1969) MfN Uña 2 possessed four double-rooted lower premolars which is in accordance with our analysis.
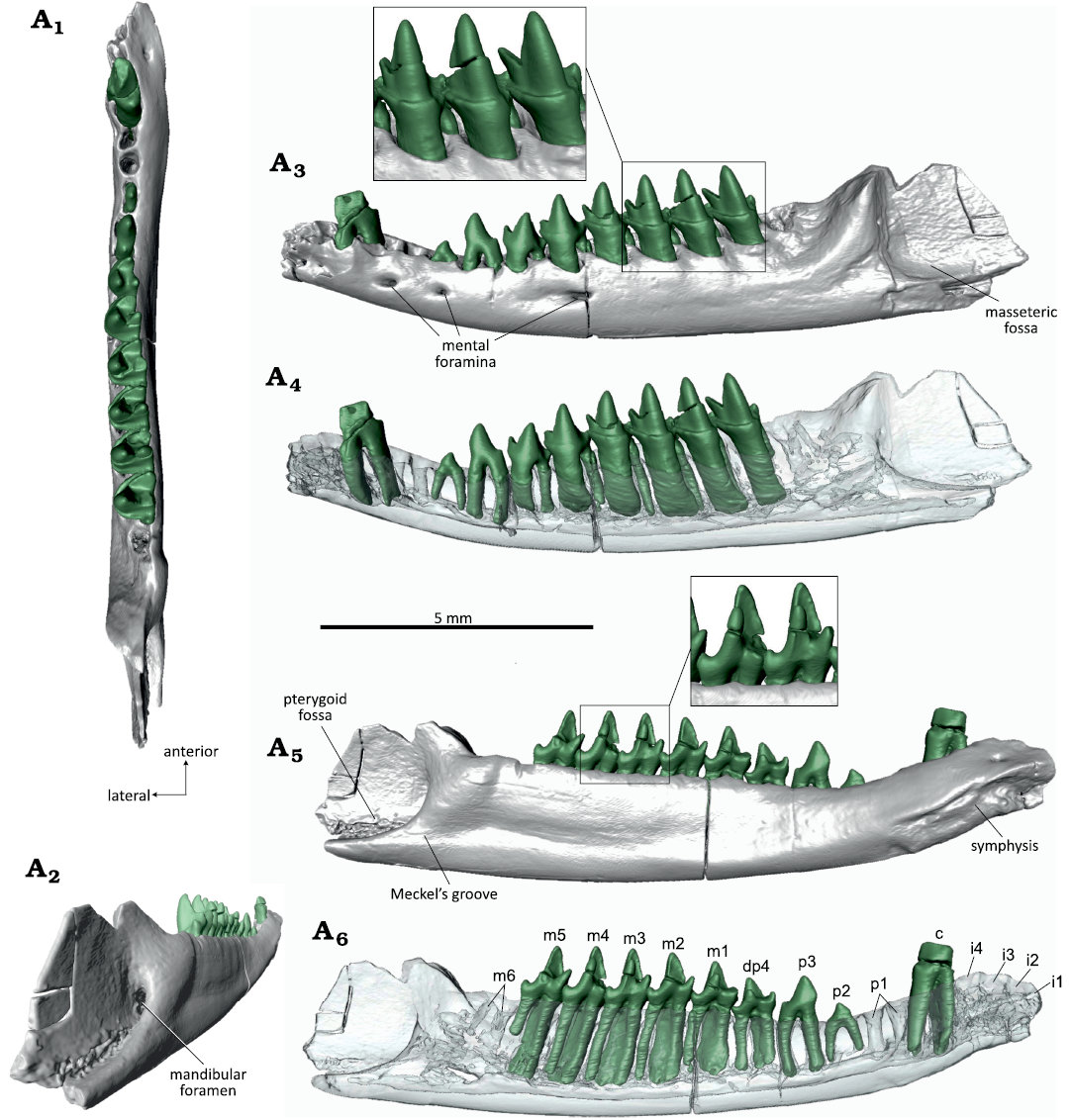
Fig. 11. Dryolestid mammal Crusafontia cuencana Henkel and Krebs, 1969, MfN Uña 1 (holotype) from Uña locality, province of Cuenca, Spain, Barremian (Lower Cretaceous). Left mandible in occlusal (A1), linguoposterior (A2), lateral (A3, A4), and medial (A5, A6) views. Inset in A3 showing enlarged m3–m5 in labial view, inset in A5 m3–m4 in ligunal view, A6 with new interpretation of premolar and molar positions.
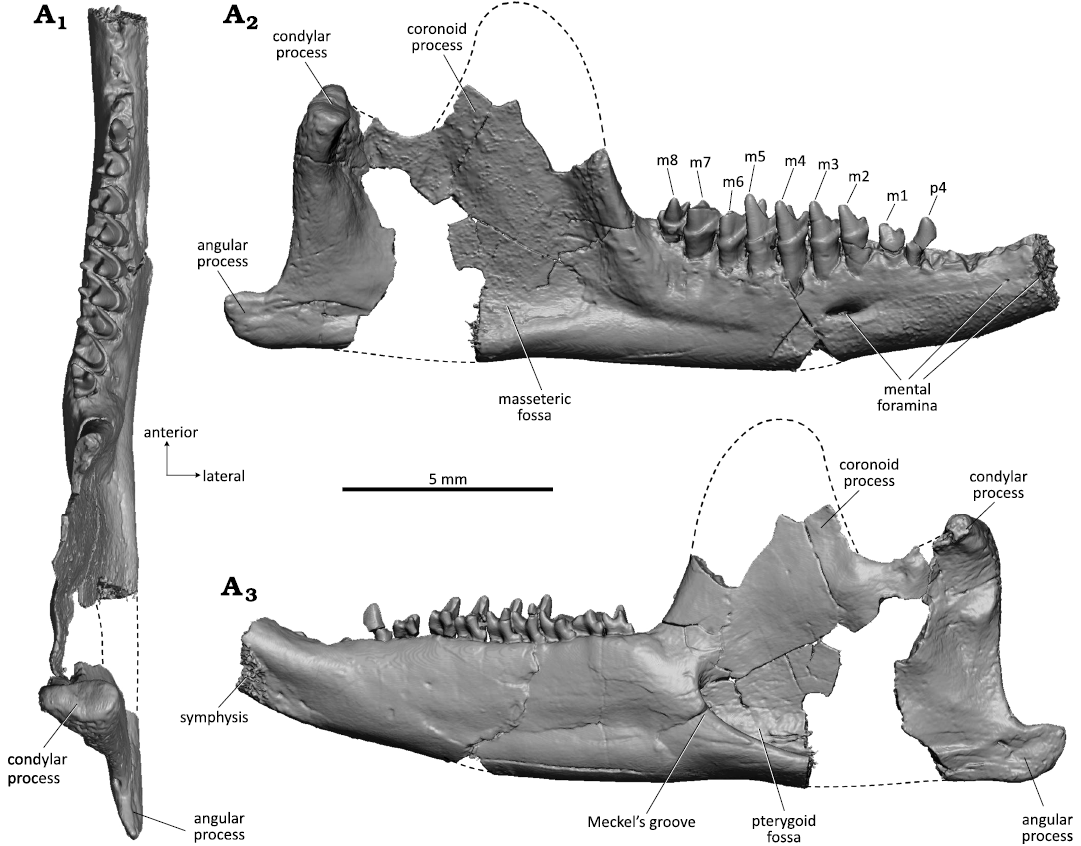
Fig. 12. Dryolestid mammal Crusafontia cuencana Henkel and Krebs, 1969, MfN Uña 2 (paratype) from Uña locality, province of Cuenca, Spain, Barremian (Lower Cretaceous). Right mandible in occlusal (A1), lateral (A2), and medial (A3) views. Reconstruction of broken parts of coronid process, ventral border of mandibular body (preserved as mold in the embedding rock) and connection of articular condyle marked by stippled lines.
The mandible of the Beckumia sinemeckelia gen. et sp. nov. type specimen (WMNM P82301) is somewhat larger than that of Crusafontia cuencana and its molars are of double size. The strongly worn molars indicate that it is from an old individual. The ultimate molar (m8) has a single root, and is directly in front of the coronoid process. Both features are adult characteristics. The small and single root of m8 suggests that m8 is the last tooth to erupt. This interpretation is based on the observation that the confluence of roots or a reduction to a single root of the ultimate molar is a typical character of the adult growth stage in other Mesozoic mammaliaforms for which the growth series are known (Schultz et al. 2019; Panciroli et al. 2019). Also, the last molar of the toothrow shifts anteriorly relative to the coronoid process during the growth of mammaliaforms (Schultz et al. 2019: fig. 4). In a growth series, the younger individuals tend to have their last molar positioned medial to the anterior edge of the coronoid process at the time of eruption; the larger and older adult specimens usually have the ultimate molar located directly in front of the coronoid process.
Stratigraphic and geographic range.—Type locality and horizon only.
Cladistic analysis
In our cladistic analysis, Minutolestes submersus gen. et sp. nov. appears as sister taxon of the clade comprising Laolestes eminens, Krebsotherium lusitanicum, Dryolestes priscus, Dryolestes leiriensis, and Guimarotodus inflatus (Fig. 13). Beckumia sinemeckelia gen. et sp. nov. appears in a different dryolestid clade, formed by Achyrodon nanus, Phascolestes mustelulus, Crusafontia cuencana, and Hercynodon germanicus, as the earliest diverging member (Fig. 13). Beckumia sinemeckelia gen. et sp. nov. exhibits a more primitive premolar pattern than C. cuencana: in the new taxon p2 is smaller than p1 and p3, and p4 is the largest premolar, a condition that is present in several other dryolestid taxa. Crusafontia has a more derived condition with three very small premolars (p1–p3) and a large p4. In Crusafontia the anterior end of the mandible is upturned (derived), whereas it is horizontal in B. sinemeckelia gen. et sp. nov. and other dryolestids. However, by the complete reduction of Meckel’s groove, B. sinemeckelia gen. et sp. nov. is more derived than Crusafontia, which shows a short Meckel’s groove, as well as all other dryolestids for which a mandible is known. The paratype mandible (MfN Uña 2) of C. cuencana represents an adult individual, as is evident from eight fully erupted molars, of which m8 is small and sits immediately in front of the ascending ramus. The molars of MfN Uña 2 exhibit considerable wear (Henkel and Krebs 1969: 459, fig. 2c–e), and the mandible possesses a short, but clearly visible Meckel’s groove (Henkel and Krebs 1969: fig. 2b). Therefore, it can be ruled out that the presence of Meckel’s groove in C. cuencana is an early ontogenetic feature. Like the mandible of C. cuencana (Henkel and Krebs 1969; Krebs 1971, 1993), the mandible WMNM P82301 is rather slender for a dryolestid; members of that clade usually have robust and vertically deep mandibular bodies. Size-wise and in proportions WMNM P82301 corresponds closely to the paratype mandible of C. cuencana from Uña.
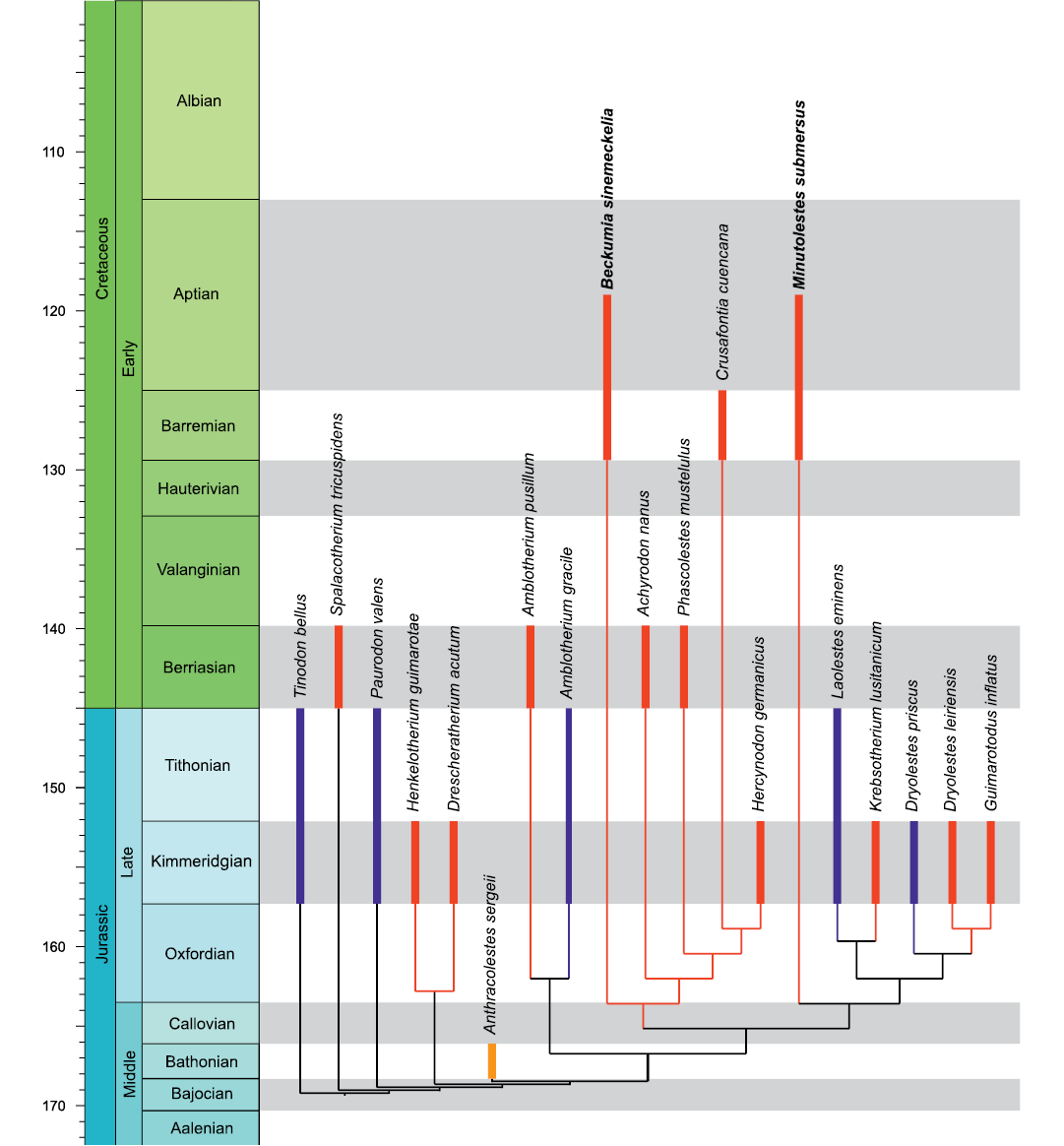
Fig. 13. Cladogram with interrelationships of the new dryolestids from the Barremian–Aptian (Early Cretaceous) of Balve. The nodes uniting taxa do not imply divergence times. Colors indicate distribution in the respective continents: red, Europe; blue, North America; yellow, Asia.
Discussion
With their Barremian–Aptian age, Cifellitherium suderlandicum gen. et sp. nov., Minutolestes submersus gen. et sp. nov., and Beckumia sinemeckelia gen. et sp. nov. represent the geologically youngest spalacotheriid and dryolestids in Europe. The next older European spalacotheriids, Yaverlestes gassoni from the Isle of Wight (Sweetman 2008), Spalacotherium henkeli from Galve (Krebs 1985), and Aliaga molinensis from Molino Alto 1 (both Spain) are of Barremian (Y. gassoni) and Hauterivian–Barremian age, respectively (Cuenca-Bescós et al. 2004). Formerly, spalacotheriids have been known in Europe only from Great Britain and the Iberian Peninsula, but C. suderlandicum gen. et sp. nov. extends the geographic distribution of Spalacotheriidae to Central Europe. The best-known spalacotheriid is the type species Spalacotherium tricuspidens Owen, 1854, which comes, like the smaller species of the genus (Spalacotherium evansae Ensom and Sigogneau-Russell, 2000, Spalacotherium taylori Clemens and Lees, 1971, and Spalacotherium hookeri Gill, 2004a), from the Purbeck Limestone Group and Wealden Supergroup and is Berriasian–Valanginian in age (Owen 1854; Clemens and Lees 1971; Ensom and Sigogneau-Russell 2000; Gill 2004a). So far, Spalacotherium has not been reported outside Europe, and C. suderlandicum gen. et sp. nov., morphologically close to the species of Spalacotherium, is a representative of this endemic European clade. In terms of their dentition, spalacolestines comprise the most derived spalacotheriids (Cifelli and Madsen 1999; Cifelli et al. 2014) and are mainly known from North America where they apparently experienced a radiation from the Aptian–Albian to early Campanian. The only European representative of Spalacolestinae is the Barremian Yaverlestes gassoni (Sweetman 2008). In contrast to medium to large sized Spalacotherium tricuspidens, S. henkeli, and C. suderlandicum gen. et sp. nov., spalacolestines are generally smaller. With a lower molar length of 2.18 mm (Krebs 1985), S. henkeli from the lower Barremian of Galve is the largest representative of Spalacotherium. Cifellitherium suderlandicum gen. et sp. nov. is about 30% smaller than S. henkeli. The spalacotheriid genera are also distinctly different from, and morphologically more derived than the Zhangheotheriidae from Siberia and northeastern China (Hu et al. 1997; Rougier et al. 2003a; Luo and Ji 2005; Lopatin et al. 2010; Bi et al. 2016; Plogschties and Martin 2019). The diverse spalacotheriid taxa and the absence of zhangheotheriid-like fossils in Europe, and the diverse and numerically abundant zhangheotheriids in Asia suggest paleobiogeographical differences in mammals during the Barremian–Aptian interval (Luo 1999; Kielan-Jaworowska et al. 2004).
Crusafontia cuencana from the early Barremian of the Spanish localities Galve and Uña (Henkel and Krebs 1969; Krebs 1985, 1993; Cuenca-Bescós et al. 2011) was previously the geologically youngest and most derived European representative of Dryolestidae. Beckumia sinemeckelia gen. et sp. nov. and Minutolestes submersus gen. et sp. nov. are younger and extend the geographical range of Cretaceous dryolestids to Central Europe. This Early Cretaceous assemblage of dryolestids from Europe is absent from the Early Cretaceous mammalian faunas of Siberia and East Asia, which are rich in other major clades (Luo 1999; Kielan-Jaworowska et al. 2004; Lopatin et al. 2010). This conspicuous absence of dryolestids corroborates the paleobiogeographic difference of mammalian distribution between Asia and Central and Western Europe.
Both dryolestids from Balve appear as the earliest divergent members of their clades despite that they are the stratigraphically youngest European representatives. Although it cannot be excluded that this is an artefact due to incomplete data, a possible explanation might be the geographical insular isolation on the European Archipelago in the Early Cretaceous that led to a long survival of early diverging taxa and endemic character evolution. Similar insular phenomena have recently been described for Late Jurassic dryolestids and other mammals from the Langenberg Quarry in the Harz Mountains (Lower Saxony) (Martin et al. 2019, 2021).
Conclusions
The new spalacotheriid and dryolestids described from Balve are the geologically youngest (Barremian–Aptian) European representatives and extend the geographical range of their clades to Central Europe. Beckumia sinemeckelia gen. et sp. nov. and Minutolestes submersus gen. et sp. nov. are late-surviving early-diverging members of their respective subclades within Dryolestidae, with some plesiomorphic character states possibly reflecting insular isolation and subsequent endemic evolution on the European Archipelago. A similar scenario was recently suggested for a Late Jurassic (Kimmeridgian) dryolestid from Lower Saxony. However, the knowledge of the Balve mammal assemblage is limited so far and artefacts caused by incomplete character information, as well as the role of ontogeny in the presentation of some character states, cannot be ruled out. Ongoing field work at the Balve locality will undoubtedly shed more light on this hitherto poorly known chapter of mammalian evolution, and the broader, emerging pattern of mammalian paleobiogeography during the Early Cretaceous.
Acknowledgements
We dedicate this study to our esteemed colleague and friend Richard L. Cifelli (Sam Noble Oklahoma Museum of Natural History, University of Oklahoma, Norman, USA) in recognition of his outstanding contributions to vertebrate paleontology and cooperativeness. We all, including many graduate students from Bonn, have greatly benefitted from his generosity, both in terms of scientific advice and selfless sharing of specimens. We thank the company Lhoist for permits and general support for working in the quarry near Balve. The excavation was technically supported by Lhoist, as well as by the building yard of the town of Balve, for which we are very grateful. Klaus-Peter Lanser (formerly WMNM), numerous preparators and students conducted the excavation in the years 2002–2016. Since then, AHS has continued the excavation, assisted by preparators and student interns. These were in 2020 Lisa Herud, Alexander Klement, Jan Matzker, Denis Theda (all Westfälische Friedrich-Wilhelms-Universität Münster, Germany) and Rebekka Nölden (Menden (Sauerland), Germany); picking sediment samples was done by Peter Jütte, Leo Hallaschka, Lisa Herud, and Alexander Klement (all Westfälische Friedrich-Wilhelms-Universität, Münster, Germany). Jannik Weidtke segmented and reconstructed the mandible (Fig. 9) and Thomas Engler (both Rheinische Friedrich- Wilhelms-Universität Bonn, Germany) plotted the cladogram of Fig. 10. We thank Daniela Schwarz (Museum für Naturkunde, Berlin, Germany) for loaning Crusafontia specimens MfN Uña 1 and Uña 2. The reviewers Zhe-Xi Luo (The University of Chicago, Chicago, USA) and Elsa Panciroli (Oxford University Museum of Natural History, Oxford, UK) provided insightful comments that greatly improved our manuscript. We thank the editors Olivier Lambert (Royal Belgian Institute of Natural Sciences, Brussels, Belgium) and Brian Davis (University of Louisville, Louisville, USA) for the invitation to contribute to this festschrift and for their careful editing of our manuscript. AA was supported by the Russian Science Foundation (project 19-14-00020), the Zoological Institute Russian Academy of Sciences (project 1021051302397-6) and a Humboldt Research Award by the Alexander von Humboldt Foundation.
References
Archibald, J.D. and Averianov, A.O. 2005. Mammalian faunal succession in the Cretaceous of the Kyzylkum Desert. Journal of Mammalian Evolution 12: 9–22. Crossref
Averianov, A.O., Martin, T, and Lopatin, A.V. 2014. The oldest dryolestid mammal from the Middle Jurassic of Siberia. Journal of Vertebrate Paleontology 34: 924–931. Crossref
Bi, S., Zheng, X., Meng, J., Wang, X, Robinson, N., and Davis, B.M. 2016. A new symmetrodont mammal (Trechnotheria: Zhangheotheriidae) from the Early Cretaceous of China and trechnotherian character evolution. Scientific Reports 6: 1–9. Crossref
Bonaparte, J.F. 1986. Sobre Mesungulatum houssayi y nuevos mammíferos cretácicos de Patagonia. 4° Congreso Argentino de Paleontología y Bioestratigrafía, Mendoza, Actas 2: 48–61.
Chen, M. and Luo, Z.-X. 2008. Morphology of dentition and postcranial skeleton of Akidolestes (in Chinese with English abstract). Acta Geologica Sinica 82 (2): 155–164.
Cifelli, R.L. 1990. Cretaceous mammals of southern Utah. III. Therian mammals from the Turonian (early Late Cretaceous). Journal of Vertebrate Paleontology 10: 332–345. Crossref
Cifelli, R.L. and Gordon, C.I. 1999. Symmetrodonts from the Late Cretaceous of southern Utah and distribution of archaic mammals in the Cretaceous of North America. Geology Studies, Bringham Young University 44: 1–16.
Cifelli, R.L. and Madsen, S.K. 1999. Spalacotheriid symmetrodonts (Mammalia) from the medial Cretaceous (upper Albian or lower Cenomanian) Mussentuchit local fauna, Cedar Mountain Formation, Utah, USA. Geodiversitas 21: 167–214.
Cifelli, R.L., Davis, B.M., and Sames, B. 2014. Earliest Cretaceous mammals from the western United States. Acta Palaeontologica Polonica 59: 31–52.
Clemens, W.A. and Lees, P.M. 1971. A review of english early Cretaceous mammals. In: D.M. Kermack and K.A. Kermack (eds.), Early Mammals. Zoological Journal of the Linnean Society 50 (Supplement 1): 117–130.
Cuenca-Bescós, G., Badiola, A., Canudo, J.I., Gasca, J.M., and Moreno-Azanza, M. 2011. New dryolestidan mammal from the Hauterivian–Barremian transition of the Iberian Peninsula. Acta Palaeontologica Polonica 56: 257–267. Crossref
Cuenca-Bescós, G., Canudo, J.I., Gasca, J.M., Moreno-Azanza, M., and Cifelli, R.L. 2004. Spalacotheriid “symmetrodonts” from the Early Cretaceous of Spain. Journal of Vertebrate Paleontology 34: 1427–1436. Crossref
Drozdzewski, G., Hartkopf-Fröder, C., Lange, F.G., Oesterreich, B., Ribbert, K.-H., Voigt, S., and Wrede, V. 1998. Vorläufige Mitteilung über unterkretazischen Tiefenkarst im Wülfrather Massenkalk (Rheinisches Schiefergebirge). Mitteilungen des Verbandes deutscher Höhlen- und Karstforscher e.V. 44: 53–66.
Drozdzewski, G., Richter, D.K., and Wrede, V. 2017. Hydrothermalkarst im nördlichen Rheinischen Schiefergebirge. Jahrbuch Karst und Höhle 2015–2017: 1–88.
Ensom, P.C. and Sigogneau-Russell, D. 2000. New symmetrodonts (Mammalia, Theria) from the Purbeck Limestone Group, Lower Cretaceous, southern England. Cretaceous Research 21: 767–779. Crossref
Fox, R.C. 1976. Additions to the mammalian local fauna from the upper Milk River Formation (Upper Cretaceous), Alberta. Canadian Journal of Earth Sciences 13: 1105–1118. Crossref
Freeman, E.F. 1979. A Middle Jurassic mammal bed from Oxfordshire. Palaeontology 22: 135–166.
Gill, P.G. 2004a. A new symmetrodont from the Early Cretaceous of England. Journal of Vertebrate Paleontology 24: 748–752. Crossref
Gill, P.G. 2004b. Kuehneotherium from the Mesozoic Fissure Fillings of South Wales. 286 pp. Ph.D. Dissertation, University of Bristol, Bristol. Available at http://research-information.bristol.ac.uk
Goloboff, P.A. and Catalano, S.A. 2016. TNT version 1.5, including a full implementation of phylogenetic morphometrics. Cladistics 32: 221–238. Crossref
Goloboff, P.A., Farris, J.S., and Nixon, K.C. 2008. TNT, a Free Program for Phylogenetic Analysis, available at https://doi.org/10.1111/j.1096-0031.2008.00217.x Crossref
Han, G. and Meng, J. 2016. A new spalacolestine mammal from the Early Cretaceous Jehol Biota and implications for the morphology, phylogeny, and palaeobiology of Laurasian “symmetrodontans”. Zoological Journal of the Linnean Society 178: 343–380. Crossref
Henkel, S. and Krebs, B. 1969. Zwei Säugetier-Unterkiefer aus der Unteren Kreide von Uña (Prov. Cuenca, Spanien). Neues Jahrbuch für Geologie und Paläontologie, Monatshefte 8: 449–463.
Hu, Y.-M., Fox, R.C., Wang, Y., and Li, C.-K. 2005. A new spalacotheriid symmetrodont from the Early Cretaceous of northeastern China. American Museum Novitates 3475: 1–20. Crossref
Hu, Y.-M., Luo, Z.-X., Li, C.-K., and Wang, Y.-Q. 1997. A new symmetrodont mammal from China and its implications for mammalian evolution. Nature 390: 137–142. Crossref
Huckriede, R. 1982. Die unterkretazische Karsthöhlen-Füllung von Nehden im Sauerland. 1. Geologische, paläozoologische und paläobotanische Datierung. Geologica et Palaeontologica 16: 183–204.
Ji, Q., Luo, Z.-X., Zheng, X., Yuan, C.-X., and Xu, L. 2009. Evolutionary development of the middle ear in Mesozoic therian mammals. Science 326: 278–281. Crossref
Kielan-Jaworowska, Z., Cifelli, R.C., and Luo, Z.-X. 2004. Mammals from the Age of Dinosaurs: Origins, Evolution and Structure. 630 pp. Columbia University Press, New York. Crossref
Krebs, B. 1969. Nachweis eines rudimentären Coronoids im Unterkiefer der Pantotheria (Mammalia). Paläontologische Zeitschrift 43: 57–63. Crossref
Krebs, B. 1971. Evolution of the mandible and lower dentition in dryolestids (Pantotheria, Mammalia). In: D.M. Kermack and K.A. Kermack (eds.), Early Mammals. Zoological Journal of the Linnean Society 50 (Supplement 1): 89–102.
Krebs, B. 1985. Theria (Mammalia) aus der Unterkreide von Galve (Provinz Teruel, Spanien). Berliner geowissenschaftliche Abhandlungen A 60: 29–48.
Krebs, B. 1993. Das Gebiß von Crusafontia (Eupantotheria, Mammalia) Funde aus der Unter-Kreide von Galve und Uña (Spanien). Berliner geowissenschaftliche Abhandlungen E 9: 233–252.
Krusat, G. 1989. Isolated molars of a triconodont and a symmetrodont (Mammalia) from the uppermost Jurassic of Portugal. Berliner gewissenschaftliche Abhandlungen A 106: 277–289.
Lanser, K.-P. and Heimhofer, U. 2013. Evidence of theropod dinosaurs from a Lower Cretaceous karst filling in the northern Sauerland (Rhenish Massif, Germany). Paläontologische Zeitschrift 89: 79–94. Crossref
Li, G. and Luo, Z.-X. 2006. A Cretaceous symmetrodont therian with some monotreme-like postcranial features. Nature 439: 195–200. Crossref
Lillegraven, J.A. and McKenna, M.C. 1986. Fossil mammals from the “Mesaverde” Formation (Late Cretaceous, Judithian) of the Bighorn and Wind River basins, Wyoming, with definitions of Late Cretaceous North American land-mammal “ages”. American Museum Novitates 2840: 1–68.
Linnaeus, C. 1758. Systema naturae per regna tria naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Vol. 1: Regnum animale. Editio decima, reformata. Laurentius Salvius, Stockholm. Crossref
Lopatin, A.V., Averianov, A.O., Maschenko, E.N., and Leshchinskiy, S.V. 2010. Early Cretaceous mammals of Western Siberia: 3. Zhangheotheriidae. Paleontological Journal 44: 573–583. Crossref
Luo, Z.-X. 1999. A refugium of relicts. Nature 400: 23–25. Crossref
Luo, Z.-X. and Ji, Q. 2005. New study on dental and skeletal features of the Cretaceous “symmetrodontan” mammal Zhangheotherium. Journal of Mammalian Evolution 12: 337–357. Crossref
Marsh, O.C. 1878. Fossil mammal from the Jurassic of the Rocky Mountains. American Journal of Science 15: 459. Crossref
Marsh, O.C. 1879. Notice of new Jurassic mammals. American Journal of Science, Series 3 20: 396–742. Crossref
Marsh, O.C. 1887. American Jurassic mammals. American Journal of Science, Series 3 33: 326–348. Crossref
Martin, T. 1995. Dryolestidae from the Kimmeridge of the Guimarota coal mine (Portugal) and their implications for dryolestid systematics and phylogeny. In: A. Sun and Y. Wang (eds.), Sixth Symposium on Mesozoic Terrestrial Ecosystems and Biota, Short Papers, 229–231. China Ocean Press, Beijing.
Martin, T. 1997. Tooth replacement in Late Jurassic Dyolestidae (Eupantotheria, Mammalia). Journal of Mammalian Evolution 4: 1–18.
Martin, T. 1998. The premolars of Crusafontia cuencana (Dryolestidae, Mammalia) from the Early Cretaceous (Barremian) of Spain. Berliner geowissenschaftliche Abhandlungen E 28: 119–126.
Martin, T. 1999. Dryolestidae (Dryolestoidea, Mammalia) aus dem Oberen Jura von Portugal. Abhandlungen der senckenbergischen naturforschenden Gesellschaft 550: 1–119.
Martin, T. 2000. The dryolestids and the primitive “peramurid” from the Guimarota Mine. In: T. Martin and B. Krebs (eds.), Guimarota: a Jurassic ecosystem, 109–120. Verlag Dr. Friedrich Pfeil, Munich.
Martin, T. 2018. Mesozoic mammals—early mammalian diversity and ecomorphological adaptations. In: F.E. Zachos and R.J. Asher (eds.), Mammalian Evolution, Diversity and Systematics. Handbook of Zoology. Mammalia, 199–299. De Gruyter, Berlin. Crossref
Martin, T., Averianov, A.O., Jäger, K.R.K., Schwermann, A.H., and Wings, O. 2019. A large morganucodontan mammaliaform from the Late Jurassic of Germany. Fossil Imprint 75: 504–509. Crossref
Martin, T., Averianov, A.O., Schultz, J.A., Schwermann, A.H., and Wings, O. 2021. A derived dryolestid mammal indicates possible insular endemism in the Late Jurassic of Germany. The Science of Nature 108: 23. Crossref
Maschenko, E.N., Lopatin, A.V., and Voronkevich, A.V. 2002. A new Early Cretaceous mammal from western Siberia. Doklady Biological Sciences 386: 475–477. Crossref
Mazin, J.-M., Billon-Bruyat, J.-P., Pouech, J., and Hantzpergue, P. 2006. The Purbeckian site of Cherves-de-Cognac (Berriasian, Early Cretaceous, southwest France): a continental ecosystem accumulated in an evaporitic littoral depositional environment. In: P.M. Barrett and S.E. Evans (eds.), 9th International Symposium on Mesozoic Terrestrial Ecosystems and Biota, Abstracts and Proceedings Volume, 84–88. Natural History Museum, London.
McKenna, M.C. 1975. Toward a phylogenetic classification of the Mammalia. In: W.P. Luckett and F.S. Szalay (eds.), Phylogeny of the Primates: a Multidisciplinary Approach, 21–46. Plenum Press, New York. Crossref
Owen, R. 1854. On some fossil reptilian and mammalian remains from the Purbecks. Quarterly Journal of the Geological Society of London 10: 420–433. Crossref
Owen, R. 1871. Monograph of the fossil Mammalia of the Mesozoic formations. Monograph of the Palaeontological Society 33 (for 1870): 1–115. Crossref
Panciroli, E.L., Benson, R.B.J., and Luo, Z.-X. 2019. The mandible and dentition of Borealestes serendipitus (Docodonta) from the Middle Jurassic of Skye, Scotland. Journal of Vertebrate Paleontology 39: e1621884. Crossref
Patterson, B. 1955. A symmetrodont mammal from the Early Cretaceous of northern Texas. Fieldiana: Zoology 37: 689–693. Crossref
Patterson, B. 1956. Early Cretaceous mammals and the evolution of mammalian molar teeth. Fieldiana: Geology 13: 1–105. Crossref
Plogschties, T. and Martin, T. 2019. New information on the maxilla, dentary, and dentition of Maotherium sinense, with comments on the zhangheotheriid dental formula. PalZ 94: 155–165. Crossref
Pouech, J., Mazin, J.-M., and Billon-Bruyat, J.-P. 2006. Microvertebrate biodiversity from Cherves-de-Cognac (Lower Cretaceous, Berriasian: Charente, France). In: P.M. Barrett and S.E. Evans (eds.), 9th International Symposium on Mesozoic Terrestrial Ecosystems and Biota, Abstracts and Proceedings Volume, 96–100. Natural History Museum, London.
Prothero, D.R. 1981. New Jurassic mammals from Como Bluff, Wyoming, and the interrelationships of non-tribosphenic Theria. Bulletin of the American Museum of Natural History 167: 281–325.
Richter, D.K., Götte, T., and Stritzke, R. 2014. Eine miozäne Hohlraumfüllung im Massenkalk der östlichen Iserlohner Kalksenke. Geologie und Paläontologie in Westfalen 86: 5–17.
Rougier, G.W., Chornogubsky, L., Casadio, S., Arango, N.P., and Giallombardo, A. 2009. Mammals from the Allen Formation, Late Cretaceous, Argentina. Cretaceous Research 30: 223–238. Crossref
Rougier, G.W., Ji, Q., and Novacek, M.J. 2003a. A new symmetrodont mammal with fur impressions from the Mesozoic of China. Acta Geologica Sinica (English Edition) 77: 7–14. Crossref
Rougier, G.W., Spurlin, B.K., and Kik, P.K. 2003b. A new specimen of Eurylambda aequicrurius and considerations on “symmetrodont” dentition and relationships. American Museum Novitates 3398: 1–15. Crossref
Schudack, M. 1987. Charophytenflora und Alter der unterkretazischen Karsthöhlen-Füllung von Nehden (NE-Sauerland). Geologie und Paläontologie in Westfalen 10: 7–44.
Schultz, J.A. and Martin, T. 2011. Wear pattern and functional morphology of dryolestoid molars (Cladotheria, Mammalia). Paläontologische Zeitschrift 85: 269–285. Crossref
Schultz, J.A., Bhullar, B.A.S., and Luo, Z.-X. 2019. Re-examination of the Jurassic mammaliaform Docodon victor by computed tomography and occlusal functional analysis. Journal of Mammalian Evolution 26: 9–38. Crossref
Simpson, G.G. 1927. Mesozoic Mammalia. VI. Genera of Morrison pantotheres. American Journal of Science, Series 5 13: 409–416. Crossref
Simpson, G.G. 1928. A Catalogue of the Mesozoic Mammalia in the Geological Department of the British Museum. 215 pp. Trustees of the British Museum, London.
Sweetman, S.C. 2008. A spalacolestine spalacotheriid (Mammalia, Trechnotheria) from the Early Cretaceous (Barremian) of southern England and its bearing of spalacotheriid evolution. Palaeontology 51: 1367–1385. Crossref
Tsubamoto, T., Rougier, G.W., Isaji, G., Manabe, M., Forasiepi, A.M. 2004. New Early Cretaceous spalacotheriid “symmetrodont” mammal from Japan. Acta Palaeontologica Polonica 49: 329–346.
Wirth, W. 1970. Eine tertiärzeitliche Karstfüllung bei Eisborn im Sauerland. Fortschritte in der Geologie von Rheinland und Westfalen 7: 403–420.
Acta Palaeontol. Pol. 67 (1): 155–175, 2022
https://doi.org/10.4202/app.00914.2021