
New craniodental material of the typotherian notoungulates from the upper Oligocene of Mendoza, central-western Argentina and their taxonomical importance
SANTIAGO HERNÁNDEZ DEL PINO, FEDERICO D. SEOANE, and ESPERANZA CERDEÑO
Hernández Del Pino, S., Seoane, F.D., and Cerdeño, E. 2022. New craniodental material of the typotherian notoungulates from the upper Oligocene of Mendoza, central-western Argentina and their taxonomical importance. Acta Palaeontologica Polonica 67 (4): 983–997.
Among the great diversity of Oligocene mammals from Quebrada Fiera (Mendoza Province, Argentina), three families of typotherian notoungulates are very abundant. Since the first paper on archaeohyracids in 2010 until the last fieldwork in 2016, numerous unpublished remains were added to the collection. The study of all these remains, together with some revised materials, allows us to provide new anatomical data and some taxonomic decisions on these groups. Within Hegetotheriidae, we present one of the most complete skulls of “Prohegetotherium schiaffinoi” known to date, providing descriptions of certain previously unknown features of the caudal cranium; MCNAM-PV 3984 is discarded as belonging to Hegetotheriopsis sulcatus, as recently suggested, and is considered as Hegetotheriinae indet., like other isolated large specimens and some teeth previously determined as Prohegetotherium sp.; and several well-preserved specimens add to the sample of the pachyrukhine Prosotherium garzoni, including a partial maxilla with erupting P2. Concerning “Archaeohyracidae”, Archaeohyrax suniensis is now better represented, recording several ontogenetic stages previously observed in the Bolivian sample of this species. Finally, within Interatheriidae, most new dental material belongs to Argyrohyrax proavus, but an isolated upper molar suggests the presence of a different Interatheriinae at Quebrada Fiera, more similar to the Bolivian Brucemacfaddenia boliviensis than to other Deseadan (late Oligocene) taxa.
Key words: Mammalia, “Archaeohyracidae”, Hegetotheriidae, Interatheriidae, Agua de la Piedra Formation, Deseadan, Mendoza Province.
Santiago Hernández Del Pino [stgo86@gmail.com] and Esperanza Cerdeño [espe@mendoza-conicet.gob.ar], Paleobiología y Paleoecología, Instituto de Nivología, Glaciología y Ciencias Ambientales, Centro Científico Tecnológico-CONICET-Mendoza. Avda, Ruiz Leal s/n, 5500 Mendoza, Argentina.
Federico D. Seoane [fseoane@gl.fcen.uba.ar], Universidad de Buenos Aires, CONICET, Instituto de Estudios Andinos Don Pablo Groeber (IDEAN), Facultad de Ciencias Exactas y Naturales, Ricardo Güiraldes 2160, C1428EGA Buenos Aires, Argentina.
Received 14 January 2022, accepted 2 June 2022, available online 24 November 2022.
Copyright © 2022 S. Hernández Del Pino et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Quebrada Fiera is a fossiliferous locality in the Malargüe Department, south of Mendoza Province, central-western Argentina (SOM 1: fig. S1 in Supplementary Online Material available at http://app.pan.pl/SOM/app67-HernandezDelPino_etal_SOM.pdf). Known since the late 1970s (Gorroño et al. 1979), the first recovered fossils were barely described (Bond and Pascual 1983; Pascual and de la Fuente 1993). The locality encompasses two areas with fossils, at each side of a creek, but the fossiliferous sediments belong to the same unit, and no faunal differences have been noticed between them (e.g., Cerdeño and Vera 2014; Pujos et al. 2021).
The fossiliferous unit (QF4 level in Pujos et al. 2021) is interpreted as the base of the Agua de la Piedra Formation, and mainly consists of white to gray tuff with oxidized minerals, angular plagioclases, and gypsiferous levels, the latter probably corresponding to short evaporitic events with no volcanic contribution. More geological and sedimentary data can be found in Gorroño et al. (1979), Combina and Nullo (2011), and Pujos et al. (2021).
From 2006, new fieldwork at Quebrada Fiera revealed a great faunal diversity. Among notoungulates, three families of small typotherians were recognized. First, “Archaeohyracidae” were mostly assigned to one species, Archaeohyrax suniensis, shared with the Bolivian site of Salla, while only one specimen was assigned to cf. Archaeotypotherium (Cerdeño et al. 2010). Instead, Hegetotheriidae shows a high diversity (Cerdeño and Reguero 2015; Kramarz and Bond 2017), with four recognized species of hegetotheriines “Prohegetotherium malalhuense”, “Prohegetotherium schiaffinoi”, Prohegetotherium cf. P. sculptum, and Hegetotheriopsis sulcatus, plus one Hegetotheriinae indet. (previously identified as Prohegetotherium sp.), and the pachyrukhine Prosotherium garzoni. This list has been updated compared to the first identifications by Cerdeño and Reguero (2015), because two teeth originally identified as Prohegetotherium sp. were later recognized as Hegetotheriopsis sulcatus by Kramarz and Bond (2017). In addition, the revision of the genera Propachyrucos and Prosotherium by Seoane et al. (2019) led to the recognition of Prosotherium garzoni at Quebrada Fiera instead of Propachyrucos cf. P. smithwoodwardi as stated by Cerdeño and Reguero (2015). In turn, Interatheriidae are represented at Quebrada Fiera by Argyrohyrax proavus, Progaleopithecus sp., and a possible third taxon described as Interatheriinae indet. (Vera et al. 2017). In contrast with these three families, the medium-size Mesotheriidae are represented by only one thrachytheriine cranial fragment identified as Trachytherus cf. T. spegazzinianus (Cerdeño 2014).
Taking into account that fieldwork at Quebrada Fiera continued until 2016, many recovered remains are still unpublished. Concerning the small typotherians, new dental material mainly belongs to “Archaeohyracidae” and Hegetotheriidae, providing specimens that increase the knowledge of intraspecific variation of the recognized taxa and even new cranial morphological information for the species “Prohegetotherium schiaffinoi”. A smaller number of new specimens of Interatheriidae also provide further information on the diversity of this group. The anatomical analysis of all this new cranial (skull, mandible, and teeth) material is the focus of the present contribution.
Institutional abbreviations.—MACN A/Pv, Museo Argentino de Ciencias Naturales “Bernardino Rivadavia”, Ameghino Collection/Vertebrate Paleontology collection, Buenos Aires, Argentina; MCNAM-PV, Museo de Ciencias Naturales y Antropológicas “J.C. Moyano”, Vertebrate Paleontological collection, Mendoza, Argentina; MLP, Museo de La Plata, La Plata, Argentina; MNHN-SAL, Muséum national d’Histoire naturelle, Salla Collection, Paris, France; MNHN, Museo Nacional de Historia Natural, Montevideo, Uruguay; UF, Florida Museum of Natural History, University of Florida, Gainesville, USA; YPM-PU, Yale Peabody Museum, Yale University, New Haven, USA.
Other abbreviations.—C/c, upper/lower canine; D/d, deciduous; EAM, external acoustic meatus; H, height; I/i, upper/lower incisor; M/m, upper/lower molar; P/p, upper/lower premolar; SALMA, South American land mammal age; W, width.
Material and methods
Most specimens studied herein (except for some revised ones) are unpublished. Material recovered at Quebrada Fiera since 2006 belongs to the Vertebrate Paleontological collection of the Museo de Ciencias Naturales y Antropológicas “J.C. Moyano”, in Mendoza, but previous findings were stored in the Museo de La Plata, La Plata (Buenos Aires Province).
This study presents morphological and metrical comparisons of the unpublished material. Descriptions are not exhaustive due to the previously published papers on each analyzed group and follow these papers for anatomical terminology (see references throughout the text). Dimensions of specimens were taken with a caliper and photographs were taken with a CANON EOS REBEL T7 camera and a stereoscopic microscope LEICA M60 with camera DMC 2900. The text focuses on the best-preserved or most informative specimens, but others are detailed and figured in SOM 1–3.
Systematic palaeontology
Mammalia Linnaeus, 1758
Notoungulata Roth, 1903
Typotheria Zittel, 1893
Hegetotheriidae Ameghino, 1894
Hegetotheriinae Ameghino, 1894
Genus Prohegetotherium Ameghino, 1897
Type species: Prohegetotherium sculptum Ameghino, 1897; Sarmiento Formation, Cabeza Blanca, Chubut Province, Argentina; upper Oligocene, Deseadan SALMA.
Remarks.—Kramarz and Bond (2017) proposed that Prohegetotherium sculptum should be restricted to its type material and rejected other remains assigned to this species over years. They also rejected some other species as belonging to Prohegetotherium, because the genus was recovered as paraphyletic in the performed phylogenetic analysis (as in other analyses, see Seoane and Cerdeño 2019, and references therein). Thus, pending an in-depth revision of all materials/species previously assigned to Prohegetotherium, we place for these species the name in quotation marks, assuming that they could belong to a different, yet unnamed genus. Only a few isolated specimens from Quebrada Fiera were recognized as Prohegetotherium cf. P. sculptum (Cerdeño and Reguero 2015). Concerning the Miocene species Hegetotherium novum, Kramarz and Bond (2017) relocated it into Prohegetotherium, but Seoane and Cerdeño (2019) supported its assignment to Hegetotherium.
“Prohegetotherium schiaffinoi (Kraglievich, 1932)”
Figs. 1, 2A, B, 3A–F; SOM 1: figs. S2, S3A–E.
Holotype: MNHN-DP-186, left partial maxilla with P2–M2.
Type locality: Cañada de las Mulas, Santa Lucía River, Canelones Department, Uruguay.
Type horizon: Fray Bentos Formation, late Oligocene, Deseadan SALMA.
Emended diagnosis (after Reguero and Cerdeño 2005).—Hegetotheriine far smaller than Hegetotherium mirabile, Hemihegetotherium trilobus, and Hemihegetotherium achathaleptum, and smaller than Prohegetotherium sculptum and Sallatherium altiplanense, with shorter and wider nasals. Wide and bulky tympanic bullae, far more extended laterally than in Hegetotherium mirabile and Hemihegetotherium achathaleptum, and relatively larger foramen magnum. Upper cheek teeth slightly imbricated; the canine is not labially placed unlike in P. sculptum and Hemihegetotherium trilobus; distal face of M3 without notch, unlike in P. sculptum, “Prohegetotherium malalhuense”, and Hegetotherium mirabile. Lower cheek teeth not imbricated; anterior lobe of p3 relatively longer and squarer than in S. altiplanense, and with a wide but shallow lingual groove; talonid of m3 without labial groove, unlike in Hegetotherium mirabile, Hemihegetotherium achathaleptum, Hemihegetotherium trilobus, and Hegetotheriopsis sulcatus, and with a posterolingual inflexion.
Material.—35 specimens, including a rather complete skull, maxillary and mandibular fragments, upper and lower teeth (detailed list in SOM 2) from Quebrada Fiera, Mendoza Province, Argentina, upper Oligocene, Deseadan SALMA.
Description.—MCNAM-PV 5076 (Figs. 1, 2A, 3A; SOM 1: fig. S2) includes a rather complete skull and associated mandible, preserving most upper and lower teeth. The skull has an estimated length of 80–85 mm (preserved portion, 78.5 mm), closer to MACN Pv 16609 (holotype of “Ethegotherium carettei (Minoprio, 1947)”, 80.8 mm; Reguero and Cerdeño, 2005) than to the Bolivian remains from Salla of “Prohegetotherium schiaffinoi” (UF 91661, 92 mm, and UF 172445, 92 mm; Reguero and Cerdeño 2005). The preserved cranial roof is mostly formed by frontals and parietals (Fig. 1A3, A4). The damage of this area prevents for testing the degree of contribution of the interparietals to the cranial vault, but a clear contribution as observed in Paedotherium sp. (MacPhee 2014: fig. 8D) seems unlikely, considering the condition in closer taxa (Hegetotherium mirabile Ameghino, 1887, and Hemihegetotherium achathaleptum Rovereto, 1914). The remnant of the postorbital process suggests it was probably small, with its tip slightly curved backwards, different from the large, acute, and bilobate process of Hegetotherium mirabile (Sinclair, 1909). The temporal lines are discernible, despite the postmortem damage, and converge far posteriorly in a short sagittal crest; even though it is incomplete, we infer that this crest is relatively shorter than in Hegetotherium mirabile, Hemihegetotherium achathaleptum, and Hemihegetotherium trilobus Croft and Anaya, 2006, and probably also lower. In MACN Pv 16609 assigned to “P. schiaffinoi” (Reguero and Cerdeño 2005), the scarcely preserved parietals evidence a smooth sagittal crest (López 2002: 298). In turn, Reguero and Cerdeño (2005: 679) described the sagittal crest of “P. schiaffinoi” from Salla as being “short, low and acute, similar to Hegetotherium”, but it was not figured. The exposure of the anterior portion of the epitympanic theca (squamosal) is visible on both sides (Fig. 1A5–A8; SOM 1: fig. S2C–F). Due to the damage, there are no identifiable foramina in the skull vault. On the occipital face (Fig. 1A7, A8), the foramen magnum is large (foramen/occiput height ratio: Hf/Ho = 0.4) and wider than tall (width/height: W/H = 1.24). Compared to other hegetotheriines, the foramen magnum is relatively higher than in Hegetotherium mirabile (Hf/Ho = 0.36 in YPM-PU 15542) and Hemihegetotherium achathaleptum (Hf/Ho = 0.27 in MACN Pv 8491), narrower than in the former (W/H = 1.36) and wider than in the latter (W/H = 1.17). Above the foramen, a faint occipital crest does not project backwards; there are no remnants of the exoccipital crests due to breakage. The preserved right lateral depression of the exoccipital is little concave, but it was probably deeper than what is observed. The occipital condyles are narrower and longer than in Hegetotherium mirabile. The paracondylar processes of the exoccipital are long (their tips are lacking) and laterally compressed, but they are relatively shorter and more conical than in Hegetotherium mirabile. The preserved left infraorbital foramen (3.6 mm × 4.2 mm) is morphologically similar to that in Hegetotherium mirabile and similarly placed above M1. The zygomatic arch and the orbit are barely preserved. Due to breakage, part of the external auditory canal is observable (SOM 1: fig. S2C–E); considering that the preserved auditory canal is laterally directed at the level of the exposure, the EAM (ectotympanic) would be directed lateroposteriorly, as in other hegetotheriids (e.g., Hegetotherium mirabile, Hemihegetotherium achathaleptum, and species of Paedotherium). Anterior to the exposure of the left epitympanic theca, the dorsal border of the petrosal is exposed due to breakage. Considering the preserved area of the epitympanic theca, MCNAM-PV 5076 confirms the well-developed epitympanic sinus described by Reguero and Cerdeño (2005) based on the Bolivian “P. schiaffinoi”, being similar to, or even larger than in, Hegetotherium mirabile. In ventral view (Fig. 1A1, A2), the incomplete palate is wide and concave, and the posterior border of the left incisive foramen is preserved. At the left posterior portion of the palate, a distinctive, long sulcus extends anteriorly to a small palatine foramen; although varying in location and development, palatine foramina are present in other hegetotheriines. The palatines have a posterior W-shaped outline, contacting posteromedially with the anterior part of the alisphenoids, with ipsilateral pterygoid processes diverging laterally. Apparently, postmortem deformation broke most of the platform below the choanae, mostly formed by the palatines and alisphenoids, leaving no remnant of the pterygoid bones. In the midline of the caudal cranium, damage removed the basisphenoid completely; the basioccipital is trapezoidal, narrower anteriorly than posteriorly, and a medial crest runs anteroposteriorly, being taller and sharper at its posterior end (Fig. 1A1, A2; SOM 1: fig. S2G, H). This crest is relatively shorter and bulkier than the long and acute median keel present in Hegetotherium mirabile and Hemihegetotherium achathaleptum. Two conspicuous posterior foramina, lateral to the medial crest, probably correspond to the exits of venous vasa (located approximately in the same area as the transclival foramina identified in the interathere Cochilius volvens Ameghino, 1902; MacPhee et al. 2021). Instead, in the YPM-PU 15542 of Hegetotherium mirabile, there is a single small foramen located at the midline of the basioccipital and there is apparently none in Hemihegetotherium achathaleptum or Hemihegetotherium trilobus (Croft and Anaya 2006), while the pachyrukine genus Paedotherium has several small foramina in the basioccipital (MacPhee 2014: fig. 8). Lateral to the basioccipital, the tympanic bullae are large and relatively wide, anteroposteriorly convex and slightly inclined laterally; each bulla is oriented along a ventromedial-anterodorsally axis (Fig. 1A1, A2; SOM 1: fig. S2G, H) and shows a steep medial wall. Compared to other hegetotheriines, the bullae are larger and especially wider (SOM 3: table 1) than in Hegetotherium mirabile and Hemihegetotherium achathaleptum. Anterior to each bulla, there is an opening that corresponds to the pyriform fenestra (after MacPhee and Forasiepi 2022). The jugular foramen is located behind each bulla, on its lateral side, as usual in typotheres, with its posterior boundary defined by the exoccipitals. The hypoglossal foramen is placed anteriorly and laterally to each condyle. Further comparisons of the basicranial anatomy with other specimens of “P. schiaffinoi” are not possible due to their incompleteness or lack of detailed descriptions. Regarding the dentition of MCNAM-PV 5076, upper premolars are subtriangular, longer than wide, and do not present posterolingual grooves (Fig. 2A). In general, all cheek teeth are comparable in size (SOM 3: table 1) and morphology to those of MCNAM-PV 3959 (“P. schiaffinoi”; Cerdeño and Reguero 2015: fig. 4B), although the centered enamel line has already disappeared with wear (for further data on this feature, see Seoane and Cerdeño 2019, and references therein).
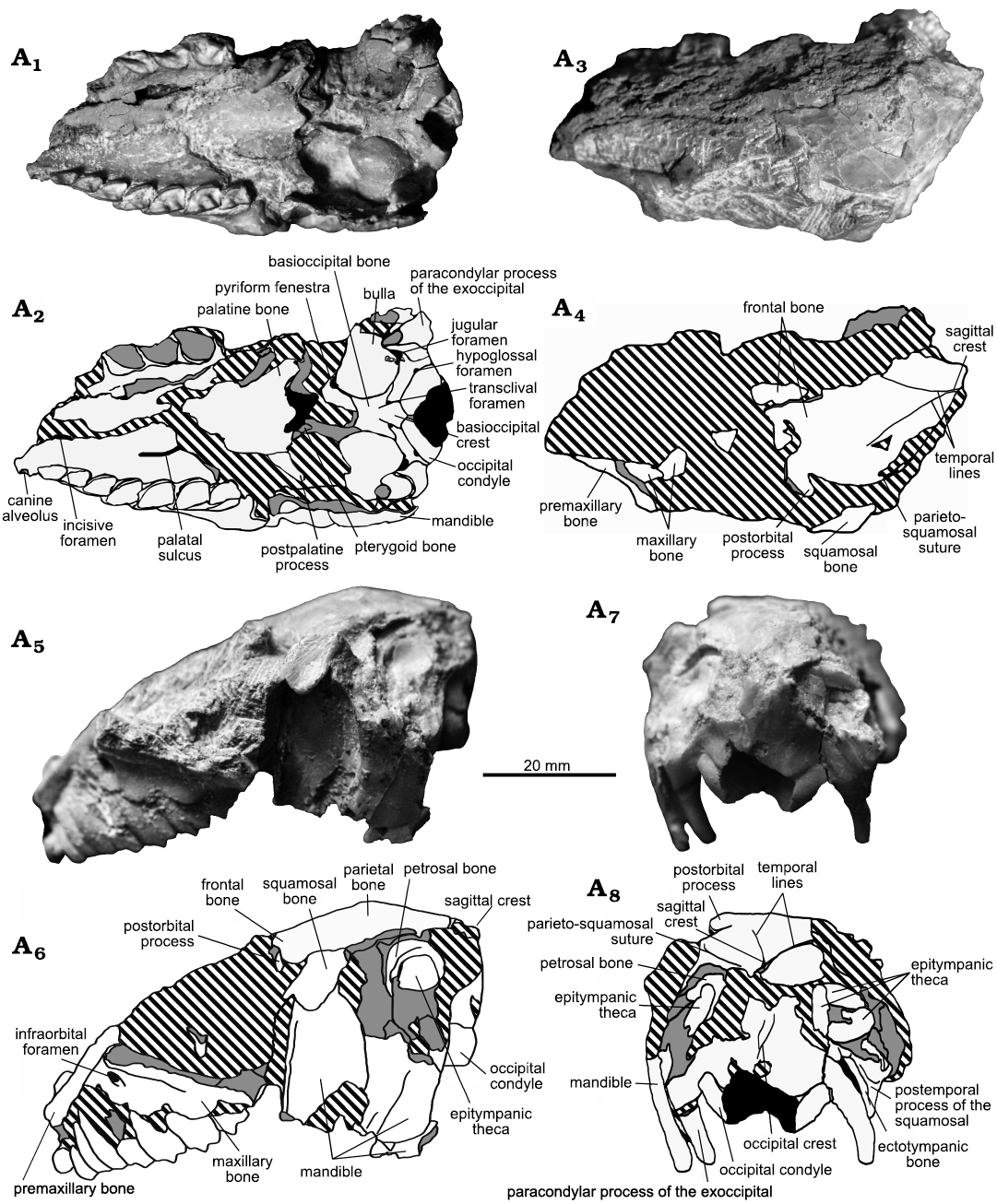
Fig. 1. Hegetotheriid notoungulate “Prohegetotherium schiaffinoi (Kraglievich, 1932)” from Quebrada Fiera, Mendoza, Argentina; upper Oligocene, Deseadan SALMA. MCNAM-PV 5076, skull in ventral (A1, A2), dorsal (A3, A4), left lateral (A5, A6), and occipital (A7, A8) views. White and gray areas indicate well-preserved bone or teeth and broken bone or teeth respectively; black areas indicates foramina, sulci and other natural depresions while striped areas indicate matrix. Photographs (A1, A3, A5, A7) and explanatory drawings (A2, A4, A6, A8).
The MCNAM-PV 5086 (Figs. 2B, 3B) also preserves associated right partial maxilla and mandible. The maxillary fragment preserves P4–M1 in situ and associated isolated tooth fragments, as well as another bone fragment with the M3. Both upper and lower teeth present a centered enamel line on the occlusal surface, indicating a younger wear stage than MCNAM-PV 5076. P4–M1 show a long parastyle and a well-differentiated paracone fold. The M3 shows an undulated ectoloph, with centered concavity and widely convex paracone fold; a smooth concavity is present on the posterior face. This morphology of the M3 coincides with that of specimens of “P. schiaffinoi” already published (see Cerdeño and Reguero 2015: fig. 4). The profile of the ectoloph is also similar to that of “Prohegetotherium malalhuense Cerdeño and Reguero, 2015”, but this species differs in the much more marked and labially directed parastyle and the lesser posterior concavity.
Many other partial maxillae and isolated upper teeth are also assigned to “P. schiaffinoi”, being comparable to those previously published (SOM 1: fig. S3A–D, SOM 2, SOM 3: table 1). Some of them show a different outline between the occlusal and the opposite ends; this is the case for P1 MCNAM-PV 4684 (SOM 1: fig. S3B) and P2 MCNAM-PV 4763 (SOM 1: fig. S3C). P4 MCNAM-PV 4947 (SOM 1: fig. S3D) has a faint posterolingual groove, usually absent in “P. schiaffinoi”; however, some materials assigned to the species show this groove, such as the Bolivian specimen from Salla UF 91661 (Reguero and Cerdeño 2005: fig. 5A).
Concerning mandible and lower teeth of “P. schiaffinoi”, MCNAM-PV 5076 (Fig. 3A) shows a wide mandibular symphysis, its posterior border reaching the level of p3; the height of the horizontal ramus increases from the level of p1 to that of m2; at this level, the ventral border shows an inflexion and elevates posteriorly, although the height of the ramus does not decrease. At the level of m3, the lateroventral masseteric crest is well developed and marks the start of the lateral concavity of the masseteric fossa. There is a mental foramen at the level of p4/m1, close to the ventral border, and another one placed anterior to the p1 level (SOM 1: fig. S2I). These characteristics are also observed in the mandible UF 91661 from Salla (Reguero and Cerdeño 2005: fig. 5B, C). Recently, Reguero et al. (2021) described an incomplete mandible from the Bolivian Petaca Formation (late Oligocene–Early Miocene) as “Prohegetotherium cf. P. schiaffinoi” (originally without quotation marks). These authors highlighted the low horizontal ramus at the level of p4 (height increasing backward) and the thick ventral border. Even though some difference in height is measured at that level (9.6 mm vs. 12.9/12.7 mm; SOM 3: table 2), the specimen is rather close to MCNAM-PV 5076, in which the ventral border is also thick, especially at the level of p4–m2 (Fig. 3A3). Instead, the Bolivian specimen differs in the position of the mental foramen (at m2/m3 level vs. p4/m1 level, and more basally placed, but this feature can vary, e.g., species of Hegetotherium [Sinclair, 1909] and Pachyrukhos [Seoane and Cerdeño, 2019]), the less marked ventral inflexion, and more divergent horizontal rami (Reguero et al. 2021: fig. 4).
The p1 of MCNAM-PV 5076 is procumbent in contrast to the vertical p2; this condition is observable on the holotype of “Ethegotherium carettei” (López 2002; SHDP, FD S, and EC personal observation). The p1 is a short tooth, but a wide and shallow lingual groove differentiates a short anterior lobe from a little longer posterior lobe; the labial face is convex with a subtle angle. The p2 is a much longer tooth, with a long trigonid and even longer and subtriangular talonid; the labial wall makes an inflexion at the limit of both lobes, and there is a well-defined labial groove. In p3 and p4, the talonid is clearly more triangular, wider than the trigonid, which remains squarer in p3 and more rounded in p4; p3 presents a smooth lingual groove between trigonid and talonid, while p4 has a more regularly convex lingual face. Molars are rather similar to p4, but with a deeper labial groove and larger talonid in m1–m2, with a posterolingual fold; the m3 has a longer talonid, with a convex labial face and a posterolingual inflexion. Morphology (Fig. 3A2) and size (SOM 3: table 2) coincide with published remains of “P. schiaffinoi” (Cerdeño and Reguero 2015: table 2).
The mandible MCNAM-PV 5086 preserves (in two fragments) both p3–m3 series, and teeth (Fig. 3B) are very similar to those of MCNAM-PV 5076. Other remains (e.g., MCNAM-PV 5018, PV 5019, and PV 5083; Fig. 3C–E) present m1–m2 with a posterolingual projection and m3 with a lingual fold, as in “P. schiaffinoi” from both Quebrada Fiera and Fray Bentos Formation (Cerdeño and Reguero 2015; Cerdeño et al. 2020). All represent young adult individuals, as they still present the bottom of fossettids.
In turn, MCNAM-PV 5022, with p4–m2 (Fig. 3F), shows the general hegetotheriid tooth morphology, but presents some particularities. The labial sulcus forms a widely open angle, there is a rounded fossettid on the trigonid of p4, an oval fossetid on the most anterior part of the talonid of m2, an occlusal centered line of enamel in m1–m2, and the trigonid is relatively long (wider and more triangular in p4). At first glance, this specimen could be interpreted as a different hegetotheriid species, but it seems more plausible that these differences correspond to juvenile features. Wide labial angles have been observed in deciduous teeth and permanent molars of juvenile individuals of Paedotherium (e.g., Cerdeño et al. 2017: fig. 4M, 4R) and fossettids can be also present in little worn permanent (e.g., Prosotherium garzoni, Seoane et al. 2019: fig. 2C, D) or deciduous teeth (species of Paedotherium, Cerdeño et al. 2017). Therefore, MCNAM-PV 5022 may represent a rather young ontogenetic stage of “Prohegetotherium schiaffinoi”, being tentatively assigned to this species based on the subtle posterolingual projection in m1–m2 and the abundance of this taxon at Quebrada Fiera, although the posterolingual projection shows some variation and is also present in “Prohegetotherium malalhuense” and the Miocene species Hegetotherium mirabile (e.g., Kramarz and Bond 2017: fig. 5). Other lower cheek teeth (SOM 1: fig. S3E, SOM 2, 3) are morphologically coincident with previously published material of “P. schiaffinoi”.
Stratigraphic and geographic range.—Upper Oligocene, Deseadan SALMA; Uruguay: Fray Bentos Formation; Bolivia: Salla, “Upper Salla Beds”; and Argentina: Corrientes and Entre Ríos, Fray Bentos Formation; Mendoza, Agua de la Piedra Formation. ?Middle Member of the Mariño Formation, ?Lower Miocene, Santacrucian SALMA (for “Ethegotherium carettei”, following Reguero and Cerdeño 2005; Cerdeño et al. 2008; Cerdeño and Reguero 2015).
“Prohegetotherium cf. P. schiaffinoi (Kraglievich, 1932)”
Fig. 2C.
Material.—MCNAM-PV 4982, right maxillary fragment with M1–M3 from Quebrada Fiera, Mendoza Province, Argentina, upper Oligocene, Deseadan SALMA.
Description.—MCNAM-PV 4982 cannot be unambiguously attributed to any of the recognized species. This specimen (Fig. 2C) is especially characterized by a M3 that presents a conspicuous posterolingual groove, a marked concavity on its posterior wall, and an undulated ectoloph with a marked paracone fold. The posterolingual groove does not appear in the other recognized taxa of Quebrada Fiera, particularly in the most abundant and closer in size “Prohegetotherium schiaffinoi”, in which the posterior concavity is smoother, though M1–M2 are rather similar to those of MCNAM-PV 4982. In the M3 of “Prohegetotherium malalhuense” (MCNAM-PV 3848, Cerdeño and Reguero 2015), a posterior concavity is observed, but it is very shallow and the posterolingual groove is barely outlined; besides, the particularly differentiated parastyle of “P. malalhuense” is not seen in MCNAM-PV 4982. In turn, a much less conspicuous posterolingual groove and posterior notch are present in the M3 of the Bolivian species Sallatherium altiplanense Reguero and Cerdeño, 2005, from Salla, but M3 is clearly narrower than M1–M2 and the latter are more lingually convex than in MCNAM-PV 4982. The lingual groove in Hegetotheriopsis sulcatus has a different condition, is more anteriorly placed, and is present in the three molars (Kramarz and Paz 2013: fig. 3.1). The M1–M3 of Hemihegetotherium achathaleptum and Hemihegetotherium trilobus have a strongly convex lingual wall and there is neither a posterior notch nor a posterolingual groove in their M3. Thus, due to the established differences, we provisionally keep MCNAM-PV 4982 as “Prohegetotherium cf. P. schiaffinoi”, without discarding a possible intraspecific variation within “P. schiaffinoi”. This specimen is clearly different from the material from Quebrada Fiera formerly identified as Prohegetotherium sp. (Cerdeño and Reguero 2015), material that is now mostly determined as Hegetotheriinae indet. (see below).
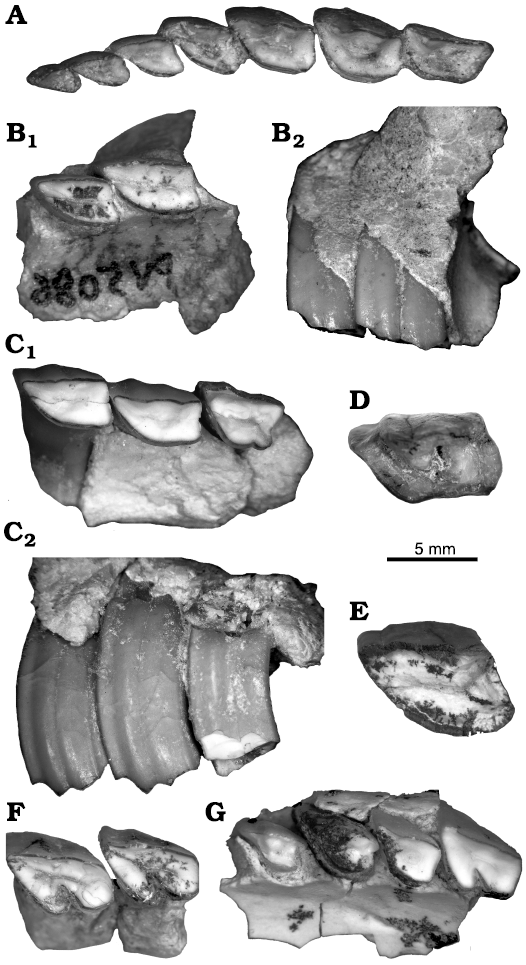
Fig. 2. Upper dentition (anterior to the left) of hegetotheriid notoungulates from Quebrada Fiera, Mendoza, Argentina; upper Oligocene, Deseadan SALMA. A, B. “Prohegetotherium schiaffinoi (Kraglievich, 1932)”. A. MCNAM-PV 5076, left P1–M3 in occlusal view. B. MCNAM-PV 5086, right maxillary fragment with P4–M1 (reversed), in occlusal (B1) and labial (B2) views. C. “Prohegetotherium cf. P. schiaffinoi”, MCNAM-PV 4982, right maxillary fragment with M1–M3 (reversed) in occlusal (C1) and labial (C2) views. D, E. Hegetotheriinae indet. D. MCNAM-PV 4185, left M1 or M2 in occlusal view. E. MCNAM-PV 4612, left P4 or M1 in occlusal view. F, G. Prosotherium garzoni Ameghino, 1897. F. MCNAM-PV 4991, right maxillary fragment with M1–M2 (reversed) in occlusal view. G. MCNAM-PV 5016, right maxillary fragment with P2–M1 (reversed) in occlusal view.
Genus Hegetotheriopsis Kramarz and Paz, 2013
Type species: Hegetotheriopsis sulcatus Kramarz and Paz, 2013; upper Oligocene–Lower Miocene, Argentina.
Hegetotheriopsis sulcatus Kramarz and Paz, 2013
Fig. 3G.
Holotype: MACN Pv CH2015, left portion of rostrum and palate with P1–M3.
Type locality: La Cantera, Chubut Province, Argentina.
Type horizon: Sarmiento Formation, Lower Miocene, Colhuehuapian SALMA.
Material.—MCNAM-PV 3852, left lower premolar (p4?) from Quebrada Fiera, Mendoza Province, Argentina, upper Oligocene, Deseadan SALMA.
Remarks.—Out of the two specimens from Quebrada Fiera reinterpreted by Kramarz and Bond (2017) as Hegetotheriopsis sulcatus, we agree with this determination for the m3 MCNAM-PV 4620. We add the lower premolar MCNAM-PV 3852 (Fig. 3G) to this species. It is similar to the p4 of Hegetotheriopsis sulcatus from the Lower Miocene of the Sarmiento Formation in Bryn Gwyn (Chubut Province), because the trigonid is subcircular and the slightly larger talonid is subtriangular, with the labial groove being perpendicular to the flat lingual wall (Kramarz and Paz 2013: fig. 3), but it differs in having a deeper labial groove, a condition that could be due to the greater degree of wear of MCNAM-PV 3852.
Stratigraphic and geographic range—Upper Oligocene–Lower Miocene, Deseadan–Colhuehuapian SALMA; Mendoza, Río Negro, Neuquén, and Chubut provinces, Argentina.
Hegetotheriinae indet.
Fig. 2D, E; SOM 1: fig. S3F–H.
Material.—11 specimens, mostly isolated upper and lower teeth (detailed list in SOM 2) from Quebrada Fiera, Mendoza Province, Argentina, upper Oligocene, Deseadan SALMA.
Description.—Among the new material, the upper molar MCNAM-PV 4185 (Fig. 2D) varies in contour from the occlusal surface (little worn, with remnants of central fossette or valley) to the opposite preserved end, where the anterolingual face is less angled, with a shorter lingual portion. The parastyle also shortens toward the basal end (SOM 3: table 1). The paracone fold remains along the labial face whereas the metacone fold fades along the crown, disappearing with wear. This opposite outline is similar to “Prohegetotherium schiaffinoi”, but general dimensions are greater. The same happens with M1 MCNAM-PV 4809. MCNAM-PV 4612 (P4 or M1, Fig. 2E) has a narrow parastylar groove throughout the height of the crown. This groove and its occlusal dimensions (SOM 3: table 1) are similar to those in MCNAM-PV 3984 (see remarks below and Cerdeño and Reguero 2015), but the latter differs in having a posterolingual groove. The m3 MCNAM-PV 4621 (SOM 1: fig. S3F) differs from MCNAM-PV 4620, recognized as Hegetotheriopsis sulcatus by Kramarz and Bond (2017), as well as from MACN Pv CH2014 (Hegetotheriopsis sulcatus Kramarz and Paz, 2013), in having a straighter lingual wall, with a smooth posterior concavity, shallower labial groove between trigonid and talonid, and longer and better differentiated “third lobe”. It neither corresponds to the other recognized hegetotheriines with a “third lobe” and similar size: Hemihegetotherium trilobus has more angled lobes and a deeper labial groove, and Hegetotherium mirabile presents a wider second lobe, and a triangular trigonid.
The lower molar MCNAM-PV 4798 (SOM 1: fig. S3H) is somewhat different from teeth assigned to “P. schiaffinoi” due to the presence of two lingual folds, one anterior and one posterior, together with a trigonid–talonid fossettid. However, this tooth is similar to the m1 of the mandible from Salla figured by Cerdeño and Reguero (2015: fig. 5G); the greater plication of the anterior lingual groove and the presence of the fossettid could be due to the younger ontogenetic stage of MCNAM-PV 4798; in fact, the fossettid is not present on the opposite side of the tooth (SOM 1: fig. S3H2; the area of the fold is broken at this side and its possible variation cannot be checked). A roughly comparable plication (but posterolingually placed) has been observed in the p3 of the juvenile mandible MACN A 52-461 of Prosotherium garzoni (Seoane et al. 2019: fig. 2C, D). MCNAM-PV 4798 also differs from Hegetotheriopsis sulcatus, as the latter lacks this kind of lingual folds. The size of MCNAM-PV 4798 is a little greater than that of the m1 of “P. schiaffinoi” (SOM 3: table 2), and we provisionally keep it outside this species.
Remarks.—We here consider several isolated cheek teeth of greater size within the hegetotheriid sample, similar to those previously identified as Prohegetotherium sp. (Cerdeño and Reguero 2015). Taking into account the restricted concept of Prohegetotherium proposed by Kramarz and Bond (2017) (see details above), we opt for determining them as Hegetotheriinae indet. The material is too scarce, not associated, to be confidently defined, and even to positively assure that it represents just one taxon. In this sense, MCNAM-PV 3984, 4406, 4652, 4656, and 4666, published as Prohegetotherium sp. (Cerdeño and Reguero 2015) are also considered now as Hegetotheriinae indet. With respect to MCNAM-PV 3984 (Cerdeño and Reguero 2015: fig. 3A), it was later reinterpreted as Hegetotheriopsis sulcatus (Kramarz and Bond 2017) based on the conspicuous lingual sulcus of this upper molar. However, this groove is more posteriorly placed in MCNAM-PV 3984 than in Hegetotheriopsis sulcatus (Kramarz and Paz 2013: fig. 3.1), which adds to other differences (parastyle labially oriented and delimited by a fine sulcus, and a slightly greater size). So, we consider more appropriate to cautiously remove MCNAM-PV 3984 from Hegetotheriopsis sulcatus and determine it as Hegetotheriinae indet.
Pachyrukhinae Kraglievich, 1934
Genus Prosotherium Ameghino, 1897
Type species: Prosotherium garzoni Ameghino, 1897; Oligocene, Argentina.
Prosotherium garzoni Ameghino, 1897
Figs. 2F, G, 3H, I; SOM 1: fig. S3I–M.
Type material: Lectotype: MACN A 52-455, skull fragment with right and left I1, right P1–P2, and left P1–P3, M1–M3. This specimen was informally proposed as a lectotype by Patterson (1952) and Reguero (1999), and published as such by Seoane et al. (2019). Paralectotype: MACN A 52-456, left mandibular fragment with p2–p4 and m2–m3.
Type locality: Chubut Province, Argentina.
Type horizon: Sarmiento Formation, upper Oligocene, Deseadan SALMA.
Material.—Nine maxillary and mandibular fragments (detailed list in SOM 2) from Quebrada Fiera, Mendoza Province, Argentina, upper Oligocene, Deseadan SALMA.
Description.—MCNAM-PV 5016 (Fig. 2G) corresponds to a young individual, in which P2 is erupting, whereas P3–P4 are already in use; the three premolars present a long parastyle according to their low wear degree. MCNAM-PV 5016, 4991 (Fig. 2F) and 4818 (SOM 1: fig. S3I) preserve bilobed molars and lack a mesostyle, unlike in Medistylus dorsatus (Ameghino, 1903) (see Reguero et al. 2007), which allows them to be recognized as Prosotherium garzoni. All are comparable to published material (Cerdeño and Reguero 2015; first identified as Propachyrucos smithwoodwardi Ameghino, 1897, and later reinterpreted as Prosotherium garzoni by Seoane et al. 2019).
The mandibular fragment MCNAM-PV 5017 (Fig. 3H) presents a mental foramen at the level of p4 (SOM 1: fig. S3J), where the bone is broken; the foramen is deep and the opening extends forwards, but it cannot be checked whether it would form a long groove, as stated by Seoane et al. (2019) for Prosotherium garzoni. The p4 has a shorter talonid than that of m1–m2; the lingual wall is rather straight in p4–m2, but forms a slight posterolingual concavity in m3, at the level of the third lobe; the latter is labially rounded and well defined by the lingual groove. In MCNAM-PV 4992 (Fig. 3I; SOM 1: fig. S3K), the mental foramen is at the posterior level of p3 and a depression extends to p4 and the level of breakage. MCNAM-PV 4993 (SOM 1: fig. S3L) differs in the absence of a clear mental foramen. In MCNAM-PV 4716 (SOM 1: fig. S3M), the two preserved molars are imbricated and their morphology is identical, like all listed specimens, to previously published remains of Prosotherium garzoni (Cerdeño and Reguero 2015); their size (SOM 3) is larger than that of other pachyrukhines, except for Pr. garzoni (Reguero et al. 2007; Seoane et al. 2019).
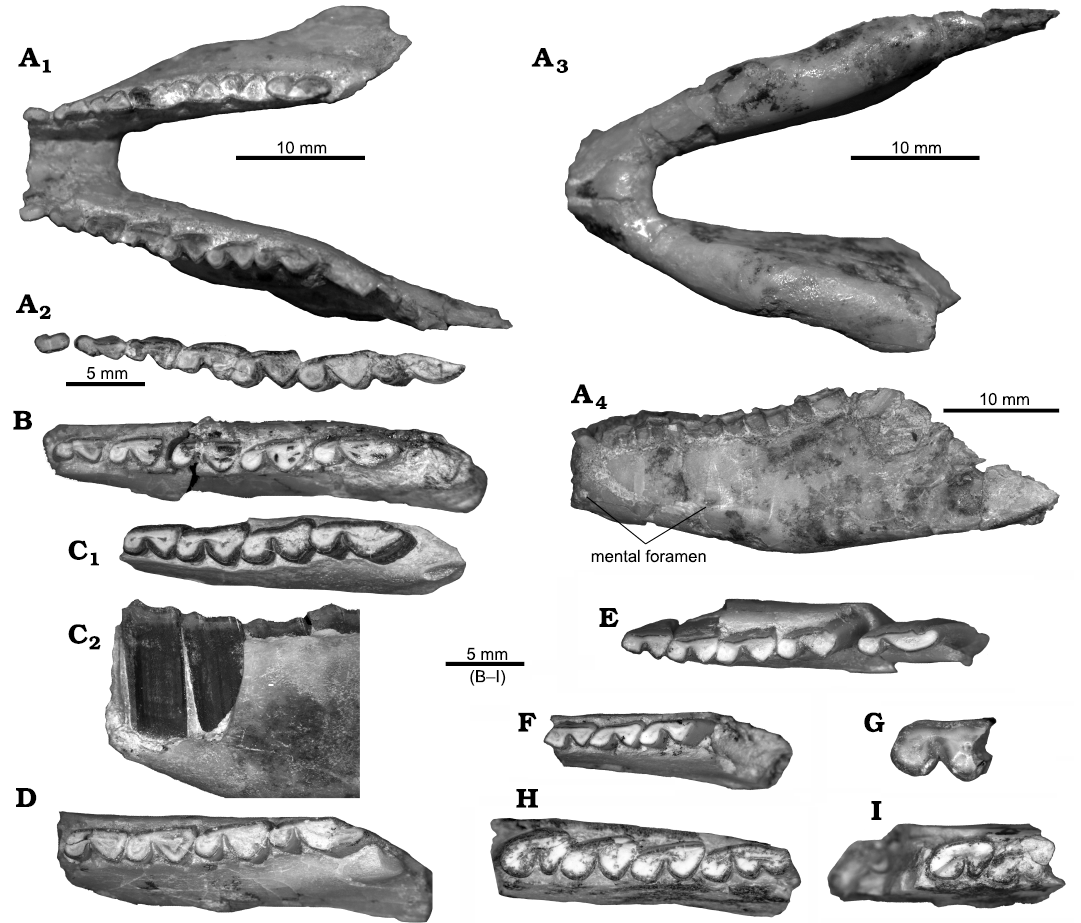
Fig. 3. Lower dentition (anterior to the left) of hegetotheriid notoungulates from Quebrada Fiera, Mendoza, Argentina; upper Oligocene, Deseadan SALMA. A–F. “Prohegetotherium schiaffinoi (Kraglievich, 1932)”. A. MCNAM-PV 5076, mandible with left and right p1–m3 in occlusal (A1), ventral (A3), and left labial (A4) views, occlusal detail of left p1–m3 (A2). B. MCNAM-PV 5086, right mandibular fragment with p3–m3 (reversed) in occlusal view. C. MCNAM-PV 5018, left mandibular fragment with p4–m3 in occlusal (C1) and lingual (reversed) (C2) views. D. MCNAM-PV 5019, left mandibular fragment with p4–m3 in occlusal view. E. MCNAM-PV 5083, left mandibular fragment with p3–m3 in occlusal view. F. MCNAM-PV 5022, left mandibular fragment with p4–m2 in occlusal view. G. Hegetotheriopsis sulcatus Kramarz and Paz, 2013, MCNAM-PV 3852, left premolar (p4?) in occlusal view. H, I. Prosotherium garzoni Ameghino, 1897. H. MCNAM-PV 5017, right mandibular fragment with p4–m3 (reversed) in occlusal view. I. MCNAM-PV 4992, right mandibular fragment with p4–m1 (reversed) in occlusal view.
Stratigraphic and geographic range—Upper Oligocene, Deseadan SALMA; Sarmiento Formation, Chubut Province, and Agua de la Piedra Formation, Mendoza Province, Argentina.
“Archaeohyracidae Ameghino, 1897”
Genus Archaeohyrax Ameghino, 1897
Type species: Archaeohyrax patagonicus Ameghino, 1897, Cabeza Blanca, Chubut Province, Argentina, upper Oligocene, Deseadan SALMA.
Archaeohyrax suniensis
Billet, Patterson, and
de Muizon, 2009
Fig. 4; SOM 1: fig. S4.
Holotype: MNHN-SAL 4: skull and mandible of a young adult, with right I1, I3–M2, left dI1–dI2, I3, P1–M3, right p1–m2 (m3 is covered with matrix), left p2–m3.
Type locality: Salla-Luribay, Bolivia.
Type horizon: Late Oligocene, Deseadan SALMA.
Material.—57 specimens, including cranial, maxillary and mandibular fragments, as well as isolated teeth (detailed list in SOM 2) from Mendoza Province, Argentina, upper Oligocene, Deseadan SALMA.
Description.—The new material of Archaeohyrax suniensis from Quebrada Fiera (SOM 2, 3) allows us to complement the ontogenetic series already observed by Cerdeño et al. (2010) by comparison with the Salla sample (Bolivia; Billet et al. 2009). Concerning upper teeth, MCNAM-PV 4357 (Fig. 4A) is an anterior cranial fragment, in which the premolars and M1 are subtriangular, wider than long, while M2–M3 are still longer than wide, with a central fossette; this specimen corresponds to the ontogenetic stage 9 in Billet et al. (2009: appendix 1A). In contrast, MCNAM-PV 4379 (Fig. 4B) belongs to a juvenile individual, preserving deciduous dentition (left DC and both series DP2–DP4), together with little worn M1 and erupting M2. The small DC is lingually and labially convex, with a centered cusp; it is entirely covered by enamel, while the permanent canine of A. suniensis lacks lingual enamel (Billet et al. 2009); DP2–DP4 are rooted, with a wide paracone fold, an anteriorly extended parastyle, and a central elongated fossette (in DP4, the lophs are in contact but not yet lingually fused); the molars are long and narrow at this stage of wear, and with three fossettes (still open in M2). This specimen corresponds to the ontogenetic stage 3, a little more worn than YPM-PU 22061 from Salla (Billet et al. 2009: fig. 12C), in which DP2–DP4 still retain an anterolabial fossette and the central valley of DP4 is lingually open. Other young individuals are: MCNAM-PV 3983 (Fig. 4C), in which DP3 and DP4 show a protoloph and a metaloph that are not merged, the lingual groove disappearing to the base of the crown; this specimen is similar to MLP 96-XI-20-7, assigned to the wear stage 2 (Cerdeño et al. 2010: fig. 2.4, table 1); and the DP4 MCNAM-PV 4204 (Fig. 4D), with three roots, trapezoid–subtriangular outline, short ectoloph, inclined anterior face, and parastyle not projected; this DP4 is similar to MNHN-SAL 471, from Salla, assigned to the wear stage 4 (Billet et al. 2009: fig. 12B). Other upper teeth of several ontogenetic stages are detailed in SOM 2 and figured in SOM 1: fig. S4A–J, K1, among which MCNAM-PV 4630 and 4945 could correspond to the ontogenetic stage 8.

Fig. 4. Dental remains (anterior to the left) of the archaeohyracid notoungulate Archaeohyrax suniensis Billet, Patterson, and de Muizon, 2009, from Quebrada Fiera, Mendoza, Argentina; upper Oligocene, Deseadan SALMA. A. MCNAM-PV 4357, rostrum and palate with right P2–M3 and left P2–M2 in dorsal (A1), palatal (A2), and right lateral (reversed) (A3) views. B. MCNAM-PV 4379, palate with left DC and both series DP2–DP4–M1–M2 in occlusal view. C. MCNAM-PV 3983, right DP3 (C1) and DP4 (C2) (reversed) of the same individual in occlusal view. D. MCNAM-PV 4204, left DP4 in occlusal (D1) and posterior (D2) views. E. MCNAM-PV 4605, left mandibular fragment with remnants of c–p1 and p2–p4 in labial (E1) and occlusal (E2) views. F. MCNAM-PV 4719, left mandibular fragment with p4–m3 in occlusal view. G. MCNAM-PV 4721, right mandibular fragment with m1–m3 (reversed) in occlusal view. H. MCNAM-PV 4199, left mandibular fragment with alveoli of anterior teeth and fragments of p2–p3 in occlusal view (H1), and associated left m3 in occlusal-labial view (H2). I. MCNAM-PV 4231, left mandibular fragment with p3–p4 in occlusal view. J. MCNAM-PV 4608, left mandibular fragment with m1–m2 in occlusal view. K. MCNAM-PV 4794, associated right premolars (reversed), possible p2 (K1), p3 (K2), and m3 (K3).
Among lower dentition, MCNAM-PV 4605 (Fig. 4E) preserves remnants of c–p1 (procumbent) and little worn p2–p4; the lingual fossettid is already formed in p4, but a short lingual groove is still present in p3 and p2; this specimen corresponds to the stage 6 of Billet et al. (2009: App. 1B). MCNAM-PV 4352 (SOM 3: table 3) belongs to an aged individual and includes maxillary (SOM 1: fig. S4K1) and mandibular (SOM 1: fig. S4K2–K4) fragments that could correspond to ontogenetic stages 11 or 12 (most teeth are incomplete); in the horizontal ramus of the mandible, there are four successive mental foramina at the same height and others much smaller (SOM 1: fig. S4K3), while Billet et al. (2009: 472) mentioned only two constant mental foramina in the material from Salla. Among younger individuals, the m1 MCNAM-PV 3980 (SOM 1: fig. S4N) and the m2 MCNAM-PV 4603 (SOM 1: fig. S4O) show a posterior groove in the entolophid that disappears toward the base of the crown; similarly, the groove between meta- and entolophid closes about 2–3 mm below the occlusal surface, which would originate a fossettid with wear and consequently a flattened lingual wall.
Other specimens (Fig. 4F–K; SOM 3: table 2) can also be correlated with different ontogenetic stages. In summary (see SOM 2), we add the stage 9 (MCNAM-PV 4357) and a juvenile individual of stage 3 (MCNAM-PV 4379) among upper dentitions; besides, comparisons with other specimens from both Quebrada Fiera and Salla allow interpreting more incomplete specimens, such as MCNAM-PV 3983 as stage 2 and MCNAM-PV 4204 as stage 4. With respect to the lower dentition, we add the stage 6 (MCNAM-PV 4605, Fig. 4E) and likely stages 7 (MCNAM-PV 4199, Fig. 4H) and 8 (MCNAM-PV 4794, Fig. 4K); other specimens increase the representation of stages 11 and 12.
Stratigraphic and geographic range—Upper Oligocene, Deseadan SALMA; Salla, Bolivia and Quebrada Fiera, Mendoza, Argentina.
Interatheriidae Ameghino, 1897
Interatheriinae Ameghino, 1897
Genus Argyrohyrax Ameghino, 1897
Type species: Argyrohyrax proavus Ameghino, 1897; upper Oligocene, Argentina.
Argyrohyrax proavus Ameghino, 1897
Fig. 5A–K; SOM 1: fig. S5.
Holotype: MACN A 52-472, palate with left I1–M3 and isolated right I1–I3, C, P1, P3–M3.
Type locality: Cabeza Blanca, Chubut Province, Argentina.
Type horizon: Upper Oligocene, Deseadan SALMA.
Material.—26 specimens including maxillary and mandibular fragments, and isolated teeth (detailed list in SOM 2) from Mendoza Province, Argentina, upper Oligocene, Deseadan SALMA.
Description.—All assigned teeth (SOM 2–3) are comparable in size and morphology to the sample of Argyrohyrax proavus from Quebrada Fiera previously studied (Vera et al. 2017). The new material includes a juvenile individual, MCNAM-PV 4962 (Fig. 5A), preserving DP1–DP4–M1, with more advanced wear than the previously published MCNAM-PV 4059 and 3968 (Vera et al. 2017: fig. 5d, f). These authors interpreted the first premolar as a P1, while Fernández et al. (2021) supported that there is a DP1 without replacement in the Miocene genera Interatherium and Protypotherium. In both MCNAM-PV 4962 and 4059, the first premolar is rooted (a little piece of bone has been removed in both cases for better observation; SOM 1: fig. S5A), like DP2–DP4, but with a higher crown and embedded deeper in the alveolus, which is evidence that it erupted later; this eruption pattern is also observed in the younger MCNAM-PV 3968 (Vera et al. 2017: fig. 5f) and the posterior to anterior eruption pattern has been previously established for interatheriines (Fernández et al. 2021, and references therein). The DP1 (following Fernández et al. 2021) of MCNAM-PV 4962 is simpler than DP2–DP4, subtriangular, longer than wide, with a main labial cusp (paracone) and a short (in height and length) parastyle. DP2–DP3 are characterized by the long parastyle directed anteriorly, less individualized from the paracone fold in DP4; the low crowns of MCNAM-PV 4962 contrast with the observed high crown of P3 in MCNAM-PV 4961 (Fig. 5B). This specimen preserves the left erupting canine (SOM 1: fig. S5A, B) together with the DP1–P2–M2 series in use. The canine has a short parastyle and a low lingual cingulum, as described for DI–DC of MCNAM-PV 3968 (Vera et al. 2017: 616); its paracone fold is more posteriorly placed than in the following premolars; the parastyle projects labially in P3 and to a lesser extent in P4, while it does not project in DP1–P2 or M1–M2. Dental dimensions of MCNAM-PV 4961 fall in the variation range of Argyrohyrax proavus as provided by Vera et al. (2017), although the premolars are slightly smaller (SOM 3: table 5). MCNAM-PV 4961 also preserves the anterior border of the orbit and a large, circular infraorbital foramen at the level of P4, similar to MCNAM-PV 4233 and MACN A 52-475 (Vera et al. 2017: 615, fig. 5a, b). In MCNAM-PV 4986 (Fig. 5C), the parastylar groove is fully visible on M1, reaching the base of the tooth; the paracone fold is more labially extended in P4.
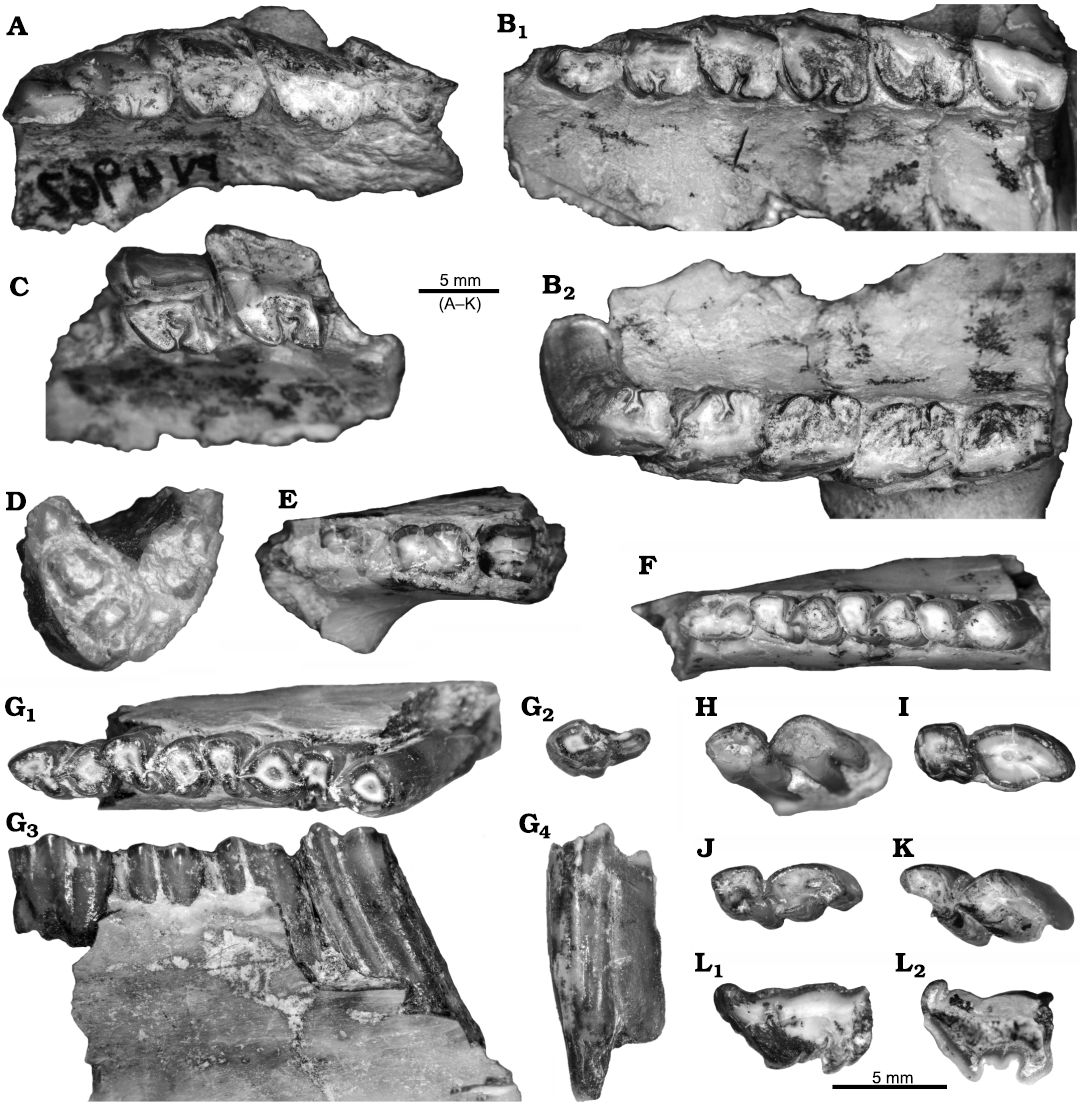
Fig. 5. Dental remains (anterior to the left, except for D) of interatheriid notoungulates from Quebrada Fiera, Mendoza, Argentina; upper Oligocene, Deseadan SALMA. A–K. Argyrohyrax proavus Ameghino 1897. A. MCNAM-PV 4962, right maxillary fragment with DP1–DP4–M1, occlusal view (reversed). B. MCNAM-PV 4961, maxillary fragments with left C–M2 in occlusal view (B1), see erupting C in SOM 1: fig. S5A, B and right P2–M2 in occlusal view (B2). C. MCNAM-PV 4986, left maxillary fragment with M1–M2 in occlusal view. D. MCNAM-PV 4245, mandibular symphysis with broken teeth in rostral view. E. MCNAM-PV 4646, right mandibular fragment with broken dp1–p2–p3 in occlusal view. F. MCNAM-PV 5082, right mandibular fragment with p2–m1 in occlusal view. G. MCNAM-PV 4963, right mandibular fragment with dp3–p4–m2 and separated m3 in occlusal (G1, G2) and lingual (G3, G4) views. H. MCNAM-PV 4308, right lower molar (unerupted m3?) in occlusal view. I. MCNAM-PV 4622, left m3 (reversed) in occlusal view. J. MCNAM-PV 4623, right m3 in occlusal view. K. MCNAM-PV 5081, left m1 or m2 (reversed) in occlusal view. L. Interatheriidae indet. MCNAM-PV 4724, right upper molar in occlusal (reversed) (L1) and opposite (L2) views.
Concerning the lower dentition, MCNAM-PV 4245 (Fig. 5D) preserves fragments of i1–i3, i2 being the largest incisor. MCNAM-PV 4646 (Fig. 5E) bears dp1–p2–p3, p2 clearly larger than dp1 (following the same interpretation as for the upper first premolar); the posterior border of the mandibular symphysis reaches the level of p2 (as in MCNAM-PV 5082, Fig. 5F), broadly coinciding with MACN A 52-474 (Vera et al. 2017: fig. 3a). MCNAM-PV 4676–4678 are p2s (SOM 1: fig. S5C, D) of different individuals, rooted, with a V-shaped trigonid with a convex labial face, and a short talonid; they are similar in morphology and size (SOM 3: table 2) to the p2 of MCNAM-PV 4697 (Vera et al. 2017: fig. 6d, erroneously identified as a p1 in the figure caption). MCNAM-PV 4963 (Fig. 5G) shows dp3 (with the start of roots) and p4–m2 in situ, and m3 separated; both m2 and m3 show a high crown and part of the roots. In MCNAM-PV 4970, the preserved p4 is high, little worn (narrow union between trigonid and talonid), and the open base of the tooth shows a curiously wrinkled enamel in the labial wall of the trigonid (SOM 1: fig. S5E). In MCNAM-PV 4607 (SOM 1: fig. S5F), trigonid and talonid are still unfused in the m3; its U-shaped trigonid is similar to that of the young individual MACN A 52-473 (Vera et al. 2017: fig. 3l, holotype of “Notohyrax conicus”). MCNAM-PV 4308 (Fig. 5H) is a barely worn molar (m3?); occlusally, each lobe shows a centered irregular depression that would be obliterated soon with wear; the length of the trigonid increases to the crown base and the metaconid area extends a little backward, delimited by an anterior groove; the talonid presents two lingual grooves. Similarly, the m3s MCNAM-PV 4622 and 4623, and the m1 or m2 MCNAM-PV 5081 (Fig. 5I–K) are little worn, with a narrow contact between the trigonid and talonid.
Stratigraphic and geographic range.—Upper Oligocene, Deseadan SALMA; Sarmiento Formation, Santa Cruz and Chubut provinces; base of the Agua de la Piedra Formation, Mendoza Province; and Fray Bentos Formation, Corrientes Province, Argentina.
Interatheriinae indet.
Fig. 5L.
Material.—MCNAM-PV 4724, isolated right upper molar (M2 or M3) from Mendoza Province, Argentina, upper Oligocene, Deseadan SALMA.
Description.—MCNAM-PV 4724 is a little worn cheek tooth of a juvenile or young adult individual. It is long and narrow, with labially extended parastyle (Fig. 5L1); it has an anterolabial fossette and a posterolingual groove on the lingual edge of the metaloph, but this groove disappears toward the base. Looking at the opposite end (Fig. 5L2), the outline changes, with an open-Y-shaped median valley, the metaloph lingually shorter, and the parastyle greatly curved posterolabially, separated from the paracone fold by a marked groove. The direction of the parastyle on the occlusal face resembles that of a M2 of Argyrohyrax proavus (e.g., Vera et al. 2017: fig. 2a, 3b), but more extended labially; in addition, the marked groove and the direction of the parastyle at the opposite end do not correspond to this species, as far as it can be observed in its holotype (Vera et al. 2017: fig. 3b). A second interatheriine was recognized at Quebrada Fiera as Progaleopithecus sp. (Vera et al. 2017: fig. 6r), based on lower teeth. The upper teeth of Progaleopithecus sp. are closer to MCNAM-PV 4724 in the labial projection of the parastyle, in both premolars and molars, as in Progaleopithecus tournoueri Ameghino, 1904 (Vera et al. 2017: fig. 7k, l), but these teeth differ in the absence of a median lobe and the greater projection of the paracone fold. Instead, MCNAM-PV 4724 appears closer to the interatheriines from Salla, Bolivia (Hitz et al. 2008). Both Brucemacfaddenia boliviensis Hitz, Billet, and Derryberry, 2008, and Federicoanaya sallaensis Hitz, Billet, and Derryberry, 2008, present upper cheek teeth with a median lobe and a labially extended parastyle in molars; B. boliviensis also has a greatly extended parastyle in premolars, an anterolabial fossette in little worn molars, and its size is closer to MCNAM-PV 4724 (F. sallaensis is a smaller species). Due to the lack of a total similarity, we determine MCNAM-PV 4724 as Interatheriinae indet., but keeping in mind its possible relationship with B. boliviensis. On the other hand, this specimen is not comparable with the other undetermined interatheriid from Quebrada Fiera described by Vera et al. (2017: 622), because the latter (MCNAM-PV 4742) is a mandibular fragment with p4–m1.
Conclusions
The present paper provides an updated overview of the diversity of small typotherians (Notoungulata) from the Deseadan (late Oligocene) fauna of Quebrada Fiera (southern Mendoza Province, Argentina). A total of 141 specimens are added to the previously described material, 57 specimens belonging to Hegetotheriidae, 57 to “Archaeohyracidae”, and 27 to Interatheriidae, adding novelties for each of these groups.
Among hegetotheriids, a relatively complete skull improves our knowledge of cranial characters of “Prohegetotherium schiaffinoi”, particularly for what concerns the caudal cranium. Only a few comparisons are possible with previously described cranial fragments of “P. schiaffinoi”; in turn, differences are established with the better-known Miocene hegetotheriids Hegetotherium mirabile and Hemihegetotherium achathaleptum, as well as other members of the group: relatively larger foramen magnum; postorbital process slightly curved backwards unlike in Hegetotherium mirabile and Hemihegetotherium trilobus; paracondylar processes shorter and more conical than in Hegetotherium mirabile; bulky tympanic bullae, extended far more laterally than in Hegetotherium mirabile, Hemihegetotherium achathaleptum, and Hemihegetotherium trilobus; crest in the basioccipital relatively shorter and bulkier than in Hegetotherium mirabile, Hemihegetotherium achathaleptum, and Hemihegetotherium trilobus, and with two lateral foramina unlike in the three latter species and Pachyrukhinae. A maxillary fragment displays some differences with “P. schiaffinoi” in the morphology of M3, being also different from the other currently recognized hegetoheriines, which leads to maintaining MCNAM-PV 4982 in open nomenclature (“Prohegetotherium cf. P. schiaffinoi”), without discarding the possibility that it might be part of the intraspecific variation of “P. schiaffinoi”. New, relatively large isolated teeth add to those previously identified as Prohegetotherium sp. (Cerdeño and Reguero 2015); this determination is herein reassessed, and this material is assigned to Hegetotheriinae indet., taking into account the still unresolved taxonomy of Prohegetotherium. The revision of some specimens interpreted by Kramarz and Bond (2017) as belonging to Hegetotheriopsis sulcatus leads to discard this attribution for the specimen MCNAM-PV 3984, identifying it instead as Hegetotheriinae indet. A newly described lower premolar is now assigned to Hegetotheriopsis sulcatus, thus supporting the presence of this species at Quebrada Fiera. Partial upper and lower dental series add to the published sample of the pachyrukhine Prosotherium garzoni.
Concerning archaeohyracids, the known ontogenetic sequence of the species Archaeohyrax suniensis is complemented with specimens representing several stages not previously observed within the Quebrada Fiera sample (Cerdeño et al. 2010). No new specimen is attributed to the second archaeohyracid species previously recorded at Quebrada Fiera (“Archaeohyracidae” indet. cf. Archaeotypotherium; Cerdeño et al. 2010).
Finally, among interatheriids, new juvenile and adult specimens add to the sample of Argyrohyrax proavus. No new material is attributed to Progaleopithecus (Vera et al. 2017). In contrast, an isolated upper molar seems to represent a different taxon; its morphology is closer to that of the Bolivian interatheriines from Salla (Hitz et al. 2008), especially Brucemacfaddenia boliviensis, although an accurate determination cannot be achieved; this similarity strengthens the links between the mammal faunas from Quebrada Fiera and Salla. Despite this, the number of shared taxa between these two localities is low, taking into account that out of the 39 mammals recognized at Quebrada Fiera, only four coincide at the species level with those from Salla: Incamys bolivianus, Archaeohyrax suniensis, “Prohegetotherium schiaffinoi”, and Pyrotherium romeroi (see Croft 2016; Pujos et al. 2021; Candela et al. 2021; and references therein); general similarities are greater at the genus level. As a whole, the mammal fauna from Quebrada Fiera has more affinities with Patagonian faunas than with lower latitudinal faunas, and also includes several exclusive taxa (e.g., Cerdeño and Vera 2015; Candela et al. 2021, and references therein).
Acknowledgements
Thanks are due to the editorial staff and reviewers: Marcelo Reguero (Museo de La Plata, Argentina), Alejandro Kramarz (Museo Argentino de Ciencias Naturales “Bernardino Rivadavia”, Buenos Aires, Argentina), and Daniel García-López (Facultad de Ciencias Naturales e Instituto Miguel Lillo, Tucumán, Argentina), whose comments allowed improving this manuscript. We especially thank Mercedes Fernández (Universidad de Luján, Buenos Aires, Argentina), for a fruitful discussion on interatheriine teeth. We acknowledge all participants in the fieldwork at Quebrada Fiera throughout years. This work is a contribution to the scientific project ANPCyT-PICT 2017-0797 (Argentina) and R-430 FDS’s contribution to the IDEAN (Instituto de Estudios Andinos “Don Pablo Groeber”, Buenos Aires, Argentina).
References
Ameghino, F. 1887. Enumeración sistemática de las especies de mamíferos fósiles coleccionados por Carlos Ameghino en los terrenos eocenos de Patagonia austral y depositados en el Museo de La Plata. Boletín del Museo de La Plata 1: 1–26.
Ameghino, F. 1894. Enumération synoptique des espèces de mammifères fossiles des formations éocènes de Patagonie. 196 pp. Imprenta P.E. Coni & hijos, Buenos Aires. Crossref
Ameghino, F. 1897. Les mammifères crétacés de l’Argentine. Deuxième contribution à la connaissance de la faune mammalogique des couches à Pyrotherium. Boletín del Instituto Geográfico Argentino 18: 1–117.
Ameghino, F. 1902. Première contribution à la connaissance de la fauna mammalogique des couches à Colpodon. Boletin de la Academia Nacional de Ciencias de Córdoba 17: 71–141.
Ameghino, F. 1903. Nuevas especies de mamíferos cretáceos y terciarios de la República Argentina. Anales de la Sociedad Científica Argentina 56: 193–208.
Ameghino, F. 1904. Nuevas especies de mamíferos cretáceos y terciarios de la República Argentina. Anales de la Sociedad Científica Argentina 57: 162–175; 327–341; 58: 35–42, 56–71, 182–192, 225–291.
Billet, G., Patterson, B., and de Muizon, C. 2009. Craniodental anatomy of late Oligocene archaeohyracids (Notoungulata, Mammalia) from Bolivia and Argentina and new phylogenetic hypotheses. Zoological Journal of Linnean Society 155: 458–509. Crossref
Bond, M. and Pascual, R. 1983. Nuevos y elocuentes restos craneanos de Proborhyaena gigantea Ameghino, 1897 (Marsupialia, Borhyaenidae, Proborhyaeninae) de la Edad Deseadense. Un ejemplo de coevolución. Ameghiniana 20: 47–60.
Candela, A.M., A., Pérez, M.E., Rasia, L., and Cerdeño, E. 2021. New caviomorph rodents from the late Oligocene of Mendoza Province, central-western Argentina. Journal of Vertebrate Paleontology 41: e1929264. Crossref
Cerdeño, E. 2014. First record of Mesotheriidae in the late Oligocene of Mendoza Province, Argentina. Ameghiniana 51: 366–370. Crossref
Cerdeño, E. and Reguero, M. 2015. The Hegetotheriidae (Mammalia, Notoungulata) assemblage from the late Oligocene of Mendoza, central-western Argentina. Journal of Vertebrate Paleontology 35: e907173. Crossref
Cerdeño, E. and Vera, B. 2014. New data on diversity of Notohippidae from the Oligocene of Mendoza, Argentina. Journal of Vertebrate Paleontology 34: 941–950. Crossref
Cerdeño, E. and Vera, B. 2015. A new Leontinidae (Notoungulata) from the late Oligocene beds of Mendoza Province, Argentina. Journal of Systematic Paleontology 13: 943–962. Crossref
Cerdeño E., López, G., and Reguero, M. 2008. Biostratigraphic considerations of the Divisaderan faunal assemblage. Journal of Vertebrate Paleontology 28: 574–577. Crossref
Cerdeño, E., Montalvo, C.I., and Sostillo, R. 2017. Deciduous dentition and eruption pattern in late Miocene Pachyrukhinae (Hegetotheriidae, Notoungulata) from La Pampa Province, Argentina. Historical Biology 29: 359–375. Crossref
Cerdeño, E., Reguero, M., and Vera, B. 2010. Deseadan Archaeohyracidae (Notoungulata) from Quebrada Fiera (Mendoza, Argentina) in the paleobiogeographic context of the South American late Oligocene. Journal of Paleontology 84: 1177–1187. Crossref
Cerdeño, E., Schmidt, G.I., Miño-Boilini, A.R., and Zurita, A.E. 2020. New data on the diversity of Notoungulata (Mammalia) from the Fray Bentos Formation (late Oligocene) in Corrientes Province, Argentina. Ameghiniana 57: 443–461. Crossref
Combina, A.M. and Nullo, F. 2011. Ciclos tectónicos, volcánicos y sedimentarios del Cenozoico del sur de Mendoza-Argentina (35°–37°S y 69°300W). Andean Geology 38: 198–218.
Croft, D.A. 2016. Horned Armadillos and Rafting Monkeys. The Fascinating Fossil Mammals of South America. 304 pp. Indiana University Press, Bloomington.
Croft, D.A. and Anaya, F. 2006. A new middle Miocene hegetotheriid (Notoungulata: Typotheria) and a phylogeny of the Hegetotheriidae. Journal of Vertebrate Paleontology 26: 387–399. Crossref
Fernández, M., Fernicola, J.C., and Cerdeño, E. 2021. Deciduous dentition and dental eruption sequence in Interatheriinae (Notoungulata, Interatheriidae): implications in the systematics of the group. Journal of Paleontology 95: 861–885. Crossref
Gorroño, R., Pascual, R., and Pombo, R. 1979. Hallazgo de mamíferos eógenos en el sur de Mendoza. Su implicancia en las dataciones de los “rodados lustrosos” y del primer episodio orogénico del Terciario en esa región. In: VII Congreso Geológico Argentino, Neuquén (1978), Relatorio, 475–487. Asociación Geológica Argentina, Neuquén.
Hitz, R.B., Billet, G., and Derryberry, D. 2008. New interatheres (Mammalia, Notoungulata) from the late Oligocene Salla beds of Bolivia. Journal of Paleontology 82: 447–469. Crossref
Kraglievich, L. 1932. Nuevos apuntes para la geología y paleontologia uruguayas. Anales del Museo de Historia Natural de Montevideo 3: 1–65.
Kraglievich, L. 1934. La antigüedad pliocena de las faunas de Monte Hermoso y Chapadmalal, deducidas de su comparación con las que le precedieron y sucedieron. 168 pp. El Siglo Ilustrado, Montevideo.
Kramarz, A. and Bond, M. 2017. Systematics and stratigraphical range of the hegetotheriids Hegetotheriopsis sulcatus and Prohegetotherium sculptum (Mammalia: Notoungulata). Journal of Systematic Palaeontology 15: 1027–1036. Crossref
Kramarz, A. and Paz, E.R. 2013. Un Hegetotheriidae (Mammalia, Notoungulata) basal del Mioceno temprano de Patagonia. Revista Mexicana de Ciencias Geológicas 30: 186–195.
Linnaeus, C. 1758. Systema naturæ per regna tria naturæ, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Tomus I. Editio decima, reformata. 824 pp. Laurentius Salvius, Stockholm. Crossref
López G.M. 2002. Redescripción de Ethegotherium carettei (Notoungulata, Hegetotheriidae) de la Formación Divisadero Largo de la provincia de Mendoza, Argentina. Ameghiniana 39: 295–306.
MacPhee, R.D.E. 2014. The serrialis bone, interparietals, “X” elements, entotympanics, and the composition of the Notoungulate caudal cranium. Bulletin of the American Museum of Natural History 384: 1–69. Crossref
MacPhee, R.D.E. and Forasiepi, A.M. 2022. Re-evaluating cranial pathways of the internal carotid artery in Notoungulata (Mammalia, Panperissodactyla). Ameghiniana 59: 141–161. Crossref
MacPhee, R.D.E., Hernández Del Pino, S., Kramarz, A., Forasiepi, A.M., Bond, M., and Sulser, R.B. 2021. Cranial morphology and phylogenetic relationships of Trigonostylops wortmani, an Eocene South American Native Ungulate. Bulletin of the American Museum of Natural History 449: 1–183. Crossref
Minoprio, J.L. 1947. Fósiles de la Formación del Divisadero Largo. Anales de la Sociedad Científica Argentina 146: 365–378.
Pascual, R. and de la Fuente, M.S. 1993. Vertebrados fósiles cenozoicos. In: V. Ramos (Ed.), XII Congreso Geológico Argentino y II Congreso de Exploración de Hidrocarburos (Mendoza, 1993). Geología y Recursos Naturales de Mendoza. Relatorio 2: 357–363.
Patterson, B. 1952. Catálogo de los mamíferos del Deseadiano y Colhuehapiano. Informe Sección Paleozoológica Vertebrados Museo Argentino de Ciencias Naturales “Bernardino Rivadavia”. 4 pp. Unpublished report, Museo Argentino de Ciencias Naturales “Bernardino Rivadavia”, Buenos Aires.
Pujos, F., Ciancio, M.R., Forasiepi, A.M., Pujos, M., Candela, A.M., Vera, B., Reguero, M.A., Combina, A.M., and Cerdeño, E. 2021. The Late Oligocene xenarthran fauna of Quebrada Fiera (Mendoza, Argentina) and its implications for sloth origins and the diversity of Paleogene cingulates. Papers in Palaeontology 7: 1613–1656. Crossref
Reguero, M.A. 1999. El problema de las relaciones sistemáticas y filogenéticas de los Typotheria y Hegetotheria (Mammalia, Notoungulata): análisis de los taxones de Patagonia de la edad-mamífero Deseadense (Oligoceno). 350 pp. Unpublished Ph.D. dissertation, Universidad Nacional de Buenos Aires, Buenos Aires.
Reguero, M.A. and Cerdeño, E. 2005. New Late Oligocene Hegetotheriidae (Mammalia, Notoungulata) from Salla, Bolivia. Journal of Vertebrate Paleontology 25: 674–684. Crossref
Reguero M.A., Dozo, M.T., and Cerdeño, E. 2007. A poorly known rodent-like mammal (Pachyrukhinae, Hegetotheriidae, Notoungulata) from the Deseadan (Late Oligocene) of Argentina. Paleoecology, biogeography and radiation of the rodent-like ungulates in South America. Journal of Paleontology 81: 1298–1304. Crossref
Reguero, M.A., Tineo, D.E., Bona, P., Pérez, L.M., Vergani, G.D., González Ruiz, G., and Poiré, D.G. 2021. A singular Hegetotheriinae (Notoungulata, Typotheria) from the late Oligocene–Early Miocene of the Subandean Region of Bolivia. Brazilian Journal of Geology 51: e20200067. Crossref
Roth, S. 1903. Noticias preliminares sobre nuevos mamíferos fósiles del Cretáceo superior y Terciario inferior de la Patagonia. Revista del Museo de La Plata 11: 133–158.
Rovereto, C. 1914. Los estratos araucanos y sus fósiles. Anales del Museo Nacional de Historia Natural de Buenos Aires 25: 1–250.
Seoane, F.D. and Cerdeño, E. 2019. Systematic revision of Hegetotherium and Pachyrukhos (Hegetotheriidae, Notoungulata) and a new phylogenetic analysis of Hegetotheriidae. Journal of Systematic Palaeontology 17: 1415–1443. Crossref
Seoane, F.D., Cerdeño, E., and Singleton, H. 2019. Re-assessment of the Oligocene genera Prosotherium and Propachyrucos (Hegetotheriidae, Notoungulata). Comptes Rendus Palevol 18: 643–662. Crossref
Sinclair, W.J. 1909. Typotheria of the Santa Cruz Beds. In: W.B. Scott (ed.), Reports of the Princeton University Expeditions to Patagonia, 1896–1899, Vol. 6, Paleontology. Mammalia of the Santa Cruz Beds, Part 1, 1–110. Schweizerbart’sche Verlagshandlung, Stuttgart.
Vera, B., Cerdeño, E., and Reguero, M. 2017. The Interatheriinae from the Late Oligocene of Mendoza (Argentina), with comments on some Deseadan Interatheriidae. Historical Biology 29: 607–626. Crossref
Zittel, K.A. 1893. Handbuch der Palaeontologie, I. Palaeozoologie. IV, Vertebrata (Mammalia). 799 pp. R. Oldenbourg, Munich.
Acta Palaeontol. Pol. 67 (4): 983–997, 2022
https://doi.org/10.4202/app.00974.2022