
Palaeobiogeographical implications of the first fossil wood flora from the Jurassic of Turkey
ÜNAL AKKEMİK, RAİF KANDEMİR, MARC PHILIPPE, YILDIRIM GÜNGÖR, and FATİH KÖROĞLU
Akkemik, Ü., Kandemir, R., Philippe, M., Güngör, Y., and Köroğlu, F. 2022. Palaeobiogeographical implications of the first fossil wood flora from the Jurassic of Turkey. Acta Palaeontologica Polonica 67 (3): 745–766.
We describe Jurassic fossilized woods from the Gümüşhane and Erzurum regions of Turkey that represent the eastern Sakarya Zone (eSZ) terrestrial biota. We collected 27 fossil wood fragments in total. All 27 fossil wood specimens represent coniferous trees. We assigned ten specimens to Agathoxylon sp. type 1, fourteen to Agathoxylon sp. type 2, two to Protelicoxylon asiaticum and one to Xenoxylon hopeiense. Middle Jurassic woods are represented by Agathoxylon only what does not allow for any palaeobiogeographic inferences. The Late Jurassic wood flora evidences a continuity of Gondwanan eSZ terrestrial areas with the Laurasian ones. The occurrence of Xenoxylon within this Late Jurassic wood flora suggests an abundant water supply under a relatively cool/humid climate. Overall, both floras show important similarities to contemporary fossil wood flora from Iran and, to a lesser extent, to those from Georgia.
Key words: Agathoxylon, Protelicoxylon, Xenoxylon, fossil wood, Jurassic, Anatolia, Turkey.
Ünal Akkemik [uakkemik@istanbul.edu.tr], İstanbul University-Cerrahpaşa, Faculty of Forestry, Department of Forest Botany, 34473 Bahçeköy, İstanbul, Turkey.
Raif Kandemir [raif.kandemir@erdogan.edu.tr], Recep Tayyip Erdoğan University, Engineering and Architecture Faculty, Department of Geological Engineering, 53100, Fener, Rize, Turkey.
Marc Philippe [marc.philippe@univ-lyon1.fr], Univ Lyon, Université Claude Bernard Lyon 1, CNRS, ENTPE, UMR 5023 LEHNA, F-69622, Villeurbanne, France.
Yıldırım Güngör [yildirimgungor@gmail.com], İstanbul University-Cerrahpasa, Faculty of Engineering, Department of Geological Engineering, İstanbul, Turkey.
Fatih Köroğlu [fkoroglu1985@gmail.com], Ankara University, Faculty of Engineering and Graduate School of Natural and Applied Sciences, Department of Geological Engineering, Ankara, Turkey.
Received 15 November 2021, accepted 20 January 2022, available online 17 August 2022.
Copyright © 2022 Ü. Akkemik et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
The Lower–Middle Jurassic volcano-sedimentary successions (Olurdere Formation in Olur region and Şenköy Formation in Gümüşhane region, NE Turkey) of the eastern Sakarya Zone (eSZ) (Yilmaz 1985; Kandemir 2004) are critical for understanding the tectonics, palaeogeography, and palaeobiogeography of eSZ, which at that time was located at the northern rim of the Neotethys Ocean. The fossil wood fragments found in the eSZ could be considered as one of the most important indicators for the presence of terrestrial and coastal environments at that time. The fossil wood fragments found from the Middle–Late Jurassic-aged units are mainly conifer woods (e.g., Fakhr and Marguerier 1977; Philippe et al. 2006, 2019; Oh et al. 2015), and yielded important information to understand palaeogeography and palaeobiogeography of terrestrial environments. Oh et al. (2015) and Philippe et al. (2019) demostrated that both Protelicoxylon and Xenoxylon are strictly Laurasian genera, and the nearest floras containing representatives of these genera are known from the Middle Jurassic of Iran (Fakhr and Marguerier 1977; Philippe 1995). Protelicoxylon and Xenoxylon were also identified in the Liassic (Lower Jurassic) of north-eastern France (Philippe 1995).
In Turkey, many palaeobotanical studies including a large diversity of woody species were performed mainly on fossil woods from Eocene to Miocene (e.g., Akkemik et al. 2016, 2017, 2018, 2019, 2020, 2021) while information on Jurassic woods remained rather restricted. The wood occurrences in the Gümüşhane and Erzurum regions have not yet been properly investigated though it was known that they may have some palaeobiogeographical potential (Philippe et al. 2006, 2017). Some fossil wood fragments from the Gümüşhane-Akçakale region were previously described by Kandemir (2004) as species of Agathoxylon. Additional three wood specimens collected by Nurdan Yavuz (Gümüşhane, Turkey) from Gümüşhane locality in Bayburt region, and later sent to Erdei Boglarka (Budapest, Hungary), have been also assigned to Agathoxylon by Philippe et al. (2006). The palynoflora of the wood-bearing Middle Jurassic Şenköy Formation has been studied by Akgün et al. (2006) who identified a presence of a mix of dry coastal cheirolepidiacean and wet fern lowland vegetation.
Additionally, Kutluk et al. (2012), identified Agathoxylon sp. in wood fragments from the Oltu Gemstone of Erzurum and dated them as Late Cretaceous. According to Bozkuş (1992) and Kara-Gülbay et al. (2018), however, the age of this material is most likely the earliest Late Jurassic.
This study constitutes the first detailed study of Jurassic wood floras from Gümüşhane and Erzurum regions of northeastern Anatolia. The purpose of the present study is to define the taxonomic identity of the Jurassic woods and to discuss their palaeobiogeographical and palaeocological implications.
Institutional abbreviations.—CNRS, Centre National de la Recherche Scientifique, Paris, France; ISTO, Istanbul University, Faculty of Forestry Herbarium, Turkey.
Other abbreviations.—DUT, Dutlu village of the city of Oltu in the province of Erzurum, Turkey; eSZ, Sakarya Zone; GED, the site of Edire in the province of Gümüşhane, Turkey; GUM, the Beyçam district of the province of Gümüşhane, Turkey; YES, Yeşilbağlar village of the city of Oltu in the province of Erzurum, Turkey.
Geological setting
Fossil woods were collected from Gümüşhane and Oltu-Olur (Erzurum) regions. Both regions are located in the eastern Sakarya Zone, NE Turkey (Fig. 1A). Locations, geological backgrounds of the regions and stratigraphic positions of the samples are given in Figs. 1B, C, 2.
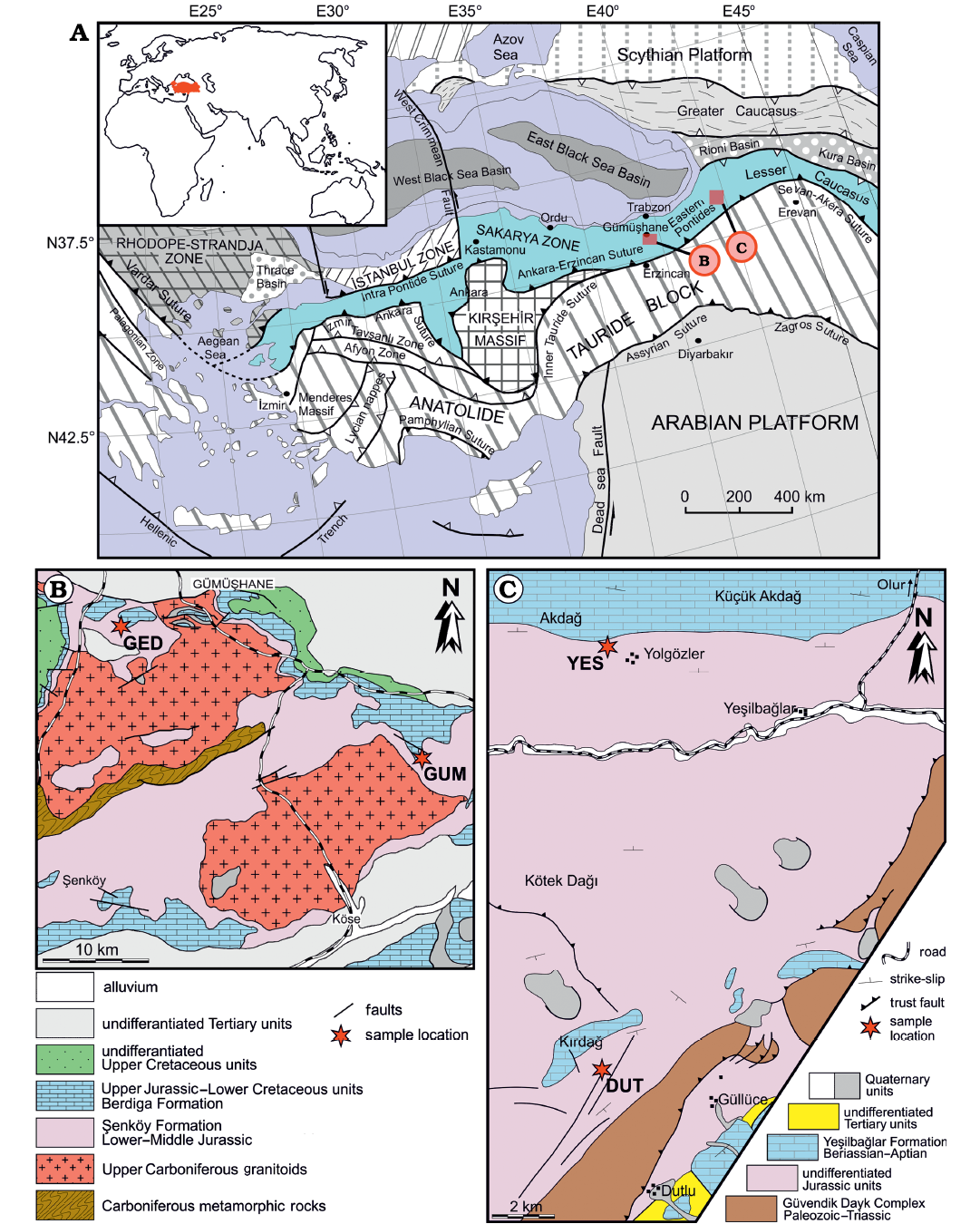
Fig. 1. A. Geographic location (inset) and regional tectonic setting of Turkey (after Okay and Tüysüz 1999). B, C. Simplified geological map of the Gümüşhane region (B, after Kandemir 2004), the Olur-Oltu region (C, after Konak and Hakyemez 2008) and location of the Jurassic wood sites.
The eastern Sakarya Zone (eSZ) in Turkey experienced a complex Mesozoic geodynamic evolution, which is not yet fully understood. The Early–Middle Jurassic period in the eSZ was characterized by the presence of a rift system which resulted in rock units of variable lithology and facies. The Lower–Middle Jurassic volcano-sedimentary successions (Olurdere Formation in Olur region and Şenköy Formation in Gümüşhane region; Yilmaz 1985; Kandemir 2004) are regarded as critical for understanding the tectonic, palaeogeography and palaeobiogeography of eSZ, which at that time was at the northern rim of the Neotethys Ocean.
These sequences are generally known as rift-related deposits in an intra-arc or a back-arc setting along the southern margin of Paleotethys. This rifting caused the opening of Neotethys in the south of Cimmerian Continent (Şengör and Yilmaz 1981; Görür et al. 1983; Kandemir 2004; Dokuz and Tanyolu 2006; Dokuz and Sünnetçi 2019; Kandemir et al. 2021). However, some researchers have attributed the deposition of these successions to an extensional basin of the fore-arc tectonic setting (Ustaömer and Robertson 2010; Okay et al. 2014; Akdoğan et al. 2018). Although there are different tectonic models of the evolution of Early–Middle Jurassic basins in the eSZ, the existence of terrestrial and coastal environments in the Jurassic have been agreed upon in all palaeogeographic reconstructions (Golonka 2004; Barrier et al. 2018; Torsvik 2019; Hinsbergen et al. 2020). The fossil woods from the eSZ, NE Turkey described herein present a direct evidence on the presence of terrestrial environments at that time.
Gümüşhane region.—The samples from Gümüşhane region were collected close to the contact between Berdiga and Şenköy formations (Fig. 2: GED and GUM). The Şenköy Formation is dated as Hettangian/Sinemurian–Bathonian and rest on a heterogeneous Paleozoic basement (Kandemir 2004).
The Şenköy Formation is ~2200 m thick and shows vertical and lateral facies changes within a short distance (Fig. 2). Facies variations resulted from the blocky-faulted topography during the deposition of its sediments (Kandemir and Yilmaz 2009). Kandemir (2004) also emphasized that the Şenköy Formation may represent typical rift-related sediments. This topographic differentiation is exemplified by the Gümüşhane section (see Fig. 2). The formation consists of basaltic and andesitic lithic tuffite, volcanogenic sandstone, shale, basaltic and andesitic lavas, conglomerate (Kandemir 2004; Dokuz and Tanyolu 2006) and Ammonitico Rosso limestone horizons (Kandemir and Yilmaz 2009). The uppermost part of the Şenköy Formation is dominated by shale, claystone, and sandstone interbedded with coals.
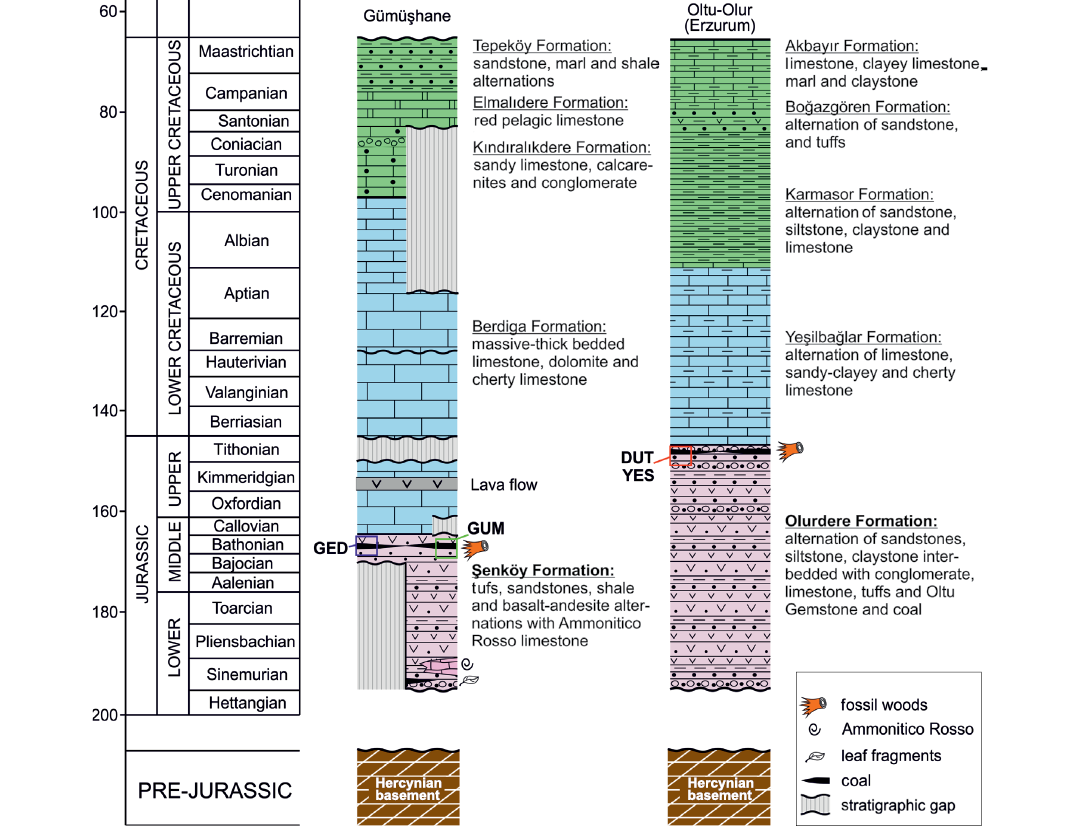
Fig. 2. Generalized columnar section of the studied regions (Gümüşhane section modified from Vincent et al. 2018; Olur-Oltu section modified from Konak and Hakyemez 2008 and Kara-Gülbay et al. 2018). Abbreviations: DUT, Dutlu village of the city of Oltu in the province of Erzurum, Turkey; GED, the site of Edire in the province of Gümüşhane, Turkey; GUM, the Beyçam district of the province of Gümüşhane, Turkey; YES, Yeşilbağlar village of the city of Oltu in the province of Erzurum, Turkey. Triassic rocks are not observed in the Eastern Sakarya zone. So we left this section blank in the figure.
Akgün et al. (2006) showed that the palynomorph assemblage from the upper part of the Şenköy Formation is typical of the Bathonian (Middle Jurassic) and is characterized by the presence of the dinoflagellate cyst Ctenidodinium continuum, miospores Matonisporites equiexinus, Dictyophyllidites harrisii, Gleicheniidites senonicus, Cerebropollenites mesozoicus, and bisaccate pollen as well as the common pollen of Cyathidites minor and Classopollis torosus. The Middle Jurassic palynoflora of the upper part of the Şenköy Formation indicates the existence of locally wetlands, coastal conditions and a hot and temporarily dry subtropical-tropical climate. The Şenköy Formation is commonly conformably overlain by Upper Jurassic–Lower Cretaceous Berdiga Formation (Pelin 1977) in the Gümüşhane region largely characterized by platform type carbonates.
The Beyçam wood specimens (GUM) were collected from the lowest part of the Berdiga Formation in the Gümüşhane region. Although this formation has been extensively studied (Kirmaci et al. 1996; Tasli et al. 1999; Koçyiğit and Altiner 2002; Koch et al. 2008; Vincent et al. 2018) its age, however, remains poorly constrained, largely because of the paucity of biostratigraphically important fossils. The authors have variously suggested that its sedimentation commenced during the Aalenian–Bajocian or Callovian (Middle Jurassic; Pelin 1977; Kirmaci 1992; Robinson et al. 1995) or during the Oxfordian or Kimmeridgian (Late Jurassic; Koch et al. 2008; Tasli et al. 1999; Dokuz and Tanyolu 2006). Detrital zircons from the Şenköy Formation (Akdoğan et al. 2018), close to the Berdiga Formation type section near Alucra, more precisely constrain a latest Oxfordian or younger depositional onset age for the formation in this region.
The Edire wood specimens (GED) were collected from Şenköy Formation in the Gümüşhane province. These fossil wood-bearing deposits correspond to the upper levels of the Şenköy Formation. Kandemir (2004) determined their age as Bathonian (Middle Jurassic) based on the pollen and spores in these deposits at the Gümüşhane in Şenköy section. In the Edire region the sediments are mostly represented by matrix and sometimes grain supported conglomerate and sandstone with lenticular structure. These conglomerate and sandstone levels have no lateral continuity. The pebbles are well rounded but poorly sorted and their origin is volcanic. Above this terrigenous level the sediments mostly consist of clay and silt-grained rocks. These include organic-rich dark layers in their intermediate levels. These layers supported coal mining in the 18th century. Fossil wood fragments were found in lateral equivalents of these coal-bearing levels. Fossil wood fragments have different sizes and are scattered at this level. Above these levels, there are red-colored, extremely brittle and weathered gravel beds of volcanoclastic rocks. Cross-bedding is observed at some levels in these volcaniclastics. The volcaniclastics pass upward into clayey-marly deposits. The sequence is overlain by the cherty limestone beds of the Berdiga Formation (Kandemir 2004). The wood-bearing sequence of the Şenköy Formation may have been deposited in a terrestrial environment and is commonly interpreted as marsh deposit. The presence of fossil wood fragments, coal-bearing levels and sedimentological properties of the sediments suggest that the depositional environment of the Şenköy Formation was terrestrial in the Edire region.
Olur-Oltu region.—In Erzurum region, the samples were collected from the upper levels of the Jurassic sequence defined as the Olurdere Formation (Fig. 2). The sampling sites are in Dutlu village of the city of Oltu in the province of Erzurum (DUT) and Yeşilbağlar village of the city of Oltu in the province of Erzurum (YES). The Jurassic strata rest on Palaeozoic–Triassic gneiss and metabasics intruded by various veins and dykes (Fig. 1C) and are represented mostly by undifferentiated volcano-sedimentary rocks. This Jurassic unit is covered by the Olurdere Formation (Figs. 1C, 2) yielding wood fragments, which are known locally as Oltu Gemstone. The formation mainly consists of claystone, sandstone, conglomerate intercalations and volcanics with thin coal bands and Oltu Gemstone occurrences (Fig. 2). The Olurdere Formation is concordantly overlain by the Upper Jurassic–Lower Cretaceous Yeşilbağlar Formation (Yilmaz 1985; Bozkuş 1992). The Formation mainly consists of thin bedded, white and grey colored, micrite and clayey micritic limestone with chert bands and chert concretions in its upper part (Figs. 1C, 2).
Kara-Gülbay et al. (2018) showed that Oltu Gemstone coal consists of several lenses of only a few meters of lateral extension each and 50 cm of maximum thickness within the Lower–Upper Jurassic Olurdere Formation in Erzurum region. These lenses were locally mined for the Oltu Gemstone resulting in numerous small galleries developed along their exposure on the southern flanks of Dutlu Mountain (Fig. 1C). Kara-Gülbay et al. (2018) suggested that Oltu Gemstone coal was deposited in an environment changing from shallow marine to deep shelf where volcanism was effective and coals were formed by deposition of organic matter of mainly terrestrial type under anoxic conditions in an environment which occasionally changed to ephemeral swamps with resin producing trees. Ağirman Aktürk (2016) suggested that Olurdere Formation is characterized by the conglomerate, sandstone, mudstone, siltstone as well as thin coal seams (Oltu Stone) alternations and describe the depositional environment of the formation as a fan delta.
Material and methods
The fossil wood fragments were taken from four different sites (Figs. 1, 2; Table 1). These are the Beyçam district of the province of Gümüşhane (GUM), the site of Edire also in the province of Gümüşhane (GED), Dutlu village of the city of Oltu in the province of Erzurum (DUT) and Yeşilbağlar village of the city of Oltu in the province of Erzurum (YES). All 27 fossil wood specimens have been coded as ISTO-FW-00217 to ISTO-FW-0043 and identified taxonomically.
Table 1. The identified fossil species from the Jurassic of eastern Anatolia, Turkey.
|
Identification |
Family |
Specimen |
Locality |
|
Agathoxylon sp. (type 1) |
Araucariaceae |
ISTO-FW-00217 |
Beyçam district of the province of Gümüşhane |
|
ISTO-FW-00218–00226 |
Edire in the province of Gümüşhane |
||
|
Agathoxylon sp. (type 2) |
ISTO-FW-00227–00230 |
Dutlu of the city of Oltu in the province of Erzurum |
|
|
ISTO-FW-00231–00237 |
Beyçam district of the province of Gümüşhane |
||
|
ISTO-FW-00238 |
Edire in the province of Gümüşhane |
||
|
ISTO-FW-00239–00241 |
Yeşilbağlar of the city of Oltu in the province of Erzurum |
||
|
Protelicoxylon asiaticum (Serra, 1969) Philippe, 1995 |
Taxaceae |
ISTO-FW-00242 |
|
|
Xenoxylon hopeiense Chang, 1929 |
unknown |
ISTO-FW-00243 |
Three thin sections were prepared from each fossil specimen in transversal, tangential longitudinal and radial longitudinal orientations. For tracheid and ray dimensions, at least 30 measurements were performed. As all woods belong to conifers, we followed IAWA criteria for gymnosperm woods (IAWA Committee 2004) and Philippe and Bamford (2008) for terminology and generic identifications. For comparison and species descriptions we used the following references: Philippe (1995), Philippe et al. (2006, 2013, 2019), Bamford and Philippe (2001), Philippe and Bamford (2008), Kloster and Gnaedinger (2018), and Boura et al. (2021).
Systematic palaeontology
Order Pinales Dumortier, 1829
FamilyAraucariaceae Henkel and Hochstetter, 1865
Genus Agathoxylon Hartig, 1848
Type species: Agathoxylon cordaianum Hartig, 1848; Coburg, Germany; Keuper, Upper Triassic.
Stratigraphic and geographic range.—Middle Jurassic for Şenköy Formation and Berdiga Formation, and Upper Jurassic for Olurdere Formation. Agathoxylon is widely distributed worldwide throughout the Jurassic.
Agathoxylon sp.
Figs. 3, 5.
Material.—Twenty-six samples (ISTO-FW-00217–00241) from Jurassic of Turkey localities GUM, GED, DUT, and YES (see Table 1 for details).
Description.—The silicified wood fragments of at least 8 cm in diameter. Transition from the earlywood to the latewood gradual and indistinct. Growth ring borders distinct, with 1–5 rows of narrowed latewood tracheids, or indistinct. Tracheid sections circular to polygonal. Tangential and radial diameters of earlywood tracheids 49 (23–93) μm and 55 (18–92) μm, respectively. These are 31 (8–67) μm and 22 (8–57) μm in the latewood. Double cell wall thickness 19 (6–39) µm in the earlywood, and 20 (8–35) µm in the latewood. Resin canals and axial parenchyma not observed (Figs. 3A, B, 5A).
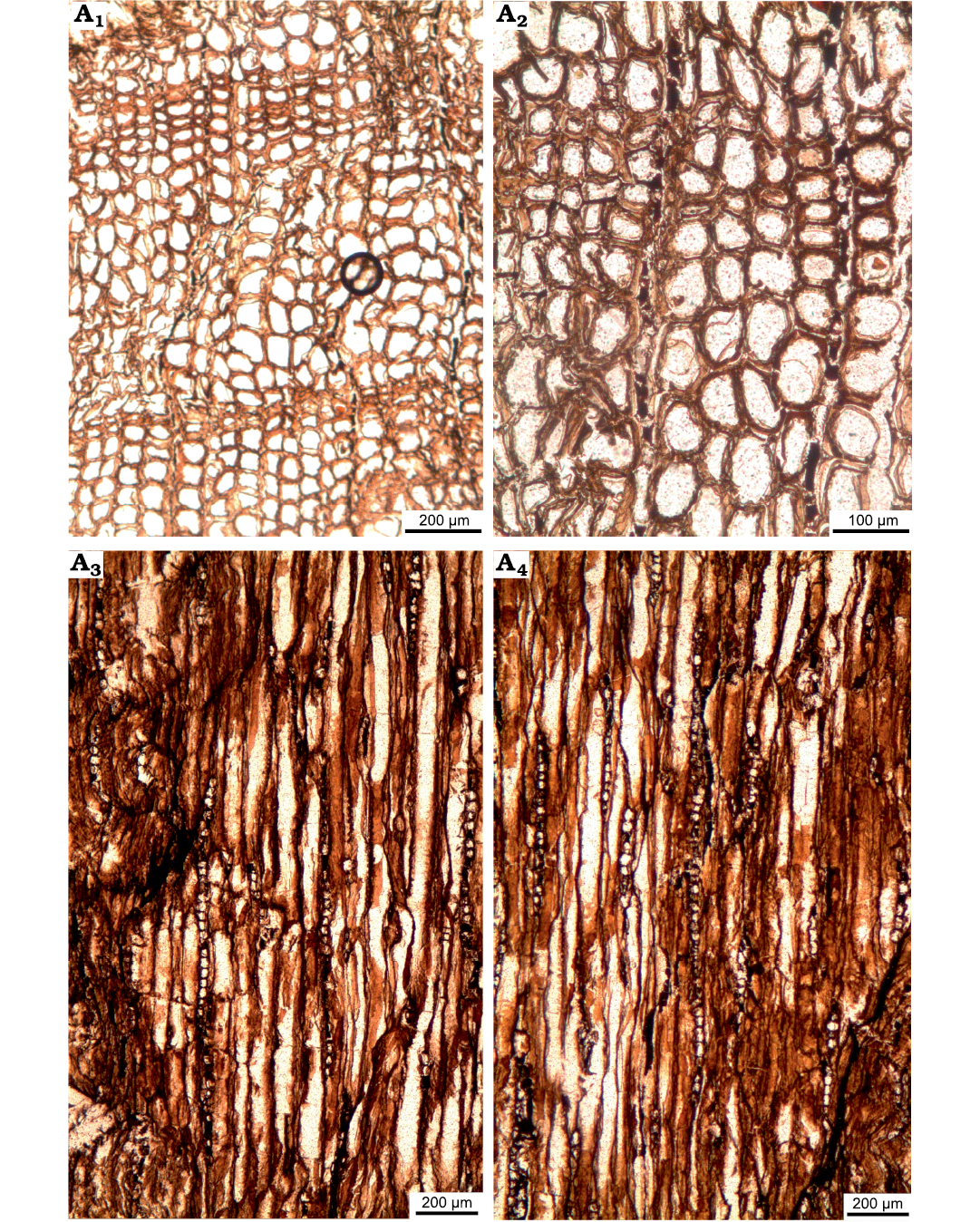
Fig. 3. Araucariacean wood sections of Agathoxylon sp. type 1 (ISTO-FW-00217) from Jurassic of the Beycam locality of the province of Gümüshane, Turkey. A1, A2, transversal sections with distinct growth ring boundary; A3, A4, tangential sections with high rays.
Rays uniseriate, rarely partly biseriate for some levels. Ray heights 10–20 (1–45) cells, 348 (50–1240) μm, ray widths 34 (14–58) μm, and 4–5 (3–7) rays per mm in type 1 (Fig. 3C, D), and average 2–6 cells in type 2 (Fig. 5B). Intertracheary pits on tangential walls, helical thickenings and axial parenchyma not observed (Figs. 3C, D, 4). Intertracheary pits on tangential walls present in type 2 (Fig. 5C).
Intertracheary pits on radial walls uni, bi-, or triseriate, contiguous, araucarian, predominantly vertically flattened to circular and rarely hexagonal. Heights of biseriate intertracheary pits on radial walls are 16 (12–20) μm, and their widths are 21 (12–33) μm, those in triseriate pits are slightly smaller, 15 (13–17) μm and 16 (12–20) μm, respectively. Rims of Sanio absent. Cross-field pits of the cupressoid type, 6 (4–9) μm in diameter. Cross-field araucarioid, with 2–13 pits per cross-field. Crystals absent (Figs. 4A–E, 5D–F).
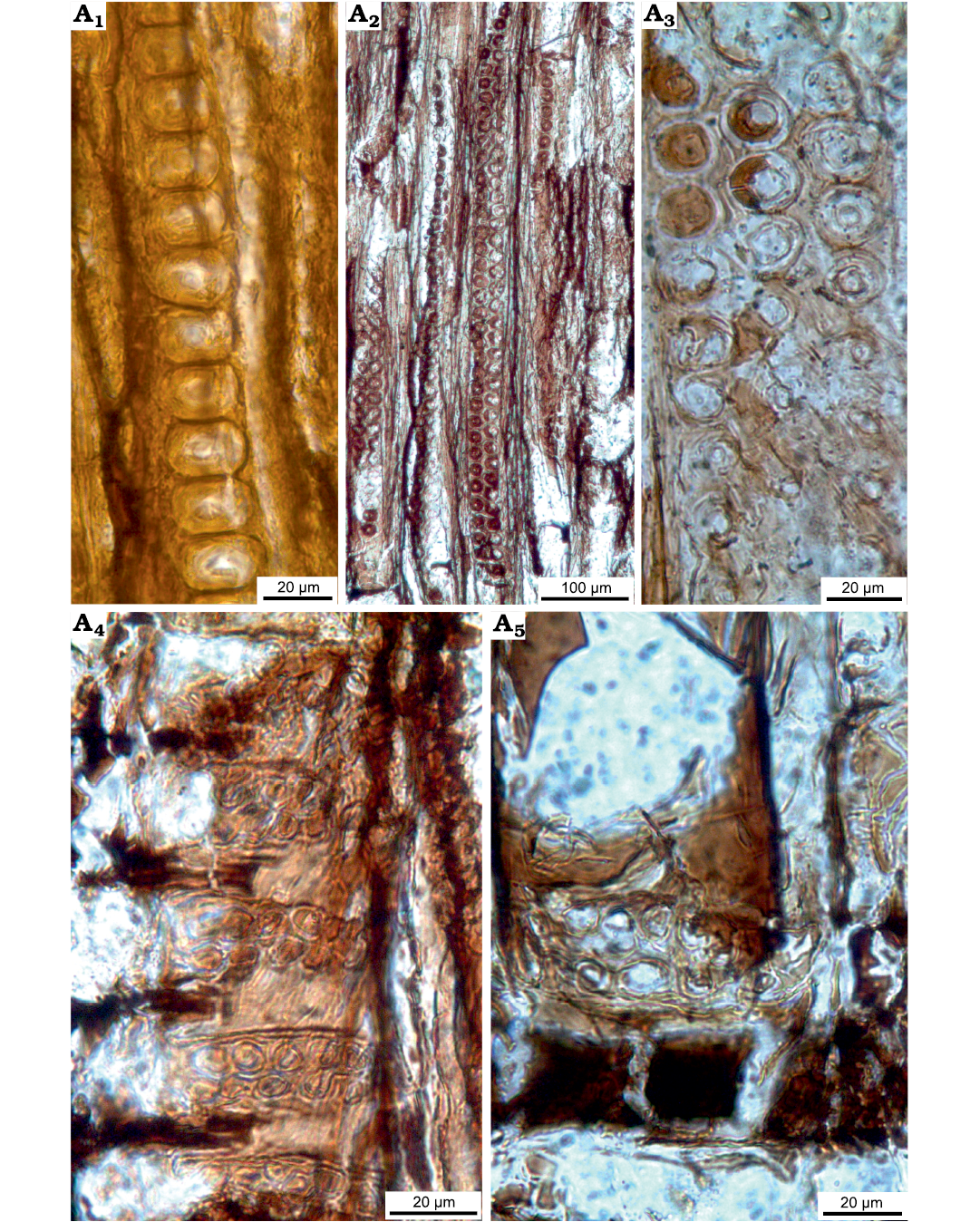
Fig. 4. Araucariacean wood sections of Agathoxylon sp. type 1 (ISTO-FW-00217) from Jurassic of the Beycam locality of the province of Gümüshane, Turkey. A1–A3, uni- to triseriate of intertracheary radial pits; A4, A5, predominantly araucariod type cross-field pits.
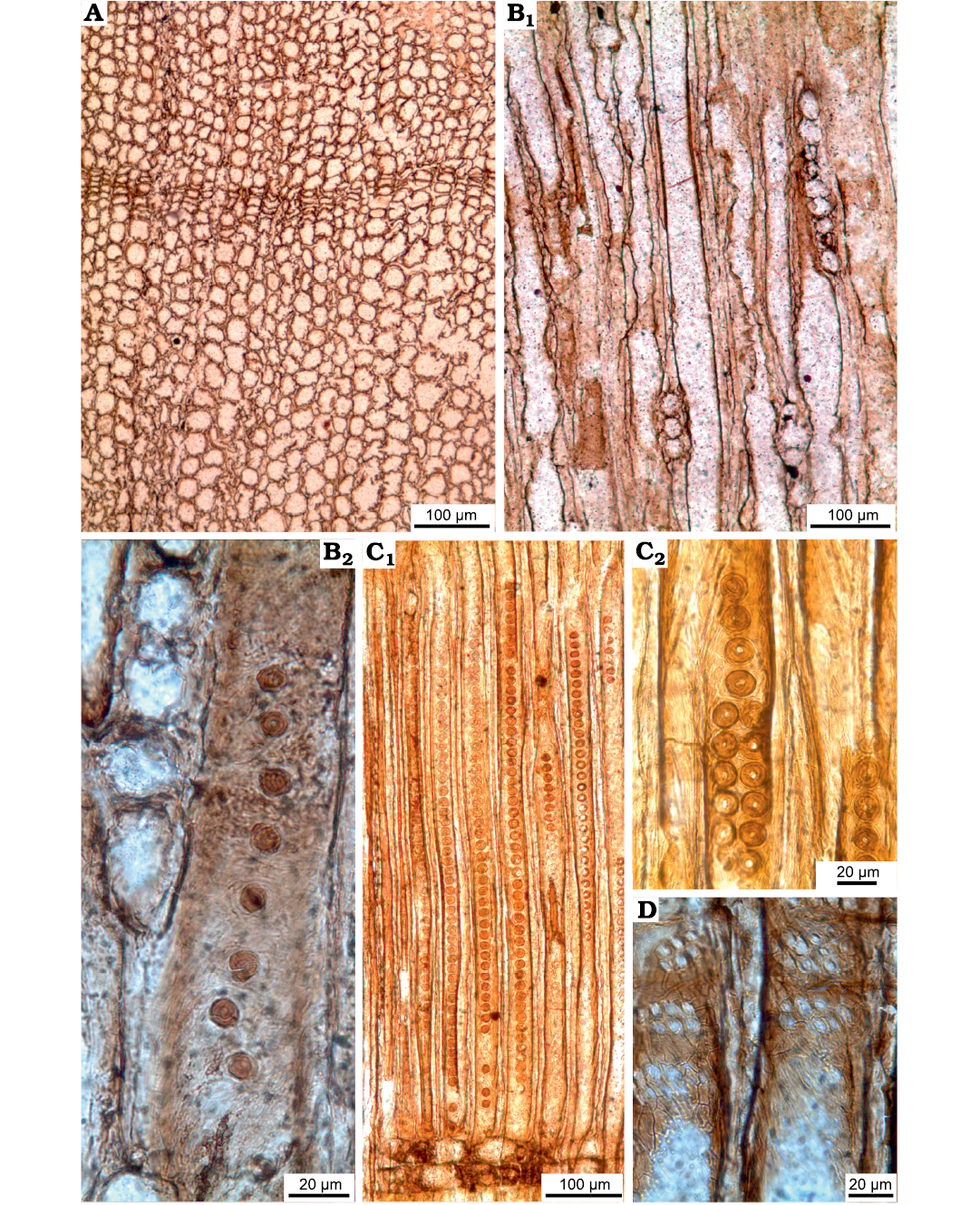
Fig. 5. Araucariacean wood sections of Agathoxylon sp. type 2 from Jurassic of the Beycam locality of the province of Gümüshane, Turkey. A. ISTO-FW-00235, transversal sections with distinct growth ring boundary. B. ISTO-FW-00234, short rays in tangential section (B1), intertracheary pits on tangential walls (B2). C. ISTO-FW-00239, predominantly uniseriate of intertracheary radial wall pits (C1, C2). D. ISTO-FW-00232, araucarioid type cross-field pits.
Remarks.—The features of the fossil wood, which are uni-, bi- or triseriate of araucarian type radial intertracheary pits, and araucarioid type of cross-field pits with numerous cupressoid oculipores are those of Agathoxylon (Philippe and Bamford 2008).
Numerous species of Agathoxylon were described from the Mesozoic (e.g., Torres and Philippe 2002; Poole and Ataabadi 2005; Zamuner 1996; Zamuner and Falaschi 2005; Kutluk et al. 2012; Pujana et al. 2015). In recent years, a detailed comparison of Gondwanan Agathoxylon woods of Permian to Paleogene age has been provided by Kloster and Gnaedinger (2018) who suggested that these woods could be classified into groups on the basis of the shape and seriation of the intertracheary pits as well as the number of pits per cross-field. Beyond the relevance of the prevalence of geographic logic over stratigraphic logic, this work is based on a discretization of the shapes and seriation of the intertracheary pits that may seem confusing to anyone accustomed to the variability of these characters, in present-day wood as well as in fossil wood, coupled with the taphonomic bias that can profoundly modify the appearance of the pits. Shape and seriation of the pits as well as their number per cross-field are not continuous variables, and any systematics is only a combination of the recognized modalities (Philippe 2011; Booi et al. 2014). Attempts at multivariate analysis are of limited interest because they overlook that most xylological variables are sub-discrete variables, which can only take on a limited number of values.
At present more than 463 species of Agathoxylon were identified, of which the generic assignment of more than 240 can be confirmed as correct from their protologue. Most of these species are rather similar to each other, and therefore a comprehensive revision is needed. It is unlikely that any taxonomic approach could create taxa within Agathoxylon that have biological, palaeoecological, or palaeobiogeographical significance. For that reason, we prefer to leave these specimens in open nomenclature, as Agathoxylon sp.
The fossil woods identified as Agathoxylon in this study can be divided into two types. In the first type (ISTO-FW- 00217–00226), the woods have higher rays (average 10–20 cells) and 1–3 seriate of intertracheary radial walls pits (Figs. 3–5). In the second type (ISTO-FW-00227–00241), the woods have shorter rays (average 2–6 cells) and predominantly uniseriate of intertracheary radial wall pits (Fig. 5).
Family Taxaceae Gray, 1821
Genus Protelicoxylon Philippe, 1995
Type species: Protelicoxylon feriziense (Fahkr and Marguerier, 1977) Philippe, 1995; Iran; Middle Jurassic.
Protelicoxylon asiaticum (Serra, 1969) Philippe, 1995
Figs. 6–8.
Material.—One silicified wood fragment numbered as ISTO-FW-00242 from Jurassic of Turkey, locality Yeşilbağlar of the city of Oltu in the province of Erzurum. It is deposited at the Department of Forest Botany, Faculty of Forestry, Istanbul University-Cerrahpasa, Istanbul, Turkey.
Description.—Growth ring boundaries distinct, with 2–4 rows of flattened latewood tracheids. Transition from earlywood to latewood gradual. Tracheid cross sections circular to polygonal (Fig. 6A, B). Tangential and radial widths of tracheids are 27 (18–36) µm and 23 (15–32) µm in the earlywood, and 22 (16–27) µm and 6 (3–10) µm in latewood, respectively. Double wall thicknesses are 8 (5–10) µm in earlywood and 4 (2–7) µm in latewood, respectively. Axial resin canals absent (Fig. 6A, B).
Rays exclusively uniseriate, very rarely partly biseriate for some levels. Rays 3–7 (1–10) cells and 104 (32–217) µm high, and 28 (17–46) µm wide (Fig. 6C, D). Axial parenchyma present (Fig. 7A, B). Intertracheary pits present, sparse and uniseriate on tangential walls (Fig. 7 C). Rays 7 (5–9) per mm. Spiral thickenings common, often paired or even grouped by three, thin (1–4 µm wide), with a low angle to the horizontal (30°) (Fig. 7D, E). A few isolated or paired callitroid thickening were observed.
Rays homogenous (Fig. 7F). Cross-fields are araucarian, with 3–13 cupressoid pits per cross-field. Diameters of cross-field pits are 4 (3–6) µm. Intertracheary pits on radial walls predominantly araucarian and uniseriate, very rarely biseriate, and then opposite (abietinean). The radial intertracheary pitting is of the mixed-type. Diameters of intertracheary pits on radial walls are 14 (11–18) µm in diameter (Figs. 7F, 8A–D).
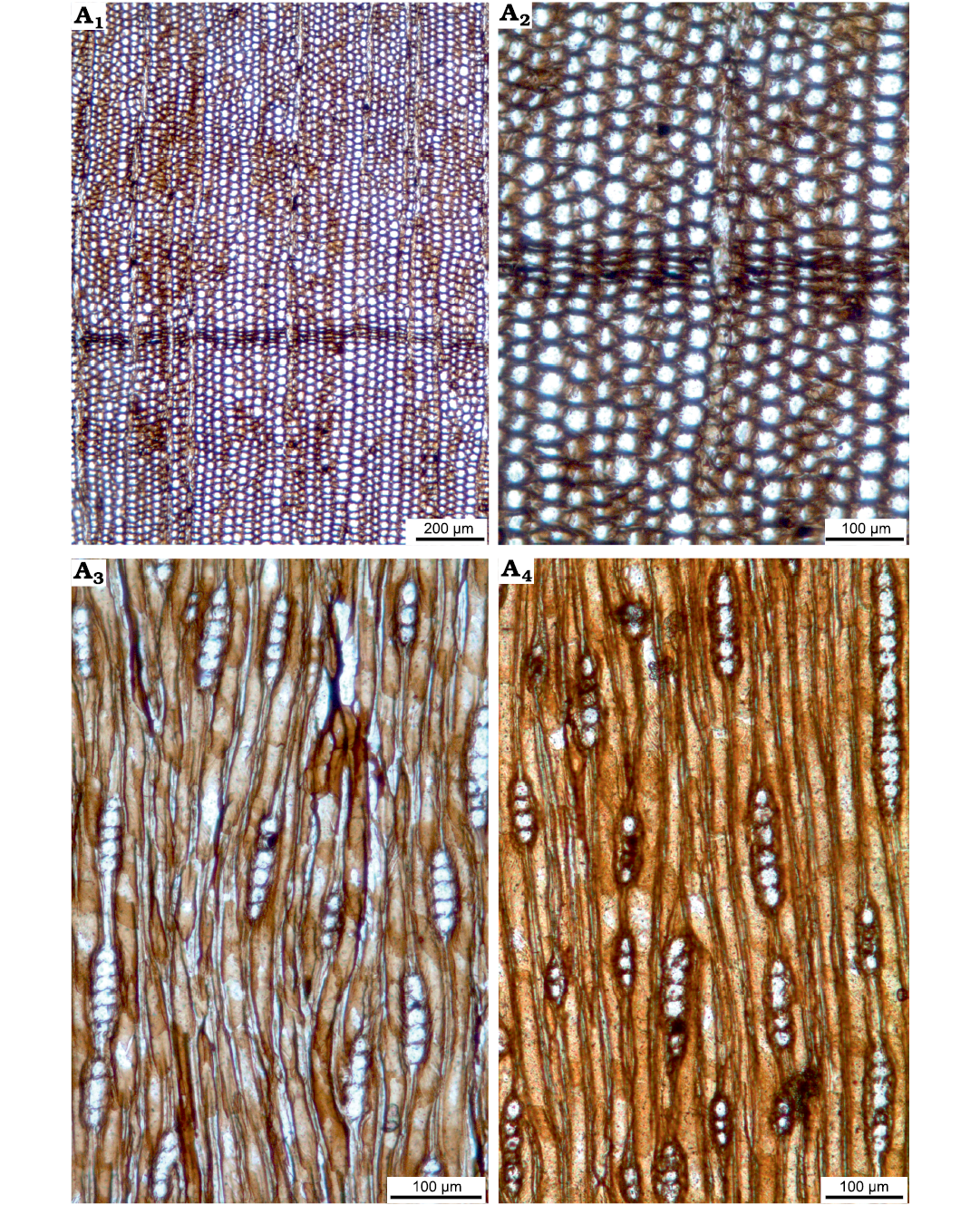
Fig. 6. Taxacean wood sections of Protelicoxylon asiaticum (Serra, 1969) Philippe, 1995 (ISTO-FW-00242) from Jurassic of the Yesilbaglar locality of the province of Erzurum, Turkey. A1, A2, transversal sections with distinct growth ring boundary; A3, A4, tangential sections with predominantly uniseriate and short rays.
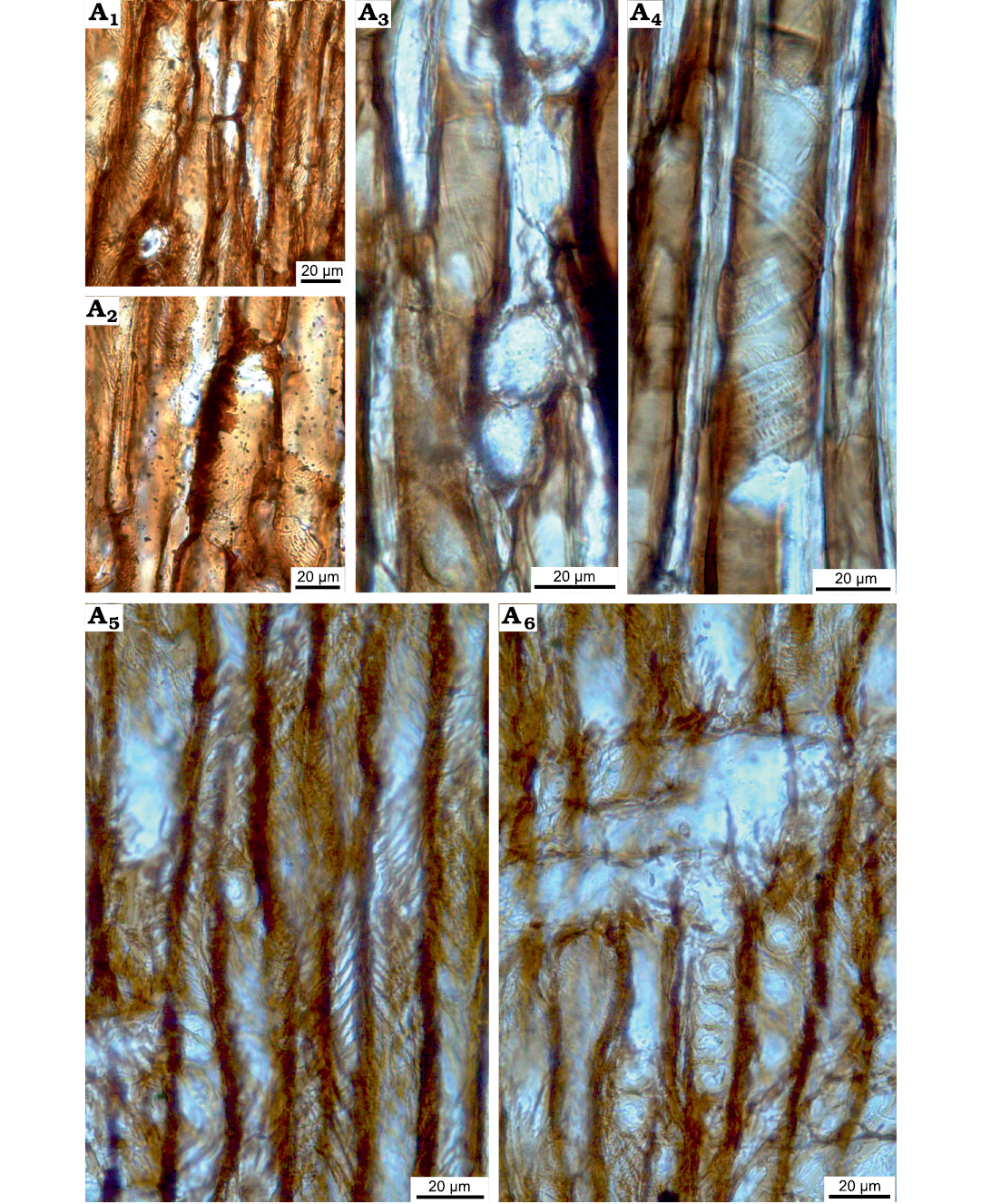
Fig. 7. Taxacean wood sections of Protelicoxylon asiaticum (Serra, 1969) Philippe, 1995 (ISTO-FW-00242) from Jurassic of the Yesilbaglar locality of the province of Erzurum, Turkey. A1, A2, axial parenchyma in tangential sections; A3, intertracheary pits in tangential section; A4, A5, spiral thickening on tracheid walls; A6, homogenous ray, araucarian and uniseriate intertracheary pits on radial walls.
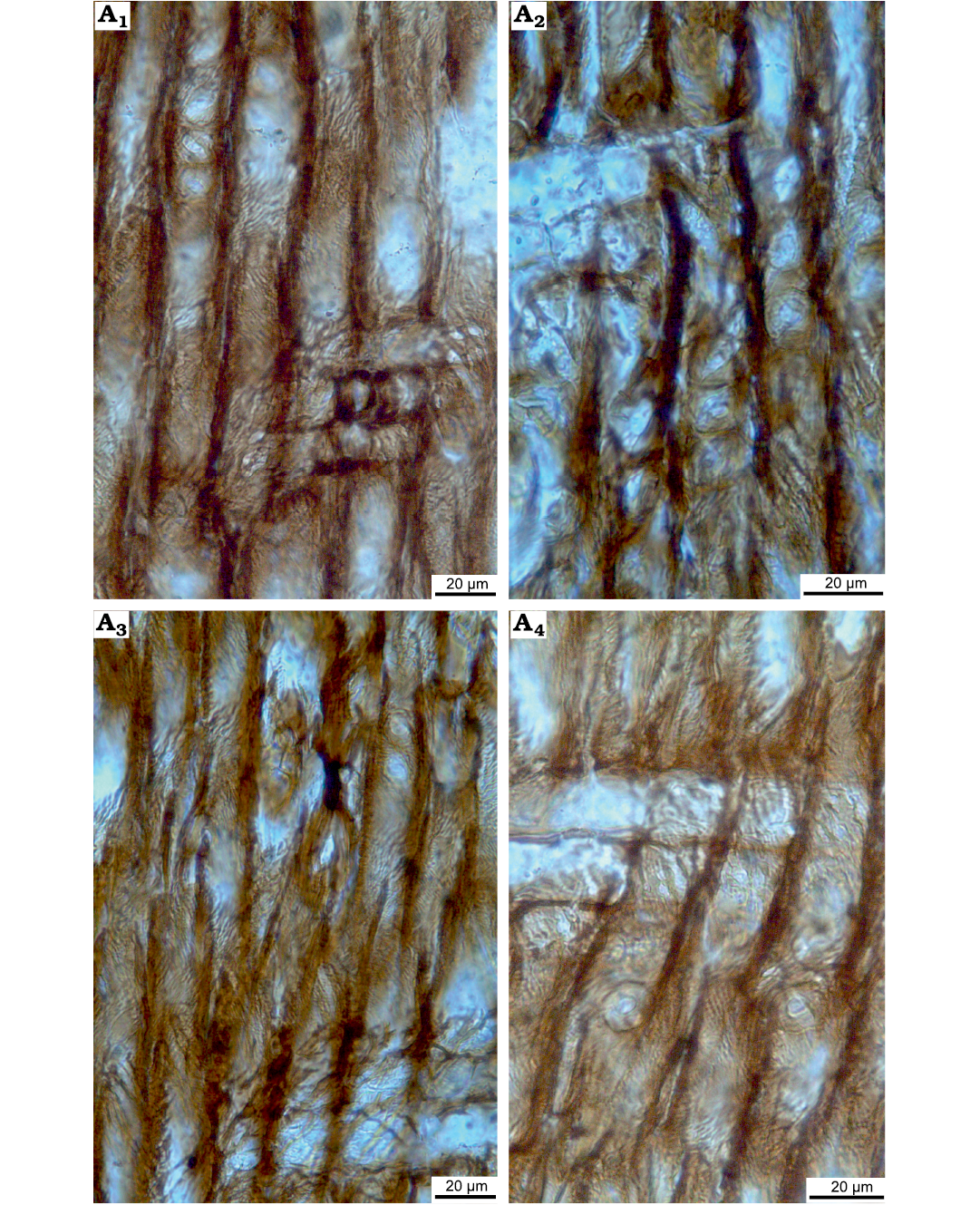
Fig. 8. Taxacean wood sections of Protelicoxylon asiaticum (Serra, 1969) Philippe, 1995 (ISTO-FW-00242) from Jurassic of the Yesilbaglar locality of the province of Erzurum, Turkey. A1–A4, homogenous ray, araucarian and uniseriate intertracheary pits on radial walls; A1, A3, A4, araucarian cross-fields pits with 3-13, each of them cupressoid in cross-fields.
Remarks.—Protelicoxylon Philippe, 1995, is a genus designed for woods that are characterized by mixed-type pitting on the radial walls of the tracheids and different types of tertiary thickenings: spirals, crassulae or thin horizontal bars. The genus has cross-fields with cupressoid to dacrydioid pits and might features axial parenchyma. This genus was recently reviewed by Philippe (1995) (Table 2).
Our wood is most similar to Protelicoxylon asiaticum (Serra, 1969) Philippe, 1995 in having predominantly araucarian type of radial intertracheary pitting, presence of axial parenchyma, 2–5 cupressoid oculipores per field, and presence of callitroid thickenings (Table 3). The wood described by Nadjafi (1982), from the Middle Jurassic of Elburz (Iran) is also strikingly similar to our material.
Table 2. Fossil wood species assigned to the Protelicoxylon (adapted from Philippe et al. 2019). The name Prototaxoxylon persicum was mentioned in an unpublished thesis (Nadjafi 1982) and is not validly published.
|
Taxon name, authority |
Reference |
Age |
Country |
|
Protelicoxylon asiaticum (Serra, 1969) Philippe, 1995 |
Early Cretaceous |
Cambodia |
|
|
Protelicoxylon feriziense (Fakhr and Marguerier, 1977) Philippe, 1995 |
Middle Jurassic |
Iran |
|
|
Protelicoxylon lepennecii Philippe, 1995 |
Early Jurassic (Toarcian) |
France |
|
|
Protelicoxylon parenchymatosum (Greguss, 1967) Philippe, 1995 |
Early Jurassic (Toarcian) |
Hungary |
Table 3. Comparison of the published fossil species of Protelicoxylon with our wood identified as Protelicoxylon asiaticum (Serra, 1969) Philippe, 1995 (ISTO-FW-00242) (adapted from Gnaedinger 2007).
|
Country, age |
Vietnam, |
Iran, |
France, |
Hungary, |
Turkey, |
|
Species |
Protelicoxylon asiaticum (Serra, 1969) Philippe, 1995 |
Protelicoxylon feriziense (Fahkr and Marguerier, 1977) Philippe, 1995 |
Protelicoxylon lepennecii Philippe, 1995 |
Protelicoxylon parenchymatosum (Greguss, 1967) Philippe, 1995 |
Protelicoxylon
asiaticum |
|
Radial pitting |
uniseriate (exceptionally
biseriate), araucarian, rarely abietinean |
uniseriate (exceptionally
biseriate), araucarian or |
predominantly |
uniseriate (exceptionally biseriate), araucarian, rarely abietinean (biseriate opposite) |
uniseriate (exceptionally
biseriate), araucarian, rarely abietinean |
|
Axial parenchyma |
present |
sparse |
abundant |
present |
present |
|
Cross fields |
2–5 cupressoid |
1 (rarely 2–3) |
2–5 cupressoid to dacrydioid oculipores |
1–2 (rarely
3) |
3–8 (rarely up to 13) cupressoid oculipores, araucarian |
|
Spiral thickenings |
present, often paired, |
present, often paired |
present |
present |
present, often paired or even by three |
|
Crassulae |
– |
? |
present |
present |
sometimes present |
|
Callitroid thickenings |
present |
present |
present |
present |
present |
Stratigraphic and geographic range.—Upper Jurassic for Olurdere Formation, Turkey from Southeast Asia (Vietnam) westward to Iran.
Family unknown
Genus Xenoxylon Gothan, 1905
Type species: Xenoxylon latiporosum (Cramer, 1868) Gothan, 1905; the exact locality and age unknown (Philippe and Cantrill 2007).
Xenoxylon hopeiense Chang, 1929
Figs. 9–11.
Material.—One silicified wood fragment coded as ISTO- FW-00243 from Jurassic of Turkey, locality Yeşilbağlar of the city of Oltu in the province of Erzurum. It is deposited at the Department of Forest Botany, Faculty of Forestry, Istanbul University-Cerrahpasa, Istanbul, Turkey.
Description.—Growth ring boundary faint but distinct with only 1–2 rows of flattened latewood tracheids. Axial resin canals absent. Transition from earlywood to latewood abrupt. Growth rings generally less than 0.5 mm wide generally. Tracheid cross-section hexagonal, rounded or rectangular (Fig. 9A, B). Tangential and radial widths of tracheids are 46.7 (30.2–57.6) µm and 56.9 (32.5–111.7) µm in the earlywood, and 27.9 (10.0–44.3) µm and 21.8 (14.2–32.5) µm in the latewood, respectively. Double wall thicknesses are 10.7 (7.7–14.2) µm in earlywood and 14.1 (9.1–20.8) µm in latewood, respectively.
Rays exclusively uniseriate, rarely partly biseriate and ray heights 5–15 (2–47) cells (314.8 [119.1–1218] µm, and their average width is 18.1 [12.9–27.0] µm) (Figs. 9C, 10A–C). Intertracheary pits present, sparse and uniseriate on tangential walls and their diameters are 11.0 (8.8–12.8) µm (Fig. 10D). Rays are 4–5 (3–7) per mm. Axial parenchyma not observed.
Rays homogenous. Intertracheary pits on radial walls uniseriate or in some cases biseriate opposite. Intertracheary pits locally xenoxylean, mostly strongly flattened, pit rows locally interrupted. Rims of Sanio commonly present between both uniseriate and biseriate pits. Heights of intertracheary pits are 17.2 (12.3–23.7) µm, their widths are 27.1 (20.0–33.3) µm, and the ratio of height to the width of pits is 0.64. Cross-fields feature predominantly one large fenestriform oopore in the earlywood, and the ray cells thin-walled. Very rarely two oopores may be seen in a single cross-field. Heights of oopores are 14.1 (9.3–19.2) µm, and their widths are 31.6 (14.4–46.6) µm, and the ratio of height of oopores to the width is 0.45. Axial parenchyma absent (Figs. 10E, F, 11A–F).
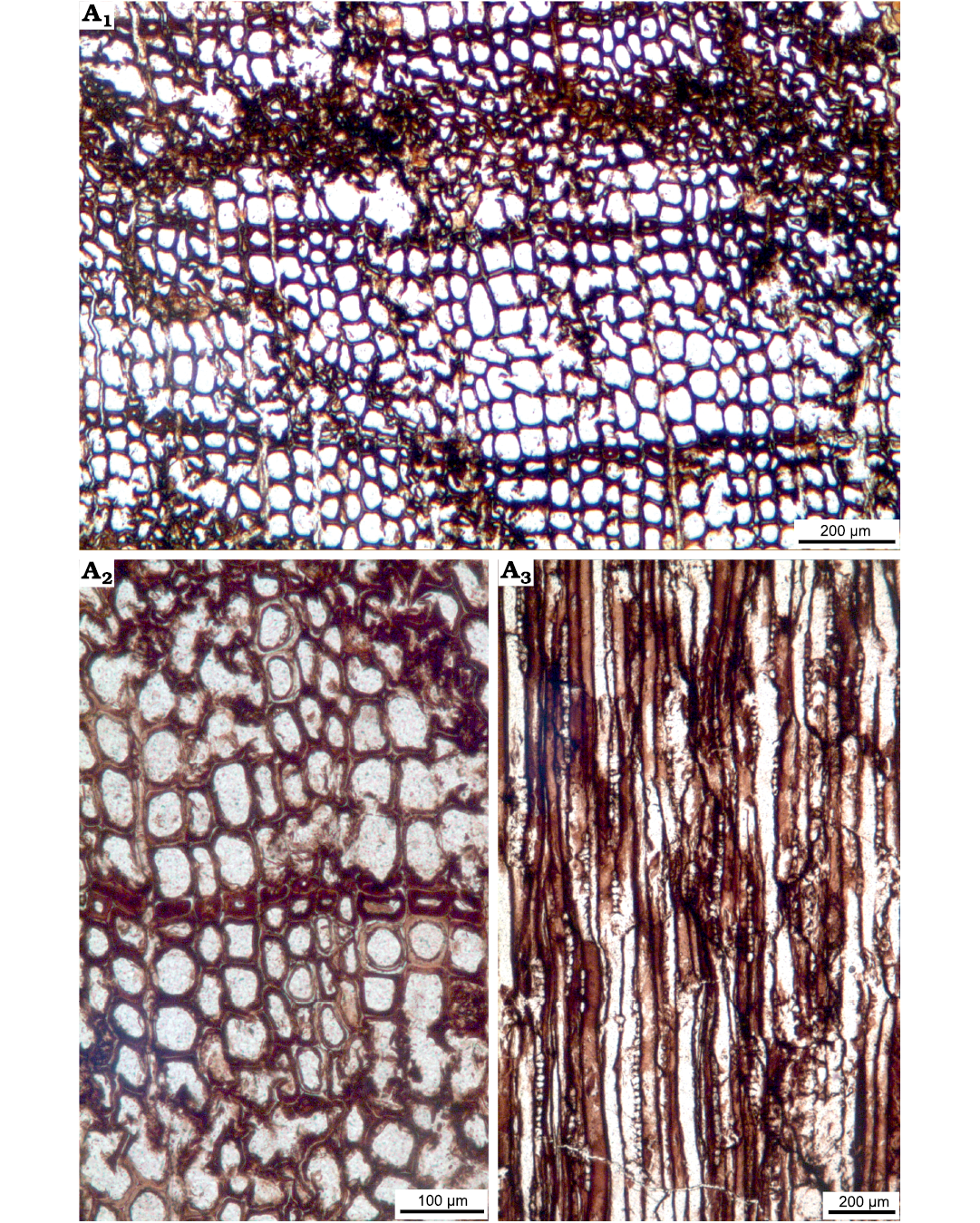
Fig. 9. Wood sections of Xenoxylon hopeiense Chang, 1929 (ISTO-FW-00243) from Jurassic of the Yesilbaglar locality of the province of Erzurum, Turkey. A1–A2, transversal sections with distinct growth ring boundary; A3, predominantly uniseriate rays in tangential section.
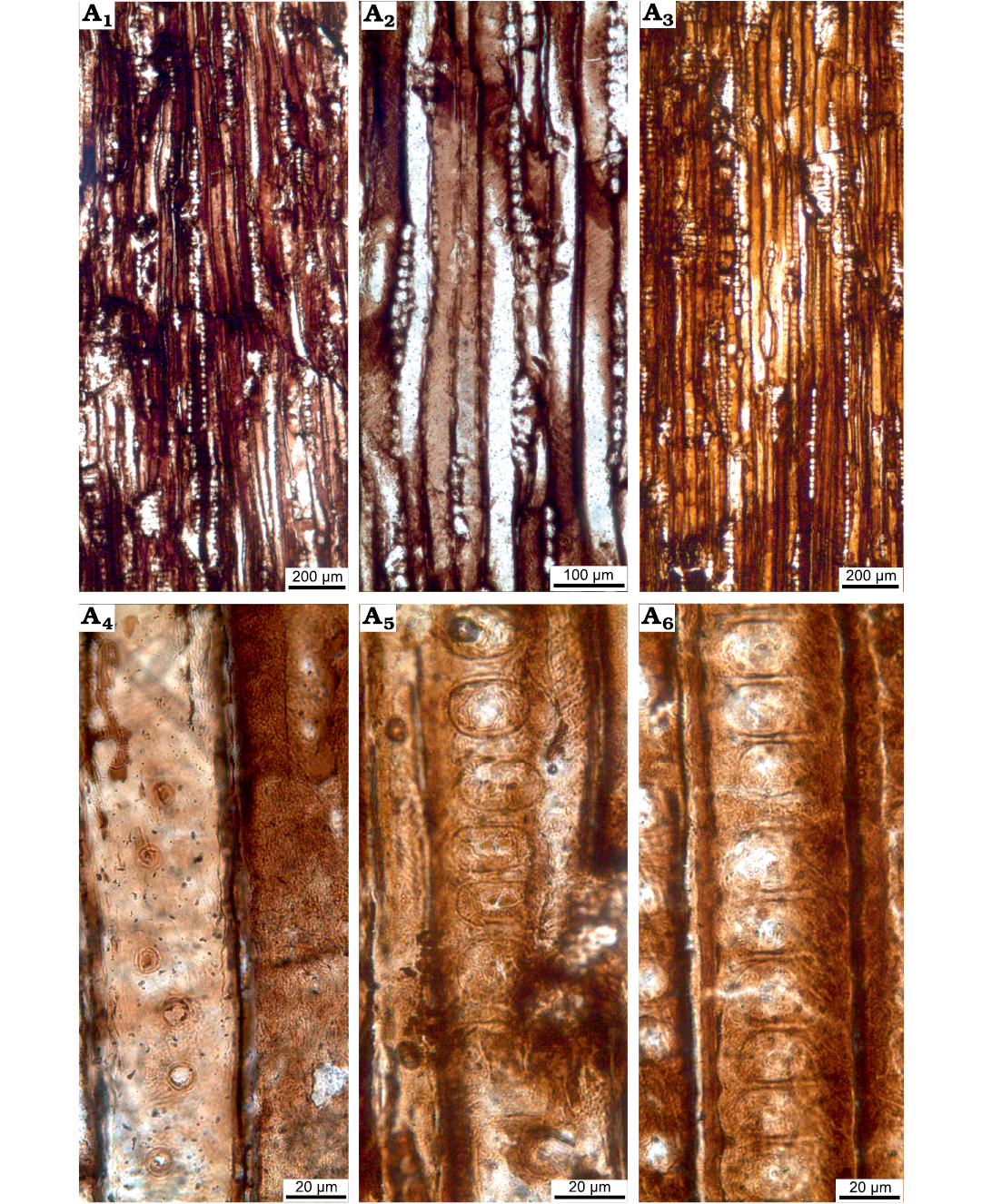
Fig. 10. Wood sections of Xenoxylon hopeiense Chang, 1929 (ISTO-FW-00243) from Jurassic of the Yesilbaglar locality of the province of Erzurum, Turkey. A1–A3, tangential sections with predominantly uniseriate and rarely partly biseriate rays; A4, intertracheary pits on the tangential walls of tracheids; A5, A6, mostly flattened xenoxylean type intertracheary pits on radial walls of tracheids.
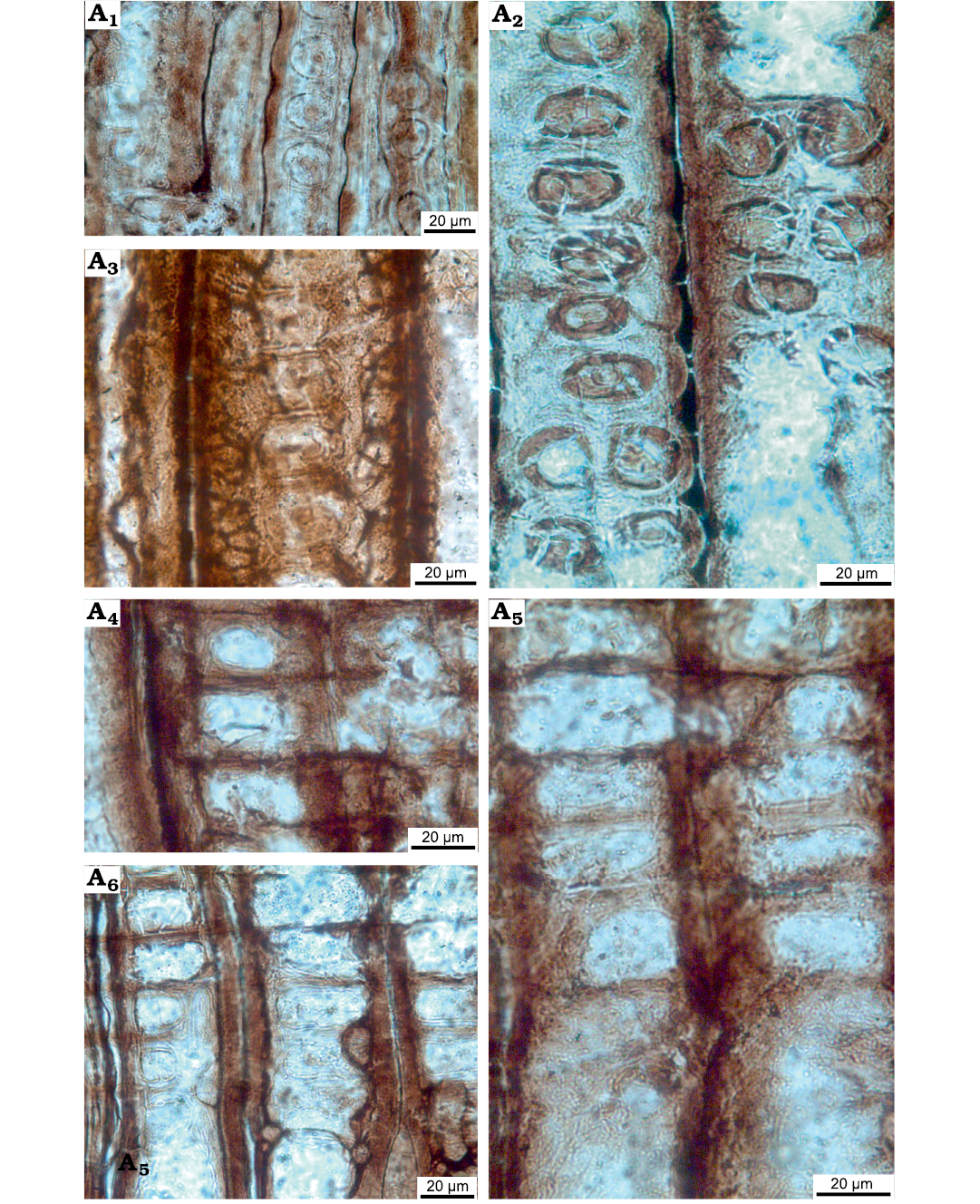
Fig. 11. Wood sections of Xenoxylon hopeiense Chang, 1929 (ISTO-FW-00243) from Jurassic of the Yesilbaglar locality of the province of Erzurum, Turkey. A1–A3, intertracheary pits on radial walls uniseriate or in some cases biseriate opposite, intertracheary pits locally xenoxylean, mostly strongly flattened, pit rows locally interrupted, rims of Sanio commonly present between both uniseriate and biseriate pits; A4–A6, window-like cross-field pits.
Remarks.—Xenoxylon was established by Gothan in 1905 based on the unusual features of some fossil gymnosperm woods from the Mesozoic, which have large rectangular earlywood cross-fields and strongly flattened more than twice as wide as high, contiguous intertracheary pits on radial walls of tracheids. This type of intertracheary pitting is called “xenoxylean radial pitting” (Müller-Stoll 1951; Suzuki and Terada 1992; Philippe and Bamford 2008; Boura et al. 2021). In similar fossil genera, cross-field oopores are more phyllocladoid (pointed and oblique, as in Protophyllocladoxylon Kräusel, 1939 [nomen conservandum]), or round and narrowly bordered all around (as in Circoporoxylon Kräusel, 1949). Due to its large and single oopores per cross-field and its locally xenoxylean pitting on radial walls of tracheids, our wood is assigned to Xenoxylon Gothan,1905.
Philippe et al. (2013) proposed an identification key for the Xenoxylon Gothan, 1905, species, and divided them into three informal groups. One of the groups is the Xenoxylon phyllocladoides Gothan, 1906, which is characterized by less strictly xenoxylean pitting, with some spaced, round or elliptic, intertracheary pits on the radial tracheid walls and rare pits on tangential walls. Our fossil clearly belongs to this group. In this group, there are: Xenoxylon hopeiense Chang, 1929; Xenoxylon huttonianum (Witham, 1833) Philippe and Hayes, 2010; Xenoxylon jakutiense Shilkina, 1986, and Xenoxylon phyllocladoides Gothan, 1906. Having Sanio’s rims, no axial parenchyma, and both uni- to biseriate radial intetracheary pitting, the present specimen differs from all other fossil species within the Xenoxylon phyllocladoides group. As axial parenchyma could have been overlooked, and as Xenoxylon jakutiense Shilkina, 1986, intertracheary radial pitting is always uniseriate, we consider that greatest similarities are with Xenoxylon hopeiense Chang, 1929.
Stratigraphic and geographic range.—Upper Jurassic for Olurdere Formation. As all Xenoxylon this is a boreal species. It has a wide distribution, ranging from Asia (China and Korea) northward to the Arctic.
Discussion
We could distinguish two fossil wood assemblages from north-eastern Anatolia (Fig. 12A). First, the Middle Jurassic assemblage, consists only of Agathoxylon. The second, the Late Jurassic assemblage, is much more diverse and comprise Agathoxylon, Protelicoxylon, and Xenoxylon. The respective compositions of the two assemblages have palaeobiogeographic and palaeoecological implications.
The monogeneric Middle Jurassic assemblage is less informative. Indeed, the genus Agathoxylon was widespread worldwide at that time. It cannot be unequivocally linked to a botanical taxon (Rößler et al. 2014), nor to any particular environment. The wood of Agathoxylon was reported from several Middle Jurassic localities that are now in relative proximity of Turkey and located on both margins of the Neotethys. The Gondwanan side includes occurrences in Israel (Philippe et al. 2004) and Tunisia (Philippe et al. 2004), while the Laurasian side includes Bulgaria, Poland, and Romania (Philippe et al. 2006) (Fig. 12A). Agathoxylon was also reported from Middle Jurassic localities the palaeobiogeography of which at that time is less clearly established, as for Eastern Pontides: Iran (Kerman Basin, Central Iran; Poole and Ataabadi 2005) and Sardinia (Costamagna et al. 2018). It is worth to note that Agathoxylon is not yet reported from the Jurassic Shemshak Group in Alborz, Iran (Fig 12A). However, the existing data on fossil woods from this unit are uncertain as limited merely to an unpublished thesis (Nadjafi 1982), where the stratigraphic position of the studied specimens is not precisely indicated.
Although the absence of data is always a sensitive matter in palaeontology, it is noteworthy that Xenoxylon is absent from the eSZ Middle Jurassic assemblage, whereas it is abundantly documented on the northern Tethyan margin at the same time (Crimea, Georgia, and Poland; Philippe et al. 2006). Xenoxylon is also documented in the Middle Jurassic of Iran, from the Dansirit Formation and Dalichai Formation in Alborz (Mohammed Gadivel and Tayebe Farahani, unpublished data, 2008 and 2017, respectively) and from the Hodjek Formation in Central Iran (Poole and Ataabadi 2005). Similarly, it is also worthwhile that the wood of Metapodocarpoxylon, which was so widely distributed in northern Gondwana in the Middle Jurassic (Philippe et al. 2003), is missing not only in the Middle but also the Late Jurassic wood assemblages of northeastern Turkey.
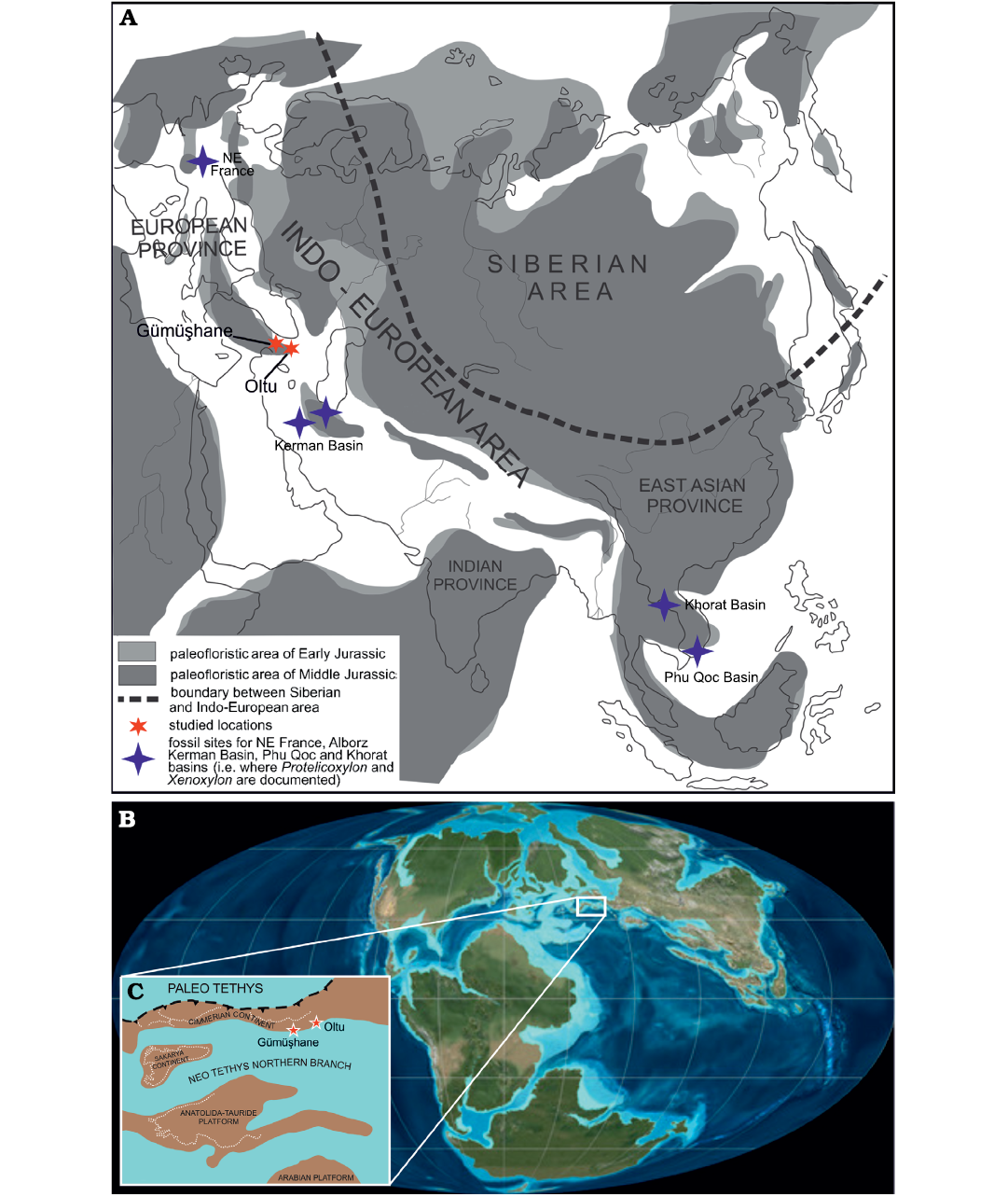
Fig. 12. The sampling localities of the identified fossil genera of Protelicoxylon and Xenoxylon from the Jurassic age of the World. A. The palaeofloristic divisions of the Early–Middle Jurassic and the palaeo-locations of the fossil sites (modified after Vakhrameev 1991) and the fossil sites for NE France, Alborz, Kerman Basin, Phu Qoc, and Khorat Basins (i.e., where Protelicoxylon and Xenoxylon are documented). B. Global palaeogeographic reconstruction of the Earth in the Late Jurassic period (from Ron Blakey https://paleonerdish.wordpress.com/2015/06/01/the-real-jurassic-world/). C. Detailed palaeogeographic map of the eastern Sakarya Zone (after Şengör and Yilmaz 1981).
Using a sedimentological approach Garcia et al. (1998) demonstrated that in the Middle–Late Jurassic of Western Europe Agathoxylon source trees did not grow in coastal environments, but rather inland. The palaeolatitudes of Agathoxylon occurrences in the Jurassic of Western Europe suggest a southern chorology for that area (Philippe et al. 2017) . It suggests that thus wood belong to relatively thermophilous trees, able to withstand dry seasons. This inference supports earlier interpretation of Akgün et al. (2006) based on palynofloras and other geological evidences.
The two Late Jurassic fossil wood assemblages described here are more informative. As in the case of the Middle Jurassic assemblages they both comprise woods of Agathoxylon. In Yeşilbağlar, besides Agathoxylon, the fossil assemblage also consists of woods of Protelicoxylon and Xenoxylon, which is an uncommon association. Both Protelicoxylon and Xenoxylon are strictly Laurasian (Oh et al. 2015; Philippe et al. 2019), and their occurrence in the eSZ demonstrates that at that time the terrestrial ecosystems of the area were connected to the Laurasian Realm (Fig. 12B, C). The spatially closest occurrence for Protelicoxylon is Ferizi, in Alborz Mountains, northern Iran (Fakhr and Marguerier 1977; Philippe 1995). In this locality the Dansirit Formation crops out and is dated as Middle Jurassic (early Bajocian; Vaez-Javadi 2010). The Dansirit Formation in Iran also yielded Xenoxylon (see above).
Genera Protelicoxylon and Xenoxylon are rarely found occurring together. Such co-occurrence is documented from the Lower Jurassic of North-eastern France (Philippe 1995), from the lower Middle Jurassic of Northern Iran (see above), here from the Upper Jurassic of eSZ, and in the Upper Jurassic–Lower Cretaceous of Southeastern Asia (Khorat Basin and its coeval Phu Qoc Basin; Serra 1969; Philippe et al. 2019; Boonchai et al. 2020). If such co-occurrence reflects that the source plants were part of the same vegetation, data suggest that corresponding vegetation originated in western Europe during the Early Jurassic, and subsequently shifted eastwards.
The boreal genus Xenoxylon is related to high palaeolatitudes throughout its time range, from the Carnian (Late Triassic) to the Maastrichtian (Late Cretaceous), with occasional extensions of its distribution southwards, probably related to overall cooler and/or wetter periods (Philippe and Thévenard 1996; Marynowski et al. 2008; Philippe et al. 2009, 2017; Oh et al. 2015; Tian et al. 2016; Richmond et al. 2019). This relation was recently questioned (Xie et al. 2021) on the basis of growth-ring analysis by the Mean Annual Sensitivity method (Fritts 1976). Wide and regular rings have already been described for Xenoxylon (Philippe et al. 2009), and while they imply greater water availability, they do not necessarily indicate that the climate was warm. Similar Mean Annual Sensitivity values are also observed in temperate to cool oceanic climates (Fritts 1976). Moreover, the weak differentiation of the latewood by Xenoxylon could well be genetically determined rather than climatically induced (Brison et al. 2001). In the present state of knowledge, these tree-ring observations do not seem to call into question the inferences from the latitudinal distribution of the genus and the correlation of southward extensions during cooler time intervals (Amiot et al. 2015, 2021). In the Eastern Pontides, as in Georgia (Delle 1960), Alborz (Vaez-Javadi 2010), Uzbekistan (Shilkina and Khudayberdyev 1971), Hunan (Miao et al. 1989), and Liaoning (Philippe et al. 2019), Xenoxylon is associated with coal, which is indicative of a humid and not too warm palaeoenvironment.
Regarding the discussion on the presence of terrestrial and shoreline environment Hoş-Çebi (1998) suggests that the coals of Jurassic coal-bearing beds in Gümüşhane region were deposited in shoreline environment connected with the sea intermittently based on the Inductively Coupled Plasma Atomic Emission Spectroscopy (ICP-AES) analyses, which is an analytical technique based on the principles of atomic spectroscopy for the determination of more than 70 elements. As for the Erzurum region, Kara-Gülbay et al. (2018) studied the organic geochemistry and depositional environment of the Oltu Gemstone in the Erzurum region. They suggest that Oltu Gemstone coals were deposited in an environment changing from shallow marine to deep shelf conditions where volcanism was effective. They also imply that coal occurrences were formed by deposition of organic matter of mainly terrestrial type under anoxic conditions in an environment which occasionally changed to ephemeral wetlands where resin-rich trees were the source of resin.
Conclusions
Fossil woods from the Middle and Upper Jurassic of the Sakarya Zone (eSZ) revealed three genera of conifers Agathoxylon, Protelicoxylon, and Xenoxylon. For the Middle Jurassic only Agathoxylon is documented, which allows no palaeobiogeographic inference. Palaeoecologically these woods may support the previous palynomorph-based investigations which suggested at least seasonally dry climate. The Late Jurassic wood flora evidences a continuity of the Gondwanan eSZ terrestrial areas with the Laurasian ones. The occurrence of Xenoxylon within this Late Jurassic wood flora suggests abundant water supply under a relatively cool/humid climate. Overall, both floras show important similarities with a contemporaneous fossil wood flora of Iran and, to a lesser extent, those of Georgia. New studies of Sakarya Zone fossil wood assemblages would have a great potential to clarify the palaeobiogeographic scenario of the Sakarya Zone terrestrial ecosystems, with important implications for the knowledge of tectonics and associated palaeoclimate.
Acknowledgements
Erdei Boglarka (Hungarian Natural History Museum, Budapest, Hungary), Tayebe Farahani (University of Isfahan, Iran), Mohammad Gadivel-Svooki (Tehran University, Iran) kindly supplied samples or information which were used here. The senior author (ÜA) was supported by the Research Fund of Istanbul University-Cerrahpaşa, Project number: FBA-2021-35698. We thank two anonymous referees for their valuable suggestions.
References
Ağirman Aktürk, E. 2016. Investigation of Sedimentary Facies and Depositional Environments of the Oxfordian-Berriasian Aged Olurdere Formation [in Turkish, with English abstract]. 117 pp. Unpublished Master Thesis, Karadeniz Technical University the Graduate School of Natural and Applied Sciences, Trabzon.
Akdoğan, R., Okay, A.I., and Dunkl, I. 2018. Triassic–Jurassic arc magmatism in the Pontidesas revealed by U-Pb detritalzircon ages in Jurassic sandstones, Northeastern Turkey. Turkish Journal of Earth Sciences 27: 89–109. Crossref
Akgün, F., Kandemir, R., and Yilmaz, C. 2006. A paleoecological investigation of Middle Jurassic palynoflora from NE Turkey. In: J. Kvaček (ed.), 7th European Palaeobotany-Palynology Conference, Prague, 6–11 September 2006, Program and Abstract Book, 1–2. National Museum, Prague.
Akkemik, Ü., Acarca, N.N., and Hatipoğlu, M. 2017. The first Glyptostroboxylon from the Miocene of Turkey. IAWA Journal 38 (4): 561–570. Crossref
Akkemik, Ü., Akkılıç, H., and Güngör, Y. 2019. Fossil wood from the Neogene of the Kilyos coastal area in Istanbul, Turkey. Palaeontographica B 299 (4–6): 133–185. Crossref
Akkemik, Ü., Arslan, M., Poole, I., Tosun, S., Köse, N., Karlıoğlu Kılıç, N., and Aydın, A. 2016. Silicified woods from two previously undescribed early Miocene forest sites near Seben, northwest Turkey. Review Palaeobotany and Palynology 235: 31–50. Crossref
Akkemik, Ü., Atıcı, G., Poole, I., and Çobankaya, M. 2018. Three new silicified woods from a newly discovered earliest Miocene forest site in the Haymana Basin (Ankara, Turkey). Review of Palaeobotany and Palynology 254: 49–64. Crossref
Akkemik, Ü., Mantzouka, D., and Kıran Yıldırım, D. 2020. The first report of a new Lesbosoxylon species from the early–middle Miocene of eastern Anatolia. Geodiversitas 42: 427–441. Crossref
Akkemik, Ü., Mantzouka, D., Tunç, U., and Koçbulut, F. 2021. The first paleoxylotomical evidence from the Mid-Eocene Climate Optimum from Turkey. Review Palaeobotany and Palynology 285: 104356. Crossref
Amiot, R., Kusuhashi, N., Saegusa, H., Shibata, M., Ikegami, N., Shimojima, S., Sonoda, T., Fourel, F., Ikeda, T., Lécuyer, C., Philippe, M., and Wang, X. 2021. Paleoclimate and ecology of Cretaceous continental ecosystems of Japan inferred from the stable oxygen and carbon isotope compositions of vertebrate bioapatite. Journal of Asian Earth Sciences 204: 104602. Crossref
Amiot, R., Wang, X., Zhou, Z., Wang, X., Lécuyer, C., Buffetaut, E., Fluteau, F., Ding, Z., Kusuhashi, N., Mo, J., Philippe, M., Suteethorn, V., Wang, Y., and Xu, X. 2015. Environment and ecology of East Asian plant-eating dinosaurs during the Early Cretaceous inferred from stable oxygen and carbon isotopes in apatite. Journal of Asian Earth Sciences 98: 358–370. Crossref
Bamford, M. and Philippe, M. 2001. Jurassic–Early Cretaceous Gondwanan homoxylous woods: a nomenclatural revision of the genera with taxonomic notes. Review of Palaeobotany and Palynology 113: 287–297. Crossref
Barrier, E., Vrielynck, B., Brouillet, J.F., and Brunet, M.F. 2018. Paleotectonic Reconstruction of the Central Tethyan Realm: Tectonosedimentary-palinspastic Maps from Late Permian to Pliocene: Atlas of 20 Maps. Commission for the Geological Map of the World, Paris.
Booi, M., van Konijnenburg-van Cittert, J., and van Waveren, I. 2014. Wood anatomical variability in Early Permian “Araucarioids”. International Association of Wood Anatomists Journal 35: 307–331. Crossref
Boonchai, N., Suteethorn, S., Sereeprasirt, W., Suriyonghanphong, C., Amiot, R., Cuny, G., Legrand, J., Thévenard, F., and Philippe, M. 2020. Xenoxylon, a boreal fossil wood in the Mesozoic redbeds of Southeast Asia: potential for the stratigraphy of the Khorat Group and the palinspatic reconstruction of Southeast Asia. Journal of Asian Earth Sciences 189: 104153. Crossref
Boura, A., Bamford, M., and Philippe, M. 2021. Promoting a standardized description of fossil tracheidoxyls. Review of Palaeobotany and Palynology 295: 104525. Crossref
Bozkuş, C. 1992, Stratigraphy of the Olur (Erzurum) region. Geological Bulletin of Turkey 35: 103–119.
Brison, A.L., Philippe, M., and Thévenard, F. 2001. Are Mesozoic growth-ring patterns climate-induced? Palaeobiology 27: 531–538. Crossref
Costamagna, L.G., Kustatscher, E., Scanu, G.G. Del Rio, M., Pittau, P., and van Konjinenburg-van Cittert, J.H.A. 2018. A palaeoenvironmental reconstruction of the Middle Jurassic of Sardinia (Italy) based on integrated palaeobotanical, palynological and lithofacies data assessment. Palaeobiodiversity and Palaeoenvironments 98: 111–138. Crossref
Delle, G.V. 1960. Recent data on the Jurassic flora of Dtkvarcheli [in Russian, with English abstract]. Doklady Akademii Nauk SSSR 133: 1150–1153.
Dokuz, A. and Tanyolu, E. 2006. Geochemical constraints on the provenance, mineral sorting and subaerial weathering of Lower Jurassic and Upper Cretaceous clastic rocks from the Eastern Pontides, Yusufeli (Artvin), NE Turkey. Turkish Journal of Earth Sciences 15: 181–209.
Dokuz, A. and Sünnetçi, K. 2019. Jurassic acidic magmatism in a back-arc setting, eastern Sakarya Zone, Turkey: Geochemical constraints and an evolutionary model. Lithos 332: 312–327. Crossref
Fakhr, M.S. and Marguerier, J. 1977. Prototaxoxylon feriziense n. sp., bois fossile du Juras sique moyen de lʼIran. In: M.S. Fakhr (ed.), Contribution à lʼétude de la flore Rhéto-liassique de la formation de Shemshak de lʼElbourz (Iran). Bibliotheque Nationale de France Paris Mémoire Science 5: 146–150.
Fritts, H.C. 1976. Tree Rings and Climate. 567 pp. Academic Press, London.
Garcia, J.P., Philippe, M., and Gaumet, F. 1998. Fossil-wood in Middle–Upper Jurassic marine sedimentary cycles of France: relations with climate, sea-level dynamics, and carbonate-platform environments. Palaeogeography, Palaeoclimatology, Palaeoecology 141: 199–214. Crossref
Gnaedinger, S. 2007. Planoxylon Stopes, Protelicoxylon Philippe y Herbstiloxylon nov. gen. (Coniferales) de la Formación La Matilde (Jurásico Medio), provincia de Santa Cruz, Argentina. Ameghiana 44: 321–335.
Golonka, J. 2004. Plate tectonic evolution of the southern margin of Eurasia in the Mesozoic and Cenozoic. Tectonophysics 381: 235–273. Crossref
Görür, N., Şengör, A.M.C., Akkök, R., and Yilmaz, Y. 1983. Sedimentological data on the opening of the northern branch of the Neotethys in the Pontides [in Turkish, with English abstract]. Türkiye Jeoloji Kurumu Bülteni 26 (1): 11–20.
Greguss, P. 1967. Fossil Gymnosperm Woods in Hungary from the Permian to the Pliocene. 222 pp. Akadémiai Kiadó, Budapest.
Hinsbergen, D.J.J.V., Torsvik, T.H., Schmid, S.M., Matenco, L.C., Maffione, M., Vissers, R.L.M., Gürer, D., and Spakman, W. 2020. Orogenic architecture of the Mediterranean region and kinematic reconstruction of its tectonic evolution since the Triassic. Gondwana Research 81: 79–229. Crossref
Hoş-Çebi, F. 1998. The Trace Element Geochemistry of Jurassic Coals in the Kelkit-Şiran (Gümüşhane) and Yusufeli (Artvin) areas [in Turkish, with English abstract]. 84 pp. Master Thesis, Karadeniz Technical University, Trabzon.
IAWA 2004. List of microscopic features for softwood identification. In: H.G. Richter, D. Grosser, I. Heinz, and P.E. Gasson (eds.), International Association Wood Anatomist Journal 25: 1–70. Crossref
Kandemir, R. 2004. Sedimentary Characteristics and Depositional Conditions of Lower Middle Jurassic Şenköy Formation in and around Gümüşhane [in Turkish, with English abstract]. 293 pp. Ph.D. Thesis, Karadeniz Technical University, Trabzon.
Kandemir, R. and Yilmaz, C. 2009. Lithostratigraphy, facies, and depositional environment of the Lower Jurassic Ammonitico Rosso type sediments (ARTS) in the Gümüşhane area, NE Turkey: implications for the opening of the northern branch of the NeoTethys Ocean. Journal of Asian Earth Science 34: 586–598. Crossref
Kandemir, R., Özyurt, M., and Karsli, O. 2021. Sedimentological and geochemical characteristics of Lower Jurassic Sandstones from Gümüşhane, NE Turkey: implications for source to sink processes, paleoenvironmental conditions, provenance and tectonic settings. International Geology Review [published online https://doi.org/10.1080/00206814.2021.1958383] Crossref
Kara-Gülbay, R., Korkmaz, S., Yaylali-Abanuz, G., and Erdoğan, M.S. 2018. Organic geochemistry and depositional environment of the Oltu Gemstone (Coal) in the Erzurum Area, NE Anatolia, Turkey. Energy Fuels 32: 1451–1463. Crossref
Kirmaci, M.Z. 1992. Sedimentological Investigation of the Upper Jurassic–Lower Cretaceous Berdiga Limestone in the Alucra-Gümüşhane-Bayburt Regions Eastern Pontides, NE, Turkey [in Turkish, with English abstract]. 256 pp. Unpublished Ph.D. Thesis, Karadeniz Technical University, Trabzon.
Kirmaci, M.Z., Koch, R., and Bucur, I.I. 1996. An Early Cretaceous section in the Kircaova area (Berdiga limestone, NE-Turkey) and its correlation with platform carbonates in W-Slovenia. Facies 34: 1–22. Crossref
Kloster, A.C. and Gnaedinger, S.C. 2018. Coniferous wood of Agathoxylon from the La Matilde Formation, (Middle Jurassic), Santa Cruz, Argentina. Journal of Paleontology 92: 546–567. Crossref
Kutluk, H., Kir, O., and Akkemik, Ü. 2012. First Report of Araucariaceae wood (Agathoxylon sp.) from the Late Cretaceous of Turkey. International Association of Wood Anatomists Journal 33: 319–326. Crossref
Koch, R., Bucur, I.I., Kirmaci, M.Z., Eren, M., and Tasli, K. 2008. Upper Jurassic and Lower Cretaceous carbonate rocks of the Berdiga Limestone—Sedimentation on an onbound platform with volcanic and episodic siliciclastic influx. Biostratigraphy, facies and diagenesis (Kircaova, Kale-Gümüshane area; NE-Turkey). Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 247: 23–61. Crossref
Koçyiğit, A. and Altiner, D. 2002. Tectonostratigraphic evolution of the North Anatolian palaeorift (NAPR): Hettangian–Aptian passive continental margin of the northern Neo-Tethys, Turkey. Turkish Journal of Earth Science 11: 169–191.
Konak, N. and Hakyemez, H.Y. 2008. Geological Maps of Turkey. 1:100000 scale, Tortum G-48 Sheet, Map and Explanatory Text. 69 pp. MTA Publications, Ankara.
Marynowski, L., Philippe, M., Zatoń, M., and Hautevelle, Y. 2008. Systematic relationships of Mesozoic wood genus Xenoxylon: integrative biomolecular and palaeobotanical approach. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 247: 177–189. Crossref
Miao, F., Qian, L., and Zhang, X. 1989. Peat-forming materials and evolution of swamp sequences-case analysis of a Jurassic inland coal basin in China. In: P.C. Lyons and B. Alpern (eds.), Peat and coal: origin, facies and depositional models. International Journal of Coal Geology 12: 733–765. Crossref
Müller-Stoll, W.R. 1951. Mikroskopie des zersetzen und fossilierten Holzes. In: H. Freund (ed.), Handbuch der Mikroskopie in der Technik, Vol. 5, 727–816. Umschau Verlag, Frankfurt am Main.
Nadjafi, A. 1982. Contribution à la connaissance de la flore ligneuse du Jurassique d’Iran. 441 pp. Unpublished Ph.D. Thesis, Paris VI University, Paris.
Oh, C., Philippe, M., and Kim, K. 2015. Xenoxylon synecology and palaeoclimatic implications for the Mesozoic of Eurasia. Acta Palaeontologica Polonica 60: 245–256.
Okay, A.I. and Tüysüz, O. 1999. Tethyan sutures of northern Turkey. Geology Society Special Publications 156: 475–515. Crossref
Okay, A.I., Sunal, G., Tüysüz, O., Sherlock, S., Keskin, M., and Kylander-Clark, A.R.C. 2014. Low-pressure–high-temperature metamorphism during extension in a Jurassic magmatic arc, Central Pontides, Turkey. Journal of Metamorphic Geology 32: 49–69. Crossref
Pelin, S. 1977. Alucra (Giresun) Güneydoğu Yöresinin Petrol Olanakları Bakımından Jeolojik İncelenmesi [with English abstract]. 103 pp. Karadeniz Technical University Publication No: 87/13, Trabzon.
Philippe, M. 1995. Bois fossiles du Jurassique de Franche-Comté (nord-est de la France): systématique et biogéographie. Palaeontographica B 236: 45–103.
Philippe, M. 2011. How many species of Araucarioxylon? Comptes Rendus Palevol 10: 201–208. Crossref
Philippe, M. and Bamford, M. 2008. A key to morphogenera used for Mesozoic conifer-like woods. Review of Palaeobotany and Palynology 148: 184–207. Crossref
Philippe, M. and Cantrill, D.J. 2007. Nomenclatural types and taxonomy of Gothan’s Arctic fossil conifer wood. Taxon 56: 551–566. Crossref
Philippe, M. and Thévenard, F. 1996. Repartition and palaeoecology of the Mesozoic wood genus Xenoxylon: palaeoclimatological implications for the Jurassic of Western Europe. Review of Palaeobotany and Palynology 91: 353–370. Crossref
Philippe, M., Afonin, M., Delzon, S., Jordan, G.J., Terada, K., and Thiébaut, M. 2019. A palaeobiogeographical scenario for the Taxaceae based on a revised fossil wood record and embolism resistance. Review of Palaeobotany and Palynology 263: 147–158. Crossref
Philippe, M., Bamford, M., Mcloughlin, S., Da Rosa Alves, L.S., Falcon-Lang, H., Gnaedinger, S., Ottone, E., Pole, M., Rajanikanth, A., Shoemaker, R.E., Torres, T., and Zamuner, A. 2004. Biogeography of Gondwanan terrestrial biota during the Jurassic–Early Cretaceous as seen from fossil wood evidence. Review of Palaeobotany and Palynology 129: 141–173. Crossref
Philippe, M., Barbacka, M., Gradinaru, E., Iamandei, E., Iamandei, S., Kázmér, M., Popa, M., Szakmány, G., Tchoumatchenco, P., and Zatoń, M. 2006. Fossil wood and Mid-Eastern Europe terrestrial palaeobiogeography during the Jurassic–Early Cretaceous interval. Review of Palaeobotany and Palynology 142: 15–32. Crossref
Philippe, M., Cuny, G., Bamford, M., Jaillard, E., Barale, G., Gomez, B., Ouaja, M., Thévenard, F., Thiébaut, M., and von Sengbusch, P. 2003. The palaeoxylological record of Metapodocarpoxylon libanoticum (Edwards) Dupéron-Laudoueneix et Pons and Late Jurassic–Early Cretaceous continental biogeography. Journal of Biogeography 30: 389–400. Crossref
Philippe, M., Jiang, H.-E., Kim, K.-S., Oh, C., Gromyko, D., Harland, M., Paik, I.-S., and Thévenard, F. 2009. Structure and diversity of the Mesozoic wood genus Xenoxylon in Far-East Asia: implications for terrestrial palaeoclimates. Lethaia 42: 393–406. Crossref
Philippe, M., Puijalon, S., Suan, G., Mousset, S., Thévenard, F., and Mattioli, E. 2017. The palaeolatitudinal distribution of fossil wood genera as a proxy for European Jurassic terrestrial climate. Palaeogeography, Palaeoclimatology, Palaeoecology 466: 373–381. Crossref
Philippe, M., Thévenard, F., Nosova, N., Kim, K., and Naugolnykh, S. 2013. Systematics of a palaeoecologically significant boreal Mesozoic fossil wood genus, Xenoxylon Gothan. Review of Palaeobotany and Palynology 193: 128–140. Crossref
Poole, I. and Ataabadi, M.M. 2005. Conifer woods of the middle Jurassic Hojedk formation (Kerman Basin) Central Iran. International Association of Wood Anatomists Journal 26: 489–505. Crossref
Pujana, R.R., Marenssib, S.A., and Santillana, S.N. 2015. Fossil woods from the Cross Valley Formation (Paleocene of Western Antarctica): Araucariaceae dominated forests. Review of Palaeobotany and Palynology 222: 56–66. Crossref
Richmond, D., Lupia, R., Philippe, M., and Klimek, J. 2019. First occurrence of the boreal fossil wood Xenoxylon meisteri from the Jurassic of North America: Morrison Formation of central Montana, USA. Review of Palaeobotany and Palynology 267: 39–53. Crossref
Robinson, A.G., Banks, C.J., Rutherford, M.M., and Hirst, J.P.P. 1995. Stratigraphic and structural development of the Eastern Pontides, Turkey. Journal of the Geology Society 152: 861–872. Crossref
Rößler, R., Philippe, M., van Konijnenburg-Van Cittert, J.H.A., Mcloughlin, S., Sakala, J., Zijlstra, G., Bamford, M., Booi, M., Brea, M., Crisafulli, A., Decombeix, A.-L., Dolezych, M., Dutra, T., Esteban, L.G., Falaschi, P., Feng Zhuo, Gnaedinger, S., Guerra Sommer, M., Harland, M., Herbst, R., Iamandei, E., Iamandei, S., Jiang Hongen, Kunzmann, L., Kurzawe, F., Merlotti, S., Naugolnykh, S., Nishida, H., Noll, R., Oh, C., Orlova, O., De Palacios, P.P., Poole, I., Pujana, R.R., Rajanikanth, A., Ryberg, P., Terada, K., Thevenard, F., Torres, T., Vera, E., Zhang, W., and Zheng, S. 2014. Which name(s) should be used for Araucaria-like fossil wood?—Results of a poll. Taxon 63: 177–184. Crossref
Serra, C. 1969. Sur les bois fossiles de l’archipel de Thô-Châu (Golfe de Thaïlande). Archives Géologique du Vietnam 12: 1–15.
Shilkina, I.A. and Khudayberdyev, R.H. 1971. Protocedroxylon and Xenoxylon [in Russian with English abstract]. Paleobotanica Uzbekistana 2: 117–133.
Suzuki, M. and Terada, K. 1992. Xenoxylon fossil woods from the lower Cretaceous Akaiwa subgroup of Shiramine, central Japan. Journal of Phytogeography and Taxonomy 40: 91–97.
Şengör, A.C. and Yilmaz, Y. 1981. Tethyan evolution of Turkey, a plate tectonic approach. Tectonophysics 75: 181–241. Crossref
Tasli, K., Özer, E., and Yilmaz, C. 1999. Biostratigraphic and environmental analysis of the Upper Jurassic–Lower Cretaceous carbonate sequence in the Basoba Yayla area (Trabzon, NE Turkey). Turkish Journal of Earth Science 8: 125–135.
Tian, N., Wang, Y., Philippe, M., Li, L., Xıe, X., and Jiang, Z. 2016. New record of fossil wood Xenoxylon from the Late Triassic in the Sichuan Basin, southern China and its palaeoclimatic implications. Palaeogeography, Palaeoclimatology, Palaeoecology 464: 65–75. Crossref
Torres, T. and Philippe, M. 2002. Nuevas espécies de Agathoxylon y Baieroxylon del Liásico de La Ligua, Chile, y evaluación de antecedentes paleoxilológicos en el Jurásico de América del Sur: Revista Geológica de Chile 29: 151–165. Crossref
Torsvik, T.H. 2019. Earth history: A journey in time and space from base to top. Tectonophysics 760: 297–313. Crossref
Ustaömer, T. and Robertson, A.H. 2010. Late Palaeozoic–Early Cenozoic tectonic development of the Eastern Pontides (Artvin area), Turkey: Stages of Closure of Tethys along the Southern Margin of Eurasia. Geological Society London Special Publications 340: 281–327. Crossref
Vaez-Javadi, F. 2010. Middle Jurassic flora from the Dansirit Formation of Shemshak Group, Alborz, north Iran. Alcheringa 35: 77–102. Crossref
Vakhrameev, V.A. 1991. Jurassic and Cretaceous Floras and Climates of the Earth. 318 pp. Cambridge University Press, Cambridge.
Vincent, S.J., Guo, L., Flecker, R., BouDagher-Fadel, M.K., Ellam, R.M., and Kandemir, R. 2018. Age constraints on intra-formational unconformities in Upper Jurassic–Lower Cretaceous carbonates in northeast Turkey; geodynamic and hydrocarbon implications. Marine and Petroleum Geology 91: 639–657. Crossref
Xie, A., Gee, C.T., Bennis, M.B., Gray, D., and Sprinkel, D.A. 2021. A more southerly occurrence of Xenoxylon in North America: X. utahense Xie et Gee sp. nov. from the Upper Jurassic Morrison Formation in Utah, USA, and its paleobiogeographic and paleoclimatic significance. Review of Palaeobotany and Palynology 291: 104451. Crossref
Yilmaz, H. 1985. The Geology of the Olur (Erzurum) Region. Black Sea University Bulletin Geology 4: 23–43.
Zamuner, A.B. 1996. Araucarioxylon petriellae n. sp. una posible Glossopteridal de la Formación Melo (Pérmico inferior) Uruguay. Ameghiniana 33: 77–82.
Zamuner, A. and Falaschi, P. 2005. Agathoxylon matildense n. sp., leño araucariáceo del Bosque petrificado del cerro Madre e Hija, Formación La Matilde (Jurásico medio), provincia de Santa Cruz, Argentina. Ameghiniana 42: 339–346.
Acta Palaeontol. Pol. 67 (3): 745–766, 2022
https://doi.org/10.4202/app.00962.2021