
Bivalve-barnacle pseudoplanktonic colonisation of wood from the Toarcian, Lower Jurassic, Strawberry Bank Lagerstätte, Somerset, UK
CRISPIN T.S. LITTLE, ANDY GALE, MATT WILLIAMS, ØYVIND HAMMER, and VINCENT FERNANDEZ
Little, C.T.S., Gale, A., Williams, M., Hammer, Ø., and Fernandez, V. 2023. Bivalve-barnacle pseudoplanktonic colonisation of wood from the Toarcian, Lower Jurassic, Strawberry Bank Lagerstätte, Somerset, UK. Acta Palaeontologica Polonica 68 (1): 133–142.
Pseudoplankton are organisms that are adapted for a mode of life attached to floating objects. In modern oceans common examples are lepadid barnacles, which attach themselves to man-made and natural objects, especially wood logs. In the fossil record, pseudoplankton examples are commonly found in black shales, such as the lower Toarcian Posidonia Shale Formation of Germany. Here there are occasional large logs of fossil wood covered in specimens of the inoceramid bivalve Pseudomytiloides dubius, with or without specimens of the pentacrinitid crinoid Seirocrinus subangularis. Some Posidonia Shale logs show evidence of for intra-species succession with younger S. subangularis specimens attached onto the stems of older ones. Another example of pseudoplankton from the Posidonia Shale is the occurrence of the numerous disarticulated specimens of the phosphatic-shelled eolepadid barnacle Toarcolepas mutans associated with a piece of fossil wood, which constitutes the oldest example of pseudoplanktonic barnacles in the fossil record. Here we report a limestone concretion from the lower Toarcian Strawberry Bank Lagerstätte (Ilminster, Somerset, UK) that preserves a piece of fossil wood with a pseudoplanktonic colony comprising at least a hundred specimens of T. mutans that attached onto a layer of P. dubius bivalves, that had already attached onto the wood. This is one of very few examples of temporal succession for pseudoplankton in the Toarcian and is also unusual in being preserved in a mixed carbonate-siliciclastic facies, rather than a black shale. The occurrence of T. mutans in the Strawberry Bank Lagerstätte concretion represents the second record of the species and also the equal oldest example of pseudoplanktonic barnacles in the fossil record.
Key words: Bivalvia, Cirripedia, Crinoidea, fossil wood, pseudoplankton, Strawberry Bank Lagerstätte, Toarcian, Early Jurassic, UK.
Crispin T.S. Little [earctsl@leeds.ac.uk; ORCID: https://orcid.org/0000-0002-1917-4460 ], School of Earth and Environment, University of Leeds, Woodhouse Lane, Leeds LS2 9JT, UK; Life Sciences Department, Natural History Museum, London, Cromwell Road, London SW7 5BD, UK.
Andy Gale [andy.gale@port.ac.uk; ORCID: https://orcid.org/0000-0002-2075-3689 ], School of Earth and Environmental Sciences, University of Portsmouth, Burnaby Building, Burnaby Road, Portsmouth PO1 3QL, UK; Earth Sciences Department, Natural History Museum, London, Cromwell Road, London SW7 5BD, UK.
Matt Williams [matt.williams@brlsi.org; ORCID: https://orcid.org/0000-0001-7409-2655 ], Bath Royal Literary and Scientific Institution, 16-18 Queen Square, Bath BA1 2HN, UK.
Øyvind Hammer [oyvind.hammer@nhm.uio.no; ORCID: https://orcid.org/0000-0001-9951-0630 ], Natural History Museum, University of Oslo, Postboks 1172, Blindern, 0318 Oslo, Norway.
Vincent Fernandez [vinfernand@gmail.com; ORCID: https://orcid.org/0000-0002-8315-1458 ], Imaging and Analysis Centre, Natural History Museum, London, Cromwell Road, London SW7 5BD, UK; European Synchrotron Radiation Facility, 71 Avenue des Martyrs, 38000 Grenoble, France.
Received 23 August 2022, accepted 23 November 2022, available online 24 January 2023.
Copyright © 2023 C.T.S. Little et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Pseudoplankton are organisms that have morphological adaptations for a mode of life attached to floating objects (Wignall and Simms 1990), and can be obligate (only occurring as pseudoplankton), facultative (also being able to live as benthic organisms) or accidental. In modern oceans some the most visually striking pseudoplanktonic organisms are lepadid barnacles, which attach themselves to man-made and natural objects (principally wood logs and flotsam), sometimes in very large numbers (1000s of individuals). The barnacles hang down from the object they are attached to using a fleshy stalk called the peduncle and filter particles from the seawater to feed (Southward 2008). Wignall and Simms (1990) classified this attachment strategy as pendent. If an object is large and remains floating for a period of time (e.g., years), then it can support a diversity of pseudoplanktonic organisms, with a number of different attachment strategies, and temporal succession may occur, where larvae of one pseudoplanktonic species settle onto and then overgrow existing pseudoplankton (Tsikhon-Lukanina et al. 2001). Eventually all floating objects either wash up onto a shoreline, break up into small pieces unable to support pseudoplankton, or sink onto the seafloor. In most of these cases any attached pseudoplankton colonies perish, but teredinid bivalves boring into floating wood can survive on the sea floor (Huggett and Gale 1995).
In the fossil record the pseudoplanktonic mode of life is quite difficult to recognize and, as Wignall and Simms (1990) pointed out, quite a number of claimed pseudoplankton occurrences are actually examples of benthic organisms. Bone-fide examples are commonly associated with black shales (organic-rich fine-grained sediments), which is because this facies was the result of deposition in sedimentary basins with low oxygen levels at the seafloor (Wignall 1994). This prevented the floating objects and their pseudoplankton load from being scavenged and/or mechanically disturbed by borrowing organisms after they had sunk onto the seafloor. In the Mesozoic, famous black shale pseudoplankton examples come from the lower Toarcian (Lower Jurassic) Posidonia Shale Formation of Germany. In the Posidonia Shale there are occasional large logs of fossil wood covered in specimens of the inoceramid bivalve Pseudomytiloides dubius, with or without specimens of the pentacrinitid crinoid Seirocrinus subangularis (e.g., Hauff and Hauff 1981). Both these species had pendant modes of attachment: P. dubius via byssal threads, S. subangularis with an attachment disc (Simms 1986). Pseudomytiloides dubius is usually interpreted as a facultative pseudoplanktonic species, because, as Kauffman (1981) pointed out, “in many horizons it is much too abundant for all specimens to have been derived from the relatively rare examples of driftwood” (see also Etter 1996; Harries and Little 1999; Caswell et al. 2009). Seirocrinus subangularis, in contrast, is now usually interpreted as an obligate pseudoplanktonic species, as it is almost always found in association with fossil wood (e.g., Seilacher et al. 1968; Hauff and Hauff 1981; Simms 1986; Matzke and Maisch 2019; Hunter et al. 2020). The lack of post-seafloor emplacement disturbance of Posidonia Shale logs has been used to elucidate fine details of settlement patterns by S. subangularis onto floating wood (e.g., Matzke and Maisch 2019; Hunter et al. 2020) and how they finally sank onto the seafloor (Seilacher et al. 1968). Matzke and Maisch (2019) found that in the Posidonia Shale of the Holcim Quarry, Dotternhausen, Germany, 20–25% of the preserved logs were encrusted by P. dubius only and approximately 25% of the logs were encrusted by P. dubius and S. subangularis. They also discovered evidence for intra-species succession in some of these examples, with younger S. subangularis specimens attached onto the stems of older ones, which they suggested was a consequence of all the space on the log surface being occupied during earlier settlement events. On one log Matzke and Maisch (2019) identified at least two generations of crinoids, and thus a minimum of two settlement events, and possibly even four. For Posidonia Shale logs encrusted with both P. dubius and S. subangularis, no studies have explicitly made clear the temporal settlement relationships between the two taxa, which either means this is difficult to discern, or that there was co-settlement of these taxa on logs.
Another example of pseudoplankton from the Posidonia Shale comes from Zell unter Aichelberg, southern Germany, with the occurrence of the numerous disarticulated specimens of the phosphatic-shelled eolepadid barnacle Toarcolepas mutans associated with a piece of fossil wood (Gale and Schweigert 2015). Indeed, these authors interpreted these to be the oldest example of pseudoplanktonic barnacles in the fossil record. Here we report on a limestone concretion (BRLSI.M3941) from the Strawberry Bank Lagerstätte, Somerset, UK, that contains a piece of fossil wood encrusted with numerous specimens of P. dubius and T. mutans. This constitutes an example of bivalve-barnacle pseudoplanktonic colonization of wood from the same time period as the Posidonia Shale, but in a mixed carbonate-siliciclastic sedimentary facies. The fossil association in BRLSI.M3941 also provides rare unambiguous evidence for inter-specific temporal successional growth in pseudoplanktonic colonies in the fossil record, with pseudoplanktonic barnacles settling on pseudoplanktonic bivalves.
Institutional abbreviations.—BRLSI, Bath Royal Literary and Scientific Institution, UK; TTNCM, South West Heritage Trust, Taunton, UK.
Geological setting
The Strawberry Bank Lagerstätte is a Toarcian-aged fossiliferous horizon located on the north side of the town of Ilminster, Somerset, UK (Williams et al. 2015; Fig. 1A). Charles Moore, a local geologist, worked the site from 1842 until 1860 and the majority of the fossils he excavated are held by the Bath Royal Literary and Scientific Institution (BRLSI), with a smaller sub collection in the South West Heritage Centre, Taunton, UK. The fauna of the Strawberry Bank Lagerstätte consists mostly of marine nektonic animals, many of which are exceptionally preserved, including articulated, three-dimensional specimens containing phosphatized soft tissue. These include ichthyosaurs (Caine and Benton 2011; Marek et al. 2015), crocodyliomorphs (Peirce and Benton 2006; Ballell et al. 2019), fishes (Cawley et al. 2019) and coleoids (Sinha et al. 2021). Also present in the Strawberry Bank fauna are insects (Williams et al. 2015), crustaceans (Audo et al. 2017) and molluscs, including ammonites, belemnites, bivalves, and gastropods. In his list of taxa from the Strawberry Bank locality Moore (1886: 191) noted “To the above list of crustacea are only to be added a colony of Cirrhipedia [sic] of many hundreds in number, attached to a piece of wood throughout its entire length, or rather to another colony of Mytilidae, which had previously taken possession of its surface.” This description correlates with specimen BRLSI.M3941, the subject of this study.
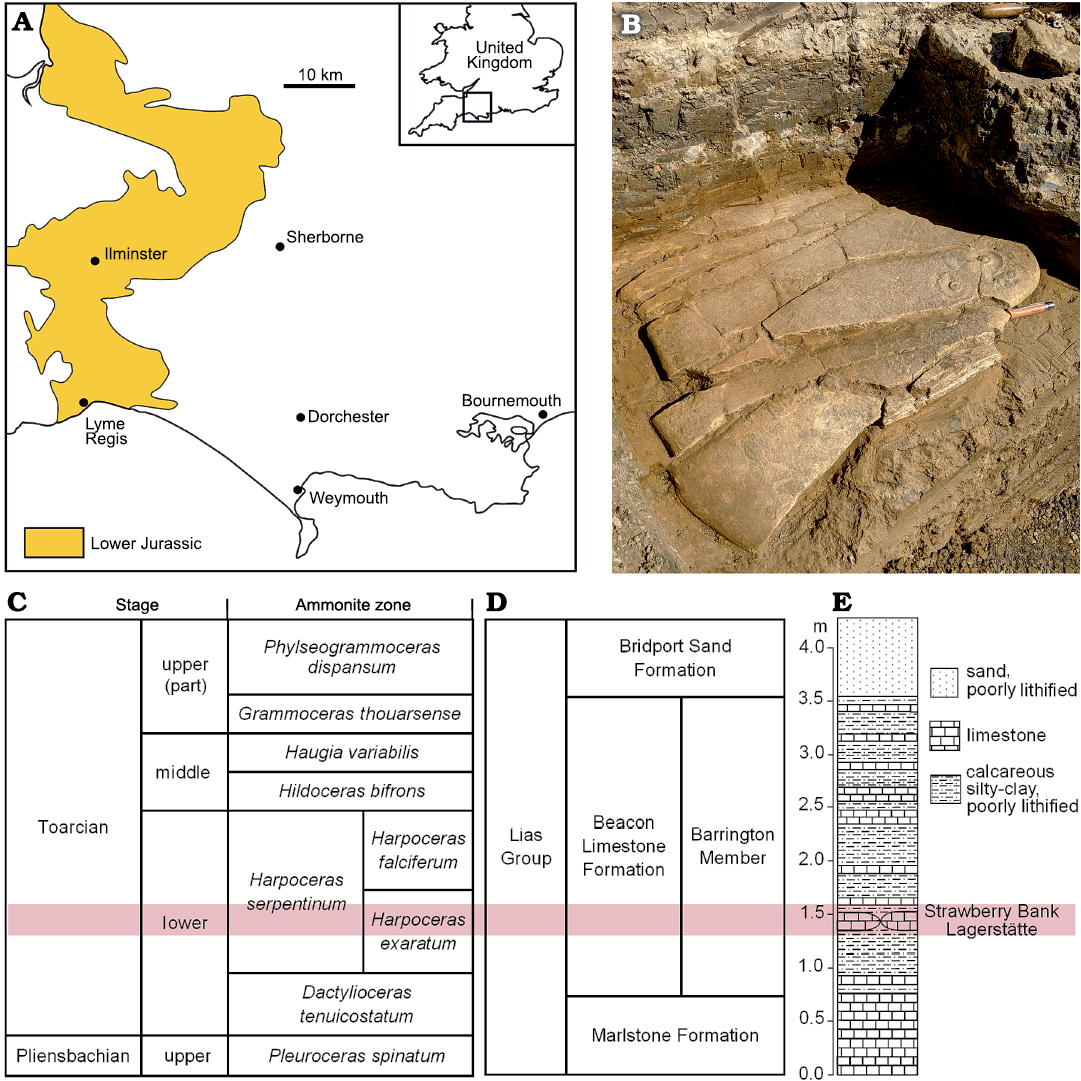
Fig. 1. Location and stratigraphy of the Strawberry Bank Lagerstätte and associated beds. A. Location of Ilminster, Somerset, England, UK and Lower Jurassic outcrop pattern in the area. B. Limestone concretion of the Strawberry Bank Lagerstätte exposed in situ north of Ilminster during excavations of the site in 2019. Folding knife is 9 cm long. C. Stage information, and ammonite zones and subzones. D. Local lithostratigraphic scheme for the same timescale. E. Simplified sedimentary log at Strawberry Bank with the Strawberry Bank Lagerstätte shaded in pink (after Williams et al. 2015 and Boomer et al. 2021).
The Strawberry Bank Lagerstätte comes from a single layer of pavement-like concretions, of variable thickness (Fig. 1B), which Moore (1866) called the fish bed of his Saurian and Fish Zone. The Strawberry Bank Lagerstätte has been correlated to the lower Toarcian Harpoceras serpentinum Zone, Elegantuliceras elegantulum Subzone in nearby sections (see Williams et al. 2015 for details) and belongs to the Barrington Member of the Beacon Limestone Formation (Fig. 1C–E). The Strawberry Bank Lagerstätte concretions represent a very localized lithology, as they are absent from the Toarcian sequence at the Seavington St Michael, only 3 km to the east (Boomer et al. 2021). The Beacon Limestone Formation in the central Somerset area comprises a mixture of carbonate, siltstone and clay beds, a facies that differs considerably from the time equivalent black shale of the Mulgrave Shale Member of the Cleveland Basin in Yorkshire, UK, which is also much more expanded (Powell 1984). Boomer et al. (2021) interpret the Beacon Limestone Formation at Seavington St Michael to have been deposited in a relatively deep-water (mid- to outer shelf) environment, yet note that the palynological evidence points to a more restricted, or proximal, setting. The presence of abundant insects in the Strawberry Bank Lagerstätte was used by Williams et al. (2015) to suggest that the Strawberry Bank Lower Toarcian sequence was deposited in a nearshore setting.
Methods
Measurements and digital photography of BRLSI.M3941 and associated macrofossils was done at the University of Leeds and the BRLSI. The bivalves were measured along the maximum visible shell length. Statistical analysis of bivalve shell length was made using PAST ver. 4.11 software (Hammer et al. 2001) and Microsoft Excel. X-ray computed tomography (µCT) of BRLSI.M3941 was performed with a Nikon XTH 225 ST system (Nikon Metrology, Leuven, Belgium) at the Imaging and Analysis Centre, Natural History Museum, London. The instrument setup comprised: a 225 kV rotating reflection tungsten target; tube voltage of 215 kV and current of 930 µA; filtration with 1 mm of tin; 4100 projections over a 360° rotation; 8 frames averaging; 250 ms exposure per frame; detector gain set to 24 dB; source-object distance of 519.7 mm and object-detector distance of 508.4 mm, resulting in a magnification of 1.978, generating data with an isotropic voxel size of 75.83 µm; the minimise ring artefact option was enabled. As the sample was too large to fit in the field of view for this magnification setup, two separate acquisitions were performed, moving the sample vertically with the motorised sample manipulator between the scans. The tomographic reconstruction was performed using CT pro 3D 6.2 (Nikon metrology, Leuven, Belgium). It included a beam hardening correction using the automatic option. The polynomial coefficients for beam hardening correction were calculated for one dataset only and applied to the second one, ensuring identical modification of the datasets. Each dataset was first reconstructed as 32-bit float. The two datasets were then merged following the protocol described in Butler et al. (2022) and resulting in a single 16-bit unsigned dataset. Volume visualization of the scan data was done with Avizo, ver. 2020.2 (Thermo Fisher Scientific) with the Volume Rendering module in Standard mode.
Results
Specimen BRLSI.M3941 comprises a roughly rectangular piece of limestone concretion, a maximum of 325 mm long, 132 mm wide and 45 mm deep (Fig. 2). All edges of the concretion are fractured, which indicates it was originally part of a larger structure (cf. Fig. 1). One surface of the concretion has a piece fossil wood running across its length, from midway to the shorter edge (which for orientation purposes we call the concretion short edge), as well as many specimens of Pseudomytiloides dubius and Toarcolepas mutans. The opposite surface of the concretion is devoid of macrofossils (SOM: fig. 1A, Supplementary Online Material available at http://app.pan.pl/SOM/app68-Little_etal_SOM.pdf). There is no information associated with the concretion to indicate which of these surfaces was stratigraphically uppermost, but we suspect, following size analysis of the bivalves (see below), that the surface with the visible macrofossils was uppermost, so we therefore call this the upper surface, and the opposite surface the lower surface. Both the lower surface and the long edge of the concretion have been trimmed (SOM: fig. 1A). The limestone forming the concretion is fine grained overall, but there is a zone, around 9 mm thick, running centrally lengthways through it, where there are some larger, darker grains (SOM: fig. 1C). It is important to note that this zone does not correspond to the upper surface of the concretion, with the wood, bivalves and barnacles.

Fig. 2. Fossilized driftwood association, BRLSI.M3941, the Toarcian of Strawberry Bank, Somerset, upper surface of limestone concretion showing wood mould and associated specimens of Pseudomytiloides dubius (Sowerby, 1828) and Toarcolepas mutans Gale and Schweigert, 2015. See SOM: fig. 1 for orientation imagery.
The fossil wood on the upper surface of the concretion exists almost exclusively as an external mould. The greater amount of this runs in a band around 40 mm thick across the centre of the concretion (Fig. 2: Band A), and also in a band around 10 mm thick along the bottom edge (Fig. 2: Band B). Between this is a band around 30 mm thick with no wood. We surmise the two bands of wood represent the same piece, and that the wood-free band represents a concavity in the original piece of wood. Running along the top side of the wood is a zone 20–30 mm thick comprising articulated Pseudomytiloides dubius specimens, with a variety of orientations (Fig. 2, SOM: fig. 2). Some, especially those against the wood, have long edges of their shells nearly parallel to the wood, but the majority have shells orientated at high angles to the edge of the wood, some at 90° (Fig. 3). There are between two and five layers of bivalves within this zone (Fig. 3A). Articulated bivalves are present in the wood-free band, and these have variable orientations (Fig. 3B). A few articulated bivalves are also observable projecting through the wood bands, and there are many bivalve-shaped convexities, that indicate bivalves are also present stratigraphically beneath the wood (Fig. 2). This is proved by the 3D rendering of the CT dataset, which shows numerous articulated bivalves within the concretion, with a variety of orientations with respect to the wood mouldic surface, including many at high angles (Fig. 4, SOM: fig. 2). The orientation of the bivalve specimens in the zone along the top edge of the wood is also variable with respect to the plane of the upper surface of the concretion. Some specimens show one valve, others display the commissure of the articulated specimen (Fig. 3C and D). This can also be seen for the bivalves underneath the wood in Fig. 4.
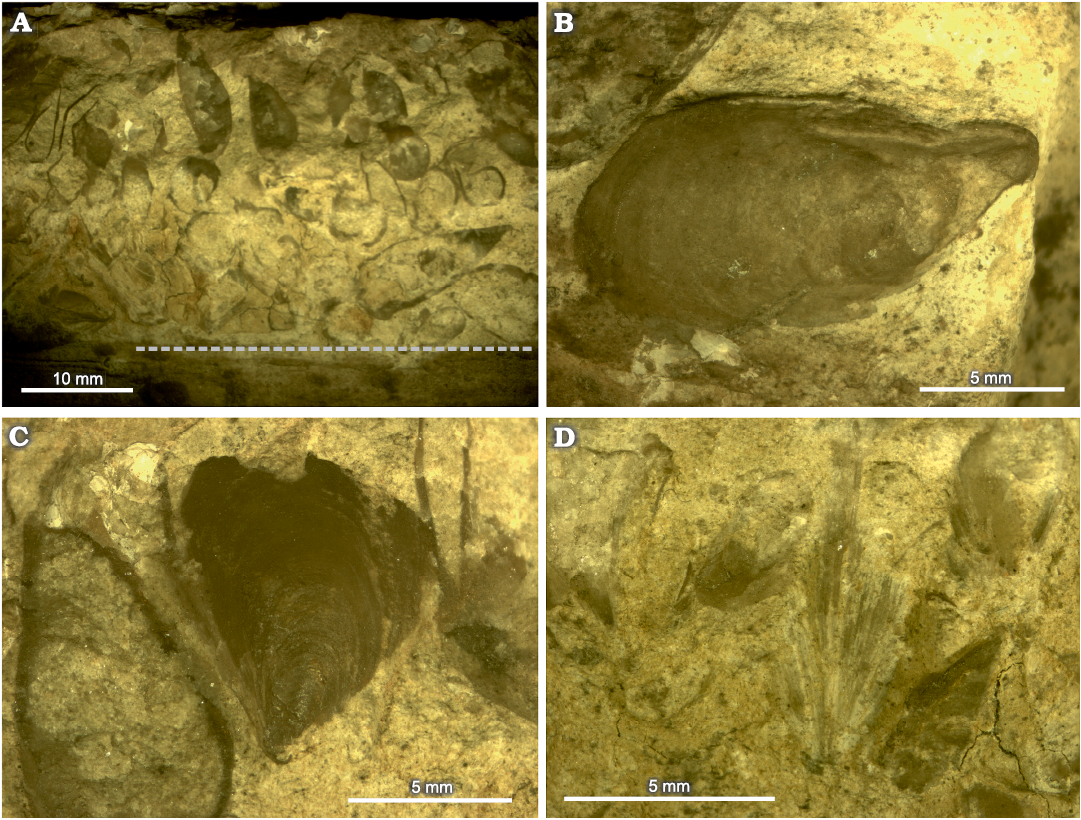
Fig. 3. Detail of Pseudomytiloides dubius (Sowerby, 1828) specimens from upper surface of BRLSI.M3941, Toarcian of Strawberry Bank, Somerset, showing stacking of individuals, articulation and orientation in relation to the wood. Dashed line in A marks the top edge of the wood mould. Locations of specimens on concretion correspond to numbered specimens on SOM: fig. 2 as follows: A, specimens 40 to 44; B, specimen 1; C, specimen 20; D, specimen 38.
On the upper surface of the concretion there are 95 bivalves in the zone along the top surface of the wood (i.e., the surface closest to the concretion long edge, Fig. 2), 35 in the wood bands, and five articulated specimens observable on two of the concretion edges, underneath the wood (SOM: fig. 1B and C). A further 94 bivalves can be counted confidently in the CT data (Fig. 4), giving a total of 229 bivalve specimens in the concretion. This should be considered a maximum value, as a number of the same bivalve specimens are seen on both the concretion surface and the CT data, although we have tried to avoid double counting where this is obvious (SOM: fig. 3). The bivalves in the zone along the top edge of the wood are significantly smaller than those in wood bands, mean shell length 10.4 mm along top of wood (n = 27) and 15.5 mm in wood bands (n = 13), Welch’s t = 3.2; p = 0.006 (SOM: fig. 4, table 1).
Above the bivalve zone along the top of the wood is a zone around 10–15 mm thick formed of at least hundred articulated and partially articulated specimens of the barnacle Toarcolepas mutans (Fig. 2). The total number of specimens is difficult to count, given the preservation. Although there is generally a clear demarcation of the bivalve and the barnacle zones, one bivalve is present in the barnacle zone, and a few barnacle valves occur within the bivalve zone (Fig. 2). In addition to the barnacles on top of the wood, a few isolated valves are present in the wood bands (Fig. 2). No barnacle specimens are apparent in the CT data (Fig. 4). In the zone along the top of the wood some barnacle specimens are “kippered”, with paired terga that are side-by-side, representing parts of individuals which have split along the sagittal plane, but with halves still attached (Fig. 5).
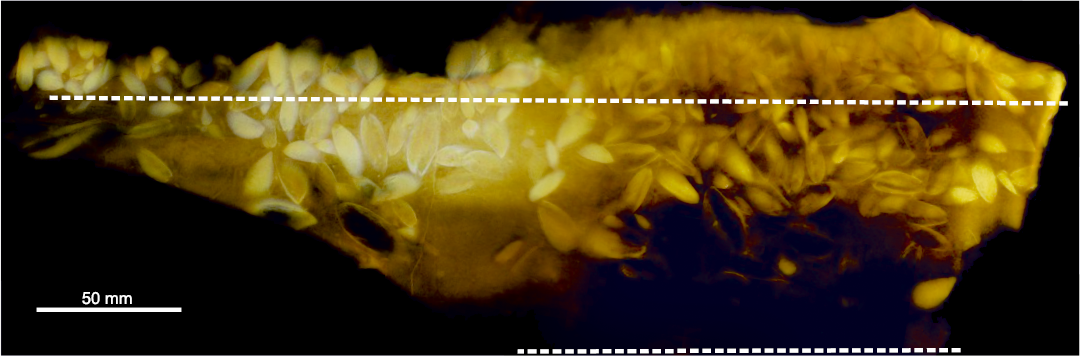
Fig. 4. Three dimensional rendering of the X-ray computed tomographic data of fossilized driftwood association, BRLSI.M3941 from the Toarcian of Strawberry Bank, Somerset. Dashed lines delineate the top and bottom edge of the wood mould (see SOM: fig. 1 for orientation imagery).
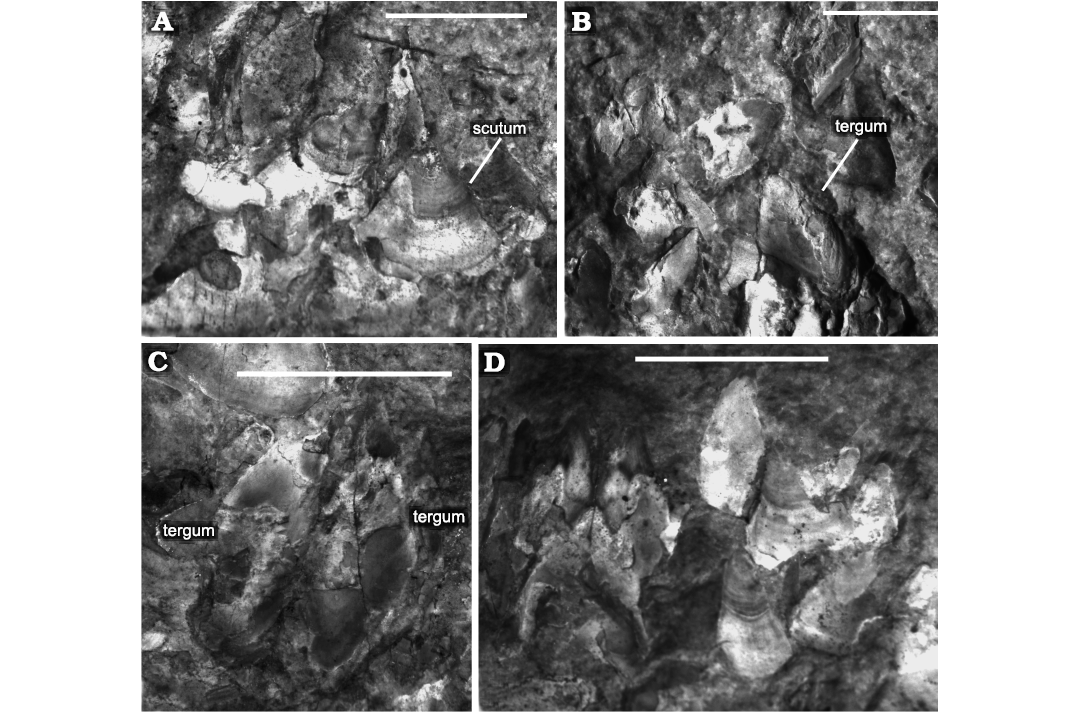
Fig. 5. Detail of the barnacle Toarcolepas mutans Gale and Schweigert, 2015, from upper surface of BRLSI.M3941, the Toarcian of Strawberry Bank, Somerset. In C, D, individuals are “kippered”—split along the sagittal plane. Locations of specimens on concretion shown in dashed boxes in SOM: fig. 2. Scale bars 5 mm.
Discussion
BRLSI.M3941 as a pseudoplanktonic association.—The majority of the bivalves in relation to the edge and underside of the wood piece have a pendant orientation, showing that they were hanging from the wood during life, presumably each attached by a byssus. In turn this indicates the wood was floating when the bivalves settled onto it and thus the bivalves were pseudoplanktonic. The multiple layers of bivalves on the edge of the wood (Fig. 3A) might imply multiple generations of bivalves, or, more likely, crowding of specimens from a single generation, as seen by the specimen packing in the CT data (Fig. 4). The small variation in measured shell length for the specimens on the wood edge (standard deviation 2.8 mm; SOM: table 1) may support this inference, as multiple generations would be expected to result in a greater specimen shell length variation. The larger average size of the bivalves in the wood bands on the concretion surface may be interpreted in two ways: an older generation of bivalve settlement onto the wood, or more optimum growth conditions, as might be expected on the underside of a floating log, with more continuous seawater immersion. Growth rate studies could be used to distinguish between these scenarios, but this would require better preservation of a greater number of the bivalve specimens in BRLSI.M3941. If the latter scenario is correct then this can be used to interpret the orientation of the wood in the concretion, confirming that what have called the upper surface of the concretion was indeed uppermost stratigraphically. This interpretation would also match with the observation of a greater range of orientations of the bivalves associated with the wood bands (Fig. 2) compared to those on the wood edge (Figs. 2, 3C, D) because this is what would be expected when the wood settled onto the seafloor, compressing the layer (or layers) of bivalves growing on its underside, altering some (but not all, as can be seen in Fig. 4) of their orientations from pendent to having long axes parallel to the bottom of the wood (e.g., Fig. 3B).
The presence of the barnacle layer outside of the bivalve layer on the wood edge strongly indicates temporal succession, with the barnacle larvae settling onto the shells on outer edge of the bivalve layer, then developing short, flexible, scaleless peduncles (none of which are preserved in BRLSI.M3941). The first settlement of barnacles may have taken advantage of the inhalant and exhalent currents generated by the gills of the bivalves they settled onto. Thus, the barnacles were also pseudoplanktonic, and represent a second occurrence of the oldest pseudoplanktonic barnacles (Gale and Schweigert 2015). The single bivalve on the outside the barnacle layer on the wood edge (Fig. 2) may even represent a third settlement event, of bivalves onto barnacles. There is no unambiguous evidence of settlement of barnacles directly onto the wood in BRLSI.M3941, as the few barnacle specimens present in the wood layer occur as disarticulated plates and could represent fragments of specimens that grew on the bivalve layer(s) hanging from the underside of the wood, and which were then partially compacted when the wood came to rest on the seafloor. Unfortunately, this scenario cannot be proved or disproved because barnacle plates do not show up in the CT data. This could be due to a lack of resolution, as the barnacle plates are considerably thinner that the bivalve shells; and/or because the X-ray attenuation of both their phosphatic mineralogy and the surrounding matrix is too similar with the experimental setup used here, resulting in undetectable contrast in the images, although apatite has comparable or slightly higher X-ray attenuation coefficient than calcite.
After successive settlement of the bivalves and barnacles on its immersed surfaces, the wood would have floated in the surface layers of the sea for a period of time (likely several years), with the increasing weight of epifauna possibly keeping the wood in a constant orientation. At some point the wood would have become waterlogged and this, together perhaps with the weight of the epifauna, would have overcome its buoyancy, causing it to sink and settle gently onto the seafloor, with reorientation of some bivalves (and possibly barnacles) on the wood surface first in contact with the fine grained and probably soupy sediment. After seafloor emplacement there was decay of the animal soft tissues, including the scutal adductor muscles of some of the barnacles (causing kippering), all the barnacle peduncles, and all the byssal threads and internal soft tissues of the bivalves. However, there was no large-scale post-emplacement disturbance of the pseudoplanktonic association by scavengers or bioturbating organisms. This was because the seafloor during the formation of the Strawberry Bank fish bed was dysoxic or even anoxic, as evidenced by the exceptional preservation of vertebrate and cephalopod soft tissues in the same layer. This lack of disturbance meant that the bivalves remained articulated, even after their adductor muscles decayed. Subsequently, some thickness of fine grained carbonate sediment was deposited on the wood and associated fauna and at this time some of the empty articulated bivalves were completely or partially filled by sediment (Fig. 3A, C), whilst others remained empty. Fairly soon after this carbonate cements precipitated in the sediments enclosing the wood to form the concretions. This must have occurred very early on during diagenesis, and definitely pre-compaction, to account for the exceptional three-dimensional preservation of many of the vertebrate fossils in the fish bed concretions (e.g., Williams et al. 2015; Sinha et al. 2021), and may have been accelerated by decay of the animal soft tissues (e.g., Yoshida et al. 2015). It was probably at this stage that the remaining empty shells of the bivalves surrounding the wood became filled by calcite cement.
Comparisons with other Toarcian pseudoplankton examples.—To our knowledge BRLSI.M3941 is the only example of a pseudoplanktonic association of bivalves and barnacles from the Toarcian, or indeed the Jurassic. The other occurrence of Toarcolepas mutans, from the time equivalent Posidonia Shale, is associated with fossil wood, but without bivalves (Gale and Schweigert 2015). A younger occurrence of the Toarcolepas from the Kimmeridgian of Germany has specimens attached to the ammonite Lithacoceras sp. (Gale et al. 2019). Given this substrate colonisation variation within T. mutans (and Toarcolepas more generally), it is not possible to be sure if the colonisation of bivalves on wood by T. mutans, as seen in BRLSI.M3941, represents an obligatory, highly specialised palaeoecological succession or just an opportunistic encounter. The high levels of selectivity of some other Jurassic barnacles to specific substrata, for example, different ammonite taxa (Gale 2019; Gale et al. 2019) might support the latter interpretation, but it should also be noted that the cypris larvae of some barnacles, such as the common goose barnacle Lepas spp. are unselective and attach to almost any floating material (Southward 2008).
The pseudoplanktonic attachment of Pseudomytiloides dubius on wood in BRLSI.M3941 is consistent with observations from the Posidonia Shale in Germany (e.g., Hauff and Hauff 1981; Matzke and Maisch 2019) and the Mulgrave Shale from the UK (CTSL, personal observation). The presence of P. dubius specimens not associated with wood or other potentially floating substrates (e.g., ammonites) in the fish bed concretions in the BRLSI collection seems to confirm the facultative pseudoplanktonic palaecological determination for this species (Kauffman 1981; Etter 1996; Caswell et al. 2009). The temporal succession of bivalves then barnacles (and possibly bivalves again) onto wood in BRLSI.M3941 is one of few such examples for this phenomenon in pseudoplankton for the Jurassic; it has been shown for Seirocrinus subangularis intra-specifically (Matzke and Maisch 2019), and might also have occurred with P. dubius and S. subangularis, although this needs to be proven.
The occurrence of the pseudoplankton association in the Strawberry Bank mixed carbonate-siliciclastic facies is unusual in a Toarcian context, as other examples occur in black shale facies (Seilacher et al. 1968; Hauff and Hauff 1981; Matzke and Maisch 2019; Hunter et al. 2020), but this is entirely in keeping with the principles of pseudoplankton, in that they should be independent of facies (Simms 1986; Wignall and Simms 1990). It cannot be coincidental that all these occurrences are in rocks that were deposited during times of reduced oxygen levels at the seafloor, thus preserving both delicate fossils (i.e., the barnacles) and also the physical relationship with their previously floating substate. Finally, we note that eight pieces of carbonate concretion with co-occurring specimens of Pseudomytiloides dubius and Toarcolepas mutans exist in the collections of the South West Heritage Trust, Taunton, UK (TTNCM.39/2011/0327–0334). These concretions come from the original Moore collection, so it is highly probable that they are pieces of the same concretion as BRLSI.M3941, although none of them have any associated wood.
Conclusions
Specimen BRLSI.M3941 is a limestone concretion from the Lower Toarcian Strawberry Bank Lagerstätte (Ilminster, Somerset, UK) that preserves a piece of fossil wood with a pseudoplanktonic colony comprising hundreds of eolepadid barnacles (Toarcolepas mutans) that attached onto a layer of inoceramid bivalves (Pseudomytiloides dubius), that had already attached onto the wood. This is one of very few examples of temporal succession for pseudoplankton in the Toarcian and is also unusual in being preserved in a mixed carbonate-siliciclastic facies, rather than a black shale. The occurrence of T. mutans in BRLSI.M3941 represents the second record of the species and also the equal oldest example of pseudoplanktonic barnacles in the fossil record.
Acknowledgements
We thank the directors of the Bath Royal Literary and Scientific Institution for permitting access to BRLSI.M3941, and also Steffen Kiel (Naturhistoriska riksmuseet, Stockholm, Sweden) and Aaron Hunter (University of Western Australia, Crawley, Australia) for their review comments on the paper.
References
Audo, D., Williams, M., Charbonnier, S., and Schweigert, G. 2017. Gabaleryon, a new genus of widespread early Toarcian polychelidan lobsters. Journal of Systematic Palaeontology 15: 205–222. Crossref
Ballell, A., Moon, B.C., Porro, L.B., Benton, M.J., and Rayfield, E.J. 2019. Convergence and functional evolution of longirostry in crocodylomorphs. Palaeontology 62: 867–887. Crossref
Boomer, I., Copestake, P., Page, P., Huxtable, J., Loy, T., Bown, P., Dunkley Jones, T., O’Callaghan, M., Hawkes, S., Halfacree, D., Reay, H., and Caughtry, N. 2021. Biotic and stable-isotope characterization of the Toarcian Ocean Anoxic Event through a carbonate-clastic sequence from Somerset, UK. In: M. Reolid, L.V. Duarte, E. Mattioli, and W. Ruebsam (eds.), Carbon Cycle and Ecosystem Response to the Jenkyns Event in the Early Toarcian (Jurassic). Geological Society of London Special Publications 514: 239–268. Crossref
Butler, R., Fernandez, V., Nesbitt, S.J., Leite, J.V., and Gower, D. 2022. A new pseudosuchian archosaur, Mambawakale ruhuhu gen. et sp. nov., from the Middle Triassic Manda Beds of Tanzania. Royal Society Open Science 9: 211622. Crossref
Caine, H. and Benton, M.J. 2011. Ichthyosauria from the Upper Lias of Strawberry Bank, England. Palaeontology 54: 1069–1093. Crossref
Caswell, B.A., Coe, A.L., and Cohen, A.S. 2009. New range data for marine invertebrate species across the early Toarcian (Early Jurassic) mass extinction. Journal of the Geological Society 166: 859–872. Crossref
Cawley, J.J., Kriwet, J., Klug, S., and Benton, M.J. 2019. The stem group teleost Pachycormus (Pachycormiformes: Pachycormidae) from the Upper Lias (Lower Jurassic) of Strawberry Bank, UK. Paläontologische Zeitschrift 93: 285–302. Crossref
Etter, W. 1996. Pseudoplanktonic and benthic invertebrates in the Middle Jurassic Opalinum Clay, northern Switzerland. Palaeogeography, Palaeoclimatology, Palaeoecology 126: 325–341. Crossref
Gale, A.S. 2019. Stalked barnacles (Cirripedia, Thoracica) from the Late Jurassic (Tithonian) Kimmeridge Clay of Dorset, UK; palaeoecology and bearing on the evolution of living forms. Proceedings of the Geologists’ Association 130: 355–365. Crossref
Gale, A.S. and Schweigert, G. 2015. A new phosphatic-shelled cirripede (Crustacea, Thoracica) from the Lower Jurassic (Toarcian) of Germany—the oldest epiplanktonic barnacle. Palaeontology 59: 59–70. Crossref
Gale, A.S., Schweigert, G., Keupp, H., and Roper, M. 2019. Thoracican cirripedes (Crustacea) from the Kimmeridgian of Brunn and Nusplingen (southern Germany), and their bearing on the origin of calanticid and scalpellid barnacles. Neues Jahrbuch fur Geologie und Palaontologie Abhandlungen 293: 1–17. Crossref
Hammer, Ø., Harper, D.A.T., and Ryan, P.D. 2001. PAST: Paleontological Statistics software for education and data analysis. Palaeontologia Electronica 4: 1–9.
Harries, P.J. and Little, C.T.S. 1999. The early Toarcian (Early Jurassic) and the Cenomanian–Turonian (Late Cretaceous) mass extinctions: similarities and contrasts. Palaeogeography, Palaeoclimatology, Palaeoecology 154: 39–66. Crossref
Hauff, B. and Hauff, R.B. 1981. Das Holzmadenbuch. 136 pp. Repro-Druck GmbH, Fellbach.
Huggett, J.M. and Gale, A.S. 1995. Palaeoecology and diagenesis of concretions containing bored wood from the London Clay of Sheppey. Proceedings of the Geologists’ Association 106: 119–136. Crossref
Hunter, A.W., Casenove, D., Mayers, C., and Mitchell, E.G. 2020. Reconstructing the ecology of a Jurassic pseudoplanktonic raft colony. Royal Society Open Science 7: 200142. Crossref
Kauffman, E.G. 1981. Ecological reappraisal of the German Posidonienschiefer (Toarcian) and the stagnant basin model. In: W. Berry, A. Boucot, and J. Gray (eds.), Communities of the Past, 311–381. Hutchinson Ross, Stroudsburg.
Marek, R.D., Moon, B.C., Williams, M., and Benton, M.J. 2015. The skull and endocranium of a Lower Jurassic ichthyosaur based on digital reconstructions. Palaeontology 58: 723–742. Crossref
Matzke, A.T. and Maisch, M.W. 2019. Palaeoecology and taphonomy of a Seirocrinus (Echinodermata: Crinoidea) colony from the early Jurassic Posidonienschiefer Formation (Early Toarcian) of Dotternhausen (SW Germany). Neues Jahrbuch für Geologie und Palaontologie 291: 89–107. Crossref
Moore, C. 1866. On the Middle and Upper Lias of the South West of England. Proceedings of Somerset Archaeological and Natural History Society 13: 119–244.
Pierce, S. and Benton, M.J. 2006. Pelagosaurus typus Bronn, 1841 (Mesoeucrocodylia: Thalattosuchia) from the Upper Lias (Toarcian; Lower Jurassic) of Somerset, England. Journal of Vertebrate Paleontology 26: 621–635. Crossref
Powell, J.H. 1984. Lithostratigraphical nomenclature of the Lias Group in the Yorkshire Basin. Proceedings of the Yorkshire Geological Society 45: 51–57. Crossref
Seilacher, A,, Drozdzewski, G., and Haude, R. 1968, Form and function of the stem in a pseudoplanktonic crinoid (Seirocrinus). Palaeontology 11: 275–282.
Simms, M.J. 1986. Contrasting lifestyles in Lower Jurassic crinoids: a comparison of benthic and pseudopelagic Isocrinida. Palaeontology 29: 475–493.
Sinha, S., Muscente, A.D., Schiffbauer, J.D., Williams, M., Schweigert, G., and Martindale, R.C. 2021. Global controls on phosphatization of fossils during the Toarcian Oceanic Anoxic Event. Scientific Reports 11: 24087. Crossref
Southward, A.J. 2008. Barnacles. In: J.H. Crothers and P.J. Hayward (eds.), Synopses of the British Fauna (New Series) 57, 1–140. Field Studies Council, Shrewsbury.
Sowerby, J. de C. 1828. The mineral conchology of Great Britain; or coloured figures and descriptions of those remains of testaceous animals or shells, which have been preserved at various times and depths in the Earth. Vol. VI, 157–200. Privately published, London.
Tsikhon-Lukanina, E.A., Reznichenko, O.G., and Nikolaeva, G.G. 2001. Ecology of invertebrates on the oceanic floating substrata in the Northwest Pacific Ocean. Russian Academy of Sciences, Oceanology 41: 525–530.
Wignall, P.B. 1994. Black Shales. 127 pp. Oxford University Press, Oxford.
Wignall, P.B. and Simms, M.J. 1990. Pseudoplankton. Palaeontology 33: 359–378.
Williams, M., Benton, M.J., and Ross, A. 2015. The Strawberry Bank Lagerstätte reveals insights into Early Jurassic life. Journal of the Geological Society 172: 683–692. Crossref
Yoshida, H., Ujihara, A., Minami, M., Asahara, Y., Katsuta, N., Yamamoto, K., Sirono, S., Maruyama, I., Nishimoto, S., and Metcalfe, R. 2015. Early post-mortem formation of carbonate concretions around tusk-shells over week-month timescales. Scientific Reports 5: 14123. Crossref
Acta Palaeontol. Pol. 68 (1): 133–142, 2023
https://doi.org/10.4202/app.01018.2022