
Post-collection taphonomy, sampling effects and the role of the collector in palaeontological collections: A case study from an early Late Triassic bone accumulation in southernmost Brazil
FRANCESCO BATTISTA, ANA MARIA RIBEIRO, FERNANDO ERTHAL, and CESAR L. SCHULTZ
Battista, F., Ribeiro, A.M., Erthal, F., and Schultz, C.L. 2023. Post-collection taphonomy, sampling effects and the role of the collector in palaeontological collections: A case study from an early Late Triassic bone accumulation in southernmost Brazil. Acta Palaeontologica Polonica 68 (2): 359–372.
One of the main “databases”, on which palaeontologists carry on their studies, is constituted by palaeontological collections. These collections are the final result of fieldwork and surveys, sampling activities, preparation and curatorial processes, and analyses. However, the content of a palaeontological collection can also be strongly biased, leading researchers to post-collection skewed results. Post-collection biases (e.g., breakage, loss of fragments, etc.) are directly linked to human activities, occurring during excavation, transport, preparation, and storage. Here, we present the case of the vertebrate remains from the Brazilian lower Carnian Santacruzodon Assemblage Zone (Santa Cruz Sequence, Santa Maria Supersequence, Paraná Basin). The studied specimens came from the Schoenstatt Sanctuary fossil site, a key outcrop for both the sequence and Santacruzodon AZ. We evaluated vertebrate remains from three Brazilian scientific collections, compiled through more than 25 years of fieldwork. The specimens housed in the three collections present high degrees of post-collection fragmentation, as well as significant differences in the bone elements present, when comparing cranial vs. post-cranial elements. Moreover, some differences in curatorial attitude have also been noticed, especially in restoration choices, leading to “discrimination” in post-collection fossil quality and highlighting the existence of the “craniocentrism” problem.
Key words: Anthropogenic bias, anthropogenic breakage, Schoenstatt Sanctuary site, Santacruzodon Assemblage Zone, Carnian, Triassic, South America.
Francesco Battista [francesco.battista87@gmail.com; ORCID: https://orcid.org/0000-0002-6154-1906 ], Fernando Erthal [fernando.erthal@ufrgs.br; ORCID: https://orcid.org/0000-0001-8036-192X ], and Cesar L. Schultz [cesar.schultz@ufrgs.br; ORCID: https://orcid.org/0000-0001-7121-0409 ], Programa de Pós-Graduação em Geociências, Instituto de Geociências, Universidade Federal do Rio Grande do Sul, Avenida Bento Gonçalves 9500, Agronomia, 91501-970 Porto Alegre, Rio Grande do Sul, Brazil
Ana Maria Ribeiro [ana-ribeiro@sema.rs.gov.br; ORCID: https://orcid.org/0000-0003-4167-8558 ], Museu de Ciências Naturais do Rio Grande do Sul, Secretária Estadual do Meio Ambiente e Infraestrutura, Rua Dr. Salvador França, 1427, Jardim Botânico, 90690-000 Porto Alegre, Rio Grande do Sul, Brazil; and Programa de Pós-Graduação em Geociências, Instituto de Geociências, Universidade Federal do Rio Grande do Sul, Avenida Bento Gonçalves 9500, Agronomia, 91501-970 Porto Alegre, Rio Grande do Sul, Brazil.
Received 22 December 2022, accepted 25 April 2023, available online 23 May 2023.
Copyright © 2023 F. Battista et al 2023. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Palaeontological collections represent a very important source of data, for understanding past biodiversity, including macroevolutionary studies and biotic changes related to extinctions and biogeographic evolution. They also provide a lot of information useful to revealing and reconstructing palaeoecological relationships ( Ponder et al. 2001; Allmon 2005; Hunter and Donovan 2005; Mannion et al. 2013; Wosik and Guenther 2016; Dean et al. 2020). As is known, however, collections are the results of several different processes that can generate further biases in our geological and palaeontological documentation (in terms of knowledge), since the geo-palaeontological record itself is incomplete (Raup 1972; Sepkoski et al. 1981; Signor and Lipps 1982; Kidwell and Behrensmeyer 1988; Behrensmeyer et al. 2000; Benton et al. 2000; Crampton et al. 2003; Dibble et al. 2005; Alroy 2010; Smith and Mcgowan 2011; Whitaker and Kimmig 2020). During the last five-six decades, at the least, several researchers have worked on these aspects, researching and proposing many statistical models in order to reduce the bias problem and better understand evolutionary trends (see Sepkoski et al. 1981; Signor and Lipps 1982; Alroy et al. 2001; Crampton et al. 2003; Mannion et al. 2013; Benton et al. 2014; Dean et al. 2020 among others).
Nevertheless, palaeontological collections can also be affected by anthropogenic biases, such as historical resampling, sampling by different institutions, selection during sampling activities, as well as different collection methods and techniques applied in the outcrop, during the field-to-lab transport, preparation and storage (Clark and Kietzke 1967; Wolff 1975; Flessa et al. 1992; Tang 2000; Kowalewski and Hoffmeister 2003; Dibble et al. 2005; Whitaker and Kimmig 2020). Sampling by different research teams can also result in collections with different numbers of specimens, and direct studies, as they do not necessarily represent the actual biodiversity of a given outcrop or assemblage zone. Identifying, recognizing, and estimating the effects of the human actions and the resulting biases is necessary in order to reduce them, to improve the quality of the palaeontological collections, and, possibly, to obtain new information, be it taphonomic or taxonomic.
In this work, we analyse quantitatively the results of anthropogenic action on fossil material coming from a unique fossil site from the early Late Triassic (Carnian), the “Santuário Schoenstatt”, located in the central-eastern portion of the Rio Grande do Sul State (Brazil). This specific outcrop plays a leading role for Brazilian and Gondwanan palaeontology and biostratigraphy since its discovery, a specific faunal association known as Santacruzodon Assemblage Zone (AZ) has been recognized for the first time (Abdala and Ribeiro 2010; Soares et al. 2011), which allowed both the refinement of the Brazilian Triassic biostratigraphic framework and a better and more detailed correlation with the rest of Gondwana. The studied specimens have been collected during the last 26 years by researchers from three different Brazilian institutions. The main purpose of this contribution is to evaluate the “weight” of samplers and researchers on a palaeontological collection, and to what extent this can determine differences between different collections, such as the curatorial attention or the faunal representativeness of the same collection.
Institutional abbreviations.—MCN-PV, Museu de Ciências Naturais, Secretaria Estadual do Meio Ambiente e Infraestrutura, Porto Alegre, Rio Grande do Sul, Brazil; MCP-PV, Museu de Ciências e Tecnologia da Pontifícia Universidade Católica do Rio Grande do Sul, Porto Alegre, Rio Grande do Sul, Brazil; UFRGS-PV-T, Paleontologia de Vertebrados, Universidade Federal do Rio Grande do Sul, Porto Alegre, Rio Grande do Sul, Brazil.
Other abbreviations.—AZ, Assemblage Zone; ifr, indeterminate fragments; ith, isolated teeth; ND, not determinable; SCS, Carnian Santa Cruz Sequence; SMS, Santa Maria Supersequence; VG, Voorhies Groups.
Geological setting
The Paraná Basin (Fig. 1) is a large intracratonic depositional basin NE-SE oriented, filled by sediments dating from Ordovician to Cretaceous, and occupying the southernmost part of Brazil, eastern Paraguay, northern Argentina, and the northwestern Uruguay (Holz et al. 2010). In Brazil, Milani (1997) recognized six depositional supersequences, including the Permian/Triassic Boundary, as well as the evidence of the climate evolution from the Mississippian glacial phase (early Carboniferous) towards the late Permian and Triassic aridification. Into the Brazilian portion of the Paraná Basin, the Triassic deposits are found basically in the central part of the Rio Grande do Sul State, with an orientation east-west, whose sediments have accumulated in tectonically controlled basins (Milani 1997; Zerfass et al. 2004, 2005; Milani et al. 2007). The Triassic packages are recognized as belonging to two second-order depositional supersequences, the Induan–Olenekian Sanga do Cabral Supersequence and the Ladinian–Rhaetian Santa Maria Supersequence (Zerfass et al. 2003), separated from each other, as well as from the underlying and overlying units, by depositional gaps (Horn et al. 2014; Milani and Ramos 1998; Soares et al. 2011; Zerfass et al. 2003, 2004).

Fig. 1. Maps showing position of the Santa Maria Supersequence in South America (southern Brazil, Paraná Basin) (A) and the Rio Grande do Sul State (B). C. Location of the outcrops in which the Santacruzodon AZ has been recognized (stars).
The Middle–Upper Triassic Santa Maria Supersequence (SMS) is represented by about 200 m thick ephemeral lacustrine and fluvial red bed deposits (Horn et al. 2014), mainly accumulated in semi-arid to arid environmental conditions evolving towards more humid conditions after the early to late Carnian transition (Horn et al. 2014; Mancuso et al. 2021), as also demonstrated by geochemical evidences (Corecco et al. 2020), perhaps linked to the Carnian Pluvial Episode (CPE) (Dal Corso et al. 2018; Simms and Ruffell 1989, 1990). Four third-order sequences have been recognized within the SMS, characterized by four distinctive faunal associations (Horn et al. 2014; Zerfass et al. 2003), favouring worldwide biostratigraphic correlations (see Martinelli et al. 2017; Melo et al. 2015; Schultz et al. 2020). From oldest to youngest: the Pinheiros-Chiniquá Sequence Dinodontosaurus AZ, dated as Ladinian–?early Carnian; the early Carnian Santa Cruz Sequence (SCS), characterized by the Santacruzodon AZ. These first two units would have been deposited in increasingly more arid conditions (Corecco et al. 2020; Horn et al. 2014). The third sedimentary package, dated Carnian to Norian, is known as Candelária Sequence, in turn divided into two portions according to two different faunal content (Hyperodapedon and Riograndia assemblage zones, respectively lower and upper portion). Finally, the Mata Sequence, possibly Rhaetian in age, is devoid of a diagnostic faunal content, but is rich in plant remains.
Between the depositional sequences of the SMS, the early Carnian SCS is the smallest of the four depositional sequences of the SMS, and only identified in the area between the cities of Venâncio Aires, Santa Cruz do Sul and Vera Cruz, in the central-eastern portion of the Rio Grande do Sul (Horn et al. 2014) (Fig. 1). For this sequence, a maximum age of 236.6±1.5 Ma has been provided by Philipp et al. (2018) through U-Pb method in dating detrital zircons. According to recent studies (Corecco et al. 2020; Mancuso et al. 2021), this sedimentary package, as well as the underlying Pinheiros-Chiniquá Sequence, is deposited in an ephemeral braided river system dominated by aeolian deposition in an arid to semi-arid context, as suggested by apparently massive loess deposits alternating with commonly less-thick sandstone and conglomeratic sandstone layers, and palaeosoils (Horn et al. 2014, 2018). In a biostratigraphic point of view, due to the great abundance of traversodontid non-mammalian cynodonts, Abdala et al. (2001), has defined the Traversodontidae Biozone. Later, the biozone was renamed as the Santacruzodon AZ by Abdala and Ribeiro (2010) and formalized by Soares et al. (2011), based on the high abundance of the traversodontid Santacruzodon hopsoni Abdala and Ribeiro, 2003, found in all known localities (Horn et al. 2014).
The specimens found in the Santacruzodon AZ are generally characterized by the presence of a thin dark red to blackish layer of manganese oxides and carbonate concretions (Reichel et al. 2005), as well as by the near absence volumetric distortion linked to permineralization processes and common phenomenon in other assemblage zones of the SMS (Holz and Schultz 1998; Reichel et al. 2005).
The studied specimens came from the Schoenstatt Sanctuary fossil site (“Santuário Schoënstatt/Schoënstadt”, in Portuguese, henceforth simply Schoenstatt), an extremely cynodont-rich fossiliferous deposit discovered by one of the authors (CLS) and Max C. Langer in 1995, in the Santa Cruz do Sul municipality in the Rio Grande do Sul State (Brazil), about 150 km from the capital city Porto Alegre (Fig. 1), during a palaeontological survey.
A taphonomic analysis was carried out by Bertoni-Machado (2004) and Bertoni-Machado and Holz (2006). In detail, Bertoni-Machado and Holz (2006), based on an apparent abundance of skull and mandibular remains, in comparison to axial and appendicular remains (Bertoni-Machado and Holz 2006: table 1), suggest that the Schoenstatt is the result of a strong biogenic control by selective predation and scavenging activities. These predatory activities were probably favoured by the presence of ephemeral and/or seasonal waterholes, like lakes or ponds, and whose level and presence was mainly controlled by the water table (according with more historical and recent studies, as Holz and Schultz 1998; Horn et al. 2014, 2018; Reichel et al. 2005).
Material and methods
The studied specimens are stored in the three biggest palaeontological collections of the Rio Grande do Sul, in Porto Alegre: Museu de Ciências Naturais, Secretaria do Meio Ambiente e Infraestrutura (MCN/SEMA-RS), Museu de Ciências e Tecnologia da Pontifícia Universidade Católica do Rio Grande do Sul (MCP), and Universidade Federal do Rio Grande do Sul (UFRGS).
The term “specimen” is understood here as a piece to which a catalog number has been assigned. Each specimen may be represented by a single bone element, by multiple associated bone elements (in anatomical connection or as a whole), or contain different bone elements whether complete or fragmentary as well as a group of indeterminate fragments.
The material has been collected over the past 26 years, beginning with the deposit discovery in 1995. Information about activities and time of sampling, researchers participating in it, and taxonomic attribution, are based on the archiving catalogues of the three collections, as well as on filed notes, when available.
Whenever possible, the sampling dates were indicated in full (yy-dd-mm), although this was not always possible (SOM 1, Supplementary Online Material available at http://app.pan.pl/SOM/app68-Battista_etal_SOM.pdf). Thus, also taking into account less complete information, such as “yy-mm” and only “yy”, in order to verify if there is a trend in sampling linked, for example, to different generations of researchers. Thus, sampling dates has been summarized in chronological order, as shown in Fig. 2.

Fig. 2. Timeline of the sampling activities carried out at the Schoenstatt site by the three institutions.
Voorhies (1969) observes that the skeletal remains of vertebrates, once disarticulated, when subjected to water flows, have different hydraulic behaviours and susceptibility to fluvial transport. In according to their shape, size and density, Voorhies (1969) identifies three main groups plus two intermediate ones, the so-called “Voorhies Groups” (sensu Behrensmeyer 1975; henceforth VG), from immediately moved (VG I; e.g., ribs and vertebrae), to gradually removed (VG II; long bones), up to the last ones that are more difficult to remove forming a lag deposit (VG III; skulls and mandibles). Here, we build on these groupings to visually quantify the effect of sampling and how it affects the resulting collections, by graphically dividing the bone elements. In the obtained scheme (Fig. 3), articulated remains, concretionary blocks containing bones, and indeterminate fragments are also counted. Due to a great abundance of indeterminate fragments and concretionary blocks with embedded bones, including many articulated and semi-articulated skeletal portions, additional “groups” have been included, such as “indeterminate fragments” (ifr) and “not determinable” (ND). All bone fragments whose anatomical origin is not certain are considered as ifr, including those whose fragmentation is of anthropogenic origin. In ND are included articulated and semi-articulated skeletal segments, embedded bones, and indeterminable elements (elements and fragments in which nothing can be recognized due to concretions). Isolated teeth (ith), wholes or fragments, root or occlusal portions, although commonly included into the VG III (together with skulls and lower jaws), even were counted separately. This choice is due to the fact that a lot of maxillae and dentaries present empty alveoli. At the same time, several teeth in each collection are broken, and complementary counterparts could be stored in different institutions. Therefore, the quantities here reported do not represent the totality of identified bone remains from the outcrop, since articulated, semi-articulated and embedded elements are considered as single units.
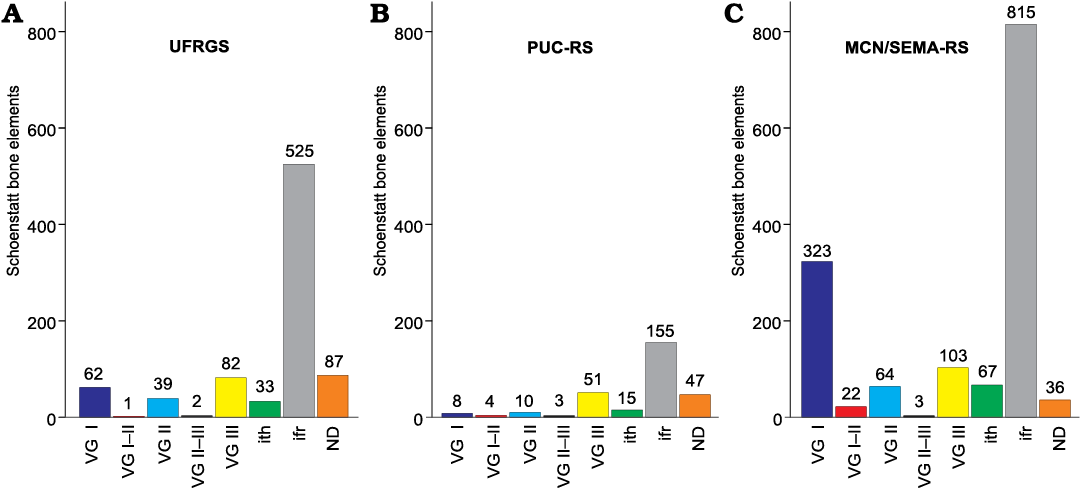
Fig. 3. Results of the sampling by three institutions (A–C) schematized into the Voorhies Groups (Voorhies 1969, sensu Behrensmeyer 1975; modified), favouring biases in data interpretations and palaeoecological reconstructions. Abbreviations: ifr, indeterminate fragments; ith, isolated teeth; ND, not determinable; VG, Voorhies Groups.
To quantify anthropogenic damages and destructive effects of excavation, transport, preparation and storage procedures, each specimen, whether catalogued or not, has been analysed, in order to recognize fresh fractures due to human actions and not related to fossilization. Several catalogued specimens contain a variable number of elements. Thus, these data have been grouped in the SOM 1. Here, the percentages of the anthropogenically-fractured elements were calculated in comparison to the total, as well as the percentage of the restored (“glued”) elements have been compared to the fractured ones. Moreover, according to the sampling dates, the samples have been plotted in line graphs according to the following groupings: catalogued specimens (all the numbered and registered specimens in the archives of the collections), elements per specimens (the real number of bony elements, whole or fragmentary elements, determinable or not, present within the same catalogue number), anthropogenic breakage (all elements showing fresh, post-discovery fracturing due to the human action), and glued elements (all those elements that, after fracturing, have been the subject of preparation, even partial, and reunification with the corresponding fragmented parts). The comparison among the three collections is shown in Fig. 4.
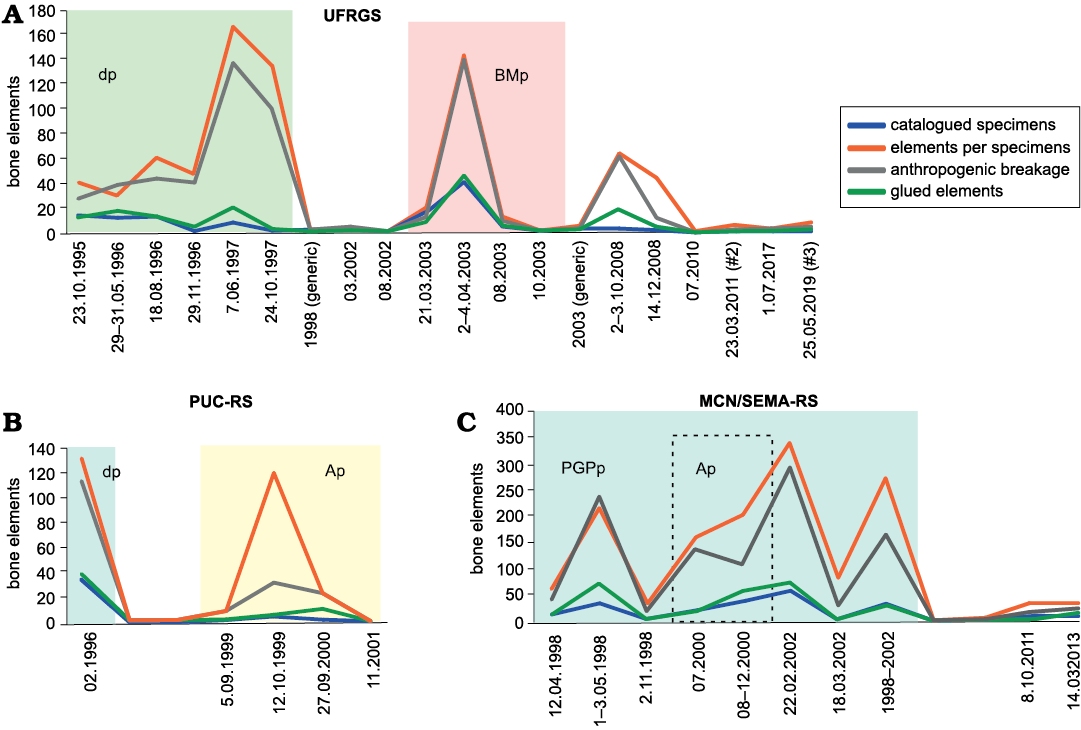
Fig. 4. Comparison between the three institutions (A–C) and relationship with sampling pulses over the years. Histograms are aligned and calibrated with regards to timeline. The dashed rectangle in C indicates the period of collaboration between MCN/SEMA-RS and PUC-RS. Abbreviations: Ap, Abdala pulse; BMp, Bertoni-Machado pulse; dp, discovery pulse; PGPp, “Programa Pró-Guaíba” (“Pró-Guaíba” Program) pulse.
Finally, a comparison between the restored elements was carried out (SOM 2), in order to verify if there was any type of selection (cranial vs. post-cranial vs. others) during preparation and potentially subsequent study or archiving, as well as between the recognized taxonomic groups (non-mammalian cynodonts vs. archosauriforms vs. others vs. rhizoliths), or not, indicating that there was no discrimination during sampling and preparation (SOM 3). In other words, we erected the following hypothesis for statistical purposes: H0, there was no biased selection between cranial vs. post-cranial vs. others (or taxonomic group), and all sampling effort since 1995 reflects random collection of fossils from the outcrop. H1, the recovery of fossils, as well as their treatment and storage were biased towards some cranial element (or taxonomic group). These null and alternative statistical hypotheses were tested using contingency tables and a Chi-squared test, using the free access software PAST v.4.11 (Hammer et al. 2001).
The photographs were taken with a digital camera Canon PowerShot A3150 IS 12.1 megapixels.
Results
Sampling timeline.—Since the discovery in October 1995, the Schoenstatt was an object of approximately continuous palaeontological surveys, excavation and sampling activities (Fig. 2; SOM 1) carried out by researchers of the three different institutions mentioned above.
The first sampling was carried out by UFRGS researchers, when the outcrop has been discovered. During that time, the sampling was mainly on the surface of the outcrop, producing a total of 14 catalogued specimens.
In February 1996, only sampling by PUC researchers was carried out, producing a total number of 131 collected remains grouped in 34 catalogued specimens, including both whole and fragmentary bones, and a segment of backbone composed by four articulated vertebrae, including ribs fragments also articulated/semi-articulated.
Between 1996 and 1998, six palaeontological surveys were carried out by UFRGS. During this time, 36 specimens were added to the collection. From April 1998 to March 2002, several surveys were carried out by the PUC-RS and MCN/SEMA-RS teams, some of these carried out in close collaboration (AMR, personal observation). Over four years, the researchers of the PUC-RS performed three expeditions, producing seven catalogued specimens, which contain an amount of 152 elements, including 22 rhizolith samples. Meanwhile, 1358 elements were collected by MCN/SEMA-RS, divided into 190 catalogued specimens.
During March 2002, UFRGS researchers carried out two different expeditions and, during almost all of the following 10 years, all the subsequent palaeontological surveys. During this long time, the main and richest samplings took place in 2003, a period during which this bone accumulation was the subject of a Master’s thesis with a taphonomic focus. A total of 77 catalogued specimens and 300 elements were collected by UFRGS in one decade (2002–2011).
Between late 2011 and early 2013, two more palaeontological surveys were carried out by MCN/SEMA-RS team, producing 14 new catalogue numbers and 66 elements.
Finally, the last known sampling was done by UFRGS in May 2019.
Summing up, over the last 26 years a great quantity of fossil material was collected in the Schoenstatt outcrop, resulting in 407 catalogued specimens (including some not catalogued specimens, n = 4), and 2562 elements. This amount includes 63 uncatalogued elements, some unprepared/partially-prepared blocks, and many articulated/semi-articulated bones (portions of axial and appendicular skeleton), as well as some rhizoliths (n = 23). As shown in the SOM 1, the total of the sampled material in this outcrop is divided as follows among the three palaeontological collections: 151 specimens per 831 elements for the UFRGS, 45 and 298 for the PUC-RS, and 211 and 1433 for the MCN/SEMA-RS.
Sampling products.—As introduced above, to observe the results of more than 25 years of sampling in the outcrop, we have summarized the samples into the “Voorhies Groups” (Voorhies 1969, sensu Behrensmeyer 1975), as summarized in the Fig. 3 and Table 1.
It is important to state that, although in a general point of view we refer to bony elements, during the palaeontological surveys some rhizolith fragments and coprolites (with doubt about the latter: Heitor Francischini, personal communication 2020) were collected. However, their quantity is not such as to alter the results and considerations of this work. In any case, as these samples are part of the collected material, they have been included here in ND group.
Table 1. Bone elements collected by the three institutions between 1995 and 2019, grouped in the Voorhies Groups (VG; Voorhies 1969, sensu Behrensmeyer 1975; modified). Abbreviations: ith, isolated teeth; ifr, indeterminate fragments; ND, not determinable elements.
|
Institution |
VG I |
VG I–II |
VG II |
VG II–III |
VG III |
ith |
ifr |
ND |
Total |
|
UFRGS |
62 |
1 |
39 |
2 |
82 |
33 |
525 |
87 |
831 |
|
PUC-RS |
8 |
4 |
10 |
3 |
51 |
15 |
160 |
47 |
298 |
|
MCN/SEMA-RS |
323 |
22 |
64 |
3 |
103 |
67 |
815 |
36 |
1433 |
|
TOTAL |
393 |
27 |
113 |
8 |
236 |
115 |
1500 |
170 |
2562 |
Anthropogenic breakage.—During the review and analysis of the studied specimens, the presence and abundance of a great number of fractured elements was noticed. In an attempt to recognize the various types of fractures (pre-burial, post-burial, and/or post-discovery), we have seen that most of the elements, of all three institutions, present an extremely important human contribution. On a total of 2562 elements, 1940 are affected by human action, corresponding to 75.72% of all the collected material. The anatomical regions most affected by anthropogenic fractures were found to be cranial (both skulls and jaws), axial (apophyseal and spinal regions of the vertebrae) and thoracic (ribs).
On an amount of 831 elements present in the UFRGS collection, 689 elements (82.91%) are characterized by fresh fractures while, in the MCN/SEMA-RS, 1072 elements on 1433, the 74.81%.
Albeit with much smaller quantities than the previous two collections, a different path seems to have been followed by the PUC-RS, which has much lower percentages of damage due to human action, with only 60.07% of broken elements (179 of 298).
A schematized summary of this phenomenon is shown in SOM 1, including each sampling event with corresponding date, undated catalogued specimens, undated uncatalogued specimens, and “not found” catalogued specimens. SOM 1 also shows the ratio, expressed in percentage, between the broken and glued elements.
“Glued” elements.—As mentioned above, a high number of broken elements were recognized (75.72% of total). Despite most of the material still needing additional preparation, a partial restoration and relocation of the broken elements with their counterparts is already underway. In fact, both during this study (a small part) and during the more than 25 years since the first sampling, several broken bone elements whose counterparts were recognized, damaged and fractured during sampling-transport-preparation-storage processes, were re-associated, repositioned and glued together. The identified fragments were treated and bonded with Paraloid B72 resin, diluted in different concentrations of acetone. Considering the number of anthropogenically affected elements (n = 1940), re-associated elements (n = 509) represent just over a quarter of the damaged elements (SOM 1). Nevertheless, this value decreases up to a little less than 20% when compared with the total number of sampled elements.
Observing in detail, the collection of the PUC-RS contains roughly 32% re-associated fragments (n = 58), in relation to broken elements, decreasing up to less than 20% if compared with the total amount. Meanwhile, MCN/SEMA-RS and UFRGS collection shows less difference between partial and total values. In the MCN/SEMA-RS collection, 273 glued samples represent 25% of the damaged bones, decreasing to less than 20% in comparison with the total from this collection. A similar variation is also observed in UFRGS collection. A total of 178 elements were restored over time, 25% of the 689 broken units. In comparison with the total amount, this percentage decreases to 21%.
Although the number of glued elements is far from the totality of the broken ones, in some cases, the recognition of corresponding parts has favoured a better osteological and anatomical identification, as summarized in SOM: table S1 and decreased the number of indeterminate fragments.
In a general point of view (SOM 2), however, the PUC-RS seems to be the one with the most “neutral” attitude, roughly paying the same curatorial attention (H0) to the three examined categories (Χ2 = 7.5849, p = 0.02254). On the other hand, with respect to curatorial activities, UFRGS (Χ2 = 134.85, p < 0.000001.) and MCN/SEMA-RS (Χ2 = 101.65, p < 0.00001) researchers have generally treated these classes differently. This disproportion increases (H1) on the UFRGS-side if considering the difference with the MCN/SEMA-RS collection with respect to the total number of specimens/elements-per-specimen (number of specimens: 151 vs. 211, elements: 831 vs. 1433).
On the contrary, examining the curatorial attention paid in order to reduce the trephic factors (post-collection and curatorial factors), but with respect to the taxonomic groups (SOM 3) kept in the collections, the UFRGS would seem more biased in its choices and restoration activities (Χ2 = 32.014, p < 0.000001) than both MCN/SEMA-RS (Χ2 = 9.0752, p = 0.010699) and PUC-RS (Χ2 = 3.5378, p = 0.3159). These results suggest a different way to approach the various groups by UFRGS, with more attention paid to a specific fossil group (H1). On the contrary, both PUC-RS and MCN/SEMA-RS seem to apply a more neutral operation mode (H0).
Discussion
The content of a collection is commonly affected by the interest of the human activity and research. This kind of bias is generally known as “sampling selection bias” or, briefly, “sampling bias” (Qin 2017), and this is due to sampling in a non-random mode. In naturalistic collections, this bias has been seen to be related even to other factors, such as animal behaviour (Biro and Dingemanse 2009; Stuber et al. 2013), “showier” morphologies of males compared to females (Cooper et al. 2019), proximity of the sampling point with respect to communication routes as well as altitude, climate, etc. (Hijmans et al. 2000), among others. Palaeontological collections do not vary far from this. On the contrary, if the problem of the behaviour and reactions of living organisms does not arise, the nature and extent of the outcrops can cause considerable variations in the completeness of the fossil record. This, together with various factors related to geological and fossilization processes, or the ease of conservation of some fossil remains compared to others, also affects sampling (Raup 1972; Kidwell and Holland 2002; McGowan and Smith 2011; Mannion et al. 2013). Other sources of bias that can affect the content of a palaeontological collection, especially and above all, a collection of fossil vertebrates, is directly linked to the choices, conscious or not, of the sampler. This phenomenon, which sees the exclusion of the more fragmentary or incomplete specimens in favour of the most complete or articulated, has been defined as Ugly Fossil Syndrome by Tang (2000), and it is easily and clearly applicable to the cranial/post-cranial “dualism” (but this can to be valid also within invertebrate collections: Walker 1989). Even historical re-sampling on the same outcrop, as well as the sampling works carried out by different and several institutions, and the use of different sampling techniques due to different purposes, are a source of bias already known for a long time. For example, Clark and Kietzke (1967) recognize notable differences in the sampled material obtained through different methods of moving and prospecting (on horseback, on foot, or kneeling on the ground), or by comparing distinct prospecting campaigns that took place over several years, or by different sampling methods (block sample to be prepared or bags of sediment to be screen-washed). All these sources of possible bias are grouped into the sullegic factors by Clark and Kietzke (1967). Obviously, it is not to be excluded that during and after the discovery and collection of a fossil specimen, and subsequently, during its packaging, transport, preparation, study, and storage, other phenomena may occur and interfere with the quality of the fossil record, as well as with the quantity and quality of recoverable information. Clark and Kietzke (1967) identify these phenomena of post-collection taphonomy and define them as trephic factors. For example, Flessa et al. (1992) demonstrated how the kind of transport vehicle and the conditions of the communication route travelled, as well as the distance and time used, can cause alteration (in their case, fragmentation) of the collected material. Although the Flessa et al. (1992) experience was made by observing fossil remains of invertebrate organisms (bivalve molluscs), the anthropogenic phenomenon of post-collection fragmentation can also affect vertebrate remains, especially depending on their degree of fragility and resistance to vibrations, jolts and shocks, which can occur during transport, resulting in generation of additional taphonomic biases. Also, post-collection damage and fragmentation due to curation is a common phenomenon known in other areas, such as archaeology and zooarchaeology, for example, leading archaeologist to have to do “archaeologies of archaeology” (Gifford-Gonzalez 2018: 149).
The case presented here represents a good example of how much the actions and attentions of collectors and researchers can affect a collection, determining important biases that can to lead to incorrect palaeoecological reconstructions, for example, being based on incomplete data or because the ensuing datasets have been strongly influenced by human choices and actions.
Since the discovery of the Schoenstatt Sanctuary fossil site in 1995, more than 2500 bone elements have been collected, corresponding to about 400 catalogued specimens (SOM 1). Nevertheless, since the sampling was performed by three different institutions, at different moments, using different techniques and organization of the field activities, much of the information is extremely poor and fragmentary. This is valid for the stratigraphic sampling position, for taphonomic aspects, as well as concerning the actual biodiversity contained in the deposit, the available information are extremely poor and fragmented within the collections themselves. In our study, we focused on specific aspects of post-collection taphonomy, such as the sampling timing and results post-collection bone fragmentation, and subsequent curatorial restoration activity, in an attempt to quantify how much our actions, as researchers, can influence the content of a given scientific (palaeontological) collection and, consequently, generate bias in our studies and in the obtained results.
In Fig. 4, for each institution two or three “sampling pulses” are recognizable, including “peaks” which representing the amount of collected materials. The first peaks identify the first three years of sampling (1995–1998), during which a large number of samples (n = 911) were collected (UFRGS = 477, PUC-RS = 131, MCN/SEMA-RS = 303). The overlapping of sampling activities, shared between UFGRS and PUC-RS during the first years, is due to the fact that students, from the both institutions, were trained and co-oriented by tutors from the both institutions. The second pulse carried out by researchers of the UFRGS is linked to the fieldwork related to the Master’s thesis project of Cristina Bertoni-Machado (see Bertoni-Machado 2004), focusing on the taphonomic analysis of the deposit, representing the most continuous sequence of sampling operated by UFRGS. A separate discussion is made about the main sampling pulses carried out by MCN/SEMA-RS. Although they occurred over three different and not continuous years (1998, 2000, 2002) and at different times, in Fig. 4 we have grouped them as a whole, as they are linked to a single project developed by MCN/SEMA-RS between 1998 and 2002 (“Programa Pró-Guaíba”). Furthermore, in those same years there was a close collaboration between MCN/SEMA-RS and PUC-RS, under the direction of one of the authors (AMR) and Fernando Abdala (currently at CONICET, Argentina), highlighted in Fig. 4C by the dashed rectangle. During this five-year-long pulse, the vast majority of the specimens kept by the MCN/SEMA-RS were sampled (1358 on 1433). Now, if on the one hand it is obvious to consider that the attention of the three institutions was not concentrated on a single—although extremely rich in fossils—outcrop, on the other it is evident that a higher and more continuous field work, such as those carried out by a financially supported Master's project or by MCN/SEMA-RS, are strongly linked to access to funding. This source of bias was pointed out also by Whitaker and Kimmig (2020) in their evaluation of the biases that can affect a natural history collection, due to researcher activities, choices, and economic limitations.
In their work about post-collection taphonomy and breaking. Flessa et al. (1992) pointed out the differences that occur through two different modes of transport. In our case, there is no difference in the type of transport, with transport on wheels the only one practicable in the study area. Nevertheless, the effects of excavations, sampling activities, transport, and preparation are clear. In SOM 1 are shown the percentages of anthropogenic breakage recognized in the collections, relating both to each samples as to their whole, in the three collections. In some cases, the number of anthropogenically broken elements is the same as the number of elements per catalogued specimens (% anthropogenic breakage = 100%), while others greatly exceed the number of any intact elements (up to 133%). Such high percentages (above 100%) are due to the fact that specimens represented by single bone elements, as well as specimens containing multiple elements catalogued together, present multiple breaks, generating one or more main bodies plus a quantity of fragments deriving from them, but counted individually for the purposes of the study.
With respect to the glued elements, there also appears to be some difference in curatorial attitude by the three institutions. We consider “cranial” elements as the sum of [(VG II–III + VG III + ith) + all the anthropogenically broken fragments belonging to previous one], “post-cranial” as [(VG I + VG I–II + VG II) + all the anthropogenically broken fragments belonging to previous one], and “others” as [(ifr + ND) + all the anthropogenically broken fragments belonging to previous one]. Analysing it makes it possible to recognize differences among the three institutions. Relative to cranial fragments, of 130 bone fragments, more than 50% were the object of curatorial attention and preparation by the crew of the Vertebrate Palaeontology Laboratory of UFRGS, while this percentage decreases to nearly 30% in the case of both Palaeontology Laboratory of the PUC-RS and Palaeontology Section of the MCN/SEMA-RS. This trend is different considering the attention paid to post-cranial broken elements, in which just over 50% of the elements have been the subject of restoration processes in the UFRGS and PUC-RS labs, while approximately 40% from MCN/SEMA-RS (145 of 354; see SOM 2) were restored. A positive gap achieved by the PUC-RS crew compared to the other two, results from the attention placed on the category “others”, even if the amount is at least three times lower.
However, the same results presented here could be affected by bias, due to the historically recognized abundance of non-mammalian cynodonts remains found at the Schoenstatt (and, in general, attributed to the entire Santacruzodon AZ: Schultz et al. 2020, and reference therein). This is possible because, due to this extremely high abundance (approx. 93%: Schmitt et al. 2019, based on published species/specimens plus unpublished specimens kept in the UFRGS collection), a lot of indeterminate fragments may still be erroneously attributed to Cynodontia. During the revision of the Schoenstatt material, both complete and fragmentary bone elements previously associated with cynodonts have been anatomically identified as non-cynodonts remains and separated from Therapsida, already being the subject of further analysis.
Between the consequences of post-collection breakage, there also is a high degree of secondary fragmentation, followed by subsequent erroneous cataloguing. In SOM: table S1 we show some specimens and bone elements altered after revision, identification, and restoration. Obviously, they are not the only elements subject to post-revision modification, but those reconstituted starting from different archiving numbers. There are a lot of bone elements (e.g., vertebrae, ribs, lower jaws portions, limb bones) assembled from various fragments from the same “specimen/catalogue number”, and which continue to be grouped under the same identification number and belong to same original “specimen”.
A great abundance of trephic effects have been recognized into the UFRGS collection. The most striking examples are provided by two blocks containing a lot of cranial and post-cranial bone elements still embedded in the carbonate concretion, the specimens UFRGS-PV-0453-Ta and UFRGS-PV-0453-Tb (Fig. 5). Parts of these, previously identified as “indeterminate fragments”, were identified among several others contained in eight different boxes. Among these, an extreme example of bias due to trephic factors is provided by a fragment stored in the UFRGS-PV-0456-T, a group of fossil materials collected in the “Caixa de Abelhas” fossil site, in Venâncio Aires municipality, but belonging to UFRGS-PV-0453-Ta. This fact does not necessarily indicate little curatorial attention by UFRGS researchers, since both outcrops (“Schoenstatt Sanctuary” and “Caixa de Abelhas”) contain fossil remains of the Santacruzodon AZ, presenting basically the same preservation and diagenetic features, as well as faunal content. This mixture may have occurred during years of study and manipulation of the material of this AZ (the specimens are kept in the same shelf, although in different boxes), and the possibility of error related to this disassociation is to be taken into consideration. Quite the opposite, this further “re-discovery” must represent a new “stimulus-and-alarm” in carrying out a new and in-depth revision of the collection, of both most complete and representative specimens, as well as of the most fragmentary, through a careful and painstaking comparison between the sources of information available (catalogue book, datasheet, field notes, and historical memory of the collectors).
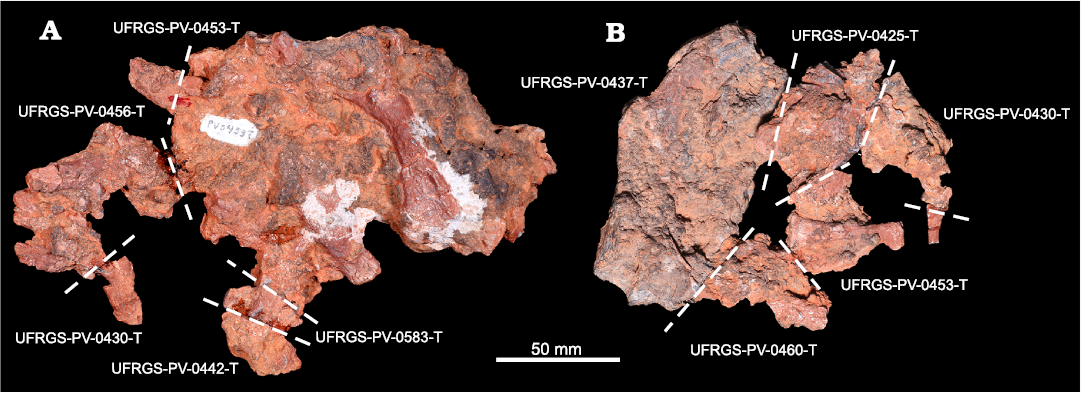
Fig. 5. Selected specimens from the UFRGS’ collection showing the effect of trephic factors. The blocks UFRGS-PV-0453-Ta (A) and UFRGS-PV-0453-Tb (B) were affected by fragmentation and the several fragments stored with different catalogue numbers.
Another example is represented by the actual UFRGS-PV-0876-T, a fragmentary lower jaw referred to Santacruzodon hopsoni Abdala and Ribeiro, 2003, by Melo et al. (2022). This specimen (Fig. 6A) was previously divided between three different catalogue numbers: UFRGS-PV-0874-T, an indeterminate fragment, UFRGS-PV-0876-T, a lower jaw fragment, and UFRGS-PV-0950-T, an indeterminate fragment grouped with several other indeterminate fragments. The same problem has been noticed for the specimen UFRGS-PV-0911-T (Fig. 6B), a fragmentary non-mammalian cynodont lower jaw, reassembled with the complementary part UFRGS-PV-0933-T (lower jaw fragment) and with some fragments identified in the UFRGS-PV-0915-T, which contained more than 40 fragmentary elements (including a tooth, a humerus, a neural arch, and an ilium).
With respect to the MCN/SEMA-RS collection, in addition to a large number of individual elements, both whole and fragmentary, the main example of curatorial bias is represented by the set consisting of MCN-PV-10114 (thoracic ribs), MCN-PV-10128 (femur, vertebrae, and ribs), MCN-PV-10129 (rib and two vertebrae), and MCN-PV-10156 (rib fragment) (Fig. 6C). These specimens, although separated during preparation and cataloguing, if repositioned in their original reciprocal position, could represent one of the clearest evidences of possible hydraulic process and orientation, in which femur and ribs appear almost iso-oriented, and the set of vertebrae grouped, aligned, and blocked by the femur (attritional accumulation). From a taphonomic point of view, this information can represent an important datapoint for deciphering the genesis of the deposit. On the contrary, the analysis of these four specimens, in a separate way, will lead to a biased interpretation. This phenomenon may be linked to the fact that the preparation of the specimen, and consequent isolation of the individual bone elements, was performed with greater attention to the systematic attribution of the specimen rather than its taphonomic context (AMR personal observation).
Although its fragmentation is not attributable to human action, an interesting case is provided by the specimens MCP-PV 4017 and MCP-PV 4028 (Fig. 6D), until now catalogued as a proximal end of right femur and an indeterminate fragment, respectively, kept in the PUC-RS collection. After revision, MCP-PV 4028 has been identified as a distal end of a right femur and complementary to the specimen MCP-PV 4017. In this case, however, the breakage is not referable to the time of sampling, nor to lack of attention during transport or preparation. Indeed, the two parts of the femur are not characterized by fresh breakage, while there are recognizable evidences of post-exhumation reworking, possibly occurring in recent times. So, this specimen represents a mixture of recent reworking processes and trephic factors, the curatorial and the archiving work being the source of the problem.
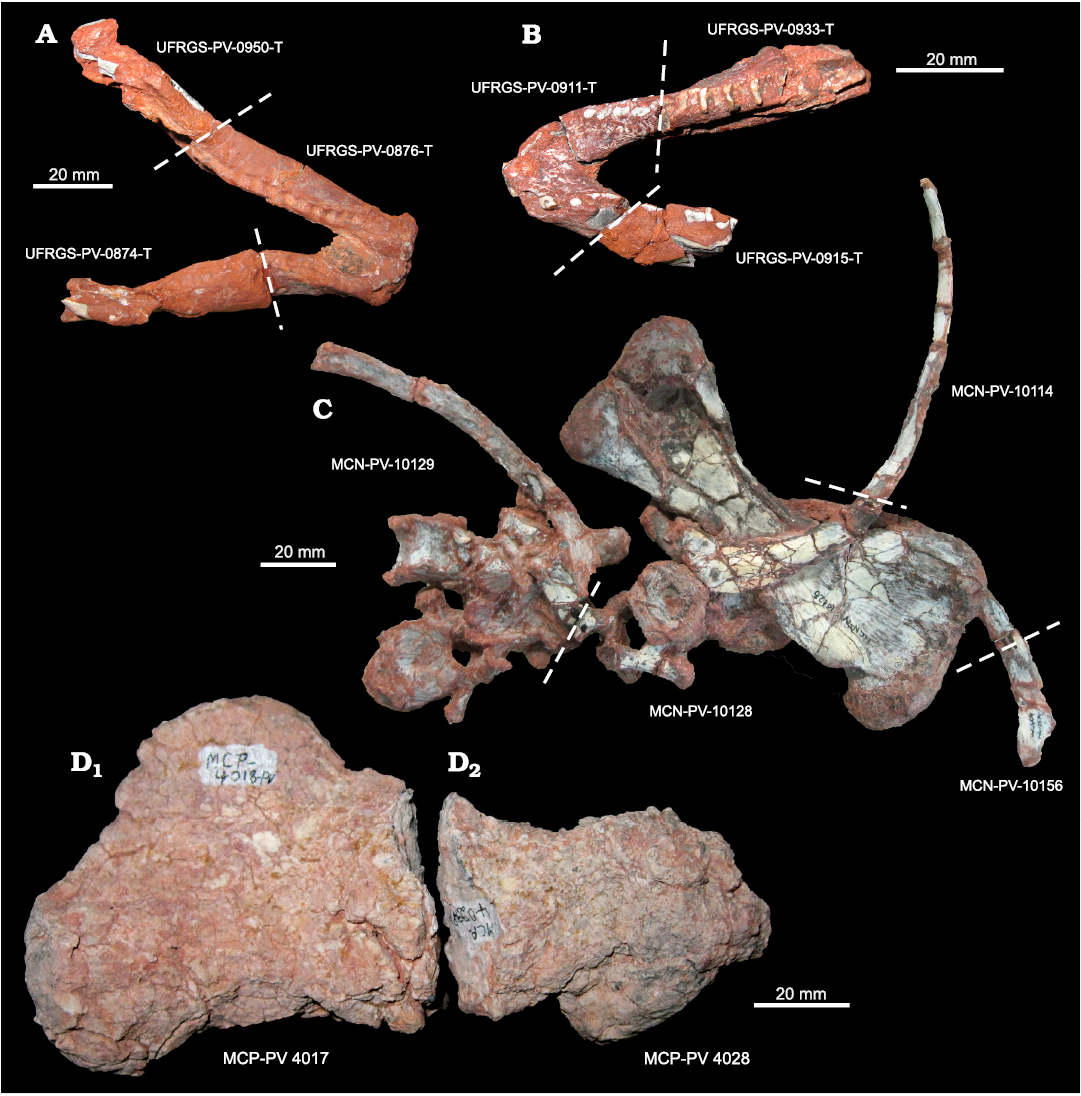
Fig. 6. Selected specimens from the three collections showing effects of sullegic (i.e., during sampling activity), trephic (i.e., from transport to preparation and storage) and “taxonomical” factors. A and B show two mandibles of traversodontid cynodonts, of which previously just two out of ten fragments were considered as such, while the rest as indeterminate fragments. In C, several specimens from a single original association, numbered separately, and now together again, still being able to provide taphonomic information. Some of them (e.g., rib fragments) had been associated with similar others and analyzed with systematic interest, losing the taphonomic information. D shows two parts of the same bone (D1, MCP-PV 4017; D2, MCP-PV 4028), a right femur, encountered already separated and isolated, and catalogued as two different specimens, part in D2 being identified as an indeterminate fragment.
One last thought is related to what is observed and described in Fig. 4 and SOM 1, that leads us to think about the content of the three collections. Any consideration on the Schoenstatt faunal content should be strongly biased, if the total amount were not availed in full. Those differences are easily recognizable in reading the works published by Abdala et al. (2001), in the first description of the new cynodont-rich fauna identified at the Schoenstatt outcrop and the proposal of the “Traversodontid Biozone”, resulting in 82 cynodont specimens, in comparison with (Bertoni-Machado and Holz 2006: table 1, lower jaw n = 12), and Soares et al. (2011: table 1, cynodont specimens n = 116), which describe a new probainognathian cynodont unearthed in 1998. Comparing the results obtained and proposed by Abdala et al. (2001) and Soares et al. (2011), it is possible to recognize a “linear” increase of the number of the specimens, which is what we would expect as consequence of new excavations over the years. The results offered by Bertoni-Machado and Holz (2006), on the other hand, appear to be against the trend, who evidently did not consider in their entirety all the specimens already collected and stored in the other collections. This remarkable difference is perhaps linked with the focus of the research, since the Bertoni-Machado and Holz (2006) work focuses on taphonomic features, while the other two focus on biostratigraphy (Abdala et al. 2001) and taxonomy (Soares et al. 2011), both paying greater attention to the total faunal content (although this aspect should also be abundantly included in a taphonomic work). If we consider that until 2003, during which sampling was carried out by Bertoni-Machado and UFRGS team, the majority of the specimens were already sampled, considering the three institutions (2303 on 2562), so the results published by Bertoni-Machado and Holz (2006) could be biased and not representative with respect to the bone accumulation. In this sense, Fig. 3 shows the differences in fossil content kept by the three institutions, schematized in the Voorhies Groups (Voorhies 1969, sensu Behrensmeyer 1975; modified). The difference between the collections is remarkable and shown in Fig. 3. If we analyse separately the content of UFRGS, PUC-RS, and MCN/SEMA-RS, we can get two or three different results: (i) a skull-dominated bone accumulation (almost a lag deposit), (ii) a nearly skull-dominated concentration, or (iii) an accumulation in which some anatomical elements have already been transported away (but not enough to suggest a high energy). Therefore, it cannot be excluded that the work of Bertoni-Machado and Holz (2006) was not influenced by the results proposed in (i). However, a more in-depth review of taphonomic evolution of the Schoenstatt bone accumulation is in progress, and is not the subject of this study.
Conclusions
Palaeontological collections are the final product of the sampling effort and one of the most important “databases” for palaeontologists. Even a long time after a specimen has been collected, it can still continue to provide new information, in accordance with the techniques applied, which are constantly evolving. However, being the product of human action, they can be affected by biases and suggest erroneous results (e.g., in palaeoecological reconstruction). These types of biases are known for a long time and researchers have to pay close attention to them, trying to reduce them. However, this is not always so easily applicable, since anthropogenic biases do not arise only during sampling. In fact, problems such as secondary fragmentation, or displacement from one box to another, are quite common phenomena.
In this study, we have analysed in a quantitive way how the effects of sullegic and trephic factors can affect a palaeontological collection. In the “Schoenstatt Sanctuary” outcrop, since its discovery in 1995, more than 2500 complete and fragmentary bone elements have been collected. The sampling was carried out by three different institutions, producing a first very important bias: the division of the fossil content. Moreover, several remains have been collected over the years by undergraduate students, especially during didactic fieldtrips of the degree course, concurring to generate biases in specimen “quality” (e.g., completeness, size, etc.) and deficit in data sampling (e.g., stratigraphic position, orientation, etc.). This aspect takes on even greater dimensions if we consider that the three collections now contain very different quantities of specimens. So, any consideration, any study about the faunal content, or of a taphonomic or palaeoecological type, will be strongly biased, if done without considering the total amount of the collected material. Thus, a comprehensive review of the taphonomy was also needed in order to reduce the existing biases related to sampling methods, historical resampling and fractioning, so as to provide a better palaeoecological reconstruction of Schoenstatt Sanctuary fossil site, which will be discussed in another study.
During the revision of the Schoenstatt material, we noticed how post-collection fragmentation affected the collections, generating up to 75% breakage. Such a high percentage of anthropogenic breakage can lead to misinterpretation, for example, as in the case of determining the number of individuals collected (MNI; Badgley 1986). The case of the specimens UFGRS-PV-0911-T, UFGRS-PV-0915-T, and UFGRS-PV-0933-T represent a good example of this, both the former and the latter being catalogued as “non-mammalian cynodont lower jaw fragment”, while they actually are complementary parts of the same bone element, now an almost complete lower jaw, representing just one and not two possible individuals. The absence of an individual dedicated to the care of the collections, in all its aspects, such as a curator, may have favoured phenomena such as displacement and mixture of bone fragments into different boxes. Although attention during the handling of fossils is always high, the absence of an official curator leads to an inevitable difficulty in respecting uniform rules and procedures (from storage and preparation, up to cataloguing and study of the specimens).
Analysing the curatorial attention paid to restoration for each institution with respect to anthropogenic breakage, our analyses highlight some differences between UFRGS, PUC-RS and MCN/SEMA-RS. If we do not consider that this same result can be affected by bias, due to the faunal content of the Schoenstatt and its differences in fossil abundance, the number of restored elements kept in the UFRGS palaeovertebrate collection indicates a cynodonts-orientated-tendency carried out by UFRGS over time, while the researcher of the other two institutions would seem to have adopted a more equilibrated way to approach the post-collection breaking problem. At the same time, however, both UFRGS and MCN/SEMA-RS researchers apply some “discrimination and selection” between cranial vs. post-cranial/others highlighting the possibility of the existence of the “craniocentrism”, a problem already known in the palaeontological literature (see Marjanović and Laurin 2019) affecting the former more than the latter.
As previously mentioned, the numbers here reported, and so all the specimens and the bone elements that constitute them, need further and deeper revision, in order to reduce both the number of anthropogenically broken elements and the effects linked to sullegic and trephic factors. New revisions of this fossil material, will certainly lead to a further decreasing of the anatomically identified broken elements, as well as of the indeterminate one, too. Furthermore, several indeterminate bone fragments, as well as bones previously attributed to indeterminate cynodonts, were recognized as belonging to other taxa. As an additional consequence, this will contribute to a more refined and detailed knowledge about the biodiversity contained in the outcrop and, as a corollary, of the entire Santacruzodon AZ.
Acknowledgements
For access to collections, we acknowledge Marco Brandalise de Andrade (at that time at MCP/PUC-RS). Heitor Francischini, Voltaire Paes-Neto, André Barcelos (all UFRGS), Jorge Ferigolo (MCN-RS/SEMA), Marina Soares (Museu Nacional/Universidade Federal do Rio de Janeiro, Brazil), Paula Dentzien-Dias (Universidade Federal do Rio Grande, Brazil), and Agustín Martinelli (CONICET/Museo Argentino de Ciencias Naturales “Bernardino Rivadavia”, Argentina) are strongly acknowledged for interesting discussion about the topics here discussed. Special thanks to Tomaz Melo (UFRGS) for help in the laboratory and for helpful discussions. We warmly thank the associate editors Eli Amson (Naturkunde Museum Stuttgart, Germany) and Daniel Barta (Oklahoma State University, USA), and the two reviewers Stephanie K. Drumheller-Horton (University of Tennessee, USA) and Jahn Hornung (Niedersächsisches Landesmuseum, Hannover, Germany) for their comments and suggestions, which helped us to greatly improve the manuscript. This work was supported by Conselho Nacional de Desenvolvimento Científico e Tecnológico-CNPq via a grant awarded to Francesco Battista (CNPq proc.: 168678/2018-7), to AMR (CNPq proc.: 310023/2021-1), to FE (CNPq proc.: 404343/2021-0), and to CLS (CNPq proc.: 311251/2021-8), and by Fundação de Amparo à Pesquisa do Estado do Rio Grande do Sul-FAPERGS to Fernando Erthal (FAPERGS Proc.: 22/2551-0000639-6).
References
Abdala, F. and Ribeiro, A.M. 2003. A new traversodontid cynodont from the Santa Maria Formation (Ladinian–Carnian) of southern Brazil, with a phylogenetic analysis of Gondwanan traversodontids. Zoological Journal of the Linnean Society 139: 529–545. Crossref
Abdala, F. and Ribeiro, A.M. 2010. Distribution and diversity patterns of Triassic cynodonts (Therapsida, Cynodontia) in Gondwana. Palaeogeography, Palaeoclimatology, Palaeoecology 286: 202–217. Crossref
Abdala, F., Ribeiro, A.M., and Schultz, C.L. 2001. A rich cynodont fauna of Santa Cruz do Sul, Santa Maria Formation (Middle–Late Triassic), southern Brazil. Neues Jahrbuch für Geologie und Paläontologie – Monatshefte 2001 (11): 669–687. Crossref
Allmon, W.D. 2005. The importance of museum collections in paleobiology. Paleobiology 31: 1–5. Crossref
Alroy, J. 2010. Geographical, environmental and intrinsic biotic controls on Phanerozoic marine diversification. Palaeontology 53: 1211–1235. Crossref
Alroy, J., Marshall, C.R., Bambach, R.K., Bezusko, K., Foote, M., Fürsich, F.T., Hansen, T.A., Holland, S.M., Ivany, L.C., Jablonski, D., Jacobs, D.K., Jones, D.C., Kosnik, M.A., Lidgard, S., Low, S., Miller, A.I., Novack-Gottshall, P.M., Olszewski, T.D., Patzkowsky, M. E., Raup, D.M., Roy, K., Sepkoski, J.J., Sommers, M.G., Wagner, P.J., and Webber, A. 2001. Effects of sampling standardization on estimates of phanerozoic marine diversification. Proceedings of the National Academy of Sciences of the United States of America 98: 6261–6266. Crossref
Badgley, C.E. 1986. Counting individuals in mammalian fossil assemblages from fluvial environments. Palaios 1: 328–338. Crossref
Behrensmeyer, A.K. 1975. The taphonomy and paleoecology of Plio-Pleistocene vertebrate assemblage east of lake Rudolf, Kenya. Bulletin of The Museum of Comparative Zoology 146: 473–578.
Behrensmeyer, A.K., Kidwell, S.M., and Gastaldo, R.A. 2000. Taphonomy and paleobiology. Paleobiology 26: 103–147. Crossref
Benton, M.J., Forth, J., and Langer, M.C. 2014. Models for the rise of the dinosaurs. Current Biology 24: R87–R95. Crossref
Benton, M.J., Wills, M.A., and Hitchin, R. 2000. Quality of the fossil record through time. Nature 403: 534–537. Crossref
Bertoni-Machado, C. 2004. Concentrações fossilíferas controladas pelo nível freático: um modelo tafonômico para reconstruções paleoambientais. 88 pp. Master Thesis, Universidade Federal do Rio Grande do Sul, Porto Alegre.
Bertoni-Machado, C. and Holz, M. 2006. Biogenic fossil concentration in fluvial settings: An example of a cynodont taphocoenosis from the Middle Triassic of southern Brazil. Revista Brasileira de Paleontologia 9: 273–282. Crossref
Biro, P.A. and Dingemanse, N.J. 2009. Sampling bias resulting from animal personality. Trends in Ecology and Evolution 24: 66–67. Crossref
Clark, J. and Kietzke, K.K. 1967. Paleoecology of the lower Nodular Zone, Brule Formation, in the Big Badlands of South Dakota. In: J. Clark, J.R. Beerbower, and K.K. Kietzke (eds.), Oligocene Sedimentation, Stratigraphy, Paleoecology and Paleoclimatology in the Big Badlands of South Dakota. Fieldiana Geology Memoirs 5: 111–155. Crossref
Cooper, N., Bond, A.L., Davis, J.L., Miguez, R.P., Tomsett, L., and Helgen, K.M. 2019. Sex biases in bird and mammal natural history collections. Proceedings of the Royal Society B: Biological Sciences 286: 20192025. Crossref
Corecco, L., Pereira, V.P., Soares, M.B., and Schultz, C.L. 2020. Geochemical study of the vertebrate assemblage zones of the Santa Maria Supersequence (Middle to Late Triassic), Paraná Basin, Brazil. Brazilian Journal of Geology 50: e20200014. Crossref
Crampton, J.S., Beu, A.G., Cooper, R.A., Jones, C.M., Marshall, B., and Maxwell, P.A. 2003. Estimating the rock volume bias in paleobiodiversity studies. Science 301: 358–360.Crossref
Dal Corso, J., Ruffell, A., and Preto, N. 2018. The Carnian Pluvial Episode (Late Triassic): New insights into this important time of global environmental and biological change. Journal of the Geological Society 175: 986–988. Crossref
Dean, C.D., Chiarenza, A.A., and Maidment, S.C.R. 2020. Formation binning: a new method for increased temporal resolution in regional studies, applied to the Late Cretaceous dinosaur fossil record of North America. Palaeontology 63: 881–901. Crossref
Dibble, H.L., Raczek, T.P., and McPherron, S.P. 2005. Excavator bias at the site of Pech de l’Azé IV, France. Journal of Field Archaeology 30: 317–328. Crossref
Flessa, K.W., Kowalewski, M., and Walker, S.E. 1992. Post-collection taphonomy; shell destruction and the Chevrolet. Palaios 7: 553–554. Crossref
Gifford-Gonzalez, D. 2018. An Introduction to Zooarchaeology. 604 pp. Springer, Cham. Crossref
Hammer, Ø., Harper, D.A.T., and Ryan, P.D. 2001. PAST: paleontological statistics software package for education and data analysis. Palaeontologia Electronica 4: 1–9.
Hijmans, R.J., Garrett, K.A., Huamán, Z., Zhang, D.P., Schreuder, M., and Bonierbale, M. 2000. Assessing the geographic representativeness of genebank collections: The case of Bolivian wild potatoes. Conservation Biology 14: 1755–1765. Crossref
Holz, M. and Schultz, C.L. 1998. Taphonomy of the south Brazilian Triassic herpetofauna: Fossilization mode and implications for morphological studies. Lethaia 31: 335–345. Crossref
Holz, M., França, A.B., Souza, P.A., Iannuzzi, R., and Rohn, R. 2010. A stratigraphic chart of the Late Carboniferous/Permian succession of the eastern border of the Paraná Basin, Brazil, South America. Journal of South American Earth Sciences 29: 381–399. Crossref
Horn, B.L.D., Goldberg, K., and Schultz, C.L. 2018. Interpretation of massive sandstones in ephemeral fluvial settings: A case study from the Upper Candelária Sequence (Upper Triassic, Paraná Basin, Brazil). Journal of South American Earth Sciences 81: 108–121. Crossref
Horn, B.L.D., Melo, T.P., Schultz, C.L., Philipp, R.P., Kloss, H.P., and Goldberg, K. 2014. A new third-order sequence stratigraphic framework applied to the Triassic of the Paraná Basin, Rio Grande do Sul, Brazil, based on structural, stratigraphic and paleontological data. Journal of South American Earth Sciences 55: 123–132. Crossref
Hunter, A. and Donovan, S. 2005. Field sampling bias, museum collections and completeness of the fossil record. Lethaia 38: 305–314. Crossref
Kidwell, S.M. and Behrensmeyer, A.K. 1988. Overview: Ecological and evolutionary implications of taphonomic processes. Palaeogeography, Palaeoclimatology, Palaeoecology 63: 1–13. Crossref
Kidwell, S.M. and Holland, S.M. 2002. The quality of the fossil record: Implications for evolutionary analyses. Annual Review of Ecology and Systematics 33: 561–588. Crossref
Kowalewski, M. and Hoffmeister, A.P. 2003. Sieves and fossils: Effects of mesh size on paleontological patterns. Palaios 18: 460–469. Crossref
Mancuso, A.C., Horn, B.L.D., Benavente, C.A., Schultz, C.L., and Irmis, R.B. 2021. The paleoclimatic context for South American Triassic vertebrate evolution. Journal of South American Earth Sciences 110: 103321. Crossref
Mannion, P.D., Benson, R.B.J., and Butler, R.J. 2013. Vertebrate palaeobiodiversity patterns and the impact of sampling bias. Palaeogeography, Palaeoclimatology, Palaeoecology 372: 1–4. Crossref
Marjanović, D. and Laurin, M. 2019. Phylogeny of Paleozoic limbed vertebrates reassessed through revision and expansion of the largest published relevant data matrix. PeerJ 6: e5565. Crossref
Martinelli, A.G., Kammerer, C.F., Melo, T.P., Paes-Neto, V.D., Ribeiro, A.M., Da-Rosa, Á.A.S., Schultz, C.L., and Soares, M.B. 2017. The African cynodont Aleodon (Cynodontia, Probainognathia) in the Triassic of southern Brazil and its biostratigraphic significance. PLoS ONE 12: e0177948. Crossref
McGowan, A.J. and Smith, A.B. 2011. Comparing the geological and fossil records: Implications for biodiversity studies. Geological Society, London, Special Publications 358: 1–254. Crossref
Melo, T.P., Abdala, F., and Soares, M.B. 2015. The Malagasy cynodont Menadon besairiei (Cynodontia; Traversodontidae) in the Middle–Upper Triassic of Brazil. Journal of Vertebrate Paleontology 35: e1002562. Crossref
Melo, T.P., Martinelli, A.G., and Soares, M.B. 2022. New occurrences of massetognathine traversodontids and chiniquodontids (Synapsida, Cynodontia) from the early Late Triassic Santacruzodon Assemblage Zone (Santa Maria Supersequence, southern Brazil): Geographic and biostratigraphic implications. Journal of South American Earth Sciences 115: 103757. Crossref
Milani, E.J. 1997. Evolução tectono-estratigráfica da Bacia do Paraná e seu relacionamento com a geodinâmica fanerozóica do Gondwana Sul-Ocidental. 255 pp. Ph.D. Thesis, Universidade Federal do Rio Grande do Sul, Porto Alegre.
Milani, E.J. and Ramos, V.A. 1998. Orogenias paleozoicas no domínio sul-ocidental do Gondwana e os cicios de subsidência da bacia do Paraná. Revista Brasileira de Geociências 28: 473–484. Crossref
Milani, E.J., Melo, H.J.G., Souza, P.A., Fernandes, L.A., and França, A.B. 2007. Bacia do Paraná. Boletim de Geociências Da Petrobras 15: 265–287.
Philipp, R.P., Schultz, C.L., Kloss, H.P., Horn, B.L.D., Soares, M.B., and Basei, M.A.S. 2018. Middle Triassic SW Gondwana paleogeography and sedimentary dispersal revealed by integration of stratigraphy and U-Pb zircon analysis: The Santa Cruz Sequence, Paraná Basin, Brazil. Journal of South American Earth Sciences 88: 216–237. Crossref
Ponder, W.F., Carter, G.A., Flemons, P., and Chapman, R.R. 2001. Evaluation of museum collection data for use in biodiversity assessment. Conservation Biology 15: 648–657. Crossref
Qin, J. 2017. Biased Sampling, Over-identified Parameter Problems and Beyond. 624 pp. Springer, Singapore, Crossref
Raup, D.M. 1972. Taxonomic diversity during the phanerozoic. Science 177: 1065–1071. Crossref
Reichel, M., Schultz, C.L., and Pereira, V.P. 2005. Diagenetic pattern of vertebrate fossils from the Traversodontidae Biozone, Santa Maria Formation (Triassic), southern Brazil. Revista Brasileira de Paleontologia 8: 173–180. Crossref
Schmitt, M.R., Martinelli, A.G., Melo, T.P., and Soares, M.B. 2019. On the occurrence of the traversodontid Massetognathus ochagaviae (Synapsida, Cynodontia) in the early late Triassic Santacruzodon Assemblage Zone (Santa Maria Supersequence, southern Brazil): Taxonomic and biostratigraphic implications. Journal of South American Earth Sciences 93: 36–50. Crossref
Schultz, C.L., Martinelli, A.G., Soares, M.B., Pinheiro, F.L., Kerber, L., Horn, B.L.D., Pretto, F.A., Müller, R.T., and Melo, T.P. 2020. Triassic faunal successions of the Paraná Basin, southern Brazil. Journal of South American Earth Sciences 104: 102846. Crossref
Sepkoski, J.J., Bambach, R.K., Raup, D.M., and Valentine, J.M. 1981. Phanerozoic marine diversity and the fossil record. Nature 293: 435–437. Crossref
Signor, P.W., and Lipps, J.H. 1982. Sampling bias, gradual extinction patterns and catastrophes in the fossil record. Geological Society of America Special Papers 190: 291–296. Crossref
Simms, M.J. and Ruffell, A.H. 1989. Synchroneity of climatic change and extinctions in the Late Triassic. Geology 17: 265–268. Crossref
Simms, M.J. and Ruffell, A.H. 1990. Climatic and biotic change in the late Triassic. Journal of the Geological Society 147: 321–327. Crossref
Smith, A.B. and McGowan, A.J. 2011. The ties linking rock and fossil records and why they are important for palaeobiodiversity studies. In: A.J. McGowan and A.B. Smith (eds.), Comparing the Geological and Fossil Records: Implications for Biodiversity Studies. Geological Society London Special Publications 358 (1): 1–5. Crossref
Soares, M.B., Abdala, F., and Bertoni-Machado, C. 2011. A sectorial toothed cynodont (Therapsida) from the Triassic Santa Cruz do Sul fauna, Santa Maria Formation, Southern Brazil. Geodiversitas 33: 265–278. Crossref
Stuber, E.F., Araya-Ajoy, Y.G., Mathot, K.J., Mutzel, A., Nicolaus, M., Wijmenga, J.J., Mueller, J.C., and Dingemanse, N.J. 2013. Slow explorers take less risk: A problem of sampling bias in ecological studies. Behavioral Ecology 24: 1092–1098. Crossref
Tang, C.M. 2000. Ugly fossil syndrome. Palaios 15: 175–176. Crossref
Voorhies, M.R. 1969. Taphonomy and population dynamics of an early Pliocene vertebrate fauna, Knox County, Nebraska. Contribution to Geology 1: 1–69. Crossref
Walker, S.E. 1989. Hermit crabs as taphonomic agents. Palaios 4: 439–452. Crossref
Whitaker, A.F. and Kimmig, J. 2020. Anthropologically introduced biases in natural history collections, with a case study on the invertebrate paleontology collections from the middle Cambrian Spence Shale Lagerstätte. Palaeontologia Electronica 23: a58. Crossref
Wolff, R.G. 1975. Sampling and sample size in ecological analyses of fossil mammals. Paleobiology 1: 195–204. Crossref
Wosik, M. and Guenther, M.F. 2016. Examination of a historic collection of isolated cranial and appendicular hadrosaurid material from the lower Kirtland Formation of the San Juan Basin, New Mexico. Journal of Paleontology 90: 763–770. Crossref
Zerfass, H., Chemale, F., and Lavina, E. 2005. Tectonic control of the Triassic Santa Maria supersequence of the Paraná Basin, southernmost Brazil, and its correlation to the Waterberg Basin, Namibia. Gondwana Research 8: 163–176. Crossref
Zerfass, H., Chemale, F., Schultz, C.L., and Lavina, E. 2004. Tectonics and sedimentation in Southern South America during Triassic. Sedimentary Geology 166: 265–292. Crossref
Zerfass, H., Lavina, E.L., Schultz, C.L., Garcia, A.J.V., Faccini, U.F., and Chemale, F. 2003. Sequence stratigraphy of continental Triassic strata of Southernmost Brazil: A contribution to Southwestern Gondwana palaeogeography and palaeoclimate. Sedimentary Geology 161: 85–105. Crossref
Acta Palaeontol. Pol. 68 (2): 359–372, 2023
https://doi.org/10.4202/app.01050.2022