
The reinstated identity of agglutinated foraminifer Campanellula capuensis from the Lower Cretaceous of southern Italy by means of a 3D model investigation
FELIX SCHLAGINTWEIT, SABRINA AMODIO, FILIPPO BARATTOLO, and MICHEL SEPTFONTAINE
Schlagintweit, F., Amodio, S., Barattolo, F., and Septfontaine, M. 2023. The reinstated identity of agglutinated foraminifer Campanellula capuensis from the Lower Cretaceous of southern Italy by means of a 3D model investigation. Acta Palaeontologica Polonica 68 (2): 245–260.
Campanellula capuensis was described as belonging to the Trochamminacea (trochospiral tests) and later transferred to the genus Orbitolinopsis of the Orbitolinidae (uniserial tests). Challenging its identity as a species of Orbitolinopsis, the most widely accepted classifications of agglutinated foraminifera reinstate Campanellula but retain its inclusion within the Orbitolinidae, subfamily Dictyoconinae. New material from the type locality and the San Lorenzello section (Matese Mountains, southern Apennines, Italy) as well as the construction of a 3D model allow to reinstate the original description as a low- to high-trochospirally (conical) coiled foraminifer with numerous chambers per whorl and to display an overall conical test morphology. An orbitolinid test construction including uniserial chambers (throughout the test or in its adult part) is absent. The conflicting opinions on taxonomic status of Campanellula are discussed leading to the removal from the order Loftusiina and the suborder Orbitolinina. Instead, Campanellula should be included into the order Lituolida and the suborder Verneuilinina. Campanellula capuensis represents a biostratigraphic and palaeobiogeographic marker taxon, restricted to upper Hauterivian–lower Barremian inner platform carbonates of the southern Neotethyan margin.
Key words: Foraminifera, Trochamminacea, biostratigraphy, calcareous agglutinated test, taxonomy, 3D reconstruction, Lower Cretaceous, Italy.
Felix Schlangintweit [felix.schlagintweit@gmx.de; ORCID: https://orcid.org/0000-0002-4024-4312 ], Lerchenauerstrasse 167, 80935 München, Germany.
Sabrina Amodio [sabrina.amodio@uniparthenope.it; ORCID: https://orcid.org/0000-0002-8241-1262 ] (corresponding author), Dipartimento di Scienze e Tecnologie, Università degli Studi di Napoli “Parthenope”, Centro Direzionale Isola C4, 80143 Napoli, Italy.
Filippo Barattolo [lippolo@unina.it; ORCID: https://orcid.org/0000-0003-4194-4764 ], Dipartimento di Scienze della Terra, dell’Ambiente e delle Risorse, Università degli Studi di Napoli Federico II, Complesso Universitario di Monte Sant’Angelo, via Cintia 21, 80126 Napoli, Italy.
Michel Septfontaine [septfontaine.m@bluewin.ch], Chemin des Pochattes 4, 1055 Froideville, Switzerland.
Received 13 December 2022, accepted 27 April 2023, available online 29 May 2023.
Copyright © 2023 F. Schlangintweit et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
This
research is dedicated to the memory
of Piero De Castro (1929–2023)
Introduction
When it was introduced, genus Campanellula De Castro, 1964, was included into the family Trochamminidae, uniting forms with trochospirally coiled tests. Some years later, based on material from Algeria, the species was assigned to the orbitolinid genus Orbitolinopsis by Schroeder in Macoin et al. (1970), assuming adult uniserial chamber arrangement and an internal test construction of this genus. Therefore, Schroeder in Macoin et al. (1970) considered Campanellula as a junior synonym of Orbitolinopsis Henson, 1948. Loeblich and Tappan (1987: 157), in turn, reinstated the Campanellula but considered it a member of family Orbitolinidae, subfamily Dictyoconinae. This suprageneric affiliation, however, has not been accepted by all subsequent workers (e.g., Arnaud-Vanneau and Sliter 1995; Gheiasvand et al. 2019). In the most recent classification of the agglutinated foraminifera, the concept of Loeblich and Tappan (1987) with Campanellula representing a genus of the Orbitolinidae is still followed (Kaminski 2014). New material from the upper Hauterivian–lower Barremian (Lower Cretaceous) San Lorenzello section (Mt. Monaco di Gioia, southern Apennines, Campania Region, Italy) and from the De Castro’s type locality (Fig. 1), has been studied in detail.
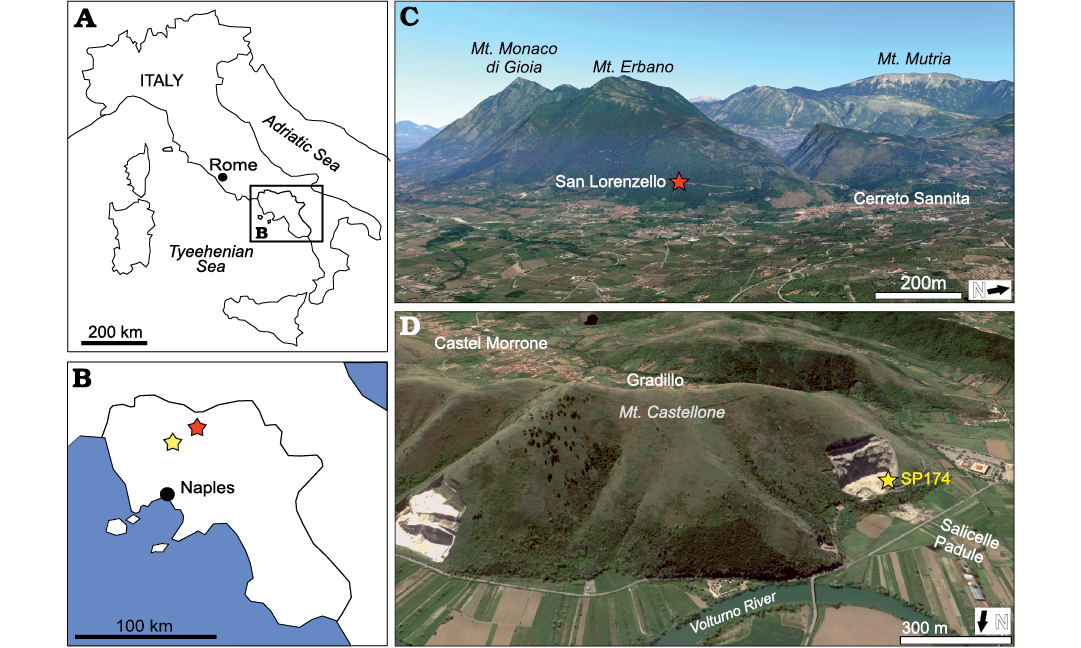
Fig. 1. Position of the studied localities. A. A contour map of the Italian Peninsula showing location of the Campania Region. B. Enlargement of the Campania Region with the two studied localities: San Lorenzello section (Benevento Province; red star) and the type locality of Campanellula capuensis (De Castro, 1964) near Castel Morrone village (Caserta Province; yellow star). C. Panoramic view of the San Lorenzello section (from Google Earth). D. Panoramic view of the Campanellula capuensis type locality along the SP174 road (from Google Earth).
A new investigation perspective on the type species, Campanellula capuensis De Castro, 1964, is here offered by the reconstruction of a 3D model that can be confronted with the test structure in thin section. The application of this method allows shedding further light into the internal test construction and the implied suprageneric position. The small-sized benthic foraminifer Campanellula capuensis represents a biostratigraphic and palaeobiogeographic marker taxon restricted to the upper Hauterivian–lower Barremian platform carbonates of the southern Tethyan margin (De Castro 1964; Macoin et al. 1970; Cherchi et al. 1981; Schroeder et al. 1978; Husinec and Sokač 2006, Velić 2007).
Institutional abbreviations.—DiST, Department of Science and Technology, Parthenope University, Naples, Italy; DiSTAR, Department of Earth Sciences, Environment and Resources, Federico II University, Naples, Italy; IP, Istituto di Paleontologia, Università di Napoli Federico II, Naples, Italy; Gmm, Geosciences Museum of Mashhad in the Geological Survey of North-Iran, East territory, Iran; IGZ, Institute of Geology, Zagreb, Croatia.
Geological setting
The studied locality is positioned in the San Lorenzello section (41°16’49.16”N, 14°32’23.24”E; Fig. 1A, B), about 70 km to the north of Naples and exposed along a roadcut on the southern slope of the Monte Monaco di Gioia (Matese Mountains, Benevento Province, Italy, Fig. 1C). This mountain is a part of the southern Apennines, a N-E verging fold and thrust belt derived from Apula-Ionian composite plate migration (Malinverno and Ryan 1986; Patacca and Scandone 2007), caused by African-Eurasian plate collision. The shallow-water carbonates, belonging to the Apennine Carbonate Platform (e.g., Patacca and Scandone 2007), have been investigated by a high-resolution integrated stratigraphy (D’Argenio et al. 1997; Ferreri et al. 2004; Amodio 2006; Amodio et al. 2008, 2013, 2018; Martino et al. 2019; Barattolo et al. 2021; Amodio et al. 2020, 2023; Parente et al. 2022). This study focuses on the Hauterivian–Barremian interval containing the local distribution range of Campanellula capuensis (78–151 m interval from the base of the section), labelled as D biostratigraphic unit (Fig. 2; see also Amodio et al. 2020: fig. 8). This interval corresponds to the Campanellula capuensis Biozone (De Castro 1991) and to the central part of the “Cuneolina scarsellai” and “Cuneolina camposaurii” biozones of Chiocchini et al. (2008). According to De Castro (1991) and Chiocchini et al. (2008), the stratigraphic range of Campanellula capuensis encompasses the Hauterivian–Barremian boundary, placed in this section at 100–110 m from the base of the section, above the first occurence of Salpingoporella melitae Radoičić, 1975 and Salpingoporella muehlbergii (Lorenz, 1902) indicated by Carras et al. (2006) as upper Hauterivian and below the appearance of Salpingoporella piriniae Carras and Radoičić, 1991, and the last occurence of Salpingoporella annulata Carozzi, 1953, in the upper Barremian (Carras et al. 2006). Campanellula capuensis is considered a key marker characterising the southern Tethys bio-province (Cherchi et al. 1981; Cherchi and Schroeder 1973, 2010), but information on its stratigraphic distribution vary in the literature (see among others Macoin et al. 1970; Velić and Sokač 1983; Peybernès et al. 1984; Claps et al. 1996; Husinec and Read 2018; Gheiasvand et al. 2019).

Fig. 2. Litho-biostratigraphy of the San Lorenzello section. The columnar log is based on the predominant lithofacies associations: Molluscan Limestones are open lagoonal deposits; BioPeloidal Limestones are inner shoal deposits; ForOstracod Limestones are restricted lagoonal deposits; Laminated Limestones are tidal flat deposits; and Charophyte Limestones are coastal ponds deposits. The log is juxtaposed with upper Valanginian–upper Barremian chronostratigraphic distribution of the main benthic foraminifera and green algae recognized in the section. The grey horizontal band represents the distribution range of Campanellula capuensis. See Amodio et al. (2020) for a detailed description of the lithofacies and biostratigraphic units.
Campanellula capuensis appears as an ubiquitous species in the San Lorenzello section where it occurs in almost all lithofacies associations (Molluscan Limestones, BioPeloidal Limestones, and ForOstracod Limestones, see for detailed description in Amodio et al. 2020: table 1), from open to restricted lagoonal facies as well as from sandy shoals. The biofacies is often very rich and diversified, including bivalves, gastropods, Cayeuxia-like thalli, green algae, benthic foraminifera and ostracods. The grainy textures encompass intraclasts, aggregate grains and peloids. The typical algal assemblage is characterised by Actinoporella podolica, Salpingoporella annulata, Praturlonella danilovae, Salpingoporella muehlbergii, Salpingoporella melitae, and Clypeina solkani (Fig. 2). In addition, Salpingoporella polygonalis, Salpingoporella piriniae and Clypeina parasolkani are also documented in this interval. The upper boundary of interval D is also marked by the last occurence of Salpingoporella annulata, and roughly corresponds to the last occurence of Clypeina parasolkani and Salpingoporella piriniae. The benthic foraminifera association includes Vercorsella scarsellai, Vercorsella laurentii, Vercorsella camposaurii, and Praechrysalidina infracretacea.
Material and methods
The San Lorenzello samples were collected during different field samplings (D’Argenio et al. 1997; Amodio 2006; Amodio et al. 2020). The material is partially hosted in the Sabrina Amodio collection (thin sections DiST/SL90–DiST/SL100), at DiST, Università degli Studi Parthenope of Naples, as well as in the Filippo Barattolo collection (thin sections DiSTAR-L8–DiSTAR-L41), at DiSTAR, Università degli Studi Federico II of Naples. We have followed the standard facies analysis for the carbonate rock classification, sequentially tabulating all the sedimentary features in a lithostratigraphic column (Fig. 2). The biostratigraphic analysis and age determination, mainly concerning benthic foraminifers and calcareous algae, were based on the biozonation schemes of the Lower Cretaceous platform carbonates (De Castro 1991; Carras et al. 2006; Chiocchini et al. 2008).
In addition, the type locality of Campanellula capuensis (see De Castro, 1964) was resampled in April 2022. This material is stored in the Filippo Barattolo collection (thin sections DiSTAR-BA. 4418), at DiSTAR, Università degli Studi Federico II of Naples. The original sampling site of De Castro is located at the last hairpin bend, near a quarry, on the SP174 road that leads from Gradillo locality in the municipality of Castel Morrone (Caserta Province, Campania Region, Italy) to Padule Salicelle (Fig. 1D). The samples containing Campanellula capuensis were also picked up from the base of the quarry wall, located on the opposite site of the roadcut, but at lower height of about 7–10 m than the original sampling site placed on the road SP174.
In this work, a high-resolution 3D model has been performed on the holotype supplemented by the analysis of numerous, variously oriented sections. The resulting 3D model was created by 3DS Max 2022 software. Afterwards, the 3D model has been sectioned and the resulting cuts compared with similar sections from the San Lorenzello material (see 3D rendering section below).
Systematic palaeontology
Phylum Foraminifera d’Orbigny, 1826
Class Globothalamea Pawlowski, Holzmann, and Tyszka, 2013
Subclass Textulariana Mikhalevich, 1980
Order Lituolida Lankester, 1885
Suborder Verneuilinina Mikhalevich and Kaminski in Kaminski, 2004
Genus Campanellula De Castro, 1964
Type species: Campanellula capuensis De Castro, 1964; Lower Cretaceous (Hauterivian–Barremian), southern Italy.
Species included: Type species and Campanellula herishtensis Schlagintweit, Rashidi, and Hanifzadeh, 2019 from the Gargasian, upper Aptian, Lower Cretaceous of central Iran.
Diagnosis.—From De Castro (1964): Imperforate, microgranular calcareous test. Cylindric-conical to bellflower in shape. Trochoid coiling with rather closed tours. Simple chambers, numerous in each tour (usually from six to eight in the type species). Simple aperture placed at the base of the chamber.
Remarks.—Campanellula belongs to the informal group of conical agglutinated foraminifera that include both trochospiral (e.g., Pseudochrysalidina Cole, 1941) and orbitoliniform taxa (e.g., Dictyoconus Blanckenhorn, 1900) (Hottinger and Drobne 1980; Vecchio and Hottinger 2007; Cruz Abad 2018). Orbitolinids may have an initial trochospire but the main test part is formed by uniserial chambers which later may become annular (Rat 1963; Schlagintweit 2023). With its small size, distinctly less than 0.5 mm, and simple chambers (= no exoskeleton), however, Campanellula cannot be regarded a larger benthic foraminifer. The microgranular-agglutinated wall exhibiting a thin outer hyaline calcareous layer is reported from the Orbitolinidae (e.g., Douglass 1960; Cherchi and Schroeder 1978; Schroeder 1985), but also from trochospirally coiled taxa such as the early Aptian Archaeosepta coratina Luperto Sinni and Masse, 1993, the early Cenomanian Altamirella Schlagintweit, Rigaud, and Wilmsen, 2015, or the Late Cretaceous Accordiella Farinacci, 1962, and Gyroconulina Schroeder and Darmoian, 1977 (see Schlagintweit et al. 2016). The hyaline layer is also present in the wall of Meyendorffina bathonica Aurouze and Bizon (own data of MS). In Altamirella for example, the hyaline calcitic layers “are not involved in the chamber formation process. They are secondarily added so that one layer can cover the umbilical side of several chambers but never covers the entire septa” (Schlagintweit et al. 2015: 3), what is also the case in Campanellula. The hyaline outer layer is very thin in Campanellula (thickness ~3–7 µm), compared to Altamirella (thickness ~5–20 µm) and appears functionally different. SEM observations on well preserved material would be required to decipher the nature of this “hyaline layer” (agglutinated? diagenetic induced?). For the Orbitolinidae, the finely agglutinated character of this layer has been generally assumed (e.g., Douglass 1960), and demonstrated for Mesorbitolina (Frijia et al. 2012).
De Castro (1964: 56) remarked that no other genus displays noteworthy similarities with Campanellula, an observation that appears still valid today. For an inclusion to any suprageneric category (family, superfamily, etc.) a clear morphological clarification of the internal arrangement of the chamber connections is provided by the three-dimensional model. In any case, the current taxonomic concept of treating Campanellula as a member of the Dictyoconinae (with uniserial chambers) and its subsequent inclusion into the order Orbitolinina has to be abandoned due to its trochospiral test throughout all stages, thus reinstating De Castro (1964). For the following synonymy, if not otherwise indicated, all references are from areas that were formerly parts of the southern margin of Neo-Tethys.
Campanellula capuensis De Castro, 1964
Figs. 3–10.
1964 Campanellula capuensis n. gen., n. sp.; De Castro 1964: 56, pl. 1, pls. 2–6 [in part].
1977 Orbitolinopsis capuensis (De Castro,1964); Chiocchini and Mancinelli 1977: 127, 137, pl. 24: 2.
1978 Orbitolinopsis capuensis (De Castro, 1964); Garcia-Hernandez 1978: 215, pl. 25: 1, 3–5.
1978 Orbitolinopsis capuensis (De Castro, 1964); Sokač and Velić 1978: 247, 249, pl. 8: 3, 4.
1982 Orbitolinopsis capuensis (De Castro, 1964); Peybernès 1982: pl. 2: 1, 2.
1982 Campanellula capuensis (De Castro, 1964); Barattolo 1982: 826–827, figs. 2, 3.
1984 Orbitolinopsis (?) capuensis (De Castro, 1964); Luperto Sinni and Masse 1984: 342, 346, 347, 350, pl. 40: 1–2.
1994 Campanellula cf. C. capuensis De Castro, 1964; Schindler and Conrad 1994: 72, pl. 5: 1.
1995 Campanellula capuensis De Castro, 1964; Chiocchini et al. 1995: 22, pl. 5: 16, 19, 20, 22–27.
1995 Campanellula capuensis De Castro, 1964; Carras 1995: 145, pl. 47: 1–3.
non 1995 Campanellula capuensis De Castro, 1964; Arnaud-Vanneau and Sliter 1995: 551, pl. 1: 12 (Central Pacific area).
non 1995 Campanellula cf. capuensis De Castro, 1964; Bucur et al. 1995: pl. 7: 12 (most likely an orbitolinid; Eastern Serbia = northern margin of Neo-Tethys).
1996 Campanellula capuensis De Castro, 1964; Claps et al. 1996: 10, 19–22, pl. 1: 9, 11–12.
2006 Campanellula capuensis De Castro, 1964; Husinec and Sokač 2006: 421, 425: 2, 7E, I–M.
2007 Campanellula capuensis De Castro, 1964; Bruni et al. 2007: 46, 49: 2, 3, pl. 3: 7–9.
2007 Campanellula capuensis De Castro, 1964; Velić 2007: 31, 42, pl. 11: 1–4.
2008 Campanellula capuensis De Castro, 1964; Chiocchini et al. 2008: 17, 42, tab.6, pl. 17: 4.
2012 Campanellula capuensis De Castro, 1964; Chiocchini et al. 2012: pl. 10: 8, 9.
2018 Campanellula capuensis De Castro, 1964; Cruz Abad 2018: 64, fig. 58.
non 2019 Campanellula capuensis De Castro, 1964; Gheiasvand et al. 2019: fig. 8g–h (N-Iran = northern margin of Neo-Tethys).
2020 Campanellula capuensis De Castro, 1964; Amodio et al. 2020: 6, 8–10, fig. 6: D–H.
Holotype: specimen in axial section (De Castro, 1964, pl. 1: 1), thin section IP-A.281.1.
Type locality: Castelmorrone, W slope of Mt. Castellone, Caserta Province, Campania Region, Italy.
Type horizon: White detritic limestone, De Castro (1964), Campanellula capuensis Biozone De Castro (1991), late Hauterivian–earliest Barremian, Cretaceous.
Description.—Test rather small (height ~0.2 mm), high conical to cylindroconical, with low trochospirally enrolled chambers. Adult specimens consist of up to 10 whorls (mostly 6–8; 6–11 [De Castro 1964]). Apical, spherical proloculus followed by up to seven trochospirally coiled whorls (Figs. 3A2, 6A2) each with five to eight chambers (Figs. 4H, 6H, 7B). The first whorls with a reduced number of chambers per whorl (? informal early praevalvulinid ontogenetic stage sensu Septfontaine 2020) may be slightly inclined towards the axis of the main adult stage (Figs. 5I3, 6A1, 7F1). In the adult stage the plan of one whorl in side view of the test is inclined about 30° with respect to the axis of the test, sometimes sub-horizontal (Fig. 8A, B). Chambers are irregular, subtriangular in transverse sections reaching deep into the umbilical region by means of blind tubular extensions (Figs. 6J, 7B3). The simple (= undivided) chambers only gradually increase in size as added (compare Figs. 4G, J1, 7B2); the ultimate chamber may occupy half to almost three-fourths of the test circumference (Figs. 3J, 6B2, 7A, B3, C, D1). In tangential section, the chambers arrangement recalls a fish scales pattern, vertically aligned (Figs. 5H1, I2, I3, J2, K, 6E). The foramina connecting chambers within a whorl are positioned at the inner chamber margin. The central part of the test is filled by anastomosed (labyrinthic) calcitic extensions from the chambers (= modification of a kind of a tooth-plate?), forming either a central mass or a columella filled by calcitic material (Fig. 6A2, B1, H). The test wall is dark microgranular (?finely agglutinated) exhibiting a rather thin umbilical layer made of clear agglutinated microgranular calcite or quartz grains per whorl (Figs. 3C1, F, 4O, 5D, 6D, F). Test most likely dimorphic expressed by specimens of bigger size assumed to belong to the microspheric generation (Figs. 3A, K2, 4F, K1).
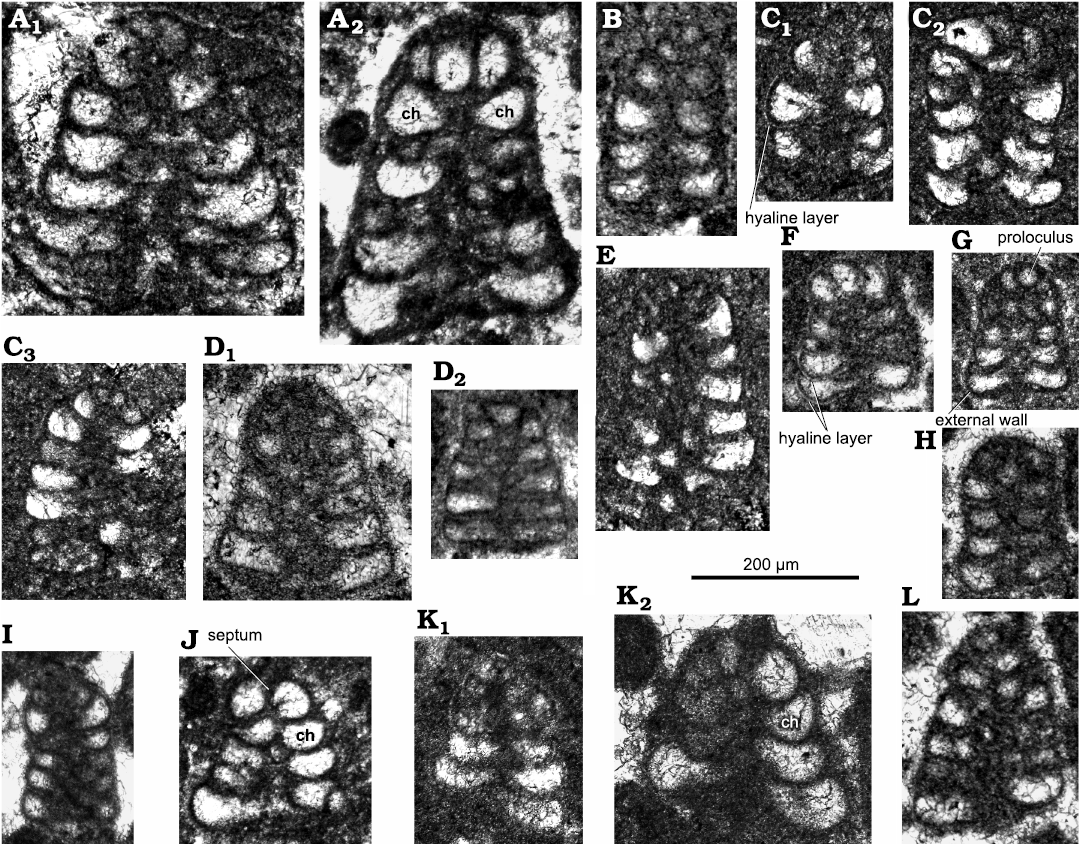
Fig. 3. Lituolinid foraminifer Campanellula capuensis De Castro, 1964, San Lorenzello section, Italy, upper Hauterivian–lower Barremian, Lower Cretaceous (A–J, L) and Križ section, Mljet Island, Croatia, Lower Cretaceous (K); various (sub)axial sections. A. Thin section DiSTAR-L29 with two individuals (A1, A2). B. Thin section DiSTAR-L8. C. Thin section DiSTAR-L39-2 with three individuals (C1–C3). D. Thin section DiSTAR-L36-1 with two individuals (D1, D2). E. Thin section DiSTAR-L39-1. F. Thin section DiST/SL90-91.1. G. Thin section DiSTAR-L35. H. Thin section DiSTAR-L36. I. Thin section DiSTAR-L41. J. Thin section DiST/SL99C.2. K. Thin section IGZ-MK with two individuals (K1, K2), after Husinec and Sokač (2006: fig. 7J and I). L. Thin section DiSTAR-L37. Rather large-sized specimens (A, K2) might belong to microspheric generation. Abbreviation: ch, chamber.
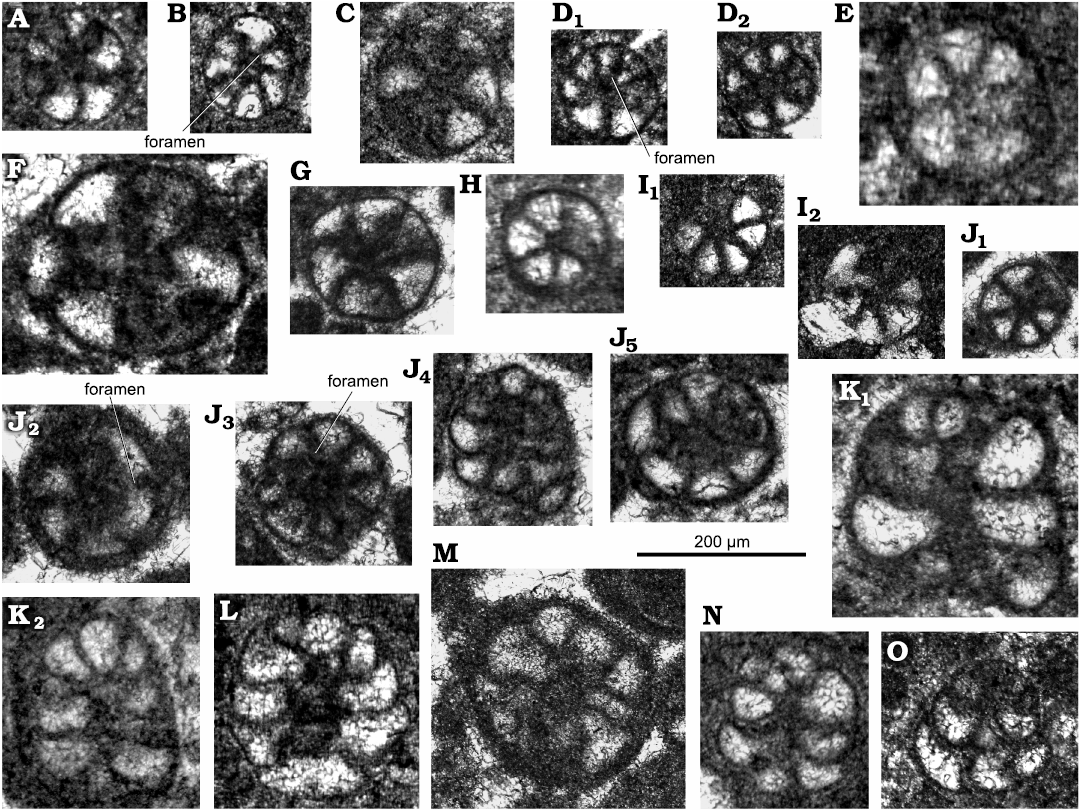
Fig. 4. Lituolinid foraminifer Campanellula capuensis De Castro, 1964, San Lorenzello section, Italy, upper Hauterivian–lower Barremian, Lower Cretaceous (A–L) and Križ section, Mljet Island, Croatia Hauterivian–lowermost Barremian, Lower Cretaceous (M–O); various transverse (A–J3, J5, M) and oblique (J4, K, L, N and O) sections. A. Thin section DiSTAR-L28. B. Thin section DiSTAR-L33-1. C. Thin section DiSTAR-L34. D. Thin section DiSTAR-L37 with two individuals (D1, D2). E. Thin section DiSTAR-L8. F. Thin section DiSTAR-L36-2. G. Thin section DiSTAR-L36. H. Thin section DiSTAR-L21-2. I. Thin section DiSTAR-L39-2 with two individuals (I1, I2). J. Thin section DiSTAR-L36-1 with five individuals (J1–J5). K. Thin section DiSTAR-L29 with two individuals (K1, K2). L. Thin section DiSTAR-L9-2. M–O. Thin sections IGZ-MK, after Husinec and Sokač (2006: unfigured specimens from Mljet Island, Croatia). Rather large-sized specimens (F and K1) might belong to microspheric generation.
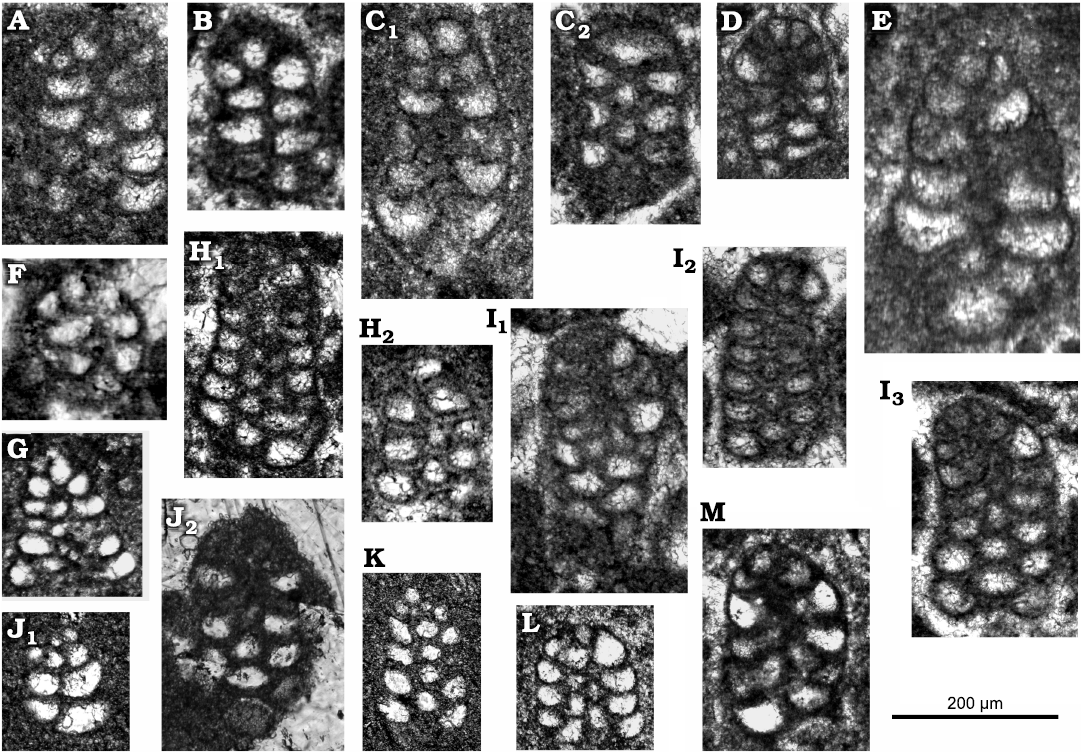
Fig. 5. Lituolinid foraminifer Campanellula capuensis De Castro, 1964, San Lorenzello section, Italy, upper Hauterivian–lower Barremian, Lower Cretaceous; various tangential sections. A. Thin section DiSTAR-L33-1. B. Thin section DiSTAR-L34. C. Thin section DiSTAR-L35 with two individuals (C1, C2). D. Thin section DiSTAR-L38-2. E. Thin section DiSTAR-L21-2. F. Thin section DiSTAR-L13. G. Thin section DiSTAR-L59. H. Thin section DiSTAR-L37 with two individuals (explain H1, H2). I. Thin section DiSTAR-L36-1 with three individuals (I1–I3). J. Thin section DiSTAR-L39-1 with two individuals (J1, J2). K. Thin section DiSTAR-L39-2. L. Thin section DiSTAR-L59. M. Thin section DiSTAR-L33-3. Rather large-sized specimen (E) might belong to microspheric generation.
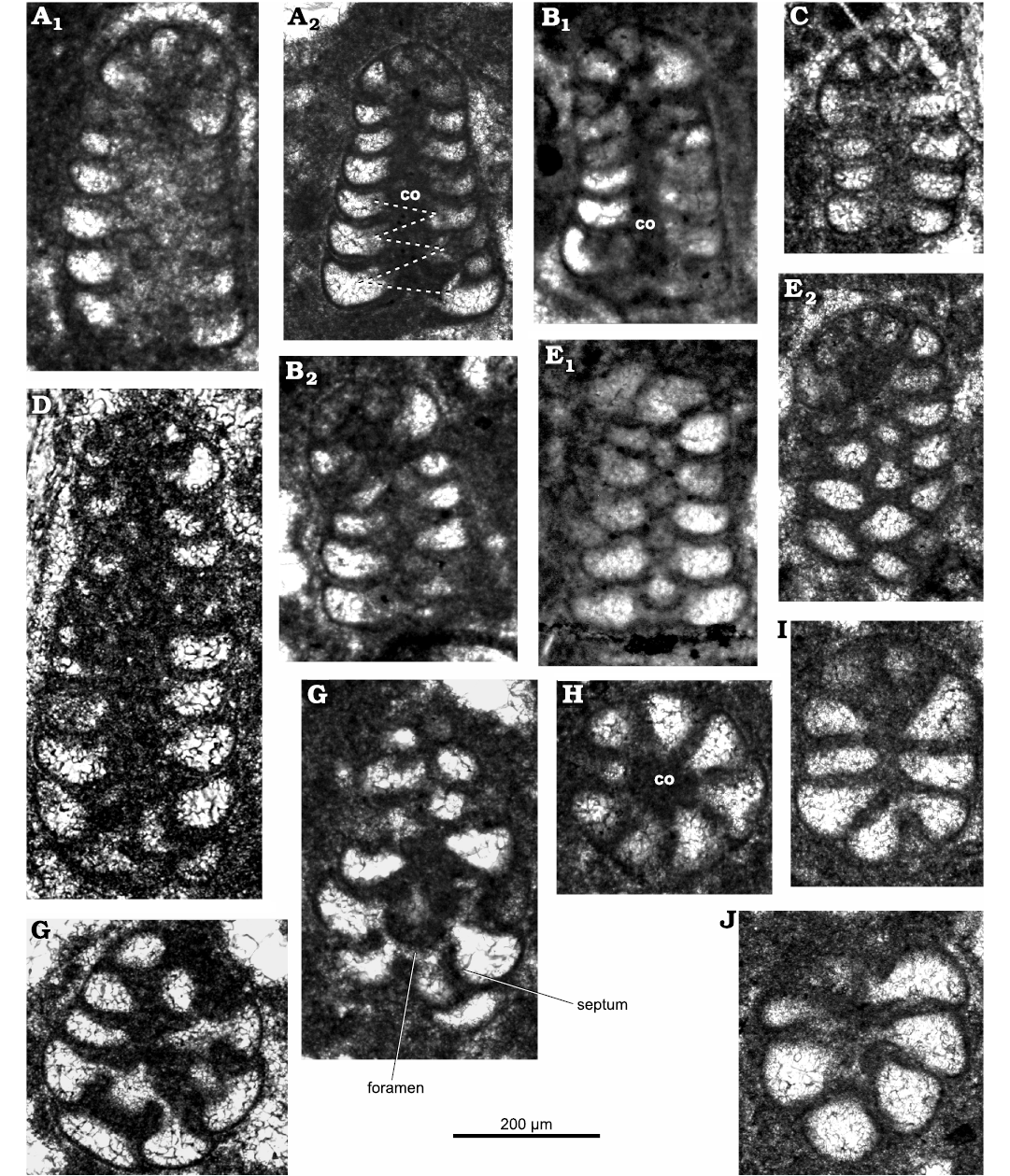
Fig. 6. Lituolinid foraminifer Campanellula capuensis De Castro, 1964, Castel Morrone, Italy, upper Hauterivian–lower Barremian, Lower Cretaceous. (Sub)axial sections (A–E), oblique sections (F, G, I, J), transverse section (H). A. Thin section DiSTAR-BA.4418.2 with two individuals (A1, A2); the dashed line in A2 points out alternated chambers resulting from the trochospiral arrangement. B. Thin section DiSTAR-BA.4418.3 with two individuals (B1, B2). C. Thin section DiSTAR-BA.4418.12. D. Thin section DiSTAR-BA.4418.15. E. Thin section DiSTAR-BA.4418.1 with two individuals (E1, E2). F. Thin section DiSTAR-BA.4418.13. G. Thin section DiSTAR-BA.4418.11. H. Thin section DiSTAR-BA.4418.3. I. Thin section DiSTAR-BA.4418.4. J. Thin section DiSTAR-BA.4418.2. Abbreviation: co, columella.
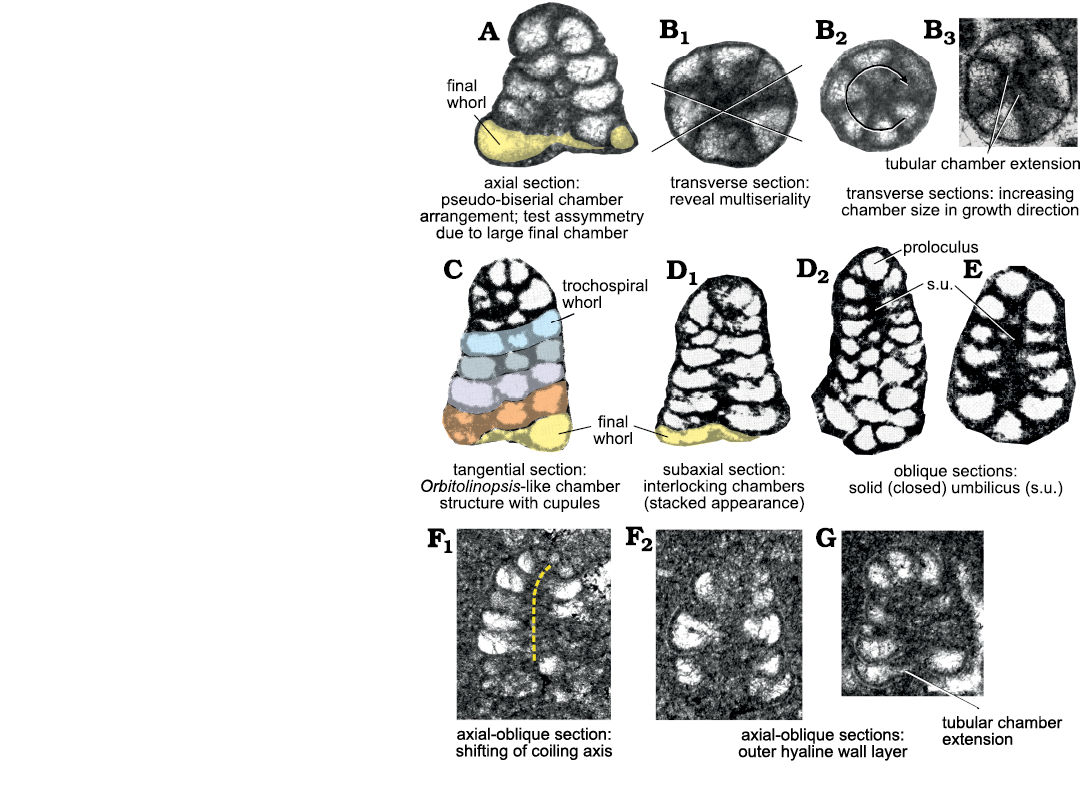
Fig. 7. Lituolinid foraminifer Campanellula capuensis De Castro, 1964, from the Lower Cretaceous of Italy, untangled; explanatory notes to the test structure based on different sections (not to scale). A, B. San Lorenzello section. A. Thin section DiST-SL99C.1, upper Hauterivian. B. Thin section DiSTAR-L36.1, lower Barremian, with three different individuals (B1, B2, B3). C. Thin section IP-A.679.1 (from De Castro 1964: pl. 1: 10), paratype, Cava Grande, Punta Orlando, Hauterivian–Barremian. D. Thin section IP-A.281.5 with two different individuals (D1, D2) (from De Castro 1964: pl. 1: 3, 5), paratype, Castel Morrone, Caserta, Hauterivian–Barremian. E. Thin section IP-A.85.3 (from De Castro 1964: pl. 1: 14), Monte La Foresta, Salerno, Hauterivian–Barremian. F, G. San Lorenzello section. F. Thin section DiSTAR-39.2 with two different individuals (F1, F2), lower Barremian; dashed line in F1 indicates the coiling axis;. G. Thin section DiST-SL90/91.1, upper Hauterivian.
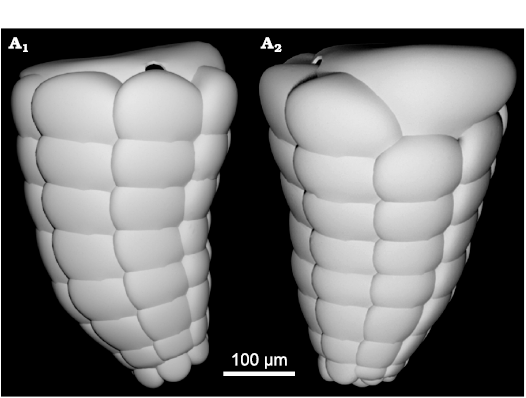
Fig. 8. Lituolinid foraminifer Campanellula capuensis De Castro, 1964, from Castel Morrone, Italy, upper Hauterivian–lower Barremian, Lower Cretaceous. IP-A.281.1, 3D test reconstruction of the holotype (De Castro 1964: pl. 1: 1:). Left (A1) and frontal right (A2) views of the test.
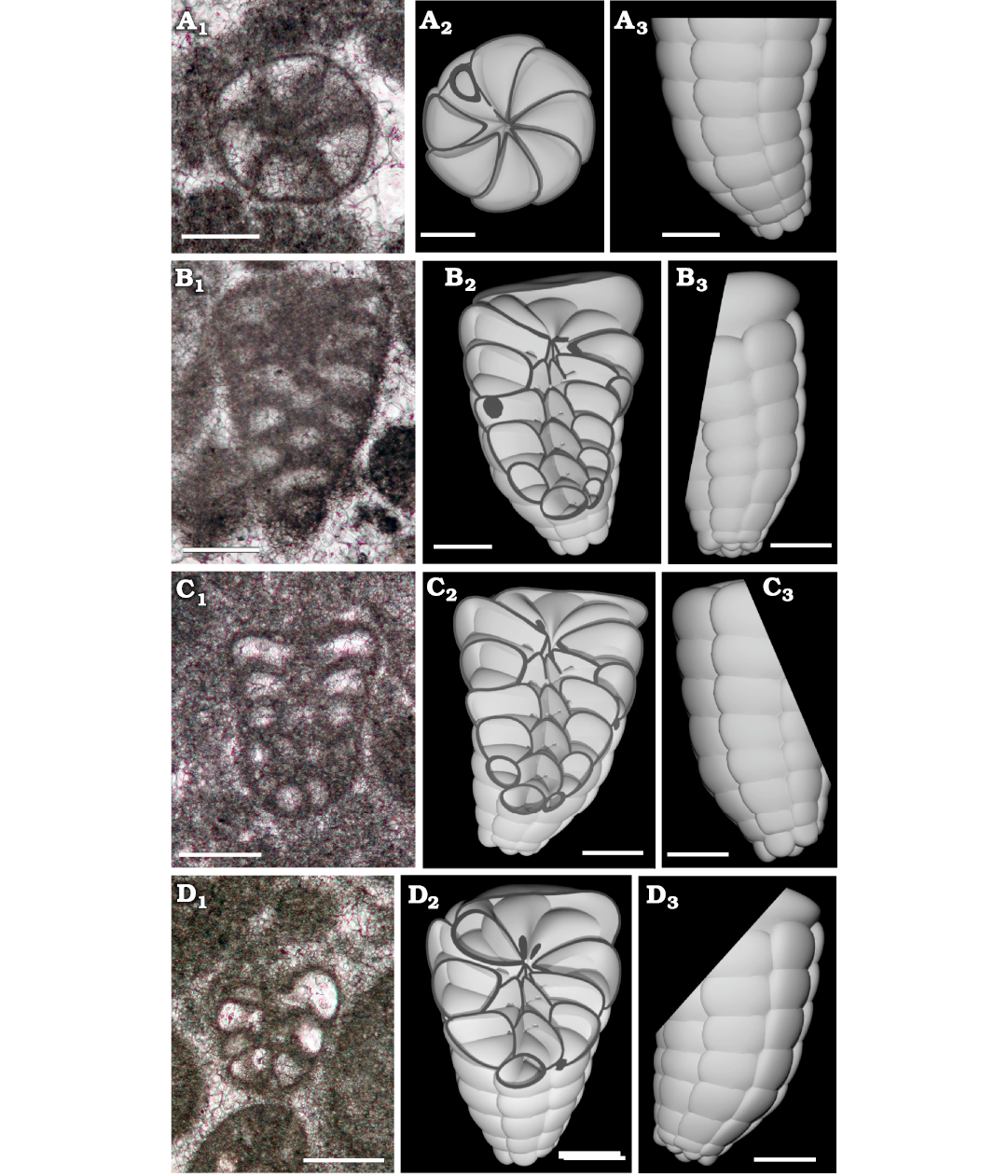
Fig. 9. Lituolinid foraminifer Campanellula capuensis De Castro, 1964, from Castel Morrone, Italy, upper Hauterivian–lower Barremian, Lower Cretaceous. A. DiSTAR-L36.1, transversal section showing six chambers per whorl (A1), compared with the 3D model transversally cut (upper view displaying eight chamber per whorl (A2), also represented in lateral view (A3). B. DiSTAR-L36.1 tangential section (B1), compared with the 3D model tangentially cut (frontal view of the section, B2), also represented in lateral view (B3). C. DiSTAR-L35.1, tangential-oblique section (C1), compared with the 3D sectioned model (cut in frontal view, C2), also represented in lateral view (C3). D. DiSTAR-L34.1, oblique section through last tours (D1), compared with the 3D sectioned model (cut in frontal view, D2), also represented in lateral view (D3). Scale bars 100 μm.
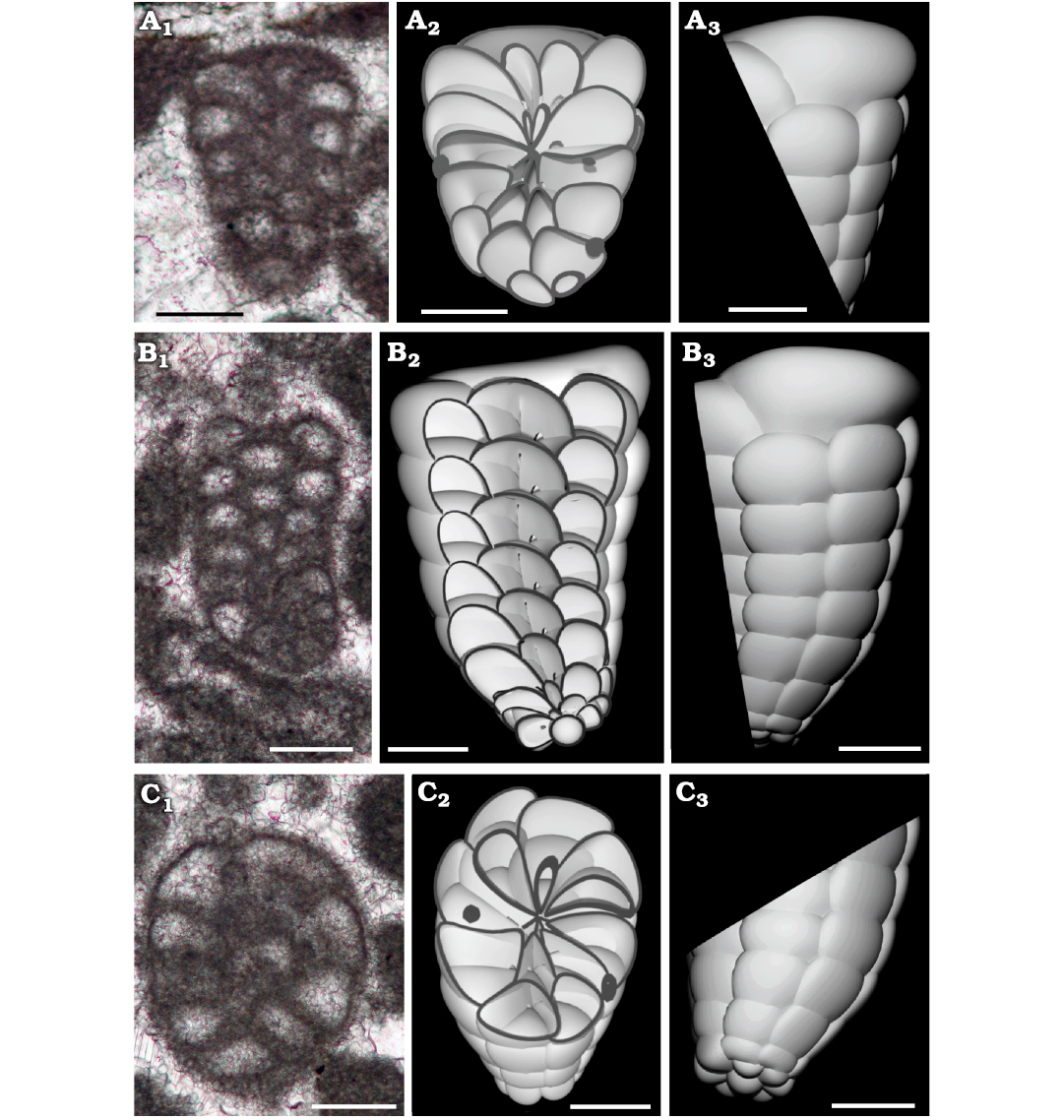
Fig. 10. Lituolinid foraminifer Campanellula capuensis De Castro, 1964, from Castel Morrone, Italy, upper Hauterivian–lower Barremian, Lower Cretaceous. A. DiSTAR-L36.1, oblique section showing (A1), compared with the 3D sectioned model (cut in frontal view, A2), also represented sideways (A3). B. DiSTAR-L36.1, longitudinal section (B1), compared with the 3D sectioned model (cut in frontal view, B2), also represented sideways (B3). C. DiSTAR-L36.1, oblique section (C1), compared with the 3D sectioned model (cut in frontal view, C2), also represented sideways (C3). Scale bars 100 μm.
3D rendering.—A three-dimensional model has been performed to obtain detailed insight of the perspective representation of the chamber arrangement. The reconstruction is based primarily on the holotype (De Castro 1964: pl. 1: 1) for the general shape in axial section. Characters such as number, shape, and size of chambers per tour are inferred by the analysis of numerous transversal and oblique sections. In Fig. 8 the reconstruction is rendered in left view (Fig. 8A) and frontal right view (Fig. 8B), positioning the early stage downwards and inner side of the curvature to be frontal. Afterwards, the 3D model has been “sliced” and the resulting “slices” compared with similar sections from the San Lorenzello material (Figs. 9, 10). However, it is a matter of fact that variability in size and shape of specimens will never allow a perfect correspondence between a sectioned specimen and a “slice” of a 3D model. Nevertheless, if the reconstruction accurately follows the constrains implied by specimens in thin section, the model can be rather faithful to the original and “slicing” predict internal structure of the foraminifer.
Remarks.—When describing Campanellula, De Castro (1964) remarked the rather low trochospirally coiled chambers in successive tight whorls and their slow increase in numbers during ontogeny. Due to this observation, De Castro (1964: 55) “did not exclude the possibility that individual whorls might exhibit uniserially-arranged chambers” (“non si esclude …. che vi possa essere qualche giro con camere disposte in serie” in the original Italian text). Further discussion on the test construction of Campanellula and the deriving suprageneric classification goes back to Schroeder in Macoin et al. (1970). In this work, specimens of a conical foraminifer attributed to Campanellula capuensis are described from the Lower Cretaceous of Algeria showing a different morphology. Its attribution to the orbitolinid genus Orbitolinopsis Henson, 1948, was based on an interpretation of the adult chambers that should be arranged rectilinear and not trochospirally coiled. In fact, the tangential sections illustrated by Macoin et al. (1970: pl. 2: 1–4, 5, 8) display an oblique cross-wise arrangement of the foramina connecting successive rectilinear chambers as in Orbitolinopsis (see also Arnaud-Vanneau 1980). Therefore, the authors introduced the new combination Orbitolinopsis capuensis (De Castro, 1964) included in the family Orbitolinidae and subfamily Dictyoconinae. As Schroeder in Macoin et al. (1970: 255) correctly stated, such an arrangement contradicts a trochospirally coiled test where direction of both coiling and foramina are the same. Such an arrangement of foramina has not been observed in the material studied from the type-area where a throughout trochospirally coiled chamber arrangement is obvious. Many of the transverse sections of Macoin et al. (1970) are equivalent to the Italian material indicating a trochospiral chamber arrangement often with the last and greatest chamber sectioned accounts for a slightly irregular (= not perfectly rounded) outline. The Algerian specimens are herein interpreted as being different in exhibiting a trochospirally coiled early and rectilinear adult parts, so that an investigation of the Algerian material is needed for further clarification.
The main differences between Campanellula capuensis De Castro, 1964, and Campanellula herishtensis Schlagintweit, Rashidi, and Hanifzadeh, 2019, are the morphology and size of the test. Campanellula capuensis displays a higher conical test and a greater size (Fig. 11).
Stratigraphic and geographic range.—Campanellula capuensis is mostly widespread in the Adriatic Plate (Apulian Plate sensu Stampfli and Borel 2004; Fig. 12). Campanellula herishtensis is restricted to the type locality (late Aptian, Central Iran).
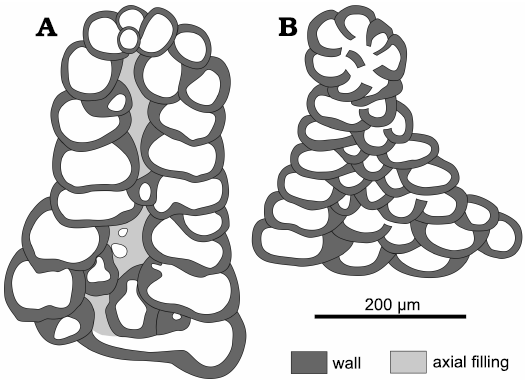
Fig. 11. Scheme of the species of Campanellula De Castro, 1964. A. Campanellula capuensis De Castro, 1964, IP-A.281.1, holotype (drawing from De Castro 1964: pl. 1: 1), Castel Morrone, Italy, upper Hauterivian–lower Barremian, Lower Cretaceous. B. Campanellula herishtensis Schlagintweit, Rashidi, and Hanifzadeh, 2019, T 72-6 (number Gmm 13950F110), holotype (drawing from Schlagintweit et al. 2019: fig. 5L), Ardakan, Province of Yazd, Central Iran, lower Gargasian, Aptian, Lower Cretaceous.
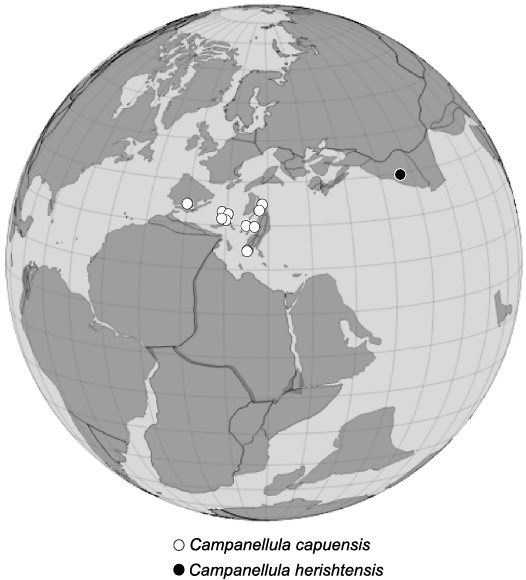
Fig. 12. Palaeobiogeographic distribution of species of Campanellula De Castro, 1964. Occurrence of species has been plotted on an Early Cretaceous (ca.120 Ma) paleogeographic map, after http://portal.gplates.org/.
Discussion
With the assumed orbitolinid test morphology including uniserial adult and subdivided chambers (see Macoin et al. 1970), Campanellula was (Loeblich and Tappan 1987), and still is included in the order Loftusiida Kaminski and Mikhalevich, 2004, suborder Orbitolinina Kaminski, 2004, respectively (Kaminski 2014) (Table 1). Based on the throughout trochospirally coiled test with simple chambers (De Castro 1964; this work), Campanellula can be included into the order Lituolida Lankester, 1885, that comprises all non-canaliculate taxa with well-defined chambers (at least in the adult stage), and a simple imperforate wall (Kaminski 2004: 240). The current classification and two further options, all based on Kaminski (2004, 2014) are summarized in Table 1. These options refer to the primary importance of the wall structure, combined with the overall test morphology (e.g., coiling type), chamber arrangement, and the presence/absence of internal chamber subdivisions. Another approach, however, considers the general test architecture of prime importance as for example proposed by Septfontaine (1988) for the Mesozoic agglutinated (larger) benthic foraminifera. With some characters (e.g., test shape and trochospiral coiling, high number of chambers per whorl) common to taxa not directly phylogenetically related like Praechrysalidina Luperto Sinni, 1979, Kilianina Pfender, 1933, Neokilianina Septfontaine, 1988, Campanellula could then also be placed within the Textulariida Delage and Hérouard, 1896.
Table 1. The Campanellula De Castro, 1964, in the context of current classifications of agglutinated Foraminifera (Pawlowski et al. 2013; Kaminski 2014). * not Moullade (1965) as indicated in Kaminski (2014).
|
Campanellula De Castro, 1964 |
|||
|
Classifications |
Current (Kaminski 2014) |
Option 1 |
Option 2 |
|
Phylum |
Foraminifera d’Orbigny, 1826 |
||
|
Class |
Globothalamea Pawlowski, Holzmann, and Tyszka, 2013 |
||
|
Subclass |
Textulariana Mikhalevich, 1980 |
||
|
Order |
Loftusiida Kaminski and Mikhalevich in Kaminski, 2004 |
Lituolida Lankester, 1885 |
|
|
Remark |
An inclusion into the order
Textulariida Delage and Hérouard, 1896, is excluded due to the
lack of a canaliculated wall |
||
|
Suborder |
Orbitolinina Kaminski, 2004 |
Ataxophragmiina Fursenko, 1958 |
Verneuilinina Mikhalevich and |
|
Definition |
Test trochospiral or conical, later stage may have reduced number of chambers per whorl, or may become uniserial and rectilinear; chamber interior of advanced taxa subdivided by vertical or horizontal exoskeletal partitions or both, by radial or transverse partitions, or with interseptal pillars (Kaminski 2004). |
Test free or attached, typically high trochospiral with an asymmetrical spire, may have a conical form, or have elongated vertical chambers inclined to previous whorls; coiling may reduce to biserial or uniserial; chambers may have complex inner structure (buttresses, pillars, radial partitions, columella), or form chamberlets; wall agglutinated typically with calcareous cement; aperture basal, a wide horizontal slit at the base of the apertural face of the last chamber, may be subdivided by a median lobe (Kaminski 2014). |
Test high trochospiral throughout or only in the initial part, later part may have an increased or decreased number of chambers per whorl or may become uniserial or cyclical; wall simple; aperture basal at least initially, later may become terminal, single or multiple, some genera with inner apertural structures (Kaminski 2004). |
|
Remarks |
“The predominantly high trochospiral to conical Ataxophragmiina and Orbitolinina possess internal partitions and interseptal pillars” (Kaminski 2004: 241). |
“The Trochamminina… comprise the low trochospirally coiled forms, while the Verneuilinina encompass the high trochospiral genera with simple walls” (Kaminski 2004: 241). |
|
|
Superfamily |
Orbitolinoidea Martin, 1890 |
not assignable |
not assignable |
|
Remark |
Orbitolinoidea do not comprise taxa with trochospiral chambers throughout (Loeblich and Tappan 1987). |
no remarks |
no remarks |
|
Family |
Orbitolinidae Martin, 1890 |
not assignable |
not assignable |
|
Subfamily |
Dictyoconinae Schubert, 1912* |
not assignable |
not assignable |
The order Textulariida is defined by taxa that possess a canaliculate wall (Kaminski 2014). It must be noted that the presence of a pseudokeriotheca (canaliculate wall) and the related possibility for observation may be related to the size of the test and the thickness of the marginal chamber wall (Vachard et al. 2004), as very small taxa like Campanellula or Kilianina (without pseudokeriotheca) have a height (vertical axis) 5–10 times less than e.g., Redmondoides lugeoni (Septfontaine, 1977) or Chrysalidina gradata d’Orbigny, 1839, both exhibiting canaliculate (pseudokeriothecal) wall (De Castro 1981). A suprageneric placing in a basket along with larger taxa aside Praechrysalidina–Chrysalidina (e.g., the informal morphogroup of the valvulinids, Septfontaine 1988, 2020) might only be a temporary solution. The question of the importance (and position) of the wall (canaliculate or not) is still pendant, and not obvious in the classification of Kaminski (2004) as “alveolar” or “canaliculate” or “non alveolar” (like Kilianina and Campanellula). These terms are not clearly defined and the corresponding morphotypes variously distributed in the different suprageneric taxa (order, sub-order) without evolutive justification. In conclusion, the suprageneric position of Campanellula is herein intentionally left open. It is concluded that Campanellula has to be removed from the order Loftusiina and the suborder Orbitolinina, instead it should be included into the order Lituolida and the suborder Verneuilinina (Table 1).
Conclusions
This work presents a comparison between the Campanellula capuensis material from the type locality (De Castro 1964) from which an original 3D model has been reconstructed and additional material was sampled at a new site of San Lorenzello (Amodio et al. 2020). This procedure unequivocally defines that Campanellula capuensis exhibits a trochospiral structure that clearly persists in all stages of shell growth, thus reinstating the original description provided by De Castro (1964). One morphological question still remains open and concerns the rectilinear evolution of the adult portion of the shell, which is only present in the Algerian specimens of Macoin et al. (1970) and which appears, at present, to be the only substantial difference from the material studied in this work. This feature would be worth considering but it is beyond the scope of the present paper. Based on our results and the present taxonomic concept of agglutinated benthic foraminifera, the genus Campanellula is excluded from the order Loftusiina, suborder Orbitolinina respectively, instead it is assigned to the order Lituolida, suborder Verneuilinina.
Acknowledgements
FB has performed the 3D reconstruction and expresses his gratitude to architect Maria Rita Cotronei (Istituto Volta, Bari, Italy) for the guidance and suggestions on the 3D software. Thanks to Antun Husinec (St. Lawrence University, New York, USA) for providing images of Campanellula capuensis from Croatia. The manuscript benefited from the valuable comments of the two reviewers Ioan Bucur (Babeş-Bolyai University, Cluj-Napoca, Romania) and Jarosław Tyszka (Institute of Geological Sciences, Polish Academy of Sciences, Kraków, Poland). FB and SA (the corresponding author) acknowledge funding by the Italian MUR under grant PRIN 2017RX9XXY “Biota resilience to global change: biomineralization of planktic and benthic calcifiers in the past, present and future”.
References
Amodio, S. 2006. Foraminifera diversity changes and paleoenvironmental analysis: the Lower Cretaceous shallow-water carbonates of San Lorenzello, Campanian Apennines, southern Italy. Facies 52: 53–67. Crossref
Amodio, S., Barattolo, F., and Martino, M. 2020. Carbonate ecosystem and depositional environment during the Valanginian–Barremian: Discussion on possible controlling factors in the Apennine Carbonate Platform record (Italy). Cretaceous Research 108: 104351. Crossref
Amodio, S., Barattolo, F., and Parente, M. 2023. The Early Cretaceous C-isotope record of shallow-marine carbonates: a transect across southern Alps, Apennines and Dinarides. Rendiconti OnLine, Società Geologica Italiana 59: 28–34. Crossref
Amodio, S., Barattolo, F., and Riding, R. 2018. Early Cretaceous dendritic shrub-like fabric in karstified peritidal carbonates from southern Italy. Sedimentary Geology 373: 134–146. Crossref
Amodio S., Ferreri V., and D’Argenio, B. 2013. Lower Cretaceous carbon-isotope records of M. Faito and S. Maria sections (central-southern Italy). Platform to basin correlation and orbital calibration. Journal of Mediterranean Earth Sciences 5: 3–6.
Amodio, S., Ferreri, V., D’Argenio, B., Weissert, H., and Sprovieri, M. 2008. Carbon-isotope stratigraphy and cyclostratigraphy of shallow-marine carbonates: the case of San Lorenzello, Lower Cretaceous of southern Italy. Cretaceous Research 29: 803-813. Crossref
Arnaud-Vanneau, A. 1980. L´Urgonien du Vercors septentrional et de la Chartreuse. Géologie Alpine Mémoire 11: 1–874.
Arnaud-Vanneau, A. and Sliter, W.V. 1995. Early Cretaceous shallow-water benthic foraminifers and fecal pellets from Leg 143 compared with coeval faunas from the Pacific Basin, Central America, and the Tethys. In: E.L. Winterer, W.W. Sager, J.V. Firth, and J.M. Sinton (eds.), Proceedings of the Ocean Drilling Program, Scientific Results 143: 537–564. Crossref
Barattolo, F. 1982. Osservazioni su Triploporella praturlonii n. sp. (alghe verdi, Dasicladali), del cretacico inferiore dell’Appennino centrale. Geologica Romana 21: 825–847.
Barattolo, F., Amodio, S., Bucur, I.I., and Martino, M. 2021. Selliporella johnsonii (Praturlon) nov. comb. and Selliporella neocomiensis (Radoičić) (green algae, Dasycladales), taxonomic reconsideration and chronostratigraphic calibration. Cretaceous Research 104848. Crossref
Bruni, R., Bucur, I.I., and Preat, A. 2007. Uppermost Jurassic–Lower Cretaceous carbonate deposits from Fara San Martino (Maiella; Italy): biostratigraphic remarks. Studia Universitatis Babeş-Bolyai, Geologia 52 (2): 45–54. Crossref
Bucur, I.I., Conrad, M.A., and Radoičić, R. 1995. Foraminifers and calcareous algae from Valanginian limestones in the Jerma River Canyon, eastern Serbia. Revue de Paléobiologie 14: 349–377.
Carras, N. 1995. La piattaforma carbonatica del Parnasso durante il Giurassico Superiore–Cretaceo inferiore [in Greek with Italian abstract]. 232 pp. Despoina Mavromatis Edition, Athens.
Carras, N., Conrad, M.A., and Radoičič, R. 2006. Salpingoporella, a common genus of Mesozoic Dasycladales (calcareous green algae). Revue de Paléobiologie 25: 457–517.
Cherchi, A. and Schroeder, R. 1973. Sur la biogéographie de l’association à Valserina du Barrémien et la rotation de la Sardaigne. Comptés Rendus de l’Académie des Sciences de Paris 277: 829–956. Crossref
Cherchi, A. and Schroeder, R. 1978. Osservazioni sul gen. Orbitolinopsis Silvestri (Foraminiferida) e sua presenza nel Barremiano della Sardegna. Bollettino della Società Sarda di Scienze Naturali 17: 159–167.
Cherchi, A. and Schroeder, R. 2010. Relazioni paleobiogeografiche nel dominio catalano-sardo-provenzale nell’intervallo Giurassico Cretacico. In: A. Cherchi, L. Simone, R. Schroeder, G. Carannante, and A. Ibba (eds.), I Sistemi carbonatici giurassici della Sardegna Orientale (G. di Orosei) ed eventi deposizionali nel sistema carbonatico giurassico-cretacico della Nurra (Sardegna Nord Occidentale). Geological Field Trips , parte II, 2 (2.1), 86–89. ISPRA and SGI, Roma.
Cherchi, A., Durand-Delga, M., and Schroeder, R. 1981. Aperçu paléogéographique sur les Provinces à grands Foraminifères du Crétacé Inférieur dans le Cadre structural Méditerranéen. Rapports et procès-verbaux des réunions de la Commission Internationale pour L’Exploration Scientifique de la Mer Méditerranée 27: 115–119.
Chiocchini, M. and Mancinelli, A. 1977. Microbiostratigrafia del Mesozoico in facies di piattaforma carbonatica dei Monti Aurunci (Lazio meridionale). Studi Geologici Camerti 3: 109–152.
Chiocchini, M., Chiocchini, R.A., Didaskalou, P., and Potetti, M. 2008. Ricerche micropaleontologiche e biostratigrafiche sul Mesozoico della piattaforma carbonatica laziale-abruzzese (Italia centrale). Memorie Descrittive della Carta Geologica d’Italia 84: 5–170.
Chiocchini, M., Farinacci, A., Mancinelli, A., Molinari, V., and Potetti, M. 1995. Biostratigrafia a foraminiferi, dasicladali e calpionelle delle successioni carbonatiche mesozoiche dell’Appennino centrale (Italia). In: A. Mancinelli (ed.), Biostratigrafia dell’Italia centrale, coord. A. Farinacci, Studi Geologici Camerti, special volume 1994 (A): 9–128.
Chiocchini, M., Pampaloni, L., and Pichezzi, R.M. 2012. Microfacies e microfossili delle succession carbonatiche mesozoiche Lazio e Abruzzo. Memorie per Servire alla descrizione della Carta Geologica d´Italia 17: 1–267.
Claps, M., Parente, M., Neri, C., and Bosellini, A. 1996. Facies and cycles of the S. Giovanni Rotondo Limestone (Lower Cretaceous, Gargano Promontory, Southern Italy): The Borgo Celano Section. Annali Università di Ferrara supplemento 7: 1–35.
Cruz Abad, E. 2018. Textura y arquitectura de los orbitolinoideos (superfamilia Orbitolinoidea): revisión y caracterización. 154 pp. Doctoral thesis Departamento de Geología de la Universitat Autònoma de Barcelona, Barcelona.
D’Argenio, B., Ferreri, V., Amodio, S., and Pelosi, N. 1997. Hierarchy of high-frequency orbital cycles in Cretaceous carbonate platform strata. Sedimentary Geology 113: 169–193. Crossref
De Castro, P. 1964. Su di un nuovo foraminifero del Cretacico inferiore dell’Appennino meridionale. Bollettino della Società dei Naturalisti in Napoli 73: 55–61.
De Castro, P. 1981. Osservazioni su Chrysalidina gradata d’Orbigny 1839 (Foraminiferida) dell’Ile Madame (Francia). Atti dell’Accademia Pontaniana (n.s.) 30: 1–25.
De Castro, P. 1991. Mesozoic. In: F. Barattolo, P. De Castro, and M. Parente (eds.), Field Trip Guide-book, “5th International Symposium on Fossil Algae”, 21–38. Giannini editore, Napoli.
Douglass, R.C. 1960. Revision of the family Orbitolinidae. Micropaleontology 6: 249–270. Crossref
Ferreri, V., Amodio, S., Sandulli, R., and D’Argenio, B. 2004. Orbital chronostratigraphy of the Valanginian–Hauterivian boundary. A cyclostratigraphic approach. In: B. D’Argenio, A.G. Fischer, I. Premoli Silva, H. Weissert, and V. Ferreri (eds.), Cyclostratigraphy: Approaches and Case Histories. SEPM Tulsa, Special Publication 81: 153–166. Crossref
Frijia, G., Di Lucia, M., Vicedo, V., Günter, Ch., Ziemann, M.A., and Mutti, M. 2012. An extra-ordinary single-celled architect: A multi-technique study of the agglutinated shell of the larger foraminifer Mesorbitolina from the Lower Cretaceous of southern Italy. Marine Micropaleontology 90–91: 60–71. Crossref
Garcia-Hernandez, M. 1978. El Juràsico terminal y el Cretàcico inferior en las Sierras de Cazorla y del Segura (Zona prébetica). Tesis Doctorales n.190, 1–334, Ph.D. Thesis, University of Granada, Granada.
Gheiasvand, M., Föllmi, K., Arnaud-Vanneau, A., Adatte, T., Spangenberg, J., Ghaderi, A., and Ashouri, A.R. 2019. New stratigraphic data for the Lower Cretaceous Tirgan Formation, Kopet-Dagh Basin, NE Iran. Arabian Journal of Geosciences 12: art. 142. Crossref
Henson, F.R.S 1948. Larger imperforate Foraminifera of south-western Asia. Families Lituolidae, Orbitolinidae and Meandropsinidae. Monograph British Museum (Natural History) 1948: i–xii + 1–128.
Hottinger, L. and Drobne, K. 1980. Early Tertiary conical imperforate foraminifera. Razprave IV razreda SAZU 22: 188–276.
Husinec, A. and Read, J.F. 2018. Cyclostratigraphic and δ13C record of the Lower Cretaceous Adriatic Platform, Croatia: Assessment of Milankovitch-forcing. Sedimentary Geology 373: 11–31. Crossref
Husinec, A. and Sokač, B. 2006. Early Cretaceous benthic associations (foraminifers and calcareous algae) of a shallow tropical-water platform environment (Mljet Island, southern Croatia). Cretaceous Research 27: 418–441. Crossref
Kaminski, M.A. 2004. The year 2000 classification of the agglutinated foraminifera. In: M. Bubik, and M.A. Kaminski (eds.), Proceeding of the 6th international workshop on agglutinated foraminifera. Grzybowski Foundation Special Publication 8: 237–255.
Kaminski, M.A. 2014. The year 2010 classification of the agglutinated foraminifera. Micropaleontology 60: 89–108. Crossref
Loeblich, A.R. Jr. and Tappan, H. 1987. Foraminiferal Genera and Their Classification. Vol. 1 + 2, 970 pp. + 847 pls. Van Nostrand Reinhold, New York. Crossref
Luperto Sinni, E. and Masse, J.P. 1984. Données nouvelles sur la micropaléontologie et la stratigraphie de la partie basale du “Calcare di Bari” (Crétacè inférieur) dans la région des Murges (Italie méridionale). Rivista Italiana di Paleontologia e Stratigrafia 90: 331–374.
Luperto Sinni, E. and Masse, J.P. 1993. Specie nuove di foraminiferi bentonici dell’Aptiano inferiore carbonatico delle Murge (Italia Meridionale). Rivista Italiana di Paleontologia 99: 213–224.
Macoin, P., Schroeder, R., and Vila, J.M. 1970. Campanellula capuensis De Castro (Foram.), position systematique et répartition stratigraphique en Algerie. Actes du IV Colloque Africain de Micropaléontologie 1970: 248–260.
Malinverno, A. and Ryan, W.B.F. 1986. Extension in the Tyrrhenian Sea and shortening in the Apennines as result of arc migration driven by sinking of the lithosphere. Tectonics 5: 227–245. Crossref
Martino, M., Barattolo, F., and Amodio, S. 2019. Early Cretaceous neritic carbonate community and facies response to palaeonvironmental and sea-level changes: new evidences from the Apennine Carbonate Platform of southern Italy. In: B. Granier (ed.), JK2018 International Meeting Around Jurassic/Cretaceous Boundary. Carnets Géologie Book 2019/01: 55–58.
Moullade, M. 1965. Contribution au problème de la classification des Orbitolinidae (Foraminiferida, Lituolacea). Comptes rendus de l'Académie des Sciences 260: 4031–4034.
Parente, M., Amodio, S., Iannace, A., and Sabbatino, M. 2022. Stratigraphy and facies of the Apennine Carbonate Platform (southern Italy): the record of Mesozoic OAEs and Miocene transgression. Geological Field Trips and Maps 14/2.3: 1–75. Crossref
Patacca, E. and Scandone, P. 2007. Geology of the Southern Apennines. Bollettino della Società Geologica Italiana Special Issue 7: 75–119.
Pawlowski, J., Holzmann, M., and Tyszka, J. 2013. New supraordinal classification of Foraminifera: Molecules meet morphology. Marine Micropaleontology 100: 1–10. Crossref
Peybernès, B. 1982. Les orbitolinides Crétacés d’Afrique: essai de synthèse. 8éme Colloque Africain de Micropaléontologie, Paris 1980. Cahiers de Micropaléontologie 2: 13–28.
Peybernès, B., Ciszak, R., Cugny, P., and Damotte, R.R. 1984. Le Crétacé inférieur et moyen des Monts de Daïa (Oranie, Algérie occidentale): analyse micropaléontologique et paléoécologique de l’intervalle Barrémien–Cénomanien. Géologie Méditerranéenne 11 (1–2): 147–158. Crossref
Rat, P. 1963. L’accroissement de taille et les modifications architecturales corrélatives chez les orbitolines. In: G.H.B. Koenigswald, J.D. Emeis, W.L. Buning, and C.W. Wagner (eds.), Evolutionary Trends in Foraminifera, 93–109. Elsevier, Amsterdam.
Schindler, U. and Conrad, M.A. 1994. The Lower Cretaceous Dasycladales from the northwestern Friuli Platform and their distribution in chronostratigrapohic and cyclostratigraphic units. Revue de Paléobiologie 13: 59–96.
Schlagintweit, F. 2023. Annular chambers in Cretaceous Orbitolinidae (Larger Benthic Foraminifera): An overview. Acta Palaeontologica Romaniae 19: 45–52. Crossref
Schlagintweit, F., Rashidi, K., and Barani, F. 2016. First record of Gyroconulina columellifera Schroeder & Darmoian, 1977 (larger benthic foraminifera) from the Maastrichtian Tarbur Formation of SW Iran (Zagros Fold-Thrust-Belt). Geopersia 6 (2): 169–185.
Schlagintweit, F., Rashidi, K., and Hanifzadeh, R. 2019. Campanellula herishtensis n. sp. (Foraminiferida; Orbitolinidae?) from the upper Aptian (Gargasian) of Central Iran. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 294: 251–161. Crossref
Schlagintweit, F., Rigaud, S., and Wilmsen, M. 2015. The benthic foraminifer Altamirella biscayana n. gen., n. sp. from the lower Cenomanian of Cantabria, N-Spain: A post-Palaeozoic fusulinan? Cretaceous Research 52: Part A, 1–8. Crossref
Schroeder, R. 1985. Genre Simplorbitolina Ciry & Rat, 1953. In: R. Schroeder and M. Neumann (eds.), Les Grands Foraminifères du Crétacé Moyen de la région Méditerranénne. Géobios Mémoire Special 7: 46–50.
Schroeder, R., Cherchi, A., Guellal, S., and Vila, J.M. 1978. Biozonation par les grands Foraminifères du Jurassique supérieur et du Crétacé inférieur et moyen des séries néritiques en Algérie du Nord-Est. Considérations paléobiogéographiques. Annales Mines Géologique 28 (2): 243–253.
Schubert, R.J. 1912. Über Lituonella und Coskinolina liburnica Stache sowie deren Beziehungen zu den anderen Dictyoconinen. Jahrbuch der K. K. Geologischen Reichsanstalt 62 (2): 195–208.
Septfontaine, M. 1988. Vers une classification évolutive des Lituolidés (foraminifères) jurassiques en milieu de plate-forme carbonatée. Revue de Paléobiologie, vol. spéc. 2: 229–256.
Septfontaine, M. 2020. Steps of morphogenesis and iterative evolution of imperforate larger foraminifera in shallow carbonate shelves during Mesozoic times: Possible relations to symbiotic and abiotic factors. In: J. Guex, J.S. Torday, and W.B. Miller (eds.), Morphogenesis, Environmental Stress and Reverse Evolution, 129–173. Springer International Publishing, Berlin. Crossref
Sokač, B. and Velić, I. 1978. Biostratigraphic investigations of the Lower Cretaceous of the Outer Dinarides I. The Neocomian of western Istria. Geološki vjesnik 30: 243–250.
Stampfli, G.M. and Borel, G.D. 2004. The TRANSMED Transects in Space and Time: Constraints on the Paleotectonic Evolution of the Mediterranean Domain. In: W. Cavazza, F. Roure, W. Spakman, G.M. Stampfli, and P.A. Ziegler (eds.), The TRANSMED Atlas. The Mediterranean Region from Crust to Mantle, 53–80. Springer, Berlin. Crossref
Vachard, D., Munnecke, A., and Servais, T. 2004. New SEM observations of keriothecal walls: implications for the evolution of Fusulinida. Journal of Foraminiferal Research 34: 232–242. Crossref
Vecchio, E. and Hottinger, L. 2007. Agglutinated conical foraminifera from the Lower–Middle Eocene of the Trentinara Formation (southern Italy). Facies 53: 509–533. Crossref
Velić, I. 2007. Stratigraphy and palaeobiogeography of Mesozoic benthic foraminifera of the Karst Dinarides (SE Europe). Geologia Croatica 60: 1–113. Crossref
Velić, I. and Sokač, B. 1983. Stratigraphy of the Lower Cretaceous indexfossils in the Karst Dinarides (Yugoslavia). Zitteliana 10: 485–491.
Acta Palaeontol. Pol. 68 (2): 245–260, 2023
https://doi.org/10.4202/app.01047.2022