
Morphological variations and geographic distribution of the rare Middle Jurassic ammonite Oecoptychius refractus
SREEPAT JAIN, MARIUSZ A. SALAMON, GÜNTER SCHWEIGERT, and BARTOSZ J. PŁACHNO
Jain, S., Salamon, M.A., Schweigert, G., and Płachno, B.J. 2023. Morphological variations and geographic distribution of the rare Middle Jurassic ammonite Oecoptychius refractus. Acta Palaeontologica Polonica 68 (2): 321–335.
The rare Middle Jurassic ammonite Oecoptychius refractus is revisited based on material collected in southern Poland (Ogrodzieniec quarry) and France (St.-Laon near Loudun, western France). Based on available data and an evaluation of the literature, O. refractus ranges from the middle Callovian Kosmoceras jason Zone to the upper Callovian Quenstedtoceras lamberti Zone. Additionally, two specimens from Kachchh (western India) were re-evaluated and are now assigned to the lower part of the upper Callovian Peltoceras athleta Zone, similar to specimens from southern Germany. In the present study, O. refractus displays large morphological variation in the shape of the body-chamber, with a gradation from V- to U-shape. Additionally, the smaller upper Callovian specimens from Poland are morphologically closer to the lectotype (more evolute and compressed) and form a separate grouping as compared to the much larger middle Callovian specimens from France. Based on available data, the authors tentatively propose Phlycticeras polygonium var. waageni [M] as the dimorphic partner of O. refractus [m]; both dimorphs have similar morphology (ribbing pattern and striations), suture line and co-occur from middle to upper Callovian. Oecoptychius refractus maintains its morphological variability throughout the middle and upper Callovian, before its final dissapearance in the Q. lamberti Zone. Oecoptychius refractus is better documented from western Tethyan localities (Poland, Germany and France) as compared to those from the eastern Tethys (Madagascar and India). Recurrent sea level rises in the Middle Jurassic might be one of the plausible factors for its extensive palaeobiogeographic range.
Key words: Ammonoidea, Oecoptychius, morphological variation, Callovian, Jurassic, Tethys, Poland.
Sreepat Jain [sreepatjain@gmail.com; ORCID https://orcid.org/0000-0002-7679-9248 ], Adama Science and Technology University, Department of Applied Geology, School of Applied Natural Sciences, P.O. Box 1888, Adama, Ethiopia.
Mariusz A. Salamon [paleo.crinoids@poczta.fm; ORCID https://orcid.org/0000-0001-9399-2798 ], University of Silesia in Katowice, Faculty of Natural Sciences, Institute of Earth Sciences, ul. Będzińska 60, 41-200 Sosnowiec, Poland;
Günter Schweigert [guenter.schweigert@smns-bw.de; ORCID https://orcid.org/0000-0003-3798-8609 ], State Museum of Natural History Stuttgart, Rosenstein 1, 70191 Stuttgart, Germany.
Bartosz J. Płachno [bartosz.plachno@uj.edu.pl; ORCID https://orcid.org/0000-0001-5579-5101 ], Jagiellonian University in Kraków, Faculty of Biology, Institute of Botany, ul. Gronostajowa 9, 30-387 Kraków, Poland.
Received 27 September 2022, accepted 6 February 2023, available online 17 May 2023.
Copyright © 2023 S. Jain et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
The species of the Middle Jurassic ammonite genus Oecoptychius Neumayr, 1878, are quite rare forms spanning from the upper Bajocian Garantiana garantiana Zone up to the basal part of the upper Callovian Peltoceras athleta Zone (Schweigert and Dietze 1998; Schweigert et al. 2003). However, their occurrences in the P. athleta Zone are excessively rare with only two records from the basal part of the P. athleta Zone; one from Sengenthal in the eastern Franconian Alb, southern Germany (Callomon et al. 1987: pl. 3: 7) and the other one from Kachchh, western India (Spath 1928: 278, pl. 81: 5). To date there are no published records younger than the upper Callovian Hecticoceras trezeense Subzone in the basal part of the P. athleta Zone (Schweigert and Dietze 1998).
Here we document the first Oecoptychius refractus (Reinecke, 1818) from southern Poland found in a condensed bed (upper Callovian–lower Oxfordian) of the Ogrodzieniec quarry (Fig. 1A‒D) (see also Jain et al. 2022a, b). The paper also re-describes the morphology of O. refractus in detail and the morphological variation noted within this species based on specimens from the condensed bed of the Ogrodzieniec quarry and from the middle Callovian Kosmoceras jason Zone of western France. Other occurrences from southern Germany as well as from Madagascar and western India are also taken into account. The present paper also re-evaluates the age of its occurrence in Kachchh (western India).
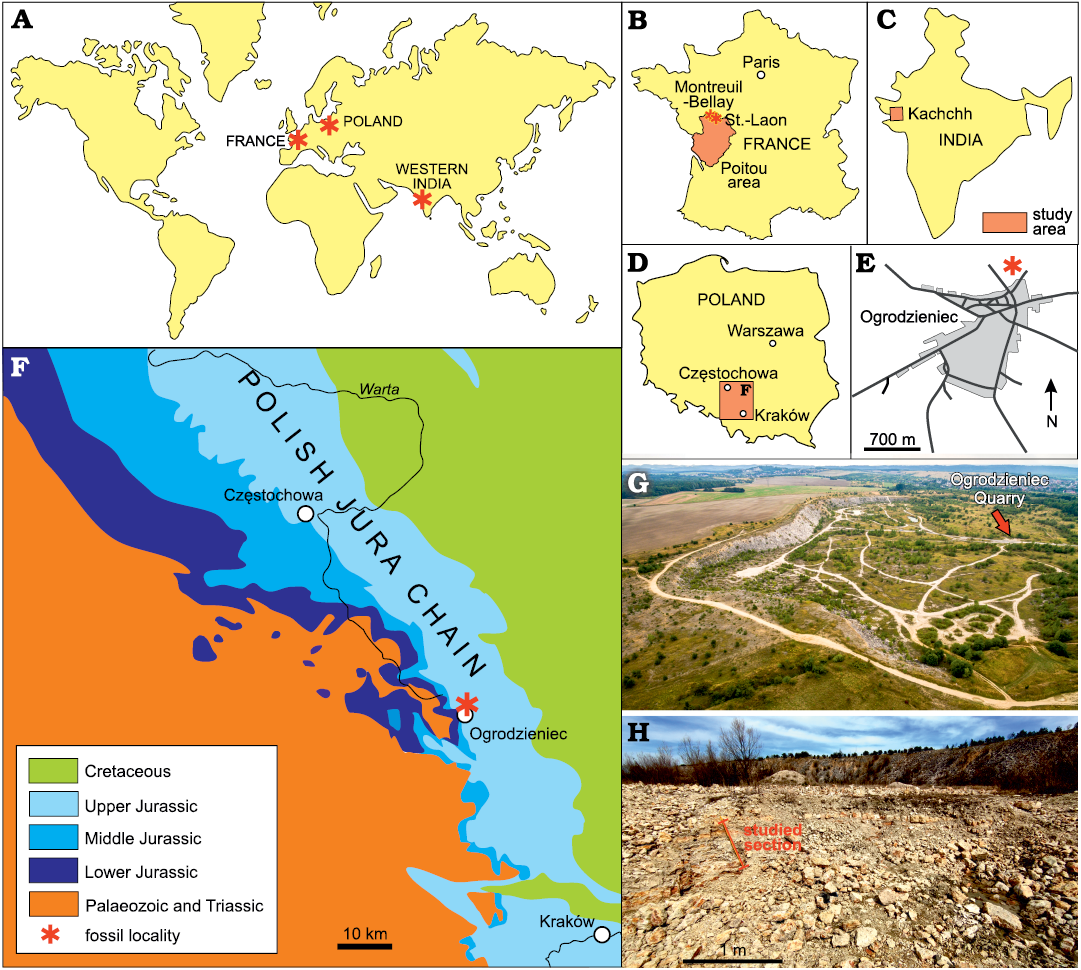
Fig. 1. Geological map and the abandoned Ogrodzieniec quarry. A. Map showing the studied localities in Poland, western France, and Kachchh (western India). B. Map of France showing the position of St.-Laon and Montreuil-Bellay. C. Map of India showing the position of Kachchh. D. Map of Poland showing the the position of Ogrodzieniec. E. Detailed location of Ogrodzieniec quarry (modified after Jain and Mazur 2021). F. Geological map of the Polish Jura Chain showing the location of the studied Ogrodzieniec quarry. G. Aerial view of the Ogrodzieniec quarry; arrow marks the location of the studied section. H. Exposure of the studied section.
Oecoptychius refractus is the only species of Oecoptychius recorded from the upper Callovian above which the genus dissapears from the fossil record (Schweigert and Dietze 1998). Thus, understanding the morphological changes of O. refractus from the middle to upper Callovian is of interest as it may provide some better understanding of the morphological changes within the genus during its demise.
Institutional abbreviations.—BSPM, Bayerische Staatssammlung für Paläontologie, München, Germany; GIUS, Institute of Earth Sciences of the University of Silesia in Katowice, Poland; NMC, Naturkundemuseum Coburg, Germany; SMNS, Staatliches Museum für Naturkunde Stuttgart, Germany.
Other abbreviations.—D, diameter of the shell; H, height of the whorl; [m], microconch; [M], macroconch; T, thickness of the whorl; U, umbilicus.
Geological setting
Ogrodzieniec quarry, Poland.—The Ogrodzieniec quarry displays a section with a stratigraphic condensation in glauconitic marl and glauconitic limestone beds ranging the upper Callovian to lower Oxfordian (see also Barski et al. 2004; Dembicz et al. 2006; see Fig. 2).

Fig. 2. Lithostratigraphy and biostratigraphy at the Ogrodzieniec quarry (modified after Dembicz et al. 2006). Arrow shows the occurrence of Oecoptychius refractus (Reinecke, 1818) within the general stratigraphy of the Ogrodzieniec quarry. Bed numbers (1, 1a, 2, 3, 18, and 37) after Dembicz et al. (2006).
The present study is based on new collections from this locality, from the condensed glauconitic nodular oolitic limestone bed β of Głowniak (2012) (Fig. 1). Associated ammonites indicate Quenstedtoceras lamberti Subzone (Q. lamberti Zone; upper Callovian) to Cardioceras bukowskii Subzone (Cardioceras cordatum Zone; lower Oxfordian), and include Peltoceras schroederi, Hibolithes hastatus (a large specimen attached to P. schroederi), Quenstedtoceras lamberti, Kosmoceras duncani, Peltoceratoides (Peltoceratoides) williamsoni, Peltoceratoides (Parawedekindia) arduennensis, Euaspidoceras babeanum, Bukowskites minax, Hecticoceras (Putealiceras) punctatum, and Euaspidoceras subbabeanum sensu Jeannet (1951) and several fragmentary specimens of Cardioceras sp. (Fig. 3) (see also Jain et al. 2022a, b).
Jain and Mazur (2021), recorded the dimorphic pairs of Rollierites biplicatum from the upper part of bed “a” and Kosmoceras rotundum in its lower part (bed β of Głowniak 2012). They assigned theirbed “a” to the Quenstedtoceras lamberti Zone (upper Callovian; Fig. 3A).

Fig. 3. Biostratigraphy of the studied section. A. Geological column, correlation of studied beds with these of Głowniak (2012) and the occurrence of specimens noted in the present study (modified after Jain and Salamon 2023). B. Field photo of the condensed bed where the studied specimens of Oecoptychius refractus (Reinecke, 1818) [m] were recorded. C. Close-up of the block with Quenstedtoceras lamberti (Q) and Hibolithes hastatus (H).
Later, Jain et al. (2022a, b) recorded the presence of Peltoceras schroederi with Kosmoceras rotundum and K. duncani from the condensed bed between bed α and β within bed “a” of Głowniak (2012). This condensed bed also yielded lower Oxfordian C. bukowskii Subzone, C. cordatum Zone (lower Oxfordian) forms such as Euaspidoceras babeanum, Bukowskites minax, Peltoceratoides (Peltoceratoides) arduennensis, P. (P.) williamsoni, and Cardioceras sp. Based on the strength of P. schroederi with Kosmoceras rotundum and K. duncani from the lower part of bed “a”, Jain et al. (2022a, b) assigned bed “a” to the Peltoceras athleta Subzone (Peltoceras athleta Zone, upper Callovian). However, both P. schroederi and K. duncani have been documented to occur within the above Quenstedtoceras lamberti Zone of the upper Callovian as well (Bonnot 1995; Courville and Bonnot 1998; Bonnot and Cariou 1999; Fortwengler et al. 1997; Lebrun and Courville 2013; Pellenard et al. 2014).
Hence, based on new field data and published ammonite records, we concur with Jain and Mazur (2021) in the assignment of the lower part of bed “a” exposed in the Ogrodzieniec quarry (i.e., bed β of Głowniak 2012) to the Quenstedtoceras lamberti Subzone, Q. lamberti Zone of the upper Callovian (Fig. 3).
Głowniak (2012) noted that in the Ogrodzieniec section, the contact between the upper Callovian and the lower Oxfordian runs within bed α (see Figs. 1, 3). Bed α is equivalent to bed 16 of Dembicz et al. (2006) which yielded reworked ammonites of both the upper Callovian Quenstedtoceras lamberti Zone and the lower Oxfordian Quenstedtoceras mariae Zone, i.e., Quenstedtoceras lamberti and Quenstedtoceras mariae, respectively. Głowniak (2012) assigned the basal sediments (bed γ) exposed in the Ogrodzieniec quarry to the Peltoceras athleta Zone but provided no information about the faunal content (Fig. 3). In this regard, current study could assume that the occurrence of Oecoptychius refractus in the Ogrodzieniec quarry (Fig. 3) could be the highest occurrence of Oecoptychius as this genus does not range above the Hecticoceras trezeense Subzone, Peltoceras athleta Zone, upper Callovian (see Schweigert and Dietze 1998). However, the ammonite assemblage occuring with O. refractus within the bed “a” in Ogrodzieniec quarry is overwhelmingly in favour of the Q. lamberti Subzone, Q. lamberti Zone, upper Callovian (Fig. 3). This could mean that O. refractus occurrence in the Ogrodzieniec quarry is stratigraphically the highest worldwide. Alternatively, it could mean faunal elements within cendensed horizon between beds β and α indicate also Peltoceras athleta Zone, upper Callovian as well. More sampling across the Ogrodzieniec quarry is required to answer this question. For now, the range of O. refractus is extended well into the Q. lamberti Zone of the upper Callovian.
Poitou area, western France.—The Callovian succession Poitou area consists of a thin ferruginous oolite beds which are very rich in ammonites and other invertebrates. At St.-Laon near Loudun (see Fig. 1B), oolitic limestone beds of the middle Callovian Kosmoceras jason and Erymnoceras coronatum zones have been especially well sampled. Specimens of O. refractus, as well as their dimorphic counterpart Phlycticeras spp. are rather common there and often preserved as inner moulds, sometimes even with impression of the lappeted aperture (Schweigert and Dietze 1998). The sample of O. refractus studied herein originates from the Kosmoceras jason Zone from St.-Laon near Loudun (see Fig. 1B). Oecoptychius refractus is present in the basal horizon of the Peltoceras athleta Zone, upper Callovian, which is best developed in the vicinity of Montreuil-Bellay (see Fig. 1B), but has become excessively rare (Schweigert et al. 2003; Bonnot et al. 2005). In higher levels of the P. athleta Zone, upper Callovian, Oecoptychius and Phlycticeras apparently disappear.
Kachchh, western India.—Spath (1928: 278, pl. 81: 5a‒c) recorded two specimens of Oecoptychius refractus [m](V-shaped body-chamber) with Phlycticeras waageni [M] (Phlycticeras polygonium var. waageni of Jain 2017: fig. 13f‒n) (see also Waagen 1875: 40, pl. 9: 2a‒c; Spath 1928: 91). The specimens were collected from the bed no. 2 of Raj Nath (1942), a characteristic marker bed within the Kachchh Basin that has also yielded Peltoceras athleta (see also Jain and Pandey 2000; Fürsich et al. 2001; Jain 1997, 2017). Beds just above this bed no. 2 have yielded the characteristic form of Collotia fraasi of the Collotia collotiformis Zone (see also Jain and Pandey 2000; Jain 2017), thus, limiting the stratigraphic occurrences of Phlycticeras polygonium var. waageni [M]‒Oecoptychius refractus [m] to the Hecticoceras trezeense Subzone, lowermost Peltoceras athleta Zone, upper Callovian, in Kachchh, as also suggested by Schweigert and Dietze (1998) from Europe.
Material and methods
A total of 29 specimens of Oecoptychius refractus were studied (Figs. 4–10; see Table 1); 12 specimens from the Ogrodzieniec quarry (MAS collection, GIUS 8-3657/1‒12; Figs. 4–6) housed at the University of Silesia in Katowice, Faculty of Natural Sciences, Institute of Earth Sciences, Sosnowiec, Poland, and 17 specimens from St.-Laon near Loudun, Poitou area, France (Rolf Mitschelen collection, SMNS 70655/1–17; Fig. 7B) housed at the State Museum of Natural History, Stuttgart, Germany.
Table 1. Dimensions of Oecoptychius refractus (Reinecke, 1818), specimens considered in the present study. Abbreviations: D, diameter of the shell; H, height of the whorl; T, thickness of the whorl; U, umbilicus; U/D, coiling ratio (umbilicus/diameter); T/H, relative whorl thickness (thickness of whorl/height of whorl).
|
Specimen no. |
D |
H |
T |
U |
U/D |
T/H |
Illustration (if any) |
|
Poland (upper Callovian) |
|||||||
|
GIUS 8-3657/1 |
20.6 |
6.9 |
9.8 |
0.60 |
0.03 |
1.42 |
Fig. 4A1‒A4 |
|
GIUS 8-3657/6 |
11.3 |
5.6 |
7.2 |
0.90 |
0.08 |
1.27 |
Fig. 4B1‒B3 |
|
GIUS 8-3657/7 |
18.7 |
6.2 |
8.7 |
0.94 |
0.05 |
1.42 |
Fig. 4C1‒C4 |
|
GIUS 8-3657/8 |
14.1 |
4.9 |
7.1 |
0.60 |
0.04 |
1.46 |
Fig. 4D1‒D4 |
|
GIUS 8-3657/9 |
12.7 |
5.0 |
7.8 |
0.50 |
0.04 |
1.55 |
Fig. 5A1‒A4 |
|
GIUS 8-3657/10 |
15.7 |
– |
– |
– |
– |
– |
Fig. 5B1, B2 |
|
GIUS 8-3657/11 |
15.9 |
5.6 |
8.6 |
– |
– |
1.54 |
Fig. 5C1‒C4 |
|
GIUS 8-3657/2 |
15.5 |
6.8 |
9.3 |
0.80 |
0.05 |
1.38 |
Fig. 5D1‒D4 |
|
GIUS 8-3657/12 |
12.5 |
6.5 |
8.0 |
– |
– |
1.23 |
Fig. 6A1‒A3 |
|
GIUS 8-3657/5 |
11.5 |
5.5 |
7.2 |
0.60 |
0.05 |
1.31 |
Fig. 6B1‒B3 |
|
GIUS 8-3657/3 |
13.0 |
5.1 |
9.8 |
0.40 |
0.03 |
1.93 |
Fig. 6C1‒C3 |
|
GIUS 8-3657/4 |
15.1 |
5.6 |
8.1 |
0.70 |
0.05 |
1.44 |
Fig. 6D1‒D3 |
|
France (middle Callovian) (after Schweigert and Dietze 1998) |
|||||||
|
Lectotype, NMC 52/1 |
12.1 |
6.8 |
8.8 |
0.50 |
0.04 |
1.29 |
Schweigert and Dietze 1998: pl. 9: 10 |
|
SMNS 63459/1 |
22.5 |
7.0 |
9.0 |
0.50 |
0.02 |
1.29 |
Schweigert and Dietze 1998: pl. 9: 4 |
|
SMNS 63458/1 |
21.2 |
6.0 |
9.0 |
– |
– |
1.50 |
Schweigert and Dietze 1998: pl. 9: 6 |
|
SMNS 63460/1 |
20.5 |
5.7 |
8.5 |
– |
– |
1.49 |
Schweigert and Dietze 1998: pl. 9: 7 |
|
SMNS 63457/1 |
16.8 |
6.2 |
8.0 |
0.30 |
0.02 |
1.29 |
Schweigert and Dietze 1998: pl. 9: 8 |
|
BSPM 1989 J 37 |
23.0 |
6.0 |
9.3 |
0.20 |
0.01 |
1.55 |
Schweigert and Dietze 1998: pl. 9: 11 |
|
SMNS 63458/2 |
21.0 |
7.3 |
11.0 |
0.50 |
0.02 |
1.51 |
– |
|
SMNS 63460/2 |
20.3 |
6.3 |
8.5 |
0.50 |
0.02 |
1.35 |
– |
|
SMNS 63460/3 |
17.5 |
6.3 |
8.2 |
0.50 |
0.03 |
1.30 |
– |
|
SMNS 63460/4 |
17.0 |
6.7 |
9.8 |
– |
– |
1.46 |
– |
|
SMNS 63457/2 |
16.0 |
7.0 |
8.3 |
– |
– |
1.19 |
– |
The Kachchh (western India) specimens of Phlycticeras polygonium var. waageni [M] (see Jain 2017: fig. 13l‒n, f‒h; refigured here as Fig. 11B1‒B4, G, H, respectively) are cataloged as SJ/JA/99-00/23 and Ju/B44/1, respectively, and kept at the Department of Geology, Rajasthan University (Rajasthan, India).
All measurements (Table 1) were taken using a caliper from the specimens and photographed using Canon D530 camera. Images were prepared using Adobe Photoshop CS6 and CorelDRAW version 21.

Fig. 4. Strigoceratid ammonoid Oecoptychius refractus (Reinecke, 1818) [m] from Ogrodzieniec quarry, Quenstedtoceras lamberti Zone, upper Callovian. A. GIUS 8-3657/1 in lateral (A1), opposite lateral (A2), ventral (A3), and dorsal (A4) views. B. GIUS 8-3657/6 in lateral (B1) and ventral (B2) views. C. GIUS 8-3657/7 in lateral view (C1), detail showing the smooth umbilical region (C2), opposite lateral (C3) and dorsal (C4) views. D. GIUS 8-3657/8 in lateral (D1), opposite lateral (D2), ventral (D3), and dorsal (D4) views.
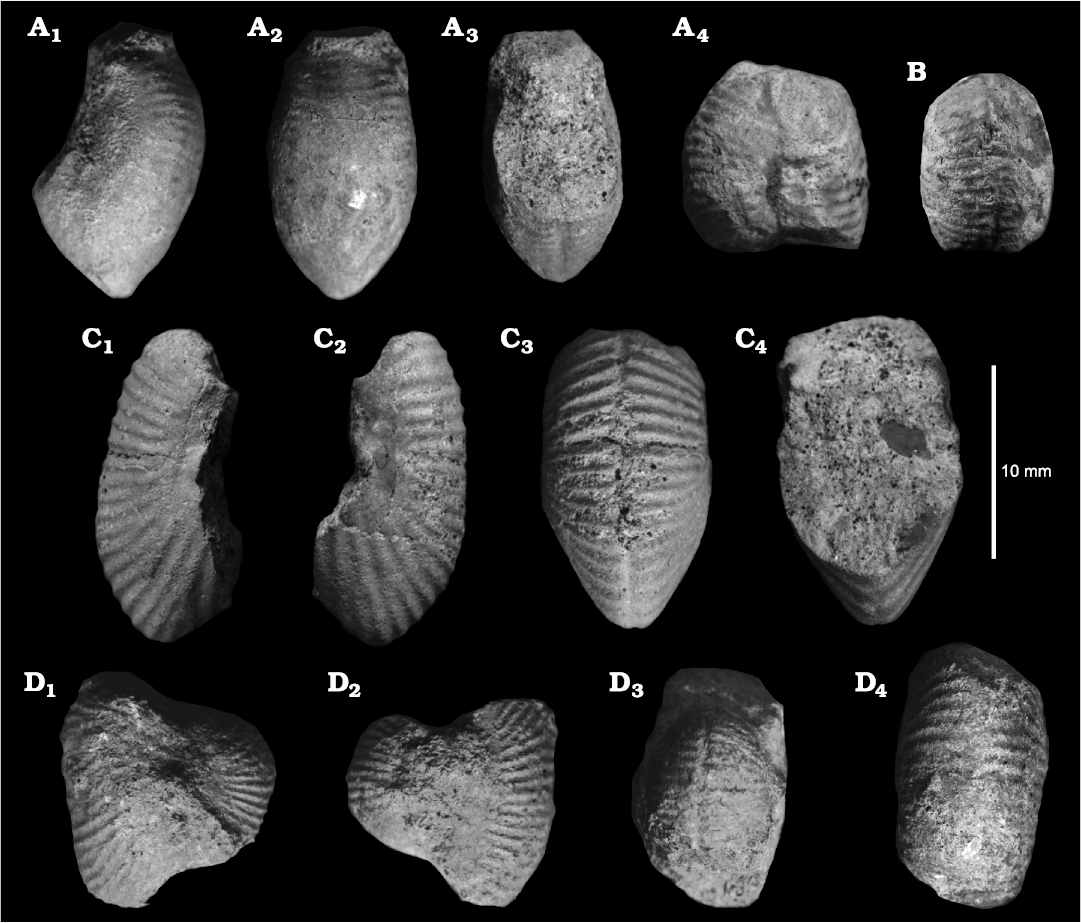
Fig. 5. Strigoceratid ammonoid Oecoptychius refractus (Reinecke, 1818) [m] from Ogrodzieniec quarry, Quenstedtoceras lamberti Zone, upper Callovian. A. GIUS 8-3657/9 in lateral (A1), dorsal (A2), ventral (A3), and dorsal (A4, note the ribbing pattern) views. B. GIUS 8-3657/10 in ventral view. C. GIUS 8-3657/11 in lateral (C1), opposite lateral (C2), ventral (C3), and dorsal (C4) views. D. GIUS 8-3657/2 in lateral (D1), opposite lateral (D2), ventral (D3), and dorsal (D4) views.

Fig. 6. Strigoceratid ammonoid Oecoptychius refractus (Reinecke, 1818) [m] from Ogrodzieniec quarry, Quenstedtoceras lamberti Zone, upper Callovian. A. GIUS 8-3657/12 in lateral (A1), dorsal (A2), and dorsal (A3, note the ribbing pattern) views. B. GIUS 8-3657/5 in lateral (B1) and ventral (B2, B3, note the ribbing pattern) views. C. GIUS 8-3657/3 11 in lateral (C1), opposite lateral (C2), and ventral (C3) views. D. GIUS 8-3657/4 in lateral (D1), opposite lateral (D2), and ventral (D3, D4, note the ribbing pattern) views.
Systematic palaeontology
Family Strigoceratidae Buckman, 1924
Subfamily Phlycticeratinae Spath, 1925
Genus Oecoptychius Neumayr, 1878
Type species: Nautilus refractus Reinecke, 1818, middle Callovian, Erymnoceras coronatum Zone, eastern Franconian Alb (Trockau, Germany).
Oecoptychius refractus (Reinecke, 1818)
Figs. 4–10.
1818 Nautilus refractus; Reinecke 1818: fig. 27‒30.
1830 Ammonites refractus Reinecke; Zieten 1830: 14, pl. 10: 9.
1846 Ammonites refractus Reinecke; Quenstedt 1846: 150, pl. 11: 12.
1857 Ammonites refractus; Quenstedt 1857: 524, pl. 69: 25, 26, 28‒30.
1878 Oecoptychius refractus; Neumayr 1878: 73.
1887 Ammonites refractus; Quenstedt 1887: 762f, pl. 86: 37‒50.
1887 Oecoptychius refractus Quenstedt; Zakrzewski 1887: 122, pl. 1: 3.
1897 Oecoptychius refractus Haan; Glangeaud 1897: 104, pl. 3: 7‒10.
1908 Ammonites (Oecoptychius) refractus Reinecke sp.; Engel 1908: pl. 4: 6.
1924 Oecoptychius refractus Haan; Roman and Debrun 1924: 106, pl. 11: 5.
1928 Oecoptychius refractus Haan; Sayn and Roman 1928: 150, pl. 10: 12.
1928 Oecoptychius refractus (Reinecke); Spath 1928: 278, pl. 81: 5.
1932 Oecoptychius refractus Reinecke; Corroy 1932: 120, pl. 10: 10‒11.
1952 Oecoptychius refractus (Haan); Makowski 1952: 31, pl. 3: 3.
1958 Oecoptychius refractus Reinecke; Collignon 1958: 28, fig. 115.
1985 Oecoptychius refractus (Reinecke); Schlegelmilch 1985: 137, pl. 54: 4.
1987 Oecoptychius refractus (Reinecke); Callomon et al. 1987: 26, pl. 3: 7.
1994 Oecoptychius refractus (Reinecke); Fischer 1994: 153, pl. 60: 2.
1998 Oecoptychius refractus (Reinecke); Schweigert and Dietze 1998: pls. 6: 5, 8: 5, 9: 4‒11.
Type material: Lectotype: NMC 52/1, original of Reinecke (1818: fig. 29), designated by Zeiss in Heller and Zeiss (1972). Paralectotype: NMC 52/2, original of Reinecke (1818: fig. 27) (see Schweigert and Dietze 1998).
Type locality: Probably eastern Franconian Alb (area of Trockau) (for further details, see Schweigert and Dietze 1998).
Type horizon: Erymnoceras coronatum Zone, middle Callovian, Middle Jurassic.
Measurements.—See Table 1.
Description.—The inner and the early part of the middle whorls not visible in any of the studied specimens. The middle and outer whorls are characterized by: (i) primaries long and sharp, bifurcate at mid- to outer third of whorl height into two forwardly inclined equally sharp secondaries, (ii) straight primaries arising from the rounded umbilical margin, (iii) with increasing shell diameter, the rounded umbilical region developing a smooth area; hence, with secondaries only visible at the outer third of whorl height, (iv) a distinct kink in the body-chamber, i.e., at larger shell sizes (on the phragmocone and in front of the body-chamber kink, in the case of a mold), a ventral groove (furrow) is noted, (v) morphological variation in the shape of the body-chamber with agradation from V- to U-shape in longitudinal section (Fig. 7).
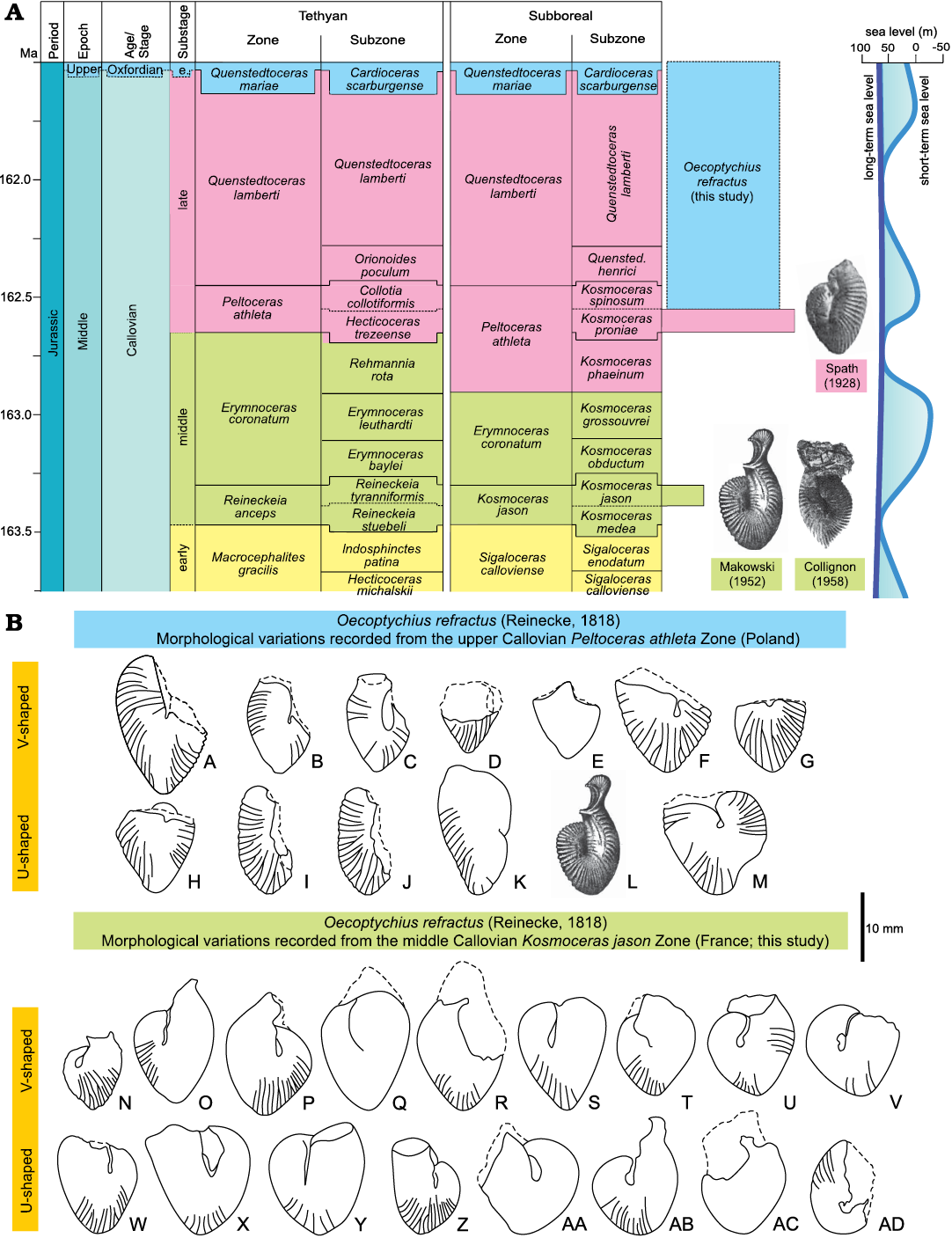
Fig. 7. Biozonation and morphological variation in Oecoptychius refractus (Reinecke, 1818) [m]. A. Biozonation used in the present study with the occurrence of O. refractus (Reinecke, 1818) noted in Kachchh (western India; Spath 1928), Poland (Makowski 1952), and Madagascar (Collignon 1958). B. Morphological variation in O. refractus (Reinecke, 1818) [m]. A, GIUS 8-3657/9; B, GIUS 8-3657/9; C, GIUS 8-3657/8; D, GIUS 8-3657/6; E, GIUS 8-3657/12; F, GIUS 8-3657/4; G, GIUS 8-3657/5; H, GIUS 8-3657/3; I, GIUS 8-3657/11; J, GIUS 8-3657/10; K, GIUS 8-3657/7; L, specimen of Makowski (1952: 31, pl. 3: 3; no mention of specimen number); M, GIUS 8-3657/2; N to AD, SMNS 70655/1–17, respectively.
In specimens preserved with the shell especially in the area of the body-chamber, and rarely on the phragmocone, a strigoceratid spiral striation is present (see Schweigert and Dietze 1998: pls. 6: 5; 9: 8, 9). A specimen with remains of the shell with striation, from the Ornatenton Formation (Kosmoceras jason Zone) of Neidlingen, SW Germany is illustrated (Fig. 8). Even if the shell is preserved, it often remains in the surrounding rock matrix and only the molds lacking the shell are collected. On molds, the spiral striae are occasionally discernible in large specimens; none of the present specimens (Dmax = 21 mm) preserve this feature. In rare cases when the shell is preserved, a smooth band is also observed which, like the furrow, dissapears at the end of the body-chamber.
The upper Callovian specimens from Poland are more similar to the lectotype by means of the dimension parameters (more evolute and compressed; average U/D = 0.05; T/H = 1.44, respectively) and form a separate grouping when compared to the much larger middle Callovian French specimens (Fig. 9, Table 1). The whorl thickness versus coiling ratio also displays separate grouping (Fig. 9D, Table 1).

Fig. 8. Strigoceratid ammonoid Oecoptychius refractus (Reinecke, 1818) [m], SMNS 63457/1, from Ornatenton Formation, Kosmoceras jason Zone, Neidlingen, SW Germany, middle Callovian, Middle Jurassic. Note the strigations, a character noted only in well-preserved specimens and reminiscent of species attributed to Phlycticeras polygonium (Zieten, 1831) var. waageni [M] and considered herein to represent macroconch of O. refractus (Reinecke, 1818).
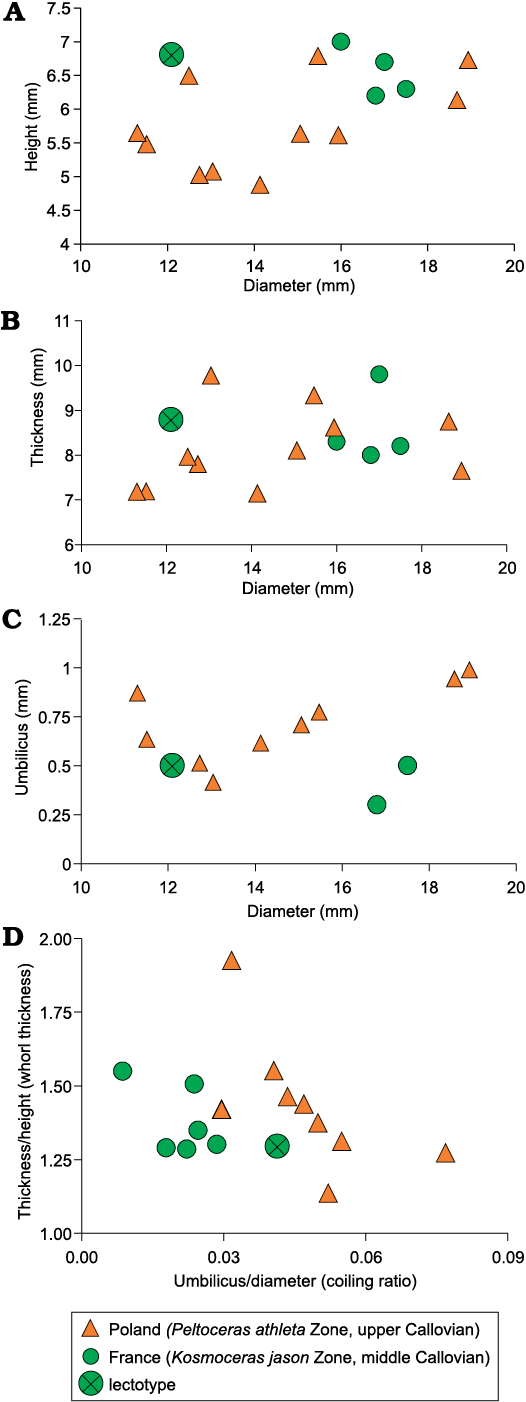
Fig. 9. Dimensional proportions of Oecoptychius refractus (Reinecke, 1818) [m]. The French and Polish specimens are compared with a reference point of the lectotype. A. Whorl height vs. shell diameter. B. Whorl thickness vs. shell diameter. C. Umbilicus vs. shell diameter. D. Whorl thickness vs. coiling ratio. For all given dimensional proportions, the French and Polish specimens form different groups.
Remarks.—Oecoptychius grossouvrei Brasil, 1894 (Parkinsoni Zone, upper Bajocian, Middle Jurassic; see also Schweigert and Dietze 1998: 22, pl. 1: 3) is morphologically close to Oecoptychius refractus (Reinecke, 1818) described herein, but lacks the distinctive body-chamber kink, has a somewhat oval-shaped body-chamber and finer secondaries restricted to the outer third of the whorl height.
Oecoptychius subrefractus (Buckman, 1920) (Morphoceras macrescens Subzone, Zigzagiceras zigzag Zone, lower Bathonian, Middle Jurassic; see also Dietl 1986; Dietze et al. 1997; Schweigert and Dietze 1998) has a similar but somewhat less developed body-chamber kink (Schweigert and Dietze 1998: 22, pl. 1: 3); it is also relatively compressed and evolute (Fig. 9, Table 1) and with a finer ornamentation.
Oecoptychius globosus Schweigert and Dietze, 1998 (Proplanulites koenigi Zone, lower Callovian; original of Quenstedt 1886: pl. 86: 51) is a very large form (27 mm shell diameter, Couffon 1919; see Table 1) but resembles O. refractus in having the mid-ventral furrow, primaries splitting into two ribs and the smooth umbilical region but lacks the characteristic and pronounced body-chamber kink; the mid-ventral furrow is less developed in O. globosus (see Schweigert and Dietze 1998: pl. 4: 6‒7).
In terms of dimensional proportions (whorl thickness vs. coiling ratio; Fig. 10), the O. grossouvrei (Parkinsonia parkinsoni Zone, upper Bajocian) falls within the O. refractus range, whereas both the O. subrefractus (Zigzagiceras zigzag Zone, lower Bathonian) and O. globosus (P. koenigi Zone, lower Callovian) are outliers (Fig. 10) and morphologically quite different (Schweigert and Dietze 1998).
Stratigraphic and geographic range.—Kosmoceras jason Zone (middle Callovian) to Quenstedtoceras lamberti Zone (upper Callovian); Poland (middle and upper Callovian; the latter is from this study), Germany (middle Callovian), France (middle and upper Callovian), Madagascar (middle Callovian), Kachchh (upper Callovian).
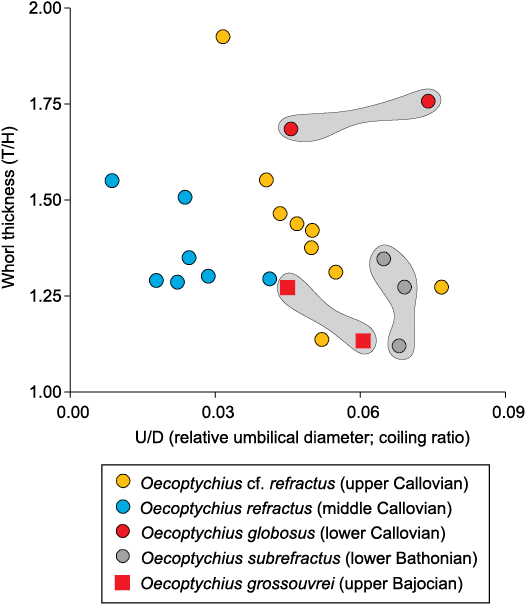
Fig. 10. Comparative dimensional proportions of various species of Oecoptychius Neumayr, 1878. Shaded areas display the range of variation in dimensional proportions.
Discussion
Morphological variation.—Schweigert and Dietze (1998), based largely on their data from southern Germany and France, noted that Oecoptychius refractus displays (i) a range of variation of adult size at the stage-level, and (ii) that the average shell size is reduced from the middle (Kosmoceras jason and Erymnoceras coronatum zones) to the upper Callovian (basal part of the Peltoceras athleta Zone). In the present study, based on a more comparative look at the middle and upper Callovian specimens (Fig. 7), three characteristic features of O. refractus are noted: (i) the changes in the body-chamber are gradiational between V- to U-shape, (ii) there is no reduction in morphological variation (relative abundance of V- to U-shaped body-chambers) over time from middle to late Callovian, although forms with the V-shaped body-chamber do slightly dominate (53‒54%), and (iii) there is indeed a general reduction in size (shell diameter) from the larger middle Callovian (average: 18.9 mm) to the smaller late Callovian (average: 14.6 mm) forms.
This illustrated morphological variation within the specimens of O. refractus from different localities and from different time intervals (Fig. 9) has a bearing on the widely applicable concept of morphology-based species assignments and reliability of these species and their varieties where there are no gradational forms. Here, we document a series of gradational forms of O. refractus that remains the same through time until its demise in the late Callovian (Fig. 9).
Dimorphic pairing.—Schweigert and Dietze (1998: 31, fig. 2) indicated the following dimorphic pairing: Oecoptychius cf. refractus [m]‒Phlycticeras polygonium [M], Oecoptychius refractus [m]‒Phlycticeras pustulatum [M] and Oecoptychius cf. refractus [m]‒Phlycticeras waageni [M] (see also Schweigert et al. 2003). Here, based on the variation in O. refractus (Fig. 7), O. refractus, and O. cf. refractus of Schweigert and Dietze (1998) are considered as O. refractus.
Both in southern Germany and in western France, O. refractus [m] occurs in the beds of the Kosmoceras jason Zone (middle Callovian, Middle Jurassic), independent from different lithologies in both regions. In this zone, a highly variable Phlycticeras polygonum [M] is common, represented by almost smooth (var. laevigatum of Quenstedt 1887) to stronger ornamented variants (var. suevicum of Quenstedt 1887); most of them share a rather slender whorl section, but some of which (var. nodosum of Quenstedt 1887) are transitional to the stratigraphically younger and stronger ornamented P. pustulatum [M]. The co-occurrence and acme of Phlycticeras polygonium [M] and Oecoptychius refractus [m] in the K. jason Zone has contributed much to the recognition of the dimorphic pairing between these genera.
Spath (1928) noted the co-occurrence of Phlycticeras waageni [M] (P. polygonium var. waageni [M] of Jain 2017: fig. 13f‒n) (see Fig. 11B–H) and O. refractus [m]. The suture line given by Spath (1928: pl. 81: 5c) (see Fig. 11I) resembles that of P. polygonium var. waageni [M] as illustrated by Jain (2017), and also the general ribbing pattern and the striation (see Fig. 11C‒H). However, Schweigert and Dietze (1998: 22, fig. 1) considered the suture lines of their O. refractus [m]‒P. polygonium [M] pair closer to P. waageni (see Fig. 11J, K). Jain (1997: figs. 2, 3) recorded the presence of Phlycticeras gr. pustulatum [M] in the middle Callovian of Jumara Dome (Kachchh, western India) (Phlycticeras polygonium [Zieten, 1831] var. nodosum [Quenstedt, 1887] [M]; see also Schweigert and Dietze 1998: 16). Based on this and the presence of longitudinal striae in O. refractus (Fig. 8) these authors considered them as dimorphic pairs.
However, here, based on available data, Phlycticeras polygonium var. waageni [M] is considered as the dimorphic partner of O. refractus [m] (Fig. 11). This assignment is provisional, as the inner whorls of the var. waageni are not known, although, both dimorphs have similar stratigraphical ranges, from the base of the middle Callovian to the base of the upper Callovian (Fig. 11) (see also Jain 2017). These dimorphs have large differences in their shell diameters; the macroconch measures a maximum of 170 mm (Schweigert and Dietze 1998), whereas the microconch measures a maximum of 27 mm (Couffon 1919).
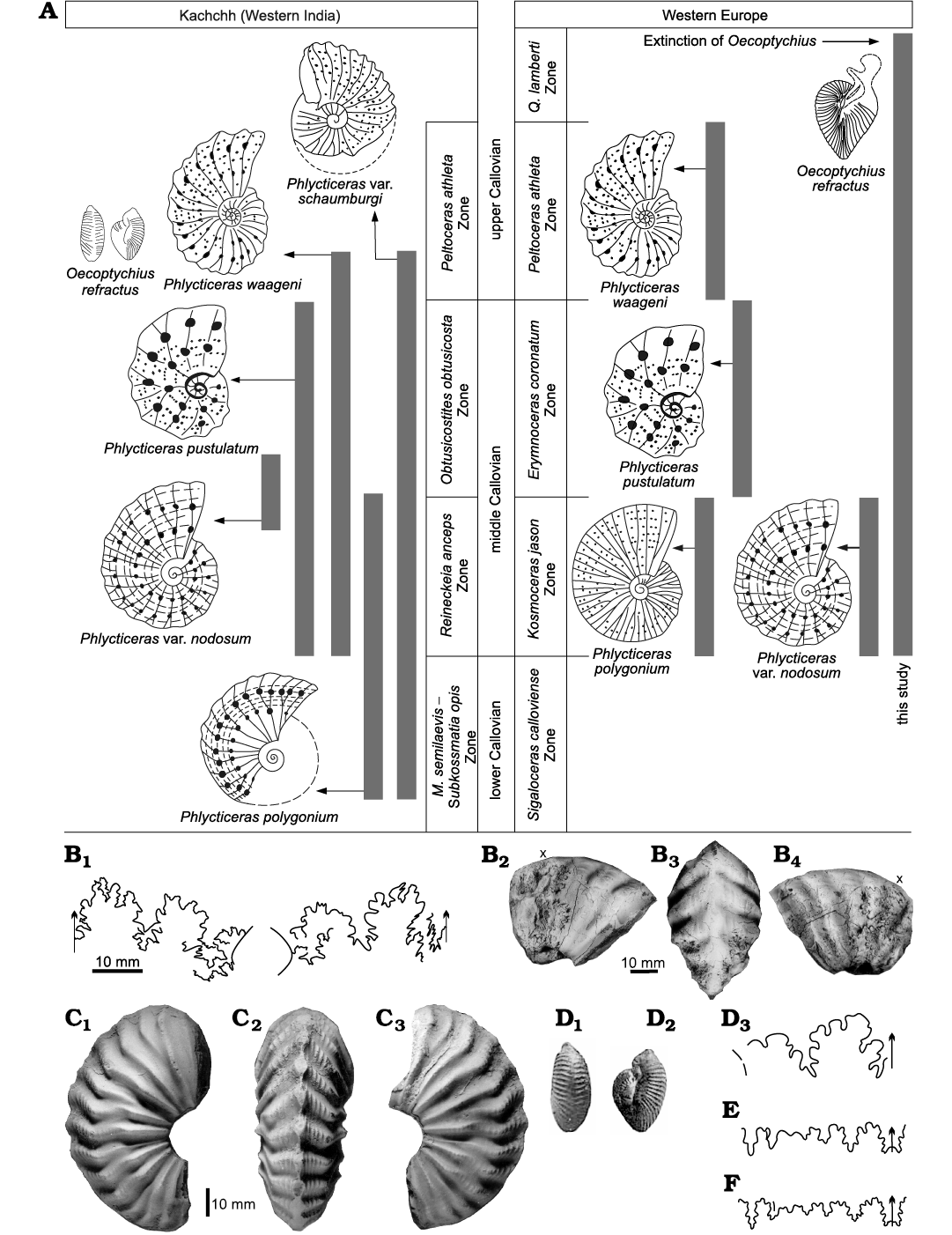
Fig. 11. Relationship of Phlycticeras [M] Hyatt, 1900, with Oecoptychius Neumayr, 1878 [m]. A. Middle–upper Callovian occurrences of species of Phlycticeras [M] with their corresponding microconch, Oecoptychius refractus (Reinecke, 1818). B, C. Phlycticeras polygonium var. waageni Buckman, 1914, from Jara, 10 km north of Jumara, Kachchh, western India, middle Callovian (after Jain 2017). B. SJ/JA/99-00/23, suture line (B1), lateral (B2), ventral (B3), and opposite lateral (B4) views. C. Ju/B44/1, lateral (C1), ventral (C2), and opposite lateral (C3) views. D, E. Oecoptychius refractus (Reinecke, 1818). D. G.S.I. No. 2/88 (Raj Nath collection) from Nurrah, Kachchh, western India, upper Callovian, ventral (D1) and lateral (D2) views, and suture line (after Spath 1928: 278, pl. 81: 5c) (D3). E. Suture line after Schweigert and Dietze 1998. F. Phlycticeras waageni Buckman, 1914, suture line after Schweigert and Dietze 1998: 22, fig. 1b. E, F, based on the similarity of suture line and similar strigations (see Fig. 8), Schweigert and Dietze (1998) considered Phlycticeras polygonium (Zieten, 1831), Phlycticeras pustulatum (Reinecke, 1818), and Phlycticeras waageni Buckman, 1914, as the macroconch of Oecoptychius refractus (Reinecke, 1818) [m]. However, in this study, based on similarly of suture lines (B1, D3), morphological characteristics (B2–B4, C), and stratigraphic occurrences of Oecoptychius refractus (Reinecke, 1818) and Phlycticeras polygonium var. waageni (= Phlycticeras waageni), are considered as a dimorphic pair. See text for further explanation. A, D–F, not to scale. Abbreviations: M., Macrocephalites; Q., Quenstedtoceras. Stratigraphic ranges for Kachchh (Western India) are after Jain (2017) and for Western Europe are after Schweigert and Dietze (1998); x marks the end of the phragmocone stage.
Palaeobiogeography.—Most records of Oecoptychius refractus (Reinecke, 1818) come from western Tethyan localities (Poland, Germany, and France) as compared to the eastern Tethyan ones (Madagascar and India). Possible reasons include (i) collecting bias due to much longer and more extensive sampling in France and Germany as compared to other areas where only random samples were picked up, (ii) collecting bias due to exposure availability in Europe, and (iii) an actual phenomenon explained by role of global eustasy; rising sea level (transgression) would result in more available niches (Fig. 7A; see also Haq 2018). Rising sea levels would have favored the formation of new niches and seaways to facilitate increased species distribution (Fig. 7A). However, only, more sampling can provide a conclusive answer.
Conclusions
Oecoptychius refractus (Reinecke, 1818) ranges from the Kosmoceras jason Zone (middle Callovian) to the Quenstedtoceras lamberti Zone (upper Callovian). The Q. lamberti Zone occurrence from southern Poland is probably the youngest record of this species worldwide. The two Kachchh (western India) specimens reported by Spath (1928) are assigned to the lower part of the upper Callovian Peltoceras athleta Zone; globally, this is now the second upper Callovian occurrence of O. refractus. Based on the present study, O. refractus is characterized by large morphological variation in the shape of the body-chamber, displaying a gradation from V- to U-shaped body-chamber. In terms of dimensional proportions, the smaller upper Callovian specimens from southern Poland are closer to the lectotype (more evolute and compressed) and also form a separate grouping as compared to the much larger middle Callovian specimens from western France. Additionally, Phlycticeras polygonium var. waageni [M] is considered here as the dimorphic partner of O. refractus [m]; both dimorphs have similar stratigraphic ranges, from the base of the middle Callovian to the base of the upper Callovian. The O. refractus samples are recorded dominantly from western Tethyan localities (Poland, Germany, and France) and less so from eastern Tethyan localities (Madagascar and India). However, more sampling is needed in the eastern Tethyan region to better understand the palaeobiogeographic distribution of O. refractus and its causal factors.
Authors’ contributions
SJ carried out the study, participated in the conceptualization and drafted the manuscript. GS participated in drafting and reviewing the manuscript. MAS carried out the fieldwork, geological data collection, participated in the fossil preparation and measurements. BJP carried out part of the fieldwork. All authors read and approved the final manuscript.
Acknowledgements
The authors thank Horacio Parent (Universidad Nacional de Rosario, Argentina) and a second anonymous reviewer, for their constructive comments and suggestions that greatly improved the manuscript. We also gratefully acknowledge Damian Kuźma (Sosnowiec, Poland) for his help during field work and to Wojciech Krawczyński (University of Silesia in Katowice, Sosnowiec, Poland) for photographs of the Polish specimens. MAS would like to thank the “Small Grants 2022” (Faculty of Natural Sciences, University of Silesia in Katowice, Poland) for financial support. This research was supported financially by the “Fast Track Grants 2.0 Back2Mobility” (University of Silesia in Katowice, Faculty of Natural Sciences, Sosnowiec, Poland) for SJ.
References
Barski, M., Dembicz, K., and Praszkier, T. 2004. Biostratigraphy and the Mid-Jurassic environment from the Ogrodzieniec quarry. Tomy Jurajskie 2: 61–68.
Bonnot, A. 1995. Les Aspidoceratidae (Ammonitina) en Europe occidentale au Callovien supérieur et a l’Oxfordien inférieur. 454 pp. Thése Université Dijon (unpublished), Dijon.
Bonnot, A. and Cariou, E. 1999. Réinterprétation de Peltoceratoides athletoides (Lahusen), 1883 (Ammonitina, Aspidoceratidae). Conséquences sur la biozonation du Callovien supérieur et de l’Oxfordien inférieur. Annales de Paleontologie 85: 155‒171. Crossref
Bonnot, A., Boursicot, P.-Y., and Ferchaud, P. 2005. Peltoceras marysae, une nouvelle espèce de Peltoceratinae (Ammonitina, Aspidoceratidae) dans lʼhorizon à Leckenbyi (Callovien supérieur, zone à Athleta) de Montreuil-Bellay (Maine-et-Loire, France). Revue de Paléobiologie 24: 75–95.
Brasil, L. 1894. Céphalopodes nouveaux ou peu connus des étages Jurassiques de Normandie. Bulletin de la Société géologique de Normandie 16: 27–46.
Buckman, S.S. 1920. Yorkshire Type Ammonites, Vol. 3 (2), 9–16. William Wesley and Son, London.
Buckman, S.S. 1924. Yorkshire Type Ammonites, Vol. 5 (44–49), 5–48. William Wesley and Son, London.
Callomon, J.B., Dietl, G., Galácz, A., Gradl, H., Niederhöfer, H.-J., and Zeiss, A. 1987. Zur Stratigraphie des Mittel- und unteren Oberjuras in Sengenthal bei Neumarkt/Opf. (Fränkische Alb). Stuttgarter Beiträge zur Naturkunde, Serie B 132: 1–53.
Collignon, M. 1958. Atlas des fossiles caractéristiques de Madagascar, Tome 2, Bathonien, Callovien, pls. 6–33. Service géologique, Tananarive.
Corroy, G. 1932. Le Callovien de la bordure Orientale du Bassin de Paris. Mémoires Carte géologique de la France. 337 pp. Imprimerie Nationale, Paris.
Couffon, O. 1919. Le Callovien du Chalet, commune de Montreuil-Bellay (Marne-et-Loire). Bulletin de la Société d’Études Scientifiques d’Angers 49: 15–97.
Courville, P. and Bonnot, A. 1998. Faunes ammonitiques et biochronologie de la zone à Athleta et de la base de la zone à Lamberti (Callovien supérieur) de la Côte de Meuse (France). Intérets des faunes nouvelles d’Aspidoceratidae. Revue de Paléobiologie 17: 307‒346.
Dembicz, K., Głowniak, E., Matyja, B., and Praszkier, T. 2006. Stop B1.4. Ogrodzieniec quarry, uppermost Bathonian to Middle Oxfordian ammonite succession. In: A. Wierzbowski, R. Aubrecht, J. Golonka, J. Gutowski, M. Krobicki, B.A. Matyja, G. Pieńkowski, and A. Uchman (eds), Jurassic of Poland and Adjacent Slovakian Carpathians. Field Trip Guidebook of 7th International Congress on the Jurassic System, Poland, Kraków, September 6–18, 2006, 144–148. Polish Geological Institute, Warszawa.
Dietl, G. 1986. Erstnachweis von Oecoptychius subrefractus (S. Buckm.) (Ammonoidea) aus dem Unter-Bathonium (Mittl. Jura) von SW-Deutschland. Stuttgarter Beiträge zur Naturkunde, Serie B 119: 1–13.
Dietze, V., Krieger, T., and Schweigert, G. 1997. Über Oecoptychius subrefractus (Buckman), Asphinctites tenuiplicatus (Brauns) und Polysphinctites secundus (Wetzel) (Ammonoidea) aus dem Unter-Bathonium (Mittlerer Jura) der Oberpfalz (Nordost-Bayern, Süddeutschland). Stuttgarter Beiträge zur Naturkunde, Serie B 245: 1–25.
Engel, T. 1908. Geognostischer Wegweiser durch Württemberg, 3rd edition. 675 pp. Schweizerbart, Stuttgart.
Fischer, J.-C. 1994. Revision critique de la Paleontologie française d’Alcide d’Orbigny. 1. Cephalopodes jurassiques. 352 pp. Masson, Paris.
Fortwengler, D., Marchand, D., and Bonnot, A. 1997. Ammonites et limite Callovien–Oxfordien dans les “Terres Noires” du Diois (Bassin du Sud-Est, France): exemples des coupes de Thuoux et Savournon. Géobios 30: 519–540. Crossref
Fürsich, F.T., Pandey, D.K., Callomon, J.H., Jaitly, A.K., and Singh, I.B. 2001. Marker beds in the Jurassic of the Kachchh Basin, Western India: their depositional environment and sequence stratigraphic significance. Journal of the Palaeontological Society of India 46: 173–198.
Glangeaud, P. 1897. Sur la forme de l’ouverture de quelques ammonites. Bulletin de la Société Géologique de France, sér. 3 25: 99–107.
Głowniak, E. 2012. The perisphinctid genus Prososphinctes Schindewolf (Ammonoidea, subfamily Prososphinctinae nov.): an indicator of palaeoecological changes in the Early Oxfordian Submediterranean sea of southern Poland. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 264: 117–179. Crossref
Haq, B.U. 2018. Jurassic Sea-Level Variations: A Reappraisal. GSA Today 28: 4–10. Crossref
Heller, F. and Zeiss, A. 1972. J.M.C. Reinecke und sein Werk: Des Urmeeres Nautili und Argonautae aus dem Gebiet von Coburg und Umgebung. Erlanger Geologische 90: 1–42.
Hyatt, A. 1900. Cephalopodes. In: K.A. Zittel and C.R. Eastman (eds.), Textbook of Paleontology, 502–604. Macmillan & Co., London.
Jain, S. 1997. On the earliest occurrence of Genus Phlycticeras Hyatt in Kachchh, Western India. Journal of Geological Society of India 49: 75–80.
Jain, S. 2017. Occurrence, age and paleobiogeography of rare genera Phlycticeras and Pachyerymnoceras from South Tethys. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 283: 119–149. Crossref
Jain, S. and Marek, M. 2021. New dimorphic species of the genus Rollierites Jeannet from southern Poland. Annales Societatis Geologorum Poloniae 91: 405‒418. Crossref
Jain, S. and Pandey, D.K. 2000. Middle Jurassic Ammonite Biozonation in Kachchh, western India. Bulletin of the Indian Geologistsʼ Association 33: 1–12.
Jain, S. and Salamon, M.A. 2023. First occurrence of the rare middle Jurassic ammonite genus Erymnocerites Jeannet from southern Poland. Historical Biology [available online, https://doi.org/10.1080/08912963.2023.2183857] Crossref
Jain, S., Salamon, M.A., and Brachaniec, T. 2022a. On the occurrence of the ammonite aptychi Lamellaptychus cf. rectecostatus (Peters, 1854) from the Ogrodzieniec quarry (southern Poland). Annales de Paléontologie 108: 102534. Crossref
Jain, S., Salamon, M.A., Schweigert, G., Kuźma, D., and Duda, P. 2022b. Genetic abnormality, parasitism and a new forma type of paleopathy from the upper Callovian ammonite genus Peltoceras Waagen. Lethaia 55: 1–17. Crossref
Jeannet, A. 1951. Stratigraphie und Palaeontologie des oolithischen Eisenerzlagers von Herznach und seiner Umgebung. Beiträge zur Geologie der Schweiz, Geotechnische Serie 13: 1–240.
Lebrun, P. and Courville, P. 2013. Le Jurassique des falaises des Vaches-Noires. Fossiles, 2013, Les falaises des Vaches-Noires. Un gisement emblématique du Jurassique à Villers-sur-Mer, Normandie. Fossiles, Hors série 4: 16–27.
Makowski, H. 1952. La faune Callovienne de Łuków en Pologne. Palaeontologia Polonica 4: 1–98.
Neumayr, M. 1878. Über unvermittelt auftretende Cephalopoden-Typen im Jura Mittel-Europas. Jahrbuch der kaiserlich-königlichen geologischen Reichsanstalt 28: 37–80.
Pellenard, P., Fortwengler, D., Marchand, D., Thierry, J., Bartolini, A., Boulila, S., Collin, P.-Y., Enay, R.R., Galbrun, B., Gardin, S., Huault, V., Huret, E., Martinez, M., and Smith, C.S. 2014. Integrated stratigraphy of the Oxfordian global stratotype section and point (GSSP) candidate in the Subalpine Basin (SE France). Volumina Jurassica 12: 1–44.
Quenstedt, F.A. 1846. Petrefaktenkunde Deutschlands. 1/1. Cephalopoden. 150 pp. Fues, Tübingen. Crossref
Quenstedt, F.A. 1857. Über die Rückenhöhle in der Schale gewisser Ammoniten (Dorso-cavati). Neues Jahrbuch für Mineralogie, Geognosie, Geologie und Petrefaktenkunde 1857: 544–548.
Quenstedt, F.A. 1887. Die Ammoniten des Schwäbischen Jura. 2. Der Braune Jura 524 pp. Schweizerbart, Stuttgart.
Rajnath 1942. The Jurassic rocks of Cutch—their bearing on some problems of Indian Geology (Presidential Address, Geology Section). Proceedings of the 29th Indian Science Congress 2: 93–106.
Reinecke, J.C.M. 1818. Maris protogaei Nautilos et Argonautas vulgo Cornua Ammonis in agro Coburgico et vicino reperiundos. 90 pp. Ahl, Coburg.
Roman, F. and Debrun, P. 1924. Etudes sur le Callovien de la vallée du Rhône. I. Le Callovien de Naves. Travaux du Laboratoire de Géologie de la Faculté des Sciences de Lyon. Ancienne série 6: 1–128.
Sayn, G. and Roman, F. 1928. Études sur le Callovien de la Vallée du Rhône. II. Monographie stratigraphique et paléontologique du Jurassique moyen de la Voulte-sur-Rhône. Travaux du Laboratoire de Géologie de la Faculté des Sciences de Lyon 13: 1–165.
Schlegelmilch, R. 1985. Die Ammoniten des süddeutschen Doggers. 284 pp. Fischer, Stuttgart. Crossref
Schweigert, G. and Dietze, V. 1998. Revision der dimorphen Ammonitengattungen Phlycticeras Hyatt – Oecoptychius Neumayr (Strigoceratidae, Mitteljura). Stuttgarter Beiträge zur Naturkunde, Serie B 269: 1–59.
Schweigert, G., Dietl, G., and Dietze, V. 2003. Neue Nachweise von Phlycticeras und Oecoptychius (Ammonitina: Strigoceratidae: Phlycticeratinae). Stuttgarter Beiträge zur Naturkunde, Serie B 335: 1–21.
Spath, L.F. 1925. Ammonites and aptychi. In: B.N.K. Wyllie and W.E. Smellie (eds.), The Collection of Fossils and Rocks From Somaliland. Monographs of the Hunterian Museum, Glasgow University 7: 111–164.
Spath, L.F. 1928. Revision of the Jurassic Cephalopod fauna of Kachh (Cutch). Memoirs of the Geological Survey of India, Palaeontologia Indica, New Series 9: 73–278.
Waagen, W. 1875. Jurassic fauna of Kutch. The Cephalopoda. Memoirs of the Geological Survey of India, Palaeontologia Indica 9: 1–40.
Zakrzewski, A.J.A. 1887. Die Grenzschichten des Braunen zum Weißen Jura in Schwaben. Jahreshefte des Vereins für vaterländische Naturkunde in Württemberg 43: 87–141.
Zieten, C.H. von 1830. Die Versteinerungen Württembergs, 2. Heft. 8 pp. Schweizerbart, Stuttgart.
Zieten, K.H. 1831. Die Versteinerungen Württembergs, 3. Heft. 17–24. Schweizerbart.
Acta Palaeontol. Pol. 68 (2): 321–335, 2023
https://doi.org/10.4202/app.01026.2022