
Early–middle Cambrian stratigraphy and faunas from northern Siberia
ARTEM KOUCHINSKY, RUARIDH ALEXANDER, STEFAN BENGTSON, FRED BOWYER, SÉBASTIEN CLAUSEN, LARS E. HOLMER, KIRILL A. KOLESNIKOV, IGOR V. KOROVNIKOV, VLADIMIR PAVLOV, CHRISTIAN B. SKOVSTED, GALINA USHATINSKAYA, RACHEL WOOD, and ANDREY Y. ZHURAVLEV
Kouchinsky, A., Alexander, R., Bengtson, S., Bowyer, F., Clausen, S., Holmer, L.E., Kolesnikov, K.A., Korovnikov, I.V., Pavlov, V., Skovsted, C.B., Ushatinskaya, G., Wood, R., and Zhuravlev, A.Y. 2022. Early–middle Cambrian stratigraphy and faunas from northern Siberia. Acta Palaeontologica Polonica 67 (2): 341–464.
New assemblages of skeletal fossils chemically extracted from carbonates of the Cambrian Stage 2–Drumian Stage are reported from the lower reaches of the Lena River as well as from the Khorbusuonka, Malaya Kuonamka, and Bol’shaya Kuonamka rivers in northern part of the Siberian Platform. The fauna studied with scanning electron microscopy includes brachiopods, molluscs, hyoliths, halkieriids, chancelloriids, tommotiids, lobopodians, palaeoscolecidans, bradoriids, echinoderms, anabaritids, hyolithelminths, and sponges showing similarity to previously described fossil assemblages from Siberia, Laurentia, and Gondwana. The material includes emended descriptions of Halkieria proboscidea, Hadimopanella knappologica, Archaeopetasus typicus, and first descriptions of Hadimopanella foveata Kouchinsky sp. nov. and Archaeopetasus pachybasalis Kouchinsky sp. nov. Affinity of Archaeopetasus to chancelloriids is suggested. Finding of an in-place operculum in a planispiral shell of Michniakia minuta enables reinterpretation of this form as a hyolith, not a mollusc. The cambroclavids Cambroclavus sp. and Zhijinites clavus and the earliest echinoderms belonging to the Rhombifera and Ctenocystoidea are reported respectively from the lower Botoman stage and Botoman–Toyonian transitional beds, correlated with Cambrian Stage 4. Carbon isotopes are analysed from sections of the Chuskuna (upper Kessyusa Group), Erkeket, Kuonamka, Olenyok, Yunkyulyabit-Yuryakh, Tyuser and Sekten formations. A major part of the δ13C record is obtained from the Cambrian Stage 4–Drumian Stage strata which remain incompletely characterised by chemostratigraphy. The Lower Anomocarioides limbataeformis Carbon isotope Excursion (LACE) from the Drumian Stage of the Khorbusuonka River is introduced herein. New chemostratigraphic data are used for regional and global correlation and facilitate study of the evolutionary development of animals and faunas through the “Cambrian explosion”.
Key words: Small shelly fossils, carbon isotopes, stratigraphy, Cambrian, Siberia, Russia.
Artem Kouchinsky [artem.kouchinsky@gmail.com], Stefan Bengtson [stefan.bengtson@nrm.se], and Christian B. Skovsted [christian.skovsted@nrm.se], Department of Palaeobiology, Swedish Museum of Natural History, Box 50007, SE-10405 Stockholm, Sweden.
Ruaridh Alexander [ruaridh.alexander@ed.ac.uk], Fred Bowyer [fred.bowyer@ed.ac.uk], and Rachel Wood [rachel.wood@ed.ac.uk], Grant Institute, School of GeoSciences, University of Edinburgh, James Hutton Road, Edinburgh, EH9 3FE, UK.
Sébastien Clausen [sebastien.clausen@univ-lille.fr], Université des Sciences et Technologies de Lille, CNRS, UMR 8198-Evo-Eco-Paleo, F-59000 Lille, France.
Lars E. Holmer[lars.holmer@pal.uu.se], Department of Earth Sciences, Palaeobiology, Uppsala University, Villavägen 16, SE-752 36 Uppsala, Sweden.
Kirill A. Kolesnikov [kolesnikovk-msu@mail.ru] and Andrey Y. Zhuravlev [ayzhur@mail.ru], Department of Biological Evolution, Faculty of Biology, Lomonosov Moscow State University, Leninskie Gory 1(12), Moscow 119234, Russia.
Igor Korovnikov [korovnikovIV@ipgg.sbras.ru], Trofimuk Institute of Petroleum Geology and Geophysics, Siberian Branch, Russian Academy of Sciences, pr. Akademika Koptyuga 3, 630090, Novosibirsk, Russia and Novosibirsk State University, ul. Pirogova 1, Novosibirsk, 630090, Russia.
Vladimir Pavlov [pavlov.ifz@gmail.com], Schmidt Institute of Physics of the Earth, Russian Academy of Sciences, Bol’shaya Gruzinskaya ul. 10(1), Moscow 123242, Russia and Kazan Federal University, ul. Kremlyovskaya 18, Kazan 420008, Russia.
Galina Ushatinskaya [gushat@paleo.ru], Borissiak Paleontological Institute, Russian Academy of Sciences, Profsoyuznaya ul. 123, 117997 Moscow, Russia.
Received 27 July 2021, accepted 26 November 2021, available online 14 June 2022.
Copyright © 2022 Kouchinsky et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Assemblages of skeletal fossils (“small shelly fossils”), i.e., mineralised skeletal parts of diverse animals known in microfossil assemblages, complemented by exceptionally preserved soft-bodied fossils in Konservat–Lagerstätten, ichnofossils and organic-walled microfossils (“small carbonaceous fossils”) reveal an astonishing diversity and uniqueness of the animal world of the early–middle Cambrian. “Lower/early” and “middle” Cambrian herein represent informal divisions, respectively including the Terreneuvian Series/Epoch–Cambrian Series/Epoch 2 and the Miaolingian Series/Epoch accepted by the International Subcomission on Cambrian Stratigraphy (see Geyer 2019; Landing et al. 2021b and references therein). The Siberian Platform is recognised for its well-preserved and continuous record of “small shelly fossils”, in addition to the archaeocyaths, brachiopods, and trilobites traditionally used for Cambrian biostratigraphy.
Updated studies of trilobites and brachiopods from the Erkeket, Yunkyulyabit-Yuryakh, and Tyussala formations of the Khorbusuonka River (Korovnikov 1998, 2002; Ushatinskaya and Korovnikov 2014, 2016; Korovnikov and Tokarev 2018) refined the regional stratigraphy summarised by Astashkin et al. (1991 and references therein). Karlova and Vodanyuk (1985) first provided lists of fossils from the upper Kessyusa Group on the Khorbusuonka River and reported tubular problematica from the underlying Turkut Formation (see also Knoll et al. 1995 and Pelechaty et al. 1996a, b). Sarsembaev and Marusin (2022) described conulariids from the upper Mattaia Formation. Ichnofossils and stratigraphy of the Kessyusa Group in the region were investigated (Dzik 2005; Nagovitsin et al. 2015; Rogov et al. 2015; Grazhdankin et al. 2020, and references therein), but with the exception of trilobites and brachiopods, other skeletal fossils from Cambrian deposits along the Khorbusuonka River have not been previously described.
Trilobite-based biostratigraphy of the Bulkur Anticline (lower reaches of the Lena River) elaborated by Repina et al. (1974), Lazarenko and Repina (1983) was summarised by Astashkin et al. (1991) and updated by Bushuev et al. (2014). Although descriptions of the early skeletal microfaunas (in addition to trilobites and brachiopods) from the Bulkur Anticline (Ulakhan-Ald’yarkhay section) were also available (Repina et al. 1974; Zinchenko et al. 1993; Vasiljeva 1998), they were not adequately illustrated.
“Small shelly fossils” and brachiopods are described herein from the Cambrian Stage 2–Drumian Stage interval of the Bulkur Anticline (lower reaches of the Lena River) and eastern flanks of the Olenek Uplift (middle reaches of the Khorbusuonka River). They derive from carbonate-siliciclastic facies at the margin of the Yudoma-Olenyok normal marine facies region defined for the lower–middle Cambrian of the Siberian Platform and located near the transitional successions (i.e., between normal marine and highly saline) of the Anabar-Sinsk facies region. In combination with new chemostratigraphic data from the Kuonamka and lowermost Olenyok formations, this report complements earlier studies of fossils from the eastern flanks of the Anabar Uplift (Gubanov et al. 2004b; Kouchinsky et al. 2011, 2015a, b) by descriptions of lingulate brachiopods and refines the δ13C record through the Cambrian strata of the Siberian Platform (Kouchinsky et al. 2001, 2005, 2007, 2008, 2015a, 2017).
Institutional abbreviations.—SMNH, Swedish Museum of Natural History, Stockholm, Sweden (Br, brachiopods; Mo, molluscs; Ar, arthropods; Ec, echinoderms; Sp, sponges; X, other fossils).
Nomenclatural acts.—This published work and the nomenclatural acts it contains have been registered in ZooBank: urn:lsid:zoobank.org:pub:D1EB034B-09AC-4578-B0FC-84FB2D63B0BC
Geological and geographical setting
Material described herein derives from sections in the northern part of the Siberian Platform (Figs. 1–4). The western set of sections is located along the Malaya Kuonamka River (sections 96-1, 96-2, and 96-3) and Bol’shaya Kuonamka River (sections 96-6, 96-7, and 96-8) on the eastern flanks of the Anabar Uplift, visited by AK and SB in 1996 and described in Gubanov et al. (2004b) and Kouchinsky et al. (2011, 2015a, b) as sections 1, 2, 3, 6, 7, and 8. Those are designated 96-1, 96-2, 96-3, 96-6, 96-7, and 96-8, respectively, by Kouchinsky et al. 2001, 2017 (see also in Kouchinsky et al. 2017: SOM) and herein.
Section 96-1 (70°08.5’ N, 114°00’ E) is situated at the left bank of the Malaya Kuonamka River, near and downstream of the village Dzhilinda. Section 96-2 (70°10’N, 114°03’E) is located on the right bank of the Malaya Kuonamka River, ca. 2 km downstream of the village Dzhilinda. Section 96-3 (70°13’N, 113°42’E) is exposed along the left bank of the Malaya Kuonamka River, 1.5–2 km downstream of the mouth of the Maspaky Brook. Section 96-6 (70°43’N, 112°50’E) crops out along the right bank of the Bol’shaya Kuonamka River, 1–2.5 km downstream of the mouth of the Ulakhan-Tyulen Brook. Section 96-7 (70°43’N, 112°52.5’E) is situated along the left bank of the Bol’shaya Kuonamka River, 3.5–4 km downstream of the mouth of the Ulakhan-Tyulen Brook. Section 96-8 (70°44.2’N, 112°58.8’E) is exposed along the left bank of the Bol’shaya Kuonamka River, ca. 7 km downstream of the mouth of the Ulakhan-Tyulen Brook.
The eastern set of sections is situated at the Olenyok Uplift (Khorbusuonka River) and Bulkur Anticline (Lena River) visited by AK, SB, IVK, and VP in 2000 and includes the following localities.
Section 11 (71°19’ N, 123°59.5’ E) herein, reported as section 12 (Knoll et al. 1995: figs. 1, 2), crops out at the right bank of the Khorbusuonka River, at the right side of the mouth of the Yuettekh Brook (ca. 1 km downstream the mouth of Yuesse-Yuettekh Brook; Fig. 1). Section 15 (71°24.5’ N, 123°48.5’ E) is situated at the right bank of the Khorbusuonka River, ca. 1 km upstream and opposite the mouth of the Mattaia River (left tributary of the Khorbusuonka River). It is described as section 17 (Karlova and Vodanyuk 1985), and represents the upper part of sections 14 (Knoll et al. 1995: figs. 1, 2), 17 (Dzik 2005: fig. 1D), and 0708–0709 (Grazhdankin et al. 2020: figs. 1, 2). Sections 11 and 15 are mainly siliciclastic facies of the uppermost Mattaia and Chuskuna formations of the Kessyusa Group (Nagovitsin et al. 2015; Rogov et al. 2015; Fig. 3). A calcimicrobial thrombolite, ca. 2 m in visible thickness, crops out at the base of section 11 (at the water level of the Khorbusuonka River) and includes embedded calcitic tubular fossils, probably anabaritids Cambrotubulus conicus Missarzhevsky, 1989 (see Kouchinsky et al. 2009), up to 5 mm in diameter and 20 mm long (SOM 1, Supplementary Online Material available at http://app.pan.pl/SOM/app67-Kouchinsky_etal_SOM.pdf). The tubes have rounded and circular cross-sections and smooth walls visible on the weathered surface of the basal bed. The latter can be attributed to the equivalent of Suordakh Member (Meshkova et al. 1973; Missarzhevsky 1980; Zinchenko 1985), separated from the overlying Chuskuna Formation by a flooding surface (Grazhdankin et al. 2020). The upper member of the Mattaia Formation in section 15 consists of sandstone beds, from which a 0.2 m layer of light yellowish-grey volcanic tuff with an age of 529.7±0.3 Ma was reported, ca. 3 m below the Chuskuna Formation (Kaufman et al. 2012; Nagovitsin et al. 2015; Grazhdankin et al. 2020).
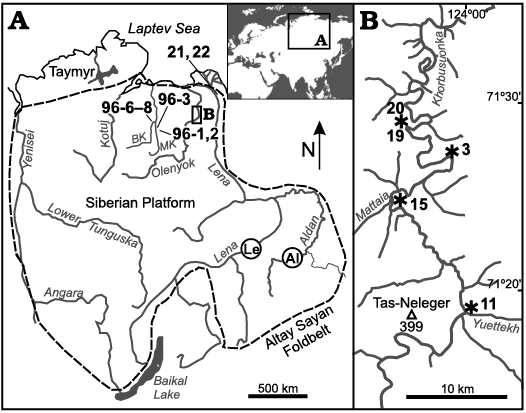
Fig. 1. Locality maps. A. Map of the Siberian Platform. B. Khorbusuonka River. Localities referred to in the main text: 96-1, 96-2, 96-3, Malaya Kuonamka (MK) and 96-6, 96-7, 96-8, Bol’shaya Kuonamka (BK) rivers, eastern flank of the Anabar Uplift; 3, 11, 15, 19, 20, Khorbusuonka River, Olenyok Uplift; 21, 22, Lena River lower reaches, Ulakhan-Ald’yarkhay Brook; Le, Lena River middle reaches, Tiktirikteekh and Bydyangaya Brooks; Al, Aldan River, between sections “Dvortsy” and Ulakhan-Sulugur. Margin of the Siberian Platform is shown by a dashed line in A.
The Chuskuna Formation belongs to the uppermost part of the Kessyusa Group and was earlier referred to as the Upper Member of the Kessyusa Formation (now Group). It represents a depositional sequence separated from the underlying Mattaia Formation and overlying Erkeket Formation by flooding surfaces (Grazhdankin et al. 2020). The lower part of the Chuskuna Formation in section 11 consists of wave-rippled sandstone with carbonate cement. The upper part is oolitic sandstone with intercalated gravelstone and oolitic carbonates. In section 15, the Chuskuna Formation is ca. 12 m thick. It consists of hummocky and wave-rippled sandstone, crossbedded oolitic sandstone, siltstone with sandstone interbeds, and intraclastic-oolitic limestone (Fig. 3). The upper part of the Chuskuna Formation (by comparison with its stratotype) is truncated in both sections (Knoll et al. 1995; Grazhdankin et al. 2020). Erosional truncation of the Kessyusa Group is confirmed herein by a sharp drop in δ13C values at the boundary with the overlying red-coloured lime mudstone and wackestone of the Erkeket Formation (Fig. 3).
Section 3 (71°27.3’ N, 123°57.5’ E), previously described as section 2 (Korovnikov 2002) and section 1 (Korovnikov and Tokarev 2018) is exposed at the right bank of the Khorbusuonka River, ca. 10 km downstream of the mouth of Mattaia (Fig. 3) and comprises carbonates of the Erkeket, Kuonamka and lower Yunkyulyabit-Yuryakh formations.
Section 19 (71°28.8’N, 123°50.0’E), on the left bank of the Khorbusuonka River (Fig. 3) is situated ca. 20 km downstream of the mouth of Mattaia River. It has been described as section 4 (Korovnikov 2002) and section 2 (Korovnikov and Tokarev 2018) and studied for palaeomagnetism by Gallet et al. (2003, 2019), who revealed a very high magnetic reversal frequency during Cambrian Age 4–Drumian (Toyonian–Mayan regional ages). The section exposes carbonates of the upper part of the Erkeket Formation and overlying Kuonamka, Yunkyulyabit-Yuryakh, and lower Tyussala formations. The Erkeket Formation is also accessible in adjacent section 20 herein (71°29’N, 123°50.8’E) representing section 5 of Korovnikov (2002).
Sections 21 and 22 are situated on the right bank, in the lower reaches of the Lena River, 0.1–0.5 km upstream (section 21) and at the left side (section 22) of the mouth (71°35’N, 127°20.5’E) of the Ulakhan-Ald’yarkhay Brook (Fig. 1; SOM 2: fig. 1). This section, situated in the Bulkur Anticline, was previously investigated by Repina et al. (1974), Lazarenko and Repina (1983), Vasiljeva (1998), Bushuev et al. (2014), and others. Samples for fossil extraction and geochemical analyses herein were collected from carbonates of the Tyuser and Sekten formations (Fig. 4).
The Lower Member of the Tyuser Formation, up to 21 m thick, includes two dolerite sills (SOM 2: figs. 2–4), with commonly occurring pillow-shaped structures (Repina et al. 1974 and herein). The latter suggest rapid cooling, probably by the seawater, during extrusion of basaltic lavas. The lower sill is ca. 7 m thick, separated from the underlying upper Ediacaran argillaceous and silty bituminous limestone of the Kharayutekh Formation (Pelechaty et al. 1996a, b) and from the upper sill by quartz-feldspathic sandstone. The upper sill varies in thickness from 1 to 7 m and shows a deep erosional cold upper contact brecciated by ca. 1 m deep fractures and hosting lenses of cemented clastic material from the overlying basal (transgressive) unit of the Middle Member of the Tyuser Formation (SOM 2: figs. 2–4).
The basal Middle Member contains polymictic conglomerate with fragments of dolerites and carbonates, sandstone with carbonate cement passing up the section into reddish bedded silty and argillaceous limestone (SOM 2: fig. 2). The lower conglomerate and sandstone beds of the basal Middle Member vary in thickness from 2 to 4 m depending on the depth of erosion in the upper dolerite. The overlying beds of the Middle Member consist of silty limestone (ca. 14 m herein and after Bushuev et al. (2014), but 18–20 m, after Repina et al. (1974) of Unit 5, calcimicrobial mud mounds surrounded by argillaceous bioclast grainstone-packstone of Unit 6 (see also Kruse et al. 1995: 314), and argillaceous and brecciated silty limestone of Unit 7 (numbering of units throughout after Repina et al. 1974). The Middle Member is ca. 35 m thick.
The Upper Member is gray-colored and ca. 70 m thick (Repina et al. 1974; Bushuev et al. 2014). It comprises argillaceous bioclastic limestone and dolomitic limestone, brecciated in the upper part of Unit 8, dolostone and dolomitic limestone of Unit 9, and bioclastic sandy and silty limestone with polymict gravelstone and argillaceous limestone of Unit 10, containing oolitic dolomite in its upper part.
The Tyuser Formation in sections 21–22 is 128 m thick and overlain by the ca. 80 m thick Sekten Formation. The latter generally consists of argillaceous limestone subdivided by Repina et al. (1974) into Units 11–13 (Units 1–3 by Lazarenko and Repina 1983). A poorly exposed 2.5 m thick unit of marl and argillite at the top of the formation is indicated by Lazarenko and Repina (1983). The highest sample from the Sekten Formation was taken at 208 m from the bottom of section 22, below a ca. 8 m interval of the outcrop covered by scree. Reddish limestone layers typical of the Mayaktakh Formation (Mayan stage) are observed immediately above the scree. Biostratigraphic subdivision of the upper Tyuser and Sekten formations is provided by Astashkin et al. (1991; Fig. 4). Updated trilobite-based biostratigraphy of the Tyuser Formation is further discussed by Bushuev et al. (2014).

Fig. 2. Stratigraphic columns of sections 96-1 and 96-2, Malaya Kuonamka River and section 96-8, Bol’shaya Kuonamka River, Siberia, Russia (after Kouchinsky et al. 2011: fig. 2). Arrows with numbers indicate derivation of fossiliferous samples. Botoman Stage is correlated herein with the Cambrian Stage 4, in accordance with Geyer (2019). Asterisk A, indicates occurrences of Davidonia anabarica, Enigmaconus? pyramidalis, Pelagiella sp. 1 and 2, Majatheca tumefacta, Tetratheca clinisepta, “Lenatheca groenlandica”, Conotheca cf. mammilata, Parkula cf. esmeraldina (see Kouchinsky et al. 2015a). Asterisk B, indicates occurrences of Anabaroconus sibiricus, Eotebenna viviannae, Kuonamkaella oliva, Leptostega cf. hyperborea, Pelagiella sp. cf. Cambretina mareki, Pelagiella sp. cf. Costipelagiella zazvorkai, Pojetaia dentifera, Protowenella sp., Pseudomyona queenslandica, and Yochelcionella angustiplicata (see Gubanov et al. 2004b and Kouchinsky et al. 2011). Abbreviations: Ac., Acadoparadoxides; An., Anabaraspis; B., Bergeroniellus expansus; P., Paramicmacca; Fm., Formation; H., Homotreta; O-S, Ovatoryctocara–Schistocephalus; T., Triplagnostus.
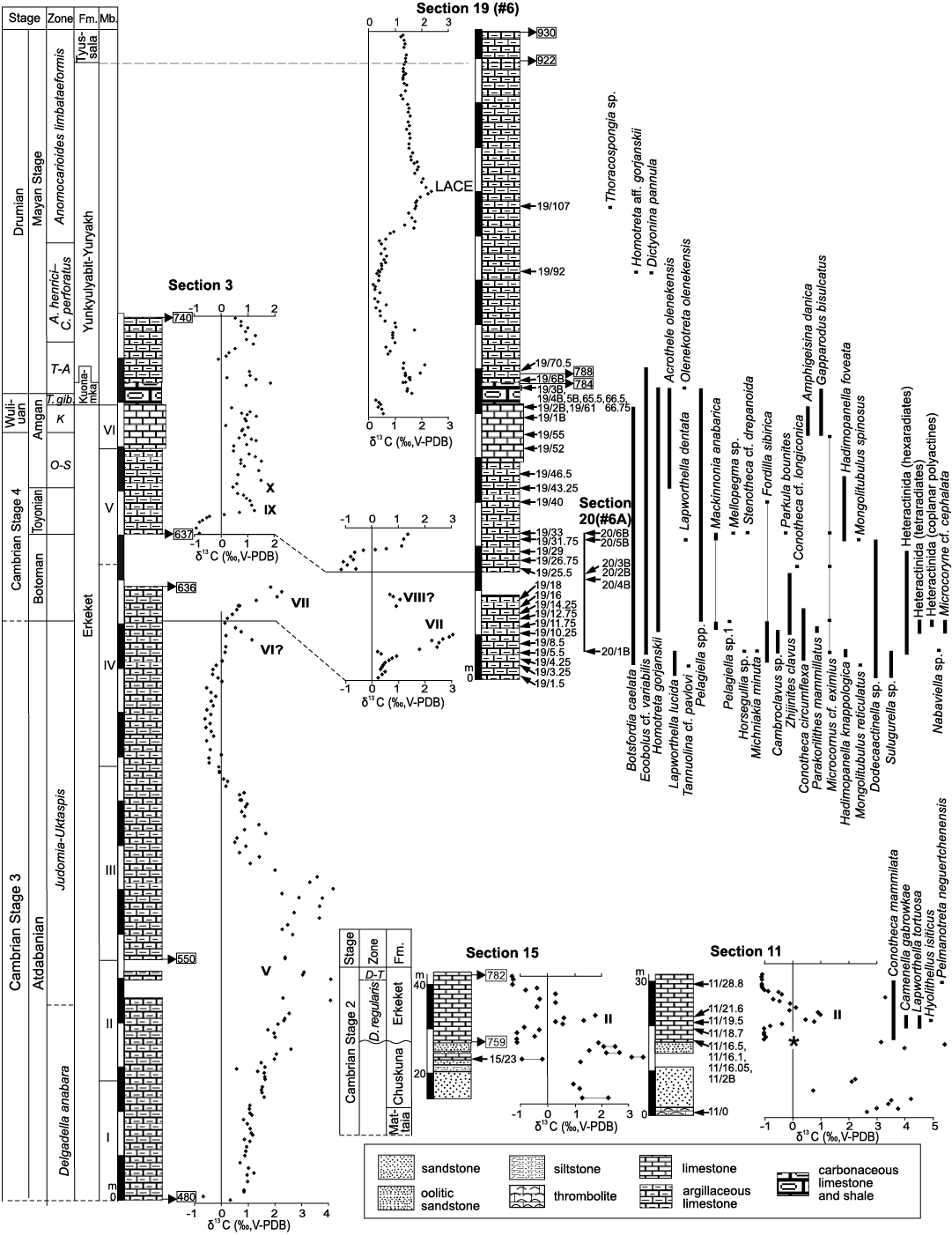
Fig. 3. Stratigraphic columns of sections 3, 11, 15, 19, and 20, Khorbusuonka River, Siberia, Russia. Arrows with numbers indicate derivation of fossiliferous samples. Roman numbers designate carbon isotope peaks after Brasier et al. (1994). Numbers in rectangles indicate position of selected samples analyzed for δ13Ccarb (SOM 4). Asterisk indicates occurrences of Bemella jacutica, Watsonella crosbyi, Aldanella sibirica, Halkieria proboscidea, Stellispinella lenaica, Archaeopetasus typicus, Egdetheca aldanica, and Markuelia secunda. Abbreviations: A., Anopolenus; C., Corynexochus; D., Dokidocyathus; D-T, Dokidocyathus lenaicus–Tumuliolynthus primigenius; Fm., Formation; K, Kounamkites; LACE, Lower Anomocarioides limbataeformis Carbon isotope Excursion; Mb., Member; O-S, Ovatoryctocara–Schistocephalus; T. gib., Tomagnostus gibbus; T-A, Tomagnostus fissus–Acadoparadoxides sacheri.
Chemostratigraphy and correlation
The upper Kessyusa Group shows positive δ13Ccarb values (Fig. 3; Knoll et al. 1995; Grazhdankin et al. 2020) and can be correlated with the lower part of Cambrian Stage 2 in other regions of the Siberian Platform (Landing and Kouchinsky 2016; Kouchinsky et al. 2017, and references therein). Two significant positive δ13Ccarb excursions reaching values higher than +4‰ are reported, respectively, from the upper Mattaia and Chuskuna formations from the Olenyok Uplift (Grazhdankin et al. 2020). The positive δ13C excursion in the upper Mattaia Formation can be correlated with the δ13C peak Iʹ described from the Anabar Uplift (Kaufman et al. 1996; Kouchinsky et al. 2017) and designating the Fortunian–Cambrian Stage 2 boundary (Landing and Kouchinsky 2016). Correlation of peak Iʹ with the δ13Ccarb positive excursion I from the dolomitic facies of the upper Ust’-Yudoma Formation in southeastern part of the Siberian Platform is plausible (Pokrovsky and Missarzhevsky 1993; Brasier et al. 1996: 469; Kouchinsky et al. 2017, but Kouchinsky et al. 2001, 2007). The presence of index-fossil Watsonella crosbyi below the δ13C peak with the absolute age 529.7 ± 0.3 Ma in the upper Mattaia Formation (Grazhdankin et al. 2020) as well as in the overlying Chuskuna Formation (Fig. 3 herein) indicates the Watsonella Zone, Cambrian Stage 2. It is thus possible to correlate the Chuskuna Formation and its younger positive δ13C values (Fig. 3) with those known between peaks Iʹ and II from the Anabar Uplift. The equivalent beds described from South China and containing ZHUCE (ZHUjiaqing Carbon isotope Excursion, after the Zhujiaqing Formation, South China, Zhu et al. 2006) are constrained between ca. 527 and 526 Ma (Tsukui et al. 2017).
A stratigraphic unconformity between the Kessyusa Group and Erkeket Formation in sections 11 and 15 along the Khorbusuonka River (Fig. 3) is marked by a sharp, 3–4‰ drop in δ13C values at their sequence boundary. This isotopic shift is similar in magnitude to that available within the Nochoroicyathus sunnaginicus Zone, Tommotian stage (correlated with Cambrian Stage 2), at the boundary between the Ust’-Yudoma and Pestrotsvet formations at the Aldan River (Magaritz et al. 1991; Brasier et al. 1994), reaching ca. -2‰ (Brasier et al. 1994) or ca. -3‰ (He et al. 2019: fig. 1). In combination with palaeontological evidence, the basal part of Erkeket Formation containing ca. -1‰ negative δ13C values belongs to the lower Dokidocyathus regularis Zone. This negative excursion can be correlated with SHICE (SHIyantou Carbon isotope Excursion, after the Shiyantou Formation, South China; Zhu et al. 2006), in the middle part of Cambrian Stage 2.
The lowermost positive peak in the Erkeket Formation, sections 11 and 15 (Fig. 3), is correlated with the carbon isotope excursion II of the Siberian reference scale (Brasier et al. 1994; Rozanov et al. 2008) corresponding to the Dokidocyathus regularis Zone, Tommotian stage (correlated with the upper part of Cambrian Stage 2). Peak II reaches +1.5‰ at Malaya Kuonamka and Bol’shaya Kuonamka (Kouchinsky et al. 2015a), Aldan (Brasier et al. 1994), Selinde (Kouchinsky et al. 2005) and Gonam, Altitude 1291 m (AK, unpublished data), whereas at the Sukharikha River section (Kouchinsky et al. 2007), the peak is ca. +1 ‰. A similar isotopic excursion is shown for the lower Erkeket Formation by Grazhdankin et al. (2020: fig. 6). The lowermost peak at the base of the Ulakhan-Ald’yarkhay section (Fig. 4) does not reach 0‰ and in combination with palaeontological data indicating correlation with the Dokidocyathus regularis Zone can be interpreted as excursion III (Brasier et al. 1994) in the upper part of the D. regularis Zone, Tommotian stage (correlated with the upper part of Cambrian Stage 2). Thus, basal beds of the Middle Member of the Tyuser Formation suggest transgressive onlap ca. 1 Myr younger than similar facies of the basal Erkeket Formation in the Khorbusuonka localities.
The boundary between the Tommotian and Atdabanian stages (~Terreneuvian–Cambrian Series 2 boundary) coincides with the first trilobites found in the Ulakhan-Ald’yarkhay section (Bushuev et al. 2014). The level is situated in the rising trend of δ13C values following the negative trough attributed to the upper Tommotian Dokidocyathus lenaicus–Tumuliolynthus primigenius Zone (Figs. 3, 4). The lower Atdabanian boundary at the Khorbosuonka River is drawn at ca. 11 m from the base of the Erkeket Formation in section 15, where the trilobite Profallotaspis sp. is reported (but not illustrated; Astashkin et al. 1991; Korovnikov 2002). Carbon isotope analyses indicate, however, that the studied interval in sections 11 and 15 (Fig. 3), ca. 15 m of the lower Erkeket Formation belongs to the Tommotian stage (correlated with the upper part of Cambrian Stage 2).
Peak IV of the Siberian reference scale is the first post-Terreneuvian maximum in the δ13C record located in the Repinaella Zone and marks the basal Delgadella anabara Zone of the Atdabanian stage (Brasier et al. 1994; Kouchinsky et al. 2015a), correlated with the Cambrian Stage 3. The peak is named EAREZE (Early Atdabanian/REpinaella Zone Excursion) by Geyer (2019). Peak IV has been proposed as a global horizon to define the base of Series 2 (Landing et al. 2013 and references therein; Landing and Kouchinsky 2016) and has a U-Pb volcanic zircon age of ca. 520 Ma (Landing et al. 2021c).
Carbon isotope excursion V of the Siberian reference scale continues through ca. 100 m, from Member 1 at the bottom of section 3 at Khorbusuonka to Member 3 of the Erkeket Formation (Fig. 3) reaching ca. +4‰ δ13C in that section. This is nearly equal to the thickness of corresponding strata in the upper Krasnoporog Formation at Sukharikha (Kouchinsky et al. 2007: fig. 2). The excursion is well-defined, ca. +3‰ in magnitude and extending over ca. 50 m from base of the Upper Member of the Tyuser Formation at the Ulakhan-Ald’yarkhay section (Fig. 4), which is similar to excursion V in the reference section from the middle reaches of the Lena River (Brasier et al. 1994). The same excursion is recorded within just ca. 20 m of the uppermost Emyaksin Formation (Kouchinsky et al. 2015a: fig. 2). This peak represents CARE (Cambrian Arthropod Radiation isotope Excursion) in the Cambrian Stage 3 of South China claimed to be recognizable worldwide (Zhu et al. 2006). Peak values of the excursion span the Judomia–Uktaspis (Prouktaspis) Zone lower boundary. A less prominent excursion VI is not as clearly recognisable (Figs. 3, 4) as in the Siberian reference section (Brasier et al. 1994), eastern Anabar Uplift (Kouchinsky et al. 2001, 2015a), and Sukharikha (Kouchinsky et al. 2007). Its upper boundary separating the Atdabanian and Botoman stages (correlated herein with the boundary between Cambrian stages 3 and 4) in the Siberian reference section (Brasier et al. 1994; Rozanov et al. 2008) is marked by a trough between excursions VI and VII.
The positive excursion VII belongs to the Bergeroniellus micmacciformis–Erbiella Zone, Botoman stage (correlated with the Cambrian Stage 4) and can be correlated across the Siberian Platform (Kouchinsky et al. 2001, 2007, 2015a). The lower Botoman peak VII is well expressed and reaches ca. +3‰ in the Khorbusuonka (Fig. 3) and reference sections (Brasier et al. 1994), and ca. +2‰ in the Bol’shaya Kuonamka (Kouchinsky et al. 2015a; Fig. 2) and Ulakhan-Ald’yarkhay (Fig. 4) sections. The position of this peak at the boundary between the Tyuser and Sekten formations supports biostratigraphic correlation of the boundary within the lower Botoman Bergeroniellus micmacciformis–Erbiella Zone (Astashkin et al. 1991: 60). This interval of heavier carbon isotope values is also known from South China (Gondwana) as MICE (MIngxinsi Carbon isotope Excursion), a prominent positive excursion in the Mingxinsi Formation, Guizhou, China, attributed to the lower part of Cambrian Stage 4 (Zhu et al. 2006). MICE was initially correlated with the VIII interval in the Sinsk Formation (Brasier et al. 1994; Rozanov et al. 2008; He et al. 2019), which is neither as prominent as VII nor apparent elsewhere, but may well represent a South Chinese equivalent to the merged Siberian excursions VII–VIII. There is no identifiable peak VIII in the Sekten Formation compared to the reference section (Fig. 4). The position of MICE within the Cambrian Stage 4 depends on understanding of the boundary between Cambrian stages 3 and 4, which is not yet ratified. Herein (but in Kouchinsky et al. 2015a), the lower Botoman stage is correlated with the lower part of Cambrian Stage 4, in accordance with presence of Calodiscus and Serrodiscus on the Siberian Platform selected as possible Stage 4 indicators (e.g., Geyer 2019).
The Toyonian stage is marked by the onset of a rising trend in carbon isotope values, from a negative trough, ca. -1‰ in the Khorbusuonka section (Fig. 3) and ca. -2‰ at Ulakhan-Ald’yarkhay (Fig. 4). This is also the case in the reference sections, where peaks IX and X mark, respectively, the Bergeroniellus ketemensis Zone of the Toyonian stage and Toyonian–Amgan stages boundary (Brasier et al. 1994; Brasier and Sukhov 1998; He et al. 2019: fig. 1). Carbon isotope values indicative of the peak X on the southeastern Siberian Platform are herein distinguished in the lowermost Amgan stage, from the upper Erkeket (Fig. 3) and Sekten (Fig. 4) formations. The δ13C trough between the paired peaks VII–VIII and IX–X recognised herein can be correlated with the AECE (Archaeocyathid Extinction Carbon isotope Excursion) identified in South China within the Cambrian Stage 4 (Zhu et al. 2006; Chang et al. 2017).
Subsequently, decreasing values are registered through the Kuonamka Formation of the eastern Anabar Uplift (Fig. 2; Brasier and Sukhov 1998), Khorbusuonka (Fig. 3) and section Chekurovka in lower reaches of the Lena River (IVK unpublished data) as well as in the same formation of the Olenek Uplift and in the Elanskoe Formation of the middle Lena River (Wotte et al. 2011). Within this interval, carbon isotope values fluctuate through the Kuonamka Formation at Malaya Kuonamka River, where cherts and bituminous shale predominate and oxidised organic carbon may have altered the carbon isotopic signal (Fig. 2). Carbon isotope record through the carbonate-predominated Kuonamka Formation at the Bol’shaya Kuonamka River shows a considerably smaller amplitude of oscillations, which is compatible with better preservation of the original δ13Ccarb values (Fig. 2). The carbon isotope trough across the boundary between the Wuliuan and Drumian stages corresponds to DICE, Drumian Carbon isotope Excursion (Zhu et al. 2006). The boundary is correlated in Siberia with the boundary between regional Amgan and Mayan stages (Zhamoida 2016).
The Yunkyulyabit-Yuryakh Formation, assigned to the Drumian Stage, conformably overlies the Kuonamka Formation at Khorbusuonka and records positive excursions in its lowermost part, 1.5–2‰ in magnitude (Fig. 3). Similar positive values are also available from the uppermost Kuonamka and lower Olenyok formations along the Malaya Kuonamka and Bol’shaya Kuonamka rivers (Fig. 2; Brasier and Sukhov 1998), lower Mayaktakh Formation in Chekurovka (IVK unpublished data) and can be correlated with an unnamed interval of positive excursion in the lower Drumian Stage by Zhu et al. (2006). The upper part of the Anomocarioides limbataeformis Zone (Drumian Stage) shows δ13C values near 0‰ in the Kulyumbe section (Kouchinsky et al. 2008) but the lower A. limbataeformis Zone (not present in the Kulyumbe section) demonstrates rising values, from a ca. 0‰ trough to a 2.5‰ peak, through the Yunkyulyabit-Yuryakh Formation in the Khorbusuonka section (Fig. 3). The latter excursion is termed herein LACE, Lower Anomocarioides limbataeformis Carbon isotope Excursion. The record remains fairly steady throughout the rest of section 19 (Fig. 3) and corroborates observations of near zero values in the upper part of the Drumian Stage from the Global Standard Chronostratigraphic Scale for the Cambrian System (Zhu et al. 2006).

Fig. 4. Stratigraphic columns of sections 21 and 22, Ulakhan-Ald’yarkhay Brook, lower reaches of the Lena River, Siberia, Russia. Arrows with numbers indicate derivation of fossiliferous samples. Asterisk indicates occurrences of Lapworthella tortuosa, Halkieria proboscidea, Stellispinella lenaica, Archaeopetasus pachybasalis, Conotheca mammilata, Egdetheca aldanica, Hyolithellus isiticus. Units after Repina et al. (1974). Abbreviations: A-L, Anabaraspis splendens–Lermontovia grandis; B-E, Bergeroniellus micmacciformis–Erbiella; D., Dokidocyathus; D-T, Dokidocyathus lenaicus–Tumuliolynthus primigenius; O-S, Ovatoryctocara–Schistocephalus; P, Profallotaspis; R, Repinaella.
Material and methods
Insoluble residues containing microfossils were extracted from 1–3 kg calcareous samples with a buffered solution of ca. 10% acetic acid. Residues with initially high argillaceous content were then separated from water-clay suspension by heating it to the boiling point and subsequent careful decanting. This simple purification technique yields cleaner microfossils almost without clay particles attached to their surface and enables better imaging. The microfossils were mounted and coated with gold for examination with scanning electron microscopes at the Swedish Museum of Natural History, including a Hitachi S4300 in 2005, 2017–2018 and a Quanta 650 FEG in 2018–2021. Abundances of fossils in samples are provided in SOM 3.
Carefully selected micritic carbonate was microdrilled from 675 cut, polished and cleaned hand samples and simultaneously analysed for δ13Ccarb and δ18Ocarb following reaction with 100% orthophosphoric acid at 75°C. The amount of powder prepared for a single analysis was 100–400 μg, with 1–3 spots analysed per sample. The analyses were obtained with a Finnigan MAT 252 equipped with an automated online Kiel Device at the Department of Geology and Geochemistry, Stockholm University in 1999–2000 (Malaya Kuonamka and Bol’shaya Kuonamka sections 96-1, 96-2, 96-3, 96-6, 96-7, and 96-8), and 2004 (Ulakhan-Ald’yarkhay sections 21 and 22). Samples from the Khorbusuonka sections 3, 11, 15, 19, and 20 were analysed with a Finnigan MAT 253 equipped with a ThermoFinnigan Gasbench II at the Department of Earth and Space Sciences, University of California, Los Angeles in 2004 and 2009. Data are reported in per mil (‰) notation relative to the Vienna Pee Dee Belemnite standard (VPDB). Secondary standards used were NBS-19, IAEA-CO-1, IAEA-CO-8, and an internal laboratory standard, CARM-1. The accuracy of the analyses for δ13Ccarb and δ18Ocarb was always better than ±0.1‰. Additional analyses of samples from the Khorbusuonka sections were performed in 2021 on an Elementar PRECISION stable isotope ratio mass spectrometer, using an Elementar iso FLOW system at the Wolfson Laboratory, School of Geosciences, Grant Institute, University of Edinburgh, UK. The standard deviation for replicate analyses (n = 27) of an in-house coral standard (Reference COR1D) measured alongside samples was better than ±0.06‰ for δ13Ccarb and ±0.06‰ for δ18Ocarb.
All measurements are available in SOM 4. It is accepted herein that very low δ18O values indicate resetting of δ18O during diagenetic alteration, e.g., by meteoric waters and hydrothermal fluids (Kaufman and Knoll 1995). Such results showing δ18O <-11‰ are available from cements of the Chuskuna Formation, mixed siliciclastic and carbonate deposits of the upper Kessyusa Group (marked in red in SOM 4). Samples with δ18O >-11‰ are used herein for chemostratigraphy.
Faunal composition
Representatives of several groups of skeletal fossils, including brachiopods, molluscs, arthropods, echinoderms, sponges, and taxa with problematic affinities are described herein from the Cambrian Stage 2 through the Drumian Stage (SOM 3). Brachiopods are represented by the class Lingulata Gorjansky and Popov, 1985, and Paterinata Williams, Carlson, Brunton, Holmer, and Popov, 1996. Most abundant are lingulates of the order Acrotretida Kuhn, 1949, including several thousand calcium phosphatic shells of Homotreta gorjanskii (Pelman, 1973) (Figs. 8–10) recovered from all of the sections through the Botoman–lower Mayan stages (correlated with the Cambrian Stage 4–lower Drumian Stage). Morphologically different H. cf. gorjanskii (Fig. 11) and H. aff. gorjanskii (Fig. 12) from the Drumian Stage are left in open nomenclature pending revision. Linnarssonia bulcurica Pelman in Pelman and Pereladov, 1986 (Fig. 13), is described from eastern flanks of the Anabar Uplift, from the Wuliuan–Drumian transitional beds. The aforementioned acrotretids have not been described beyond the Siberian Platform.
Among the lingulids (Order Lingulida Waagen, 1885), Eoobolus variabilis (Pelman, 1977) (Figs. 6, 7) is the most abundant form herein. Hundreds of specimens are recovered from all of the sections, from the Botoman–lower Mayan stages. Eoobolus priscus (Poulsen, 1932) (Fig. 5) is only identified herein from the Judomia–Uktaspis (Prouktaspis) Zone, Atdabanian stage (correlated with the Cambrian Stage 3) of the eastern flanks of the Anabar Uplift, where it is represented by several dozens of valves. Beyond Siberia, both forms are known from the Cambrian Stage 3–Drumian Stage of Laurentia, South Australia and Antarctica. Over a hundred calcium phosphatic valves of the lingulids Acrothele olenekensis Pelman in Pelman and Pereladov, 1986 (Fig. 15) and Botsfordia caelata (Hall, 1847) (Fig. 14) are found from the Amgan and lower Mayan stages (Cambrian Stage 4–lower Drumian Stage), from sections of the Khorbusuonka River and eastern Anabar Uplift. Botsfordia caelata has also been collected from the Botoman stage herein and described from the Cambrian Stage 4–Guzhangian Stage of Laurentia.

Fig. 5. Lingulid brachiopod Eoobolus priscus (Poulsen, 1932) from the lower Cambrian Emyaksin Formation, Bol’shaya Kuonamka River, Siberia, Russia; samples 6/66.2 (A–I, L) and 7/55 (J, K). A–E, L. Ventral valves, SMNH Br151173–151177, 151272, respectively. E2, close up of E1; L2, L3, close-ups of the brephic shell. F–K. Dorsal valves, SMNH Br151178–151183, respectively. Scale bar: 1 mm (A–E1, F–K), 500 µm (L1), 250 µm (E2), 60 µm (L2), and 8 µm (L3).
The oldest paterinid brachiopods in the material are represented by several ventral valves of Paterinida gen. and sp. indet. (Fig. 19) from the Dokidocyathus regularis Zone, Tommotian stage (correlated with the Cambrian Stage 2) of the Khorbusuonka and Lena rivers. Several dozens of calcium phosphatic shells and fragments of Pelmanotreta neguertchenensis (Pelman, 1977) (Fig. 16) are also collected from the Dokidocyathus lenaicus–Tumuliolynthus primigenius Zone, Tommotian stage–Repinaella Zone, Atdabanian stage (correlated with the upper part of Cambrian Stage 2 and Stage 3), eastern Anabar Uplift and Khorbusuonka. Olenekotreta olenekensis Ushatinskaya, 1997 (Fig. 17), is available as ten disarticulated calcium phosphatic valves and fragments in a single sample from eastern Anabar Uplift, Kounamkites Zone, lower Amgan stage (correlated with the Wuliuan Stage). A single sample from the Anopolenus henrici–Corynexochus perforatus Zone, Mayan stage (correlated with the Drumian Stage) from the Khorbusuonka River yielded five shells of Dictyonina pannula (White, 1874) (Fig. 18). The latter species is also known from the Cambrian Stage 4–Wuliuan Stage of Laurentia.
A new representative of the stem-group Brachiopoda from the lower Botoman stage (correlated with Cambrian Stage 4) of Khorbusuonka is described in open nomenclature owing to fragmentary preservation (Fig. 20). It shows a previously unknown combination of features typical of tannuolinids, mickwitziids and linguliform brachiopods and is most similar to Oymurania Ushatinskaya in Kouchinsky et al., 2015b. The latter occurs within the Atdabanian and lower Botoman stages of the Siberian Platform. Two fragmentary mitral sclerites of Tannuolina cf. pavlovi Kouchinsky, Bengtson, and Murdock, 2010 (Fig. 21), are found in a single sample from the Khorbusuonka River, from the lower Botoman stage. Sclerites of other tommotiids, Camenella Missarzhevsky in Rozanov and Missarzhevsky, 1966 (Fig. 22), and Lapworthella Cobbold, 1921 (Fig. 23), occur in several samples from the Khorbusuonka and Lena rivers, but they are not common. Those include several dozens of calcium phosphatic sclerites of Camenella garbowskae Missarzhevsky in Rozanov and Missarzhevsky, 1966, from the Tommotian stage (correlated with the upper part of Cambrian Stage 2) and a few other sclerite types, probably, morphological varieties of C. garbowskae, such as C. cf. garbowskae (Fig. 22D; Kouchinsky et al. 2015a: fig. 39), C. aff. garbowskae (Fig. 22K; Kouchinsky et al. 2015a: fig. 43), and Camenella plana (Missarzhevsky in Rozanov et al., 1969) (Fig. 22F; Kouchinsky et al. 2015a: fig. 40). Sclerites of Lapworthella Cobbold, 1921, are represented by three species, with several dozens of specimens each. Lapworthella tortuosa Missarzhevsky in Rozanov and Missarzhevsky, 1966, occurs in eastern Anabar Uplift, Khorbusuonka and lower Lena River localities, in the Dokidocyathus regularis Zone, Tommotian stage. Lapworthella lucida Meshkova, 1969, is reported herein from the uppermost Atdabanian and lower Botoman stages (correlated with the uppermost part of Cambrian Stage 3 and lower part of Cambrian Stage 4, respectively) of the Khorbusuonka River and lower reaches of the Lena River. Lapworthella dentata Missarzhevsky in Rozanov et al., 1969, described as Lapworthella schodackensis by Rozanov et al. (2010: fig. 58) from the southeastern Siberian Platform and reported from Atdabanian–Botoman strata by Kouchinsky et al. (2015a), ranges into the Toyonian stage at Khorbusuonka.
Conchiferan molluscs are extracted from carbonates as calcium phosphatic internal moulds or coatings of millimeter-sized cap-shaped shells of helcionellids, coiled forms, such as aldanellids and pelagiellids, and bivalves. Rare silicified shells of helcionellids occur within Wuliuan–Drumian strata of the Kuonamka Formation. The most abundant form Aldanella sibirica Parkhaev and Karlova, 2011 (Figs. 28, 29), is represented by hundreds of specimens and occurs in the basal Erkeket Formation, Dokidocyathus regularis Zone, Tommotian stage. A similar form A. cf. sibirica, was described from the lower part of Cambrian Stage 2 on eastern flanks of the Anabar Uplift (Kouchinsky et al. 2017).
Several hundred specimens of the helcionellid Davidonia anabarica (Parkhaev, 2005) (Fig. 24A, B), are recovered from lower Botoman strata of the Erkeket Formation at the Khorbusuonka River as well as from the upper Emyaksin Formation of the eastern Anabar Uplift (Kouchinsky et al. 2015a). The species probably ranges into the Toyonian Stage (correlated with the Cambrian Stage 4) of the Khorbusuonka River and up to the Kounamkites Zone, Amgan Stage (correlated with the Wuliuan Stage) of the Lena River (SOM 3). Davidonia anabarica is very similar to Davidonia rostrata (Zhou and Xiao, 1984) reported from the Cambrian stages 3–4 and lower Wuliuan Stage of Gondwana (Bengtson et al. 1990; Wotte 2006; Claybourn et al. 2019; Li et al. 2019) and Laurentia (Skovsted 2004).
Bivalves, Fordilla sibirica Krasilova, 1977 (Figs. 26, 27A–C), are collected from the Delgadella anabara–Nevadella Zone, Atdabanian stage of the lower Lena River and the lower Botoman–Toyonian stages (correlated with the Cambrian Stage 4) of Khorbusuonka. Because of preservation, however, most of the internal moulds cannot be reliably distinguished from the similar species Pojetaia runnegari Jell, 1980, available worldwide from the Cambrian Stage 3. Several internal moulds of a recently described species, Pojetaia dentifera Kouchinsky, Bengtson, Clausen, Gubanov, Malinky, and Peel, 2011 (Fig. 27D–F), can be identified from the upper Toyonian Stage (correlated with the Cambrian Stage 4) of the lower Lena River, extending the range of this species from the lower Drumian Stage of Siberia as originally described (Kouchinsky et al. 2011). In addition, two moulds and a replica of the external shell surface of Watsonella crosbyi Grabau, 1900 (Fig. 25), from the Cambrian Stage 2 are illustrated from the Chuskuna Formation at Khorbusuonka (Fig. 25).
Other conchiferans are rare in the collection (Fig. 24; SOM 3), such as Bemella jacutica (Missarzhevsky in Rozanov and Missarzhevsky, 1966) and Obtusoconus cf. mirabilis Vasiljeva, 1990, from the Dokidocyathus regularis Zone, Tommotian stage, which are known to occur in the Cambrian Stage 2 elsewhere on the Siberian Platform. Stenotheca cf. drepanoida from the Toyonian stage herein is similar to Stenotheca drepanoida (He and Pei in He et al., 1984) described from the Cambrian Series 2 of South China, South Australia, and Siberia (Vendrasco et al. 2011b). At least two unidentified species of Pelagiella Matthew, 1895, are reported herein from Cambrian Series 2 beds in sections of the Khorbusuonka and lower Lena rivers. Those are compared to phosphatised and silicified material from the Cambrian Series 2, Wuliuan and lower Drumian stages, described from the eastern Anabar Uplift (Kouchinsky et al. 2011, 2015a). An assemblage of conchiferan molluscs from the eastern Anabar Uplift includes other species and is more diverse probably owing to an originally more diverse and now phosphatised fauna in the lower Botoman beds of the upper Emyaksin Formation (Kouchinsky et al. 2015a) and better preservation of fossils due to silicification in the Kuonamka Formation (Gubanov et al. 2004b; Kouchinsky et al. 2011).
Two forms can be attributed to multiplated molluscs, stem-group chitons, such as problematic shell 1 (Fig. 31) from the Toyonian stage and problematic shell 2 (Fig. 32) from the lower Botoman stage (correlated with the Cambrian Stage 4). Other representatives of the aculiferan molluscan lineage, Halkieria proboscidea (Meshkova, 1969) (Fig. 33), are a common form in the Tommotian stage of Siberia. Similar halkieriid sclerites have also been described from the Cambrian Stage 2 of South China (Parkhaev and Demidenko 2010) and France (Devaere et al. 2013).
Planispiral evolute secondarily phosphatised conchs and internal moulds of Michniakia minuta Missarzhevsky in Rozanov and Missarzhevsky, 1966 (Fig. 42) occur within the upper Cambrian Stage 3 and lower Stage 4 beds of the Siberian Platform (upper Atdabanian and lower Botoman stages). The only specimen reported from outside Siberia was derived from Spain (Gubanov et al. 2004a) and was broadly attributed to the Atdabanian stage. The apical morphology with a septum, fibrous microstructure of the porous wall and an in-place operculum in Michniakia are compatible with those in hyoliths, among which planispirally coiled forms are also reported (Feng et al. 2001; Kouchinsky et al. 2017; Peel 2021b).
The diversity of hyoliths (phylum uncertain, class Hyolitha Marek, 1963) herein can be assessed on the base of their opercula and conchs, although original associations between these two shell parts are rarely known. Hyoliths described herein are a few millimeters in size. Larger specimens, up to 150 mm long occur in rock but they are not phosphatised and cannot be extracted with chemical preparation. It is, thus, possible that some small phosphatised individuals represent not only separate species but also juveniles of larger forms.
Among hyoliths of the order Orthothecida Marek, 1966 (see Skovsted et al. 2020), most abundant forms herein are several hundred phosphatised conchs and opercula of Conotheca mammilata Missarzhevsky in Rozanov et al. 1969 (Fig. 40A–D) from the Cambrian stages 2 and 3 (see Kouchinsky et al. 2015a, 2017), silicified internal moulds of Triplicatella sp. (Fig. 46) with casts of opercula from the lowermost Drumian strata of the Kuonamka Formation at Khorbusuonka, and Cupitheca sp. (Fig. 44) represented by phosphatised conchs and probably opercula from the Cambrian Stage 4. Generally, Cupitheca-like hyoliths are interpreted herein as a polyphyletic group of orthothecid hyoliths with a circular cross-section and an apically convex terminal septum (Kouchinsky et al. 2015a, 2017, and herein).
Other orthothecids found herein are less abundant and represented by up to several dozen each of phosphatised conchs and opercula. Among them the following taxa can be defined. Conotheca cf. longiconica (Qian, 1978) (Fig. 40E, F), from the basal Toyonian stage (correlated with the Cambrian Stage 4) of the Khorbusuonka River and the Delgadella anabara–Nevadella Zone, Atdabanian stage (correlated with the Cambrian Stage 3) of the Lena River. Similar conchs are illustrated by Kouchinsky et al. (2015a: figs. 25E–G, 27E; 2017: fig. 52) from the Cambrian stages 2 and 3 of the eastern Anabar Uplift and reported from South China (Qian and Bengtson 1989). Conotheca circumflexa Missarzhevsky in Rozanov et al. 1969 (Fig. 40G, H), a hyolith with a helically twisted conch, occurs in the upper Atdabanian–lower Botoman strata (see also Kouchinsky et al. 2009, 2015a). Egdetheca aldanica Missarzhevsky in Rozanov et al. 1969 (Fig. 40I–K), is a characteristic form from the Tommotian stage of Siberia (correlated with the Cambrian Stage 2). Phosphatised opercula of Majatheca sp. are also found in our samples from the Dokidocyathus regularis Zone, Tommotian stage as well as six (three common and three rare) types of unidentified orthothecid opercula (SOM 3; Figs. 43, 47–50).
The first appearance of hyolith appendages (helens) is registered herein in the upper Dokidocyathus regularis Zone (Fig. 55D). It has been suggested by Skovsted et al. (2020) that the order Hyolithida Sysoev, 1957, represents a monophyletic lineage of hyoliths with opercular appendages, thereby distinguished from a polyphyletic array of other hyoliths (orthothecids) without helens (Zhang et al. 2018). The most abundant forms of hyolithids are represented by several dozens of calcium phosphatic internal moulds of conchs, Microcornus cf. eximius Duan, 1984, from the upper Cambrian Stage 2–Cambrian Stage 4. Microcornus eximius Duan, 1984 is known from the Cambrian Series 2 of South China (Duan 1984), North China (Pan et al. 2019), Australia (Bengtson et al. 1990), Antarctica (Wrona 2003), and Laurentia (Malinky and Skovsted 2004). Parakorilithes mammillatus He and Pei in He et al., 1984 (Fig. 53) occurs in the lower Botoman stage of the Khorbusuonka River. It was previously reported from the Cambrian Series 2 of Australia (Bengtson et al. 1990; Gravestock et al. 2001), Antarctica (Wrona 2003), North China (Pan et al. 2019), and Laurentia (Malinky and Skovsted 2004). Finally, Parkula bounites Bengtson in Bengtson et al., 1990 (Fig. 51A, B) is identified from the upper Toyonian and lower Amgan stages (Cambrian Stage 4–lowermost Wuliuan Stage) of the lower Lena River. The species is thought to be widespread and previously described from the Cambrian Series 2 of Australia (Bengtson et al. 1990; Gravestock et al. 2001), Antarctica (Wrona 2003), North China (Pan et al. 2019), Laurentia (Malinky and Skovsted 2004; Skovsted 2006a; Skovsted and Peel 2007; Devaere et al. 2019), and Siberia (Kouchinsky et al. 2015a).
Phosphatised sclerites of the problematic order Cambroclavida Conway Morris and Chen, 1991, are found on the Siberian Platform (see also Novozhilova and Korovnikov 2019). They occur in the lower Cambrian Stage 4 and are represented by both cambroclavid and zhijinid morphs, Cambroclavus sp. (Fig. 34A–J) and Zhijinites clavus (Mambetov in Mambetov and Repina, 1979; Fig. 34K–U), respectively. The latter is previously known from coeval beds of Kazakhstan.
Chancelloriides (incertae sedis order Chancelloriida Walcott, 1920) are mostly preserved as internal moulds of disarticulated sclerites in calcium phosphate and glauconite or less frequently occurring phosphatised sclerites and coatings or rarely silicified (from the Kuonamka Formation). Thousands of disarticulated sclerites are available from samples ranging in age from the lower Dokidocyathus regularis Zone of the Tommotian stage (correlated with the Cambrian Stage 2), wherefrom Stellispinella lenaica (Zhuravleva and Korde, 1955) is described (Fig. 35), to the Bergeroniellus expansus Zone of the Botoman stage (correlated with the Cambrian Stage 4) (SOM 3). Chancelloriid sclerites are especially abundant in the Delgadella anabara Zone of the Atdabanian stage (correlated with the Cambrian Stage 3) and the lower Amgan stage (correlated with the Cambrian Stage 4). Most of the sclerites are generally referable to Chancelloria spp., whereas Allonnia sp. and Archiasterella sp. can also be distinguished in some cases. A new species, Archaeopetasus pachybasalis Kouchinsky sp. nov. as well as Archaeopetasus typicus (Sipin, 1999) are described herein from the Dokidocyathus regularis Zone, Tommotian stage (Figs. 37, 38). They are reinterpreted as external sclerites of animals, probably allied with chancelloriids.
Calcium phosphatic spiniform sclerites of protoconodonts are relatively rare and represented mainly by Gapparodus bisulcatus (Müller, 1959) from Khorbusuonka (Fig. 56A, C, D). The species is widespread and known from the Toyonian–lower Mayan stages of the Siberian Platform (correlated with the Cambrian Stage 4–lower Drumian Stage) (Kouchinsky et al. 2011, 2015a), ranging from the Cambrian Series 2 through the Furongian Series elsewhere.
Palaeoscolecidans are represented by thousands of disarticulated calcium phosphatic sclerites of the form genus Hadimopanella Gedik, 1977, and belong to several species. Identified species are Hadimopanella knappologica (Bengtson, 1977) from the lower Botoman stage (Fig. 57), the Siberian Platform and Hadimopanella foveata Kouchinsky sp. nov. (Fig. 58) from the Toyonian and lower Amgan stages (correlated with the Cambrian Stage 4) of Khorbusuonka.
Except for trilobites and agnostoids, which are beyond the scope of this paper, other ecdysozoan fossils are rare. They include the phosphatised globular embryonic stage microfossils, Markuelia secunda Val’kov in Val’kov and Karlova, 1984, from the Dokidocyathus regularis Zone, Tommotian stage of Khorbusuonka (Fig. 60). Markuelia has been previously reported from the Cambrian Stage 2 of the southeastern Siberian Platform, the Miaolingian Series of China and Australia, the Furongian Series of China, and the Lower Ordovician of Laurentia (Dong et al. 2010). Calcium phosphatic lobopodian sclerites of Microdictyon robisoni Bengtson, Matthews, and Missarzhevsky, 1986 (Fig. 61A) are identified from the Kounamkites Zone, Amgan stage (correlated with the Wuliuan Stage) of the lower Lena River, also known from the Wuliuan Stage in Siberia (Kouchinsky et al. 2011) and Laurentia (Bengtson et al. 1986; Peel et al. 2016). Bradoriid arthropods are represented by the calcium phosphatic carapace of Cambria sibirica Neckaja and Ivanova, 1956 (Fig. 61D) from the Delgadella anabara–Nevadella Zone, Atdabanian stage (correlated with the Cambrian Stage 3), hollow spines of Mongolitubulus spinosus (Hinz, 1987) (Fig. 61B) from the Toyonian stage (correlated with the Cambrian Stage 4) and Mongolitubulus reticulatus Kouchinsky, Bengtson, Clausen, Gubanov, Malinky, and Peel, 2011 (Fig. 61C). Mongolitubulus spinosus is currently known from the Cambrian Series 2 of Siberia, the Great Britain, and Sweden (Kouchinsky et al. 2015a; Skovsted et al. 2021 and herein). Mongolitubulus reticulatus is described from the lower Cambrian Stage 4–Wuliuan Stage of the Siberian Platform (Kouchinsky et al. 2011 and herein).
The Echinodermata are represented by several thousand phosphatised and recrystallised ossicles from the Cambrian Stage 4–Drumian Stage (see also Kouchinsky et al. 2011, 2015a). Those include brachiolar plates of eocrinoids, such as biserial ones, ranging from the middle Atdabanian stage (Kouchinsky et al. 2015a) and more rare uniserial unbranched brachial ossicles, ranging from the lower Botoman stage (Kouchinsky et al 2015a and herein). Monomeric cylindrical ossicles representing stem holomeric columnals occur in the Botoman–Mayan stages (correlated with the Cambrian Stage 4–Drumian Stage) herein (Fig. 63). The earliest holomeric columnals occur in the lower Botoman stage (herein), probably slightly older than those known from Morocco and Laurentia. Together with co-occurring proximal stem plates they may represent the earliest rhombiferans (Fig. 63). Edrioasteroids occur in a form of rare ambulacral flooring and thecal plates (Fig. 63), ranging from the lower Botoman stage (correlated with the Cambrian Stage 4; see also Kouchinsky et al. 2011, 2015a; Clausen and Peel 2012; Zamora et al. 2013; Peel et al. 2016). Some rare but distinct ctenocystoid plates are found in the Botoman-Toyonian transitional beds of the Khorbusuonka River (correlated with the Cambrian Stage 4). Although preserved as disarticulated oral elements, these represent the earliest finds of the group, reducing the incongruence between their stratigraphic range and their position as basalmost, almost bilateral echinoderms. Numerous thecal plates of uncertain affinity are also available in the material (Fig. 64).
Calcium phosphatic tubes belonging to the order Hyolithelminthida Fisher, 1962 are represented by Hyolithellus isiticus Missarzhevsky in Rozanov et al. 1969 (Fig. 65B–E) from the Dokidocyathus regularis Zone, Tommotian stage (correlated with the Cambrian Stage 2) and several hundred fragmentary tubes of Hyolithellus sp. (Fig. 65A, F and H), ranging up to the Toyonian stage (correlated with the Cambrian Stage 4), as well as Torellella spp. (Figs. 65G, I–L, 66). Tubes of several species of problematic anabaritids are preserved as calcium phosphatic internal moulds (Fig. 67A–F). Among them, Anabarites korobovi (Tiksitheca korobovi Missarzhevsky in Rozanov and Missarzhevsky, 1966) is the most abundant and occurs in the basal Tommotian beds. Diverse and abundant anabaritids were previously described from sections of the Anabar Uplift (see Kouchinsky et al. 2015a, 2017), from the Fortunian–Cambrian Stage 3. Problematic tubes, Coleoloides trigeminatus Missarzhevsky in Rozanov et al., 1969 and Coleoloides cf. paucistriatus Poulsen, 1967, including their initial parts, are preserved as calcium phosphatic and glauconised internal moulds in the Tommotian stage (correlated with the Cambrian Stage 2) herein and elsewhere on the Siberian Platform (Fig. 68).
Fossil sponges are common in the Cambrian strata. They occur as intact compressions in Lagerstätten and as isolated spicules in residues of dissolved rocks. Spiculate “pharetronids”, heavily mineralised heteractinids and “lithistids” (mostly, anthaspidellids) and hypercalcified aspiculate archaeocyaths are immured in reefal facies. Quantitative analyses of taphocoenosis structure reveal an important role of poriferans in Cambrian reefal and level-bottom palaeocommunities where these animals comprise a significant part of individuals in terms of both the diversity and biovolume (Conway Morris 1986; Ivantsov et al. 2005a; Zhao et al. 2009; Zhuravlev et al. 2015). Interestingly, while sponges of Cambrian Lagerstätten, but “soft-bodied” horny sponges, are relatively simple in spicular morphology and mostly built of monaxons and stauractines (e.g., Walcott 1920; Rigby 1986; Chen et al. 2004; Rigby and Collins 2004; Ivantsov et al. 2005b; García-Bellido et al. 2007; Wu et al. 2014; Botting and Peel 2016), spicule assemblages acid-extracted from a number of coeval sections display a much higher diversity of morphologies (e.g., Mostler and Mosleh-Yazdi 1976; Bengtson 1986; Fedorov and Pereladov 1987; Zhang and Pratt 1994; Dong and Knoll 1996; Sugai et al. 2004; Castellani et al. 2012; Kouchinsky et al. 2015a; Peel 2019). Sponge spicules are common in samples throughout the sections. They are described under formal names and the senior synonyms are selected for morphologically similar elements. Most of the spicules can be identified as hexactinellids and calcareans. The Hexactinellida includes relatively rare siliceous spicules identified as Thoracospongia sp. (Fig. 70) from the Mayan stage (correlated with the Drumian Stage) and Abnormisella sp. (Fig. 71K, L) from the Tommotian stage. Calcareans are represented by rarely preserved triactines of Dodecaactinella sp. from Botoman and Toyonian stages (correlated with the Cambrian Stage 4) of the Khorbusuonka River (Fig. 72A–H). These spicules are replaced by dolomite. Other spicules have a more problematic affinity. For instance, a fibrous microstructure of some mutirayed spicules heteractinids is indicative of their primary aragonite mineralization in acicular radiating units (Fig. 75F, H). Such a microstructure and composition are not typical of any extant sponges but occur in radiocyaths (Kruse et al. 2015).
Finally, fragments of phosphatised archaeocyaths frequently occur in the basal Erkeket and Tyuser formations of the studied region, belonging to the Dokidocyathus regularis Zone, Tommotian stage (correlated with the upper part of Cambrian Stage 2), such as Cryptoporocyathus junicanensis Zhuravleva, 1960 (Fig. 79B), Cambrocyathellus sp. (Fig. 79F), and Dictyocyathus sp. (Fig. 79C). A fragment similar to C. junicanensis is also found in the uppermost Kessyusa Group of Khorbusuonka River, in beds attributed to the Watsonella Zone, Cambrian Stage 2 (Fig. 79E). Single fragments of archaeocyaths, such as Carinacyathus sp. (Fig. 79D) and Ajacicyathina family, gen. and sp. indet. (Fig. 79A, H, I), occur in the lower Botoman stage (correlated with the lower part of Cambrian Stage 4) of the Khorbusuonka River. Preservation in phosphate replicas complements previous descriptions of archaeocyaths in thin-sections.
Faunal assemblages
The most abundant and diverse fossil assemblages derive from the basal Erkeket Formation and the Middle Member of the Tyuser formations, Dokidocyathus regularis Zone, Tommotian stage (correlated with the upper part of Cambrian Stage 2) as well as from the middle Erkeket Formation, lower Botoman stage (correlated with the lower part of Cambrian Stage 4). Tommotian assemblages of phosphatised skeletal fossils were previously reported from these strata by Repina et al. (1974), Astashkin et al. (1991), and Vasiljeva (1998: 31), from lenses of limestone sandwiched within polymictic conglomerate and sandstone beds of the lowermost Middle Member (middle Sub-Formation) of the Tyuser Formation, from the Ulakhan-Ald’yarkhay (section 21 herein; SOM 2: figs. 3, 4). Those strata provided a continuous δ13C record allowing a correlation with the Tommotian stage of Siberia (correlated with the upper part of Cambrian Stage 2). According to the chemostratigraphy, the fossil assemblages restricted to the basal Erkeket Formation at Khorbusuonka (section 11) with the Siberian δ13C excursion II appear to be somewhat older than those from the basal Middle Member of the Tyuser Formation characterised by the δ13C excursion III (Figs. 3, 4).
The “small shelly fossils” from sections 11 and 21 are dominated by diverse and abundant orthothecid hyoliths, frequently occurring sclerites of chancelloriids, such as Stellispinella lenaica, and halkieriids, Halkieria proboscidea, as well as coiled shells of the conchiferan mollusc, Aldanella sibirica. The fauna also includes paterinid brachiopods, hyolithelminths, such as Hyolithellus spp. and tubular problematica, Coleoloides trigeminatus. By contrast with section 21, Anabarites korobovi frequently occurs in section 11, as well as sclerites of tommotiids, such as Camenella gabrowski and Lapworthella tortuosa, and the problematic Archaeopetasus typicus. Archaeopetasus pachybasalis sp. nov. typically occurs in section 21, but is unknown from Khorbusuonka. In both localities, fragments of archaeocyaths, spicules of hexactinellids and calcareans occur.
By contrast, secondarily phosphatised calcareous fossils are rare in coeval Tommotian strata in the eastern Anabar sections 96-3 and 96-6 (Kouchinsky et al. 2015a). Tommotian assemblages from latter sections yield almost exclusively originally organophosphatic forms, such as hyolithelminths, Hyolithellus cf. insolitus and Torellella cf. laevigata, tommotiids Camenella garbowskae, and siliceous sponge spicules (Kouchinsky et al. 2015a). On the contrary, phosphatisation of originally calcareous fossils and phosphatic moulds are common in lower beds of the Emyaksin Formation, where rich but older assemblages from Cambrian Stage 2 occur (Kouchinsky et al. 2017).
Assemblages of diverse and abundant phosphatised “small shelly fossils” are also present in strata embracing the Calodiscus–Erbiella Zone (Bergeroniellus micmacciformis–Erbiella Zone), lower Botoman stage, and recording the Siberian δ13C excursion VII (Figs. 3, 4). Fossils from sections 19 and 20 of the Erkeket Formation are similar to the fauna from the uppermost Emyaksin Formation of the eastern flanks of the Anabar Uplift (Kouchinsky et al. 2015a: fig. 2), sections 96-1 (samples K1/2B, K1/3B, 1/5, and 1/7.8, Malaya Kuonamka River) and 96-7 (sample 7/70, Bol’shaya Kuonamka River), although molluscs and hyoliths are more diverse in the latter localities (Kouchinsky et al. 2015a: SOM).
Skeletal fauna from sections 19 and 20 at the Khorbusuonka River (SOM 3) is represented by lingulate brachiopods (most abundant in sample 19/10.25), such as Eoobolus variabilis, Homotreta gorjanskii, and Botsfordia caelata, tommotiids Lapworthella lucida (common in sample 19/5.5), helcionellids, such as Davidonia anabarica (sample 19/12.75), the bivalve Fordilla sibirica and at least two species of Pelagiella, sclerites of problematic cambroclavids, Cambroclavus sp. and Zhijinites clavus (unknown from the Anabar Uplift, Kouchinsky et al. 2015a), and sclerites of chancelloriids (abundant in several samples). Hyoliths are mostly millimeter-sized orthothecids and less frequent hyolithids, including their phosphatised appendages. Other fossils are represented by abundant Hadimopanella knappologica sclerites (samples 19/5.5 and 20/1B), diverse echinoderm ossicles (including frequent blastozoan brachiolars, rare rhombiferans, and edrioasteroids), calcium phosphatic hyolithelminth tubes (Hyolithellus and Torellella), trilobite fragments, hexactinellids and heteractinid spicules. Calcarean spicules are represented by rare Dodecaactinella (not found in the Kuonamka localities). Fossil assemblages from the lower Botoman stage are attributed herein to the lower Cambrian Stage 4 and can be compared with the upper Cambrian Stage 3–lower Stage 4 assemblages from Gondwana and Laurentia: South Australia (Bengtson et al. 1990; Gravestock et al. 2001), Antarctica (Wrona 2003, 2004), North China (Liu et al. 2021), Mexico (Devaere et al. 2019), Newfoundland (Skovsted and Peel 2007), and Greenland (Skovsted 2006a).
The material under discussion also includes skeletal fossil assemblages from the Toyonian–lower Amgan stages (correlated with the Cambrian Stage 4), which are yet poorly characterised in Siberia. They can be compared with well-preserved phosphatised Ovatoryctocara granulata fauna from Greenland (Ovatoryctocara Zone; Peel et al. 2016; Peel 2021a). The Siberian assemblages of this age derive from the Khorbusuonka River (samples 19/25.5 through 19/55 and 20/2B through 20/6B, Fig. 3), the lower Lena River (samples 22/50 and 22/55.5, Fig. 4), and the Malaya Kuonamka River (samples 1/9, K1/5B, K1/6B, K1/7B, Fig. 2). The lowermost beds of this interval are marked by a negative trough in δ13C values (Figs. 3, 4), observed at the boundary between the Botoman and Toyonian stages. Subsequent onset of a rising trend in isotopic values in the Toyonian stage is associated with the peak IX (Figs. 3, 4; Brasier et al. 1994; Wotte et al. 2011; He et al. 2019).
Originally organo-phosphatic fossils herein are well-preserved, whereas primarily calcareous skeletons occurring within this interval from Siberia are rarely phosphatised and well-preserved. They are preserved in a recrystallised calcite form and cannot be obtained by maceration in a usual way. The extractable fossils are either silicified or glauconitised (mostly internal moulds) or rarely incrusted by pyrite (e.g., Hyolithida sp. indet. 2, pyritised conch, with operculum attached, from sample 1/9). Primarily calcium phosphatic skeletons extracted from samples are mainly represented by frequent to abundant lingulate brachiopods Homotreta gorjanskii and Eoobolus variabilis, with much less abundant Botsfordia caelata (section 96-1) and Acrothele olenekensis (section 19). Other organophosphatic skeletons are represented by frequent to abundant sclerites of Hadimopanella spp., including Hadimopanella foveata Kouchinsky sp. nov. (sections 19 and 20), rare Lapworthella dentata (samples 19/31.75, 20/5B), Mongolitubulus spinosus (sample 19/31.75), and hyolithelminth tubes. Originally aragonitic sclerites of chancelloriids are abundant in some samples and are mostly represented by glauconitised internal moulds. Several forms of conchiferan molluscs rarely occur in samples; those are Davidonia anabarica (samples 19/31.75, 19/33), Stenotheca cf. drepanoida, Yochelcionella sp., Pelagiella spp., Parailsanella sp., Pojetaia dentifera (sample 22/50), as well as orthothecid and hyolithid hyoliths.
Fossil assemblages from the Kounamkites Zone were previously described by Gubanov et al. (2004b) and Kouchinsky et al. (2011) from the Kuonamka Formation of the eastern flanks of the Anabar Uplift, the Bol’shaya Kuonamka River (samples 8/14.8 through 8/23.1) and the Malaya Kuonamka River (samples 1/16.5 through 1/28, and K1/17B, K1/8B; Fig. 2). Those are complemented herein by a few samples from the uppermost Erkeket Formation (samples 19/1B, 19/2B, 19/61) and from the Sekten Formation (sample 22/67) belonging to the Wuliuan Stage. Lingulate brachiopods, Homotreta gorjanskii and Eoobolus variabilis dominate the samples, whereas Botsfordia caelata and Acrothele olenekensis are rare to frequent in several samples (SOM 3). Other originally organo-phosphatic fossils are represented by rare disarticulated sclerites of Microdictyon robisoni Bengtson, Matthews, and Missarzhevsky, 1986, and Hadimopanella spp., protoconodonts, such as Gapparodus bisulcatus (Müller, 1959) and Amphigeisina danica (Poulsen, 1966). Protoconodonts occur more frequently in samples from the Kuonamka Formation of the Anabar Uplift (Kouchinsky et al. 2011), where Amphigeisina danica predominates. Protoconodonts are distributed throughout this interval and are represented by Gapparodus bisulcatus, Amphigeisina danica, Hagionella cultrata (Missarzhevsky, 1977), and undetermined forms (Kouchinsky et al. 2011). Therein, bradoriids represented by deformed carapaces and spines of Mongolitubulus reticulatus Kouchinsky, Bengtson, Clausen, Gubanov, Malinky, and Peel, 2011, occur. Echinoderm plates are rare and preserved as phosphatised and glauconitised disarticulated ossicles, occasionally with a pyritic cover. Hexactinellid spicules are present in the Kuonamka Formation of the Anabar Uplift (Kouchinsky et al. 2011), where they can be locally abundant (sample K1/17B). Secondarily phosphatised calcareous skeletal fragments are generally poorly preserved and represented by abundant disarticulated sclerites of chancelloriids; relatively rare conchiferan molluscs and hyoliths occur in the upper Sekten and uppermost Erkeket formations of the lower Lena and Khorbusuonka rivers, respectively. Extracted residues from samples of the Kuonamka Formation of the Anabar Uplift yield silicified and, thus, better preserved, more abundant and diverse fossils of the same groups (Gubanov et al. 2004b; Kouchinsky et al. 2011).
Faunal assemblages of the Tomagnostus fissus–Acadoparadoxides sacheri Zone, lower Mayan stage, which is correlated with the lowermost Drumian Stage, occur in the upper Kuonamka Formation of the Anabar Uplift (Kouchinsky et al. 2011: sample 8/32 therein; Fig. 2) and the Khorbusuonka River (Fig. 3, SOM 3). In the upper Kuonamka Formation of both regions, this interval shows a rising and swift δ13C trend (Figs. 2, 3). Fossil preservation via silicification is more common at this level than in older beds and includes a variety of originally calcareous skeletal fossils, otherwise poorly preserved or unextractable if silicification does not occur (Kouchinsky et al. 2011). Molluscs and hyoliths are rare in such samples, but common in 8/32. In addition, glauconitisation of moulds and pyrite incrustation can be developed. Originally organo-phosphatic skeletons are preserved here, such as lingulate brachiopods Olenekotreta olenekensis, Acrothele olenekensis, Homotreta gorjanskii, Eoobolus variabilis, Botsfordia caelata, and Linnarssonia bulcurica as well as palaeoscolecidan sclerites, Hadimopanella spp. and the protoconodont Gapparodus bisulcatus. Hexactinellid siliceous spicules of Thoracospongia cf. follispiculata Mehl, 1996 (Kouchinsky et al. 2011: fig. 39) and other pentactins and hexactins are present (Kouchinsky et al. 2011: fig. 40).
Systematic palaeontology
Phylum Brachiopoda Duméril, 1806
Subphylum Linguliformea Williams, Carlson, Brunton, Holmer, and Popov, 1996
Class Lingulata Gorjansky and Popov, 1985
Order Lingulida Waagen, 1885
Family Eoobolidae Holmer, Popov, and Wrona, 1996
Genus Eoobolus Matthew, 1902
Type species: Obolus triparilis Matthew, 1902; Miaolingian; MacLean Brook Group, Avalonian Cape Breton Island, Nova Scotia, Canada.
Eoobolus priscus (Poulsen, 1932)
Fig. 5.
Material.—Several tens of calcium phosphatic dorsal and ventral valves, including figured SMNH Br151173–151183, from samples 6/66.2 and 7/55, upper Emyaksin Formation, Bol’shaya Kuonamka, Siberia, Russia. Judomia–Uktaspis (Prouktaspis) Zone, Atdabanian stage (correlated with the Cambrian Stage 3).
Description.—Shell biconvex, outer surface with fine concentric growth lines with occasional sets of drapes and radial striation. Brephic shell, 250–300 μm wide and long, covered with numerous densely spaced pits, 1–3 μm in diameter. Ventral valve longitudinally elongated oval in outline, narrowing towards posteromarginal umbo, with apical angle 85–100°. Ventral pseudointerarea slightly raised and divided by a deep pedicle groove, which widens anteriorly. Dorsal valve oval to subcircular with rounded posterior margin. Pseudointerarea consists of anteriorly curved median plate passing into propareas. Median ridge well defined, low and widens anteriorly.
Remarks.—Eoobolus priscus is very similar to Eoobolus variabilis (Pelman, 1977), described below, but differs in having narrower outer parts of the ventral propareas, a distinct radial striation on the outer surface of the ventral valve; moreover, the dorsal valve has an almost isometrical outline as well as a more prominent and undivided dorsal median ridge.
Stratigraphic and geographic range.—Cambrian Stage 3–Wuliuan Stage; Laurentia, Avalonia, Siberia, South Australia, and Antarctica.
Eoobolus variabilis (Pelman, 1977)
Figs. 6, 7.
Material.—Several hundred calcium phosphatic dorsal and ventral valves and complete shells, including figured SMNH Br151184–151196, from samples K1/6B, K1/7B, 1/22, 1/28, Malaya Kuonamka and 7/60, 7/70, 8/22.5, 8/23.1, Bol’shaya Kuonamka, upper Emyaksin and Kuonamka formations, and from samples 19/4B, 19/5B, 19/10.25, 19/66.5, Erkeket and Kuonamka formations, Khorbusuonka River. Siberia, Russia; Botoman stage (correlated with the Cambrian Stage 4)–Tomagnostus fissus–Acadoparadoxides sacheri Zone, Drumian Stage.
Description.—Shell biconvex, outer surface covered with concentric growth lines with occasional sets of drapes, concentric rows of tubercles up to 5 μm in diameter and fine radial striation. Radial striation on dorsal valve developed better than on the ventral valve, where it is almost indistinguishable. Brephic shell, 200–250 μm wide and long, covered with densely spaced numerous pits 1–3 μm in diameter. Ventral valve longitudinally elongated oval in outline, narrows towards the posteromarginal umbo, with apical angle 90–110°. Ventral pseudointerarea slightly raised and divided by a deep pedicle groove, which widens anteriorly. Dorsal valve oval with rounded posterior margin. Pseudointerarea consists of anteriorly curved median plate passing into propareas. Median ridge consists of three subparallel low and narrow branches slightly diverging anteriorly.
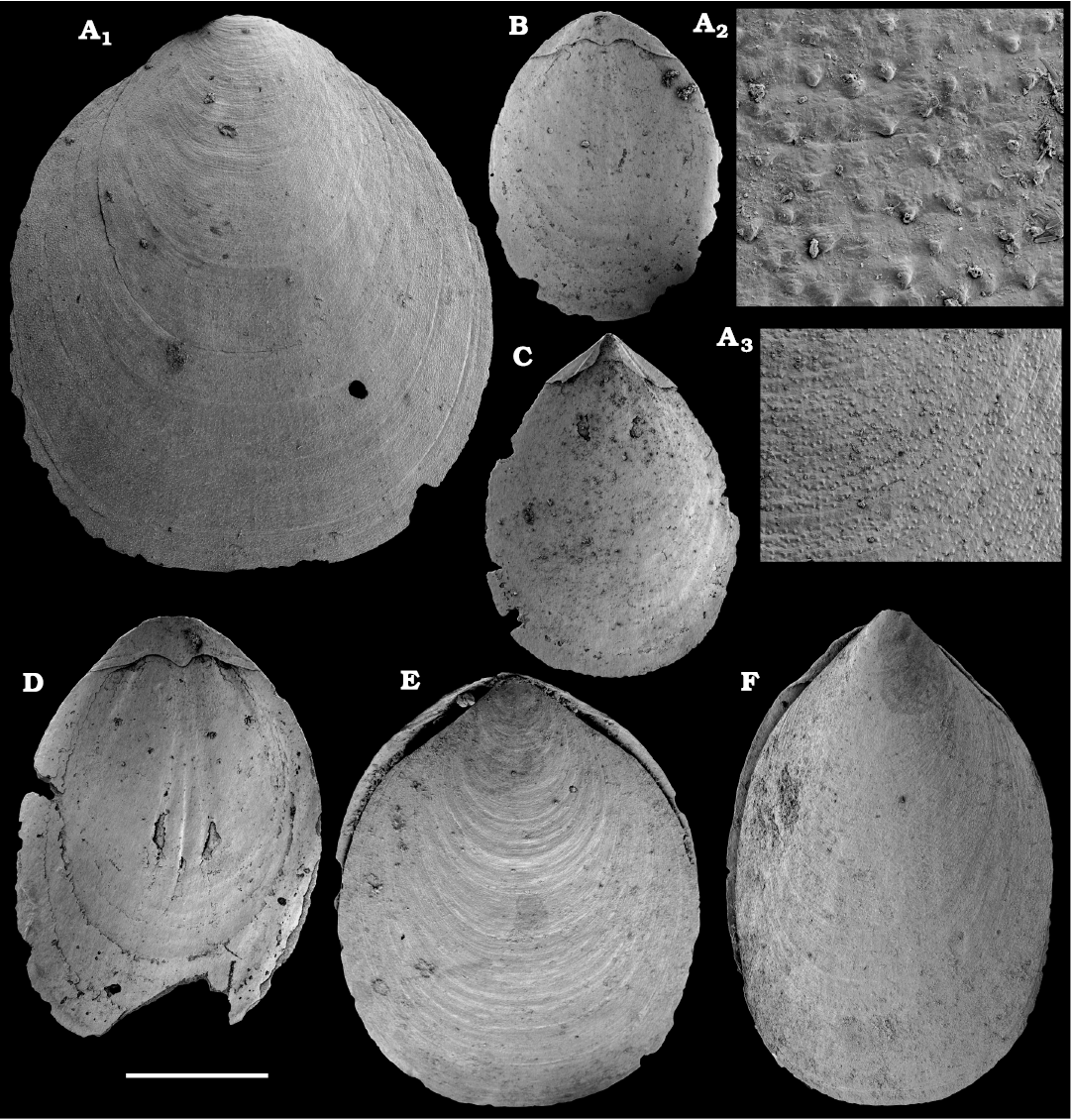
Fig. 6. Lingulid brachiopod Eoobolus variabilis (Pelman, 1977) from the middle Cambrian Kuonamka Formation, Bol’shaya Kuonamka River, Siberia, Russia, sample 8/23.1. A, B, D. Dorsal valves, SMNH Br151184–151186, respectively. A2, A3, close-ups of tuberculate surface. C. Ventral valve, SMNH Br151187. E, F. Specimens with both valves preserved, view on the ventral valve, SMNH Br151188 and 151189, respectively. Scale bar: 1 mm (A1, B–F), 250 µm (A3), 50 µm (A2).
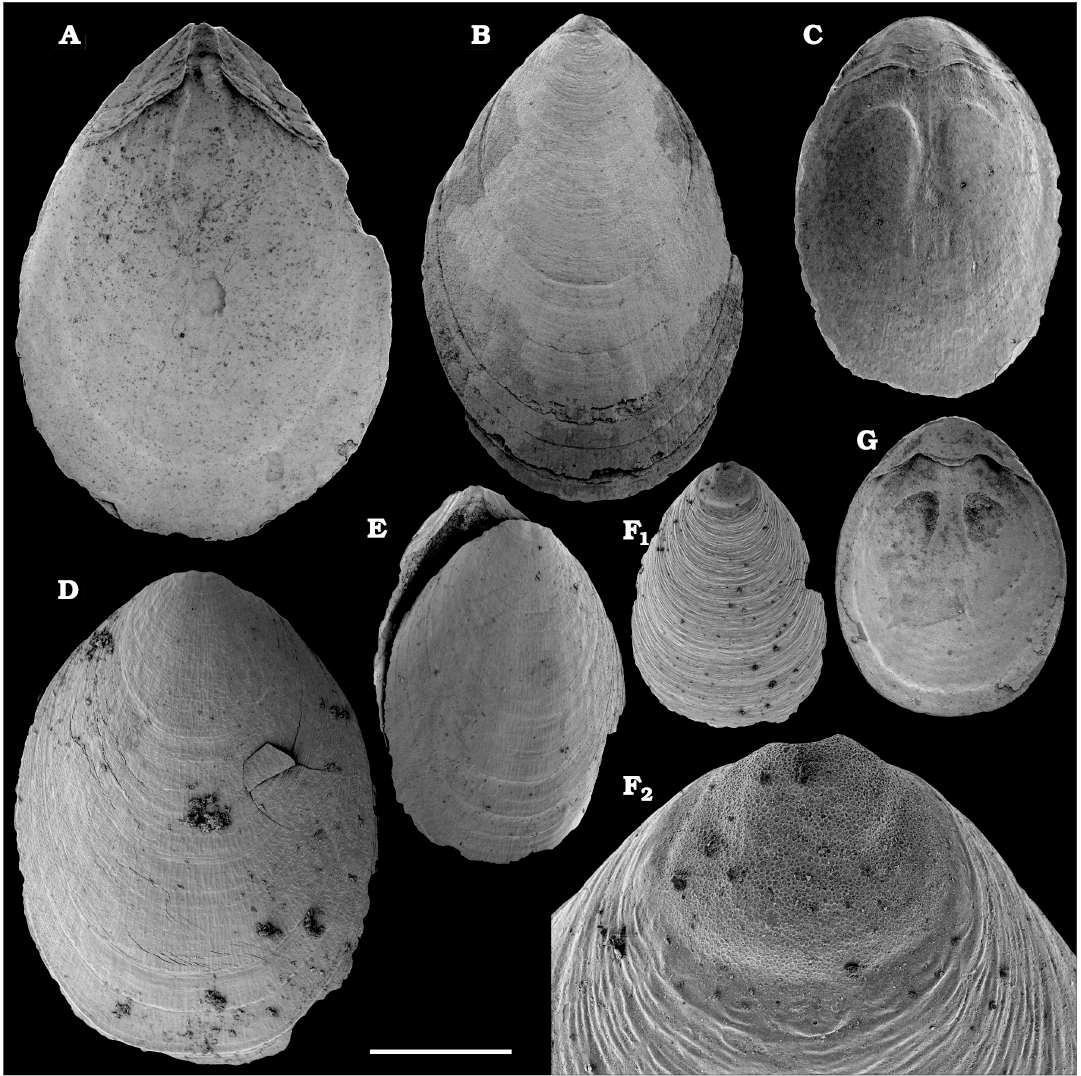
Fig. 7. Lingulid brachiopod Eoobolus variabilis (Pelman, 1977) from the Cambrian Emyaksin Formation, Bol’shaya Kuonamka River (A–C), Erkeket Formation, Khorbusuonka River (D–F), and Kuonamka Formation, Malaya Kuonamka River (G) , Siberia, Russia; sample 7/70 (A–C), samples 19/4B, 19/66.5, 19/10.25 (D–F, respectively), and sample K1/7B (G). A, B. Ventral valves, SMNH Br151190 and 151191, respectively. C, D, F, G. Dorsal valves, SMNH Br151193–151196, respectively. F2, close-up showing brephic shell. E. Specimen with both valves preserved, view on the dorsal valve, SMNH Br151192. Scale bar: 1 mm (A–E, G), 500 µm (F1), 100 µm (F2).
Remarks.—Eoobolus variabilis is closely similar to E. pelmani, E. priscus, and E. siniellus, also described from the Siberian Platform; it differs in having more massive and wider outer parts of ventral propareas. The ventral apical angle of E. variabilis is usually wider and varies between 95–110° (the angle does not exceed 100° in E. priscus, after Ushatinskaya and Korovnikov 2014).
Stratigraphic and geographic range.—Cambrian Stage 3–Drumian Stage; Laurentia, Avalonia, Siberia, South Australia, and Antarctica.
Order Acrotretida Kuhn, 1949
Family Acrotretidae Schuchert, 1893
Genus Homotreta Bell, 1941
Type species: Homotreta interrupta Bell, 1941; Miaolingian; Pagoda Limestone, Montana, USA.
Homotreta gorjanskii (Pelman, 1973)
Figs. 8–10.
Material.—Several thousand calcium phosphatic dorsal and ventral valves, including figured SMNH Br151197–151211, from samples K1/1B, K1/4B, K1/5B, K1/6B, K1/7B, 1/5, 1/9, 1/22, 1/28, 7/70, 8/14.8, 8/17.6, 8/18.5, 8/22.5, 8/23.1, upper Emyaksin and Kuonamka formations, Bol’shaya Kuonamka and Malaya Kuonamka rivers, samples 19/10.25, 19/14.25, 19/16, 19/25.5, 19/26.75, 19/29, 19/31.75, 19/33, 19/40, 19/43.25, 19/46.5, 19/52, 19/55, 19/61, 19/66.75, 19/1B, 19/3B, 19/4B, 19/5B, 19/6B, 20/1B, 20/2B, 20/3B, 20/4B, Erkeket and Kuonamka formation, Khorbusuonka River and samples 22/55.5 and 22/67, Sekten Formation, Lena River. Siberia, Russia; Lower Botoman–lower Mayan stages (correlated with the Cambrian Stage 4–lower Drumian Stage).
Description.—Shell rounded in outline, covered with fine concentric growth lines with occasional sets of drapes. Brephic shell broadly oval in outline, 150–200 μm long and 200–250 μm wide, covered with scattered pits, 0.5–1.0 μm in diameter, becoming less distinct towards the margin of the brephic shell. Circular foramen, 25–30 μm in diameter, opens at the tip of a conical extension of brephic shell at its posterior margin. Ventral valve low conical with anteromarginal umbo and steeply procline pseudointerarea. External pedicle opening enclosed within brephic shell at its margin. Prominent apical process situated directly anterior to the foramen on the ventral interior. Dorsal valve gently convex, rounded with a marginal umbo. Dorsal pseudointerarea orthocline, with a broad triangular median groove and short narrow propareas. Dorsal interior with well-defined blade-like median ridge.
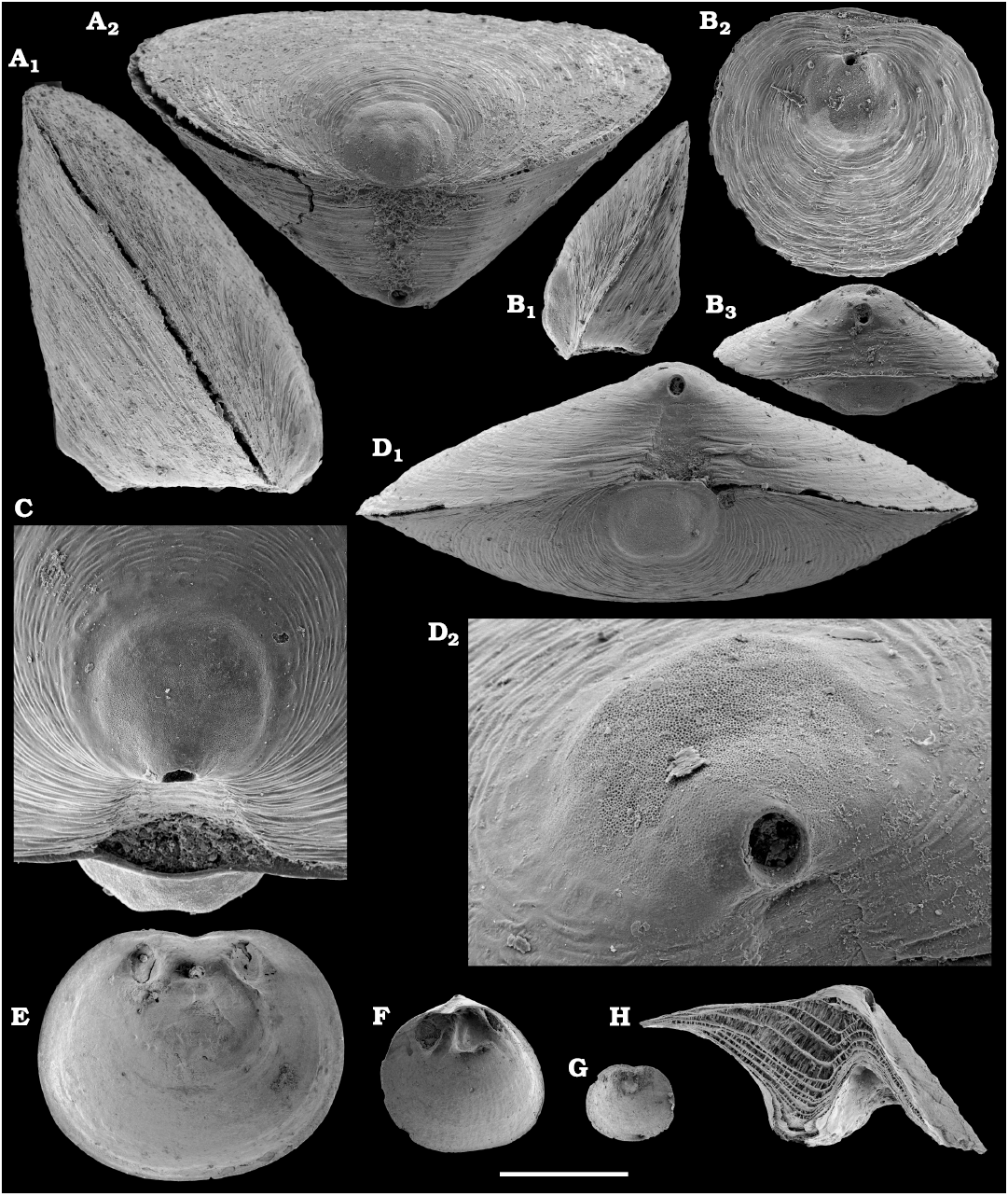
Fig. 8. Lingulid brachiopod Homotreta gorjanskii (Pelman, 1973) from the middle Cambrian Kuonamka Formation, Malaya Kuonamka and Bol’shaya Kuonamka rivers, Siberia, Russia; samples 1/28 (A, B) and 1/22 (H) and samples 8/14.8 (C, D), 8/18.5 (F), and 8/23.1 (E, G). A–D. Specimens with both valves preserved, SMNH Br151197–151200, respectively. A1, B1, lateral; A2, B3, D1, posterior; and B2, ventral views; C, D2, view on the ventral valve showing brephic shell. E–G. Ventral valves, SMNH Br151201–151203, respectively. H. Ventral valves, SMNH Br151204, sagittal cross-section showing lamination. Scale bar: 1 mm (E–G), 500 µm (H), 250 µm (A, B, D1), 125 µm (C), 60 µm (D2).
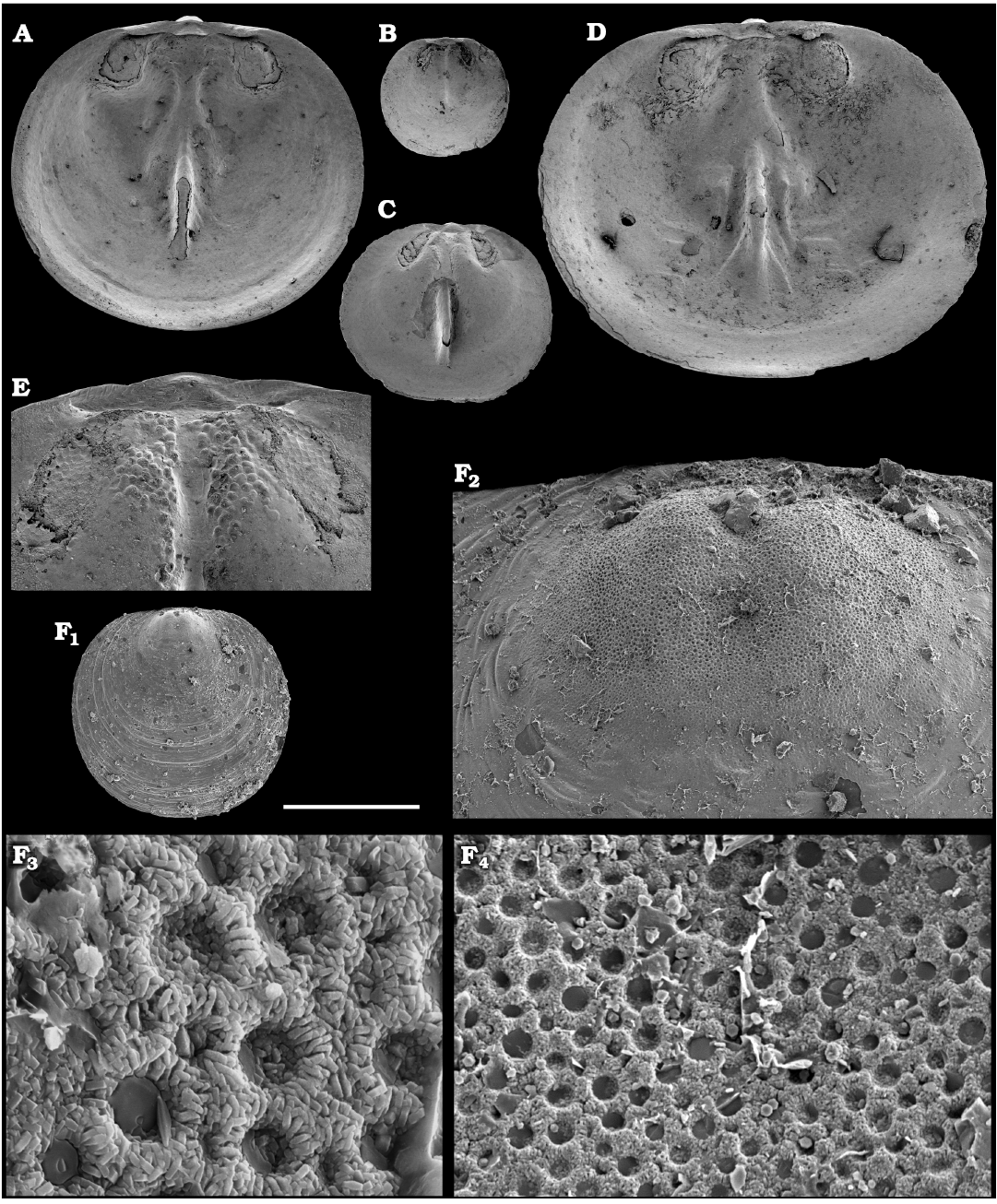
Fig. 9. Lingulid brachiopod Homotreta gorjanskii (Pelman, 1973) from the middle Cambrian Kuonamka Formation, Malaya Kuonamka and Bol’shaya Kuonamka rivers, Siberia, Russia; samples 8/22.5 (A), 8/23.1 (B, D), 8/18.5 (C, E), and 1/28 (F). A–F. Dorsal valves, SMNH Br151205–151210, respectively. F2, close-up showing brephic shell; F3, F4, enlargements with pits on the surface. Scale bar: 1 mm (A–D), 500 µm (F1), 250 µm (E), 60 µm (F2), 8 µm (F4), and 2.5 µm (F3).
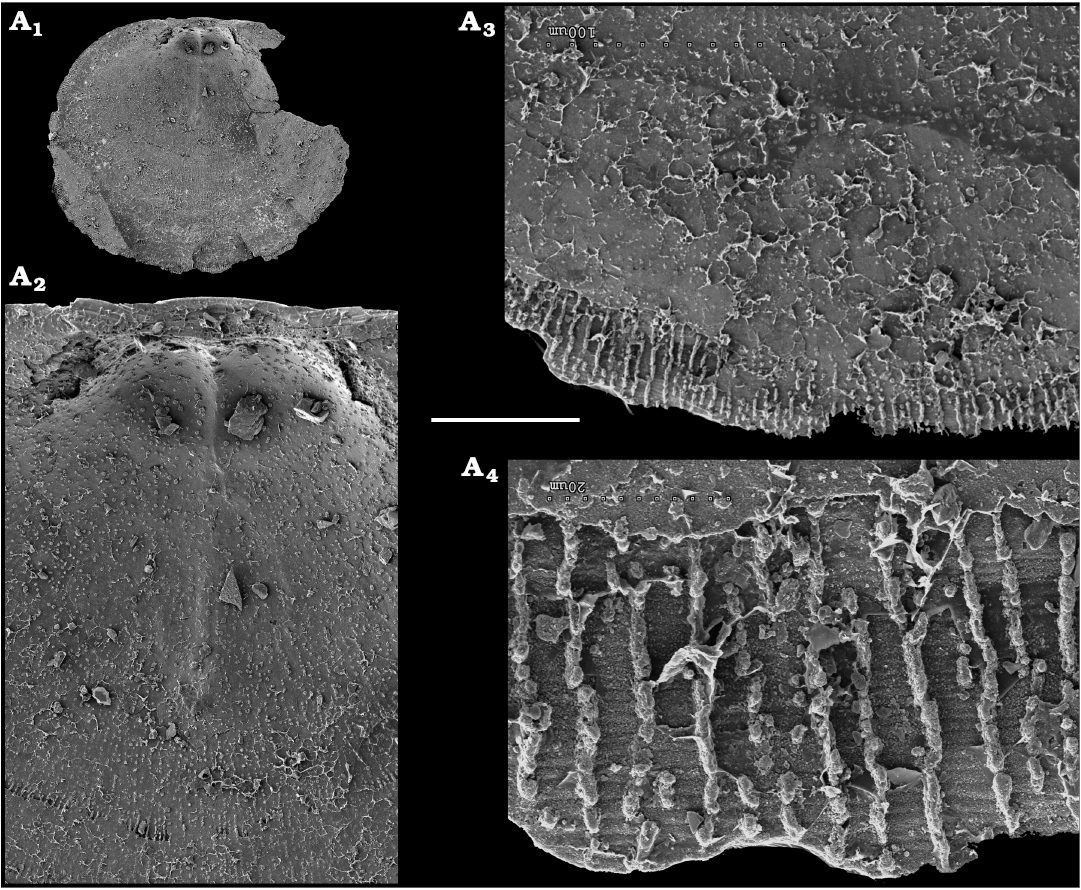
Fig. 10. Lingulid brachiopod Homotreta gorjanskii (Pelman, 1973), SMNH Br151211, from the middle Cambrian Kuonamka Formation, Malaya Kuonamka River, Siberia, Russia; sample 1/28. A1, dorsal valve; A2, close-up of A1; A3, A4, enlargements of anterior margin with mantle canals. Scale bar: 500 µm (A1), 125 µm (A2), 60 µm (A3), 17 µm (A4).
Remarks.—See comparative discussion in Ushatinskaya and Korovnikov (2019).
Stratigraphic and geographic range.—Botoman–lower Mayan stages (correlated with the Cambrian Stage 4–lower Drumian Stage), Siberian Platform.
Homotreta cf. gorjanskii (Pelman, 1973)
Fig. 11.
Material.—Several tens of calcium phosphatic dorsal and ventral valves, including figured SMNH Br151212–151222, from samples 2/0, 2/10, and 8/32, Kuonamka and Olenyok formations, Bol’shaya Kuonamka and Malaya Kuonamka rivers, Siberia, Russia. Tomagnostus fissus–Acadoparadoxides sacheri Zone, lower Mayan stage (Drumian Stage).
Description.—Shell rounded in outline, covered with fine concentric growth lines with occasional sets of drapes. Ventral valve conical with marginal umbo. Brephic shell, ca. 200 μm in diameter, broadly oval in outline, 150–180 μm long and 180–210 μm wide, covered with scattered pits, 0.5–2.0 μm in dimeter, becoming less distinct towards the margin of the brephic shell. Circular foramen, 25–30 μm in diameter, opens at the tip of a conical extension of brephic shell at a distance from its posterior margin. Ventral pseudointerarea steeply procline. Apical foramen in a short tube, directed posteriorly and upward. External pedicle opening enclosed within brephic shell. Dorsal valve gently convex, rounded with a marginal umbo. Dorsal pseudointerarea with a broad triangular median groove and broad propareas. Dorsal interior with faint broad median ridge. Brephic shell broadly oval in outline, covered with pits, ca. 1 μm in diameter.

Fig. 11. Lingulid brachiopod Homotreta cf. gorjanskii (Pelman, 1973) from the middle Cambrian Olenyok Formation, Malaya Kuonamka River, Siberia, Russia; sample 2/10. A–E. Ventral valves, SMNH Br151212–151216, respectively. A2, close-up of brephic shell, A3, enlargement showing pitted surface; E1, lateral view; E2, posterior view. F–K. Dorsal valves, SMNH Br151217–151222, respectively. G, brephic shell. Scale bar: 250 µm (A1, B–F, H–K), 125 µm (A3, G), and 12.5 µm (A2).
Remarks.—In Homotreta gorjanskii, the foramen closes at the boundary between the brephic and adult shells and is directed posteriorly, whereas in Homotreta cf. gorjanskii, the foramen appears to be enclosed within the brephic shell and directed posteriorly and upwards. The ventral brephic shell in H. cf. gorjanskii has asymmetrically spaced tuberсles and a more prominent cone-like extension from the foramen. The maximum diameter of the pits on the brephic shell of H. cf. gorjanskii is twice as large as the maximum diameter of pits in H. gorjanskii.
Homotreta cf. gorjanskii probably represents a new species, but the systematics of the Siberian forms of the family Acrotretidae is currently under revision and it is left under open nomenclature here.
Homotreta aff. gorjanskii (Pelman, 1973)
Fig. 12.
Material.—Several hundred calcium phosphatic dorsal and ventral valves, including figured SMNH Br151223–151232, from sample 19/92, Yunkyulyabit-Yuryakh Formation, Khorbusuonka River, Siberia, Russia. Anopolenus henrici–Corynexochus perforatus Zone, Mayan stage (correlated with the Drumian Stage).
Description.—Shell rounded in outline, covered with fine concentric growth lines with occasional sets of drapes. Ventral valve high conical with umbo situated anteriorly to the posterior margin. Ventral pseudointerarea steeply procline. Apical foramen in a short tube, external pedicle opening enclosed within brephic shell at its margin. Dorsal valve gently convex, rounded with a marginal umbo. Dorsal pseudointerarea orthocline, with a broad triangular median groove and short propareas. Dorsal interior a broad indistinct median ridge. Brephic shell broadly oval in outline, 150–170 μm long and 150–170 μm wide, covered with pits, ca. 1 μm in diameter.

Fig. 12. Lingulid brachiopod Homotreta aff. gorjanskii (Pelman, 1973) from the middle Cambrian Erkeket Formation, Khorbusuonka River, Siberia, Russia; sample 19/92. A. Specimen with both valves preserved, SMNH Br151223. A1, posterior view; A2, lateral view. B, D, E, I. Ventral valves, SMNH Br151224–151227, respectively. I1, oblique posterior view, I2, inner surface. C, F–H, J. Dorsal valves, SMNH Br151228–151232, respectively. H1, inner surface; H2, oblique inner view; J2, close-up showing brephic shell. Scale bar: 500 µm (A–E, G, H, I1, J1), 250 µm (F, I2), and 60 µm (J2).
Remarks.—This form is different from Homotreta gorjanskii in having anteriorly placed umbo and a higher ventral valve. Similar specimens are attributed to H. gorjanskii by Pelman (1977: pl. 8: 2а, 2в, 2г).
Genus Linnarssonia Walcott, 1885
Type species: Obolella transversa Hartt in Dawson, 1868; middle Cambrian; Avalonian New Brunswick, Canada.
Linnarssonia bulcurica Pelman in Pelman and Pereladov, 1986
Fig. 13.
Material.—Over one hundred calcium phosphatic dorsal and ventral valves, including figured SMNH Br151233–151239, from samples 1/16.5, K1/8B, 8/32, Kuonamka Formation, Bol’shaya Kuonamka and Malaya Kuonamka rivers, Siberia, Russia. Kounamkites and Tomagnostus fissus–Acadoparadoxides sacheri zones, Amgan and lower Mayan stages (correlated with the Wuliuan–lower Drumian stages).
Description.—Shell rounded in outline, covered with fine concentric growth lines with sets of drapes. Ventral valve conical. Umbo situated anteriorly to the posterior margin of ventral valve. Ventral pseudointerarea steeply procline. Apical foramen in a short tube. External pedicle opening not enclosed within brephic shell. Dorsal valve gently convex, rounded with a marginal umbo. Dorsal pseudointerarea orthocline, with a broad median groove and short propareas. Dorsal interior with low median ridge. Brephic shell broadly oval in outline, 200–220 μm long and 220–270 μm wide, covered with pits, ca. 1 μm in diameter, becoming less distinct towards the margin of the brephic shell.
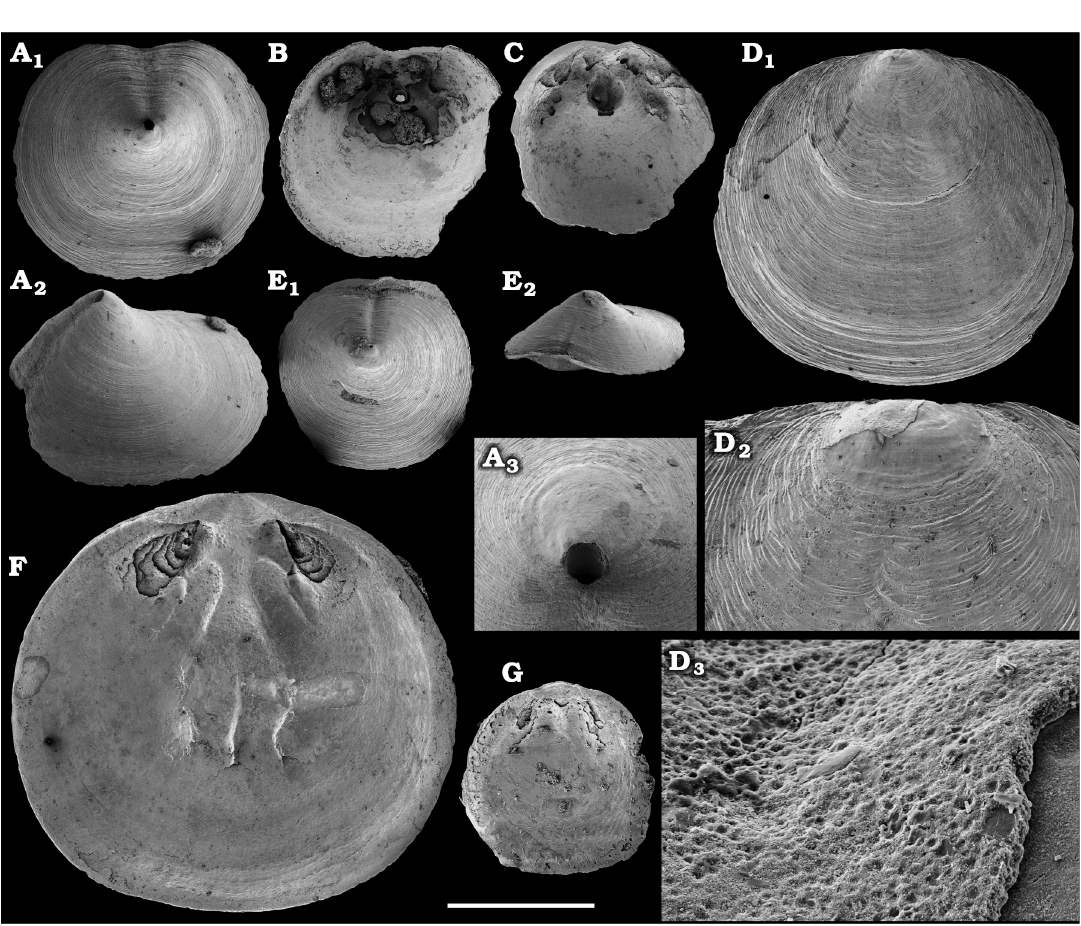
Fig. 13. Lingulid brachiopod Linnarssonia bulcurica Pelman in Pelman and Pereladov, 1986 from the middle Cambrian Kuonamka Formation, Malaya Kuonamka and Bol’shaya Kuonamka rivers, Siberia, Russia; sample 1/16.5 (A–D, F, G) and sample 8/32 (E). A–C. Ventral valves, SMNH Br151233–151235, respectively. A1, external surface; A2, lateral view; A3, close-up showing brephic shell. D, F, G. Dorsal valves, SMNH Br151237–151239, respectively. D2, close-up of the brephic shell; D3, enlargement with pits on the surface. E. Specimen with both valves preserved, SMNH Br151236. E1, external surface; E2, oblique lateral view. Scale bar: 1 mm (A1, A2, B, C, D1, E, F), 500 µm (G), 250 µm (A3, D2), and 12.5 µm (D3).
Remarks.—See comparative discussion in Ushatinskaya and Korovnikov (2019).
Stratigraphic and geographic range.—Amgan and lower Mayan stages (correlated with the Wuliuan and lower Drumian stages), Siberian Platform.
Family Botsfordiidae Schindewolf, 1955
Genus Botsfordia Matthew, 1891
Type species: Obolus pulcher Matthew, 1889; middle Cambrian; eastern North America, Avalonian New Brunswick.
Botsfordia caelata (Hall, 1847)
Fig. 14.
Material.—Over one hundred calcium phosphatic dorsal and ventral valves, including figured SMNH Br151240–151248, from samples K1/5B, K1/7B, 1/7.8, 1/9, 7/70, 8/23.1, 8/32, upper Emyaksin and Kuonamka formations, Bol’shaya Kuonamka and Malaya Kuonamka rivers, samples 19/3.25, 19/10.25, 19/1B, 19/2B, Erkeket Formation, Khorbusuonka River, Siberia, Russia. Botoman to lower Mayan stages (correlated with the Cambrian Stage 4–lower Drumian Stage).
Description.—Shell biconvex flattened, rounded in outline, up to 4 mm long and wide, with slightly undulating growth lines. Numerous tubercles ca. 1 μm in diameter, arranged along the concentric growth lines developed on the surface of adult shell. In larger specimens radial costellae are present. Brephic shell oval, 350–400 μm long and 450–500 μm wide, distinctly separated from the adult shell by a halo. Brephic shell surface with numerous tiny pits, ca. 1 μm in diameter. Ventral brephic valve bears a tubercle near umbo and two low tubercles situated anteriorly. Dorsal brephic valve with a pair of distinct tubercles at the posterior margin and respective elevations situated anteriorly. Ventral valve has developed propareas, short pseudointerarea divided by delthyrium, which forms a pedicle opening. Dorsal valve with a narrow pseudointerarea and divided by a low median septum.
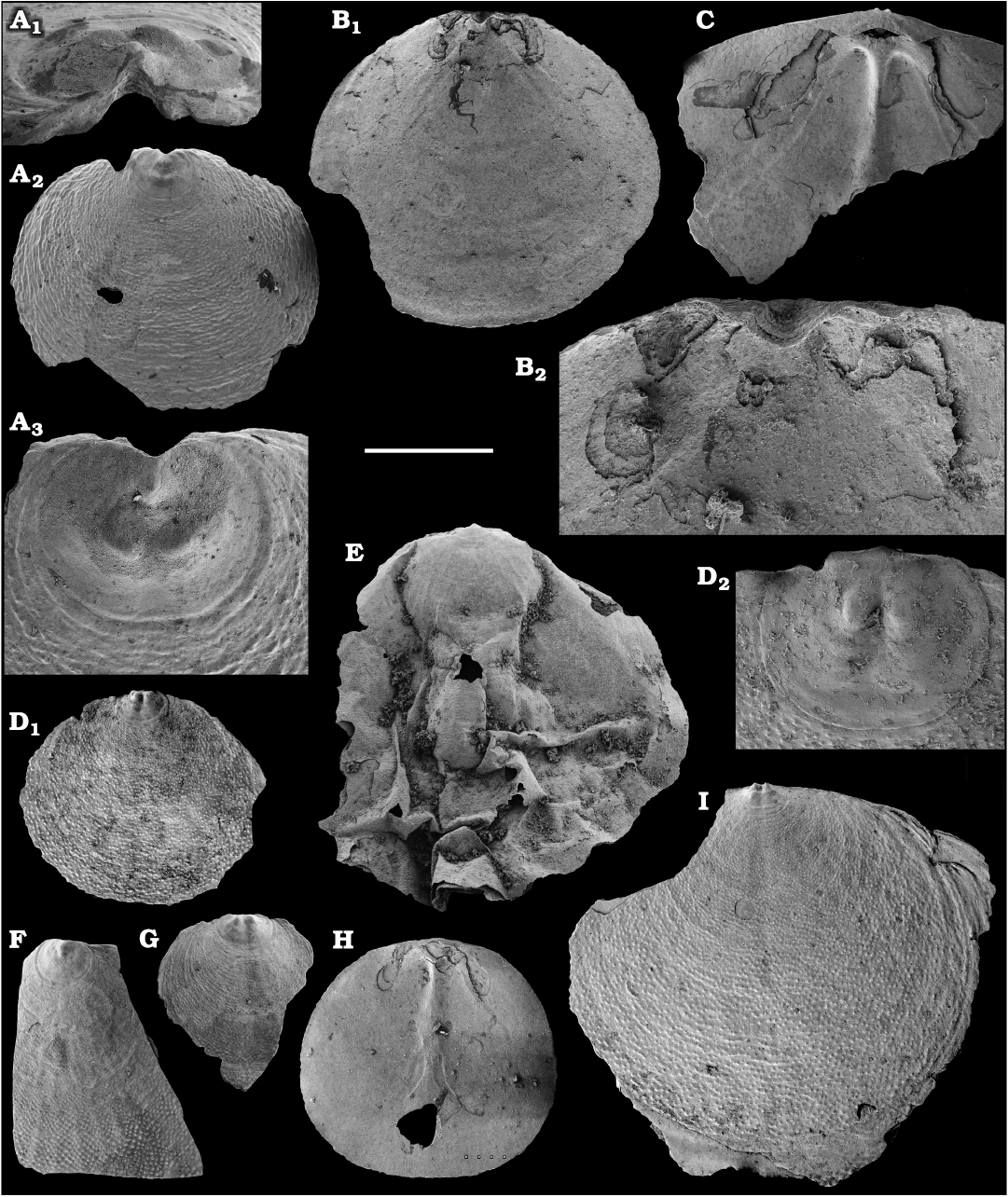
Fig. 14. Lingulid brachiopod Botsfordia caelata (Hall, 1847) from the Cambrian Emyaksin (F, G) and Kuonamka (A–E, H, I) formations, Malaya Kuonamka and Bol’shaya Kuonamka rivers, Siberia, Russia; samples K1/7B (A, B, H) and 1/9 (C–E, I) and 7/70 (F, G). A, B, F. Ventral valves, SMNH Br151240–151242, respectively. A1, A3, B2, close-ups showing brephic shell. C, D, G–I. Ventral valves, SMNH Br151243–151247, respectively. D2, enlargement showing brephic shell. E. Deformed specimen with both valves preserved, SMNH Br151248, view on the ventral valve. Scale bar: 1 mm (A2, B1, C, D1, E–I) and 250 µm (A1, A3, B2, D2).
Remarks.—See comparative discussion in Ushatinskaya and Korovnikov (2016).
Stratigraphic and geographic range.—Cambrian Stage 4–lower Drumian Stage, Siberian Platform and Laurentia.
Family Acrothelidae Walcott and Schuchert in Walcott, 1908
Genus Acrothele Linnarsson, 1876
Type species: Acrothele coriacea Linnarsson 1876; middle Cambrian; Sweden.
Acrothele olenekensis Pelman in Pelman and Pereladov, 1986
Fig. 15.
Material.—Over one hundred calcium phosphatic dorsal and ventral valves, including figured SMNH Br151249–151253, from samples 19/43.25, 19/52, 19/55, 19/65.5, 19/66.5, 19/66.75, 19/2B, 19/5B, 19/6B, Erkeket and Kuonamka formations, Khorbusuonka River, samples 1/22 and 8/18.5, Kuonamka Formation, Malaya Kuonamka and Bol’shaya Kuonamka rivers, Siberia, Russia. Amgan and lower Mayan stages (correlated with the Cambrian Stage 4–lower Drumian Stage).
Description.—Shell slightly biconvex flattened, rounded in outline, with coarse concentric growth lines. Numerous tubercles ca. 1 μm in diameter are scattered on the surface of adult shell. Ventral valve with anterior position of umbo relative to the posterior margin. Brephic shell broadly oval, 400–500 μm long and 550–600 μm wide, distinctly separated from the adult shell by a halo. Brephic shell surface with numerous tiny pits, a few μm in diameter. Marginal umbo of dorsal valve. Ventral brephic valve with a sub-rectangular elevated, 50–80 μm wide foramen contacting the adult shell. Two distinct tubercles situated anteriorly. Dorsal brephic valve with one pair of prominent tubercles at the posterior margin and another pair of less prominent tubercle situated anteriorly. Pseudointerarea pass gradually into the lateral margins of ventral valve. Dorsal valve with a narrow pseudointerarea, short and narrow propareas. Median septum low.
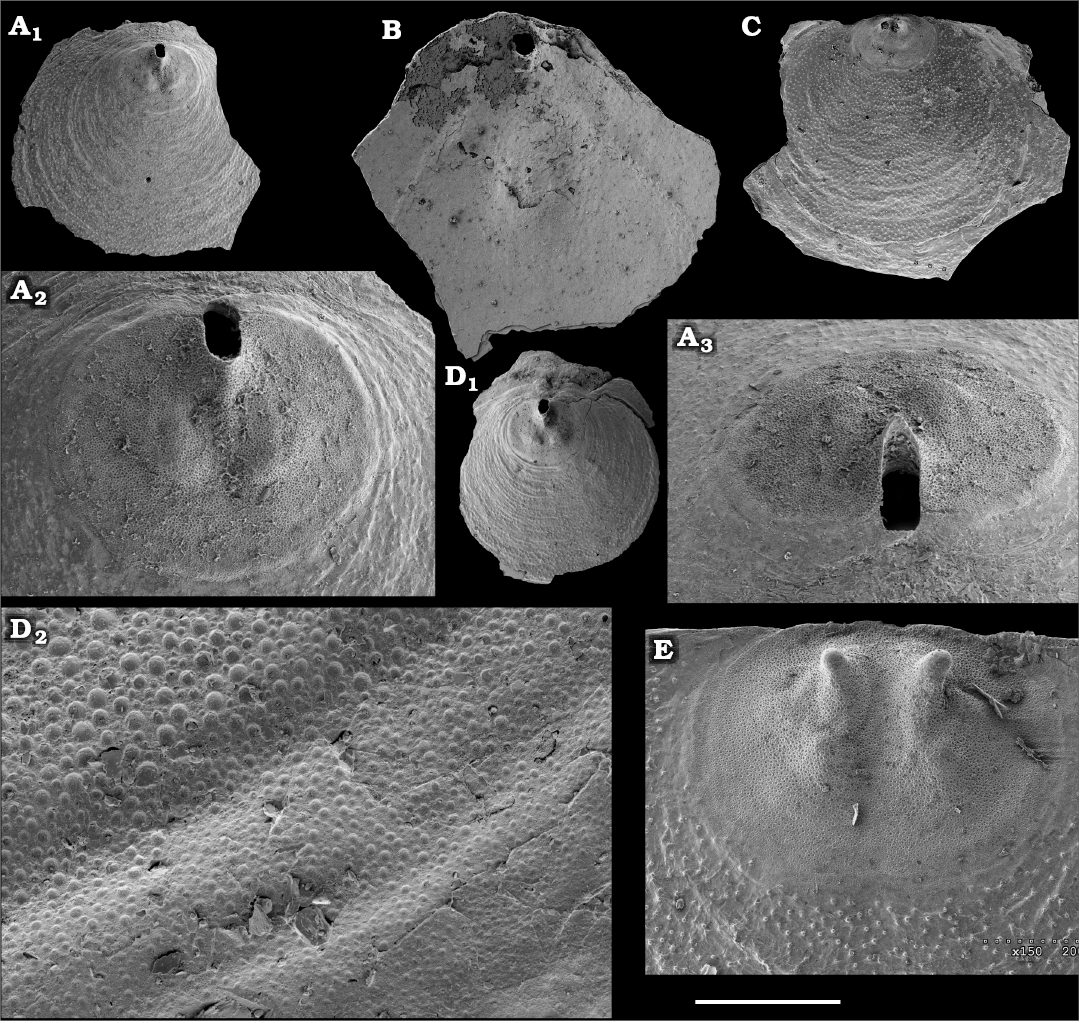
Fig. 15. Lingulid brachiopod Acrothele olenekensis Pelman in Pelman and Pereladov, 1986 from the middle Cambrian Kuonamka Formation, Malaya Kuonamka and Bol’shaya Kuonamka rivers, Siberia, Russia; samples 1/22 (A, B, D), K1/17B (C), and 8/18.5 (E). A, B, D. Ventral valves, SMNH Br151249–151251, respectively. A2, A3, close-ups showing brephic shell; D2, enlargement showing pits on the surface of brephic shell. C, E. Dorsal valves, SMNH Br151252 and 151253, respectively. E, dorsal valve of the brephic shell. Scale bar: 1 mm (A1, C, D1), 500 µm (B), 250 µm (A2, A3, E), and 25 µm (D2).
Remarks.—See the comparative study in Ushatinskaya and Korovnikov (2016).
Stratigraphic and geographic range.—Amgan and lower Mayan stages (correlated with the Wuliuan–lower Drumian stages), Siberian Platform.
Class Paterinata Williams, Carlson, Brunton, Holmer, and Popov, 1996
Order Paterinida Rowell, 1965
Family Paterinidae Schuchert, 1893
Genus Pelmanotreta Skovsted, Ushatinskaya, Holmer, Popov, and Kouchinsky, 2014
Type species: Cryptotreta neguertchenensis Pelman, 1977; upper Tommotian stage (Cambrian Stage 2); Siberian Platform, Russia.
Pelmanotreta neguertchenensis (Pelman, 1977)
Fig. 16.
Material.—Fifty fragmentary and complete calcium phosphatic dorsal valves and fragments of ventral valves, including figured SMNH Br151254–151258, from samples 3/23, 3/25B, 3/34, 7/16, 7/20, 7/22.6B, 7/25.5, 7/25.6, 7/27.5, Emyaksin Formation, Malaya Kuonamka and Bol’shaya Kuonamka rivers and sample 11/28.8, Erkeket Formation, Khorbusuonka River. Siberia, Russia; Dokidocyathus lenaicus–Tumuliolynthus primigenius Zone, Tommotian stage (correlated with the upper part of the Cambrian Stage 2)–Repinaella Zone, Atdabanian stage (correlated with the Cambrian Stage 3).
Description.—Dorsal valves, up to 5 mm wide, semicircular in outline, slightly convex, with broad and shallow sulcus, fine concentric growth lines, interrupted by radially oriented series of nick points. A narrow interarea situated along the entire posterior margin. Interior surface with radiating imprints of pinnate mantle canals. Dorsal brephic shell semicircular, ca. 200 μm long and 300 μm wide, with radial folds.
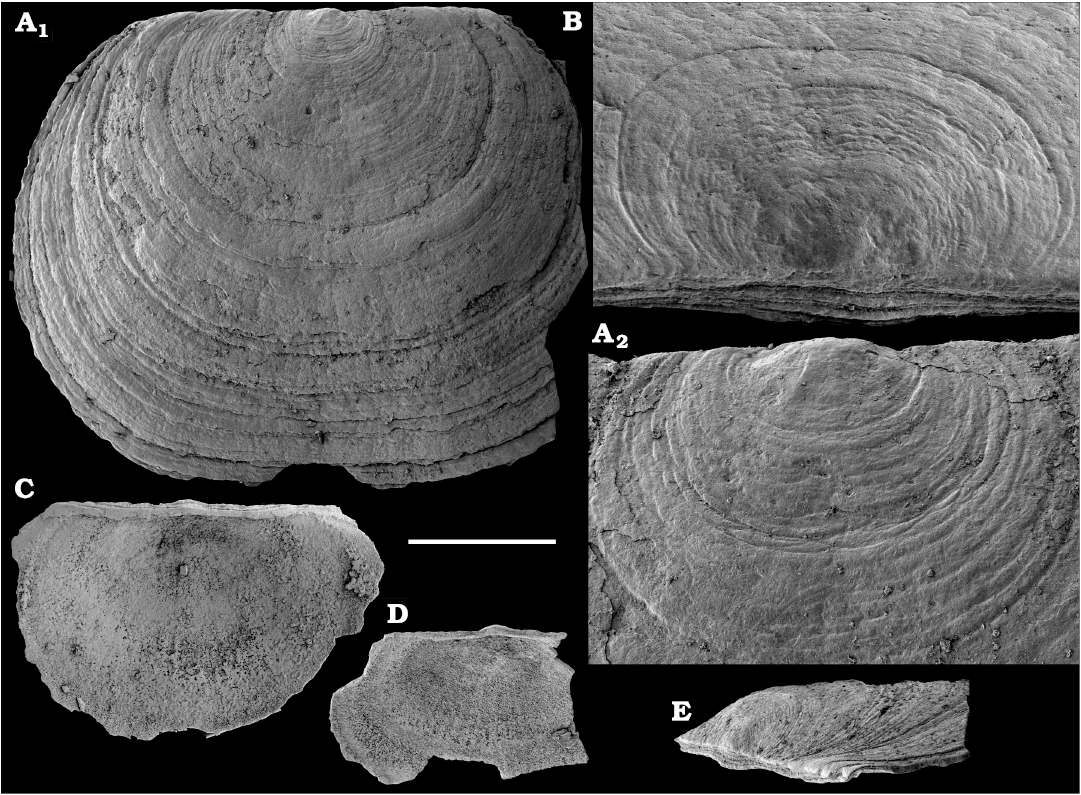
Fig. 16. Paterinid brachiopod Pelmanotreta neguertchenensis (Pelman, 1977) from the lower Cambrian Emyaksin Formation, Bol’shaya Kuonamka River, Siberia, Russia; sample 7/27.5. A–E. Dorsal valves, SMNH Br151254–151258, respectively. A2,close-up of the brephic shell. Scale bar: 1 mm (A1, C–E) and 250 µm (A2, B).
Remarks.—See Skovsted et al. (2015) for discussion of this species and genus.
Stratigraphic and geographic range.—Dokidocyathus regularis Zone, Tommotian stage (correlated with the upper part of Cambrian Stage 2)–Nochoroicyathus kokoulini Zone, Atdabanian stage (correlated with the Cambrian Stage 3), Siberian Platform.
Genus Olenekotreta Ushatinskaya, 2016
Type species: Olenekina olenekensis Ushatinskaya, 1997; Agnostus pisiformis–Proagnostus bulbus Zone, upper Mayan stage (correlated with the Guzhangian Stage); middle reaches of the Kotuj River, Eira Formation.
Olenekotreta olenekensis (Ushatinskaya, 1997)
Fig. 17.
Material.—Ten calcium phosphatic dorsal and ventral valves and apical fragments, including figured SMNH Br151259–151264, from sample 8/17.6, Kuonamka Formation, Bol’shaya Kuonamka River, Siberia, Russia. Kounamkites Zone, Amgan stage (correlated with the Wuliuan Stage).
Description.—Fragmentary ventral and dorsal valves, up to 2 mm wide, with the outer surface ornamented with concentric wavy growth lines superimposed with radially oriented plication. Ventral valve convex, with apex overhanging posteriorly, pseudointerarea orthocline with an open broad subtriangular delthyrium. Dorsal valve gently convex with straight posterior part and wide notothyrium. Brephic shell, 400–450 µm wide and 250–350 µm long, with numerous flattened tubercles ca. 5 µm in diameter. Brephic ventral valve with a single bulge at the apex and a pair of lateral bulges anteriorly. Brephic dorsal valve with a single bulge at the apex and two pairs of bulges in the middle and anterior parts.

Fig. 17. Paterinid brachiopod Olenekotreta olenekensis Ushatinskaya, 1997 from the middle Cambrian Kuonamka Formation, Bol’shaya Kuonamka River, Siberia, Russia; sample 8/17.6. A, C, E. Ventral valves, SMNH Br151259–151261, respectively. B, D, F. Dorsal valves, SMNH Br151262–151264, respectively. Scale bar: 500 µm (A–D) and 250 µm (E, F).
Remarks.—See description and comparison by Ushatinskaya (1997, 2016).
Stratigraphic and geographic range.—Amgan and Mayan stages (correlated with the Wuliuan–Guzhangian stages), Siberian Platform.
Genus Dictyonina Cooper, 1942
Type species: Trematis pannulus White, 1874 (see Cooper 1942: 228); Cambrian Stage 4–Wuliuan Stage; Pioche Shale, Nevada, USA.
Dictyonina pannula (White, 1874)
Fig. 18.
Material.—Single dorsal and four ventral calcium phosphatic valves, including figured SMNH Br151265–151268, from sample 19/92, Yunkyulyabit-Yuryakh Formation, Khorbusuonka River, Siberia, Russia. Anopolenus henrici–Corynexochus perforatus Zone, Mayan stage (correlated with the Drumian Stage).
Description.—Ventral valve semicircular in outline, up to 1.4 mm wide and 1.2 mm long, strongly convex with prominent apex overhanging the posterior margin. Ventral interarea with a broad subtriangular delthyrium. Dorsal valve slightly convex, notothyrium broadly triangular. The outer surface of both valves ornamented with concentric wavy folds superimposed with radially oriented plication. Brephic shell, 400–550 μm wide and 250–350 μm long, with a smooth surface. Brephic ventral valve with a single bulge at the apex. Brephic dorsal valve semicircular and without distinct elevations.

Fig. 18. Paterinid brachiopod Dictyonina pannula (White, 1874) from the middle Cambrian Yunkyulyabit-Yuryakh Formation, Khorbusuonka River, Siberia, Russia; sample 19/92. A, B, D. Ventral valves, SMNH Br151265–151267, respectively. A1, B3, view on the external surface; A2, B1, lateral view; A3, oblique lateral view; B2, posterior view. C. Dorsal valve, SMNH Br151268. C1, posterior view; C2, external surface. Scale bar 500 µm.
Remarks.—See description and comparison by Rowell (1980), Basset-Butt (2016), Pegel et al. (2016).
Stratigraphic and geographic range.—Wuliuan–Guzhangian stages, Siberian Platform and Laurentia.
Paterinida gen. and sp. indet.
Fig. 19.
Material.—Five fragmentary calcium phosphatic ventral valves, including figured SMNH Br151269–151272, from samples 11/2B and 11/16.05, basal Erkeket Formation, Khorbusuonka River, Siberia, Russia. Dokidocyathus regularis Zone, Tommotian stage (correlated with the upper part of Cambrian Stage 2).
Description.—Ventral valve semicircular in outline, up to 1.4 mm wide and 1.2 mm long, strongly convex with prominent apex overhanging the posterior margin. Ventral interarea with a broad subtriangular delthyrium. The outer surface ornamented with concentric wavy folds superimposed with radially oriented plication and radial striation. Brephic ventral valve smooth almost circular, 700–800 μm in diameter, with straight posterior and rounded anterior margins, carries radial furrows terminating at the margins and probably representing setal canals.
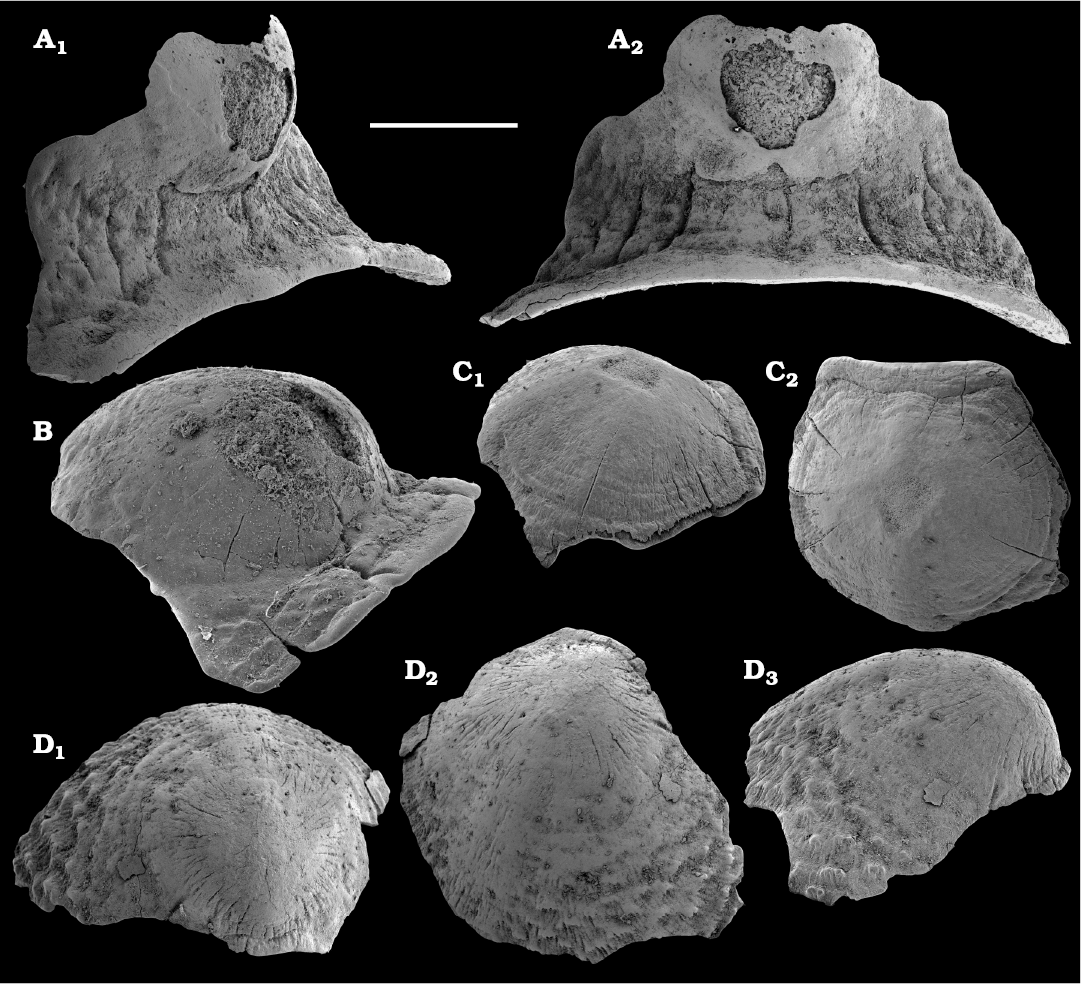
Fig. 19. Paterinida gen. and sp. indet., ventral valves, from the lower Cambrian Erkeket Formation, Khorbusuonka River, Siberia, Russia; samples 11/2B (A, C, D) and 11/16.05 (B). A. SMNH Br151269. Apical part and subapical proximal margin of the shell. A1, oblique posterior view; A2, posterior view. B. SMNH Br151270. Apical fragment with subapical margin. C. SMNH Br151271. Juvenile specimen showing brephic shell. C1, lateral view; C2, plan view. D. SMNH Br151362. Fragment with distal part of the shell. D1, posterior view; D2, external surface of the fragment; D3, lateral view. Scale bar 500 µm.
Phylum, class, order, and family uncertain
Stem group Brachiopoda indet.
Fig. 20.
Material.—Two shell fragments consisting of calcium phosphate, SMNH X11001–11002, sample 20/1B. Erkeket Formation, Khorbusuonka River, Siberia, Russia. Lower Botoman Stage (correlated with the lower part of Cambrian Stage 4).
Description.—Both fragments have the same prismatic columnal microstructure consisting of tightly packed continuous prismatic columns, ca. 10 μm in cross-section, oriented perpendicularly to the shell surface. Prisms tightly packed, with density ca. 100 prisms per 0.01 mm2, up to 200 µm in length, do not branch or anastomose, tetragonal (mostly rectangular) or rarely triangular in cross-section. In the plan view, adjacent prisms tend to be aligned and form parallel and concentric sets on the shell surface. Imbricated growth increments have serrated margins, where protruded parts house openings of the tangential canals, 40–50 μm in diameter, running sub-parallel to the surface of growth lamellae in radial direction with respect to the shell margin. Narrower orthogonal canals, 10–15 µm in diameter, are sub-perpendicular to lamination. Their openings are scattered on the shell exterior and some of them are located within openings of the tangential canals.
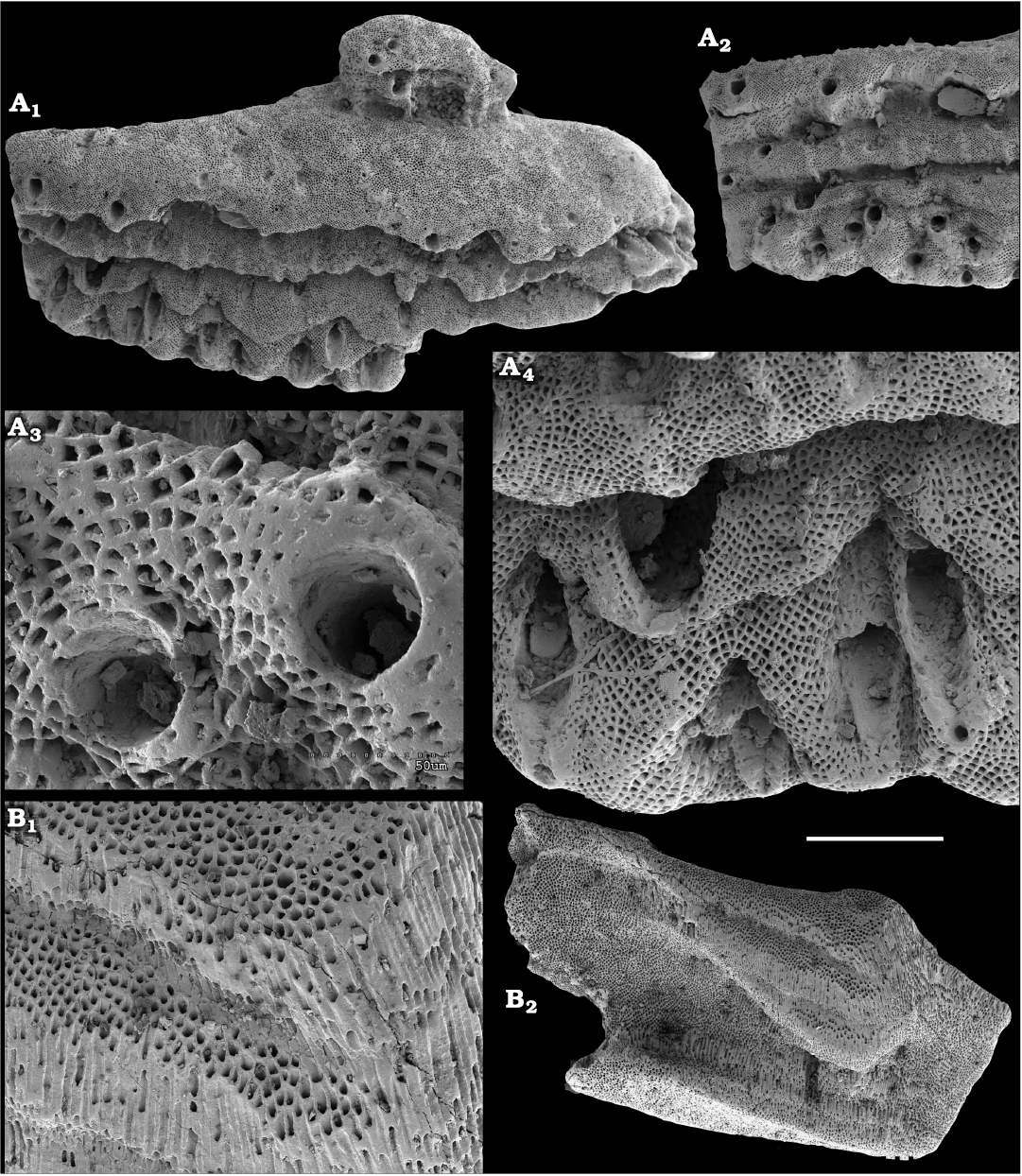
Fig. 20. Stem-group Brachiopoda indet. from the lower Cambrian Erkeket Formation, Khorbusuonka River, Siberia, Russia; sample 20/1B. Fragments of shell, SMNH X11001 (A) and SMNH X11002 (B). A1, B2, general view; A3, A4, close-ups showing prismatic walls and openings of presumably setigerous canals; B1, enlargement showing prismatic wall. Scale bar: 500 µm (A1, A2, B2), 125 µm (A4, B1), and 50 µm (A3).
Remarks.—The fossil is represented by presumably marginal anterior (Fig. 20A) and subapical posterior (Fig. 20B) shell fragments. The shell fragments are ca. 2 mm each and their morphology suggests ca. 10 mm diameter of the shell. The prismatic microstructure of the wall is homologous to the layer with compact lamination in the wall of Oymurania gravestocki Ushatinskaya in Kouchinsky et al., 2015 (Kouchinsky et al. 2015b; Kouchinsky and Bengtson 2017). Shell prisms in Oymurania are however mainly hexagonal and irregularly spaced in the plan view. In having prismatic microstructure of the wall and two sub-perpendicular sets of canals, the new form is most similar to Oymurania, but the overall shell morphology is yet unknown. Unlike other stem-group brachiopods, such as Oymurania Ushatinskaya in Kouchinsky et al., 2015 and Setatella Skovsted, Streng, Knight, and Holmer, 2010, and linguliform brachiopods (Williams and Holmer 1992; Cusack et al. 1999; Williams and Cusack 1999) as well as more basal representatives of stem group brachiopods, such as tannuolinids Tannuolina Fonin and Smirnova, 1967, and Micrina Laurie, 1986, the acrotretoid columnar microstructure was not observed in this study. However, a comparable prismatic shell structures lacking acrotretoid columns have been documented from tommotiids such as Eccentrotheca Landing, Nowlan, and Fletcher, 1980, and also in paterinid brachiopods such as Salanygolina Ushatinskaya, 1987 (Balthasar et al. 2009; Holmer et al. 2009). Tangential and orthogonal canals are known from shells of tannuolinids and mickwitziids. The tangential canals represent setigerous tubes similar to the epipunctae (Jin et al. 2007), whereas the Micrina–Setatella type orthogonal canals are interpreted as punctae with mantle outgrowths (caeca) reaching the periostracum (Pérez-Huerta et al. 2009). The new form is interpreted herein to be a stem group brachiopod with phosphatic shell and demonstrates a novel combination of features known from tannuolinids and mickwitziids as well as linguliform brachiopods. Canal openings scattered on the outer shell surface have single apertures. In Oymurania, however, the tangential canals are not found within the inner shell layer of tightly packed prismatic columns (Kouchinsky and Bengtson 2017). Subdivision of prisms into stacked tablets is not observed in the new form described herein.
Phylum and class uncertain
Order Tommotiida Missarzhevsky, 1970
Family Tannuolinidae Fonin and Smirnova, 1967
Genus Tannuolina Fonin and Smirnova, 1967
Type species: Tannuolina mullifora Fonin and Smirnova 1967; Atdabanian stage; Altay Sayan Foldbelt, Russia.
Tannuolina cf. pavlovi Kouchinsky, Bengtson, and Murdock, 2010
Fig. 21.
Material.—Two fragmentary mitral sclerites, SMNH X11003, 11004 and a fragment SMNH X11005, from sample 19/3.25. Erkeket Formation, Khorbusuonka River, Siberia, Russia. Lower Botoman stage (correlated with the lower part of Cambrian Stage 4).
Description.—The apical fragments covered with co-marginal undulating folds with circular pores, up to 50 μm in diameter scattered on the accrescent side. The openings increase in diameter from the apical part towards the aperture. The apical part is flattened, with pores, ca. 5 μm in diameter along its periphery. The decrescent side is not preserved. The accrescent side passes into a flattened lateral side. Pores numerous on the accrescent side, but scarce and small, ca. 5 μm on the lateral side. The inner surface of sclerites smooth.
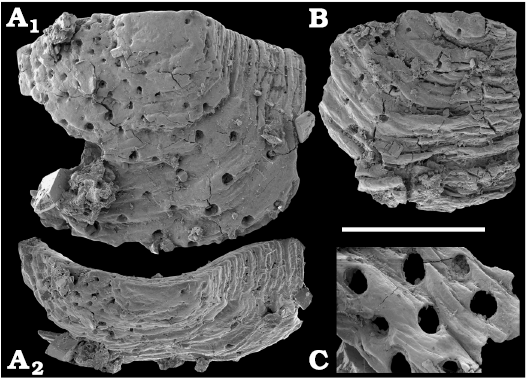
Fig. 21. Tommotiid Tannuolina cf. pavlovi from the lower Cambrian Erkeket Formation, Khorbusuonka River, Siberia, Russia; sample 19/3.25. A, B. Mitral sclerites, SMNH X11003 and 11004, respectively. A1, view on the accrescent side; A2, apical view. C. Fragment of sclerite with large pores, SMNH X11005. Scale bar 250 µm.
Remarks.—The fossils are similar to apical parts of mitral sclerites of Tannuolina pavlovi Kouchinsky, Bengtson, and Murdock, 2010, from the lower Tommotian stage of Siberian Platform, but further comparison is hampered by incomplete preservation.
Family Tommotiidae Bengtson, 1970
Genus Camenella Missarzhevsky in Rozanov and Missarzhevsky, 1966
Type species: Camenella garbowskae Missarzhevsky in Rozanov and Missarzhevsky, 1966; Dokidocyathus regularis Zone, Tommotian stage; Chekurovka Village, lower Lena River, Siberia, Russia.
Camenella garbowskae Missarzhevsky in Rozanov and Missarzhevsky, 1966
Fig. 22.
Material.—Several tens of calcium phosphatic sellate and mitral sclerites, including figured SMNH X11006–11017, from samples: 11/18.7, 11/19.5, 11/21.6, 11/28.8. Erkeket Formation, Khorbusuonka River, Siberia, Russia. Dokidocyathus regularis Zone, Tommotian stage (correlated with the upper part of Cambrian Stage 2).
Description.—Sellate sclerites (Fig. 22G–J, L) asymmetrical, up to 1.2 mm long and wide in planar view. Sella, a deep longitudinal depression, 1/4 to 1/3 of the sclerite width, divides sclerites into larger and smaller lobes. The larger lobe is higher, approximately twice as long and more than four times the width of the smaller lobe and carries 10–15 variously expressed radial ridges. Smaller lobe ornamented with longitudinal striations. Co-marginal folds well expressed on the dorsal surface of sclerites. Apex slightly coiled. Duplicature adpressed to the inner surface near the apex. Inner surface of sclerites smooth with gently expressed co-marginal folds.
Mitral sclerites (Fig. 22A, B, E) asymmetrical pyramidal, up to 1 mm long, from apex to the aperture and up to 1.2 mm wide, covered with distinct concentric growth lines. Obplicate side carries several faint radial ridges and striation (see description of morphological terms in Kouchinsky et al. 2015a: comments to fig. 37A). Plicate side with four prominent radial ridges. Accrescent and decrescent sides without radial ridges. Apex slightly inclined over the decrescent side.

Fig. 22. Tommotiid Camenella Missarzhevsky in Rozanov and Missarzhevsky, 1966 from the lower Cambrian Erkeket Formation, Khorbusuonka River, Siberia, Russia; samples 11/19.5 (A, E, G), 11/18.7 (B), 11/21.6 (C, D, F, H, I, J, L), and 11/28.8 (K). A, B, E. Camenella garbowskae Missarzhevsky in Rozanov and Missarzhevsky, 1966, mitral sclerites, SMNH X11006–11008, respectively. F. Camenella plana (Missarzhevsky in Rozanov et al., 1969), mitral sclerite SMNH X11009, tentatively synonymized with C. garbowskae. C, G–J, L. Camenella garbowskae Missarzhevsky in Rozanov and Missarzhevsky, 1966, sellate sclerites, SMNH X11010–11015, respectively. D. Camenella cf. C. garbowskae, sellate sclerite, SMNH X11016. K. Camenella aff. C. garbowskae, sellate sclerite, SMNH X11017. Scale bar: 500 µm (A–D, F–I, K, L) and 1 mm (E, J).
Remarks.—For emended diagnosis and discussion of the Camenella see Bengtson (1986) and Skovsted et al. (2009). For emended diagnosis of C. garbowskae see Kouchinsky et al. (2015a).
Sellate sclerites with a flared margin (Fig. 22D) identical to Camenella cf. C. garbowskae in Kouchinsky et al. (2015a: fig. 39) may well also belong to the C. garbowskae scleritome.
Camenella aff. C. garbowskae Missarzhevsky, 1966 in Rozanov and Missarzhevsky, 1966 (Fig. 22K) is represented by a single dextral sellate sclerite from sample 11/28.8. The sclerite has five prominent radial ribs and distinct commarginal folds. Similar sclerites are described from the Tommotian and Atdabanian stages of Siberia, from the Emyaksin and Tyuser formations (Rozanov and Missarzhevsky 1966: pl. 13: 11–12; Rozanov et al. 1969: 169; Kouchinsky et al. 2015a: fig. 43). Planiform mitrals (Fig. 22F), similar to Camenella plana (Missarzhevsky in Rozanov et al., 1969) known to co-occur with C. garbowskae and can probably be included in C. garbowskae scleritome (Bengtson 1986; Kouchinsky et al. 2015a).
Stratigraphic and geographic range.—Tommotian stage, Siberia, Russia.
Family Lapworthellidae Missarzhevsky in Rozanov and Missarzhevsky, 1966
Genus Lapworthella Cobbold, 1921
Type species: Lapworthella nigra Cobbold, 1921; Callavia Zone, lower Cambrian; Comley, Shropshire, Avalonian Britain.
Lapworthella lucida Meshkova, 1969
Fig. 23A–F, J, K.
1969 Lapworthella lucida Meshkova sp. nov.; Meshkova 1969: 167, pl. 53: 5.
1974 Lapworthella lucida; Repina et al. 1974: 81, pl. 18: 7, 8.
Material.—Several tens of calcium phosphate sclerites, including figured SMNH X11018–11025, from samples 19/1.5, 19/3.5, 19/5.5, 20/1B, lower Botoman stage (correlated with the lower part of Cambrian Stage 4), Khorbusuonka River. Several sclerites from sample 21/112, upper Judomia–Uktaspis (Prouktaspis) Zone, Atdabanian stage (correlated with the Cambrian Stage 3), lower Lena River. Siberia, Russia.
Description.—Straight or irregularly curved sclerites up to 2 mm long with rounded isometrical aperture, up to 0.5 mm in diameter. Wall with transversal ribs covered with growth lines and tubercles directed towards the apex of sclerite alternating with relatively much broader intervening rings covered with tubercles of two distinctly different sizes. The apical end of sclerite is extended, tapered and tuberculated.
Remarks.—Different from other species in having broader intervening spaces (rings) between ridges and tubercles of two distinctly different sizes scattered on the surface of the rings and apical part.
Stratigraphic and geographic range.—Upper part of Cambrian Stage 3–lower Stage 4, Siberian Platform.
Lapworthella tortuosa Missarzhevsky in Rozanov and Missarzhevsky, 1966
Fig. 23G–I, L–N.
Material.—Several tens of sclerites from samples 11/18.7, 11/19.5, 11/21.6, Khorbusuonka River and several sclerites, including illustrated SMNH X11026–11031, from sample 21/22.7, Lena River. Siberia, Russia; Dokidocyathus regularis Zone, Tommotian stage (correlated with the upper part of Cambrian Stage 2).
Description.—Elongated conical sclerites, up to 1 mm long with rounded isometrical, subrectangular or more extended oval aperture, up to 0.5 mm wide. Almost straight or gently curved in one plane, sometimes irregularly curved at the pointed tip. External ornamentation consists of distinct transversal denticulated ridges bearing growth lines. Distance between ridges ca. 100 µm. Rings between ridges are concave with numerous tubercles and are wider than smooth surface of the ridges.

Fig. 23. Tommotiid Lapworthella Cobbold, 1921 from the Cambrian Erkeket Formation, Khorbusuonka River (A–D, F–L, O–Q) and Tyuser Formation, lower reaches of the Lena River (E, M, N), Siberia, Russia; samples 11/18.7 (G, L), 11/21.6 (H, I), 19/1.5 (D), 19/5.5 (F, J, K), 19/31.75 (Q), 20/1B (A–C), 20/5B (O, P), 21/22.7 (M, N), 21/112 (E). A–F, J, K. Lapworthella lucida Meshkova, 1969 sclerites, SMNH X11018–11025, respectively. G–I, L–N. Lapworthella tortuosa Missarzhevsky in Rozanov and Missarzhevsky, 1966 sclerites, SMNH X11026–11031, respectively. O–Q. Lapworthella dentata Missarzhevsky in Rozanov et al., 1969 sclerites, SMNH X11032–11034, respectively. Scale bar 250 µm.
Remarks.—Forms with isometrical and flattened cross-sections are known from the same samples. Irregularly curved tip in a single sclerite attributed to the same species has not been previously reported.
Stratigraphic and geographic range.—Tommotian stage, Siberian Platform.
Lapworthella dentata Missarzhevsky in Rozanov et al., 1969
Fig. 23O–Q.
Material.—Single sclerite, SMNH X11032, from sample 19/31.75 and several tens of sclerites, including figured SMNH X11033 and 11034, from sample 20/5B, Khorbusuonka River, Siberia, Russia. Toyonian stage (correlated with Cambrian Stage 4).
Description.—Pyramidal, slightly curved sclerites, up to 1 mm long, with oval or subrectangular aperture, up to 0.5 mm wide. External ornamentation consists of distinct transversal ridges bearing short denticles. The density of denticles is 35–50 per mm. Distance between ridges 50–100 µm. Rings between ridges are concave with circumferential growth lines.
Stratigraphic and geographic range.—Cambrian Series 2, Siberian Platform. Occurrence beyond the Siberian Platform depending on synonymy with Lapworthella schodackensis (Lochman, 1956) from Laurentia (Landing, 1984) and Lapworthella cornu (Wiman, 1903) from Baltica and Avalonia (see discussion by Devaere and Skovsted 2017).
Phylum Mollusca Cuvier, 1797
Class uncertain (stem-group Conchifera)
Order Helcionellida Geyer, 1994
Family Helcionellidae Wenz, 1938
Genus Davidonia Parkhaev, 2017
Type species: Mellopegma rostrata Zhou and Xiao, 1984; Cambrian stages 3–4, Houjiashan Formation; Huoqiu, Anhui Province, North China.
Davidonia anabarica (Parkhaev, 2005)
Fig. 24A, B.
Material.—Several hundred calcium phosphatic internal moulds, including figured specimen SMNH Mo194701 and 194702, from samples 19/11.75 and 19/12.75, lower Botoman stage (correlated with the lower part of Cambrian Stage 4), Erkeket Formation, Khorbusuonka River. Additional material attributed to Davidonia cf. anabarica represents two internal moulds, from samples 19/31.75 and 19/33, Toyonian stage (correlated with the Cambrian Stage 4), Erkeket Formation, Khorbusuonka River and several internal moulds from sample 22/67, Sekten Formation, lower Lena River, Kounamkites Zone, Amgan stage (correlated with the Wuliuan Stage). Siberia, Russia.
Remarks.—The fossils are identical to those from the lower part of the Botoman stage, eastern flanks of the Anabar Uplift described by Kouchinsky et al. (2015a). As pointed out by Kouchinsky et al. (2015a: 430), Davidonia anabarica has tuberculated internal moulds, otherwise similar to Davidonia rostrata. The latter is reported from the Cambrian Stages 3–4 and lower Wuliuan Stage and occurs in Gondwana (Bengtson et al. 1990; Wotte 2006; Claybourn et al. 2019; Li et al. 2019) and Laurentia (Skovsted 2004).
Stratigraphic and geographic range.—Lower Botoman stage (correlated with the lower part of Cambrian Stage 4), Siberian Platform.
Genus Bemella Missarzhevsky in Rozanov et al., 1969
Type species: Helcionella jacutica Missarzhevsky in Rozanov and Missarzhevsky, 1966; Dokidocyathus regularis Zone, Tommotian stage; Tiktirikteekh, near Churan village, middle Lena River, Siberia, Russia.
Bemella jacutica (Missarzhevsky in Rozanov and Missarzhevsky, 1966)
Fig. 24M, N.
Material.—Several calcium phosphatic internal moulds, including figured SMNH Mo194703–194704, from sample 11/16.1, basal Erkeket Formation, Khorbusuonka River, Siberia, Russia. Dokidocyathus regularis Zone, Tommotian stage (correlated with the upper part of Cambrian Stage 2).
Remarks.—Smooth or having broad concentric folds internal moulds of rapidly expanding shells coiled less than 1/2 revolution, with broad oval aperture and rounded blunt initial part (see Kouchinsky et al. 2015a). Other low cyrtoconic forms from samples 11/2B and 11/16.1 represented by internal moulds, SMNH Mo194715 and Mo194716, with broad apertures and transversal ribs cannot be undoubtedly defined herein. They are referred to as Helcionellidae indet. (Fig. 24J, K).
Stratigraphic and geographic range.—Cambrian Stage 2, Siberian Platform.
Genus Obtusoconus Yu, 1979
Type species: Obtusoconus paucicostatus Yu, 1979; middle Meishucunian stage (upper Fortunian–lower part of Cambrian Stage 2); Yichang County, western Hubei Province, South China.
Obtusoconus cf. mirabilis Vasiljeva, 1990
Fig. 24E, F.
Material.—Two calcium phosphatic internal moulds, SMNH Mo194705 and 194706, from samples 11/2B and a single mould from sample 11/16.5, basal Erkeket Formation, Khorbusuonka River, Siberia, Russia. Dokidocyathus regularis Zone, Tommotian stage (correlated with the upper part of Cambrian Stage 2).
Remarks.—Bilaterally symmetrical, rapidly expanding orthoconic laterally compressed shells with oval aperture (length/width ratio ca. 2) and a globular, slightly laterally displaced protoconch. The inner shell surface (as replicated on the internal moulds) with regularly spaced broad transverse folds. Specimens described as Obtusoconus cf. mirabilis Vasiljeva, 1990, by Kouchinsky et al. (2017: fig. 13A, C) have irregularly spaced transverse folds. See Kouchinsky et al. (2017) for comparison with other species of Obtusoconus.
Genus Parailsanella Zhegallo in Voronova et al., 1987
Type species: Parailsanella acris Zhegallo in Voronova et al., 1987; Fallotaspis Zone, Montezuman stage; Mackenzie Mountains, Northwest Territories, Canada.
Parailsanella sp.
Fig. 24G, H.
Material.—Several calcium phosphatic internal moulds from sample 21/54, Tyuser Formation, Delgadella anabara–Nevadella Zone, Atdabanian stage, 22/0 from top Tyuser Formation, Bergeroniellus micmacciformis–Erbiella Zone, lower Botoman stage (correlated with the lower Cambrian Stage 4), and 22/50 from the Sekten Formation, upper Toyonian stage (correlated with the Cambrian Stage 4) and including two figured specimens, SMNH Mo194707 and 194708, from sample 22/50. Ulakhan-Ald’yarkhay, lower Lena River. Siberia, Russia.
Remarks.—Slightly cyrtoconic, laterally moderately compressed conical calcium phosphatic internal moulds with oval aperture (length/width ratio ca. 2), regular transverse ribs and bulbous apex. Those steinkerns may represent different species, but their overall morphology is compatible with species of Parailsanella, notably Parailsanella sp. 2 from the lower Cambrian Stage 4 (herein) of Siberian Platform (Kouchinsky et al. 2015a: fig. 7) and Parailsanella luonanensis Li, Zhang, Skovsted, Yun, Pan, and Li, 2019, from Series 2 of North China.
Genus Horsegullia Parkhaev, 2004
Type species: Yorkiella horsegulliensis Parkhaev in Gravestock et al., 2001; Cambrian stages 3–4, Mernmerna Formation, Horse Gully, Yorke Peninsula, South Australia (Gravestock et al. 2001: pl. 33: 1). See discussion by Parkhaev (2004) on the preoccupied name Yorkiella Parkhaev in Gravestock et al., 2001.
Horsegullia sp.
Fig. 24L, O, P.
Material.—Several tens of phosphatic internal moulds, including figured SMNH Mo194709–194711, from sample 20/1B, Erkeket Formation, Khorbusuonka River, Siberia, Russia. Lower Botoman stage (correlated with the lower Cambrian Stage 4).
Description.—Smooth internal moulds coiled through 1/4 of a whorl and moderately laterally compressed. Initial part indistinct. Apex bluntly rounded and gently hooked, projecting beyond the apertural margin. The aperture has a drop-like outline, with length/width ratio ca. 1.8, with rounded margins tapering towards the subapical side of the shell. The parietal train on the subapical side is low, narrow and short.

Fig. 24. Molluscs from the lower Cambrian Erkeket Formation, Khorbusuonka River (A–F, J–N) and Sekten Formation, lower reaches of the Lena River (G–I), Siberia, Russia; samples 11/2B (E, F, J), 11/16.1 (K, M, N), 19/12.75 (A), 19/11.75 (B), 20/1B (L, O, P), 20/6B (C, D), and 22/50 (G–I). A, B. Davidonia anabarica (Parkhaev, 2005), internal moulds, SMNH Mo194701 and 194702, respectively. C. Mellopegma sp., internal mould, SMNH Mo194713. D. Stenotheca cf. drepanoida (He and Pei in He et al., 1984), silicified shell, SMNH Mo194714. D1, lateral view; D2, oblique apertural view. E, F. Obtusoconus cf. mirabilis Vasiljeva, 1990, internal moulds, SMNH Mo194705 and 194706, respectively. G, H. Parailsanella sp., internal moulds, SMNH Mo194707 and Mo194708, respectively. I. Protowenella flemingi Runnegar and Jell, 1976, internal mould, SMNH Mo194712. M, N. Bemella jacutica (Missarzhevsky in Rozanov and Missarzhevsky, 1966), internal moulds, SMNH Mo194703 and 194704, respectively. J, K. Helcionellidae indet., internal moulds, SMNH Mo194715 and 194716, respectively. K1, subapical view; K2, upper view. L, O, P. Horsegullia sp., internal moulds, SMNH Mo194709–194711. L1, subapical; L2, O1, lateral; L3, upper; and O2, oblique apertural views. Scale bar: 1 mm (K2), 500 µm (A–C, E–H, J, K1, L –P), and 250 µm (D, I).
Remarks.—The form is different from Horsegullia horsegulliensis by a less extended but more rounded blunt apex as well as by a more elongated aperture and a less distinct parietal train. The latter species is described from Cambrian stages 3–4 of Gondwana (Australia and North China; Gravestock et al. 2001; Li et al. 2021).
Family Stenothecidae Runnegar and Jell, 1980
Genus Mellopegma Runnegar and Jell, 1976
Type species: Mellopegma georginense Runnegar and Jell, 1976; Drumian Stage; Gowers Formation, Georgina Basin, South Australia.
Mellopegma sp.
Fig. 24C.
Material.—Single internal mould, SMNH Mo194713, from sample 20/6B, Erkeket Formation, Khorbusuonka River, Siberia, Russia. Toyonian stage (correlated with the Cambrian Stage 4).
Remarks.—Laterally compressed internal mould with faint transverse folds and deep subapical sinus. The apex is slightly shifted dorsally (supra-apically) and abut with a bulbous initial part. Different from the most similar Mellopegma indecorum (Missarzhevsky in Rozanov et al., 1969) in having a prominent bulbous initial part and lacking subapical curvature.
Genus Stenotheca Salter in Hicks, 1872
Type species: Stenotheca cornucopia Salter in Hicks, 1872. See remarks on the type species by Vendrasco et al. (2011b).
Stenotheca cf. drepanoida (He and Pei in He et al., 1984)
Fig. 24D.
Material.—Single specimen, SMNH Mo194714 represented by a silicified shell from sample 20/6B, Erkeket Formation, Khorbusuonka River, Siberia, Russia. Toyonian stage (correlated with the Cambrian Stage 4).
Remarks.—Shell strongly laterally compressed and coiled through 1/4 of a whorl, fragmentarily preserved without apex. The fossil is similar to Stenotheca drepanoida (He and Pei in He et al. 1984) known from the upper part of Cambrian Stage 3–lower Stage 4 of South China, South Australia, and Siberia (Vendrasco et al. 2011a).
Class, order, and family uncertain
Watsonella crosbyi Grabau, 1900, represents a transitional form between laterally compressed univalved molluscs such as Anabarella plana Vostokova, 1962 and the earliest bivalves-fordillids, Fordilla Barrande, 1881 and Pojetaia Jell, 1980 (e.g., Vendrasco et al. 2011a, b; Kouchinsky et al. 2017; Guo et al. 2021).
Genus Watsonella Grabau, 1900
Type species: Watsonella crosbyi Grabau, 1900; Camenella baltica Zone?, upper part of Cambrian Stage 2–lower part of Cambrian Stage 3; Boulders at Sandy Cove and Pleasant Beach, Cohasset, Massachusetts, USA.
Watsonella crosbyi Grabau, 1900
Fig. 25.
Material.—Two calcium phosphatic internal moulds and a single external mould, including figured SMNH Mo194717 and 194718, from samples 15/23, upper Kessyusa Group, Chuskuna Formation and 11/16.1, basal Erkeket Formation, Khorbusuonka River, Siberia, Russia. Cambrian Stage 2.

Fig. 25. Mollusc Watsonella crosbyi Grabau, 1900, from the lower Cambrian Erkeket Formation, Khorbusuonka River, Siberia, Russia; sample 15/23. Internal (A) and external (B) moulds, SMNH Mo194717 and 194718, respectively, of probably the same specimen. A1, lateral view; A2, dorsal view; B1, B3, close-ups showing replicated sculpture on the shell surface. Scale bar: 1 mm (A, B2), 250 µm (B3), and 80 µm (B1).
Remarks.—External calcium phosphatic mould representing cast of the left side of a laterally compressed ca. 3 mm long shell. The mould shows aciculate texture representing external surface of the shell. The texture forms radially oriented series of perpendicular to irregularly spaced concentric continuous growth lines. Phosphatic casts of endolith microboring within now dissolved shell attach to the surface of the mould. The canals tend to be radially oriented. Same canals are present on the surface of internal mould from the same sample. Watsonella crosbyi occurs in the upper Kessyusa Group on the Olenyok Uplift (see e.g., Kouchinsky 1999), wherefrom it ranges into the overlying Erkeket Formation.
Stratigraphic and geographic range.—Cambrian Stage 2, Siberia, western Mongolia, East Gondwana (South China, Australia), West Gondwana (Armorican France) and Avalonia (Massachusetts, Cape Breton Island, eastern Newfoundland). Cambrian Stage 3, Avalonia (Landing et al. 1989, 2013) and South Australia (Gravestock et al. 2001).
Class Bivalvia Linnaeus, 1758
Order Fordillida Pojeta, 1975
Family Fordillidae Pojeta, 1975
Genus Fordilla Barrande, 1881
Type species: Fordilla troyensis Barrande, 1881; Upper Elliptocephala asaphoides assemblage, upper Stage 3 or 4; Middle Granville Formation, continental slope debris flow in Taconian allochthon, eastern New York State, USA.
Fordilla sibirica Krasilova, 1977
Figs. 26, 27A–C.
Material.—Several calcium phosphatic internal moulds, including figured SMNH Mo167596 (Fig. 26B), Mo194719, 194720, 194723, and 194724 from samples 19/4.25, 19/11.75, 19/12.75, 19/40, 20/1B, from the Erkeket Formation, Khorbusuonka River, Botoman–Toyonian stages (correlated with the Cambrian Stage 4). Several dozens of calcium phosphatic internal moulds including figured SMNH Mo194721 and 194722, from samples 21/46.1 and 21/54, Tyuser Formation, Lena River, Delgadella anabara–Nevadella Zone, Atdabanian stage (correlated with the Cambrian Stage 3). Siberia, Russia.

Fig. 26. Bivalve mollusc Fordilla sibirica Krasilova, 1977, from the lower Cambrian Erkeket Formation, Khorbusuonka River, Siberia, Russia; sample 19/12.75. A–C. Internal moulds, SMNH Mo194719, 167596, and 194720, respectively. A1, B1, C, lateral view; A2, dorsal view, A3–A5, B2–B4, close-ups showing surface textures. Scale bar: 830 µm (C), 500 µm (A1, A2, B1), 100 µm (A3–A5, B2–B4).
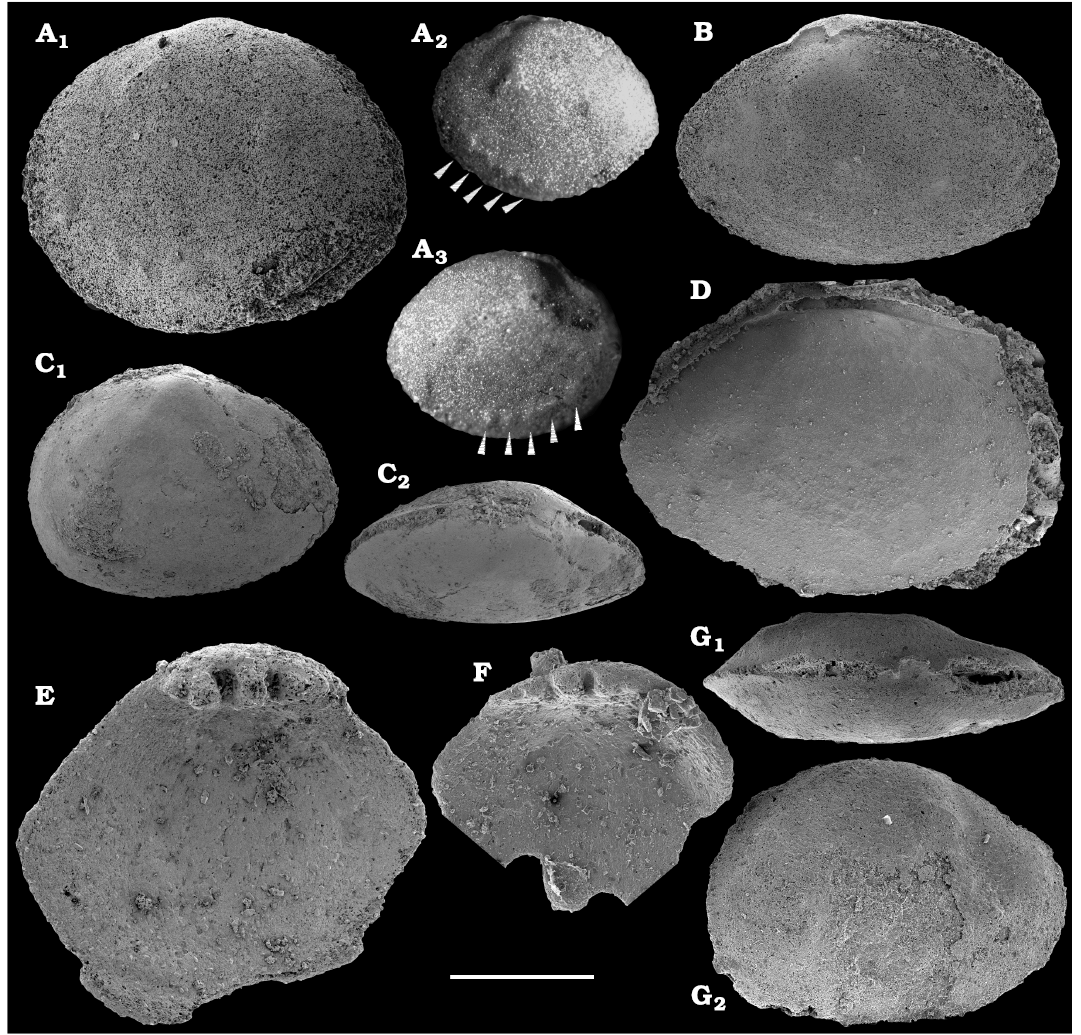
Fig. 27. Bivalve mollusc Fordilla sibirica Krasilova, 1977 from the lower Cambrian Tyuser (A, B) and Erkeket (C, D) formations and Pojetaia dentifera Kouchinsky, Bengtson, Clausen, Gubanov, Malinky, and Peel, 2011, from Sekten Formation (E–G), lower reaches of the Lena River (A, B, E–G) and Khorbusuonka River (C, D), Siberia, Russia; samples 21/54 (A, B), 20/1B (C), 19/40 (D), and 22/50 (E–G). A–G. Internal moulds, SMNH Mo194721–194727, respectively. A1, Scanning electron microscope image showing muscle scars on the surface of internal mould, left valve. A2, A3, light-microscope micrographs showing muscle scars (arrowed), left and right sides of internal mould corresponding, respectively, to inner surfaces of left and right valves; G1, dorsal; C2, oblique dorsal; and C1, G2, lateral views. Scale bar: 500 µm (A1, B–G) and 830 µm (A2, A3).
Remarks.—Shell ca. 2.0 mm in length, equivalved. Valves suboval with anteriorly shifted umbones. Dentition represented by casts of a single weakly impressed tooth and corresponding socket on each valve. Ligament area straight, opisthodetic, along more than half of the dorsal margin. Anterior pallial muscles are evidenced by five nodular scars on the left and right sides of an internal mould as well as a larger anterior adductor is preserved (Fig. 27A: arrowed). Muscular scars are similar to those reported from Fordilla sibirica by Krasilova (1977, 1987). According to Elicki and Gürsu (2009), Fordilla can be distinguished from Pojetaia Jell, 1980, in consistently larger size and having the umbo shifted towards the anterior margin. For that reason, the internal mould (SMNH Mo167596 from sample 19/12.75, Fig. 26B) described by Vendrasco et al. (2011a: pl. 1: 1, 3–6) as Pojetaia runnegari is reinterpreted herein as Fordilla sibirica, in a way similar to other specimens from the same sample. Microstructures of Pojetaia and Fordilla differ therefore more significantly than suggested by Vendrasco et al. (2011a: 838). Fordilla (Fig. 26A, B) displays a staircase-like texture throughout the entire surface, in contrast with Pojetaia with a more developed polygonal aspect of the texture. The texture on the available moulds of the left and right valves has the same clockwise imbricating pattern. Such a consistent arrangement of microstructural elements with respect to antero-posterior direction of the mollusc is presumably similarly expressed in the lineage from Anabarella via Watsonella to Fordilla (see discussion by Vendrasco et al. 2011a), and Watsonella can consequently be interpreted as belonging to the stem group Bivalvia (not Rostroconchia, see also Peel 2021c).
Stratigraphic and geographic range.—Cambrian Series 2, Siberian Platform.
Genus Pojetaia Jell, 1980
Type species: Pojetaia runnegari Jell, 1980; Cambrian Stage 3, Parara Limestone; South Australia.
Pojetaia dentifera Kouchinsky, Bengtson, Clausen, Gubanov, Malinky, and Peel, 2011
Fig. 27D–F.
Material.—Five phosphatised shell fragments, including figured SMNH Mo194725–194727, from sample 22/50, Sekten Formation, Ulakhan-Ald’yarkhay, Lena River, Siberia, Russia. Upper Toyonian stage (correlated with the Cambrian Stage 4).
Remarks.—Shell fragments with four peg-like hinge teeth. Dentition is identical to that in Pojetaia dentifera described from the Drumian Stage of Siberia by Kouchinsky et al. (2011: fig. 3), consisting of 3–5 peg-like teeth decreasing in size anteriorly. A single internal mould of a fordillid bivalve (Fig. 27F) elliptical in outline with centrally placed umbo co-occurs and probably represents the same species, although its dentition consisting of at least two teeth is incompletely preserved.
Stratigraphic and geographic range.—Upper Toyonian and Amgan stages (correlated with the upper Stage 4 to lower Drumian Stage), Siberian Platform.
Class uncertain
Order Pelagiellida Mackinnon, 1985
Family Aldanellidae Linsley and Kier, 1984
Genus Aldanella Vostokova, 1962
Type species: Pleurotomaria (Raphistoma) attleborensis Shaler and Foerste, 1888; Lower Cambrian; Hoppin Hill, Massachusetts, USA.
Aldanella sibirica Parkhaev and Karlova, 2011
Figs. 28, 29.
Material.—About 200 calcium phosphatic internal moulds with partially phosphatised shells and phosphatic crusts, including figured SMNH Mo194728–194737, from samples 11/16.05, 11/16.1, 11/2B, lower Erkeket Formation, Khorbusuonka River and samples 21/21, 21/22, Tyuser Formation, Lena River. Siberia, Russia; Dokidocyathus regularis Zone, Tommotian stage (correlated with the upper part of Cambrian Stage 2).
Description.—Dextrally coiled moderately expanding shell. Available specimens include up to two whorls and are less than 1 mm in diameter. Spire does not protrude above the last whorl. Apertural cross-section represents an extended oval with width/height ratio 1.6–1.8. The last whorl has gently convex adapical (upper) and umbilical (lower) surfaces. Gently rounded lateral periphery of the whorls spirally ornamented with fine undulating plication perpendicular to the growth lines (Fig. 29A1). Protoconch, ca. 100 μm in diameter, not clearly separated from the adult shell (Fig. 29), has an extended apical termination (Fig. 29A).
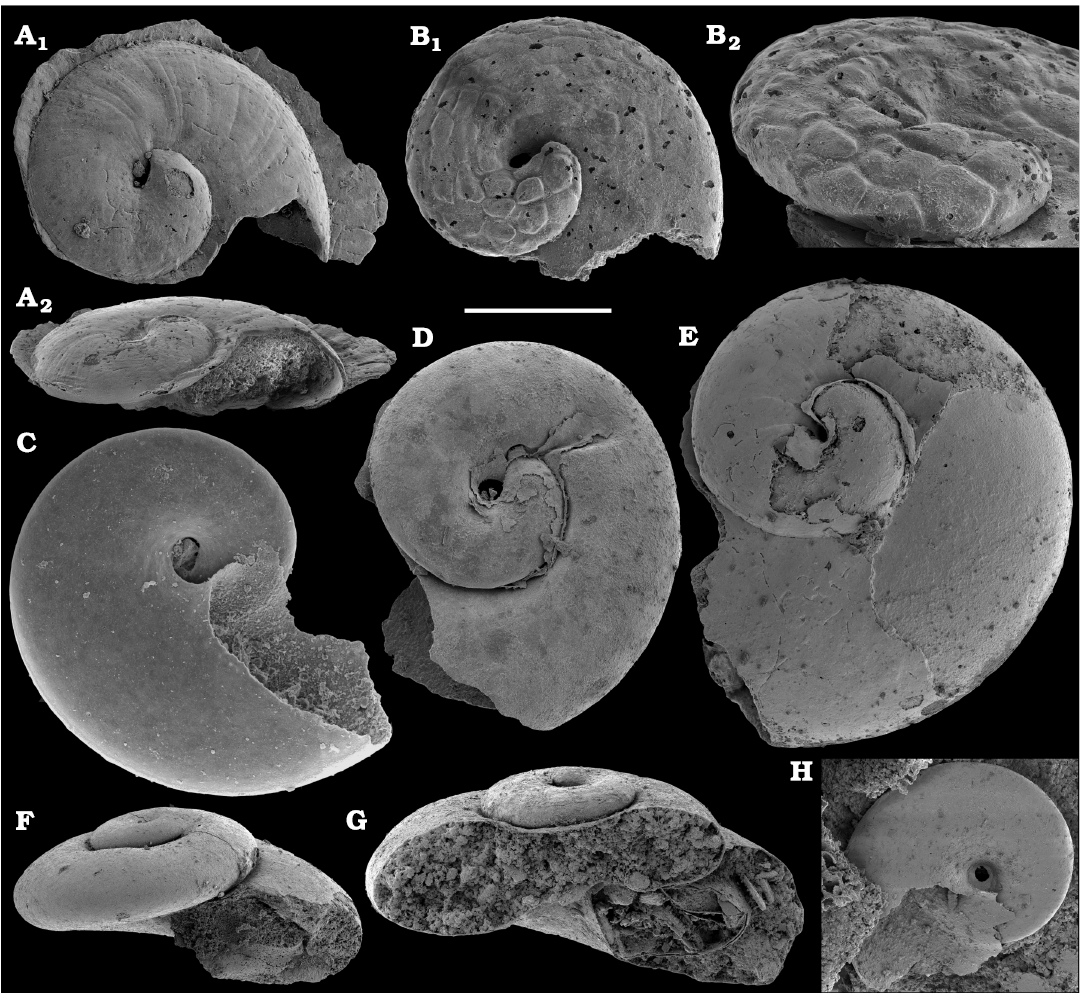
Fig. 28. Pelagiellid mollusc Aldanella sibirica Parkhaev and Karlova, 2011, from the lower Cambrian Erkeket Formation, Khorbusuonka River, Siberia, Russia; samples 11/16.05 (A, B, E–H) and 11/2B (C, D). A–H. Phosphatised shells, phosphatic coats and partially preserved phosphatic interal moulds, SMNH Mo194728–194735, respectively. A1, adapical view; A2, apertural view. B. Internal mould with probably diagenetic polygonal structures; B2, close-up showing polygons. Scale bar: 500 µm (F–H), 250 µm (A, B1, C–E), and 125 µm (B2).
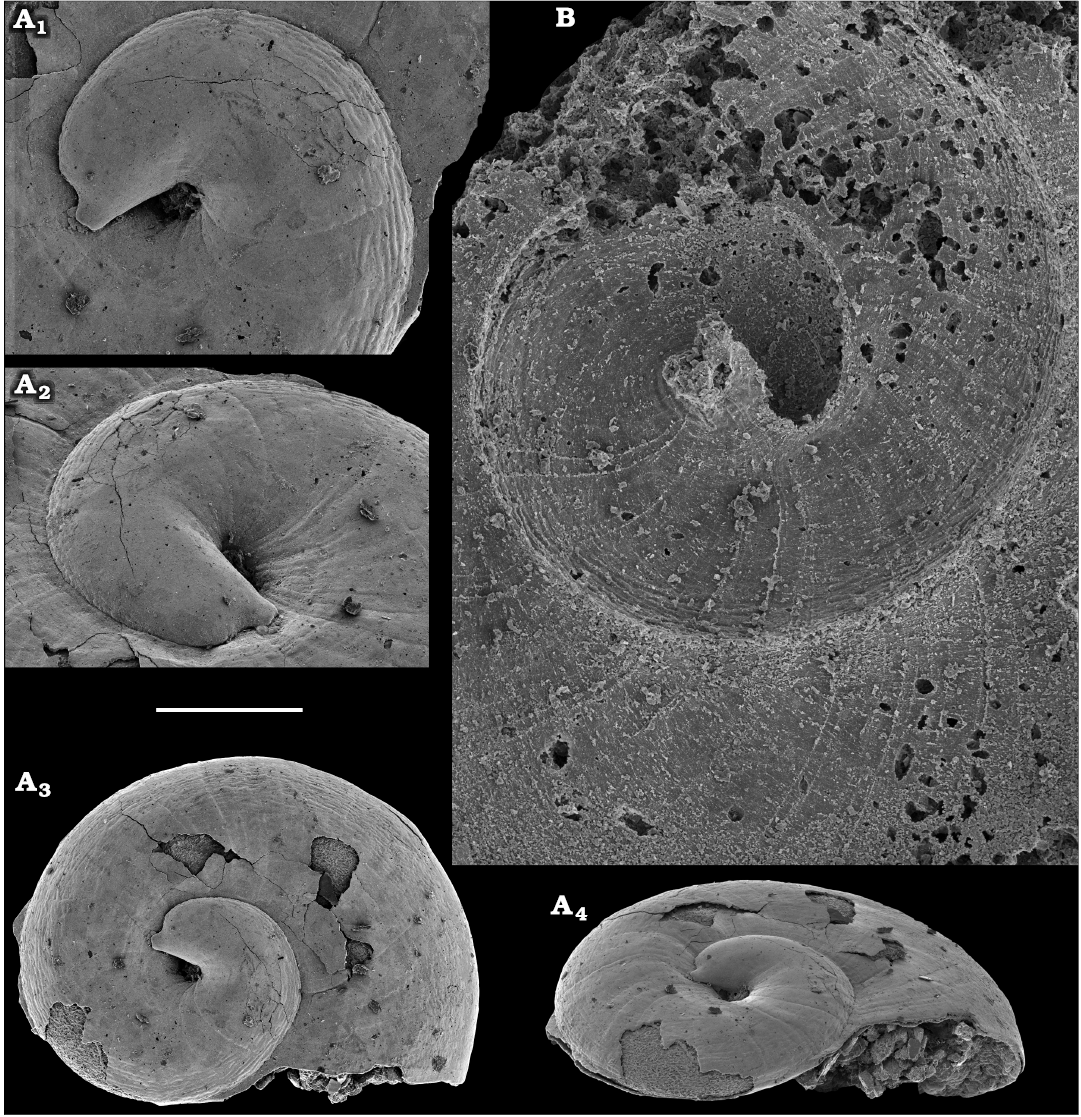
Fig. 29. Pelagiellid mollusc Aldanella sibirica Parkhaev and Karlova, 2011, from the lower Cambrian Tyuser Formation, lower reaches of the Lena River (A) and Erkeket Formation, Khorbusuonka River (B), Siberia, Russia; samples 21/21 (A) and 11/16.1 (B). A. Phosphatised shell, SMNH Mo194736. A1, A2, enlargements showing protoconch; A3, adapical view; A4, oblique adapical view. B. External mould, SMNH Mo194737, with sculpture of the shell surface. Scale bar: 250 µm (A3, A4), 125 µm (A1, A2, B).
Remarks.—In the holotype, the spiral ornament cover the entire whorl surface excluding protoconch (Parkhaev and Karlova 2011: pl. 11: 9). In the specimens described herein, spiral plication is available near the sutural lines of the whorls and is probably eroded on the rest of the shell surface. The spiral ornament probably represents phosphatised periostracum, which is rarely preserved. A polygonal texture, with units having variable relief and 20–40 μm in diameter, present on the surface of the first whorl in some internal moulds of Aldanella (Fig. 28B) cannot therefore derive from periostracal texture, nor from a prismatic microstructure in the ca. 5 μm thick wall (see also Kouchinsky et al. 2017: figs. 18A, B, 20A, F, 23H).
Stratigraphic and geographic range.—Tommotian stage (correlated with the upper part of Cambrian Stage 2), Siberia, Russia.
Family Pelagiellidae Knight, 1956
Remarks.—Based on observation of chetae in Pelagiella from the Kinzers Formation (Thomas et al. 2020), Landing et al. (2021a) concluded that pelagiellids, except for Pelagiella atlantoides, probably belong to the order Sabellida Levinsen, 1883, of the class Polychaeta Grube, 1850. Sabellids appeared, however, by the end of the Permian, while almost all Palaeozoic tubeworms are probable lophophorates judging by their tube microstructure (Ippolitov et al. 2014). Pelagiella has a more typical molluscan shell microstructure (Li et al. 2017), morphology of the early shell and protoconch (e.g., Kouchinsky et al. 2011: figs. 8–10), and invariability in the degree of coiling (Vinn and Mutvei 2009; Ippolitov et al. 2014), as would not be expected in annelid tubeworms. Moreover, chitin synthesis is maintained by the same genetic machinery in molluscs as that in annelids and brachiopods (e.g., Zakrzewski et al. 2014; Schiemann et al. 2017). Thus, the original interpretation of pelagiellids as molluscs is in better agreement with the entire complex of features being observed (Thomas et al. 2020).
Genus Pelagiella Matthew, 1895
Type species: Cyrtolites atlantoides Matthew, 1894; Lower Cambrian; New Brunswick, Canada.
Pelagiella sp. 1
Fig. 30A, D.
Material.—Two calcium phosphatic internal moulds, SMNH Mo194738 and 194739, from sample 19/12.75. Erkeket Formation, Khorbusuonka River, Siberia, Russia. Lower Botoman stage (correlated with the Cambrian Stage 4).
Description.—Shell represented by smooth very rapidly expanding internal moulds consisting of up to 3/4 whorl without preserved aperture. Peripheral surface of the whorl rounded and strongly extended distally, separated from the spiral and umbilical surfaces by an angle. Spiral surface concave, umbilical surface convex. Cross-section of the whorl extended drop-shaped.

Fig. 30. Pelagiellid and khairkhaniid molluscs from the lower Cambrian Erkeket Formation, Khorbusuonka River (A, B, D, E, G, H), Sekten Formation, lower reaches of the Lena River (C, F, I) formations and Kessyusa Group, Khorbusuonka River (J), Siberia, Russia; samples 19/12.75 (A, D, G, H), 19/29 (B), 19/40 (E), 22/50 (C, F, I), and 15/23 (J). A, D. Pelagiella sp. 1, internal moulds, SMNH Mo194738 and 194739, respectively. B, C, E–H. Pelagiella spp., internal moulds, SMNH Mo194740–194745, respectively. E1, apertural view; E2, adapical view. I. Phosphatised wall of Pelagiella sp. cf. Cambretina mareki Horný, 1964, SMNH Mo194746. J. Nomgoliella sinistrovolubilis Missarzhevsky, 1981, internal mould, SMNH Mo194747. J1, apertural view; J2, adapical view. Scale bar: 1 mm (J) and 500 µm (A–I).
Remarks.—Pelagiella sp. 1 described herein is very similar to Pelagiella sp. 1 from the upper Emyaksin Formation (Kouchinsky et al. 2015a: fig. 19A) and is different from Pelagiella spp. below in a strongly extended peripheral part of the whorl and depressed spiral side. External shell sculpture not preserved. Internal moulds are most similar to Pelagiella madianensis (Zhou and Xiao, 1984) from Cambrian Series 2 of Gondwana. Ornamentation of P. madianensis consists of granules arranged in spiral rows (Parkhaev in Gravestock et al. 2001; Li et al. 2017, 2019), whereas the ornamentation of probably the same species, Pelagiella sp. 1 (see Kouchinsky et al. 2015a: fig. 18) consists of commarginal ribs.
Pelagiella spp.
Fig. 30B, C, E–H.
Material.—Several tens of calcium phosphatic internal moulds, including figured SMNH Mo 194740–194745, from samples 19/12.75, 19/29, 19/40, 20/1B, 20/6B, Erkeket Formation, Khorbusuonka River, Botoman–Toyonian stages (correlated with the Cambrian Stage 4) and several calcium phosphatic internal moulds from sample 21/57.75, Tyuser Formation, Delgadella anabara–Nevadella Zone, Atdabanian stage (correlated with the Cambrian Stage 3). Several calcium phosphatic internal moulds from samples 22/50, 22/55.5, and 22/67, Sekten Formation, Lena River, uppermost Toyonian and Amgan stages (correlated with the Cambrian Stage 4 and lowermost Wuliuan Stage). Siberia, Russia.
Remarks.—Smooth rapidly expanding internal moulds consisting of up to one whorl. Peripheral surface of the whorl rounded, passing into convex spiral and umbilical surfaces. Cross-section of the whorl rounded triangular to ovaliform. The moulds probably belong to several species. They are similar to internal moulds described from the Emyaksin Formation (Kouchinsky et al. 2015a: figs. 19B, C, 20). In the latter forms, the outer shell surface bears transversal ribs also described from Costipelagiella nevadense Skovsted, 2006b. A phosphatised wall fragment found in sample 22/50 has a cancellate ornament identical to that described from Cambretina mareki Horný, 1964 (see Kouchinsky et al. 2011) and consists of rounded 30–40 μm wide polygons. It is not however clear if the fragment belongs to the same species as internal moulds and if this type of external surface ornamentation is unique to Cambretina mareki.
Class and order uncertain
Family Khairkhaniidae Missarzhevsky, 1989
Genus Nomgoliella Missarzhevsky, 1981
Type species: Nomgoliella sinistrovolubilis Missarzhevsky, 1981; Beds with Anabarella plana, Cambrian Stage 2; Khasagt-Khajrkhan-Uul Ridge, Salaany-Gol Rivulet, Western Mongolia.
Nomgoliella sinistrovolubilis Missarzhevsky, 1981
Fig. 30J.
Material.—Single internal mould, SMNH Mo194747, from sample 15/23, Chuskuna Formation, upper Kessyusa Group, Khorbusuonka River. Siberia, Russia. Lower part of Cambrian Stage 2.
Description.—Sinistrally coiled low-spired internal mould, ca. 2 mm in diameter, consisting of a single rapidly expanding whorl with broad oval cross-section. Initial part in contact with the expanded whorl. Dorsal side of the internal mould representing ca. 1/2 revolution of the whorl is somewhat flattened.
Remarks.—Dorsally flattened internal mould of a single specimen described herein can be interpreted as preservational difference from the Mongolian forms. Barskovia Golubev, 1976, and Nekolenia Vasiljeva, 1998 (probably synonymous after Parkhaev 2002: 34) are different in being loosely coiled and having almost circular cross-section of the whorl, although B. hemisymmetrica (Kouchinsky et al. 2017: fig. 23) may show broadly oval outlines.
Stratigraphic and geographic range.—Cambrian Stage 2 of the Siberian Platform and western Mongolia.
Problematic shell 1
Fig. 31.
Material.—Single calcium phosphatic internal mould, SMNH X11035, from sample 19/40. Erkeket Formation, Khorbusuonka River, Siberia, Russia. Toyonian stage (correlated with the Cambrian Stage 4).
Description.—Bilaterally symmetrical rapidly expanding slightly cyrtoconic shell, ca. 0.8 mm wide and long, and ca. 0.6 mm tall, with a broadly oval aperture. Subapical part flattened. Mould covered with radial striae and pits.

Fig. 31. Problematic shell 1, from the lower Cambrian Erkeket Formation, Khorbusuonka River, Siberia, Russia; sample 19/40. Internal mould with pitted surface, SMNH X11035; A1, upper; A2, sub-apical; A3, and A4, lateral views. Scale bar 250 µm.
Remarks.—Generally similar fossils described from the lower Cambrian were attributed to the Tianzhushanellida Conway Morris in Bengtson et al., 1990. Those are often interpreted as stem-group brachiopods with calcareous shells, probably originally bivalved, although no firm evidence exists (Liu 1979; Bengtson et al. 1990; Esakova and Zhegallo 1996; Parkhaev in Gravestock et al. 2001; Li et al. 2014; Kouchinsky et al. 2015a). They are mainly different from the problematic shell herein in having two bilaterally symmetrical umbonal cavities separated by a medial septum.
Problematic shell 2
Fig. 32.
Material.—Single fragment of shell, SMNH X11042, from samples 19/10.25, and four fragments, SMNH X11039–11041, 11043, from 19/11.75. Erkeket Formation, Khorbusuonka River, Siberia, Russia. Lower Botoman stage (correlated with the lower part of Cambrian Stage 4).
Description.—Arcuate fragments of shell showing accretionary growth, with continuous prominent parallel ridges curved towards the shell margin. Spaces between ridges are perpendicularly striated. Several aligned prominent spines inclined in the direction of curvature of the growth ridges, increase in size in the same direction. The spines are somewhat flattened with their thick bases elongated parallel to the growth ridges. Each spine situated in the depression between two growth ridges and carries a central canal.

Fig. 32. Problematic shell 2, from the lower Cambrian Erkeket Formation, Khorbusuonka River, Siberia, Russia; samples 19/11.75 (A–C, E) and 19/10.25 (D). A–E. Phosphatised fragments of shells with spines, SMNH X11039–11043, respectively. A1, D1, plain view; A2, lateral view of the shell fragment with cyanobacterial filaments (SMNH X11337, see Fig. 69A) attached to its inner side (bottom); A3, close-up showing spines; D2, lateral view. C, E, fragments of presumably subapical part of the shell. E1, E2, view on the same fragment, ca 90º rotated around the longitudinal axis. Scale bar: 250 µm (C, E), 165 µm (A1, A2, B1, D), and 50 µm (A3, B2).
Remarks.—It is not clear if the central canal it opened at the tip of the spines, because tips are broken off. The fragments resemble Paracarinahites spp. The latter representing arcuate imbricated plates, each carrying a spine and fused in an elongated curved structure. Another similarity exists with spiny subapical zones of Ocruranus–Eohalobia group sclerites described by Siegmund (1997). Both forms can likely be attributed to multiplated molluscs.
Order Sachitida He in Yin et al., 1980
Family Halkieriidae Poulsen, 1967
Genus Halkieria Poulsen, 1967
Type species: Halkieria obliqua Poulsen, 1967; Læså Formation; lower Cambrian; Bornholm Island, Denmark.
Halkieria proboscidea (Meshkova, 1969)
Fig. 33.
Material.—Several tens of phosphatised sclerites and calcium phosphatic internal moulds, including figured SMNH X11044–11059, from samples 11/2B, 11/16.05, 11/16.1, 11/16.5, basal Erkeket Formation, Khorbusuonka River, and samples 21/21 and 21/22, Tyuser Formation, Lena River. Siberia, Russia; Dokidocyathus regularis Zone, Tommotian stage (correlated with the upper part of Cambrian Stage 2).
Description.—Cultrate sclerites, 2.5–3.5 times longer than wide, with a broadly triangular cross-section, void axial inner part (lumen) and coalescent lateral zones. Lumen represented by internal moulds, 2–3 times narrower than the blade of sclerite, rounded at the base of sclerite and becoming lenticular towards the apex. Lateral zones house blind channels running parallel to each other and perpendicularly to the respective sclerite margin. Sclerites are slightly sigmoidally curved in lateral view, with dorsally directed distal end and can be slightly helically twisted. Dorsal surface divided by a well-defined median ridge into two almost bilaterally symmetrical sides with 3–4 less prominent parallel longitudinal ridges each, as well as faint striation subperpendicular to the longitudinal sculpture. Basal part rounded triangular in the plan view, extended and deflected ca. 90° with respect to the blade of sclerite. Oval foramen corresponds to ca. 1/4 of the basal part width. Foramen is delimited by a concentric buttress and situated in the middle of basal part.
Palmate sclerites are 0.6–2.0 mm long and deflected ca. 90° at the basal part. They are slightly twisted, have a narrow lenticular cross-section and a pointed apex. A slit-like foramen is situated at the basal end of sclerite delimited by a concentric buttress. Rough and frequent transversal ridges of growth increments are situated on the deflected basal part of sclerite. Blade is slightly arcuate longitudinally, carries a medial low ridge and several faint parallel ridges on the dorsal surface. The surface of the sclerite is covered with transversal growth lines and flattened tubercles.

Fig. 33. Sclerites of sachitid Halkieria proboscidea (Meshkova, 1969), from the lower Cambrian Erkeket Formation, Khorbusuonka River, Siberia, Russia; samples 11/16.05 (A, C, D, H, J, K, M, N), 11/2B (B, E, F, I, L, O), 11/16.1 (G). A–K. Cultrate sclerites, SMNH X11044–11054, respectively. A1, oblique lateral view; A2, oblique view on the upper surface of sclerite; A3, oblique view on the lower surface of sclerite; B1, C1, K2, views on the lower surface of sclerites; B2, C2, K1, lateral views. F, close-up of the sclerite and its internal mould. G, internal mould. L–O. Palmate sclerites, SMNH X11056–11059, respectively. Scale bar: 1 mm (A2, A3, B, F, G, I), 500 μm (A1, C, D, E, J, K, N, O), and 250 μm (H, L, M).
Remarks.—Apparently conspecific cultrate sclerites were described by Kouchinsky et al. (2017) as Halkieria sp. B, represented by narrow internal moulds from the lower Emyaksin Formation on the eastern flanks of the Anabar Uplift in Siberia. Palmates of the species are probably represented by Halkieria sp. type C (Kouchinsky et al. 2017). Similar cultrate and palmate forms are described by Parkhaev and Demidenko (2010) and Devaere et al. (2013) from the Cambrian Stage 2 of South China and France, respectively, where they are attributed to Halkieria sacciformis (Meshkova, 1969). Herein we infer that Sachites proboscideus Meshkova, 1969, represents internal moulds (lumen) of cultrate sclerites (Meshkova 1974: pl. 25: 1, 2) and Sachites sacciformis represents palmates and cultrates of the same species. Meshkova (1969) first described both forms in the same publication from the Tommotian stage of Siberia. The system of canals in sclerites of Halkieria proboscidea and other sachitids is interpreted to be homologous with the aestetes of the Aculifera (Bengtson 1992; Vinther and Nielsen 2005; Vinther 2009; Peel 2020).
Stratigraphic and geographic range.—Cambrian Stage 2, Siberia, South China, and France.
Phylum and class uncertain
Order Cambroclavida Conway Morris and Chen, 1991
Family Zhijinitidae Qian, 1978
Genus Cambroclavus Mambetov in Mambetov and Repina, 1979
Type species: Cambroclavus antis Mambetov in Mambetov and Repina, 1979; Rhombocorniculum cancellatum Zone, upper part of Cambrian Stage 3; Beshtash Formation, Beshtash Creek, Talass Alatau Range, Kazakhstan.
Cambroclavus sp.
Fig. 34A–J.
Material.—Single phosphatised sclerite from sample 19/5.5 and 15 phosphatic internal moulds and partial phosphatic casts of the external surface of sclerites, including figured SMNH X11060–11069, from samples 19/10.25, 19/11.75, 20/1B, Erkeket Formation, Khorbusuonka River, Siberia, Russia. Lower Botoman stage (correlated with the lower part of Cambrian Stage 4).
Description.—Isolated sclerites, 0.3–0.7 mm in length, representing elongated basal shields with growth increments on the surface and imperfect bilateral symmetry. They are slightly sigmoidally curved in the plan view and flattened. Lateral margins subparallel, occasionally with a variably developed constriction. Dorsal side distinguished by a rounded elevated facet situated on its presumably anterior part. Facet delimited by a circular rim and anteriorly inclined at ca. 45º towards the longitudinal axis of sclerite. The facet representing attachment site of a spine carries a single almost circular hole, 30–60 μm in diameter. Posterior part of the shield elongated with blunt termination. A broad median ridge is present on the dorsal surface in some specimens. Ventral (lower) surface of the shield smooth. Wall of the basal shield composed of aciculate elements radiating from attachment site of the spine, perpendicularly to growth increments of the shield. The aciculate elements are longitudinally oriented in the spine.
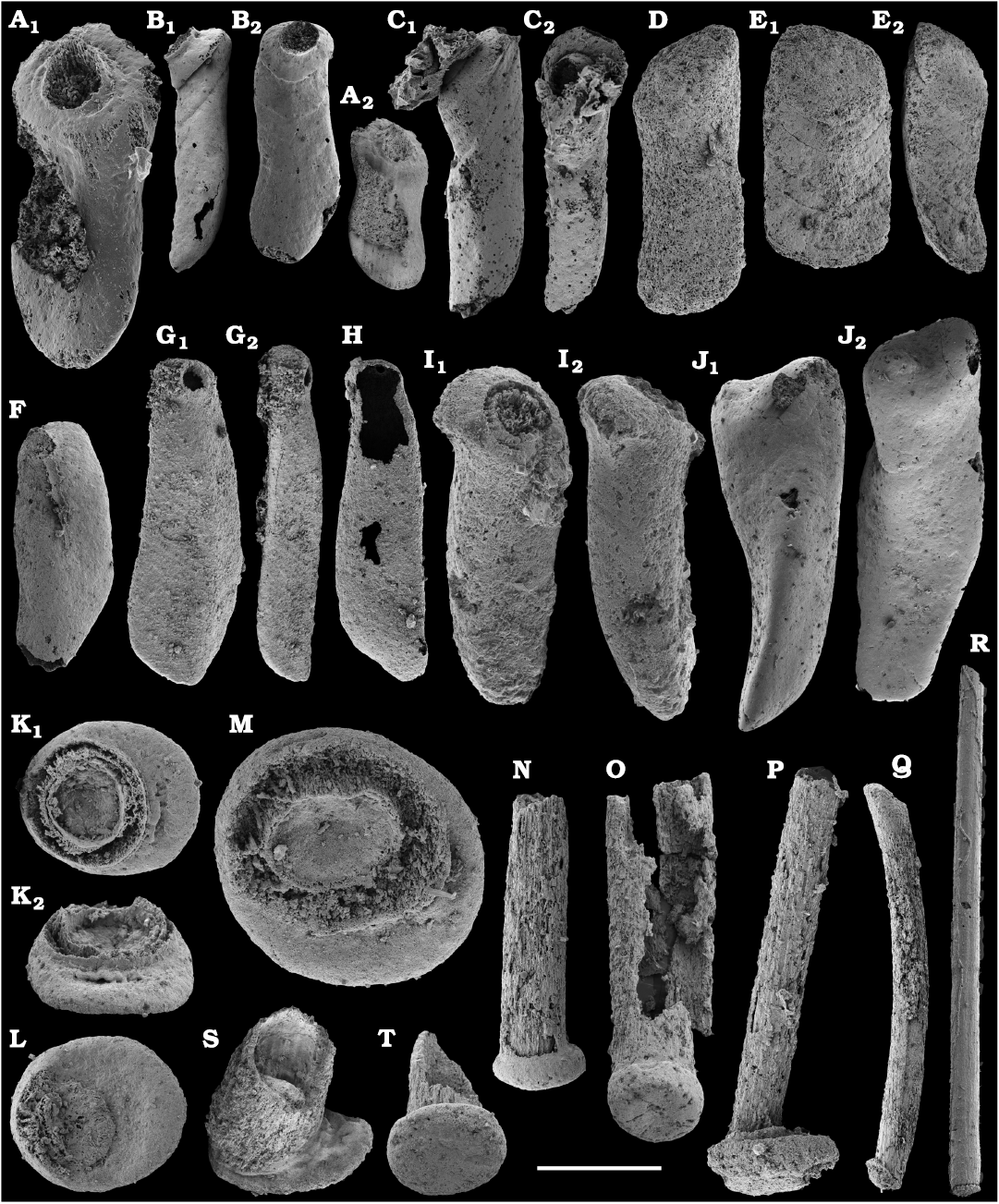
Fig. 34. Sclerites of cambroclavids from the lower Cambrian Erkeket Formation, Khorbusuonka River, Siberia, Russia; samples 19/5.5 (I), 19/10.25 (A–E, J), 19/11.75 (K–T), and 20/1B (F–H). A–J. Cambroclavus sp., SMNH X11060–11069, respectively. A1, A2, B2, C2, E1, G1, I1, J2, view on the upper surface of sclerites; B1, C1, E2, G2, I2, J1, lateral view. K–T. Zhijinites clavus (Mambetov in Mambetov and Repina, 1979), SMNH X11070–11079, respectively. K1, view on the upper surface of sclerite; K2, lateral view. Scale bar: 500 µm (Q), 250 µm (A2, B–H, J–L, N, O, Q–T), 125 µm (A1, I, M, P).
Comparison.—Given morphological variation within the same fossil assemblage of sclerites, it is premature to describe a new species with the material currently available, until more specimens representative of the entire morphological variation will be accumulated. It seems the Siberian forms are different from Cambroclavus absonus Conway Morris in Bengtson et al., 1990, in the absence of flaring of the posterior part of the basal shield which has in C. absonus a distinct hourglass-like shape. Prong-like extensions available in C. bicornis (Qian and Xiao, 1984) have not been found either. The type species, Cambroclavus antis exhibit prominent longitudinal ridges on the upper and lower surfaces of the basal shield (Mambetov and Repina 1979). It was also noted that inflated sclerites can be minor components of Cambroclavus scleritomes (Duan 1984; Qian and Xiao 1984; Wotte 2009; Na and Li 2011).
Remarks.—Cambroclavids are documented herein from the Siberian Platform. Cambroclavid sclerites were presumably calcareous with a fibrous composition of the wall, closely and regularly spaced on the available fragments of their articulated arrays (Bengtson et al. 1990; Conway Morris and Chen 1991).
The nature of cambroclavid sclerite-bearing animals is yet poorly known. A possible cambroclavid scleritome described by Conway Morris and Peel (2010) from Sirius Passet in Greenland represents a compression of an externally segmented worm-like body. The latter is mainly covered with rows of cambroclavid-type sclerites and presumably zhijinid-type sclerites on its only preserved (anterior?) end. The transition between these two zones is abrupt, which agrees with presence of distinct and often co-occurring cambroclavid and zhinzhinid morphs of sclerites in the small shelly fossil assemblages.
Right- and left-handed sclerites cannot be distinguished (but Qian et al. 2000), which rather agrees with a supposed worm-like body rounded in cross-section, rather than with a flattened slug-like animal with externally well-defined left and right sides. Laterally alternating longitudinal rows of diagenetically phosphatised imbricated sclerites are rarely preserved articulated, as reported from Deltaclavus Conway Morris and Chen, 1991 (text-figs 8, 9, 11 therein). Similarly preserved fossils reported by Brock and Cooper (1993: fig. 11) and Zhang et al. (2021) may represent folded parts of cambroclavid scleritomes consisting of Deltaclavus-like sclerites lacking spines.
Genus Zhijinites Qian, 1978
Type species: Zhijinites longistriatus Qian, 1978; Meishucunian stage; Yunnan, Sichuan, and Guizhou provinces, South China.
Zhijinites clavus (Mambetov in Mambetov and Repina, 1979)
Fig. 34K–T.
1979 Cambroclavus clavus Mambetov sp. nov.: Mambetov and Repina 1979: 122–123, pl. 13: 3, 5, 7–9.
1981 Cambroclavus clavus Mambetov in Mambetov and Repina, 1979; Missarzhevsky and Mambetov 1981: 71, pl. 9: 1, 2, 8.
Material.—Over 20 phosphatic internal moulds, phosphatised sclerites and their partial phosphatic casts, including figured SMNH X11070–11079, from samples 19/10.25, 19/11.75, 20/1B, 20/4B. Erkeket Formation, Khorbusuonka River, Siberia, Russia. Botoman stage (correlated with the Cambrian Stage 4).
Description.—Disarticulated nail-shaped sclerites consisting of a circular or oval tablet-like basal shield and relatively thick and long shaft, straight or slightly curved in one plane. The shaft is circular in cross-section. Basal shield with a slightly concave upper (dorsal) surface bearing an articulation facet for the shaft. The latter has a circular attachment site adpressed into the upper surface of basal shield. Walls relatively thick delimiting hollow space inside the spine. Two concentric rims delimit inner and outer surfaces of a relatively thick wall that often preserves acicular units. The units are elongated longitudinally, subparallel to the surface of the shaft. The latter can reach at least 1.6 mm in length in incompletely preserved specimens. The shaft is inclined at ca. 80º towards the upper surface of the basal shield. Its tip is not preserved. The external surface was probably longitudinally striated or smooth. Articulation facet in the anterior part occupies more than half of the length of the basal shield. Its posterior part is rounded and smooth. The lower (ventral) surface of the shield is smooth.
Remarks.—Similar to Parazhijinites guizhouensis Qian and Yin, 1984, and material illustrated by Conway Morris et al. (1997: 179, pl. 3) as P. cf. guizhouensis, but the latter has a radial or nodular ornamentation, angle of inclination of the shaft to the basal disk from 55° (where the base is broader) to 90° (in association with a reduced base). There are neither transversally compressed spines in our material, nor longitudinal ribbing, nor a median ridge.
Stratigraphic and geographic range.—Rhombocorniculum cancellatum and Microcornus parvulus zones, Kazakhstan and lower Botoman stage (correlated with the Cambrian Stage 4), Siberia, Russia.
Phylum and class uncertain
Order Chancelloriida Walcott, 1920
Family Chancelloriidae Walcott, 1920
Genus Stellispinella Vasiljeva in Vasiljeva and Sayutina, 1993
Type species: Chancelloria lenaica Zhuravleva and Korde, 1955; Tommotian Stage; Siberian Platform, Russia.
Stellispinella lenaica (Zhuravleva and Korde, 1955)
Fig. 35.
1955 Chancelloria lenaica Zhuravleva and Korde sp. nov.; Zhuravleva and Korde 1955: 476–477, text-fig. 1и–у (Russian).
1955 Chancelloria aldanica Zhuravleva and Korde sp. nov.; Zhuravleva and Korde 1955: 476, text-fig. 1ф–ю (Russian).
1985 Stellaria lenaica (Zhuravleva and Korde, 1955); Vasiljeva 1985: 121, pl. 44: 7, 8.
1985 Stellaria aldanica (Zhuravleva and Korde, 1955): Vasiljeva 1985: 121–122, text-fig. 2, pl. 44: 4, 5.
1989 Ginospina araniformis Missarzhevsky sp. nov.: Missarzhevsky 1989: 205–206, pl. 21: 2 and pl. 22: 10, 13.
1998 Stellispinella lenaica (Zhuravleva and Korde, 1955): Vasiljeva 1998: 100, pl. 19: 10, 11, 13.
1998 Stellispinella aldanica (Zhuravleva and Korde, 1955): Vasiljeva 1998: 100–101, pl. 19: 3, 8, 12, 14.
Material.—Over a hundred phosphatised articulated sclerites, their fragments and phosphatic coatings, including figured SMNH X11081–11093, from samples 11/2B, 11/16.05,11/16.1, basal Erkeket Formation, Khorbusuonka River, and samples 21/21 and 21/22, Tyuser Formation, Lena River. Siberia, Russia; Dokidocyathus regularis Zone, Tommotian stage (correlated with the upper part of Cambrian Stage 2).
Description.—Articulated sclerites consist of 9–12 lateral rays curved up to 30º away from the plane of the basal facet and a central disk, which is a subdued central ray. Surface covered with scattered granules or short rods, ca. 10 µm in diameter. Lateral rays taper from massive bases. Base of the central disk is up to twice larger in diameter than bases of lateral rays. Basal facet of each ray flattened, with a central circular foramen.

Fig. 35. Sclerites of chancelloriids from the lower Cambrian Erkeket Formation, Khorbusuonka River (F–I) and Tyuser Formation, lower reaches of the Lena River (A–E, J–L), Siberia, Russia; samples 11/16.1 (F), 11/2B (G), 11/16.05 (H, I), and 21/21 (A–E, J–M). A–D, F–H, K. Stellispinella lenaica (Zhuravleva and Korde, 1955), sclerites with a central disk, SMNH X11081–11088, respectively. A1, plain view; A2, oblique lateral view. E, I, J. Stellispinella lenaica (Zhuravleva and Korde, 1955), sclerites without central disk, SMNH X11089–11091, respectively. I1, lateral view; I2, plain view. L, M. Diplospinella zintchenkoi Vasiljeva in Vasiljeva and Sayutina, 1988, SMNH X11092 and X11093, respectively. L1, L2, lateral view. Scale bar 500 µm.
Remarks.—Central disk present in this species is not known from any other species and genera of chancelloriids (see discussion on synonymies below). The type species of Stellispinella was not well-defined or described by Zhuravleva and Korde (1955). Chancelloria lenaica was redescribed by Vasiljeva (1985), who defined its neotype and selected it as a type species of a new genus Stellaria Vasiljeva, 1985. The latter proved to be preoccupied and renamed Stellispinella Vasiljeva in Vasiljeva and Sayutina, 1993. The main distinction between two available and simultaneously described species, Chancelloria lenaica Zhuravleva and Korde, 1955, and Chancelloria aldanica Zhuravleva and Korde, 1955, is that their articulated sclerites are, respectively, rounded in the plan view with 10–12 lateral rays versus ovaliform with 7–10 lateral rays (Vasiljeva 1985). Otherwise similar in morphology, size and granular ornamentation, both forms co-occur in the material from the Tommotian stage of the Siberian Platform and can be attributed herein to the same species, Stellispinella lenaica (Zhuravleva and Korde, 1955) as well as Ginospina araniformis Missarzhevsky, 1989 (see Vasiljeva 1998).
Several other types of co-occurring sclerites without central discs may represent parts of the same scleritome. Those consist of 5 to 8 lateral rays connected with each other without centrally differentiated unit (Fig. 35E, I, J). Frequently occurring disarticulated and partially articulated sclerites of those types attributed herein to Stellispinella lenaica are available from the basal beds of the Erkeket and the middle Tyuser formations.
Such other sclerites, which may also be parts of the same scleritome are known to co-occur with Stellispinella lenaica in the Ulakhan-Ald’yarkhay section are represented by Diplospinella zintchenkoi Vasiljeva in Vasiljeva and Sayutina, 1988. The latter is the type and only species of Diplospinella Vasiljeva in Vasiljeva and Sayutina, 1988, with its younger synonym, Sissospina irregularis Missarzhevsky, 1989. There are two such sclerites from the type locality, from sample 21/21 (Fig. 35L, M). They have straight basal facet, 1–2 relatively massive long gently curved rays and 3–5 intermediate rays subperpendicularly situated with respect to the basal facet of sclerite. Much smaller and shorter rays are attached to the base of sclerite. These forms are known from the Tommotian stage of the Siberian Platform.
Stratigraphic and geographic range.—Tommotian stage (correlated with the upper part of Cambrian Stage 2), Siberia, Russia.
Genus Chancelloria Walcott, 1920
Type species: Chancelloria eros Walcott, 1920; middle Cambrian; Burgess Shale, British Columbia, Canada.
Chancelloria spp.
Fig. 36.
Material.—Thousands of disarticulated sclerites from many samples (SOM 3), including figured specimens, SMNH X11094–11105, from samples 11/2B, 11/16.1, 19/11.75, 21/21, 21/51, and 21/54; ranging in age from the Tommotian stage (correlated with the upper part of Cambrian Stage 2) to the Toyonian stage (Cambrian Stage 4), in the Khorbusuonka River and Ulakhan-Ald’yarkhay sections, Siberia, Russia.
Remarks.—Articulated sclerites from the lower Tommotian stage D. regularis Zone, from the basal Erkeket Formation and from basal part of the Middle Member of the Tyuser Formation are represented by rare 7+1, 8+1, and 9+1 sclerites and a single 11+1 sclerite (SOM 3). The majority of sclerites from Cambrian Stage 3–lower Stage 4 have nine lateral rays with one central ray (9+1) and five rays without a central ray (5+0). Sclerites composed of six, seven or ten lateral rays and a central ray (6+1, 7+1, and 10+1) are relatively rare. Chancelloria spp. sclerites have lateral rays gently curved away from the plane of the basal facet (central ray incompletely preserved; Fig. 36A–D) and there is no firm evidence to support the presence of a particular number of species. Chancelloria sclerites have phosphatised walls composed of longitudinally oriented along the surface aciculate laths (Fig. 36P–S), similar to those described from chancelloriids previously (Kouchinsky 2000; Kouchinsky et al. 2015a: fig. 34A).
A single sclerite (Fig. 36E) with three rays bent away from the basal facet (3+0 type) may represent a species of Allonnia Doré and Reid, 1965 (Bengtson et al. 1990), which has not been previously reported from the Tommotian strata of the Siberian Platform. Single sclerites with three rays also attributable to Allonnia sp. occur in samples from the upper part of Cambrian Stage 3 (21/54) and lower Mayan Stage (19/5B) (Fig. 36N, O). Diameter of the basal facet in each ray of Allonnia sp. (?) from the Tommotian stage is about half of that in younger forms.
Several tens of articulated sclerites from samples 19/11.75 and 19/12.75 have four or five rays with one of the rays bent outwards. Those can be attributed to Archiasterella Sdzuy, 1969. Archiasterella-type sclerites occur in a relatively few samples from Cambrian Stage 3 compared to Chancelloria spp. and some of them may correspond to the 4+0 sclerites known from Chancelloria. In samples 19/11.75 and 19/12.75, the 7+1 and 9+1-type sclerites of Chancelloria spp. are found in association with the Archiasterella sclerites.
Genus Archaeopetasus Conway Morris and Bengtson in Bengtson et al., 1990
Type species: Archaeopetasus excavatus Conway Morris and Bengtson in Bengtson et al., 1990; Cambrian Stage 3; South Australia.
Archaeopetasus pachybasalis Kouchinsky sp. nov.
Figs. 37, 38.
Zoobank LSID: urn:lsid:zoobank.org:act:FCA618F0-6F22-4EEA-8E 16-9BBEE18FDE6B
Etymology: From the Greek pachys, thick and Latin basalis, having a pedestal; for the thick basal part of sclerite.
Type material: Phosphatised disarticulated sclerites: holotype SMNH X11113 (Fig. 37) and paratypes SMNH X11114‒11125 (Fig. 38), from the type locality and horizon.
Type locality: locality 21/21 (71°35’N, 127°21’E) on the right bank of the Lena River, ca. 500 m upstream from the mouth of the Ulakhan-Ald’yarkhay Brook, northern Siberia, Russia.
Type horizon: Basal Middle Member of the Tyuser Formation, 21–22 m from the base of the the Tyuser Formation, Dokidocyathus regularis Zone, Tommotian stage, correlated herein with the upper part of Cambrian Stage 2.
Material.—Several hundred phosphatised sclerites and phosphatic coatings, including type material and figured SMNH X11126–11129, from samples 21/21 and 21/22, Tyuser Formation, Lena River, Siberia, Russia. Dokidocyathus regularis Zone, Tommotian stage.
Diagnosis.—Broadly flaring smooth or tuberculated conical sclerite having undulating brim and thick protruding basal part, which is folded and funnelling towards the middle.
Description.—Sclerites, 0.5–1.5 mm in diameter, having funnel- or hat-shaped smooth upper surface and doughnut- or flower-shaped plicated basal part. The upper surface narrows abruptly into a single centrally situated tubular extension with a pointed tip. The upper surface is covered with flattened tubercles which tend to be elongated and aligned along the undulating margin of sclerite. Fragmentarily preserved spines are 0.3–0.6 mm in diameter and 0.5–1.5 mm long. They are also covered concentrically with elongated tubercles. Inner surface of spines is covered in places with tightly located flattened tubercles, ca. 10 μm in diameter. Basal part subdivided by several irregular but generally radially directed folds into several partially separated sectors funnelling towards the center of the lower side of sclerite. There is a circular deep pit in the middle of the lower side leading to a channel, ca. 0.3 mm in diameter inside the tubular extension of the upper surface of sclerite, thus forming inner layer of the tube. Interspace between phosphatic coatings of the opposite sides of sclerite often preserve aciculated elements oriented subperpendicular to the inner surface of phosphatic coat/wall.

Fig. 36. Sclerites of chancelloriids from the lower Cambrian Erkeket Formation, Khorbusuonka River (A, K, N, P–S) and Tyuser Formation, lower reaches of the Lena River (B–J, L, M, O), Siberia, Russia; samples 11/16.1 (A), 19/5B (N), 19/11.75 (P–S), 19/12.75 (K), 21/21 (B–H), 21/51 (I, J), 21/54 (L, M, O). A–D, P, R, S. Chancelloria spp. with a central ray, SMNH X11094–11100, respectively. F–H, M, Q. Chancelloria spp. without a central ray, SMNH X11101–11105, respectively. E, N, O. Allonnia sp., SMNH X11106–11108, respectively. I–L. Archiasterella sp., SMNH X11109–11112, respectively. Scale bar: 500 µm (A–O, Q) and 250 µm (P, R, S).

Fig. 37. Sclerites of chancelloriid Archaeopetasus pachybasalis Kouchinsky sp. nov., from the lower Cambrian Tyuser Formation, lower reaches of the Lena River, Siberia, Russia; sample 21/21. Holotype specimen, SMNH X11113. A1, upper view; A2, A3, lateral view; A4, lower surfaces of sclerite. Scale bar 500 µm.

Fig. 38. Sclerites of chancelloriid Archaeopetasus pachybasalis Kouchinsky sp. nov., from the lower Cambrian Tyuser Formation, lower reaches of the Lena River, Siberia, Russia; sample 21/21. A–H, J–L, P. Basal part of phosphatized sclerites, SMNH X11114–11125, respectively. A1, upper surface; A2, J2, oblique view on the upper surface; J1, L1, lateral view; C1, F1, lower surface; C2, F2, L2, oblique view on the lower surface of sclerites. I, M–O. Apical spines detached from the basal part of sclerites, SMNH X11126–11129, respectively. Scale bar 500 µm.
Remarks.—Archaeopetasus excavatus Conway Morris and Bengtson in Bengtson et al., 1990, different in having isometrical granules on the external surface of sclerite, cone-shaped central spine, and relatively thin basal part. Archaeopetasus typicus (Sipin, 1999) has a central boss-like elevation which never extends as a spine and a well-defined central pit with granulated surface on the lower smooth surface of sclerite.
Similar fossils from the Tommotian stage of Aldan and Lena rivers were figured by Missarzhevsky in Rozanov et al. (1969: pl. 7: 3, 15, 16) and Missarzhevsky (1989: pl. 24: 12) and tentatively interpreted as opercula of Coleolella billingsi (Sysoev, 1962), a problematic calcareous tubular fossil with circular cross-section and transversal flanges (Fig. 67G). The fossil from the same locality is considered by Repina et al. (1974: pl. 19: 11) an attachment disk of Hyolithellus vladimirovae Missarzhevsky in Rozanov and Missarzhevsky, 1966.
Reinterpreted herein as a sclerite, the fossil is similar to sclerites of Nidelric pugio Hou, Williams, Siveter, Siveter, Gabbott, Holwell, and Harvey, 2014, from Chengjiang Lagerstätte and Nidelric gaoloufangensis Zhao, Li, and Selden, 2018, from Guanshan biota, Cambrian Series 2, Stage 4, Palaeolenus Zone (Zhao et al. 2018). The latter forms show single-element spines with rounded bases, ca. 0.5 mm in diameter and belong to a group probably allied to chancelloriids.
Stratigraphic and geographic range.—Known from the type horizon and locality only.
Archaeopetasus typicus (Sipin, 1999)
Fig. 39.
1999 Sukharikhina typica Sipin 1999: 8–10, fig. 1а–k.
Material.—Over one hundred calcium phosphatic coatings of sclerites, including figured SMNH X11130–11135, from sample 11/2B, basal Erkeket Formation, Khorbusuonka River, Siberia, Russia. Dokidocyathus regularis Zone, Tommotian stage (correlated with the upper part of Cambrian Stage 2).
Description.—Low-conical isometrical plate, 0.5–1.5 mm in diameter and 0.1–0.5 mm thick, with slightly undulating margin. The margin is usually gently arched at one or two sides but any bilateral symmetry is not evident. The convex (upper) side with a central domal elevation having a single blunt tip. The upper surface with scattered tubercles, ca. 10 μm in diameter. The opposite (lower) side concave smooth with a central circular cavity, 0.1–0.3 mm in diameter covered with tightly located low-relief tubercles, ca. 10 μm in diameter.
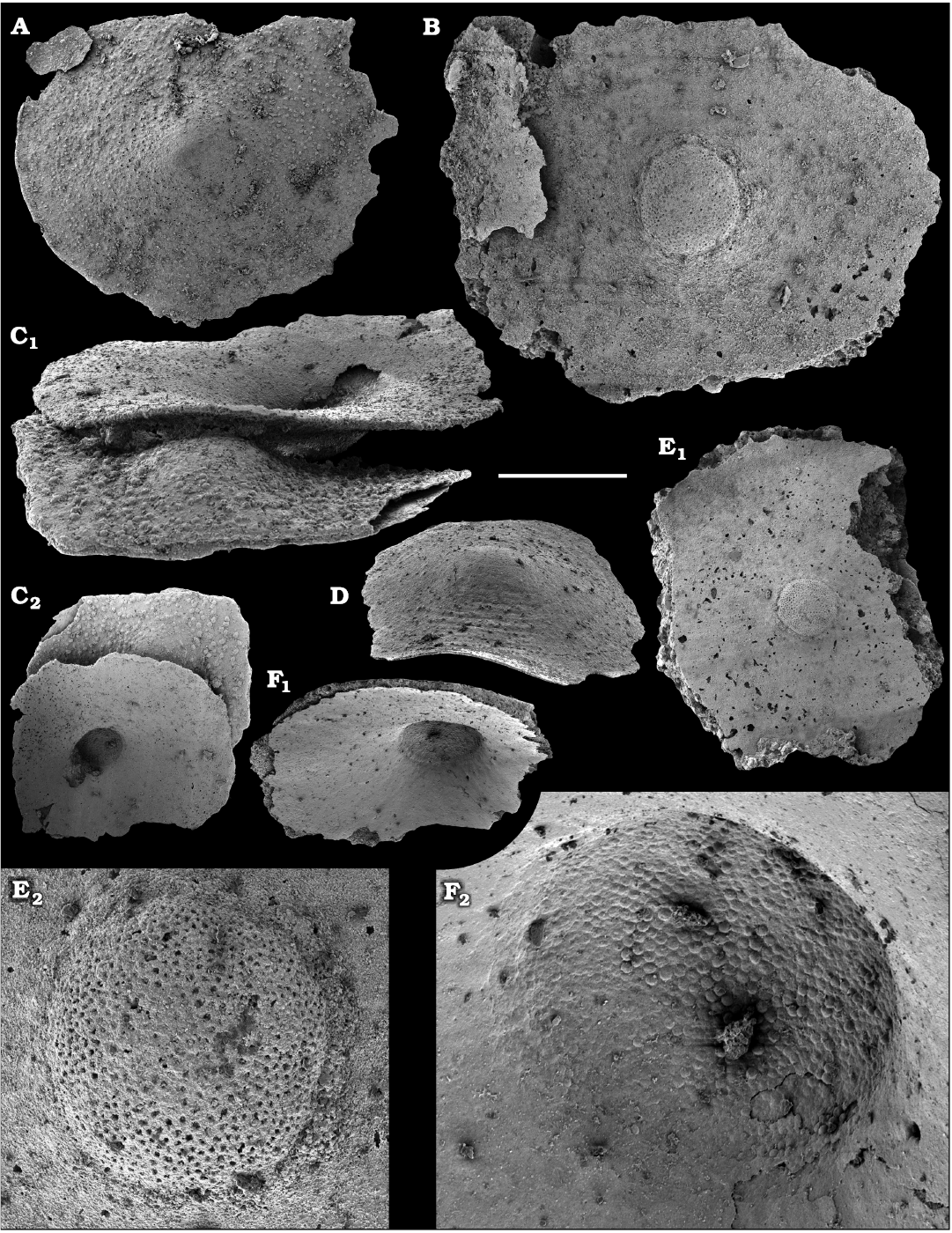
Fig. 39. Sclerites of chancelloriid Archaeopetasus typicus (Sipin, 1999) from the lower Cambrian Erkeket Formation, Khorbusuonka River, Siberia, Russia; sample 11/2B. A–F. Sclerites preserved as phosphatic coats on their convex and concave sides, SMNH X11130–11135, respectively. A. Positive replica of the convex outer surface of sclerite showing scattered tubercles. B. Negative replica of the concave inner surface of sclerite (note a partially preserved positive replica of the convex outer surface with scattered tubercles in the left). C. Two sclerites merged along their convex outer surfaces; C1, close-up showing two merged sclerites in lateral view. D. Oblique lateral view on the convex outer surface of sclerite. E. Phosphatic coat representing a negative replica of the concave inner surface of sclerite (E1), enlargement showing central elevation with pits (E2). F. Phosphatic coat representing a positive replica of the concave inner surface of sclerite with a central cavity (F1), close-up showing the central cavity with flattened tubercles (F2). Scale bar: 500 µm (A, C2, D, E1, F1), 250 µm (B, C1), and 100 µm (E2, F2).
Remarks.—The interspace between the phosphatic coating of the opposite sides of the sclerite is void, probably due to dissolution of the filling material during chemical extraction. In insoluble residues, calcium phosphate coatings representing both sides often occur disarticulated. The upper side of the lower coating, when disarticulated or broken, exhibits a circular tubercle, ca. 0.2 mm in diameter. The latter is a counterpart of the cavity on the lower surface of the sclerite. The circular tubercle bears numerous pits tightly arranged over the entire surface. As was mentioned by Sipin (1999, 2001), the pits correspond to tiny flattened tubercles scattered on the opposite side. Sometimes two different sclerites are stacked or merge with their concave sides. It was interpreted as a “bivalved” fossil by Sipin (1999) in description of a new genus and species, Sukharikhina typica Sipin, 1999, but reinterpreted herein as a taphonomic feature.
Stratigraphic and geographic range.—Nochoroicyathus sunnaginicus and Dokidocyathus regularis zones, Tommotian stage (correlated with the upper part of Cambrian Stage 2), Siberian Platform, Russia.
Phylum uncertain
Class Hyolitha Marek, 1963
Order Orthothecida Marek, 1966
Family uncertain
Genus Conotheca Missarzhevsky in Rozanov et al., 1969
Type species: Conotheca mammilata Missarzhevsky in Rozanov et al. 1969; upper Tommotian–lower Atdabanian stages (correlated with the upper part of Cambrian Stage 2 and lower Stage 3); Tiktirikteekh, near Churan village, middle Lena River, Siberian Platform, Russia.
Conotheca mammilata Missarzhevsky in Rozanov et al.,1969
Fig. 40A–D.
Material.—Several hundred phosphatised conchs from samples 11/2B, 11/16.05, 11/16.1, 11/16.5, 11/18.7, 11/19.6, 11/21.6, 11/28.8, including figured SMNH X11136–11139, from the Erkeket Formation, Khorbusuonka River and sample 21/21, Tyuser Formation, Lena River. Siberia, Russia; Dokidocyathus regularis Zone, Tommotian stage (correlated with the upper part of Cambrian Stage 2).
Description.—Conchs almost straight or gently dorso-ventrally curved, with moderate expansion rate and apical angle up to 15° in dorsal view. Aperture straight and circular. Juvenile conch blunt and somewhat bulbous. External surface with straight thin transverse growth lines. Wall consists of inner layer of transversally oriented fibers and outer layer of longitudinally oriented fibers.
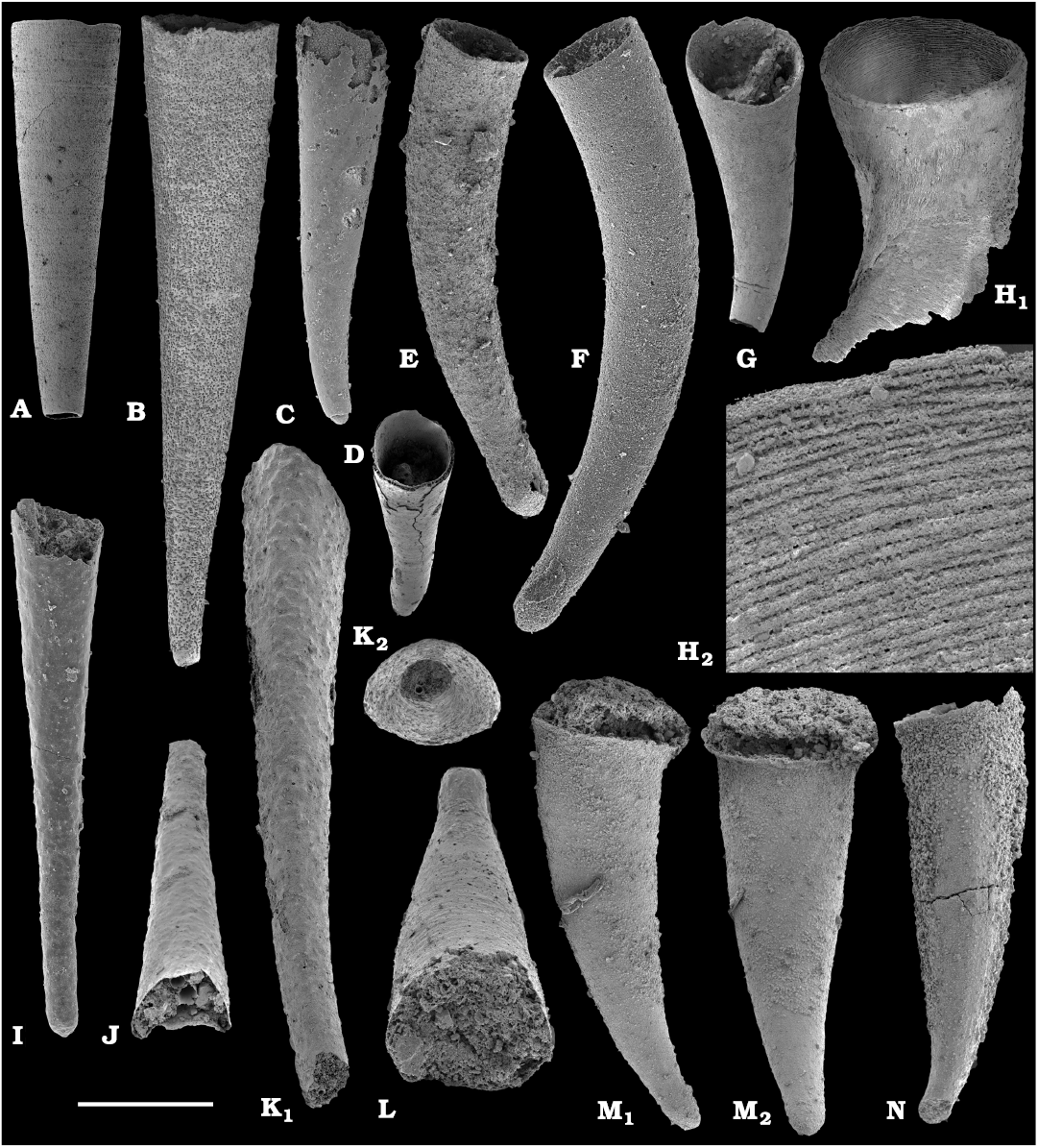
Fig. 40. Hyolith conchs from the lower Cambrian Erkeket Formation, Khorbusuonka River (A–D, G, H, K–N) and Tyuser Formation, lower reaches of the Lena River (E, F, I, J), Siberia, Russia; samples 11/16.05 (K), 11/16.1 (A), 11/16.5 (B, C), 11/2B (D), 19/4.25 (H), 19/11.75 (L), 19/12.75 (M, N), 19/14.25 (G), 21/21 (I, J), and 21/51 (E, F). A, B. Conotheca mammilata Missarzhevsky in Rozanov et al., 1969, phosphatised conchs, SMNH X11136 and 11137, respectively. C, D. Conotheca mammilata Missarzhevsky in Rozanov et al., 1969, phosphatised conchs with initial part preserved, SMNH X11138 and 11139, respectively. E, F. Conotheca cf. longiconica (Qian, 1978), phosphatised conchs, SMNH X11142 and 11143, respectively. G, H. Conotheca circumflexa Missarzhevsky in Rozanov et al., 1969, phosphatised conchs, SMNH X11140 and 11141, respectively. H2, close-up showing transversal fibers in the wall. I–K. Egdetheca aldanica Missarzhevsky in Rozanov et al., 1969, calcium phosphatic internal moulds, SMNH X11144–11146, respectively. K1, lateral view; K2, apical view. L–N. Orthothecida sp. indet., calcium phosphatic internal moulds, SMNH X11147–11149, respectively. M1, lateral view; M2, dorsal view. Scale bar: 1 mm (A), 500 µm (B–D, G–K), 250 µm (E, F, L–N), and 50 µm (H2).
Remarks.—Co-occurring circular orthothecid opercula (Fig. 41) fitting circular straight aperture of Conotheca mammilata conchs may belong to the same species. The outer surface of the operculum is flattened, with a rounded slightly protruding initial part displaced somewhat dorsally. The inner surface of the operculum carries a circumferential bilaterally symmetrical circular ridge, offset slightly from the margin of the operculum and delimiting a circular depression in the middle of the operculum. The outer face of the ridge dips ca. 90º towards the opercular margin. The inner face slopes towards the center of operculum and carries bilaterally symmetrical lateral sets of radial folds. The crest of the ridge undulates: it arches ventrally to the opercular margin, produces bilaterally symmetrical elevated folds laterally and a dorsal pair of closely situated short prongs. The lateral folds are reminiscent of clavicules of hyolithids, whereas prongs can be considered cardinal processes. Although attached opercula are not found and cannot be at present unequivocally paired with conchs in our material, similar opercula with an offset marginal zone are previously described from Conotheca australiensis Bengtson in Bengtson et al., 1990, Conotheca rotunda (Duan, 1984), and Conotheca brevica Qian, Xie, and He, 2001. They have similar clavicle-like structures and paired processes on the internal side (Devaere et al. 2014).
Stratigraphic and geographic range.—Tommotian stage (correlated with the upper part of Cambrian Stage 2), Siberian Platform, Russia.
Conotheca circumflexa Missarzhevsky in Rozanov et al., 1969
Fig. 40G, H.
Material.—Twenty phosphatised conchs, including figured SMNH X11140 and 11141, from samples 19/4.25, 19/5.5, 19/8.5, 19/14.25. Erkeket Formation, Khorbusuonka River, Siberia, Russia. Lower Botoman stage (correlated with the lower part of Cambrian Stage 4).
Description.—Conch helically curved about 1/4 revolution clockwise in the direction of growth, with apical angle ca. 20°. Aperture straight and slightly oval. Wall consists of transversally oriented fibrous bundles running around the conch in the inner layer and longitudinally oriented in the outer layer.
Remarks.—See also Kouchinsky et al. (2009: fig. 45 and 2015a: fig. 27F).
Stratigraphic and geographic range.—Atdabanian and Botoman stages (correlated with the Cambrian Stage 3 and lower part of Stage 4), Siberian Platform, Russia.
Conotheca cf. longiconica (Qian, 1978)
Fig. 40E, F.
Material.—Over 20 phosphatised conchs, including figured SMNH X11142 and 11143, from sample 19/25.5, Erkeket Formation, Khorbusuonka River, Toyonian stage (correlated with the Cambrian Stage 4) and samples 21/46.1, 21/51, 21/52, 21/54, Tyuser Formation, Lena River, Delgadella anabara–Nevadella Zone, Atdabanian stage (correlated with the Cambrian Stage 3). Siberia, Russia.
Description.—Conch gently curved in sagittal plane, with circular cross-section and straight aperture. Apical angle ca. 10°. Juvenile conch rounded blunt, not clearly separated. Wall porous consisting of a network of fibers, similarly to other hyoliths.
Remarks.—This form is more curved and slimmer than the holotype of C. mammilata. It occurs in the Cambrian Stages 2 and 3 of the Siberian Platform. Similar conchs are illustrated by Kouchinsky et al. (2015a: figs. 25E–G, 27E; 2017: fig. 52). Qian and Bengtson (1989) synonymised Circotheca longiconica Qian, 1978, with Conotheca subcurvata (Yu, 1974). We support herein placement within Conotheca, but find difference between these two species. Conotheca longiconica (Qian, 1978) has a conch regularly curved in a single plane, whereas Conotheca subcurvata shows irregular curvature (see Luo et al. 1984: pl. 7: 8).
Genus Michniakia Missarzhevsky in Rozanov and Missarzhevsky, 1966
Type species: Michniakia minuta Missarzhevsky in Rozanov and Missarzhevsky, 1966; monotypic; Atdabanian stage (correlated with the Cambrian Stage 3); Chekurovka section, lower reaches of the Lena River, Siberian Platform, Russia.
Michniakia minuta Missarzhevsky in Rozanov and Missarzhevsky, 1966
Fig. 42.
1966 Michniakia minuta sp. nov.; Missarzhevsky in Rozanov and Missarzhevsky 1966: 106–107, pl. 10: 5, 6.
1986 Michniakia minuta Missarzhevsky in Rozanov and Missarzhevsky, 1966; Ermak and Pelman 1986: 189–190, pl. 26: 1, 2.
2004 Michniakia minuta Missarzhevsky in Rozanov and Missarzhevsky, 1966; Gubanov et al. 2004a: 212, fig. 6: 14–18, 33.
Material.—Several tens of calcium phosphatic internal moulds with fragments of phosphatised shells, including SMNH X11353–11357, from sample 20/1B, Erkeket Formation, Khorbusuonka River, Siberia, Russia. Lower Botoman stage (correlated with the lower Cambrian Stage 4).
Description.—Planispiral slowly expanding shell, up to 1 mm in diameter, coiled to 3/4 revolution, with blunt flattened apex and circular cross-section, ca. 350 µm at the aperture. Shell and internal moulds form a loop bringing the apex into contact with the later growth stage at ca. 3/4 of a whorl. A single conch is slightly anisostrophically (dextrally) coiled (Fig. 42A4).
The wall consists of at least two layers. The outer layer exposed on the shell surface is longitudinally striated perpendicularly to faint growth lines near the apex (Fig. 42C1). Near the aperture, striation is tangentially oriented and aligned with circular to ovaliform pores, up to 10 μm in diameter (Fig. 42A1). The inner layer has an apparently smooth surface and forms the apical septum (Fig. 42D, E1). The aperture with a gentle sinus in its outer (ventral) margin (Fig. 42E3). A single specimen (Fig. 42B) is preserved with a broadly oval, nearly circular operculum sealing the aperture. The operculum and conch are bilaterally symmetrical along the same (dorso-ventral) plane. The ventral margin of operculum is gently incurved to fit the ventral sinus of the aperture. The dorsal margin of operculum is almost straight and aligned with the dorsal edge of the aperture. The operculum has a slight radial plication in its dorsal part. The distal part of the operculum is flattened and overlaps the apertural margin. The initial part of the operculum is situated in its centre.
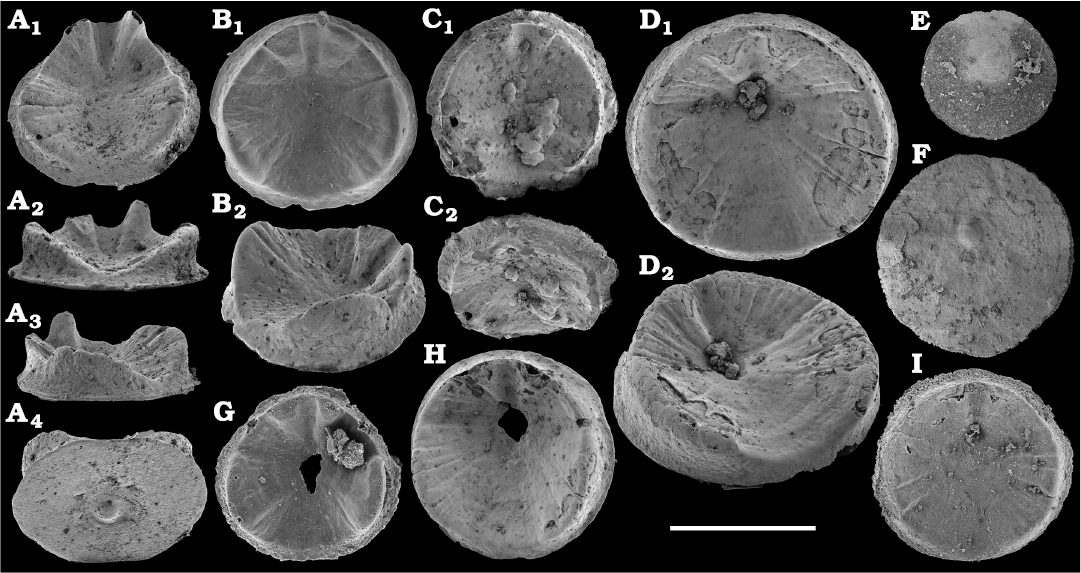
Fig. 41. Hyolith opercula from the lower Cambrian Tyuser Formation, lower reaches of the Lena River (A, B, G) and Erkeket Formation, Khorbusuonka River (C–F, H, I), Siberia, Russia; samples 11/16.05 (C, H), 11/16.1 (I), 11/2B (D–F), and 21/21 (A, B, G). A–I. Phosphatised opercula tentatively attributed to Conotheca mammilata Missarzhevsky in Rozanov et al., 1969, SMNH X11150–11158, respectively. A1, B1, C1, D1, inner surface; A2, ventral view; A3, lateral view; B2, oblique lateral view; A4, C2, oblique dorsal view on the opercula. Scale bar 500 µm.
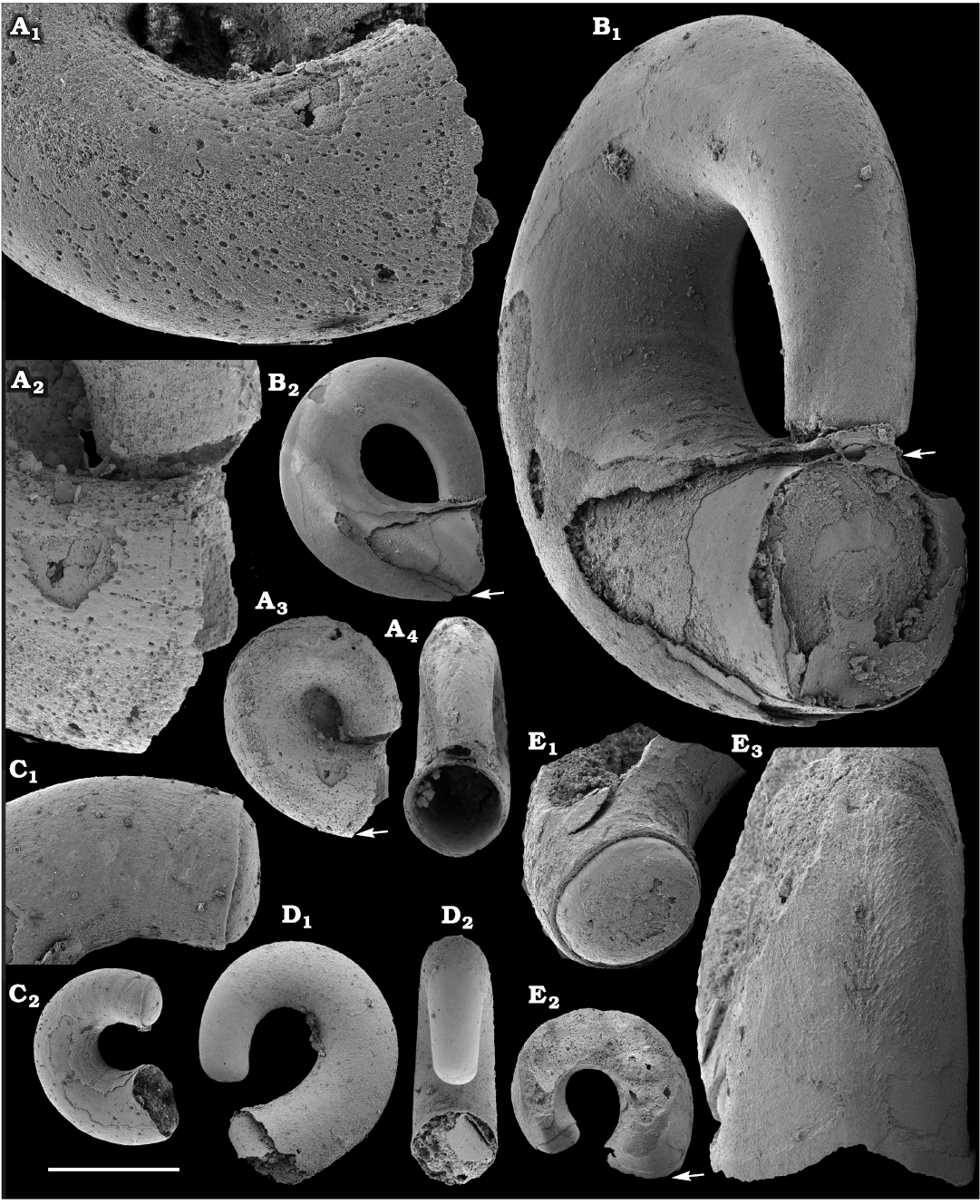
Fig. 42. Hyolith Michniakia minuta Missarzhevsky in Rozanov and Missarzhevsky, 1966, from the lower Cambrian Erkeket Formation, Olenyok Uplift, Siberia, Russia; sample 20/1B. A–E. Internal moulds with partially phosphatised shell and phophatic coatings, SMNH X X11353–11357, respectively. A. Phosphatised shell with longitudinal and tangential fibrosity and pores in the wall. A3, lateral view showing ventral sinus of the aperture (arrow); A4, pertural view; A1, A2, enlargements of the apical part and aperture. B. Shell sealed with operculum (contact between the edge of the aperture and dorsal margin of the operculum arrowed). B1,oblique apertural view; B2, lateral view showing ventral sinus of the aperture (arrows). C. Shell showing septum at the apical end (C1, close-up). D. Shell preserved as inner phosphatic coating including that of the apical septum; D1, lateral view; D2, apertural view. E. Shell with apertural sinus; E2, ventral side arrowed; E1, enlargement of the apical end; E3, ventral side. Scale bar: 500 µm (A3, A4, B2, C2, D, and E2) and 165 µm (A1, A2, B1, C1, E1, and E3).
Remarks.—The septum-like structure suggests decollating growth mode assumed for Cupitheca—type conchs of hyoliths (see below). The fibrous striation with aligned pores in the wall, flattened apical part with a rounded septum, aperture closed by operculum suggest a hyolithan affinity of Michniakia previously reported as a mollusc. Based on hyolith-type microstructures, it has been suggested that hyolith morphologies included planispirally coiled forms (Feng et al. 2001). Other coiled mollusc-like shells such as isostrophic Khairkhania, anisostrophic Barskovia, Nomgoliella, and Philoxenella demonstrate features compatible with the same affinity (Kouchinsky et al. 2017). Planispiral, evolute shells of Ardrossania paveyi Runnegar in Bengtson et al., 1990, from the Mernmerna Formation of South Australia have shells up to 4 revolutions and 1.5 mm in diameter, 300–400 µm in cross-section diameter at the aperture (Gravestock et al. 2001). It has been suggested to be similar to gastropod-like molluscs and hyoliths (Bengtson et al. 1990). Parkhaev in Gravestock et al. (2001) attributed Ardrossania to the family Khairkhaniidae Missarzhevsky, 1989.
Stratigraphic and geographic range.—Cambrian Series 2, Siberia (Russia) and Spain.
Family Protowenellidae Peel, 2021b
Genus Protowenella Runnegar and Jell, 1976
Type species: Protowenella flemingi Runnegar and Jell, 1976; Drumian Stage; Gowers Formation, Georgina Basin, South Australia.
Protowenella flemingi Runnegar and Jell, 1976
Fig. 24I.
Material.—Several calcium phosphatic and glauconised internal moulds, including figured SMNH Mo194712, from sample 22/50, Sekten Formation, Lena River, Siberia, Russia. Upper Toyonian stage (correlated with the Cambrian Stage 4).
Description.—Planispiral calcium phosphatic internal mould, ca. 0.5 mm in size, coiled through half of a whorl, with rounded cross-section and globose initial part. Subapical side of the mould is delimited by a symmetrically located pair of shallow diverging grooves, termed circumbilical channels.
Remarks.—Peel (2021b) described Protowenella flemingi from the Wuliuan Stage of Greenland with an in-place operculum of a hyolith-type and suggested affinity with orthothecid hyoliths. This is corroborated by presence of planispirally coiled forms among early–middle Cambrian hyoliths (Feng et al. 2001; Kouchinsky et al. 2011; and herein).
Stratigraphic and geographic range.—Cambrian Stage 4–Drumian Stage, Siberian Platform (Russia), Cambrian Series 2 of North China and Miaolingian Series of Australia, Laurentia and Bornholm (Baltica) (see Li et al. 2019 for discussion).
Family uncertain
Genus Egdetheca Missarzhevsky in Rozanov et al., 1969
Type species: Egdetheca aldanica Missarzhevsky in Rozanov et al. 1969; Dokidocyathus regularis Zone, Tommotian stage (upper part of Cambrian Stage 2); Tiktirikteekh Creek, Churan Village, middle Lena River, Siberian Platform, Russia.
Egdetheca aldanica Missarzhevsky in Rozanov et al., 1969
Fig. 40I–K.
Material.—Over 20 calcium phosphatic internal moulds, including figured SMNH X11144–11146, from samples 11/2B, 11/16.05, 11/16.1, 11/16.5, basal Erkeket Formation, Khorbusuonka River, and sample 21/21, Tyuser Formation, Lena River; Siberia, Russia. Dokidocyathus regularis Zone, Tommotian stage (upper part of Cambrian Stage 2).
Remarks.—Conchs slowly expanding and slightly curved in sagittal plane, with a semi-oval or rounded trapezoidal cross-section. Initial part extended concave on the ventral side and slightly bulbous at the end. The surface of the internal mould is usually covered with ca. 10 µm low tubercles, which in combination with the general morphology of the mould is diagnostic for the species.
Stratigraphic and geographic range.—Nochoroicyathus sunnaginicus and Dokidocyathus regularis zones, Tommotian stage (correlated with the upper part of Cambrian Stage 2), Siberian Platform, Russia.
Genus Majatheca Missarzhevsky in Rozanov et al., 1969
Type species: Majatheca tumefacta Missarzhevsky in Rozanov et al., 1969; upper Tommotian stage, lower Cambrian; Tiktirikteekh Creek, Churan Village, middle Lena River, Siberian Platform, Russia.
Majatheca sp.
Fig. 43.
Material.—Several tens of phosphatised opercula, including figured SMNH X11159–11163, from samples 11/16.05 and 11/2B, basal Erkeket Formation, Khorbusuonka River and sample 21/21, Tyuser Formation, lower Lena River. Siberia, Russia; Dokidocyathus regularis Zone, Tommotian stage (correlated with the upper part of Cambrian Stage 2).
Description.—Outer surface of the operculum flattened to slightly concave, gently bending in sagittal direction (Fig. 43C2), showing a rounded trapezoidal or rounded pentagonal aspect in the outline with slightly undulating margin. Initial part centrally placed, rounded and slightly protruding on the outer surface of operculum. The latter carries continuous concentric growth lines. Inner surface of the operculum carries a circumferential bilaterally symmetrical slightly dorso-ventrally extended rounded pentagonal ridge, offset from the margin of the operculum, and a gently sloping rim between the ridge and opercular margin. Outer face of the ridge vertical with respect to the rim. Inner face slopes gently towards the center of operculum and carries bilaterally symmetrical radial folds. Crest of the ridge undulates. In ventral part of the operculum it arches broadly towards the opercular margin, forms two elevated folds laterally and produces a narrow fold dorsally towards the opercular margin.
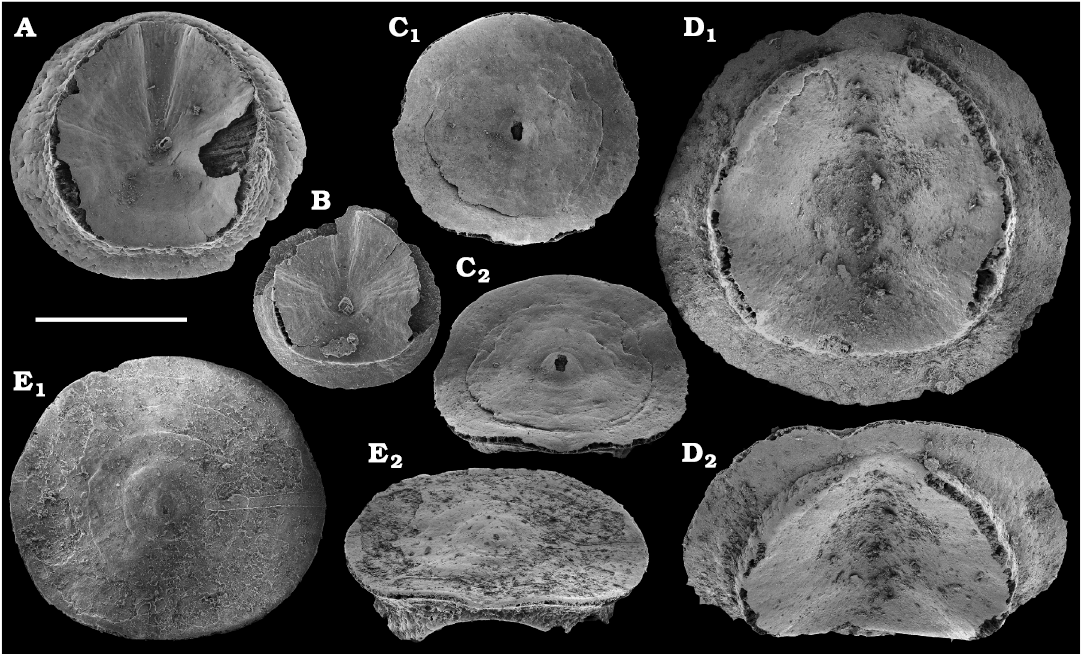
Fig. 43. Hyolith opercula Majatheca sp. from the lower Cambrian Erkeket Formation, Khorbusuonka River (A, C–E) and Tyuser Formation, lower reaches of the Lena River (B), Siberia, Russia; samples 11/16.05 (A, C, E), 11/2B (D), and 21/21 (B). A–E. Phosphatised opercula, SMNH X11159–11163, respectively. C1, E1, outer surface; C2, E2, oblique ventral view; D1, inner surface; D2, oblique dorsal view. Scale bar 500 µm.
Remarks.—Similar opercula, Circotheca sp. 1, are illustrated by Meshkova (1974: pl. 11: 2, 3, 9) from the Tyuser Formation, Chekurovka section (Lena River). A larger specimen probably of the same species is defined by Meshkova (1974: pl. 11: 12) as Allatheca sp. The opercula lack processes and are generally similar to Majatheca tumefacta (Fig. 51F herein and Kouchinsky et al. 2015a: fig. 24) from lower Cambrian Stage 4 beds of the Siberian Platform. Majatheca tumefacta opercula have, however, more laterally extended broad oval shape and a clearly developed rounded trapezoidal aspect of the circumferential ridge on the inner side of the operculum.
Genus Cupitheca Duan in Xing et al., 1984
Type species: Cupittheca brevituba Duan in Xing et al., 1984; Meishucunian stage, Kuanchuanpu Formation and Qiongzhusian stage, Xihaoping Formation; China.
Cupitheca sp.
Fig. 44.
Material.—Several hundred phosphatised conchs and calcium phosphatic internal moulds, including figured SMNH X11164–11172, from samples 19/5.5, 19/8.5, 19/10.25, 19/11.75, 19/12.75, 19/25.5, 19/26.75, 19/29, 19/33, 20/1B, Erkeket Formation, Khorbusuonka River, Botoman and Toyonian stages (correlated with the Cambrian Stage 4) and sample 21/51, Tyuser Formation, Delgadella anabara–Nevadella Zone, Atdabanian stage (correlated with the Cambrian Stage 3). Siberia, Russia.
Remarks.—Represented by phosphatised fragments of conchs gently curved in sagittal plane, with circular cross-section and straight aperture. Apical end of the fragments with a hemispherical, apically convex septum. Wall carries irregular thin growth lines and circumferentially running densely arranged fibers in the inner layer. The outer layer and surface of the septum has a porous structure. Deeper portion of the septum preserved as parallel columns perpendicularly oriented to the outer and inner surfaces. The columns are phosphatic infills of the porous channels through the septum. Apices of the conchs are detached and a few co-occurring fragments can herein be interpreted as initial parts broken off along the septal margin. They are goblet-shaped, with an extended tip having a blunt termination, abruptly expanding towards a circular aperture.
Opercula (Fig. 45) are found associated with the conchs, but they cannot be unequivocally interpreted as belonging to the same species. They are circular with flat, slightly undulating outer side carrying concentric growth lines. Bilaterally symmetrical structures on the inner side represent elevated lateral undulating ridges with radial folds. Ridges dip steeply towards the margin of the operculum and slope gently towards the initial part. The initial part is centrally situated in the operculum and represents an elevated cone on the exterior surface opposed by a deep pit on the interior. There is a sinus between the elevated dorsal parts of the ridges, on the dorsal side of the operculum. The ridges level off towards the ventral one-third part of the operculum, where the ventral sector is gently convex on the interior.
Forms described under such names as Cupitheca Duan in Xing et al., 1984, and Actinotheca Xiao and Zhou, 1984, recovered as conch fragments with circular to broadly oval cross-section and a hemispherical, apically convex terminal septum can be interpreted as a mode of preservation in a polyphyletic group of orthothecid hyoliths with a septum in the apical part and a circular cross-section. A similar mode of preservation occurs, e.g. in Majatheca tumefacta Missarzhevsky in Rozanov et al., 1969, with a broken off apical part (Kouchinsky et al. 2015a: fig. 22B3).
Genus Triplicatella Conway Morris in Bengtson et al., 1990
Type species: Triplicatella disdoma Conway Morris in Bengtson et al., 1990; Abadiella huoi Zone, Cambrian Stage 3; Curramulka, Yorke Peninsula, Australia.
Triplicatella sp.
Fig. 46.
Material.—Five silicified internal moulds of conchs with casts of inner surfaces of opercula (situated in closed position with respect to the aperture of the conch), including figured specimens SMNH X11178–11181, from sample 19/65.5, and probably several hundred silicified moulds of conchs without traces of opercula belonging to the same species, from samples 19/66.5, 19/66.75, 19/3B, 19/4B, Kuonamka Formation, Khorbusuonka River, Siberia, Russia. Tomagnostus fissus–Acadoparadoxides sacheri Zone, lower Mayan stage (correlated with the lowermost Drumian Stage).
Description.—Conchs rounded trapezoidal in cross-section, gently curved in sagittal plane, moderately expanding from the initial part, at ca. 25° in the dorsal view and ca. 20° in lateral view. The aperture carries a bilaterally symmetrical cast of a folded operculum, rounded trapezoidal to oval in the frontal view. The operculum, 0.7–1 mm wide is flattened to slightly concave externally. It carries two deep folds forming a V-shaped groove diverging at ca. 40° on the surface viewed in dorso-ventral orientation of the conch. Invagination of the dorsal margin of the operculum produces two distinct embayments in the dorsal margin of the conch mould. The folds continue ventrally, converging and shallowing towards the ventral margin of the operculum.
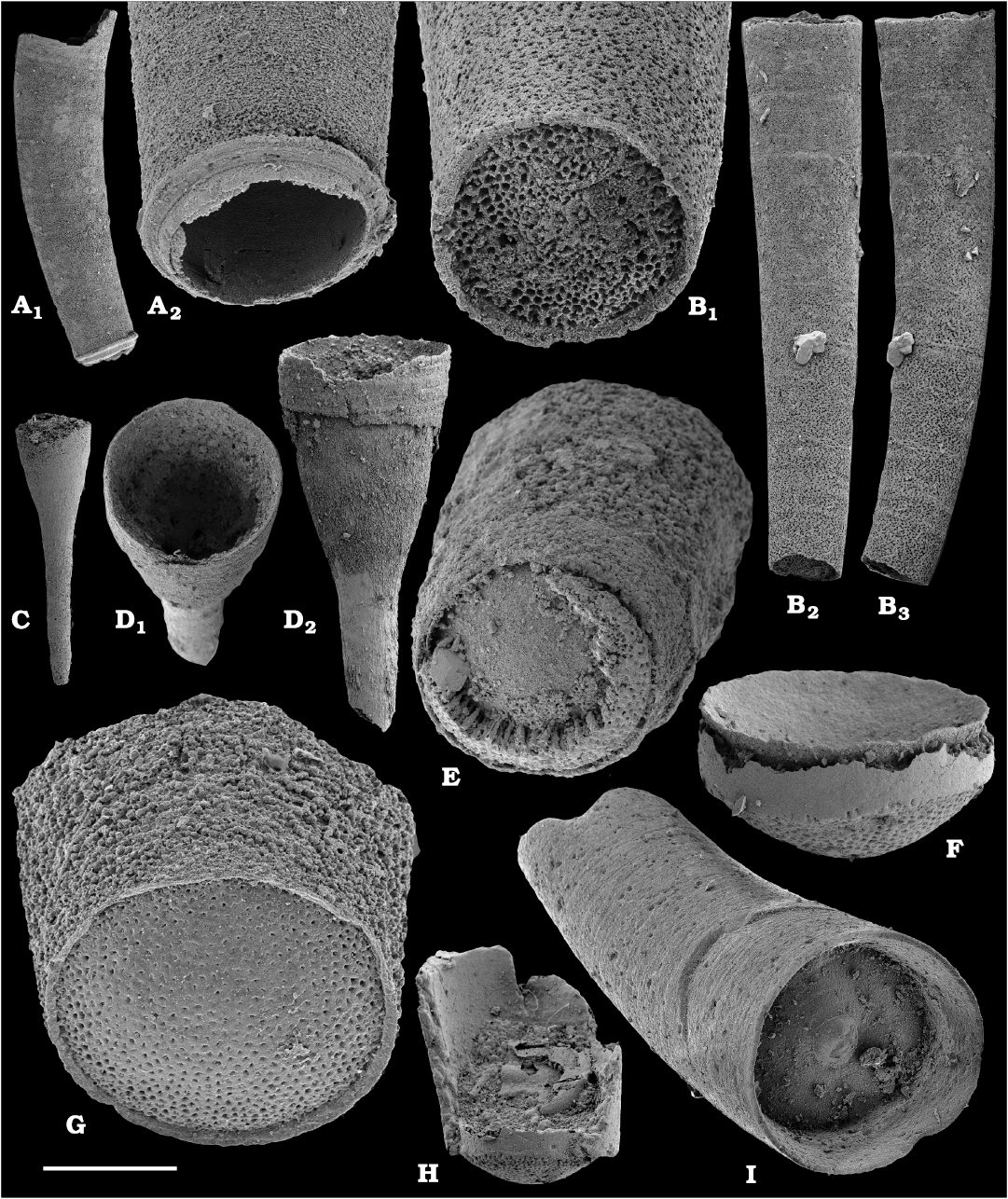
Fig. 44. Hyolith conchs Cupitheca sp., from the lower Cambrian Erkeket Formation, Khorbusuonka River, Siberia, Russia; samples 19/12.75 (E–I) and 19/33 (A–D). A, B, E–I. Phosphatised conchs, SMNH X11164–11170, respectively. A1, B3, lateral view; A2, close-ups of the apical end; B2, dorsal view. B1, enlarged apical end showing septum. E–H, fragments of conch with septum. I, probable operculum of the same species within apertural end of conch. C, D. Apical ends of conch, presumably of the same species, with initial parts preserved, SMNH X11171 and 11172, respectively. D1, oblique apertural view; D2, lateral view. Scale bar: 500 µm (A1, B2, B3, C), 250 µm (D, H, I), and 125 µm (A2, B1, E–G).
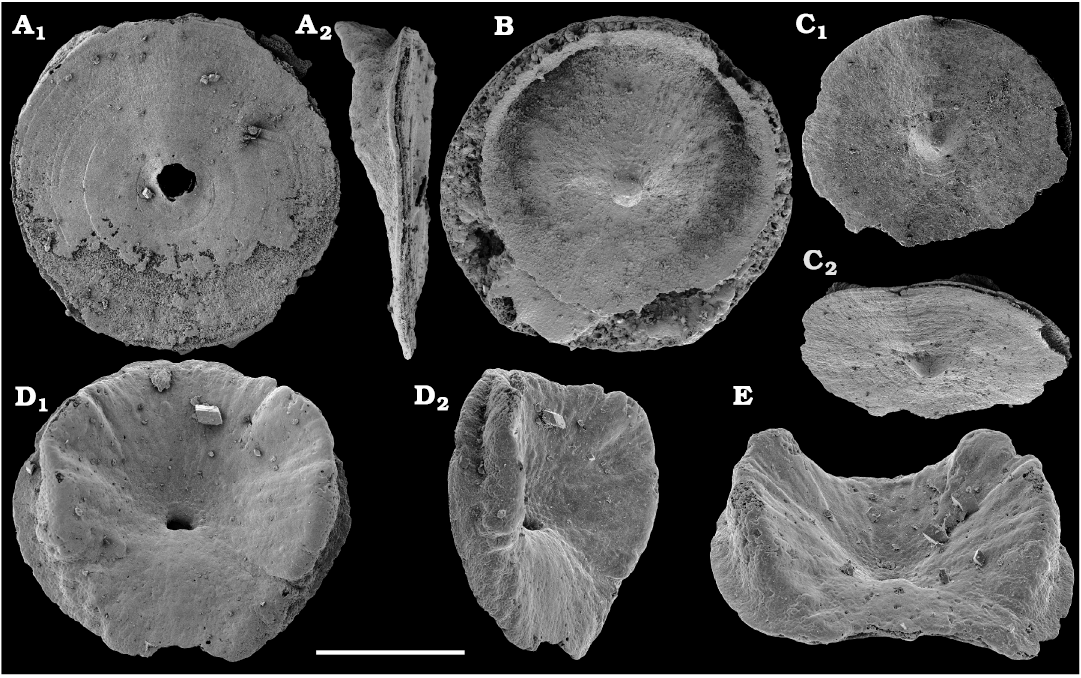
Fig. 45. Hyolith opercula, presumably Cupitheca sp. (see Fig. 44), from the lower Cambrian Erkeket Formation, Khorbusuonka River, Siberia, Russia; sample 19/12.75. A–E. Phosphatised opercula, SMNH X11173–11177, respectively. A1, C1, outer surface; A2, lateral view; C2, oblique dorsal view; D1, inner surface; D2, oblique lateral view. Scale bar 250 µm.

Fig. 46. Hyolith conchs and opercula, Triplicatella sp., from the middle Cambrian Kuonamka Formation, Khorbusuonka River, Siberia, Russia; sample 19/65.5. A–D. Silicified internal moulds of conch closed with operculum, SMNH X11178–11181, respectively. A1, lateral view; A2, apertural view. Scale bar: 1 mm (B) and 500 µm (A, C, D).
Remarks.—The form described above is similar to that figured by Kouchinsky et al. (2011: fig. 11A), although a silicified operculum and conch therein may not belong to the same species, and to Triplicatella sinuosa figured by Kouchinsky et al. (2015a: fig. 29C). Four species of Triplicatella Conway Morris in Bengtson et al., 1990, are described, such as Triplicatella disdoma Conway Morris in Bengtson et al., 1990 (type species), Triplicatella sinuosa Skovsted, Peel, and Atkins, 2004, Triplicatella peltata Skovsted, Peel, and Atkins, 2004, and Triplicatella papilio Kouchinsky in Kouchinsky et al., 2015a, are known from acid isolated material while a fifth species, Triplicatella opimus (Yu, 1974) with preserved soft parts, is known from the Chengjiang fauna of South China (Liu et al. 2020, 2021).
Orthothecida sp. indet.
Fig. 40L–N.
Material.—Several tens of calcium phosphatic internal moulds, including SMNH X11147–11149, from 19/5.5, 19/11.75, 19/12.75. Erkeket Formation, Khorbusuonka River, Siberia, Russia. Lower Botoman stage (correlated with the lower Cambrian Stage 4).
Description.—Moderately expanding conchs, slightly dorso-ventrally curved at the initial part. Aperture rounded trapezoidal and straight. In dorsal view, the apical angle is ca. 30°. Apical part rounded blunt, not separated.
Orthothecid operculum type A
Fig. 47.
Material.—Several tens of phosphatised opercula, including figured SMNH X11182–11189, from sample 21/21, Tyuser Formation, Lena River, Siberia, Russia. Dokidocyathus regularis Zone, Tommotian stage (correlated with the upper part of Cambrian Stage 2).
Description.—Bilaterally symmetrical operculum with a sub-circular outline and flattened outer side. Initial part circular flattened situated in a shallow depression of the outer side of the operculum, about twice as close to the dorsal side of the operculum as to the ventral one. A bilaterally symmetrical pair of radial folds beginning at the initial part divide the outer surface into two unequal sectors (lobes) situated at ca. 90° to each other. The opposite surface of the operculum carries a circular ridge with two closely situated prongs, elevated lateral parts and sinus in the ventral portion. The outer side of the ridge is steep towards the margin of the operculum. The inner side slopes towards the initial part.
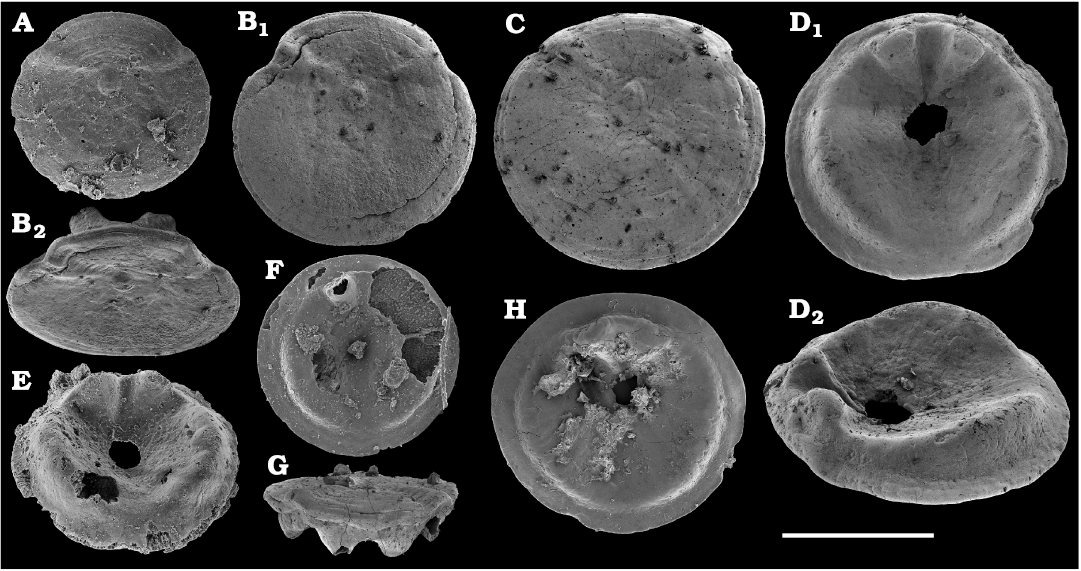
Fig. 47. Hyolith opercula “Orthothecid operculum type A”, from the lower Cambrian Tyuser Formation, lower reaches of the Lena River, Siberia, Russia; sample 21/21. A–H. Phosphatised opercula, SMNH X11182–11189, respectively. B1, outer surface; B2, oblique dorsal; D1, inner surface; and D2, oblique lateral views. Scale bar 500 µm.
Remarks.—Most similar to the Conotheca mammilata? operculum (Fig. 41), but differs from it in having outer side divided by а bilaterally symmetrical pair of radial folds into ventral and dorsal lobes.
Orthothecid operculum type B
Fig. 48.
Material.—Several tens of phosphatised opercula, including figured SMNH X11190–11195, from samples 11/2B and 11/16.05, basal Erkeket Formation, Khorbusuonka River and 21/21, Tyuser Formation, Lena River. Siberia, Russia; Dokidocyathus regularis Zone, Tommotian stage (correlated with the upper part of Cambrian Stage 2).
Description.—Broadly ovaliform (smaller specimens) to rounded rectangular operculum somewhat extended in sagittal plane, with thick wall and low domal external surface covered with distinct growth increments. Initial part elevated and blunt, slightly shifted towards the dorsal side of the operculum. The external surface flattens laterally. Inner surface concave and smooth separated from the external surface by a thick rim consisting of growth increments. The concavity is somewhat extended in the sagittal plane of the operculum and has a shallow sinus dorsally.
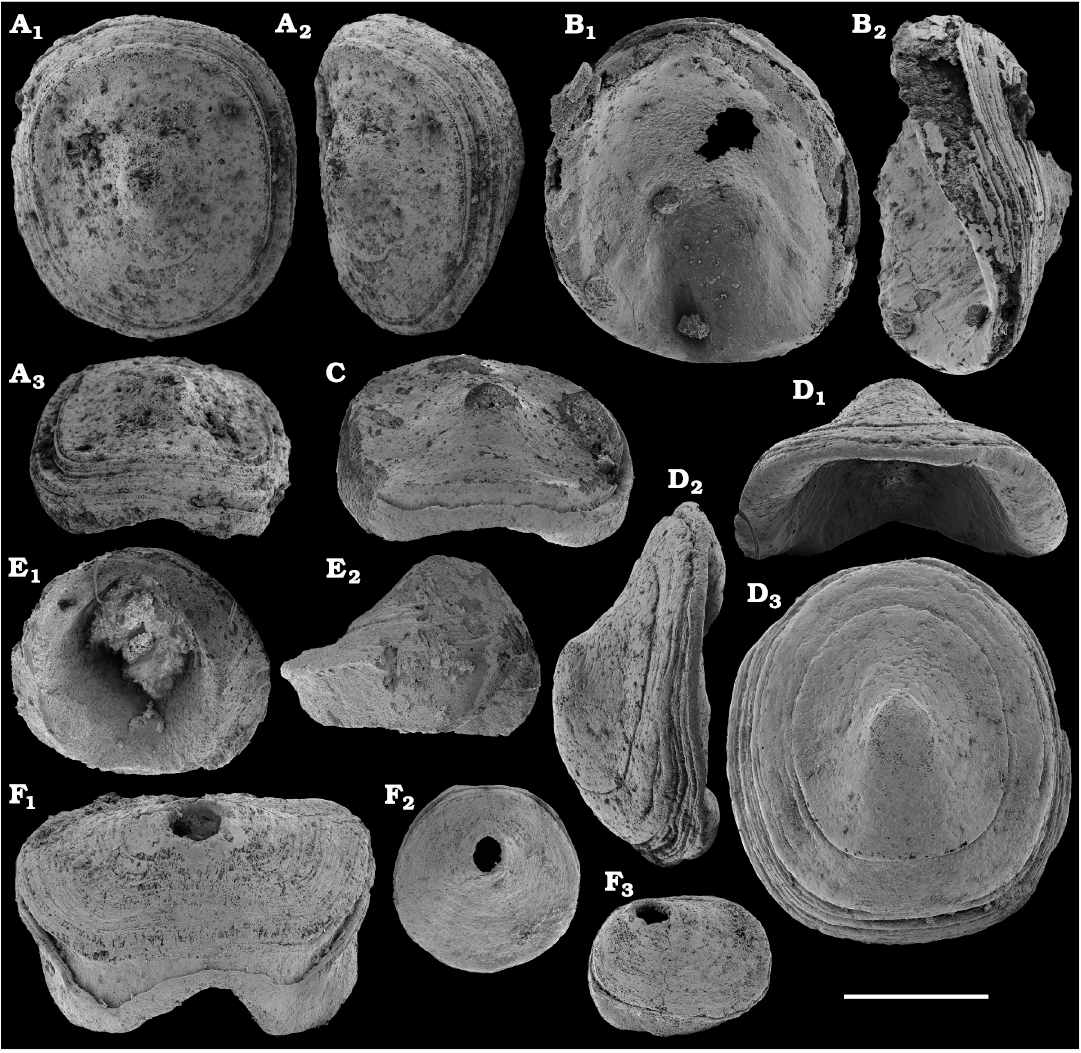
Fig. 48. Hyolith opercula “Orthothecid operculum type B”, from the lower Cambrian Erkeket Formation, Khorbusuonka River (A, C–F) and Tyuser Formation, lower reaches of the Lena River (B), Siberia, Russia; samples 11/16.5 (A, C, F), 11/2B (D, E), and 21/21 (B). A–D. Phosphatised opercula, SMNH X11190–11193, respectively. E, F. Phosphatised opercula of presumably the same species, SMNH X11194 and 11195, respectively. A1, D3, F2, outer surface; A2, B2, F3, oblique lateral; D2, E2, lateral; A3, D1, F1, dorsal; B1, E1, inner surface views. Scale bar: 500 µm (A–E, F2, F3) and 250 µm (F1).
Orthothecid operculum type C
Fig. 49.
Material.—Several tens of phosphatised opercula, including figured SMNH X11196–11202, from samples 11/2B and 11/16.05, basal Erkeket Formation, Khorbusuonka River and 21/21, Tyuser Formation, Lena River. Siberia, Russia; Dokidocyathus regularis Zone, Tommotian stage (correlated with the upper part of Cambrian Stage 2).
Description.—Operculum broadly oval slightly extended in the sagittal plane with a rounded initial part situated closer to the dorsal margin, at ca. 1/3 of the opercular diameter. Domal in the lateral view, with a deep sinus of the dorsal margin. The outer surface covered with concentric growth lines and thicker growth increments at the margin. Inner side concave and smooth, with radial striae and pitted surface. Walls thick consisting of fibrous bundles.
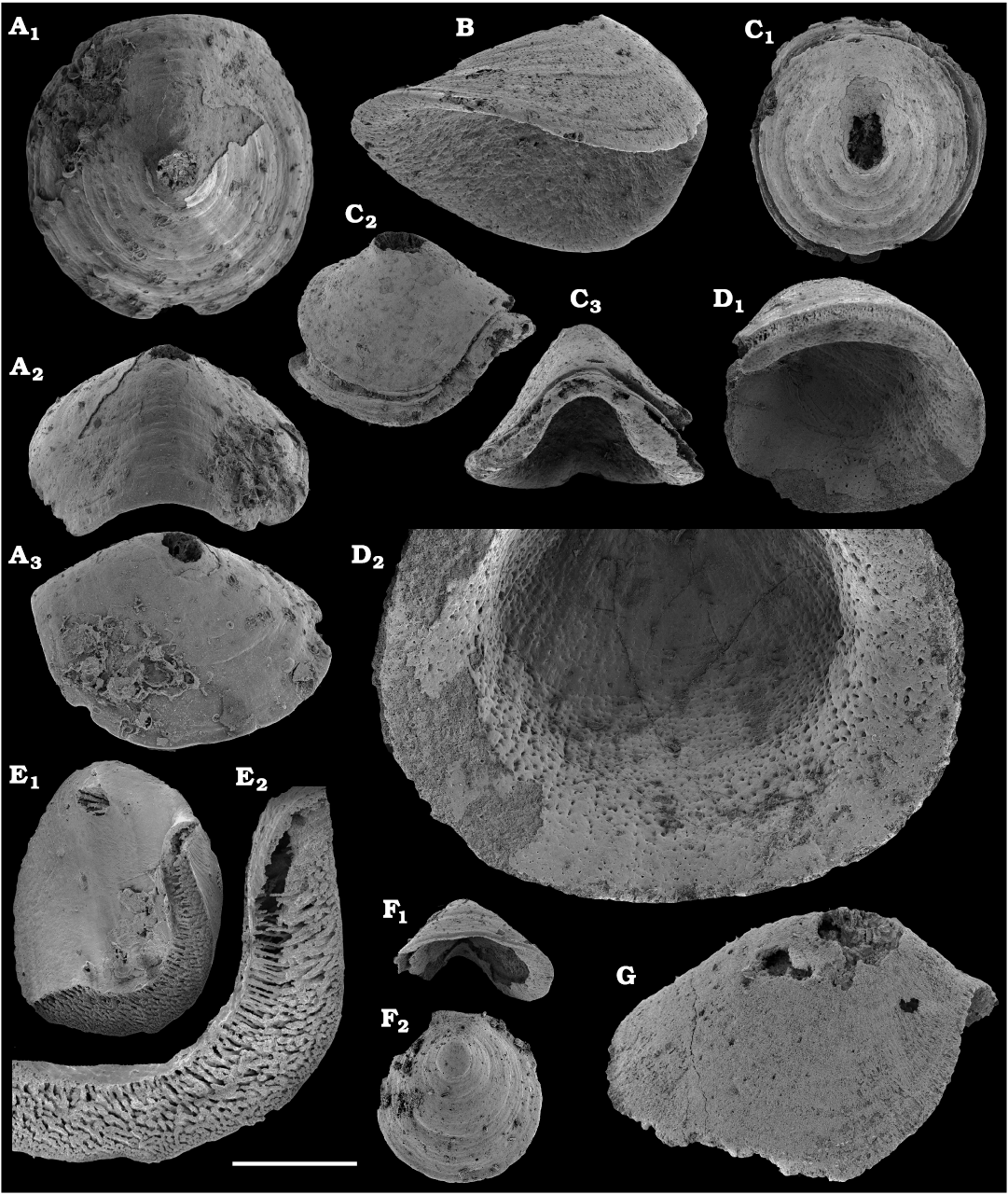
Fig. 49. Hyolith opercula “Orthothecid operculum type C”, from the lower Cambrian Erkeket Formation, Khorbusuonka River (A–D) and Tyuser Formation, lower reaches of the Lena River (E–G), Siberia, Russia; samples 11/2B (A–D) and 21/21 (E–G). A–G. Phosphatised opercula, SMNH X11196–11202, respectively. E1, inner surface and transverse broken view; E2, enlargement showing fibrous composition of the wall; F1, ventral view; A1, C1, F2, outer surface; A2, C3, dorsal view; A3, lateral view; C2, oblique lateral view; D1, oblique inner surface view; D2, enlargement showing pitted inner surface. Scale bar: 500 µm (A, C, D1, E1, F) and 250 µm (B, D2, E2, G).
Remarks.—Other undetermined phosphatised orthothecid opercula of different types occur in the Dokidocyathus regularis Zone, Tommotian stage: SMNH X11203 (Fig. 50A), from sample 11/16.05, SMNH X11205 (Fig. 50C) and SMNH X11206 (Fig. 50D), from sample 11/2B (basal Erkeket Formation, Khorbusuonka River) and SMNH X11204 (Fig. 50B), from sample 21/21 (Tyuser Formation, Lena River).
Order Hyolithida Sysoev, 1957
Family uncertain
Genus Microcornus Mambetov, 1972
Type species: Microcornus parvulus Mambetov, 1972; Rhombocorniculum cancellatum Zone, lower Cambrian; Geres Member of the Shabakty Group, Ushbas River, Lesser Karatau, Kazakhstan.
Microcornus cf. eximius Duan, 1984
Fig. 52A, B, D, F–H.
Material.—Several tens of calcium phosphatic internal moulds, including SMNH X11215–11220, from samples 21/33, 21/46.1, 21/51, 21/52, 21/54, Tyuser Formation, Dokidocyathus lenaicus–Tumuliolynthus primigenius Zone, Tommotian stage–Delgadella anabara Zone, Atdabanian stage (upper Stage 2–Stage 3), 22/50, Sekten Formation, Lena River, upper Toyonian stage (correlated with the Cambrian Stage 4) and from 20/1B, 19/12.75, 19/25.5, 19/33, 19/55, Erkeket Formation, Khorbusuonka River, Botoman–Amgan stages (correlated with the Cambrian Stage 4–lower Wuliuan Stage). Siberia, Russia.
Description.—Internal moulds representing straight moderately expanding conchs. In dorsal view, the apical angle is ca. 30°. Cross-section rounded triangular. Aperture orthogonal with a long semi-circular ventral ligula, dorsal and a pair of lateral sinuses. Dorsal side arched, with flattened flanks, passes with angulation into a flattened ventral side. Aperture with slightly protruding ventral side. Larger internal moulds with a flared aperture suggesting presence of lateral sinuses in the conch. Initial part bulbous separated from the main part by a shallow constriction, flattened dorso-ventrally, slightly curved dorsally and carrying a pointed tip.
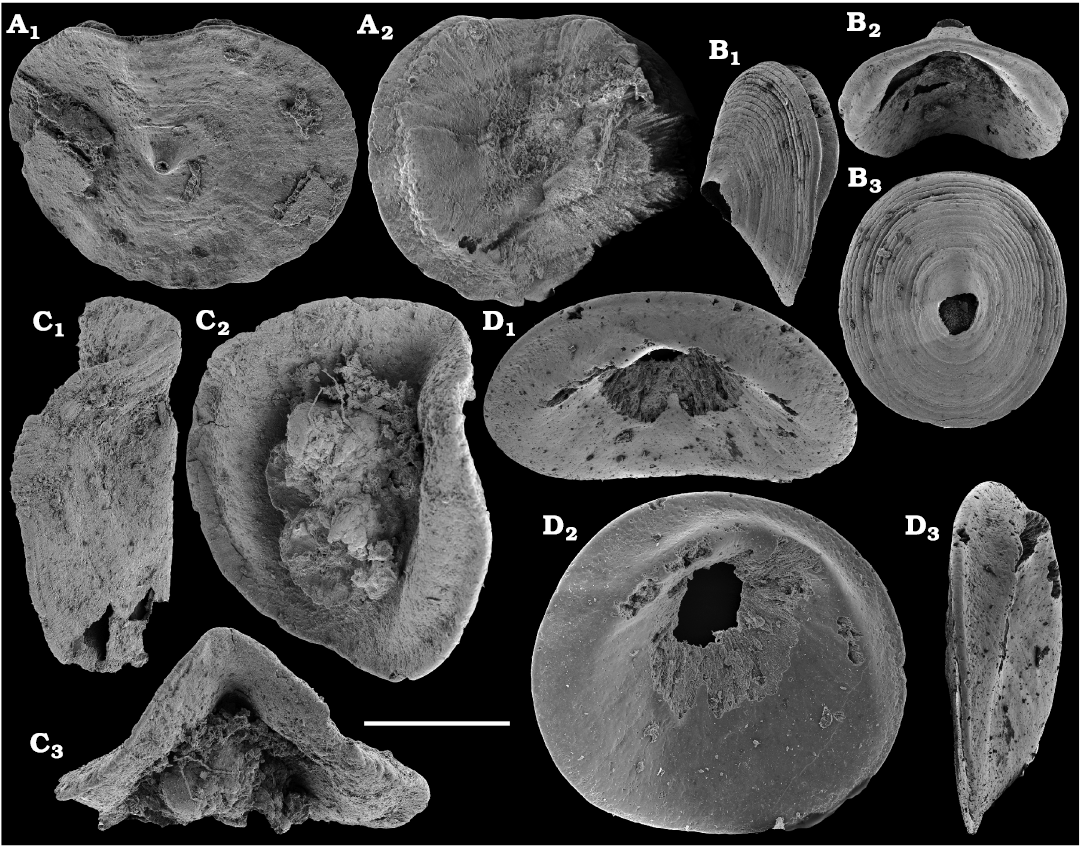
Fig. 50. Hyolith opercula from the lower Cambrian Erkeket Formation, Khorbusuonka River (A, C, D) and Tyuser Formation, lower reaches of the Lena River (B), Siberia, Russia; samples 11/16.05 (A), 11/2B (C, D), and 21/21 (B). A–D. Phosphatised orthothecid opercula of four different types, SMNH X11203–11206, respectively. A1, B3, outer surface; A2, D2, inner surface; B1, C1, D3, lateral view; B2, ventral view; C2, oblique inner surface; C3, D1, dorsal view. Scale bar: 625 µm (D), 500 µm (B), and 250 µm (A1, A2, C).
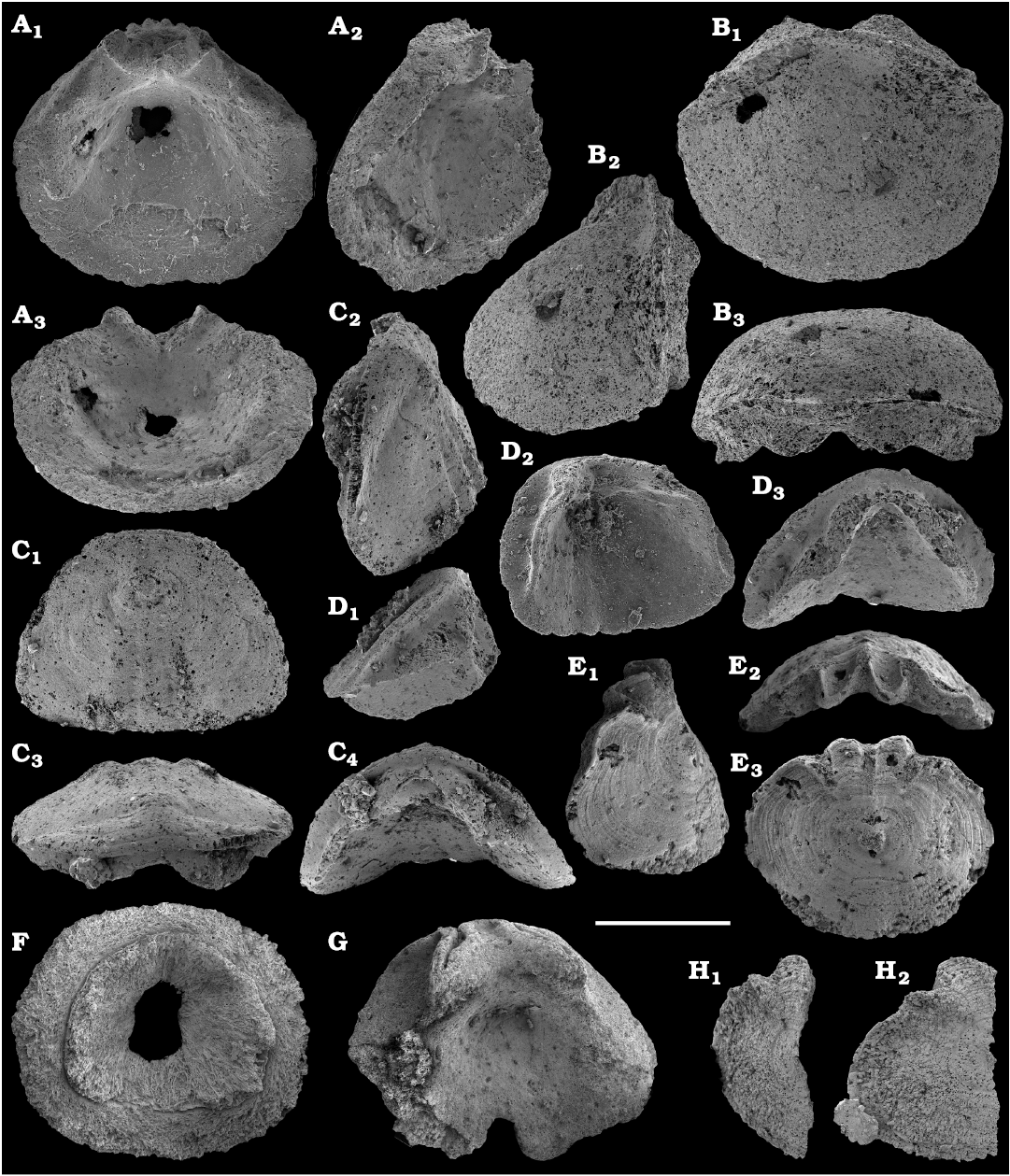
Fig. 51. Phosphatised hyolith opercula from the lower Cambrian Sekten Formation, lower reaches of the Lena River (A–D) and Erkeket Formation, Khorbusuonka River (E–H), Siberia, Russia; samples 19/10.25 (E, H), 19/11.75 (F, G), and 22/50 (A–D). A, B. Parkula bounites Bengtson in Bengtson et al., 1990, SMNH X11207 and 11208, respectively. C, D. Hyolithid operculum 1, SMNH X11209 and X11210, respectively. E, H. Triplicatella sp., SMNH X11211 and X11212, respectively. F. Majatheca tumefacta Missarzhevsky in Rozanov et al., 1969, SMNH X11213. G. Triplicatella papilio Kouchinsky in Kouchinsky et al., 2015, SMNH X11214. A1, D2, inner surface; B1, C1, E3, H2, outer surface; A2, oblique lateral; B2, C2, D1, E1, H1, lateral; C3, E2, dorsal; and A3, C4, ventral views. Scale bar 500 µm.
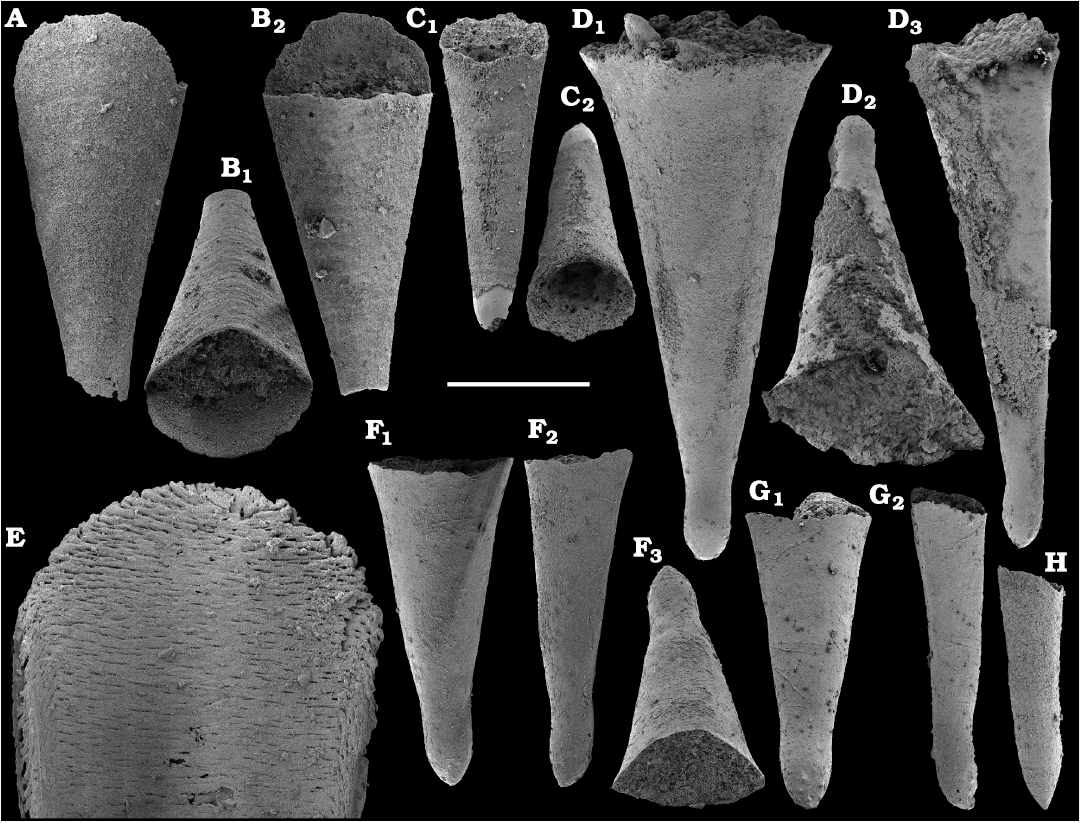
Fig. 52. Hyolith conchs from the lower Cambrian Erkeket Formation, Khorbusuonka River (C, D, F–H) and Tyuser (A, B) and Sekten (E) formations, lower reaches of the Lena River, Siberia, Russia; samples 19/12.75 (D, F, G), 19/25.5 (H), 19/33 (C), 21/51 (A, B), and 22/50 (E). A, B, D, F–H. Microcornus cf. eximius Duan, 1984, phosphatised conchs (A, B) and calcium phosphatic internal moulds (D, F, G), SMNH X11215–11220, respectively. C, E. Parkula bounites? Bengtson in Bengtson et al., 1990, phosphatised conchs, SMNH X11221–11222, respectively. B1, C2, D2, F3, oblique apertural, B2, C1, D1, F1, G1, dorsal; and D3, F2, G2, lateral views; E, enlargement showing inner surface of ventral side of a fragmentary conch. Scale bar: 500 µm (A–D, F–H) and 250 µm (E).
Remarks.—Microcornus eximius Duan, 1984, is most similar but its conch has a well-developed median sinus, which is not evident on the internal moulds herein and weakly expressed in the available phosphatised conchs (Fig. 52A, B). Flared lateral parts at the aperture of internal moulds suggesting presence of lateral sinuses in the conch. The form is similar to Hyolithida indet. 3 (Kouchinsky et al. 2011: fig. 11G–I), from sample 8/32, Drumian Stage of the Anabar Uplift. It is distinguished from Microcornus sp. (Fig. 52C) by a larger apical angle. The latter form is in its turn similar to Microcornus sp. described by Kouchinsky et al. (2011: fig. 12L–Q) preserved as silicified conchs from sample 8/18.5, from lower Wuliuan Stage beds of the Kuonamka Formation. Such a morphology can also be attributed to Parkula bounites, reported herein from opercula. M. eximius is also reported from Series 2 of South China (Duan 1984), North China (Pan et al. 2019), Australia (Bengtson et al. 1990), Antarctica (Wrona 2003), and Laurentia (Malinky and Skovsted 2004).
Genus Parakorilithes He and Pei in He et al., 1984
Type species: Parakorilithes mammillatus He and Pei in He et al., 1984; lower Cambrian; North China.
Parakorilithes mammillatus He and Pei in He et al., 1984
Fig. 53.
Material.—Several phosphatised conchs, including figured SMNH X11223–11225, and 11351, from 19/10.25 and 19/11.75; Khorbusuonka River, Erkeket Formation, Siberia, Russia. Lower Botoman stage (correlated with the lower part of Cambrian Stage 4).
Description.—Moderately expanding conch gently curved in sagittal plane, with an oval aperture showing protruding dorsal and ventral margins (lips). The dorsal lip is longer than the ventral one. Ventral and dorsal sides of the conch covered with growth lines curved towards the aperture following curvature of the lips. Laterally, the growth lines show angulation between the ventral and dorsal curves. Initial part rounded blunt slightly separated from the main conch by a shallow constriction.
Remarks.—Pan et al. (2019) emended diagnosis of the genus and considered the species as a senior synonym to Hyptiotheca karraculum Bengtson in Bengtson et al. 1990. Bengtson et al (1990) interpreted the dorsal side in this form as shorter than the ventral one and the sagittal ventrally directed curvature of the conch. The curvature is herein reinterpreted as the normal one, dorsally directed, but with the dorsal lip longer than the ventral one.
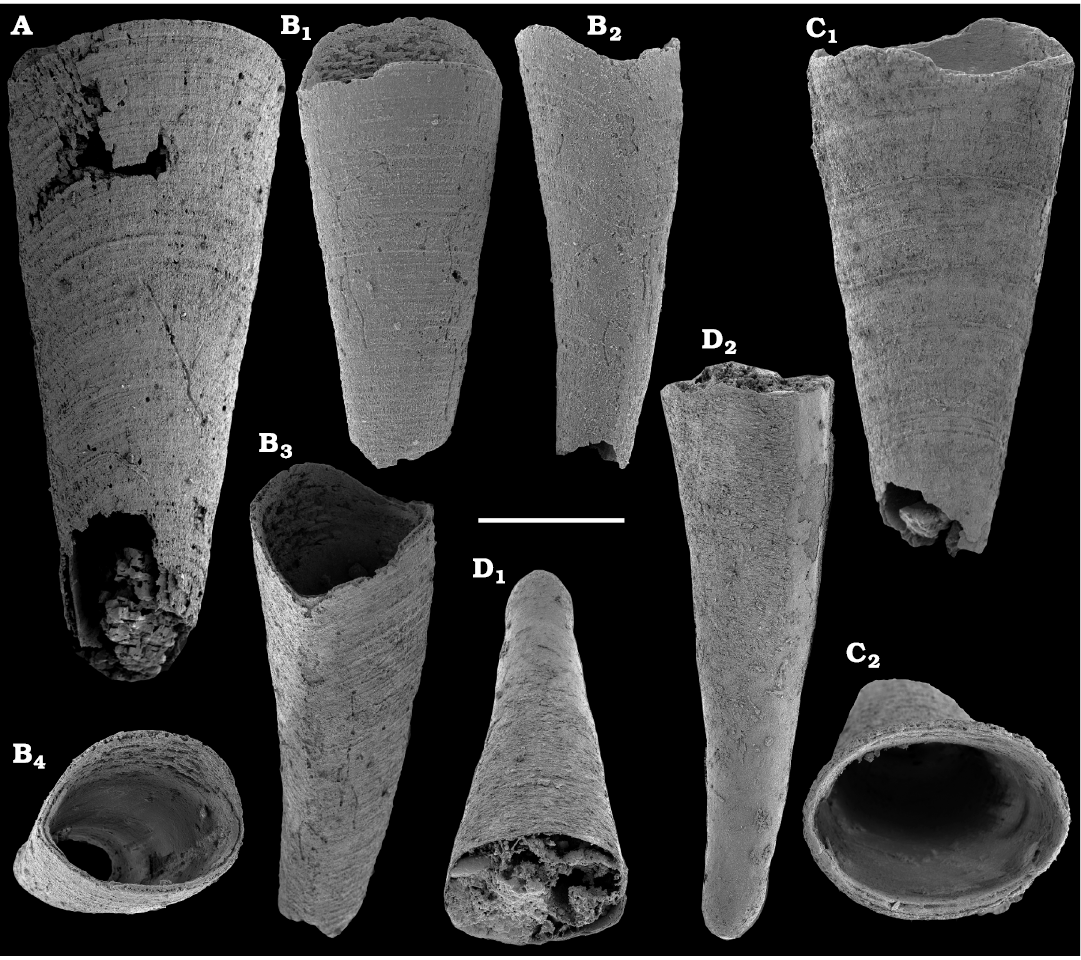
Fig. 53. Hyolith conchs Parakorilithes mammillatus He and Pei in He et al., 1984, from the lower Cambrian Erkeket Formation, Khorbusuonka River, Siberia, Russia; sample 19/10.25. A–D. Phosphatised conchs, SMNH X11223–11225, and 11351 respectively. B1, C1, dorsal; B2, D2, lateral; B3, oblique lateral; C2, B4, apertural; and D1, oblique apertural views. Scale bar 250 µm.
Stratigraphic and geographic range.—Series 2 of Australia (Bengtson et al. 1990; Gravestock et al. 2001), Antarctica (Wrona 2003), North China (Pan et al. 2019), and Laurentia (Malinky and Skovsted 2004).
Genus Parkula Bengtson in Bengtson et al., 1990
Type species: Parkula bounites Bengtson in Bengtson et al., 1990; Abadiella huoi Zone, lower Cambrian; Kulpara, Yorke Peninsula, Australia.
Parkula bounites Bengtson in Bengtson et al., 1990
Figs. 51A, B, 52C, E.
Material.—Several phosphatised opercula and probably conchs, including figured SMNH X11207, 11208, 11221, and 11222, from sample 19/33, Erkeket Formation, Khorbusuonka River, and samples 22/50 and 22/67, Sekten Formation, Lena River. Siberia, Russia; upper Toyonian and lower Amgan stages (correlated with the Cambrian Stage 4–lowermost Wuliuan Stage).
Description.—Operculum broadly oval in the plan view, with a crescent-shaped dorsal lobe (cardinal shield) and convex semi-circular ventral lobe (conical shield). The dorsal margin is inclined at ca. 60° with respect to the ventral lobe. Initial part flattened. The dorsal lobe bears short blade-like clavicles situated along the margin between two lobes at edge of conical shield. A single pair of short cardinal processes diverging at ca. 90°.
Remarks.—Fragments of the hyolithid conchs with semi-circular ventral lip occurring in sample 22/50 (Fig. 53E) can be attributed to the same species.
Stratigraphic and geographic range.—Cambrian Series 2 of Australia (Bengtson et al. 1990; Gravestock et al. 2001), Antarctica (Wrona 2003), North China (Pan et al. 2019), Laurentia (Malinky and Skovsted 2004; Skovsted 2006b; Skovsted and Peel 2007; Devaere et al. 2019), and Siberia (Kouchinsky et al. 2015a).
Hyolithid operculum 1
Fig. 51C, D.
Material.—Two phosphatised opercula, SMNH X11209 and 11210, from sample 22/50, Sekten Formation, Lena River, Siberia, Russia. Upper Toyonian stage (correlated with the Cambrian Stage 4).
Description.—Operculum rounded trapezoidal in plane view, with a narrow semi-circular dorsal lobe and convex trapezoidal ventral lobe. The dorsal lobe bears short rounded blade-like cardinal processes. Clavicles indistinct. Three distinct radial folds diverge slightly from the apex towards ventral margin being bilaterally symmetrically arranged with respect the sagittal plane of the operculum.
Hyolithid operculum 2
Fig. 54.
Material.—Ten phosphatised opercula, including figured SMNH X11226–11229, from sample 19/33, Erkeket Formation, Khorbusuonka River, Siberia, Russia. Botoman stage (correlated with the Cambrian Stage 4).
Description.—Operculum almost circular in frontal view and domal in side view. Apex displaced towards the dorsal margin at ca. 1/4 of the diameter of operculum. The outer surface of the operculum has thin continuous growth lines. Its ventral part forms a radial fold producing a sinus of the ventral margin of the operculum. A smooth crescent-shaped lobe forms the inner side of the dorsal margin. There is a deep concavity on the inner side towards the apex of the operculum. The ventral and lateral margins produce a semi-circular elevated ridge with smooth exterior steeply sloping inwards and outwards.
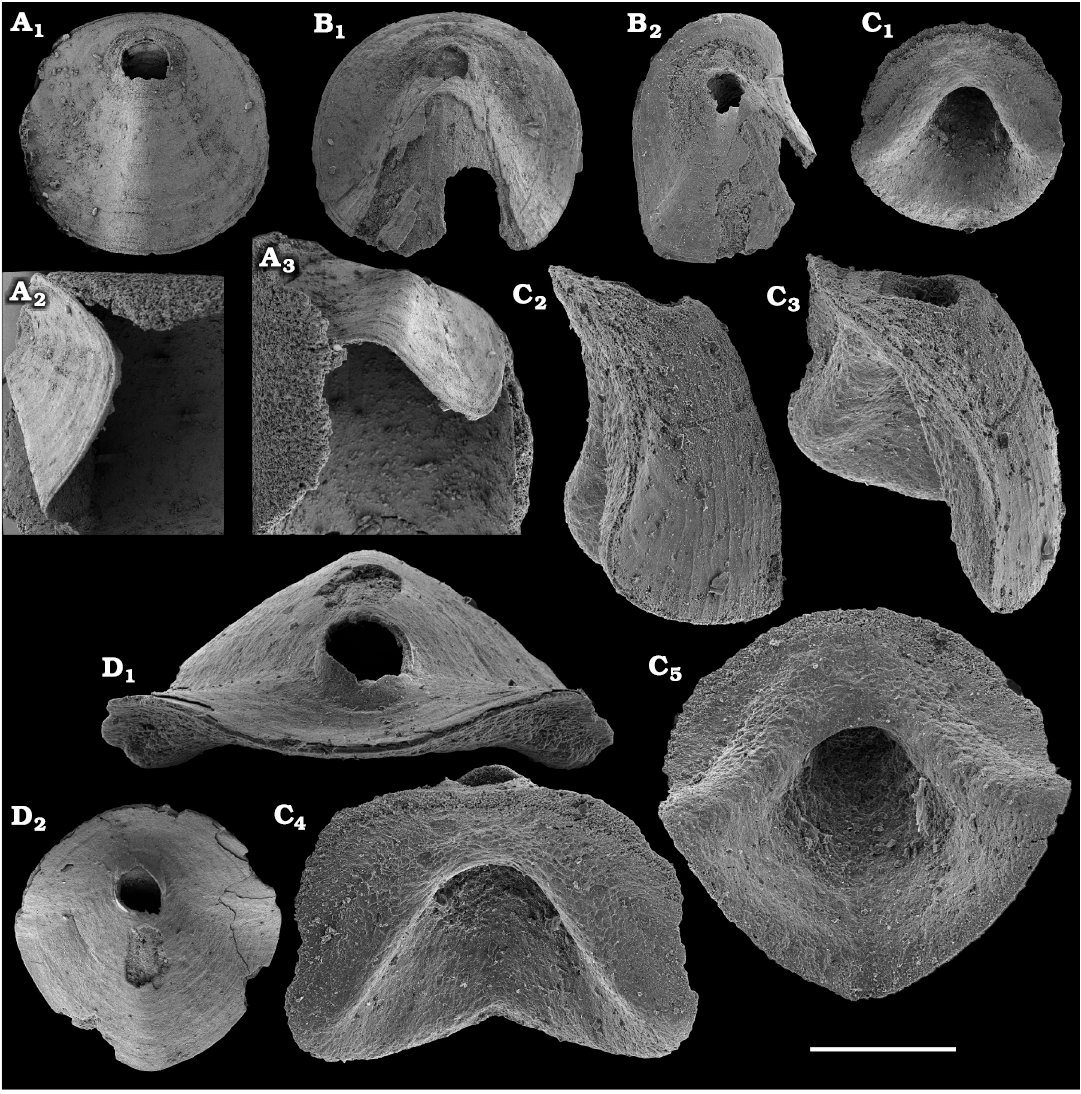
Fig. 54. Hyolithid operculum 2 from the lower Cambrian Erkeket Formation, Khorbusuonka River, Siberia, Russia; sample 19/33. A–D. SMNH X11226–11229, respectively. A1, B1, D2, outer surface; A2, C2, lateral; A3, ventral; B2, oblique outer surface C1, C5, inner surface; C3, oblique lateral; and D1, C4, dorsal views. Scale bar: 250 µm (A, B, C1, D2) and 125 µm (C2–C5, D1).
Hyolithid helens
Fig. 55.
Material.—Over ten phosphatised hyolithid appendages, including figured SMNH X11230–11235, from samples 19/10.25 and 20/1B, Erkeket Formation, Khorbusuonka River, Protolenus–Nelegeria Zone, Botoman stage (correlated with the Cambrian Stage 4), sample 22/50, Sekten Formation, Lena River, Toyonian stage (correlated with the Cambrian Stage 4) and a single fragment of proximal part of a helen from sample 21/21, Tyuser Formation, Dokidocyathus regularis Zone, Tommotian stage (correlated with the upper part of Cambrian Stage 2), Lena River. Siberia, Russia.
Remarks.—The appendages from samples 19/10.25 and 20/1B probably belong to the same species. They are elongated blade-like, somewhat helically twisted around longitudinal axis, with extended elliptical cross-section. They consist of distal part ornamented with continuous transversal terraces and smooth basal part, the latter is ca. 1/5 of the total appendage length. The terraces are inclined towards the distal end (see Kouchinsky et al. 2015a: fig. 32 and Skovsted et al. 2020: fig. 2). The fossil contains radially fanning longitudinally oriented fibers reminiscent to those in hyoliths described herein, by Kouchinsky et al. (2015a: fig. 32) and Skovsted et al. (2020). Appendage from sample 22/50 is smooth, probably due to preservation. A fragment from sample 21/21 is questionably interpreted as a helen. It derives from the Tommotian stage and co-occur with circular opercula having a pair of radial folds between dorsal and ventral parts (Fig. 47).
Phylum Chaetognatha Leuckart, 1854
Class, order, and family uncertain
Remarks.―Two orders, Protoconodontida and Pseudoconodontida are proposed by Landing (1995) as histologically distinct phyletic/taxonomic groups to encompass organo-phosphatic spiniform sclerites described herein. The order Protoconodontida Landing, 1995, with such forms as Amphigeisina danica (Poulsen, 1966), Gapparodus bisulcatus (Müller, 1959), and Hagionella cultrata (Missarzhevsky, 1977) among others, includes, according to Landing (1995), sclerites that grew by basal-internal accretion of microcrystalline laminae. By contrast, the order Pseudoconodontida Landing, 1995, is characterised by fibers aligned subparallel with the surface of the wall, such as those in Protohertzina Missarzhevsky, 1973. A fibrous structure of the wall (arranged in growth lamellae) was later reported from G. bisulcatus and H. cultrata (Kouchinsky et al. 2011, 2015a) and contrasts with the lamellar microcrystalline structure of the wall in A. danica (see Kouchinsky et al. 2011 and herein). Pseudoconodontida originally refer to the same group (sensu Landing 1995) other histologically and morphologically different body-surface sclerites (not grasping spines) of problematic Fomitchella (see Kouchinsky et al. 2017) and Rhombocorniculum (see e.g., Kouchinsky et al. 2015a). It appears therefore that the orders Protoconodontida and Pseudoconodontida are neither histologically distinct nor monophyletic and cannot be used herein in their original formal meaning.
Genus Gapparodus Abaimova, 1978
Type species: Hertzina? bisulcata Müller, 1959; upper Cambrian; Sweden.
Gapparodus bisulcatus (Müller, 1959)
Fig. 56A, C, D.
Material.—Over twenty calcium phosphatic sclerites, including figured SMNH X11236–11238, from samples 19/2B, 19/3B, 19/5B, 19/55, 19/65.5, 19/66.5, upper Erkeket and Kuonamka formations, Khorbusuonka River, Siberia, Russia. Amgan and lower Mayan stages (correlated with the upper part of Cambrian Stage 4–lower Drumian Stage).
Remarks.—Slender, gently curved bilaterally symmetrical calcium phosphatic hollow sclerites with an oval or figure-of-eight shaped cross-section extended in sagittal plane. Two longitudinal lateral furrows divide the sclerite into two halves having almost the same width. One with a strongly convex surface and convex in lateral view, whereas the opposite one with a gently convex or flattened surface. Wall with lamellar fibrous composition.
Stratigraphic and geographic range.—Globally distributed and known from Siberia in Cambrian Stage 4 and Wuliuan Stage (Müller et al. 1995; Kouchinsky et al. 2011, 2015a), ranges from Series 2 through the Furongian Series in South and North China (Dong and Bergström 2001; Babcock et al. 2007; Steiner et al. 2007), Baltica (Bengtson 1976; Müller and Hinz 1991), Gondwana (Özgül and Gedik 1973; Lee 1975), Laurentia (Clark and Miller 1969; Clark and Robison 1969; Landing 1974; Peel et al. 2016), and Kazakhstan (Abaimova and Ergaliev 1975).
Genus Protohertzina Missarzhevsky, 1973
Type species: Protohertzina anabarica Missarzhevsky, 1973; Anabarites trisulcatus Zone, lower Cambrian (lower Fortunian Stage); mouth of the Kotujkan River, Siberia, Russia.
Protohertzina sp.
Fig. 56B.
Material.—Single phosphatic sclerite, SMNH X11240 from sample 11/2B, basal Erkeket Formation, Khorbusuonka River, Siberia, Russia. Dokidocyathus regularis Zone, Tommotian stage (correlated with the upper part of Cambrian Stage 2).
Description.—Bilaterally symmetrical, spine-shaped apatitic sclerite, ca. 1.5 mm long fragment, gently curved in the sagittal plane, with oval cross-section and internal cavity. Convex side of sclerite is rounded and smooth. Opposite, longitudinally concave side of sclerite flattened, separated from the convex side by distinct lateral angulations. Wall composed of multiple layers of fibers longitudinally oriented on outer surface of sclerites.
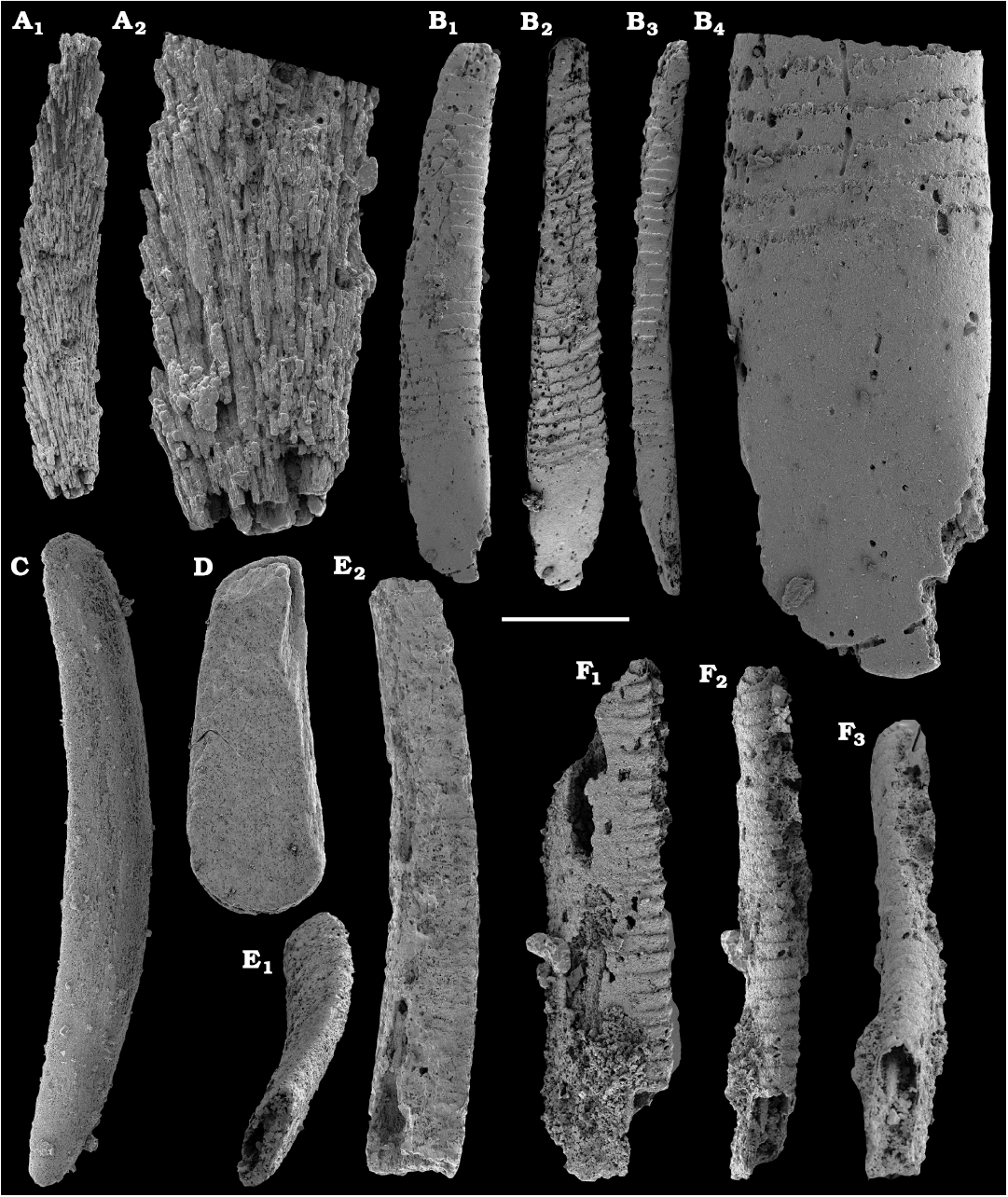
Fig. 55. Hyolithid helens from the lower Cambrian Erkeket Formation, Khorbusuonka River (A, B, E, F) and Tyuser (D) and Sekten (C) formations, lower reaches of the Lena River, Siberia, Russia; samples 19/10.75 (A, B, E), 20/1B (F), 21/21 (D), and 22/50 (C). A–F. Phosphatized hyolithid appendages (helens), SMNH X11230–11235, respectively. A2, enlargement of A1 showing proximal end; B1, upper surface; B2, oblique view on the upper and frontal (left side in B1) surfaces; B3, view on the rear margin of the helen (right side in B1); B4, close-up of proximal portion; E1, oblique lateral view; E2, F1, plain view; F2, F3, view on the margin (left side in F1). Scale bar: 500 µm (C, D), 250 µm (A1, B1–B3, E, F), 80 µm (A2, B4).

Fig. 56. Sclerites of protoconodonts (A–E) and problematic spines (F–I) from the early and middle Cambrian, Khorbusuonka River, Siberia, Russia; samples 11/2B (B), 19/55 (E), and 19/61 (F–I) from the Erkeket Formation and 19/3B (C) and 19/5B (A, D) from the Kuonamka Formation. A, C. Gapparodus bisulcatus (Müller, 1959), SMNH X 11236 and X11237, respectively. B. Protohertzina sp., SMNH X11240. D. Sclerite tentatively assigned to Gapparodus bisulcatus (Müller, 1959), SMNH X11238. D1, view on the concave side; D2, lateral view. E. Amphigeisina danica (Poulsen, 1966), SMNH X11239. F–I. Calcium phosphatic internal moulds of problematic spines with corrugated walls, SMNH X11241–11244, respectively. Scale bar: 500 µm (A–E) and 250 µm (E–I).
Remarks.—The sclerite is incompletely preserved and its exact affinity is difficult to determine given morphological variability in Protohertzina and absence of basal part.
Phylum, class, order, and family uncertain
Genus Amphigeisina Bengtson, 1976
Type species: Hertzina? danica Poulsen, 1966; lower middle Cambrian; Bornholm, Danmark.
Amphigeisina danica (Poulsen, 1966)
Fig. 56E.
Material.—Three fragmentary calcium phosphatic sclerites, including figured SMNH X11239, from samples 19/55 and 19/2B, upper Erkeket Formations, Khorbusuonka River, Siberia, Russia. Ovatoryctocara–Schistocephalus and Kounamkites zones, Amgan stage (correlated with the uppermost Stage 4–Wuliuan Stage).
Remarks.—Fragments of slender, gently curved bilaterally symmetrical hollow sclerites with a relatively narrow concave side separated from the opposite convex side by longitudinal carinated angulations. Cross-section broad crescent-shaped. Outer surface smooth. Despite lamination in the wall, growth lines cannot be distinguished, nor fibrous composition of the wall (Bengtson 1976; Kouchinsky et al. 2011), by contrast with such other forms of protoconodonts described from Siberia and elsewhere as Protohertzina, Gapparodus, Gumella, Hagionella, and Phakelodus. Such a difference in biomineralisation suggests that Amphigeisina does not belong to the same clade of animals but probably nektonic ecdysozoans, although its affinity is pending further study.
Stratigraphic and geographic range.—Upper Botoman stage through the upper Kounamkites Zone, Amgan stage (correlated with the Cambrian Stage 4–Wuliuan Stage; Kouchinsky et al. 2011, 2015a, herein), Siberia (Russia). Cambrian Series 2 to 3 of South and North China (Qian et al. 2004), Avalonia (Great Britain; Hinz 1987), Laurentia (Landing 1974; Peel et al. 2016), Baltica (Poulsen 1966; Bengtson 1976), and Kazakhstan (Gridina 1991).
Problematic spines
Fig. 56F–I.
Material.—Over ten calcium phosphatic internal moulds and fragments, including figured SMNH X11241–11244, from sample 19/61, uppermost Erkeket Formation, Khorbusuonka River, Siberia, Russia. Kounamkites Zone, Amgan stage (correlated with the Wuliuan Stage).
Remarks.—Internal moulds of straight and longitudinally flattened spines with a rounded to drop-shaped cross-section of the lumen. Walls of the spines were transversely plicated more or less regularly by rugae of varying amplitude. Corrugation appears to be more prominent along one of the narrower sides of the spine. They are asymmetrical and one of the wider sides can be more flattened than the other one, suggesting a dorso-ventral difference in morphology. The fossils can be interpreted as internal moulds of spines of trilobites or other ecdysozoan animals.
Superphylum Ecdysozoa Aguinaldo, Turbeville, Linford, Rivera, Garey, Raff, and Lake, 1997
Group Scalidophora Lemburg, 1995
Phylum uncertain
Class Palaeoscolecida Conway Morris and Robison, 1986
Order Palaeoscolecida Conway Morris and Robison, 1986
Family Palaeoscolecidae Whittard, 1953
Genus Hadimopanella Gedik, 1977
Type species: Hadimopanella oezgueli Gedik, 1977; middle Cambrian; Taurus Mountains, Turkey.
Hadimopanella knappologica (Bengtson, 1977)
Fig. 57.
Material.—Hundreds of individual calcium phosphate sclerites, including SMNH X11245–11256, from samples 19/5.5 and 20/1B, Erkeket Formation, Khorbusuonka River, Siberia, Russia. Lower Botoman stage (correlated with the Cambrian Stage 4).
Description.—Disk-like, 100–300 μm in diameter, circular to broadly oval sclerites with a gently convex smooth lower side and the upper side representing the conical surface capped with a flattened nodular face. The latter includes numerous tubercles (nodes), ca. 10 μm in diameter. The outermost tubercles form a ring, whereas other tubercles are less orderly distributed inside the ring. The conical surface carries radially oriented folds situated between the outer margin of sclerite and the outermost ring of tubercles of the nodular face.
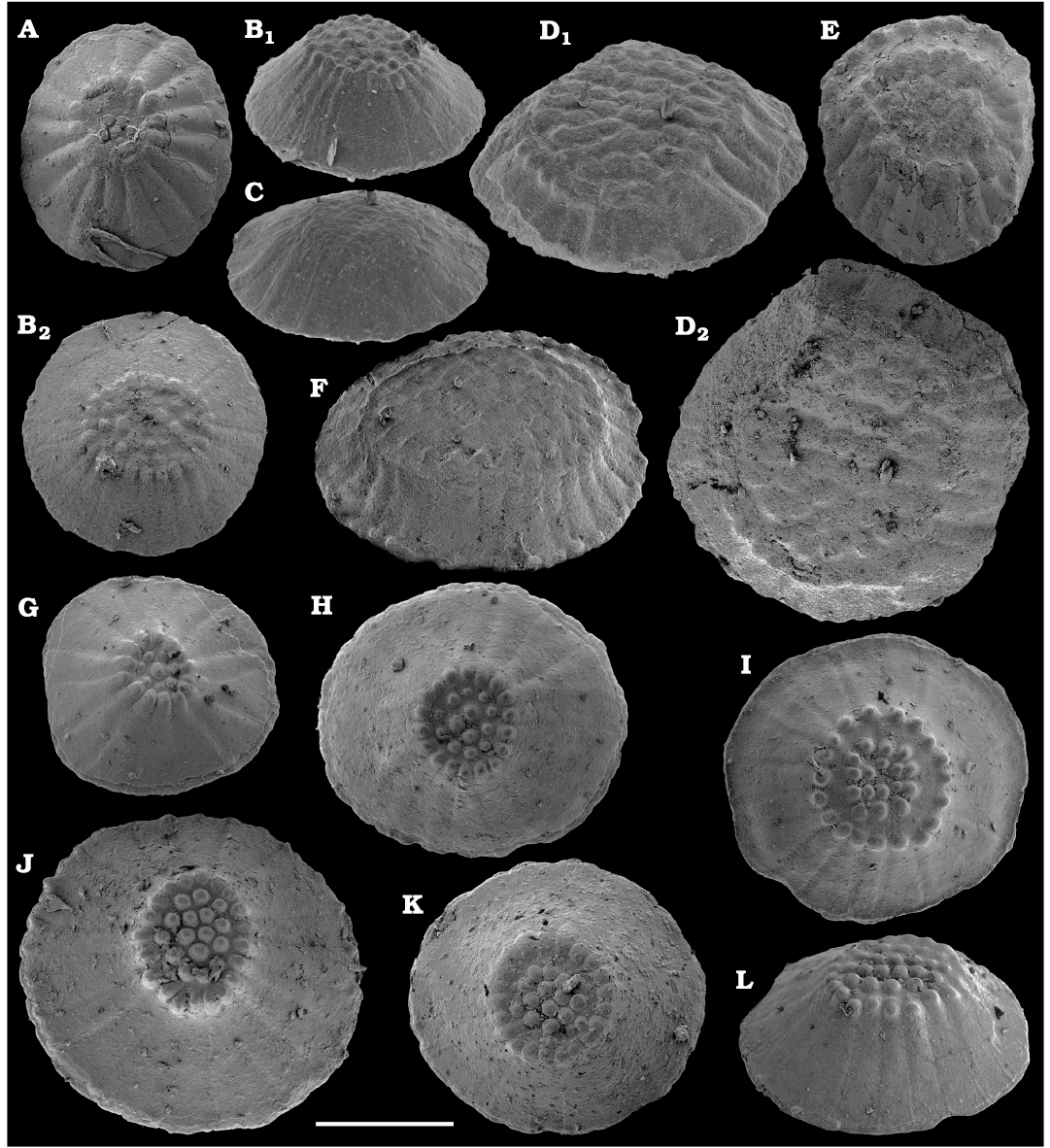
Fig. 57. Paleoscolecid Hadimopanella knappologica (Bengtson, 1977) from the lower Cambrian Erkeket Formation, Khorbusuonka River, Siberia, Russia; samples 19/5.5 (A–F) and 20/1B (G–L). A–L. Disarticulated sclerites, SMNH X11245–11256, respectively. A, B2, D2, E–K, apical view; B1, C, D1, L, oblique apical view. Scale bar 100 µm.
Remarks.—The holotype of the species shows numerous nodes on the nodular face and distinct radial folds on the conical surface (Bengtson 1977). The number of nodes can vary but presence of radial folds remains a diagnostic feature of the species. Type material occurs in the Judomia–Uktaspis (Prouktaspis) Zone, Atdabanian Stage, Perekhod Formation, middle Lena River (Astashkin et al. 1991). Previously described range of the fossil from the Emyaksin Formation is correlated with the lower Judomia–Uktaspis (Prouktaspis) Zone, Atdabanian stage–Calodiscus–Erbiella Zone, lower Botoman stage. Forms described by Kouchinsky et al. (2011: fig. 32) from the Kounamkites Zone, Amgan stage (correlated with the Wuliuan Stage) are reinterpreted herein as Hadimopanella sp. (see below). Only those sclerites figured by Kouchinsky et al. (2015a: fig. 57G–N) from the Judomia–Uktaspis (Prouktaspis) Zone, Atdabanian stage with diagnostic radial folds are interpreted herein as H. knappologica. Those with a single apical tip (Kouchinsky et al. 2015a: fig. 57B, E) in the middle of conical side are similar to H. apicata Wrona, 1982, from the upper Stage 3–Stage 4 of Gondwana and Laurentia. The latter species has sclerites up to 150 μm in diameter and up to four nodes in larger specimens.
Stratigraphic and geographic range.—Judomia–Uktaspis (Prouktaspis) Zone, Atdabanian stage–Calodiscus–Erbiella Zone, lower Botoman stage (correlated with the upper part of Cambrian Stage 3–lower Cambrian Stage 4); Siberian Platform.
Hadimopanella foveata Kouchinsky sp. nov.
Fig. 58.
Zoobank LCID: urn:lsid:zoobank.org:act:1BC57845-B56E-467B-9A C6-BE6E97BD5BC9
Etymology: From Latin fovea, pit, refers to the pitted upper face of sclerites.
Type material: Disarticulated calcium phosphatic sclerites: Holotype SMNH X11257 (Fig. 58A) and paratypes SMNH X11258–11260, 11262–11265 (Fig. 58B–D, F–I), from the type locality and horizon.
Type locality: Locality 19/31.75 (71°28.8’N, 123°50.0’E) on the left bank of the Khorbusuonka River, ca. 20 km downstream of the mouth of the Mattaia River, northern Siberia, Russia.
Type horizon: Erkeket Formation, ca. 30 m below the Kuonamka Formation, Toyonian Stage (correlated with the Cambrian Stage 4).
Material.—Several hundred calcium phosphate sclerites, in addition to the type series, from samples 19/31.75, 19/33, 19/40, 19/43.25, 19/46.5, 20/5B, 20/6B, Khorbusuonka River, Siberia, Russia. Toyonian–lower Amgan stages (correlated with the Cambrian Stage 4).
Diagnosis.—Almost circular disk-like calcium phosphatic sclerites with a flattened circular upper face bearing closely situated pits. The latter are arranged in a circlet at the margin of the upper face.
Description.—Disk-like, almost circular sclerites, 100–200 μm in diameter, with a gently convex smooth lower side. The upper side represents a conical surface capped with a flattened circular upper face with closely situated pits, ca. 10 μm in diameter. The outermost part of the upper face represents a ring of 5–12 pits, whereas pits of the same type inside the ring are less orderly distributed. Conical surface carries radially oriented furrows and intervening folds situated at the margin of sclerite and granulation surrounding the upper face. The number of pits increases with size of sclerites.
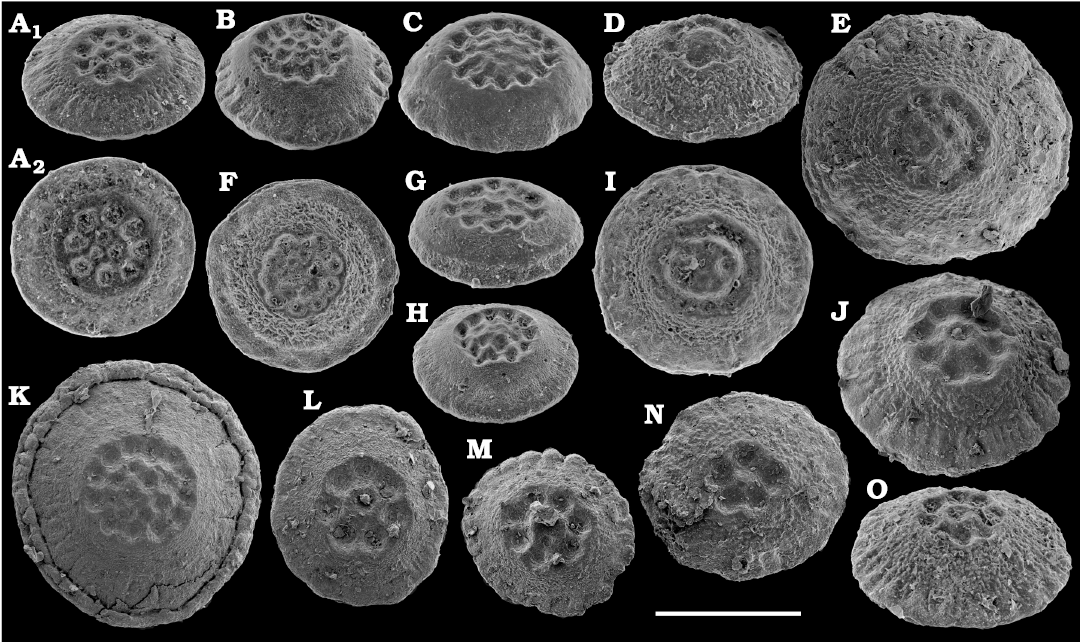
Fig. 58. Paleoscolecid Hadimopanella foveata sp. nov., from the lower Cambrian Erkeket Formation, Khorbusuonka River, Siberia, Russia; samples 19/31.75 (A–D, F–I), 19/43.25 (E, J, N, O), 20/5B (L, M), and 20/6B (K). A–O. Disarticulated sclerites, SMNH X11257–11268, 11352–11354, respectively. A, holotype. A1, B–D, G, H, O, oblique apical view; A2, E, F, I–N, apical view. Scale bar 100 µm.
Stratigraphic and geographic range.—Toyonian–lower Amgan stages (correlated with the Cambrian Stage 4), Siberian Platform, Russia.
Hadimopanella spp.
Fig. 59.
Material.—Several hundred calcium phosphate sclerites, including SMNH X11269–11292, from samples 19/26.75, 19/31.75, 19/33, 19/40, 19/43.25, 19/46.5, 19/55, 20/5B, 20/6B, Erkeket Formation, Toyonian–Amgan stages (correlated with the Cambrian Stage 4–Wuliuan Stage), 19/65.5, 19/66.5 from the Kuonamka Formation and 19/70.5 from the Yunkyulyabit-Yuryakh Formation, lower Mayan Stage (correlated with the lower Drumian Stage). Khorbusuonka River, Siberia, Russia.
Description.—Disk-like, 100–250 μm in diameter, almost circular or ovaliform sclerites with a gently convex smooth lower side and the upper side representing the conical surface capped with a nodular face with nodes 10–15 μm in diameter. The nodular face with an oval outline ornamented by up to 40 tightly arranged nodes (Fig. 59A–E) or having a circular outline ornamented by up to 30 tightly arranged nodes (Fig. 59J–R, T, U, X). In some of the sclerites a circlet of nodes surrounds a central group of 1–4 nodes (Fig. 59K, L, T, X). Conical surface carries a marginal rim with concentric zones (see Fig. 59A–D) and short radial plication with faint radial folds and furrows in the periphery (see Fig. 59J–L, N, V) as well as covered with fine granulation (Fig. 59T–V). Marginal zone of sclerite can be very narrow in some forms (Fig. 59O–R). Conical surface can be displaced, with nodes inclined in the same direction (Fig. 59G–I, S, W).

Fig. 59. Paleoscolecids Hadimopanella spp. from the Cambrian Erkeket (A–L, N–X) and Yunkyulyabit-Yuryakh (M) formations, Khorbusuonka River, Siberia, Russia; samples 19/31.75 (B, H–K, N, P–W), 19/43.25 (A, E–G, L, O), 19/70.5 (M), 20/5B (C, X), and 20/6B (D). A–X. Disarticulated sclerites, SMNH X11269–11292, respectively. A–D, F, G1, L, M2, S–V, X, apical; E, G2, K, M1, N–P, R, W2, oblique apical; Q, oblique basal; and W1, lateral views. Scale bar 100 µm.
Phylum, class, order, and family uncertain
Remarks.—Markuelia is included in an informal supraphyletc group Scalidophora Lemburg, 1995 (Dong et al. 2010).
Genus Markuelia Val’kov, 1983
Type species: Markuelia secunda Val’kov in Val’kov and Karlova, 1984; Basal Pestrotsvet Formation, Nochoroicyathus sunnaginicus Zone, Tommotian stage (correlated with the upper part of Cambrian Stage 2); Altitude 1291 m, Gonam River (Aldan River Basin), southeastern part of the Siberian Platform, Russia.
Markuelia secunda Val’kov in Val’kov and Karlova, 1984
Fig. 60.
Material.—Single specimen, SMNH X11293, from sample 11/16.05, basal Erkeket Formation, Khorbusuonka River, Siberia, Russia. Dokidocyathus regularis Zone, Tommotian stage (correlated with the upper part of Cambrian Stage 2).
Remarks.—The fossil represents a phosphatised enrolled embryo (as described by Bengtson and Yue, 1997), whose worm-like segmented body is tightly coiled into a sphere, ca. 0.5 mm in diameter. Although tail spines are not revealed, as in the previously described material, presence of short, conical spines on trunk annuli suggests affinity with Markuelia secunda Val’kov in Val’kov and Karlova, 1984 (see Dong et al. 2010). Annuli are ca. 30 µm thick at the middle. The spines are 30–40 µm long. Different species of Markuelia reportedly occurs in the Cambrian Stage 2 of Siberia, Miaolingian Series of China and Australia, Furongian Series of China, and Lower Ordovician of Laurentia (Dong et al. 2010).
Globula of unknown affinity, 0.2–0.3 mm in diameter is found in the Kuonamka Formation, lower Mayan Stage beds (sample 19/5B, correlated with the lowermost Drumian Stage). It is phosphatised and deformed into a tri-lobate shape (Fig. 69J). The fossil is identical to the globulae reported from the Tommotian and Atdabanian Stages of the Anabar Uplift (Kouchinsky et al. 2015a: fig. 73B).
Stratigraphic and geographic range.—Markuelia secunda is known from the upper part of Cambrian Stage 2, Siberian Platform, Russia.
Phylum Tardipolypoda Chen and Zhou, 1997
Class Xenusia Dzik and Krumbiegel, 1989
Order Scleronychophora Hou and Bergström, 1995
Family Eoconchariidae Hao and Shu, 1987
Genus Microdictyon Bengtson, Matthews, and Missarzhevsky, 1986
Type species: Microdictyon effusum Bengtson, Matthews, and Missarzhevsky, 1986; Rhombocorniculum cancellatum Zone, upper part of Cambrian Stage 3; Geres Member of the Shabakty Group, Ushbas River, Lesser Karatau, Kazakhstan.
Microdictyon robisoni Bengtson, Matthews, and Missarzhevsky, 1986
Fig. 61A.
Material.—Single sclerite, SMNH X11294, from sample 22/67, Sekten Formation, Lena River, Siberia, Russia. Kounamkites Zone, Amgan stage (correlated with the Wuliuan Stage).
Description.—Dome-shaped fragmentary sclerite with a maximum dimension of ca. 1 mm in the plan view and ca. 0.3 mm in height. Distinct, almost circular holes ranging from ca. 20 μm in diameter at the margin to 80 μm at the middle of sclerite, surrounded by 5–7 nodes. The nodes are expressed as cones with flattened tops shifted towards the centre of the sclerite. There is a narrow closed channel in each cone open at the base. Openings of the holes through the sclerite are each surrounded by a basal ring.
Stratigraphic and geographic range.—Wuliuan Stage in Siberia (Russia) (Kouchinsky et al. 2011) and Laurentia (Bengtson et al. 1986; Peel et al. 2016).
Phylum Arthropoda von Siebold in von Siebold and Stannius, 1848
Class uncertain
Order Bradoriida Raymond, 1935
Family Cambriidae Lee, 1975
Genus Cambria Neckaja and Ivanova, 1956
Type species: Cambria sibirica Neckaja and Ivanova, 1956; Atdabanian stage; Lena River, Siberia, Russia.
Cambria sibirica Neckaja and Ivanova, 1956
Fig. 61D.
Material.—Two conjoined deformed valves of a phosphatised carapace, SMNH Ar75001, from sample 21/57.75, Tyuser Formation, Ulakhan-Ald’yarkhay, lower reaches of Lena River, Siberia, Russia. Delgadella anabara–Nevadella Zone, Atdabanian Stage (correlated with the Cambrian Stage 3).
Description.—Carapace (dorsal shield) is ca. 3.2 mm long. Prominent anterior lobe situated in the anterior cardinal area of each valve and forms a granulated cone with a blunt tip. Posterior lobe is lower, more extended and gently curved. Connecting lobe indistinct. Continuous admarginal ridge confluent with valve margin. The valve outer surface with granulose ornament.

Fig. 60. Scalidophoran Markuelia secunda Val’kov in Val’kov and Karlova, 1984 from the Cambrian Erkeket Formation, Khorbusuonka River, Siberia, Russia; sample 11/16.05. Phosphatized enrolled embryo in different views (A1–A3), SMNH X11293. Scale bar 250 µm.

Fig. 61. Lobopodian (A) and bradoriid (B–D) calcium phosphatic fragments, from the lower Cambrian Erkeket Formation, Khorbusuonka River (B, C) and Tyuser (D), and Sekten (A) formations, lower reaches of the Lena River, Siberia, Russia; samples 19/3.25 (C), 19/31.75 (B), 21/57.75 (D), and 22/67 (A). A. Microdictyon robisoni Bengtson, Matthews, and Missarzhevsky, 1986, sclerite, SMNH X11294. B. Mongolitubulus spinosus (Hinz, 1987), spine, SMNH Ar75002. C. Mongolitubulus reticulatus Kouchinsky, Bengtson, Clausen, Gubanov, Malinky, and Peel, 2011, spine, SMNH Ar75003. D. Cambria sibirica Neckaja and Ivanova, 1956, carapace, SMNH Ar75001; D1, anterior view; D2, D3, dorsal view. Scale bar: 250 µm (A–C) and 1.25 mm (D).
Remarks.—For description of C. sibirica and comparison with other species see Williams et al. (1994), Vannier et al. (2005), Hinz-Schallreuter et al. (2008). Cambriid bradoriids are the oldest bradoriids known from Cambrian Stages 3 and 4 of Siberia, Gondwana and Laurentia (e.g., Melnikova et al. 1997).
Stratigraphic and geographic range.—Atdabanian stage (Cambrian Stage 3), Siberian Platform, Russia.
Family Mongolitubulidae Topper, Skovsted, Harper, and Ahlberg, 2013
Genus Mongolitubulus Missarzhevsky, 1977
Type species: Mongolitubulus squamifer Missarzhevsky, 1977; Botoman stage; Salany-Gol River, Mongolia.
Mongolitubulus spinosus (Hinz, 1987)
Fig. 61B.
Material.—Single fragment of calcium phosphatic spine with broken off tip, SMNH Ar75002, from sample 19/31.75, Erkeket Formation, Khorbusuonka River, Siberia, Russia. Toyonian stage (correlated with the Cambrian Stage 4).
Remarks.—Hollow spine fragment nearly circular in cross-section and covered with longitudinally extended scaly polygons with raised ends pointed towards the apex of the spine. The specimen is 0.8 mm long and 0.2 mm wide and similar to the spines described from the Botoman Stage, Emyaksin Formation of the Anabar Uplift (Kouchinsky et al. 2015a, see description therein). Currently available material extends the stratigraphic range of Mongolitubulus spinosus into the Toyonian stage (Cambrian Stage 4). .
Stratigraphic and geographic range.—Cambrian Series 2, Siberian Platform (Russia) and Britain.
Mongolitubulus reticulatus Kouchinsky, Bengtson, Clausen, Gubanov, Malinky, and Peel, 2011
Fig. 61C.
Material.—Single calcium phosphate apical fragment of a specimen SMNH Ar75003, from sample 19/3.25, Erkeket Formation, Khorbusuonka River, Siberia, Russia. Lower Botoman stage (correlated with the Cambrian Stage 4).
Remarks.—Short apical fragment of a hollow tubular spine with nearly circular cross-section. Outer surface covered with subdued longitudinally extended polygons. Tip of the spine recurved and pointed. The specimen ca. 0.7 mm long and ca. 0.2 mm in diameter. Mongolitubulus reticulatus is different from Mongolitubulus spinosus (see above) in ornamentation and in having a bent apical part.
Stratigraphic and geographic range.—Lower Botoman stage (correlated with the Cambrian Stage 4)–Wuliuan Stage (type material from the Kuonamka Formation, Anabar Uplift, Kounamkites Zone, Amgan stage, see Kouchinsky et al. 2011), Siberian Platform, Russia.
Phylum Echinodermata Klein, 1734
Subphylum Blastozoa Sprinkle, 1973
Class, order, and family uncertain
Brachiolar plates
Fig. 62A, B.
Material.—Several thousand phosphatised ossicles, including figured SMNH Ec36852 and 36853, from samples 19/5.5, 19/10.25, 19/11.75, 19/12.75, 19/25.5, 19/26.75, 19/29, 19/31.75, 19/33, 19/40, 19/43.25, 19/46.5, 20/1B, 20/2B, 20/3B, 20/4B, 20/5B, 20/6B, Erkeket Formation, Botoman–Toyonian stages (correlated with the Cambrian Stage 4) and sample 19/70.5, Yunkyulyabit-Yuryakh Formation, lower Mayan stage (Drumian Stage), Khorbusuonka River, and samples 22/0, top of the Tyuser Formation, 22/50 and 22/67, Sekten Formation, Botoman–Toyonian stages (Cambrian Stage 4), lower reaches of the Lena River. Siberia, Russia.
Description.—The brachiolar plates are preserved as isolated elements crescent-shaped in cross-section (Fig. 62B) or articulated hoof-shaped pairs (Fig. 62A) delimiting a slightly concave oral and convex aboral surfaces. Disarticulated (isolated) brachiolars are up to 0.5 mm long, with a length-to-width ratio of 0.5–2, and may slightly taper distally. Smaller brachiolars tend to be more elongated than the bigger ones. Articulation in a pair of opposed plates takes place along two flattened articulation surfaces (longitudinal ridges) delimiting a large, oval aboral canal (Fig. 62A1: arrowed). The oral surface is variably concave, and consist of a median furrow flanked by two poorly developed but continuous ridges delimiting two variably developed lateral depressions. The external surface of the plates (latera) are smooth, and straight. Distal and proximal articulation surfaces are parallel and consist of a peripheral flat wall delimiting crescent shaped depressions around the aboral canal. Oral articulation between two opposite plates is straight and submedian (along the median oral furrow), while aboral articulation is either longitudinal or oblique. Articulated elements are aligned and opposite.
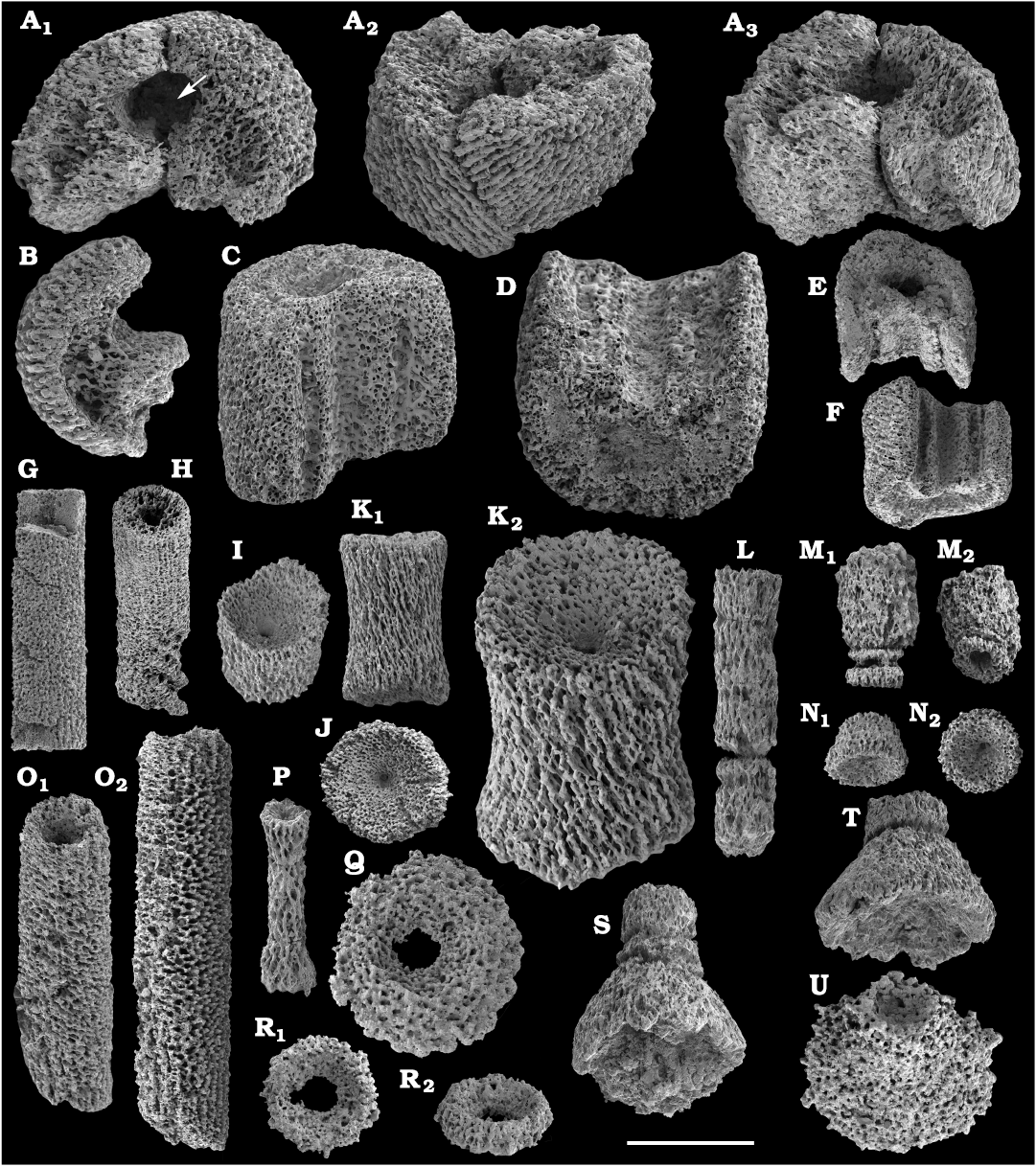
Fig. 62. Blastozoan plates from the lower Cambrian Erkeket Formation, Khorbusuonka River, Siberia, Russia; samples 20/2B (A, B), 20/5B (I, J, K, U), 19/12.75 (C, F), 19/11.75 (D, E), 19/10.25 (G, H), 19/107 (L, S, T), 19/46.5 (M, N, P, Q), 19/65.5 (O), and 19/2B (R). A, B. Brachiolar, biserial plates, SMNH Ec36852 and 36853. A1, articulation surface and aboral canal, arrowed; A2, aboral surface; A3, oral surface. C–F. Brachial, uniserial plates, SMNH Ec36854–36857, respectively. G–R. Stem holomeric columnals, SMNH Ec36858–36869, respectively. K1, lateral view; K2, oblique view showing articulation facet; M1, lateral view of a stem fragment consisting of three articulated ossicles; M2, oblique view showing articulation facet; N1, fragmentary stem ossicle in oblique lateral view; N2, view on the articulation facet; O1, elongated stem ossicle, oblique lateral view showing articulation facet; O2, lateral view; R1, discoidal stem ossicle, view on the articulation facet; R2, oblique lateral view. S–T. Distal fragments of holomeric stem including columnals articulated with a holdfast, SMNH Ec36870–36871, respectively. U. Holdfast?, SMNH Ec36872. Scale bar: 500 µm (E–J, K1, M–O) and 250 µm (A–D, K2, L, P–U).
Remarks.—Brachiolar plates represent biserial feeding appendages of blastozoans. They are here considered as eocrinoids sensu lato, although some may represent the distal arm of Rhombiferans (see below). The plates are grossly similar to Brachiolar type B described by Kouchinsky et al. (2015a: figs. 62A–H, 63A–D) from the Judomia Zone, Atdabanian stage (correlated with the Cambrian Stage 3) and Calodiscus–Erbiella Zone, lower Botoman stage (correlated with the lower part of Cambrian Stage 4).
Class, order, and family uncertain
Brachial plates
Fig. 62C–F.
Material.—Four phosphatised ossicles, SMNH Ec36854–36857, from samples 19/11.75 and 19/12.75, Erkeket Formation, Khorbusuonka River, Siberia, Russia. Lower Botoman stage (correlated with the lower part of Cambrian Stage 4).
Description.—The semi-cylindrical, uniserial unbranched ossicles, ca. 0.6 mm in length and width, crescent- or hoof-shaped in cross-section (Fig. 62C–F) have parallel, straight to slightly concave and smooth latus. The longitudinal food groove runs along the entire oral surface, flanked by elevated lateral walls. It occupies almost 3/4 of the oral surface in width. It consists of a central furrow (1/3 of the groove in width) separated from two narrower lateral furrows (1/2 the central furrow in width) by two small, longitudinal ridges. The proximal and distal hoof-shaped articulation facets are delimited by a raised peripheral rim and have a central oval-shaped concavity which leads to a more or less distinct aboral canal. The canal, oval in cross-section when visible, is more or less filled by claustrum-like stereom and, or exogenous material. The distal and proximal articulations are indistinct, flat to irregularly undulose.
Remarks.—The plates are very similar to uniserial brachitaxial plates described by Kouchinsky et al. (2015a: fig. 63E–H) from the Calodiscus–Erbiella Zone (lower Botoman stage) of the Anabar Uplift, with which they represent the earliest uniserial elements of blastozoan feeding appendages known so far. These uniserial plates have an overall similarity to fused biserial brachiolar plates, but the latter always show an open aboral canal joining the proximal to the distal facet. Although the nomenclature used herein is based on the terminology dedicated to the description of the crinoid arm-ossicles, their affinity is still debated. Considering the presence of uniserial elements in some early rhombiferan arms (Zamora and Smith 2012), and the age of described plates, these uniserial elements are here assigned to the subphylum Blastozoa. These elements might represent the distal portion of a biserial appendage (as observed in the Ordovician blastozoan Haimacystis; Sumrall et al. 2001), and therefore may have been associated with some of the previously described brachiolars. Alternatively, they may represent different taxa.
Stem holomeric columnals and holdfasts
Fig. 62G–U.
Material.—Tens of phosphatised ossicles, including figured SMNH Ec36858–36872, from samples 19/10.25, 19/46.5, 19/65.5, 19/70.5, 19/107, 20/4B, 20/5B, 20/6B, Erkeket, Kuonamka, and Yunkyulyabit-Yuryakh formations, Khorbusuonka River, Siberia, Russia. Botoman–Mayan stages (correlated with the Cambrian Stage 4–Drumian Stage).
Description.—The flat discoidal to tall holomeric columnals are circular in transverse section 70–400 µm in diameter, up to 1.7 mm long, with length/width ratio 1.7–5.0. Laterali are flat in discoidal elements, flat to concave in taller, cylindrical elements. Distal and proximal facets distinctly concave with a narrow, smooth, planar to slightly convex, peripheral articulum. Narrow convex perifacet is present in the discoidal elements, but absent in the taller ossicles. A concave, deep areola surrounds the central, circular lumen. Lumen is narrow in tall elements and larger in flat, discoidal ossicles, 50–100 µm in diameter. Some stem fragments consisting of articulated ossicles (Fig. 62L, M, S, T) are described, including two distal stem fragments preserving their conical holdfast (Fig. 62S, T). Holdfasts are conical, with straight to faintly convex latus. Their lower, distal surface is slightly to strongly concave with an irregular margin. Their proximal facet (visible on isolated elements) is concave and bordered by a flat, narrow epifacet.
Remarks.—Recrystallization in the recovered columnals precludes any analysis of their stereom microstructure. Two specimens preserve stem portions comprising several articulated ossicles. The first one (Toyonian, correlated with the Cambrian Stage 4; Fig. 62M), is composed of 3 distinct ossicles. The proximal columnal is slightly wider and clearly taller. It is followed distally by two discoidal elements, the intermediate being the flattest. The lumen is large and luminal index high. This fragment, based on the luminal index, may represent the proximal portion of a heteromorphic or xenomorpic stem (definition hampered by incomplete preservation). The second stem-ossicle (Mayan, correlated with the Drumian Stage; Fig. 62L) most probably represents a homeomorphic portion of a stem consiting of 4–5 small, cylindrical ossicles (height index about 1). The articulation between the ossicles is faint, most probably due to diagenetic overgrowth of the ossicles, but those were undoubtedly originally articulated. Cylindrical elements from sample 19/10.25 (Fig. 62G, H) may represent the oldest holomeric columnals recovered so far, from the lower Botoman stage (correlated with the lower part of Cambrian Stage 4). Although relatively rare, holomeric columnals are now continuously recovered from lowermost Cambrian Stage 4 through the Drumian Stage, along the Khorbusuonka River. Those include the earliest known holomeric columnals, older than those reported from Morocco (Clausen and Smith 2008; Clausen et al. 2014) and Laurentia (Clausen and Peel 2012). These reports show the pelmatozoan echinoderms were adapted to hard substrate attachment at the earliest stage of their evolution (Clausen et al. 2014; Zamora et al. 2017), and that their stem quickly diversified from polyplated holdfast to homeomorphic and xenomorphic or heteromorphic holomeric stems. However, none were reported from the Kuonamka River (Kouchinsky et al. 2011), wherefrom disarticulated echinoderm elements were described. Some ossicles may represent stem parts of the earliest rhombiferans (see below).
Class Rhombifera (Zittel, 1879)
Order uncertain
Family Dibrachicystidae? Zamora and Smith, 2012
Proximal stem plates
Fig. 63A, B.
Material.—Two phosphatised plates, SMNH Ec36873 and 36874, from sample 19/10.25, Erkeket Formation, Khorbusuonka River, Siberia, Russia. Lower Botoman stage (correlated with the lower Cambrian Stage 4).
Description.—Plates with triangular chevron-like outline, ca. 0.3 and 0.6 mm wide, with arcuate adaxial surface (bordering the stem lumen) bordered by a rim. The abaxial, triangular part of the plate is relatively flat, slightly curved, and made of coarse galleried to fasciculated stereom.
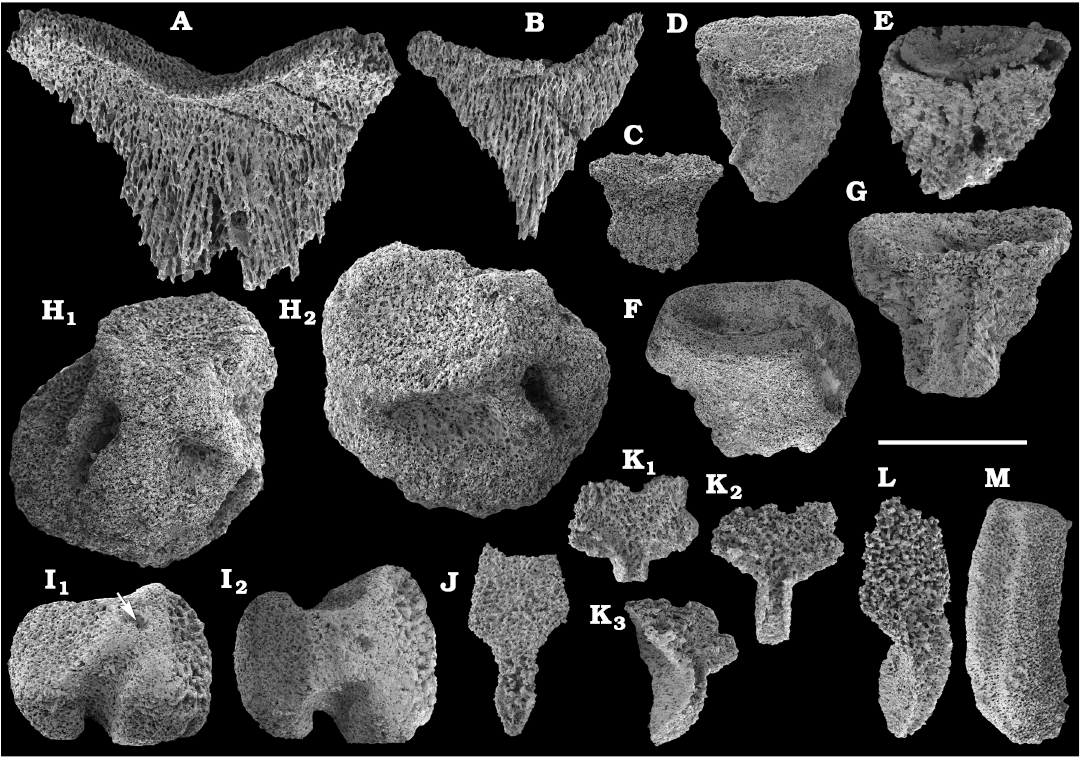
Fig. 63. Echinoderm plates from the lower Cambrian, Erkeket Formation, Khorbusuonka River, Siberia, Russia; samples 19/10.25 (A, B), 19/11.75 (E, G–I), 19/12.75 (C, F), 19/25.5 (J, L), 20/2B (K), and 20/6B (D, M). A, B. Proximal stem plates of Rhombifera, SMNH Ec36873 and 36874, respectively. C–G. Edrioasteroid thecal plates (from basal attachment disc?), SMNH Ec36876–36880, respectively. H. Plate from edrioasteroid mouth-frame, SMNH Ec36881. I. Ambulacral flooring plate of Edrioasteroidea, SMNH Ec36875. I1, oblique view, a shallow pit for presumably insertion of a ligament for a cover-plate is arrowed; I2, upper view. J–L. Ctenidial plates (Echinodermata, Ctenocystoidea), SMNH Ec36882–36884. K1, plain view on a trapezoidal dorsal blade; K2, plain view of the plate showing dorsal blade and hook-shaped process; K3, plate in lateral view. M. Echinodermata, thecal frame-plate of unknown affinity, SMNH Ec36885. Scale bar: 1 mm (D, F, I, M) and 500 µm (A–C, E, G, H, J–L ).
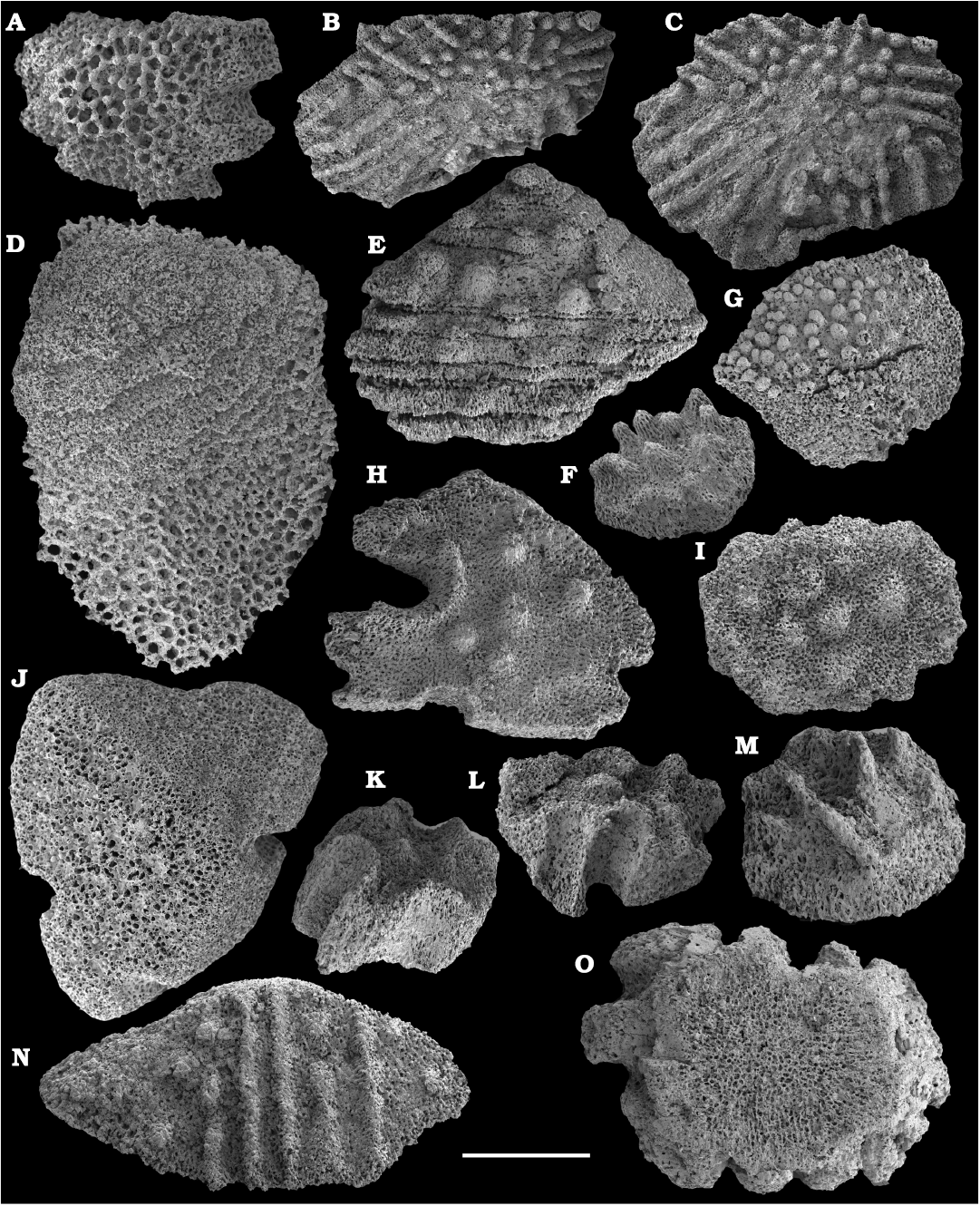
Fig. 64. Thecal plates of Echinodermata from the lower Cambrian Erkeket Formation of Khorbusuonka River (B, C, E–O) and the middle Cambrian Sekten Formation, lower reaches of the Lena River (A, D), Siberia, Russia; samples 19/11.75 (O), 19/12.75 (J), 20/2B (I, K–M), 20/5B (B, C, F, G, N), 20/6B (E, H), and 22/67 (A, D). A–O. Thecal plates, SMNH Ec36886–36900, respectively. Scale bar 500 µm.
Remarks.—The proximal stem plates described herein are similar to those described from the Drumian Stage of Siberia (Kouchinsky et al. 2011: fig. 38A–F) and identified by Zamora et al. (2013: fig.13.5 f, g) as proximal stem plates of dibrachicystids (Zamora and Smith 2012). Dibrachicistidae is a basal group of the Rhombifera, the earliest undoubted representatives of which are reported from Wuliuan Stage of Gondwana (Zamora 2010; Zamora and Smith 2012; Zamora et al. 2013). Articulated specimens of dibrachicystids show a tripartite stem with multiple holomeric columnals (Zamora and Smith 2012), similar to those (Fig. 62G, H) found together with the plates described herein.
Class Edrioasteroidea Billings, 1858
Order and family uncertain
Ambulacral flooring plate
Fig. 63I.
Material.—Single phosphatised ossicle, SMNH Ec36875, from sample 19/11.75, Erkeket Formation, Khorbusuonka River, Siberia, Russia. Lower Botoman stage (correlated with the lower Cambrian Stage 4).
Description.—Plate is hourglass-shaped in plane view, ca. 1.4 mm long, ca. 1.0 mm wide, and ca. 0.7 mm thick. Its external (upper) side is subdivided into wider abradial facet (right parts of Fig. 63I) and more flattened adradial surface (see Kouchinsky et al. 2011: fig. 36) connected by a short median area. The latter carries a circular shallow pit (Fig. 63I1: arrow), ca. 120 µm in diameter, a presumable insertion of a ligament for a cover-plate. Two semi-elliptical lateral gaps in median area represent sutural pores between adjacent ambulacral plates.
Remarks.—The plate is of the same age as the one described by Kouchinsky et al. (2015a) from the Anabar Uplift in Siberia (see also Kouchinsky et al. 2011; Clausen and Peel 2012; Zamora et al. 2013; Peel et al. 2016).
Edrioasteroid thecal plates
Fig. 63C–H.
Material.—Seven disarticulated plates, SMNH Ec36876–36881, from samples 19/11.75, 19/12.75, 20/6B. Erkeket Formation, Khorbusuonka River, Siberia, Russia. Lower Botoman and Toyonian stages (correlated with the Cambrian Stage 4).
Description.—Nearly bilaterally symmetrical ossicles, up to 0.8 mm across, curved in the lateral view. The plate has a smooth convex and ridged concave triangular or trapezoidal surfaces perpendicular to a slightly concave and smooth ovaliform basal face surrounded by a rounded rim. The two sides converge towards an embayed edge of the plate. The ridges radiate from a terminal face of the plate. A different grossly bilaterally symmetrical plate (Fig. 63H; interradial oral frame plate) is polygonal to oval in upper view, about 800 µm in proximal–distal length and about 600 µm in perpendicular extension. Three distinct clefts mark the abradial surface of the plate. The distal portion of the plate slopes down distally in upper view, whereas its inner face is flat and almost perpendicular to the aradial surface.
Remarks.—Plates similar in morphology and age were illustrated by Landing and Bartowski (1996: fig. 8 [13–19]), Kouchinsky et al. (2015a: fig. 64; see also references and discussion therein) and Peel et al. (2016: fig. 17) and were tentatively interpreted as possible plates from basal attachment disc of edrioasteroids.
Class Ctenocystoidea Robison and Sprinkle, 1969
Order Ctenocystoida Ubaghs and Robison, 1988
Family Ctenocystidae Sprinkle and Robison, 1978
Ctenidial plates
Fig. 63J–L.
Material.—Three secondarily phosphatised plates, all figured, SMNH Ec36882–36884, from samples 20/2B and 19/25.5 Erkeket Formation, Khorbusuonka River, Siberia, Russia. Uppermost Botoman–basal Toyonian stages (correlated with the Cambrian Stage 4).
Description.—Anterolateral plates of the lower-surface (ventral) marginal-frame (ctenidial plate, from about 500 to 750 µm in length) composed of an anterior hook-shaped process and a trapezoidal to squared dorsal blade (about 450 µm in maximum width). The gutter-like anterior process is lanceolate and arcuate in lateral view with a concavo-convex, U-shaped transverse section. The lateral walls of the posterior process decrease laterally and posteriorly at the junction with flat to slightly concave posterior blade, the latter being bordered with posteriorly decreasing walls.
Remarks.—These ctenidium plates are the oldest ctenoid evidences reported so far. Similar hook-shaped ctenidial plates are reported in most ctenocystoid species (Zamora et al. 2012). Ctenidium plates of Ctenocystis (Robison and Sprinkle 1969) have similar gutter-like, hook-shape anterior blade with the gutter oriented outwards (Rahman and Clausen 2009). The ctenocystoids are interpreted as basalmost, bilateral to slightly asymmetrical echinoderms (Rahman and Clausen 2009; Zamora et al. 2012), although they occur relatively late in the early echinoderm record (Wuliuan Stage). This discovery from the Cambrian Stage 4 of the Khorbusuonka River thus significantly reduces the incongruence between the phylogenetic position of ctenocystoids and their first appearance data. Along with other remains reported herein, they place the Siberian Platform as one of the diversification centres of basal echinoderms (SOM 5).
Class, order, and family uncertain
Thecal plates
Fig. 64.
Material.—Several thousand secondarily phosphatised plates and ossicles, including figured SMNH Ec36886–36900, from samples 19/11.75, 19/12.75, 19/25.5, 19/26.75, 19/29, 19/31.75, 19/33, 19/40, 19/43.25, 19/46.5, 19/55, 20/1B, 20/2B, 20/3B, 20/4B, 20/5B, 20/6B, Erkeket Formation, Khorbusuonka River, samples 22/0, top of the Tyuser Formation, 22/50, 22/55.5, 22/67, Sekten Formation, Lena River, Siberia, Russia; Botoman–Amgan stages (correlated with the Cambrian Stage 4–Wuliuan Stage).
Remarks.—The systematic affinity of disarticulated thecal plates of echinoderms are generally difficult to determine. Assemblages of plates herein contain those with irregular shape and well-defined epispires. External ornamentation of the plates is represented by rather prominent bumps sometimes in combination with groups of parallel ridges or alternatively a smooth surface with a polygonal network. The plates either thin out at the margin suggesting an imbricated arrangement or have flattened abutment faces.
During fossilization, the stereom becomes phosphatised via encrustation or/and infill of canals. Alternatively, cavities in the stereom can be filled in with calcite or dolomite in crystalline continuity with the surrounding material. In some cases, such diagenetic alteration of stereom co-occurs with glauconitisation (Kouchinsky et al. 2011). Silicification of echinoderm ossicles also takes place (Kouchinsky et al. 2011). Replacement of the original high-magnesial calcite ossicles by low-magnesium calcite or dolomite results in formation of monocrystals. The latter, either of calcite or dolomite, have a perfect rhombohedral cleavage in three directions. Distinctive cleavage patterns can be seen but rarely visible in all of the directions in the whole ossicle. More often, cleavage results in splitting of recrystallised ossicles into fragments bounded by plane surfaces (Fig. 64B, D, E). Isolated flat fragments can occur and mimic those of archaeocyaths or plates of Microdictyon that may lead to confusion in identification and biostratigraphy.
Phylum and class uncertain
Order Hyolithelminthida Fisher, 1962
Family Hyolithellidae Walcott, 1886
Genus Hyolithellus Billings, 1871
Type species: Hyolithes micans Billings, 1871; lower Cambrian; Taconic allochthon, Quebec, Canada.
Hyolithellus isiticus Missarzhevsky in Rozanov et al., 1969
Fig. 65B–E.
Material.—Several fragmentary tubes in calcium phosphate, including figured SMNH X11295–11298, from samples 11/19.5, lower Erkeket Formation, Khorbusuonka River and 21/21, Tyuser Formation, Lena River. Siberia, Russia; Dokidocyathus regularis Zone, Tommotian stage (correlated with the upper part of Cambrian Stage 2).
Remarks.—The tubes described by Rozanov et al. (1969: 150) have a smooth initial part similar to Hyolithellus tenuis Missarzhevsky in Rozanov and Missarzhevsky, 1966, intermediate part with faint transversal ribs as in Hyolithellus vladimirivae Missarzhevsky in Rozanov and Missarzhevsky, 1966, and well-developed flanges inclined towards the aperture at later growth stages. The available fragments suggest a diameter ca. 1 mm. External surface with faint transverse folds perpendicular to the longitudinal axis of the tube. The holotype derives from section Isit’ in the middle reaches of the Lena River, from the Dokidocyathus regularis Zone. Hyolithellus insolitus Grigor’eva in Voronin et al., 1982 (see Kouchinsky et al. 2015a) differs in having weaker concentric ribs without flanges and faint undulating longitudinal ridges and furrows between the ribs.
Stratigraphic and geographic range.—Dokidocyathus regularis Zone, Tommotian stage (correlated with the upper part of Cambrian Stage 2) through the Botoman stage (correlated with the Cambrian Stage 4), Siberia, Russia and equivalent beds in Mongolia, South China, India, Antarctica and Britain (Rozanov et al. 2010: 59).
Hyolithellus sp.
Fig. 65A, F, H.
Material.—Several hundred fragmentary calcium phosphatic tubes, including figured SMNH X11299–11301, from samples 15/23, Chuskuna Formation (Kessyusa Group), Watsonella Zone, Cambrian Stage 2, 11/16.1, 11/16.5, 11/18.7, 11/21.6, Erkeket Formation, Khorbusuonka River and 21/22, Tyuser Formation, Lena River, Dokidocyathus regularis Zone, Tommotian stage (correlated with the upper part of Cambrian Stage 2), 19/3.25, 19/4.25, 19/14.25, 19/16, Botoman stage (Cambrian Stage 4), 19/25.5, 19/26.75, 19/29, 19/46.5, 20/4B, 20/5B, Toyonian stage (Cambrian Stage 4), Khorbusuonka River, 21/51, Delgadella anabara–Nevadella Zone, Atdabanian stage (Stage 3), and 22/0, Tyuser Formation, Bergeroniellus micmacciformis–Erbiella Zone, Botoman stage (Stage 4), lower reaches of the Lena River. Siberia, Russia.
Remarks.—Slightly irregularly curved very slowly expanding tubes with circular cross-section, up to 300 µm in diameter. External surface smooth or with faint transverse folds perpendicular to the longitudinal axis of the tube. Hyolithellus sp. lack longitudinal striation and regularly spaced ribs.
Family Torellellidae Holm, 1893
Genus Torellella Holm, 1893
Type species: Hyolithes laevigatus Linnarsson, 1871; lower Cambrian; Sweden.
Torellella spp.
Figs. 65G, I–L, 66.
Material.—Several hundred calcium phosphatic tubes and fragments, including figured SMNH X11302–11314, from the Erkeket and Tyuser (sample 22/0) formations. Samples 11/18.7, 11/21.6, 11/28.8, Khorbusuonka River, 21/21, 21/22, 21/22.7, 21/33, Lena River, Tommotian stage (correlated with the upper part of Cambrian Stage 2), 19/1.5, 19/3.25, 19/4.25, 19/5.5, 19/14.25, 20/1B, 20/2B, Khorbusuonka River, and 22/0, Lena River, Botoman stage (Cambrian Stage 4). Siberia, Russia.
Remarks.—Specimens in Fig. 65K, L are similar to Torellella biconvexa Missarzhevsky in Rozanov et al., 1969. They have lenticular cross-section and frequent weak transversal growth lines, diverging at ca. 10°, slightly undulating and curved predominantly in a single plane of their narrow sides. Specimens in Fig. 65I, J are similar to Torellella mutila Missarzhevsky, 1989 (Missarzhevsky 1989: 195, pl. 24: 8) from the Bergeroniellus micmacciformis–Erbiella Zone, Botoman stage of the middle Lena River. The form is also similar to T. biconvexa, but is predominantly irregularly curved in a single plane. A longitudinal furrow can be observed on narrow sides of the tube (Fig. 65I) and a fragment (Fig. 65G). Similar furrows are described from Torellella cf. laevigata by Kouchinsky et al. (2015a).
Tubes of Hyolithellus sp. and Torellella sp. are perforated by circular, oval and rounded rectangular holes, 20–60 µm across (Fig. 66, see also Kouchinsky et al. 2015a). Rare perforated tubes of Hyolithellus sp. and Torellella sp., otherwise identical to non-perforated ones, were first mentioned and illustrated by Repina et al. (1974: 72, 73, pl. 19: 3, 4; pl. 15: 1, 3) from the upper Dokidocyathus regularis Zone, Tommotian stage of the Chekurovka section in lower reaches of the Lena River. Other co-occurring fossils do not show such perforations, implying that Hyolithellus and Torellella had similar properties of their organophosphatic tubes corroborating their affinity to the same animal clade.
Bifurcating and irregularly curved, ca. 10 µm thick galleries are found on a single fragment of Torellella sp. from lower Botoman stage (Fig. 66H). The galleries are generally subparallel to the tube surface, with only a few branches directed perpendicularly to the surface. The latter type of bioerosion has not yet been described from hyolithelminths and was likely bored by an organism different from a producer of thicker perpendicular canals (holes) available in other fragments. The shape of the holes in Hyolithellus and Torellella tubes depend on plywood arrangement of multiple layers with orthogonal orientation of the constituent fibres. In thinner walls in Hyolithellus, rounded and circular holes predominate. Rectangular pores occur in thicker tubes of Torellella, where they penetrate through multiple alternating laminae having mutually orthogonal orientation of fibers.
Phylum and class uncertain
Order Angustiochreida Val’kov and Sysoev, 1970
Family Anabaritidae Missarzhevsky, 1974
Genus Anabarites Missarzhevsky in Voronova and Missarzhevsky, 1969
Type species: Anabarites trisulcatus Missarzhevsky in Voronova and Missarzhevsky, 1969; Anabarites trisulcatus Zone, Nemakit-Daldynian stage (Fortunian Stage); mouth of the Kotujkan River, Anabar Uplift, Siberian Platform, Russia.
Anabarites tristichus Missarzhevsky in Rozanov et al., 1969
Fig. 67A.
Material.—Single calcium phosphatic internal mould, SMNH X11315, from sample 11/16.05, basal Erkeket Formation, Khorbusuonka River, Siberia, Russia. Dokidocyathus regularis Zone, Tommotian stage (correlated with the upper part of Cambrian Stage 2).
Remarks.—Represented by a gently curved internal mould replicating lumen of a tube. Three radially symmetrical longitudinal rows of elongated notches are situated in shallow depressions between flattened convex lobes. Faint replicas of parallel growth lines run transversally. See Kouchinsky et al. (2009) for additional description and discussion.
Stratigraphic and geographic range.—Terreneuvian Series and lower Stage 3; Siberia (Russia) and Gondwana (South China).
Anabarites hexasulcatus (Missarzhevsky, 1974)
Fig. 67B.
Material.—Single calcium phosphatic internal mould, SMNH X11316, from sample 15/23, Chuskuna Formation (upper part of the Kessyusa Group), Khorbusuonka River, Siberia, Russia. Watsonella Zone, Cambrian Stage 2.
Remarks.—Fragment of internal mould with six longitudinal lobes and radially symmetrical cross-section, ca. 0.5 mm in diameter. Rounded lobes delimited by alternating deeper first- and shallower second-order longitudinal furrows.
Stratigraphic and geographic range.—Uppermost Fortunian and lower part of Cambrian Stage 2, Siberian Platform (Russia), western Mongolia (Esakova and Zhegallo 1996: 96, pl. 3.17), and probably South China (Kouchinsky et al. 2009).
Anabarites korobovi (Missarzhevsky in Rozanov and Missarzhevsky, 1966)
Fig. 67E, F.
Material.—Several hundred calcium phosphatic internal moulds, including SMNH X11319 and 11320 figured herein and those illustrated by Kouchinsky et al. 2009: fig. 37, from samples 11/2B, 11/16.05, and 11/16.1, basal Erkeket Formation, Khorbusuonka River, Siberia, Russia. Dokidocyathus regularis Zone, Tommotian stage (correlated with the upper part of Cambrian Stage 2).
Remarks.—Represented by internal moulds gently curved in one plane, slowly expanding with rounded triangular cross-section, covered with straight transverse growth lines. The type material in Rozanov et al. (1966) is described from the Chekurovka locality in lower reaches of the Lena River, ca. 60 km upstream from the Ulakhan-Ald’yarkhay sections. Beyond Siberia, such forms are reported from the Cambrian Stage 2 of South China and Avalonia (see Kouchinsky et al. 2009).
Stratigraphic and geographic range.—Nochorojcyathus sunnaginicus and Dokidocyathus regularis zones of the Tommotian stage (correlated with the upper part of Cambrian Stage 2), Siberian Platform, Russia; Cambrian Stage 2, Avalonia (Landing 1988).
Genus Selindeochrea Val’kov, 1982
Type species: Selindeochrea tecta Val’kov, 1982; lower part of Cambrian Stage 2; upper reaches of the Selinde River, Siberia, Russia.
Selindeochrea ternaria (Missarzhevsky in Rozanov et al. 1969)
Fig. 67C.
Material.—Single calcium phosphatic internal mould, SMNH X11317, from sample 15/23, Chuskuna Formation (upper part of the Kessyusa Group), Khorbusuonka River, Siberia, Russia. Watsonella Zone, Cambrian Stage 2.
Remarks.—Fragment of internal mould radially symmetrical in cross-section, with three prominent narrow lobes separated by V-shaped grooves.
Stratigraphic and geographic range.—Upper Fortunian Stage and lower part of Cambrian Stage 2 of the Siberian Platform (Russia) and South China.
Phylum, class, order, and family uncertain
Genus Coleoloides Walcott, 1889
Type species: Coleoloides typicalis Walcott, 1889; lower Cambrian, Brigus Formation; Manuel’s Brook, Conception Bay, Newfoundland, Canada.
Coleoloides trigeminatus Missarzhevsky in Rozanov et al., 1969
Fig. 68A–C, K–M.
Material.—Over ten calcium phosphatic and glauconised internal moulds, including figured SMNH X11326 (C), sample 11/2B, lower Erkeket Formation, Khorbusuonka River and SMNH X11329 (M), sample 21/22, Tyuser Formation, lower reaches of the Lena River, Dokidocyathus regularis Zone, Tommotian stage (upper part of Cambrian Stage 2), Siberia, Russia. Additional material from the Tommotian stage, lower Pestrotsvet Formation of southeastern Siberian Platform includes: SMNH X11325 (B) and SMNH X11328 (L), from sample M303/2, Tiktirikteekh Brook, middle Lena River, D. regularis Zone (see Rozanov et al. 1969: 31, text-fig. 6), SMNH X11327 (K), sample 479a, section “Dvortsy”, Aldan River, Nochoroicyathus sunnaginicus Zone (collected by Anatolij K. Val’kov), and SMNH X11324 (A), sample 510, section Ulakhan-Sulugur, Aldan River, N. sunnaginicus Zone (collected by Anatolij K. Val’kov). Siberia, Russia.
Description.—Somewhat irregularly curved and helically twisted or straight internal moulds representing fragments of very slowly expanding thin-walled tubes, 0.1–0.2 mm in diameter, with 6–12 longitudinal lobes. The lobes rounded in cross-section and separated by narrower intervening grooves. Initial part circular in cross-section, ca. 0.1 mm in diameter (Fig. 68K). At ca. 0.5 mm from probably broken off and incompletely preserved apical end, the cylindrical initial part transforms abruptly into a tube with six lobes, helically twisted anticlockwise. The lobes bifurcate at a later growth stage. Tube fragments greater than 0.2 mm in diameter have up to 12 lobes (Fig. 68B, L).
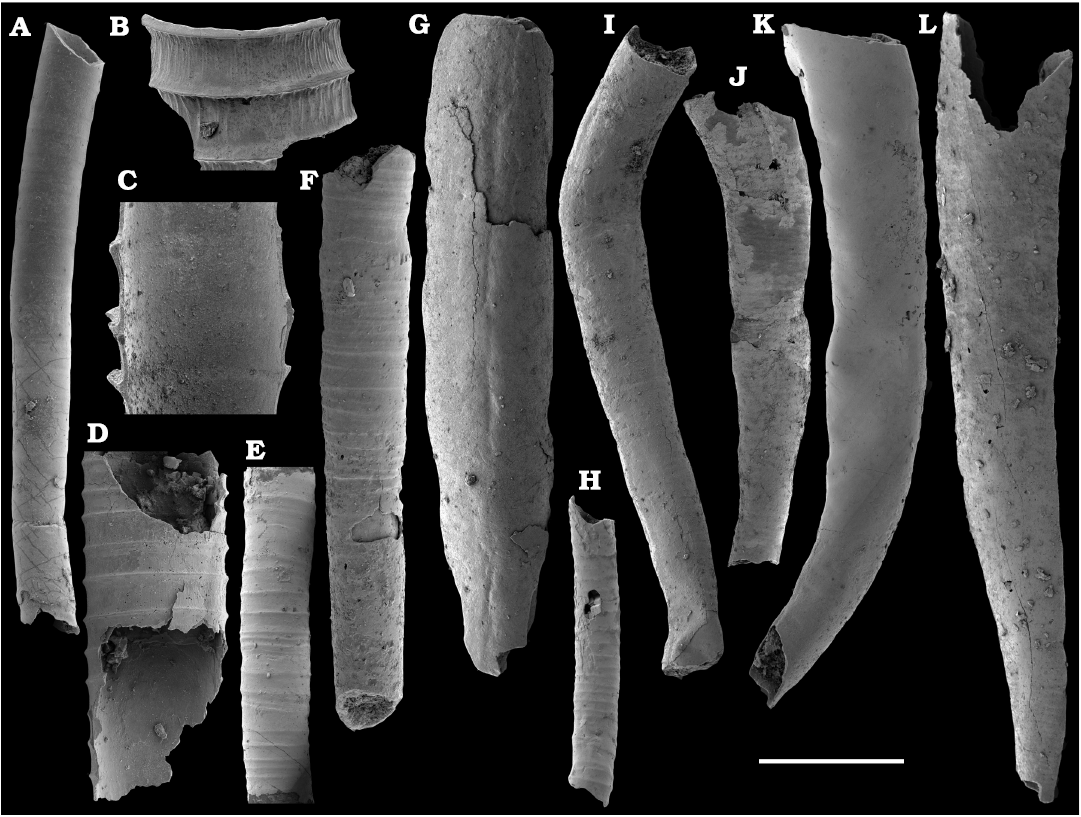
Fig. 65. Calcium phosphatic tubes of hyolithelminths from the lower Cambrian Erkeket Formation, Khorbusuonka River (A–C, G–J) and Tyuser Formation, lower reaches of the Lena River (D–F, K, L), Siberia, Russia; samples 11/19.5 (B, C), 19/3.25 (H, J), 19/4.25 (I), 19/5.5 (G), 19/16 (A), 21/21 (D, E, K), and 21/22 (F, L). A, F, H. Hyolithellus sp., SMNH X11299–11301, respectively. B–E. Hyolithellus isiticus Missarzhevsky in Rozanov et al., 1969, SMNH X11295–11298, respectively. G, I–L. Torellella spp., SMNH X11302–11306, respectively. Scale bar: 1 mm (D, L), 500 µm (A–C, E, F, H, I–K), and 250 µm (G).
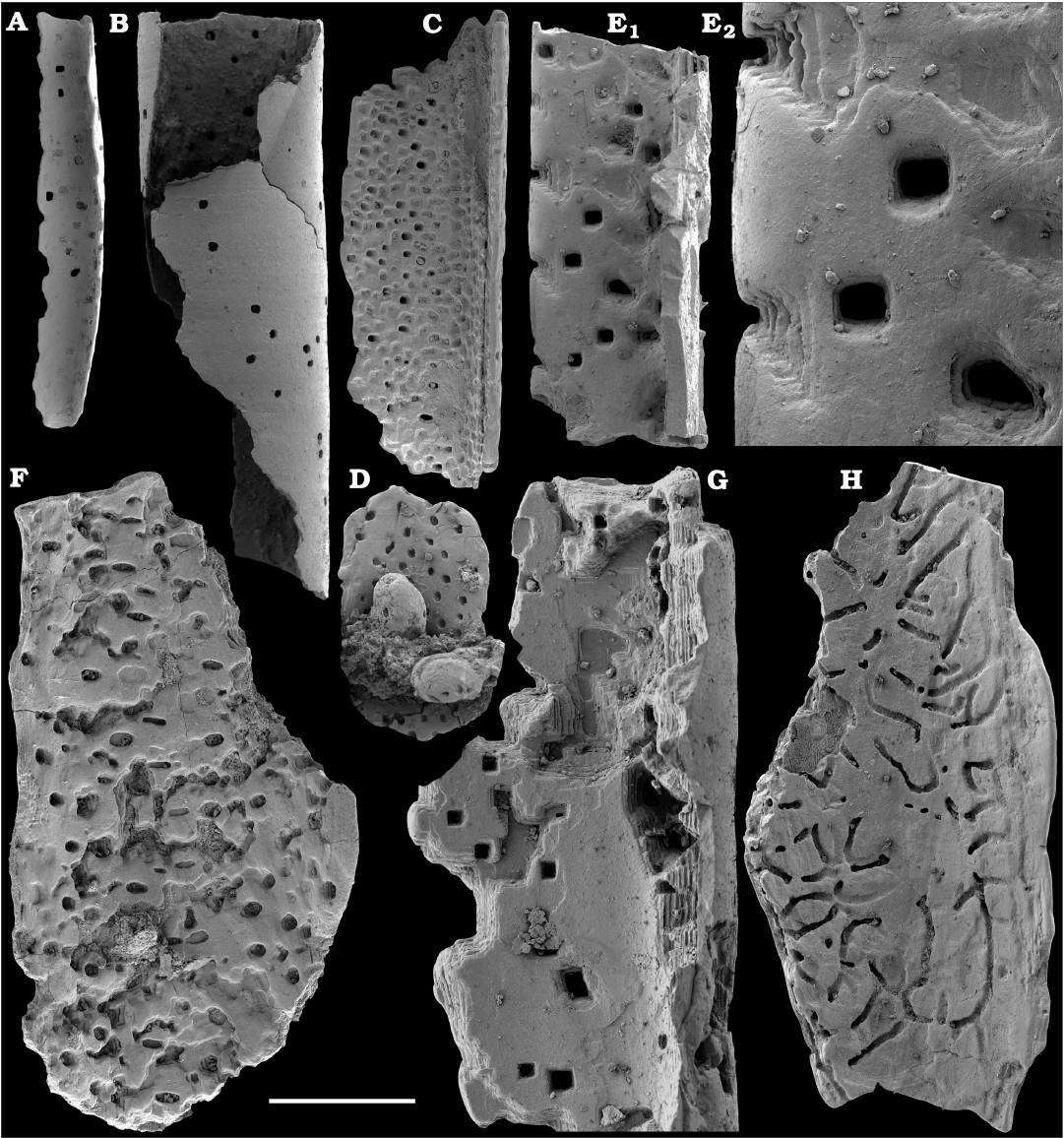
Fig. 66. Torellella spp. with evidences of bioerosion, from the lower Cambrian Erkeket Formation, Khorbusuonka River (A–C, E, G) and Tyuser (F, H) and Sekten (D) formations, lower reaches of the Lena River, Siberia, Russia; samples 19/4.25 (A), 19/5.5 (E), 19/14.25 (B), 20/1B (C, G), 21/129 (D), and 22/0 (F, H). A–G. Fragments of calcium phosphatic tubes penetrated by sub-vertical canals, SMNH X11307–11313, respectively. E2, close-up of E1 showing perforations. H. Fragment of tube with canals sub-parallel to the surface, SMNH X11314. Scale bar: 500 µm (A–D), 250 µm (E1, F–H), and 80 µm (E2).
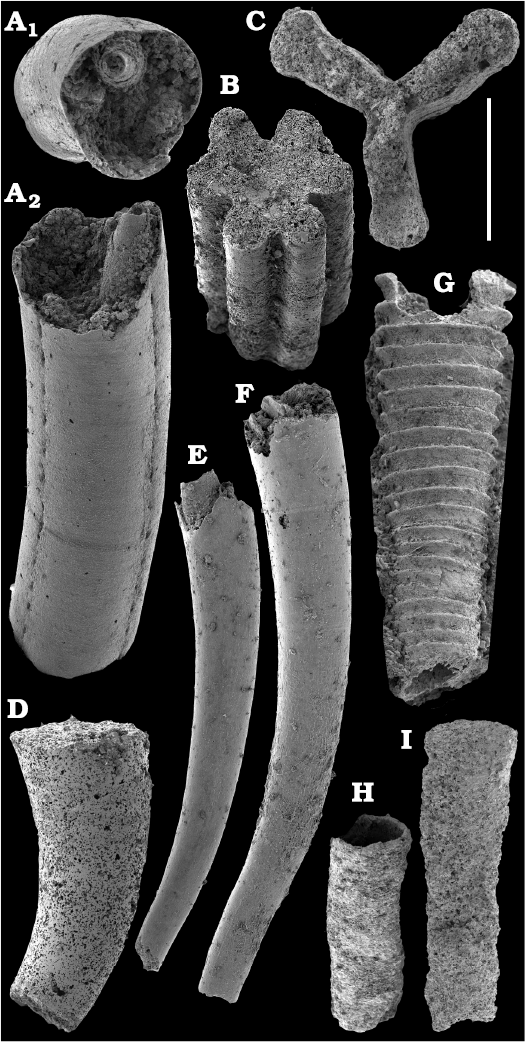
Fig. 67. Anabaritids, internal moulds (A–F) and other problematic tubes (G–I) from the lower Cambrian Kessyusa Group (B–D, H, I) and Erkeket Formation (A, E–G), Khorbusuonka River, Siberia, Russia; samples 11/16.05 (A), 11/2B (E, F), 15/23 (B–D, H, I), and 20/1B (G). A. Anabarites tristichus Missarzhevsky in Rozanov et al., 1969, SMNH X11315. A1, transversal view of the tube; A2, lateral view. B. Anabarites hexasulcatus (Missarzhevsky, 1974), SMNH X11316. C. Selindeochrea ternaria (Missarzhevsky in Rozanov et al., 1969), SMNH X11317. D. Cambrotubulus sp., SMNH X11318. E, F. Anabarites korobovi (Missarzhevsky in Rozanov and Missarzhevsky, 1966), SMNH X11319 and 11320, respectively. G. Coleolella billinsi (Sysoev, 1962), SMNH X11321. H, I. Platysolenites antiquissimus Eichwald, 1860, SMNH X11322 and 11323, respectively. Scale bar: 250 µm (A) and 500 µm (B–I).

Fig. 68. Problematicum Coleoloides Walcott, 1889, internal moulds of tubes, from the lower Cambrian Erkeket (C, D), Tyuser (M), and Pestrotsvet (A, B, E–L) formations, Siberia, Russia; samples 11/2B (C) and 11/16.1 (D) from Khorbusuonka River; sample 21/22 (M) from lower reaches of Lena River; sample M303/2 (B, H, J, L) from Tiktirikteekh Brook (Rozanov et al. 1969: 31, text-fig. 6), sample 171–174 (E) from Bydyangaya Brook (Val’kov 1982: 27, text-fig. 8) middle reaches of the Lena River, sample 479a (K) from section “Dvortsy”, Aldan River, and sample 510 (A, F, G, I) from section Ulakhan-Sulugur, Aldan River (Fig. 1). A–C, K–M. Coleoloides trigeminatus Missarzhevsky in Rozanov et al., 1969, SMNH X11324–11329, respectively. D. Coleoloides sp., SMNH X11330. E–J. Coleoloides cf. paucistriatus Poulsen, 1967, SMNH X11331–11336, respectively. A1, B2, C2, E1, F2, G1, H2, I1, L2, lateral view; A2, B1, C1, F1, H1, L1, transversal view; E2, G2, I2, transversal view in the apical part. Scale bar: 500 µm (M) and 250 µm (A–L).
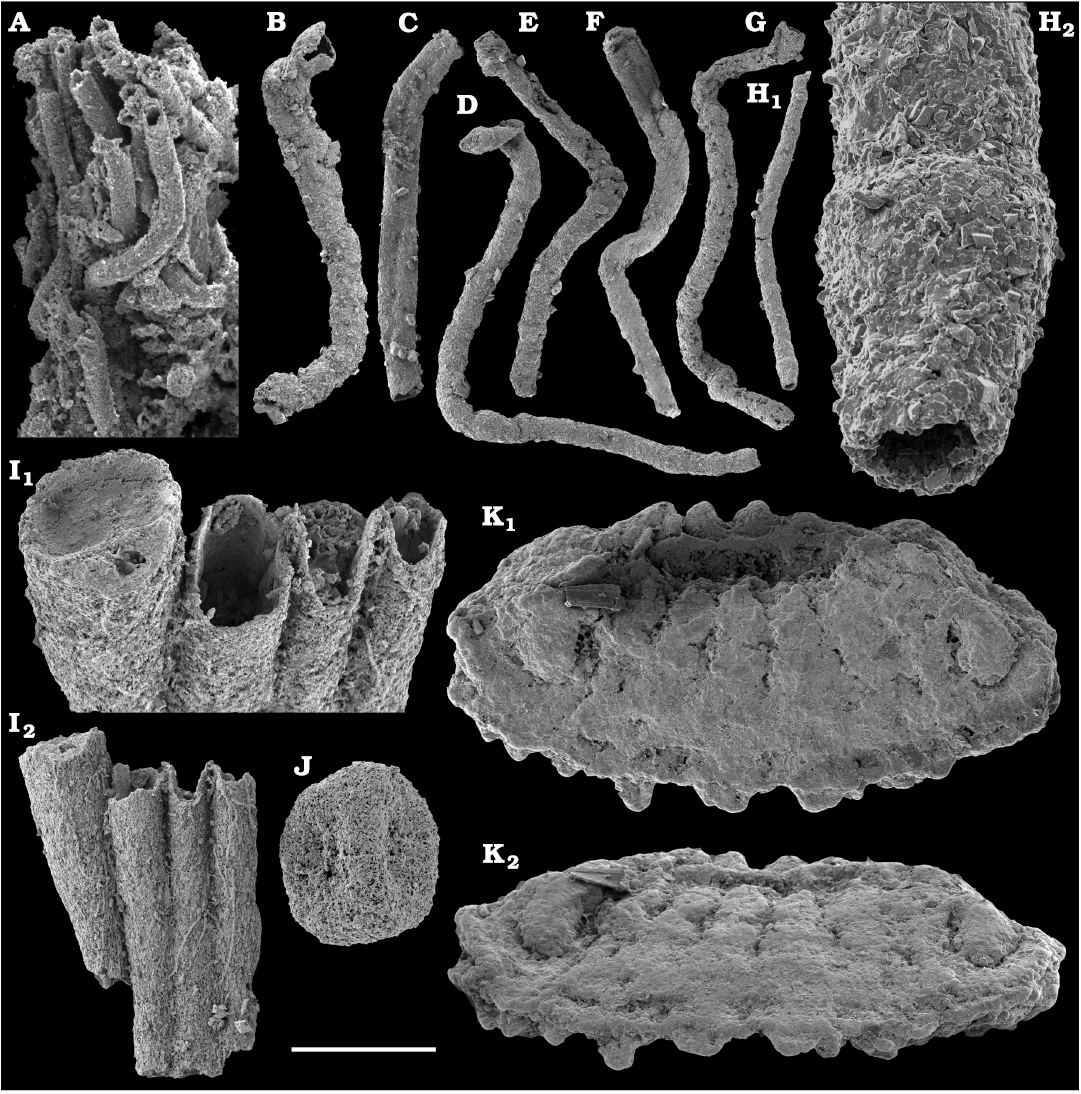
Fig. 69. Problematic fossils from the lower Cambrian Erkeket Formation, Khorbusuonka River, Siberia, Russia; samples 11/16.1 (K), 19/11.75 (A, I), 19/12.75 (B), 19/16 (D, F, G, H), 19/18 (C, E), and 19/5B (J). A. Tubular diagenetically phosphatised cyanobacterial trichomes, SMNH X11337. B–H. Problematic silicified tubes, SMNH X11338–11344, respectively. H2, enlargement showing presumably initial part of the tube. I. Problematic phosphatized tubes longitudinally merged, SMNH X11345. I1, close-up of I2, showing apertures of the tubes. J. Globula, SMNH X11346. K. Disk-like fossil, SMNH X11347. K1, plain view; K2, lateral view. Scale bar: 500 µm (B–G, H1), 250 µm (I2, J, K), 125 µm (I1), 50 µm (A, H2).
Remarks.—Coleoloides trigeminatus Missarzhevsky in Rozanov et al., 1969, was first described from the Dokidocyathus regularis Zone of the Tommotian stage, Siberian Platform as elongated, somewhat helically twisted and irregularly curved phosphatic internal moulds, ca. 0.1 mm in diameter, typically with six longitudinal ribs. The latter commonly run spirally around the longitudinal axis of the mould. The initial stage of growth has not been previously illustrated. Missarzhevsky (1982, 1989) indicated C. trigeminatus among anabaritids, but the affinity of Coleoloides to a particular group of the earliest tubular fossils remains problematic. Although cross-sections of C. trigeminatus and Anabaritellus hexasulcatus are very similar in shape, they are markedly different in diameter, which is 3–4 times greater in A. hexasulcatus and retains its six-folded cross-section at later stages of growth (Kouchinsky et al. 2009: fig. 33; Kouchinsky et al. 2017: fig. 76D). Brasier (1984: 234) and Qian and Bengtson (1989: 130–131) synonymised C. trigeminatus with C. typicalis Walcott, 1889, with the latter showing a considerable variability in number of thin longitudinal ridges on the surface of internal moulds. However, this synonymy is not supported herein.
Stratigraphic and geographic range.—Tommotian stage (correlated with the upper part of Cambrian Stage 2), Siberian Platform (Russia), and, probably, Cambrian Stage 2 of Avalonia and South China (see Kouchinsky et al. 2009).
Coleoloides cf. paucistriatus Poulsen, 1967
Fig. 68E–J.
Material.—Over ten calcium phosphatic and glauconised internal moulds from sample 11/2B, basal Erkeket Formation, Khorbusuonka River, Siberia, Russia; Dokidocyathus regularis Zone, Tommotian Stage (upper part of Cambrian Stage 2). Additional material from the Tommotian Stage, lower Pestrotsvet Formation of southeastern Siberian Platform (Russia) includes: SMNH X11334 and X11336, from sample M303/2, Tiktirikteekh Brook, middle Lena River, D. regularis Zone (see Rozanov et al. 1969: 31, text-fig. 6), SMNH X11331, from sample 171–174, Bydyangaya Brook, middle reaches of the Lena River, Nochoroicyathus sunnaginicus Zone (see Val’kov 1982: 27, text-fig. 8), and SMNH X11332, X11333, and X11335, from sample 510, section Ulakhan-Sulugur, Aldan River, N. sunnaginicus Zone (collected by Anatolij K. Val’kov).
Description.—Irregularly curved and helically twisted internal moulds with initial part (Fig. 68E) circular in cross-section and cylindrical in shape, ca. 0.5 mm long and 50–70 µm in diameter. The tubes expand slightly from the initial part and acquire two (Fig. 68E1), four and then eight longitudinal lobes running spirally clockwise around the longitudinal axis of the mould. The lobes are crest-shaped in cross-section and separated by flattened faces. New longitudinal ribs gradually intervene in the middle of the faces between the available ribs. A specimen with diameter ca. 100 µm shows 4 ribs (Fig. 68I), those ca. 150 µm in diameter have 8 ribs (Fig. 68F), and at a ca. 250 µm diameter the number of longitudinal ribs reaches 12 (Fig. 68H, J). Ribs evenly spaced and separated from each other by flattened faces, ca. twice the width of the ribs.
Remarks.—Similar to Coleoloides paucistriatus Poulsen, 1967, in having up to 12 longitudinal ribs and wide flattened faces between them. Different from co-occurring Coleoloides trigeminatus in having ridged lobes and wide flattened faces between them. There are six lobes in C. trigeminatus at the initial part, whereas two spiral lobes in C. cf. paucistriatus. Superficially similar to internal moulds of Anabarites isiticus Missarzhevsky (see Kouchinsky et al. 2009: fig. 35) from the Tommotian stage of south-eastern Siberian Platform, but A. isiticus has ca. 3 times greater diameter, higher expansion rate and consistent tri-radial symmetry. By contrast, cross-section of C. cf. paucistriatus can be four-ray symmetrical. It suggests that the general symmetry of the body in Coleoloides, of either a stable radial or bilateral type, is unlikely reflected by morphology of the tube.
Phylum Foraminifera d’Orbigny, 1826
Class, order, and family uncertain
Genus Platysolenites Pander, 1851
Type species: Platysolenites antiquissimus Eichwald, 1860; lower Cambrian (Terreneuvian Series); near Kunda, Estonia.
Platysolenites antiquissimus Eichwald, 1860
Fig. 67H, I.
Material.—SMNH X11322 and X11323, from sample 15/23, Chuskuna Formation (upper part of the Kessyusa Group), Khorbusuonka River, Siberia, Russia. Watsonella Zone, Cambrian Stage 2.
Remarks.—Somewhat compressed fragments of tubes, 0.5–1 mm long and 0.2–0.3 mm in original diameter, with granular (presumably agglutinated) composition of the wall, ca. 20 μm thick. Similar fossils are described from the Fortunian stage and lower part of Cambrian Stage 2 outcrops of the Nemakit-Daldyn, Manykay, Medvezhya and lower Emyaksin formations on the Anabar Uplift (Kouchinsky et al. 2017).
Stratigraphic and geographic range.—Uppermost Ediacaran? (Kontorovich et al. 2008; Zhuravlev et al. 2012), Terreneuvian Series of Siberia, Baltica, Laurentia, and Avalonia; Cambrian Stage 3 of Baltica, Laurentia, Avalonia, and Armorica (McIlroy et al. 2001; Streng et al. 2005, Kouchinsky et al. 2017).
Problematic tubular fossils
Fig. 69A–I.
Remarks.—Other problematic tubular fossils are represented by silicified tubes, 50–120 µm in diameter and up to 3 mm long. Their walls are smooth but showing occasional swellings probably produced or enhanced by diagenetic silicification. Originally, the tubes were not rigid that is suggested by deformations, such as irregular curvature and obliteration in places. The initial part of the tubes is open and goblet-shaped (Fig. 69H2).
An array of irregularly curved phosphatised tubes (Fig. 69A) loosely arranged in the same direction is found attached to a shell fragment (Fig. 32A2). The tubes may represent cyanobacterial trichoms. They are 5–10 µm in diameter and up to 200 µm in visible length, smooth, without visible partitioning.
A set of four incompletely preserved straight and smooth phosphatised tubes, 100–150 µm in diameter, attached to each other longitudinally may represent a fragment of tabulatomorph colonial coral (Fig. 69I). The fragment is ca. 600 µm long. Distinct septation is not revealed. One of the tubes is apparently closed by a gently concave tabula (Fig. 69I1).
Phylum Porifera Grant, 1836
Class Hexactinellida Schmidt, 1870
Order and family uncertain
Genus Thoracospongia Mehl, 1996
Type species: Thoracospongia follispiculata Mehl, 1996; Florian–Undillan stages (Wuliuan–Drumian stages), Miaolingian Series, Cambrian; Georgina Basin, Queensland, Australia.
Thoracospongia sp.
Fig. 70.
Material.—Ten siliceous spicules, including figured SMNH Sp11409–11413, from sample 19/107, Yunkyulyabit-Yuryakh Formation, Khorbusuonka River, Siberia, Russia. Anomocarioides limbataeformis Zone, Mayan stage (correlated with the Drumian Stage).
Description.—Hexact follipinules with a modified obese axial ray rounded in cross-section. The ray is strongly inflated to form a straight or curved tear-shaped ellipsoid or a globule, 0.2–0.5 mm in diameter across the widest part and 0.4–0.5 mm long. The surface of this ray ornamented with multiple longitudinal undulating ribs bearing diminutive denticles. Ribs become more prominent distally and are spaced 15–30 µm apart. Other rays, incompletely preserved and probably broken off in two of the specimens (Fig. 70C, D) are slender, tapering, bearing an axial canal circular in cross-section. The four minute paratangential rays, 15–30 µm in diameter at their junction, are crossed at the same plane at 90° from each other. An additional ray, up to 50 µm in diameter is developed on the spicule distal end, perpendicularly to the paratangential rays.
Remarks.—Hexact follipinules identical in size and morphology are common in a more diverse and abundant assemblage of spicules described as Thoracospongia cf. follispiculata Mehl, 1996, from the lower Mayan stage (Drumian Stage) Kuonamka Formation of the Anabar Uplift (Kouchinsky et al. 2011), including pentactines with robust paratangential rays and stauractins with robust conical rays. The latter may partly represent pentactine follipinules with broken-off axial ray. Stauractines have not been however recognised in the material described herein that is different from the type species and from Thoracospongia cf. follispiculata. Thoracospongia lacrimiformis Peel, 2017b, from the Henson Gletscher Formation of North Greenland (Wuliuan Stage) differs by finer but more prominent ridges (Peel 2017b, 2019) and resembles “pinnular hexactines” from the upper part of Cambrian Stage 3 of the Altay Sayan Foldbelt (Sugai et al. 2004). Unnamed obese pentactines from the Wilberns Formation of the Llano Uplift, Texas (Furongian Series) are smooth and possess more prominent paratangential rays (Rigby 1975). In addition to Thoracospongia Mehl, 1996, follipinules of broadly the same habit but with additional paratangential and secondary rays were described under the name Rigbyella Mostler and Mosleh-Yazdi, 1976, from Iran. This generic name has been proved to be preoccupied and subsequently replaced by Rigbykia Özdikmen, 2009. The set of Thoracospongia-type spicules described as Rigbyella sp. from the Miaolingian Burj Formation of Jordan includes both swollen and relatively slim varieties (Elicki 2011). In younger beds, acanthose hexactine and pentactine follipinnules occur in association with regular triaxon spicules in the Guzhangian Stage equivalent of Australia (Bengtson 1986), Cambrian Stage 10 of Baltica (Castellani et al. 2012) as well as in the Ordovician (Carrera and Maletz 2014; for further discussion see also Peel 2017b, 2018, 2019).
Despite drastic differences of local spicule assemblages, hexactinellid and demosponge macroscleres dominate and more species-specific microscleres are distinguished in rare cases. A few early Cambrian microscleres were described by Reitner and Mehl (1995) and Botting et al. (2015) in thin sections but none of them are convincing enough due to difficulty to distinguish microscleres from polycystine tests and broken macroscleres. Macroscleres in Cambrian spicule assemblages appear to have microsclere counterparts from the Furongian, Ordovician and later periods, such as Thoracospongia follipinule-like elements, Tanjaella equinate hexactines, Asterodesma aster-like sphaeroclones and many others (Rigby 1975; Castellani et al. 2012; Carrera and Maletz 2014; Kruse and Reitner 2014). All these spicules are at least one order larger than their microsclere twins. Thus, either microsclere-bearing sponges originated from their macrosclere-bearing predecessors or Cambrian poriferans belonged mostly to extinct lineages being replaced during the latest Cambrian–Ordovician radiation. The finds of intermediate in size spicules in the middle Cambrian (Chen and Dong 2008), however, supports the suggestion of a gradual transition from sponges having larger skeletal elements to those with skeletons strengthened by microscleres.
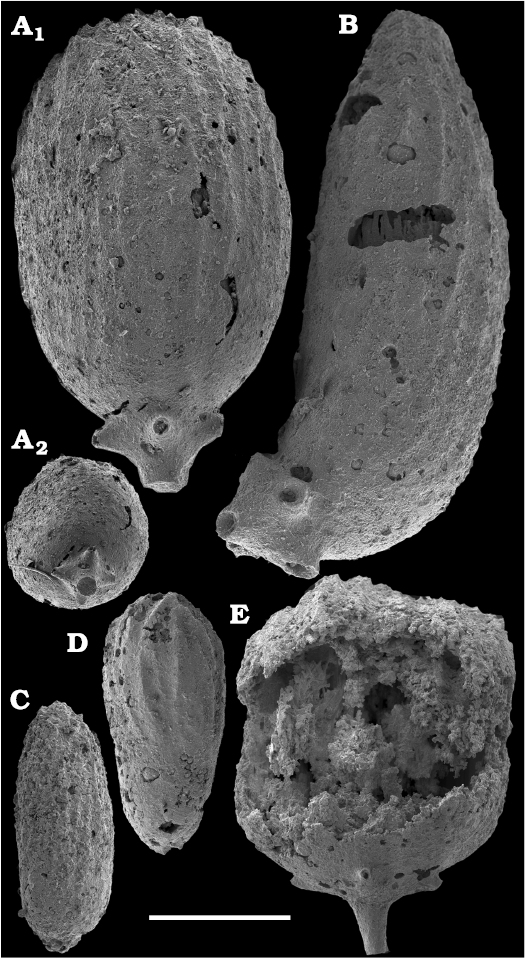
Fig. 70. Spicules of hexactinellid Thoracospongia sp. from the middle Cambrian Yunkyulyabit-Yuryakh Formation, Khorbusuonka River, Siberia, Russia; sample 19/107. A–E. Follipinules with a modified obese axial ray, SMNH Sp11409–Sp11413, respectively. A1, lateral view on the axial ray; A2, four paratangential rays. Scale bar: 125 µm (A) and 250 µm (B–F).
Genus Abnormisella Fedorov in Fedorov and Pereladov, 1987
Type species: Abnormisella insperata Fedorov in Fedorov and Pereladov, 1987; Kounamkites Zone, Amgan stage (correlated with the Wuliuan Stage), Cambrian, Kuonamka Formation; Siberian Platform, Russia.
Abnormisella sp.
Fig. 71K, L.
Material.—Single spicules, SMNH Sp11416, from sample 11/18.7, lower Erkeket Formation, Khorbusuonka River and SMNH Sp11415, from sample 6/0.3, Emyaksin Formation, Bol’shaya Kuonamka River (see Kouchinsky et al. 2015a), Siberia, Russia. Dokidocyathus regularis Zone, Tommotian stage (correlated with the upper part of Cambrian Stage 2).
Description.—Pentactine megascleres with four paratangential rays diverging from their intersection at 150° with the axial ray and at 50–60° from each other. Rays with smooth surface, gently tapering and circular in cross-section.
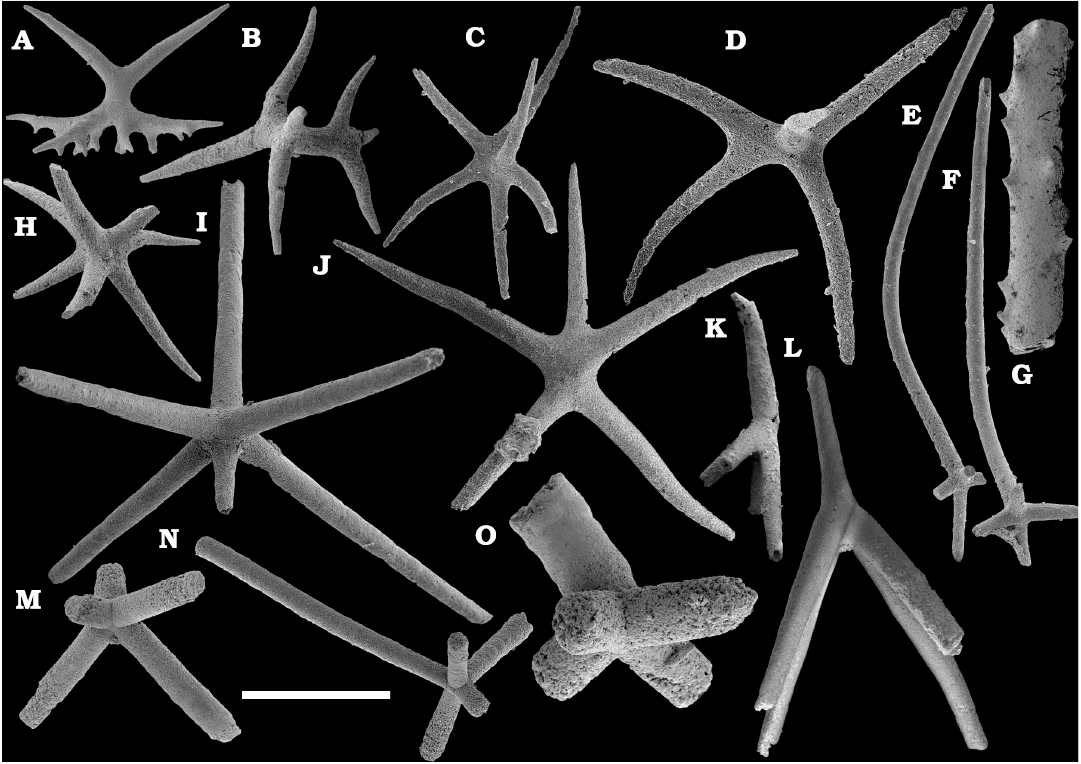
Fig. 71. Hexactinellid spicules from the lower Cambrian Erkeket (A–F, H–J, L–O) and Yunkyulyabit-Yuryakh (G) formations, Khorbusuonka River and Emyaksin Formation, Bol’shaya Kuonamka River (K), Siberia, Russia; samples 11/18.7 (A–F, H–J, L), 11/16.05 (M, N), 19/107 (G), 20/1B (O), and 6/0.3 (K). A–D, H, J. Polyactine siliceous megascleres with acutely tapering slim curved rays, SMNH Sp11424–11429, respectively. E, F, I, M–O. Pentactines and hexactines with thin smooth rays, SMNH Sp11417–11422, respectively. G. Fragment of a pinnular pentactine, SMNH Sp11423. K, L. Abnormisella sp., SMNH Sp11415 (K) and 11416 (L). Scale bar: 250 µm (A–D, H–J, O) and 500 µm (E–G, K–N).
Remarks.—Without defining a holotype, Fedorov and Pereladov (1987) attributed a number of co-occurring pentactines and stauractines from the middle Cambrian Kuonamka Formation to Abnormisella insperata Fedorov in Fedorov and Pereladov, 1987, including pentactines with a long axial ray and four paratangential rays diverging at 90–180° with respect to the axial ray. Subsequently, Peel (2019) selected a lectotype and emended diagnosis of the genus and its type species. The emended diagnosis included only pentactines with inclined axial ray, thus appearing bilaterally symmetrical. Forms found in Cambrian Stage 4–Guzhangian Stage of North Greenland attributed by Peel (2019) to A. insperata have an acanthose axial ray of a greater diameter than those of paratangential rays arranged in two pairs having different cross-sectional diameters, that is not the case in the spicules described herein and by Kouchinsky et al. (2015a). Peel (2019) attributed Speciosuspongia wangcunensis Chen and Dong, 2008, from the Drumian (middle Cambrian) of South China to the same genus, but they have thicker and swollen acanthose axial rays. The latter form has more in common with “anchorate” spicules from the Atdabanian (Cambrian Stage 3) Bagrad Formation of the Batenev Range, Altay Sayan Foldbelt (Sugai et al. 2004), which in addition to pentactines include elements with short duplicated paratangential rays. Spear-shaped spicules occur in the lower Tommotian stage Emyaksin and Erkeket formations (Kouchinsky et al. 2015a: fig. 70A and Fig. 71L herein), but their morphology varies including hexactines, with an additional ray directed away from the axial ray (Kouchinsky et al. 2015a: fig. 70B), as well as pentactines with their axial ray diverging from the axis of symmetry of the spicule (Fig. 71K).
Other siliceous spicules in the assemblage from sample 11/18.7 herein include polyactine megascleres with acutely tapering slim slightly curved rays (Fig. 71A–D, H, J). Among them, there are tetractines (Fig. 71A) with three acutely tapering straight rays arranged in the same plane at an angle 120° from each other (paratangential rays) and a thicker straight axial ray perpendicular to them. This ray, as well as one of the radial rays, branch dichotomously at 120° to each other. Additional short stubby spines may be present at the external surface of paratangential rays (Fig. 71A).
Fedorov in Pegel et al. (2016) compared these spicules with plagiotriaenes and phyllotriaenes of “lithistid” sponges and with diactines but these spicules are modified tetractines. Some demosponge ectosomal phyllotriaene megascleres resemble closely Bistella Fedorov in Pegel et al., 2016, in both morphology and size range (e.g., Schuster et al. 2015: fig. 4C). Morphologically these elements are comparable with homosclerophorid tetractines and polycystine (radiolarian) point-centered spicules but are much larger following the data of Uriz (2006) on extant sponges and of Maletz (2011) for the oldest siliceous polycystines. However, their co-occurrence with typical hexactinellid spicules in the same sampling sets does not exclude the association of both spicule types to the same sponge skeletons.
Pentactines and hexactines with thin smooth rays
Fig. 71E, F, I, M–O.
Material.—Several tens of spicules, including SMNH Sp11417–11422, from samples 11/16.05 and 11/18.7, Dokidocyathus regularis Zone, Tommotian stage (correlated with the upper part of Cambrian Stage 2) and 20/1B, lower Botoman stage. Erkeket Formation, Khorbusuonka River, Siberia, Russia.
Remarks.—Siliceous spicules consisting of five to six thin smooth rays. The rays are almost cylindrical and taper very slowly. Four paratangential rays directed away from the axial ray, at 100–110° and at 80–100° to each other in planar view. Axial ray can be much longer than tangential rays and curved (Fig. 71E, F). A single spicule (Fig. 71I) with sixth ray situated on the same axis as the axial ray and opposite to the latter.
The specimens described herein are most similar to those illustrated by Kouchinsky et al. (2015a: fig. 70D), from the Tommotian Stage, Emyaksin Formation and Kouchinsky et al. (2011: fig. 40R–V), from the Amgan Stage (correlated with the Wuliuan Stage), Kuonamka Formation, Anabar Uplift on the Siberian Platform, although their relatively simple morphology precludes undoubted assignment of such Tommotian and younger spicules to the same species.
Specimens with a long curved axial ray (Fig. 71E, F) resemble pentactines with an acanthose axial ray, described from the Atdabanian (correlated with the Cambrian Stage 3) beds of the Emyaksin Formation (Kouchinsky et al. 2015a: fig. 70M).
Pinnular pentactine
Fig. 71G.
Material.—Single broken spicule, figured SMNH Sp11423, from sample 19/107, Yunkyulyabit-Yuryakh Formation, Khorbusuonka River, Siberia, Russia. Anomocarioides limbataeformis Zone, Mayan stage (correlated with the Drumian Stage).
Description.—Pentactine-type megasclere with a long axial ray bearing multiple distally directed barbs. The ray is roughly square in cross section. Ray diameter 0.75 mm.
Remarks.—Despite fragmentary preservation, the pinnular axial ray is typical of pentactines widely distributed in Cambrian strata (Bengtson 1986; Mehl 1998; Ivantsov et al. 2005a; Kouchinsky et al. 2011, 2015a; Peel 2018). Pegel et al. (2016) formally described such spicules from the Atdabanian (correlated with the Cambrian Stage 3) Udachny Formation of the Anabar Uplift as Tanchajella Fedorov in Pegel et al., 2016. Of a great interest, are pinnular pentiactines having been acid-extracted from the Cambrian Stage 4 Forteau Formation of Newfoundland, Canada, which are preserved as wholly carbonaceous films and interpreted as the remains of robust organic spicule sheaths of the calcarean type (Harvey 2010).
Class Calcarea Bowerbank, 1864
Subclass Calcinea Bidder, 1899
Order uncertain
Family Polyactinellida Mostler, 1985
Genus Dodecaactinella Reif, 1968
Type species: Dodecaactinella oncera Reif, 1968; Porkuni stage (Hirnantian Stage), Upper Ordovician; Estonia.
Dodecaactinella sp.
Fig. 72A–H.
Material.—Over a hundred of dolomitised spicules and fragments from samples 19/1.5, 19/3.25, 19/4.25, 19/5.5, 19/8.5, 19/10.25, 19/11.75, 19/25.5, 19/31.75, including figured SMNH Sp 11430–11437, Erkeket Formation, Khorbusuonka River, Siberia, Russia. Botoman and Toyonian stages (correlated with the Cambrian Stage 4).
Description.—Equiangular symmetrical triactine-type spicules, typically ca. 0.5 mm in size, up to 1 mm in the available material. The main rays diverge at 120° and branch dichotomously (Fig. 72B, D, E, G) or trichotomously (Fig. 72A, C) in the same plane at ca. 50 µm from the center of the spicule to its periphery. All rays 20–80 µm in diameter, do not taper and are smooth.
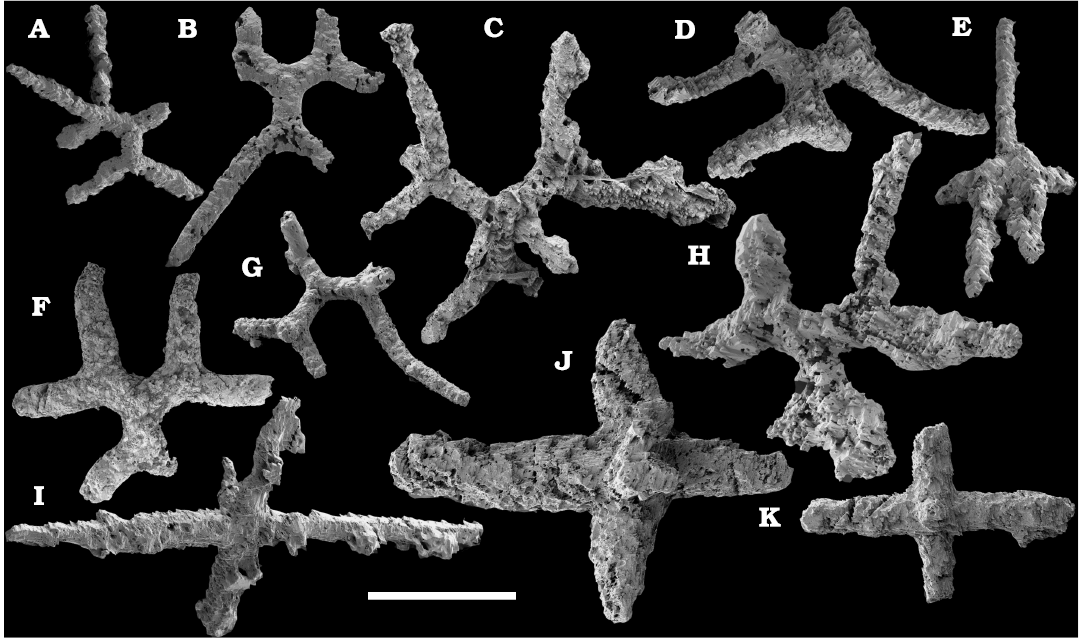
Fig. 72. Calcareous (dolomitised) spicules from the lower Cambrian Erkeket Formations, Khorbusuonka River, Siberia, Russia; samples 19/4.25 (H, I), 19/5.5 (A, J), 19/8.5 (D), 19/10.25 (C), 19/11.75 (E, G), 19/25.5 (B, F), and 20/1B (K). A–H. Dodecaactinella sp., SMNH Sp11430–11437, respectively. I–K. Sulugurella sp., SMNH Sp11438–11440, respectively. Scale bar 250 µm.
Remarks.—Such acid-extracted spicules from different localities (Mostler 1985, 1996; Shabanov et al. 1987; Bengtson et al. 1990; Dong and Knoll 1996; Peel 2019) display faces of corroded crystals of calcite or dolomite constituting their recrystallised, originally high-magnesium calcite rays. Such a composition of spicules as well as their distinct triactine symmetry fits to those of the Calcarea (e.g., Jones and Jenkins 1970; Uriz 2006).
Reif (1968) assigned dodecaactinellid spicules to hexactinellids, but Rigby and Toomey (1978) correctly attributed them to calcareans. Dodecaactinella-like triactines are globally distributed in the Cambrian strata beginning from the Cambrian Stage 2 (Reif 1968; Mostler 1985; Shabanov et al. 1987; Bengtson et al. 1990; Wood et al. 1993; Elicki 1994; Kruse et al. 1995; Dong and Knoll 1996; Mehl 1998; Sugai et al. 2004; Wrona 2004; Ivantsov et al. 2005a; Skovsted and Peel 2010; Peel 2019). Besides Dodecaactinella Reif, 1968, such spicules are known as Phobetractinia Reif, 1968, Sardospongia Mostler, 1985, Polyactinella Mostler, 1985, Bengtsonella Mostler, 1996 (preoccupied name replaced by Mostlerhella Özdikmen, 2009), and Pseudosardospongia Fedorov in Pegel et al., 2016. They differ by the presence or absence of the third- and fourth-order branching and additional rays, probably resulting from incomplete fragmentation of fused pharetronid-type skeletons. The latter are known from Bottonaecyathus Vologdin, 1940, and Gravestockia Reitner, 1992, and may therefore belong to different species and genera. While relatively large branching Bottonaecyathus participated in early Cambrian reef building in Tuva and Mongolian microcontinents, small Gravestockia was a cryptobiont in metazoan reefs and microbialites of Australia (Zhuravlev 2001).
Class incertae sedis
Order Heteractinida Hinde, 1887
Hexaradiate spicules
Fig. 73.
Material.—Several hundred phosphatised spicules, including figured SMNH Sp11441–11459, from samples 19/5.5, 19/8.5, 19/10.25, 19/11.75, 19/12.75, 19/29, Erkeket Formation, Khorbusuonka River, 22/0 and 22/50, from the uppermost Tyuser and Sekten formations, lower reaches of the Lena River. Siberia, Russia; lower Botoman and Toyonian stages (correlated with the Cambrian Stage 4).
Description.—Spicules with six paratangential rays regularly radiating from a central disk at ca. 60º between each other in the plan view and with a slight angle in the lateral view with respect to the plane of the central disk. One side of the central disk is slightly concave and smooth. The opposite, convex side can be smooth (Fig. 73B), or bear a short axial ray with a robust base slightly inclined towards the central disk (Fig. 73F1, H, I, J, O: arrow). In other forms, the convex side of the spicule carries 10–15 accessory rays radiating from the central disk (Fig. 73G, K–S). The accessory rays can be short and blunt or more prominent and pointed.

Fig. 73. Heteractinid spicules from the lower Cambrian Erkeket Formations, Khorbusuonka River, Siberia, Russia; samples 19/10.25 (A–F, I), 19/11.75 (H, J, P, R), and 19/12.75 (G, K–O, Q, S). A–S. Hexaradiate spicules, SMNH Sp11441–11459, respectively. C–E, S, spicules with dissolved internal parts of the rays reminiscent of axial canals. F1, spicule in lateral view; F2, concave side of the spicule with radiating six paratangential rays. H, I, strongly eroded spicules. G, K–S, hexaradiate spicules with accessory rays. Arrow in O points to fused spicules. Arrow in S points to dissolved inner portion of the ray reminiscent of axial canal. Scale bar: 250 µm (C–G, J, M–O, Q, S) and 500 µm (A, B, H, I, K, L, P, R).
Remarks.—In a number of publications on Cambrian spicules, such hexaradiates are ascribed to the heteractinid sponge Eiffelia Walcott, 1920 (e.g., Bengtson et al. 1990; Skovsted 2006a). The type material of Eiffelia is represented by complete skeletons consisting of tetraradiate spicules and flat hexaradiate spicules with long lateral rays and central disks (Walcott 1920; Botting and Butterfield 2005). Rays perpendicular to the plane of the other four rays were observed in some of the tetraradiates, whereas all hexaradiate spicules had no evidence of perpendicular rays (Botting and Butterfield 2005: 1555). Bengtson et al. (1990), who described very similar hexaradiate spicules from upper Cambrian Stage 3–lower Stage 4 of Australia as Eiffelia araniformis (Missarzhevsky in Mambetov and Missarzhevsky, 1981), synonymised several species of Lenastella Missarzhevsky in Missarzhevsky and Mambetov, 1981, from coeval deposits of Kazhakhstan with E. araniformis, as well as Actinoites Duan, 1984, and Niphadus Duan, 1984, from South China. Mehl (1998) described such hexaradiates from the Drumian Stage of Australia. Spicules from Kazakhstan and Siberia have more massive bases of their rays and larger central disks compared to slender rays and relatively smaller central disks in Australian forms. Nodes on the convex side of central disk observed in the specimens from Australia are not revealed in the material from Kazakhstan and Siberia. Hexaradiate spicules of E. araniformis (Missarzhevsky in Missarzhevsky and Mambetov, 1981) do not have accessory rays but occasionally a short single distal ray.
Peel (2016, 2019) described spicules of Eiffelia floriformis Peel, 2019, from Cambrian Stage 4 Henson Gletscher Formation, Peary Land, North Greenland and suggested that six- and four-rayed spicules of undetermined heteractinids from the lower Botoman stage Emyaksin Formation of the Anabar Uplift in Siberia (Kouchinsky et al. 2015a: fig. 72A, C–E, G, H) can be attributed to the same species. Hexaradiates and co-occurring tetraradiate forms with accessory rays (see below) described herein are, however, different in having rays radiating at a slight angle to the central disk, longer accessory rays, and long axial rays in tetraradiates.
In some chemically extracted specimens, a star-like hollow depression is present (Fig. 73C–E). Probably, the hollow space within the central discs of hexaradiates and axial parts of their rays appeared owing to dissolution of a primary labile carbonate mineral, such as aragonite, and incomplete phosphatisation of the core. This suggestion is also corroborated by broken off and subsequently rounded and phosphatised tips of rays. A fibrous microstructure of these elements is indicative of their probable primary aragonite mineralization in the form of acicular radiating needles (Fig. 73C, D). Such a microstructure and composition are not typical of any extant sponges but occur in radiocyaths, which two-walled globular and pyriform rigid skeletons are built of giant spicule-like elements, meroms, consisting of distal and proximal star-like nesasters connected by long shafts (Kruse et al. 2015).
Spicules of the heteractinid Eiffelia Walcott, 1920, and the protomonaxonid sponge Lenica Gorjansky, 1977, were interpreted as bimineralic, namely calcitic with a silica core (Botting and Butterfield 2005; Bengtson and Vinther 2006; Bengtson and Collins 2015; Nadhira et al. 2019; Drozdov et al. 2022). Such a complex mineralogy could potentially lead to a selective dissolution and phosphatisation, but appears to be a less plausible explanation of preservational features observed herein.
Tetraradiate spicules with accessory rays
Fig. 74A–H, J, L–P, S.
Material.—Several hundred phosphatised spicules, including figured SMNH Sp11460–11474, from samples 19/10.25, 19/11.75, 19/12.75, Erkeket Formation, Khorbusuonka River and sample 22/0, uppermost Tyuser Formation, Lena River. Siberia, Russia; lower Botoman stage (correlated with the Cambrian Stage 4).
Description.—Spicules with four rays radiating from central disk, respectively, at ca. 90º between each other in the plan view, with a slight angle in the lateral view. A single long ray emerges perpendicular to the central disk. On the opposite side of the spicule, 10–15 accessory rays radiate from the central disk. Those can be knobby or more prominent and pointed. A single central prominent and pointed ray can be present among them.

Fig. 74. Heteractinid spicules from the lower Cambrian Erkeket Formation, Khorbusuonka River (A–E, G, I, K–Q, V, Y) and Tyuser Formation, lower reaches of the Lena River (F, H, J, U, W, X, Z) , Siberia, Russia; samples 11/16.05 (Y), 19/10.25 (E, I, K, L, O–Q), 19/11.75 (A–D, R–T, V), 19/12.75 (G, M, N), 21/21 (U, W, X), and 22/0 (F, H, J, Z). A–H, J, L–P, S. Polyactinal spicules with an axial ray, SMNH Sp11460–11474, respectively. U, W–Y. Inflated polyactinal spicules from the Tommotian Stage, SMNH Sp11475–11478, respectively. I, K, Q, R, T, V, Z. Microcoryne cf. cephalata Bengtson in Bengtson et al., 1990, SMNH Sp11479–11485, respectively. Scale bar: 250 µm (A–J, N, O, Q, R, V, Y, Z) and 500 µm (K–M, P, S, T, U, W, X).
Remarks.—In general such skeletal elements resemble basic heteractinid sponge spicules possessing a longer and more massive proximal ray, which is directed inwards the sponge wall (Pickett 2002). Fused spicules and mostly broken rays indicate that these elements have formed relatively rigid and moderately thick-walled skeletons, which are typical of a number of heteractinids (Pickett 2002).
Polyactinal spicules with multiple coplanar rays
Fig. 75.
Material.—Several tens of phosphatised spicules, including figured SMNH Sp11486–11494, from samples 19/11.75 and 19/12.75, Erkeket Formation, Khorbusuonka River, Siberia, Russia. Lower Botoman stage (correlated with the Cambrian Stage 4).
Description.—Discoidal polyactinal stellate spicules, 0.3 to 0.7 mm in diameter, consisting of coplanar flattened and tapering rays with blunt apical terminations. The number of rays varies from 6 to 15. The disc surface is smooth, with one side slightly concave and the other one slightly convex. In some cases, a circlet of ca. 6 primary, longer rays can be distinguished from secondary, shorter and usually thinner rays radiating on the concave side of the spicule.

Fig. 75. Heteractinid spicules from the lower Cambrian Erkeket Formation, Khorbusuonka River, Siberia, Russia; samples 19/11.75 (C–I), and 19/12.75 (A, B). A–I. Polyactinal spicules with multiple coplanar rays, SMNH Sp11486–11494, respectively. C1, plainview; C2, lateral view. Scale bar: 250 µm (A–H) and 500 µm (I).
Remarks.—A fibrous microstructure of these elements is indicative of their probable primary aragonitic mineralization in the form of acicular radiating units (Fig. 75F, H). Such a microstructure and composition are not typical of any extant sponges but occur in radiocyaths. Among them, forms without proximal nesasters occur as well as those with a single layer in the wall consisting of nesasters without shafts (Zhuravlev and Sayutina 1985). The earliest radiocyath, Gonamispongia ignorabilis Korshunov, 1968, derives from the Pestrotsvet Formation of Uchur-Maya region, Siberian Platform, from the Dokidocyathus lenaicus–Tumuliolynthus primigenius Zone of the Tommotian stage (Korshunov 1968; Zhuravlev and Sayutina 1985). Their wall consists of distal nesasters with proximal shafts. The forms discussed herein apparently lack shafts originally, because their attachment sites cannot be determined. Overall morphology also suggests a similarity with heteractinid spicules described above.
Genus Microcoryne Bengtson in Bengtson et al., 1990
Type species: Microcoryne cephalata Bengtson in Bengtson et al., 1990; Cambrian Stage 3, Ajax Limestone; Arrowie Basin, South Australia.
Microcoryne cf. cephalata Bengtson in Bengtson et al., 1990
Fig. 74I, K, Q, R, T, V, Z.
Material.—Several tens of phosphatised spicules, including figured SMNH Sp11479–11485, from samples 19/10.25, 19/11.75, and 19/12.75, Erkeket Formation, Khorbusuonka River and from sample 22/0, Tyuser Formation, Lena River. Siberia, Russia; lower Botoman stage (correlated with the lower part of Cambrian Stage 4).
Description.—Mace-shaped phosphatised polyactinal spicules, up to 1 mm long and 0.4 mm wide, consisting of a tuberculated or spinose head and an axial (proximal?) ray. The axial ray is 70–130 µm in diameter and reaches 0.7 mm in length, but may be indistinguishable in some forms (Fig. 74I). Up to three (in the available specimens) elongated and pointed paratangential rays are inclined towards to the axial ray, at ca. 30º (Fig. 74Q). They are slender and shorter than the axial ray and radiate from a common part of spicule, the head. The entire surface of the head is covered with numerous short blunt tubercles and longer spines. They tend to be radially grouped in 3–4 circlets, but may also be disorderly arranged on the entire head surface (Fig. 74Z). The spicules, now phosphatised, consist of aciculate longitudinally aligned elements.
Remarks.—Bengtson et al. (1990) suggested that these microfossils from the Cambrian Series 2 of South Australia can belong to octocorals, based on their microstructure of acicular needle-shaped radiating crystals. Similar spicules were reported from other parts of Gondwana, such as Germany and Jordan (Elicki 1994, 2011) and occur through the Cambrian Stage 3–Miaolingian Series. These microfossils are also ascribed to heteractinid spicules (Kouchinsky et al. 2015a). Peel (2019: fig. 4A, J) suggested sponge affinities based on the presence of axial canal in a single specimen of Microcoryne sp. found in the Cambrian Stage 4 of North Greenland. If high morphological variability of Microcoryne is not taken into consideration, formal genera and species are described such as Conwaymorrisispongia bengtsoni (Kozur et al. 2008), which type material even is derived from the type localities of Microcoryne cephalata (Bengtson et al. 1990). Some spicules may also be thickened by a secondarily phosphatised mass consisting of aciculate elements with a probably originally aragonitic composition (Fig. 74T). We interpret such spicules as belonging to heteractinids with originally aragonitic polycrystalline composition and occasionally aragonitic cement overgrowth.
Heteractinid elements co-occur in the lower Botoman samples with fragments of archaeocyaths represented by phosphatised infills of canals in the walls (Fig. 78A–G) and with presumably eroded and rounded fusiform fragments of polyactinal spicules (Fig. 78H–P).
Class, order, and family incertae sedis
Genus Nabaviella Mostler and Mosleh-Yazdi, 1976
Type species: Nabaviella elegans Mostler and Mosleh-Yazdi, 1976; Miaolingian–Furongian Series, Cambrian, Mila Formation; Alborz Mountains, Iran.
Nabaviella sp.
Fig. 76.
Material.—Four calcium phosphatic internal moulds and coatings, SMNH Sp11495–11498, from samples 19/5.5 and 20/1B, Erkeket Formation, Khorbusuonka River, Siberia, Russia. Lower Botoman stage (correlated with the lower part of Cambrian Stage 4).
Description.—Clavulate spicules consisting of a prominent straight slowly tapering axial ray (shaft), which bears a bulbous swelling (Fig. 76C arrowed) at presumably proximal end of the spicule, and a star-like disc (umbel) at the other (presumably distal) end. The disc, up to 0.5 mm in diameter, consists of 11–13 short rays slightly flattened proximally and recurved distally. The shaft is up to 1.3 mm long (Fig. 76C), circular in cross-section, up to 0.2 mm in diameter at the disc (Fig. 76A). A transversally broken spicule (Fig. 76B1) displays a wide axial canal having a circular cross-section, ca. 0.1 mm in diameter at the disc. There is a short distal knob, probably representing a reduced axial ray on the opposite side of the disc (Fig. 76A2 and D1 arrowed). The moulding surface of the spicule displays an irregular scalloped ornamentation (Fig. 76B).
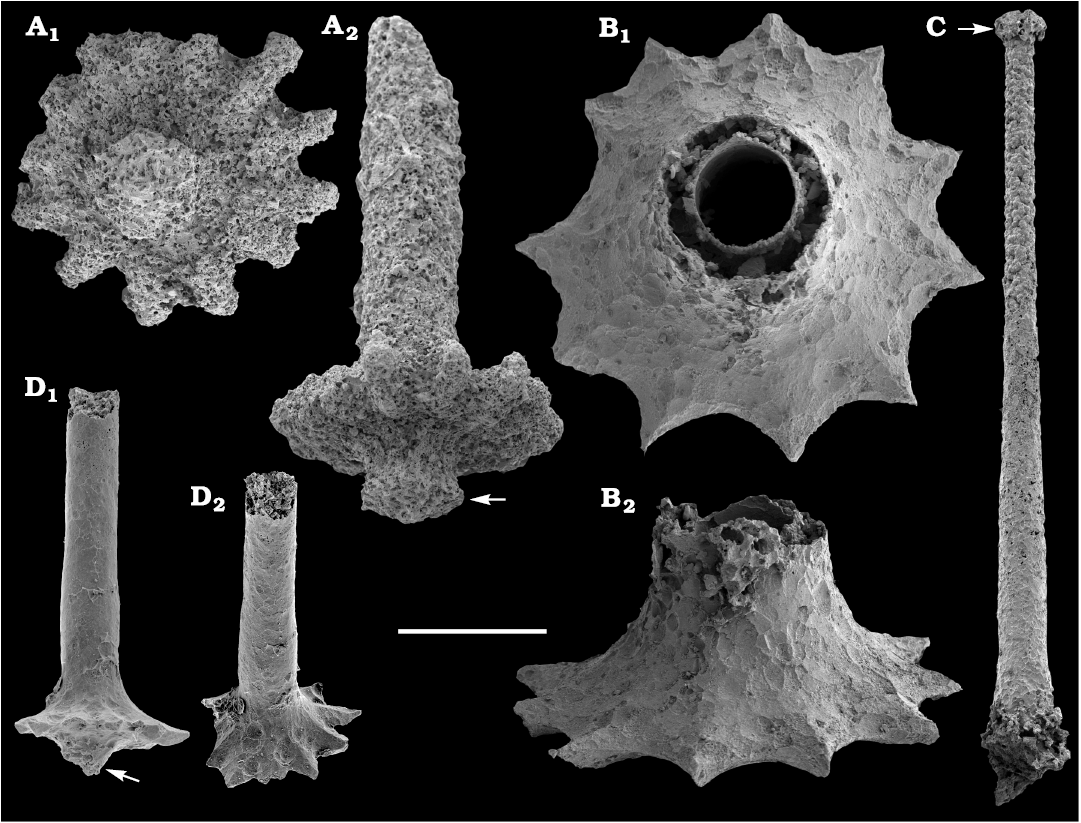
Fig. 76. Spicules of Nabaviella sp. from the lower Cambrian Erkeket Formation, Khorbusuonka River, Siberia, Russia; samples 20/1B (A) and 19/5.5 (B–D). A. Calcium phosphatic mould of the spicule with the proximal (proximal part of the spicule) shaft and a terminal knob (arrowed in A2) at the opposite surface of the distal disc (distal part of the spicule) consisting of radiating rays (A1), SMNH Sp11495. B. Calcium phosphatic coatings incompletely replicating the proximal surface of the disc with basal parts of the rays and internal axial canal in the spicule, SMNH Sp11496. B1, plain view; B2, lateral view. C. Internal mould of the proximal shaft with terminal proximal swelling (arrow), SMNH Sp11497. D. Calcium phosphatic internal mould of the axial canal and internal cavity of the distal disc and terminal knob (arrowed in D1), SMNH Sp11498. Scale bar: 125 µm (B) and 250 µm (A, C, D).
Remarks.—Mostler and Mosleh-Yazdi (1976) figured spicules with 7–11 rays as syntypes of their type species, including a clavulate anchoring spicule with eight curved paratangential rays as the holotype. This microfossil is compared with either chancelloriid sclerites (Mostler and Mosleh-Yazdi 1976) or with hexactinellid spicules, including hemidiscs (Mostler 1985), clavules (Webby and Trotter 1993) and tylodiscs, and demosponge paraclavules (Reitner and Mehl 1995). All these spicule types, however, are microscleres. Peel (2019) erroneously quoted “N. gracilis Mostler and Mosleh-Yazdi, 1976” as the type species, but there was no such a species in the original paper. Nabaviella spicules can be also morphologically compared to calcareous meroms of radiocyaths. By contrast, meroms lack axial canals and distal knobs but have straight or slightly recurved radiating and bifurcating coplanar rays at both proximal and distal ends (Zhuravlev and Sayutina 1985).
Genus Sulugurella Fedorov in Pelman et al., 1990
Type species: Sulugurella sulugurica Fedorov in Pelman et al., 1990; Nochoroicyathus sunnaginicus Zone, Tommotian stage (correlated with the upper part of Cambrian Stage 2); Aldan River, Siberian Platform, Russia.
Sulugurella sp.
Fig. 72I–K.
Material.—Several dolomitised spicules and fragments from samples 19/4.25, 19/5.5, 20/1B, including figured SMNH Sp11438–11440, Erkeket Formation, Khorbusuonka River, Siberia, Russia. Lower Botoman stage (correlated with the lower part of Cambrian Stage 4).
Description.—Incomplete pentact spicules with four rays situated in the same plane at an angle 90° between each other. The paratangential rays are 50–100 µm in diameter and somewhat thinner than a perpendicular axial ray.
Remarks.—The surface of spicules is strongly corroded and displays crystal faces of secondary dolomite replacing original high-magnesium calcite. Calcareous pentactines as well as hexactines and tetractines with the overall spicule morphology typical of hexactinellids are relatively common in lower Cambrian strata and described under the generic name Calcihexactina Sdzuy, 1969 (e.g., Brock and Cooper 1993; Peel 2019). In the original description of the type species, Calcihexactina franconia Sdzuy, 1969, from the Wuliuan Stage of the Franconian Forest, Germany, Sdzuy (1969) indicated axial canals that is absent from calcarean spicules. Thus, the calcareous composition in Sulugurella-type spicules is secondary and they can be attributed to hexactinellids. On the other hand, they can be originally calcitic and represent remains of some stem sponges possessing hexactine spicules composed partly or wholly of calcite (Botting and Muir 2018).
Inflated hexactines
Fig. 77.
Material.—Several tens of hexactines, including figured SMNH Sp11500–11518, from samples 19/10.25, 19/11.75, 19/12.75, Erkeket Formation, Khorbusuonka River, lower Botoman Stage (correlated with the lower part of Cambrian Stage 4), sample 11/16.05, basal Erkeket Formation and sample 21/21, Tyuser Formation, Lena River, Dokidocyathus regularis Zone, Tommotian Stage (correlated with the upper part of Cambrian Stage 2). Siberia, Russia.
Description.—Phosphatised spicules, 0.4–1.0 mm in size, composed of six tapering rays with smooth surface. The opposite rays are situated along three axes crossed at nearly right angles with each other. Presence of axial canals is not evident owing to preservation. Some of the spicules have relatively slim rays and regular hexact morphology (Fig. 77A, G, I). In other spicules (Fig. 77C–F, J–S), the central part is swelled and can be almost spherical (Fig. 77P), 300–500 µm in diameter. Strongly inflated spicules with conical rays lacking axial canals occur (Fig. 77D). In some of these forms, rays are underdeveloped or eroded and form short knobs.

Fig. 77. Inflated hexactines from the lower Cambrian Erkeket Formation, Khorbusuonka River (A–N) and Tyuser Formation, lower reaches of the Lena River (O–S), Siberia, Russia; samples 19/10.25 (C, D, H, K), 19/11.75 (A, B, E, F, J, L, M), 19/12.75 (G, I), 11/16.05 (N), and 21/21 (O–S). A, B, G–I. Hexactines with smooth tapering rays, SMNH Sp11500–11504, respectively. C–F, J–S. Strongly inflated hexactines with smooth tapering rays, SMNH Sp11505–11518, respectively. Scale bar 500 µm.
Remarks.—Inflated hexactines with conical rays are described as Cjulanciella asimmetrica Fedorov in Fedorov and Pereladov, 1987 from the Ovatoryctocara–Schistocephalus Zone, Amgan stage (correlated with the uppermost Stage 4–lowermost Wuliuan Stage), Kuonamka Formation, Siberia. Cjulanciella asimmetrica demonstrates a high variety of siliceous spicules in their assemblages. The type species syntypes include diactines, tetractines and pentactines (Fedorov and Pereladov 1987). The oldest allied spicules are detected in the uppermost Atdabanian (correlated with the Cambrian Stage 3) Emyaksin Formation in the south-western Anabar Uplift of the Siberian Platform (Shabanov et al. 1987). Similar spicules were reported from the Cambrian Stage 3 of the Altay Sayan Foldbelt in Siberia (Sugai et al. 2004) and Wuliuan–Drumian stages of the Siberian Platform (Kouchinsky et al. 2011: fig. 40E–G), lower Cambrian of Laurentia (Skovsted and Holmer 2006: fig. 5G, H), middle–upper Cambrian of Australia (Bengtson 1986: fig. 9E; Mehl 1998: pl. 1: 1), South China (Ding et al. 1992; Dong and Knoll 1996: fig. 7.1; Steiner et al. 2007), and upper Cambrian of Argentina (Heredia et al. 1987). Mehl (1996, 1998) considered such spicules as a part of a single dermal sponge skeleton built in conjunction with Thoracospongia-type follipinules but at present in situ associations have been found only in the Wuliuan Henson Gletscher Formation of North Greenland (Peel 2017b, 2019).
Phosphatised hexactines with simple rays co-occur with hexa- and tetraradiated spicules of heteractinids in our material (Fig. 77) and some of them may derive from the same scleritomes. Hexactines vary in size 2–3 times and some of them have a swelled axial zone. Presence of such spicule types in the heteractinid assemblages from the lower Botoman stage of northern Siberian Platform is noted by Kouchinsky et al. (2015a). Such spicules also occur in the Dokidocyathus regularis Zone, Tommotian stage (correlated with the upper part of Cambrian Stage 2), wherefrom hexaradiate heteractinid spicules are unknown. Although such hexactins with conical and often swelled rays may represent siliceous hexactinellid spicules diagenetically replaced by carbonate and phosphate minerals, axial cavities may have originated from dissolution of carbonate under phosphatic coatings during chemical extraction. The origin of such spicules is thus not clear, but some of them (Fig. 74U, W, X, Y) are very similar to tetraradiates with accessory distal rays described above from heteractinids. It cannot therefore be excluded that first heteractinids with tetraradiate spicules appeared in the upper part of Cambrian Stage 2.
Class Archaeocyatha Bornemann, 1884
Calcareous, originally consisting of high-magnesium calcite cups of archaeocyaths are prone to dissolution either during diagenesis or rock treatment. Internal moulds of canals and cavities in the cups or calcium phosphatic veneers replicating skeletal structures allow, however, recognition of the original morphology of archaeocyaths in three-dimensional details. Phosphatised fragments of archaeocyaths are preserved as a part of small shelly fossil assemblages, found in the Tommotian stage of Siberia (herein), Cambrian Stage 3 Salaany-Gol Formation of the Mongolian Zavkhan Terrane (Debrenne et al. 1990; Pruss et al. 2019), Shuijingtuo Formation of the Three Gorges area of the Yangtze Platform (Zhang et al. 2016), and Puerto Blanco Formation of Mexico (Laurentia, Devaere et al. 2019: fig. 23), Cambrian Stage 4 Browns Pond Formation of the Eastern Laurentian Taconic allochthon (Landing and Bartowski 1996; AZ own observation in Ed Landing’s collection 2008), Bastion Formation of North-East Greenland (Skovsted 2006a) and glacial erratics of Antarctica (Wrona 2004). Archaeocyaths cups from Cambrian Stage 4–Wuliuan Stage, Henson Gletscher Formation were interpreted as an octocoral by Peel (2017a: fig. 4) or as problematic fossils from Series 2 of Australia by Laurie (1986: fig. 10). Archaeocyaths, preserved in calcium phosphate, resemble in some cases superficially other groups and can be confused with ecdysozoan moults (i.e., Microdictyon), ossicles of echinoderms preserved in calcite and fragmented along the cleavage planes in monocrystals (Fig. 64) and phosphatised fragments of hyolithid helens (Kouchinsky et al. 2015a: fig. 32 and Fig. 55 herein).
Order Ajacicyathida R. Bedford and J. Bedford, 1939
Suborder Ajacicyathina R. Bedford and J. Bedford, 1939
Superfamily Ethmophylloidea Okulitch, 1937
Family Carinacyathidae Krasnopeeva, 1953
Genus Carinacyathus Vologdin, 1932
Type species: Carinacyathus loculatus Vologdin, 1932; Botoman stage (correlated with the Cambrian Stage 4); Lebed’ River, Altay Mountains, Altay Sayan Foldbelt, Russia.
Carinacyathus sp.
Fig. 79D.
Material.—A single phosphatised fragment, SMNH Sp11542, from sample 20/1B, Erkeket Formation, Khorbusuonka River, Siberia, Russia. Lower Botoman stage (correlated with the lower part of Cambrian Stage 4).
Remarks.—Phosphatic crust replicating a fragment of a thick cup wall with geniculate tubular canals, elliptical in cross-section and 30–50 µm across. Among forms of archaeocyaths described from the lower Botoman part of the Erkeket Formation, tubular noncommunicating (lacking lateral pores) canals from the inner wall are known only from Carinacyathus, while other genera are characterised either by simple inner walls or flattened and S-shaped in vertical section scales. From investigation of Carinacyathus cups in thin sections, it was suggested that all of the species have the inner wall with downwardly projecting, straight canals, connected to the central cavity by supplementary structures, bracts (Debrenne et al. 2015). The new material indicates that the inner wall is pierced by geniculate (V-shaped in longitudinal section) canals without bracts.
Ajacicyathina family, genus, and species indet.
Fig. 79A, H, I.
Material.—Three phosphatised fragments, SMNH Sp11535–11537, from sample 20/1B, Erkeket Formation, Khorbusuonka River, Siberia, Russia. Lower Botoman stage (correlated with the lower part of Cambrian Stage 4).
Remarks.—Phosphatic crusts replicating up to 6 mm long fragments of double walled initial parts of cups with simple rounded pores on the outer and inner walls and on the straight septa (vertical plate-like elements connecting the walls). Even distribution of rounded pores in combination with strait septa in the intervallum (cup space limited by walls) and absence of tabulae (horizontal plate-like intervallar elements) are indicative of the suborder Ajacicyathina. This suborder existed during the Tommotian–Toyonian stages (correlated with the Cambrian stages 2–4) and most of its representatives are known to have simple and almost undistinguishable morphologies at early stages of growth.
Order Capsulocyathida Zhuravleva in Zhuravleva et al., 1964
Suborder Capsulocyathina Zhuravleva in Zhuravleva et al., 1964
Family Cryptoporocyathidae Zhuravleva, 1960
Genus Cryptoporocyathus Zhuravleva, 1960
Type species: Cryptoporocyathus junicanensis Zhuravleva, 1960; Medvezh’ya Formation, Tommotian stage (correlated with the upper part of Cambrian Stage 2); Moyero River, Siberian Platform, Russia.
Cryptoporocyathus junicanensis Zhuravleva, 1960
Fig. 79B.
Material.—Three phosphatised fragments, including SMNH Sp11538, from sample 21/21, Tyuser Formation, Lena River, Siberia, Russia. Dokodocyathus regularis Zone, Tommotian stage (correlated with the upper part of Cambrian Stage 2).
Description.—Phosphatic crusts replicating wall fragments penetrated by evenly distributed pores of two distinct sizes. Larger pores are elliptical in vertical direction, up to 200 × 100 µm in diameters. Smaller pores are rounded or elongated, up to 40 µm long and 20 µm wide. Inner (Fig. 79B1) and outer (Fig. 79B2) surfaces of the wall are similar. Traces of septa or tabula attachment are not observed.
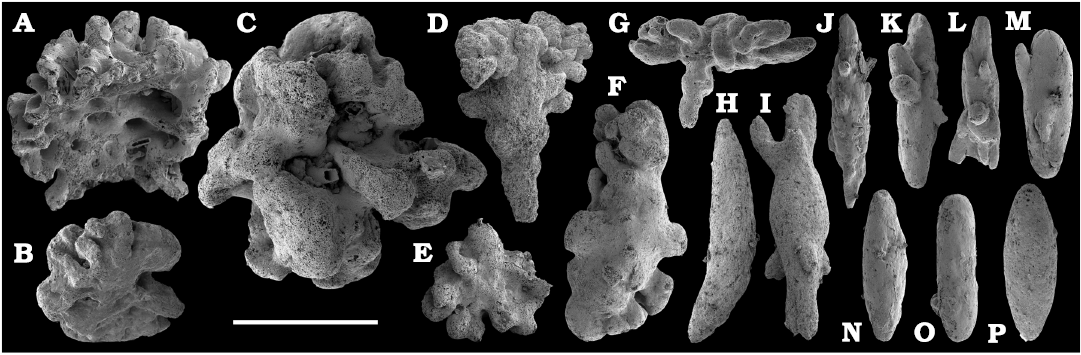
Fig. 78. Sponge fragments from the lower Cambrian Erkeket Formation, Khorbusuonka River (A–F, H, I, M, N) and Tyuser Formation, lower reaches of the Lena River (G, J–L, O, P), Siberia, Russia; samples 19/10.25 (B–F, M, N), 19/11.75 (A, H, I), and 22/0 (G, J–L, O, P). A–G. Fragments of archaeocyaths, SMNH Sp11519–11525, respectively. H–P. Fragments of possible heteractinid spicules, SMNH Sp11526–11534, respectively. Scale bar: 250 µm (A, C) and 500 µm (B, D–P).
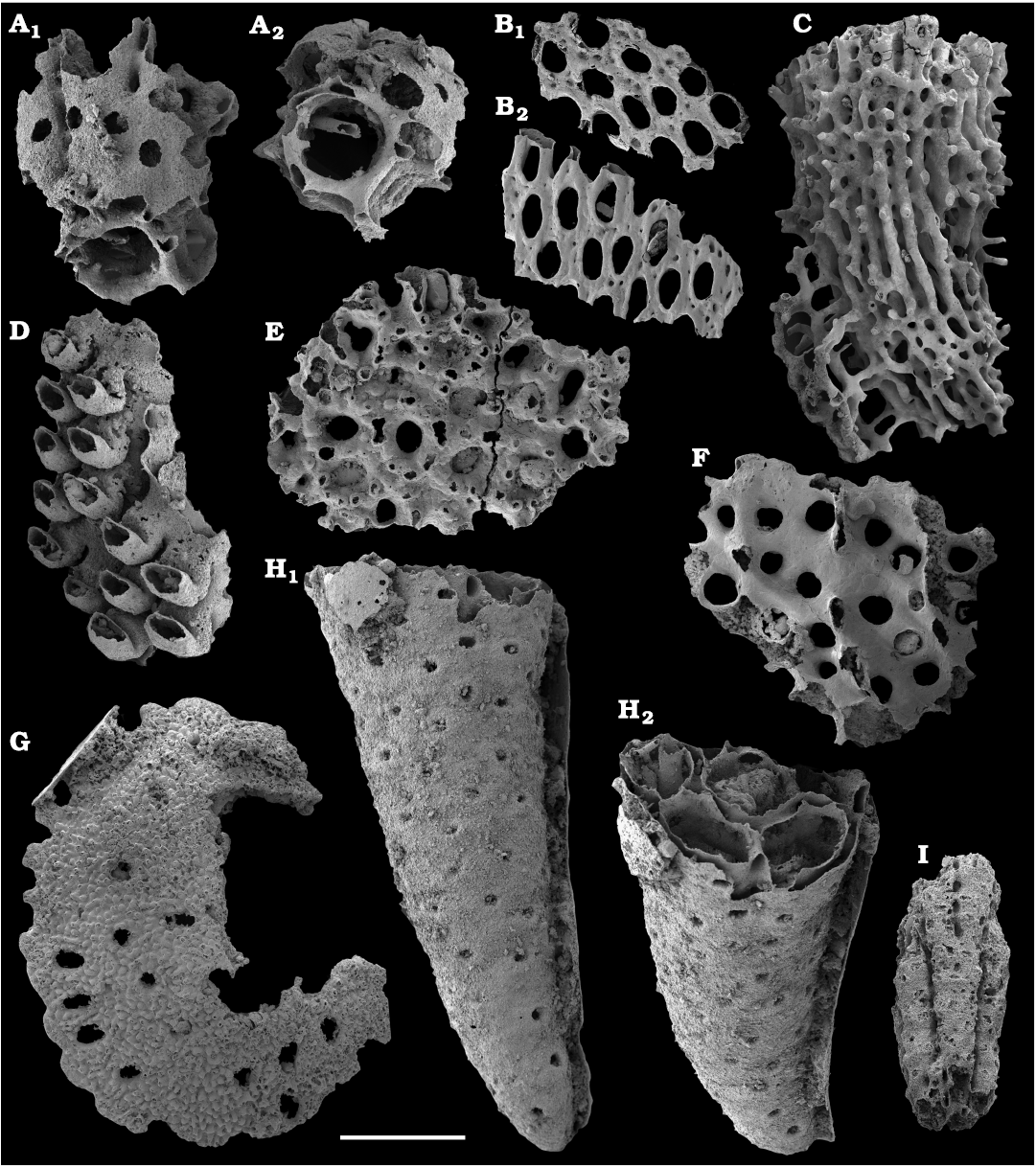
Fig. 79. Archaeocyatha, phosphatized fragments from the lower Cambrian Erkeket (A, D, H, I) and Chuskuna (E) formations, Khorbusuonka River and Tyuser Formation, lower reaches of the Lena River (B, C, F, G), Siberia, Russia; samples 15/23 (E), 20/1B (A, D, H, I), and 21/21 (B, C, F, G). A, H, I. Ajacicyathina family, gen. and sp. indet., fragments of cups with simple rounded pores, SMNH Sp11535–11537, respectively. A1, H1, lateral view; A2, basal view; H2, oblique lateral view showing inner part of the cup. B, E. Cryptoporocyathus junicanensis Zhuravleva, 1960. B. Wall fragments, SMNH Sp11538–11539, respectively. B1, inner surface of the wall. B2, outer surface of the wall. E. Wall fragment, SMNH Sp11540. C. Dictyocyathus sp., fragment of an intervallar dictyonal network, SMNH Sp11541. D. Carinacyathus sp., fragment of a cup wall with geniculate tubular canals, SMNH Sp11542. F. Cambrocyathellus sp., fragment of a cup, SMNH Sp11543. G. Possible archaeocyath pelta (upper covering structure of the cup), SMNH Sp11544. Scale bar: 1.25 mm (C), 500 µm (B, F–I), and 250 µm (A, D, E).
Remarks.—The only archaeocyath possessing an outer wall with such a type of the porosity and lacking any intervallar elements on the entire Siberian Platform is Cryptoporocyathus junicanensis Zhuravleva, 1960, belonging to single chambered capsulocyathines (Zhuravleva 1960; Debrenne et al. 2015). A similar form occurring in the Tommotian and lowermost Atdabanian strata but studied in thin sections only, Korshunovicyathus melnikovi (Korshunov and Zhuravleva, 1967) is characterised by an attached microporous sheath on the outer wall, rather than smaller pores situated between larger ones (Debrenne et al. 2015).
A similar wall fragment, SMNH Sp11539 (Fig. 79E), from sample 15/23 is found in the Chuskuna Formation, upper Kessyusa Group, at the Khorbusuonka River, beds attributed to the Watsonella Zone, Cambrian Stage 2. The fragment shows irregularly distributed rounded and elliptical pores, up to 70 µm across, smaller in diameter than those in Cryptoporocyathus junicanensis fragments from sample 21/21. They are surrounded by a pustulose structures with smaller pores. Archaeocyath fragments from the upper 10–12 m of the Kessyusa Group were previously reported without illustration by Missarzhevsky (1989) and Astashkin et al. (1991: 70) (see Kouchinsky et al. 2012: SOM, appendix 1).
Stratigraphic and geographic range.—Cambrian Stage 2, Siberia, Russia.
Order Archaeocyathida Okulitch, 1935
Suborder Loculicyathina Zhuravleva, 1955
Superfamily Loculicyathoidea Zhuravleva, 1954
Family Loculicyathidae Zhuravleva, 1954
Genus Cambrocyathellus Zhuravleva, 1960
Type species: Cambrocyathellus tschuranicus Zhuravleva, 1960; Pestrotsvet Formation, Tommotian stage (correlated with the upper part of Cambrian Stage 2); Churan Village, Lena River, Siberian Platform, Russia.
Cambrocyathellus sp.
Fig. 79F.
Material.—A single phosphatised fragment SMNH Sp11543 from sample 21/21, Tyuser Formation, Lena River, Siberia, Russia. Dokodocyathus regularis Zone, Tommotian stage (correlated with the upper part of Cambrian Stage 2).
Remarks.—Fragment of the outer wall pierced by relatively large pores of rounded shapes and uneven sizes, 80–140 µm in dimeter. Ridges representing remains of vertical plate-like elements are observed on the inner surface of the wall. Outer walls with irregularly situated pores of variable sizes are more characteristic of archaeocyathids than ajacicyathids, while the presence of septum-like elements infers the affinity to the Loculicyathina (Debrenne et al. 2015). Two vertical rows of pores per each interseptal space is typical of Cambrocyathellus Zhuravleva, 1960, which is known on the Siberian Platform from the Tommotian and lowermost Atdabanian (correlated with the lower part of the Cambrian Stage 3) strata (Zhuravleva 1960; Zhuravlev et al. 1983).
Suborder Archaeocyathina Okulitch, 1935
Superfamily Dictyocyathoidea Taylor, 1910
Family Dictyocyathidae Taylor, 1910
Genus Dictyocyathus Bornemann, 1891
Type species: Dictyocyathus tenerrimus Bornemann, 1891; Matoppa Formation, Cambrian Stage 4; Cuccuru Contu, Sardinia, Italy.
Dictyocyathus sp.
Fig. 79C.
Material.—A single phosphatised fragment SMNH Sp11541 from sample 21/21, Tyuser Formation, Lena River, Siberia, Russia. Dokodocyathus regularis Zone, Tommotian stage (correlated with the upper part of Cambrian Stage 2).
Remarks.—Phosphatic crust replicating fragment of intervallum built of relatively regular dictyonal network. The latter consists of evenly spaced vertical and radial interpore lintels connected by synapticulae (rod-like horizontal elements of the same size). Such a structure within the entire intervallar space is typical of a number of forms within the Archaeocyathina, but of them, Dictyocyathus is the only one genus known on the Siberian Platform from the Tommotian and Atdabanian strata (Debrenne et al. 2015).
Conclusions
Carbon isotope results are first reported herein from the upper Kessyusa Group and lower Erkeket Formation from two sections studied along the Khorbusuonka River. The upper part of the Kessyusa Group contains two positive excursions and is correlated with the lower part of Cambrian Stage 2 in other regions of Siberia and probably worldwide. Finds of Watsonella crosbyi at the base of the second (upper) excursion provides a biostratigraphic marker of the Watsonella Zone. Carbon isotope excursion II in lower beds of the Erkeket Formation in those sections is characteristic of the Dokidocyathus regularis Zone of the Tommotian stage. New carbon isotope data presented herein from the Bulkur anticline in lower reaches of the Lena River suggest the upper D. regularis Zone, with excursion III for basal beds of the middle Tyuser Formation. Updated lowermost occurrence of trilobites in the Ulakhan-Ald’yarkhay section by Bushuev et al. (2014) corresponds to the lower boundary of the Atdabanian stage situated in the stratotype at the rising trend of the δ13C excursion IV. Positive peaks II–VII revealed in the sections studied can be correlated with respective excursions known from the stratotype area in the Aldan–middle Lena rivers’ region, as well as from the Sukharikha River and eastern flanks of the Anabar Uplift on the Siberian Platform. The δ13C record from the Khorbusuonka and Lena rivers reported herein can be correlated with globally recognised carbon isotopic features. Those include δ13C troughs, SHICE (SHIyantou Carbon isotope Excursion), AECE (Archaeocyathid Extinction Carbon isotope Excursion), and DICE (Drumian Carbon isotope Excursion) corresponding, respectively, to values preceding Siberian peak II, between the paired peaks VII–VIII and IX–X, and across the Wuliuan-Drumian stages boundary. Positive excursions EAREZE (Early Atdabanian/REpinaella Zone Excursion), CARE (Cambrian Arthropod Radiation isotope Excursion) and MICE (MIngxinsi Carbon Isotope Excursion) are recognised herein as, respectively, Siberian peak IV, peak V, and merged peaks VII and VIII. The lower Judomia Zone boundary in section 3 following our new chemostratigraphy is tentatively placed higher than that suggested by biostratigraphy. A positive excursion named herein LACE (Lower Anomocarioides limbataeformis Carbon isotope Excursion) is revealed from the Yunkyulyabit-Yuryakh Formation of the Khorbusuonka River.
Combined chemo- and biostratigraphic correlation facilitates study of evolutionary radiation of animals and faunas through the taphonomic window provided by early diagenetic phosphatisation in the Terreneuvian and Cambrian Series 2 of the Siberian Platform and elsewhere. Phosphatisation preserved diverse skeletal faunas in similar facial environments associated with the onlap deposits near commonly diachronous transgressive boundaries. Skeletal fossils chemically extracted from Cambrian normal marine carbonates are reported herein from the Tommotian–Mayan regional stages (correlative with Cambrian Stage 2–Drumian Stage) of the Siberian Platform. In addition to higher diversity assemblages from the Dokidocyathus regularis Zone of the Tommotian stage (correlated with the upper part of Cambrian Stage 2) and Calodiscus–Erbiella Zone (Bergeroniellus micmacciformis–Erbiella Zone) of the lower Botoman stage (correlated with the lower part of Cambrian Stage 4), the material includes lower-diversity fossil assemblage from the Toyonian–lower Amgan stages (Cambrian Stage 4) yet poorly characterised from Siberia. No species in common from the middle Tommotian and lower Botoman rich assemblages herein are identified, separated from each other by ca. 7 Ma. Such a profound difference is corroborated by previously suggested episode of extinction in the late Tommotian, spanned by the III/IV trough in δ13C record (see e.g., Landing and Kouchinsky 2016: fig. 1), prior to rebound diversification in the early Atdabanian and during Cambrian Age 3 (Zhuravlev and Wood 1996; Zhuravlev 2001).
New representative of the stem group Brachiopoda with previously unknown combination of features typical of tannuolinids, mickwitziids, linguliform brachiopods and most similar to Oymurania (Kouchinsky et al. 2015b; Kouchinsky and Bengtson 2017) is recovered from the lower Botoman Stage of Khorbusuonka. From the same beds, first findings of tannuolinids, Tannuolina cf. pavlovi Kouchinsky, Bengtson, and Murdock, 2010, from the lower Cambrian Stage 4 of the Siberian Platform are illustrated. Sclerites of Lapworthella lucida Meshkova, 1969, first described from the upper Tyuser Formation by Meshkova (1969: 167, pl. 53: 5), are reported herein from the type locality and from the lower Botoman stage of the Khorbusuonka River. Lapworthella dentata Missarzhevsky in Rozanov et al. 1969, previously known from Cambrian Stage 3 strata on the Siberian Platform (see Kouchinsky et al. 2015a) is herein reported from the Toyonian stage beds (Stage 4) at Khorbusuonka River.
The bivalve Pojetaia dentifera Kouchinsky, Bengtson, Clausen, Gubanov, Malinky, and Peel, 2011, is identified from the Toyonian stage (correlated with the Cambrian Stage 4) of the lower Lena River and extends significantly the lower range of this species previously known from the lower Drumian Stage (Kouchinsky et al. 2011). Reinterpretation of preservational mode of sclerites of an aculiferan mollusc, Halkieria proboscidea (Meshkova, 1969) prompted emended description of the species originated from the Tommotian stage of Siberia and Gondwana.
Apical septum, apertural operculum and fibrous microstructure of the wall with pores in planispiral evolute shells of Michniakia minuta Missarzhevsky in Rozanov and Missarzhevsky, 1966, from the lower Botoman stage of Khorbusuonka is similar to those found in hyoliths (Feng et al. 2001; Kouchinsky et al. 2017). Phosphatised hyolithid appendages (helens) are found in the lower Botoman (correlated with the lower part of Cambrian Stage 4) strata of Khorbusuonka River and the first appearance of appendages is registered in the upper Dokidocyathus regularis Zone, Tommotian stage (correlated with the upper part of Cambrian Stage 2) that is consistent with the early diversification of hyolithid hyoliths in the upper Stage 2.
First finds of problematic cambroclavid sclerites in Siberia, including Cambroclavus sp. and Zhijinites clavus (Mambetov in Mambetov and Repina, 1979), are reported from the Botoman stage (correlated with the lower part of Cambrian Stage 4) of the Khorbusuonka River.
Archaeopetasus pachybasalis Kouchinsky sp. nov. and Archaeopetasus typicus (Sipin, 1999) from the Dokidocyathus regularis Zone, Tommotian stage (correlated with the upper Cambrian Stage 2) are reinterpreted as external sclerites attributed to chancelloriid-like animals. Co-occurring chancelloriids are represented by sclerites of Stellispinella lenaica (Zhuravleva and Korde, 1955) and other forms.
New species of palaeoscolecidans, Hadimopanella foveata Kouchinsky sp. nov. is described from the Toyonian–lower Amgan stages (correlated with the Cambrian Stage 4) of the Khorbusuonka River, where Hadimopanella knappologica (Bengtson, 1977) also occur in the lower Botoman stage. The bradoriid Mongolitubulus reticulatus Kouchinsky, Bengtson, Clausen, Gubanov, Malinky, and Peel, 2011, is reported from the Cambrian Stage 4, thus extending the range of the species described previously from the Wuliuan Stage (Kouchinsky et al. 2011).
Chevron-like proximal stem plates and a few associated columnals may represent the earliest rhombiferans reported herein from the lower Botoman stage (lower part of Cambrian Stage 4). Together with ambulacral flooring and thecal plates of edrioasteroids, brachial uniserial and brachiolar biserial, a mosaic of problematic thecal plates co-occurs in the lower Botoman assemblages of echinoderm ossicles and illustrates rapid radiation in the Echinodermata by the early Cambrian Age 4 (Kouchinsky et al. 2011, 2015a; Clausen and Peel 2012; Zamora et al. 2013; Peel et al. 2016). Ctenidium plates of ctenocystoids are first reported herein from the middle part of the Cambrian Stage 4.
Two species of problematic tubicolous fossils, Coleoloides trigeminatus Missarzhevsky in Rozanov et al., 1969, and Coleoloides cf. paucistriatus Poulsen, 1967, from the Tommotian stage of the Siberian Platform are distinguished following different morphology of their initial parts and later growth stages.
Acknowledgements
Our field trips to Malaya Kuonamka and Bol’shaya Kuonamka rivers in 1996 and to the Khorbusuonka and Lena rivers in 2000 were financially supported by grants to SB from the Royal Swedish Academy of Sciences and the Swedish Research Council. We thank Shane Pelechaty (Shell, Calgary, Canada) for collecting isotope samples from outcrops of the Malaya Kuonamka and Bol’shaya Kuonamka rivers in 1996, Stefan Ohlsson (Swedish Museum of Natural History, Stockholm, Sweden) and Klara Hajnal (Stockholm University, Sweden) for technical assistance with sample preparation and analyses. AK is grateful to Ilya Kuchinsky (International English School in Täby, Sweden) for assistance with SEM imaging. Colin Chilcott is thanked for technical support at the University of Edinburgh, UK. The authors are grateful to Ed Landing (New York State Museum, Albany, USA) and Sarah Jacquet (University of Missouri, Columbia, USA) for their invaluable comments on the manuscript. LH’s work was supported by a grant from the Swedish Research Council (VR 2018-03390). AZ’s study is carried out as a part of Scientific Project 04-1-21 of the State Order of the Government of the Russian Federation to the Lomonosov Moscow State University No. 121031600198-2. RW and FB are supported by a NERC Discovery Grant (NE/T008458/1). VP has been supported by the Kazan Federal University Strategic Academic Leadership Program.
References
Abaimova, G.P. 1978. Late Cambrian conodonts of central Kazakhstan [in Russian]. Paleontologičeskij žurnal 1978 (4): 77–87.
Abaimova, G.P. and Ergaliev, G.K. 1975. Finds of conodonts in the Middle and Upper Cambrian of Malyj Karatau [in Russian]. Trudy Instituta Geologii i Geofiziki Sibirskogo Otdeleniâ Akademii Nauk SSSR 333: 390–394.
Aguinaldo, A., Turbeville, J., Linford, L., Rivera, M.C., Garey, J.R., Raff, R.A., and Lake, J.A. 1997. Evidence for a clade of nematodes, arthropods and other moulting animals. Nature 387: 489–493. Crossref
Astashkin, V.A., Pegel, T.V., Repina, L.N., Rozanov, A.Y., Shabanov, Y.Y., Zhuravlev, A.Y., Sukhov, S.S., and Sundukov, V.M. 1991. Cambrian System on the Siberian Platform. Correlation chart and explanatory notes. International Union of Geological Sciences Publication 27, 1–133.
Babcock, L.E., Robison, R.A., Rees, M.N., Peng, S., and Saltzman, M.R. 2007. The Global boundary Stratotype Section and Point (GSSP) of the Drumian Stage (Cambrian) in the Drum Mountains, Utah, USA. Episodes 30: 85–95. Crossref
Balthasar, U., Holmer, L.E., Skovsted, C.B., and Brock, G.A. 2009. Homologous skeletal secretion in tommotiids and brachiopods. Geology 37: 1143–1146. Crossref
Barrande, J. 1881. Systême Silurien du centre de la Bohême, Vol. 6, Classe des Mollusques. Ordre des Acéphalés. 342 pp. Charles Bellmann, Prague.
Bedford, R. and Bedford, J. 1939. Development and classification of Archaeos (Pleospongia). Memoirs of the Kyancutta Museum 6: 67–82.
Bell, W.C. 1941. Cambrian Brachiopoda from Montana. Journal of Paleontology 16: 193–255.
Bengtson, S. 1970. The Lower Cambrian fossil Tommotia. Lethaia 3: 363–392. Crossref
Bengtson, S. 1976. The structure of some Middle Cambrian conodonts, and the early evolution of conodont structure and function. Lethaia 9: 185–206. Crossref
Bengtson, S. 1977. Early Cambrian button-shaped phosphatic microfossils from the Siberian Platform. Palaeontology 20: 751–762.
Bengtson, S. 1986. Siliceous microfossils from the Upper Cambrian of Queensland. Alcheringa 10: 195–216. Crossref
Bengtson, S. 1992. The cap-shaped Cambrian fossil Maikhanella and the relationship between coeloscleritophorans and molluscs. Lethaia 25: 401–420. Crossref
Bengtson, S. and Collins, D. 2015. Chancelloriids of the Cambrian Burgess Shale. Palaeontologia Electronica 18: 1–67.
Bengtson, S. and Vinther, J. 2006. The role of Eiffelia in early sponge evolution. Geological Society of America, Abstracts with Program 38: 304.
Bengtson, S. and Yue, Z. 1997. Fossilized metazoan embryos from the earliest Cambrian. Science 277: 1645–1648. Crossref
Bengtson, S., Conway Morris, S., Cooper, B.J., Jell, P.A., and Runnegar, B.N. 1990. Early Cambrian fossils from South Australia. Memoirs of the Association of Australasian Palaeontologists 9: 1–364.
Bengtson, S., Matthews, S.C., and Missarzhevsky, V.V. 1986. The Cambrian netlike fossil Microdictyon. In: A. Hoffman and M.H. Nitecki (eds.), Problematic Fossil Taxa. Oxford Monographs on Geology and Geophysics 5, 97−115. Oxford University Press, New York.
Bidder, G.P. 1899. The skeleton and classification of calcareous sponges. Proceedings of the Royal Society of London 64: 61–76. Crossref
Billings, E. 1858. On the Asteriadae of the Lower Silurian rocks of Canada. Geological Survey of Canada, Figures and Descriptions of Canadian Organic Remains 3: 75–85. Crossref
Billings, E. 1871. On some new species of Palaeozoic fossils. Canadian Naturalist 6: 213–223, 240.
Bornemann, J.G. 1884. Bericht über die Fortsetzung seiner Untersuchungen cambrischer Archaeocyathus-Formen und verwandter Organismen von der Insel Sardinien. Zeitschrift der Deutschen Geologische Gesellschaft 36: 702–706.
Bornemann, J.G. 1891. Die Versteinerungen des cambrischen Schichtensystem der Insel Sardinien. Zweite Abteilung. Nachschrift, III. Archaeocyathinae. Nova Acta Academiae Caesareae Leopoldino-Carolinae Germanicae Naturae Curiosorum 56: 495–500.
Botting, J.P. and Butterfield, N.J. 2005. Reconstructing early sponge relationships by using the Burgess Shale fossil Eiffelia globosa Walcott. Proceedings of the National Academy of Sciences 102: 1554–1559. Crossref
Botting, J.P. and Muir, L.A. 2018. Early sponge evolution: A review and phylogenetic framework. Palaeoworld 27: 1–29. Crossref
Botting, J.P. and Peel, J.S. 2016. Early Cambrian sponges of the Sirius Passet Biota, North Greenland. Papers in Palaeontology 2: 463–487. Crossref
Botting, J.P., Yuan, X., and Lin, J.P. 2015. Tetraradial symmetry in early poriferans. Chinese Science Bulletin 59: 639–644. Crossref
Bowerbank, J.S. 1864. A Monograph of the British Spongiadae. Vol. 1, 290 pp. Ray Society. London. Crossref
Brasier, M.D. 1984. Microfossils and small shelly fossils from the Lower Cambrian Hyolithes Limestone at Nuneaton, English Midlands. Geological Magazine 121: 229–253. Crossref
Brasier, M.D. and Sukhov, S. S. 1998. The falling amplitude of carbon isotopic oscillations through the Lower to Middle Cambrian: northern Siberia data. Canadian Journal of Earth Sciences 35: 353–373. Crossref
Brasier, M.D., Rozanov, A.Y., Zhuravlev, A.Y., Corfield, R.M., and Derry, L.A. 1994. A carbon isotope reference scale for the Lower Cambrian succession in Siberia: report of IGCP project 303. Geological Magazine 131: 767–783. Crossref
Brock, G.A. and Cooper, B.J. 1993. Shelly fossils from the Early Cambrian (Toyonian) Wirrealpa, Aroona Creek, and Ramsay limestones of South Australia. Journal of Paleontology 67: 758–787. Crossref
Bushuev, E., Goryaeva, I., and Pereladov, V. 2014. New discoveries of the oldest trilobites Profallotaspis and Nevadella in the northeastern Siberian Platform, Russia. Bulletin of Geosciences 89: 347–364. Crossref
Carrera, M.G. and Maletz, J. 2014. Ordovician sponge spicules from the Spitsbergen, Nevada and Newfoundland: new evidence for hexactinellid and demosponge early diversification. Journal of Systematic Palaeontology 12: 961–981. Crossref
Castellani, C., Maas, A., Haug, C., Haug, J.T., and Waloszek, D. 2012. Isolated sponge spicules from the late Cambrian Alum Shale Formation (“Orsten” nodules) of Sweden. Bulletin of Geosciences 87: 443–460. Crossref
Chang, C., Hu, W., Wang, X., Yu, H., Yang, A., Cao, J., and Yao, S. 2017. Carbon isotope stratigraphy of the lower to middle Cambrian on the eastern Yangtze Platform, South China. Palaeogeography, Palaeoclimatology, Palaeoecology 479: 90–101. Crossref
Chen, F. and Dong, X. 2008. Middle Cambrian highly modified sponge spicules from western Hunan, South China. Acta Scientiarum Naturalium Universitatis Pekinensis 44: 883–889.
Chen, J.-Y. and Zhou, G.-Q. 1997. Biology of the Chengjiang fauna. In: J.-Y. Chen, Y.-N. Cheng, and H. Van Iten (eds.), The Cambrian Explosion and the Fossil Record. Bulletin of the National Museum of Natural Science 10: 11–105.
Chen, Z., Hu, J., Zhou, C., Xiao, S., and Yuan, X. 2004. Sponge fossil assemblage from the Early Cambrian Hetang Formation in southern Anhui. Chinese Science Bulletin 49: 1625–1628. Crossref
Clark, D.L. and Miller, J.S. 1969. Early evolution of conodonts. Geological Society of America Bulletin 80: 125–134. Crossref
Clark, D.L. and Robison, R.A. 1969. Oldest conodonts in North America. Journal of Paleontology 43: 1044–1046.
Clausen, S. and Peel, J.S. 2012. Middle Cambrian echinoderm remains from the Henson Gletscher Formation of North Greenland. GFF 134: 173–200. Crossref
Clausen, S. and Smith, A.B. 2008. Stem structure and evolution in the earliest pelmatozoan echinoderms. Journal of Paleontology 82: 737–748. Crossref
Clausen, S., Álvaro, J.J., and Zamora, S. 2014. Replacement of benthic communities in two Neoproterozoic–Cambrian subtropical-to-temperate rift basins, High Atlas and Anti-Atlas, Morocco. Journal of African Earth Sciences 98: 72–93. Crossref
Claybourn, T.M., Jacquet, S.M., Skovsted, C.B., Topper, T.P., Holmer, L.E. and Brock, G.A. 2019. Mollusks from the upper Shackleton Limestone (Cambrian Series 2), Central Transantarctic Mountains, East Antarctica. Journal of Paleontology 93: 437–459. Crossref
Cobbold, E.S. 1921. The Cambrian horizons of Comley and their Brachiopoda, Pteropoda, Gastropoda, etc. Quarterly Journal of the Geographical Society of London 76: 325–386. Crossref
Conway Morris, S. 1986. The community structure of the Middle Cambrian phyllopod bed (Burgess Shale). Palaeontology 29: 423–467. Crossref
Conway Morris, S. and Chen, M. 1991. Cambroclaves and paracarinachitids, early skeletal problematica from the Lower Cambrian of South China. Palaeontology 34: 357–397.
Conway Morris, S. and Peel, J.S. 2010. New palaeoscolecidan worms from the lower Cambrian: Sirius Passet, Latham Shale and Kinzers Shale. Acta Palaeontologica Polonica 55: 141–156.
Conway Morris, S. and Robison, R.A. 1986. Middle Cambrian priapulids and other soft-bodied fossils from Utah and Spain. Paleontological Contributions from the University of Kansas Papers 117: 1–22.
Conway Morris, S., Crampton, J.S., Xiao, B., and Chapman, A.J. 1997. Lower Cambrian cambroclaves (incertae sedis) from Xinjiang, China, with comments on the morphological variability of sclerites. Palaeontology 40: 167–189.
Cooper, G.A. 1942. New genera of North American brachiopods. Journal of the Washington Academy of Sciences 32: 228–235.
Cusack, M., Williams, A., and Buckman, J.O. 1999. Chemico-structural evolution of the shell of linguloid brachiopods. Palaeontology 42: 799–840. Crossref
Cuvier, G. 1797. Tableau élementaire de l’historie naturelle des animaux. 710 pp. Baudouin, Paris. Crossref
d’Orbigny, A.D. 1826. Tableau méthodique de la classe des Céphalopodes. Annales des Sciences Naturelles, sér. 1 7: 245–314.
Dawson, J.W. 1868. Acadian Geology; the Geological Structure, Organic Remains and Mineral Resources of Nova Scotia, New Brunswick and Prince Edward Island. 694 pp. Macmillan, London. Crossref
Debrenne, F., Rozanov, A., and Zhuravlev, A. 1990. Regular Archaeocyaths. 218 pp. Éditions du Centre National de la Recherche Scientifique, Cahiers de Paléontologie, Paris.
Debrenne, F., Zhuravlev, A.Y., and Kruse, P.D. 2015. General features of Archaeocyatha. In: P.A. Selden (ed.), Treatise on Invertebrate Paleontology, Part E, Porifera (Revised), Hypercalcified Porifera, Volume 5, 845–992. University of Kansas, Lawrence. Crossref
Devaere, L. and Skovsted, C.B. 2017. New early Cambrian sclerites of Lapworthella schodackensis from NE Greenland: advancements in knowledge of lapworthellid taxonomy, sclerite growth and scleritome organization. Geological Magazine 154: 1061–1072. Crossref
Devaere, L., Clausen, S., Álvaro, J.J., Peel, J.S. and Vachard, D. 2014. Terreneuvian orthothecid (Hyolitha) digestive tracts from northern Montagne Noire, France: Taphonomic, ontogenetic and phylogenetic implications. PLoS ONE 9 (2): e88583. Crossref
Devaere, L., Clausen, S., Sosa-Leon, J.P., Palafox-Reyes, J.J., Buitrón-Sánchez, B.E., and Vachard, D. 2019. Early Cambrian Small Shelly Fossils from northwest Mexico: Biostratigraphic implications for Laurentia. Palaeontologia Electronica 22.2.41A: 1–60. Crossref
Devaere, L., Clausen, S., Steiner, M., Álvaro, J.J., and Vachard, D. 2013. Chronostratigraphic and palaeogeographic significance of an early Cambrian microfauna from the Heraultia Limestone, northern Montagne Noire, France. Palaeontologia Electronica 16 (2): 17A, 91. Crossref
Ding, L., Zhang, L., Li, Y., and Dong, J. 1992. The Study of the Late Sinian –Early Cambrian Biota from the Northern Margin of Yangtze Platform. 156 pp. Scientific and Technical Documents Publishing House, Beijing.
Dong, X-P. and Bergström, S.M. 2001. Middle and Upper Cambrian protoconodonts and paraconodonts from Hunan, South China. Palaeontology 44: 949–985. Crossref
Dong, X.-P. and Knoll, A.H. 1996. Middle and Late Cambrian sponge spicules from Hunan, China. Journal of Paleontology 70: 173–184. Crossref
Dong, X.-P., Bengtson, S., Gostling, N.J., Cunningham, J.A., Harvey, T.H.P., Kouchinsky, A., Val’kov, A.K., Repetski, J.E., Stampanoni, M., Marone, F., and Donoghue, P.C.J. 2010. The anatomy, taphonomy, taxonomy and systematic affinity of Markuelia: Early Cambrian to Early Ordovician scalidophorans. Palaeontology 53: 1291–1314. Crossref
Doré, F. and Reid, R.E. 1965. Allonnia tripodophora nov.gen., nov. sp., nouvelle Éponge du Cambrien inférieur de Carteret (Manche). Compte Rendu Sommaire des Séances de la Société Géologique de France 1965: 20–21.
Drozdov, A.L., Zhuravlev, A.Yu., Karpenko, A.A., Kolesnikov, K.A., and Ivantsov, A.Yu. 2022. The morphology and chemical composition of siliceous spicules of the Early Cambrian sponge Lenica unica Gorjansky, 1977. Russian Journal of Marine Biology 48 (1): 33–38. Crossref
Duan, C. 1984 [date of article biblid 1983]. Small shelly fossils from the Lower Cambrian Xihaoping Formation in the Shennongjia District, Hubei Province—hyoliths and fossil skeletons of unknown affinities [in Chinese with English summary]. Bulletin of the Tianjin Institute of Geology and Mineral Resources 7: 143–188.
Duméril, A.M.C. 1806. Zoologie analytique ou method naturelle de classification des animaux, rendue plus facile à l’aide de tableaux synoptiques. 344 pp. Allais, Paris Crossref
Dzik, J. 2005. Behavioral and anatomical unity of the earliest burrowing animals and the cause of the “Cambrian explosion”. Paleobiology 31: 503–521. Crossref
Dzik, J. and Krumbiegel, G. 1989. The oldest “onychophoran” Xenusion: a link connecting phyla? Lethaia 22: 169–181. Crossref
Eichwald, E. 1860. Lethaea rossica ou palaeontologie de la Russie. 1657 pp. Schweizerbart, Stuttgart.
Elicki, O. 1994. Lower Cambrian carbonates from eastern Germany: palaeontology, stratigraphy and palaeogeography. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 191: 69–93.
Elicki, O. 2011. First skeletal microfauna from the Cambrian Series 3 of the Jordan Rift Valley (Middle East). Memoirs of the Association of Australasian Palaeontologists 42: 153–173.
Elicki, O. and Gürsu, S. 2009. First record of Pojetaia runnegari Jell, 1980 and Fordilla Barrande, 1881 from the Middle East (Taurus Mountains, Turkey) and critical review of Cambrian bivalves. Paläontologische Zeitschrift 83: 267–291. Crossref
Ermak,V.V. and Pelman,Y.L. 1986. Some Cambrian molluscs and brachiopods of the northern Kharaulakh [in Russian]. In: I.T. Žuravleva (ed.), Biostratigrafiâ i paleontologiâ kembriâ Severnoj Azii, 188–200. Nauka, Moskva.
Esakova, N.V. and Zhegallo, E.A. [Žegallo, E.A.] 1996. Biostratigraphy and fauna of the Lower Cambrian of Mongolia [in Russian]. In: A.Û. Rozanov (ed.), Trudy Sovmestnoj Rossijsko-Mongol’skoj Paleontologičeskoj Ekspeditsii 46: 1–216.
Fedorov, A.B. and Pereladov, V.S. 1987. Spicules of siliceous sponges from the Kuonamka Formation of the North-East of the Siberian Platform [in Russian]. In: S.P. Bulynnikova and I.G. Klimova (eds.), Novye vidy drevnih rastenij i bespozvonočnyh fanerozoâ Sibiri, 36–46. SNIGGiMS, Novosibirsk.
Feng, W., Xinan M., and Kouchinsky, A. 2001. Hyolith-type microstructure in a mollusc-like fossil from the Early Cambrian of Yunnan, China. Lethaia 34: 303–308. Crossref
Fisher, D.W. 1962. Other small conoidal shells. In: R.C. Moore (ed.), Treatise on Invertebrate Paleontology. Part W. Miscellanea, W98–W143. Geological Society of America and University of Kansas Press, Lawrence.
Fonin, V.D. and Smirnova, T.N. 1967. New group of problematic Early Cambrian organisms and methods of preparing them [in Russian]. Paleontologičeskij žurnal 1967 (2): 7–18.
Gallet, Y., Pavlov, V., and Courtillot, V. 2003. Magnetic reversal frequency and apparent polar wander of the Siberian platform in the earliest Palaeozoic, inferred from the Khorbusuonka river section (northeastern Siberia). Geophysical Journal International 154: 829–840. Crossref
Gallet, Y., Pavlov, V., and Korovnikov, I. 2019. Extreme geomagnetic reversal frequency during the Middle Cambrian as revealed by the magnetostratigraphy of the Khorbusuonka section (northeastern Siberia). Earth and Planetary Science Letters 528: 115823. Crossref
García-Bellido, D.C., Gozalo, R., Chirivella Martorell, J.B., and Liñán, E. 2007. The demosponge Leptomitus and a new species from the Middle Cambrian of Spain. Palaeontology 50: 467–478. Crossref
Gedik, I. 1977. Conodont stratigraphy in the Middle Taurus [In Turkish with English abstract]. Bulletin of the Geological Society of Turkey 20: 35–48.
Geyer, 1994 Order Helcionellida Geyer, G. 1994. Middle Cambrian molluscs from Idaho and early conchiferan evolution. In: E. Landing (ed.), Studies in Stratigraphy and Paleontology in Honor of Donald W. Fisher. New York State Museum, Bulletin 481: 69–86.
Geyer, G. 2019. A comprehensive Cambrian correlation chart. Episodes 42: 321–332. Crossref
Golubev, S.N. 1976. Ontogenetic changes and evolutionary trends in Early Cambrian spiral gastropods of the superfamily Pelagiellacea [in Russian]. Paleontologičeskij žurnal 1976 (10): 143–149.
Gorjansky, V.Y. [Gorânskij, V.Û.] 1977. First find of sponge remains in the Lower Cambrian of eastern Siberia [in Russian]. Ežegodnik Vsesoûznogo Paleontologičeskogo Obŝestva 20: 274–276.
Gorjansky, V.Y. [Gorânskij, V.Û.] and Popov, L.E. 1985. The morphology, systematic position, and origin of inarticulate brachiopods with carbonate shells [in Russian]. Paleontologičeskij žurnal 1985 (3): 3–13.
Grabau, A.W. 1900. Palaeontology of the Cambrian terranes of the Boston Basin. Occasional Papers of the Boston Society of Natural History 4: 601–694.
Grant, R.E. 1836. Animal kingdom. In: R.B. Todd (ed.), The Cyclopedia of Anatomy and Physiology 1, 107–118. Gilbert and Piper, London.
Gravestock, D.I., Alexander, E.M., Demidenko, Y.E., Esakova, N.V., Holmer, L.E., Jago, J.B., Lin, T., Melnikova, L.M., Parkhaev, P.Y., Rozanov, A.Y., Ushatinskaya, G.T., Zang, W., Zhegallo, E.A., and Zhuravlev, A.Y. 2001. The Cambrian biostratigraphy of the Stansbury Basin, South Australia. Transactions of the Palaeontological Institute 282: 1–344.
Grazhdankin, D.V., Marusin, V.V., Izokh, O.P., Karlova, G.A., Kochnev, B.B., Markov, G.E., Nagovitsin, K.E., Sarsembaev, Z., Peek, S., Cui, H., and Kaufman, A.J. 2020. Quo vadis, Tommotian? Geological Magazine 157: 22–34. Crossref
Gridina, N.M. 1991. Lower Cambrian conodonts and phosphatic problematica from central Kazakhstan [in Russian]. Paleontologičeskij žurnal 1991 (3): 102–108.
Grube, A.E. 1850. Die Familien der Anneliden. Archiv für Naturgeschichte, Berlin 16 (1): 249–364.
Gubanov, A.P., Fernández Remolar, D.C., and Peel, J.S. 2004a. Early Cambrian molluscs from Sierra de Córdoba (Spain). Geobios 37: 199–215. Crossref
Gubanov, A., Kouchinsky, A., Peel, J.S., and Bengtson, S. 2004b. Middle Cambrian molluscs of “Australian” aspect from northern Siberia. Alcheringa 28: 1–20. Crossref
Guo, J.-F., Li, G.-X., Qiang, Y.-Q., Song, Z.-C., Zhang, Z.-F., Han, J., and Wang, W.-Z. 2021. Watsonella crosbyi from the lower Cambrian (Terreneuvian, Stage 2) Yanjiahe Formation in Three Gorges Area, South China. Palaeoworld 30: 1–19. Crossref
Hall, J. 1847. Palaeontology of New York, Volume 1, Containing Descriptions of the Organic Remains of the Lower Division of the New-York System. 338 pp. Geological Survey of the State of New York, Albany.
Hao, Y.-C. and Shu, D.-G. 1987. The oldest well-preserved Phaeodaria (Radiolaria) from southern Shaanxi [in Chinese with English abstract]. Geoscience 1: 301–310.
Harvey, T.H.P. 2010. Carbonaceous preservation of Cambrian hexactinellid sponge spicules. Biology Letters 6: 834–837. Crossref
He, T., Pei, F., and Fu, G. 1984. Some small shelly fossils from the Lower Cambrian Xinji Formation in Fangcheng County, Henan Province [in Chinese with English abstract]. Acta Palaeontologica Sinica 23: 350–357.
He, T., Zhu, M., Mills, B.J.W., Wynn, P.M., Zhuravlev, A.Y., Tostevin, R., Pogge von Strandmann, P.A.E., Yang, A., Poulton, S.W., and Shields, G.A. 2019. Possible links between extreme oxygen perturbations and the Cambrian radiation of animals. Nature Geoscience 468: 468–474. Crossref
Heredia, S., Bordonado, O., and Matteoda, E. 1987. Espiculas de poriferos de la formacion la Cruz, Cambrio Superior, Departamento Las Heras, provincia de Mendosa. Ameghiniana 24: 17–20.
Hicks, H. 1872. On some undescribed fossils from the Menevian Group of Wales. Quarterly Journal of the Geological Society of London 27: 41–42. Crossref
Hinde, G.J. 1887. A monograph of the British fossil sponges, Part 1, Vol. 1, Sponges of Palaeozoic and Jurassic strata. Monographs of the Palaeontographical Society, London 40: 1–92. Crossref
Hinz, I. 1987. The Lower Cambrian microfauna of Comley and Rushton, Shropshire/England. Palaeontographica A 198: 41–100.
Hinz-Schallreuter, I., Gozalo, R., and Liñán, E. 2008. New bradorid arthropods from the Lower Cambrian of Spain. Micropalaeontology 53: 497–510. Crossref
Holm, G. 1893. Sveriges kambrisk-siluriska Hyolithidae och Conulariidae. Sveriges Geologiska Undersökning, Afhandlingar och Uppsatser C 112: 1–172.
Holmer, L.E., Popov, L.E., and Wrona, R. 1996. Early Cambrian brachiopods from King George Island (South Shetland Islands), West Antarctica. Palaeontologica Polonica 55: 37–50.
Holmer, L.E., Pettersson Stolk, S., Skovsted, C.B., Balthasar, U., and Popov, L.E. 2009. The enigmatic Early Cambrian Salanygolina—a stem group of rhynchonelliform chileate brachiopods? Palaeontology 52: 1–10. Crossref
Horný, R. 1964. The Middle Cambrian Pelagiellacea of Bohemia (Mollusca). Sbornik Národniho Muzea v Praze 20B: 133–140.
Hou, X-G., Williams, M., Siveter, D.J., Siveter, D.J., Gabbott, S., Holwell, D., and Harvey, T.H.P. 2014. A chancelloriid-like metazoan from the early Cambrian Chengjiang Lagerstätte, China. Scientific Reports 4: 7340. Crossref
Hou X.-G. and Bergström, J. 1995. Cambrian lobopodians—ancestors of extant onychophorans? Zoological Journal of the Linnean Society 114: 3–19. Crossref
Ippolitov, A.P., Vinn, O., Kupriyanova, E.K., and Jäger, M. 2014. Written in stone: history of serpulid polychaetes through time. Memoirs of Museum Victoria 71: 123–159. Crossref
Ivantsov, A.Y. [Ivancov, A.Û.], Zhuravlev, A.Y. [Žuravlev, A.Û.], Krasilov, V.A., Leguta, A.V., Melnikova, L.M. [Mel’nikova, L.M.], Urbanek, A., Ushatinskaya, G.T. [Ušatinskaâ, G.T.], and Malakhovskaya, Y.E. [Malahovskaâ, Â.E.] 2005a. Unique Sinsk localities of Early Cambrian organisms (Siberian Platform) [in Russian]. Trudy Paleontologičeskogo Instituta Rossijskoj Academii Nauk 284: 1–143.
Ivantsov, A.Y., Zhuravlev, A.Y., Leguta, A.V., Krassilov, V.A., Melnikova, L.M., and Ushatinskaya G.T. 2005b. Palaeoecology of the Early Cambrian Sinsk biota from the Siberian Platform. Palaeogeography, Palaeoclimatology, Palaeoecology 220: 69–88. Crossref
Jell, P.A. 1980. Earliest known pelecypod on Earth—a new early Cambrian genus from South Australia. Alcheringa 4: 233–239. Crossref
Jin, J., Zhan, R., Copper, P., and Caldwell, W.G.E. 2007. Epipunctae and phosphatized setae in Late Ordovician plaesiomyid brachiopods from Anticosti Island, eastern Canada. Journal of Paleontology 81: 666–683. Crossref
Jones, W.C. and Jenkins, D.A. 1970. Calcareous sponge spicules: a study of magnesian calcites. Calcified Tissue Research 4: 314–329. Crossref
Kuhn, O. 1949. Lehrbuch der Paläozoologie. 326 pp. Schweizerbart’sche Verlagsbuchhandlung, Stuttgart.
Karlova, G.A. and Vodanyuk, S.A. [Vodanûk, S.A.] 1985. New data on the deposits transitional to the Cambrian from the Khorbosuonka River basin (Olenyok Uplift) [in Russian]. In: V.V. Homentovskij (ed.), Stratigrafiâ pozdnego dokembriâ i rannego paleozoâ Sibiri. Vend i rifej, 3–13. Institut Geologii i Geofiziki SO AN SSSR, Novosibirsk.
Kaufman, A.J. and Knoll, A.H. 1995. Neoproterozoic variations in the C-isotopic composition of seawater: Stratigraphic and biogeochemical implications. Precambrian Research 73: 27–49. Crossref
Kaufman, A.J., Knoll, A.H., Semikhatov, M.A., Grotzinger, J.P., Jacobsen, S.B., and Adams, W. 1996. Integrated chronostratigraphy of Proterozoic–Cambrian boundary beds in the western Anabar region, northern Siberia. Geological Magazine 133: 509–533. Crossref
Kaufman, A.J., Peek, S., Martin, A.J., Cui, H., Grazhdankin, D., Rogov, V., Xiao, S., Buchwaldt, R., and Bowring, S. 2012. A shorter fuse for the Cambrian Explosion? Geological Society of America Abstracts with Programs 44 (7): 326.
Klein, J.T. 1734. Naturalis dispositio Echinodermatum. Accessit lucubratiuncula de aculeis Echinorum marinorum, cum spicilegio de Belemnitis. 79 pp. Gedani, Schreiber. Crossref
Knight, J.B. 1956. New families of gastropods. Journal of the Washington Academy of Sciences 46: 241–242.
Knoll, A.H., Grotzinger, J.P., Kaufman, A.J., and Kolosov, P. 1995. Integrated approaches to terminal Proterozoic stratigraphy: An example from the Olenek Uplift, northeastern Siberia. Precambrian Research 73: 251–270. Crossref
Kontorovich, A.E., Varlamov, A.I., Grazhdankin, D.V., Karlova, G.A., Klets, A.G., Kontorovich, V.A., Saraev, S.V., Terleev, A.A., Belyaev, S.Y., Varaksina, I.V., Efimov, A.S., Kochnev, B.B., Nagovitsin, K.E., Postnikov, A.A., and Filippov, Y.F. 2008. A section of Vendian in the east of the West Siberian Plate (based on data from the Vostok 3 Borehole). Russian Geology and Geophysics 49: 932–939. Crossref
Korovnikov, I.V. 1998. Trilobite assemblages and biostratigraphy of the Lower Cambrian section in the Khorbosuonka River valley (Olenek Uplift, northeastern Siberian Platform). Russian Geology and Geophysics 39: 1378–1392.
Korovnikov, I.V. 2002. New data on biostratigraphy of the Lower and Middle Cambrian series in the Northeastern Siberian Platform. Russian Geology and Geophysics 43: 826–836.
Korovnikov, I.V. and Tokarev, D.A. 2018. New data on biostratigraphy of the Middle Cambrian section at the Khorbusuonka River, northeastern Siberian Platform. Stratigraphy and Geological Correlation 26: 599–610. Crossref
Korshunov, V.I. [Koršunov, V.I.] 1968. Gonamispongia, a new genus of sponges of family Chancelloriidae [in Russian]. Paleontologičeskij žurnal 1968 (3): 127–129.
Korshunov, V.I. [Koršunov, V.I.] and Zhuravleva, I.T. [Žuravleva, I.T.] 1967. New archaeocyath species from the Lower Cambrian of Yakutia [in Russian]. In: A.B. Ivanovskij and B.S. Sokolov (eds.), Novye dannye po biostratigrafii nižnego paleozoâ Sibirskoj platformy, 3–11. Nauka, Novosibirsk.
Kouchinsky, A. 1999. Shell microstructures of the Early Cambrian Anabarella and Watsonella as new evidence on the origin of the Rostroconchia. Lethaia 32: 173–180. Crossref
Kouchinsky, A. 2000. Shell microstructures in Early Cambrian molluscs. Acta Palaeontologica Polonica 45: 119–150.
Kouchinsky, A. and Bengtson, S. 2017. X-ray tomographic microscopy tightens affinity of the early Cambrian Oymurania to the brachiopod stem group. Acta Palaeontologica Polonica 62: 39–43. Crossref
Kouchinsky, A., Bengtson, S., and Murdock, D. 2010. A new tannuolinid problematic from the lower Cambrian of the Sukharikha River in northern Siberia. Acta Palaeontologica Polonica 55: 321–331. Crossref
Kouchinsky, A., Bengtson, S., Clausen, S., and Vendrasco, M.J. 2015a. An early Cambrian fauna of skeletal fossils from the Emyaksin Formation, northern Siberia. Acta Palaeontologica Polonica 60: 421–512.
Kouchinsky, A., Bengtson, S., Clausen, S., Gubanov, A. P., Malinky, J.M., and Peel, J.S. 2011. A Middle Cambrian fauna of skeletal fossils from the Kuonamka Formation, northern Siberia. Alcheringa 35: 123–189. Crossref
Kouchinsky, A., Bengtson, S., Feng, W., Kutygin, R., and Val’kov, A. 2009. The Lower Cambrian fossil anabaritids: affinities, occurrences and systematics. Journal of Systematic Palaeontology 7: 241–298. Crossref
Kouchinsky, A., Bengtson, S., Gallet, Y., Korovnikov, I., Pavlov, V., Runnegar, B., Shields, G., Veizer, J., Young, E., and Ziegler, K. 2008. The SPICE carbon isotope excursion in Siberia: a combined study of the upper Middle Cambrian–lowermost Ordovician Kulyumbe River section, northwestern Siberian Platform. Geological Magazine 145: 609–622. Crossref
Kouchinsky, A., Bengtson, S., Landing, E., Steiner, M., Vendrasco, M., and Ziegler, K. 2017. Terreneuvian stratigraphy and faunas from the Anabar Uplift, Siberia. Acta Palaeontologica Polonica 62: 311‒440. Crossref
Kouchinsky, A., Bengtson, S., Missarzhevsky, V., Pelechaty, S., Torssander, P., and Val’kov, A. 2001. Carbon isotope stratigraphy and the problem of a pre-Tommotian Stage in Siberia. Geological Magazine 138: 387–396. Crossref
Kouchinsky, A., Bengtson, S., Pavlov, V., Runnegar, B., Torssander, P., Young, E., and Ziegler, K. 2007. Carbon isotope stratigraphy of the Precambrian–Cambrian Sukharikha River section, northwestern Siberian platform. Geological Magazine 144: 1–10. Crossref
Kouchinsky, A., Bengtson, S., Pavlov, V., Runnegar, B., Val’kov, A., and Young, E. 2005. Pre-Tommotian age of the lower Pestrotsvet Formation in the Selinde section on the Siberian platform: Carbon isotopic evidence. Geological Magazine 142: 319–325. Crossref
Kouchinsky, A., Bengtson, S., Runnegar, B., Skovsted, C., Steiner, M., and Vendrasco, M.J. 2012. Chronology of early Cambrian biomineralisation. Geological Magazine 149: 221–251. Crossref
Kouchinsky, A., Holmer, L.E., Steiner, M., and Ushatinskaya, G.T. 2015b. The new stem-group brachiopod Oymurania from the lower Cambrian of Siberia. Acta Palaeontologica Polonica 60: 963–980. Crossref
Kozur, H.W., Mostler, H., and Repetski, J.E. 2008. A new heteractinellid sponge from the lowermost Ordovician of Nevada and a discussion of the suborder Heteractinellidae. Geo.Alp 5: 53–67.
Krasilova, I.N. 1977. Fordillids (Bivalvia) from the early Palaeozoic of the Siberian Platform [in Russian]. Paleontologičeskij žurnal 1987 (4): 24–30.
Krasilova, I.N. 1987. The oldest representatives of the bivalve mollusks [in Russian]. Paleontologičeskij žurnal 1977 (2): 42–48.
Krasnopeeva, P.S. 1953. Features of the Kameshki archaeocyathan assemblage in the facies of effusive-sedimentary deposits exemplified by the archaeocyaths of the western part of Tuva [in Russian]. Trudy Tomskogo Gosudarstvennogo Universiteta, Seriâ Geologičeskaâ 124: 51–62.
Kruse, P.D. and Rietner, J.R. 2014. Northern Australian microbial-metazoan reefs after the mid-Cambrian mass extinction. Memoirs of the Association of Australasian Palaeontologists 45: 31–53.
Kruse, P.D., Zhuravlev, A.Y., and Debrenne, F. 2015. Radiocyaths and potentially allied taxa: Systematic descriptions. In: P.A. Selden (ed.), Treatise on Invertebrate Paleontology, Part E, Porifera (Revised), Hypercalcified Porifera, Volume 5, 1083–1093. University of Kansas; Lawrence, Kansas.
Kruse, P.D., Zhuravlev, A.Y., and James, N.P. 1995. Primordial metazoan-calcimicrobial reefs: Tommotian (Early Cambrian) of the Siberian Platform. Palaios 10: 291–321. Crossref
Landing, E. 1974. Early and Middle Cambrian Conodonts from the Taconic Allochthon, Eastern New York. Journal of Paleontology 48: 1241–1248.
Landing, E. 1984. Skeleton of lapworthellids and the suprageneric classification of tommotiids (Early and Middle Cambrian phosphatic problematica). Journal of Paleontology 58: 1380–1398.
Landing, E. 1988. Lower Cambrian of eastern Massachusetts: stratigraphy and small shelly fossils. Journal of Paleontology 62: 661–695.
Landing, E. 1995. Upper Placentian–Branchian Series of mainland Nova Scotia (middle–upper Lower Cambrian): faunas, paleoenvironments, and stratigraphic revision. Journal of Paleontology 69: 475–495. Crossref
Landing, E. 1996. Reconstructing the Avalon continent: marginal-to-inner platform transition in the Cambrian of Avalonian New Brunswick. Canadian Journal of Earth Sciences 33: 623–632. Crossref
Landing, E. and Bartowski, K.E. 1996. Oldest shelly fossils from the Taconic Allochthon and late Early Cambrian sea-levels in eastern Laurentia. Journal of Paleontology 70: 741–761. Crossref
Landing, E. and Kouchinsky, A. 2016. Correlation of the Cambrian Evolutionary Radiation: geochronology, evolutionary stasis of earliest Cambrian (Terreneuvian) small shelly fossil (SSF) taxa, and chronostratigraphic significance. Geological Magazine 153: 750–756. Crossref
Landing, E., Nowlan, G.S., and Fletcher, T.P. 1980. A microfauna associated with Early Cambrian trilobites of the Callavia Zone, northern Antigonish Highlands, Nova Scotia. Canadian Journal of Earth Sciences 17: 400–418. Crossref
Landing, E., Geyer, G., Brasier, M.D., and Bowring, S.A. 2013. Cambrian Evolutionary Radiation: context, correlation, and chronostratigraphy—overcoming deficiencies of the first appearance datum (FAD) concept. Earth-Science Reviews 123: 133–172. Crossref
Landing, E., Geyer, G., Jirkov, I.A., and Schiaparelli, S. 2021a. Lophotrochozoa in the Cambrian evolutionary radiation and the Pelagiella problem. Papers in Palaeontology 7: 2227–2244. Crossref
Landing, E., Geyer, G., Schmitz, M.D., Wotte, T., and Kouchinsky, A. 2021b. (Re)proposal of three Cambrian Subsystems and their Geochronology. Episodes 44: 273–283. Crossref
Landing, E., Schmitz, M.D., Geyer, G., Trayler, R.B., and Bowring, S.A. 2021c. Precise early Cambrian U-Pb zircon dates bracket the oldest trilobites and archaeocyaths in Moroccan West Gondwana. Geological Magazine 158: 219–238. Crossref
Laurie, J.R. 1986. Phosphatic fauna of the Early Cambrian Todd River Dolomite, Amadeus Basin, central Australia. Alcheringa 10: 431–454. Crossref
Lazarenko, N.P. and Repina, L.N. 1983. Section of the Lower and Middle Cambrian strata on the eastern slope of the Bulkur Antecline (Tuora-Sis Ridge, northern Verkhoyan’e) [in Russian]. In: L.N. Repina and N.P. Suvorova (eds.), Biostratigrafiâ i fauna pograničnyh otloženij nižnego i srednego kembriâ Sibiri. Trudy Instituta Geologii i Geofiziki Sibirskogo Otdeleniâ Akademii Nauk SSSR 548: 22–35.
Lee, H.-Y. 1975. Conodonts from the Upper Cambrian Formation, Kangweon-Do, South Korea and its stratigraphical significance. The Graduate School, Yonsei University, Seoul, Korea 12: 71–89.
Lee, Y.-W. 1975. On the Cambrian Ostracoda with new materialfrom Sichuan, Yunnan and souhern Shaanxi, China) [in Chinese]. Professional Papers on Stratigraphy and Palaeontology 2: 37–72.
Lemburg, C. 1995. Ultrastructure of the introvert and associated structures of the larvae of Halicryptus spinulosus (Priapulida). Zoomorphology 115: 11–29. Crossref
Leuckart, R. 1854. Salpen und Verwandte. Zoologische Untersuchungen Giessen 2: 47–63. Crossref
Levinsen, G.M.R. 1883. Systematisk-geografisk Oversigt over de nordiske Annulata, Gephyrea, Chaetognathi og Balanoglossi. Videnskabelige Meddelelser fra Dansk naturhistorisk Forening i Köbenhavn 1882: 160–251. Crossref
Li, G., Zhang, Z., Hua, H., and Yang, H. 2014. Occurrence of the enigmatic bivalve fossil Apistoconcha in the lower Cambrian of southeast Shaanxi, North China Platform. Journal of Paleontology 88: 359–366. Crossref
Li, L., Zhang, X., Skovsted, C.B., Yun, H., Pan, B., and Li, G. 2021. Revisiting the molluscan fauna from the Cambrian (Series 2, Stages 3–4) Xinji Formation of North China. Papers in Palaeontology Vol. 7, Part 1, 521–564. Crossref
Li, L., Zhang, X., Yun, H., and Li, G. 2017. Complex hierarchical microstructures of Cambrian mollusk Pelagiella: insight into early biomineralization and evolution. Scientific Reports 7: 1935. Crossref
Linnaeus, C. 1758. Systema naturae per regna tria naturae: secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Editio decima, reformata, Vol. 1. 824 pp. Laurentius Salvius, Holmiae. Crossref
Linnarsson, J.G.O. 1871. Om några försteningar från Sveriges och Norges “Primordialzon”. Öfversikt af Kongliga Vetenskaps-Akademiens Förhandlingar 6: 789–796.
Linnarsson, J.G.O. 1876. On the Brachiopoda of the Paradoxides Beds of Sweden. Bihang till Kongliga Svenska Vetenskaps-Akademiens Handlingar 3 (12): 1–34. Crossref
Linsley, R.M. and Kier, W.M. 1984. The Paragastropoda: a proposal for a new class of Paleozoic Mollusca. Malacologia 25: 241–254.
Liu, D. 1979. Earliest Cambrian brachiopods from southwest China [in Chinese]. Acta Palaeontologica Sinica 18: 505–511.
Liu, F., Skovsted, C.B., Topper, T.P., and Zhang, Z.-F. 2020. Are hyoliths Palaeozoic lophophorates? National Science Review 7: 453–469. Crossref
Liu, F., Skovsted, C.B., Topper, T.P., and Zhang, Z.-F. 2021. Revision of Triplicatella (Orthothecida, Hyolitha) with preserved digestive tracts from the early Cambrian Chengjiang Lagerstätte, South China. Historical Biology 33: 1857–1871. Crossref
Lochman, C. 1956. Stratigraphy, paleontology and paleogeography of the Elliptocephala asaphoides strata in Cambridge and Hoosick quadrangles, New York. Bulletin of the Geological Society of America 67: 1331–1396. Crossref
Luo, H., Jiang, Z., Wu, X., Song, X., Ouyang, L., Xing, Y., Liu, G., Zhang, S., and Tao, Y. 1984. Sinian–Cambrian Boundary Stralotype Seclion at Meishucun, Jinning, Yunnan, China (in Chinese and English). 154 pp. People’s Publishing House, Yunnan.
Mackinnon, D.I. 1985. New Zealand late Middle Cambrian molluscs and the origin of Rostroconchia and Bivalvia. Alcheringa 9: 65–81. Crossref
Magaritz, M., Latham, A.J., Kirschvink, J.L., Zhuravlev, A.Y., and Rozanov, A.Y. 1991. Precambrian–Cambrian boundary problem I: Carbon isotope correlations for Vendian and Tommotian time between Siberia and Morocco. Geology 19: 847–850. Crossref
Maletz, J. 2011. Radiolarian skeletal structures and biostratigraphy in the early Palaeozoic (Cambrian–Ordovician). Palaeoworld 20: 116–133. Crossref
Malinky, J.M. and Skovsted, C.B. 2004. Hyoliths and small shelly fossils from the Lower Cambrian of North-East Greenland. Acta Palaeontologia Polonica 49: 551–578.
Mambetov A.M. 1972. A new Lower Cambrian hyolithid genus from Mally Karatau (Northwestern Tien Shan) [in Russian]. Paleontologičeskij žurnal 1972 (2): 268–269.
Mambetov, A.M. and Repina, L.N. 1979. The Lower Cambrian of Talasskij Ala-Too and its correlation with the sections of Malyj Karatau and the Siberi an Platform [in Russian]. In: I.T. Žuravleva and N.P. Meškova (eds.), Biostratigfiâ i paleontologiâ nižnego kembriâ Sibiri. Trudy Instituta Geologii i Geofiziki Sibirskogo Otdeleniâ Akademii Nauk SSSR 406: 98–138.
Marek, L. 1963. New knowledge on the morphology of Hyolithes. Sborník geologickych ved, řada Paleontologie 1: 53–72.
Marek, L. 1966. New hyolithid genera from the Ordovician of Bohemia. Časopis národního muzea 135: 89–92.
Matthew, G.F. 1889. On the classification of the Cambrian rocks in Acadia. Canadian Record of Science 3: 303–315.
Matthew, G.F. 1891. Illustrations of the fauna of the St. John Group, No. 5. Royal Society of Canada, Proceedings and Transactions (Series 1, Section 4) 8: 123–166.
Matthew, G.F. 1894. Illustrations of the fauna of the St. John Group. Transactions of the Royal Society of Canada 11: 85–129.
Matthew, G.F. 1895. The Protolenus fauna. Transactions of the New York Academy of Sciences 14: 101–153. Crossref
Matthew, G.F. 1902 Notes on Cambrian faunas. Royal Society of Canada, Transactions, Series 3, Section 4 8: 93–112. Crossref
McIlroy, D., Green, O.R., and Brasier, M.D. 2001. Palaeobiology and evolution of the earliest agglutinated Foraminifera: Platysolenites, Spirosolenites and related forms. Lethaia 34: 13–29. Crossref
Mehl, D. 1996. Phylogenie und Evolutionsökologie der Hexactinellida (Porifera) im Paläozoikum. Geologische Paläontologische Mitteilungen der Universität Innsbruck, Sonderband 4: 1–55.
Mehl, D. 1998. Porifera and Chancelloriidae from the Middle Cambrian of the Georgina basin, Australia. Palaeontology 41: 1153–1182.
Melnikova, L., Siveter, D.J., and Williams, M. 1997. Cambrian Bradoriida and Phosphatocopida of the former Soviet Union. Journal of Micropalaeontology 16: 179–191. Crossref
Meshkova, N.P. [Meškova, N.P.] 1974. Cambroscleritida incertae sedis – a new order of Cambrian fossils. In: I.T. Žuravleva and A.Û. Rozanov (eds.), Biostratigfiâ i paleontologiâ nižnegokembriâ Evropy i Severnoj Azii, 190–193. Nauka, Moskva.
Meshkova, N.P. [Meškova, N.P.] 1969. On the paleontological characteristics of the Lower Cambrian strata of the Siberian Platform [in Russian]. In: I.T. Žuravleva (ed.), Biostratigfiâ i paleontologiâ nižnego kembriâ Sibiri i Dal’nego Vostoka, 158–172. Nauka, Moskva.
Meshkova, N.P. [Meškova, N.P.], Zhuravleva, I.T. [Žuravleva, I.T.], and Luchinina, V.A. [Lučinina, V.A.]. 1973. Lower Cambrian and the lower part of the Middle Cambrian of the Olenek Uplift [in Russian]. In: I.T. Žuravleva (ed.), Problemy paleontologii i biostratigrafii nižnego kembriâ Sibiri i Dal’nego Vostoka, 194–214. Nauka, Novosibirsk.
Missarzhevsky, V.V. [Missarževskij, V.V.] 1970. New generic name Tommotia, nom. nov. [in Russian]. Paleontologičeskij žurnal 1970 (2): 100.
Missarzhevsky, V.V. [Missarževskij, V.V.] 1973. Conodontomorph organisms from the Precambrian‒Cambrian boundary beds of the Siberian Platform and Kazakhstan [in Russian]. In: I.T. Žuravleva (ed.), Problemy paleontologii i biostratigrafii nižnego kembriâ Sibiri i Dal’nego Vostoka, 53–58. Nauka, Novosibirsk.
Missarzhevsky, V.V. [Missarževskij, V.V.] 1974. New data on the oldest fossils of the early Cambrian of the Siberian Platform [in Russian]. In: I.T. Žuravleva and A.Û. Rozanov (eds.), Biostratigfiâ i paleontologiâ nižnego kembriâ Evropy i severnoj Azii, 179–189. Nauka, Moskva.
Missarzhevsky, V.V. [Missarževskij, V.V.] 1977. Conodonts (?) and phosphatic problematica from the Cambrian of Mongolia and Siberia [in Russian]. In: L.P. Tatarinov (ed.), Bespozvonočnye paleozoâ Mongolii, 10–19. Nauka, Moskva.
Missarzhevsky, V.V. [Missarževskij, V.V.] 1980. Precambrian–Cambrian boundary strata in the western slope of the Olenek Uplift (Olenek River) [in Russian]. Bûlleten’ Moskovskogo Obŝestva Ispytatelej Prirody (Otdel Geologičeskii) 55: 23–34.
Missarzhevsky, V.V. [Missarževskij, V.V.] 1982. Subdivision and correlation of the Precambrian–Cambrian boundary beds based on some ancient groups of skeIetal organisms [in Russian]. Bûlleten’ Moskovskogo Obŝestva Ispytatelej Prirody (Otdel Geologičeskii) 57: 52–67. [English translation published in International Geological Review 25: 745–759]. Crossref
Missarzhevsky, V.V. [Missarževskij, V.V.] 1981. Early Cambrian hyoliths and gastropods from Mongolia [in Russian]. Paleontologičeskij žurnal 1981 (1): 21–28.
Missarzhevsky, V.V. [Missarževskij, V.V.] 1989. Oldest skeletal fossils and stratigraphy of Precambrian and Cambrian boundary beds [in Russian]. Trudy Geologičeskogo Instituta Akademii Nauk SSSR 443: 1–237.
Missarzhevsky, V.V. [Missarževskij, V.V.] and Mambetov, A.M. 1981. Stratigraphy and fauna of the Cambrian and Precambrian boundary beds of the Lesser Karatau Range [in Russian]. Trudy Geologičeskogo Instituta Akademii Nauk SSSR 326: 1–92.
Mostler, H. 1985. Neue heteractinide Spongien (Calcispongea) aus dem Unter- und Mittelkambrium Südwestsardiniens. Berichte des Naturwissenschaftlich-Medizinischen Vereins in Innsbruck 72: 7–32.
Mostler, H. 1996. Polyactinellid Schwämme, eine auf das Paläozoikum beschränkte Calcispongien-Gruppe. Geologische Paläontologische Mitteilungen der Universität Innsbruck 21: 223–243.
Mostler, H. and Mosleh-Yazdi, A. 1976. Neue Poriferen aus oberkambrischen Gesteinen der Milaformation im Elburzgebirge (Iran). Geologische Paläontologische Mitteilungen der Universität Innsbruck 5: 1–36.
Müller, K.J. 1959. Kambrische Conodonten. Zeitschrift der Deutschen Geologische Gesellschaft 111: 434–485. Crossref
Müller, K.J. and Hinz, I. 1991. Upper Cambrian conodonts from Sweden. Fossils & Strata 28: 1–153.
Müller, K.J., Walossek, D., and Zakharov, A. 1995. “Orsten” type phosphatised soft-integument preservation and a new record from the Middle Cambrian Kuonamka Formation in Siberia. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 197: 101–118. Crossref
Na, L. and Li, G.-X. 2011. Nail-shaped sclerite fossils from the Lower Cambrian Xihaoping Member of Fangxian, Hubei Province [in Chinese, with English abstract]. Acta Micropalaeontologica Sinica 28: 284–300.
Nadhira, A., Sutton, M.D., Botting, J.P., Muir, L.A., Gueriau, P., King, A., Briggs, D.E.G., Siveter, D.J., and Siveter, D.J. 2019. Three-dimensionally preserved soft tissues and calcareous hexactins in a Silurian sponge: implications for early sponge evolution. Royal Society Open Science 6: 190911. Crossref
Nagovitsin, K.E., Rogov, V.I., Marusin, V.V., Karlova, G.A., Kolesnikov, A.V., Bykova, N.V., and Grazhdankin, D.V. 2015. Revised Neoproterozoic and Terreneuvian stratigraphy of the Lena-Anabar Basin and north-western slope of the Olenek Uplift, Siberian Platform. Precambrian Research 270: 226–245. Crossref
Neckaja, A.I. [Neckaâ, A.I.] and Ivanova, V.A. 1956. The first ostracode find in the Lower Cambrian of East Siberia [in Russian]. Doklady Akademii Nauk SSSR 111: 1095–1097.
Novozhilova, N.V. and Korovnikov, I.V. 2019. Small shelly fossils in the Cambrian basement of the West Siberian Geosyncline. Stratigraphy. Geological Correlation 27: 3–11. Crossref
Okulitch, V.J. 1935. Cyathospongia—a new class of Porifera to include the Archaeocyathinae. Transactions of the Royal Society of Canada, Series 3, Section 4 29: 75–106.
Okulitch, V.J. 1937. Changes in nomenclature of Archaeocyathi (Cyathospongia). Proceedings of the Geological Society of America 1936: 358.
Özdikmen, H. 2009. Substitute names for eight sponge genus group names (Porifera). Munis Entomology & Zoology 4: 212–218.
Özgül, N. and Gedik, I. 1973. New information about stratigraphy and conodont fauna of the Lower Paleozoic Çaltepe and Seydişehir Formations, Central Taurus [in Turkish]. Bulletin Geological Society of Turkey 14: 25–101.
Pan, B., Skovsted, C.B., Sun, H.J., and Li, G.X. 2019. Biostratigraphical and palaeogeographical implications of Early Cambrian hyoliths from the North China Platform. Alcheringa 43: 351–380. Crossref
Pander, C.H. 1851. Sur une découverte de fossiles faite dans la partie inférieure du terrain silurien de Russie. Bulletin de la Société Géologique de France 8: 251–259.
Parkhaev, P.Y. 2002. Phylogenesis and the system of the Cambrian univalved mollusks. Paleontological Journal 36: 25–36.
Parkhaev, P.Y. 2004. Horsegullia nom. nov.—a replacement name for Yorkiella Parkhaev, 2001, a genus of Cambrian gastropods. Paleontological Journal 38: 576.
Parkhaev, P.Y. 2005. Two new species of the Cambrian helcionelloid mollusks from the northern part of the Siberian Platform. Paleontological Journal 39: 615–619.
Parkhaev, P.Y. 2017. Davidonia nom. nov., a new substitute name for a Cambrian gastropod genus. Paleontological Journal 51: 574. Crossref
Parkhaev, P.Y. and Demidenko, Y.E. 2010. Zooproblematica and Mollusca from the Lower Cambrian Meishucun section (Yunnan, China) and taxonomy and systematics of the Cambrian small shelly fossils of China. Paleontological Journal 44: 883–1161. Crossref
Parkhaev, P.Y. and Karlova, G.A. 2011. Taxonomic revision and evolution of Cambrian mollusks of the genus Aldanella Vostokova, 1962 (Gastropoda: Archaeobranchia). Paleontological Journal 45: 1145–1205. Crossref
Peel, J.S. 2017a. A problematic cnidarian (Cambroctoconus; Octocorallia?) from the Cambrian (Series 2–3) of Laurentia. Journal of Paleontology 91: 871–882. Crossref
Peel, J.S. 2017b. First record from Laurentia of some middle Cambrian (Series 3) sponge spicules. Alcheringa 41: 306–314. Crossref
Peel, J.S. 2018. Sponge spicules from the Holm Dal Formation (Cambrian Series 3, Guzhangian) of North Greenland (Laurentia). GFF 140: 306–317. Crossref
Peel, J.S. 2019. Sponge spicule assemblages from the Cambrian (Series 2–3) of North Greenland (Laurentia): systematics and biogeography. GFF 141: 133–161. Crossref
Peel, J.S. 2020. The oldest palaeoloricate mollusc (Cambrian Series 2, Stage 4; North Greenland) and its bearing on aculiferan evolution. Bulletin of Geosciences 95: 127–144. Crossref
Peel, J.S. 2021a. An outer shelf shelly fauna from Cambrian Series 2 (Stage 4) of North Greenland (Laurentia). Journal of Paleontology 95: 1–41. Crossref
Peel, J.S. 2021b. In-place operculum demonstrates that the Middle Cambrian Protowenella is a hyolith and not a mollusc. Alcheringa [published online https://doi.org/10.1080/03115518.2021.2004225]. Crossref
Peel, J.S. 2021c. Pseudomyona
from the Cambrian of North Greenland (Laurentia) and the early
evolution of bivalve molluscs. Bulletin of
Geosciences 96: 195–215. Crossref
Peel, J.S., Streng, M., Geyer, G., Kouchinsky, A., and Skovsted, C.B. 2016. Ovatoryctocara granulata assemblage (Cambrian Series 2–Series 3 boundary) of Løndal, North Greenland. Australasian Palaeontological Memoirs 49: 241–282.
Pegel, T.V. [Pegel’, T.V.], Egorova, L.I., Shabanov, Y.Y. [Šabanov, Û.Y.], Korovnikov, I.V., Luchinina, V.A. [Lučinina, V.A.], Salikhova, A.K. [Salihova, A.K.], Sundukov, V.M., Fedorov, A.B., Zhuravlev, A.Y. [Žuravlev, A.Û.], Parkhaev, P.Y. [Parhaev, P.Û.], and Demidenko, Y.E. [Demidenko, Û.E.] 2016. Stratigrafiâ neftegazonosnyh bassejnov Sibiri. Kembrij Sibirskoj platformy. Tom 2: Paleontologiâ. 344 pp. IPGG SB RAS, Novosibirsk.
Pelechaty, S.M., Grotzinger, J.P., Kashirtsev, V.A., and Zhernovsky, V.P. 1996a. Chemostratigraphic and sequence stratigraphic constraints on Vendian–Cambrian basin dynamics, northeast Siberian Craton. Journal of Geology 104: 543–564. Crossref
Pelechaty, S.M., Kaufman, A.J., and Grotzinger, J.P. 1996b. Evaluation of δ13C chemostratigraphy for intrabasinal correlation: Vendian strata of northeast Siberia. GSA Bulletin 108: 992–1003. Crossref
Pelman, Y.L. [Pel’man, Û.L.] 1973. Some inarticulate brachiopods from the Lower and Middle Cambrian of the Olenek River basin (lower reaches) [in Russian]. In: I.T. Žuravleva (ed.), Problemy paleontologii i biostratigrafii nižnego kembriâ Sibiri i Dal’nego Vostoka. Trudy Instituta Geologii i Geofiziki Sibirskogo Otdeleniâ Akademii Nauk SSSR 49: 69–80.
Pelman, Y.L. [Pel’man, Û.L.] 1977. Early and Middle Cambrian Inarticulate Brachiopods of the Siberian Platform [in Russian]. Trudy Instituta Geologii i Geofiziki Sibirskogo Otdeleniâ Akademii Nauk SSSR 316: 1–168.
Pelman, Y.L. [Pel’man, Û.L.] and Pereladov, V.S 1986. Stratigraphy and brachiopods of the Lower–Middle Cambrian of the Arga–Sala River (southern Anabar River Region) [in Russian]. In: I.T. Žuravleva (ed.), Biostratigfiâ i paleontologiâ kembriâ Severnoj Azii. Trudy Instituta Geologii i Geofiziki Sibirskogo Otdeleniâ Akademii Nauk SSSR 669: 119–154.
Pelman, Y.L. [Pel’man, Û.L.], Ermak, V.V., Fedorov, A.B., Luchinina, V.A. [Lučinina, V.A.], Zhuravleva, I.T. [Žuravleva, I.T.], Repina, L.N., Bondarev, V.I., and Borodaevskaya, Z.V. [Borodaevskaâ, Z.V.] 1990. New data on the upper Precambrian and lower Cambrian stratigraphy and palaeontology of the r. Dzhanda (right tributary of the r. Aldan) [in Russian]. In: L.N. Repina (ed.), Biostratigfiâ i paleontologiâ kembriâ Severnoj Azii. Trudy Instituta Geologii i Geofiziki Sibirskogo Otdeleniâ Akademii Nauk SSSR 765: 3–43.
Pérez-Huerta, A., Cusack, M., McDonald, S., Marone, F., Stampanoni, M., and MacKay, S. 2009. Brachiopod punctae: A complexity in shell biomineralisation. Journal of Structural Biology 167: 62–67. Crossref
Pickett, J. 2002. Order Heteractinida Hinde, 1887. In: J.N.A. Hooper and R.W.M. Van Soest (eds.), Systema Porifera: A Guide to the Classification of Sponges. Vol. 2, 1121–1140. Kluwer Academic/Plenum Publishers, New York. Crossref
Pojeta, J.P., Jr. 1975. Fordilla troyensis Barrande and early bivalve phylogeny. Bulletins of American Paleontology 63: 363–384.
Poulsen, C. 1932. The Lower Cambrian faunas of East Greenland. Meddelelser om Grønland 87: 1–66.
Poulsen, C. 1967. Fossils from the Lower Cambrian of Bornholm. Kongelige Danske Videnskabernes Selskab, Matematisk-Fysiske Meddelelser 36: 1–46.
Poulsen, V. 1966. Early Cambrian distacodontid conodonts from Bornholm. Biologiske Meddelelser Danske Videnskabernes Selskab 23, 1–10.
Pruss, S.B., Dwyer, C.H., Smith, E.F., Macdonald, F.A., and Tosca, N.J. 2019. Phosphatised early Cambrian archaeocyaths and small shelly fossils (SSFs) of southwestern Mongolia. Palaeogeography, Palaeoclimatology Palaeoecology 513: 166–177. Crossref
Qian, Y. 1978. The Early Cambrian hyolithids in central and southwest China and their stratigraphical significance [in Chinese with English abstract]. Memoirs of Nanjing Institute of Geology and Palaeontology, Academia Sinica 11: 1–38.
Qian, Y. and Bengtson, S. 1989. Palaeontology and biostratigraphy of the Early Cambrian Meishucunian Stage in Yunnan Province, south China. Fossils and Strata 24: 1–156.
Qian, J.-X. and Xiao, B. 1984. An Early Cambrian small shelly fauna from Aksu–Wushi region, Xinjiang [in Chinese, with English abstract]. Professional Papers of Stratigraphy and Palaeontology 13: 65–90.
Qian, Y., Li, G.-X., Zhu, M.-Y., Steiner, M., and Erdtmann, B.-D. 2004. Early Cambrian protoconodonts and conodont-like fossils from China: taxonomic revisions and stratigraphic implications. Progress in Natural Science 14: 173–180. Crossref
Qian, Y., Xie, Y., and He, T. 2001. Hyoliths of the Lower Cambrian Chiungchussuan Stage in South Shaanxi Province. Acta Palaeontologica Sinica 40: 31–43.
Qian, Y., Zhu, M.-Y., and Li, G.-X. 2000. Reconsiderations of the phylogenetic affinities of zhijinitids [in Chinese, with English abstract]. Acta Micropalaeontologica Sinica 17: 307–316.
Rahman, I.A. and Clausen, S. 2009. Re-evaluating the palaeobiology and affinities of the Ctenocystoidea (Echinodermata). Journal of Systematic Palaeontology 7: 413–426. Crossref
Raymond, P.E. 1935. Leanchoilia and other mid-Cambrian Arthropoda. Bulletin of the Museum of Comparative Zoology, Harvard 46: 202–230.
Reif, W.-E. 1968. Schwämmreste aus dem oberen Ordovizium von Estland und Schweden. Neues Jahrbuch für Geologie und Paläontologie, Monatshefte 1968 (12): 733–744.
Reitner, J. 1992. “Coralline Spongien”. Der Versuch einer phylogenetisch-taxonomischen Analyse. Berliner Geowissenschaftliche Abhandlungen, Reihe E 1: 1–352.
Reitner, J. and Mehl, D. 1995. Early Paleozoic diversification of sponges: New data and evidences. Geologische Paläontologische Mitteilungen der Universität Innsbruck 20: 335–347.
Repina, L.N., Lazarenko, N.P., Meshkova, N.P. [Meškova, N.P.], Korshunov, V.I. [Koršunov, V.I.], Nikiforov, N.I., and Aksarina, N.A. 1974. Biostratigraphy and fauna of the Lower Cambrian of Kharaulakh (Tuora-Sis ridge) [in Russian]. Trudy Instituta Geologii i Geofiziki Sibirskogo Otdeleniâ Akademii Nauk SSSR 235: 1–299.
Rigby, K. 1975. Some unusual hexactinellid sponge spicules from the Cambrian Wilberns Formation of Texas. Journal of Paleontology 49: 412–415.
Rigby, J.K. 1986. Sponges of the Burgess shale (Middle Cambrian), British Columbia. Palaeontographica Canadiana 2: 1–105.
Rigby, J.K. and Collins, D. 2004. Sponges of the Middle Cambrian Burgess Shale and Stephen Formations, British Columbia. Royal Ontario Museum Contributions in Science 1: 1–164.
Rigby, J.K. and Toomey, D.F. 1978. A distinctive sponge spicule assemblage from organic buildups in the Lower Ordovician of southern Oklahoma. Journal of Paleontology 52: 501–506.
Robison, R.A. and Sprinkle, J. 1969. Ctenocystoidea: new class of primitive echinoderms. Science 166: 1512–1514. Crossref
Rogov, V.I., Karlova, G.A., Marusin, V.V., Kochnev, B.B., Nagovitsin, K.E., and Grazhdankin, D.V. 2015. Duration of the first biozone in the Siberian hypostratotype of the Vendian. Russian Geology and Geophysics 56: 501–511. Crossref
Rowell, A.J. 1965. Inarticulata. In: R.C. Moore (ed.), Treatise on Invertebrate Paleontology. Part H, Brachiopoda, Volume 1, H260–H296. Geological Society of America, New York and University of Kansas Press, Lawrence.
Rowell, A.J. 1980. Inarticulate brachiopods of the Lower and Middle Cambrian Pioche Shale of the Pioche district, Nevada. University of Kansas, Paleontological Contributions, Paper 98: 1–26.
Rozanov, A.Y. [Rozanov, A.Û.] and Missarzhevsky, V.V. [Missarževskij, V.V.] 1966. Biostratigraphy and fauna of the Cambrian lower horizons [in Russian]. Trudy Geologičeskogo Instituta Akademii Nauk SSSR 148: 1–126.
Rozanov, A.Y., Khomentovsky, V.V., and Shabanov, Y.Y., Karlova, G.A., Varlamov, A.I., Luchinina, V.A., Pegel’, T.V., Demidenko, Y.E., Parkhaev, P.Y., Korovnikov, I.V., and Skorlotova, N.A. 2008. To the problem of stage subdivision of the Lower Cambrian. Stratigraphy and Geological Correlation 16: 1–19.
Rozanov, A.Y. [Rozanov, A.Û.], Missarzhevsky, V.V. [Missarževskij, V.V.], Volkova, N.A., Voronova, L.G., Krylov, I.N., Keller, B.M., Korolyuk, I.K. [Korolûk, I.K.], Lendzion, K., Michniak, R., Pyhova, N.G., and Sidorov, A.D. 1969. The Tommotian Stage and the Cambrian lower boundary problem [in Russian]. Trudy Geologičeskogo Instituta Akademii Nauk SSSR 206: 1–380.
Rozanov, A.Y. [Rozanov, A.Û.], Parkhaev, P.Y. [Parhaev, P.Û.], Demidenko, Y.E. [Demidenko, Û.E.], Karlova, G.A., Korovnikov, I.V., Shabanov, Y.Y. [Šabanov, Û.â], Ivantsov, A.Y. [Ivancov, A.Û.], Luchinina, V.A. [Lučinina, V.A.], Malakhovskaya, Y.E. [Malahovskaâ, Â.E.], Mel’nikova, L.M., Naimark, E.B. [Najmark, E.B.], Ponomarenko, A.G., Skorlotova, N.A., Sundukov, V.M., Tokarev, D.A., Ushatinskaya, G.T. [Ušatinskaâ, G.T.], and Kipriyanova, L.D. [Kipriânova, L.D.] 2010. Iskopaemye stratotipov ârusov nižnego kembriâ. 228 pp. PIN RAN, Moskva.
Runnegar, B. and Jell, P. A. 1976. Australian Middle Cambrian Molluscs and their bearing on early molluscan evolution. Alcheringa 1: 109–138. Crossref
Runnegar, B. and Jell, P.A. 1980. Australian Middle Cambrian molluscs: corrections and additions. Alcheringa 4: 111–113. Crossref
Sarsembaev, Z.A. and Marusin, V.V. 2022. Nonmineralized triradial conulariids from the lowermost Cambrian Stage 2 of the Olenek Uplift, Siberian Platform. Journal of Paleontology [published online, https://doi.org/10.1017/jpa.2022.21]. Crossref
Schiemann S.M., Martín-Durán, J.M., Børve, A., Vellutini, B.C., Passamaneck, Y.J., and Hejnol, A. 2017. Clustered brachiopod Hox genes are not expressed collinearly and are associated with lophotrochozoan novelties. Proceedings of the National Academy of Sciences of the USA 114: E1913–E1922. Crossref
Schindewolf, O.H. 1955. Über einige kambrische Gattungen inartikulater Brachiopoden. Neues Jahrbuch für Geologie und Paläontologie, Monatshefte 1955 (12): 530–557.
Schmidt, O. 1870. Grundzüge einer Spongienfauna des atlantischen Gebietes. 88 pp. Wilhelm Engelmann, Leipzig.
Schuchert, C. 1893. Classification of the Brachiopoda. American Geologist 11: 141–167.
Schuster, A., Erpenbeck, D., Pisera, A., Hooper, J., Bryce, M., Fromont, J., and Wörheide, G. 2015. Deceptive desmas: Molecular phylogenetics suggests a new classification and uncovers convergent evolution of lithistid sponges. PLoS ONE 10: e116038. Crossref
Sdzuy, K. 1969. Unter- und mittelkambrische Porifera (Chancelloriida und Hexactinellida). Paläontologische Zeitschrift 43: 115–147. Crossref
Shabanov, Y.Y. [Šabanov, Û.â], Astashkin, V.A. [Astaškin, V.A.], Pegel, T.V. [Pegel’, T.V.], Egorova, L.I., Zhuravleva, I.T. [Žuravleva, I.T.], Pelman, Y.L. [Pel’man, Û.L.], Sundukov, V.M., Stepanova, M.V., Sukhov, S.S. [Suhov, S.S.], Fedorov, A.B., Shishkin, B.B. [Šiškin, B.B.], Vaganova, N.V., Ermak, V.I., Ryabukha, K.V. [Râbuha, K.V.], Yadrenkina A.G. [Âdrenkina, A.G.], Abaimova, G.P., Lopushinskaya, T.V. [Lopušinskaâ, T.V.], Sychev, O.V. [Syčev, O.V.], and Moskalenko, T.A. 1987. Nižnij paleozoj ûgo-zapadnogo sklona Anabarskoj anteklizy (po materialam bureniâ). 208 pp. Nauka, Novosibirsk.
Shaler, N.S. and Foerste, A.F. 1888. Preliminary description of North Attleborough fossils, Bulletin of the Museum of Comparative Zoology 16: 27–41.
Sipin, D.P. 1999. New genus of problematic organisms from the Lower Cambrian of north-western Siberian Platform [in Russian]. Paleontologičeskij žurnal 1999 (2): 8–10.
Sipin, D.P. 2001. Hiolity, problematičnye organizmy i stratigrafiâ nižnego kembriâ severo-zapada Sibirskoj platformy. 201 pp. Unpublished Doctoral Candidate Thesis, Institut Geologii Nefti i Gaza SO RAN, Novosibirsk.
Siegmund, H. 1997. The Ocruranus–Eohalobia group of small shelly fossils from the Lower Cambrian of Yunnan. Lethaia 30: 285–291. Crossref
Skovsted, C.B. 2004. The mollusc fauna of the Early Cambrian Bastion Formation of North-East Greenland. Bulletin of the Geological Society of Denmark 51: 11–37. Crossref
Skovsted, C.B. 2006a. Small shelly fauna from the upper Lower Cambrian Bastion and Ella Island formations, north-east Greenland. Journal of Paleontology 80: 1087–1112. Crossref
Skovsted, C.B. 2006b. Small shelly fossils from the basal Emigrant Formation (Cambrian, uppermost Dyeran Stage) of Split Mountain, Nevada. Canadian Journal of Earth Sciences 43: 487–496. Crossref
Skovsted, C.B. and Holmer, L.E. 2006. The Lower Cambrian brachiopod Kyrshabaktella and associated shelly fossils from the Harkless Formation, southern Nevada. GFF 128: 327–337. Crossref
Skovsted, C.B. and Peel, J.S. 2007. Small shelly fossils from the argillaceous facies of the Lower Cambrian Forteau Formation of western Newfoundland. Acta Palaeontologica Polonica 52: 729–748.
Skovsted, C.B. and Peel, J.S. 2010. Early Cambrian brachiopods and other shelly fossils from the basal Kinzers Formation of Pennsylvania. Journal of Paleontology 84: 754–762. Crossref
Skovsted, C.B., Balthasar, U., Brock, G.A., and Paterson, J.R. 2009. The tommotiid Camenella reticulosa from the lower Cambrian of South Australia, scleritome reconstructions and tommotiid phylogeny. Acta Palaeontologica Polonica 54: 525–540. Crossref
Skovsted, C.B., Martí Mus, M., Zhang, Z., Pan, B., Li, L., Liu, F., Li, G., and Zhang, Z. 2020. On the origin of hyolith helens. Palaeogeography, Palaeoclimatology, Palaeoecology 555: 109848. Crossref
Skovsted, C.B., Streng, M., Knight, I., and Holmer, L. E. 2010. Setatella significans, a new name for mickwitziid stem group brachiopods from the lower Cambrian of Greenland and Labrador. GFF 132: 117–122. Crossref
Skovsted, C.B., Topper, T.P., McLoughlin, S., Johansson, O., Liu, F., and Vajda, V. 2021. First discovery of Small Shelly Fossils and new occurrences of brachiopods and trilobites from the early Cambrian (Stage 4) of the Swedish Caledonides, Lapland. GFF 143: 134–150. Crossref
Skovsted, C.B., Ushatinskaya, G., Holmer, L.E., Popov, L.E., and Kouchinsky, A. 2015. Taxonomy, morphology, shell structure and early ontogeny of Pelmanotreta nom. nov. from the lower Cambrian of Siberia. GFF 137: 1–8. Crossref
Sprinkle, J. 1973. Morphology and Evolution of Blastozoan Echinoderms. 296 pp. Harvard University Museum of Comparative Zoology, Cambridge. Crossref
Sprinkle, J. and Robison, R.A. 1978. Addendum to subphylum Homalozoa. Ctenocystoids. In: R. C. Moore and C. Teichert (eds.), Echinodermata 2, Crinoidea. Part T. Treatise on invertebrate paleontology, T998–T1002. Geological Society of America, New York
Steiner, M., Li, G., Qian, Y., Zhu, M., Erdtmann, B.D. 2007. Neoproterozoic to early Cambrian small shelly fossil assemblages and a revised biostratigraphic correlation of the Yangtze Platform (China). Palaeogeography, Palaeoclimatology, Palaeoecology 254: 67–99. Crossref
Streng, M., Babcock, L.E., and Hollingsworth, J.S. 2005. Agglutinated protists from the Lower Cambrian of Nevada. Journal of Paleontology 79: 1214–1218. Crossref
Sugai, Y., Iwata, K., Sennikov, N.V., Obut, O.T., and Khlebnikova T.V. 2004. Sponge spicules from the Lower Cambrian of Bateny Ridge and Gorny Altai (Bagrad and Shashkunar formations). Novosti paleontologii i stratigrafii, Priloženie k žurnalu “Geologiâ i geofizika” 45 (6–7): 59–73.
Sumrall, C.D., Sprinkle, J., and Guensburg, T.E. 2001. Comparison of flattened blastozoan echinoderms: Insights from the new early Ordovician eocrinoid Haimacystis rozhnovi. Journal of Paleontology 75: 985–992. Crossref
Sysoev, V.A. 1957. On the morphology, systematics and affinity of hyoliths [in Russian]. Doklady Akademii Nauk SSSR 116: 304–307.
Sysoev, V.A. 1962. Hiolity kembri severnogo sklona Aldanskogo ŝita. 66 pp. Izdatel’stvo Akademii Nauk SSSR, Moskva.
Taylor, T.G. 1910. The Archaeocyathinae from the Cambrian of South Australia with an account of the morphology and affinities of the whole class. Memoirs of the Royal Society of South Australia 2: 55–188.
Thomas, R.D.K., Runnegar, B., and Matt, K. 2020. Pelagiella exigua, an early Cambrian stem gastropod with chaetae: lophotrochozoan heritage and conchiferan novelty. Palaeontology 63: 601–627. Crossref
Topper, T.P., Skovsted, C.B., Harper, D.A.T., and Ahlberg, P. 2013. A bradoriid and brachiopod dominated shelly fauna from the Furongian (Cambrian) of Västergötland, Sweden. Journal of Paleontology 87: 69–83. Crossref
Tsukui, K., Isozaki, Y., Zhu, M., Ramezani, J., Sato, T., Zhang, X., and Bowring, S.A. 2017. High-precision U-Pb temporal constraints on the early Cambrian diversification of animal life from eastern Yunnan, China. JpGU-AGU Joint Meeting 2017. Abstract [published online, https://confit.atlas.jp/guide/event-img/jpguagu2017/BCG09-01/public/pdf?type=in]
Ubaghs, G. and Robison, R.A. 1988. Homalozoan echinoderms of the Wheeler Formation (Middle Cambrian) of western Utah. University of Kansas Paleontological Contributions, Paper 120: 1–17.
Uriz, M.-J. 2006. Mineral skeletogenesis in sponges. Canadian Journal of Zoology 84: 322–356. Crossref
Ushatinskaya, G.T. [Ušatinskaâ, G.T.] 1987. Unusual inarticulate brachiopods from the Lower Cambrian of Mongolia [in Russian]. Paleontologičeskij žurnal 1987 (2): 62–68.
Ushatinskaya, G.T. 1997. New Middle and Late Cambrian lingulids and paterinids (Brachiopoda) from the north of the Siberian Platform. Paleontological Journal 31: 489–497.
Ushatinskaya, G.T. 2016. Protegulum and brephic shell of the earliest organophosphatic brachiopods. Paleontological Journal 50: 141–152. Crossref
Ushatinskaya, G.T. and Korovnikov, I.V. 2014. Revision of the Early–Middle Cambrian Lingulida (Brachiopoda) from the Siberian Platform, Paleontological Journal 48: 26–40. Crossref
Ushatinskaya, G.T. and Korovnikov, I.V. 2016. Revision of the Superfamily Acrotheloidea (Brachiopoda, Class Linguliformea, Order Lingulida) from the Lower and Middle Cambrian of the Siberian Platform. Paleontological Journal 50: 450–462. Crossref
Ushatinskaya, G.T. and Korovnikov, I.V. 2019. Revision of the Early and Middle Cambrian Acrotretids (Brachiopoda, Linguliformea) from the Siberian Platform. Paleontological Journal 53: 689–714. Crossref
Val’kov, A.K. 1982. Biostratigrafiâ nižnego kembriâ vostoka Sibirskoj platformy (učuro-majskij rajon). 91 pp. Nauka, Moskva.
Val’kov, A.K. 1983. Distribution of the oldest skeletal fossils and correlation of the Cambrian lower boundary in south-eastern part of the Siberian Platform [in Russian]. In: V.V. Homentovskij, M.S. Âkšin, and G.A. Karlova (eds.), Pozdnij dokembrij i rannij paleozoj Sibiri. Vendskie otloženiâ, 37–48. Institut Geologii i Geofiziki SO AN SSSR, Novosibirsk.
Val’kov, A.K. and Karlova, G.A. 1984. Fauna from the Vendian–Cambrian transition beds in the lower reaches of the Gonam River [in Russian]. In: V.V. Homentovskij, V.Û. Šenfil’, and M.S. Âkšin (eds.), Stratigrafiâ pozdnego dokembriâ i rannego paleozoâ. Srednââ Sibir’, 12–41. Institut Geologii i Geofiziki SO AN SSSR, Novosibirsk.
Val’kov, A.K. and Sysoev, V.A. 1970. Cambrian angustiochreids of Siberia [in Russian]. In: A.K. Bobrov (ed.), Stratigrafiâ i paleontologiâ proterozoâ i kembriâ vostoka Sibirskoj platformy, 94–100. Âkutskoe Knižnoe Izdatel’stvo, Âkutsk.
Vannier, J., Williams, M., Alvaro, J.J., Vicaïno,
D., Monceret, S., and Monceret, E. 2005. New Early Cambrian bivalved
arthropods from southern France. Geological
Magazine 142: 751–763. Crossref
Vasiljeva, N.I. 1985. A contribution to the systematics of the Order Chancelloriida Walcott, 1920 (incertae sedis) from Lower Cambrian deposits of the East Siberian Platform [in Russian]. In: B.S. Sokolov and I.T. Žuravleva (eds.), Problematiki pozdnego dokembriâ i paleozoâ. Trudy Instituta Geologii i Geofiziki Sibirskogo Otdeleniâ Akademii Nauk SSSR 632: 115–126.
Vasiljeva, N.I. 1990. New Early Cambrian gastropod mollusks from the Siberian Platform and some questions of their systematics [in Russian]. In: A.I. Nikolaev (ed.), Microfauna SSSR: Voprosy sistematiki i biostratigrafii, 4–21. VNIGRI, Leningrad.
Vasiljeva, N.I. 1998. Melkaâ rakovinnaâ fauna i biostratigrafiâ nižnego kembriâ Sibirskoj platformy. 189 pp. Vserossijskij Naučno-Issledovatel’skij Geologo-Razvedočnyj Institut, Sankt-Petersburg.
Vasiljeva, N.I. and Sayutina, T.A. [Saûtina, T.A.] 1988 Morphological diversity of chancelloriid sclerites [in Russian]. In: I.T. Žuravleva and L.N. Repina (eds.), Kembrij Sibiri i Srednej Azii. Trudy Instituta Geologii i Geofiziki Sibirskogo Otdeleniâ Akademii Nauk SSSR 720: 190–208.
Vasiljeva, N.I. and Sayutina, T.A. [Saûtina, T.A.] 1993. New generic and species names of Early Cambrian sclerites of chancelloriids [in Russian]. Paleontologičeskij žurnal 1993 (1): 113–114.
Vendrasco, M.J., Checa, A.G., and Kouchinsky, A.V. 2011a. Shell microstructure of the early bivalve Pojetaia and the independent origin of nacre within the Mollusca. Palaeontology 54: 825–850. Crossref
Vendrasco, M.J., Kouchinsky, A., Porter, S.M., and Fernandez, C.Z. 2011b. Phylogeny and escalation in Mellopegma and other Cambrian molluscs. Palaeontologia Electronica 14(2): 1–44.
Vinn, O. and Mutvei, H. 2009. Calcareous tubeworms of the Phanerozoic. Estonian Journal of Earth Sciences 58: 286–296. Crossref
Vinther, J. 2009. The canal system in sclerites of Lower Cambrian Sinosachites (Halkieridae: Sachitida): Significance for the molluscan affinities of the sachitids. Palaeontology 52: 689–712. Crossref
Vinther, J. and Nielsen, C. 2005. The Early Cambrian Halkieria is a mollusk. Zoologica Scripta 34: 81–89. Crossref
Vologdin, A.G. 1932. Arheociaty Sibiri. Vypusk 2. Fauna kembrijskih izvestnâkov Altaâ. 106 pp. Gosudarstvennoe Naučno-Tehničeskoe Geologo-Razvedočnoe Izdatel’stvo, Moskva.
Vologdin, A.G. 1940. Archaeocyaths and algae of the Cambrian limestones of Mongolia and Tuva, Part 1 [in Russian]. Trudy Mongol’skoj Komissii Akademii Nauk SSSR 34: 1–268.
Von Siebold, C.T. 1848. Lehrbuch der vergleichenden Anatomie der Wirbellosen Thiere. Erster Theil. In: C.T. von Siebold and H. Stannius (eds.), Lehrbuch der vergleichenden Anatomie, 679 pp. Verlag von Veit & Comp., Berlin. Crossref
Voronin, Y.I., Voronova, L.G., Grigor’eva, N.V., Drozdova, N.A., Zhegallo, E.A. [Žegallo, E.A.], Zhuravlev, A.Y. [Žuravlev, A.Û.], Ragozina, A.L., Rozanov, A.Y. [Rozanov, A.Û.], Sayutina, T.A. [Saûtina, T.A.], Sysoev, V.A., and Fonin, V.D. 1982. The Precambrian–Cambrian boundary in the geosynclinal regions (reference section Salany-Gol, MNR) [in Russian]. Trudy Sovmestnoj Sovecko-Mongol’skoj Paleontologičeskoj Ekspedicii 18: 1–150.
Voronova, L.G. and Missarzhevsky, V.V. [Missarževskij, V.V.] 1969. Finds of algae and worm tubes in the Precambrian–Cambrian boundary beds of the northern part of the Siberian Platform [in Russian]. Doklady Akademii Nauk SSSR 184: 207–210.
Voronova, L.G., Drozdova, N.A., Esakova, N.V., Zhegallo, E.A. [Žegallo, E.A.], Zhuravlev, A.Y. [Žuravlev, A.Û.], Rozanov, A.Y. [Rozanov, A.Û.], Sayutina, T.A. [Saûtina, T.A.], and Ushatinskaya, G.T. [Ušatinskaâ, G.T.] 1987. Lower Cambrian fossils from the Mackenzie Mountains (Canada) [in Russian]. Trudy Paleontologičeskogo Instituta Akademii Nauk SSSR 224: 1–88.
Vostokova, V.A. 1962. Cambrian gastropods from Siberia and Taymyr [in Russian]. Trudy Naučno-Issledovatel’skogo Instituta Geologii Arktiki 28: 51–74.
Waagen, W. 1885. Salt Range fossils. Brachiopoda. Memoirs, Palaeontologia Indica, Series 13 (1) 5: 729–770.
Walcott, C.D. 1886. Second contribution to the studies on the Cambrian faunas of North America. Bulletin of the United States Geological Survey 30: 1–369. Crossref
Walcott, C.D. 1889. Descriptive notes on new genera and species from the Lower Cambrian or Olenellus Zone of North America. Proceedings of the U.S. National Museum 12: 33–46. Crossref
Walcott, C.D. 1908. Cambrian Brachiopoda: Description of new genera and species. Smithsonian Miscellaneous Collections 53: 53–137.
Walcott, C.D. 1920. Cambrian geology and paleontology. Middle Cambrian Spongiae. Smithsonian Miscellaneous Collections 67: 261–364.
Webby, B.D. and Trotter, J. 1993. Ordovician sponge spicules from New South Wales, Australia. Journal of Paleontology 67: 28–41. Crossref
Wenz, W. 1938. Gastropoda. Allgemeiner Teil und Prosobranchia. In: O.H. Schindewolf (ed.), Handbuch der Paläozoologie, Band 6, 1–720. Borntrager, Berlin.
White, C.A. 1874. Preliminary Report Upon Invertebrate Fossils Collected by the Expeditions of 1871, 1872, and 1873, With Descriptions of New Species. U.S. Army Geographical and Geological Explorations and Surveys West of the One Hundredth Meridian. 27 pp. Government Printing Office, Washington.
Whittard, W.F. 1953. Palaeoscolex piscatorum gen. et sp. nov., a worm from the Tremadocian of Shropshire. Quarterly Journal of the Geological Society of London 109: 125–136. Crossref
Williams, A. and Cusack, M. 1999. Evolution of a rhythmic lamination in the organophosphatic shells of brachiopods. The Journal of Structural Biology 126: 227–240. Crossref
Williams, A. and Holmer, L.E. 1992. Ornamentation and shell structure of acrotretoid brachiopods. Palaeontology 35: 657–692.
Williams, A., Carlson, S.J., Brunton, C.H.C., Holmer, L.E., and Popov, L.E. 1996. A supra-ordinal classification of the Brachiopoda. Philosophical Transactions of the Royal Society of London, Series B 351: 1117–1193. Crossref
Williams, M., Siveter, D.J., Hinz-Schallreuter, I., and Melnikova, L. 1994. On Cambria sibirica Neckaja and Ivanova. A Stereo-Atlas of Ostracod Shells 21: 9–12.
Wiman, C. 1903. Studien über das Nordbaltische Silurgebiet. I. Olenellussandstein, Obolussandstein und Ceratopygeschieffer. Bulletin of the Geological Institution of the University of Uppsala 6: 12–76.
Wood, R.A., Zhuravlev, A.Y., and Chimed Tseren, A. 1993. The ecology of Lower Cambrian buildups from Zuune Arts, Mongolia: Implications for early metazoan reef evolution. Sedimentology 40: 829–858. Crossref
Wotte, T. 2006. New Middle Cambrian molluscs from the Láncara Formation of the Cantabrian Mountains (northwestern Spain). Revista Española de Paleontología 21: 145–158. Crossref
Wotte, T. 2009. The youngest cambroclaves: Cambroclavus absonus from the Middle Cambrian of the Cantabrian zone (northwestern Spain). Journal of Paleontology 83: 128–134. Crossref
Wotte, T., Strauss, H., and Sundberg, F.A. 2011. Carbon and sulfur isotopes from the Cambrian Series 2–Cambrian Series 3 of Laurentia and Siberia. Museum of Northern Arizona Bulletin 67: 43–63.
Wrona, R. 2003. Early Cambrian molluscs from glacial erratics of King George Island, West Antarctica. Polish Polar Research 24: 181–216.
Wrona, R. 2004. Cambrian microfossils from glacial erratics of King George Island, Antarctica. Acta Palaeontologica Polonica 49: 13–56.
Wu, W., Zhu, M., and Steiner, M. 2014. Composition and tiering of the Cambrian sponge communities. Palaeogeography, Palaeoclimatology, Palaeoecology 398: 86–96. Crossref
Xiao, L. and Zhou, B. 1984. Early Cambrian Hyolitha from Huainan and Huoqiu County in Anhui Province. Professional Papers on Stratigraphy and Palaeontology 13: 141–151.
Xing, Y., Ding Q., Luo H., He, T., Wang Y., Jiang, Z., Chen, Y., Zhang, S., Liu, G., Xiong, X., Chen, P., Wang, Y., Yin, G., Zheng, S., Qian, Y., Yue, Z., Wu, X., Song, X., Ouyang, L., and Ding, L. 1984 [date of cover 1983]. The Sinian–Cambrian boundary of China [in Chinese with English summary]. Bulletin of the Institute of Geology, Chinese Academy of Geological Sciences 10: 1–262.
Yin, J., Ding, L., He, T., Li, S., and Shen, L. 1980. The Palaeontology and Sedimentary Environment of the Sinian System in Emei-Ganluo Area, Sichuan [in Chinese with English absract]. 231 pp. Sichuan People’s Publishing House, Chengdu.
Yu, W. 1974. Cambrian hyolithids. In: Nanjing Institute of Geology and Paleontology (ed.), Handbook of Stratigraphy and Palaeontology of Southwest China, 111–112. Academia Sinica, Science Press, Beijing.
Yu, W. 1979. Earliest Cambrian monoplacophorans and gastropods from western Hubei with their biostratigraphical significance [in Chinese with English absract]. Acta Palaeontologica Sinica 18: 233–270.
Zakrzewski, A.-C., Weigert A., Helm, C., Adamski, M., Adamska, M., Bleidorn, C., Raible, F., and Hausen, H. 2014. Early divergence, broad distribution, and high diversity of animal chitin synthases. Genome Biology & Evolution 6: 316–325. Crossref
Zamora, S. 2010. Middle Cambrian echinoderms from North Spain show echinoderms diversified earlier in Gondwana. Geology 38: 507–510. Crossref
Zamora, S. and Smith, A.B. 2012. Cambrian stalked echinoderms show unexpected plasticity of arm construction. Proceedings of the Royal Society of London B 279: 293–298. Crossref
Zamora, S., Deline, B., Álvaro, J.J., and Rahman, I.A. 2017. The Cambrian Substrate Revolution and the early evolution of attachment in suspension-feeding echinoderms. Earth-Science Reviews 171: 478–491. Crossref
Zamora, S., Lefebvre, B., Álvaro, J.J., Clausen, S., Elicki, O., Fatka, O., Jell, P., Kouchinsky, A., Jih-Pai, L., Nardin, E., Parsley, R., Rozhnov, S., Sprinkle, J., Sumrall, C.D., Vizcaino, D., and Smith, A.B. 2013. Cambrian echinoderm diversity and palaeobiogeography. In: D. Harper and T. Servais (eds.), Early Palaeozoic Palaeography and Biogeography. Geological Society, London, Memoirs 38: 157–171. Crossref
Zamora, S., Rahman, I.A., and Smith, A.B. 2012. Plated Cambrian bilaterians reveal the earliest stages of echinoderm evolution. PLoS ONE 7 (6): e38296. Crossref
Zhamoida, A.I. [Žamojda, A.I.] (ed.) 2016. Postanovleniâ mežvedomstvennogo stratigrafičeskogo komiteta i ego postojannyh komissij, Vol. 44. 68 pp. VSEGEI, Sankt-Petersburg.
Zhang, X.-G. and Pratt, B.R. 1994. New and extraordinary Early Cambrian sponge spicule assemblage from China. Geology 22: 43–46. Crossref
Zhang, Z.F., Zhang, Z.L., Li, G.X., and Holmer, L.E. 2016. The Cambrian brachiopod fauna from the first-trilobite age Shuijingtuo Formation in the Three Gorges area of China. Palaeoworld 25: 333–355. Crossref
Zhang, Z.L., Skovsted, C.B., and Zhang, Z.F. 2018. A hyolithid without helens preserving the oldest hyolith muscle scars; palaeobiology of Paramicrocornus from the Shujingtuo Formation (Cambrian Series 2) of South China. Palaeogeography, Palaeoclimatology, Palaeoecology 489: 1–14. Crossref
Zhang, Z.L., Zhang, Z.F., Ma, J., Taylor, P.D., Strotz, L.C., Jacquet, S.M., Skovsted, C.B., Chen, F., Han, J., and Brock, G.A. 2021. Fossil evidence unveils an early Cambrian origin for Bryozoa. Nature 599: 251–255. Crossref
Zhao, F., Caron, J.-B., Hu, S., and Zhu, M. 2009. Quantitative analysis of taphofacies and paleocommunities in the early Cambrian Chengjiang Lagerstätte. Palaios 24: 829–839. Crossref
Zhao, J., Li, G.-B., and Selden, P.A. 2018. New well-preserved scleritomes of Chancelloriida from early Cambrian Guanshan Biota, eastern Yunnan, China. Journal of Paleontology 92: 955–971. Crossref
Zhou, B. and Xiao, L. 1984. Early Cambrian monoplacophorans and gastropods from Huainan and Huoqiu counties, Anhui Province [in Chinese]. Professional Papers of Stratigraphy and Palaeontology, Chinese Academy of Geological Sciences 13: 125–140.
Zhu, M.-Y., Babcock, L.E., and Peng, S.-C. 2006. Advances in Cambrian stratigraphy and paleontology: Integrating correlation techniques, paleobiology, taphonomy and paleoenvironmental reconstruction. Palaeoworld 15: 217–222. Crossref
Zhuravlev, A.Y. 1998. Outlines of the Siberian Platform sequence stratigraphy in the Lower and lower Middle Cambrian (Lena-Aldan area). Revista Española de Paleontología No Extr. Homenaje al Prof. Gonzalo Vidal: 105–114. Crossref
Zhuravlev, A.Y. 2001. Paleoecology of Cambrian reef ecosystems. In: G.D. Stanley, Jr. (ed.), The History and Sedimentology of Ancient Reef Systems, 121–157. Kluwer Academic/Plenum Publishers, New York. Crossref
Zhuravlev, A.Y. [Žuravlev, A.Û.] and Sayutina, T.A. [Saûtina, T.A.] 1985. Radiocyaths of Mongolia. Toward the revision of the “Class” Radiocyatha [in Russian]. In: B.S. Sokolov and I.T. Žuravleva (eds.), Problematiki pozdnego dokembriâ i paleozoâ. Trudy Instituta Geologii i Geofiziki Sibirskogo Otdeleniâ Akademii Nauk SSSR 632: 52–62.
Zhuravlev, A.Y. and Wood, R.A. 1996. Anoxia as the cause of the mid-Early Cambrian (Botomian) extinction event. Geology 24: 311–314. Crossref
Zhuravlev, A.Y., Liñán, E., Gámez Vintaned, J.A., Debrenne, F., and Fedorov, A.B. 2012. New finds of skeletal fossils in the terminal Neoproterozoic of the Siberian Platform and Spain. Acta Palaeontologica Polonica 57: 205–224. Crossref
Zhuravlev, A.Y., Naimark, E.B., and Wood, R.A. 2015. Controls on the diversity and structure of earliest metazoan communities: early Cambrian reefs from Siberia. Earth-Science Reviews 147: 18–29. Crossref
Zhuravlev, A.Y. [Žuravlev, A.Û.], Zhuravleva, I.T. [Žuravleva, I.T.], Korshunov, V.I. [Koršunov, V.I.], Osadchaya, D.V. [Osadčaâ, D.V], Rozanov, A.Y. [Rozanov, A.Û.], and Sundukov, V.M. 1983. Phylum Archaeocyatha. Class Regularia [in Russian]. In: B.S. Sokolov and I.T. Žuravleva (eds.), Ârusnoe rasclenenie nižnego kembriâ Sibiri, Atlas okamenelostej. Trudy Instituta Geologii i Geofiziki Akademii Nauk SSSR 558: 22–30.
Zhuravleva, I.T. [Žuravleva, I.T.] 1954. Nastavlenie po sboru i izučeniû arheociat. 46 pp. Paleontologičeskij Institut Akademii Nauk SSSR, Moskva.
Zhuravleva, I.T. [Žuravleva, I.T.] 1955. Cambrian archaeocyaths of the eastern flank of the Kuznetsk Ala-Tau [in Russian]. Trudy Paleontologičeskogo Instituta Akademii Nauk SSSR 56: 5–56.
Zhuravleva, I.T. [Žuravleva, I.T.] 1960. Arheociaty Sibirskoj platformy. 344 pp. Akademiâ Nauk SSSR, Moskva.
Zhuravleva, I.T. [Žuravleva, I.T.] and Korde, K.B. 1955. A find of the sponge Chancelloria Walcott, 1920 in Lower Cambrian strata of Siberia [in Russian]. Doklady Akademii Nauk SSSR 104: 474–477.
Zhuravleva, I.T. [Žuravleva, I.T.], Konyushkov, K.N. [Konûškov, K.N], and Rozanov, A.Y. [Rozanov, A.Û.] 1964. Arheociaty Sibiri. Dvustennye arheociaty. 132 pp. Nauka, Moskva.
Zinchenko, V.N. [Zinčenko, V.N.] 1985. Local stratigraphic subdivisions of the Cambrian of northeastern Siberian Platform [in Russian]. In: M.L. Kokoulin and V.A. Rudavskaâ (eds.), Stratigrafiâ pozdnego dokembriâ i rannego paleozoâ Sibirskoj platformy. Sbornik naučnyh trudov, 15–22. VNIGRI, Leningrad.
Zinchenko, V.N. [Zinčenko, V.N.], Vasiljeva, N.I., and Rudavskaya, V.A. [Rudavskaâ, V.A.] 1993. Basal Cambrian strata of the north-eastern margin of the Siberian Platform [in Russian]. In: F.A. Čirva and V.N. Zinčenko (eds.), Stratigrafiâ fanerozoâ neftegazonosnyh regionov Rossii, 4–18. VNIGRI, Sankt-Petersburg.
Zittel, K.A. 1879. Protozoa, Coelenterata, Echinodermata and Moluscoidea. Handbuch der Paläontologie, Band 1, Paläozoologie. 765 pp. Druck und Verlag von R. Oldenbourg, München.
Acta Palaeontol. Pol. 67 (2): 341–464, 2022
https://doi.org/10.4202/app.00930.2021