

A new fossil from the London Clay documents the convergent origin of a “mousebird-like” tarsometatarsus in an early Eocene near-passerine bird
GERALD MAYR and ANDREW C. KITCHENER
Mayr, G. and Kitchener, A.C. 2023. A new fossil from the London Clay documents the convergent origin of a “mousebird-like” tarsometatarsus in an early Eocene near-passerine bird. Acta Palaeontologica Polonica 68 (1): 1–11.
We describe a partial skeleton of a small bird from the lower Eocene London Clay of Walton-on-the-Naze (Essex, UK), which shows close affinities to two phylogenetically controversial early Paleogene taxa, Morsoravis sedilis (lower Eocene of Denmark) and Pumiliornis tessellatus (lower/middle Eocene of Germany). Our phylogenetic analysis supports a clade including M. sedilis, P. tessellatus, and the new species, Sororavis solitarius gen. et sp. nov., and this clade is introduced as Morsoravidae fam. nov. Even though morsoravids resulted within crown group Psittacopasseres in our analysis, as the sister taxon of the Zygodactylidae and Passeriformes, they exhibit a presumably plesiomorphic tarsometatarsus morphology unlike that of zygodactyl psittacopasserines. The elongate tarsometatarsus of S. solitarius shows a resemblance to that of extant mousebirds (Coliiformes) and has a large trochlea for the second toe, which bears an extensive trochlear furrow. This distinctive foot morphology is likely to represent an adaptation for a grasping foot and may indicate a locomotory behaviour similar to that of mousebirds, which are arboreal acrobats that forage in the foliage of trees and shrubs. Since the Psittacopasseres and the Coliiformes are widely separated in current phylogenies, the “mousebird-like” tarsometatarsus of the new species represents a notable instance of convergent evolution in small arboreal birds.
Key words: Aves, Sororavis solitarius, evolution, Walton-on-the-Naze, Eocene, UK.
Gerald Mayr [gerald.mayr@senckenberg.de; ORCID: https://orcid.org/0000-0001-9808-748X ], Ornithological Section, Senckenberg Research Institute and Natural History Museum Frankfurt, Senckenberganlage 25, 60325 Frankfurt am Main, Germany.
Andrew C. Kitchener [a.kitchener@nms.ac.uk; ORCID: https://orcid.org/0000-0003-2594-0827 ], Department of Natural Sciences, National Museums Scotland, Chambers Street, Edinburgh EH1 1JF, UK, and School of Geosciences, University of Edinburgh, Drummond Street, Edinburgh EH8 9XP, UK.
Received 20 December 2022, accepted 1 February 2023, available online 7 March 2023.
Copyright © 2023 G. Mayr and A.C. Kitchener. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Morsoravis sedilis Bertelli, Lindow, Dyke, and Chiappe, 2010, is a small bird known from a three-dimensionally preserved partial skeleton from the early Eocene Fur Formation in Denmark, which was originally assigned to the Charadriiformes (Bertelli et al. 2010). Mayr (2009, 2011) challenged this classification and likened M. sedilis to Pumiliornis tessellatus Mayr, 1999, from the uppermost lower or lowermost middle Eocene of Messel in Germany and to Eocuculus cherpinae Chandler, 1999, from the lower Oligocene of Europe and North America. Eocuculus was previously assigned to the Cuculidae (Chandler 1999, but see Mayr 2006), whereas the affinities of Pumiliornis were considered uncertain (Mayr 1999, 2008).
A subsequent analysis supported close affinities between Morsoravis, Pumiliornis, and the early Eocene Psittacopes, and found these taxa to be part of the Psittacopasseres, the clade including the Psittaciformes and Passeriformes (Mayr 2015). Psittacopes was initially considered to be an archaic taxon of the Psittaciformes, but later it was hypothesised that it represents a zygodactyl stem group representative of the Passeriformes, alongside another early Cenozoic taxon, the Zygodactylidae (Mayr 2015, 2017, 2022). Psittacopes is the type genus of the taxon Psittacopedidae, which meanwhile includes several other early Paleogene taxa (Mayr 2020; Mayr and Kitchener in press).
An analysis by Ksepka et al. (2019) found Morsoravis, Psittacopes, and Pumiliornis to form a clade together with Eofringillirostrum from the lower Eocene North America Green River Formation, and this clade was the sister taxon of a clade including the Zygodactylidae and crown group Passeriformes. A clade including Morsoravis, Pumiliornis, and the psittacopedid taxa Psittacopes and Parapsittacopes was also obtained in an analysis by Mayr (2020). In a more recent analysis (Mayr and Kitchener in press), Morsoravis and Pumiliornis were recovered in a clade including the Psittacopedidae, Zygodactylidae, and crown group Passeriformes, with Eocuculus and Eofringillirostrum not being part of this clade.
However, all of the above phylogenies are rather weakly supported and poorly resolved, and critical osteological data of Morsoravis and Pumiliornis, especially concerning the morphology of the tarsometatarsus, remained unknown. Here were describe a partial skeleton of a Morsoravis-like bird from the lower Eocene London Clay of Walton-on-the-Naze (Essex, UK), which stems from the collection of the late Michael Daniels. The specimen belongs to a new genus-level taxon, and like other bird fossils from Walton-on-the-Naze (e.g., Mayr 2020; Mayr and Kitchener in press) it consists of three-dimensionally preserved isolated bones. As such it provides new osteological data that corroborate close affinities between Morsoravis and Pumiliornis and adds to a better understanding of the evolutionary significance of these distinctive Eocene birds.
Nomenclatural acts.—This published work and the nomenclatural acts it contains have been registered in ZooBank: http://zoobank.org/urn:lsid:zoobank.org:pub:40AC6727-2995-46F3-ACF8-9E5A810FE984.
Institutional abbreviations.—GMH, Geiseltalsammlung, Martin-Luther Universität of Halle-Wittenberg, Germany; MGUH, Geological Museum of the University of Copenhagen, Denmark; NMS, National Museums Scotland, Edinburgh, UK; SMF, Senckenberg Research Institute Frankfurt, Germany.
Other abbreviations: CI, consistency index; L, tree length; RI, retention index.
Material and methods
A phylogenetic analysis was performed on the basis of the emended character matrix of Mayr and Kitchener (in press; for character descriptions and character matrix see SOM 1, 2, Supplementary Online Material available at http://app.pan.pl/SOM/app68-Mayr_Kitchener_SOM.pdf). The analysis was run with the heuristic search modus of NONA 2.0 (Goloboff 1993) through the WINCLADA 1.00.08 interface (Nixon 2002), with the commands hold 10000, mult*1000, and hold/10. Bootstrap support values were calculated with 1000 replicates, ten searches holding ten trees per replicate, and TBR branch swapping without max*. The trees were rooted with the anseriform Anhimidae. Two characters were coded as additive. Tree statistics recorded are tree length (L), consistency index (CI), and retention index (RI). A constrained analysis was performed with the heuristic search modus of PAUP*4.0a169 (Swofford 2002); concerning the extant taxa a backbone constraint was enforced based on the results of current molecular analyses (Prum et al. 2015; Kuhl et al. 2021).
Systematic palaeontology
Aves Linnaeus, 1758
Telluraves Yuri, Kimball, Harshman, Bowie, Braun, Chojnowski, Han, Hackett, Huddleston, Moore, Reddy, Sheldon, Steadman, Witt, and Braun, 2013
Australaves Ericson, 2012
Family Morsoravidae nov.
LSID Zoobank: http://zoobank.org/urn:lsid:zoobank.org:act:B81B0E09 -9CC3-44E9-9FCF-BB6667A90FE0.
Type genus: Morsoravis Bertelli, Lindow, Dyke, and Chiappe, 2010.
Included genera: Morsoravis Bertelli, Lindow, Dyke, and Chiappe, 2010; Pumiliornis Mayr, 1999; Sororavis gen. nov.
Diagnosis.—The new taxon is characterised by (1) a high number of 21 praesacral vertebrae (not ascertainable for Sororavis gen. nov.); (2) a long and slender tibiotarsus; (3) a very large tuberositas musculi tibialis cranialis that is situated at the medial margin of the tarsometatarsus on the level of the foramina vascularia proximalia; (4) a trochlea metatarsi II of cylindrical shape that is plantarly deflected and has a marked and extensive trochlear furrow; (5) a fossa on the dorsal tarsometatarsus surface immediately proximal to the trochlea metatarsi III; (6) a trochlea metatarsi IV that exhibits a plantarly directed, wing-like flange and has a laterally slanted distal margin, which forms an angle of about 45° relative to the longitudinal axis of the tarsometatarsus (the presence of the flange is not ascertainable for Morsoravis); (7) a very wide first phalanx of the fourth toe (not ascertainable for Sororavis gen. nov.). Characters (4) and (7) are here considered diagnostic apomorphies of the new taxon.
Genus Sororavis nov.
LSID Zoobank: http://zoobank.org/urn:lsid:zoobank.org:act:E26943A7- 0011-4266-A5C3-1F706C9EAD18.
Type species: Sororavis solitarius sp. nov.; see below.
Etymology: From Latin soror, sister and avis, bird; in reference to the phylogenetic closeness of the new taxon to Morsoravis, to which the new name is intended to be phonetically similar.
Diagnosis.—The new taxon differs from Morsoravis Bertelli, Lindow, Dyke, and Chiappe, 2010, in that the tarsometatarsus is more elongated (ratio length of bone to minimum width of shaft 12.9 vs. 8.9 in M. sedilis). It is distinguished from Pumiliornis Mayr, 1999 in that the tibiotarsus has a pons supratendineus and the tarsometatarsus is more elongated (ratio length of bone to minimum width of shaft 12.9 vs. 8.3 in P. tessellatus).
Sororavis solitarius sp. nov.
Figs. 1–3.
LSID Zoobank: http://zoobank.org/urn:lsid:zoobank.org:act:A979CC 23-3409-4CDD-B28E-2FEA52691A86.
Etymology: From Latin solitaries, single, solitary, in reference to the fact that only one specimen of this species has been identified among the numerous birds from Walton-on-the Naze in the Daniels collection.
Holotype: NMS.Z.2021.40.75 (Fig. 1; tip of upper beak; fragments of mandibular rami; both coracoids; partial furcula; partial sternum; right humerus lacking distal end; proximal and distal ends of left humerus; proximal end of right ulna; left tibiotarsus lacking proximal end; proximal end of left tarsometatarsus; right tarsometatarsus; four pedal phalanges), collected in 1986 by M. Daniels (original collector’s number WN 86530).
Type locality: Walton-on-the-Naze, Essex, UK.
Type horizon: Walton Member of the London Clay Formation (previously Division A2; Rayner et al. 2009; Aldiss 2012); lower Eocene (lower Ypresian, 54.6‒55 million years old; Collinson et al. 2016).
Diagnosis.—As for genus. The new species is of similar size to Morsoravis sedilis (tarsometatarsus length 14.3 mm vs. ~13.0 mm in M. sedilis; Bertelli et al. 2010) and distinctly larger than Pumiliornis tessellatus (tarsometatarsus length 8.9‒9.9 mm; Mayr 1999).
Measurements (maximum length, in mm).—Right coracoid, 11.0; right humerus, length as preserved, 14.0, estimated total length, ~15‒16; left tibiotarsus, length as preserved, 16.5, estimated total length, ~23‒25; right tarsometatarsus, 14.3.
Description.—The tip of the upper beak (Figs. 1A1, 2B1) is narrow and pointed, and its shape indicates similar proportions to the beak of Morsoravis sedilis (Fig. 2A1) and extant thrushes (Turdidae, Passeriformes). The fragments of the mandibular rami show that the lower jaw was long and dorsoventrally low as it is in M. sedilis.
The coracoid (Figs. 1A3‒A5, 2B2) resembles the corresponding bone of the Psittacopedidae (Fig. 2D) and Zygodactylidae (Fig. 2C) as well as that of crown group passerines (Fig. 2E). The extremitas omalis has a hook-like shape and the facies articularis scapularis is shallow. The processus procoracoideus is broken, but judging from the larger portion preserved in the right coracoid, it appears to have been fairly long. A foramen nervi supracoracoidei is absent. The extremitas sternalis is comparatively narrow, with a short processus lateralis. The medial margin bears an incipient notch.

Fig. 1. The bones preserved in the holotype of the morsoravid bird Sororavis solitarius gen. et sp. nov. (NMS.Z.2021.40.75), from the lower Eocene London Clay of Walton-on-the-Naze, UK. A1, tip of upper beak in dorsal view; A2, fragments of mandible; A3, A4, left coracoid in dorsal (A3) and ventral (A4) views; A5, A6, right coracoid in dorsal (A5) and ventral (A6) views; A7, partial furcula; A8, A9, cranial portion of sternum in ventral (A8) and lateral (A9) views; A10, A11, partial right humerus in cranial (A10) and caudal (A11) views; A12‒A15, proximal (A12, A13) and distal (A14, A15) portions of left humerus in caudal (A12, A14) and cranial (A13, A15) views; A16, proximal end of right ulna in cranioventral view; A17, A18, partial left tibiotarsus in caudal (A17) and cranial (A18) views; A19‒A24, right tarsometatarsus in dorsal (A19), medial (A20), plantar (A21), lateral (A22), proximal (A23), and distal (A24) views; A25, A26, proximal end of left tarsometatarsus in plantar (A25) and dorsolateral (A26) views; A27, first phalanx of third toe in dorsal and plantar view; A28, second to fourth phalanges of fourth toe in different views (plantar, dorsal, and lateral, respectively).
Only a fragment of the extremitas sternalis of the furcula is preserved (Fig. 1A7), which indicates that the bone was U-shaped and had slender shafts. The apophysis furculae is broken, but appears to have been small.
The specimen includes the cranial portion of the sternum (Fig. 1A8, A9), which has a long and mediolaterally wide spina externa that is only incipiently bifurcated. The apex carinae shows little cranial projection.
The humerus (Figs. 1A10‒A15, 2B3) resembles that of the Psittacopedidae and Zygodactylidae (Fig. 2G, H; Mayr and Kitchener in press). As in the latter, the fossa pneumotricipitalis lacks pneumatic openings, which is likely to be a plesiomorphic characteristic and functionally correlated with the occurrence of pleurocoels on the thoracic vertebrae (Mayr 2021). The tuberculum dorsale is small. The crista deltopectoralis is broken, but the remaining portions indicate that it was proximodistally short. The sulcus scapulotricipitalis is very shallow. The condylus ventralis is more elongated and less globose than in the Psittacopedidae and Zygodactylidae; unlike in the latter two taxa, the dorsal margin of the bone does not form a well-defined tuberculum/processus supracondylaris dorsalis. With regard to other features, such as the size and position of the fossa musculi brachialis and tuberculum supracondylare ventrale as well as the shape of the processus flexorius, the distal end of the bone resembles the distal humerus of psittacopedids and zygodactylids (Fig. 2G, H).
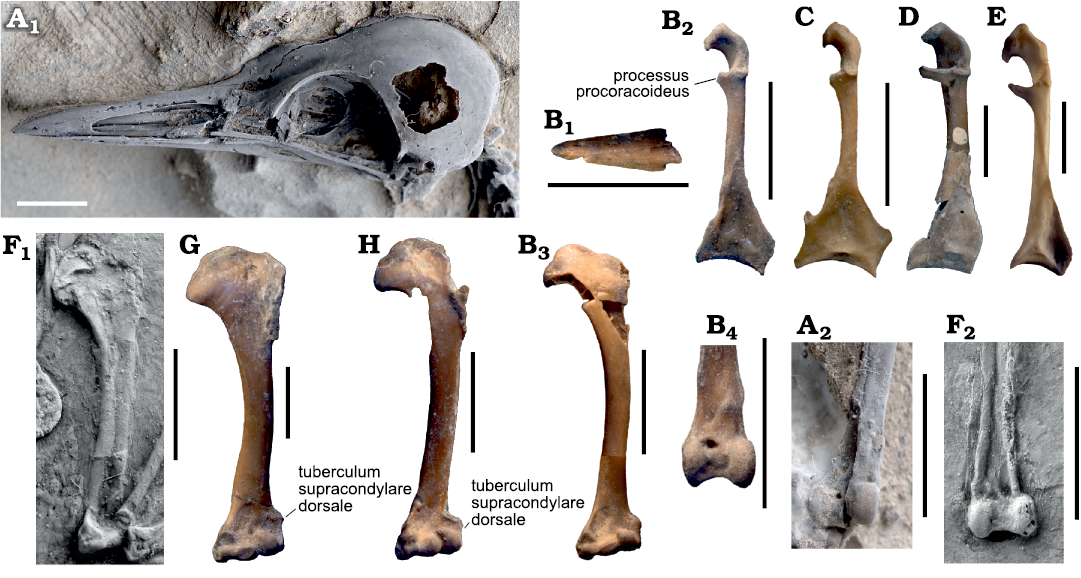
Fig. 2. Beak and selected postcranial bones of the morsoravid bird Sororavis solitarius gen. et sp. nov. in comparison to those of other Morsoravidae and the Psittacopedidae, Zygodactylidae, and Coliiformes. A. Morsoravis sedilis Bertelli, Lindow, Dyke, and Chiappe, 2010 (holotype, MGUH 28930), from the lower Eocene Fur Formation in Denmark; A1, skull in dorsolateral view (coated with ammonium chloride); A2, distal end of left tibiotarsus in cranial view. B. Sororavis solitarius gen. et sp. nov. (holotype, NMS.Z.2021.40.75), from the early Eocene London Clay of Walton-on-the-Naze, UK. B1, tip of upper beak in dorsal view; B2, right coracoid in dorsal view; B3, composite image of partial right humerus (mirrored) and distal end of left humerus in cranial view; B4, distal end of left tibiotarsus in cranial view. C. Primoscens carolinae Mayr and Kitchener, 2022 (Zygodactylidae) (holotype, NMS.2021.40.54), from the early Eocene London Clay of Walton-on-the-Naze, UK, left coracoid in dorsal view (mirrored). D. ?Psittacopes occidentalis Mayr and Kitchener, 2022 (Psittacopedidae) (holotype, NMS.Z.2021.40.44), from the early Eocene London Clay of Walton-on-the-Naze, UK, left coracoid in dorsal view (mirrored). E. The extant Myiarchus tyrannulus (Statius Müller, 1776) (Passeriformes, Tyrannidae) (SMF 9584), right coracoid in dorsal view. F. Pumiliornis tessellatus Mayr, 1999 (SMF-ME 2475B), from the luppermost lower or lowermost middle Eocene of Messel, Germany; F1, right humerus in cranial view (mirrored); F2, distal end of left tibiotarsus in cranial view. G. Parapsittacopes bergdahli Mayr, 2021 (Psittacopedidae) (NMS. Z.2021.40.43), from the early Eocene London Clay of Walton-on-the-Naze, UK, right humerus in cranial view. H. Primozygodactylus cf. danielsi Mayr, 1998 (Zygodactylidae) (NMS.2021.40.49), from the early Eocene London Clay of Walton-on-the-Naze, UK, right humerus in cranial view (mirrored). Scale bars 5 mm.
The holotype includes the proximal end of the ulna (Fig. 1A16), which shows a fairly undiagnostic morphology, with an olecranon of moderate size, a rather small cotyla dorsalis, and an average-sized cotyla ventralis with a circular outline.
The tibiotarsus (Figs. 1A17, A18, 2B4), whose proximal end is lacking, is a very long and slender bone, and its shaft is proportionally narrower than in Morsoravis. The distal end of the bone closely resembles the distal tibiotarsus of Morsoravis sedilis (Fig. 2A2). The pons supratendineus, which is absent in Pumiliornis (Fig. 2F2), is proximodistally long. Proximal to the condylus dorsalis and lateral to the distal end of the sulcus extensorius, there is an embossment of subtriangular outline. The condylus medialis is somewhat less tall proximodistally and less wide mediolaterally than the condylus lateralis.
The tarsometatarsus (Figs. 1A19‒A24, 3A1‒A3) is proportionally longer and narrower than the corresponding bone of Morsoravis sedilis (Fig. 3B) and Pumiliornis tessellatus (Fig. 3C). The shaft is very flat in dorsoplantar direction. As in P. tessellatus, the proximal margin of the bone is not oriented perpendicular to its longitudinal axis. Furthermore as in M. sedilis and P. tessellatus, the medial portion of the cotyla medialis forms a proximally projecting lip (Fig. 3A1); medial to it there is a small medial embossment (Fig. 1A23). The hypotarsus exhibits two sulci for the tendons of musculus flexor digitorum longus and musculus flexor hallucis longus, respectively. The small foramina vascularia proximalia are on the same level and are widely separated. The large tuberositas musculi tibialis cranialis is situated at the medial margin of the bone and contributes to a medial bulge of the shaft; in P. tessellatus, there is an equally large and medially situated tuberositas musculi tibialis cranialis (this feature is not visible in the M. sedilis holotype, but a large tuberositas musculi tibialis cranialis was present on the proximal end of the right tarsometatarsus; this bone was figured by Kristoffersen 2002, but it broke off subsequently and is now lost). The plantar surface of the shaft is essentially flat and lacks a crista medianoplantaris. The distal end of the bone closely resembles the tarsometatarsus of M. sedilis. The foramen vasculare distale is small and there is no sulcus between this foramen and the incisura intertrochlearis lateralis. The fossa metatarsi I is situated on the plantar surface of the bone, near to its medial margin. In distal view, the trochleae form an arch, with both the trochlea metatarsi II and the trochlea metatarsi IV being plantarly deflected. The trochlea metatarsi II is large and of subcylindrical shape; it is plantarly deflected and bears a marked trochlear furrow that extends onto its medial surface (Fig. 3A3). In the holotype and only known specimen of M. sedilis this furrow on the medial surface of the trochlea metatarsi II is clearly visible (Fig. 3B2, the plantar tarsometatarsus surface is not exposed). The distal end of the tarsometatarsus is poorly preserved in the known specimens of Pumiliornis tessellatus, but the trochlea metatarsi II appears to be of similar shape to that of S. solitarius and is much larger than in psittacopedids (Fig. 3D2). On the dorsal surface of the distal tarsometatarsus of S. solitarius, there is a depression immediately proximal to the trochlea metatarsi III, which is also present in P. tessellatus (the corresponding area of the tarsometatarsus is damaged in the M. sedilis holotype). The trochlea metatarsi III is not mediolaterally widened and has a well-developed trochlear furrow. The trochlea metatarsi IV is plantarly deflected and forms a wing-like flange of similar shape to that in extant Leptosomiformes and Strigiformes, which indicates at least semizygodactyl feet. As in Morsoravis and Pumiliornis, the distal margin of the trochlea metatarsi IV is laterally slanted, forming an angle of about 45° relative to the longitudinal axis of the tarsometatarsus.
The holotype includes four pedal phalanges, which we identify as the first phalanx of the third toe and the second to fourth phalanges of the fourth toe (Fig. 1A27, A28). The first phalanx of the third toe has a distinctive morphology in that the trochlea has very long plantar rims. The proximal phalanges of the fourth toe are not greatly abbreviated.
Stratigraphic and geographic range.—Type locality and horizon only.
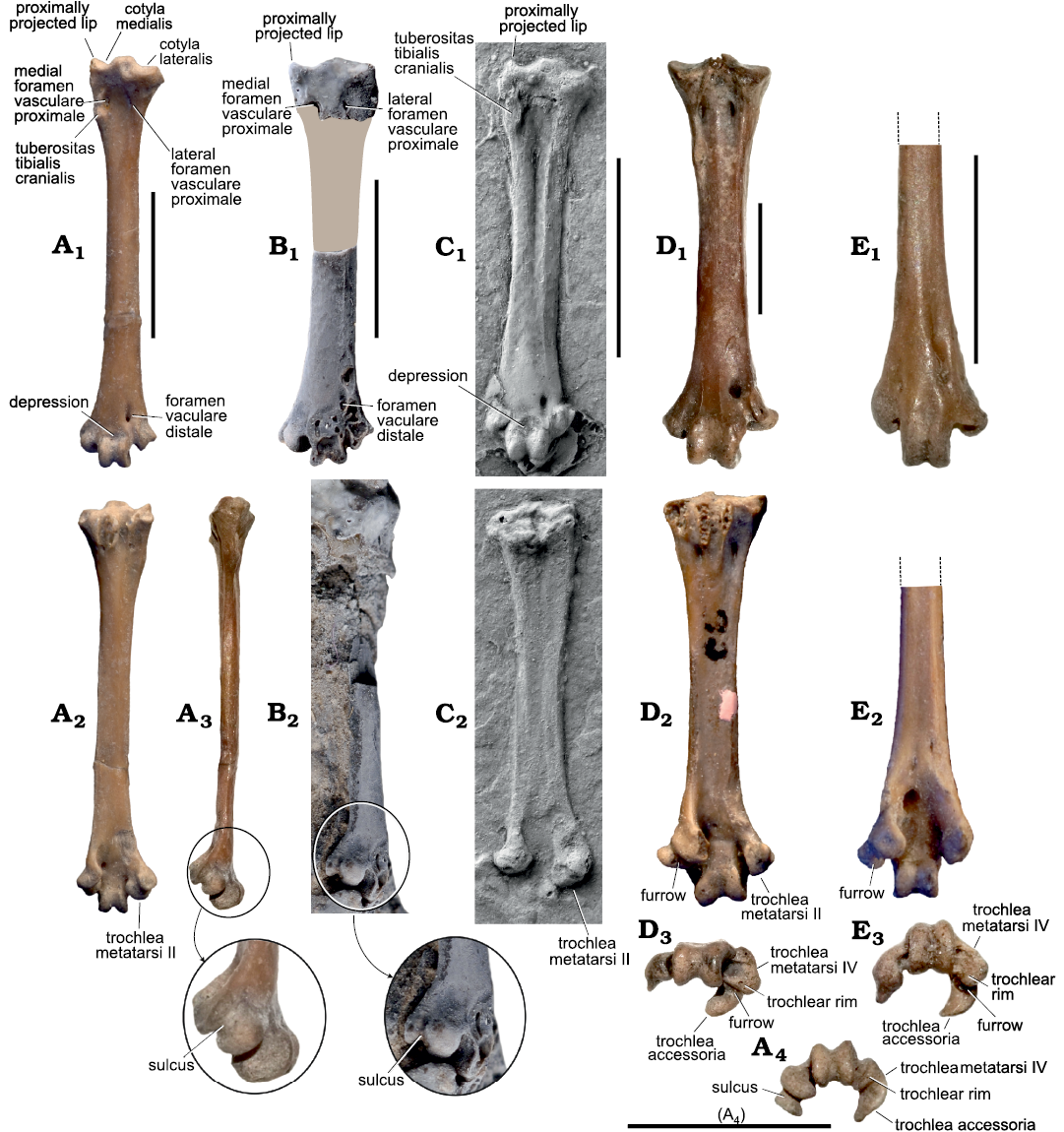
Fig. 3. Leg bones of the morsoravid bird Sororavis solitarius gen. et sp. nov. in comparison to those of other Morsoravidae and the Psittacopedidae and Zygodactylidae. A. Sororavis solitarius gen. et sp. nov. (holotype, NMS.Z.2021.40.75), from the early Eocene London Clay of Walton-on-the-Naze, UK, right tarsometatarsus (mirrored), in dorsal (A1), plantar (A2), and medial (A3) views, the arrow indicates an enlarged detail of the distal end; distal end of right tarsometatarsus (mirrored) in distal view (A4). B. Morsoravis sedilis Bertelli, Lindow, Dyke, and Chiappe, 2010 (holotype, MGUH 28930), from the early Eocene Fur Formation in Denmark, left tarsometatarsus in dorsal (B1) and medial (B2) views; coated with ammonium chloride, in B1, surrounding matrix was digitally removed and a missing portion of the shaft is highlighted by the grey-brown area, the arrow in B2, indicates an enlarged detail of the distal end. C. Pumiliornis tessellatus Mayr, 1999 (SMF-ME 2475A), from the latest early or earliest middle Eocene of Messel, Germany, left tarsometatarsus in dorsal (C1) and plantar (C2) views. D. Psittacomimus eos Mayr and Kitchener, 2022 (NMS.Z.2021.40.39), from the early Eocene London Clay of Walton-on-the-Naze, UK, left tarsometatarsus in dorsal (D1), plantar (D2), and distal (D3) views. E. Primozygodactylus cf. danielsi Mayr, 1998 (Zygodactylidae) (NMS.2021.40.49), from the early Eocene London Clay of Walton-on-the-Naze, UK, distal portion of right tarsometatarsus (mirrored), in dorsal (E1), plantar (E2), and distal (E3) views. Scale bars 5 mm.
Results of the phylogenetic analysis
Analysis of the primary character matrix in the SOM resulted in ten most parsimonious trees (L = 316, CI = 0.33, RI = 0.59), the consensus tree of which is shown in Fig. 4. The analysis supported a clade including Sororavis, Morsoravis, and Pumiliornis. This clade, the Morsoravidae fam. nov., was obtained as part of a more inclusive clade that also comprises the Psittacopedidae, Zygodactylidae, and crown group Passeriformes, with the Morsoravidae being the sister group of a clade formed by the Zygodactylidae and crown group Passeriformes. However, whereas the clade formed by Sororavis, Morsoravis, and Pumiliornis received weak bootstrap support, the more inclusive clade was not retained in the bootstrap analysis (Fig. 4). Three characters were optimised as apomorphies of a clade including Sororavis, Morsoravis, and Pumiliornis, that is, ch. 78: tarsometatarsus, trochlea metatarsi III with a deeply incised groove between the trochlear rims; ch. 82: tarsometatarsus, trochlea metatarsi IV with a laterally slanted distal margin, which forms an angle of about 45° relative to the longitudinal axis of the tarsometatarsus; and ch. 86: fourth toe, proximal phalanx greatly widened (this character was optimised as a diagnostic apomorphy with CI = 1.0).

Fig. 4. Strict consensus tree of ten most parsimonious trees (L = 316, CI = 0.33, RI = 0.59) resulting from the phylogenetic analysis. Unsupported nodes were collapsed; extinct taxa are indicated by a dagger. Bootstrap support values are given next to the internodes.
The following characters were optimised as apomorphies of a clade including Sororavis, Morsoravis, and Pumiliornis, as well as the Psittacopedidae, Zygodactylidae, and crown group passerines: Ch. 22: furcula with well-developed apophysis furculae; ch. 34: humerus, processus flexorius prominent and reaching distally well beyond distal margin of condylus ventralis; ch. 61: tibiotarsus, crista cnemialis lateralis distinctly hooked. Two further characters represent reversals into the plesiomorphic condition: ch. 62: tibiotarsus, distal end not mediolaterally wide and not craniocaudally compressed, without shallow trochlea cartilaginis tibialis; ch. 87: fourth toe, proximal three phalanges not abbreviated.
A clade including Sororavis, Morsoravis, Pumiliornis, the Zygodactylidae, and crown group passerines is supported by: Ch. 48: carpometacarpus, distinct, narrow fossa supratrochlearis running across dorsal portion of trochlea carpalis; ch. 67: tarsometatarsus, medial portion of cotyla medialis forming a proximally projecting lip. Three further characters represent reversals into the plesiomorphic condition: Ch. 74: tarsometatarsus, without dorsally open sulcus between foramen vasculare distale and incisura intertrochlearis lateralis; ch. 77: tarsometatarsus, trochlea metatarsi III not much wider in mediolateral than in dorsoplantar direction; ch. 79: tarsometatarsus, trochlea metatarsi III without distinct tubercle on lateral side.
Like other analyses of morphological data, our study did not recover major clades that received strong support in analyses of sequence data (e.g., Prum et al. 2015; Kuhl et al. 2021; Sangster et al. 2022). This is most obvious in the case of the Eufalconimorphae (the clade including the Falconiformes and Psittacopasseres), Eucavitaves (the clade including the Trogoniformes, Bucerotiformes, Coraciiformes, and Piciformes), and Coraciimorphae (the clade including the Coliiformes, Leptosomiformes, and Eucavitaves).
A second analysis was therefore run that was constrained to a molecular backbone phylogeny for the extant taxa based on current molecular phylogenies (Prum et al. 2015; Kuhl et al. 2021). This analysis resulted in 714 most parsimonious trees (L = 331, CI = 0.32, RI = 0.57), the consensus tree of which is shown in Fig. 5. Concerning the affinities of the new species, the results of this analysis are concordant with the primary data set. However, unlike in the primary analysis, halcyornithids, messelasturids, and vastanavids were found to be outside crown group Psittacopasseres.
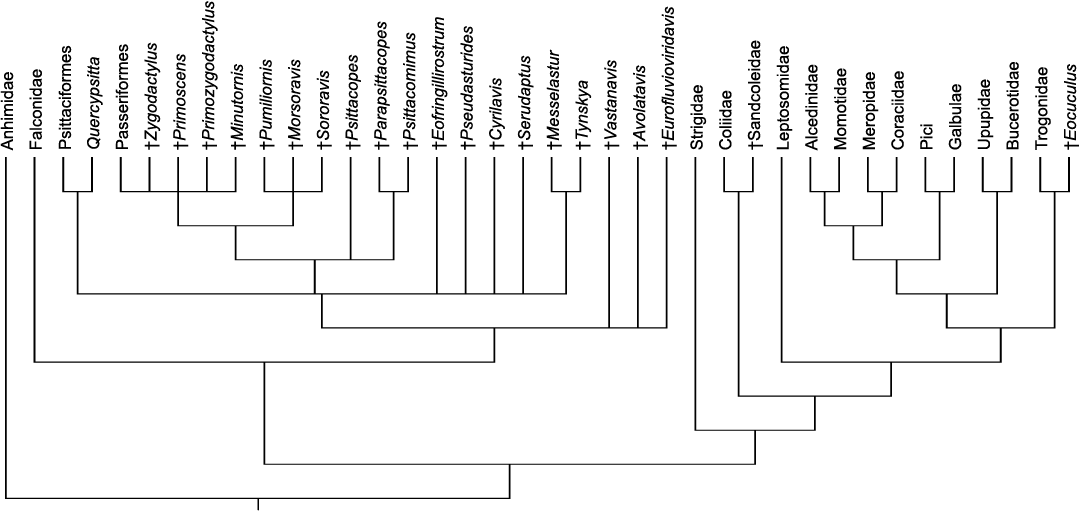
Fig. 5. Strict consensus tree of 714 most parsimonious trees (L = 331, CI = 0.32, RI = 0.57) resulting from an analysis that, concerning the extant taxa, was constrained to the results of current molecular analyses (Prum et al. 2015; Kuhl et al. 2021). Extinct taxa are indicated by a dagger.
Discussion
Phylogenetic affinities.—The new fossil corroborates close affinities between Morsoravis and Pumiliornis, with the well-preserved bones of Sororavis solitarius gen. et sp. nov. elucidating morphological features that are not clearly visible in the known specimens of Morsoravis and Pumiliornis. This is particularly true for the tarsometatarsus, whose distal end conforms to the distal tarsometatarsus of Morsoravis with respect to the marked trochlear furrow of the trochlea metatarsi II, whereas the proximal end of the bone agrees with Pumiliornis in the large and medially situated tuberositas musculi tibialis cranialis (the trochlea metatarsi II is poorly preserved in the known Pumiliornis tessellatus specimens, whereas the proximal tarsometatarsus end is damaged in the holotype of Morsoravis sedilis).
Even though all formally named taxa of the Morsoravidae fam. nov. are from European sites, these birds also occurred during the early Eocene in North America, with Grande (2013: fig. 143B) figuring a skeleton from the North American Green River Formation, which closely resembles Sororavis solitarius. This fossil was likened to Morsoravis sedilis by Grande (2013) and was the sister taxon of M. sedilis in an analysis by Ksepka et al. (2019). In contrast, a tarsometatarsus from the lower Eocene North American Nanjemoy Formation, which was tentatively referred to Pumiliornis by Mayr et al. (2022), may actually be from a representative of the Psittacopedidae (Mayr and Kitchener in press); this fossil is clearly distinguished from the tarsometatarsus of S. solitarius in that the trochlea accessoria is separated by a furrow from the trochlea metatarsi IV proper.
Our analysis recovers the Morsoravidae as the sister group of a clade formed by zygodactylids and crown group Passeriformes, and the taxon is therefore found to be nested within zygodactyl stem group Psittacopasseres. Eofringillirostrum and Eocuculus were shown to be only distantly related to the Morsoravidae, and compared to the “finch-beaked” species of Eofringillirostrum, the upper beak of S. solitarius appears to have been much narrower and more thrush-like, the humerus is more elongated and without a well-defined tuberculum supracondylare dorsale, the tarsometatarsus is proportionally longer, with the trochlea metatarsi IV reaching farther distally, and the proximal phalanges of the fourth toe are more slender and less abbreviated. Sororavis solitarius differs from Eocuculus in that the humerus is less stout, the tibiotarsus much longer, the tarsometatarsus more elongated, and the trochlea metatarsi II wider and of sub-cylindrical shape.
A placement of morsoravids within crown group Psittacopasseres conforms to the results of previous analyses (Mayr 2015; Ksepka et al. 2019; Mayr and Kitchener in press). The coracoid and humerus of S. solitarius closely resemble the corresponding elements of early Eocene zygodactyl representatives of the Psittacopasseres, that is, the Psittacopedidae and Zygodactylidae, of which there are multiple specimens in the Daniels collection (Figs. 2, 3; Mayr and Kitchener in press). However, the distal end of the tarsometatarsus of the new species exhibits some distinct differences to psittacopedids and zygodactylids, whose distal tarsometatarsus resembles that of the Psittaciformes. The most notable derived feature associated with the tarsometatarsus morphology of psittaciforms, psittacopedids, and zygodactylids is a large accessory trochlea, which is delimited by a furrow from the trochlea metatarsi IV proper (Fig. 3D1‒E3). This furrow is absent in Sororavis, in which the trochlea accessoria is less developed and the trochlear rim of the trochlea metatarsi IV more dorsoplantarly (in the distal view of the tarsometatarsus) oriented than in psittacopedids and zygodactylids. If the Morsoravidae are nested within Psittacopasseres, the derived morphology of the distal tarsometatarsus was either lost in morsoravids, or it evolved convergently in psittaciforms, psittacopedids, and zygodactylids.
However, we note that the position of morsoravids within Psittacopasseres was not corroborated by the bootstrap analyses and is mainly based on characters that show homoplasy or represent reversals into the plesiomorphic state. Therefore, the higher-level affinities of these birds still have to be firmly established, and the morphology of the distal tarsometatarsus of Sororavis would actually be in better agreement with a position of the new taxon, as well as other morsoravids, outside the Psittacopasseres, which is defined as the least inclusive clade containing the Psittaciformes and the Passeriformes (Sangster et al. 2022).
According to current molecular phylogenies (e.g., Ericson et al. 2006; Prum et al. 2015; Kuhl et al. 2021), Telluraves, the clade including arboreal landbirds, falls into two sister taxa termed Australaves and Afroaves (Ericson 2012; Sangster et al. 2022). Together with the Cariamiformes and Falconiformes, the Psittacopasseres are part of the Australaves, whereas the Afroaves include (among others) the Accipitriformes, Coliiformes, Strigiformes, Coraciiformes, and Piciformes. Sororavis solitarius resembles extant Coliiformes in the overall morphology of some bones, which is particularly true for the humerus (Fig. 6A1, B, C1) and the long and slender tarsometatarsus (Fig. 6A2, C2). Because the Psittacopasseres and Coliiformes are widely separated in current phylogenies, these similarities are notable, but various differences in detail between Sororavis and coliiform birds indicate that they have evolved due to convergence. Concerning the humerus, for example, the tuberculum supracondylare ventrale and the condylus dorsalis of Sororavis are proportionally smaller than in coliiform birds and the condylus ventralis is less globose (Fig. 6A1, B, C1). As shown by the skeletons of Pumiliornis tessellatus and the undescribed Sororavis-like bird from the Green River Formation (see above), the ulna of morsoravids exceeds the humerus in length, whereas both bones are of subequal length in the Coliiformes. The tarsometatarsus of Sororavis is distinguished from that of fossil and extant Coliiformes in that the fossa metatarsi I is situated on its plantar face and not on the medial one as in coliiform birds. Furthermore unlike in coliiform birds, the proximal phalanges of the fourth toe of Sororavis are not greatly abbreviated. A convergent origin of the similar tarsometatarsus morphology of Sororavis and crown group Coliiformes is also indicated by the fact that the tarsometatarsi of Pumiliornis and Morsoravis are more like those of the Psittacopedidae in their proportions, and stem group Coliiformes, such as the Sandcoleidae (Houde and Olson 1992; Mayr and Peters 1998; Mayr 2018), likewise have stouter tarsometatarsi than extant mousebirds.
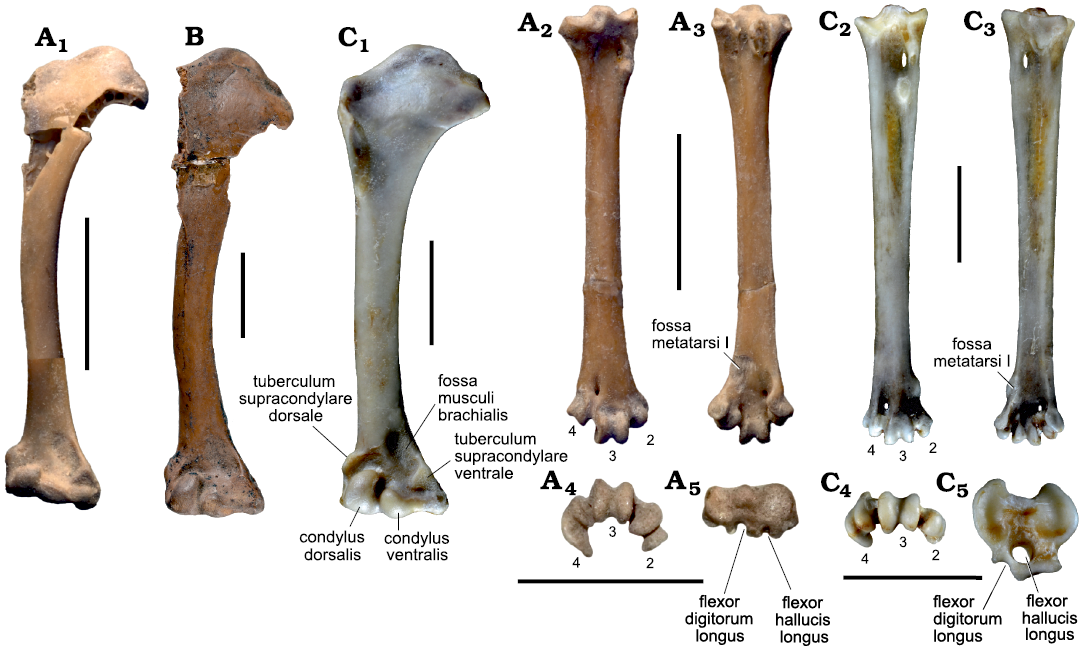
Fig. 6. Humeri and tarsometatarsi of the morsoravid bird Sororavis solitarius gen. et sp. nov. and coliiform birds. A. Sororavis solitarius gen. et sp. nov. (holotype, NMS.Z.2021.40.75), from the early Eocene London Clay of Walton-on-the-Naze, UK; A1, composite reconstruction of proximal portion of right humerus and mirrored distal end of left humerus in cranial view; right tarsometatarsus in dorsal (A2), plantar (A3), distal (A4), and proximal (A5) views. B. Selmes absurdipes Peters, 1999 (Coliiformes) (GMH XXXVI-305-1963), from the middle Eocene of the Geisel Valley in Germany; right humerus in cranial view. C. The extant Colius striatus Gmelin, 1789 (Coliiformes) (SMF 8019); C1, right humerus in cranial view; right tarsometatarsus in dorsal (C2), plantar (C3), distal (C4), and proximal (C5) views. The tarsometatarsal trochleae are numbered. Scale bars 5 mm.
Ecomorphological aspects.—The taxon Morsoravidae is characterised by a derived morphology of the tarsometatarsus, which exhibits a large, subcylindrical trochlea metatarsi II with a pronounced trochlear furrow, a trochlea metatarsi IV with a plantarly directed, wing-like flange and a large and medially situated tuberositas musculi tibialis cranialis. The laterally slanted distal margin of the trochlea metatarsi IV of Sororavis solitarius suggests a splayed fourth toe, and the wing-like plantar flange of this trochlea indicates a semi-zygodactyl foot, in which the fourth toe was at least partially reversed. A semizygodactyl foot was also assumed to have been present in Morsoravis and Pumiliornis (Mayr 2011).
The trochlea metatarsi II of S. solitarius is much larger than the corresponding tarsometatarsal trochlea of the Psittacopedidae and Zygodactylidae. Its cylindrical shape and the marked trochlear furrow (Figs. 1A24, 3A4) suggest that the articulation between this trochlea and the second toe formed a hinge-like joint, with little freedom for rotationary movements. Together with the semi-zygodactyl foot, this trochlear morphology probably enhanced the grasping capabilities of the foot. The very large tuberositas musculi tibialis cranialis indicates a strongly developed musculus tibialis cranialis inserting on it, which flexes the tarsometatarsus and lifts the body if the tarsometatarsus is fixed.
We hypothesise that morsoravids had a specialised feeding ecology, which was facilitated by this derived leg morphology. Pollen grains in the gut contents of a Pumiliornis tessellatus fossil indicate that this species visited flowers and was probably at least facultatively nectarivorous (Mayr and Wilde 2014). As noted above, S. solitarius shows a resemblance to coliiform birds with regard to tarsometatarsus morphology and the proportions of the long and slender tibiotarsus are likewise similar. Therefore, like mousebirds, the new species may have been capable of dangling from branches in order to reach fruits and inflorescences, or food items associated therewith (however, unlike in Sororavis, the proximal phalanges of the toes of mousebirds are strongly abbreviated, which indicates some functional differences in detail).
Mousebirds have a fossil record that goes back into the late Paleocene and by that time they had already evolved their characteristic foot structure (Houde and Olson 1992; Mayr 2018, 2022). Irrespective of the exact position of morsoravids within Australaves, the occurrence of a “mousebird-like” tarsometatarsus in Sororavis represents a remarkable case of parallelism and suggests that ecological niches for avian arboreal acrobats were occupied by multiple, only distantly related groups in the early Paleogene.
Conclusions
Sororavis solitarius gen. et sp. nov. forms a clade together with Morsoravis sedilis from the Fur Formation in Denmark and Pumiliornis tesselatus from Messel in Germany, and the new fossil from the London Clay adds to a better understanding of this distinctive group of early Eocene arboreal birds. Judging from their disparate tarsometatarsus morphologies, these three species appear to have occupied diverse ecological niches, and at least Sororavis and Morsoravis are likely to have been adapted to some level of direct or indirect plant-bird interactions. Their position within Psittacopasseres, the clade including the Psittaciformes and Passeriformes, may challenge previous hypotheses concerning the ancestral tarsometatarsus morphology of psittacopasserines, and the new fossil underscores the significance of the Walton-on-the-Naze site for a reconstruction of early Eocene European avifaunas and ecosystems.
Acknowledgements
We thank Sven Tränkner (Senckenberg Research Institute, Frankfurt, Germany) for taking most of the photographs (additional images are by GM). Ian Garofalo (New Mexico State University, Las Cruces, USA) is thanked for pointing out an erroneous scoring in a previous version of the character matrix regarding the taxon Tynskya. Comments from two anonymous reviewers improved the manuscript.
References
Aldiss, D.T. 2012. The stratigraphical framework for the Palaeogene successions of the London Basin, UK. British Geological Survey Open Reports OR/12/004: 1–87.
Bertelli, S., Lindow, B.E.K., Dyke, G.J., and Chiappe, L.M. 2010. A well-preserved “charadriiform-like” fossil bird from the Lower Eocene Fur Formation of Denmark. Palaeontology 53: 507–531. Crossref
Chandler, R.M. 1999. Fossil birds of Florissant, Colorado: with a description of a new genus and species of cuckoo. National Park Service, Geological Resources Division, Technical Report NPS/NRGRD/GRDTR-99: 49–53.
Collinson, M.E., Adams, N.F., Manchester, S.R., Stull, G.W., Herrera, F., Smith, S.Y., Andrew, M.J., Kenrick, P., and Sykes, D. 2016. X-ray micro-computed tomography (micro-CT) of pyrite-permineralized fruits and seeds from the London Clay Formation (Ypresian) conserved in silicone oil: a critical evaluation. Botany 94: 697–711. Crossref
Ericson, P.G.P. 2012. Evolution of terrestrial birds in three continents: biogeography and parallel radiations. Journal of Biogeography 39: 813–824. Crossref
Ericson, P.G.P., Anderson, C.L., Britton, T., Elzanowski, A., Johansson, U.S., Källersjö, M., Ohlson, J.I., Parsons, T.J., Zuccon, D., and Mayr, G. 2006. Diversification of Neoaves: integration of molecular sequence data and fossils. Biology Letters 2: 543–547. Crossref
Goloboff, P.A. 1993. NONA version 2.0. Published by the author, S.M. de Tucumán.
Grande, L. 2013. The Lost World of Fossil Lake. Snapshots from Deep Time. 432 pp. University of Chicago Press, Chicago. Crossref
Houde, P. and Olson, S.L. 1992. A radiation of coly-like birds from the early Eocene of North America (Aves: Sandcoleiformes new order). In: K.E. Campbell (ed.), Papers in Avian Paleontology Honoring Pierce Brodkorb. Natural History Museum of Los Angeles County, Science Series 36: 137–160.
Kristoffersen, A.V. 2002. The Avian Diversity in the Latest Paleocene–earliest Eocene Fur Formation, Denmark. A Synopsis. 95 pp. Ph.D. Dissertation, University of Copenhagen, Geological Institute, Copenhagen.
Ksepka, D.T., Grande, L., and Mayr, G. 2019. Oldest finch-beaked birds reveal parallel ecological radiations in the earliest evolution of passerines. Current Biology 29: 657–663. Crossref
Kuhl, H., Frankl-Vilches, C., Bakker, A., Mayr, G., Nikolaus, G., Berno, S.T., Klages, S., Timmermann, B., and Gahr, M. 2021. An unbiased molecular approach using 3’UTRs resolves the avian family-level tree of life. Molecular Biology and Evolution 38: 108–127. Crossref
Mayr, G. 1999. Pumiliornis tessellatus n. gen. n. sp., a new enigmatic bird from the Middle Eocene of Grube Messel (Hessen, Germany). Courier Forschungsinstitut Senckenberg 216: 75–83.
Mayr, G. 2006. A specimen of Eocuculus Chandler, 1999 (Aves, ?Cuculidae) from the early Oligocene of France. Geobios 39: 865–872. Crossref
Mayr, G. 2008. Pumiliornis tessellatus Mayr, 1999 revisited—new data on the osteology and possible phylogenetic affinities of an enigmatic Middle Eocene bird. Paläontologische Zeitschrift 82: 247–253. Crossref
Mayr, G. 2009. Paleogene Fossil Birds. 1st Edition. 262 pp. Springer, Heidelberg. Crossref
Mayr, G. 2011. On the osteology and phylogenetic affinities of Morsoravis sedilis (Aves) from the early Eocene Fur Formation of Denmark. Bulletin of the Geological Society of Denmark 59: 23–35. Crossref
Mayr, G. 2015. A reassessment of Eocene
parrotlike fossils indicates a previously undetected radiation of
zygodactyl stem group representatives of passerines (Passeriformes).
Zoologica Scripta 44: 587–602.
Crossref
Mayr, G. 2017. Avian Evolution: The Fossil Record of Birds and Its Paleobiological Significance. 293 pp. Wiley-Blackwell, Chichester. Crossref
Mayr, G. 2018. New data on the anatomy and paleobiology of sandcoleid mousebirds (Aves, Coliiformes) from the early Eocene of Messel. Palaeobiodiversity and Palaeoenvironments 98: 639–651. Crossref
Mayr, G. 2020. A remarkably complete skeleton from the London Clay provides insights into the morphology and diversity of early Eocene zygodactyl near-passerine birds. Journal of Systematic Palaeontology 18: 1891–1906. Crossref
Mayr, G. 2021. On the occurrence of lateral openings and fossae (pleurocoels) in the thoracic vertebrae of neornithine birds and their functional significance. Vertebrate Zoology 71: 453–463. Crossref
Mayr, G. 2022. Paleogene Fossil Birds. 2nd Edition. 239 pp. Springer, Cham. Crossref
Mayr, G. and Kitchener, A.C. (in press). Psittacopedids and zygodactylids: The diverse and species-rich psittacopasserine birds from the early Eocene London Clay of Walton-on-the-Naze (Essex, UK). Historical Biology [published online, https://doi.org/10.1080/08912963.2022.2141629]. Crossref
Mayr, G. and Peters, D.S. 1998. The mousebirds (Aves: Coliiformes) from the Middle Eocene of Grube Messel (Hessen, Germany). Senckenbergiana lethaea 78: 179–197. Crossref
Mayr, G. and Wilde, V. 2014. Eocene fossil is earliest evidence of flower-visiting by birds. Biology Letters 10: 20140223. Crossref
Mayr, G., De Pietri, V.L., and Scofield, R.P. 2022. New bird remains from the early Eocene Nanjemoy Formation of Virginia (USA), including the first records of the Messelasturidae, Psittacopedidae, and Zygodactylidae from the Fisher/Sullivan site. Historical Biology 34: 322–334. Crossref
Nixon, K.C. 2002. WinClada, Version 1.00.08. Published by the author, Ithaca.
Prum, R.O., Berv, J.S., Dornburg, A., Field, D.J., Townsend, J.P., Lemmon, E.M., and Lemmon, A.R. 2015. A comprehensive phylogeny of birds (Aves) using targeted next-generation DNA sequencing. Nature 526: 569–573. Crossref
Rayner, D., Mitchell, T., Rayner, M., and Clouter, F. 2009. London Clay Fossils of Kent and Essex. 228 pp. Medway Fossil and Mineral Society, Rochester.
Sangster, G., Braun, E.L., Johansson, U.S., Kimball, R.T., Mayr, G., and Suh, A. 2022. Phylogenetic definitions for 25 higher-level clade names of birds. Avian Research 13: 100027. Crossref
Swofford, D.L. 2002. PAUP* Phylogenetic analysis using parsimony (*and other methods), version 4.0b10. Sinauer Associates, Sunderland.
Acta Palaeontol. Pol. 68 (1): 1–11, 2023
https://doi.org/10.4202/app.01049.2022