
Further desmostylian remains from the upper Oligocene of Vancouver Island, British Columbia, Canada
BRIAN LEE BEATTY
The North Pacific rim was home to an extinct group of semiaquatic marine mammals, the order Desmostylia, which superficially resembled hippos. Desmostylians are an uncommon fossil vertebrate in most localities where they occur, and Oligocene taxa particularly so. Beyond the type dentition and two femora of Cornwallius sookensis, and the skull and postcrania of Behemotops proteus, both previously described from the region of Muir Creek locality of the upper Oligocene Sooke Formation of southern coast of Vancouver Island, British Columbia, Canada, no other records from this site have been reported. Additional specimens of teeth have now been found in the Royal British Columbia Museum and Canadian Museum of Nature, including a long-forgotten and unreported paratype. In total, three additional teeth, one of B. proteus and two possibly belonging to C. sookensis, are reported here from its type locality. More importantly, the upper molar reported here, as well as UCMP 36078 from Baja California and USNM 181740 from Oregon, all resemble each other but should be considered as belonging to Cornwallius only with caution.
Introduction
Desmostylians are an extinct group of semiaquatic marine mammals known from the North Pacific, from Japan to Baja California, Mexico. Fossils of this enigmatic order have been found in surprising abundance in one locality, Monocline Ridge, in central California, but are usually quite rare and fragmentary elsewhere, making it common for specimens of significant quality to be known by names derived from their locality (e.g., “Izumi specimen” or “Stanford Paleoparadoxia”). Despite this, it is clear that even fragmentary specimens are important to our understanding of their distributions and diversity, as some specimens are found in unusual places or are unlike anything we would expect from known ranges of variation, due to polymorphism or ontogeny observable in larger samples. Recent finds on Vancouver Island, British Columbia, Canada (Beatty and Cockburn 2015) have proven that it is wise to be cautious when first identifying specimens, as assumptions about morphology of unknown regions of the body based on similarities between known parts may be flawed, as we still have relatively little information about this group and its relationships. Here I present some newly recognized specimens of desmostylians from Vancouver Island that may ultimately represent new taxa, though the previously expressed cautions suggest that they should simply be noted so we may be cognizant of such potential diversity.
All the specimens described below were found in the area near Kirby Creek on Vancouver Island (Fig. 1). This small waterway cuts through sediments of the Sooke Formation, which is considered latest Oligocene in age (24.1–24.8 Ma; Prothero et al. 2008; Nesbitt et al. 2010; Beatty and Cockburn 2015). Other Desmostylia have been found from the Sooke Formation near Kirby Creek, including the type specimen (two molars) of Cornwallius sookensis from the beach near Muir Creek (Cornwall 1922; Hay 1923; Beatty 2006a, 2009) and Behemotops proteus (Beatty and Cockburn 2015). Kirby Creek flows into the Salish Sea less than a kilometer from Muir Creek Beach, and exposures of the Sooke Formation can be followed along the cliffs of the beach between the two. It seems that the specimens below came from the same horizon as the specimens of C. sookensis (Cornwall 1922; Hay 1923; Beatty 2006a, 2009) and B. proteus (Beatty and Cockburn 2015).
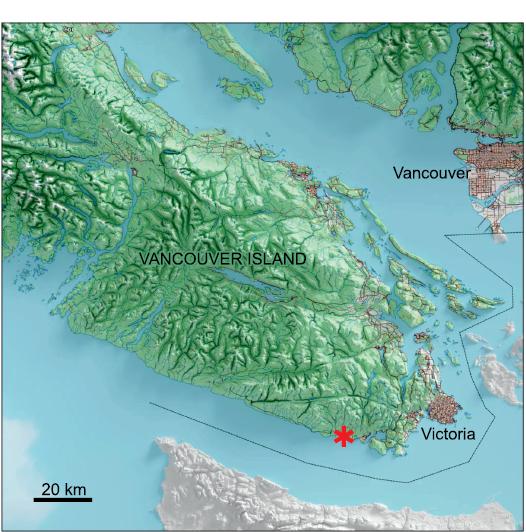
Fig. 1. Map of the Vancouver region (British Columbia, Canada) with Sooke Formation exposures that produced the desmostylian specimens reported here (marked with an asterisk). The dashed line indicates the border between the USA and Canada.
Institutional abbreviations.—CMN, Canadian Museum of Nature, Ottawa, Canada; IGM, Instituto Nacional de Investigaciones Geologico-Mineras, Bogota, Colombia; LACM, Natural History Museum of Los Angeles County, USA; RBCM, Royal British Columbia Museum, Victoria, Canada; UCMP, University of California Museum of Paleontology, Berkeley, USA; UOMNCH, University of Oregon Museum of Natural and Cultural History, Eugene, USA; USNM, National Museum of Natural History, Smithsonian Institution, Washington, DC, USA.
Other abbreviations.—We follow standard convention in abbreviating tooth families as I, C, P, and M, with upper and lower case letters referring to upper and lower teeth, respectively.
Systematic palaeontology
Class Mammalia Linnaeus, 1758
Order Desmostylia Reinhart, 1953
Family indeterminate
Genus Behemotops Domning, Ray, and McKenna, 1986
Behemotops cf. B. proteus Domning, Ray, and McKenna, 1986
Fig. 2.
Material.—LACM 124106, right lower postcanine teeth (Ray et al. 1994) from LACM locality 5123, Disque Quadrangle, Washington, Pysht Formation, late Oligocene; RBCM.EH2007. 008.0001, a partial skull, including canine tusk and two molars, and 12 thoracic vertebrae, 17 ribs (Beatty and Cockburn 2015) from Sombrio River in Juan De Fuca Provincial Park (Vancouver Island, Canada), from the late Oligocene Sooke Formation. RCBM EH2008 010 0001 (Fig. 2) is a single cusp from an upper molar from near Kirby Creek (Vancouver Island, Canada), from the late Oligocene Sooke Formation.
Description.—RCBM EH2008 010 0001 is a single cusp from an almost desmostylodont tooth (Fig. 2) that resembles the paracone of Behemotops proteus from Muir Creek (RBCM.EH2007.008.0001; Beatty and Cockburn 2015). Desmostylodont teeth are those having columnar cusps made of thick enamel, which are considered to be traits of Paleoparadoxia, Cornwallius, and Desmostylus (Clark 1991). This cusp, in isolation, appears almost columnar, particularly because of its almost vertical edge. This is the feature that most resembles the paracone or metacone of the left M2 of B. proteus, RBCM.EH2007.008.0001. The paracone and metacone of RBCM.EH2007.008.0001 and RCBM EH2008 010 0001 lack the cingulum that is otherwise characteristic of the postcanine teeth of B. proteus (Doming et al. 1986; Ray et al. 1994). The size of RCBM EH2008 010 0001 is also similar to the paracone or metacone of B. proteus. The cusp of RBCM EH2008 010 0001 is smaller than the equivalent cusps of C. sookensis, which also lack a cingulum, but are larger than this new specimen.
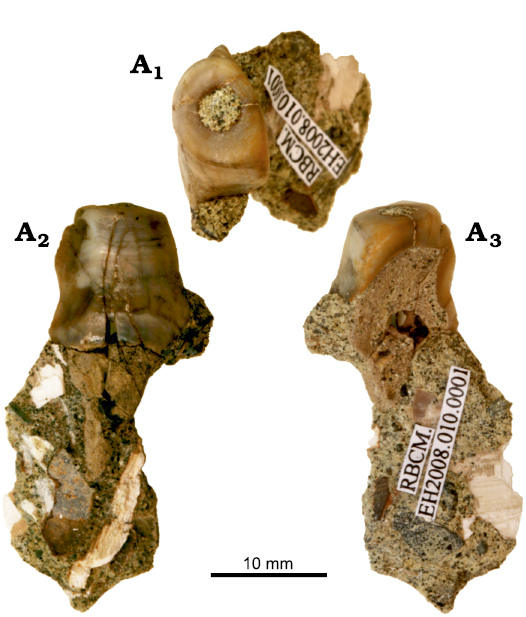
Fig. 2. Desmostylian mammal Behemotops cf. B. proteus Domning, Ray, and McKenna, 1986 (RBCM EH2008 010 0001) from Sooke Formation, late Oligocene, Muir Creek, Vancouver Island, British Columbia, Canada. Partial molar tooth, in occlusal (A1), lingual (A2), and buccal (A3) views.
Family Desmostylidae Osborn, 1905
Genus Cornwallius Hay, 1923
Cornwallius sookensis (Cornwall, 1922)
Figs. 3, 4.
Holotype: RBCM 486, a left lower molar. According to Cornwall (1922) the “type” is a tooth, the same specimen described by Lawrence M. Lambe in Kermode (1917); he also describes a second tooth found in 1921. Of these two teeth, RBCM 486 (a left lower molar) and RBCM 491 (a right M3), the former is the holotype (Cornwall 1922: fig. 1A; Hay 1924: pl. 2: 1–3), and is most likely an m1 or m2 (Beatty 2009).
Type locality: Cliffs between Coal (Kirby) and Muir creeks, Vancouver Island, British Columbia, Canada.
Type horizon: Sooke Formation, Zemorrian (late Oligocene).
Material.—RBCM 491, a right M3 (Cornwall 1922: fig. 1B; Hay 1923: fig. 4; Hay 1924: text-fig. 1, pl. 1: 4, 5, pl. 2: 4, 5) from Vancouver Island Sooke Formation, late Oligocene. UCMP 32682, a fragment of a lower molar including two cusps (Clark and Arnold 1923: fig 3a, b; VanderHoof 1937) both of which were found from between Coal (Kirby) and Muir creeks, Vancouver Island, British Columbia, Canada; UCMP 36078, a right M1 and UCMP 36079, a right m2; from Bahia San Carlos, previously reported as San Telmo Point (VanderHoof 1942), and Punta San Telmo, Baja California, Mexico (VanderHoof 1942: pl. 1). USNM 157811, an isolated partial adult lower molar, comprising two straight, moderately hypsodont columnar cusps still partially embedded in matrix and including intercuspal matrix that appears to include remains of crushed shells (Beatty 2009); USNM 181738, a nearly complete (edentulous) and only slightly laterally distorted skull of a small individual, which is most likely a subadult (Beatty 2009); USNM 181739, an adult edentulous left mandibular ramus (Beatty 2009); USNM 181740, skull of a large, presumably adult, individual with left and right C1, M2, M3 and additional roots of incisors and one M1 (Reinhart 1982; Beatty 2009); USNM 181741, a right mandible with two molariform teeth, which are either m1 and m2, or m2 and m3, is much smaller than what would fit with the larger skull (USNM 181740; Beatty 2009). USNM 181741 appears to have fit an animal the same size as that to which the smaller skull (USNM 181738) belonged, and is probably a juvenile or subadult (like USNM 181738) (Beatty 2009); USNM 181743, a heavily worn isolated lower premolar (Beatty 2009); USNM 214737, a right pelvis, mostly complete except for portions of the sacroiliac joint and ischiopubic ramus (Beatty 2009); USNM 214738, a tibial diaphysis (Beatty 2009); USNM 214740, a partial adult mandible, including the symphysis and a portion of the left tusk (Beatty 2009); USNM 244034, an edentulous left mandible from an individual of the same size as USNM 181741, possibly belonging to the same individual as they were found at the same spot in different years (Beatty 2009); USNM 244036, a left M2 in which the accessory cusp anterior to the protocone is broken off and partially restored with a waxy substance (Beatty 2009); USNM 333334, an isolated partial lower molar comprising two straight cusps, mostly free of matrix, with only a small remnant of intercuspal matrix that appears to include the fossilized remains of plant material (Beatty 2009). USNM 157811, 181738–181741, 181743, 214737, 214738, 214740, 244034, 244036, and 333334 are all from near Seal Rock State Park, Lincoln County, Oregon, USA (Beatty 2009). Newly referred RBCM.EH2005.002.0001, a right M2 or M3 (Fig. 3); CMN 9196, a right m1 (Fig. 4) from near Kirby Creek (Vancouver Island, Canada), late Oligocene Sooke Formation.
Description.—RBCM.EH2005.002.0001: A right upper second or third molar that shows very little sign of wear and is missing its roots. This specimen is rather fragile and the broken edges where the roots were attached appear thin, suggesting that the roots were incompletely formed. The cusp arrangement, including accessory cusps and styles, are identical in size and position to those of the upper left molars of the large skull of Cornwallius sookensis USNM 181740 (Beatty 2009). The cusps and styles of RBCM.EH2005.002.0001 are also identical to UCMP 36078 (VanderHoof 1942), though the small accessory cusp/style between the paracone and metacone appears to be broken off in UCMP 36078, making its size uncertain. Additionally, another tooth, IGM 3688 from the El Cien Formation at Punta San Carlos, Baja California Sur (Applegate 1986) resembles the m2 of USNM 181741 as well. It seems that RBCM.EH2005.002.0001 represents the same taxon as USNM 181740 from Oregon and UCMP 36078 and IGM 3688 from Baja California.
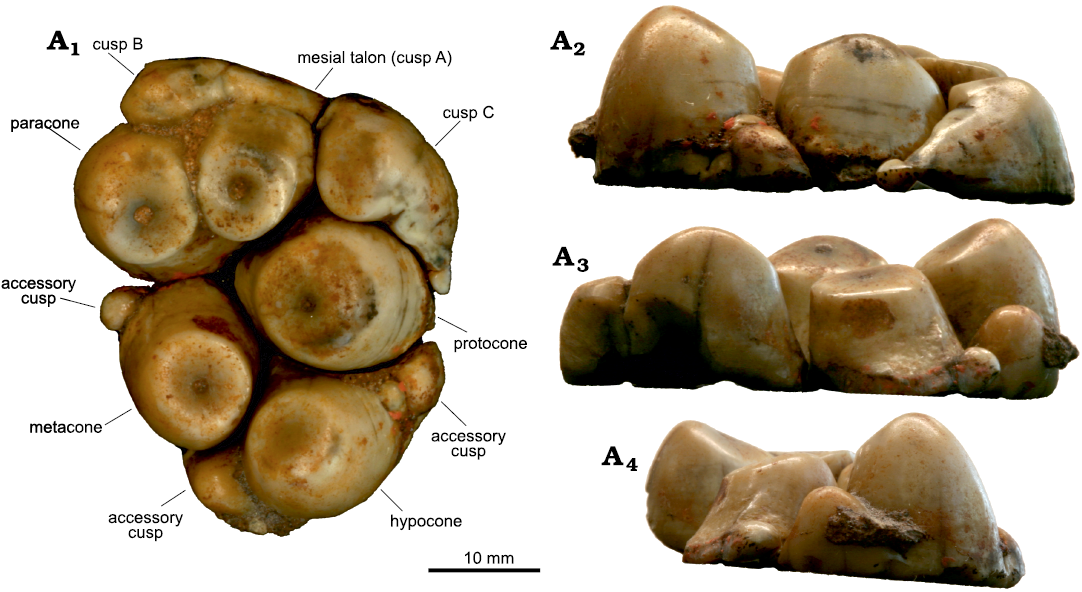
Fig. 3. Desmostylian mammal Cornwallius sookensis (Cornwall, 1922) (RBCM.EH2005.002.0001) from Sooke Formation, late Oligocene, Kirby Creek, Vancouver Island, British Columbia, Canada. Right M2 or M3, in occlusal (A1), lingual (A2), buccal (A3), and mesial (A4) views. Cusps are labeled according to Beatty (2009).
The wear on the mesial talon, paracone, protocone, and metacone all show a small exposure of dentine, whereas the wear on the hypocone has only formed a small facet with defined edges, but no dentine exposure. This is a more advanced stage of wear than the tooth of C. sookensis from Baja California Sur (UCMP 36078) (VanderHoof 1942). This resembles the wear seen on some partially erupted molars in subadult Desmostylus hesperus specimens (USNM 8191, USNM 214743, UOMNHCH F79346).
CMN 9196: This tooth is a right lower second or third molar (Fig. 4), most similar in morphology to the teeth of the mandible referred to C. sookensis, USNM 181741 (Beatty 2009). USNM 181741 is a right mandible containing a very worn m1 and less worn m2. The m1 is worn to the point where the dentine lakes of the paraconids and metaconid are merged, as are the dentine lakes of the hypoconid and entoconid. The only cusp of the m1 remaining was the accessory cusp between the paraconids and hypoconid. This seems to be only because it was close enough to the base of the crown to retain its enamel edges and evade this dentine lake merger. The m2 of the same specimen, though at a similar stage of wear as CMN 9196, notably lacks an accessory cusp between the paraconids and hypoconid like the m1 has. Thus, CMN 9196 is most likely a lower right m1 of an animal just like USNM 181741 (Beatty 2009). As far as it is possible to tell from isolated teeth, this is most likely to be the same species, C. sookensis. Moreover, CMN 9196 is an almost mirror image of the holotype first or second molar (RBCM 486). It is nearly at the same wear stage as RBCM 486 (Beatty 2009: fig. 2) and may represent the same individual animal.
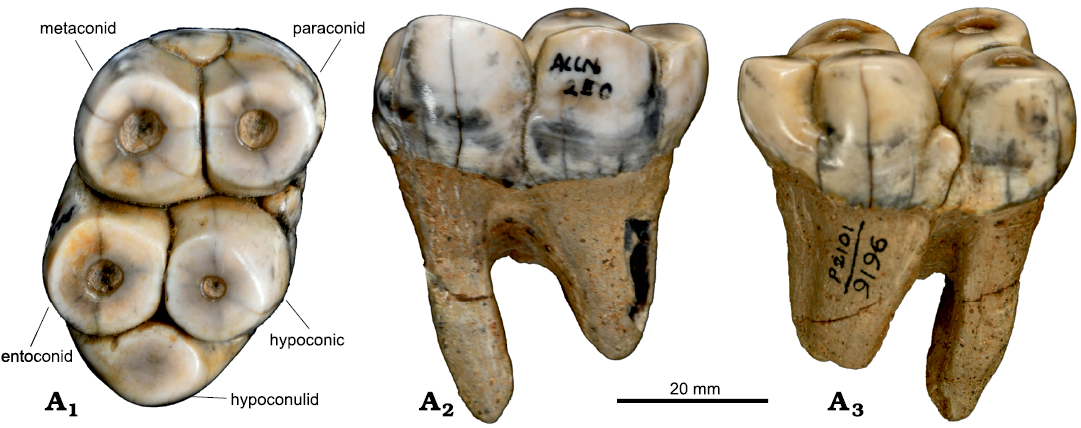
Fig. 4. Desmostylian mammal Cornwallius sookensis (Cornwall, 1922) (CMN 9196) from unknown locality, Sooke Formation, late Oligocene, Vancouver Island, British Columbia, Canada. Right m2 or m3, in occlusal (A1), lingual (A2), and buccal (A3) views. Note that this tooth is almost a perfect mirror image of the holotype of C. sookensis (RBCM 486). Cusps are labeled according to Beatty (2009).
Remarks.—Despite its small size, the holotype (RBCM 486) is virtually identical to the posterior molar of USNM 181741. USNM 181741 appears to have fit an animal the same size as that to which the smaller skull (USNM 181738) belonged, and is probably a juvenile or subadult (like USNM 181738) (Beatty 2009).
Stratigraphic and geographic range.—Zemorrian (upper Oligocene) of Vancouver Island, British Columbia (Canada); Unalaska Island, Aleutian Islands, Alaska (USA); Yaquina Bay and Beaver Creek, Lincoln County, Oregon (USA); and Bahia San Carlos, Baja California Sur (Mexico).
Discussion
Typically, additional specimens provide answers to previous questions. In the present case, it appears that these new specimens have only brought about more uncertainty, which is the prompt for this paper. These problems are twofold: (i) the mysterious additional unreported paratype of Cornwallius sookensis; (ii) recognition of taxa initially described based on isolated teeth, like C. sookensis, or lower teeth lacking association with cranial material, like Behemotops proteus.
CMN 9196 and the holotype of Cornwallius sookensis.—Curiously, CMN 9196 introduces some historical complexity. The original material of Cornwallius sookensis consisted of two teeth (Cornwall 1922; Hay 1923). According to VanderHoof (1937: 204) and Hay (1924: 5), the holotype tooth, a left m1 or m2 (BCPM 486/USNM 15165 cast) was collected by M. Egerton in 1916 and then passed along to Ira Cornwall. Another tooth, a left m3 (BCPM 491/USNM 15166 cast), was collected in 1921 from the same locality by Reverend Robert Connell and also donated to Ira Cornwall. An additional tooth was collected by B.L. Clark in 1922 from very near the type locality (UCMP 32682, a fragment of a lower molar including two cusps; VanderHoof 1937). These specimens went missing from the collection in 1928, but then reappeared in the collections in 1932 in a cigarette tin (McAnally 1993). No other teeth had been reported in scientific publications or mentioned in the notes associated with specimens at the RBCM or USNM, yet another tooth resembling the holotype left lower first or second molar (RBCM 486) is in the collection of the Canadian Museum of Nature in Ottawa, Ontario, Canada. This specimen (CMN 9196, described above) is cataloged as a type specimen, is noted to have been collected by Robert Connell and identified by Ira Cornwall (like the type at the RBCM), yet was never reported by Cornwall (1922) or Hay (1923). As stated above, CMN 9196 is a good match for the holotype of C. sookensis, and may very well be a previously unreported paratype, possibly belonging to the same individual.
Taxon identity and the problem of organ taxa.—The specimens of Cornwallius sookensis described above are identified as such because they have the same morphology as the original material of C. sookensis (Cornwall 1922; Hay 1923) and specimens later referred to C. sookensis (Beatty 2009), and they do not resemble the dentition of the other desmostylian known from this locality, Behemotops proteus (Ray et al. 1996; Beatty and Cockburn 2015) and Behemotops cf. B. proteus (the partial tooth reported here, RBCM.EH2007.008.0001). This appears sufficient, though it also highlights a deficiency in past identifications of specimens of desmostylians from this locality and others. For example, Beatty (2006a) identified specimens of femora from Muir Creek as belonging to C. sookensis. But this identification was largely based on their being a small desmostylian from this locality. Since that time another small desmostylian, B. proteus, has been reported from this site (Beatty and Cockburn 2015), and other sites with multiple desmostylian taxa have been reported, such as Seal Rock State Park in Oregon (Ray et al. 1996; Beatty 2009; Beatty and Cockburn 2015). It would be unwise to consider these isolated postcrania as belonging to C. sookensis with certainty, now that more than one small desmostylian are known to have coexisted here.
Notably, CMN 9196 resembles the holotype of C. sookensis, and could very well be a tooth collected at the same time that got disassociated at the time of collection (100+ years ago, these specimens were not collected with plaster jacketing techniques). CMN 9196 and the mandible USNM 181741 have lower molars matching the holotype, so the most conservative assessment would be to interpret them both as C. sookensis.
However, RBCM.EH2005.002.0001 (from Vancouver Island; reported here), USNM 181740 (from Oregon; Beatty 2009), and UCMP 36078 (from Baja California; VanderHoof 1942) are all referred to the same taxon because they are appropriately-sized desmostylians with similarly brachydont, desmostylodont teeth. The initial assumption about the identity of RBCM.EH2007.008.0001, before close inspection, was that it was C. sookensis because it came from the type locality of C. sookensis. Co-occurrence of different desmostylians in the same localities has been known for some time (Domning et al. 1986), and it seems wise to be cautious in making such assumptions about unassociated fossils. Perhaps RBCM.EH2005. 002.0001, USNM 181740, and UCMP 36078 represent some other, similarly brachydont desmostylian. It is also possible that, like other large mammals with much better samples of specimens known, such as brontotheres (Mihlbachler 2008), lower molars may turn out to be much less diagnostic than was previously believed, rendering C. sookensis a nomen dubium.
Thus, it would be reckless to ascribe new taxonomic names to the specimens in doubt. It is also important to remember that with more complete specimens, we may find that the diversity of desmostylians is greater than previously recognized, so that their taxonomy will need revision. Many such specimens are found meters apart, and it risks misrepresenting the diversity of this group by assuming low diversity at each locality. Desmostylians are “as rare as hens’ teeth”, particularly those from the Oligocene (VanderHoof 1942; Domning et al. 1986; Inuzuka 2000; Barnes and Goedert 2001; Beatty 2006a, b, 2009; Beatty and Cockburn 2015), making it important that their identification is conducted with a conservative and careful approach. It would be wise to continually reassess the basis of taxonomic identity of specimens being studied, and to explore all opportunities to evaluate sources of variation within each taxon and the group. Recent efforts of this sort on postcrania (Matsui 2017; Matsui et al. 2018) have been valuable and should be encouraged. It is essential that collectors and museums keep adequate records of field associations and preservational states of specimens, so that the identities of these rare taxa can be properly deciphered.
Acknowledgements.—Joan Kerik, Victoria Arbour, and Derek Larson (all RBCM), Danielle Fraser (CMN), and Nicholas Pyenson and David Bohaska (both USNM) kindly provided access and support in studying the collections in their care. Thomas Cockburn (RBCM) provided much-needed assistance with the study of the RBCM collection. Two referees provided useful comments that improved the manuscript greatly.
References
Applegate, S.P. 1986. The El Cien Formation, strata of Oligocene and Early Miocene age in Baja California Sur. Universidad Nacional Autonoma de Mexico, Instituto de Geologia, Revista 6 (2): 145–162.
Barnes, L.G. and J.L. Goedert. 2001. Stratigraphy and Paleoecology of Oligocene Desmostylian Occurrences in Western Washington State, USA. Bulletin of Ashoro Museum of Paleontology 2: 7–22.
Beatty, B.L. 2006a. Rediscovered specimens of Cornwallius (Mammalia, Desmostylia) from Vancouver Island, British Columbia, Canada. PalArch’s Journal of Vertebrate Palaentology 1: 1–6.
Beatty, B.L. 2006b. Specimens of Cornwallius sookensis (Desmostylia, Mammalia) from Unalaska Island, Alaska. Journal of Vertebrate Paleontology 26: 785–787. Crossref
Beatty, B.L. 2009. New material of Cornwallius sookensis (Mammalia: Desmostylia) from the Yaquina Formation of Oregon. Journal of Vertebrate Paleontology 29: 894–909. Crossref
Beatty, B.L. and Cockburn, T.C. 2015. New insights on the most primitive desmostylian from a partial skeleton of Behemotops (Desmostylia, Mammalia) from Vancouver Island, British Columbia. Journal of Vertebrate Paleontology 35 (5): e979939. Crossref
Clark, B.L. and Arnold, R. 1923. Fauna of the Sooke Formation, Vancouver Island. University of California Publications in Geological Sciences 14: 123–234.
Clark, J.M. 1991. A new early Miocene species of Paleoparadoxia (Mammalia: Desmostylia) from California. Journal of Vertebrate Paleontology 11: 490–508. Crossref
Cornwall, I.E. 1922. Some notes on the Sooke Formation, Vancouver Island, B.C. The Canadian Field-Naturalist 36: 121–123.
Domning, D.P., Ray, C.E., and McKenna, M.C. 1986. Two new Oligocene desmostylians and a discussion of tethytherian systematics. Smithsonian Contributions to Paleobiology 59: 1–56. Crossref
Hay, O.P. 1923. Characteristics of sundry fossil vertebrates. Pan-American Geologist 39: 101–121.
Hay, O.P. 1924. Notes on the osteology and dentition of the genera Desmostylus and Cornwallius. Proceedings of the United States National Museum 65: 1–10. Crossref
Inuzuka, N. 1987. Primitive desmostylians, Behemotops and the evolutionary pattern of the order Desmostylia [in Japanese]. In: Professor Masaru Matsui Memorial Volume, 13–25. Hokkaido University, Sapporo.
Inuzuka, N. 2000. Primitive Late Oligocene desmostylians from Japan and phylogeny of the Desmostylia. Bulletin of the Ashoro Museum of Paleontology 1: 91–124.
Kermode, F. 1917. Palaeontology. Report of the Provincial Museum of Natural History (Victoria, British Columbia) 1917: 42–43.
Linnaeus, C. 1758. Systema naturæ per regna tria naturæ, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Tomus I. Editio decima, reformata. 824 pp. Laurentius Salvius, Stockholm. Crossref
Matsui, K. 2017. How can we reliably identify a taxon based on humeral morphology? Comparative morphology of desmostylian humeri. PeerJ 5: e4011. Crossref
Matsui, K., Kimura, Y., Nagata, M., Inose, H., Ikeda, K., Beatty, B.L., Obayashi, H., Hirata, T., Otoh, S., Shinmura, T., Agematsu, S., and Sashida, K. 2018. A long-forgotten “dinosaur” bone from a museum cabinet, uncovered to be a Japan’s iconic extinct mammal, Paleoparadoxia (Desmostylia, Mammalia). Royal Society Open Science 5 (7): 172441. Crossref
McAnally, L.M. 1993. Fallacy, felony and fossils from the Sooke Formation. Vancouver Island Paleontological Society Newsletter 3: 8–9.
Mihlbachler, M.C. 2008. Species taxonomy, phylogeny, and biogeography of the Brontotheriidae (Mammalia: Perissodactyla). Bulletin of the American Museum of Natural History 311: 1–475. Crossref
Nesbitt, E.A., Martin, R.A., Carroll, N.P., and Grieff, J. 2010. Reassessment of the Zemorrian foraminiferal stage and Juanian molluscan stage north of the Olympic Mountains, Washington State and Vancouver Island. Newsletters in Stratigraphy 43: 275. Crossref
Osborn, H.F. 1905. Ten years’ progress in the mammalian palaeontology of North America. American Geologist 36: 199–229.
Prothero, D.R., Draus, E., Cockburn, T.C., and Nesbitt, E.A. 2008. Paleomagnetism and counterclockwise tectonic rotation of the Upper Oligocene Sooke Formation, southern Vancouver Island, British Columbia. Canadian Journal of Earth Sciences 45: 499–507. Crossref
Ray, C.E., Domning, D.P., and McKenna, M.C. 1994. A new specimen of Behemotops proteus (Order Desmostylia) from the Marine Oligocene of Washington. In: A. Berta and T.A. Demeré (eds.), Contributions in Marine Mammal Paleontology Honoring Frank C. Whitmore, Jr. Proceedings of the San Diego Society of Natural History 29: 205–222.
Reinhart, R. 1953. Diagnosis of the new mammalian Order, Desmostylia. Journal of Geology 61: 187. Crossref
Reinhart, R.H. 1982. The extinct mammalian order Desmostylia. National Geographic Society Research Reports 14: 549–555.
VanderHoof, V.L. 1937. A study of the Miocene sirenian Desmostylus. University of California Publications, Bulletin of the Department of Geological Sciences 24:169–262.
VanderHoof, V.L. 1942. An occurrence of the Tertiary marine mammal Cornwallius in Lower California. American Journal of Science 240: 298–301.
Brian Lee Beatty [bbeatty@nyit.edu; ORCID: https://orcid.org/0000-0002-5464-0041 ], NYIT College of Osteopathic Medicine, Northern Blvd, Old Westbury, New York 11568, USA; National Museum of Natural History, Smithsonian Institution, Washington, DC 20560, USA.
Received 6 March 2022, accepted 6 January 2023, available online 10 May 2023.
Copyright © 2023 B.L. Beatty. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Acta Palaeontol. Pol. 68 (2): 373–378, 2023
https://doi.org/10.4202/app.00990.2022