
New information on the cranial anatomy of the Middle Triassic rhynchosaurian reptile Bentonyx sidensis
THITIWOOT SETHAPANICHSAKUL, ROBERT A. CORAM, and MICHAEL J. BENTON
Sethapanichsakul, T., Coram, R.A., and Benton, M.J. 2023. New information on the cranial anatomy of the Middle Triassic rhynchosaurian reptile Bentonyx sidensis. Acta Palaeontologica Polonica 68 (1): 53–62.
Rhynchosaurs were key herbivores over much of the world in the Middle and Late Triassic, often dominating their faunas ecologically. The anatomy of the Middle Triassic forms is debated in certain phylogenetically important details. Among several genera from the Anisian of England, Bentonyx sidensis is represented by a near-complete skull. New CT scanning of the holotype skull reveals previously concealed details of the palate and braincase in particular, and this allows comparison with other Middle Triassic rhynchosaurs and confirmation that Bentonyx is a distinct taxon from others of similar age including Fodonyx, Rhynchosaurus, Langeronyx, and Stenaulorhynchus. These are part of the first wave of rhynchosaur diversification in the Middle Triassic.
Key words: Reptilia, Rhynchosauria, Bentonyx, skull anatomy, Middle Triassic, Anisian, UK.
Thitiwoot Sethapanichsakul [sethapth@gmail.com; ORCID: https://orcid.org/0000-0001-5739-4774 ] and Michael J. Benton [mike.benton@bristol.ac.uk; ORCID: https://orcid.org/0000-0002-4323-1824 ], School of Earth Sciences, University of Bristol, Bristol, BS8 1RJ, UK.
Robert Coram [rob@britishfossils.co.uk; ORCID: https://orcid.org/0000-0003-3048-6665 ], British Fossils, Unit 18-20 Bailie Gate Industrial Estate, Sturminster Marshall, Wimborne, Dorset BH21 4DB, UK; and School of Earth Sciences, University of Bristol, Bristol, BS8 1RJ, UK.
Received 26 August 2022, accepted 14 November 2022, available online 20 January 2023.
Copyright © 2023 T. Sethapanichsakul et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Rhynchosaurs were archosauromorph reptiles that appeared in the Early Triassic and became important terrestrial herbivores over much of the world during the Middle and early Late Triassic, from 247–227 Ma. During this time, they diversified across what are now Europe, Africa, the Indian subcontinent, and the Americas at a time when the Earth’s major landmasses were united in the supercontinent Pangea and terrestrial ecosystems were rebuilding themselves following the devastating end-Permian mass extinction, 252 Ma (Benton et al. 2004; Chen and Benton 2012).
Middle Triassic rhynchosaurs have been known since 1840 from the Helsby Sandstone Formation (late Anisian) of England (Murchison and Strickland 1840; Owen 1842), a series of sandstone-dominated, fluvial, terrestrial units known locally as the Bromsgrove Sandstone in the West Midlands and the Otter Sandstone in Devon (Benton et al. 1994; Benton 1990). The Otter Sandstone in particular has yielded a well-documented fauna of fishes, temnospondyl amphibians, procolophonids, stem diapsids, and rauisuchians (Benton et al. 1994; Coram et al. 2019, 2021), together with two rhynchosaurs, Bentonyx sidensis Langer, Montefeltro, Hone, Whatley, and Schultz, 2010, and Fodonyx spenceri Hone and Benton, 2008. Between them, these rhynchosaurs make up approximately 50% of identifiable macrovertebrate material in the Otter Sandstone and are the only recorded medium-large sized herbivores (Benton et al. 1994; Coram et al. 2019).
The holotype of the rhynchosaur Bentonyx sidensis is a well-preserved skull that has been described in some detail from external appearances (Hone and Benton 2008; Langer et al. 2010). CT scanning of this specimen has now revealed previously obscured anatomical details.
Institutional abbreviations.—BRSUG, University of Bristol, Geology Collections, School of Earth Sciences, UK; MANCH, Manchester Museum, University of Manchester, UK; NHMUK, Natural History Museum, London, UK; SHRBM, Shrewsbury Museum and Art Gallery, UK.
Material and methods
Specimens.—The study focuses on the complete skull of the holotype specimen of Bentonyx sidensis (BRSUG 27200). This specimen comes from near the top of the Otter Sandstone of Sidmouth, Devon, UK, dated as late Anisian (Middle Triassic; c. 242 Ma), on the basis of regional geological correlations, fossils, and magnetostratigraphy (Hounslow and McIntosh 2003). The Otter Sandstone is a sedimentary unit characterized by fluvial sandstone beds deposited under a semi-arid climate (Coram et al. 2019, 2021) and assigned to the Helsby Sandstone Formation, a division of the Sherwood Sandstone Group (Ambrose et al. 2014).
Scanning and segmentation.—The specimen was scanned using the Nikon XTH 225ST X-ray tomography scanner belonging to the Palaeobiology Research Group in the School of Earth Sciences, University of Bristol, at 144 kV which produced a dataset containing 2809 slices at a voxel resolution of 50.68 µm.
Segmentation of skeletal elements from the computed tomography (CT) datasets was performed using Avizo 2020.2 (Visualization Sciences Group) to generate three-dimensional models. In addition to segmenting bone from matrix, specific elements were digitally isolated. All surface models were generated in Avizo with an unconstrained smoothing factor of 2, and then exported in an ASCII Standard Triangle Language (STL) format. These files were then imported into Blender 2.93.3 (Blender Foundation) for the purpose of taking photographs and model manipulation. Measurements were also taken in Blender, and the unit scaling was changed to 0.00137 to calibrate the model for accurate measurements of the 3D model.
Data archiving statement.—Data for this study are available in the MorphoSource Digital Repository: https://www.morphosource.org/projects/000481699
Results
BRSUG 27200 is the holotype adult skull of Bentonyx sidensis (Langer et al. 2010), previously described by Hone and Benton (2008), who covered most of the cranial elements that had been prepared. The CT scans reveal additional details of the anatomy not seen in the earlier descriptions (Hone and Benton 2008; Langer et al. 2010). In particular, the model rendered from the scans (Figs. 1–6) displays additional details of the braincase, palate, and tooth-bearing elements.
The calcitic matrix surrounding the skeletal elements has a similar density to the bone, making the identification of matrix and bone separation difficult. Calcite granules within the matrix appear as globules and lenses in the CT slices that can easily be mistaken for bone, and sometimes overgrow the bone, especially around the braincase. These crystalline overgrowths can obscure true bone surfaces, hiding the sutures and any surface textures. Nonetheless, the scans revealed a great deal of detail of the portions of the skull previously obscured within the rock, and these are the focus of the descriptions here.
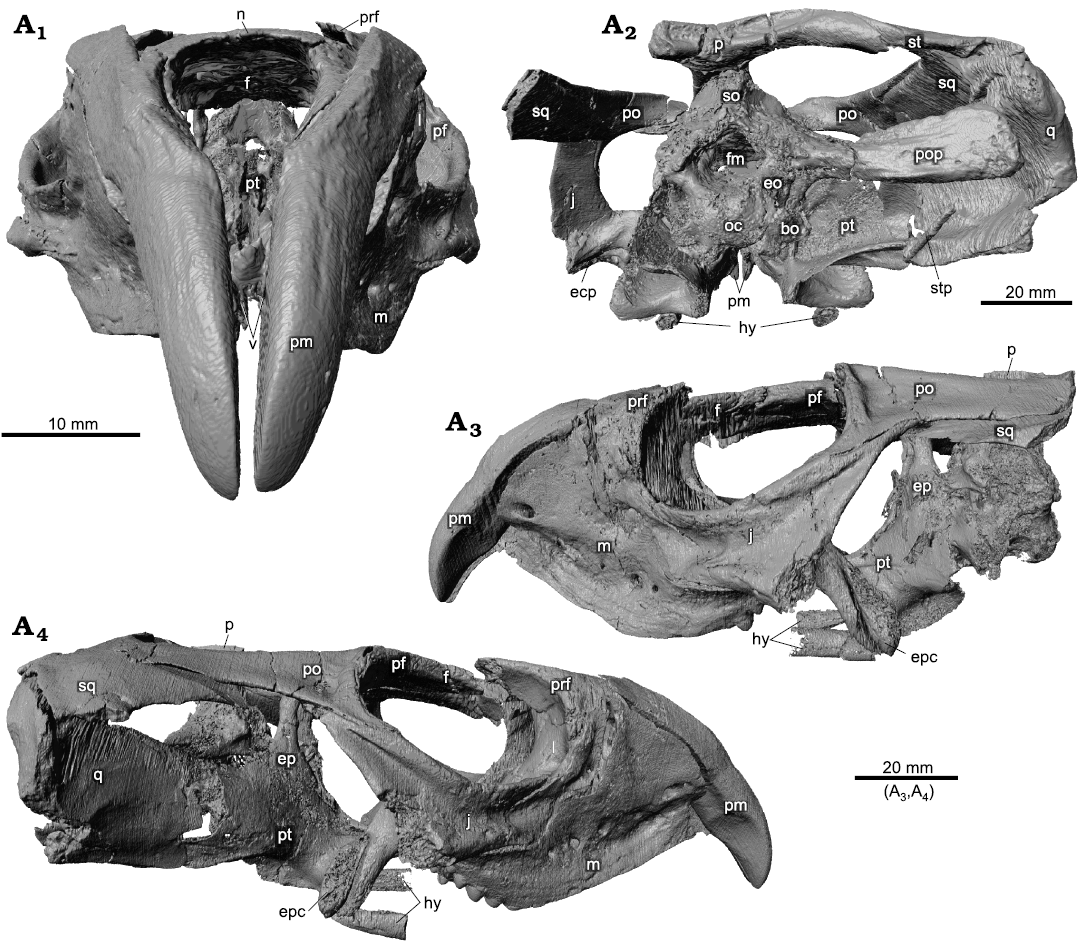
Fig. 1. Holotype of the rhynchosaurian reptile Bentonyx sidensis Hone and Benton, 2008 (BRSUG 27200) from Otter Sandstone (Middle Triassic) of Devon, UK. Skull in anterior (A1), posterior (A2), left lateral (A3), and right lateral (A4) views. Images generated from the CT scan data. Abbreviations: bo, basioccipital; ecp, ectopterygoid; eo, exoccipital; ep, epipterygoid; fm, foramen magnum; hy, hyoid; j, jugal; l, lacrimal; m, maxilla; n, nasal; oc, occipital condyle; p, parietal; pf, postfrontal; pm, premaxilla; po, postorbital; pop, paroccipital process; prf, prefrontal; pt, pterygoid; q, quadrate; so, supraoccipital; sq, squamosal; st, supratemporal; stp, stapes; v, vomer.
The skull is relatively complete, mainly lacking the posterior left-hand portion and parts of the temporal arch on the right-hand side (Figs. 1–4). The premaxillary tusks are complete in anterior view (Figs. 1A1, 2A1), and swathed on either side by the anterior processes of the maxillae (Figs. 1A1, A3, A4, 2A1, A3, A4). The maxillae, jugals, and orbital regions are complete on both sides (Figs. 1A3, A4, 2A3, A4), as are the upper portions of the temporal area. The braincase is relatively complete and in approximately correct position (Figs. 1A2, 2A2, 5, 6). The dorsal view of the skull (Figs. 3A1, 4A1) shows most elements except the posterior left-hand angle, and bones are relatively undistorted. The ventral (Figs. 3A2, 4A2) and dorsal (Figs. 3A3, 4A3) views of the palate provide a great deal of new information. We focus our description on the palate and braincase and consider the tooth-bearing elements elsewhere.
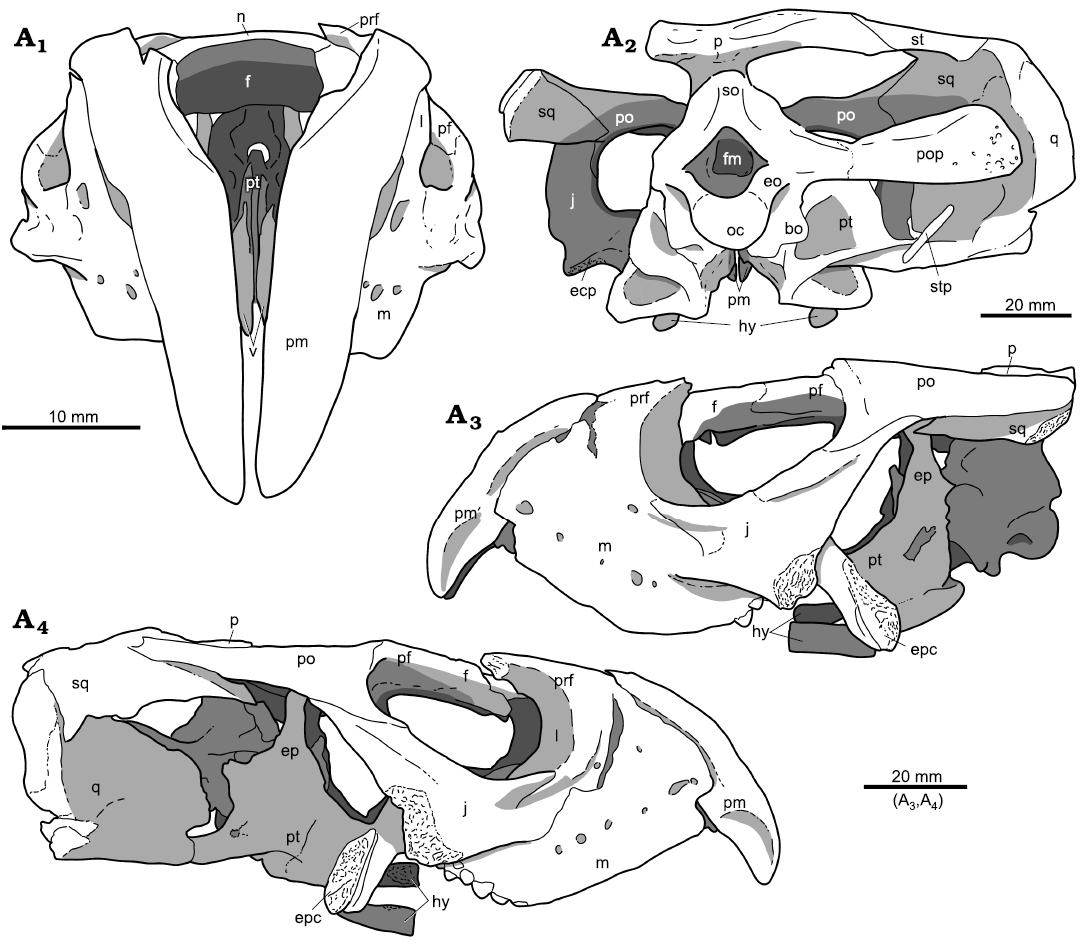
Fig. 2. Holotype of the rhynchosaurian reptile Bentonyx sidensis Hone and Benton, 2008 (BRSUG 27200) from Otter Sandstone (Middle Triassic) of Devon, UK. Skull in anterior (A1), posterior (A2), left lateral (A3), and right lateral (A4) views. Outline drawings to match CT scan images in Fig. 1. Abbreviations: bo, basioccipital; ecp, ectopterygoid; eo, exoccipital; ep, epipterygoid; f, frontal; fm, foramen magnum; hy, hyoid; j, jugal; l, lacrimal; m, maxilla; n, nasal; oc, occipital condyle; p, parietal; pf, postfrontal; pm, premaxilla; po, postorbital; pop, paroccipital process; prf, prefrontal; pt, pterygoid; q, quadrate; so, supraoccipital; sq, squamosal; st, supratemporal; stp, stapes; v, vomer.
Palate.—The palate was partially described previously (Hone and Benton 2008: 98), but here we can present it in its entirety (Figs. 3A2, A3, 4A2, A3). The palatal series is well preserved and mostly intact, but the contact between vomer and palatine is separated and damaged. We cannot detect any palatal teeth, whether on pterygoid, palatine, or vomer, based on careful scrutiny of the scans.
We could not identify a septomaxilla, an important feature in early archosauromorphs, but generally not reported in rhynchosaurs (Ezcurra 2016: 131) until it was identified in Hyperodapedon sanjuanensis (Gentil and Ezcurra 2022: figs. 3a, 9a, 10). In this taxon, the septomaxilla is a small, elongate element, visible in ventral view of the palate, located on either side of a small midline opening and meeting the anterior tips of the vomer and maxillary tooth plate, nestling at the root of the premaxillary tusk as it contacts these palatal bones. In Bentonyx sidensis, the anterior tip of the maxillary tooth plate and of the vomer end below and medially of the proximal root of the premaxillary tusk, in palatal view (Figs. 3A2, 4A2), but we cannot distinguish a separate element, a septomaxilla, linking between these elements. Whether it was 5, replace there and has been lost or is not part of the cranial anatomy of this animal cannot be said. Gentil and Ezcurra (2022: fig. 15) identify septomaxillae also in the early rhynchosaur Mesosuchus browni, as well as in the derived taxa Teyumbaita sulcognathus and Hyperodapedon mariensis, so the bone was presumably present throughout rhynchosaurs, but has proved hard to identify and might, as in B. sidensis, not survive fossilisation in some cases.
The vomer has been described, but the model reveals its dorsal aspect (Figs. 3A3, 4A3). The paired vomers have been offset vertically. The dorsal process of the vomers is thin and rises dorsally, and a canal is formed on each paired element that extends along their full length.
The palatine is broad and appears to conform and undulate in a similar fashion to the other cranial elements it contacts. The medial margin of the palatines meets the lateral margin of the anterior process of the pterygoid. The lateral wing contacts the dorsomedial surface of the jugal and the medial surface of the maxilla, its posterolateral wing meets the ectopterygoid and the pterygoid forming the margins of the infraorbital fenestra. Ventrally (Figs. 3A2, 4A2), the maxillary contacts generate a ventral ridge that curls medially conforming with the dorsomedial surface of the maxilla.

Fig. 3. Holotype of the rhynchosaurian reptile Bentonyx sidensis Hone and Benton, 2008 (BRSUG 27200) from Otter Sandstone (Middle Triassic) of Devon, UK. Skull in dorsal (A1), ventral palatal (A2), and dorsal palatal (A3) views. Images generated from the CT scan data. Abbreviations: aaf, anterior alveolar foramen; bo, basioccipital; ch, choana; ecp, ectopterygoid; ep, epipterygoid; f, frontal; hy, hyoid; iof, infraorbital foramen; j, jugal; lr, lateral ridge; m, maxilla; n, nasal; oc, occipital condyle; op, opisthotic; p, parietal; paf, posterior alveolar foramen; pal, palatine; pbs, parabasisphenoid; pf, postfrontal; pm, premaxilla; po, postorbital; pop, paroccipital process; pr, prootic; prf, prefrontal; pt, pterygoid; q, quadrate; pt(qp), quadrate process of the pterygoid; so, supraoccipital; sq, squamosal; st, supratemporal; v, vomer.
The pterygoids form the largest element of the palate, each possessing three main processes, the longest of which is the anterior process that overlaps a posterior portion of the vomers. In dorsal view (Figs. 3A3, 4A3), the anterior process possesses a canal and tall medial ridge continuous to that seen in the vomers. This morphology is retained throughout the anterior process before reducing to merge with the lateral and posterior pterygoid processes. The lateral process of the pterygoid is dorsoventrally flat and robust. It is the shortest of the three processes and sweeps posteriorly in a broad arc to form the posterolateral process. The posterolateral process extends posterolaterally and meets the ventral margins of the quadrate. The posterior process has a complex morphology, being broad and flat and twisting clockwise slightly, so appearing slightly curved in lateral view. There are several ridges on the posterior portion of the medial surface, including a dorsally trending ridge that forms the posterior margin of the pocket into which the basipterygoid process of the braincase inserts (Figs. 3A3, 4A3). The lateral ridge appears like a shelf extending along the length of the process, becoming less pronounced distally.
The ectopterygoid is robust and spans behind the maxillary tooth plate, meeting the ventromedial margin of the jugal. Its posteroventral process contacts the anterior margin of the lateral wing of the pterygoid and expands medially. The anterior portion of the ectopterygoid flattens out to contact the jugal and the entire posterior termination of the maxillary tooth plate, before curving dorsally to form part of the posterior margin of the orbit.
Posterodorsal to the basipterygoid pocket is the epipterygoid (Figs. 1A4, 2A4, 3A3, 4A3), a narrow vertical rod that rises to meet the underside of the parietal.
The anterior tip of a hyoid is preserved on the right-hand side (hy, Figs. 3A2, 4A2), nearly circular in cross section and ending anteriorly with a flat surface. The location and shape are similar to putative hyoids reported in Hyperodapedon gordoni (Benton 1983: 637–638) and Rhynchosaurus articeps (Benton 1990: 234, figs. 4c, e, 5c, 16, 44f, 45d), where they were interpreted as ossified ventrally placed ceratobranchials or ossified epihyal portions of the ceratohyal. These structures could have helped amplify hearing of ground-borne sounds or might have formed part of the anchoring for a substantial tongue.
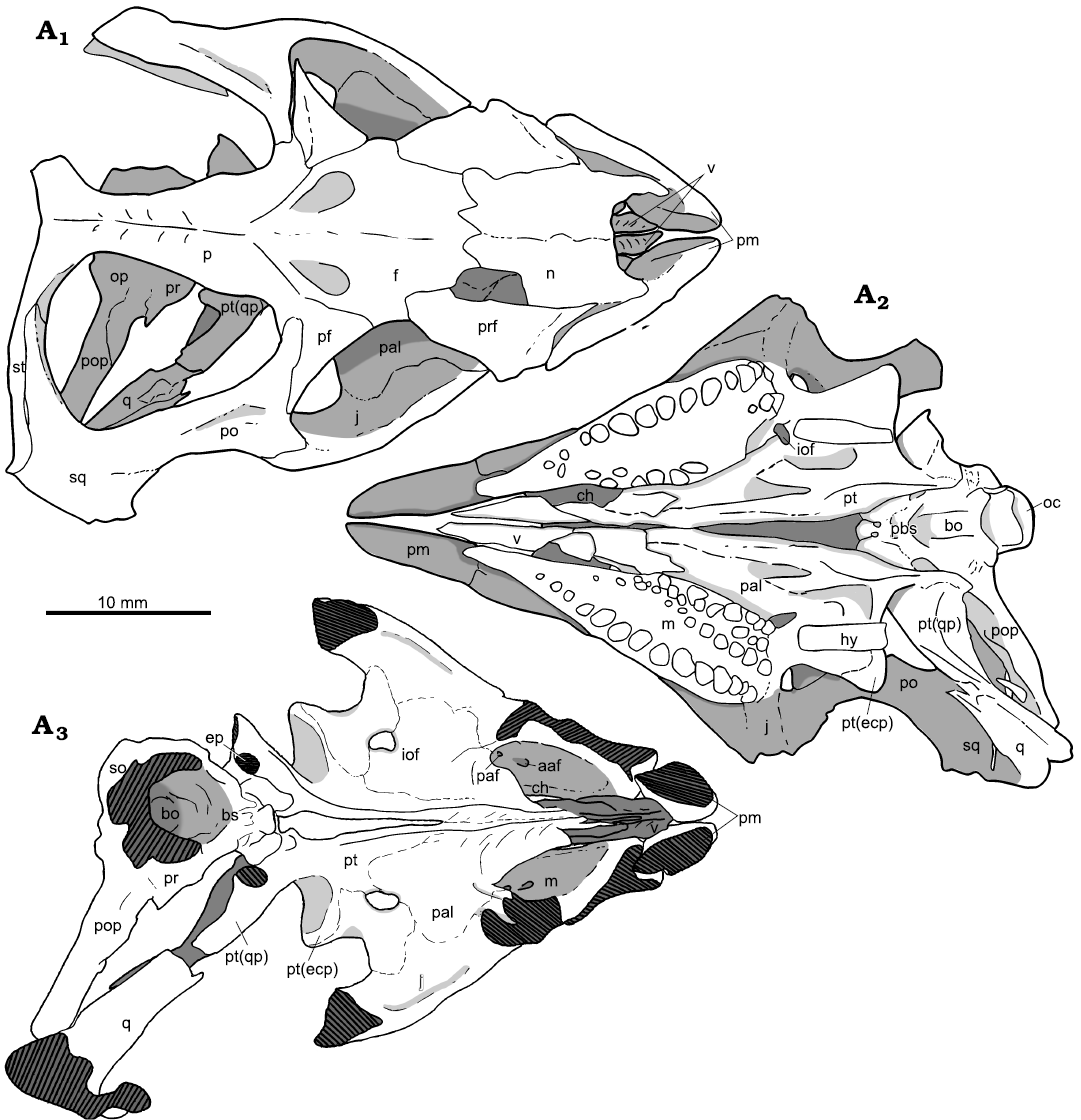
Fig. 4. Holotype of the rhynchosaurian reptile Bentonyx sidensis Hone and Benton, 2008 (BRSUG 27200) from Otter Sandstone (Middle Triassic) of Devon, UK. Skull in dorsal (A1), ventral, palatal (A2), and dorsal palatal (A3) views. Outline drawings to match CT scan images in Fig. 3. Abbreviations: aaf, anterior alveolar foramen; bo, basioccipital; ch, choana; ecp, ectopterygoid; ep, epipterygoid; f, frontal; hy, hyoid; iof, infraorbital foramen; j, jugal; lr, lateral ridge; m, maxilla; n, nasal; oc, occipital condyle; op, opisthotic; p, parietal; paf, posterior alveolar foramen; pal, palatine; pbs, parabasisphenoid; pf, postfrontal; pm, premaxilla; po, postorbital; pop, paroccipital process; pr, prootic; prf, prefrontal; pt, pterygoid; q, quadrate; pt(qp), quadrate process of the pterygoid; so, supraoccipital; sq, squamosal; st, supratemporal; v, vomer.
Braincase.—The braincase is in place within the cranium but has been shifted slightly to the right (Figs. 1A2, 5, 6). Parts of the braincase, notably the distal tip of the paroccipital process, are well preserved, but the midline elements show considerable damage by abrasion or recyrystallisation on burial. The contact between the pterygoid and the parabasisphenoid has been preserved. The braincase itself is typical of rhynchosaurs and has been partially described, but the CT scans reveal further details of the morphology of several buried braincase elements.
The basioccipital is only generally visible, as a damaged occipital condyle (Figs. 5A1, 6A1), and contracting anteriorly before expanding as the spheno-occipital tubera at the contact with the parabasisphenoid (Figs. 5A3, A4, A6, 6A3, A4, A6).
The parabasisphenoid is mostly intact with some distortion where calcite overgrowth damaged the structure, but the anterior parasphenoid process is absent, whether through true absence or lack of preservation is uncertain. The parabasisphenoid makes up the anterior section of the braincase and lies anterior to the basioccipital. In anterior view (Figs. 5A2, 6A2) the parabasisphenoid rises vertically forming a pronounced dorsum sellae. The foramina for the carotid arteries and for cranial nerve VI cannot be distinguished because of damage to the specimen. Between the basipterygoid processes lies a deep fossa identified as the site of the paired carotid foramina, but detail is absent. Ridges extend antero-posteriorly along the lateroventral side of the element. The median pharyngeal recess (Figs. 5A6, 6A6) is deep and extends almost the entire antero-posterior length of the ventral side of the parabasisphenoid.
The exoccipital is best preserved on the left side. It makes up the lateroventral margin of the foramen magnum. The sutures with the opisthotic and basioccipital are unclear, as is the opening for cranial nerve XII.
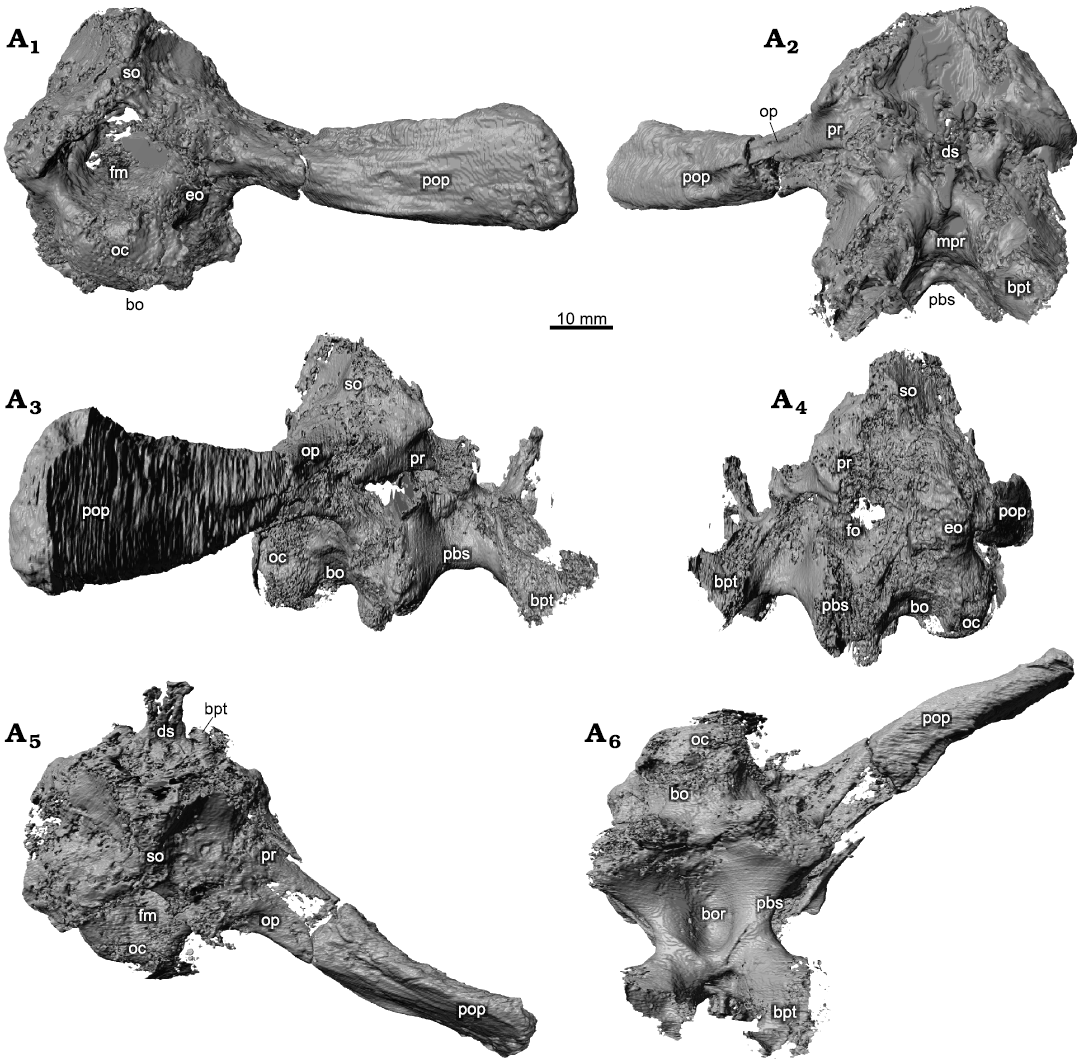
Fig. 5. Holotype of the rhynchosaurian reptile Bentonyx sidensis Hone and Benton, 2008 (BRSUG 27200) from Otter Sandstone (Middle Triassic) of Devon, UK. Braincase in posterior (A1), anterior (A2), right lateral (A3) and left lateral (A4), dorsal (A5), and ventral (A6) views. Images generated from the CT scan data. Abbreviations: bo, basioccipital; bor, basioccipital recess; bpt, basipterygoid; ds, dorsum sellae; eo, exoccipital; fm, foramen magnum; fo, fenestra ovalis; mpr, median pharyngeal recess; oc, occipital condyle; pbs, parabasisphenoid; pop, paroccipital process; pr, prootic; so, supraoccipital.
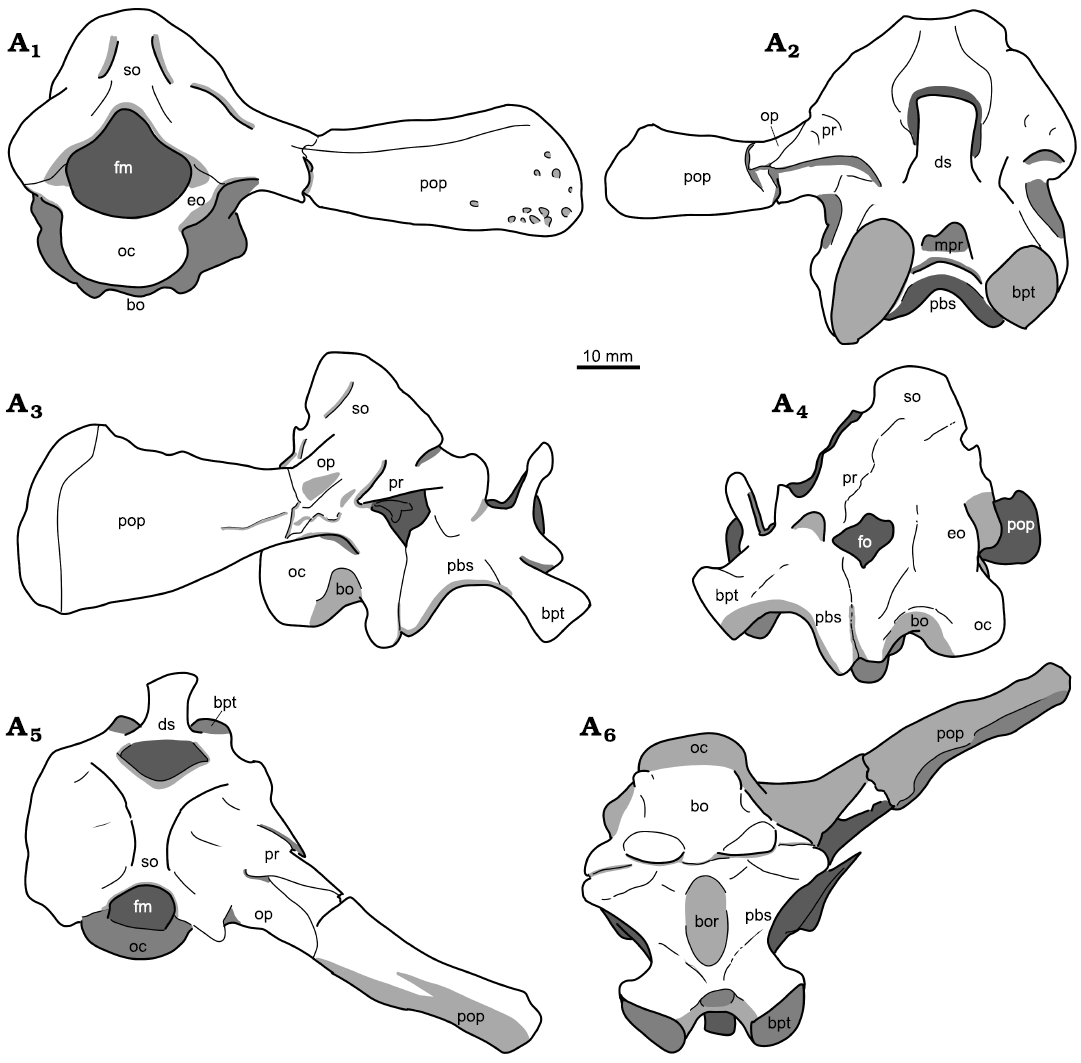
Fig. 6. Holotype of the rhynchosaurian reptile Bentonyx sidensis Hone and Benton, 2008 (BRSUG 27200) from Otter Sandstone (Middle Triassic) of Devon, UK. Braincase in posterior (A1), anterior (A2), left lateral (A3) and right lateral (A4), dorsal (A5), and ventral (A6) views. Outline drawings to match CT scan images in Fig. 5. Abbreviations: bo, basioccipital; bor, basioccipital recess; bpt, basipterygoid; ds, dorsum sellae; eo, exoccipital; fm, foramen magnum; fo, fenestra ovalis; mpr, median pharyngeal recess; oc, occipital condyle; pbs, parabasisphenoid; pop, paroccipital process; pr, prootic; so, supraoccipital.
The lateral faces of the braincase are poorly preserved. Although some cranial nerve foramina can be observed, precise identification is difficult. The prootic is best preserved on the right side. A subdued ridge extends posterolaterally along the paroccipital process of the opisthotic. The curve of the medial margin of the prootic is much less pronounced than in Hyperodapedon gordoni (Benton 1983: fig. 10). A single fragment of a stapes is preserved on the right side lying along the posterior end of the posterolateral process of the pterygoid (Figs. 1A4, 2A4).
Discussion
The most immediate comparison is with Fodonyx spenceri, also from the Otter Sandstone. In discriminating Fodonyx and Bentonyx, although the skulls in the holotypes of type species of both taxa are about the same length (140 mm), they differ greatly in posterior width, 130 mm in Bentonyx and nearly 170 mm in Fodonyx. Langer et al. (2010) regarded both as adults, and suggested Bentonyx was not a juvenile of Fodonyx.
There are some further differences between Bentonyx and Fodonyx (cf. Benton 1990: 262–267) in details of cranial anatomy. Langer et al. (2010) noted that Bentonyx has a slender anterior branch of the jugal (about the same depth as the underlying portion of the maxilla), the anterior margin of the quadrate branch of the pterygoid forms an angle of less than 50° to the midline, and the maxillary tooth plates comprise more than half the palatal length when measured along the midline. Autapomorphies of Bentonyx are “a rounded depression on the ventral surface of the basisphenoid and exceptionally large basal tubera” (Langer et al. 2010: 1884), both of which features distinguish the genus from all other rhynchosaurs with a preserved braincase, but it cannot be said whether these features do not occur in Fodonyx as its braincase is not known.
Among Middle Triassic rhynchosaurs from England, Langeronyx brodiei is known from the Bromsgrove Sandstone of Warwick, age-equivalent to the Otter Sandstone (Benton 1990; Ezcurra et al. 2016). It had an estimated skull length of 90–140 mm (holotype, 93 mm; Benton 1990: 254), the largest specimens being similar in size to Bentonyx sidensis and Fodonyx spenceri, but its skull is considerably narrower, with a width to length ratio of 0.7, compared to B. sidensis (0.9) and F. spenceri (1.2). Further, the cranium of L. brodiei differs from all other rhynchosaurs in the following combination of character states (Ezcurra et al. 2016: 14): “laterally facing and sub-circular orbit with a raised and thickened border that is less marked along the postorbital bone than other parts of the orbital margin; anterior process of the jugal dorsoventrally tall, in which the ratio between the height of the base of the anterior process and the anteroposterior length of the orbit is 0.37, with this process being exposed more broadly than the maxilla in lateral view; dorsal surface of the prefrontal deeply concave; dorsal surface of the frontal slightly concave transversely; occlusal dentition of maxilla composed of two tooth-bearing areas separated by a single well-delimited longitudinal sulcus; maxillary lateral tooth-bearing area is crest-shaped; lingual maxillary teeth scattered, do not form clear rows and are not regularly spaced; lingual dentary teeth conical and of similar size to the occlusal teeth”.
The fourth rhynchosaur from the Helsby Sandstone Formation, Rhynchosaurus articeps, is phylogenetically quite different, and less derived than the others (Langer et al. 2010; Ezcurra et al. 2016). Skull specimens are smaller than the other taxa, measuring 60–82 mm in length and with a width-to-length ratio of 0.8–0.9. The premaxillae are also relatively longer and the jugal more slender and less heavily sculpted. Likewise, the jugal-postorbital bar between orbit and temporal fenestrae, the postorbital-squamosal bar, and the squamosal and quadratojugal are all more slender than in the other English Anisian rhynchosaurs. This is seen also in dorsal view of the skull roof, where the nasals, frontals, and parietals are narrower in R. articeps. Additionally, the jugal-postorbital contact is located more ventrally in the posterior margin of the orbit in R. articeps.
Stenaulorhynchus stockleyi from the Manda Formation of Tanzania, also possibly Anisian (or maybe younger; Ezcurra et al. 2017) in age, was larger, with a skull about 240 mm long and 160 mm wide (Huene 1938). The skull width ratio is 0.67, similar to Langeronyx brodiei. Stenaulorhynchus stockleyi differs from the Middle Triassic rhynchosaurs from England in having a frontal that is broader than long, an apomorphy of the genus (Benton 1990: 299). Further, the occipital condyle is located in a very anterior position in comparison with the quadrates, associated with strongly posteriorly projected pterygoids. The teeth on the maxilla are relatively smaller than in the English species, and there is a backward-pointing spine on the second sacral rib. Stenaulorhynchus stockleyi probably had a supratemporal located between parietal and squamosal, and visible in posterior view, shown as part of an unusual “forked” split of the medio-dorsal process of the squamosal (Huene 1938: pl. 1: 1).
Ezcurra et al. (2016) suggest that these named Anisian-aged rhynchosaurs did not possess a complete lower temporal bar, as traditionally reconstructed (e.g., Huene 1938; Benton 1990). Certainly, Huene (1938: 88–89) explored the possibility that the bar was missing in Stenaulorhynchus stockleyi, and yet reconstructed the process in plaster and in his drawings. Ezcurra et al. (2016) argue that the jugal terminated as a relatively anteriorly located smooth point, and there was no articulation with the quadratojugal, in Rhynchosaurus articeps and Fodonyx spenceri. This portion of the skull is absent on both sides in Bentonyx sidensis, so it too could have had a gap in the lower temporal arcade. The same is possible also with Langeronyx brodiei, for which only the anterior portion of the left-hand side of the skull is known. New specimens with undamaged lower temporal regions would resolve the question, as would CT scanning of some specimens of R. articeps (e.g., SHRBM G132/1982; SHRBM 3; NHMUK R1236; MANCH L7642). If these Middle Triassic rhynchosaurs showed an incomplete lower temporal bar, the structure then reformed itself in the Late Triassic Hyperodapedontinae.
The other phylogenetically important trait highlighted by Ezcurra et al. (2016) is the presence and absence of palatal teeth. They confirm the presence of numerous rows of palatal teeth in the early Anisian basal rhynchosaurs Howesia and Mesosuchus, and their absence in most of the later forms, including Bentonyx and Fodonyx. However, raised ridges on the pterygoid bearing teeth had been identified in R. articeps (Huxley 1887; Woodward 1907; Huene 1929) and in Stenaulorhynchus stockleyi (Huene 1938; Chatterjee 1974), but Benton (1990: 274) suggested such pterygoid teeth were absent in both taxa. Ezcurra et al. (2016: 9–10) agree as for R. articeps, but they identify small pterygoid teeth in S. strockleyi. CT scanning of appropriate specimens of both taxa could add certainty on these points.
Acknowledgements
We thank Cesar Schultz (Universidade Federal do Rio Grande do Sul, São Paulo, Brazil) and Martin Ezcurra (Museo Argentino de Ciencias Naturales “Bernardino Rivadavia”, Buenos Aires, Argentina) for their very helpful reviews of the MS. Funding to MJB from UK Natural Environment Research Council grant (NE/P013724/1).
References
Ambrose, K., Hough E., Smith, N.J.P. and Warrington, G. 2014. Lithostratigraphy of the Sherwood Sandstone Group of England, Wales and south-west Scotland. British Geological Survey Research Report RR/14/01: 1–50.
Benton, M.J. 1983. The Triassic reptile Hyperodapedon from Elgin: functional morphology and relationships. Philosophical Transactions of the Royal Society of London, Series B 302: 605–717. Crossref
Benton, M.J. 1990. The species of Rhynchosaurus, a rhynchosaur (Reptilia, Diapsida) from the Middle Triassic of England. Philosophical Transactions of the Royal Society of London, Series B 328: 213–306. Crossref
Benton, M.J., Tverdokhlebov, V.P., and Surkov, M.V. 2004. Ecosystem remodelling among vertebrates at the Permian–Triassic boundary in Russia. Nature 432: 97–100. Crossref
Benton, M.J., Warrington, G.R., Newell, A.J., and Spencer, P.S. 1994. A review of the British Middle Triassic tetrapod assemblages. In: N.C. Fraser and H.-D. Sues (eds.), In the Shadow of the Dinosaurs, 131–160. Cambridge University Press, Cambridge.
Chatterjee, S. 1974. A rhynchosaur from the Upper Triassic Maleri Formation of India. Philosophical Transactions of the Royal Society, London, Series B 267: 209–261. Crossref
Chen, Z.-Q. and Benton, M.J. 2012. The timing and pattern of biotic recovery following the end-Permian mass extinction. Nature Geoscience 5: 375–383. Crossref
Coram, R.A., Radley, J.D., and Benton, M.J. 2019. The Middle Triassic (Anisian) Otter Sandstone biota (Devon, UK): review, recent discoveries and ways ahead. Proceedings of the Geologists’ Association 130: 294–306. Crossref
Coram, R.A., Radley, J.D., and Benton, M.J. 2021. Triassic tragedy—a bone bed in the Otter Sandstone of East Devon, south-west England. Geology Today 37: 183–190. Crossref
Ezcurra, M.D. 2016. The phylogenetic relationships of basal archosauromorphs, with an emphasis on the systematics of proterosuchian archosauriforms. PeerJ 4: e1778. Crossref
Ezcurra, M.D., Fiorelli, L.E., Martinelli, A.G., Rocher, S., von Baczko, M.B., Ezpeleta, M., Tarboda, J.R.A., Hechenleitner, E.M., Trotteyn, M.J., and Desojo, J.B. 2017. Deep faunistic turnovers preceded the rise of dinosaurs in southwestern Pangaea. Nature Ecology & Evolution 1: 1477–1483. Crossref
Ezcurra, M.D., Montefeltro, F., and Butler, R.J. 2016. The early evolution of rhynchosaurs. Frontiers in Ecology and Evolution 3: 142. Crossref
Gentil, A.R. and Ezcurra, M.D. 2022. Skull osteology of the holotype of the rhynchosaur Hyperodapedon sanjuanensis (Sill, 1970) from the Upper Triassic Ischigualasto Formation of Argentina. Anatomical Record 305: 1168–1200. Crossref
Hone, D.W.E. and Benton, M.J. 2008. A new genus of rhynchosaur from the Middle Triassic of south-west England. Palaeontology 51: 95–115. Crossref
Hounslow, M.W. and McIntosh, G. 2003. Magnetostratigraphy of the Sherwood Sandstone Group (Lower and Middle Triassic), south Devon, UK: detailed correlation of the marine and non-marine Anisian. Palaeogeography, Palaeoclimatology, Palaeoecology 193: 325–348. Crossref
Huene, F. von 1929. Über Rhynchosaurier und andere Reptilien aus den Gondwana-Ablagerungen Südamerikas. Geologische und Paläontologische Abhandlunge, Neue Folge 17: 1–62.
Huene, F. von 1938. Stenaulorhynchus, ein Rhynchosauride der ostafrikanischen Obertrias. Nova Acta Leopoldina, Neue Folge 6: 80–121.
Huxley, T.H. 1887. Further observations upon Hyperodapedon gordoni. Quarterly Journal of the Geological Society 43: 675–694. Crossref
Langer, M.C., Montefeltro,
F.C., Hone, D.W.E., Whatley, R., and Schultz, C.L. 2010. On Fodonyx spenceri and a new rhynchosaur
from the Middle Triassic of Devon. Journal
of Vertebrate Paleontology 30: 1884–1888. Crossref
Murchison, R.I. and Strickland, H.E. 1840. On the Upper Formations of the New Red Sandstone System in Gloucestershire, Worcestershire, and Warwickshire. Transactions of the Geological Society 5: 331–348. Crossref
Owen, R. 1842. Description of an extinct lacertian reptile, Rhynchosaurus articeps Owen of which the bones and foot-prints characterize the Upper New Red Sandstone at Grinsill, near Shrewsbury. Transactions of the Cambridge Philosophical Society Series 2 7: 355–369.
Woodward, A.S. 1907. On Rhynchosaurus articeps (Owen). Report of the British Association for the Advancement of Science 1907: 293–299.
Acta Palaeontol. Pol. 68 (1): 53–62, 2023
https://doi.org/10.4202/app.01019.2022