
A new look at the Emsian (Early Devonian), sarcopterygian fishes from the Holy Cross Mountains, Poland, with a special reference to porolepiforms
OLGA WILK
Wilk, O. 2023. A new look at the Emsian (Early Devonian), sarcopterygian fishes from the Holy Cross Mountains, Poland, with a special reference to porolepiforms. Acta Palaeontologica Polonica 68 (2): 343–357.
Sarcopterygian remains are relatively common in the so-called “Placoderm Sandstone” (storm-origin bone-bearing breccia) from the Emsian (Lower Devonian) of Podłazie in the Holy Cross Mountains (Poland). Previous studies have mainly referenced the presence of dipnomorphs, chiefly portrayed by porolepiforms and occurrences of dipnoans. Porolepiforms are represented by Porolepis ex gr. P. posnaniensis, based on scales, skull and shoulder-girdle elements, whereas other remains have been attributed to Heimenia sp. The aim of this paper is to present the most detailed description of the porolepiforms and other sarcopterygian fishes from the “Placoderm Sandstone” from the Holy Cross Mountains since 1960s. Here, I present a new study based on more than 200 specimens, using both microtomographic (CT-scan) and silicon casts analyses, which indicate that not two, but at least three different taxa of sarcopterygians occur in the “Placoderm Sandstone” from Podłazie. The new data presented herein depict a more diversified assemblage of sarcopterygians in the marginal-marine environment of the Lower Devonian of the Holy Cross Mountains than previously thought, and shed a new light on sarcopterygian diversity along the shores of the Old-Red continent.
Key words: Sarcopterygii, Porolepiformes, Placoderm Sandstone, Devonian, Holy Cross Mountains, Poland.
Olga Wilk [olga.wilk@pgi.gov.pl; ORCID: https://orcid.org/0000-0002-6199-4518 ], Faculty of Geology, University of Warsaw, al. Żwirki and Wigury 93, 02-089 Warszawa, Poland; Polish Geological Institute-National Research Institute, ul. Rakowiecka 4, 00-975 Warszawa, Poland.
Received 24 November 2022, accepted 1 April 2023, available online 9 May 2023.
Copyright © 2023 O. Wilk. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
The bone-bearing breccia known as the “Placoderm Sandstone” of Poland was originally established by Gürich (1896) and is now best-known form one locality in the Holy Cross Mountains, the Podłazie Hill. The Emsian (Lower Devonian) siliciclastic deposits yielded an abundant and diverse fossil fish assemblage, including acanthodians, agnathans, chondrichthyans, placoderms, and sarcopterygians (Tarlo 1957, 1961, 1964, 1965; Kulczycki 1960; Szrek et al. 2014, 2015, 2021; Szrek and Dupret 2017; Wilk et al. 2020). Due to the deposition of the carcasses during high-energy events and subsequent dissolution of the mineralized skeleton by migrating fluids, all skeletal elements such as dermal and endoskeletal bones, plates, scales, spines, and teeth are preserved as natural moulds (Szrek et al. 2014). The spatial distribution and relative abundance of remains also were analysed and show terrestrial-influenced conditions in the Kielce region with special focus on Podłazie which is the richest site in such fossils (Wilk et al. 2022).
The Porolepiformes is a group of large, robust sarcopterygians (lobe-finned fishes), exclusively found in near-shore environments across the Devonian, which consists of three families: Porolepididae Woodward, 1891, Holoptychiidae Owen, 1860, and Ventalepididae Lebedev and Lukševičs, 2018. Several characteristics may distinguish this group from the rest of the remaining sarcopterygians, including the presence of dendrodont-type teeth (Schultze 1969; Panchen and Smithson 1987; Mondéjar-Fernández and Janvier 2014), presence of sub-squamosal bones (Cloutier 1990; Ahlberg 1991; Cloutier and Schultze 1996), absence of intertemporal and supratemporal bones, contribution of the nasal series to the skull roof margin posterior to the orbit (Ahlberg 1992), and the otic part of the main lateral line canal passing through the radiation center of the postparietal bones (Ahlberg 1991; Mondéjár-Fernandez and Clément 2012; Mondéjar-Fernández et al. 2021).
The first report of porolepiforms from Poland reaches out to the 19th century, when Kade (1858) described and illustrated scale material from erratic boulders from Silesia as Gyrolepis posnaniensis (Actinopterygii) and Gyroptychius posnaniensis (Osteolepididae) (Ørvig 1969). Later, Woodward (1891) assigned them to the genus Porolepis based on similarities with scales he collected from Spitsbergen. However, the first detailed description of sarcopterygians from the Emsian “Placoderm Sandstone” was by Kulczycki (1960). He presented a thorough description of scales, skull and shoulder-girdle elements that he attributed to Porolepis ex gr. posnaniensis. Referring to previous reports (Ørvig 1969; Clément 2001b), Dec (2010) proposed to reinterpret Kulczycki’s material and suggested that the Polish material include at least two porolepiform taxa. Later, Wilk et al. (2020) proved that the material from Podłazie Hill contains scales belonging both to Porolepis sp. and Heimenia sp.
The main purpose of this paper is to show the most detailed description of sarcopterygian fishes with special reference to the porolepiforms from the “Placoderm Sandstone” from the Holy Cross Mountains and compare the biodiversity during the Early Devonian along the shores of the Old-Red continent.
Institutional abbreviations.—ANSP, Academy of Natural Sciences, Philadelphia, USA; MUT, Military University of Technology, Warsaw, Poland; MZ, Polish Academy of Sciences Museum of the Earth, Warsaw, Poland (PASME); PIG, PIG-PIB, Polish Geological Institute-National Research Institute, Warsaw, Poland; D1080aD27UW, University of Warsaw, Warsaw, Poland.
Geological setting
All fossil material presented in this study was collected from the site called Podłazie Hill (Fig. 1A), located near Daleszyce, about 12 km southeast of Kielce in the Holy Cross Mountains, Poland. Details on this locality have been provided by Szrek et al. (2014) and supplemented later by Szrek et al. (2021) and Wilk et al. (2022). The so-called “Placoderm Sandstone” is particularly well-known from this part of the Holy Cross Mountains and is remarkable by its abundance in fossil fishes. The diverse vertebrate assemblage (including acanthodians, agnathans, chondrichthyans, placoderms, and osteichthyans) is composed of remains that were transported during storms and deposited in shallow water conditions, where they mixed with reworked sediment (Szrek et al. 2014; Fig. 1B).

Fig. 1. A. Map showing location (inset) and Lower Devonian deposits of the Holy Cross Mountains (HCM), star indicates the investigated locality. B. Lithological profile of Podłazie Hill (modified from Szrek et al. 2014). C. Block of “Placoderm Sandstone” with vertebrate remains (after Szrek et al. 2014: fig. 7A).
The sandstone was exploited for road construction until the 1960s, and later it was used by the local residents for economic purposes. The quarry was closed and abandoned for many years, which resulted in overgrown vegetation, making its excavation difficult (Król et al. 2021). Therefore, a large part of the material from the present study comes from the rich collection gathered during the last sixty years by Marek Dec, Julian Kulczycki, and Piotr Szrek.
Material and methods
The bone bearing breccia (the “Placoderm Sandstone”) is an exceptional type of quartzitic sandstone, in which the original bone tissues dissolved during diagenetic processes and were sometimes re-filled with clayey substance (Fig. 1C). Because of the high-energy environmental regime during the deposition of the “Placoderm Sandstone”, all fossils are preserved as disarticulated impressions (natural moulds) of bony elements but with an excellent preservation of soft tissues imprints such as nerves or muscle attachments. Due to the absence of fossilized bone tissues the only methods for examining the material is through micro-CT-scanning and silicone casting. Herein, both methods were used. Firstly, silicon casts on the external surface of the sandstone blocks were made and photographed using a Nikon D80 camera with AF Micro Nikkor 60mm 1:2:8D lens. Next, 18 blocks were scanned at the MUT using a high-resolution μCT-scanning in order to observe the remains located inside the blocs with no exposure to the surface. Application of these two methods allowed the identification of normally hidden and thus unknown features of the fossil specimens and as a result it constitutes the most extensive insight ever into the “Placoderm Sandstone”. All the collected material of porolepiforms includes about 150 silicon casts and more than 50 specimens found inside the scanned blocks. Most of the material was fragmented or broken and could not be determined with certainty. However, after a comprehensive analysis, it was possible to distinguish more than 30 different elements. The studied material was collected mainly by J. Kulczycki in 1956 and by P. Szrek and M. Dec during field work in the Holy Cross Mountains between 2004 and 2022. The material is curated in the PASME and in the PIG-PIB.
Systematic palaeontology
Class Sarcopterygii Romer, 1955
Order Dipnomorpha Ahlberg, 1991
Suborder Porolepiformes Jarvik, 1942
Family Porolepididae Woodward, 1891
Genus Porolepis Woodward, 1891
Type species: Gyrolepis posnaniensis Kade, 1858; Devonian, Międzyrzecz area, Poland.
Porolepis sp.
Figs. 2–4.
Material.—MZ-VIII/Vp-56, 58, 400, 458, PIG 178, PIG-PIB-5P, parts of the skull and postcranial skeleton, Podłazie, Emsian.
Description.—Ethmosphenoid shield: MZ-VIII/Vp-56 (Fig. 2) is 35 mm long and 44 mm wide. The anterior margin of the premaxilla is rounded with four partly preserved teeth. The specimen displays two relatively shallow, oval (10 mm long and 6 mm wide) internasal cavities separated by a 7 mm long internasal ridge. The attachment area for the left vomer and the edges of the fossa autopalatina are visible. Posterior to the internasal cavities there is a fossa where a triangular parasphenoid should have been located, but it is not preserved.
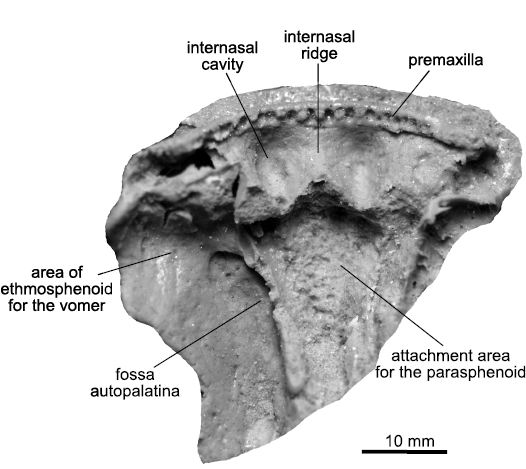
Fig. 2. Porolepidid sarcopterygian Porolepis sp. from Podłazie, Holy Cross Mountains, Poland, Emsian. MZ-VIII/Vp-56, ethmosphenoid in ventral view.
Lower jaw: The material consists of two fragmentary mandibles (MZ-VIII/Vp-58 and 400, Fig. 3A, B) that have been previously mentioned by Kulczycki (1960: pl. 5: D) and Dec (2010). They represent the anterior parts of the lower jaw. Specimen MZ-VIII/Vp-58 (Fig. 3A) is 37 mm long and 14 mm high and is gently widening posteriorly. This specimen is one of two among the whole material where the symphysis is present and it is also possible to distinguish the mentomandibular rib, the precoronoid fossa and the first and second coronoids. The coronoid fangs are not preserved and the coronoids do not display marginal denticles. The incomplete prearticular is 13 mm long, narrow and smooth with no denticles on its preserved surface. Underneath it, a wide adductor fossa is visible associated with a Meckelian foramen. Teeth on the dentary are broken.
MZ-VIII/Vp-400 (Fig. 3B) represents only a small quadrangular fragment (32 mm in length and height) of the anterior part of the jaw. The anterior margin of the dentary does not display any teeth but bears eight tooth sockets. Only the anterior coronoid without the coronoid fang is present. The precoronoid fossa is partly preserved, as well as a portion of the prearticular.
Median extrascapular: PIG 178 (Fig. 4B) is 49 mm long and 19 mm wide, tapering posteriorly. It represents an incomplete left half of the median extrascapular. The outer surface is covered with cosmine. Anteriolaterally there are larger sensory pores on the surface. The anterior margin, where the bone is slightly thinner, corresponds to the area overlapped by the postparietals. The inner side has no distinctive features, except a faintly preserved supratemporal commissural canal.

Fig. 3. Porolepidid sarcopterygian Porolepis sp. from Podłazie, Holy Cross Mountains, Poland, Emsian. Lower jaws in ventral view. A. MZ-VIII/Vp-58. B. MZ-VIII/Vp-400.
Parietal: PIG-PIB-5P (Fig. 4A) represents an incomplete part of the posterolateral visceral side of the left parietal (37 mm long and 22 mm wide). The posterior margin bears a small process resulting in a pointed posterolateral end of the bone bearing an open infraorbital sensory canal. The anterior portion of the bone is greatly eroded. Growth center can be traced and located excentrically close to the infraorbital sensory canal.
Postparietal: The anterior and posterior edges of MZ-VIII/Vp-458 (Fig. 4C) are broken. The plate is 33 mm long and 15 mm wide, with roughly parallel lateral and medial margins and becomes narrower towards the anterior margin. Two overlapping surfaces for the prespiracular antero-laterally and the tabular laterally are well preserved. No pit lines can be observed.
Remarks.—The general outline of the ethmosphenoid MZ-VIII/Vp-56 (Fig. 2) is similar to Porolepis brevis Jarvik, 1937 (Jarvik 1972: fig. 65B). As in Porolepis brevis, the shape of the internasal cavities is oval, and just next to them lies the attachment area for the vomer. Under the internasal cavities, the contour of the anterior part of the parasphenoid is triangular and anteriorly pointed, which agrees with that in Porolepis brevis. The ethmosphenoid in Heimenia ensis Ørvig, 1969 (Clément 2001a: fig. 2) is also similar to the one presented herein, but the internasal cavities are much deeper and the parasphenoid is wider. Glyptolepis groenlandica Jarvik, 1972, has entirely different internasal cavities with strongly marked margins and the internasal ridge is much broader (Jarvik 1972: pl. 14: 5).
MZ-VIII/Vp-58 and 400 probably representing the Porolepis, are characterized as follows: denticles absent on the coronoids; the first coronoid fang is long and narrow; the prearticular is smooth and relatively slim; the mentomandibular rib is rather slender and narrowing to the symphysis.
The general outline of the median extrascapular (Fig. 4B) described herein fit with the trapezoid median extrascapular of “porolepids” like Porolepis brevis (Jarvik 1972: fig. 43) and Durialepis edentatus Otto, 2007 (Mondéjar-Fernández 2020: fig. 10A). As in Porolepis brevis (Jarvik 1972: fig. 61, pl. 6: 2), the extrascapular has a trapezoid shape with a wider anterior part for the overlapping of the postparietal, and a narrower posterior edge. Among holoptychiids, Holoptychius sp. (Jarvik 1972: fig. 58) has a rounded posterior margin while in Glyptolepis groenlandica the overall shape of the bone is rounded (Jarvik 1972: fig. 41). Remarkably, the course of the supratemporal commissural canal is more posteriorly situated in this specimen than in other porolepiforms, where it runs midway (e.g., Porolepis sp.) or across the anterior third of the bone (e.g., holoptychiids like Holoptychius sp. and Glyptolepis groenlandica) (Mondéjar-Fernández 2020: fig. 10A).
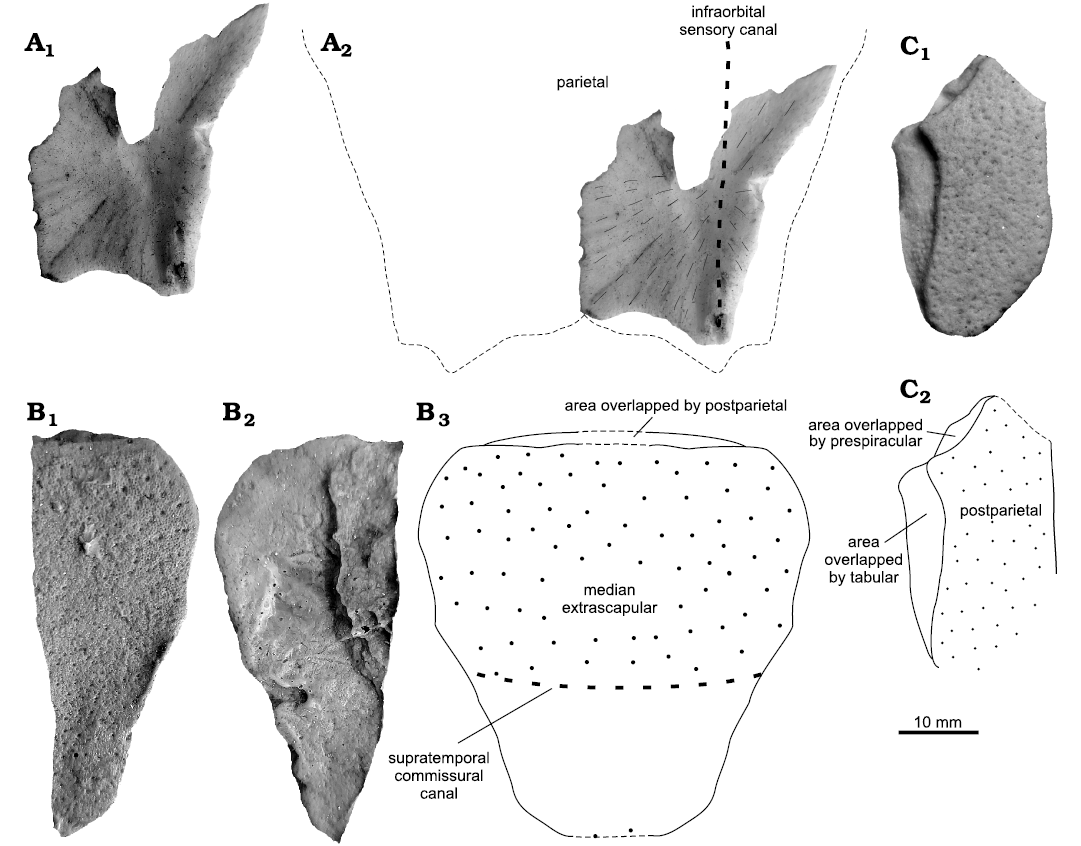
Fig. 4. Porolepidid sarcopterygian Porolepis sp. from Podłazie, Holy Cross Mountains, Poland, Emsian. A. PIG-PIB-5P, parietal in ventral view (A1), interpretative drawing of the complete element in ventral view (A2). B. PIG 178, median extrascapular in dorsal (B1) and ventral (B2) views, interpretative drawing of the complete element in dorsal view (B3). C. MZ-VIII/Vp-458, postparietal in dorsal view (C1), interpretative drawing of the dorsal view (C2).
The general outline of the posterolateral part of the parietal is similar as in Porolepis brevis (Jarvik 1972: fig. 34A, pl. 8: 1). Despite that the anterior part of the bone is eroded the posterior margin with a sharply pointed process oriented downward looks similar.
The postparietal (Fig. 4C) is similar to the one in Nasogaluakus chorni Schultze, 2000 (Schultze 2000: fig. 1; Mondéjar-Fernández 2020: fig. 10D). Although the bone is much smaller and the margin with the tabular is straighter and only gently curve. Similarly, the postparietal displays a small lateral expansion at the level of the anterior tip of the tabular, similar to numerous other porolepiforms (Schultze 2000: fig. 1; Mondéjar-Fernández 2020: fig. 10D). However, an holoptychiid attribution can be ruled out because of the presence of numerous fine cosmine pores in the presented specimen.
Genus Heimenia Ørvig, 1969
Heimenia sp.
Type species: Heimenia ensis Ørvig, 1969; Devonian, Spitsbergen.
Figs. 5, 6.
Material.—MZ-VIII/Vp-55, 61, 441, 459, PIG-PIB-7P–11P, part of the skull and postcranial skeleton; Podłazie, Emsian (Kulczycki 1960: pl. 5: A, C; pl. 6: 18, 20, 21, 23).
Description.—Coronoids and teeth: Three separate coronoids (PIG-PIB-8P–10P) and three teeth (MZ-VIII/Vp-441, 459, PIG-PIB-11P) have been identified (Fig. 5A–F). Four of these specimens (PIG-PIB-8P, 9P, 11P, MZ-VIII/Vp-459) have been previously illustrated by Kulczycki (1960: pl. 6: 18, 20, 21, 23). All three coronoids (Fig. 5A–C) have a single completely-preserved fang, slightly curved at the top and covered by fine striae along its entire length. All coronoid fangs reach a maximum length between 15–20 mm and are overally rounded in cross-section. The biggest tooth (PIG-PIB-11P, Fig. 5D) is 37 mm and the smallest one (MZ-VIII/Vp-459, Fig. 5E) is 20 mm high. The lateral margin on the coronoids contacting the dentary is covered by a single row of irregularly arranged small denticles.
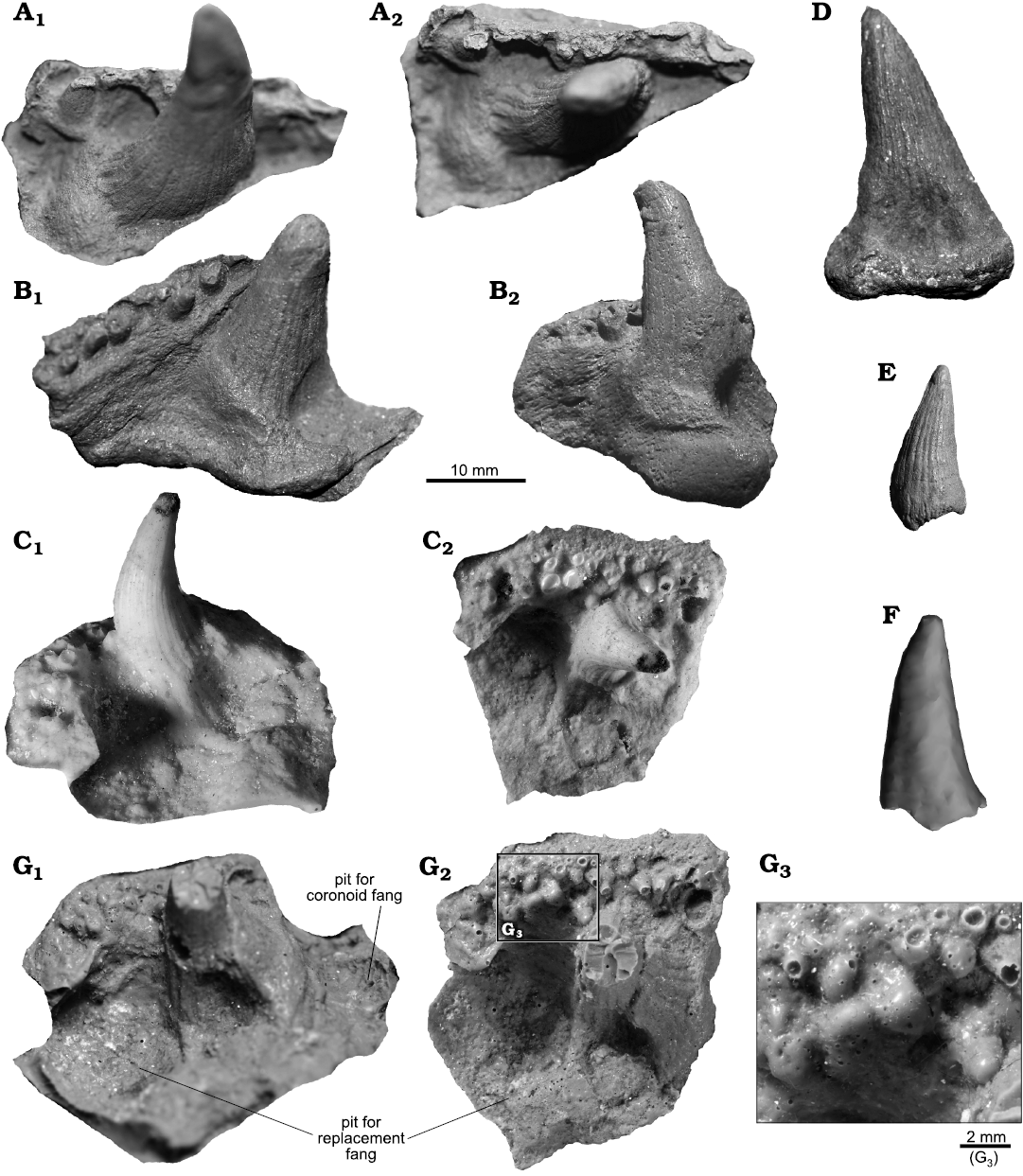
Fig. 5. Porolepidid sarcopterygian Heimenia sp. from Podłazie, Holy Cross Mountains, Poland, Emsian. Coronoids (A–C), teeth (D–F), dermopalatine (G). A. PIG-PIB-8P in lateral (A1) and ventral (A2) views. B. PIG-PIB-9P in lateral (B1) and ventral (B2) views. C. PIG-PIB-10P in lateral (C1) and ventral (C2) views. D. PIG-PIB-11P in lateral view. E. MZ-VIII/Vp-459 in lateral view. F. MZ-VIII/Vp-441 in lateral view. G. PIG-PIB-7P in lateral (G1) and ventral (G2) views, close up (G3).
Dermopalatine: PIG-PIB-7P (Fig. 5G) represents a well preserved left dermopalatine in ventral view of approximately 45 mm width. The posterior margin displays a large pit for a replacement fang. The anterior process of the dermopalatine is broken. The antero-lateral margin is covered by many little denticles, which are arranged irregularly. The fang located in the middle part of the dermopalatine is broken at one-third of its height, showing a circular cross-section. The fang is covered by fine ridges with shallow grooves, extending from the base.
Lower jaw: Two anterior parts of lower jaw were found in the material (MZ-VIII/Vp-55, 61). Both have been figured by Kulczycki (1960: pl. 5: A, C). MZ-VIII/Vp-61 (Fig. 6A) corresponds to the biggest specimen among the described material. It is 150 mm long, with a height at the level of the first coronoid just behind the coronoid fang of 74 mm and gradually wider (88 mm) at the posterior edge that ends behind the second coronoid. Across the entire length of the dentary a row of little denticles is visible. The biggest coronoid fang is 28 mm high and 13 mm wide at the base. The fangs have ridges and grooves on the whole surface and they are rounded in cross-section. One fang from the first coronoid is complete, but the second one has a slightly broken tip. Both coronoid fangs are similar in size. Denticles in the lateral vertical lamina come in two sizes, smaller and larger. The prearticular is about 80 mm long, relatively wide and narrowing anteriorly. The entire surface of the prearticular is covered by denticles, which are smaller and more eroded ventrally and bigger dorsally. The anterior part ends at the level of the first coronoid. Just anterior to it, there is a large precoronoid fossa for the upper jaw tooth.
MZ-VIII/Vp-55 (Fig. 6B) is relatively small (35 mm long, 16 mm wide) compared with the previously described lower jaw, but it is the only specimen in which all three coronoid fangs are preserved. The first two fangs are almost completely preserved, but the third one is broken at mid height, revealing a circular cross section. The lateral margin of the coronoids is characterized by the presence of a single row of little denticles. The teeth are not preserved on the dentary, only their sockets can be seen. The anterior part of the mandible is broken, thus the symphysis is not preserved. The mentomandibular rib is present, although there is no preserved prearticular.

Fig. 6. Porolepidid sarcopterygian Heimenia sp. from Podłazie, Holy Cross Mountains, Poland, Emsian. Lower jaws in ventral view. A. MZ-VIII/Vp-61. B. MZ-VIII/Vp-55.
Remarks.—The shape of the dermopalatine found in the porolepiform material is similar to that of Heimenia, holoptychiids but also in certain “osteolepiforms” (Jarvik 1980; Rosen et al. 1981; Lebedev 1995; Clément 2001a). The dermopalatine described herein has almost all the same characters (pit for replacement fangs, pit for coronoid fang, row of small denticles in the vertical lamina) as Heimenia ensis as figured by Clément (2001a: fig. 5B). However, the new specimen is bigger than described by Clement’s (2001a) and slightly more eroded.
According to Dec’s (2010) preliminary observations of the examined material, MZ-VIII/Vp-61 and 55 were suggested to belong probably to Heimenia sp. (Vorobyeva 1963; Ørvig 1969; Clément 2001a) because: the lateral vertical lamina of the coronoids is covered with small denticles; the first coronoid fang is short and wide; the prearticular is relatively wide with a row of denticles on the dorsal edge; the mentomandibular rib is rather short and massive transitioning into a spoon-shape area.
Since Kulczycki (1960) identified Porolepis ex gr. posnaniensis Kade, 1858, from Podłazie Hill, several authors cast some doubts whether the described specimen actually represents a new species (Vorobyeva 1963; Ørvig 1969; Clément 2001a). MZ-VIII/Vp-61 is relatively big and it slightly differs from other known lower jaws from the same site. Jarvik (1972: pl. 12: 6) described the lower jaw of Heimenia sp. but only from the external side, so there is no described elements of the visceral side. Because the described material from Podłazie presents only the visceral side of the lower jaw, there is no possibility to compare them with others. Clément (2001b: pl. 6: A, B) presented the lower jaw of Heimenia ensis from both, visceral and external side. However, the jaws described by both Jarvik (1972) and Clément (2001b) are much narrower than MZ-VIII/Vp-61, although the lengths are similar. Differences in the inner surface of the specimen are noticeable also. In most specimens, the prearticular extends to the level of the posterior margin of the first coronoid, unlike in MZ-VIII/Vp-61 where it ends more anteriorly, at the level of the anterior edge of the first coronoid. A similar feature found in all specimens is the rounded incision for the adductor fossa and the covering of the dorsal edge of the prearticular with irregularly arranged little denticles. Precoronoid fossa in all published specimens (Kulczycki 1960; Clément 2001b) is spoon-like shape as in specimens described herein (Fig. 5A, B)
Sarcopterygii indet.
Fig. 7–9.
Material.—MZ-VIII/Vp-66, 401–403, 437, 449, 457, 581, PIG-PIB-2P, 3P, 4P 12P, PIG 273, 349, PIG 1733.II.437, D1080aD27UW, parts of the skull and postcranial skeleton; Podłazie, Emsian (Kulczycki 1960: pl. 6: 16).
Description.—Jugal: MZ-VIII/Vp-581 was microtomographically studied (Fig. 7). It consists of an isolated right jugal (20 mm long and 12 mm high), relatively well preserved although the straight lower margin, which contacted the maxilla, appears to be slightly eroded, however, its course is easy to trace (Fig. 7A1, A3). The jugal is externally convex and gently concave viscerally (Fig. 7A5). All the contacts with the other cheek bones are well recognizable (Fig. 7A6). The anterior margin, which contacted the lachrymal, is pierced by an opening for the jugal sensory canal that runs across the bone (Fig. 7A1–A5). The orbit rim is deeply concave and is antero-dorsally located. The postero-dorsal margin is slightly concave and would have contacted the postorbital. Across this margin, a double infraorbital sensory canal opening is visible. Both openings are closely located and separated by a thin septum of about 2 mm in thickness, however may represent secondary structure among ramification of the main canal (Fig. 7A2, A3, A5). The posterior margin is straight for the contact with the squamosal and it carries an opening for the infraorbital sensory canal, visible in posterior view. The course of the lateral line canals can be followed on the visceral side through elongated thickenings revealing a relatively straight infraorbital canal running close to the ventral margin and the junction between the postorbital branch and infraorbital lateral line canals on the postero-ventral corner of the bone. Due to the poor preservation there are no visible pores or other evidences of cosmine.
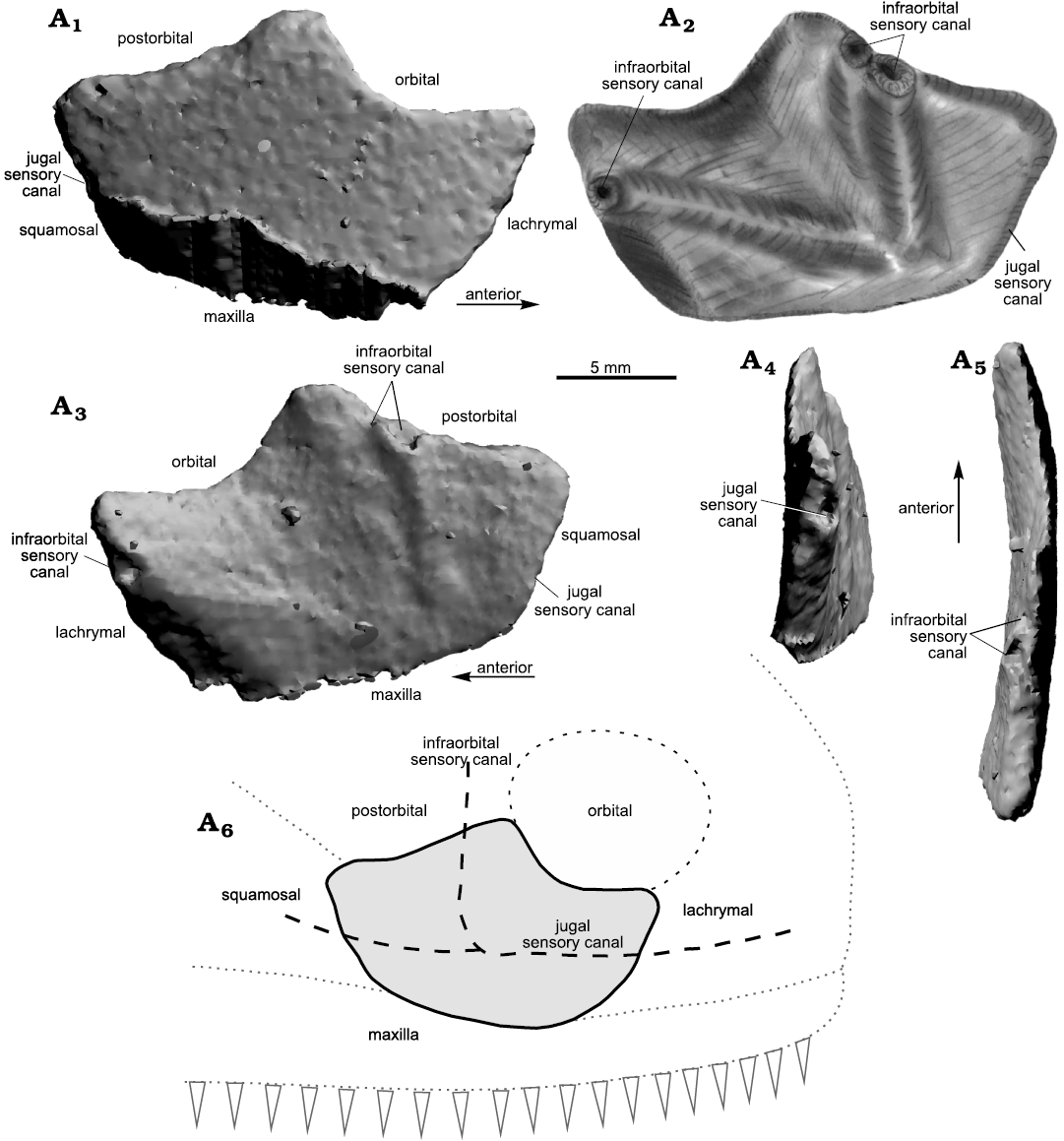
Fig. 7. Sarcopterygii indet. from Podłazie, Holy Cross Mountains, Poland, Emsian. MZ-VIII/Vp-58, right jugal in external (A1, A2), ventral (A3), and lateral (A4, A5) views, interpretative drawing (A2), interpreted outline of the original (likely) shape of the jugal (A6; not to scale).
Lower jaw: Six incomplete lower jaws (MZ-VIII/Vp-401–403, PIG-PIB-2P, PIG 349, PIG 1733.II.437) have been found (Fig. 8A–F). Some of these specimens (MZ-VIII/Vp-401–403, PIG 1733.II.437) were briefly described by Dec (2010). The smallest specimen (PIG-PIB-2P, Fig. 8A) is 24 mm long and 5 mm high and corresponds to an incomplete dentary. On the external surface, the small pores of the cosmine cover are visible. The small specimen MZ-VIII/Vp-401 (28 mm in length, Fig. 8B) preserves the symphysis anteriorly and the lamina carrying the parasymphysial dental plate is also visible. PIG 349 (Fig. 8C) is 36 mm long. On the upper edge the row of six tooth sockets is visible. The external surface is covered with cosmine, but the inner side is smooth without any characteristics. MZ-VIII/Vp-402 (Fig. 8D) is preserved from both, visceral and outer side. The 60 mm long external part is preserved to a greater extent, whereas only 20 mm of the visceral side are visible. The external surface on the specimen is covered with cosmine. On the visceral side a part of coronoid without a coronoid fang is preserved. On the external surface of MZ-VIII/Vp-403 (51 mm long) which is covered with cosmine, the pore arrangement of the lateral line network is visible (Fig. 8E). Due to partial damage to the bone, the top layer on the posterior part of the specimen has been breached so it is possible to track the main lateral line canal with short branches extending from it. PIG 1733.II.437 (Fig. 8F) represents the anterior tip of a toothless lower jaw but it is very eroded and poorly preserved.
Maxilla: Two maxillary fragments (MZ-VIII/Vp-449, PIG-PIB-3P), have been found. MZ-VIII/Vp-449 (Fig. 8G) is rectangular in outline and it is about 60 mm long and 40 mm wide. The entire external surface is covered with cosmine pores. One little tooth is preserved with only its tip visible in lateral view. Most likely the specimen represents the middle part of the maxilla. Specimen PIG-PIB-3P (Fig. 8H) is almost 90 mm long and 20 mm wide. It displays three little teeth and a visceral ridge visible on the lingual side.

Fig. 8. Sarcopterygii indet. from Podłazie, Holy Cross Mountains, Poland, Emsian. Lower jaw (A–F), maxilla (G, H). A. Lower jaw (PIG-PIB-2P) in dorsal (A1) and lateral (A2) views. B. MZ-VIII/Vp-401 in dorsal (B1) and ventral (B2) views. C. PIG 349 in dorsal (C1) and ventral (C2) views. D. MZ-VIII/Vp-402) in ventral (D1) and lateral (D2) views. E. MZ-VIII/Vp-403 in lateral view. F. PIG 1733.II.437 in dorsal (F1) and ventral (F2) views. G. MZ-VIII/Vp-449 in lateral view. H. PIG-PIB-3P in ventral view.
Ethmosphenoid: MZ-VIII/Vp-437 (Fig. 9A) is 25 mm wide and 17 mm high. The internasal cavities are bean shape with a noticeable internasal ridge. The floor of the left internasal cavity is broken. The area for the right vomer is slightly marked on the outer margin of the right internasal cavity, but it is also incomplete. A portion of the premaxilla bearing six tooth sockets is visible on the anterior margin of the specimen.
Postparietal and tabular: An incomplete left part of a postparietal (PIG-PIB-12P, Fig. 9B) is 77 mm long and 39 mm wide. Only the dorsal surface is visible. The overlapping surface for the tabular is observed on the lateral side, which is located deeper in comparison to the external surface of the postparietal. The medial edge is badly preserved. The posterior margin is almost complete, it displays a slightly sinusoid contour. The tabular is 42 mm long and 19 mm wide. The outer margin appears to be eroded on the anterolateral and bottom edge.
Other elements: The general outline of D1080aD27UW (Fig. 9C) is triangular in shape and broad-base. It is 5.2 cm long and 4 cm in the widest point. All the outer surface is ornamented. MZ-VIII/Vp-66 (Fig. 9G) similar to the previous specimen, triangular in shape is 4.8 cm long and 2.3 cm in the widest point, it has been presented by Kulczycki (1960: pl. 6: 16). Its external surface is also ornamented. Specimen PIG 273 (Fig. 9D) is a dermal bone covered entirely with cosmine. The anterolateral margin is slightly lower and has no cosmine cover, which indicates that another bone was overlapping it. There is no visible lateral line canal. The specimen is 4 cm long and 2.5 cm wide. The last one is MZ-VIII/Vp-457 (Fig. 9F). It is 4.9 cm long and 3.2 cm wide, tapering to one extreme. Both external side edges are broken, however, one part has a deep, oval depression for overlapping bone. The dorsal edge of the bone is covered by little cavities.

Fig. 9. Sarcopterygii indet. from Podłazie,
Holy Cross Mountains, Poland, Emsian. A.
MZ-VIII/Vp-437, ethmosphenoid in ventral view. B.
PIG-PIB-12P, parietal in dorsal view. C–G. Undetermined elements in lateral view.
C. D1080aD27UW. D.
PIG 273. E. PIG-PIB-4P. F. MZ-VIII/Vp-457. G.
MZ-VIII/Vp-66.
Remarks.—The isolated jugal bone displays a series of peculiar features. The shape of the orbital margin suggests relatively large eyes. The bone contacted certainly the lachrymal, maxilla, squamosal and postorbital and postero-ventrally framed the orbit (Fig. 7A1). This general arrangement fits the condition known in porolepiforms (e.g., Mondéjar-Fernández 2020: fig. 10), onychodonts (Mondéjar-Fernández 2020: fig. 17), “osteolepiforms” (Jarvik 1985: figs. 2C, 5A–C; Borgen 2011: fig. 143D, J; Borgen and Nakrem 2016: fig. 143D, J) and other tetrapodomorphs (Säve-Söderbergh 1932: text-figs. 2, 11, 12, 15; Jarvik 1996: figs. 23, 24; Daeschler et al. 2009: fig. 4A, B). However, the bone is proportionally short with a relatively large and concave orbital margin, different from porolepiforms like Porolepis brevis, Nasogaluakus chorni, Laccognathus panderi, and Durialepis edentatus (Jarvik 1972: fig. 43C; Schultze 2000: fig. 4; Mondéjar-Fernández and Clément 2012: fig. 11; Mondéjar-Fernández 2020: fig. 10) in which the jugal is relatively large and the orbital margin being comparatively shorter. Moreover, the length of each margin is almost equal in this specimen, whereas in porolepiforms the largest edges are those for the postorbital and maxilla (e.g., Jarvik 1972: fig. 5; Mondéjar-Fernández 2020: figs. 9, 10; Mondéjar-Fernández and Clément 2012: fig. 11). The specimen of Porolepis brevis described and illustrated by Jarvik (1972: pl. 6: 1) has a jugal bone with outline shape and proportions similar to the Polish specimens. However, if the position in Jarvik’s figure (1972: pl. 6: 1) is rotated the margin which contacts the maxilla includes the posterior opening of the jugal sensory canal, which is obviously incorrect. A very short orbital rim or no contact whatsoever between the orbit and the jugal occurs in “osteolepiforms”. Despite the fact that the relative length of the bone margins and general shape of the jugal in certain “osteolepiforms” like Gyroptychius (Jarvik 1985: fig. 5) are similar to this Polish specimen, the extremely reduced contact between the jugal and the orbit strongly differs. Comparisons with onychodonts do not allow to assign the specimen to this group. The jugal in onychodonts does not show a concaved eye margin and is rather straight (e.g., Andrews et al. 2006: figs. 3B, 4B; Long 2001: fig. 1). The jugal in onychodonts also possesses an overlapping surface for the lachrymal in the anterior part, which is not present in the Polish specimen. Among onychodonts, Onychodus sp. displays a ventral expansion of the infraorbital sensory canal into maxilla (Long 2001: fig. 1C), which also does not occur in the Polish specimen. Because of its unusual proportions that suggest the presence of large orbits, this jugal bone does not fit to any known taxa already identified in the “Placoderm Sandstone”. The posterior part of the bone and the anterior margin for the contact with the lachrymal suggest a robust maxilla with anteriorly high portion forming a triangle pointing towards the jugal–lachrymal suture; this feature is similar to that of the tetrapod Ichthyostega sp. (Ahlberg et al. 2005). Other features may also point towards a tetrapodomorph affinity of this bone. Daeschler et al. (2009: fig. 4A, B) illustrated two jugal plates belonging to undetermined tetrapod forms. They are similar to the specimen from Podłazie in carrying deeply concave orbital margins, especially in ANSP 22892 (Daeschler et al. 2009: fig. 4B), but the contact with the maxilla is significantly shorter in the Polish specimen. The plate seems not to be abraded as it preserves almost all of its natural margins. On the inner side, elongated elevations reveal the course of the lateral line canal. However, the external side does not show any signs of the course of the lateral line, suggesting that it was deeply enclosed in the bone and pores on the surface are unfortunately not visible due to the scan resolution. Enclosing of the retained lateral line is regarded as an advanced character of the tetrapodomorphs (e.g., Ahlberg and Milner 1994; Jarvik 1996; Clack 2012). To sum up, a precise identification of the specimen is not possible because of its preservation as an isolated element. However, based on comparisons with other putative co-eval sarcopterygian candidates, it most likely represents an undetermined tetrapodomorph form.
Described here lower jaws (MZ-VIII/Vp-401–403, PIG-PIB-2P, PIG 349, PIG 1733.II.437) are either too damaged or solely preserved on one side without any distinctive characters that allowed identifying any taxa in detail. However, the presence of cosmine on the external side of the dermal bones allows to assign them to Sarcopterygii (except Actinistia and Onychodontida; Schultze 2016).
MZ-VIII/Vp-449 and PIG-PIB-3P most likely represent parts of maxilla (Mondéjar-Fernández 2020: figs. 5, 6) of porolepiforms owing to the shape (long and narrow) and the presence of big pores on cosmine cover.
The ethmosphenoid MZ-VIII/Vp-437 (Fig. 9A) is completely different from those known in the “Porolepididae” and presented herein (Fig. 2). A rather thin internasal ridge and rounded, bean-shape internasal cavities were not observed in any known species. In most porolepiforms, the internasal cavities are rounded (Porolepis cf. spitsbergensis, Jarvik, 1972 [pl. 7: 1]; Powichthys thorsteinssoni, Jessen, 1980 [pl. 5: 1], oval (Porolepis brevis, Jarvik, 1972 [fig. 65]; Glyptolepis groenlandica, Jarvik, 1972 [fig. 31]) whereas they are more elongated in onychodonts, e.g., Qingmenodus yui Lu and Zhu, 2010 (Lu et al. 2016: fig. 1A).
PIG-PIB-12P (Fig. 9B) shows a postparietal and tabular which are similar to Glyptolepis groenlandica (Mondéjar-Fernández 2020: fig. 10H). The general outline of the described specimen is almost the same but the upper part of the postparietal is more “sinusoidal” than it is in Glyptolepis. The notch for the tabular also shows resemblance to Glyptolepis, however, due to the fact that the major part of the tabular is eroded it is hard to estimate the approximated natural shape of the bone.
Among the dozens of disarticulated remains, several bones can only be assigned to the osteichthyans (Fig. 9C–G). Some bones described herein as “other elements” belong most likely to Sarcopterygii but due to the state of the preservation they cannot be determined accurately. Only those that have cosmine (PIG-273, MZ-VIII/Vp-457) could be tentatively assigned to porolepiforms but cosmine is also present in basal tetrapodomorphs and dipnoans. PIG-PIB-4P (Fig. 9E), which probably represents urohyal has a wide anterior end, narrowing greatly towards the posterior end, thus more closely resembling the one in Eusthenopteron foordi (Jarvik, 1954 [fig. 10D]). D1080aD27UW has a very specific ornamentation, which is similar to the one occurring in Eusthenodon bourdoni (Downs, Barbosa, and Daeschler, 2021 [fig. 6]). MZ-VIII/Vp-66 has a similar ornamentation, and these characters can indicate a potential affinity to Eusthenodon sp. MZ-VIII/Vp-66 could possibly correspond to the anterior part of a clavicle (e.g., Downs et al. 2018: fig. 7B) but due to the poor state of preservation is hard to establish with any certainty.
Palaeodiversity of Porolepiformes from the Early Devonian of Laurussia
During the Early Devonian, Podłazie Hill in the Holy Cross Mountains was part of the Małopolska block and it belonged to the bigger continent called Laurussia or Euramerica (Domeier and Torsvik 2014). On the other side of this continent in the area of today’s Canadian Arctic, near the Anderson River, the Bear Rock Formation was formed. Despite the fact that both sites were located on opposite sides of the Euramerican continent, several hundred kilometers apart from each other, the fish assemblages are similar and show close spatial and environmental relations. Among the fauna from the Podłazie Hill, acanthodians, agnathans, chondrichthyans, osteichthyans, and placoderms have been identified (e.g., Tarlo 1957; Kulczycki 1960; Blieck 1980; Szrek and Dupret 2017; Wilk et al. 2022). On the other hand, the Anderson River fauna is represented by common Early Devonian fish remains belonging to acanthodians, placoderms, and osteichthyans (Schultze and Cumbaa 2017).
In both areas, the amount of the material suggests that conditions during the Emsian were favorable for the development of many fishes. Porolepiforms found in Podłazie belong to the Porolepis and Heimenia, while in the Canadian Arctic remains of Porolepis, Heimenia, Nasogaluakus have been reported. Skull elements, as well as elements of the postcranial skeleton, scales and teeth are common. The presence of at least two genera of porolepiforms (i.e., Porolepis and Heimenia in the Holy Cross Mountains of Poland and the Anderson River in the Canadian Arctic), suggest a viable open-water connection between these two areas during the late Early Devonian, consistent with a wider distribution of Heimenia, Porolepis, and other porolepiforms in the Early Devonian as suggested by Schultze and Cumbaa (2017). This is also confirmed by the fact that outside the area of the Holy Cross Mountains (Kulczycki 1960; Ørvig 1969) and the Anderson River (Jarvik 1967; Schultze 1968; Ørvig 1969; Schultze and Cumbaa 2017), the presence of Heimenia was recorded in the Lower Devonian from the Baltic region (Lyarskaya 1981; Clément 2001a, b), central Europe (Jessen 1980), China (Zhu 2000; Zhao and Zhu 2010) and possibly Vietnam (Thanh and Janvier 1987), while Porolepis is known exclusively from Europe, Spitsbergen, Australia, and Canada (Gross 1936; Obruchev 1938; Jarvik 1942; Kulczycki 1960; Vorobyeva 1963; Jessen 1980; Schultze 1993, Clément 2004; Johanson et al. 2013; Mondéjar-Fernández 2020).
Conclusions
The “Placoderm Sandstone” is an unique kind of siliciclastic sequence of the Lower Devonian in Poland known for abundant vertebrate fossils. Most of the bony elements are broken and incomplete due to the deposition in the high-energy environment. Based on high-resolution μCT and silicon casts, cranial and postcranial skeletal elements of the Emsian sarcopterygians with the special reference to porolepiforms from the Holy Cross Mountains have been described.
During research, among the numerous vertebrate material, parts of the skull and postcranial skeleton belonging to at least two genera of porolepiforms (Porolepis, Heimenia) and other sarcopterygians have been noted. In the examined material, the most common elements correspond to lower jaws and coronoids with fangs. The posterior part of the lower jaw is similar in all known porolepiforms but they differ in the articular region compared to the description provided by Ahlberg (1992). The meckelian bone, the coronoids and the prearticular are similar in both Heimenia and Porolepis. The external side is covered by a relatively high dentary with four infradentaries below. On the anterior part of the dentary of Porolepis and Heimenia, an antero-medial lamina forms the roof of the symphysial pit and extends up to the symphysis. However, in Porolepis the teeth are not present in the transverse part of the dorsal margin of the dentary, which is slightly depressed (Jarvik 1972).
The presented results indicate that there is a higher taxonomic diversity among the sarcopterygian fishes from the “Placoderm Sandstone” in the Holy Cross Mountains than previously supposed.
Acknowledgements
I would like to warmly thank Marek Dec (Warsaw, Poland) for the silicon casts he prepared in past years and which he kindly passed by to me. I am also grateful to Per Ahlberg (Uppsala University, Sweden) and Jorge Mondéjar-Fernández (Senckenber Research Institute & Natural History Museum, Fulda, Germany) for their valuable remarks and suggestions. I would like to thank Piotr Szrek (Polish Geological Institute-National Research Institute, Warsaw, Poland) for assistance in recognizing material and the revision of an early version of the manuscript. I would also like to thank the Museum of the Earth, Polish Academy of Sciences (Warsaw, Poland) for loaning the blocks of the “Placoderm Sandstone” for CT-scanning. This work was supported by the University’s Integrated Development Programme (ZIP), co-financed by the European Social Fund within the framework of Operational Programme Knowledge Education Development 2014–2020, action 3.5. This research was financed by the Polish National Science Center (project number 2019/35/B/ST10/01505 granted to Michał Ginter). The author thanks the reviewers Richard Cloutier (Université du Québec à Rimouski, Canada) and Hans-Peter Schultze (University of Kansas, Lawrence, USA) and the editor Daniel E. Barta (Oklahoma State University, Tahlequah, USA) for their comments, which significantly improved the final version of the manuscript.
References
Ahlberg P.E. 1991. A re-examination of sarcopterygian interrelationships, with special reference to the Porolepiformes. Zoological Journal of the Linnean Society 103: 241–287. Crossref
Ahlberg, P.E. 1992. The palaeoecology and evolutionary history of the porolepiform sarcopterygians. In: E. Mark-Kurik (ed.), Fossil Fishes as Living Animals, 71–90. Estonian Academy of Science, Tallinn.
Ahlberg, P.E. and Milner, A.R. 1994. The origin and early diversification of tetrapods. Nature 368: 507–514. Crossref
Ahlberg, P.E., Clack, J.A., and Blom, H. 2005. The axial skeleton of the Devonian tetrapod Ichthyostega. Nature 437: 137–140. Crossref
Andrews, S., Long, J., Ahlberg, P.E., Barwick, R., and Campbell, K. 2006. The structure of the sarcopterygian Onychodus jandemarrai n. sp. from Gogo, Western Australia: with a functional interpretation of the skeleton. Transactions of the Royal Society of Edinburgh Earth Sciences 96: 197–307. Crossref
Blieck, A. 1980. Le genre Rhinopteraspis Jaekel (Vertébrés, Hétérostracés) du Dévonien inférieur: systématique, morphologie, répartition. Bulletin du Muséum national d’Histoire naturelle 4: 25–47.
Borgen, U.J. 2011. Morphology, Phylogeny and Taxonomy of Osteolepiform Fish. 694 pp. Ph.D. Dissertation, Natural History Museum, University of Oslo, Oslo.
Borgen, U.J and Nakrem, H.A. 2016. Morphology, phylogeny and taxonomy of osteolepiform fish. Fossils and Strata 61: 1–520. Crossref
Clack, J.A. 2012. Gaining Ground: the Origin and Evolution of Tetrapods. 544 pp. Indiana University Press, Indiana.
Clément, G.2001a. Evidence for lack of choanae in the Porolepiformes. Journal of Vertebrate Paleontology 21: 795–802. Crossref
Clément, G. 2001b. Etudes anatomiques des genres Porolepis, Woodward 1891 et Heimenia Ørvig, 1969 (“Porolepididae”, Sarcopterygii) et du genre Powichthys Jessen, 1975 (Powichthyidae, Sarcopterygii) du Dévonien Inférieur et Moyen du Spitzberg: phylogénie des Dipnomorpha. 297 pp. Ph.D. dissertation. Muséum National d’Histoire Naturelle de Paris, Paris.
Clément, G. 2004. Nouvelles données anatomiques et morphologie générale des “Porolepididae” (Dipnomorpha, Sarcopterygii). Revue de Paléobiologie 9: 193–211.
Cloutier, R. 1990. Phylogenetic Interrelationships of the Actinistians (Osteichthyes: Sarcopterygii): Patterns, Trends, and Rates of Evolution. 430 pp. Ph.D. dissertation, University of Kansas, Kansas.
Cloutier, R. and Schultze, H.-P. 1996. Porolepiform fishes (Sarcopterygii). In: H.-P. Schultze and R. Cloutier (eds.), Devonian Fishes and Plants of Miguasha, Québec, Canada, 248–270. Verlag Dr. Friedrich Pfeil, Munich.
Daeschler, E.B., Clack, J.A., and Shubin, N.H. 2009. Late Devonian tetrapod remains from Red Hill, Pennsylvania, USA: how much diversity? Acta Zoologica 90: 306–317. Crossref
Dec, M. 2010. Ryby kostnoszkieletowe z dolnodewońskich piaskowców plakodermowych w Górach Świętokrzyskich. 49 pp. M.Sc. thesis, University of Warsaw, Warsaw.
Domeier, M. and Torsvik, T.H. 2014. Plate tectonic in the late Paleozoic. Geoscience Frontiers 5: 303–350. Crossref
Downs, J.P., Daeschler, E.B., Long, A.M., and Shubin, N.H. 2018. Eusthenopteron jenkinsi sp. nov. (Sarcopterygii, Tristichopteridae) from the Upper Devonian of Nunavut, Canada, and a review of Eusthenopteron taxonomy. Breviora 562: 1–24. Crossref
Downs, J.P., Barbosa, J., and Daeschler, E.B. 2021. A new species of Eusthenodon (Sarcopterygii, Tristichopteridae) from the Upper Devonian (Famennian) of Pennsylvania, USA, and a review of Eusthenodon taxonomy. Journal of Vertebrate Paleontology 41 (3): e1976197. Crossref
Gross, W. 1936. Beitraege zur Osteologie baltischer und rheinischer Devon-Crossopterygier. Paläontologische Zeitschrift 18: 129–155. Crossref
Gürich, G. 1896. Das Paläozoicum im Polnischen Mittelgebirge. Verhandlugen der Russischen-Kaiserlichen Mineralogischen Gesellschaft zu St. Petersburg 2: 1–539.
Jarvik, E. 1937. On the species of Eusthenopteron found in Russia and the Baltic States. Bulletin of the Geological Institution of the University of Upsala 27: 63–127.
Jarvik, E. 1942. On the structure of the snout of crossopterygians and lower gnathostomes in general. Zoologiska Bidrag fran Uppsala 21: 235–675.
Jarvik, E. 1954. On the visceral skeleton in Eusthenopteron with a discussion of the parasphenoid and palatoquadrate in fishes. Kungliga Svenska Vetenskaps Akademiens Handlingar 5: 1–104.
Jarvik, E. 1967. On the structure of the lower jaw in dipnoans: with a description of an early dipnoan from Canada, Melanognathus canadensis, gen. et sp. nov. Journal of the Linnean Society (Zoology) 47: 155–183. Crossref
Jarvik, E. 1972. Middle and Upper Devonian Porolepiformes from East Greenland with special reference to Glyptolepis groenlandica n. sp.: and a discussion on the structure of the head in the Porolepiformes. Meddelelser om Grønland 187: 1–295.
Jarvik, E. 1980. Basic Structure and Evolution of Vertebrates. Volume 1. 575 pp. Academic Press, London.
Jarvik, E. 1985. Devonian osteolepiform fishes from East Greenland. Meddelelser om Grønland 13: 1–52.
Jarvik, E. 1996. The Devonian tetrapod Ichthyostega. Lethaia 29: 1–76. Crossref
Jessen, H.L. 1980. Lower Devonian Porolepiformes from the Canadian Arctic with special reference to Powichthys thorsteinssoni Jessen. Palaeontographica 167: 180–214.
Kade, G. 1858. Über die devonischen Fischreste eines Diluvialblockes. Programm der Königlichen Realschule zu Meseritz 1858: 1–23.
Johanson, Z., Ahlberg, P.E., and Ritchie, A. 2013. First record of Porolepis (Sarcopterygii; Porolepiformes) from eastern Gondwana. Canadian Journal of Earth Sciences 50: 249–253. Crossref
Król, P., Fijałkowska, A., Kozak, B., and Giełżecka-Mądry, D. 2021. Piaskowce dolnego dewonu na górze Barczy (Góry Świętokrzyskie) i historia ich eksploatacji. Przegląd Geologiczny 69: 81–90.
Kulczycki, J. 1960. Porolepis (Crossopterygii) from the Lower Devonian of the Holy Cross Mountains. Acta Palaeontologica Polonica 5: 65–103.
Lebedev, O. 1995. Morphology of a new osteolepidid fish from Russia. Bulletin du Muséum national d’Histoire naturelle 4: 287–341.
Lebedev, O. and E. Lukševičs. 2018. New materials on Ventalepis ketleriensis Schultze, 1980 extend the zoogeographic area of a Late Devonian vertebrate assemblage. Acta Geologica Polonica 68: 437–454.
Long, J. 2001. On the relationships of Psarolepis and the onychodontiform fishes. Journal of Vertebrate Paleontology 21: 815–820. Crossref
Lu, J., Zhu, M., Ahlberg, P.E., Qiao, T., Zhu, Y., Zhao, W., and Jia, L. 2016. A Devonian predatory fish provides insights into the early evolution of modern sarcopterygians. Science Advances 2: 1–8. Crossref
Lyarskaya, L.A. [Lârskaâ, L.A.] 1981. Baltic Devonian Placodermi: Asterolepididae [in Russian, with English abstract]. 152 pp. Zinatne, Riga.
Mondéjar-Fernández, J. 2020. A new onychodont (Osteichthyes; Sarcopterygii) from the Middle Devonian of Morocco and its bearing on early osteichthyan evolution. Journal of Systematic Palaeontology 18: 573–606. Crossref
Mondéjar Fernández, J. and Clément, G. 2012. Squamation and scale microstructure evolution in the Porolepiformes (Sarcopterygii, Dipnomorpha) based on Heimenia ensis from the Devonian of Spitsbergen. Journal of Vertebrate Paleontology 32: 267–284. Crossref
Mondéjar-Fernández, J. and Janvier, P. 2014. Further evidence for the presence of holoptychiid porolepiforms (Sarcopterygii, Dipnomorpha) from the Frasnian of Colombia. Comptes Rendus Palevol 13 (7): 587–597. Crossref
Mondéjar‐Fernández, J., Friedman, M., and Giles, S. 2021. Redescription of the cranial skeleton of the Early Devonian (Emsian) sarcopterygian Durialepis dentates Otto (Dipnomorpha, Porolepiformes). Papers in Palaeontology 7: 789–806. Crossref
Obruchev, D.V. [Obručev, D.V.]1938. Vertebrata aus dem Obersilur und Devon des Urals [in Russian, with German abstract]. Materials of the Central Scientific Research Institute for Geological Exploration (ZNIGRI) 2: 36–43.
Ørvig, T. 1969. Vertebrates from the Wood Bay Group and the position of the Emsian Eifelian boundary in the Devonian of Vestspitsbergen. Lethaia 2: 273–328. Crossref
Owen, R. 1860. Palaeontology, or, a Systematic Summary of Extinct Animals and Their Geological Relations. 420 pp. A. and C. Black, Edinburgh. Crossref
Panchen, A.L. and Smithson, T.R. 1987. Character diagnosis, fossils and the origin of tetrapods. Biological Reviews 62: 341–436. Crossref
Romer, A.S. 1955. Herpetichthyes, Amphibioidei, Choanichthyes or Sarcopterygii? Nature 176: 126. Crossref
Rosen, D., Forey, P., Gardiner, B., and Patterson, C. 1981. Lungfishes, Tetrapods, Paleontology, and Plesiomorphy. Bulletin of the American Museum of Natural History 167: 159–276.
Säve-Söderbergh, G. 1932. Preliminary note on Devonian stegocephalians from East Greenland. Meddelelser om Grønland 94: 1–107.
Schultze, H.-P. 1968. Palæoniscoidea-Schuppen aus dem Unterdevon Australiens und Kanadas und aus dem Mitteldevon Spitzbergens. Bulletin of the British Museum (Natural History) 16: 341–368.
Schultze, H.-P. 1969. Die Faltenzähne der rhipidistiiden Crossopterygier, der Tetrapoden und der Actinopterygier-Gattung Lepisosteus; nebst einer Beschreibung der Zahnstruktur von Onychodus (struniiformer Crossopterygier). Palaeontographica Italica 65: 63–137.
Schultze, H.-P. 1993. Osteichthyes: Sarcopterygii. In: M.J. Benton (ed.), The Fossil Record 2, 657–663. Chapman & Hall, London.
Schultze, H.-P. 2000. A porolepiform rhipidistian from the Lower Devonian of the Canadian Arctic. Fossil Record 3: 99–109. Crossref
Schultze, H.-P. 2016. Scales, enamel, cosmine, ganoine, and early osteichthyans. Comptes Rendus Palevol 15: 83–102. Crossref
Schultze, H.-P. and Cumbaa, S.L. 2017. A new Early Devonian (Emsian) arthrodire from the Northwest Territories, Canada, and its significance for paleogeographic reconstruction. Canadian Journal of Earth Sciences 54: 461–476. Crossref
Szrek, P. and Dupret, V. 2017. Placoderms from the Early Devonian “placoderm sandstone” of the Holy Cross Mountains, Poland with biostratigraphical and palaeobiogeographical implications. Acta Palaeontologica Polonica 62: 789–800. Crossref
Szrek, P., Niedźwiedzki G., and Dec, M. 2014. Storm origin of bone-bearing beds in the Lower Devonian placoderm sandstone from Podłazie Hill (Holy Cross Mountains, central Poland). Geological Quarterly 58: 795–806. Crossref
Szrek, P., Dec, M., and Niedźwiedzki, G. 2015. The first placoderm fish from the Lower Devonian of Poland. Journal of Vertebrate Paleontology 35 (3): e930471. Crossref
Szrek, P., Dec, M., and Wilk, O. 2021. The First Early Devonian Dipnoi from the Holy Cross Mountains, Poland. Journal of Vertebrate Paleontology 41 (1): e1923323. Crossref
Tarlo, L.B. 1957. A preliminary note on new ostracoderms from the Lower Devonian (Emsian) of central Poland. Acta Palaeontologica Polonica 2: 225–233.
Tarlo, L.B. 1961. Rhinopteraspis cornubica (McCoy), with notes on the classification and evolution of the pteraspids. Acta Palaeontologica Polonica 6: 367–402.
Tarlo, L.B. 1964. Psammosteiformes (Agnatha)—a review with descriptions of new material from the Lower Devonian of Poland. I general part. Palaeontologia Polonica 13: 1–135.
Tarlo, L.B. 1965. Psammosteiformes (Agnatha)—a review with descriptions of new material from the Lower Devonian of Poland. II systematic part. Palaeontologia Polonica 15: 1–168.
Thanh, T.D. and Janvier, P. 1987. Les vertébrés dévoniens du Viêtnam. Annales de Paléontologie 73: 165–194.
Vorobyeva, E.I. 1963. On crossopterygians of the genus Porolepis from the Devonian of the USSR. Paleontological Journal 2: 83–92.
Wilk, O., Szrek, P., and Ginter, M. 2022. Environmental differentiation reflected in the vertebrate fauna diversity in the Lower Devonian of the Holy Cross Mountains, Poland. Geological Quarterly 66 (35): 1–9. Crossref
Wilk, O., Szrek, P., Dec, M., Glinka, B., and Ahlberg, P.E. 2020. Comments on the squamation of Polish Lower Devonian porolepiforms. Journal of Vertebrate Paleontology 39(6): e1738448. Crossref
Woodward, A.S. 1891. The Devonian fish—fauna of Spitzbergen. Annals and Magazine of Natural History 8: 1–15. Crossref
Zhao, W. and Zhu, M. 2010. Siluro-Devonian vertebrate biostratigraphy and biogeography of China. Palaeoworld 19: 4–26. Crossref
Zhu, M. 2000. Catalogue of Devonian vertebrates in China, with notes on bio-events. Courier Forschungs-Institut Senckenberg 223: 373–390.
Acta Palaeontol. Pol. 68 (2): 343–357, 2023
https://doi.org/10.4202/app.01044.2022