
New Late Cretaceous microvertebrate assemblage from the Campanian–Maastrichtian Williams Fork Formation, northwestern Colorado, USA, and its paleoenvironmental implications
NICKOLAS A. BRAND, ANDREW B. HECKERT, ISRAEL SANCHEZ, JOHN R. FOSTER, REBECCA K. HUNT-FOSTER, and JAELYN J. EBERLE
Brand, N.A., Heckert, A.B., Sanchez, I., Foster, J.R., Hunt-Foster, R.K., and Eberle, J.J. 2022. New Late Cretaceous microvertebrate assemblage from the Campanian–Maastrichtian Williams Fork Formation, northwestern Colorado, USA, and its paleoenvironmental implications. Acta Palaeontologica Polonica 67 (3): 579–600.
We describe a microvertebrate assemblage from the J&M site, of the Upper Cretaceous (Campanian–Maastrichtian) Williams Fork Formation. Breakdown of fossil bearing matrix was achieved with the use of heated dimethyl sulfoxide. Nine of the recovered taxa are new to both the J&M site and the Williams Fork Formation. The sharks Lonchidion griffisi, Chiloscyllium sp., and Cantioscyllium markaguntensis are the first non-batoid elasmobranchs reported from the Williams Fork Formation and are all represented by teeth. The rays Cristomylus and Psuedomyledaphus are also newly reported from teeth. The most common identifiable fossils were teeth of indeterminate amiids, most likely belonging to Melvius. Osteichthyan fossils new to the Williams Fork Formation include teeth of Paralbula, an indeterminate pycnodontid tooth plate fragment, and an indeterminate lungfish tooth fragment. A tooth of the teiid Peneteius is also the first reported from within the Williams Fork Formation. Alligatoroid teeth are relatively common and are extremely similar to those of the contemporaneous durophage Brachychampsa but are generically indeterminate. Terrestrial taxa were recovered in much smaller numbers. Theropod dinosaur fossils included isolated tooth fragments belonging to an indeterminate dromaeosaurid and, possibly, to Richardoestesia. We recovered both multituberculate and metatherian fossils in the form of isolated teeth. Some of these taxa are known from marine and estuarine deposits and, given that so many of these marine associated taxa have been recovered together, it seems likely that the J&M site is recording marine or estuarine influence within at least part of its depositional history. The mammalian taxa suggest a Judithian–Lancian age for the site, while records of the squamate Peneteius and the ray Myledaphus, suggest that the J&M site may be temporally transitional between other late Campanian and late Maastrichtian-aged localities.
Key words: Chondrichthyes, Osteichthyes, Dinosauria, Lepidosauria, Mammalia, Euselachii, paleoenvironment, microvertebrate, fluvial, Judithian, Lancian, North America.
Nickolas A. Brand [nbrand2@gmu.edu], George Mason University, Department of Environmental Science and Policy, 4400 University Drive, Fairfax VA, 22030, USA; Department of Geological & Environmental Sciences, ASU Box 32067, Appalachian State University, Boone, North Carolina, 28608-2067, USA.
Andrew B. Heckert [heckertab@appstate.edu] and Israel Sanchez [sanchezi@appstate.edu], Department of Geological & Environmental Sciences, ASU Box 32067, Appalachian State University, Boone, North Carolina, 28608-2067, USA.
John R. Foster [johnfoster@utah.gov], Utah Field House of Natural History State Park Museum, 496 East Main Street, Vernal, Utah 84078, USA.
Rebecca K. Hunt-Foster [Rebecca_Hunt-Foster@nps.gov], Dinosaur National Monument, P.O. Box 128, Jensen, Utah, 84035, USA.
Jaelyn J. Eberle [Jaelyn.eberle@colorado.edu], University of Colorado Museum of Natural History and Department of Geological Sciences, University of Colorado, Boulder, Colorado, 80309, USA.
Received 12 August 2021, accepted 12 December 2021, available online 21 July 2022.
Copyright © 2022 N.A. Brand et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
The Williams Fork Formation is an Upper Cretaceous stratigraphic unit in the Mesa Verde Group that crops out in northwestern Colorado, northeastern Utah, and southern Wyoming that is regionally important for coal and natural gas production (e.g., Hettinger and Kirschbaum 2002), but is relatively understudied paleontologically. These strata are generally considered late Campanian (74–70 Ma) in age, (Foster and Hunt-Foster 2015). The Mesa Verde Group encompasses a variety of facies, including fluvial, swamp, beach, floodplain, lagoon, tidal channel, and shallow marine deposits (Hettinger and Kirschbaum 2002). We collected from the J&M site of the Williams Fork Formation, which is located just outside of the town of Rangely, in northwestern Colorado (Fig. 1). Traditionally the J&M site has been interpreted as a channel sandstone (Foster and Hunt-Foster 2015), and thus has been suspected to represent a period of freshwater fluvial deposition.
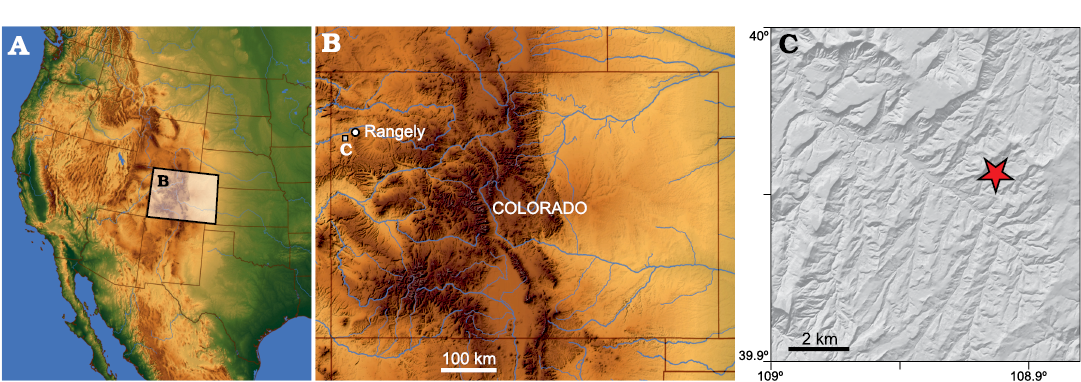
Fig. 1. Index maps showing the location of the J&M site in northwestern Colorado. A. Shaded relief map of the western United States highlighting Colorado. B. The position of the study area in northwestern Colorado. C. Digital elevation model of the study area, star denotes location of the J&M site. Precise locality data on file at MWC (see text).
Unlike many broadly contemporaneous nonmarine units, there is relatively little known about the vertebrate record of the Williams Fork Formation. J. David Archibald (San Diego State University, USA) led multiple collecting efforts to the Williams Fork Formation in the mid- to late 1980s with one of us (JRF) as a crew member in 1989. The Archibald-led crews collected a diverse assemblage of vertebrates through both traditional macrovertebrate collecting techniques and screening a number of ant hills, publishing a largely unillustrated faunal list (Archibald 1987). This paper, a master’s thesis by one of Archibald’s students (Diem 1999), and other published studies focused on the mammal (Archibald 1987; Lillegraven 1987) and dinosaur fossils from the Williams Fork Formation, which included a chasmosaurine skull (Diem and Archibald 2005), an indeterminate hadrosaur, and multiple theropod teeth (Hunt-Foster and Foster 2013; Foster and Hunt-Foster 2015). Identification of the chasmosaurine as species of Pentaceratops (Diem and Archibald 2000; Lucas et al. 2006) led Sullivan and Lucas (2006, see also Fowler 2017) to identify the Williams Fork Formation as Kirtlandian (late Campanian) in age.
To date, most descriptions of the Williams Fork Formation vertebrate assemblage, particularly the aquatic taxa, exist only in the form of numerous unpublished reports, mostly to the U.S. Bureau of Land Management, abstracts, and a thesis (e.g, Diem 1999; Diem and Archibald 2000; Hunt-Foster and Foster 2013; Vaughn et al. 2015; Brand et al. 2017), or largely unillustrated reports (Archibald 1987). Aquatic taxa recovered include species of the ray Myledaphus, the gar Lepisosteus, and the amiid Melvius (Foster and Hunt-Foster 2015). Reptiles are present, with testudines (turtles), lizards, and crocodyliform remains being especially abundant. Recently, Foster and Hunt-Foster (2015) described evidence of a giant crocodilian from the J&M site. This fossil assemblage suggests a mixture of terrestrial and freshwater environments, and supports previous preliminary reports of high concentrations of freshwater fossils within the Williams Fork Formation (Hunt-Foster and Foster 2013). A compiled vertebrate faunal list of both previously reported taxa and those reported herein from the Williams Fork Formation is provided in Appendix 1.
Institutional abbreviations.—MWC, Museum of Western Colorado, Fruita, Colorado, USA.
Geological setting
Although often ascribed to the Mesa Verde Group, the Williams Fork Formation is actually younger than the type Mesa Verde Group in southwestern Colorado (e.g., Hettinger and Kirschbaum 2002). In northwestern Colorado, northeastern Utah, and Wyoming, the Mesa Verde Group consists of, in ascending order, the Sego, Iles, and Williams Fork formations. In western Colorado the Williams Fork Formation is underlain by the Iles Formation and overlain by the Lewis Shale south of the study area, but is unconformably overlain by Paleogene strata further northward (Hettinger and Kirschbaum 2002). Although more than 260 m of Williams Formation sediments are exposed in the outcrop belt west of Rangely, Colorado, the base of the unit is not exposed and the top is truncated unconformably by Cenozoic strata in this region (Noll 1988; Raynolds and Hagadorn 2016) (Fig. 2B). In northwestern Colorado Hettinger and Kirschbaum (2002: pl. 2) considered the underlying Iles Formation to be equivalent to the lower Neslen Formation and the Williams Fork Formation stratigraphically equivalent to the upper Neslen, Farrer, and Tuscher formations, which encompass the Didymoceras nebrascaense and Didymoceras stevonsoni ammonite zones of the lowermost upper Campanian (Kirschbaum and Hettinger 2004).
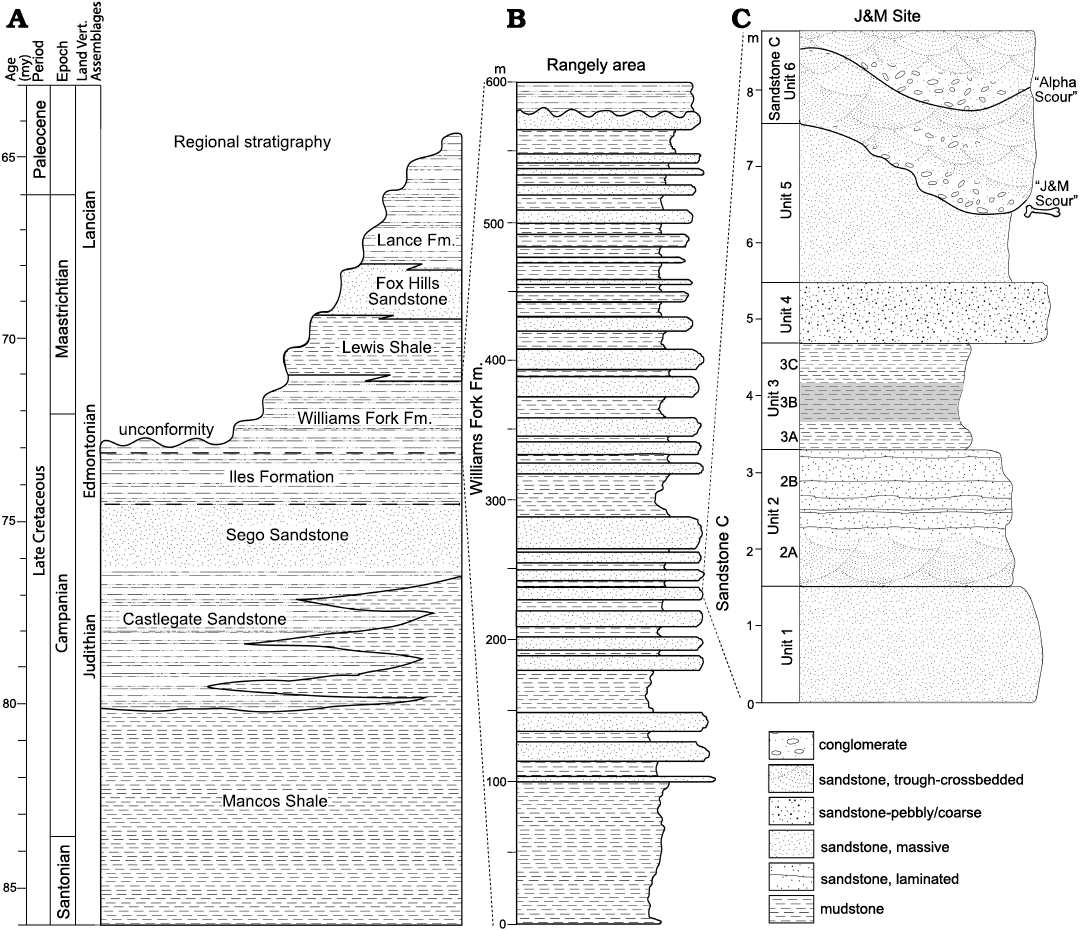
Fig. 2. Stratigraphy of the J&M site within the Williams Fork Formation. A. Regional Upper Cretaceous stratigraphy. B. Generalized stratigraphic section of the Williams Fork Formation west of Rangely (after Noll 1988). C. Stratigraphic section of Williams Fork Formation strata exposed at the J&M site, showing the stratigraphic position of the locality in Noll’s (1988) “sandstone C”. Width of stratigraphic units mimics the local weathering profile. Shading on unit 3B indicates a darker color than mudstones A and C. A detailed description of the J&M site stratigraphy is provided in the SOM: table S1 (Supplementary Online Material available at http://app.pan.pl/SOM/app67-Brand_etal_SOM.pdf).
An unpublished Master’s thesis by Noll (1988) provides the highest stratigraphic resolution for the fossils described here. Noll (1988) studied the sedimentology and stratigraphy of the Williams Fork Formation in detail, designating multiple sandstone bodies (A–H in ascending stratigraphic order, Noll 1988: fig. 10), and reported a regional dip for these sandstones of 4° to the northwest. We ascribe the J&M site to his “sandstone C”, also informally known as the “Carrotmen” sandstone because it hosts the archaeological site by the same name (Noll 1988). This is the thickest of his sandstone bodies, locally divided into a lower and upper division; the J&M site is located high in the lower division while several other locations in this area are stratigraphically lower. Noll (1988) reported paleocurrents based on trough crossbeds and ripples demonstrating paleoflow to the southeast in the lower portion of sandstone C, but a change to northerly paleocurrents in the upper part of sandstone C. Generally speaking, Noll (1988) recognized only nonmarine depositional facies for the Williams Fork Formation, including channel deposits (such as sandstone C), lateral accretion, downstream accretion, sandy bedforms, and fines from overbank deposits, designated by his facies CH, LA, SB, and OF, respectively. Noll (1988) interpreted these as representing, broadly, channel, levee, floodplain, and back swamp environments. Importantly, all of the finer-grained, non-channel facies he reports appear to represent nonmarine deposits, as indicated by abundant plant debris, including root casts and localized lignitic beds as well as nonmarine invertebrates.
Noll (1988) measured dozens of stratigraphic sections in this area, but the sections presented in his thesis figures are almost entirely composites. Thus, while his measured sections 7 and 159 bracket the J&M site to the north and south, respectively, we present our own measured section here (Fig. 2C). All of the vertebrates from the J&M site were recovered from sandstone and conglomerate in the basal 0.5 m of our unit 6 (Fig. 2C). This lenticular deposit is as much as 2.4 m thick of fining-upwards, cliff-forming conglomerate and sandstone that fills scours in the underlying fine-grained sandstone of unit 5. The “J&M scour” appears to represent an initial pulse of sedimentation, and is itself truncated by larger scours and multiple-meter-thick crossbedded sheets of the “main body” of Noll’s (1988) sandstone C. The underlying strata (units 1–5 in our measured section) match the interlayered siltstone and mudstone (with minor interbedded sandstone) that Noll (1988) interpreted as levee and floodplain deposits.
Material and methods
We collected material from the J&M site by breaking large (≥1 kg) blocks of sandstone from the cliff face using sledge hammers and chisels to collect pieces that were small enough to transfer for further processing. To the extent possible, the resulting fragments, sand and dust were collected along with the bulk rock with the aim of recovering microvertebrate fossils. The total sample weighed approximately 30 kg.
We attempted multiple methods to disaggregate the bulk rock samples and concentrate the sand. The bulk rock consisted of small (10 mm) to large (≥100 mm) rock fragments of two consistencies. Samples that were obviously laminated and fissile were more susceptible to disaggregation. Other samples were more strongly cemented and were not easily broken down even when worked with a rock hammer.
Traditional water-based screenwashing was employed first, but this proved ineffective after a week of soaking (both water and a water/detergent mix were used). A large sample of the more indurated rock that was left soaking in water for months did eventually break down, although it did not contain identifiable microfossils after this material was concentrated. We next experimented with freeze and thaw methods. A typical freeze-thaw cycle consisted of placing multiple small to medium sized rock samples into plastic containers and filling the containers with water before putting them into a freezer overnight. The next morning, the containers would be removed and allowed to thaw before repeating the freezing phase. Even after five of these freeze and thaw cycles we did not see significant breakdown, and vacuuming the air out of the pores during the soak phase (using a vacuum pump over the soaking container) before freezing did not speed up the process.
The only real success was the result of using “pharma grade” (99.995%) dimethyl sulfoxide (DMSO) to aid in disaggregation following Triplehorn (2002), which resulted in breakdown of the fissile material, but not the non-fissile rocks. Room temperature tests yielded breakdown after four to five days, while heating the DMSO to 50°C allowed breakdown of samples in just two days. We describe our exact methodology in the following paragraph.
We filled 12 oz (~350 ml) glass jars halfway with fissile sediments and then poured DMSO over the sediment until it was 90% covered. We then capped the jars and placed them on a hotplate set to 50° C in a fume hood. After two days the rock would be broken down and we filtered the DMSO, pouring the DMSO/sediment mixture into a glass funnel with a coffee filter placed inside. The DMSO would filter out slowly, and could be reused to conserve material. We could then soak up to four jars worth of post-DMSO material in large, five gallon (~20 l) buckets. Using paint sieves suspended from the top of the containers allowed us to sieve the fossils and let sediment pass through, following Avrahami et al. (2015). After one day, the water was changed to ensure that no DMSO remained and allowed to rinse for another 24 hours. The sediment was then dried and run through a set of nested sieves (4, 2, 1, and 0.5 mm sieves) to obtain a microfossil matrix concentrate. Relative abundance of reported taxa within different size classes are reported in Table 2. The senior author (NAB) and various student volunteers then picked this concentrate using binocular microscopes.
Table 2. Occurrence of vertebrate taxa in macro- and screenwash samples from the J&M site. Records of all taxa recovered from the J&M site of the Williams Fork Formation. Macrovertebrate (MV) category indicates presence of taxa in found during typical quarrying operations, preparation, or in bulk material sampled for screenwashing. 2–4 mm and 1–2 mm categories represent the presence of taxa in screenwash material of that size. Taxa in bold are new records for the Williams Fork Formation. C, common; U, uncommon; –, absent. Taxa preceded by an * are previously reported, but illustrated here for the first time.
|
Taxon |
MV |
2–4 mm |
1–2 mm |
|
Lonchidion griffisi |
U |
C |
U |
|
Lonchidiidae gen. et sp. indet. |
U |
– |
– |
|
Cantioscyllium markaguntensis |
– |
U |
– |
|
cf. Chiloscyllium sp. |
– |
C |
U |
|
*Myledaphus bipartitus |
C |
C |
– |
|
Cristomylus sp. |
– |
U |
– |
|
Pseudomyledaphus madseni |
C |
C |
– |
|
*cf. Melvius sp. |
C |
C |
– |
|
Pycnodontiformes gen. et sp. indet. |
U |
U |
– |
|
Paralbula casei |
– |
C |
– |
|
Actinopterygii gen. et sp. indet. |
U |
U |
– |
|
*Lepisosteidae gen. et sp. indet. |
– |
U |
– |
|
Dipnoi gen. et sp. indet. |
– |
U |
– |
|
*Amphibia gen. et sp. indet. |
– |
U |
– |
|
Peneteius sp. |
– |
U |
– |
|
*cf. Brachychampsa sp. |
C |
C |
– |
|
Dromaeosauridae gen. et sp. indet. |
– |
U |
– |
|
*cf. Richardoestesia |
– |
U |
– |
|
Hadrosauridae gen. et sp. indet. |
C |
– |
– |
|
Meniscoessus sp. |
– |
U |
– |
|
cf. Cimolodon nitidus |
– |
U |
– |
|
Multituberculata gen. et sp. indet. |
U |
C |
– |
|
?Leptalestes cooki |
– |
U |
– |
|
Amia sp. |
U |
– |
– |
Illustrations of microvertebrate fossils were made with a Keyence VHX-600 microscope. Fossils were photographed at 50–200 × magnification, based on the specimen size. 3D scan data was collected using the proprietary Keyence software’s automated digital image stacking process for future analysis. These scans and photomicrographs often made use of both glare-reduction and high dynamic range mode options found in the image capture software. We saved images as .tiff files in order to preserve maximum image quality. The images were saved with scale bars calculated digitally by the Keyence software. The resulting images were manipulated in Adobe Photoshop to create Figs. 3–8. The backgrounds around the fossils were cropped out, and we created a new, floating scale bar for each photograph based on the original Keyence provided scale. The individual specimen photographs were compiled in Photoshop.
All specimens were collected under permits issued to the Museum of Western Colorado (MWC) by the US Bureau of Land Management (BLM). The fossils are reposited at the Museum of Western Colorado’s Dinosaur Journey facility in Fruita, Colorado, USA, where detailed locality information is available upon request.
Systematic palaeontology
Elasmobranchii Bonaparte, 1838
Euselachi Hay, 1902
Hybodontoidea Owen, 1846
Lonchidiidae Herman, 1977
Genus Lonchidion Estes, 1964
Type species: Lonchidion selachos Estes, 1964; Lance Formation, Upper Cretaceous.
Lonchidion griffisi Case, 1987
Fig. 3A–D.
Material.—MWC 8851, numerous tooth and teeth fragments; MWC 9580, tooth fragment; MWC 9581, tooth fragment; MWC 9582, tooth fragments; MWC 9583, tooth fragments; MWC 9584, tooth; MWC 9585, tooth fragment; MWC 9967, 13 tooth fragments; all from the Campanian–Maastrichtian, Williams Fork Formation, J&M site, northwest Colorado, USA.
Emended diagnosis.—Lonchidion species with multiple (three or more) well defined “lateral cusplets” located mesio-distally, to each side, of the principal cusp. Also possesses between two and five serrations on the labial peg of the crown. Teeth may be almost perfectly straight, or may exhibit strong curvature (modified from Case 1987).
Description.—We assign chondrichthyan teeth that are gracile, narrow labiolingually and elongated laterally to Lonchidion griffisi. The crown is prominent and low, and the occlusal surface forms a crest that has well defined individual cusplets. The lateral margins of the crown are pointed and may resemble the lateral cusps.

Fig. 3. Representative non-batoid chondrichthyans from the J&M site, Colorado, USA, Williams Fork Formation, Campanian–Maastrichtian, Upper Cretaceous. A–D. Lonchidion griffisi Case, 1987. A. MWC 8851, tooth crown in labial (A1) and occlusal (A2) views. B. MWC 9580, fragmentary tooth crown in labial (B1) and occlusal (B2) views. C. MWC 9581, fragmentary tooth crown in lingual (C1), labial (C2), and occlusal (C3) views. D. MWC 9582, fragmentary tooth crown in labial (D1) and occlusal (D2) views. E, F. Cantioscyllium markaguntensis Kirkland, Eaton, and Brinkman, 2013. E. MWC 8866, tooth crown in labial (E1) and occlusal (E2) views. F. MWC 9586, tooth crown in labial (F1) and lingual (F2) views. G, H. cf. Chilloscyllium sp. G. MWC 8850, morph 1 tooth crown in mesial (G1) and labial (G2) views. H. MWC 9576, morph 2 tooth crown in mesial (H1) and labial (H2) views. I. Hybodontoidea gen. et sp. indet., MWC 9800, dorsal fin spine in lateral view.
Remarks.—We have recovered mostly crown fragments, with little to no root material remaining, totalling 19 different tooth crowns and fragments. Two of the fragments fit together to form a single tooth. Lonchidion species are a ubiquitous component of Campanian fresh and estuarine fossil assemblages in North America (Kirkland et al. 2013). Lonchidion griffisi was originally described as Lissodus griffisi by Case (1987), however, revisions by Rees and Underwood (2002) reassigned many species of “Lissodus’’ to Lonchidion, including Lissodus griffisi, an assignment followed by Fischer (2008). Material of L. griffisi is known only from the Mesaverde Formation (Case 1987; Fischer 2008). The original material was recovered from the Teapot Sandstone Member of the Mesaverde Formation, and the deposits were determined to be of estuarine nature due to the presence of both marine and nonmarine fish fossils (Case 1987).
Stratigraphic and geographic range.—Lonchidion griffisi was previously known only from the Late Campanian Teapot Sandstone Member of the Mesaverde Formation in Wyoming.
Lonchidiidae gen. et sp. indet.
Fig. 3I.
Material.—MWC 9800, dorsal fin spine; from the Campanian–Maastrichtian, Williams Fork Formation, J&M site, northwest Colorado, USA.
Description.—We interpret a short segment of bone with small, offset denticles (MWC 9800, Fig. 3I) as a fine spine fragment. The hybodont fin spine we illustrate here was found while preparing larger fossils. Best exposed in lateral view, it preserves nine posteriorly recurved denticles that are offset from each other in an alternating pattern, but do not comprise two rows. The lateral side of the spine bears many relatively fine ridges.
Remarks.—Lonchidiid fin spines are striated and possess small, hooked denticles (e.g., Duffin 1985: fig. 25). The single row of alternating denticles and relatively smooth lateral surface are features that distinguish MWC 9800 from fin spines of most other hybodonts, including Hybodus, which typically have at least two distinct rows of parallel denticles and strongly developed lateral ridges (e.g., Kirkland et al. 2013; fig. 9.4a–e) and Lissodus, which bear alternate denticles but retain the strongly developed lateral ridges (Kirkland et al. 2013). The specimen we illustrate here well-matches fin spine fragments assigned to Lonchidion by Kirkland et al. (2013: fig. 9.5j–m).
Elasmobranchii Bonaparte, 1838
Neoselachii Compagno, 1977
Galeomorphii Compagno, 1973
Orectolobiformes Applegate, 1972
Ginglymostomatidae Gill, 1862
Genus Cantioscyllium Woodward, 1889
Type species: Cantioscyllium decipens Woodward, 1889; Lower Chalk, Upper Cretaceous.
Cantioscyllium markaguntensis Kirkland, Eaton, and Brinkman, 2013
Fig. 3E, F.
Material.—MWC 8866, tooth, MWC 9586, tooth; from the Campanian–Maastrichtian, Williams Fork Formation, J&M site, northwest Colorado, USA.
Description.—Small Cantioscyllium species differentiated by the presence of two or more lateral cusplets on either side of the crown, and by very well-defined enamel folding along the labial surface (after Kirkland et al. 2013).
Remarks.—We assign small chondrichthyan teeth with a lingually inclined broad central cusp, that bear clearly distinct lateral cusps at the margins of the crown, and possess a roughly heart shaped root when viewed basally to Cantioscyllium. Because there are two or more cusplets on either side of the crown and the labial surface of the crown is heavily textured we assign these teeth to C. markaguntensis. We have collected two teeth representative of Cantioscyllium. Cantioscyllium markaguntensis is differentiated from all other known freshwater orectolobiformes collected from Utah by the strong labial surface ornamentation (Kirkland et al. 2013), and has previously been collected only from the Straight Cliffs Formation of the Markagunt Plateau in Cedar Canyon, Utah, so this is the first report of it from the Williams Fork Formation. It is possibly the oldest known freshwater orectolobiform shark from southern Utah (Kirkland et al. 2013).
Hemiscyllidae Gill, 1862
Genus Chiloscyllium Müller and Henle, 1838
Type species: Chiloscyllium greeni Cappetta, 1973; Carlile Shale, Upper Cretaceous.
cf. Chiloscyllium sp.
Fig. 3G, H.
Material.—MWC 8850, tooth with root. MWC 9576, tooth with root. MWC 9968, 17 teeth, many with roots; from the Campanian–Maastrichtian, Williams Fork Formation, J&M site, northwest Colorado, USA.
Description.—Small (less than 2 mm) chondrichthyan teeth that are shortened apico-basally. These teeth lack lateral cusplets on the crown and may be smooth or bear weak enamel folding along the labial surface (after Kirkland et al. 2013).
Remarks.—We assign chondrichthyan teeth that are small and possess strongly triangular crowns, lacking lateral cusplets, and a weakly textured or untextured labial surface to cf. Chiloscyllium sp. A centrally oriented, well-defined ridge may be present on the labial surface of the tooth from the base of the crown to the tip. We have collected 17 specimens that we attribute to this genus. The teeth we describe are most similar to Chiloscyllium missouriense Case, 1979, because they lack lateral cusplets and most have a smooth labial surface of the crown, though Kirkland et al. (2013) note that the specimens with the central ridge may belong to a new species or even genus. We report two morphs of Chiloscyllium teeth. Morph one tapers aggressively from the tip of the crown to the base, and tends to contain a raised ridge running along the center of the labial surface of the crown (Fig. 3G). Morph 2 tapers much more conservatively, to the point of being almost conical in crown shape. The lowermost portion of the labial surface of the crown extends basally and labially to further cover the root, resulting in a noticeable “drop” in the crown (Fig. 3H). While these morphs may represent different species, or simply be teeth in different positions in the jaw, such a question is beyond the scope of this paper and will necessitate further, focused study. Because the roots of many specimens were either worn or encrusted with sediment, species level identifications cannot be sufficiently demonstrated. The genus Chiloscyllium contains extant species (Cook et al. 2014), although fossil species may be in need of revision (Kirkland et al. 2013). Chiloscyllium missouriense has been recovered from the Teapot Sandstone of the “Mesaverde Formation’’ in Wyoming (Case 1987), and similar teeth have been reported from the broadly coeval St. Mary’s River Formation of Alberta, Canada (Kirkland et al. 2013). This is the first report of Chiloscyllium from the Williams Fork Formation.
Batomorphii Cappetta, 1980
Rajiformes Berg, 1940
Rhinobatoidea Müller and Henle, 1837
Genus Myledaphus Cope, 1876
Type species: Myledaphus bipartitus Cope, 1876; Fort Union Formation, Upper Cretaceous.
Myledaphus bipartitus Cope, 1876
Fig. 4A, B.
Material.—MWC 8887, two teeth; MWC 9591, tooth; from Campanian–Maastrichtian, Williams Fork Formation, J&M site, northwest Colorado, USA.
Description.—Mid-to-high crowned durophagous teeth. A low, transverse ridge divides the oral surface of the crown through the center. Labiolingually oriented enamel folds are present on the oral surface, and continue as ridges onto the vertical faces of the crown (Kirkland et al. 2013).
Remarks.—We assign batoid teeth that have a roughly hexagonal to rhomboidal shape, with both the occlusal and lateral surfaces of the crown textured, and a transverse ridge that may divide the occlusal surface of the crown to Myledaphus bipartitus. Occlusal texturing is asymmetrical across the transverse ridge. The root is often taller than the crown and may taper inwards basally. The teeth vary in crown width 1–4 mm, and combined crown plus root height of 1.5–3 mm. We have collected 19 specimens of Myledaphus bipartitus. Campanian (M. bipartitus) and Maastrichtian (M. pustulosis) species of Myledaphus bear different enamel ornamentation patterns (Kirkland et al. 2013), thus making them potentially useful biostratigraphically. The specimens from the J&M site thus far all appear to pertain to M. bipartitus rather than M. pustulosis, suggesting a Campanian age. Some specimens appear to have ridges that are more broken up, and resemble somewhat the pustules of M. pustulosis, although this patterning is not as pronounced as that of the types of M. pustulosis. Myledaphus material is particularly abundant in the late Campanian-aged rocks, when the genus appears to have originated and begun to replace the rays Pseudomyledaphus and Cristomylus (Kirkland et al. 2013).
Stratigraphic and geographic range.—The uppermost Cretaceous of New Mexico, Colorado, Utah, Wyoming, South Dakota North Dakota and Montana in the Unites States, Alberta and Saskatchewan in Canada, and Mexico.
Genus Cristomylus Kirkland, Eaton, and Brinkman, 2013
Type species: Cristomylus nelsoni Kirkland, Eaton, and Brinkman, 2013; Mussentuchit Member of the Cedar Mountain Formation, Upper Cretaceous.
Cristomylus sp.
Fig. 4C, D.
Material.—MWC 8886, tooth, MWC 9588, tooth; from the Campanian–Maastrichtian, Williams Fork Formation, J&M site, northwest Colorado, USA.
Description.—Flattened teeth with rounded lateral surfaces of the crown. A transverse crest is present that divides the crown into equal halves in occlusal view (Kirkland et al. 2013). These specimens are smaller than the teeth of species of Myledaphus and Psuedomyledaphus, and contain a pair of lateral foramina on the labial surface of the root (Kirkland et al. 2013).

Fig. 4. Representative batoid chondrichthyan teeth from the J&M site, Colorado, USA, Williams Fork Formation, Upper Cretaceous. A, B. Myledaphus bipartitus Cope, 1876. A. MWC 8887, tooth crown and root in mesial (A1), labial (A2), and occlusal (A3) views. B. MWC 9591, tooth crown and root in mesial (B1), labial (B2), and occlusal (B3) views. C, D. Pseudomyledaphus sp. C. MWC 9589, tooth crown and root in mesial (C1) and occlusal (C2) views. D. MWC 9590, tooth crown and root in mesial (D1) and occlusal (D2) views. E, F. Cristomylus sp. E. MWC 8886, tooth crown in mesial (E1) and occlusal (E2) views. F. MWC 9588, tooth crown in mesial (F1) and occlusal (F2) views.
Remarks.—We assign batoid teeth with a transverse crest but that are otherwise characterized by heavily rounded lateral faces of the crown to Cristomylus. The teeth retain a slightly hexagonal shape and the root surfaces are heavily weathered. The transverse crest may be straight or follow the curvature of the tooth, if the shape of the crown is not perfectly linear. The root is always taller than the crown in our specimens. We have collected only three specimens of Cristomylus, and the genus is previously unreported from the upper Campanian, as it is otherwise only known from older rocks (Kirkland et al. 2013). While we do not have enough specimens to provide a species level identification, it is of interest that the J&M site bears specimens of species of both Myldepahus and of Cristomylus, as Myledaphus appears to have replaced Cristomylus in many late Campanian ecosystems (Kirkland et al. 2013).
Genus Pseudomyledaphus Kirkland, Eaton, and Brinkman, 2013
Type species: Pseudomyledaphus madseni Kirkland, Eaton, and Brinkman, 2013; John Henry Member of the Straight Cliffs Formation, Upper Cretaceous.
Pseudomyledaphus madseni Kirkland, Eaton, and Brinkman, 2013
Fig. 4E, F.
Material.—MWC 8868, tooth. MWC 9589, tooth. MWC 9590, tooth. MWC 9966, 17 teeth, many with roots; from the Campanian–Maastrichtian, Williams Fork Formation, J&M site, northwest Colorado, USA.
Description.—Large, flattened teeth similar to these of species of Myledaphus, other than that they lack the characteristic enamel folds of Myledaphus, except on the mesial lingual face of the crown (after Kirkland et al. 2013)
Remarks.—We assign batoid teeth similar to those of Myledaphus, but that lack the enameloid texturing on the lateral surfaces of the crown, to Pseudomyledaphus madseni. These teeth are hexagonal to rhomboidal in shape, and lack any enamel texturing on the occlusal surface of the crown. The lateral surfaces of the crown are straight and do not curve as dramatically as those seen in Cristomylus. Pseudomyledaphus is the most common chondrichthyan we recovered from the J&M site, with a total of 45 individual teeth collected. The teeth vary in width 1.5–6 mm, and height 1.5–4 mm. Pseudomyledaphus is previously unknown from the upper Campanian (Kirkland et al. 2013). Like Crystomylus, it is believed that this genus was replaced by Myledaphus during the late Campanian (Kirkland et al. 2013), making its co-occurrence with Myledaphus in the J&M site of interest.
Stratigraphic and geographic range.—Conician–Campanian of Utah, United States and Alberta, Canada.
Osteichthyes Huxley, 1880
Actinopterygii Klein, 1885
Amiidae Bonaparte, 1838
Vidalamiinae Grande and Bemis, 1998
Vidalamiini Grande and Bemis, 1998
Genus Melvius Bryant, 1987
Type species: Melvius thomasi Bryant, 1987; Hell Creek Formation, Upper Cretaceous.
cf. Melvius sp.
Fig. 5B.
Material.—MWC 8867, tooth; MWC 8888, numerous teeth (more than 100); from the Campanian–Maastrichtian, Williams Fork Formation, J&M site, northwest Colorado, USA.
Description.—Tall, lanceolate osteichthyan crowns with well-defined carinae running from crown tip to base (Bryant 1987).
Remarks.—We assign teeth that are tall and lanceolate with oval bases and well-defined carinae present from the base of the crown to the tip to cf. Melvius sp. These teeth may be opaque to nearly translucent, and are widest at the base and get progressively narrower toward the tip. Sizes vary from 3–7 mm in height and 2–5 mm in width. The teeth are generally comparable to the crowns of both Maastrichtian Melvius thomasi Bryant, 1987, and Campanian Melvius chauliodous Grande and Bemis, 1998, and highly carinate tooth tips are a defining character of the Vidalamiinae and are not restricted to Melvius (Bryant 1987; Grande and Bemis 1998; Sullivan et al. 2011). While diagnostic characters of Melvius are all in the vertebrae and skull (Grande and Bemis 1998), Melvius is the only vidalamiine currently known from the Upper Cretaceous of North America (Grande and Bemis 1998), so we tentatively refer the isolated teeth to cf. Melvius sp. here. These amiid teeth are the most common identifiable microvertebrate fossil at the J&M site, where we have collected over one hundred of them. Originally described from Hell Creek Formation channel deposits (Bryant 1987), Melvius fossils are often found in close proximity to the Western Interior Seaway margin, thus suggesting that they may have been capable of inhabiting brackish or marine waters (Bryant 1987). Other than an abstract by Hunt-Foster and Foster (2013) that mentions a large, diagnostic Melvius centrum from a different locality, this is the first report of cf. Melvius sp. from the Williams Fork Formation.
Pycnodontiformes Berg, 1937
Pycnodontiformes gen. et sp. indet.
Fig. 5C.
Material.—MWC 8873, tooth plate fragment; from the Campanian–Maastrichtian, Williams Fork Formation, J&M site, northwest Colorado, USA.
Description.—Small, rounded and flattened teeth that may be found isolated or packed into tooth plates. Teeth are rounded and arranged in rows that are attached directly to the bone (Kriwet 2005).

Fig. 5. Representative osteichthyan teeth from the J&M site, Colorado, USA, Williams Fork Formation, Campanian–Maastrichtian, Upper Cretaceous. A, E. Paralbula casei Estes, 1969b. A. MWC 8855, tooth crown in occlusal (A1), basal (A2), and mesial (A3) views. E. MWC 8856, tooth crown in occlusal view. B. cf. Melvius sp., MWC 8867, tooth crown in labial view. C. Pycnodontiformes gen. et sp. indet., MWC 8873, tooth plate fragment in occlusal (C) view. D. Actinopterygii indet., MWC 8867, tooth crown in lateral (D) view. F. Dipnoi indet., MWC 8885, toothplate fragment in lateral view.
Remarks.—We assign one tooth plate and numerous isolated teeth as pertaining to the Pycnodontiformes. The plate is small, with individual teeth being 0.5–1 mm in diameter, and the whole tooth plate fragment is only 3 mm across at its widest point. We distinguish this specimen from species of Paralbula by the more elongate and curved shape of the teeth present in these specimens (Estes 1969b; Sahni 1972). The presence of patterning on the teeth suggests that they may have belonged to a juvenile animal, and did not have time to wear down to a perfectly smooth surface (Kriwet 2005).
Phyllodontidae Sauvage, 1875
Genus Paralbula Blake, 1940
Type species: Paralbula marylandica Blake, 1940; Piscataway Member of the Aquia Formation, Lower Eocene.
Paralbula casei Estes, 1969b
Fig. 5A, E.
Material.—MWC 8855, tooth; MWC 8856, tooth; MWC 8890, numerous teeth; from the Campanian–Maastrichtian, Williams Fork Formation, J&M site, northwest Colorado, USA.
Description.—Small, rounded teeth with light enamel texture radiating from the center outwards on the occlusal surface of the crown. The root surface is well developed, with distinct radial banding (after Sahni 1972).
Remarks.—Small rounded osteichthyan teeth with a crushing morphology and a heavily developed root surface characterized by concentric rings are ascribed to Paralbula casei. The root surface is concave towards the crown, and is as wide as the base of the crown. The occlusal surface is textured in a radial pattern that extends basally from the center of the tooth. The teeth are all rounded, and vary from 1.5–4 mm in diameter. The crowns vary from textured to smooth on the occlusal surface, which we attribute to differing levels of tooth wear. Paralbula casei has previously been reported from the Judith River Formation, where it is possibly associated with the formation’s marine deposits (Sahni 1972). Estes (1969b) noted that the most well preserved phyllodontid specimens tended to come from near-shore marine and estuarine deposits, where it is hypothesized that they may have been most prevalent. Paralbula has not previously been reported from the Williams Fork Formation.
Stratigraphic and geographic range.—Upper Cretaceous of North America, Spain and Russia; Lower Eocene of England.
Actinopterygii gen. et sp. indet.
Fig. 5D.
Material.—MWC 8857, tooth; MWC 9593, 12 teeth; from the Campanian–Maastrichtian, Williams Fork Formation, J&M site, northwest Colorado, USA.
Description.—Slender and conical osteichthyan teeth in which the tip of the crown consists of smooth enamel and may be translucent, while the lower portion has enamel striations running vertically, a constriction may be present where the striated base meets the smooth tip (after Brinkman et al. 2017).
Remarks.—We have recovered 13 teeth that we assign to Actinopterygii indet. because they are slender and have a smooth, translucent tip of the crown but a striated base. Almost all of the teeth are fragmentary and are identifiable only because of the vertical striations present on the tooth crown. Such teeth are widely distributed through Actinopterygii and so we only identify them as Actinopterygii indet.
Holostei Müller, 1844
Lepisosteiformes Hay, 1929
Lepisosteidae Cuvier, 1825
Lepisosteidae gen. et sp. indet.
Material.—MWC 8223, scales; from the Campanian–Maastrichtian, Williams Fork Formation, J&M site, northwest Colorado, USA.
Description.—Lepisosteid scales are rhomboid-shaped, possess thick bony bases beneath the external ganoin layer, and retain a distinctive sheen when not significantly weathered.
Remarks.—The most common scales found at the J&M site are also relatively large (≥2 mm in maximum dimension), rhomboid scales with thick bony bases beneath the exterior ganoin layer that we assign to Lepisosteidae indet. Lepisosteid scales are a common component of Upper Cretaceous microvertebrate assemblages (Estes 1964; Grande 2010), and are widespread among estuarine deposits of the Late Cretaceous (Case and Schwimmer 1988).
Sarcopterygii Romer, 1955
Dipnoi Müller, 1844
Dipnoi gen. et sp. indet.
Fig. 5F.
Material.—MWC 8885, toothplate fragment; from the Campanian–Maastrichtian, Williams Fork Formation, J&M site, northwest Colorado, USA.
Description.—A robust tooth fragment with two pronounced ridges that project from the body of the tooth. Due to this tooth’s fragmentary nature, it is impossible to suggest the total number of tooth ridges or the size of the complete tooth.
Remarks.—Lungfish have a reduced dentition of four complex teeth, or toothplates, with each bearing elongate ridges that emanate from the medial margin of the tooth (Denison 1974). These cuspate ridges end in a pointed surface on the labial side of each toothplate, which are formed from varieties of dentine (Harrell and Ehret 2019). We recovered an incomplete toothplate belonging to a lungfish, although it is broken, and only preserves two ridges, which is not enough to allow a precise identification. The ridges are distinct enough however, to verify it as belonging to a lungfish. Dipnoans are relatively rare in the Cretaceous of Western Interior (Kirkland 1987), and have not been reported previously from the Williams Fork Formation.
Amphibia Linnaeus, 1758
Amphibia gen. et sp. indet.
Fig. 6D.
Material.—MWC 9969, jaw fragment with teeth; from Campanian–Maastrichtian, Williams Fork Formation, J&M site, northwest Colorado, USA.
Description.—Small jaw fragment bearing three smooth, pointed teeth which are darker towards the apex of the crown. A fourth tooth is broken proximally to the jaw, revealing a hollow interior. The jaw is relatively deep, with weak but visible texturing on the lateral surface.

Fig. 6. Representative reptile (A–C, E) and amphibian (D) teeth from the J&M site, Colorado, USA, Williams Fork Formation, Campanian–Maastrichtian, Upper Cretaceous. A–C. cf. Brachychampsa sp. A. MWC 9577, gracile morph tooth crown in lateral (A1) and occlusal (A2) views. B. MWC 9578, robust morph tooth crown in lateral (B1) and occlusal (B2) views. C. MWC 9579, robust morph tooth crown in lateral (C1) and occlusal (C2) views. D. Tetrapoda indet., MWC 8877, jaw fragment with teeth in lateral (D1) and occlusal (D2) views. E. Peneteius sp., MWC 8871, tooth crown with root in mesial (E1), lateral (E2), and occlusal (E3) views.
Remarks.—We recovered a small jaw fragment with external texture and the remains of four teeth (Fig. 6D). We tentatively consider this tooth to represent a lissamphibian because of the hollow nature of the tooth crowns (Fig. 6D2) and the external texturing. While a more specific identification would require more material, it is important to note that this specimen represents the first published lissamphibian material from the Williams Fork Formation.
Reptilia Laurenti, 1768
Squamata Oppel, 1811
Polyglyphanodontinae Estes, 1983
Polyglyphanodontini Nydam, Eaton, and Sankey, 2007
Genus Peneteius Estes, 1969a
Type species: Peneteius aquilonius Estes, 1969a; Hell Creek Formation, Upper Cretaceous.
Peneteius sp.
Fig. 6E.
Material.—MWC 8871, tooth; from the Campanian–Maastrichtian, Williams Fork Formation, J&M site, northwest Colorado, USA.
Description.—Squamate teeth superficially resembling those of mammals, with a tall crown tipped with up to six cusps. The two central cusps are the largest and form a “u” shaped ridge along the occlusal surface (after Nydam et al. 2000). Peneteius aquilonius is distinguished from Peneteius saueri McCord, 1998, by the former’s comparatively narrow shaft, unconstricted bases, and a transverse blade with a smaller angle (Nydam et al. 2007),
Remarks.—We assign
one squamate tooth with six cusps to a species of Peneteius.
The two central cusps are much larger than the surrounding accessory
cusps. Each primary cusp is paired with two accessory cusps. Both pairs
of accessory cusps, and the pair of primary cusps, form peaks on
otherwise concave ridges on the occlusal surface of the tooth. The tooth
root is thick, and flares strongly outwards from the crown. Based on its
morphology this specimen represents a maxillary tooth.
We are unable to assign a species level identification for this
specimen, as it displays a mixture of character states diagnostic of
both described Peneteius species. The tooth
has a relatively narrow shaft of the crown, with a non constricted base
that compares favorably with P. aquilonius (Estes 1969a). In contrast, the main cusps are less than
half the total height of the crown, the accessory cusps are angled
outwards relative to the main cusps, and the transverse blade is obtuse
in angle (110–120°), all of which are diagnostic of P.
saueri (Nydam et al. 2000). This combination
of characters may represent a new species of Peneteius,
or alternatively, an aberrant individual or worn tooth. A precise
identification will require the collection and description of more Peneteius teeth from the Williams Fork
Formation.
This is the first record of Peneteius from
the Williams Fork Formation. Peneteius has
previously been recovered from sites of both Campanian and Maastrichtian
age (Nydam et al. 2000, 2013),
although P. saueri is thus far only known
from Campanian aged deposits (Nydam et al. 2007, 2013).
Of note for this occurrence is that it is geographically north of the
reported Peneteius distribution during the
late Campanian (Nydam et al. 2013). This specimen
therefore represents a potentially significant range extension for the
genus assuming a late Campanian age for the J&M site, or
alternatively represents a post-Campanian occurrence.
Crocodylomorpha Hay, 1929
Crocodylia Gmelin, 1788
Eusuchia Huxley, 1875
Alligatoroidea Gray, 1844
Genus Brachychampsa Gilmore, 1911
Type species: Brachychampsa montana Gilmore, 1911; Hell Creek Formation, Upper Cretaceous.
cf. Brachychampsa sp.
Fig. 6A–C.
Material.—MWC 9577, tooth; MWC 9578, tooth; MWC 9579, tooth; MWC 9592, numerous teeth; from the Campanian–Maastrichtian, Williams Fork Formation, J&M site, northwest Colorado, USA.
Description.—Crocodyliform teeth of small to intermediate size. Gracile teeth are taller, more triangular, and less blunt, while robust teeth are proportionately lower, less conical, more blunt and are larger in both width and crown height. These teeth are very similar to those of Brachychampa (Williamson 1996).
Remarks.—We assign relatively low-crowned, blunt, striated alligatoroid teeth that vary from gracile and recurved to a robust morphotype we refer to cf. Brachychampsa sp. These teeth are relatively common at the J&M site and are similar to those of the species of alligatoroid Brachychampsa, although precise identification is rendered impossible by a lack of diagnostic skull material. We have collected a variety of morphotypes, with taller, triangular, gracile teeth, intermediate, more conical teeth, and lower blunt teeth. The latter of these teeth strongly suggest a durophagous diet.
Dinosauria Owen, 1842
Theropoda Marsh, 1881
Coelurosauria Huene, 1914
Dromaeosauridae Matthew and Brown, 1922
Dromaeosauridae gen. et sp. indet.
Fig. 7B.
Material.—MWC 8872, tooth; from the Campanian–Maastrichtian, Williams Fork Formation, J&M site, northwest Colorado, USA.
Description.—We assign one tooth to Dromaeosauridae indet. This specimen is, unfortunately, broken both towards the tip of the crown and towards the root but is laterally compressed and finely serrated. The tooth is recurved, has very fine denticles on both the mesial and distal surfaces of the crown, and compares favorably to the dromaeosauridae morphotype A as figured by Williamson and Brusatte (2014).

Fig. 7. Representative theropod dinosaur teeth from the J&M site, Colorado, USA, Williams Fork Formation, Campanian–Maastrichtian, Upper Cretaceous. A. cf. Richardoestesia sp., MWC 8865, tooth crown in labial view (A1), mesial (A2) and distal (A3) serration detail views. B. Dromaeosauridae indet., MWC 8872, tooth crown fragment in lingual (B1), distal (B2) and mesial (B3) serration detail views. C. Hadrosauridae indet., MWC 8896, tooth crown in occlusal (C1) and lateral (C2) views.
Remarks.—Recurved, laterally compressed teeth where the serrations are uniform in size and shape from base to tip of the crown have been assigned to the Dromaeosauridae (Williamson and Brusatte 2014). Based on the uniform shape and small size of the serrations, we suspect that this specimen most likely represents an indeterminate dromaeosaur. Without more material, more precise identification is impossible.
Genus Richardoestesia Currie, Rigby, and Sloan, 1990
Type species: Richardoestesia gilmorei Currie, Rigby, and Sloan, 1990; Dinosaur Park Formation, Upper Cretaceous.
cf. Richardoestesia sp.
Fig. 7A.
Material.—MWC 8865, tooth; from the Campanian–Maastrichtian, Williams Fork Formation, J&M site, northwest Colorado, USA.
Description.—Laterally compressed and serrated tooth with rounded denticles on the mesial and distal crown margins. Serrations along the distal surface are larger when compared to those on the mesial surface (Williamson and Brusatte 2014). Given the size of the tooth and relative sizes of the rounded denticles on either margin, we tentatively refer this tooth to Richardoestesia sp.
Remarks.—We refer one tooth to the genus Richardoestesia. The tooth is slightly recurved. The serrations along the distal surface of the crown are relatively large and rounded. The tooth was cracked in two places all the way through the crown when recovered. The specimen has since broken, although photomicrographs were taken before this occurred (Fig. 7A). Without more material, it is impossible to ascertain a species level identification of this tooth. Teeth assigned to Richardoestesia are relatively common in the Upper Cretaceous of North America (Sullivan and Lucas 2006), and Richardoestesia gilmorei has been reported from the Williams Fork Formation by Diem (1999).
Ornithischia Seeley, 1888
Ornithopoda Marsh, 1881
Hadrosauridae Cope, 1869
Hadrosauridae gen. et sp. indet.
Fig. 7C.
Material.—MWC 8896, teeth and tooth fragments; from the Campanian–Maastrichtian, Williams Fork Formation, J&M site, northwest Colorado, USA.
Description.—Five sided, ornithischian dinosaur tooth fragments bearing medial carinae belong to Hadrosauridae (Weishampel et al. 2004). The occlusal surfaces of the teeth are rhomboidally shaped in cross section, and the teeth vary in size. Unworn teeth have tall crowns with well-defined carinae along the lateral surfaces. Worn teeth may be wider than they are tall, and the carinae are often smoothed away (Coombs 1988).
Remarks.—We assign four teeth to indeterminate Hadrosauridae. All but one are fragmentary, and heavily worn. The lone whole tooth is roughly pentagonal in cross section. All the teeth are clearly heavily worn down, and all are roughly 10 mm in diameter. They appear to be crown remnants, and do not preserve significant root material. While unidentifiable to below the family level, these teeth and other occurrences of hadrosaurids from the Williams Fork Formation (Archibald 1987; Foster and Hunt-Foster 2015) indicate that these animals were widespread at the time.
Mammalia Linnaeus, 1758
Allotheria Marsh, 1880
Cimolomyidae Marsh, 1889
Genus Meniscoessus Cope, 1882
Type species: Meniscoessus conquistus Coper, 1882; Laramie Formation, Upper Cretaceous.
Meniscoessus sp.
Fig. 8A.
Material.—MWC 8848, upper incisor; from the Campanian–Maastrichtian, Williams Fork Formation, J&M site, northwest Colorado, USA.
Description.—MWC 8848, a mitt-shaped incisor measuring 4.73 mm in length by 2.06 mm in width, appears to have the entire crown intact. The tips of both the large anterior and small posterior cusps are worn. Based on its relatively large size and mitt-like morphology, MWC 8848 is identified as an upper incisor of species of Meniscoessus (compare to Sahni 1972: fig. 12M; Clemens 1963: fig. 45).
Remarks.—Comparison with AMNH 57873 (Clemens 1963: fig. 45) indicates that MWC 8848 is considerably smaller than upper incisors of Meniscoessus robustus Marsh, 1889, from the Lance Formation. Three species of Meniscoessus smaller than M. robustus are documented from the Williams Fork Formation. Meniscoessus collomensis Lillegraven, 1987, was initially described by Lillegraven (1987), and this species along with Meniscoessus major Russell, 1936, and Meniscoessus cf. M. intermedius Fox, 1976, were subsequently reported by Diem (1999). Therefore, it is probable that MWC 8848 belongs to one of these three species.
Multituberculata Cope, 1884
Cimolodontidae Marsh, 1889
Genus Cimolodon Marsh, 1889
Type species: Cimolodon nitidus Marsh, 1889; Lance Formation, Upper Cretaceous.
cf. Cimolodon nitidus Marsh, 1889
Fig. 8C.
Material.—MWC 8860, a fragment of a left p4; from the Campanian–Maastrichtian, Williams Fork Formation, J&M site, northwest Colorado, USA.
Description.—We recovered a fragment of a left p4 that preserves the posterior half of the tooth, including worn serrations, labial and lingual ridges, and the posterior root (Fig. 8C). Based on the wear pattern, there were seven serrations on the posterior half of the blade (Fig. 8C1), implying that the complete tooth probably had double this number. The tooth fragment preserves parts of five labial and lingual ridges originating from the serrations, with the two posterior-most serrations lacking distinct ridges.

Fig. 8. Representative mammalian teeth from the J&M site, Colorado, USA, Williams Fork Formation, Campanian–Maastrichtian, Upper Cretaceous. A. Meniscoessus sp., MWC 8848, incisor crown in labial (A1) and lingual (A1) views. B. Multituberculata indet., MWC 8863, tooth crown in occlusal (B1), lingual (B2), and distal (B3) views. C. cf. Cimolodon nitidus Marsh, 1889, MWC 8860, left p4 tooth crown in occlusal (C1), labial (C2), and lingual (C3) views. D. ?Leptalestes cooki (Clemens, 1966), MWC 8859, tooth crown in occlusal (D1), labial (D2), and lingual (D3) views.
Remarks.—The profile of MWC 8860 indicates a relatively high-crowned p4 with a steep posterior slope, comparable to the posterior profile of p4s of Cimolodon nitidus figured by Clemens (1963: figs. 23–25) and Lillegraven (1969: fig. 11), a species known from both Lancian and “Edmontonian” localities (Cifelli et al. 2004). In fact, Diem (1999) documented C. nitidus from the Williams Fork Formation. MWC 8860 measures 2.63 mm in length; therefore, we estimate the complete tooth measured at least 5 mm in length, which places it at the low end of the size range of p4s of C. nitidus from the Lance Formation (see Clemens 1963: table 7). Clemens (1963) further noted that on worn p4s of C. nitidus, the enamel is removed from posterior parts of the external face of the crown; this is the case for MWC 8860 (see Fig. 8C2).
Multituberculata fam., gen. et sp. indet.
Fig. 8B.
Material.—MWC 8863, a left M2; from the Campanian–Maastrichtian, Williams Fork Formation, J&M site, northwest Colorado, USA.
Description.—MWC 8863, a left M2, is heavily worn, but appears to have had a cusp formula of 1:3:3 or 4. The tooth is triangular in shape, and has a length of 1.91 mm and width of 1.71 mm.
Remarks.—Based on size, MWC 8863 is noticeably smaller than M2s of Cimolomys gracilis and Cimolodon nitidus measured by Clemens (1963) from the Lance Formation. Sahni (1972) noted that teeth of Cimolomys clarki from the Judith River Formation are approximately 70% the size of teeth of C. gracilis, although his figured M2 (Sahni 1972: fig. 11F) is still larger than MWC 8863, whose length and width measurements are less than 2 mm. The cusp formula and size of MWC 8863 fall within that of M2s of Mesodma sp. reported by Clemens (1963: table 3) from the Lance Formation. According to Archibald (1982), however, isolated M2s of species of Mesodma and Cimexomys are indistinguishable. Diem (1999) identified six isolated M2s from the Williams Fork Formation that he simply identified as “Mesodma sp. or Cimexomys sp.” (Diem 1999: 114); two of the six M2s (UCM 57372 and 57472) have the same length and width measurements as MWC 8863.
Metatheria Huxley, 1880
Pediomyoidea Simpson, 1927
Pediomyidae Simpson, 1927
Genus Leptalestes Davis, 2007
?Leptalestes cooki (Clemens, 1966)
Fig. 8D.
Material.—MWC 8859, a left m2 or m3; from the Campanian–Maastrichtian, Williams Fork Formation, J&M site, northwest Colorado, USA.
Description.—MWC 8859, a left m2 or m3, is the only metatherian tooth thus far recovered from the J&M site (Fig. 8D). The protoconid is broken off, but the other cusps are intact. On MWC 8859, the cristid obliqua attaches to the posterior wall of the trigonid near the midline of the protoconid, as is diagnostic for pediomyoids (Davis 2007). As is characteristic of ?Leptalestes cooki (see Davis 2007; Clemens 1966), the metaconid is larger than the paraconid, and the talonid is wider than the trigonid. The entoconid and hypoconulid are twinned, and the posteriorly-projecting hypoconulid is the lowest cusp on the tooth. Anterolabial and posterolabial cingulids are present on MWC 8859, although they do not extend around the labial margins of the proto- and hypoconids, respectively.
Remarks.—MWC 8859 has a length of 2.36 mm, a trigonid width of 1.67 mm, and a talonid width of 1.75 mm. Its size is within the range of m3s of ?Leptalestes cooki (= Pediomys cooki) measured by Clemens (1966: table 22), and is 0.1 mm longer than UCM 57394, a right m2 or m3 of ?Leptalestes cooki identified by Diem (1999) from the Williams Fork Formation. ?Leptalestes cooki is known from Judithian–Lancian aged localities in the U.S. Western Interior and Alberta, Canada (Davis 2007).
Discussion
The Williams Fork Formation has generally been interpreted as fluvio-deltaic deposits (Noll 1988; Foster and Hunt-Foster 2015) and our results suggest that the J&M site is consistent with this hypothesis. We report an assemblage that contains terrestrial, marine and estuarine faunal elements. The abundance of newly reported chondrichthyans from the J&M site provide some evidence for proximity to marine waters, as many are commonly found throughout the margin of the Western Interior Seaway and are often interpreted as euryhaline (Kirkland et al. 2013). Lonchidion griffisi has previously been reported only from a series of estuarine sites (Case 1987), although other species of Lonchidion have been reported from a variety of environments, so the genus was likely euryhaline as well (Rees and Underwood 2002; Fischer 2008). The shark Chiloscyllium is the most common non-batoid chondrichthyan at the J&M site with 19 specimens. While it has been reported from freshwater deposits (Kirkland et al. 2013) it is more frequently recovered as part of marine assemblages (Becker et al. 2010; Guinot et al. 2013; Schubert et al. 2017). Cantioscyllium markaguntensis has only been reported from a freshwater deposit (Kirkland et al. 2013), although it is the rarest of the Williams Fork chondrichthyans. Myledaphus is often extremely abundant in latest Cretaceous freshwater deposits (Demar and Breithaupt 2006), but Pseudomyledaphus dominates the J&M site elasmobranch fossils by number, as has been noted for multiple sites of Campanian age in Alberta, Canada (Cullen et al. 2016). Five of these chondrichthyans are new to the Williams Fork Formation, indicating that the J&M site may be recording a different paleoenvironment than previously collected material from the formation.
The osteichthyan fauna of the J&M site points towards some marine influence. Bryant (1987) noted that the amiid Melvius is very frequently recovered in close proximity to the Western Interior Seaway, and that other, smaller amiids were fully marine during the Cretaceous, making their presence at the J&M site unsurprising. There is a possibility that some of the smaller teeth which we have referred to Melvius could belong to these smaller amiids, as all the specimens are isolated. Paralbula casei is known from marine localities, and it has been reported that most well preserved phyllodontid fossils are recovered from estuarine sites, and not purely marine or freshwater deposits (Estes 1969b). The lepisosteid fish suggest a freshwater or estuarine setting (Grande 2010). This combined fauna of chondrichthyans and osteichthyans appears to represent an estuarine or otherwise brackish water environment. While the common terrestrial and freshwater fossils (gastropods, leaves, dinosaurs, mammals, lepisosteids, crocodyliforms, and testudines) should not be overlooked when interpreting the depositional environment of the J&M site, the overall assemblage is indicative of a coastal or marginal environment, as has previously been suggested for parts of the Williams Fork Formation
While the J&M site appears to be a fluvial channel scour deposit (Foster and Hunt-Foster 2015), it is not difficult to explain the abundance of marine and brackish associated fauna found there. The dinosaurs and mammals are representative of a terrestrial environment, and their remains were likely simply transported by the river that deposited the sediments. Similarly, we recovered a leaf from the “J&M scour”, portion of the site. Marine taxa are parsimoniously explained by the proximity of the site to the margin of the Western Interior Seaway at the time. If close enough to the location of the Western Interior Seaway, euryhaline fishes and invertebrates would be free to travel between the two environments and could end up fossilized in the channel lags that likely resulted in the J&M site. Alternatively, it is possible that changing sea levels could have moved and reworked already deposited materials through time, resulting in the concentration of microfossils from multiple fluvial, estuarine, and marine environments. In either case, the paleofauna preserved at the J&M site conforms with prior reports of the Williams Fork Formation containing fluvio-deltaic, marginal or near-marine deposits.
We briefly examined the level of
post-mortem wear and erosion on tooth fossils in order to try and
constrain transport distance differences between faunal groups.
Terrestrial fossils, as expected, tended to have more breaks and were
often worn and/or abraded by transportation processes. Unfortunately,
there was no clear distinction between the groups we had deemed marine
and fluvial. Fossils from both groups exhibited variable wear, with some
specimens essentially pristine, and other specimens broken and worn,
even within the same taxa. This means that inferred transport distance
may not be an effective means of interpreting the depositional
environment of the J&M site, at least with the current collection.
The real depositional history of the J&M site is almost certainly
more complex than simply marine, fluvial, or estuarine.
The J&M site superficially shares characteristics with
channel-hosted microfossil beds as described by Rogers and Brady (2010).
The fossils appear to have been deposited within a channel scour
feature, and are overwhelmingly disarticulated or fragmentary in nature.
Both millimeter- and centimeter-scale specimens have been recovered from
the site, and there are well preserved as well as broken and abraded
fossils. While these characteristics would seem to point towards a
channel-hosted microfossil bed (Rogers and Brady 2010),
a more detailed sedimentological analysis is required of the site before
such a hypothesis can be sufficiently evaluated.
In addition to providing information about the potential paleoenvironment at the time of deposition, the microvertebrate assemblage from the J&M site may also serve as valuable tool for constraining the age of the Williams Fork Formation in the absence of numerical dates. The fossil mammals reported help constrain the age for the J&M site, as both ?Leptalestes cooki and species of Meniscoessus are found in strata that range from the Judithian to the Lancian land mammal ages. Cimolodon nitidus has been reported from “Edmontonian” through Lancian aged strata as well (Cifelli et al. 2004). Thus the mammalian fauna of the J&M site points towards a late Campanian to late Maastrichtian age of deposition. This is consistent with the findings of Diem (1999) who suggested the Williams Fork Formation fauna was intermediate in age between the Judithian and Lancian ages.
The reported teeth of Myledaphus and Peneteius are potentially significant. Some of the J&M site Myledaphus teeth may represent an intermediate between the ridge-enameled M. bipartitus and the pit-enameled M. pustolosus. The former species is common in rocks of Campanian age, while the latter is found only in the Maastrichtian (Cook et al. 2014). Similarly, the Peneteius tooth recovered from the J&M site is intermediate in both morphology and geographic location between the two previously published species of this genus. The Campanian P. saueri is recovered from sediments in the southwestern North America, while the Maastrichtian P. aquilonius is known from further north along the Western Interior Seaway (Nydam et al. 2013). Because it is intermediate both geographically and morphologically, it seems likely that this tooth belonged to an organism that is temporally intermediate between the late Campanian and late Maastrichtian. As hypothesized for the Peneteius tooth reported here (MWC 8871), this may indicate that the J&M site represents a temporal bridge between the myriad upper Campanian and upper Maastrichtian fossil-bearing localities of western North America.
Conclusions
The J&M site of the Williams Fork Formation requires a more complex screenwashing process than usual (including the use of DMSO), but has proven to be a rich microvertebrate locality. We have recovered nine taxa that are not only new to the J&M site, but new to the Williams Fork Formation as a whole. These first reports of taxa from the Williams Fork Formation include the first non-batoid sharks (Lonchidion, Chiloscyllium, Cantioscylium), two rays (Pseudomyledaphus, Cristomylus), the first lungfish and pycnodontids, and the squamate Peneteius. Together, the new osteichthyan and chondrichthyan fossils suggest an estuarine or brackish environment for the J&M site. We note that, in addition to being new records of taxa from the formation, some of these records will also have implications for the geographic distribution of diverse taxa. For example, the record of Peneteius appears to bridge both the latitudinal and temporal gap between Campanian Peneteius records to the south and Maastrichtian records to the north. Some specimens of Myledaphus from this site may also represent a transitional form between the upper Campanian Myledaphus bipartitus and the upper Maastrichtian Myledaphus pustolosus. The mammalian microvertebrate assemblage suggests a Judithian–Lancian age for the site. Continued study may have implications for the entire Williams Fork Formation as a whole, especially regarding environments that are transitional between terrestrial and marine.
Acknowledgements
We acknowledge the Appalachian State University (ASU) “Finding Fossils on Friday” (FFF) volunteers for their hard work and time picking through concentrated fossil-bearing sediment. Members of the ASU Department of Geology Summer 2016, 2018, and 2019 GLY 2857 Field and Museum Methods courses helped collect the fossils and sediment that we used for this study. Jayson Sellars (ASU undergraduate) produced the digital elevation model in Fig. 1C under the direction of Scott Marshall (ASU). Numerous persons, particularly James I. Kirkland (Utah Geological Survey, Salt Lake City, USA) and David Ward (Natural History Museum London, UK), provided useful feedback on earlier versions of this work presented at the Society of Vertebrate Paleontology meetings in 2017 and 2018. Reviews by Thomas Cullen (Carleton University, Ottawa, Canada), Charlie Underwood (University of London, Birbeck, UK), and an anonymous reviewer contributed greatly to the quality of this manuscript, and we appreciate guidance from the editor, Daniel B. Barta, as well. We would like to thank the Bureau of Land Management for the collection permit used to recover fossils from the J&M site, and the Museum of Western Colorado for serving as the repository for these fossils. We acknowledge the ASU Department of Geological and Environmental Sciences for equipment and funding used to conduct this research, as well as the ASU Office of Student Research for additional support of the student authors (NB and IS).
References
Applegate, S.P. 1972. A revision of the higher taxa of orectolobids. Journal of the Marine Biological Association of India 14: 743–751.
Archibald, J.D. 1982. A study of Mammalia and geology across the Cretaceous–Tertiary boundary in Garfield County, Montana. University of California Publications in Geological Sciences 122: 1–286.
Archibald, J.D. 1987. Late Cretaceous (Judithian and Edmontonian) vertebrates and geology of the Williams Fork Formation, N.W. Colorado. In: P.M. Currie and E.H. Koster (eds.), Fourth Symposium on Mesozoic Terrestrial Ecosystems, Short Papers, 7e11. Royal Tyrell Museum, Drumheller.
Avrahami, H.M., Heckert, A.B., and Martin, L. 2015. Comparison of nested sieves, traditional screen boxes, and paint sieves for the recovery of microvertebrate fossils. Journal of Vertebrate Paleontology 35: 81–82.
Becker, M.A., Wellner, R.W., Mallery, C.S., Jr., and Chamberlain, J.A. 2010. Chondricthyans from the lower Ferron Sandstone Member of the Mancos Shale (Upper Cretaceous: Middle Turonian) of Emery and Carbon counties, Utah, USA. Journal of Paleontology 84: 248–266. Crossref
Berg, L.S. 1937. A classification of fish-like vertebrates. Bulletin de l’Académie des Sciences de l’URSS. Classe des Sciences Mathématiques et Naturelles 4: 1277–1280.
Berg, L.S. 1940. Classification of fishes both recent and fossil. Transactions of the Zoological Academy of Sciences, USSR 5: 85–517.
Blake, S. 1940. Paralbula, a new fossil fish based on basal plates from the Eocene and Miocene of Maryland. Journal of the Washington Academy of Sciences 30: 205–209.
Bonaparte, C.L. 1838. Selachorum tabula analytica. Nouvelles Annales des Sciences Naturelles 2: 195–214.
Brand, N.A., Heckert, A.B., Foster, J.R., and Hunt-Foster, R.K. 2017. The microvertebrate fossil assemblage of the Upper Cretaceous (Campanian–Maastrichtian) Williams Fork Formation, Western Colorado. Journal of Vertebrate Paleontology 35: 85.
Brinkman, D.B., Neuman, A.G., and Divay, J.D. 2017. Non-marine fishes of the late Santonian Milk River Formation of Alberta, Canada—Evidence from vertebrate microfossil localities. Vertebrate Anatomy Morphology Paleontology 3: 7–46. Crossref
Bryant, L.J. 1987. A new genus and species of Amiidae (Holostei; Ostichthyes) from the Late Cretaceous of North America, with comments on the phylogeny of the Amiidae. Journal of Vertebrate Paleontology 7: 349–361. Crossref
Cappetta, H. 1973. Selachians from the Carlile Shale (Turonian) of South Dakota. Journal of Paleontology 47: 504–514.
Cappetta, H. 1980. Les Sélaciens du Crétacé Supérieur du Liban. II. Batoides. Palaeontographica A 168: 149–229.
Case, G.R. 1979. Additional fish records from the Judith River Formation (Campanian) of Montana. Geobios 12 (2): 223–233. Crossref
Case, G.R. 1987. A new selachian fauna from the Late Campanian of Wyoming (Teapot Sandstone Member, Mesaverde Formation, Big Horn Basin). Palaeontographica A 197 (1–3): 1–37.
Case, G.R. and Schwimmer, D.R. 1988. Late Cretaceous fish from the Blufftown Formation (Campanian) in western Georgia. Journal of Paleontology 62: 290–301. Crossref
Cifelli, R.L., Eberle, J.J., Lofren, D.L., Lillegraven, J.A., and Clemens, W.A. 2004. Mammalian biochronology of the latest Cretaceous. In: M.O. Woodburne (ed.), Late Cretaceous and Cenozoic mammals of North America, 20–42. Columbia University Press, New York. Crossref
Clemens, W.A. 1963. Fossil mammals of the type Lance Formation, Wyoming. Part I. Introduction and Multituberculata. University of California Publications in Geological Sciences 48: 1–104.
Clemens, W.A. 1966. Fossil mammals of the type Lance Formation, Wyoming. Part II. Marsupialia. University of California Publications in Geological Sciences 62: 1–122.
Compagno, L.J.V. 1973. Interrelationships of living elasmobranchs. Journal of the Linnaen Society 53: 63–98.
Compagno, L.J.V. 1977. Phyletic relationships of living sharks and rays. American Zoologist 17: 303–322. Crossref
Cook, T.D., Newbrey, M.G., Brinkman, D.B., and Kirkland, J.I. 2014. Euselachians from the freshwater deposits of the Hell Creek Formation of Montana. The Geological Society of America Special Paper 503: 229–246. Crossref
Coombs, W. P. 1988, The Status of the dinosaurian genus Diclonius and the taxonomic utility of hadrosaurian teeth. Journal of Paleontology 62: 812–817.
Cope, E.D. 1869. Synopsis of the extinct Batrachia, Reptilia, and Aves of North America. Transactions of the American Philosophical Society 14: 252. Crossref
Cope, E.D. 1876. On some extinct reptiles and Batrachia from the Judith River and Fox Hills beds of Montana. Proceedings of the Academy of Natural Sciences of Philadelphia 28: 340–359.
Cope, E.D. 1882. Mammalia in the Laramie Formation. American Naturalist 16: 830–831. Crossref
Cope, E.D. 1884. The Tertiary Marsupialia. The American Naturalist 18: 686–697. Crossref
Cullen, T.M., Fanti, F., Capobianco, C., Ryan, M.J., and Evans, D.C. 2016. A vertebrate microsite from a marine-terrestrial transition in the Foremost Formation (Campanian) of Alberta, Canada, and the use of faunal assemblage data as a paleoenvironmental indicator. Palaeogeography, Palaeoclimatology, and Palaeoecology 444: 101–114. Crossref
Currie, P.J., Rigby, J.K., Jr., and Sloan, R.E. 1990. Theropod teeth from the Judith River Formation of southern Alberta, Canada. In: K. Carpenter and P.J. Currie (eds.), Dinosaur Systematics: Approaches and Perspectives, 108–125. Cambridge University Press, Cambridge. Crossref
Cuvier, G. 1825. Recherches sur les ossemens fossiles, ou l´on rérablit les charactèrs de plusieurs animaux dont les revolutions du globe ont détrout les espèces. 4514 pp. G. Dufour et E. d’Ocagne, Paris.
Davis, B.M. 2007. A revision of “pediomyid” marsupials from the Late Cretaceous of North America: Acta Palaeontologica Polonica 52: 217–256.
Demar, D.G., Jr. and Breithaupt, B.H. 2006. The Non mammalian Vertebrate Microfossil Assemblages of the Mesaverde Formation (Upper Cretaceous, Campanian) of the Wind River and Bighorn basins, Wyoming. New Mexico Museum of Natural History and Science Bulletin 35: 33–53.
Denison, R.H. 1974. The structure and evolution of teeth in lungfishes: Fieldiana Geology 33 (3): 31–58. Crossref
Diem, S. 1999. Vertebrate Faunal Analysis of the Upper Cretaceous Williams Fork Formation, Northwestern Colorado. 188 pp. Master’s Thesis, San Diego State University, San Diego.
Diem, S.D. and Archibald, J.D. 2000. Vertebrate fauna from the Upper Cretaceous Williams Fork Formation, northwestern Colorado. Journal of Vertebrate Paleontology 20 (Supplement to 3): 38–39A.
Diem, S. and Archibald, J.D. 2005. Range extension of Southern Chasmosaurine Ceratopsian Dinosaurs into Northwestern Colorado. Journal of Paleontology 79: 251–258. Crossref
Duffin, C.J. 1985. Revision of the hybodont selachian genus Lissodus Brough (1935). Palaeontographica A 188: 105–152.
Estes, R. 1964. Fossil vertebrates from the Late Cretaceous Lance Formation, eastern Wyoming. California University Publications on Geological Sciences 49: 180.
Estes, R. 1969a. Relationships of two Cretaceous lizards (Sauria, Teiidae). Breviora 317: 1–8.
Estes, R. 1969b. Studies on fossil phyllodont fishes: interrelationships and evolution in the Phyllodontidae (Albuloidei). Copeia 1969 (2): 317–331 Crossref
Estes, R. 1983. Handbuch der Paläoherpetologie. Teil 10A. Sauria terrestria, Amphisbaenia. 245 pp. Gustav Fischer, Stuttgart.
Fischer, J.R. 2008. Brief synopsis of the hybodont form taxon Lissodus Brough, 1935, with remarks on the environment and associated fauna. Freiberger Forschungshefte C 528: 1–23.
Foster, J.R. and Hunt-Foster, R.K. 2015. First report of a giant neosuchian (Crocodyliformes) in the Williams Fork Formation (Upper Cretaceous: Campanian) of Colorado. Cretaceous Research 55: 66–73. Crossref
Fox, R.C. 1976. Cretaceous mammals (Meniscoessus intermedius, new species, and Alphadon sp.) from the lowermost Oldman Formation, Alberta. Canadian Journal of Earth Sciences 13: 1216–1212. Crossref
Gill, T. 1862. Analytical synopsis of the order of Squali: and revision of the nomenclature of the genera. Annals of the Lyceum of Natural History of New York 7 (32): 367–413. Crossref
Fowler, D.W. 2017. Revised geochronology, correlation, and dinosaur stratigraphic ranges of the Santonian–Maastrichtian (Late Cretaceous) formations of the Western Interior of North America. PLOS ONE 12 (11): e0188426. Crossref
Gilmore, C.W. 1911. A new fossil alligator from the Hell Creek beds of Montana. Proceedings of the United States National Museum 41: 297–302. Crossref
Gmelin, J.F. 1788. Systema naturae de Caolus Linnaeus. Editio decema tertia, acuta, reformate, 2, N. P. 1661 pp. Georg Emanuel Beer, Leipzig.
Grande, L. 2010. An empirical synthetic pattern study of gars (Lepisosteiformes) and closely related species, based mostly on skeletal anatomy. The resurrection of Holostei. American Society of Ichthyologists and Herpetologists Special Publication 6: 1–871.
Grande, L. and Bemis, W.E. 1998. A comprehensive phylogenetic study of amiid fishes (Amiidae) based on comparative skeletal anatomy, an empirical search for interconnected patterns of natural history. Society of Vertebrate Paleontology Memoir 4: 1–690. Crossref
Gray, J.E. 1844. Catalogue of Tortoises, Crocodilians, and Amphisbaenians in the Collection of the British Museum. 88 pp. British Museum (Natural History), London.
Guinot, G., Underwood, C.J., Cappetta, H., and Ward, D.J. 2013. Sharks (Elasmobranchii: Euselachii) from the Late Cretaceous of France and the UK. Journal of Systematic Paleontology 11: 589–671. Crossref
Harrell, T.L. and Ehret, D.J. 2019. Lungfish tooth plates (Sarcopterygii, Dipnoi) from the Late Cretaceous (Santonian) Eutaw Formation of Alabama and Mississippi, USA. Journal of Paleontology 93: 1–12. Crossref
Hay, O.P. 1902. Bibliography and catalogue of the fossil Vertebrata of North America. U.S. Geological Survey Bulletin 179: 1–868. Crossref
Hay, O.P. 1929. Second bibliography and catalogue of the fossil Vertebrata of North America. Publications of the Carnegie Institute of Washington 390: 1–2003.
Herman, J. 1977. Les sélachians des terrains néocrétacés & paléocénes de Belgique & des contrées limitrophes. Mémoires pour servir á l’esplication de Cartes Géologiques et Mini´res de la Belgique, Service Géologique de Belgique 15: 1–401.
Hettinger, R.D. and Kirschbaum, M.A. 2002. Stratigraphy of the Upper Cretaceous Mancos Shale (upper part) and Mesa Verde Group in the southern part of the Uinta and Piceance basins, Utah and Colorado. USGS Investigations Series I-2764: 1–21.
Huene, F. von 1914. Das natürliche system der Saurischia. Zentralblatt Mineralogie, Geologie, und Palaeontologie B 1914: 154–158.
Hunt-Foster, R.K. and Foster, J.R. 2013. Paleofauna of the Williams Fork Formation (Upper Cretaceous), northwestern Colorado: coastal deltaic deposits dominated by freshwater taxa. Journal of Vertebrate Paleontology 33: 146.
Huxley, T.H. 1875. On Stagonolepis robertsoni, and on the evolution of the Crocodile. Proceedings of the Geological Society of London 31: 423–438. Crossref
Huxley, T.H. 1880. On the application on the laws of evolution to the arrangement of the Vertebrata and more particularly of the Mammalia. Zoological Society of London, Scientific Memoirs 4: 457–472.
Kirkland, J.I. 1987. Upper Jurassic and Cretaceous lungfish tooth plates from the Western Interior. The last dipnoan faunas of North America. Hunteria 2 (2): 1–16.
Kirkland, J.I., Eaton, J.G., and Brinkman, D.B. 2013. Elasmobranchs from upper Cretaceous freshwater facies in southern Utah. In: A.L. Titus and M.A. Loewen (eds.), At the Top of the Grand Staircase: the Late Cretaceous of Southern Utah, 153–194. Indiana University Press, Bloomington.
Kirschbaum, M.A. and Hettinger, R.D. 2004. Facies analysis and sequence stratigraphic framework of Upper Campanian strata (Neslen and Mount Garfield Formations, Bluecastle Tongue of the Castlegate Sandstone, and Mancos Shale), eastern Book Cliffs, Colorado and Utah. USGS Digital Data Series 69G: 1–40. Crossref
Klein, E.F. 1885. Beiträge zur Bildung des Schädels der Knochenfische, 2. Jahreshefte des Vereins für Vaterländische Naturkunde in Württemberg 42: 205–300.
Kriwet, J. 2005. A comprehensive study of the skull and dentition of pycnodont fishes. Zitteliana 45: 135–188.
Lillegraven, J.A. 1987. Stratigraphic and evolutionary implications of a new species of Meniscoessus (Mutlituberculata, Mammalia) from the Upper Cretaceous Williams Fork Formation, Moffat County, Colorado. Dakoterra 3: 46–56.
Lillegraven, J.A. 1969. Latest Cretaceous mammals of upper part of Edmonton Formation of Alberta, Canada, and review of marsupial-placental dichotomy in mammalian evolution. University of Kansas Paleontological Contributions 50: 1–122.
Laurenti, J.N. 1768. Classis reptilium. Specimen medicum, exhibens synopsis Reptilium emendatum, cum experimentis circa venena et antidote Reptilium Austriacorum. 214 pp. Joan Thomae, Viennae. Crossref
Linnaeus, C. 1758. Systema naturæ per regna tria naturæ, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Tomus I. Editio decima, reformata. 824 pp. Laurentius Salvius, Stockholm. Crossref
Lucas, S.G. and Kihm, A.J. 1982. Paleontological Resource Study and Inventory of Part of the White River Resource Area and Vicinity, Piceance Creek Basin, Northeastern Colorado. 141 pp. ESCA-Technical Corporation, Albuquerque.
Lucas, S.G., Sullivan, R.M., and Hunt, A.P. 2006. Re-evaluation of Pentaceratops and Chasmosaurus (Ornithischia: Ceratopsidae) in the Upper Cretaceous of the Western Interior. New Mexico Museum of Natural History and Science Bulletin 35: 367–370.
Marsh, O.C. 1880. Notice of new Jurassic mammals representing two new orders. The American Journal of Science, Series 3 20 (28): 235–239. Crossref
Marsh, O.C. 1881. Classification of the Dinosauria. The American Journal of Science, Series 3 19: 253–259.
Marsh, O.C. 1889. Discovery of Cretaceous Mammalia. Part II. The American Journal of Science, Series 38: 81–92. Crossref
Matthew, W.D. and Brown, B. 1922. The family Deinodontidae, with notice of a new genus from the Cretaceous of Alberta. Bulletin of the American Museum of Natural History 46: 367–385.
McCord, R.D. 1998. A new genus and species of Cretaceous polyglyphanodontinen lizard (Squamata, Teiidae) from the Kaiparowits Plateau, Utah. In: Y. Yomida, L.J. Flynn, and L.L. Jacobs (eds.), Advances in Vertebrate Paleontology and Geochronology. National Science Museum Monographs 14: 281–292.
Müller, J. 1844. Über den Bau und die Grenzen der ganoiden und über das natürliche System der Fische. Abhandlungen der Akademie der Wissenschaften zu Berlin 1844: 117–216.
Müller, J. and Henle, F.G.J. 1837. On the generic characters of cartilaginous fishes with descriptions of new genera. Magazine of Natural History 2: 1–91.
Müller, J. and Henle, F.G.J. 1838. Systematische Beschreibung der Plagiostomen. xxii + 200 pp. Veit, Berlin.
Noll, M.D. 1988. Sedimentology of the Upper Cretaceous Williams Fork Formation, Rio Blanco County, Northwestern Colorado. 213 pp. Master’s Thesis, San Diego State University, Department of Geological Sciences, San Diego.
Nydam, R.L., Eaton, J.G., and Sankey, J. 2007, New taxa of transversely-toothed lizards (Squamata: Scincomorpha) and new information on the evolutionary history of “Teiids”: Journal of Paleontology 81: 538–549. Crossref
Nydam, R.L., Gauthier, J.A., and Chiment, J.J. 2000. The mammal-like teeth of the Late Cretaceous Lizard Peneteius aquilonius Estes, 1969 (Squamata, Teiidae). Journal of Vertebrate Paleontology 20: 628–631. Crossref
Nydam, R.L., Rowe, T.B., and Cifelli, R.L. 2013. Lizards and snakes of the Terlingua local fauna (late Campanian), Aguja Formation, Texas, with comments on the distribution of paracontemporaneous squamates throughout the Western Interior of North America: Journal of Vertebrate Paleontology 33: 1081–1099. Crossref
Oppel, M. 1811. Die Ordnungen, Familien und Gattungen der Reptilien als Prodrom einer Naturgeschichte derselben. 87 pp. Joseph Lindauer, Munchen. Crossref
Owen, R. 1842. Report on British fossil reptiles. Report of the British Association of Advanced Sciences 9: 60–204.
Owen, R. 1846. Lectures on the Comparative Anatomy and Physiology of the Vertebrate Animals, Delivered at the Royal College of Surgeons of England in 1844 and 1846. Part 1: Fishes. 308 pp. Longman, Brown, Green, and Longmans, London. Crossref
Raynolds, R.G. and Hagadorn, J.W. 2016. Colorado stratigraphy chart. Colorado Geological Survey, Map Series 53. Denver Museum of Nature and Science, Denver.
Rees, J. and Underwood, C.J. 2002. The status of the shark genus Lissodus Brough, 1935, and the position of the nominal Lissodus species within the Hybodontoidea (Selachii). Journal of Vertebrate Paleontology 22: 471–479. Crossref
Rogers, R.R. and Brady, M.E. 2010. Origins of microfossil bonebeds: Insights from the Upper Cretaceous Judith River Formation of north-central Montana. Paleobiology 36: 80–112. Crossref
Romer, A.S. 1955. Herpetichthyes, Amphibioidei, Choanichthyes or Sarcopterygii. Nature 176: 126. Crossref
Russell, L.S. 1936. New and interesting mammalian fossils from western Canada. Transactions of the Royal Society Canada Series 3 30: 75–80.
Sahni, A. 1972. The vertebrate fauna of the Judith River Formation, Montana. Bulletin of the American Museum of Natural History 147: 1–412.
Schubert, J.A., Wick, S.L., and Lehman, T.M. 2017. An Upper Cretaceous (Middle Campanian) marine chondrichthyan and osteichthyan fauna from the Rattlesnake Mountain Sandstone Member of the Aguja Formation in West Texas. Cretaceous Research 69: 6–33. Crossref
Sauvage, H.E. 1875. Sur la faune ichthyologique de I’ile Saint-Paul. Compte Rendu de l’Acadamie des Sciences, Paris 81: 987–989.
Seeley, H.G. 1888. The classification of the Dinosauria. Report of the British Association for the Advancement of Science 57: 698–699.
Simpson, G.G. 1927. Mammalian fauna of the Hell Creek Formation of Montana. American Museum Novitates 267: 1–7.
Sullivan, R.M. and Lucas, S.G. 2003. The Kirtlandian, a new land-vertebrate “age” for the Late Cretaceous of western North America. In: S.G. Lucas, S.C. Semken, W. Berglof, D. Ulmer-Scholle (eds.), Geology of the Zuni Plateau. New Mexico Geological Society 54th Annual Fall Field Conference Guidebook, 369–377. New Mexico Geological Society, Socorro. Crossref
Sullivan, R.M. and Lucas, S.G. 2006. The Kirtlandian land-vertebrate “age”-faunal composition, temporal position and biostratigraphic correlation in the nonmarine Upper Cretaceous of western North America. New Mexico Museum of Natural History and Science Bulletin 35: 7–29.
Sullivan, R.M., Lucas, S.G., and Jasinski, S.E. 2011. Preliminary observations on a skull of the amiid fish Melvius, from the Upper Cretaceous Kirtland Formation, San Juan Basin, New Mexico. New Mexico Museum of Natural History and Science Bulletin 53: 475–483.
Triplehorn, D.M. 2002. An easy way to remove fossils from sandstones: DMSO disaggregation. Journal of Paleontology 76: 394–395. Crossref
Vaughn, M.C., Foster, J.R., Nydam R.L., Heckert, A.B., and Hunt-Foster, R.K. 2015. Newly reported lissamphibian and squamate taxa from the Williams Fork Formation (Upper Cretaceous; Campanian), Colorado: Closing the Gap. Journal of Vertebrate Paleontology, SVP Program and Abstracts Book 35: 230.
Weishampel, D.B., Dodson, P., and Osmólska, H. 2004. The Dinosauria. 2nd Edition. xviii + 861 pp. University of California Press, Berkeley. Crossref
Williamson, T.E. 1996. ?Brachychampsa sealeyi, sp. nov., (Crocodylia, Alligatoroidea) From the Upper Cretaceous (Lower Campanian) Menefee Formation, Northwestern New Mexico. Journal of Vertebrate Paleontology 16: 421–431. Crossref
Williamson, T.E. and Brusatte, S.L. 2014. Small theropod teeth from the Late Cretaceous of the San Juan Basin, northwestern New Mexico and their implications for understanding latest Cretaceous dinosaur evolution. PLoS ONE 9 (4): e93190. Crossref
Woodward, A.S. 1889. Catalogue of the Fossil Fishes in the British Museum (Natural History). 474 pp. British Museum of Natural History, London. Crossref
Acta Palaeontol. Pol. 67 (3): 579–600, 2022
https://doi.org/10.4202/app.00934.2021
Vertebrate faunal list of the Williams Fork Formation. Taxa in bold are new and reported from the formation for the first time here. Taxa preceded by an * are previously reported, but illustrated here for the first time. Non-bold taxa reported previously and compiled from Lucas and Kihm (1982), Archibald (1987), Lillegraven (1987), Diem (1999) and Diem and Archibald (2005).
Elasmobranchii Bonaparte, 1838
Euselachii Hay, 1902
Hybodontoidea Owen, 1846
Lonchidiidae Herman, 1977
Lonchidion Estes, 1964
Lonchidion griffisi Case, 1987
Neoselachii Compagno, 1977
Galeomorphii Compagno, 1973
Orectolobiformes Applegate, 1972
Ginglymostomatidae Gill, 1862
Cantioscyllium Woodward, 1889
Cantioscyllium markaguntensis Kirkland, Eaton, and Brinkman, 2013
Hemiscyllidae Gill, 1862
Chiloscyllium Müller and Henle, 1838
Chiloscyllium sp.
Batomorphii Cappetta, 1980
Rajiformes Berg, 1940
Rhinobatoidea Müller and Henle, 1837
*Myledaphus Cope, 1876
Myledaphus bipartitus Cope, 1876
Cristomylus Kirkland, Eaton, and Brinkman, 2013
Cristomylus sp.
Pseudomyledaphus Kirkland, Eaton, and Brinkman, 2013
Pseudomyledaphus madseni Kirkland, Eaton, and Brinkman, 2013
Osteichthyes Huxley, 1880
Actinopterygii Klein, 1885
Amiidae Bonaparte, 1838
Amiinae
Amia sp.
Amiinae gen. et sp. nov.
Vidalamiinae Grande and Bemis, 1998
Vidalamiini Grande and Bemis, 1998
*Melvius Bryant, 1987
cf. Melvius sp.
Semionotiformes
Gen. et sp. indet.
Acipenseriformes(?)
Gen. et sp. indet.
Pycnodontiformes Berg, 1937
Pycnodontiformes gen. et sp. indet.
Phyllodontidae Sauvage, 1875
Paralbula Blake, 1940
Paralbula casei Estes, 1969b
Holostei Müller, 1844
Lepisosteiformes Hay, 1929
Lepisosteidae Cuvier, 1825
*Lepisosteidae gen. et sp. indet.
Sarcopterygii Romer, 1955
Dipnoi Müller, 1844
Dipnoi gen. et sp. indet.
Amphibia
Salentia
Anura indet.
Caudata
Caudata indet.
Reptilia Laurenti, 1768
Testudinata
Baenidae
Boremys sp.
Neurankylidae
Neurankylus sp.
Thescelus sp.
Compsemys(?) sp.
Trionychidae
Trionyx sp.
Dermatomydidae
Adocus sp.
Squamata Oppel, 1811
Teiidae Gray, 1827
Peneteius estes, 1969a
Peneteius sp.
Choristodera
Champsosaurus sp.
Crocodylomorpha Hay, 1930
Crocodylia Gmelin, 1788
Eusuchia Huxley, 1875
Crocodylidae
Leidyosuchus sp.
Alligatoroidea Gray, 1844
*Brachychampsa Gilmore, 1911
cf. Brachychampsa sp.
Theropoda Marsh, 1881
Coelurosauria von Huene, 1914
Troodontidae
Troodon formosus
Dromaeosauridae Matthew and Brown, 1922
Dromaeosaurus albertensis
Saurornitholestes langstoni
Gen. et sp. indet.
*Richardoestesia Currie Rigby and Sloan, 1990
cf. Richardoestesia sp.
Tyrannosauridae
aff. Daspletosaurus
aff. Albertosaruus
Gen. et sp. indet.
Ornithischia Seeley, 1888
Ankylosauria
Ankylosauridae
Ankylosaurus magniventris
Nodosauridae
Gen. et sp. indet.
Ornithopoda Marsh, 1881
Hypsilophodontidae
Thescelosaurus neglectus
Hadrosauridae Cope, 1869
Gen. et sp. indet.
Ceratopsia
Ceratopsidae
cf. Pentaceratops sternbergi
Incertae sedis
Indet. marine reptile
Mammalia Linnaeus, 1758
Allotheria Marsh, 1880
Multituberculata Cope, 1884
Neoplagiaulacidae
Mesodma thompsoni
Cimolodontidae
Cimolodon nitidus
Cimolomyidae Marsh, 1889
Meniscoessus aff. M. intermedius
Meniscoessus major
Meniscoessus collomensis
cf. Cimolomys sp.
Family indet.
?Paracimexomyes sp.
Gen. et sp. nov. 1 (Cimolomys-like)
?Gen. et sp. nov. 2 (plagiaulacid or Microcosmodontidae eucosmodontid)
?Gen. et sp. nov.
Metatheria Huxley, 1880
Family indet.
Aenigmadephys sp. nov.
“Alphadontidae”
Turgidodon rhaister
Turgidodon russelli
Alphadon marshi
Alphadon wilsoni
Pediomyidae
Pediomys cooki
Aquiladelphis incus
Stagodontidae
Eodelphis sp.
Marsupialia Illiger, 1811
Marsupialia indet.